Уровень техники, к которой относится изобретение
Настоящее изобретение касается новых рекомбинантных вакцин, обеспечивающих защитный иммунитет, в особенности против туберкулеза.
Уровень техники
Туберкулез (ТВ), который вызывается микроорганизмом Mycobacterium tuberculosis, остается существенной глобальной проблемой. Подсчитано, что каждый третий представитель мирового населения инфицирован M.tuberculosis (Kochi, 1991). Во многих странах единственным способом контроля заболеваемости ТВ являлась вакцинация с использованием бациллы M.bovis Calmette-Guerin (BCG). Однако средняя общая эффективность вакцины BCG против ТВ составляет примерно 50%, и значения ее сильно варьируются в пределах от 0% до 80% в разных конкретных случаях (Roche et al., 1995). Таким образом, BCG должна быть улучшена, например, посредством генной инженерии с целью получения вакцины, позволяющей лучше контролировать заболеваемость ТВ (Murray et al., 1996; Hess and Kaufmann, 1993). Такая широко распространенная опасность, как множественная лекарственная устойчивость штаммов M.tuberculosis, дополнительно подчеркивает срочную необходимость в новых ТВ вакцинах (Grange, 1996).
M.tuberculosis относится к группе внутриклеточных бактерий, которые размножаются внутри фагоцитарных вакуолей покоящихся макрофагов, таким образом, защита против ТВ зависит от Т-клеточного иммунитета (Kaufmann, 1993). Однако некоторые исследования, проведенные на мышах и человеке, показали, что Mycobacteria стимулируют антиген-специфичные CD4 и CD8 Т-клетки, несущие главный комплекс гистосовместимости (МНС) класса II или класса I по рестрикции соответственно (Kaufmann, 1993).
Важная роль CD8 Т-клеток, несущих МНС класса I по рестрикции, была убедительно продемонстрирована посредством неудавшегося эксперимента по контролю экспериментального инфицирования микроорганизмами M.tuberculosis мышей, дефицитных по β2-микроглобулину (β2m) (Flynn et al., 1993). Поскольку эти мутантные мыши не имеют МНС класса I, функциональные CD8 Т-клетки не могут развиваться. В отличие от инфекции M.tuberculosis β2m-дефицитные мыши способны усваивать определенные инфекционные дозы вакцины на основе штамма BCG (Flynn et al., 1993; Ladel et al., 1995). Более того, BCG вакцинация β2m-дефицитных мышей продлевала срок их выживания после последующего инфицирования M.tuberculosis, когда BCG-иммунизированные C57BL/6 демонстрировали устойчивость к ТВ (Flynn et al., 1993). Такая различная CD8 Т-клеточная зависимость между M.tuberculosis и BCG может быть объяснена следующим: антигены M.tuberculosis имеют лучший доступ в цитоплазму, нежели антигены из BCG, что приводит к более выраженной представленности МНС класса I (Hess and Kaufmann, 1993). Следовательно, более эффективный CD8 Т-клеточный ответ генерируется M.tuberculosis. Данное предположение было позже подтверждено повышенной представленностью МНС класса I в случае не относящегося к делу антигена, овальбумина, совместной с M.tuberculosis, лучше, чем с BCG, инфекцией антиген-представляющих клеток (АРС) (Mazzaccaro et al., 1996).
Секретируемые белки M.tuberculosis включают ценный источник антигенов для представления МНС класса I. Недавно, ДНК вакцина, кодирующая секретируемый антиген Ag85A, вызвала ответы CD8 Т-клеток, несущих МНС класса I по рестрикции у мышей, которые могут способствовать защите от ТВ (Huygen et al., 1996). Вообще, накопленные данные показывают, что иммунизация секретируемыми белковыми антигенами M.tuberculosis индуцирует некоторую защиту от ТВ у морских свинок и мышей (Horwitz et al., 1995; Andersen, 1994). Следовательно, важной задачей на пути к разработке улучшенных ТВ вакцин на основе BCG, является увеличение доступности цитоплазмы инфицированных АРС для секретируемых, BCG-специфичных антигенов. Последующая доставка пептидов, являющихся производными этих секретируемых белков, в последовательность событий представления МНС класса I может привести к усилению уже существующего BCG-специфичного иммунного ответа, предупреждающего заболевание ТВ.
Высвобождение из фаголизосом L.monocytogenes представляет собой уникальный механизм усиления представления листериальных антигенов МНС класса I (Berche et al., 1987; Portnoy et al., 1988). Листериолизин (Hlу), порообразующий сульфгидрил-активируемый цитолизин, необходим для высвобождения микроорганизмов L.monocytogenes из фаголизосомальных вакуолей в цитозоль клетки хозяина (Gaillard et al., 1987; Portnoy et al., 1988). Данная функция высвобождения была недавно передана штамму Bacillus subtilis и ослабленному в своей вирулентности штамму Salmonella ssp. (Bielecki et al., 1991; Gentschev et al., 1995; Hess and Kaufmann, 1997). Экспрессия Hlу мутантным штаммом неспорообразующих В. subtilis или Salmonella ssp. приводила, в результате, к высвобождению бактерий из фаголизосом в цитозоль J774 макрофаг-подобных клеток (Bielecki et al., 1991; Gentschev et al., 1995; Hess and Kaufmann, 1997).
WO 99/101496 и Hess et al. (1998) описывают рекомбинантные штаммы Mycobacterium bovis, которые секретируют биологически активные листериолизин-слитые белки. Было показано, что данные штаммы M.bovis являются эффективными вакцинами против ТВ в случае нескольких животных моделей.
Раскрытие изобретения
В соответствии с настоящим изобретением Hlу был экспрессирован в дефицитных по уреазе штаммах BCG. Эти дефицитные по уреазе штаммы BCG демонстрируют повышенную активность Hlу в фагосомах и, в свою очередь, повышенную эффективность порообразования в мембранах эндосом, что свидетельствует об их превосходной эффективности иммунной защиты. Кроме того, дефицитные по уреазе штаммы BCG-Hlу вовлечены в процессы апоптоза, которые могут также вносить свой вклад в наблюдаемое усиление иммунной защиты. Таким образом, имеются широкие возможности по созданию вакцин на основе дефицитных по уреазе штаммов BCG. Кроме того, было неожиданно обнаружено, что дефицитные по уреазе штаммы BCG являются более безопасными по сравнению с родительскими штаммами BCG и, таким образом, могут применяться, в частности, для вакцинации пациентов, страдающих иммунодефицитом.
Первым аспектом настоящего изобретения является бактериальная клетка, а именно дефицитная по уреазе клетка Mycobacterium, содержащая рекомбинантную молекулу нуклеиновой кислоты, которая кодирует слитый полипептид, включающий (а) как минимум один домен из полипептида, в котором данный полипептидный домен способен вызывать иммунный ответ у млекопитающих, и (б) домен, ответственный за высвобождение из фаголизосом. Предпочтительно, чтобы клетка была способна экспрессировать молекулу нуклеиновой кислоты, являющуюся предметом настоящего изобретения. Более предпочтительно, чтобы клетка была способна секретировать слитый полипептид и/или предоставлять его в форме, пригодной для распознавания антигена МНС класса I по рестрикции.
Бактериальная клетка, являющаяся предметом настоящего изобретения, - это клетка, дефицитная по уреазе, например грамотрицательная либо грамположительная бактериальная клетка, предпочтительно клетка Mycobacterium. Дефицитность по уреазе может быть достигнута посредством частичной, либо полной инактивации одной или нескольких клеточных молекул нуклеиновой кислоты, кодирующих субъединицы уреазы, в частности ureA, кодирующей А субъединицу уреазы, ureB, кодирующей В субъединицу уреазы, и/или ureC, кодирующей С субъединицу уреазы. Последовательности ureA, ureB и ureC у Mycobacteria, в частности у M.bovis и M.tuberculosis, а также кодируемые ими белки описаны в работах Reyrat et al. (1995) и Clemens et al. (1995), которые включены в данное описание во всей своей полноте путем отсылки.
Предпочтительно, когда дефицитный по уреазе бактериальный штамм получен посредством делеций и/или вставок одного либо нескольких нуклеотидов в последовательности нуклеиновой кислоты, которые кодируют субъединицы уреазы, и/или в последовательности, контролирующие их экспрессию. Делеции и/или вставки могут быть получены в результате гомологичной рекомбинации, вставки транспозона либо применения других подходящих методов.
В рамках особо предпочтительного воплощения настоящего изобретения инактивирована последовательность ureC, например, посредством конструирования вектора-самоубийцы, включающего ген ureC, нарушенный с помощью селективного маркерного гена, трансформации клетки мишени данным вектором и скринингом с целью отбора положительных по селективному маркеру клеток, имеющих негативный по уреазе фенотип, как описано в Reyrat et al. (1995).
Клетка, являющаяся предметом настоящего изобретения, - это предпочтительно клетка M.bovis, клетка M.tuberculosis, в частности ослабленная в своей вирулентности клетка M.tuberculosis, либо другая Mycobacteria, например M.microti, M.smegmatis, M.canettii, M.marinum или M.fortuitum или Mycobacteria, как описано в Reyrat et al. (1995).
Клетка Mycobacterium, являющаяся предметом настоящего изобретения, содержит рекомбинантную молекулу нуклеиновой кислоты, например молекулу нуклеиновой кислоты, SEQ ID No.1. Данная молекула нуклеиновой кислоты включает последовательность, кодирующую сигнальный пептид (нуклеотид 1-120), последовательность, кодирующую иммуногенный домен (нуклеотид 121-153), последовательность, кодирующую пептидный линкер (нуклеотид 154-210), последовательность, кодирующую фаголизосомный домен (нуклеотид 211-1722), еще одну последовательность, кодирующую пептидный линкер (нуклеотид 1723-1800), и последовательность, кодирующую случайный пептид (нуклеотид 1801-1870). Соответствующая аминокислотная последовательность представлена в SEQ ID No.2.
Нуклеиновая кислота содержит как минимум один иммуногенный домен из полипептида. Иммуногенный домен может быть выделен из организма рода Mycobacterium, предпочтительно из Mycobacterium tuberculosis или из Mycobacterium bovis. Этот домен имеет длину как минимум 6 или предпочтительно как минимум 8 аминокислот. Иммуногенный домен - это предпочтительно часть нативного полипептида Mycobacterium. Тем не менее, в объем настоящего изобретения включен также модифицированный иммуногенный домен, полученный из нативного иммуногенного домена посредством замены, делеции и/или вставки одной либо нескольких аминокислот.
Иммуногенный домен, однако, не ограничивается антигеном Mycobacterium и может быть выбран из числа аутоантигенов, раковых антигенов и антигенов патогена, таких как вирусные антигены, антигены паразитов, антигены бактерий вообще и их иммуногенные фрагменты. Специфические примеры подходящих раковых антигенов - это антигены рака человека, такие как продукт гена супрессора опухоли р53 (Houbiers et al., 1993) и антигены дифференциации меланоцитов, например Melan-A/MART-1 и gp100 (van Elsas et al., 1996). Специфические примеры подходящих вирусных антигенов - это антигены вируса опухоли человека, такие как антигены вируса палилломы человека, например антигены Е6 и Е7 (Bosch et al., 1991); антигены вируса гриппа, например нуклеопротеин вируса гриппа (Matsui et al., 1995; Fu et al., 1997); или антигены ретровирусов, такие как HIV антигены, например HIV-1 антигены р17, р24, RT и Env (Harrer et al., 1996; Haas et al., 1996). Конкретные примеры подходящих антигенов паразитов - это антигены Plasmodium, такие как антиген печеночной стадии (LSA-1), белок круглого спорозоита (CS (circumsporozoite) или аллельные варианты ср26 или ср29); тромбоспондин - связанный безымянный белок (TRAP), треонин- и аспарагин-богатый белок спорозоита (STARP) Plasmodium falciparum (Aidoo et al., 1995) и антигены Toxoplasma, такие как р30 из Toxoplasma gondii (Khan et al., 1991; Bulowand Boothroyd, 1991). Конкретные примеры подходящих бактериальных антигенов - это антигены Legionella, такие как Главный белок-помощник (Major secretary protein) из Legionella pneumophila (Blander and Horwitz, 1991).
Иммуногенный домен способен вызывать иммунный ответ в млекопитающих. Данный иммунный ответ может быть опосредован В-клеточным иммунным ответом. Предпочтительно, тем не менее, если иммуногенный домен способен вызывать опосредованный Т-клетками иммунный ответ, более предпочтительно CD8 Т-клеточный ответ МНС класса I по рестрикции.
Домен, способный вызывать иммунный ответ, наиболее предпочтительно выбирается из иммуногенных пептидов либо полипептидов из M.bovis или M.tuberculosis, или из их иммуногенных фрагментов. Специфические примеры подходящих антигенов - это Ag85B (р30) из M.tuberculosis (Harth et al., 1996), Ag85B (α-antigen) из BCG M.bovis (Matsuo et al., 1988), Ag85A из M.tuberculosis (Huygen et al., 1996) и ESAT-6 из M.tuberculosis (Sorensen et al., 1996, Harboe et al., 1996 and Andersen et al., 1995). Более предпочтительно, когда иммуногенный домен получен из антигена Ag85B. Наиболее предпочтительно, когда иммуногенный домен содержит последовательность из аминокислот с 41 до 51 в последовательности SEQ ID No.2.
В соответствии с настоящим изобретением рекомбинантная молекула нуклеиновой кислоты также включает домен выхода из фаголизосом, то есть полипептидный домен, который осуществляет высвобождение слитого полипептида из фаголизосом в цитозоль клетки млекопитающего. Предпочтительно, когда домен выхода из фаголизосом является доменом выхода из фаголизосом Listeria, который описан в US 5733151, который включен в данное описание во всей своей полноте путем отсылки. Более предпочтительно, когда домен выхода из фаголизосом выделен из организма L.monocytogenes. Наиболее предпочтительно, когда домен выхода из фаголизосом кодируется молекулой нуклеиновой кислоты, выбранной из: (а) последовательности нуклеотидов, включающей нуклеотиды 211-1722, как показано в SEQ ID No.1, (b) последовательности нуклеотидов, которая кодирует ту же аминокислотную последовательность, что и последовательность из (а), и (с) последовательности нуклеотидов, которая в жестких условиях гибридизуется с последовательностями из (а) или (b).
Помимо последовательности нуклеотидов, представленной как SEQ ID No.1, настоящее изобретение также включает в себя гибридизующиеся с ней последовательности нуклеиновых кислот. В рамках настоящего изобретения термин "гибридизация" используется, как это определено в Sambrook et al. (Molecular Cloning. A laboratory manual, Cold Spring Harbor Laboratory Press (1989), 1.101-1.104). В соответствии с настоящим изобретением термин "гибридизация" используется в том случае, когда положительный гибридизационный сигнал может все еще наблюдаться после отмывки в течение одного часа с использованием 1 X SSC и 0,1% SDS при 55°С, предпочтительно при 62°С и более предпочтительно при 68°С, и особенно в случае 1 часа в 0,2 X SSC и 0,1% SDS при 55°С, предпочтительно при 62°С и более предпочтительно при 68°С. Последовательность, гибридизующаяся при таких условиях отмывки с последовательностью нуклеотидов, представленной как SEQ ID No.1, является последовательностью нуклеотидов, кодирующей домен выхода из фаголизосом предпочтительной для настоящего изобретения.
Последовательность нуклеотидов, кодирующая домен выхода из фаголизосом, как описано выше, может быть прямо получена из организма Listeria, либо из любого рекомбинантного источника, например рекомбинантных клеток E.coli, содержащих соответствующую молекулу нуклеиновой кислоты Listeria или ее вариант, как описано выше.
Предпочтительно, когда рекомбинантная молекула нуклеиновой кислоты, кодирующая слитый полипептид, включает последовательность, кодирующую сигнальный пептид. Более предпочтительно, когда сигнальная последовательность является сигнальной последовательностью активной в Mycobacteria, предпочтительно в M.bovis, например нативной сигнальной последовательностью M.bovis. Предпочтительным примером подходящей сигнальной последовательности является последовательность нуклеотидов, кодирующая сигнальный пептид Ag85B, которая представлена в SEQ ID No.1 с нуклеотида 1 по 120.
Кроме того, предпочтительно, чтобы пептидный линкер был введен между иммуногенным доменом и доменом выхода из фаголизосом. Предпочтительно, когда такой пептидный линкер имеет длину от 5 до 50 аминокислот. Более предпочтительно, когда последовательность кодирует линкер, соответствующий показанному в SEQ ID No.1 с нуклеотида 154 по 210, либо когда последовательность соответствует ей с учетом вырождения генетического кода.
Нуклеиновая кислота может быть расположена в рекомбинантном векторе. Предпочтительно, когда рекомбинантный вектор является прокариотическим вектором, то есть вектором, содержащим необходимые элементы для репликации и/или интеграции в геном в клетке прокариот. Предпочтительно, когда рекомбинантный вектор несет молекулу нуклеиновой кислоты, являющуюся предметом настоящего изобретения, действенно связанную с последовательностью, контролирующей экспрессию. Предпочтительно, когда контролирующая экспрессию последовательность является контролирующей экспрессию последовательностью активной в Mycobacteria, в частности в M.bovis. Вектор может являться внехромосомным вектором, либо вектором, пригодным для интеграции в хромосому. Примеры таких векторов хорошо известны профессионалам в данной области и, кроме того, описаны в работах Sambrook et al.
Следующим аспектом настоящего изобретения являются дефицитные по уреазе клетки бактерий, например клетки Mycobacterium, предпочтительно клетки M.bovis, которые содержат хотя бы одну молекулу нуклеиновой кислоты, кодирующую пептид либо полипептид выхода из фаголизосом. Даже в том случае, когда пептид либо полипептид выхода из фаголизосом не является слитым с антигеном, обнаруживается удивительное улучшение иммуногенных свойств.
Рекомбинантная бактериальная клетка, представленная в соответствии с данным дополнительным аспектом настоящего изобретения, может содержать как минимум один дополнительный рекомбинант, например молекулу инородной нуклеиновой кислоты, кодирующую пептид либо полипептид, способный вызывать иммунный ответ в млекопитающих. Такой дополнительный иммуногенный пептид либо полипептид может быть отобран из числа антигенов Mycobacterium или, в широком смысле, из аутоантигенов, опухолевых антигенов, антигенов патогенов и их иммуногенных фрагментов. Молекула нуклеиновой кислоты, кодирующая дополнительный пептид либо полипептид, может быть расположена в том же векторе, что и слитый ген. Однако она также может располагаться, к примеру, на другой плазмиде, независимо от слитого гена, или быть интегрированной в хромосому.
Было неожиданно обнаружено, что соответствующая настоящему изобретению клетка Mycobacterium имеет такую внутриклеточную стабильность в зараженных клетках, например макрофагах, которая одинакова или меньше, нежели внутриклеточная стабильность соответствующей нативной клетки Mycobacterium, не содержащей рекомбинантную молекулу нуклеиновой кислоты.
Настоящее изобретение также касается фармацевтической композиции, содержащей в качестве активного агента описанную выше клетку, необязательно вместе с фармацевтически приемлемыми растворителями, носителями и адъювантами. Предпочтительно, когда композиция является живой вакциной, пригодной для введения млекопитающим, предпочтительно человеку. Фактически выбранный способ вакцинации зависит от выбора вакцинного вектора. Введение может осуществляться посредством введения единичной дозы либо повторяющихся доз через определенные интервалы времени. Подходящая дозировка зависит от различных параметров, таких как вакцинный вектор сам по себе, или способ введения. Может быть выбран способ введения через поверхность слизистой оболочки (например, глазной, интраназальный, оральный, желудочный, кишечный, ректальный, вагинальный или через мочевые пути), либо по парентеральному пути (например, подкожно, внутрикожно, внутримышечно, внутривенно или внутрибрюшинно).
Кроме того, настоящее изобретение касается способа приготовления рекомбинантных клеток бактерий, как описано выше. В соответствии с первым аспектом настоящего изобретения данный способ включает такие стадии как (i) получение дефицитной по уреазе бактериальной клетки, в частности клетки Mycobacterium, (ii) вставка рекомбинантной молекулы нуклеиновой кислоты в эту бактериальную клетку, причем данная молекула нуклеиновой кислоты кодирует слитый полипептид, включающий в себя (а) как минимум один домен из полипептида, в котором этот домен способен вызывать иммунный ответ в клетках млекопитающих и (б) домен выхода из фаголизосом, и (iii) культивирование клеток, полученных в соответствии со стадией (ii) при подходящих условиях. Предпочтительно, когда получена клетка, которая способна экспрессировать описанную молекулу нуклеиновой кислоты. Более предпочтительно, когда клетка является клеткой M.bovis.
В соответствии со следующим аспектом настоящего изобретения данный способ включает в себя такие стадии, как (i) получение дефицитной по уреазе бактериальной клетки, в частности клетки Mycobacterium, (ii) вставка рекомбинантной молекулы нуклеиновой кислоты в эту бактериальную клетку, причем данная молекула нуклеиновой кислоты кодирует пептид либо полипептид выхода из фаголизосом, и (iii) культивирование клеток, полученных в соответствии со стадией (ii) при подходящих условиях.
При желании, способ, являющийся предметом настоящего изобретения, включает в себя вставку в бактериальную клетку как минимум одной дополнительной рекомбинантной молекулы нуклеиновой кислоты, причем данная молекула нуклеиновой кислоты кодирует пептид либо полипептид, способный вызывать иммунный ответ у млекопитающих.
Наконец, данное изобретение касается способа приготовления живой вакцины, содержащей разработанные рекомбинантные клетки в фармацевтически эффективном количестве вместе с фармацевтически приемлемыми растворителями, носителями и/или адъювантами.
Благодаря высокой безопасности дефицитных по уреазе бактериальных клеток, которая была продемонстрирована на двух различных животных моделях (Пример 3), живая вакцина, являющаяся объектом настоящего изобретения, может быть в отдельных случаях пригодной для введения испытуемым с иммунодефицитом, например испытуемым, страдающим от инфекции или субъектам, подверженным действию иммуносупрессорных препаратов. В особо предпочтительном воплощении живая вакцина, являющаяся объектом настоящего изобретения, используется как туберкулезная вакцина для испытуемых с иммунодефицитом.
В следующем предпочтительном воплощении живая вакцина используется как опухолевая вакцина, например, как вакцина против поверхностного рака мочевого пузыря. В следующем еще более предпочтительном воплощении настоящего изобретения живая вакцина используется в области ветеринарии, например, как вакцина против листериоза, туберкулида или бычьего туберкулеза.
Настоящее изобретение будет далее проиллюстрировано нижеследующими фигурами и списком последовательностей.
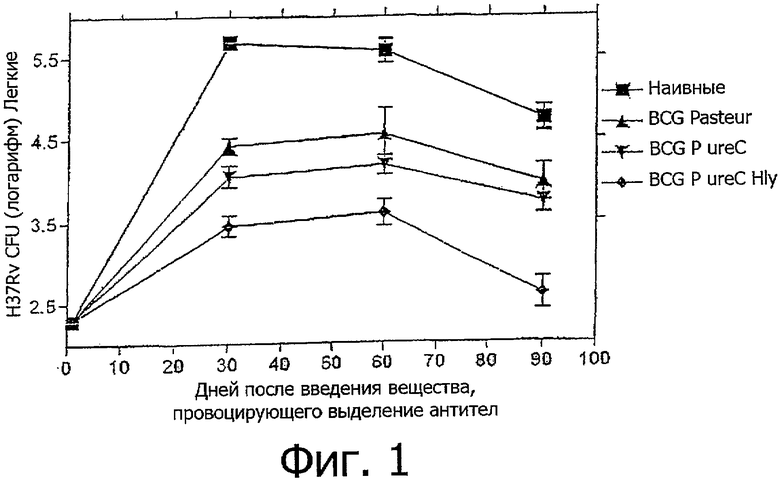
Фигура 1: демонстрирует защитную способность rBCG ureC Hlу в аэрозольной модели мышечного туберкулеза. BALB/c мыши были внутривенно иммунизованы с использованием 1×106 CFU (koe) rBCG ureC Hlу, BCG P ureC, либо нативной BCG "Pasteur". Спустя 120 дней после вакцинации в организм животных было введено H37Rv (200 организм/легкие) посредством аэрозоля. Содержание бактерий в инфицированных органах (селезенка и легкие) было оценено через 30, 60 и 90 дней после введения. Каждая точка соответствует 10 животным.
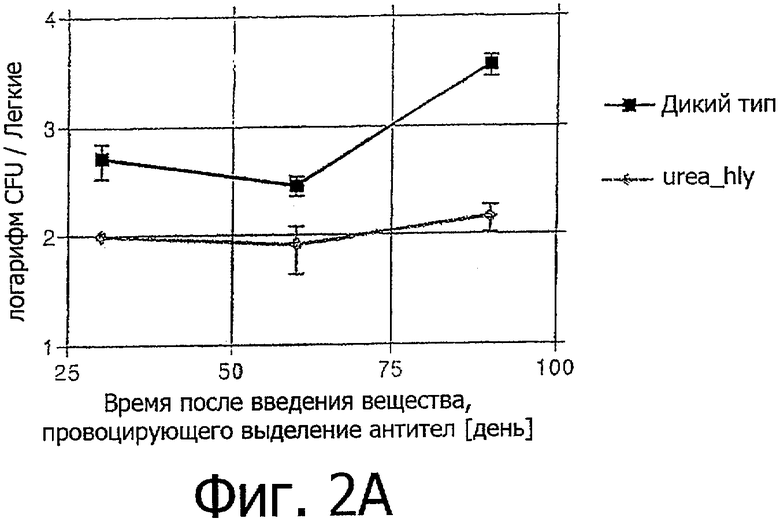
Фигура 2: демонстрирует число микроорганизмов в легких (Фигура 2А) либо в селезенке (Фигура 2B). Rag1-/- мыши были инфицированы родительским штаммом BCG (дикий тип), либо штаммом rBCG ureC Hlу (urea-Hlу). Содержание бактерий в инфицированном органе было оценено через 30, 60 и 90 дней после инфекции.
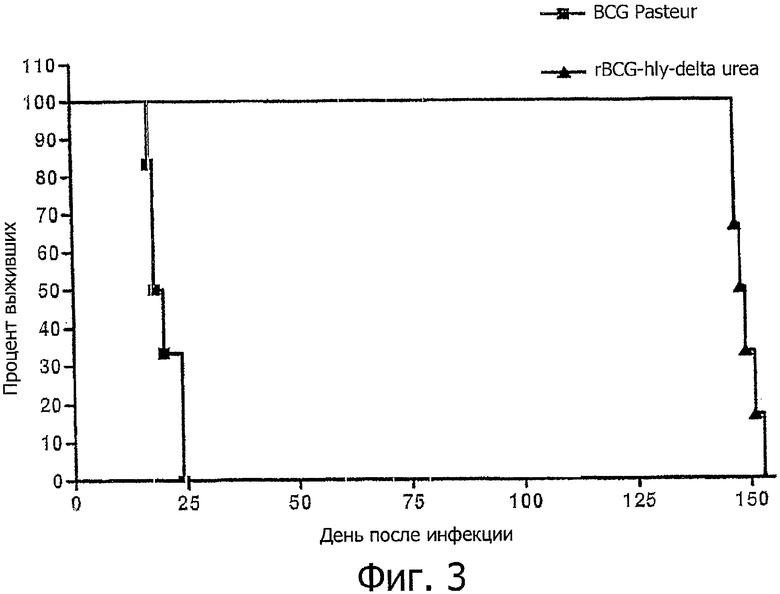
Фигура 3: демонстрирует коэффициент выживания SCID мышей, инфицированных с использованием BCG "Pasteur" и rBCG delta ureC Hlу.
SEQ ID No.1: показывает последовательность нуклеотидов молекулы нуклеиновой кислоты в соответствии с настоящим изобретением.
SEQ ID No.2: показывает соответствующую последовательность аминокислот молекулы нуклеиновой кислоты из SEQ ID No.1.
Осуществление изобретения
Пример 1. Получение дефицитных по уреазе штаммов BCG Hlу и тесты на мышиной модели
1. Инактивация уреазной активности BCG delta ureC.
С целью увеличить защитную способность штамма BCG, содержащего белок Hlу (rBCG-Hlу), уреазная активность была удалена.
Для получения дефицитного по уреазе мутанта, Reyrat et al. сконструировали вектор-самоубийцу, включающий в себя ген ureC, нарушенный канамициновым маркером (ген aph). Два микрограмма данного конструкта было линеаризовано с использованием Sac I рестриктазы и введено в М.bovis BCG путем электропорации. Устойчивые к канамицину трансформанты, подверглись отбору на негативный по уреазе фенотип (cf. Reyrat et al., 1995).
2. Конструирование "челночного" mycobacterial E. coli экспрессионного вектора pMV306:Hlу.
Для передачи функции выхода из фагосомы (опосредованной Hlу L.monocytogenes EGD Sv 1/2a) BCG Pasteur (1173 Р3) delta ureC, был использован Е. coli-mycobacterial "челночный вектор". Встраивающаяся в геном плазмида рМУ306, предшественник вектора pMV361, делает возможной стабильную хромосомную экспрессию Hlу.
Выделенный из plLH-1 1,7 - т.п.н. PstI фрагмент ДНК, кодирующий открытую рамку считывания (ORF) hly-hlyA (E. coli рHlу152-специфичный гемолизин А) был вставлен в PstI сайт плазмиды рАТ261. Полученный в результате слитый ген обеспечивал экспрессию секреторных белков, направляемых в супернатант посредством BCG-специфичного сигнального пептида Ag85B. Данный конструкт был назван рАТ261:Hlу и его ДНК экспрессионная кассета XbaI-SalI, находящаяся под транскрипционным контролем hsp60 микобактериального промотора, была последовательно использована для вставки в родительский вектор рМV306, приведшей к получению конструкта pMV306:Hlу. Была проанализирована последовательность ДНК hlу-специфичных сайтов вставки в обеих микобактериальных экспрессионных плазмидах. Предположительно зрелый Hlу-слитый белок содержит 30 аминокислот на N конце и 52 аминокислоты в С-концевой части слитого белка, которые частично совпадают с HlуA Е. coli.
3. Защитная способность в рамках мышиной модели.
Экспрессионный вектор pMV306:Hlу был трансформирован в дефицитный по уреазе штамм BCG pasteur (BCG P ureC). Полученный в результате штамм был обозначен как rBCG ureC Hlу. Защитная способность этого дефицитного по уреазе штамма микобактерий, в сравнении с родительским BCG Pasteur и BCG Pasteur ureC, в случае модели мышечного туберкулеза показана на фигуре 1. Было неожиданно обнаружено, что rBCG ureC Hlу вызывал усиленную защиту уже в ранних временных точках (день 30 после инфекции), которая продолжалась в течение всего срока наблюдения (до дня 90).
Был предпринят следующий эксперимент по длительной защите с использованием rBCG ureC Hlу. BALB/c мыши были внутривенно вакцинированы с использованием rBCG ureC Hlу, rBCG-Hlу или родительского BCG и обработаны с использованием аэрозоля на 120 день после вакцинации М. tuberculosis H37Rv, RBCG-Hlу и родительские BCG вызывали соизмеримую по эффективности защиту против М.tuberculosis H37Rv к дню 90. В отличие от них rBCG ureC Hlу вызывал усиленную защиту уже в ранних временных точках, начиная со дня 30 после инфекции. Более того, данная усиленная защита длилась в течение всего периода наблюдения и показывала уменьшение содержания М. tuberculosis H37Rv в легких на 90 день после инфекции в более чем 2 log CFU (koe), в сравнении с нативными мышами, и в более чем 1 log CFU (koe) no сравнению с мышами, вакцинированными родительским BCG.
Похожие результаты были получены после заражения клиническим изолятом М.tuberculosis Beijing. BALB/c мыши были внутривенно иммунизированы с использованием rBCG ureC Hlу, BCG-Hlу или родительского BCG и обработаны с помощью аэрозоля на 120 день после вакцинации М.tuberculosis Beijing. Вакцинация с использованием BCG ureC Hlу вызывала усиленную защиту против М.tuberculosis Beijing уже в ранних временных точках (день 30), которая продолжалась в течение всего срока наблюдения до дня 90 после инфекции. В сравнении с вакцинацией родительским BCG, вакцинация с использованием rBCG ureC Hlу вела к уменьшению содержания М.tuberculosis Beijing в легких в 1 log CPU (koe).
Пример 2. Длительная защита против М.tuberculosis H37Rv в морских свинках
Поскольку мыши являются относительно устойчивыми к инфекции М.tuberculosis, морские свинки, как более восприимчивая животная модель были использованы для проверки на вакцинационную способность rBCG ureC Hlу. Морские свинки были подкожно иммунизированы с использованием соответствующего вакцинного штамма микобактерий, rBCG ureC Hlу или родительского BCG, и привес, также как и CFU наблюдались после введения М.tuberculosis H37Rv. Морские свинки, иммунизированные с использованием rBCG ureC Hlу, показывали одинаковый привес с теми животными, которые были вакцинированы родительским штаммом BCG вплоть до дня 120, в то время как невакцинированные животные страдали от ТВ, о чем свидетельствовало падение привеса тела.
Визуальная проверка легких и селезенки перед анализом CFU показала, что туберкулы на поверхности обоих органов, взятых у BCG-иммунизированных морских свинок, были намного больше по размеру и более многочисленны чем те, что были взяты у BCG ureC Hlу-вакцинированных животных.
Пример 3. Оценка безопасности BCG ureC Hlv
Мыши Rag 1-/-, дефицитные по Т- и В-клеткам были инфицированы 106 микроорганизмами родительского штамма BCG (дикий тип) или штамма rBCG ureC Hlу. Наблюдалось присутствие микроорганизмов в легких и селезенке. Значительно уменьшенное CFU rBCG ureC Hlу было отмечено в легких (фигура 2А). В селезенке слегка уменьшенные значения CFU было отмечено после инфекции rBCG ureC Hlу в сравнении с инфекцией родительским BCG (фигура 2B).
Далее, безопасность BCG ureC Hlу была протестирована на иммунодефицитных SCID мышах. Для этой цели SCID мыши были внутривенно привиты с использованием 107-108 микроорганизмов rBCG ureC Hlу или родительского штамма BCG. В то время как SCID мыши, привитые родительским штаммом, умирали в срок до 25 после инъекции, мыши, привитые rBCG ureC Hlу, выживали до 150 дня после инъекции (фигура 3).
Данные эксперименты продемонстрировали, что BCG ureC Hlу является более безопасной по сравнению с родительским штаммом BCG.











| название | год | авторы | номер документа |
|---|---|---|---|
| СОЧЕТАНИЕ РЕКОМБИНАНТНОЙ МИКРОБАКТЕРИИ И БИОЛОГИЧЕСКИ АКТИВНОГО СРЕДСТВА В КАЧЕСТВЕ ВАКЦИНЫ | 2005 |
|
RU2495677C2 |
| Рекомбинантный штамм Mycobacterium bovis rBCG-1 на основе вакцинного штамма Mycobacterium bovis BCG strain Russia 368 | 2022 |
|
RU2807732C1 |
| РЕКОМБИНАНТНАЯ BCG, СВЕРХЭКСПРЕССИРУЮЩАЯ PhoP-PhoR | 2017 |
|
RU2760580C2 |
| ВАКЦИНА MYCOBACTERIUM TUBERCULOSIS | 2009 |
|
RU2473365C1 |
| Мультиэпитопный полипептид для иммунизации против Mycobacterium tuberculosis | 2023 |
|
RU2824195C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pTSE6, КОДИРУЮЩАЯ ГИБРИДНЫЙ ПОЛИПЕПТИД GST-ESAT-6 СО СВОЙСТВАМИ ВИДОСПЕЦИФИЧНОГО МИКОБАКТЕРИАЛЬНОГО АНТИГЕНА ESAT-6, РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ГИБРИДНОГО ПОЛИПЕПТИДА GST-ESAT-6 И РЕКОМБИНАНТНЫЙ ПОЛИПЕПТИД GST-ESAT-6 | 2004 |
|
RU2282661C2 |
| ГИБРИДИЗАЦИЯ ГЕТЕРООЛИГОМЕРНЫХ МИКОБАКТЕРИАЛЬНЫХ АНТИГЕНОВ | 2015 |
|
RU2695462C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ТУБЕРКУЛЕЗНОЙ ИНФЕКЦИИ И ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ ПОЛУЧЕНИЯ ДЕЙСТВУЮЩИХ КОМПОНЕНТОВ ЭТОЙ КОМПОЗИЦИИ | 2003 |
|
RU2262351C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pAg85A-CBD, ШТАММ Escherichia coli [pREP4, pAg85A-CBD], ХИМЕРНЫЙ БЕЛОК Ag85A-CBD И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2429292C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА CFP10-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pCFP10-DBD, ШТАММ Escherichia coli [pREP4, pCFP10-DBD], ХИМЕРНЫЙ БЕЛОК CFP10-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2546875C1 |

Изобретение относится к биотехнологии. Описана дефицитная по уреазе клетка Mycobacterium, которая является продуцентом слитого полипептида и которая содержит рекомбинантную молекулу нуклеиновой кислоты, кодирующую слитый полипептид, включающий в себя как минимум один домен из полипептида, который способен вызывать иммунный ответ у млекопитающих, и домен выхода из фаголизосом. Раскрыта фармацевтическая композиция, представляющая собой живую вакцину, содержащая в качестве активного агента описанную клетку, обеспечивающая защитный иммунитет против туберкулеза. Представлен способ живой вакцины. Раскрыт способ приготовления описанной клетки Mycobacterium. Настоящее изобретение позволяет получать более безопасную вакцину против туберкулеза. 7 н. и 28 з.п. ф-лы, 4 ил.
(а) последовательности нуклеотидов, включающей нуклеотиды 211-1722, как показано в SEQ ID No.1,
(б) последовательности нуклеотидов, которая кодирует ту же аминокислотную последовательность, что и последовательность из (а), и
(в) последовательности нуклеотидов, которая в жестких условиях гибридизуется с последовательностями из (а) или (б).
обеспечение клеток по п.1 или 16;
помещение эффективного количества указанных клеток в фармацевтически приемлемый разбавитель, носитель и адъювант.
(i) получение дефицитной по уреазе клетки Mycobacterium;
(ii) вставку рекомбинантной молекулы нуклеиновой кислоты в указанную клетку Mycobacterium, причем эта молекула нуклеиновой кислоты кодирует слитый полипептид, включающий (а) как минимум один домен из полипептида, причем этот домен способен вызывать иммунный ответ в клетках млекопитающих и (б) домен выхода из фаголизосом; и
(iii) культивирование клеток, полученных в соответствии со стадией (ii) при подходящих условиях.
(i) получение дефицитной по уреазе клетки Mycobacterium;
(ii) вставку рекомбинантной молекулы нуклеиновой кислоты в указанную клетку Mycobacterium, причем эта молекула нуклеиновой кислоты кодирует слитый пептид, содержащий как минимум один домен полипептида, причем этот домен способен вызывать иммунный ответ у млекопитающих и домен выхода из фаголизосом; и
(iii) культивирование клеток, полученных в соответствии со стадией (ii) при подходящих условиях.
| WO 9910496 04.03.1999 | |||
| Clemens et al | |||
| Purification, charaterizstion and genetic analysis of Mycobacterium tuberculosis urease, a potentially critical determinant of host-pathogen interaction J | |||
| Bacteriology, vol.177, no.19, 1995, p.5644-5652 | |||
| ПРИМЕНЕНИЕ Mycobacterium vaccae ДЛЯ РЕГУЛЯЦИИ ПО ТИПУ ОТРИЦАТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ Th2-АКТИВНОСТИ ИММУННОЙ СИСТЕМЫ | 1997 |
|
RU2197989C2 |

