Область техники, к которой относится изобретение
Изобретение относится к генной инженерии, биохимии, биотехнологии и иммунологии, в частности, к получению композиции на основе рекомбинантных псевдоаденовирусных частиц, несущих рекомбинантные гены tul4, fopA и dnaK антигенов Francisella tularensis Tul4, FopA и DnaK, и белков DBD-Tul4, 6-His-FopA и DnaK-6His, обладающей способностью продуцировать стабильный, устойчивый иммунный ответ на Francisella tularensis, и способу получения такой композиции.
Уровень техники:
Francisella tularensis является возбудителем такого заболевания человека и животных, как туляремия. Туляремия - это острое или хроническое системное природно-очаговое зоонозное заболевание, которое характеризуется лихорадкой, интоксикацией и поражением лимфатических узлов [Олсуфьев Н.Г. Таксономия, микробиология и лабораторная диагностика возбудителя туляремии - М.: Медицина, 1975 - 192 с;  . Epidemiology of tularemia // Balkan Med. J - 2014 - Vol. 31, №1 - P. 3-10]. Одним из важнейших факторов сдерживания инфекции и предупреждения вспышек является наличие надежной и безопасной вакцины, дающей длительную защиту от инфекции [Мещерякова И.С., Михайлова Т.В., Демидова Т.Н., Кормилицына М.И. Эпизоотическая и эпидемическая активность природных очагов туляремии различных ландшафтно-эпидемиологических типов в период 2009-2014 гг. // Медицинская паразитология и паразитарные болезни - 2016 - №1 - С. 42-61].
. Epidemiology of tularemia // Balkan Med. J - 2014 - Vol. 31, №1 - P. 3-10]. Одним из важнейших факторов сдерживания инфекции и предупреждения вспышек является наличие надежной и безопасной вакцины, дающей длительную защиту от инфекции [Мещерякова И.С., Михайлова Т.В., Демидова Т.Н., Кормилицына М.И. Эпизоотическая и эпидемическая активность природных очагов туляремии различных ландшафтно-эпидемиологических типов в период 2009-2014 гг. // Медицинская паразитология и паразитарные болезни - 2016 - №1 - С. 42-61].
В настоящее время имеется только одна вакцина против туляремии - живая туляремийная вакцина на основе аттенуированного штамма F. tularensis subsp. holarctica 15 [Олсуфьев Н.Г. Таксономия, микробиология и лабораторная диагностика возбудителя туляремии. - М.: Медицина, 1975 - 192 с, Гузовская Т.С., Близнюк A.M., Чистенко Г.Н. Эпидемиологический надзор за туляремией // Минск. - 2004 - 24 с.]. Данная вакцина производится ФГУП НПО «Микроген» МЗ РФ под названием «Вакцина туляремийная живая сухая».
Вакцины на основе аттенуированных штаммов обладают рядом недостатков, основными из которых являются ограниченная протективность в отношении наиболее патогенных штаммов, остаточная вирулентность и спонтанные перерождения в непротективную форму, высокая реактогенность и нестабильность штамма при хранении и пересеве из-за высокой частоты рекомбинации и появления точечных мутаций [Rohmer L., Brittnacher М., Svensson K. et al. Potential source of Francisella tularensis live vaccine strain attenuation determined by genome comparison // Infect. Immun. - 2006. - Vol. 74, №12. - P. 6895-6906; Hong K.G., Park P.G., Seo S.H. et al. Current status of vaccine development for tularemia preparedness // Clin. Exp. Vaccine Res. - 2013 - Vol. 2, №1. - P. 34-39; Rowe H.M., Huntley J.F. From the outside, in: the Francisella tularensis envelope and virulence // Front. Cell. Infect. Microbiol. - 2015. - Vol. 5. - Article 94].
Поэтому перспективным направлением является разработка субъединичной вакцины на основе белков F. tularensis. Для создания иммуногенной композиции в данной работе были выбраны антигены FopA, Tul4 и DnaK.
Tul4 - один из наиболее изученных белков наружной мембраны F. tularensis. Молекулярная масса Tul4 около 17 кДа. Последовательность белка Tul4 консервативна у различных штаммов F. tularensis, узнается Т-клетками лиц, вакцинированных F. tularensis. Определена локализация некоторых Т-клеточных детерминант белка Tul4.
FopA - белок с молекулярной массой 43 кДа находится в наружной мембране F. tularensis. Этот белок был выделен из экстрактов F. tularensis, используя сыворотки больных. Он был назван Fop (Francisella outer membrane protein), поскольку при клонировании гена fopA и его экспрессии в кишечной палочке локализовался вместе с главным белком клеточной стенки. FopA из F. tularensis LVS был клонирован в Е. coli, определена его нуклеотидная последовательность.
DnaK - белок теплового шока (70-75 кДа), возможный компонент наружной мембраны клетки F. tularensis, обнаруживаемый в периплазме. Белки теплового шока, появляющиеся в клетке в стрессовых условиях, участвуют в индукции защитного клеточного иммунитета. Белок DnaK кодируется геном dnaK, входящим в состав оперона groE Francisella. Известна последовательность туляремийного белка DnaK, во многом сходная с таковой белка DnaK Escherichia coli.
Иммуногенность данных белков и их способность защищать иммунизированных мышей от летальной инфекции была показана в ряде работ. Так, в работе [Ashtekar AR, Katz J, Xu Q, Michalek SM. A mucosal subunit vaccine protects against lethal respiratory infection with Francisella tularensis LVS // PLoS One. - 2012. - 7(11). - e50460] была сделана первая попытка создания препарата на основе комбинации двух иммуннодоминантных белков F. tularensis. В работе авторы исследовали иммуногенность препарата, содержащего рекомбинантные белки DnaK и Tul4, с GPI-0100 (полусинтетическое производное сапонина) в качестве адъюванта. Данный препарат вызывал клеточный и гуморальный иммунный ответ у иммунизированных мышей, при этом авторы показали, что иммунизация рекомбинантными белками DnaK и Tul4 вместе, но не каждым из них по отдельности, защищает 80% мышей от летальной респираторной инфекции F. tularensis LVS.
В работе [Oh Н, Kim CY, Kim СН, Hur GH, Park JH. A Synthetic Tul4 and FopA Peptide Cocktail of Francisella tularensis Induces Humoral and Cell-Mediated Immune Responses in Mice // J Microbiol Biotechnol. - 2016. - 26(9). - 1613 - 9] показано, что препарат, состоящий из рекомбинантных пептидов, являющихся фрагментами белков Tul4 и FopA, а также CpG в качестве адъюванта, вызывает иммунный ответ у мышей на клеточном и гуморальном уровне. Иммунизация проводилась трижды с интервалом в две недели. Спустя шесть недель после иммунизации, в сыворотке мышей детектировались специфические антитела к белку Tul4. Антитела к белку FopA не были обнаружены, что авторы в работе объясняют нерастворимостью использованных пептидов FopA. Таким образом, в данной работе высказана идея о важности иммунизации двумя белками внешней мембраны F. tularensis, но фактически, проведена иммунизация пептидами, представляющими собой эпитоп белка Tul4 и, в качестве адъюванта в препарат были добавлены CpG олигонуклеотиды.
При этом в работе [Hickey AJ, Hazlett KR, Kirimanjeswara GS, Metzger DW. Identification of Francisella tularensis outer membrane protein A (FopA) as a protective antigen for tularemia. Vaccine // 2011. - 29(40). - 6941-7] показано, что иммунизация мышей рекомбинантным белком FopA была способна индуцировать специфичный антительный ответ, защищать мышей от летальной дозы вакцинного штамма LVS, но не вирулентного штамма SchuS4.
В настоящее время активно разрабатываются вакцины на основе вирусных векторов. Они представляют собой рекомбинантные вирусы, в геном которых встроен целевой ген с набором регуляторных элементов. При введении в организм таких вакцин происходит попадание генетического материала в клетки организма и экспрессия в них генов целевых белков патогена. Иммунная система распознает антигены, что приводит к индукции, как гуморального, так и клеточного иммунного ответа [Draper S., Heeney J. Viruses as vaccine vectors for infectious diseases and cancer // Nat. Rev. Microbiol. - 2010. - V. 8. - c. 62-73]. Системы на основе вирусных векторов использовались для разработки субъединичных вакцин против туляремии. В работе [Banik S, Mansour АА, Suresh RV, Wykoff-Clary S, Malik M, McCormick AA, Bakshi CS. Development of a Multivalent Subunit Vaccine against Tularemia Using Tobacco Mosaic Virus (TMV) Based Delivery System // PLoS One. - 2015. - 10(6). - e0130858.] авторы использовали систему на основе вируса табачной мозаики (ВТМ) в качестве носителя иадъюванта для мультивалентной субъединичной вакцины против туляремии. В качестве белков были использованы OmpA - подобный белок, шаперон DnaK или попротеин Tul4 из высоковирулентного штамма F. tularensisSchuS4. Полученные результаты показали, что иммунизация ВТМ с рекомбинантными белками F. tularensis индуцировала гуморальный и клеточный иммунный ответ, а также обладала ограниченной протективностью, защищая 40% мышей от летальных доз штамма F. tularensis LVS.
В настоящей работе в качестве носителей использованы аденовирусы. Аденовирусы обладают рядом преимуществ. Они не патогенны, так как из их генома удалены области, ответственные за патогенность. Такие векторы способны обеспечивать высокий уровень экспрессии целевого трансгена в клетке-мишени, заражать как делящиеся, так и постмитотические клетки. Аденовирусы способны накапливаться в культуре клеток в высоких титрах. Еще одним положительным свойством таких вирусов является то, что они выводятся из организма в течение 4-5 недель. Также процесс получения нового рекомбинантного аденовируса занимает всего несколько недель, что позволяет реагировать на меняющуюся эпидемиологическую обстановку в максимально сжатые сроки [Карпов А. и др. Конструирование рекомбинантных аденовирусов птиц CELO, экспрессирующих гены гликопротеинов GB, GE, GI вируса болезни Марека // Биотехнология. - 2007. - Т. 5. - С. 38-44]. Наиболее хорошо изученным и часто используемым в качестве вакцинного вектора является рекомбинантный аденовирус человека пятого серотипа. Безопасность таких аденовирусов с делетированными Е1 и E3 областями генома подтверждается целым рядом проведенных клинических испытаний различных вакцинных и терапевтических препаратов на их основе [Hoelscher М.А. et al., Development of adenoviral-vector-based pandemic influenza vaccine against antigenically distinct human H5N1 strains in mice, Lancet, 2006, №367 (9509), c. 475-81; Van Kampen K.R. et al., Safety and immunogenicity of adenovirus-vectored nasal and epicutaneous influenza vaccines in humans // Vaccine, 2005. - №23(8). - c. 1029-36]. Применение генетических вакцин на основе данного аденовируса может быть ограничено наличием предсуществующего иммунного ответа у людей, уже встречавшихся ранее с этим вирусом [Bangari D.S. et al., Comparative transduction efficiencies of human and nonhuman adenoviral vectors in human, murine, bovine, and porcine cells in culture // Biochem. Biophys. Res. Commun. - 2005. - №327. - c. 960-966]. Такого воздействия предсуществующего иммунного ответа можно избежать при интраназальной иммунизации, вместо парентерального введения.
Разработаны быстрые и гибкие технологии получения рекомбинантных аденовирусов, позволяющие реализовать масштабное производство различных кандидатных вакцин на основе аденовирусных векторов на одной технологической линии, без ее переоборудования и изменения регламента. Вышеперечисленные свойства делают рекомбинантные аденовирусы хорошей технологической платформой для создания широкого спектра вакцин против различных заболеваний [Harrop R. et al., Recombinant viral vectors: cancer vaccines // Adv. Drug Deliv. Rev. - 2006. - №58(8). - c. 931-947; Tang D.C. et al., Overexpression of adenovirus-encoded transgenes from the cytomegalovirus immediate early promoter in irradiated tumor cells // Hum. Gene Ther. - 1997. - №8(17). - c. 2117-2124; Tang D.C. et al., Butyrate-inducible and tumor-restricted gene expression by adenovirus vectors // Cancer Gene Ther. - 1994. - №1(1). - c. 15-20].
Возможность использования аденовирусных частиц для создания вакцин против туляремии обсуждается в патенте US 9,624,510 В2 "Adenoviral vectors comprising partial deletions of e3". При этом патент описывает аденовирусный вектор с частичной делецией в домене E3, как систему для разработки вакцин и генной терапии, но не конкретный вариант иммуногенной композиции против туляремии.
Наиболее близкими аналогами данного изобретения являются описанные в заявке на изобретение US 2015/0030632 A1 и в патенте RU 2270249 С1 «субъединичные» вакцины - иммуногенные композиции на основе белка Tul4. Для композиции, включающей белок Tul4 из патента US 2015/0030632 A1 показана не только иммуногенность, но и протективность - после заражения F. Tularensis SchuS4 выжило 75% мышей, иммунизированных препаратом, содержащим белок Tul4.
Отличие настоящего изобретения от обоих аналогов состоит в том, что, во-первых, для прайм-вакцинации используются аденовирусные частицы в качестве носителей антигенов - белков Tul4, FopA и DnaK, для последующих, бустерных, вакцинаций используются рекомбинантные белки Tul4, FopA и DnaK. Кроме того, в отличие от патентов US 2015/0030632 A1 и RU 2270249 С1 для иммунизации был использован не один белок, а смесь рекомбинантных белков - Tul4, FopA и DnaK. Третьим отличием является то, что для повышения иммуногенности используются два адъюванта - гидроксид алюминия и CpG олигонуклеотиды: частицы гидроксида алюминия применяются для иммобилизации белков на их поверхности, а смесь двух олигонуклеотидов CpG - для дополнительной стимуляции иммунного ответа.
Еще одним аналогом настоящего изобретения является US 2014/0356415 A1, где раскрыта иммуногенная композиция, содержащая белки Tul4, FopA и DnaK. Отличием настоящего изобретения от указанного аналога является использование рекомбинантных белков DBD-Tul4, 6-His-FopA и DnaK-6His, что позволяет избегать нежелательных побочных эффектов при введении композиции, так как рекомбинантные белки имеют высокую степень очистки, а также стабильность, что позволяет добиваться высокой стандартизации препаратов.
Раскрытие изобретения:
Техническое решение изобретения выражается в получении композиции на основе рекомбинантных псевдоаденовирусных частиц (РПАН), несущих гены белковых антигенов F. tularensisDBD-Tul4, 6-His-FopA и DnaK-6His, а также рекомбинантных белков Tul4, FopA и DnaK, которые получают по средствам их экспрессии в клетках непатогенных прокариотических микроорганизмов (штаммов Е. coli) и их дальнейшей аффинной очистке, и двух адъювантов - гидроксида алюминия и двух олигонуклеотидов CpG.
Сущность изобретения выражается в получении композиции на основе рекомбинантных псевдоаденовирусных частиц, экспрессирующих гены tul4, fopA и dnaK белковых антигенов F. Tularensis и отдельных высокоочищенных и стабильных рекомбинантных белков DBD-Tul4, 6-His-FopA и DnaK-6His, которые характеризуются высоким уровнем экспрессии и стабильности в бактериальных экспрессионных системах. Полученные РПАН и белки обладают иммуногенностью, как показано в примерах 3 и 5, а также нетоксичны, что делает возможным их использование в составе иммуногенных композиций, направленных на специфическую активацию иммунитета и формирование иммунологической памяти.
Для реализации изобретения решены следующие задачи: (i) сконструированы рекомбинантные псевдоаденовирусные частицы, несущие гены tul4, fopA и dnaK белковых антигенов F. tularensis; (ii) проведена проверка экспрессии генов антигенов из состава рекомбинантных псевдоаденовирусных частиц методом вестерн-блот на Fc-часть химерных белков; (iii) показана иммуногенность полученных по заявленному изобретению рекомбинантных псевдоаденовирусных частиц, несущих гены tul4, fopA и dnaK белковых антигенов F. tularensis на мышах. Для решения данных задач получают три типа псевдоаденовирусных частиц, несущих гены tul4, fopA и dnaK белковых антигенов F. tularensis, соответственно. При создании конструкций бактериальный лидерный пептид удаляют, и добавляют эукариотический лидерный пептид для индукции гуморального иммунного ответа. Гены клонируют в плазмидный вектор pSh-PA-ILZ-Fc, таким образом, что полученная конструкция содержит ген DnaK (FopA или Tul4, соответственно), а также изолейциновый зиппер (ILZ) и Fc-фрагмент антитела IgG2a (Fc), которые необходимы для гексамеризации антигена, что значительно увеличивает их иммуногенность. Полученные линеаризованные векторы pSh-FopA, pSh-Tul4 и pSh-DnaK смешивают с геномной ДНК рекомбинантного аденовируса Ad5 и котрансформировали в клетки Е. coli (штамм BJ5183). Полученные плазмиды трансформируют в другой штамм Е. coli (DH5alpha), в котором в отличие от штамма BJ5183 возможно препаративное накопление рекомбинантных плазмид. На следующем этапе клетки линии 293 трансфецируют линеаризованными плазмидами, лизируют, и из полученного лизата выделяют псевдоаденовирусные частицы.
Для получения рекомбинантных белков получают плазмиды, содержащие нуклеотидные последовательности рекомбинантных генов, кодирующих рекомбинантные белки DBD-Tul4, 6-His-FopA и DnaK-6His. Синтетические ДНК с последовательностями, соответствующими генам, кодирующим белки DBD-Tul4, 6-His-FopA и DnaK-6His, спланированные таким образом, чтобы нуклеотидный состав кодонов был оптимизирован для гетерологичной экспрессии в непатогенном лабораторном штамме Е. coli.
После трансформации полученными плазмидами pTUL4, pFOPA и pDNAK получают штаммы-продуценты Е. coli М15 [pREP4, pTul4], Е. coli Ml 5 [pREP4, pFopA] и E.coli M15 [pREP4, pDnaK] с высокой продукцией рекомбинантных белков Tul4, FopA и DnaK в тельцах включения (ТВ). Расчетные молекулярные массы рекомбинантных белков Tul4, FopA и DnaK составляют 30,0, 40,5 и 70,4 кДа, соответственно. Дальнейшая очистка всех индивидуальных рекомбинантных белков DBD-Tul4, 6-His-FopA и DnaK-6His производится путем растворения ТВ в денатурирующих условиях и очистки колоночной аффинной хроматографией в денатурирующих условиях на носителях, содержащих в качестве неподвижной фазы декстран (сефадекс), в случае DBD-Tul4 или содержащих иммобилизованный металл (металл-хелатная хроматография) в случае 6-His-FopA и DnaK-6His, с последующим удалением денатурирующих агентов из раствора белков.
В результате очистки получают с высоким выходом высокой степени чистоты рекомбинантные белки DBD-Tul4, 6-His-FopA и DnaK-6His, которые характеризуются стабильностью при очистке, обладают иммуногенностью при введении лабораторным животным, что делает возможным их использование в составе иммуногенной композиции, содержащей смесь данных рекомбинантных белков и направленной на специфическую активацию иммунитета и формирование иммунологической памяти.
На этапе получения синтетической последовательности ДНК, кодирующей белок Tul4, настоящий способ отличается от аналога RU 2270249 С1 тем, что слитные (конъюгированные) белки в настоящем изобретении и в аналоге различаются: аминокислотная последовательность белка, заявленного в RU 2270249 С1, содержит последовательность Tul4, слитную с аффинным целлюлозосвязывающим доменом CBD, а в настоящем изобретении последовательность Tul4 конъюгирована с декстрансвязывающим доменом DBD, кодируемым геном белка с декстрансвязывающим доменом из Leuconostoc mesenteroides (DBD1).
Техническим результатом, достигаемым при реализации заявляемой группы изобретений, является получение оптимальных для гетерологичной экспрессии синтетических последовательностей ДНК, кодирующих псевдоаденовирусные частицы, несущие гены tul4, fopA и dnaK, а также рекомбинантные белки, обеспечивающие высокий уровень продукции белков DBD-Tul4, 6-His-FopA и DnaK-6His, в непатогенных лабораторных экспрессионных штаммах Е. coli; простую и эффективную очистку белков, обладающих высокой иммунореактивностью; создание вакцинного препарата, содержащего смесь этих белков с двумя адъювантами - гидроксидом алюминия, выступающим в качестве носителя для иммобилизации белков, и CpG, дополнительно стимулирующим иммунную реакцию. Указанный технический результат достигается за счет получения рекомбинантных псевдоаденовирусных частиц, несущих гены белков DBD-Tul4, 6-His-FopA и DnaK-6His с нуклеотидной последовательностью SEQ ID NO 1, SEQ ID NO 3 и SEQ ID NO 5, соответственно. Аминокислотная последовательность соответствующих генов SEQ ID NO 2, SEQ ID NO 4 и SEQ ID NO 6, соответственно.
Для достижения указанного технического результата также были получены рекомбинантные белки DBD-Tul4, 6-His-FopA и DnaK-6His в клетках лабораторных штаммов-продуцентов Е. coli. Для синтеза рекомбинантных белков DBD-Tul4, 6-His-FopA и DnaK-6His в клетках лабораторных штаммов-продуцентов Е. coli используются соответствующие рекомбинантные плазмиды с рекомбинантными генами с нуклеотидной последовательностью SEQ ID NO 15 или нуклеотидной последовательностью SEQ ID NO 17 или нуклеотидной последовательностью SEQ ID NO 19.
Рекомбинантный белок DBD-Tul4 имеет аминокислотную последовательность SEQ ID NO 16, включающую последовательность аминокислотных остатков декстрансвязывающего домена (DBD) из Leuconostoc mesenteroides, глицин-сериновый спейсер и аминокислотную последовательность белка Tul4 F. tularensis.
Рекомбинантный белок 6-His-FopA имеет аминокислотную последовательность SEQ ID NO 18 включающую последовательность 6His, и аминокислотную последовательность белка FopA F. tularensis.
Рекомбинантный белок DnaK-6His имеет аминокислотную последовательность SEQ ID NO 20, включающую аминокислотную последовательность белка DnaK F. tularensis и последовательность 6His.
Кроме того, техническим результатом, достигаемым при осуществлении настоящей группы изобретений, является получение высокого уровня продукции белков DBD-Tul4, 6-His-FopA и DnaK-6His (см. пример 1, фиг. 11), которые после очистки способны взаимодействовать с соответствующими антителами (см. пример 4, фиг. 14) и способны вызывать активный иммунный ответ в мышиной модели (см. пример 5, фиг. 15-17).
Наличие в рекомбинантных белках DBD-Tul4, 6-His-FopA и DnaK-6His эпитопов (антигенных детерминант), идентичных таковым у природных белков, подтверждается тем, что специфические поликлональные сывороточные антитела кроликов, полученные к рекомбинантным белкам, взаимодействуют с белковыми антигенами вакцинного штамма F. tularensis (см. пример 4, фиг. 14).
Иммуногенность препарата рекомбинантных псевдоаденовирусных частиц, несущих гены tul4, fopA и dnaK подтверждается появлением выраженной иммунной реакции in vivo в мышиной модели (см. пример 3). Что является еще одним техническим результатом.
Для получения такого технического результата как иммуногенность препарата рекомбинантных белков DBD-Tul4, 6-His-FopA и DnaK-6His, созданные рекомбинантные белки с последовательностями SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20 были включены в состав вакцинного препарата, содержащего адъюванты - гидроксид алюминия и CpG олигонуклеотиды. В части формулирования вакцинного препарата, содержащего смесь рекомбинантных белков с адъювантами - гидроокисью алюминия и олигонуклеотидов CpG, технический результат достигается за счет создания белков DBD-Tul4, 6-His-FopA и DnaK-6His с указанными последовательностями SEQ ID NO 16, SEQ ID NO 18, SEQ ID NO 20, а также за счет иммобилизации этих белков на поверхности частиц в суспензии гидроокиси алюминия и за счет дополнительной стимуляции иммунного ответа олигонуклеотидами CpG.
Иммуногенность смеси рекомбинантных белков в составе вакцинного препарата с адъювантами - гидроокисью алюминия и CpG олигонуклеотидами - была исследована in vivo в мышиной модели. Результат проявился в виде формирования выраженной иммунной реакции и стимуляции образования специфических антител ко всем трем белковым антигенам, входящим в состав вакцинной композиции (см. пример 5, фиг. 15-18).
Изобретение также позволяет специфически индуцировать иммунитет, включающий иммунизацию мышей рекомбинантными псевдоаденовирусными частицами, несущими гены tul4, fopA и dnaK белковых антигенов F. tularensis и последующую двукратную иммунизацию мышей подкожно с интервалом в две недели препаратом, содержащим рекомбинантные белки DBD-Tul4, 6-His-FopA и DnaK-6His и адъюванты - гидроксид алюминия и CpG олигонуклеотиды (см. пример 6).
Таким образом, изобретение позволяет получить стабильный и устойчивый иммунный ответ на Franciella tularensis.
Изобретение проиллюстрировано следующими примерами, приведенными ниже.
Описание фигур
На представленных фигурах обозначения Tul4, FopA и DnaK соответствуют белкам DBD-Tul4, 6-His-FopA и DnaK-6His.
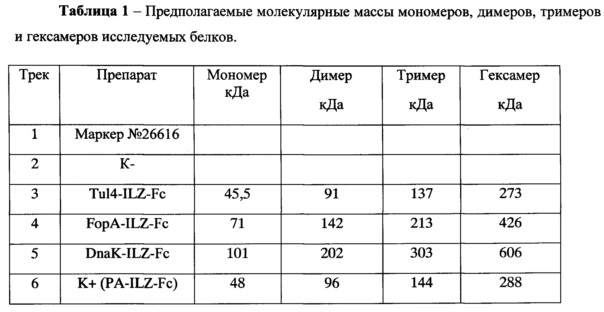
На фиг. 1 показана Схема экспрессирующей кассеты рекомбинантной псевдоаденовирусной частицы, несущей гены белковых антигенов F. tularensis (tul4 или fopA или dnaK соответственно). Где: CMV - промотор цитомегаловируса человека; трансген - ген белковых антигенов F. tularensistul4 или fopA или dnaK; рА - сигнал полиаденилирования SV40 (Simian vacuolating virus 40); Ad5 - геномная часть аденовируса человека 5 серотипа, ΔЕ1, ΔЕ3 - делеции Е1 и E3 областей аденовирусного генома.
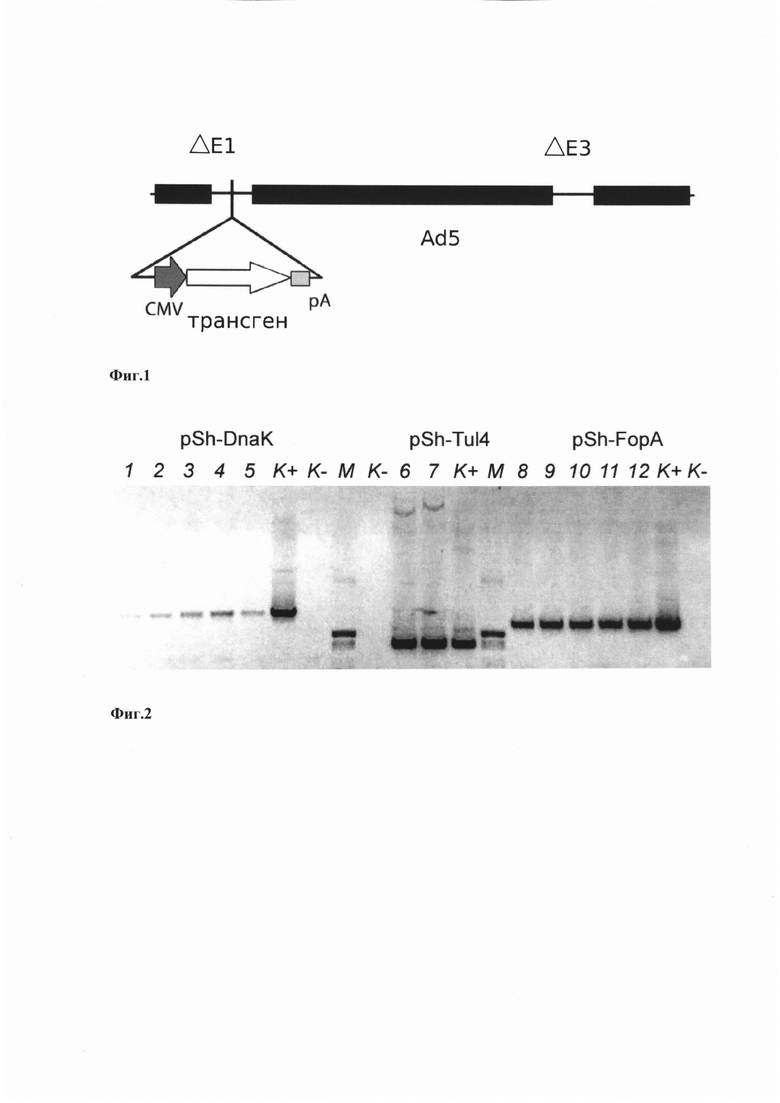
На фиг. 2 показаны результаты анализа клонов методом ПЦР на наличие генов, кодирующих антигены DnaK, Tul4, FopA. Дорожки 1-5 в виде полос детектируются клоны плазмиды pSh-DnaK, К+ - положительный контроль, К - - отрицательный контроль (полоса отсутствует), М - маркер молекулярной массы pUC19, гидролизованный по Hpall, на треках 6, 7 в виде полос детектируются клоны плазмиды pSh-Tul4, К+ - положительный контроль, К- - отрицательный контроль (полоса отсутствует), на треках 8-12 в виде полос детектируются клоны плазмиды pSh-FopA, К+ - положительный контроль, К- - отрицательный контроль (полоса отсутствует).
На фиг. 3 представлены результаты анализа клонов: с использованием эндонуклеаз рестрикции NotI+HinDIII. Дорожки 1 - pET-FopA, 2 - pSh-FopA, 3 - pET-Tul4, 4 - pSh-Tul4, 5 - pET-DnaK, 6 - pSh-DnaK; с использованием эндонуклеаз рестрикции NotI+XhoI: 7 - pET-FopA, 8 - pSh-FopA, 9 - pET-Tul4, 10 - pSh-Tul4, 11 - pET-DnaK, 12 - pSh-DnaK; не гидролизованные плазмиды: 13 - pET-FopA, 14 - pSh-FopA, 15 - pET-Tul4,16 - pSh-Tul4, 17 - pET-DnaK, 18 - pSh-DnaK.
На фиг. 4 представлены результаты анализа клонов методом ПЦР a) Ad5-DnaK, б), Ad5-FopA, в) Ad5-Tul4. Дорожки 1-16 - ПЦР-продукты, полученные при анализе, (М) Маркер молекулярной массы (ДНК фага лямбда, гидролизованная по сайту PstI).
На фиг. 5 представлены результаты анализа положительных клонов методом ПЦР. Дорожки К- - отрицательный контроль (полоса отсутствует), 3 pAd5-DnaK, 7 pAd5-DnaK, К+ - pSh-DnaK (положительный контроль), К- - отрицательный контроль (полоса отсутствует), 1 pAd5-FopA, 5 pAd5-FopA, К+ - pSh-FopA (положительный контроль), К- - отрицательный контроль (полоса отсутствует), 5 pAd5-Tul4, 9 pAd5-Tul4, К+ - pSh-Tul4 (положительный контроль).
На фиг. 6 показана электрофореграмма результатов гидролиза плазмидной ДНК эндонуклеазой рестрикции NotI. Дорожки (1) pAd5-Tul4, (2) pAd5-FopA, (3) pAd5-DnaK, (4) pAd5-EASY, M - маркер молекулярных масс (ДНК фага лямбда, гидролизованная по сайту PstI).
На фиг. 7 показаны результаты вестерн-блоттинга. Дорожки (М) Маркер молекулярной массы №266166, (1) Отрицательный контроль (К-), (2) Экспрессированный антиген Tul4 рекомбинантными псевдоаденовирусными частицами, несущими ген белкового антигена F. tularensis Tul4, (3) Экспрессированный антиген FopA рекомбинантными псевдоаденовирусными частицами, несущими ген fopA белкового антигена F. tularensis, (4) Экспрессированный антиген DnaK рекомбинантными псевдоаденовирусными частицами, несущими ген dnaK белкового антигена F. tularensis, (5) - Положительный контроль (К+) (PA-ILZ-Fc), а - тример Tul4-ILZ-Fc (137 кДа), б - тример FopA-ILZ-Fc (213 кДа), в - тример DnaK-ILZ-Fc (303 кДа), г - тример PA-ILZ-Fc (144 кДа).
На фиг. 8 показана диаграмма изменения титра антител к белку FopA, полученная при анализе пулированных сывороток мышей, иммунизированных вакцинным препаратом, содержащим аденовирусные частицы в сравнении с контрольной группой, а) Иммунный ответ на FopA в контрольной группе; б) Иммунный ответ на FopA-компоненты препарата.
На фиг. 9 показана диаграмма изменения титра антител к белку DnaK, полученная при анализе пулированных сывороток мышей, иммунизированных вакцинным препаратом, содержащим аденовирусные частицы в сравнении с контрольной группой, а) Иммунный ответ на DnaK в контрольной группе; б) Иммунный ответ на DnaK-компоненты препарата.
На фиг. 10 показана диаграмма изменения титра антител к белку Tul4, полученная при анализе пулированных сывороток мышей, иммунизированных вакцинным препаратом, содержащим аденовирусные частицы в сравнении с контрольной группой, а) Иммунный ответ на Tul4 в контрольной группе; б) Иммунный ответ на Tul4-компоненты препарата.
На фиг. 11 представлена электрофореграмма экстрактов клеток продуцентов рекомбинантных белков Tul4, FopA и DnaK до и после индукции IPTG. Дорожки (1) Е. coli M15 [pRep4, pTul4] до индукции; (2) Е. coli M15 [pRep4, pTul4] после индукции - рекомбинантный белок Tul4, 30 кДа; (3) Е. coli M15 [pRep4, pFopA] до индукции; (4) Е. coli M15 [pRep4, pFopA] после индукции - рекомбинантный белок FopA, 40 кДа; (5) Е. coli M15 [pRep4, pDnaK] до индукции; (6) Е. coli M15 [pRep4, pDnaK] после индукции - рекомбинантный белок DnaK, 70 кДа; (7) маркеры молекулярной массы.
На фиг. 12 представлена электрофореграмма образцов рекомбинантного белка DBD-Tul4 на различных стадиях очистки. Дорожки: М - маркеры молекулярной массы, (1) экстракт клеток продуцента после индукции IPTG, (2) супернатант (легко растворимые клеточные белки продуцента), (3) отмытые тельца включения, (4) элюат с колонки с сефадексом G-150.
На фиг. 13 представлена электрофореграмма образцов рекомбинантного белка FopA на различных стадиях очистки. Дорожки: (М) маркеры молекулярной массы, (1) экстракт клеток продуцента после индукции IPTG, (2) супернатант (легко растворимые клеточные белки продуцента), (3) - отмытые тельца включения, (4) элюат с колонки с сорбентом WorkBeads 40 Ni.
На фиг. 14 представлена нитроцеллюлозная мембрана с перенесенными белками, окрашенная мышиными антисыворотками, полученными после иммунизации препаратом трех рекомбинантных белков Tul4, FopA, DnaK с адъювантами, и субстратами пероксидазы - ДАБ и перекисью водорода. Дорожки 1, 2 - лабораторный образец раствора рекомбинантного белка Tul4 0,5 и 0,05 мг/мл, соответственно; 4,5 - лабораторный образец раствора рекомбинантного белка FopA 0,5 и 0,05 мг/мл, соответственно; 7,8 - лабораторный образец раствора рекомбинантного белка DnaK 0,5 и 0,05 мг/мл, соответственно; 3,6,9 - экстракт вакцинного штамма F. tularensis; М - белковые маркеры молекулярной массы.
На фиг. 15 представлена диаграмма изменения титра антител к FopA. а) в ответ на введение раствора адъювантов в контрольной группе животных; б) в сыворотке мышей после двукратного введения препарата трех рекомбинантных белков Tul4, FopA, DnaK с адъювантами.
На фиг. 16 представлена диаграмма изменения титра антител к DnaK. а) в ответ на введение раствора адъювантов в контрольной группе животных; б) в сыворотке мышей после двукратного введения препарата трех рекомбинантных белков Tul4, FopA, DnaK с адъювантами.
На фиг. 17 представлена диаграмма изменения титра антител к Tul4. а) в ответ на введение раствора адъювантов в контрольной группе животных; б) в сыворотке мышей после двукратного введения препарата трех рекомбинантных белков Tul4, FopA, DnaK с адъювантами.
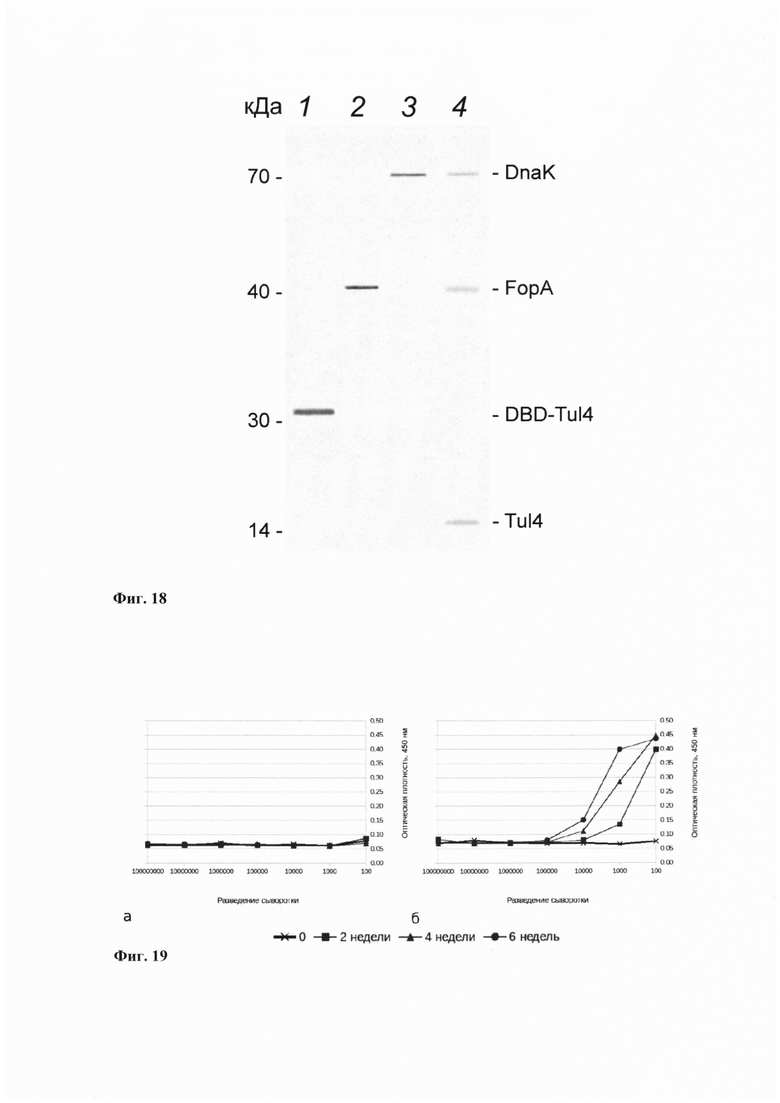
На фиг. 18 представлен результат иммуноблоттинга - нитроцеллюлозная мембрана с перенесенными белками, окрашенная мышиными антисыворотками, полученными после иммунизации препаратом трех рекомбинантных белков Tul4, FopA, DnaK с адъювантами, и субстратами пероксидазы - ДАБ и перекисью водорода. (1) лабораторный образец раствора рекомбинантного белка Tul4, (2) лабораторный образец раствора рекомбинантного белка FopA, (3) лабораторный образец раствора рекомбинантного белка DnaK, (4) экстракт клеток вакцинного штамма.
На фиг. 19 представлена диаграмма изменения титра антител к FopA. а) в ответ на введение раствора адъювантов в контрольной группе животных; б) в сыворотке мышей после иммунизации рекомбинантными псевдоаденовирусными частицами в качестве прайм-компонента и двукратной иммунизации препаратом трех рекомбинантных белков Tul4, FopA, DnaK с адъювантами в качестве бустерного компонента.
На фиг. 20 представлена диаграмма изменения титра антител к DnaK. а) в ответ на введение раствора адъювантов в контрольной группе животных; б) в сыворотке мышей после иммунизации рекомбинантными псевдоаденовирусными частицами в качестве прайм-компонента и двукратной иммунизации препаратом трех рекомбинантных белков Tul4, FopA, DnaK с адъювантами в качестве бустерного компонента.
На фиг. 21 представлена диаграмма изменения титра антител к Tul4. а) в ответ на введение раствора адъювантов в контрольной группе животных; б) в сыворотке мышей после иммунизации рекомбинантными псевдоаденовирусными частицами в качестве прайм-компонента и двукратной иммунизации препаратом трех рекомбинантных белков Tul4, FopA, DnaK с адъювантами в качестве бустерного компонента.
Конкретно настоящее изобретение относится к иммуногенной композиции, включающей (i) прайм-компонент, состоящий из рекомбинантных псевдоаденовирусных частиц: частиц, несущих ген tul4 Francisella tularensisc последовательностью SEQ ID NO 1, частиц, несущих ген fopA Francisella tularensis с последовательностью SEQ ID NO 3 и частиц, несущих ген dnaK Francisella tularensis с последовательностью SEQ ID NO 5, при этом указанные гены оптимизированы для экспрессии в эукариотических клетках, и/или (ii) бустерный компонент, включающий рекомбинантные белки: белок DBD-Tul4 с последовательностью SEQ ID NO 16, молекулярной массой 30,0 кДа, содержащий декстрансвязывающий домен декстрансукразы из L. citreum КМ20, представленный аминокислотными остатками 1-140 SEQ ID NO 16, Gly-Ser спейсер, представленный остатками 141-150 SEQ ID NO 16 и последовательность белка Tul4 Franciella tularensis, представленной остатками 151-282 SEQ ID NO 16; белок 6-His-FopA с последовательностью SEQ ID NO 18, молекулярной массой 40,5 кДа; белок DnaK-6His с последовательностью SEQ ID NO 20, молекулярной массой 70,4 кДа, а также содержащая фармацевтически приемлемый носитель или разбавитель.
Предпочтительно иммуногенная композиция представляет собой вакцину, индуцирующую иммунный ответ против Franciella tularensis.
Предпочтительно иммуногенная композиция используется для индукции специфического клеточного и гуморального иммунитета против Franciella tularensis, по средствам введения прайм-компонента интраназально и введения бустерного компонента подкожно, дважды с интервалом в две недели.
Настоящее изобретение также относится к способу получения указанной иммуногенной композиции, включающему получение трех типов псевдоаденовирусных частиц, несущих гены tul4, fopA и dnaK белковых антигенов F. tularensis, соответственно, при этом бактериальный лидерный пептид удаляют, и добавляют эукариотический лидерный пептид, каждый ген клонируют в свой плазмидный вектор pSh-PA-ILZ-Fc, куда также клонируют изолейциновый зиппер (ILZ) и Fc-фрагмент антитела IgG2a (Fc), полученные линеаризованные векторы pSh-FopA, pSh-Tul4 и pSh-DnaK смешивают с геномной ДНК рекомбинантного аденовируса Ad5 и котрансформируют в клетки Е. coli штамма BJ5183, полученные плазмиды трансформируют в штамм Е. coli DH5alpha, затем клетки линии 293 трансфецируют линеаризованными плазмидами, лизируют, и из полученного лизата выделяют псевдоаденовирусные частицы; получают рекомбинантные белки Tul4, FopA и DnaK, для чего получают плазмиды, содержащие, синтетические ДНК с последовательностями, соответствующими генам, кодирующим белки DBD-Tul4, 6-His-FopA и DnaK-6His, где нуклеотидный состав кодонов оптимизирован для гетерологичной экспрессии в непатогенном лабораторном штамме Е. coli, на основе указанных плазмид pTUL4, pFOPA и pDNAK получают штаммы-продуценты Е. coli М15 [pREP4, pTul4], Е. coli М15 [pREP4, pFopA] и E.coli M15 [pREP4, pDnaK] с высокой продукцией рекомбинантных белков Tul4, FopA и DnaK в тельцах включения (ТВ), дальнейшая очистка всех индивидуальных рекомбинантных белков DBD-Tul4, 6-His-FopA и DnaK-6His производится путем растворения ТВ в денатурирующих условиях и проведении колоночной аффинной хроматографией в денатурирующих условиях на носителях, содержащих в качестве неподвижной фазы декстран (сефадекс), в случае DBD-Tul4 или содержащих иммобилизованный металл (металл-хелатная хроматография) в случае 6-His-FopA и DnaK-6His, с последующим удалением денатурирующих агентов из раствора белков, на основе полученных псевдовирусных частиц и белков готовят композицию, содержащую либо псевдовирусные частицы, либо белки, либо комбинацию псевдовирусных частиц и белков.
В одном из вариантов воплощения настоящего способа псевдовирусные частицы и белки подвергают лиофильной сушке.
Примеры
Пример 1. Конструирование и получение рекомбинантных псевдоаденовирусных частиц, несущих гены белковых антигенов F. tularensis Tul4, FopA и DnaK.
В данном примере показан ход конструирования рекомбинантных псевдоаденовирусных частиц, несущих гены tul4, fopA и dnaK белковых антигенов F. tularensis, и процесс их получения.
Первым этапом является процесс разработки генетических конструкций и в дальнейшем получение соответствующих рекомбинантных аденовирусов, несущих гены tul4, fop А и dnaK белковых антигенов F. tularensis Tul4, FopA и DnaK (Ad-tul4, Ad-fop A, Ad-dnaK) (см. Фиг. 1).
Аминокислотные последовательности антигенов Francisella tularensis были получены из базы данных UniProt: DnaK - Q14GW9, FopA - Q5NH85, Tul4 - P18149. Бактериальный лидерный пептид в генах fopA и tul4 был удален. Для индукции гуморального иммунного ответа ко всем трем генам был добавлен эукариотический лидерный пептид. Кодонный состав генов был оптимизирован под экспрессию в клетках Mus musculus. В ЗАО «Евроген» были синтезированы гены fopa, dnak и tul4 и включены в составе плазмид рЕТ28а. Нуклеотидные и аминокислотные последовательности генов белков Tul4 (SEQ ID NO 1 и SEQ ID NO 2), FopA (SEQ ID NO 3 и SEQ ID NO 4), DnaK (SEQ ID NO 5 и SEQ ID NO 6) приведены в списке последовательностей.
Гены были клонированы по рестрикционным сайтам NotI и XhoI в плазмидный вектор pSh-PA-ILZ-Fc, таким образом, удален ген РА (ген протективного антигена) и вместо него вставлен ген dnaK (fopA или tul4 соответственно). Изолейциновый зиппер (ILZ) и Fc-фрагмент антитела IgG2a (Fc) необходимы для гексамеризации антигена, что значительно увеличивает иммуногенность антигенов [Shcherbinin D.N. et al. Protective immune response against Bacillus anthracis induced by intranasal introduction of a recombinant Adenovirus Expressing the Protective Antigen Fused to the Fc-fragment of IgG2a // Acta Naturae - 2014, - V. 6 - №1 - c. 76-84].
С целью определения наличия антигена был проведен анализ клонов методом ПЦР. Результаты представлены на фиг. 2. При анализе использовали следующие пары праймеров, представленные в перечне последовательностей, для определения dnaK были использованы праймеры SEQ ID NO 7 и SEQ ID NO 8, fopA SEQ ID NO 9 и SEQ ID NO 10, tul4 - SEQ ID NO 11 и SEQ ID NO 12.
Также был проведен анализ клонов по молекулярному весу и рестрикционным картированием. Результаты представлены на фиг. 3.
Полученные векторы pSh-FopA, pSh-Tul4 и pSh-DnaK линеаризовали с помощью эндонуклеазы рестрикции Pmel, смешивали с геномной ДНК рекомбинантного аденовируса Ad5 и котрансформировали в клетки Е. coli (штамм BJ5183). Далее полученные клоны анализировали методом ПЦР на наличие гексона Ad5. Результаты представлены на фиг. 4. Для постановки ПЦР использовали праймеры на наличие гексона Ad5 (SEQ ID NO 13 и SEQ ID NO 14). Положительные клоны анализировали методом ПЦР на наличие антигена. Результаты представлены на фиг. 5.
Полученные клоны трансформировали в штамм Е. coli (DH5 alpha), в котором возможно препаративное накопление рекомбинантных плазмид. Выделенную плазмидную ДНК анализировали рестрикцией по сайту NotI (Фиг. 6).
На следующем этапе клетки линии 293 были трансфецированы плазмидами, гидролизованными с помощью эндонуклеазы рестрикции PacI. Трансфекцию проводили в 24-луночном планшете с использованием реактива Lipofectamine 2000 ("Invitrogen"). Через 10 дней после трансфекции клетки собирали, замораживали-оттаивали и полученным лизатом, содержащим рекомбинантные аденовирусы, заражали клетки 293 линии в 35 мм чашке. Через 5 дней наблюдали специфический лизис клеток, обусловленный цитопатическим действием вирусов. Из полученного препарата рекомбинантные вирусы были размножены на клетках 293 линии и получено препаративное количество данного вируса. Для этого клетки 293 линии, выращенные на 150 мм культуральных чашках, были инфицированы рекомбинантными аденовирусами в дозе 107 БОЕ/чашку (бляшкообразующих единиц на чашку). Инфицированную культуру инкубировали 72 часа, после чего клетки были собраны, путем перемораживания был приготовлен лизат. Лизированные клетки отделяли от жидкости, содержащей вирус, центрифугированием при 2000 об/мин. Концентрирование и очистку вируса проводили двукратным ультрацентрифугированием в градиенте хлористого цезия. Титр препарата вирусов определяли методом бляшкообразования в культуре клеток линии 293. В результате было получено около 50 о.е. (оптических единиц) каждого вируса с титром 5×109 БОЕ/мл (бляшкообразующих единиц на чашку).
Пример 2. Проверка экспрессии антигенов из состава рекомбинантных псевдоаденовирусных частиц методом вестерн-блоттинга со специфическими антителами к Fc-части химерных белков.
Клетки А549 на 25 см матрасе были трансдуцированы рекомбинантными аденовирусами. Через 24 часа среду меняли на среду DMEM, содержащую 2% бычьей сыворотки, и инкубировали 6 дней. Среду собирали, концентрировали в 10 раз, добавляли 1/3 объема буфера образца, содержащего дитиотреитол, и прогревали 10 минут при 98°С. Для детекции использовали мышиные антитела анти-IgG, меченные пероксидазой хрена. Обнаружение полос проводили методом хемилюминесценции.
Поскольку антиген может быть в мультимерной форме, были рассчитаны молекулярные массы для каждой из форм, представленные в Табл. 1.
По результатам вестерн-блоттинга были обнаружены тримерные формы антигенов (Фиг. 7). Ниже 100 кДа не наблюдалось полос ни в одном из треков. Цистеиновые S-S связи в Fc-фрагменте восстановились дитиотреитолом до SH-групп. Благодаря гидрофобным взаимодействиям изолейциновый зиппер после прогревания, по-видимому, вновь образовал тримеры. Вестерн-блоттинг образцов в неденатурирующих условиях не проводили, так как из-за большой молекулярной массы возможно не вхождение белков в гель даже с минимальной процентным содержанием акриламида/метиленбисакриламида. Наблюдаемые в некоторых пробах при специфическом окрашивании полосы выше зон, соответствующих трехмерным формам, свидетельствуют о возможной гексамеризации антигенов.
Таким образом, в данном примере продемонстрирована эффективная экспрессия DBD-Tul4, 6-His-FopA и DnaK-6His полученными конструкциями (псевдоаденовирусными частицами, несущими гены tul4, fopA и dnaK белковых антигенов F. tularensis Tul4, FopA, DnaK).
Пример 3. Определение в опытах на мышах иммуногенности полученных по заявленному изобретению рекомбинантных псевдоаденовирусных частиц, несущих гены белковых антигенов F. tularensis Tul4, FopA и DnaK.
Иммуногенность рекомбинантных аденовирусов, содержащих гены tul4, fopA и dnaK F. tularensis Tul4, FopA и DnaK оценивали на мышиной модели. Объектом исследования служили мыши (Mus musculus) линии Balb/c, самки, в возрасте ~4-5 недель.
Животные распределялись по группам по 10 особей по критерию массы тела таким образом, чтобы индивидуальное значение массы не отклонялось от среднего значения более чем на ±10%. В экспериментальной группе животные были иммунизированы интраназально по 50 мкл в каждую ноздрю препаратом, содержащим аденовирусные частицы трех типов. Одна доза вакцинного препарата (100 мкл) содержала: РПАН, несущие ген белка 6-His-FopA F. tularensis - 1,4×108 БОЕ; РПАН, несущие ген белка DBD-Tul4F. tularensis - 1,2×108 БОЕ; РПАН, несущие ген белка DnaK-6HisF. tularensis - 1,3×108 БОЕ. В контрольной группе мыши были иммунизированы физиологическим раствором.
Оценка состояния здоровья во время клинического осмотра проводилась по демонстрации животными неспровоцированного поведения, внешним признакам благополучного состояния здоровья, изменению массы тела, изменениям измеряемых клинических показателей и проявлению ожидаемой ответной реакции на стимул. Оценка выраженности боли, дистресса и страданий осуществлялась в соответствии с международными руководствами [Kohn D.F. et al. Public statement: guidelines for the assessment and management of pain in rodents and rabbits // J Am Assoc Lab Anim Sci, 2007. - №46(2). - c. 97-108; Recognition and Alleviation of Distress in Laboratory Animals, Washington, D.C., THE NATIONAL ACADEMIES PRESS, 2008, 132 p.],
В течение всего эксперимента состояние животных соответствовало физиологическому. Отклонений в поведении не отмечено. Проявлений болевого синдрома, признаков дистресса не выявлено. Животные на протяжении всего исследования были активные, реагировали на внешние стимулы. Выделение кала и мочи было нормальное. Шерстный покров был чистый и опрятный. Гибели животных зарегистрировано не было, вынужденная эвтаназия не проводилась. Для оценки иммуногенности проводили определение титров специфичных антител к каждому компоненту в сыворотке крови иммунизированных мышей до иммунизации, спустя 2 и 4 недели после иммунизации. Титр антител к белку FopA со временем менялся незначительно и к финалу эксперимента достигал значений немногим более 1:10000, как видно из фиг. 8б. В случае с DnaK, даже на ранних стадиях эксперимента титр к DnaK превышал 1:10000, а финальный титр достигал значений более 1:100000 (Фиг. 9б). Иммунный ответ на препарат, содержащий три вида аденовирусных частиц, продемонстрировал близкие показатели титра специфичных антител к Tul4, который составлял примерно 100000 в финальной пункции сыворотки (Фиг. 10б).
В контрольной группе животных не наблюдалось повышения уровня антител ни к одному из трех белков, как видно из фиг. 8а, 9а, 10а.
Таким образом, продемонстрирована иммуногенность полученных по заявленному изобретению рекомбинантных псевдоаденовирусных частиц, несущих гены белковых антигенов F. tularensis Tul4, FopA и DnaK.
Пример 4. Получение экспрессионных плазмид pTUL4, pFOPA и pDNAK.
а) Получение синтетических генов, кодирующих белки DBD-Tul4, 6-His-FopA и DnaK-6His.
Генно-инженерные и микробиологические манипуляции, амплификацию и секвенирование ДНК проводили по стандартным методикам [Маниатис Т., Фрич Э., Сэмбрук Дж. Молекулярное клонирование, М.: Мир. 1984. Клонирование ДНК. Методы. Под ред. Д. Гловера, Пер. с англ., М.: Мир. 1988; Saiki R.K., Gelfand D.H., Stoffel S., Scharf S.J., Higuchi R., Horn G.T., Mullis K.B., Erlich H.A. Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase // Science - 1988. Vol. 239 - P. 487-491; Sanger F., Nicklen S., Coulson A.R. DNA sequencing with chain-terminating inhibitors (DNA polymerase/nucleotide sequences/bacteriophage ФХ174) // Proc. Nat. Acad. Sci. - 1977. - Vol. 74, - Р. 5463-5467].
Синтетическая последовательность ДНК, соответствующая гену, кодирующему белок Tul4 (WP 003014685.1), была спланирована таким образом, чтобы нуклеотидный состав кодонов был оптимизирован для гетерологичной экспрессии в непатогенном лабораторном штамме Е. coli. Оптимизацию кодонного состава синтетического гена проводили с помощью программы JCat (http://www.jcat.de/), корректировку вторичной структуры транскрибируемой РНК - с помощью веб-сервера DINAMelt (http://mfold.rna.albany.edu/?q=DINAMelt/Two-state-folding). Синтетический ген tul4 был фланкирован на 5'-конце сайтом Ncol, а на 3'-конце - сайтом Kpn2I. Аналогичным образом были получены последовательности генов dnaK и fopA.
Нуклеотидные последовательности трех генов tul4, fopA, dnaK были синтезированы твердофазным амидофосфитным методом (ЗАО «Евроген», Россия) на синтезаторе Applied Biosystems ABI 3900.
б) Получение и клонирование рекомбинантных генов tul4, fopA, dnaK в плазмидные вектора.
Последовательности генов tul4, fopA, dnaK были встроены в плазмиду pQE6 под промотор фага Т5. Затем последовательность гена tul4 была клонирована в плазмиду pR1073, содержащую последовательность декстрансвязывающего домена из Leuconostoc mesenteroides (DBD1) и нуклеотидную последовательность GS-спейсера на С-конце. Полученная рекомбинантная плазмида pTul4 содержит нуклеотидную последовательность SEQ ID NO 15, включающую ген, кодирующий DBD, последовательность, кодирующую GS-спейсер, и нуклеотидную последовательность гена, кодирующего белок Tul4. Аминокислотная последовательность SEQ ID NO 16 соответствующего рекомбинантного белка DBD-Tul4 приведена в списке последовательностей.
Последовательности гена fopA была переклонирована в плазмиду pQE13, содержащую последовательность, кодирующую 6His на N-конце. Полученная рекомбинантная плазмида pFopA содержит нуклеотидную последовательность SEQ ID NO 17, включающую последовательность гена белка FopA и последовательность, кодирующую 6His на N-конце. Аминокислотная последовательность рекомбинантного белка 6-His-FopA SEQ ID NO 18 приведена в списке последовательностей.
Последовательность гена dnaK была переклонирована в плазмиду pQE16, содержащую нуклеотидную последовательность, кодирующую 6His на С-конце. Полученная рекомбинантная плазмида pDnaK содержит SEQ ID NO 19, включающую нуклеотидную последовательность гена белка DnaK и нуклеотидную последовательность, кодирующую 6His на С-конце. Аминокислотная последовательность рекомбинантного белка DnaK-6His SEQ ID NO 20 приведена в списке последовательностей.
Полученными конструкциями pTul4, pFopA и pDnaK трансформировали штамм Е. coli M15 [pREP4] и получали штаммы-продуценты Е. coli М15 [pREP4, pTul4], Е. coli М15 [pREP4, pFopA] и E.coli M15 [pREP4, pDnaK] с уровнем продукции белка Tul4 30%, уровнем продукции белка FopA 25% и уровнем продукции белка DnaK 20% от тотального белка клетки (фиг. 11).
Пример 5. Очистка рекомбинантного белка DBD-Tul4.
Для получения рекомбинантного белка DBD-Tul4 биомассу продуцента Е. coli М15 [pREP4, pTul4] ресуспендировали в 5 - 10-кратном объеме буферного раствора. Для лизиса продуцента DBD-Tul4 использовали буфер: 10 мМ Tris-HCl, рН 8,0; 50 мМ NaCl; 0,15 мМ PMSF; 0,1% Тритон Х-100; 0,5 мг/мл лизоцима. Инкубировали 1 ч при комнатной температуре, периодически перемешивая. Обрабатывали полученную суспензию ультразвуком для дезинтеграции клеток с помощью ультразвукового дезинтегратора. Центрифугировали 30 мин при 10000 g и температуре 5°C. Целевой белок DBD-Tul4 находился в тельцах включения (ТВ), т.е. в осадочной фракции. Осадок, полученный после лизиса биомассы и центрифугирования (ТВ), отмывали 10-кратным объемом буферного раствора: 10 мМ Трис-HCl, рН 8,0, 500 мМ NaCl, 1% Тритон Х-100 и растворяли в буфере: 10 мМ Tris-HCl, рН 8,0; 50 мМ NaCl; 8 М мочевины; 0,1% Тритон X-100. Центрифугировали при 12000 об/мин в течение 30 мин. Очистку белка проводили методом аффинной хроматографии на декстран содержащем сорбенте (сефадекс G-150). Раствор белка перед адсорбцией на сорбенте разбавляли в 4 раза буфером без мочевины 10 мМ Tris-HCl, рН 8,0; 50 мМ NaCl; 0,1% Тритон Х-100. Элюцию целевого белка проводили раствором 8 М мочевины в 10 мМ Tris-HCl, рН 8,0. Инкубировали полученные белковые фракции в буфере для элюции в течение суток для обеспечения конформационной гомогенности белка. Для удаления мочевины образцы диализовали против дистиллированной воды с двукратной сменой при температуре 4°C. Полученный раствор белка DBD-Tul4 расфасовывали в полипропиленовые пробирки и замораживали при температуре минус 20°C. Далее их хранили в замороженном виде при -70°C. Электрофореграмма образцов рекомбинантного белка DBD-Tul4 на различных стадиях очистки представлена на фиг. 12.
Пример 6. Очистка рекомбинантного белка 6-His-FopA.
Для получения рекомбинантного белка 6-His-FopA биомассу продуцента Е. coli М15 [pREP4, pFopA] ресуспендировали в 5 - 10-кратном объеме буферного раствора. Для лизиса использовали буфер: 20 мМ имидазол-HCl, рН 8,0; 50 мМ NaCl; 0,15 мМ PMSF; 0,1% Тритон Х-100; 0,5 мг/мл лизоцима. Инкубировали 1 ч при комнатной температуре, периодически перемешивая. Обрабатывали полученную суспензию ультразвуком для дезинтеграции клеток с помощью ультразвукового дезинтегратора. Центрифугировали 30 мин при 10000 g и температуре 5°C. Целевой белок 6-His-FopA находился в тельцах включения, т.е. в осадочной фракции. Осадок, полученный после лизиса и дезинтеграции 1 г биомассы и центрифугирования (ТВ), отмывали 10-кратным объемом буферного раствора: 10 мМ Трис-HCl, рН 8,0, 500 мМ NaCl, 1% Тритон Х-100 и растворяли в буфере: 20 мМ имидазол-HCl, рН 8,0; 0,5 М NaCl; 8 М мочевины; 0,1% Тритон Х-100. Центрифугировали при 12000 об/мин в течение 30 мин. Очистку белка проводили методом колоночной металл-хелатной аффинной хроматографии на сорбенте WorkBeads 40 Ni в денатурирующих условиях. Элюцию целевого белка проводили градиентом имидазола от 20 до 500 мМ в 7 М мочевине.
Инкубировали полученные белковые фракции в том же буфере, в котором проводилась элюция, в течение суток для обеспечения конформационной стабилизации белка. Для удаления имидазола и мочевины элюат диализовали против 1 л 10 мМ Tris-HCl, 150 мМ NaCl, рН 8,0 при температуре 4°C с двукратной сменой буфера. Раствор белка FopA расфасовывали в полипропиленовые пробирки и замораживали при температуре минус 20°C. Далее их хранили в замороженном виде при -70°C.
Электрофореграмма образцов рекомбинантного белка 6-His-FopA на различных стадиях очистки представлена на фиг. 13.
Пример 7. Иммунноблоттинг рекомбинантных белков DBD-Tul4, 6-His-FopA и DnaK-6His.
Для подтверждения сходства антигенных детерминант (эпитопов) рекомбинантных белков и соответствующих белков F. tularensis был проведен электрофорез рекомбинантных белков в полиакриламидном геле с последующим иммуноблоттингом с применением специфических кроличьих антисывороток (поликлональных антител), полученных к высокоочищенным рекомбинантным белкам DBD-Tul4, 6-His-FopA и DnaK-6His, и белкового экстракта аттенуированного вакцинного штамма туляремии (F. tularensis вакцинный штамм 15 НИИЭГ). Кроличьи антисыворотки к высокоочищенным рекомбинантным белкам DBD-Tul4, 6-His-FopA и DnaK-6His были получены с высокими титрами (в ИФА от 1:100000 до 1:500000), что позволило выявить присутствие соответствующих природных белков-антигенов в экстракте аттенуированного вакцинного штамма туляремии (F. tularensis вакцинный штамм 15 НИИЭГ), и, с другой стороны, подтвердило идентичность антигенных свойств природных белков и соответствующих рекомбинантных белков. В качестве отрицательного контроля использовался экстракт белков, приготовленный из суспензии клеток нетрансформированного штамма Е. coli M15, вкачестве положительного - экстракт клеточных белков вакцинного штамма F. tularensis 15 НИИЭГ. После электрофореза по Лэммли в полиакриламидном геле рекомбинантные белки перенесли на нитроцеллюлозную мембрану. Далее мембрану разделили на части, соответствующие отдельным белкам (как показано на фиг. 14) и инкубировали с растворами соответствующих кроличьих антисывороток (первичных антител) в разведении 1:2500 в TBS-TW с 0,5 мг/мл БСА. После инкубации мембрану промыли и инкубировали на шейкере при комнатной температуре в течение часа с козлиными антителами к иммуноглобулинам кролика, коньюгированными с пероксидазой хрена (p-GAR, ИМТЕК, Россия) в разведении 1:5000 и окрасили в растворе субстратов пероксидазы (диаминобензидина - ДАБ и перекиси водорода) в темноте до проявления четко видимых коричневых полос. Результат окрашивания ДАБ представлен на фиг. 14. В контрольном образце - экстракте белков штамма Е. coli M15, производными которого являются штаммы-продуценты рекомбинантных белков, окраска отсутствует.
Пример 8. Проверка иммуногенности рекомбинантных белков DBD-Tul4, 6-His-FopA и DnaK-6His.
Для эксперимента были использованы две группы самок мышей линии Balb/c с SPF-статусом здоровья, по 10 штук в каждой. Экспериментальной группе подкожно два раза с интервалом две недели вводили 50 мкл препарата, содержащего три типа рекомбинантных белков, а также адъювант. Препарат содержал по 10 мкг каждого рекомбинантного белка (DBD-Tul4, 6-His-FopA и DnaK-6His), в качестве адъюванта были использованы гидроксид алюминия - 1 мг и CpG олигонуклеотиды - 0,3 мг (по 0,15 мг CpG ODN 1585 и ODN 1826). Контрольной группе вводили раствор, содержащий адъюванты без рекомбинантных белков (гидроксид алюминия - 1 мг и CpG олигонуклеотиды - 0,3 мг (по 0,15 мг CpG ODN 1585 и ODN 1826)). Перед введением препарата и спустя 2 недели после каждого введения у всех животных брали кровь для оценки гуморального иммунного ответа. Для оценки иммуногенности компонентов препарата было проведено определение титров антител, специфичных к каждому из рекомбинантных белков, в контрольной и опытной группах.
Как видно на фиг. 15а, 16а, 17а, в течение всего эксперимента в пулированных сыворотках контрольной группы, которой был введен адъювант без рекомбинантных белков, не происходит нарастания титра антител, специфичных к FopA, DnaK и Tul4 - компонентам препарата, соответственно.
В то же время после двукратной иммунизации препаратом наблюдались высокие титры антител, специфичных к белку FopA (фиг. 15б). Столь же высокий финальный титр наблюдался для белка DnaK (см. фиг. 16б) и Tul4 (как показано на фиг. 17б).
Специфичность иммунного ответа к выбранным антигенам подтверждена с помощью иммуноблоттинга к рекомбинантным белкам и белкам лизата вакцинного штамма (см. фиг. 18). Нитроцеллюлозные мембраны после переноса белков инкубировали с первичной мышиной антисывороткой в разведении 1:2500. Затем мембраны инкубировали со вторичными антителами (конъюгированными с пероксидазой хрена кроличьими антителами против иммуноглобулинов мыши, IMTEK) в разведении 1:5000. Проявляли реакцией с диаминобензидином (ДАБ) и перекисью водорода.
Неспецифичной реакции в ответ на введение растворов и адъювантов, включенных в вакцину, не было обнаружено (фиг. 18). Результаты свидетельствуют о том, что полученные рекомбинантные белковые антигены F. tularensis (Tul4, DnaK, FopA) в совокупности с выбранными адъювантами обладают высокой иммуногенностью.
Пример 9. Введение рекомбинантных псевдоаденовирусных частиц, несущих гены белковых антигенов F. tularensis Tul4, FopA и DnaK в качестве прайм-компонента и препарата, содержащего рекомбинантные белки DBD-Tul4, 6-His-FopA и DnaK-6HisH адъювант в качестве бустерного компонента.
Для эксперимента были использованы две группы самок мышей линии Balb/c с SPF-статусом здоровья. Экспериментальной группе (10 животных) вводили прайм-компонент интраназально, по 50 мкл в каждую ноздрю, затем спустя две недели подкожно в качестве бустерной иммунизации вводили 50 мкл препарата, содержащего три типа рекомбинантных белков, а также адъювант.Бустерную иммунизацию повторяли спустя две недели.
Одна доза прайм-компонента (100 мкл) содержала: РПАН, несущие ген белка FopA F. tularensis - 1,4×108 БОЕ; РПАН, несущие ген белка Tul4 F. tularensis - 1,2×108 БОЕ; РПАН, несущие ген белка DnaK F. tularensis - 1,3×108 БОЕ.
Буст-компонент содержал по 10 мкг каждого рекомбинантного белка (Tul4, FopA и DnaK), в качестве адъюванта были использованы гидроксид алюминия - 1 мг и CpG олигонуклеотиды - 0,3 мг (по 0,15 мг CpG ODN 1585 и ODN 1826). Контрольной группе (10 животных) вводили физиологический раствор, затем, спустя две недели, раствор, содержащий адъюванты без рекомбинантных белков. На каждом этапе и спустя две недели после последней иммунизации у всех животных брали кровь для оценки гуморального иммунного ответа. Для оценки иммуногенности компонентов препарата было проведено определение титров антител, специфичных к каждому из рекомбинантных белков, в контрольной и опытной группах.
Как видно на фиг. 19а, 20а, 21а, в течение всего эксперимента в пулированных сыворотках контрольной группы, которой был введен адъювант без рекомбинантных белков, не происходит нарастания титра антител, специфичных к FopA, DnaK и Tul4 белкам, соответственно.
При иммунизации полной вакциной по 3-этапной схеме для белка FopA был получен титр, превышающий 1:100000 (см. фиг. 19), что заметно больше значений титров, полученных при введении прайм- и буст-компонентов по-отдельности (см. примеры 3 и 8, фиг. 8 и 15, соответственно).
В случае с DnaK также наблюдался высокий финальный титр, достигавший уровня 1:100000, как показано на фиг. 20.
Для белка Tul4, как и для антигенов FopA и DnaK, при иммунизации псевдоаденовирусными частицами с последующей двукратной буст-иммунизацией рекомбинантными антигенами в количестве 10 мкг (адъювант: CpG и А1) был получен высокий финальный титр, как показано на фиг. 21.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, ШТАММ ESCHERICHIA COLI M15 [pREP4, pTUL4spCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, РЕКОМБИНАНТНЫЙ БЕЛОК TUL4spCBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ TUL4spCBD | 2004 |
|
RU2270249C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА Ag85A-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pAg85A-DBD, ШТАММ Escherichia coli [pREP4, pAg85A-DBD], ХИМЕРНЫЙ БЕЛОК Ag85A-DBD | 2013 |
|
RU2520078C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА pESAT6-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pESAT6-DBD, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК ESAT6-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2520737C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ТРЕХ ГИБРИДНЫХ БЕЛКОВ ОБОЛОЧКИ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА, ОПРЕДЕЛЯЮЩИХ ПРИНАДЛЕЖНОСТЬ К СИБИРСКОМУ (DBD2-D3S), ЕВРОПЕЙСКОМУ (DBD2-D3E) И ДАЛЬНЕВОСТОЧНОМУ (DBD2-D3D) ПОДТИПАМ ВИРУСА; РЕКОМБИНАНТНЫЕ ПЛАЗМИДЫ pDBD2-D3S, pDBD2-D3E И pDBD2-D3D; ШТАММЫ-ПРОДУЦЕНТЫ Escherichia coli M15 [pREP4]; ХИМЕРНЫЕ БЕЛКИ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2560588C1 |
| Штамм рекомбинантной псевдоаденовирусной частицы, экспрессирующий химерный ген MBL-CT666 Chlamydia trachomatis, способ его получения, иммуногенная композиция для защиты от урогенитального хламидиоза человека | 2019 |
|
RU2721123C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pESAT6-CFP10-DBD, РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli M15 [pREP4, pESAT6-CFP10-DBD], СПОСОБ ПОЛУЧЕНИЯ, ИММОБИЛИЗАЦИИ, КОНЦЕНТРИРОВАНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОГО БЕЛКА ESAT6-CFP10-DBD НА ДЕКСТРАНЕ, РЕКОМБИНАНТНЫЙ БЕЛОК ESAT6-CFP10-DBD И ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ БЕЛОК ESAT6-CFP10-DBD | 2013 |
|
RU2539026C1 |
| Штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа Ad5-tetOFF-E3-HA125, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов H1, H2, H5 для создания противогриппозных иммуногенных препаратов, способ получения гена | 2018 |
|
RU2713722C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА CFP10-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pCFP10-DBD, ШТАММ Escherichia coli [pREP4, pCFP10-DBD], ХИМЕРНЫЙ БЕЛОК CFP10-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2546875C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pD4spGBD, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА D4-GBD, РЕКОМБИНАНТНЫЙ БЕЛОК D4-GBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ИССЛЕДОВАНИЯ СВЯЗЫВАНИЯ БЕЛКА D4-GBD С АНТИТЕЛАМИ СЫВОРОТОК БОЛЬНЫХ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ D4-GBD | 2008 |
|
RU2401305C2 |
| Универсальная противогриппозная вакцина | 2015 |
|
RU2618918C2 |

















Изобретение относится к биотехнологии. Представлена иммуногенная композиция, включающая (i) прайм-компонент, состоящий из рекомбинантных псевдоаденовирусных частиц: частиц, несущих ген tul4 Francisella tularensis с последовательностью SEQ ID NO 1, частиц, несущих ген fopA Francisella tularensis с последовательностью SEQ ID NO 3, и частиц, несущих ген dnaK Francisella tularensis с последовательностью SEQ ID NO 5, при этом указанные гены оптимизированы для экспрессии в эукариотических клетках, и/или (ii) бустерный компонент, включающий рекомбинантные белки: белок DBD-Tul4 с последовательностью SEQ ID NO 16, молекулярной массой 30,0 кДа, содержащий декстрансвязывающий домен декстрансукразы из L. citreum КМ20, представленный аминокислотными остатками 1-140 SEQ ID NO 16, Gly-Ser спейсер, представленный остатками 141-150 SEQ ID NO 16, и последовательность белка Tul4 Francisella tularensis, представленную остатками 151-282 SEQ ID NO 16; белок 6-His-FopA с последовательностью SEQ ID NO 18, молекулярной массой 40,5 кДа; белок DnaK-6His с последовательностью SEQ ID NO 20, молекулярной массой 70,4 кДа, а также содержащая фармацевтически приемлемый носитель или разбавитель и способ ее получения. Изобретение позволяет получить стабильный и устойчивый иммунный ответ на Francisella tularensis. 2 н. и 3 з.п. ф-лы, 21 ил., 1 табл., 9 пр.
1. Иммуногенная композиция, включающая (i) прайм-компонент, состоящий из рекомбинантных псевдоаденовирусных частиц: частиц, несущих ген tul4 Francisella tularensis с последовательностью SEQ ID NO 1, частиц, несущих ген fopA Francisella tularensis с последовательностью SEQ ID NO 3, и частиц, несущих ген dnaK Francisella tularensis с последовательностью SEQ ID NO 5, при этом указанные гены оптимизированы для экспрессии в эукариотических клетках, и/или (ii) бустерный компонент, включающий рекомбинантные белки: белок DBD-Tul4 с последовательностью SEQ ID NO 16, молекулярной массой 30,0 кДа; белок 6-His-FopA с последовательностью SEQ ID NO 18, молекулярной массой 40,5 кДа; белок DnaK-6His с последовательностью SEQ ID NO 20, молекулярной массой 70,4 кДа, а также содержащая фармацевтически приемлемый носитель или разбавитель.
2. Иммуногенная композиция по п. 1, где указанная композиция представляет собой вакцину, индуцирующую иммунный ответ против Francisella tularensis.
3. Иммуногенная композиция по пп. 1 и 2, где указанная композиция используется для индукции специфического клеточного и гуморального иммунитета против Franciella tularensis посредством введения прайм-компонента интраназально и введения бустерного компонента подкожно, дважды с интервалом в две недели.
4. Способ получения иммуногенной композиции по п. 1, включающий получение трех типов псевдоаденовирусных частиц, несущих гены белковых антигенов F. tularensis Tul4, FopA и DnaK соответственно, при этом бактериальный лидерный пептид удаляют и добавляют эукариотический лидерный пептид, каждый ген клонируют в свой плазмидный вектор pSh-PA-ILZ-Fc, куда также клонируют изолейциновый зиппер (ILZ) и Fc-фрагмент антитела IgG2a (Fc), полученные линеаризованные векторы pSh-FopA, pSh-Tul4 и pSh-DnaK смешивают с геномной ДНК рекомбинантного аденовируса Ad5 и котрансформируют в клетки Е. coli штамма BJ5183, полученные плазмиды трансформируют в штамм Е. coli DH5alpha, затем клетки линии 293 трансфецируют линеаризованными плазмидами, лизируют и из полученного лизата выделяют псевдоаденовирусные частицы; получают рекомбинантные белки DBD-Tul4, 6-His-FopA и DnaK-6His, для чего получают плазмиды, содержащие синтетические ДНК с последовательностями, соответствующими генам, кодирующим белки DBD-Tul4, 6-His-FopA и DnaK-6His, где нуклеотидный состав кодонов оптимизирован для гетерологичной экспрессии в непатогенном лабораторном штамме Е. coli, на основе указанных плазмид TUL4, pFOPA и pDNAK получают штаммы-продуценты Е. coli М15 [pREP4, pTul4], Е. coli M15 [pREP4, pFopA] и E. coli M15 [pREP4, pDnaK] с высокой продукцией рекомбинантных белков DBD-Tul4, 6-His-FopA и DnaK-6HisB тельцах включения (ТВ), дальнейшая очистка всех индивидуальных рекомбинантных белков DBD-Tul4, 6-His-FopA и DnaK-6His производится путем растворения ТВ в денатурирующих условиях и проведения колоночной аффинной хроматографии в денатурирующих условиях на носителях, содержащих в качестве неподвижной фазы декстран (сефадекс) в случае DBD-Тil4 или содержащих иммобилизованный металл (металл-хелатная хроматография) в случае 6-His-FopA и DnaK-6His, с последующим удалением денатурирующих агентов из раствора белков, на основе полученных псевдовирусных частиц и белков готовят композицию, содержащую либо псевдовирусные частицы, либо белки, либо комбинацию псевдовирусных частиц и белков.
5. Способ по п. 4, где псевдовирусные частицы и белки подвергают лиофильной сушке.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, ШТАММ ESCHERICHIA COLI M15 [pREP4, pTUL4spCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, РЕКОМБИНАНТНЫЙ БЕЛОК TUL4spCBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ TUL4spCBD | 2004 |
|
RU2270249C1 |
| US 20140356415 A1, 04.12.2014. | |||

