Все документы, приведенные в контексте данного документа, включены полностью путем ссылки.
ОБЛАСТЬ ТЕХНИКИ
Данное изобретение лежит в области вакцинных композиций и способов иммунизации с применением вакцинных композиций.
УРОВЕНЬ ТЕХНИКИ
Первое введение вакцинной композиции, содержащей антиген патогенного микроорганизма, вызывает первичный иммунный ответ на антиген в виде как активированных клеток, так и клеток памяти. Последующее подвергание антигену (например, подвергание патогенному микроорганизму) вызывает развитие клеток памяти и вторичный иммунный ответ, который является более быстрым и сильным, чем первичный иммунный ответ, обеспечивая защиту от патогенного микроорганизма.
Хотя клетки памяти могут сохраняться в течение месяцев или даже лет после первичного подвергания антигену, обычно необходимо обеспечить повторную иммунизацию антигеном, для того чтобы гарантировать поддержание долговременной иммунологической памяти. Таким образом, режимы вакцинации часто включают несколько повторяющихся через короткие интервалы инъекций, для того чтобы обеспечить исходный банк клеток памяти, и последующие вторичные инъекции антигена с увеличенными интервалами, для того чтобы поддерживать иммунологическую память. Необходимость нескольких повторяющихся через короткие интервалы инъекций и частота, с которой требуется вторичная инъекция антигена, варьируется в зависимости от вакцины и возраста пациента.
Была бы преимущественной способность уменьшать число возбуждающих доз и частоту и число вторичных инъекций антигена, не подвергая опасности поддержание иммунологической памяти.
В идеале было бы предпочтительным полностью устранить необходимость дополнительных возбуждающих доз и повторных доз антигена и вводить вакцины в виде однократной дозы. Следовательно, объектом изобретения является получение иммуногенных композиций, которые вызывают долговременную иммунологическую память в отсутствие повторных доз антигена и/или в отсутствие множественных возбуждающих доз.
Кроме того, объектом настоящего изобретения является получение иммуногенных композиций, содержащих антигены вируса гриппа, стрептококка группы B и менингококка серологической группы B.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Вакцины часто содержат вспомогательные вещества для усиления иммунной активности. Примеры известных вспомогательных веществ включают соли алюминия, эмульсии типа «масло в воде», сапонины, цитокины, липиды и олигонуклеотиды CpG. В настоящее время для применения у человека одобрены только соли алюминия, 3-де-O-ацилированный монофосфорил-липид A ('3dMPL') и MF59.
Другой молекулой, которая, как известно, имеет свойства вспомогательного вещества, является α-галактозилцерамид (α-GalCer или α-GC), гликолипид, точнее сказать, гликозилцерамид, первоначально выделенный из морских губок [I]. α-GalCer является лигандом класса MHC I-подобных молекул, CD1d, и представляет собой молекулы CD1d к инвариантным природным клеткам T киллерам (NKT). α-GalCer был первоначально исследован на его способность вызывать NKT-клеточный иммунный ответ на опухолевые клетки [2]. Было также показано, что инвариантные NKT-клетки вызывают активацию B-клеток, усиление пролиферации B-клеток и выработку антител [3,4]. Было показано, что α-GalCer действует в качестве вспомогательного вещества для многообразия вводимых одновременно белковых антигенов [5]. Было показано, что одновременное введение α-GalCer с облученными спорозоитами или рекомбинантными вирусами, экспрессирующими антиген малярии, увеличивает уровень защитного иммунитета против малярии у мышей [6]. Было также показано, что α-GalCer действует в качестве вспомогательного вещества для ДНК вакцины, кодирующей HIV-1 ген капсидных белков (gag) и ген белка оболочки (env) [7], и вызывает гуморальный и клеточный иммунный ответ на вирус гриппа HA при введении интраназально [8].
Как не удивительно, но было обнаружено на данный момент, что применение лиганда CD1d, такого как α-GalCer, в качестве вспомогательного вещества вакцины не только значительно усиливает иммунный ответ на антигены в вакцине, но и также вызывает увеличение специфического B-клеточного пула памяти против данных антигенов. В частности, было обнаружено, что введения однократной дозы композиции, содержащей α-GalCer и антиген, достаточно, для того чтобы способствовать увеличению специфического B-клеточного пула памяти, который усиливает иммунный ответ на контрольное заражение антигеном даже через один год. Способность данного лиганда CD1d содействовать увеличению специфического B-клеточного пула памяти показывает, что применение CD1d лигандов в качестве вспомогательных веществ вакцины может уменьшить число и частоту возбуждающих и повторных доз антигена, необходимых для получения долговременной иммунологической памяти.
Также было обнаружено, что лиганды CD1d являются, как не удивительно, эффективными вспомогательными веществами для антигенов, полученных из стрептококка группы B, менингококка серологической группы B и для антигенов определенного вируса гриппа.
Способы получения долговременной иммунологической памяти
Изобретение обеспечивает способ получения долговременной иммунологической памяти к антигену у пациента, который в этом нуждается, включающий введение указанному пациенту композиции, содержащей:
a) указанный антиген; и
b) лиганд CD1d, таким образом, что число и/или частота доз указанной композиции, необходимые для того, чтобы указанный пациент был способен на иммунный ответ при последующем подвергании указанному антигену, понижены по сравнению с введением указанного антигена в отсутствие лиганда CD1d.
Предпочтительно способ данного изобретения уменьшает число и/или частоту доз указанной композиции, необходимой указанному пациенту, для того чтобы быть способным усиливать защитный иммунный ответ при последующем подвергании указанному антигену, по сравнению с введением указанного антигена в отсутствие лиганда CD1d. Под «защитным иммунным ответом» подразумевается, что усиленный иммунный ответ при последующем подвергании антигену является достаточным, для того чтобы предотвратить приобретение пациентом заболевания, связанного с антигеном. Уменьшение числа и/или частоты доз данной композиции, необходимой, для того чтобы усилить защитный иммунный ответ на антиген, можно измерить посредством стандартных способов, известных на данном уровне техники.
Способ изобретения может уменьшать число и/или частоту доз композиции, содержащей антиген, необходимые, для того чтобы вызвать защитный иммунный ответ при последующем подвергании данному антигену. Некоторые иммунизации в настоящее время требуют трех или четырех возбуждающих доз антигена, для того чтобы усилить защитный иммунный ответ при последующем подвергании антигену. Предпочтительно способ изобретения уменьшает число доз, необходимое, для того чтобы вызвать защитный иммунный ответ на антиген, до однократной возбуждающей дозы.
Современные способы иммунизации также часто требуют повторных иммунизаций с увеличенными интервалами, для того чтобы поддерживать защитный иммунный ответ при последующем подвергании данному антигену. Например, иммунизации, проведенные в грудном возрасте, обычно предполагают повторные дозы антигена, вводимые через месяцы или годы после введения первоначальной дозы. Предпочтительно способ изобретения уменьшает частоту повторных доз композиции, содержащей антиген, необходимую, для того чтобы поддерживать защитный иммунный ответ при последующем подвергании данному антигену. Предпочтительно способ изобретения позволяет вводить повторные дозы антигена через интервалы более одного года, предпочтительно более двух лет, предпочтительно более 5 лет, предпочтительно более 10 лет. Согласно предпочтительному варианту осуществления изобретения необходимость в повторных дозах антигена полностью исключена и, для того чтобы вызвать защитный иммунный ответ при последующем подвергании данному антигену, достаточно однократной дозы антигена.
Согласно одному аспекту изобретения обеспечивается способ вызывания иммунного ответа на антиген у пациента, включающий введение указанному пациенту:
a) указанного антигена; и
b) лиганда CD1d,
причем указанный антиген и лиганд CD1d вводили также указанному пациенту предварительно более чем за год.
Изобретение также обеспечивает применение антигена и лиганда CD1d в производстве лекарственного средства, для того чтобы вызывать защитный иммунный ответ у пациента, причем указанный антиген и лиганд CD1d вводили также указанному пациенту предварительно более чем за 1 год.
Предпочтительно иммунный ответ является защитным иммунным ответом. Предпочтительно указанный антиген и лиганд CD1d вводили указанному пациенту предварительно более чем за 18 месяцев, предпочтительно предварительно более чем за 2 года, 5 лет или 10 лет.
Данный антиген и лиганд CD1d, вводимые пациенту, согласно данному аспекту изобретения можно вводить в виде смеси, т.е. в виде единой композиции, содержащей как антиген, так и лиганд CD1d. В качестве альтернативы данный антиген и лиганд CD1d можно вводить пациенту последовательно в одном и том же месте, причем первым вводится либо данный антиген, либо лиганд CD1d. Данный антиген и лиганд CD1d можно также вводить пациенту раздельно в разных местах, например, в разные конечности. Первоначальную дозу лиганда CD1d и антигена, вводимую пациенту предварительно более чем за 1 год, можно также вводить в виде единой композиции лиганда CD1d и антигена, или лиганд CD1d и антиген можно было вводить последовательно или раздельно.
Количество лиганда CD1d, вводимое пациенту, для того чтобы вызывать иммунный ответ, может варьироваться в зависимости от возраста и веса пациента, которому вводят композицию, но будет обычно содержать между 1-100 мкг/кг массы тела пациента. Как не удивительно, но было обнаружено, что низкие дозы лиганда CD1d являются достаточными, для того чтобы усиливать иммунный ответ при одновременном введении антигена и поддерживать долговременную иммунологическую память на данный антиген. Количество лиганда CD1d, включенного в композиции изобретения, может, следовательно, быть меньше 50 мкг/кг массы тела пациента, меньше 20 мкг/кг, меньше 10 мкг/кг, меньше 5 мкг/кг, меньше 4 мкг/кг или меньше 3 мкг/кг.
Согласно дополнительному аспекту изобретения обеспечивается способ вызывания иммунного ответа на антиген у пациента, включающий введение указанному пациенту:
a) указанного антигена; и
b) лиганда CD1d,
причем количество лиганда CD1d, включенного в композицию, меньше 10 мкг/кг массы тела пациента, предпочтительно меньше 5 мкг/кг, меньше 4 мкг/кг или меньше 3 мкг/кг.
Изобретение также обеспечивает применение антигена и лиганда CD1d в производстве лекарственного средства, для того чтобы вызывать иммунный ответ у пациента, причем количество лиганда CD1d меньше 10 мкг/кг массы тела пациента, предпочтительно меньше 5 мкг/кг, меньше 4 мкг/кг или меньше 3 мкг/кг.
Данный антиген и лиганд CD1d, вводимые пациенту, согласно данному аспекту изобретения можно: вводить в виде смеси; вводить пациенту последовательно в одном и том же месте (причем первым вводится либо данный антиген, либо лиганд CD1d); или вводить пациенту раздельно в разных местах, например, в разные конечности.
Лиганды CDld
Лигандом CD1d, включенным в композиции изобретения, может быть любая молекула, которая связывается с молекулой CD1d. Молекулы CD1d находятся на инвариантных клетках NKT (iNKT), B-клетках, дендритных клетках, мононуклеарных клетках и обычных T-клетках, а лиганды CD1d изобретения могут связываться с молекулами CD1d, находящимися на любой из данных клеток. Связывание лигандов CD1d изобретения с молекулами CD1d может активировать клетки iNKT, B-клетки, дендритные клетки, мононуклеарные клетки и обычные T-клетки. Предпочтительно связывание лигандов CD1d с молекулами CD1d активирует клетки iNKT. Способность молекулы связываться с молекулой CD1d можно определить стандартными способами, известными в данной области техники. Способность лиганда CD1d активировать клетки, в частности инвариантные клетки NKT, можно определить измерением уровней цитокинов, высвобождающихся из клеток в присутствии лиганда CD1d, по сравнению с уровнями цитокинов, высвобождающихся в отсутствие лиганда CD1d. Предпочтительно лиганды CD1d, включенные в композиции изобретения, повышают уровень секреции цитокинов инвариантными клетками NKT по сравнению с уровнем секреции цитокинов инвариантными клетками NKT в отсутствие лиганда CD1d. Лиганды CD1d данного изобретения могут способствовать высвобождению цитокинов или Th2 цитокинов. Предпочтительно лиганды CD1d данного изобретения повышают уровни IFN-γ, IL-4 и IL-13, секретируемых инвариантными клетками NKT, по сравнению с уровнями IFN-γ, IL-4 и IL-13, секретируемых инвариантными клетками NKT в отсутствие лиганда CD1d.
Подходящие молекулы, которые можно исследовать на способность действовать в качестве лигандов CD1d, которые активируют инвариантные клетки NKT, включают пептиды и сахариды. Предпочтительно лигандами CD1d данного изобретения являются гликолипиды. Обзор гликолипидных антигенов, которые, как известно, действуют в качестве лигандов CD1d, которые могут быть включены в композиции данного изобретения, предоставляется в ссылке 9.
Примеры подходящих лигандов CD1d для применения в композициях данного изобретения включают α-гликозилцерамиды. α- Гликозилцерамидами, применяемыми в композициях данного изобретения, предпочтительно являются соединения формулы (I):
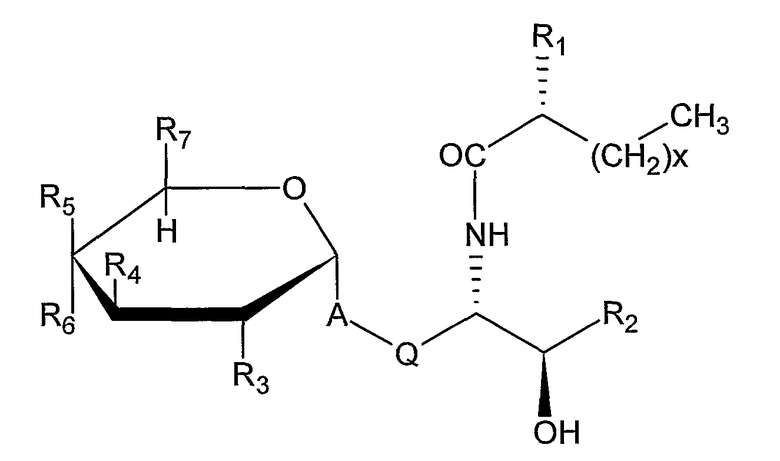 ,
,
где
A представляет собой O, CH2, -CH2CH=CH, -CH=CHCH2,
Q представляет собой (CH2)n, причем n представляет собой целое число 0 или 1,
R1 представляет собой H или OH,
X представляет собой целое число между 1 и 30,
R2 представляет собой заместитель, выбранный из группы, состоящей из следующих от (a) до (e) (причем Y представляет собой целое число между 5 и 17);
(a) -CH2(CH2)YCH3
(b) -CH(OH)(CH2)YCH3
(c) -CH(OH)(CH2)YCH(CH3)2
(d) -CH=CH(CH2)YCH3
(e) -CH(OH)(CH2)YCH(CH3)CH2CH3,
R3 представляет собой H, OH, NH2, NHCOCH3 или моносахарид,
R4 представляет собой OH или моносахарид,
R5 представляет собой H, OH или моносахарид,
R6 представляет собой H, OH или моносахарид, и
R7 представляет собой H, CH3, CH2OH или -CH2-моносахарид.
X предпочтительно представляет собой между 7 и 27, более предпочтительно между 9 и 24 и более предпочтительно между 13 и 20. Y предпочтительно представляет собой между 7 и 15 и более предпочтительно между 9 и 13.
Понятие «моносахарид» означает молекулу сахара, имеющую цепь из 3-10 атомов углерода в форме альдегида (альдоза) или кетона (кетоза). Подходящие моносахариды для применения в данном изобретении включают как природные, так и синтетические моносахариды. Типовые моносахариды включают триозы, такие как глицероза и дигидроксиацетон; тетрозы, такие как эритроза и эритрулаза; пентозы, такие как ксилоза, арабиноза, рибоза, ксилулоза, рибулоза; метилпентозы (6-деоксигексозы), такие как рамноза и фруктоза; гексозы, такие как глюкоза, манноза, галактоза, фруктоза и сорбоза; гептозы, такие как глюкогептоза, галаманногептоза, седогептулоза и манногептулоза. Предпочтительными моносахаридами являются гексозы.
Моносахаридные группы могут быть присоединены к структуре при R3, R4, R5, R6 или R7 положении для образования гликозидной связи. Обычно моносахарид присоединяется к R3, R4, R5, R6 или R7 положению через кислород, связанный с C-1 атомом углерода моносахарида, образуя гликозидную связь.
Если R3 представляет собой моносахарид, то он предпочтительно выбирается из α-D-галактопиранозы, β-D-галактопиранозы, α-D-глюкопиранозы или β-D-глюкопиранозы.
Если R4 представляет собой моносахарид, то он предпочтительно выбирается из β-D-галактофуранозы или N-ацетил α-D-галактопиранозы.
Если R5 представляет собой моносахарид, то он предпочтительно выбирается из α-D-галактопиранозы, β-D-галактопиранозы, α-D-глюкопиранозы или β-D-глюкопиранозы.
Если R6 представляет собой моносахарид, то он предпочтительно выбирается из α-D-галактопиранозы, β-D-галактопиранозы, α-D-глюкопиранозы или β-D-глюкопиранозы.
Если R7 представляет собой моносахарид, то он предпочтительно выбирается из метил α-D-галактопиранозида, метил β-D-галактопиранозида, метил α-D-глюкопиранозида или метил β-D-глюкопиранозида.
Предпочтительно R5 и R6 отличаются. Предпочтительно один из R5 и R6 представляет собой H.
Дополнительные примеры α-гликозилцерамидов, подходящих для включения в композиции данного изобретения, предоставлены в ссылке 2.
Предпочтительно α-гликозилцерамид представляет собой α-галактозилцерамид (α-GalCer), имеющий формулу, данную ниже, или его аналог:

α-GalCer и его аналоги, включенные в композиции данного изобретения, могут быть выделены непосредственно из морских губок или могут быть химически синтезированными продуктами.
Примеры аналогов α-GalCer, подходящих для применения в композициях данного изобретения, и способы синтеза данных продуктов предоставлены в ссылках 10 и 11. Предпочтительным аналогом α-GalCer является KRN7000, который имеет формулу (2S,3S,4R)-l-O-(α-D-галактопиранозил)-2-(N-гексакозаноиламино)-l,3,4-октадекантриол. Синтез KRN7000 описывается в ссылке 12.
Кроме того, предпочтительными аналогами α-GalCer являются C-сопряженные аналоги α-GalCer, такие как аналоги, описанные в ссылках 13, 14 и 15. Предпочтительным C-сопряженным аналогом α-GalCer является CRONY-101, синтез которого описывается в ссылке 13.
Усеченные аналоги α-GalCer, в которых жирная ацильная цепь и/или сфингозиновая цепь усечены по сравнению с α-GalCer, также можно применять в данном изобретении. Примеры усеченных аналогов α-GalCer предоставлены в ссылке 16. Предпочтительным усеченным аналогом α-GalCer является 'OCH', в котором жирная ацильная цепь имеет усечение двух углеводородов, а сфингозиновая цепь имеет усечение девяти углеводородов по сравнению с предпочтительным α-GalCer (т.е. R1=H, X=21, R2=CH(OH)(CH2)4CH3, R3=OH, R4=OH, R5=OH, R6=H и R7=CH2OH).
Предпочтительные дополнительные усеченные аналоги α-GalCer включают аналоги, в которых жирная ацильная цепь имеет усечение двух углеводородов, а сфингозиновая цепь имеет усечение семи или трех углеводородов по сравнению с α-GalCer (т.е. R1=H, X=21, R3=OH, R4=OH, R5=OH, R6=H, R7=CH2OH, а R2 представляет собой либо CH(OH)(CH2)6CH3, либо CH(OH)(CH2)10CH3).
α-GalCer, KRN7000 и OCH все являются фитосфингозин-содержащими α-гликозилцерамидами. Однако данное изобретение также включает применение сфинганин-содержащих аналогов KRN700, OCH и других α-гликозилцерамидов, описанных выше. Синтез сфинганин-содержащих аналогов KRN700 и OCH описывается в ссылке 17.
Лиганды CD1d, применяемые в композициях данного изобретения, могут также включать сульфатидные аналоги, такие как аналоги, описанные в ссылке 18. Предпочтительным аналогом α-GalCer является 3"-O-сульфогалактозилцерамид.
Хотя α-GalCer первоначально был выделен из морских губок, лиганды CD1d структуры, подобной α-GalCer, недавно были выделены из грамотрицательных бактерий. Дополнительными лигандами CD1d, которые могут быть включены в композиции данного изобретения, являются, таким образом, гликолипиды бактериального происхождения и, в частности, бактериальные гликозилцерамиды, выделенные из внешней мембраны Sphingomonas и Ehrlichia. Примеры подобных гликозилцерамидов включают α-глюкуронозилцерамид и α-галактуронозилцерамид из Sphingomonas, получение которых описывается в ссылке 19. Получение дополнительных лигандов CD1d из Sphingomonas и из Borrelia описывается в ссылке18.
Изобретение также включает применение лигандов CD1d, которые не принадлежат к гликосфинголипидному семейству. В частности, изобретение включает применение лигандов CD1d, которые являются гликоглицеролипидами. Гликоглицеролипиды, которые можно применять в данном изобретении, включают диацилглицерины, в частности моногалактозилдиацилглицерины. Подходящие моногалактозилдиацилглицерины для применения в данном изобретении описываются в ссылке 20.
Антигенные компоненты композиции
Антигеном, включенным в композицию для вызывания долговременной иммунологической памяти, описанным выше, может быть любой антиген, известный для применения в вызывании иммунного ответа. Антиген может содержать белковый антиген или сахаридный антиген.
Сахаридные антигены
Если антиген представляет собой сахаридный антиген, то он предпочтительно соединен с белком-носителем. Предпочтительно сахаридный антиген представляет собой бактериальный сахарид и, в частности, бактериальный капсулярный сахарид.
Примеры бактериальных капсулярных сахаридов, которые могут быть включены в композиции данного изобретения, включают капсулярные сахариды из Neisseria meningitidis (серологические группы A, B, C, W135 или Y), Streptococcus pneumoniae (серологические типы 4, 6B, 9V, 14, 18C, 19F или 23F), Streptococcus agalactiae (типы Ia, Ib, II, III, IV, V, VI, VII или VIII), Haemophilus influenzae (типируемые штаммы: a, b, c, d, e или f), Pseudomonas aeruginosa, Staphylococcus aureus и т.д. Другие сахариды, которые могут быть включены в композиции данного изобретения, включают глюканы (например, грибковые глюканы, такие как глюканы в Candida albicans), и грибковые капсулярные сахариды, например, из мембраны Cryptococcus neoformans.
Мембрана N. meningitidis серологической группы A (MenA) представляет собой гомополимер (α1→6)-связанного N-ацетил-D- маннозамин-1-фосфата с частичным O-ацетилированием в C3 и C4 положениях. Мембрана N. meningitidis серологической группы B (MenB) представляет собой гомополимер (α2→8)-связанной сиаловой кислоты. Капсулярный сахарид N. meningitidis серологической группы C (MenC) представляет собой гомополимер (α2→9)-связанной сиаловой кислоты с переменным O-ацетилированием в положениях 7 и/или 8. Сахарид N. meningitidis серологической группы W135 представляет собой полимер, состоящий из звеньев дисахарида сиаловая кислота-галактоза [→4)-D-Νeup5Ac(7/9OAc)-α-(2→6)-D-Gal-α-(l→]. Он имеет переменное O-ацетилирование в 7 и 9 положениях сиаловой кислоты [21]. Сахарид N. meningitidis серологической группы Y представляет собой сахарид, подобный сахариду серологической группы W135, за исключением того, что повторяющееся дисахаридное звено включает глюкозу вместо галактозы [→4)-D-Νeup5Ac(7/9OAc)-α-(2→6)-D-Glc-α-(l→]. Он также имеет переменное O-ацетилирование в 7 и 9 положениях сиаловой кислоты.
Капсулярный сахарид H. influenzae типа b (Ηib) представляет собой полимер рибозы, рибитола и фосфата ['PRP', (poly-3-β-D-рибоза-(l,l)-D-рибитол-5-фосфат)].
Композиции данного изобретения могут содержать смеси конъюгатов сахаридных антигенов. Предпочтительно композиции данного изобретения содержат сахаридные антигены более чем из одной серологической группы N. meningitidis, например, композиции могут содержать сахаридные конъюгаты из серологических групп A+C, A+W135, A+Y, C+W135, C+Y, W135+Y, A+C+W135, A+C+Y, C+W135+Y, A+C+W135+Y и т.д. Предпочтительные композиции содержат сахаридные конъюгаты из серологических групп C и Y. Другие предпочтительные композиции содержат сахаридные конъюгаты из серологических групп C, W135 и Y.
Если смесь содержит менингококковые сахариды из серологической группы A и, по крайней мере, один сахарид другой серологической группы, соотношение (w/w) MenA сахарида и сахарида любой другой серологической группы может быть больше 1 (например, 2:1, 3:1, 4:1, 5:1, 10:1 или выше). Предпочтительные соотношения (w/w) для сахаридов из серологических групп A:C:W135:Y представляют собой: 1:1:1:1; 1:1:1:2; 2:1:1:1; 4:2:1:1; 8:4:2:1; 4:2:1:2; 8:4:1:2; 4:2:2:1; 2:2:1:1; 4:4:2:1; 2:2:1:2; 4:4:1:2 и 2:2:2:1.
Кроме того, предпочтительные композиции данного изобретения содержат Hib сахаридный конъюгат и сахаридный конъюгат, по крайней мере, из одной серологической группы N. meningitidis, предпочтительно более чем из одной серологической группы N. meningitidis. Например, композиция данного изобретения может содержать Hib конъюгат Hib и конъюгаты из N. meningitidis серологических групп A, C, W135 и Y.
Изобретение, кроме того, включает композиции, содержащие сахаридные конъюгаты Streptococcus pneumoniae. Предпочтительно композиции содержат сахаридные конъюгаты более чем из одного серологического типа Streptococcus pneumoniae. Предпочтительные композиции содержат сахаридные конъюгаты из Streptococcus pneumoniae серологических типов 4, 6B, 9V, 14, 18C, 19F и 23F (7-валентные). Композиции могут, кроме того, содержать сахаридные конъюгаты из Streptococcus pneumoniae серологических типов 4, 6B, 9V, 14, 18C, 19F, 23F, 1 и 5 (9-валентные) или могут содержать сахаридные конъюгаты из Streptococcus pneumoniae серологических типов 4, 6B, 9V, 14, 18C, 19F, 23F, 1, 5, 3 и 7F (11-валентные).
Кроме того, предпочтительные композиции данного изобретения содержат пневмококковые сахаридные конъюгаты и сахаридные конъюгаты из Hib и/или N. meningitidis. Предпочтительно композиции данного изобретения могут содержать сахаридные конъюгаты из S. pneumoniae серологических типов 4, 6B, 9V, 14, 18C, 19F и 23F и Hib сахаридный конъюгат. Предпочтительно композиции данного изобретения могут содержать сахаридные конъюгаты из S. pneumoniae серологических типов 4, 6B, 9V, 14, 18C, 19F и 23F и сахаридные конъюгаты из N. meningitidis серологических групп A, C, W135 и Y. Композиции согласно данному изобретению могут также содержать сахаридные конъюгаты из S. pneumoniae серологических типов 4, 6B, 9V, 14, 18C, 19F и 23F, Hib сахаридный конъюгат и сахаридные конъюгаты из N. meningitidis серологических групп A, C, W135 и Y.
Предпочтительным является, что защитная эффективность индивидуальных конъюгатов сахаридных антигенов не устраняется посредством их комбинирования, хотя фактическая иммуногенность (например, ELISA титры) может уменьшаться.
Получение капсулярных сахаридных антигенов
Способы получения капсулярных сахаридных антигенов хорошо известны. Например, ссылка 22 описывает получение сахаридных антигенов из N. meningitidis. Получение сахаридных антигенов из H. influenzae описывается в главе 14 ссылки 86. Получение сахаридных антигенов и конъюгатов из S. pneumoniae описывается на данном уровне техники. Например, Prevenar™ представляет собой вакцину 7-валентного пневмококкового конъюгата. Процесс получения сахаридных антигенов из S. agalactiae описывается подробно в ссылках 23 и 24.
Сахаридные антигены можно химически модифицировать. Например, они могут быть модифицированы, чтобы заместить одну или более гидроксильных групп защитными группами. Это, в частности, применимо для менингококковой серологической группы A, причем ацетильные группы можно заместить защитными группами, для того чтобы предотвратить гидролиз [25]. Подобные модифицированные сахариды представляют собой все-таки сахариды серологической группы A в пределах содержания изобретения.
Капсулярные сахариды можно применять в форме олигосахаридов. Такие олигосахариды обычно образуются посредством фрагментации очищенного капсулярного полисахарида (например, посредством гидролиза), после которой будет обычно следовать очистка фрагментов необходимого размера.
Фрагментация полисахаридов осуществляется предпочтительно для достижения результирующей средней степени полимеризации (СП) в олигосахариде менее 30. СП можно обычно измерить посредством ионообменной хроматографии или посредством колориметрических анализов [26].
Если осуществляется гидролиз, то гидролизат обычно будет отсортирован по размеру, для того чтобы удалить короткие олигосахариды [27]. Этого можно достигнуть различными способами, такими как ультрафильтрация, за которой следует ионообменная хроматография. Олигосахариды со степенью полимеризации менее чем или равной около 6 предпочтительно удаляют для серологической группы A, а олигосахариды с СП менее чем около 4 предпочтительно удаляют для серологических групп W135 и Y.
Носители
Предпочтительно носителем является белок. Предпочтительными белками-носителями, к которым присоединяются сахаридные антигены в композициях данного изобретения, являются бактериальные токсины, такие как дифтерийный токсин или столбнячный токсин. Подходящие белки-носители включают CRM197 мутант дифтерийного токсина [28-30], дифтерийный токсин, белок внешней мембраны N. meningitidis [31], синтетические пептиды [32,33], белки теплового шока [34,35], белки коклюша [36,37], цитокины [38], лимфокины [38], гормоны [38], факторы роста [38], синтетические белки, содержащие множественные человеческие CD4+ T-клеточные эпитопы из различных антигенов, полученных из патогенных микроорганизмов [39], такие как белок N19 [40], белок D из H. influenzae [41,42], пневмококковый поверхностный белок PspA [43], пневмолизин [44], белки, связывающие железо [45], токсин A или B из C. difficile [46], человеческий сывороточный альбумин (предпочтительно рекомбинант) и т.д.
Присоединение сахаридного антигена к носителю осуществляется предпочтительно через -NH2 группу, например, в боковой цепи лизинового остатка в белке-носителе или аргининового остатка. Если сахарид имеет свободную альдегидную группу, то он может реагировать с амином в носителе, для того чтобы образовать конъюгат посредством восстановительного аминирования. Присоединение также может осуществляться через -SH группу, например, в боковой цепи цистеинового остатка.
Если композиция содержит более одного сахаридного антигена, то становится возможным применение более одного носителя, например, для того чтобы уменьшить риск подавления носителя. Таким образом, различные носители можно применять для различных сахаридных антигенов, например, сахариды Neisseria meningitidis серологической группы A можно было бы присоединить к CRM197, тогда как сахариды типа C можно было бы присоединить к столбнячному токсину. Также является возможным применение более одного носителя для конкретного сахаридного антигена. Сахариды могли бы существовать в двух группах, те, которые соединены с CRM197, и другие, соединенные со столбнячным токсином. Обычно, однако, предпочтительным является применение одного и того же носителя для всех сахаридов.
Один единственный белок-носитель мог бы нести более одного сахаридного антигена [47,48]. Например, один единственный белок-носитель мог бы иметь присоединенные к нему сахариды из различных патогенных микроорганизмов или из различных серологических групп одного и того же патогенного микроорганизма. Для достижения данной цели можно смешать различные сахариды перед реакцией конъюгации. Обычно, однако, предпочтительным является иметь отдельные конъюгаты для каждой серологической группы, причем различные сахариды смешивают после конъюгации. Отдельные конъюгаты могут располагаться на одном и том же носителе.
Предпочтительными являются конъюгаты с соотношением сахарид:белок (w/w) между 1:5 (т.е. избыток белка) и 5:1 (т.е. избыток сахарида). Предпочтительными являются соотношения между 1:2 и 5:1, такие как соотношения между 1:1,25 и 1:2,5.
Конъюгаты можно применять для соединения со свободным носителем [49]. Если данный белок-носитель присутствует как в свободной, так и в связанной формах в композиции данного изобретения, то предпочтительным является наличие несвязанной формы не более 5% от общего количества белка-носителя в композиции в целом, а более предпочтительно наличие менее 2% по массе.
Можно применять любую подходящую реакцию конъюгации с любым подходящим сшивающим агентом при необходимости.
Обычно сахарид нужно будет активировать или функционализировать перед конъюгацией. Активация может подразумевать, например, реагенты цианирования, такие как CDAP (например, l-циано-4-диметиламинопиридиниум тетрафторборат [50, 51 и т.д.]). Другие подходящие методики применяют карбодиимиды, гидразиды, активные эфиры, норборан, п-нитробензойную кислоту, N-гидроксисукцинимид, S-NHS, EDC, TSTU (смотри также введение к ссылке 52).
Связи через линкерную группу можно осуществлять, применяя любую известную методику, например, методики, описанные в ссылках 53 и 54. Один тип связи подразумевает восстановительное аминирование полисахарида, соединение конечной аминогруппы с одним концом линкерной группы адипиновой кислоты, а затем соединение белка с другим концом линкерной группы адипиновой кислоты [55, 56]. Другие линкеры включают B-пропионамидо [57], нитрофенилэтиламин [58], галоацилгалогениды [59], гликозидные связи [60], 6-аминокапроновая кислота [61], ADH [62], фрагменты от C4 до C12 [63] и т.д. В качестве альтернативы применению линкера можно использовать непосредственную связь. Непосредственные связи с белком могут образовываться окислением полисахарида, за которым следует восстановительное аминирование с белком, как описано, например, в ссылках 64 и 65.
Предпочтительным является процесс, подразумевающий введение аминогрупп в сахарид (например, посредством замещения терминальных =0 групп на -NH2), за которым следует получение производных с адипиновым диэфиром (например, N-гидроксисукцинимид диэфиром адипиновой кислоты) и реакция с белком-носителем.
После конъюгации можно разделить свободные и связанные сахариды. Существует множество подходящих способов, включая гидрофобную хроматографию, тангенциальную ультрафильтрацию, диафильтрацию и т.д. [смотри ссылки 66 & 67 и т.д.].
Поскольку композиция данного изобретения содержит деполимеризованный сахарид, то предпочтительным является, что деполимеризация предшествует конъюгации.
Получение подходящих сахаридных конъюгатов антигенов, подходящих для включения в композиции данного изобретения, описывается в ссылке 68.
Белковые антигены
Поскольку антиген, включенный в композиции данного изобретения, является белковым антигеном, он может быть выбран из:
- белкового антигена из N. meningitidis серологической группы B, такого как антигены в ссылках с 69 по 75. Применяя стандартную номенклатуру ссылки 73, NMB2132, NMB1870 и NMB0992 представляют собой три предпочтительных белка, которые можно применять в качестве основы подходящего антигена;
- белкового антигена из S. pneumoniae (например, из PhtA, PhtD, PhtB, PhtE, SpsA, LytB, LytC, LytA, Spl25, Sp101, Spl28, Spl30 и Spl33, как раскрыто в ссылке 76);
- антигена из вируса гепатита A, такого как инактивированный вирус [например, 77, 78; глава 15 ссылки 86];
- антигена из вируса гепатита B, таких как мембранного и/или ядерного антигенов [например, 78,79; глава 16 ссылки 86];
- антигена из вируса гепатита C [например, 80]. Антигены вируса гепатита C, которые можно применять, могут включать один или более из следующих: HCV E1 и/или E2 белки, E1/E2 гетеродимерные комплексы, капсидные белки и неструктурные белки или фрагменты данных антигенов, причем неструктурные белки могут необязательно быть модифицированы, для того чтобы исключить ферментативную активность, но сохранить иммуногенность (например, 81, 82 и 83);
- антигена из Bordetella pertussis, такого как голотоксин коклюша (ГК) и филаментный гемагглютинин (ФГА) из B. pertussis, необязательно также в комбинации с пертактином и/или агглютиногенами 2 и 3 [например, ссылки 84 & 85; глава 21 ссылки 86];
- дифтерийного антигена, такого как дифтерийный токсин [например, глава 13 ссылки 86];
- столбнячного антигена, такого как столбнячный токсин [например, глава 27 ссылки 86];
- антигена из N. gonorrhoeae [например, 69, 70, 71];
- антигена из Chlamydia pneumoniae [например, 87, 88, 89, 90, 91, 92, 93];
- антигена из Chlamydia trachomatis [например, 94];
- антигена из Porphyromonas gingivalis [например, 95];
- полиоантигена(ов) [например, 96, 97; глава 24 ссылки 86], такого как IPV;
- антигена(ов) бешенства [например, 98], такого как лиофилизованный инактивированный вирус [например, 99, RabAvert™];
- антигенов кори, паротита и/или коревой краснухи [например, главы 19, 20 и 26 ссылки 86];
- антигенов из Helicobacter pylori, таких как CagA [от 100 до 103], VacA [104, 105], NAP [106, 107, 108], HopX [например, 109], HopY [например, 109] и/или уреаза;
- антигена(ов) вируса гриппа [например, главы 17 & 18 ссылки 86], такого как гемагглютинин и/или поверхностные белки нейраминидазы;
- антигена из Moraxella catarrhalis [например, 110];
- белкового антигена из Streptococcus agalactiae (стрептококк группы B) [например, 111, 112];
- антигена из Streptococcus pyogenes (стрептококк группы A) [например, 112, 113, 114];
- антигена из Staphylococcus aureus [например, 115];
- антигена(ов) из парамиксовируса, такого как респираторно-синцитиальный вирус (RSV [116, 117]) и/или вирус парагриппа (PIV3 [118]);
- антигена из Bacillus anthracis [например, 119, 120, 121];
- антигена из вируса семейства флавивирусов (род флавивирус), такого как из вируса желтой лихорадки, вируса японского энцефалита, четырех серологических типов вируса лихорадки денге, вируса клещевого энцефалита, вируса лихорадки Западного Нила;
- антигена пестивируса, такого как из вируса классической свиной лихорадки, вируса диареи крупного рогатого скота и/или вируса пограничной болезни овец;
- антигена парвовируса, например, из парвовируса B19;
- антигена вируса простого герпеса (HSV). Предпочтительным антигеном HSV для применения в данном изобретении является мембранный гликопротеин gD. Предпочтительным является применение gD из HSV-2 штамма ('gD2' антиген). Композиция может применять форму gD, в которой C-терминальная мембранная «якорная» область была удалена [122], например, усеченный gD, содержащий аминокислоты 1-306 натурального белка с дополнением аспарагина и глутамина на C-конце. Подобная форма белка содержит сигнальный пептид, который расщепляется, давая зрелый белок из 283 аминокислот. Делеция «якоря» позволяет получить белок в растворимой форме;
- антигена человеческого папиломавируса (HPV). Предпочтительными антигенами HPV для применения в данном изобретении являются капсидные белки L1, которые могут собираться с образованием структур, известных как вирусоподобные частицы (VLP). VLP можно получить посредством рекомбинантной экспрессии L1 в дрожжевых клетках (например, в S. cerevisiae) или в клетках насекомых (например, в клетках Spodoptera, таких как S. frugiperda, или в клетках Drosophila). Для дрожжевых клеток плазмидные векторы могут нести ген(ы) L1; для клеток насекомых бакуловирусные векторы могут нести ген(ы) L1. Более предпочтительно данная композиция содержит L1 VLP как из HPV-16, так и из HPV-18 штаммов. Было показано, что данная бивалентная комбинация является высокоэффективной [123]. Кроме штаммов HPV-16 и HPV-18, также возможно включать L1 VLP из HPV-6 и HPV-11 штаммов. Также возможно применение онкогенных штаммов HPV. Вакцина может содержать между 20-60 мкг/мл (например, около 40 мкг/мл) L1 на HPV штамм.
Композиция может содержать один или более данных антигенов, которые могут быть при необходимости детоксифицированы (например, детоксификация коклюшного токсина посредством химических и/или генетических способов).
Поскольку в смесь включен дифтерийный антиген, то предпочтительным является также включение столбнячного антигена и коклюшного антигена. Аналогично, поскольку включен столбнячный антиген, то предпочтительным является также включение дифтерийного и коклюшного антигенов. Аналогично, поскольку включен коклюшный антиген, то предпочтительным является также включение дифтерийного и столбнячного антигенов.
Антигены в смеси будут обычно присутствовать в концентрации, по крайней мере, 1 мкг/мл каждый. Обычно концентрации любого данного антигена будет достаточно, для того чтобы вызвать иммунный ответ на данный антиген.
В качестве альтернативы применению белковых антигенов в смеси, можно применять нуклеиновую кислоту, кодирующую данный антиген. Таким образом, белковые компоненты смеси можно заменить нуклеиновой кислотой (предпочтительно ДНК, например, в виде плазмиды), которая кодирует белок. Аналогично композиции данного изобретения могут содержать белки, которые имитируют антигены, например мимеотопы [124] или антиидиотипические антитела.
В качестве альтернативы или кроме антигенов, перечисленных выше, композиция может содержать препарат везикулы внешней мембраны (ВВМ) из N. meningitidis серологической группы B, такой как препараты, раскрытые в ссылках 125, 126, 127, 128 и т.д.
Дополнительные композиции
Дополнительным объектом данного изобретения является получение вакцинных композиций, которые обеспечивают защиту от стрептококка группы B, N. meningitidis серологической группы B и/или вируса гриппа. Было обнаружено, что лиганды CD1d, как ни удивительно, являются эффективными вспомогательными веществами для антигенов из данных патогенных микроорганизмов.
Композиции, описанные ниже, включают, по крайней мере, один антиген из стрептококка группы B, N. meningitidis серологической группы B или вируса гриппа. Данные композиции могут содержать дополнительные антигены. Например, данные композиции могут также включать один или более сахаридных антигенов, связанных с одним или более носителями, такими как носители, описанные выше для включения в композиции для применения в получении долговременной иммунологической памяти. В качестве альтернативы, или кроме того, данные композиции могут содержать один или более белковых антигенов, описанных выше.
Стрептококк группы B
Изобретение, следовательно, обеспечивает композицию, содержащую: a) лиганд CD1d; и b) антиген из стрептококка группы B.
Примеры антигенов из стрептококка группы B (Streptococcus agalactiae) для включения в данную композицию находятся в ссылках 111 & 112. Таким образом, композиция может содержать белок, содержащий одну или более: (i) аминокислотных последовательностей S. agalactiae в ссылке 112 (с четными номерами SEQ ID ΝO: от 2 до 10960 ссылки 112); (ii) аминокислотную последовательность, имеющую, по крайней мере, 80% идентичность последовательности по отношению к аминокислотной последовательности S. agalactiae (i); аминокислотную последовательность, содержащую эпитоп из аминокислотной последовательности S. agalactiae (i). Предпочтительно композиция содержит один или более белков из от GBSl до GBS689, как описано в ссылке 112 (смотри Таблицу IV в контексте). Более предпочтительно композиция содержит белковый антиген GBS80.
Менингококк
Изобретение также обеспечивает композицию, содержащую: a) лиганд CD1d; и b) антиген из Neisseria meningitidis.
Антиген из N. meningitidis, включенный в данную композицию, может быть белковым антигеном или препаратом везикулы внешней мембраны (ВВМ). Примеры препаратов ВВМ, которые можно включать в данную композицию, включают препараты ВВМ из N. meningitidis серологической группы A, B, C, W135 или Y. Примеры белковых антигенов из N. meningitidis, которые можно включать в данную композицию, также предоставлены выше. Предпочтительно белковый антиген получают из N. meningitidis серологической группы B, и он при введении пациенту вызывает иммунный ответ, который дает перекрестную реакцию с клетками N. meningitidis серологической группы B. Предпочтительные белковые антигены, которые вызывают иммунный ответ, который дает перекрестную реакцию с клетками N. meningitidis серологической группы B, включают белковые антигены 'ΔG287nz-953', '936-741' и '961c' [129]. Предпочтительно композиция содержит более одного антигена из N. meningitidis. Предпочтительно композиция содержит все три белковых антигена 'ΔG287nz-953', '936-741' и '961c'. Другие применяемые белковые антигены базируются на ΝMB2132, NMB1870 и NMB0992.
Вирус гриппа
Изобретение также обеспечивает композицию, содержащую: a) лиганд CD1d; и b) антиген вируса гриппа.
Антиген вируса гриппа обычно получают из вирионов гриппа, но, в качестве альтернативы, антигены, такие как гемагглютинин, могут экспрессироваться в рекомбинантном носителе (например, в клеточной линии насекомого с применением бакуловирусного вектора) и применяться в очищенной форме [130,131]. Обычно, однако, антигены будут из вирионов.
Антиген может принимать форму живого вируса или более предпочтительно инактивированного вируса. Поскольку применяется инактивированный вирус, то вакцина может содержать целый вирион, расщепленный вирион или очищенные поверхностные антигены (включая гемагглютинин и обычно также включая нейраминидазу). Антигены вируса гриппа могут также быть представлены в виде виросом [132].
Вирус гриппа может быть ослабленным. Вирус гриппа может быть температурочувствительным. Вирус гриппа может быть холодовым. Данные три возможности применимы, в частности, для живых вирусов.
Штаммы вируса гриппа для применения в вакцине меняются от сезона к сезону. В текущий межпандемический период вакцины обычно включают два штамма вируса гриппа A (H1N1 и H3N2) и один штамм вируса гриппа B, а тривалентная вакцина является обычной. Данное изобретение также может применять вирусы пандемических штаммов (т.е. штаммов, к вакцине которых реципиент и основное население являются иммунологически неподготовленными), таких как штаммы подтипов H2, H5, H7 или H9 (в частности, вируса гриппа A), а противогриппозные вакцины для пандемических штаммов могут быть моновалентными или могут базироваться на нормальной тривалентной вакцине, дополненной пандемическим штаммом. Однако, в зависимости от сезона и от природы антигена, включенного в вакцину, данное изобретение может обеспечивать защиту от одного или более HA подтипов H1, H2, H3, H4, H5, H6, H7, H8, H9, H10, H11, H12, H13, H14, H15 или H16.
Другими штаммами, которые можно успешно включать в композиции, являются штаммы, которые резистентны к противовирусной терапии (например, резистентные к осельтамивиру [133] и/или занамивиру), включая резистентные пандемические штаммы [134].
Адъювантные композиции данного изобретения применяются, в частности, для проведения вакцинации против пандемических штаммов. Характеристиками штамма вируса гриппа, которые дают ему возможность вызывать пандемическую вспышку инфекции, являются: (a) он содержит новый гемагглютинин по сравнению с гемагглютининами в циркулирующих на данный момент человеческих штаммах, т.е. гемагглютинин, который не проявлялся у населения более десяти лет (например, H2) или вообще не встречался прежде у населения (например, H5, H6 или H9, которые обычно обнаруживали только в популяциях птиц), так, что население будет иммунологически не подготовлено к гемагглютинину данного штамма; (b) он способен передаваться горизонтально у населения; и (c) он является патогенным для людей. Вирус с гемагглютинином типа H5 является предпочтительным для проведения вакцинации против пандемического вируса гриппа, такого как штамм H5N1. Другие возможные штаммы включают H5N3, H9N2, H2N2, H7N1 и H7N7 и любые другие, являющиеся потенциально пандемическими штаммами.
Композиции данного изобретения могут включать антиген(ы) из одного или более (например, 1, 2, 3, 4 или более) штаммов вируса гриппа, включая вирус гриппа A и/или вирус гриппа B. Если вакцина включает более одного штамма вируса гриппа, то различные штаммы обычно культивируют отдельно и смешивают, после того как вирусы собраны из культуры и получены антигены. Таким образом, способ данного изобретения может включать стадию смешивания антигенов более чем из одного штамма вируса гриппа.
Вирус гриппа может быть реассортантным штаммом и может быть получен методиками «обратной генетики». Методики «обратной генетики» [например, 135-139] позволяют получить in vitro вирусы гриппа с необходимыми геномными сегментами, применяя плазмиды. Обычно они включают экспрессию (a) молекул ДНК, которые кодируют необходимые вирусные молекулы РНК, например, из промоторов polI, и (b) молекул ДНК, которые кодируют вирусные белки, например, из промоторов polII, таким образом, что экспрессия обоих типов ДНК в клетке приводит к сборке полного интактного инфекционного вириона. ДНК предпочтительно обеспечивает все вирусные РНК и белки, а также возможным является применение вируса-помощника для получения некоторых РНК и белков. Предпочтительными являются способы на основе плазмидных векторов, применяющие отдельные плазмиды для получения каждой вирусной РНК [140-142], и данные способы будут также включать применение плазмид для экспрессии всех или некоторых (например, только PB1, PB2, PA и NP белки) вирусных белков, причем в некоторых способах применяются вплоть до 12 плазмид. Для того чтобы уменьшить число необходимых плазмид, новый подход [143] объединяет множество транскрипционных кассет РНК-полимеразы I (для синтеза вирусной РНК) на одной и той же плазмиде (например, последовательности, кодирующие 1, 2, 3, 4, 5, 6, 7 или все 8 сегментов вируса гриппа A вРНК) и множество областей, кодирующих белок, с промоторами РНК-полимеразы II на другой плазмиде (например, последовательности, кодирующие 1, 2, 3, 4, 5, 6, 7 или все 8 транскриптов вируса гриппа A мРНК). Предпочтительные аспекты способа ссылки 143 включают: (a) PB1, PB2 и PA мРНК-кодирующие области на единой плазмиде; и (b) все 8 вРНК-кодирующих сегментов на единой плазмиде. Возможным является применение двойных промоторов polI и polII для одновременного кодирования вирусных РНК и для мРНК, экспрессируемых из одной матрицы [144,145].
Таким образом, вирус может включать один или более сегментов РНК из A/PR/8/34 вируса (обычно 6 сегментов из A/PR/8/34, причем сегменты HA и N из штамма вакцины, т.е. 6:2 реассортант), в частности, если вирусы культивируют в яйцеклетках. Он также может включать один или более РНК сегментов из A/WSN/33 вируса или из любого другого вирусного штамма, применимого для получения реассортантных вирусов для вакцинного препарата. Обычно данное изобретение обеспечивает защиту от штамма, который способен передаваться от человека к человеку, и, таким образом, геном штамма будет обычно включать, по крайней мере, один сегмент РНК, который происходит из вируса гриппа млекопитающего (например, человека).
Вирусы, применяемые в качестве источника антигенов, можно культивировать как в яйцеклетках, так и в клеточной культуре. Современный стандартный способ культивирования вируса гриппа применяет куриные яйца с развивающимся эмбрионом, причем вирус очищают от содержимого яйца (аллантоисной жидкости). Совсем недавно, однако, вирусы культивировали в клеточной культуре животного, и ввиду скорости и аллергий пациентов данный способ культивирования является предпочтительным.
Клеточным субстратом будет обычно клеточная линия млекопитающего, такая как MDCK; CHO; 293T; BHK; Vero; MRC-5; PER.C6; WI-38 и т.д. Предпочтительные клеточные линии млекопитающего для культивирования вирусов гриппа включают: клетки MDCK [146-149], полученные из собачьей почки Madin Darby; клетки Vero [150-152], полученные из почки африканской зеленой мартышки (Cercopithecus aethiops); или клетки PER.C6 [153], полученные из человеческих эмбриональных ретинобластов. Данные клеточные линии широко доступны, например, из американской коллекции типовых культур (АКТК) [154], из клеточного депозитария Coriell [155] или из европейской коллекции культивируемых клеток животных (ЕККЖ). Например, АКТК предоставляет многообразие различных клеток Vero под каталожными номерами CCL-81, CCL-81.2, CRL-1586 и CRL-1587, и она предоставляет клетки MDCK под каталожным номером CCL-34. PER.C6 доступна из ЕККЖ под депозитным номером 96022940. В качестве менее предпочтительной альтернативы клеточным линиям млекопитающего, вирус можно культивировать на птичьих клеточных линиях [например, ссылки 156-158], включая клеточные линии, полученные из уток (например, утиной сетчатки глаза) или куриц, например фибробластов эмбриона цыпленка (ФЭЦ), и т.д.
Если вирус культивировали на клеточной линии млекопитающего, то композиция преимущественно не будет содержать яичных белков (например, овальбумина и овомукоида) и ДНК цыпленка, таким образом, уменьшая аллергенность.
Для культивирования на клеточной линии, такой как клетки MDCK, вирус можно культивировать на клетках в суспензии [146] или в вязкой культуре. Одной подходящей MDCK клеточной линией для суспензионной культуры является MDCK 33016 (депонированная как DSM ACC 2219). В качестве альтернативы можно применять культуру на микроносителях.
Если вирус культивировали на клеточной линии, то культура для культивирования предпочтительно не будет содержать (т.е. будет исследована и даст отрицательный результат на загрязнение) вирус простого герпеса, респираторно-синцитиальный вирус, вирус парагриппа 3, ОРВИ коронавирус, аденовирус, риновирус, реовирусы, полиомавирусы, бирнавирусы, цирковирусы и/или парвовирусы. В частности, предпочтительным является отсутствие вируса простого герпеса.
Если вирус культивировали на клеточной линии, то композиция предпочтительно содержит менее 10 нг (предпочтительно менее 1 нг, и более предпочтительно менее 100 пг) остаточной ДНК клетки-носителя на дозу, хотя следовые количества ДНК клетки-носителя могут присутствовать. Обычно ДНК клетки-носителя, которую необходимо исключить из композиций данного изобретения, является ДНК, которая длинней 100 пар оснований.
Измерение остаточной ДНК клетки-носителя является в настоящее время стандартным обязательным требованием для биологических препаратов и лежит в рамках стандартных возможностей специалиста. Анализ, применяемый для измерения ДНК, обычно будет ратифицированным анализом [159,160]. Характеристики выполнения ратифицированного анализа можно описать в математических и количественных выражениях, и его возможные источники ошибки будут установлены. Анализ будет главным образом проверен на характеристики, такие как точность, воспроизводимость, специфичность. Как только анализ откалиброван (например, по известным стандартным количествам ДНК клетки-носителя) и проверен, тогда можно в плановом порядке представить количественные измерения ДНК. Можно применять три принципиальных способа количественного выражения ДНК: способы гибридизации, такие как Саузерн-блоттинг или слот-блоттинг [161]; способы иммуноанализа, такие как система Threshold™ [162]; и количественная ПЦР [163]. Все данные способы хорошо знакомы специалисту, хотя точные характеристики каждого способа могут зависеть от клетки-носителя, о которой идет речь, например выбор проб для гибридизации, выбор праймеров и/или проб для амплификации и т.д. Система Threshold™ для молекулярных устройств является количественным анализом пикограммовых уровней общего количества ДНК и применялась для контроля уровней загрязняющей ДНК в биофармацевтических лекарствах [162]. Обычный анализ подразумевает не сайт-специфическое образование реакционного комплекса между биотинилированным связывающим белком однонитевой ДНК, связанным с уреазой антителом неоднонитевой ДНК и ДНК. Все компоненты анализа включены в полный набор для анализа общего количества ДНК, поставляемый производителем. Различные коммерческие производители предлагают количественные ПЦР-анализы для определения остаточной ДНК клетки-носителя, например AppTec™ Laboratory Services, BioReliance™, Althea Technologies и т.д. Сравнение хемилюминесцентного гибридизационного анализа и системы общего количества ДНК Threshold™ для измерения загрязнения человеческой вирусной вакцины ДНК клетки-носителя можно найти в ссылке 164.
Загрязняющую ДНК можно удалить в процессе получения вакцины, применяя стандартные процедуры очистки, например хроматографию и т.д. Удаление остаточной ДНК клетки-носителя можно увеличить посредством обработки нуклеазой, например применением ДНКазы (дезоксирибонуклеазы). Общеизвестным способом уменьшения загрязнения ДНК клетки-носителя является раскрытый в ссылках 165 & 166, подразумевающий двухстадийную обработку, сначала применяя ДНКазу (например, Benzonase), а затем катионный детергент (например, CTAB).
Вакцины, содержащие <10 нг (например, <1 нг, <100 пг) ДНК клетки-носителя на 15 мкг гемагглютинина, являются предпочтительными, так как они являются вакцинами, содержащими <10 нг (например, <1 нг, <100 пг) ДНК клетки-носителя на 0,25 мл объема. Вакцины, содержащие <10 нг (например, <1 нг, <100 пг) ДНК клетки-носителя на 50 мкг гемагглютинина, являются более предпочтительными, так как они являются вакцинами, содержащими <10 нг (например, <1 нг, <100 пг) ДНК клетки-носителя на 0,5 мл объема.
Клеточные линии, поддерживающие репликацию вируса гриппа, предпочтительно культивируются в бессывороточной культуральной среде и/или безбелковой среде. Предпочтительной для нас является среда, бессывороточная среда в контексте настоящего изобретения, в которой нет примесей сыворотки человеческого или животного происхождения. Подразумевается, что «безбелковая» означает культуры, в которых размножение клеток происходит при исключении белков, факторов роста, других белковых примесей и несывороточных белков, но может необязательно включать белки, такие как трипсин или другие протеазы, которые могут быть необходимыми для вирусного культивирования. Сами клетки, выращиваемые в подобных культурах, естественно, содержат белки.
Клеточные линии, поддерживающие репликацию вируса гриппа, предпочтительно культивируются при температуре ниже 37°C [167], например 30-36°C.
Гемагглютинин (ГА) является основным иммуногеном в инактивированных вакцинах вируса гриппа, а дозы вакцины стандартизуются, исходя из уровней ГА, которые обычно измеряют посредством анализа простой радиальной иммунодиффузии (ПРИД). Вакцины обычно содержат около 15 мкг ГА на штамм, хотя также применяются и более низкие дозы, например, для детей или в пандемических ситуациях. Применяли фракционированные дозы, такие как 1/2 (т.е. 7,5 мкг ГА на штамм), 1/4 и 1/8 [168,169], так как они имеют более высокие дозы (например, 3x или 9x дозы [170,171]). Таким образом, вакцины могут содержать между 0,1 и 150 мкг ГА на штамм вируса гриппа, предпочтительно между 0,1 и 50 мкг, например 0,1-20 мкг, 0,1-15 мкг, 0,1-l0 мкг, 0,1-7,5 мкг, 0,5-5 мкг и т.д. Конкретные дозы содержат, например, около 15, около 10, около 7,5, около 5, около 3,8, около 1,9, около 1,5 и т.д. на штамм. Таким образом, вакцины могут содержать между 0,1 и 20 мкг ГА на штамм вируса гриппа, предпочтительно между 0,1 и 15 мкг, например 0,1-10 мкг, 0,1-7,5 мкг, 0,5-5 мкг и т.д. Конкретные дозы содержат, например, около 15, около 10, около 7,5, около 5, около 3,8, около 1,9 и т.д. Данные более низкие дозы наиболее применимы при наличии вспомогательного вещества в вакцине, как и в случае данного изобретения.
ГА, применяемый в данном изобретении, может быть природным ГА, как полученный из вируса, или может быть модифицированным. Например, известно, что ГА модифицируют, для того чтобы удалить детерминанты (например, участки повышенной основности), которые служат причиной того, что вирус является высокопатогенным в птичьих видах, так как данные детерминанты могут в противном случае предотвращать развитие вируса в яйцах.
Инактивированная, но нецельноклеточная вакцина (например, вакцина расщепленного вируса или вакцина очищенного поверхностного антигена) может содержать матричный белок, для того чтобы пользоваться дополнительными T-клеточными эпитопами, которые находятся в данном антигене. Таким образом, нецельноклеточная вакцина (в частности, субъединичная вакцина), которая содержит гемагглютинин и нейраминидазу, может дополнительно содержать Ml и/или M2 матричный белок. Если присутствует матричный белок, то предпочтительным является включение определимых уровней матричного белка M2. Также может присутствовать нуклеопротеин.
Технология приготовления фармацевтических композиций
Антигены и лиганды CD1d, описанные выше, подходят, в частности, для включения в иммуногенные композиции и вакцины. Способ изобретения может, следовательно, включать стадию приготовления антигена и лиганда CD1d в виде иммуногенной композиции и вакцины. Изобретение обеспечивает композицию или вакцину, получаемую данным способом.
Иммуногенные композиции и вакцины данного изобретения будут содержать обычно, кроме антигена(ов) и лигандов CD1d, 'фармацевтически приемлемые носители', которые включают любой носитель, который сам по себе не вызывает выработку антител, оказывающих превратное влияние на индивида, получающего композицию. Подходящими носителями обычно являются большие, медленно метаболизирующие макромолекулы, такие как белки, полисахариды, полимолочные кислоты, полигликолиевые кислоты, полимерные аминокислоты, аминокислотные сополимеры, трегалоза [172], липидные агрегаты (такие как масляные капельки или липосомы) и неактивные вирусные частицы. Подобные носители хорошо известны специалистам в данной области техники. Вакцины могут также содержать разбавители, такие как вода, соляной раствор, глицерин и т.д. Кроме того, могут присутствовать вспомогательные вещества, такие как увлажняющее средство или эмульгатор, pH буферные вещества и тому подобное. Детальное обсуждение фармацевтически приемлемых вспомогательных веществ представлено в ссылке 173.
Иммуногенные композиции, применяемые в качестве вакцин, содержат иммунологически эффективное количество антигена, а также любой другой из указанных компонентов при необходимости. Под 'иммунологически эффективным количеством' подразумевается, что введение данного количества индивиду либо в однократной дозе, либо в виде части серии является эффективным для лечения или профилактики. Данное количество варьируется в зависимости от здоровья и физического состояния индивида, которого необходимо лечить, возраста, таксономической группы индивида, которого необходимо лечить (например, нечеловеческого примата, примата и т.д.), способности иммунной системы индивида синтезировать антитела, степени необходимой защиты, технологии приготовления вакцины, лечебной оценки врачом медицинской обстановки и других значимых факторов. Ожидается, что количество будет попадать в относительно широкий диапазон, который можно определить посредством стандартных исследований.
Вакцину можно вводить совместно с другими иммунорегуляторными агентами. Лиганд CD1d действует в качестве вспомогательного вещества в рамках иммуногенных композиций данного изобретения. Вакцина может включать дополнительные вспомогательные вещества. Подобные вспомогательные вещества включают, но не ограничиваются следующим.
Вспомогательные вещества, которые можно применять в данном изобретении включают, но не ограничиваются:
• Минерал-содержащую композицию, включающую кальциевые соли и алюминиевые соли (или их смеси). Кальциевые соли включают фосфат кальция (например, "CAP" частицы, раскрытые в ссылке 174). Алюминиевые соли включают гидроксиды, фосфаты, сульфаты и т.д., причем соли находятся в любой подходящей форме (например, гель, кристаллическая форма, аморфная форма и т.д.). Предпочтительной является адсорбция данных солей. Минерал-содержащие композиции также можно приготовить в виде частицы соли металла [175]. Вспомогательные вещества соли алюминия описываются более подробно ниже.
• Эмульсию типа масло в воде (прямую эмульсию), более подробно описанную ниже.
• Иммуностимулирующий олигонуклеотид, такой как олигонуклеотид, содержащий CpG мотив (динуклеотидная последовательность, содержащая неметилированный цитозин, связанный фосфатной связью с гуанозином), TpG мотив [176], двухцепочечную РНК, олигонуклеотид, содержащий палиндромную последовательность, или олигонуклеотид, содержащий поли(dG) последовательность. Иммуностимулирующие олигонуклеотиды могут включать нуклеотидные модификации/аналоги, такие как фосфоротиоатные модификации, и могут быть двухцепочечными или (за исключением РНК) одноцепочечными. Ссылки с 177 по 179 раскрывают замещения аналогов, например, замещение гуанозина 2'-деокси-7-деазагаунозином. Эффект олигонуклеотидов CpG в качестве вспомогательного вещества дополнительно обсуждается в ссылках 180-185. Последовательность CpG может быть направлена на TLR9, такой как мотив GTCGTT или TTCGTT [186]. Последовательность CpG может быть специфичной для вызывания Th1 иммунного ответа, такая как CpG-A ODN (олигодеоксинуклеотид), или она может быть более специфичной для вызывания B-клеточного иммунного ответа, такая как CpG-B ODN. CpG-A и CpG-B ODN обсуждаются в ссылках 187-189. Предпочтительно CpG представляет собой CpG-A ODN. Предпочтительно CpG олигонуклеотид построен таким образом, что 5'-конец является доступным для рецепторного распознавания. Необязательно две олигонуклеотидные CpG последовательности могут быть соединены на их 3'-концах, для того чтобы образовывать "иммуномеры". Смотри, например, ссылки 190-192. Применимым CpG вспомогательным веществом является CρG7909, также известный как ProMune™ (Coley Pharmaceutical Group, Inc.). Иммуностимулирующие олигонуклеотиды будут обычно содержать, по крайней мере, 20 нуклеотидов. Они могут содержать менее 100 нуклеотидов.
• 3-O-деацилированный монофосфорил-липид A ('3dMPL', также известный как 'MPL™') [193-196]. 3dMPL был получен из безгептозного мутанта Salmonella minnesota и является химически подобным липиду A, но не содержит кислотолабильную фосфорильную группу и лабильную к основаниям ацильную группу. Получение 3dMPL было первоначально описано в ссылке 197. 3dMPL может находиться в форме смеси родственных молекул, отличающихся их ацилированием (например, имеющих 3, 4, 5 или 6 ацильных цепей, которые могут быть различной длины). Два глюкозаминовых (также известных как 2-деокси-2-аминоглюкоза) моносахарида представляют собой N-ацилированные по его атомам углерода в 2-положении (т.е. по положениям 2 и 2'), а также существует O-ацилирование в положении 3'.
• Соединение имидазохинолина, такое как Imiquimod ("R-837") [198,199], Resiquimod ("R-848") [200] и их аналоги; и их соли (например, соли гидрохлориды). Дополнительные детали об иммуностимулирующих имидазохинолинах можно найти в ссылках с 201 по 205.
• Соединение тиосемикарбазона, такое как соединения, раскрытые в ссылке 206. Способы приготовления, производства и скрининга активных соединений также описываются в ссылке 206. Тиосемикарбазоны, в частности, являются эффективными в стимуляции человеческих мононуклеаров периферической крови для выработки цитокинов, таких как TNF-α.
• Соединение триптантрина, такое как соединения, раскрытые в ссылке 207. Способы приготовления, производства и скрининга активных соединений также описываются в ссылке 207. Триптантрины, в частности, являются эффективными в стимуляции человеческих мононуклеаров периферической крови для выработки цитокинов, таких как TNF-α.
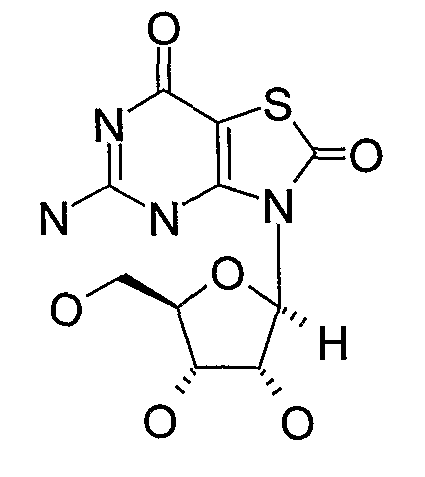
• Нуклеозидный аналог, такой как: (a) Isatorabine (ANA-245; 7-тиа-8-оксогуанозин):
и его пролекарства; (b) ANA975; (c) ANA-025-1; (d) ANA380; (e) соединения, раскрытые в ссылках с 208 по 210; (f) соединение, имеющее формулу:
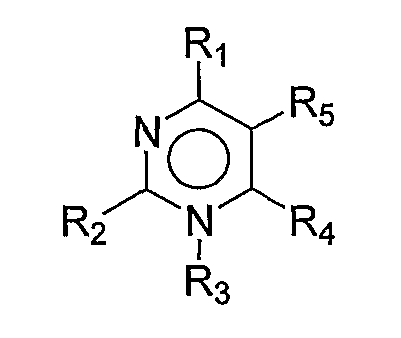 ,
,
где:
R1 и R2 каждый независимо представляют собой H, галоген, -NRaRb, -OH, C1-6-алкокси, замещенную C1-6-алкокси, гетероциклил, замещенный гетероциклил, C6-10-арил, замещенный C6-10-арил, C1-6-алкил или замещенный C1-6-алкил;
R3 отсутствует, представляет собой H, C1-6-алкил, замещенный C1-6-алкил, C6-10-арил, замещенный C6-10-арил, гетероциклил или замещенный гетероциклил;
R4 и R5 каждый независимо представляют собой H, галоген, гетероциклил, замещенный гетероциклил, -C(O)-Rd, C1-6-алкил, замещенный C1-6-алкил или связаны вместе с образованием 5-членного кольца, как в R4-5:
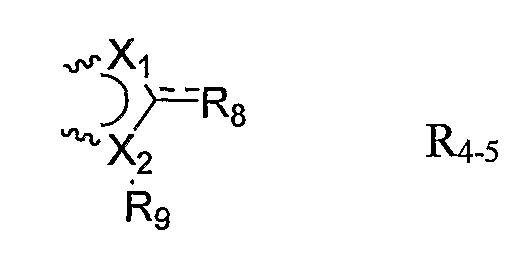 ,
,
причем связывание достигается связями, обозначенными ~~~~;
X1 и X2 каждый независимо представляют собой N, C, O или S;
R8 представляет собой H, галоген, -OH, C1-6-алкил, C2-6-алкенил, C2-6-алкинил, -OH, -NRaRb, -(CH2)n-0-Rc, -O-(C1-6-алкил), -S(O)pRe или -C(O)-Rd;
R9 представляют собой H, C1-6-алкил, замещенный C1-6-алкил, гетероциклил, замещенный гетероциклил или R9a, причем R9a представляет собой:
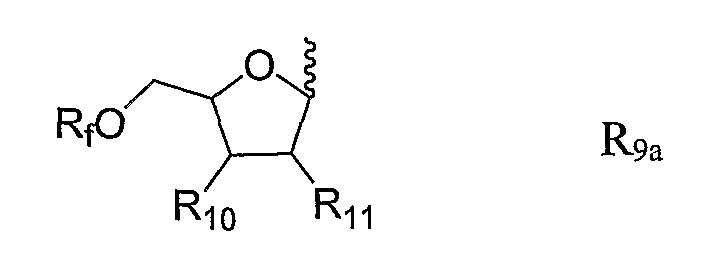 ,
,
причем связывание достигается связями, обозначенными ~~~~;
R10 и R11 каждый независимо представляют собой H, галоген, C1-6-алкокси, замещенную C1-6-алкокси, -NRaRb или -OH;
каждый Ra и Rb независимо представляет собой H, C1-6-алкил, замещенный C1-6-алкил, -C(O)Rd, C6-10-арил;
каждый Rc независимо представляет собой H, фосфат, дифосфат, трифосфат, C1-6-алкил или замещенный C1-6-алкил;
каждый Rd независимо представляет собой H, галоген, C1-6-алкил, замещенный C1-6-алкил, C1-6-алкокси, замещенную C1-6-алкокси, -NH2, -NH(C1-6-алкил), -NH(замещенный C1-6-алкил), -N(C1-6-алкил)2, -N(замещенный C1-6-алкил)2, C6-10-арил или гетероциклил;
каждый Re независимо представляет собой H, C1-6-алкил, замещенный C1-6-алкил, C6-10-арил, замещенный C6-10-арил, гетероциклил или замещенный гетероциклил;
каждый Rf независимо представляют собой H, C1-6-алкил, замещенный C1-6-алкил, -C(O)Rd, фосфат, дифосфат или трифосфат;
каждое n независимо представляет собой 0, 1, 2 или 3;
каждое p независимо представляет собой 0, 1 или 2; или
(g) фармацевтически приемлемая соль любого соединения от (a) до (f), таутомер любого соединения от (a) до (f) или фармацевтически приемлемая соль таутомера.
• Локсорибин (7-аллил-8-оксогуанозин) [211].
• Соединения, раскрытые в ссылке 212, включающие: соединения ацилпиперазина, индолдионовые соедиения, тетрагидроизохинолиновые (ТГИХ) соединения, бензоциклодионовые соединения, соединения аминоазавинила, аминобензимидазол хинолиновые (АБИХ) соединения [213,214], соединения гидрапталамида, соединения бензофенона, соединения изоксазола, стериновые соединения, соединения хиназилинона, соединения пиррола [215], антрахиноновые соединения, хиноксалиновые соединения, триазиновые соединения, соединения пиразолопиримидина и соединения бензазола [216].
• Соединения, раскрытые в ссылке 217, включающие 3,4-ди(lH-индол-3-ил)-lH-пиррол-2,5-дионы, аналоги стауроспорина, дериватизованные пиридазины, хромен-4-оны, индолиноны, хиназолины и нуклеозидные аналоги.
• Производное аминоалкилглюкозаминид фосфата, такое как RC-529 [218,219].
• Фосфазен, такой как поли[ди(карбоксилатофенокси)фосфазен] ("PCPP"), описанный, например, в ссылках 220 и 221.
• Низкомолекулярные иммунопотенциаторы (SMIPs), такие как:
N2-метил-1-(2-метилпропил)-1H-имидазо[4,5-c]хинолин-2,4-диамин
N2,N2-диметил-1-(2-метилпропил)-1H-имидазо[4,5-c]хинолин-2,4-диамин
N2-этил-N2-метил-l-(2-метилпропил)-1H-имидазо[4,5-c]хинолин-2,4-диамин
N2-метил-1-(2-метилпропил)-N2-пропил-1H-имидазо[4,5-c]хинолин-2,4-диамин
1-(2-метилпропил)-N2-пропил-1H-имидазо[4,5-c]хинолин-2,4-диамин
N2-бутил-1-(2-метилпропил)-1H-имидазо[4,5-c]хинолин-2,4-диамин
N2-бутил-N2-метил-1-(2-метилпропил)-1H-имидазо[4,5-c]хинолин-2,4-диамин
N2-метил-1-(2-метилпропил)-N2-пентил-1H-имидазо[4,5-c]хинолин-2,4-диамин
N2-метил-1-(2-метилпропил)-N2-проп-2-енил-1H-имидазо[4,5-c]хинолин-2,4-диамин
1-(2-метилпропил)-2-[(фенилметил)тио]-1H-имидазо[4,5-c]хинолин-4-амин
1-(2-метилпропил)-2-(пропилтио)-1H-имидазо[4,5-c]хинолин-4-амин
2-[[4-амино-1-(2-метилпропил)-1H-имидазо[4,5-c]хинолин-2-ил](метил)амино]этанол
2-[[4-амино-1-(2-метилпропил)-1H-имидазо[4,5-c]хинолин-2-ил](метил)амино]этилацетат
4-амино-1-(2-метилпропил)-1,3-дигидро-2H-имидазо[4,5-c]хинолин-2-он
N2-бутил-1-(2-метилпропил)-N4,N4-бис(фенилметил)-1H-имидазо[4,5-c]хинолин-2,4-диамин
N2-бутил-N2-метил-1-(2-метилпропил)-N4,N4-бис(фенилметил)-1H-имидазо[4,5-c]хинолин-2,4-диамин
N2-метил-1-(2-метилпропил)-N4,N4-бис(фенилметил)-1H-имидазо[4,5-c]хинолин-2,4-диамин
N2,N2-диметил-1-(2-метилпропил)-N4,N4-бис(фенилметил)-1H-имидазо[4,5-c]хинолин-2,4-диамин
1-{4-амино-2-[метил(пропил)амино]-1H-имидазо[4,5-c]хинолин-1-ил}-2-метилпропан-2-ол
1-[4-амино-2-(пропиламино)-1H-имидазо[4,5-c]хинолин-1-ил]-2-метилпропан-2-ол
N4,N4-дибензил-1-(2-метокси-2-метилпропил)-N2-пропил-1H-имидазо[4,5-c]хинолин-2,4-диамин.
• Сапонины [глава 22 ссылки 249], которые представляют собой гетерологическую группу стериновых гликозидов и тритерпеноидных гликозидов, которые находятся в коре, листьях, стеблях, корнях и даже цветах широкого диапазона видов растений. Сапонин из коры дерева Quillaia saponaria Molina был широко изучен в качестве вспомогательного вещества. Сапонин также можно получить в промышленности из Smilax ornata (сарсапарилла), Gypsophilla paniculata (гипсофила ползучая) и Saponaria officianalis (мыльный корень). Составы сапонинового вспомогательного вещества включают очищенные составы, такие как QS21, а также липидные составы, такие как ISCOM. QS21 продается под названием Stimulon™. Сапониновые композиции очищали с применением ВЭЖХ и обращенно-фазовой ВЭЖХ. Посредством применения данных способов были идентифицированы специфические очищенные фракции, включая QS7, QS17, QS18, QS21, QH-A, QH-B и QH-C. Предпочтительно сапонин представляет собой QS21. Способ получения QS21 раскрыт в ссылке 222. Сапониновые составы могут также содержать стерин, такой как холестерин [223]. Комбинации сапонинов и холестеринов можно применять для образования особых частиц, называемых иммуностимулирующими комплексами (ISCOM) [глава 23 ссылки 249]. ISCOM также обычно содержат фосфолипид, такой как фосфатидилэтаноламин или фосфатидилхолин. Любой известный сапонин можно применять в ISCOM. Предпочтительно ISCOM содержит один или более QuilA, QHA & QHC. ISCOM, кроме того, описываются в ссылках 223-225. Необязательно ISCOM могут быть лишены дополнительного детергента [226]. Обзор разработки вспомогательных веществ, основанных на сапонине, можно найти в ссылках 227 & 228.
• Бактериальные АДФ-рибозил токсины (например, E. coli термолабильный энтеротоксин "LT", холерный энтеротоксин "CT" или коклюшный токсин "PT") и их детоксифицированные производные, такие как мутантные токсины, известные как LT-K63 и LT-R72 [229]. Применение детоксифицированных АДФ-рибозил токсинов в качестве слизистых вспомогательных веществ описывается в ссылке 230, а в качестве парэнтеральных вспомогательных веществ - в ссылке 231.
• Биоадгезивы и мукоадгезивы, такие как микросферы этерифицированной гиалуроновой кислоты [232] или хитозан и его производные [233].
• Микрочастицы (т.е. частица диаметром от ~400 нм до ~150 мкм, более предпочтительно диаметром от ~200 нм до ~30 мкм или диаметром от ~500 нм до ~10 мкм), образованные из веществ, которые являются биоразлагаемыми и нетоксичными (например, поли(α-гидроксикислота), полигидроксимасляная кислота, полиортоэфир, полиангидрид, поликапролактон и т.д.), причем предпочтительным является сополимер лактида с гликолидом, необязательно обработанный, таким образом, чтобы иметь отрицательно заряженную поверхность (например, SDS) или положительно заряженную поверхность (например, катионным детергентом, таким как CTAB).
• Липосомы (главы 13 & 14 ссылки 249). Примеры липосомных составов, подходящих для применения в качестве вспомогательных веществ, описываются в ссылках 234-236.
• Простые эфиры полиоксиэтилена и сложные полиоксиэтиленовые эфиры [237]. Подобные составы, кроме того, содержат сурфактанты полиоксиэтиленсорбитанового сложного эфира в комбинации с октоксинолом [238], а также сурфактанты полиэтиленалкильных простых или сложных эфиров в комбинации, по крайней мере, с одним дополнительным неионным сурфактантом, таким как октоксинол [239]. Предпочтительные простые эфиры полиоксиэтилена выбираются из следующей группы: полиоксиэтилен-9-лауриловый эфир (лаурет 9), полиоксиэтилен-9-стеариловый эфир, полиоксиэтилен-8-стеариловый эфир, полиоксиэтилен-4-лауриловый эфир, полиоксиэтилен-35-лауриловый эфир и полиоксиэтилен-23-лауриловый эфир.
• Мурамиловые пептиды, такие как N-ацетилмурамил-L-треонил-D-изоглутамин ("thr-MDP"), N-ацетил-нормурамил-L-аланил-D-изоглутамин (nor-MDP), N-ацетилглюкозаминил-N-ацетилмурамил-L-Al-D-изо-Glu-L-Ala-дипальмитоксипропиламид ("DTP-DPP" или "Theramide™), N-ацетилмурамил-L-аланил-D-изоглутаминил-L-аланин-2-(1'-2'дипальмитоил-sn-глицеро-3-гидроксифосфорилокси)-этиламин ("MTP-PE").
• Препарат протеосомы наружного белка мембраны, приготовленный из первой грамотрицательной бактерии, в комбинации с препаратом липосахарида (LPS), полученном из второй грамотрицательной бактерии, причем протеосома наружного белка мембраны и препараты LPS образуют устойчивый нековалентный комплекс вспомогательного вещества. Подобные комплексы включают "IVX-908", комплекс, содержащий внешнюю мембрану Neisseria meningitidis и LPS.
• Метилинозин 5'-монофосфат ("MIMP") [240].
• Соединение полигидроксилированного пирролизидина [241], такое как соединение, имеющее формулу:
 ,
,
где R выбирается из группы, содержащей водород, линейные или разветвленные, незамещенные или замещенные, насыщенные или ненасыщенные ацильную, алкильную (например, циклоалкил), алкенильную, алкинильную и арильную группы или их фармацевтически приемлемую соль или производное. Примеры включают, но не ограничиваются: казуарин, казуарин-6-α-D-глюкопиранозу, 3-эпи-казуарин, 7-эпи-казуарин, 3,7-диэпи-казуарин и т.д.
• Гамма-инулин [242] или его производные, такие как альгаммулин.
• Соединение формулы I, II или III или его соль:
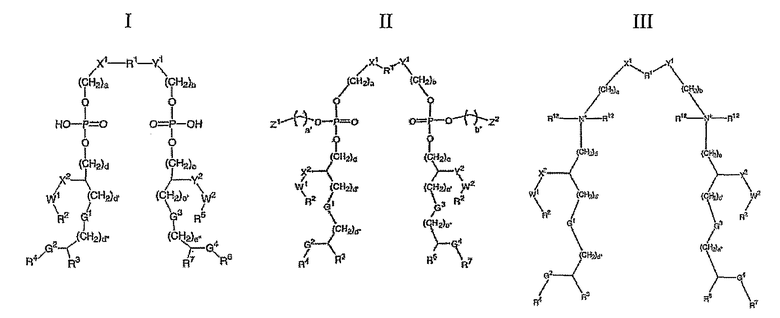 ,
,
определенное в ссылке 243, такое как 'ER 803058', 'ER 803732', 'ER 804053', 'ER 804058', 'ΕR 804059', 'ER 804442', 'ER 804680', 'ΕR 804764', 'ER 803022' или 'ER 804057', например:
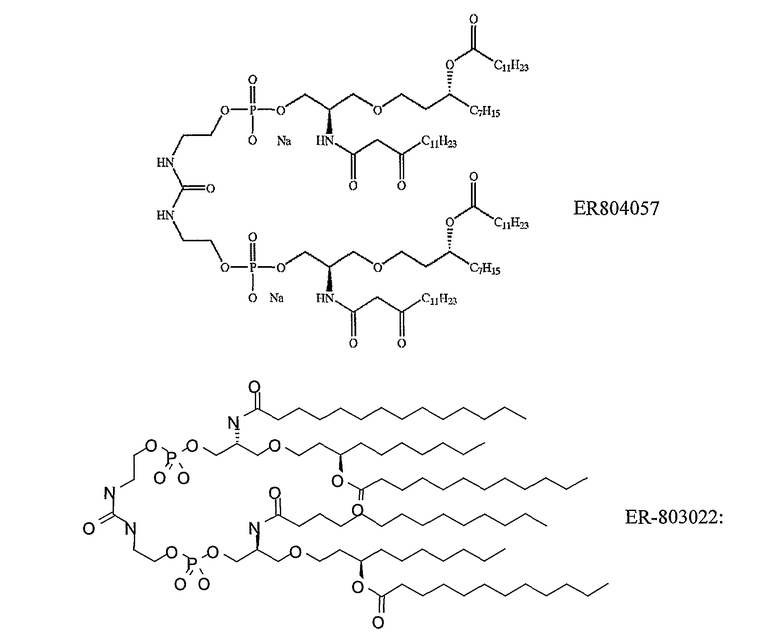
• Производные липида A из Escherichia coli, такие как OM-174 (описанные в ссылках 244 & 245).
• Состав катионного липида и (обычно нейтрального) солипида, такого как аминоприл-диметил-миристолеилокси-пропанаминиум бромид-дифитаноилфосфатидилэтаноламин ("Vaxfectin™") или аминоприл-диметил-бис-додецилокси-пропанаминиум бромид-диолеоилфосфатидилэтаноламин ("GAP-DLRIE:DOPE"). Предпочтительными являются составы, содержащие соли (±)-N-(3-аминоприл)-N,N-диметил-2,3-бис(син-9-тетрадеценилокси)-1-пропанаминиума [246].
• Соединения, содержащие липиды, связанные с фосфат-содержащим ациклическим остовом, такие как TLR4 антагонист E5564 [247,248]:

Данные и другие, активные в качестве вспомогательных вещества обсуждаются более подробно в ссылках 249 & 250.
Медицинские способы и применения
После приготовления композиции данного изобретения можно вводить непосредственно пациенту. Пациентами, которых нужно лечить, могут быть животные; в частности, можно лечить человеческих пациентов. Вакцины, в частности, применимы для вакцинации детей и подростков. Было показано, что вакцины являются эффективными в MHC II-/- животных моделях, а следовательно, полагают, что они будут применимы для лечения пациентов с иммунной недостаточностью. Они могут доставляться системным и/или слизистым путями.
Обычно иммуногенные композиции приготавливают в виде инъекций, либо в виде жидких растворов, либо в виде суспензий; также можно приготовить твердые формы, подходящие для растворения или суспензирования в жидких средах перед инъекцией. Препарат также можно эмульгировать или инкапсулировать в липосомы для усиления вспомогательного эффекта. Непосредственной доставкой композиций обычно является парентеральная (например, посредством инъекции, либо подкожно, внутрибрюшинно, внутривенно, либо внутримышечно или доставка во внутрипоровое пространство ткани). Данные композиции также можно вводить в очаг повреждения. Другие пути введения включают пероральное и пульмональное введение, суппозитории и трансдермальные или чрескожные наружные применения (например, смотри ссылку 251), шприцы и безыгольные шприцы. Дозированное лечение может быть схемой однократной дозы или схемой многократного приема (например, включающей повторные дозы антигена).
Вакцины данного изобретения предпочтительно являются стерильными. Они предпочтительно являются апирогенными. Их предпочтительно буферизуют, например, между pH 6 и pH 8, обычно около pH 7.
Вакцины данного изобретения могут содержать детергент (например, твин, такой как твин 80) в низких содержаниях (например, <0,01%). Вакцины данного изобретения могут содержать спирт из сахара (например, маннитол) или трегалозу, например, около 15 мг/мл, в частности, если они лиофилизованы.
Оптимальные дозы индивидуальных антигенов можно определить эмпирически. Обычно, однако, антигены данного изобретения будут вводить в дозе между 0,1 и l00 мкг каждого антигена на дозу при обычном объеме дозировки 0,5 мл. Доза обычно представляет собой между 5 и 20 мкг каждого антигена на дозу.
Количество лиганда CD1d, вводимого пациенту, для того чтобы вызвать иммунный ответ, может варьироваться в зависимости от возраста и веса пациента, которому вводят композицию, но будет обычно содержать между 1-100 мкг/кг массы тела пациента. Как не удивительно, было обнаружено, что низкие дозы лиганда CD1d являются достаточными, для того чтобы усилить иммунный ответ на вводимый одновременно антиген и поддержать долговременную иммунологическую память к данному антигену. Количество лиганда CD1d, включенного в композиции данного изобретения, следовательно, может быть менее 50 мкг/кг массы тела пациента, менее 20 мкг/кг, менее 10 мкг/кг, менее 5 мкг/кг, менее 4 мкг/кг или менее 3 мкг/кг.
Вакцины согласно данному изобретению могут быть либо профилактическими (т.е. для того, чтобы предотвратить инфицирование), либо терапевтической (т.е. для того, чтобы лечить заболевание после инфицирования), но обычно будет профилактической.
Данное изобретение обеспечивает лиганд CD1d и антиген из стрептококка группы B для применения в медицине. Данное изобретение обеспечивает лиганд CD1d и антиген из N. meningitidis серологической группы B для применения в медицине. Данное изобретение обеспечивает лиганд CD1d и антиген из вируса гриппа, выбранного из штамма вируса гриппа, который способен или имеет возможность вызывать пандемическую вспышку, для применения в медицине.
Данное изобретение также обеспечивает способ повышения иммунного ответа у пациента, включающий введение пациенту вакцины, согласно данному изобретению. В частности, данное изобретение также обеспечивает способ повышения иммунного ответа у пациента, включающий введение пациенту лиганда CD1d и антигена из стрептококка группы B. Данное изобретение также обеспечивает способ повышения иммунного ответа у пациента, включающий введение пациенту лиганда CD1d и антигена из N. meningitidis серологической группы B. Данное изобретение также обеспечивает способ повышения иммунного ответа у пациента, включающий введение пациенту лиганда CD1d и антигена из вируса гриппа, выбранного из штамма вируса гриппа, который способен или имеет возможность вызывать пандемическую вспышку.
Антиген и лиганд CD1d можно вводить одновременно, последовательно или отдельно. Например, лиганд CD1d можно вводить, для того чтобы подготовить млекопитающее, перед введением антигена или после введения антигена, для того чтобы усиливать иммунный ответ млекопитающего на данный конъюгат. Если вводят более одного антигена, то антигены можно вводить одновременно с лигандом CD1d, который вводится отдельно, одновременно или последовательно со смесью антигенов. Способ повышения иммунного ответа может включать введение первой дозы антигена и лиганда CD1d, а затем введение необязательной второй невспомогательной дозы антигена. Первую дозу антигена и лиганда CD1d можно вводить одновременно, последовательно или отдельно.
Иммунный ответ является предпочтительно защитным иммунным ответом и может включать гуморальный иммунный ответ и/или клеточный иммунный ответ. Пациент может быть взрослым или ребенком. Пациент может быть в возрасте 0-6 месяцев, 6-12 месяцев, 1-5 лет, 5-15 лет, 15-55 лет или старше 55 лет. Предпочтительно пациент является ребенком.
Пациент может быть с иммунной недостаточностью. Пациент может иметь нарушение, связанное с недостатком функции иммунной системы, а в частности, нарушение, связанное с недостатком функции в CD4 T-клеточных ответах. Примеры подобных нарушений включают, но не ограничиваются, СПИД, атаксию-телеангиэктазию, синдром Ди Георге, пангипогаммаглобулинемию, синдром Вискотта-Олдрича и недостаточности комплемента.
Данное изобретение обеспечивает применение антигена из стрептококка группы B в производстве лекарственного средства для повышения иммунного ответа у пациента, причем лекарственное средство вводят с лигандом CD1d. Данное изобретение обеспечивает применение лиганда CD1d в производстве лекарственного средства для повышения иммунного ответа у пациента, причем лекарственное средство вводят с антигеном из стрептококка группы B. Данное изобретение обеспечивает применение антигена из стрептококка группы B и лиганда CD1d в производстве лекарственного средства для повышения иммунного ответа у пациента. Данное изобретение также обеспечивает применение антигена из стрептококка группы B в производстве лекарственного средства для повышения иммунного ответа у пациента, причем пациент был предварительно обработан лигандом CD1d. Данное изобретение обеспечивает применение лиганда CD1d в производстве лекарственного средства для повышения иммунного ответа у пациента, причем пациент был предварительно обработан антигеном из стрептококка группы B. Данное изобретение обеспечивает применение антигена из стрептококка группы B в производстве лекарственного средства для повышения иммунного ответа у пациента, причем пациент был предварительно обработан антигеном из стрептококка группы B и лигандом CD1d.
Данное изобретение также обеспечивает применение антигена из N. meningitidis серологической группы B в производстве лекарственного средства для повышения иммунного ответа у пациента, причем лекарственное средство вводят с лигандом CD1d. Данное изобретение также обеспечивает применение лиганда CD1d в производстве лекарственного средства для повышения иммунного ответа у пациента, причем лекарственное средство вводят с антигеном из N. meningitidis серологической группы B. Данное изобретение также обеспечивает применение антигена из стрептококка группы B (N. meningitidis серологической группы B) и лиганда CD1d в производстве лекарственного средства для повышения иммунного ответа у пациента. Данное изобретение также обеспечивает применение антигена из N. meningitidis серологической группы B в производстве лекарственного средства для повышения иммунного ответа у пациента, причем пациент был предварительно обработан лигандом CD1d. Данное изобретение, кроме того, обеспечивает применение лиганда CD1d в производстве лекарственного средства для повышения иммунного ответа у пациента, причем пациент был предварительно обработан антигеном из N. meningitidis серологической группы B. Данное изобретение также обеспечивает применение антигена из N. meningitidis серологической группы B в производстве лекарственного средства для повышения иммунного ответа у пациента, причем пациент был предварительно обработан антигеном N. meningitidis серологической группы B и лигандом CD1d.
Данное изобретение также обеспечивает применение антигена из вируса гриппа (как описано выше) в производстве лекарственного средства для повышения иммунного ответа у пациента, причем лекарственное средство вводят с лигандом CD1d. Данное изобретение также обеспечивает применение лиганда CD1d в производстве лекарственного средства для повышения иммунного ответа у пациента, причем лекарственное средство вводят с антигеном из вируса гриппа (как описано выше). Данное изобретение также обеспечивает применение антигена из вируса гриппа (как описано выше) и лиганда CD1d в производстве лекарственного средства для повышения иммунного ответа у пациента. Данное изобретение также обеспечивает применение антигена из вируса гриппа (как описано выше) в производстве лекарственного средства для повышения иммунного ответа у пациента, причем пациент был предварительно обработан лигандом CD1d. Данное изобретение, кроме того, обеспечивает применение лиганда CD1d в производстве лекарственного средства для повышения иммунного ответа у пациента, причем пациент был предварительно обработан антигеном из вируса гриппа (как описано выше). Данное изобретение также обеспечивает применение антигена из вируса гриппа (как описано выше) в производстве лекарственного средства для повышения иммунного ответа у пациента, причем пациент был предварительно обработан антигеном из вируса гриппа (как описано выше) и лигандом CD1d.
Лекарственное средство предпочтительно представляет собой иммуногенную композицию (например, вакцину). Лекарственное средство предпочтительно представляет собой средство для предотвращения и/или лечения заболевания, вызванного стрептококком группы B, Neisseria meningitidis (например, менингит, сепсис и т.д.) или вирусом гриппа.
Вакцины можно исследовать в стандартных животных моделях (например, смотри ссылку 252).
Данное изобретение, кроме того, обеспечивает набор реагентов, включающий: стрептококковый антиген группы B и лиганд CD1d. Данное изобретение, кроме того, обеспечивает набор реагентов, включающий антиген из N. meningitidis серологической группы B и лиганд CD1d. Данное изобретение, кроме того, обеспечивает набор реагентов, включающий антиген из вируса гриппа и лиганд CD1d. Антиген и лиганд предпочтительно обеспечиваются в виде отдельных компонентов набора, таким образом, чтобы они подходили для раздельного введения, например, в различные конечности.
Определения
Понятие "содержащий" заключает в себе понятие "включающий", а также "состоящий", например, композиция "содержащая" X может состоять исключительно из X или может включать что-нибудь дополнительное, например, X+Y.
Понятие "около" по отношению к числовому значению x означает, например, x±10%.
Понятие "по существу" не исключает значения "совершенно", например, композиция, которая "по существу не содержит" Y, может совершенно не содержать Y. При необходимости понятие "по существу" можно опустить из определения данного изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 показывает геометрические Ig титры.
A) Сывороточные титры белок-специфических антител в C57/BL6 мышах, иммунизированных внутримышечно α-GC и бактериальными (TT, столбнячный токсин, или DT, дифтерийный токсин) или вирусными белками (H3N2, гемагглютинин-нейраминидазная субъединица из штамма вируса гриппа A). Мыши, иммунизированные белками и α-GC (закрашенные прямоугольники), показали более высокие титры антител, чем мыши, иммунизированные одними белками (пустые прямоугольники). Ось X показывает время в днях.
B) Мыши, несущие iNKT-клетки (Ja18+/+), иммунизированные H3N2 и α-GC (закрашенные прямоугольники), показывают увеличение сывороточного титра H3N2-специфических антител по сравнению с мышами, несущими клетки iNKT, иммунизированными только H3N2 (пустые прямоугольники). Мыши, не содержащие клеток iNKT (Ja18-/-), иммунизированные H3N2 и α-GC (закрашенные прямоугольники), не показывают увеличение сывороточного титра H3N2-специфических антител по сравнению с мышами, не содержащими клеток iNKT, иммунизированными только H3N2 (пустые прямоугольники). Все иммунизации были подкожными.
Фигура 2 показывает геометрическое значение IgG титров.
A) α-GC является таким же сильнодействующим, как и CFA, CpG, MF59 и квасцы, в получении как IgG1 и IgG2a антител. Антигеном был TT.
B) Мыши MHC-II-/-, иммунизированные подкожно H3N2 (и α-GC), повысили детектируемые титры антител (IgG), тогда как мыши MHC-II-/-, иммунизированные подкожно только H3N2 или с квасцами, не повысили.
Фигура 3: Сравнение α-GC и MF59 в мышиной модели инфицирования вирусом гриппа. Все иммунизации были внутримышечными.
A) H1N1-специфические титры IgG (геометрическое значение). Мыши, иммунизированные H1N1 и α-GC, имеют титры антител, которые сравнимы с титрами мышей, иммунизированных H1N1 и MF59, и которые значительно выше, чем титры, обнаруженные в мышах, иммунизированных только белковой вакциной.
B) Процент выживаемости, а именно дни после контрольного заражения. 80% мышей, иммунизированных H1N1 и α-GC, и 100% мышей, иммунизированных H1N1 и MF59, живы после контрольного заражения вирусом гриппа.
Фигура 4 показывает H3N2 титры Ig (геометрическое значение). Белые прямоугольники без вспомогательного вещества; заштрихованные прямоугольники имеют α-GC:
A) IgGl и IgG2a иммунный ответ WT, IL-4-/- и IFN-γR-/- мышей на подкожную иммунизацию H3N2 и α-GC (наличие показано темным) или подкожную иммунизацию только H3N2 (наличие показано белым). Иммунизация мышей дикого типа только H3N2 вызвала IgG1 иммунный ответ (Th2), тогда как иммунизация H3N2 и α-GC вызвала IgGl и IgG2a иммунный ответ (Th0). Иммунизация мышей IL4-/- только H3N2 не вызвала IgG иммунного ответа, тогда как иммунизация H3N2 и α-GC вызвала IgG1 и IgG2a иммунный ответ (Th0). Иммунизация мышей IFN-γR-/- только H3N2 вызвала IgGl иммунный ответ (Th2), а иммунизация H3N2 и α-GC вызвала значительно более сильный иммунный ответ IgG1 (Th2). Пунктирная линия показывает минимальное разбавление исследуемой сыворотки.
B) Мыши, обработанные моноклональным антителом анти-CD40L перед и во время подкожной иммунизации H3N2 и α-GC, показывают H3N2 титры антител, которые значительно ниже, чем титры, наблюдаемые в мышах, обработанных контролем IgG.
Фигура 5:
A) Мышей первично обрабатывали на 0 и 2 неделях только H3N2 или H3N2 и α-GC. Через 30 недель после первой иммунизации обе группы мышей повторно иммунизировали только H3N2 белком. Фигура показывает H3N2-Ig титр (геометрическое значение), а именно время (недели). Стрелки показывают время иммунизаций. Мыши, первично обработанные двумя дозами H3N2 и α-GC, а затем повторно обработанные только белком (закрашенные прямоугольники), показали титры антител значительно выше, чем мыши, первично обработанные и повторно иммунизированные только H3N2 (пустые прямоугольники). Иммунизации были подкожными.
B) Частота встречаемости клеток-предшественников, секретирующих антитело H3N2, в мышах, первично обработанных согласно Фигуре 5A (H3N2-IgG ASC предшественники на миллион B-клеток). Частота встречаемости клеток-предшественников, секретирующих антитело H3N2, на 30 неделе была значительно выше в мышах, иммунизированных дважды H3N2 и α-GC (заштрихованный), чем в мышах, иммунизированных дважды только H3N2 (белый).
Фигура 6: H3N2-Ig титр (геометрическое значение), а именно время (недели). Спад H3N2-специфических антител в мышах, не содержащих клетки iNKT (Jaα 18-/-), и мышах, несущих клетки iNKT (Jaα 18+/+), иммунизированных дважды подкожно только H3N2. Спад антиген-специфических антител быстрее в мышах, не содержащих клетки iNKT (треугольники), чем в мышах, несущих клетки iNKT (круги).
Фигура 7 показывает частоту встречаемости ASC предшественников (т.е. B-клетки памяти) в C57BL/6 мышах через 6 недель после последней иммунизации (из двух) столбнячным токсином +/- вспомогательными веществами, в виде числа на миллион B-клеток. C57BL/6 мышей иммунизировали внутримышечно на 0 день и на 14 день столбнячным токсином без вспомогательного вещества, α-GC или квасцов. Мыши, иммунизированные столбнячным токсином и α-GC, показали частоту встречаемости TT-специфических B-клеток памяти значительно выше, чем мыши, иммунизированные только столбнячным токсином, все мыши, иммунизированные столбнячным токсином в квасцах, не показали. * показывает p<0,05, а именно безантигеновое введение, а ** показывают p<0,01. *** показывают p<0,05, а именно TT без вспомогательного вещества.
Фигура 8: Схема иммунизации, использованная, для того чтобы определить, является ли необходимым присутствие α-GC во всех вакцинных дозах, применяемых для первичной обработки. 20 C57BL/6 мышей женского пола в возрасте 6-8 недель разделили на 4 группы из 5 мышей. Группу 1) иммунизировали H3N2 в ФБР на 0 неделе и H3N2 + α-GC через 2 недели. Группу 2) иммунизировали H3N2 + α-GC на 0 неделе и H3N2 в ФБР через 2 недели. Группу 3) иммунизировали H3N2 в ФБР на 0 неделе и на 2 неделе. Группу 4) иммунизировали H3N2 и α-GC на 0 неделе и через 2 недели. Через 56 недель после первоначальной иммунизации мышей из всех групп подвергли обработке 3 мкг H3N2 в ФБР, а вызванный повторно иммунный ответ определяли на 58 неделе. Все иммунизации были внутримышечными.
Фигура 9: Сравнение иммунного ответа H3N2-антител мышей в группе 3 Фигуры 8 (иммунизированных дважды H3N2 в ФБР) с: A) иммунными ответами в мышах 4 группы Фигуры 8 (иммунизированных дважды H3N2 в α-GC); B) иммунным ответoм мышей в группе 1 Фигуры 8 (иммунизация H3N2 в ФБР, а затем H3N2 в α-GC); и C) иммунными ответами в мышах 2 группы Фигуры 8 (иммунизация H3N2 в α-GC, а затем H3N2 в ФБР). Различий во временах полужизни антител не было.
Фигура 10: Сравнение иммунного ответа H3N2-антител, наблюдаемого после: A) иммунизации дважды α-GC (группа 4) по отношению к α-GC только во второй иммунизации (группа 1); B) мышей, иммунизированных дважды α-GC (группа 4) по отношению к α-GC только в первой иммунизации (группа 2); и C) иммунизации α-GC только в первой иммунизации (группа 2) по отношению к иммунизации α-GC только во второй иммунизации (группа 1).
Фигура 11: Повторно вызванный иммунный ответ мышей, иммунизированных, как описано в Фигуре 8. Через две недели после повторной иммунизации только H3N2 (проведенной на 56 неделе), мыши, первично обработанные одной или двумя дозами α-GC, показали более высокие повторно вызванные иммунные ответы, чем мыши, первично обработанные 2 дозами только H3N2. Данные представляют собой H3N2 Ig титры (геометрическое значение) на 56 & 58 неделях.
Фигура 12: Были определены частоты встречаемости MenB-специфических B-клеток памяти в селезенке мышей, иммунизированных, как описано в Фигуре 13. Более высокие частоты встречаемости MenB-специфических B-клеток памяти были обнаружены в селезенках мышей, иммунизированных α-GC или MF59 по сравнению с квасцами. График показывает MenB-специфические IgG B-клетки памяти на миллион B-лимфоцитов. * & **: p<0,05 & p<0,01, а именно без вспомогательного вещества.
Фигура 13: Мышей иммунизировали смесью 3 MenB антигенов, ΔG287nz-953, 936-741 и 961c (20 мкг/доза, 5 мкг/доза или 2,5 мкг/доза каждый) с примесями 0,1 мкг α-GC, 0,6 мг квасцов, l00 мкл MF59 или без вспомогательного вещества. Была проведена серия иммунизаций на 0, 21 и 35 дни, а IgG титры к каждому антигену определяли после каждой иммунизации, вплоть до 105 дня. Как α-GC, так и MF59 вызывали более высокие бактерицидные титры антител, чем квасцы.
Фигура 14: Сравнение CD4 T-клеточного иммунного ответа на рекомбинантные MenB антигены в мышах, иммунизированных внутримышечно на 0 и 21 день: a) комбинированной вакциной, содержащей антигены 3 MenB и α-GC; b) комбинированной вакциной, содержащей антигены 3 MenB и квасцы; или c) комбинированной вакциной, содержащей только антигены 3 MenB. CD4 T-клеточный ответ определяли через две недели после второй иммунизации посредством выдерживания всех спленоцитов с определенным количеством рекомбинантных MenB белков в течение 16 часов (последние 14 из которых в присутствии Брефелдин A). Число CD4 T-клеток, вырабатывающих TNFa, определяли посредством внутриклеточного окрашивания и FACS анализа. Мыши, иммунизированные комбинацией из трех MenB антигенов и α-GC, согласованно показали более сильный CD4 иммунный ответ по сравнению с мышами, иммунизированными комбинацией из MenB антигенов и квасцов или без вспомогательного вещества. В качестве положительного контроля исследовали иммунный ответ всех трех групп мышей на поликлональную стимуляцию. Все три группы мышей показали одинаковый иммунный ответ на поликлональную стимуляцию анти-CD3 антитела (IaCD3), как показано во вставке Фигуры. Ось Y показывает CD4 T-клетки, вырабатывающие TNFα, в виде процентного содержания от всех CD4+ T-клеток.
Фигура 15: Титры (геометрическое значение). Сравнение титров IgG, IgG1 и IgG2a в мышах, иммунизированных антигенами GBS. Мыши, иммунизированные 1 мкг GBS80 и α-GC, показали значительно более высокие титры IgG1 и IgG2a, чем мыши, иммунизированные 1 мкг только GBS80, тогда как мыши, иммунизированные GBS80 в квасцах, не показали. Мыши, иммунизированные 20 мкг GBS80 и α-GC, показали титры IgG1, равные по значению титрам мышей, иммунизированных 20 мкг GBS80 и квасцами, и более высокие титры IgG2a. *, ** p<0,05, p<0,01, а именно GBS80 без вспомогательного вещества.
Фигура 16: Мышей иммунизировали на 0 день и на 21 день одним из трех антигенов MenB (DG287nz-953, 936-741 или 961c) или смесью всех трех антигенов в комбинации с: a) α-GC; b) квасцами; или c) без вспомогательного вещества. Уровни бактерицидных антител против MenB штаммов MC58, 2996, H44/76 и NZ98/254 определяли через две недели после второй иммунизации и через две недели после третьей иммунизации. Бактерицидных антител было значительно больше, если комбинированную вакцину вводили α-GC в качестве вспомогательного вещества, по сравнению с квасцами. Все иммунизации были внутримышечными.
Фигура 17: Частоты встречаемости B-клеток памяти в селезенке матери, иммунизированной GBS80. Частоты встречаемости плазмоцитов, вырабатывающих GBSSO-специфические антитела, были значительно выше в селезенках матерей, иммунизированных GBS80 и α-GC, чем в селезенках матерей, иммунизированных только GBS80 или с квасцами. График показывает число GBS80 IgG плазмоцитов на миллион лимфоцитов.
Фигура 18: Сравнение частот встречаемости плазмоцитов матерей, иммунизированных GBS80 и α-GC, и матерей, иммунизированных GBS80 и квасцами. Частоты встречаемости плазмоцитов были значительно выше у матерей, иммунизированных GBS80 и α-GC. График показывает число GBS80 IgG плазмоцитов на миллион B-лимфоцитов.
Фигура 19: Мышей иммунизировали только смесью из 3 MenB антигенов, ΔG287nz-953, 936-741 и 961c (20 мкг/доза каждый), с примесями 0,1 мкг α-GC или с примесями 0,6 мг квасцов. Серию из трех иммунизаций осуществляли на 0, 21 и 35 дни, а титры IgG на каждый антиген определяли после каждой иммунизации. α-GC был также эффективен в усилении иммунного ответа антител, как и квасцы, для всех трех антигенов MenB в комбинированной вакцине. Все иммунизации были внутримышечными.
СПОСОБЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Дополнительную информацию, касающуюся способов осуществления данного изобретения, можно найти в ссылке 253.
Пример 1: Инвариантные клетки NKT содействуют защитным иммунным ответам in vivo и вносят вклад в поддержание B-клеточной памяти
Сущность изобретения
Рестриктированные по CD1d естественные инвариантные T-клетки киллеры (iNKT) представляют собой врожденные лимфоциты, которые распознают гликолипидные антигены, такие как α-галактозилцерамид (α-GC). Для того чтобы исследовать эффекты врожденной иммунной системы на адаптивные иммунные ответы in vivo, авторы определяли, влияют ли клетки iNKT на определяющие характеристики иммунного ответа, такие как защита от инфекций и B-клеточная память. Авторы иммунизировали мышей бактериальными или вирусными белками в комбинации с α-GC и обнаружили, что мыши, иммунизированные белками и α-GC, вырабатывают титры антител, которые на от одного до двух порядков выше, чем титры, вызванные только белками, и, что важнее, они больше защищают от инфекций, таких как вирус гриппа. Мыши, которые не содержат MHC класса II, не вырабатывают антитела при иммунизации белками и общепринятыми вспомогательными веществами, однако, иммунизация данных мышей белками и α-GC вызывает детектируемый IgG, специфичный для данного белка, показывая, что клетки iNKT могут частично восстановить содействие B-клеткам посредством класса II рестриктированных CD4+ T-клеток. Наконец, авторы обнаружили, что мыши, иммунизированные белками и α-GC, имеют частоту встречаемости белок-специфических B-клеток памяти, которая выше, чем частота встречаемости, наблюдаемая в мышах, иммунизированных только белками. Кроме того, мыши, не содержащие клетки iNKT, показывают спад титров циркулирующих антител, который быстрее, чем спад, наблюдаемый в мышах дикого типа, наводя на мысль о непредвиденном влиянии клеток iNKT на продолжительность жизни плазмоцитов. Итак, данные результаты исследования указывают на важную роль клеток iNKT в регулировании иммунного ответа и поддержке B-клеточной памяти in vivo.
Результаты
Активация клеток iNKT усиливает иммунный ответ на белковые антигены in vivo.
Авторы недавно показали, что человеческие клетки iNKT могут содействовать B-лимфоцитам при размножении и выработке иммуноглобулинов in vitro. Для того чтобы определить in vivo значимость данного открытия, авторы иммунизировали C57/BL6 мышей бактериальными (TT, столбнячный токсин, или DT, дифтерийный токсин) или вирусными белками (H3N2, гемагглютинин-нейраминидазная субъединица из штаммов вируса гриппа A) с или без NKT специфического гликолипида, α-GC, и определяли сывороточные титры белок-специфических антител в различные моменты времени. Фигура 1A показывает, что со всеми антигенами мыши, иммунизированные белками и α-GC (закрашенные прямоугольники), показали титры антител на от одного до двух порядков выше, чем титры мышей, иммунизированных только белками (пустые прямоугольники). Подобные результаты были получены в BALB/c, CD1 и C3H/HeJ мышах (данные не показаны).
Для того чтобы доказать, что активность вспомогательного вещества α-GC была обусловлена активацией клеток iNKT, авторы иммунизировали мышей, несущих (Ja18+/+ и Ja18+/-) или не содержащих (Ja18-/-) клетки iNKT, гриппозными белками H3N2 с или без α-GC. Как показано на Фигуре 1B, все мыши, иммунизированные только H3N2 (пустые прямоугольники), проявляли сравнимые иммунные ответы независимо от наличия клеток iNKT. Однако Фигура 1B показывает, что если иммунизация осуществляется H3N2 и α-GC (закрашенные прямоугольники), то мыши, несущие клетки iNKT, показывают значительное увеличение сывороточного титра H3N2-специфических антител, тогда как мыши, не содержащие iNKT, не способны показать подобный результат. Данные результаты были подкреплены таким открытием (данные не показаны), что α-GC не проявлял активность вспомогательного вещества в мышах, не содержащих CD1d (CD1-/-), рестрикционный элемент, который представляет α-GC к T-клеточному рецептору клеток iNKT.
Для того чтобы сравнить активность α-GC с активностью более общепринятых вспомогательных веществ, мышей иммунизировали повышенными дозами TT, вводимыми отдельно, с α-GC или с оптимальной дозой одного из следующих вспомогательных веществ: CFA (одно из самых сильных вспомогательных веществ, которое применяется в мышах [254]), CpG (сильный Th0/Th1 иммуностимулятор, который в настоящее время исследуется у человека 255), MF59 и квасцы (два вспомогательных вещества, лицензированные для применения у человека [256, 257], оба рассматриваемые Th0/Th2 индуцирующие факторы). Как показано на Фигуре 2A, α-GC в основном находится как можно выше эталонных вспомогательных веществ в содействии выработки как антител IgG1, так и IgG2a.
Наконец, авторы определяли, могли ли иммунные ответы на белковые антигены вырабатываться при содействии iNKT лимфоцитов в отсутствие общепринятого CD4+T-клеточного хелпера, в положении, когда общепринятые вспомогательные вещества не способны обеспечить содействие. Таким образом, две группы C57BL/6 мышей, которые не содержат молекулы MHC класса II (MHC-II-/-), иммунизировали дважды H3N2, вводимым отдельно или с α-GC или с квасцами. Как ожидается, MHC-II-/- мыши, иммунизированные только H3N2 или в квасцах, не показали никаких антиген-специфических антител (Фигура 2B). В таком случае MHC-II-/- мыши, иммунизированные H3N2 и α-GC, установили детектируемые титры антитела (IgG).
Итак, данные результаты показывают, что клетки iNKT, активированные in vivo посредством α-GC, делают возможными иммунные ответы на белковые антигены, в некоторой степени, сравнимые с иммунными ответами общепринятых вспомогательных веществ. В противоречии с данными вспомогательными веществами, α-GC не нуждается в MHC класса II рестриктированных CD4 T-лимфоцитах, для того чтобы вырабатывать иммунный ответ.
Вспомогательный иммунитет iNKT-клеток
Показав, что α-GC усиливает иммунный ответ на патогенные белки, авторы обратились к качеству иммунного ответа и задались вопросом, способны ли данные антитела защитить от инфекций. С этой целью авторы сравнили вспомогательный эффект α-GC с вспомогательным эффектом MF59 (вспомогательное вещество, лицензированное для применения у человека с гриппозными вакцинами) в мышиной модели инфицирования вирусом гриппа. Взрослых мышей C57BL/6 иммунизировали на 0 и 15 дни только H1N1 белками (из человеческого вируса гриппа A/NewCaledonia/20/99), с α-GC или с MF59. Через две недели после последней иммунизации мышей подвергли 90% летальной дозе (LD) адаптированного к мышам A/WS/33 вируса гриппа, и их выживаемость отслеживали в течение двух недель. Как показано на Фигуре 3A, за один день до контрольного заражения мыши, иммунизированные H1N1 и α-GC, имеют титры антител, сравнимые с титрами мышей, иммунизированных H1N1 и MF59, и они значительно выше, чем титры, обнаруженные в мышах, иммунизированных только белковой вакциной. Кроме того, Фигура 3B показывает, что через две недели после контрольного заражения 80% мышей, иммунизированных H1N1 и α-GC, и 100% мышей, иммунизированных белками и MF59, были живы, тогда как только 10% мышей, которые были иммунизированы вакциной, основанной только на белках, были все еще живы на момент окончания отслеживания.
Итак, из данных результатов авторы заключили, что α-GC-зависимая iNKT-клеточная активация может увеличивать эффективность вакцин против инфекционнных заболеваний.
Механизм iNKT-клеточного содействия B-клеткам
Затем авторы исследовали механизмы, управляющие iNKT-клеточным содействием B-клеткам in vivo.
Во-первых, для того чтобы исследовать роль цитокинов, авторы определяли вспомогательный эффект α-GC как в C57BL/6 мышах, так и в конгенных мышах, не содержащих цитокин IL-4 или рецептор для IFN-γ (IFN-γR). Фигура 4A (левая панель) показывает, что в мышах дикого типа иммунизация только гриппозными белками 3HN2 (показано белым) вызвала Th2 иммунный ответ, но что указывает IgG1 и отсутствие IgG2a, тогда как иммунизация белком и α-GC вызвала сбалансированный Th0 иммунный ответ, что показывается наличием как IgG1, так и IgG2a (показано черным). Средняя панель Фигуры 4A показывает, что мыши, не содержащие IL-4, не имеют никакого иммунного ответа при иммунизации только белком, тогда как они установили сбалансированный Th0 иммунный ответ при иммунизации белком и α-GC (показано черным). Наконец, мыши, не содержащие IFN-γ рецептора (Фигура 4A, правая панель), показывают Th2 иммунный ответ (IgG1 антитела) при иммунизации только белком (показано белым). Хотя титры IgG1 значительно увеличиваются в мышах, иммунизированных белком и α-GC (показано черным), увеличения антител IgG2a выше естественных уровней не происходит. Итак, данные результаты исследований показывают, что отдельно взятый IL-4 является необязательным для α-GC-зависимой клетки iNKT, для того чтобы содействовать B-лимфоцитам, тогда как IFN-γ является существенным для сбалансированного (Th0) хелперного эффекта клеток iNKT.
Во-вторых, авторы задались вопросом, являлись ли CD40/CD40L взаимодействия необходимыми для α-GC-зависимого iNKT-клеточного хелпера in vivo. Мы, следовательно, определяли иммунные ответы на H3N2 в мышах, обработанных насыщающими количествами нейтрализующего анти-CD40L mAb. Как показано на Фигуре 4B, после иммунизации H3N2 и α-GC мыши, обработанные анти-CD40L mAb, показали титры H3N2 антител, которые значительно ниже, чем титры, наблюдаемые в мышах, обработанных контрольным IgG.
α-GC усиливает повторные иммунные ответы и вносит вклад в поддержание B-клеточной памяти
Ключевой характеристикой адаптивной иммунной системы является способность устанавливать быстрый "повторно вызванный" иммунный ответ на антиген, который встречался ранее. Для того чтобы определить, оказывал ли вспомогательный эффект α-GC влияние на повторно вызванные иммунные ответы, мышей иммунизировали дважды на 0 и 2 неделе только H3N2 или с α-GC. Затем осуществили третью (повторную) иммунизацию всех мышей только H3N2 на 30 неделе. Фигура 5A показывает, что в соответствии с данными, представленными на Фигуре 1, после первых двух доз мыши, иммунизированные H3N2 и α-GC, показали титры антител значительно выше, чем титры из мышей, получающих только H3N2. H3N2-специфические антитела спадали с течением времени, достигая естественных уровней в обеих группах на около 30 неделе, если все мыши были повторно подвергнуты третьей иммунизации только H3N2. Через две недели авторы определяли иммунные ответы и обнаружили, что мыши, которым вводили белок и α-GC во время первых двух иммунизаций, показали титры антител после третьей значительно выше, чем титры мышей, которые были иммунизированы во время всех трех иммунизаций только белком (Фигура 5A). В соответствии с данными результатами, в параллельном эксперименте авторы обнаружили, что частота встречаемости клеток-предшественников, секретирующих H3N2-антитела (ASC), определенная на 30 неделе (непосредственно перед третьей иммунизацией) в селезенке мышей, иммунизированных дважды H3N2 и α-GC, была значительно выше, чем частота, наблюдаемая в селезенке мышей, иммунизированных дважды только H3N2 (Фигура 5B).
Для дальнейшего исследования роли клеток iNKT в регулировании B-клеточной памяти авторы определяли устойчивость антиген-специфических антител, вызванных только белком (H3N2) в сыворотке мышей, несущих (Ja18+/+ и Ja18+/-) или не содержащих (Ja18-/-) клетки iNKT. Во всех группах мышей титры антиген-специфических антител достигали сравнимых уровней через две недели после второй иммунизации. Однако Фигура 6 показывает, что в то время как в мышах, несущих клетки iNKT, антиген-специфические антитела спадали с равномерной низкой скоростью, спад титра антител в мышах, не содержащих клетки iNKT, был значительно быстрее. Так как ни одна из данных мышей не получала α-GC, то авторы заключили, что некоторый уровень "спонтанной" iNKT активности может оказывать влияние на время полужизни циркулирующих антител.
Итак, данные результаты исследования показывают, что iNKT-клеточная активация приводит к более сильному иммунному ответу на повторную иммунизацию, и что данный факт обусловлен усиленным расширением антиген-специфического B-клеточного пула памяти. Кроме того, спонтанная iNKT активность in vivo, как оказалось, играет гомеостатическую роль в поддержании уровней циркулирующих антител.
Пример 2: Примирование a-GalCer в отсутствие повторного стимулирования значительно усиливает иммунный ответ
Как отмечалось выше, частота встречаемости клеток-предшественников, секретирующих H3N2-антитела (ASC) (т.е. B-клеток памяти), определенная на 30 неделе (непосредственно перед третьей иммунизацией) в селезенке мышей, иммунизированных дважды H3N2 и α-GC, была значительно выше, чем частота, наблюдаемая в селезенке мышей, иммунизированных дважды только H3N2 (Фигура 5B).
Схожие результаты были получены в экспериментах, проведенных на мышах, иммунизированных столбнячным токсином (Фигура 7 и 18). Фигура 7 показывает частоту встречаемости клеток-предшественников ASC в C57BL/6 мышах через 6 недель после последней из двух иммунизаций на 0 день и на 14 день столбнячным токсином без вспомогательного вещества, с вспомогательным веществом α-GC или с вспомогательным веществом квасцами. Применение α-GC в качестве вспомогательного вещества значительно увеличило частоту встречаемости клеток-предшественников ASC по сравнению с применением вспомогательного вещества квасцов. Аналогично Фигура 18 показывает, что частота встречаемости клеток предшественников ASC в CD1 мышах была значительно выше через три месяца после последней из двух иммунизаций GBS80 и α-GC по сравнению с иммунизациями GBS80 и квасцами.
Способность α-GC значительно увеличивать частоту встречаемости B-клеток памяти по сравнению с квасцами при введении в качестве вспомогательного вещества в серии из двух иммунизаций навело на мысль, что α-GC мог бы быть способным вызывать увеличение специфических B-клеток памяти при применении в качестве вспомогательного вещества в однократной иммунизации. Эксперимент, следовательно, проводили, для того чтобы определить эффект однократной дозы α-GC и H3N2 антигена на специфический B-клеточный пул памяти (Фигуры 8-11).
Фигура 8 показывает схему иммунизации, применяемую в эксперименте. 20 C57BL/6 женских особей мышей в возрасте 6-8 недель разделили на 4 группы по 5 мышей. Группу 1) иммунизировали H3N2 в ФБР на 0 неделе и H3N2 + α-GC через две недели. Группу 2) иммунизировали H3N2 + α-GC на 0 неделе и H3N2 в ФБР через две недели. Группу 3) иммунизировали H3N2 в ФБР на 0 неделе и на 2 неделе. Группу 4) иммунизировали H3N2 и α-GC на 0 неделе и через 2 недели. Через 56 недель после первоначальной иммунизации мышей из всех групп подвергли обработке 3 мкг H3N2 в ФБР. Все иммунизации были внутримышечными.
Фигура 9 сравнивает H3N2-иммунный ответ мышей в группе 3 (иммунизированных H3N2 в ФБР) с иммунными ответами в мышах, иммунизированных α-GC во время обеих иммунизаций (панель A), α-GC только во время первой иммунизации (панель B) и α-GC только во время второй иммунизации (панель C). Было обнаружено, что α-GC усиливает иммунный ответ даже при введении только во время первой или второй иммунизации. Различий между четырьмя группами во времени полужизни антитела не наблюдалось.
Фигура 10 обеспечивает парные сравнения иммунного ответа, наблюдаемого после: A) иммунизации дважды α-GC по отношению к α-GC только во время второй иммунизации; B) мышей, иммунизированных дважды α-GC, а именно α-GC только во время первой иммунизации; и C) иммунизации дважды α-GC C только во время первой иммунизации по отношению к иммунизации α-GC только во время второй иммунизации. Максимальная эффективность наблюдалась при введении α-GC в первой дозе вакцины. Поставка α-GC в первой дозе вакцины произвела сильный иммунный ответ по отношению к поставке его во второй дозе вакцины (Фигура 10C), и иммунный ответ при поставке α-GC в первой дозе вакцины был схож с иммунным ответом, полученным при поставке α-GC в обеих дозах вакцины (Фигура 10B).
Фигура 11 подтверждает, что мыши, первично обработанные α-GC, показывают сильный повторный иммунный ответ на вакцинацию, даже если α-GC включается только в первую или вторую дозу из двух примирующих инъекций. Данные результаты наводят на мысль о том, что включение α-GC в качестве вспомогательного вещества в вакцинные композиции может уменьшить число примирующих иммунизаций, необходимых, для того чтобы достигнуть долговременной иммунологической памяти и уменьшить частоту встречаемости и число повторных иммунизаций.
Пример 3: α-GC усиливает иммунные ответы в мышиной модели неонатального сепсиса, вызванного Streptococcus agalatiae
α-GC исследовали на его способность усиливать защитные иммунные ответы в мышиной модели неонатального сепсиса, вызванного инфекцией Streptococcus agalatiae.
Женские особи мышей разделили на 3 группы. Группу 1 первоначально обрабатывали на 0 день 20 мкг GBS80 в отсутствие вспомогательного вещества и повторно обрабатывали той же самой композицией на 21 день. Мышей спаривали на 23 день и забирали кровь на 43-36 дни, для того чтобы сделать возможным определение титров GBS80-IgG перед рождением потомства на 50-53 дни. Потомство подвергли обработке 90% летальной дозой S. agalactiae в возрасте 0-48 часов от роду. Через 3 месяца после повторной дозы матерей умертвили, выделили селезенки и определили частоты встречаемости предшественников GBS80-IgG плазмоцитов. Такой же схеме иммунизации следовали в группах 2 и 3, за исключением того, что мышей в группе 2 первично и повторно обрабатывали GBS80 и квасцами, а мышей в группе 3) первично и повторно обрабатывали GBS80 и 0,1 мкг α-GC.
Результаты были следующими:
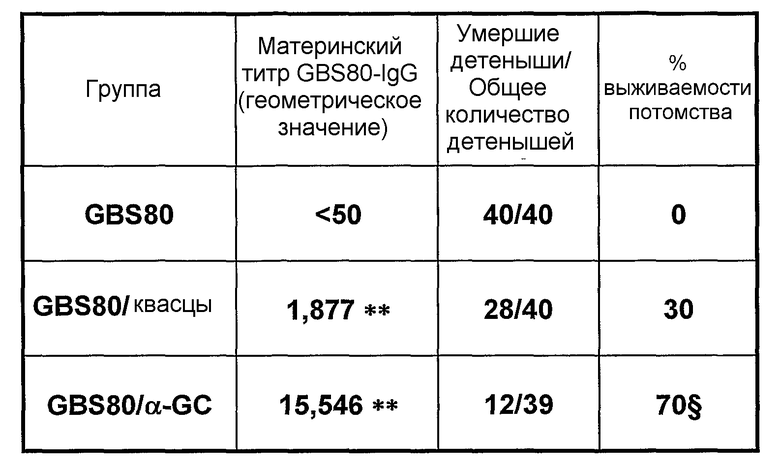
Таким образом, применение α-GC в качестве вспомогательного вещества вызвало GBS80-IgG иммунный ответ у матерей, который был в 8 раз сильнее, чем иммунный ответ IgG, вызванный квасцами. Более сильный иммунный ответ у матери привел к усиленной защите ее потомства от GBS инфекции. 70% потомства от матери, иммунизированной GBS80 и α-GC, выжило при подвергании обработке S. galactiae по сравнению со всего 30% потомства матерей, иммунизированных GBS80 и квасцами.
Эксперимент повторили с мышами, которые были иммунизированы либо 20 мкг, либо 1 мкг GBS80. Всех мышей первично обрабатывали на 0 день, повторно обрабатывали на 20 день, спаривали на 34 день и забирали кровь на 48 день перед рождением потомства на 54-58 дни. Потомство немедленно подвергли обработке 90% летальной дозой S. Agalactiae и определяли выживаемость около 48 часов. Матерей умертвили через 3 месяца после повторной иммунизации, и селезенки удалили для определения частот встречаемости предшественников плазмоцитов GBSSO-IgG. Мышей иммунизировали: 1 мкг GBS80 с квасцами, α-GC или без вспомогательного вещества; 20 мкг GBS80 с квасцами, α-GC или без вспомогательного вещества; или ФБР или только квасцовым вспомогательным веществом.
Как показано на Фигуре 15, мыши, иммунизированные 1 мкг GBS80 и α-GC, показали значительно более высокие титры IgG1 и IgG2a, чем мыши, иммунизированные 1 мкг GBS80 и квасцами. Мыши, иммунизированные 20 мкг GBS80 и α-GC, показали титры IgG1, равные по значению титрам мышей, иммунизированных 20 мкг GBS80 и квасцами, и более высокие титры IgG2a. Результаты были следующими:
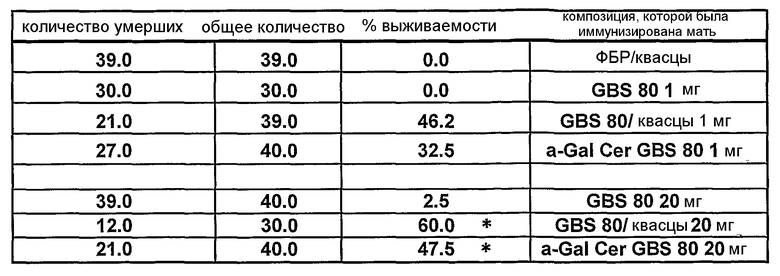
Таким образом, % выживаемости у мышей, иммунизированных GBS80 и α-GC, был равным по значению выживаемости у мышей, иммунизированных GBS80 и квасцами.
Селезенки матерей, иммунизированных GBS80 и α-GC, также содержали значительно более высокие частоты встречаемости плазмоцитов GBS80 IgG и B-клеток памяти (Фигуры 17 и 18, соответственно). Частоты встречаемости GBS80-специфических плазмоцитов и B-клеток памяти находили путем определения наличия GBS80-специфических антител в 10-дневных супернатантах из культур спленоцитов низкой плотности, инкубированных только в культуральной среде или в присутствии CpG и IL-2, как описано в ссылке 258.
В дополнительном эксперименте беременных CD1 женских особей мышей иммунизировали ФБР, GBS80, GBS80 + квасцы или GBS80 + α-GC. Сыворотку, взятую за неделю до родов из каждой группы иммунизированных CD1 женских особей или из неподвергнутых никакому воздействию CD1 женских особей, объединили и ввели подкожно (3 мкл/доза в конечном объеме 20 мкл) новорожденным детенышам 24-часового возраста, рожденным от неподвергнутых никакому воздействию CD1 матерей. Через 3 часа всех новорожденных детенышей подвергли обработке внутрибрюшинно 1 LD90 живого Streptococcus agalactiae. Выживаемость детенышей отслеживали в течение 2 дней. Детеныши, иммунизированные сывороткой матерей, иммунизированных GBS80, все умерли (28 детенышей из 28), и только 1 из 27 детенышей, иммунизированных сывороткой матерей, иммунизированных ФБР, выжил. Наличие вспомогательных веществ (квасцы или α-GC) улучшило выживаемость при иммунизации α-GC, который был более эффективным, чем квасцы, в увеличении выживаемости. Выживаемость детенышей, иммунизированных сывороткой матерей, иммунизированных GBS+α-GC, была на 165% выше, чем выживаемость детенышей, иммунизированных сывороткой матерей, иммунизированных GBS+квасцы.
Подобные данные показывают, что α-GC является, как не удивительно, значительно более эффективным, чем квасцы, в вызывании защитного иммунного ответа на S. agalactiae.
Пример 4: a-GC усиливает иммунный ответ на комбинированную вакцину, содержащую несколько белковых антигенов из N. meningitidis серологической группы B
Определяли способность α-GC действовать в качестве вспомогательного вещества для комбинаций антигенов из N. meningitidis серологической группы B, (MenB) антигенов.
Мышей иммунизировали смесью из 3 MenB антигенов, ΔG287nz-953, 936-741 и 961c (20 мкг/доза каждый) с примесями 0,1 мкг α-GC или с примесями 0,6 мг квасцов. Была осуществлена серия из трех иммунизаций на 0, 21 и 35 дни, и определяли IgG титры к каждому антигену после каждой иммунизации. Как показано на Фигуре 19, α-GC был также эффективен, как и квасцы в усилении иммунного ответа на все три MenB антигена в комбинированной вакцине.
α-GC также усилил бактерицидный иммунный ответ на данные антигены. Фигура 16 сравнивает бактерицидные иммунные ответы на MenB штаммы MC58, 2996, H44/76 и NZ98/254 в образцах сыворотки мышей, иммунизированных смесью из 3 MenB антигенов ΔG287nz-953, 936-741 и 961c или каждым из данных антигенов отдельно в комбинации с квасцами, α-GC или без вспомогательного вещества. Бактерицидные иммунные ответы на иммунизацию антигенами MenB и α-GC были согласованно выше, чем ответ на иммунизацию антигенами MenB и квасцами.
Второй эксперимент проводили, для того чтобы определить ex vivo CD4 T-клеточный ответ на антигены MenB. Группы из 6 CD1 женских особей мышей иммунизировали дважды смесью антигенов MenB, приготовленной в ФБР, квасцах или α-GC. Через 10 дней после 2-ой иммунизации 3 мышей/группа умертвили, а их селезенки удалили. Все спленоцитные суспензии из отдельных мышей культивировали с антигенами MenB в течение 16 часов, последние 12 из которых были в присутствии Брефелдина A, для того чтобы сделать возможным внутриклеточную аккумуляцию цитокинов. Стимулированные спленоциты зафиксировали, нарушили проницаемость мембраны и окрасили анти-CD3, анти-Cd4, анти-CD69, анти-IFNg и анти-TNFa моноклональными антителами. Процентные содержания CD3+CD4+CD69+цитокина+клеток в общей CD4+ клеточной популяции определяли посредством FACS анализа.
Данные результаты показаны на Фигуре 14. Было обнаружено, что α-GC является более эффективным, чем квасцы, в размножении CD4 T-клеток, вырабатывающих TNFα в ответ на MenB антигены, показывая, что α-GC способен вызывать клеточно-опосредованный иммунный ответ на MenB антигены, который является, по крайней мере, равным иммунному ответу, вызываемому квасцами. В качестве положительного контроля был исследован иммунный ответ всех трех групп мышей на поликлональную стимуляцию. Все три группы мышей показали одинаковый иммунный ответ на поликлональную стимуляцию анти-CD3 антителом (IaCD3), как показано во вставке Фигуры 14.
Дополнительный эксперимент провели, для того чтобы определить способность α-GC действовать в качестве вспомогательного вещества для одних и тех же трех антигенов MenB по сравнению с квасцами или MF59. Мышей иммунизировали смесью 3 MenB антигенов, ΔG287nz-953, 936-741 и 961c (20 мкг/доза, 5 мкг/доза или 2,5 мкг/доза каждый) с примесями 0,1 мкг α-GC, 0,6 мг квасцов, 100 мкл MF59 или без вспомогательного вещества. Серию из трех иммунизаций провели на 0, 21 и 35 дни, а титры IgG к каждому антигену определяли после каждой иммунизации. Как показано на Фигуре 13, как α-GC, так и MF59 вызывали более высокие бактерицидные титры антител, чем квасцы. Также определяли частоты встречаемости MenB-специфических B-клеток памяти в селезенке иммунизированных мышей. Как показано на Фигуре 12, более высокие частоты встречаемости MenB-специфических B-клеток памяти были обнаружены в селезенках мышей, иммунизированных α-GC или MF59 по сравнению с квасцами. Подобные данные показывают, что α-GC является более эффективным, чем квасцы, и таким же эффективным, как и MF59, в вызывании бактерицидного иммунного ответа на антигены MenB и в вызывании Men-B специфических B-клеток памяти, необходимых для долговременной иммунологической памяти.
Будет понятно, что данное изобретение было описано только посредством примеров, и возможно осуществить изменение деталей, не отклоняясь от сущности и объема изобретения.
ССЫЛКИ
[1] Natori et al., Tetrahedron, 1994, 50: 2771-2783
[2] EP-A-1018548
[3] GaIIi et al., Vaccine, 2003, 21: S2/48-S2/54
[4] Galli et al., J. Exp. Med., 2003, 197: 1051-1057
[5] WO03/009812
[6] Gonzalez-Aseguinolaza et al., J. Exp Med, 2002, 5: 617-624
[7] Huang et al., 2003, Abstract 396, 10th Conference on retroviruses and opportunistic infection
[8] Ko et al., J. Immunol, 2005, 175: 3309-3317
[9] De Libero et al., Nature Reviews Immunology, 2005, 5: 485-496
[10] US 5936076
[11] WO03/105769
[12] Oki et al., J. Clin. Investig., 113: 1631-1640
[13] US2005 0192248
[14] Yang et al., Angew. Chem. Int. Ed, 2004, 43: 3818-3822
[15] WO2005/102049
[16] Goff et al., J. Am. Chem., Soc, 2004, 126: 13602-13603
[17] Ndonye et al., J. Org. Chem., 2005, 70: 10260-10270
[18] Wu et al., PNAS, 2005, 102(5): 1351-1356
[19] Mattner et al., Nature, 2005, 434: 525-529
[20] Kinjo et al., Nature, 2006, 7(9): 978-986
[21] WO2005/033148.
[22] WO03/007985
[23] Wessels et al., (1990) J Clin Invest 86:1428-33.
[24] Wessels et al., (1989) Infect Immun 57:1089-94.
[25] Международная патентная заявка PCT/IB03/01436.
[26] Ravenscroft et al., (1999) Vaccine 17:2802-2816.
[27] Costantino et al., (1999) Vaccine 17:1251-1263.
[28] Anonymous (Jan 2002) Research Disclosure, 453077.
[29] Anderson (1983) Infect Immun 39(l):233-238.
[30] Anderson et al., (1985) J Clin Invest 76(l):52-59.
[31] EP-A-0372501.
[32] EP-A-0378881.
[33] EP-A-0427347.
[34] WO93/17712
[35] WO94/03208.
[36] WO98/58668.
[37] EP-A-0471177.
[38] WO91/01146
[39] Falugi et al., (2001) Eur J Immunol 31:3816-3824.
[40] Baraldo et al., (2004) Infect Immun. 72:4884-7
[41] EP-A-0594610.
[42] WO00/56360.
[43] WO02/091998.
[44] Kuo et al., (1995) Infect Immun 63:2706-13.
[45] WO01/72337
[46] WO00/61761.
[47] WO99/42130.
[48] WO2004/011027.
[49] WO96/40242.
[50] Lees et al., (1996) Vaccine 14:190-198.
[51] WO95/08348.
[52] WO98/42721
[53] Патент США 4882317
[54] Патент США 4695624
[55] Mol. Immunol., 1985, 22, 907-919
[56] EP-A-0208375
[57] WO00/10599
[58] Gever et al., Med. Microbiol. Immunol, 165: 171-288 (1979).
[59] Патент США 4057685.
[60] Патент США 4673574; 4761283; 4808700.
[61] Патент США 4459286.
[62] Патент США 4965338
[63] Патент США 4663160.
[64] Патент США 4761283
[65] Патент США 4356170
[66] Lei et al., (2000) Dev Biol (Basel) 103:259-264.
[67] WO00/38711; Патент США 6146902.
[68] WO2006/027685.
[69] WO99/24578.
[70] WO99/36544.
[71] WO99/57280.
[72] WO00/22430.
[73] Tettelin et al., (2000) Science 287:1809-1815.
[74] WO96/29412.
[75] Pizza et al., (2000) Science 287:1816-1820.
[76] WO02/22167.
[77] Bell (2000) Pediatr Infect Dis J 19:1187-1188.
[78] Iwarson (1995) APMIS 103:321-326.
[79] Gerlich et al., (1990) Vaccine 8 Suppl:S63-68 & 79-80.
[80] Hsu et al., (1999) Clin Liver Dis 3:901-915.
[81] WO03/002065
[82] WO01/37869
[83] WO04/005473
[84] Gustafsson et al., (1996) N. Engl. J. Med. 334:349-355.
[85] Rappuoli et al., (1991) TIBTECH 9:232-238.
[86] Vaccines (2004) eds. Plotkin & Orenstein. ISBN 0-7216-9688-0.
[87] WO02/02606.
[88] Kalman et al., (1999) Nature Genetics 21:385-389.
[89] Read et al., (2000) Nucleic Acids Res 28:1397-406.
[90] Shirai et al., (2000) J. Infect. Dis. 181(Suppl 3):S524-S527.
[91] WO99/27105.
[92] WO00/27994.
[93] WO00/37494.
[94] WO99/28475.
[95] Ross et al., (2001) Vaccine 19:4135-4142.
[96] Sutter et al., (2000) Pediatr Clin North Am 47:287-308.
[97] Zimmerman & Spann (1999) Am Fam Physician 59:113-118, 125-126.
[98] Dreesen (1997) Vaccine 15 Suppl:S2-6.
[99] MMWR Morb Mortal WkIy Rep 1998 Jan 16; 47(1):12, 19.
[100] Covacci & Rappuoli (2000) J. Exp. Med. 19:587-592.
[101] WO93/18150.
[102] Covacci et al., (1993) Proc. Natl Acad. ScL USA 90: 5791-5795.
[103] Tummuru et al., (1994) Infect. Immun. 61:1799-1809.
[104] Marchetti et al., (1998) Vaccine 16:33-37.
[105] Telford et al., (1994) J. Exp. Med. 179:1653-1658.
[106] Evans et al., (1995) Gene 153:123-127.
[107] WO96/01272 & WO96/01273, в частности, SEQ ID NO:6.
[108] WO97/25429.
[109] WO98/04702.
[110] McMichael (2000) Vaccine 19 Suppl l:S101-107.
[111] Schuchat (1999) Lancet 353(9146):51-6.
[112] WO02/34771.
[113] Dale (1999) Infect Dis Clin NorthAm 13:227-43, viii.
[114] Ferretti et al., (2001) PNAS USA 98: 4658-4663.
[115] Kuroda et al., (2001) Lancet 357(9264):1225-1240; see also pages 1218-1219.
[116] Anderson (2000) Vaccine 19 Suppl l:S59-65.
[117] Kahn (2000) Curr Opin Pediatr 12:257-262.
[118] Crowe (1995) Vaccine 13:415-421.
[119] J Toxicol Clin Toxicol (2001) 39:85-100.
[120] Demicheli et al., (1998) Vaccine 16:880-884.
[121] Stepanov et al., (1996) J Biotechnol 44:155-160.
[122] EP-A-0139417.
[123] Harper et al., (2004) Lancet 364(9447): 1757-65.
[124] Charalambous & Feavers (2001) J Med Microbiol 50:937-939.
[125] WO01/52885.
[126] Bjune et al., (1991) Lancet 338(8775):1093-1096.
[127] Fukasawa et al., (1999) Vaccine 17:2951-2958.
[128] Rosenqvist et al., (1998) Dev. Biol. Stand. 92:323-333.
[129] WO2004/032958.
[130] WO96/37624.
[131] WO98/46262.
[132] Huckriede et al., (2003) Methods Enzymol 373:74-91.
[133] Herlocher et al., (2004) J Infect Dis 190(9): 1627-30.
[134] Le et al., (2005) Nature 437(7062):l108.
[135] Hoffmann et al., (2002) Vaccine 20:3165-3170.
[136] Subbarao et al., (2003) Virology 305:192-200.
[137] Liu et al., (2003) Virology 314:580-590.
[138] Ozaki et al., (2004) J. Virol. 78:1851-1857.
[139] Webby et al., (2004) Lancet 363:1099-1103.
[140] WO00/60050.
[141] WO01/04333.
[142] US 6649372.
[143] Neumann et al., (2005) Proc Natl Acad Sci USA 102:16825-9.
[144] WO01/83794.
[145] Hoffmann et al., (2000) Virology; 267(2):310-7.
[146] WO97/37000.
[147] Brands et al., (1999) Dev Biol Stand 98:93-100.
[148] Halperin et al., (2002) Vaccine 20:1240-7.
[149] Tree et al., (2001) Vaccine 19:3444-50.
[150] Kistner et al., (1998) Vaccine 16:960-8.
[151] Kistner et al., (1999) Dev Biol Stand 98:101-110.
[152] Bruhl et al., (2000) Vaccine 19:1149-58.
[153] Pau et al., (2001) Vaccine 19:2716-21.
[154] http://www.atcc.org/
[155] http://locus.umdnj.edu/
[156] WO03/076601.
[157] WO2005/042728.
[158] WO03/043415.
[159] Lundblad (2001) Biotechnology and Applied Biochemistry 34:195-197.
[160] Guidance for Industry: Bioanalytical Method Validation. U.S. Department of Health and Human Services Food and Drug Administration Center for Drug Evaluation and Research (CDER) Center for Veterinary Medicine (CVM). May 2001.
[161] Ji et al., (2002) Biotechniques. 32:1162-7.
[162] Briggs (1991) J Parenter Sci Technol. 45:7-12.
[163] Lahijani et al., (1998) Hum Gene Ther. 9:1173-80.
[164] Lokteff et al., (2001) Biologicals. 29:123-32.
[165] EP-B-0870508.
[166] US 5948410.
[167] WO97/37001.
[168] WO01/22992.
[169] Hehme et al., (2004) Virus Res. 103(l-2):163-71.
[170] Treanor et al., (1996) J Infect Dis 173:1467-70.
[171] Keitel et al., (1996) Clin Diagn Lab Immunol 3:507-10.
[172] WO00/56365.
[173] Gennaro (2000) Remington: The Science and Practice of Pharmacy. 20th ed ISBN: 0683306472
[174] Патент США 6355271.
[175] WO0023105.
[176] WO0122972.
[177] Kandimalla et al., (2003) Nucleic Acids Research 31:2393-2400.
[178] WO02/26757.
[179] WO99/62923.
[180] Krieg (2003) Nature Medicine 9:831-835.
[181] McCluskie et al., (2002) FEMS Immunology and Medical Microbiology 32:179-185.
[182] WO98/40100.
[183] Патент США 6207646.
[184] Патент США 6239116.
[185] Патент США 6429199.
[186] Kandimalla et al., (2003) Biochemical Society Transactions 31 (part 3):654-658.
[187] Blackwell et al., (2003) J Immunol 170:4061-4068.
[188] Krieg (2002) Trends Immunol 23:64-65.
[189] WO01/95935.
[190] Kandimalla et al., (2003) BBRC 306:948-953.
[191] Bhagat et al., (2003) BBRC 300:853-861.
[192] WO03/035836.
[193] Myers et al., (1990) pages 145-156 of Cellular and molecular aspects of endotoxin reactions.
[194] Ulrich (2000) Chapter 16 (pages 273-282) of reference 250.
[195] Johnson et al., (1999) J Med Chem 42:4640-9.
[196] Baldrick et al., (2002) Regulatory Toxicol Pharmacol 35:398-413.
[197] UK patent application GB-A-2220211.
[198] US 4680338.
[199] US 4988815.
[200] WO92/15582.
[201] Stanley (2002) Clin Exp Dermatol 27:571-577.
[202] Wu et al., (2004) Antiviral Res. 64(2):79-83.
[203] Vasilakos et al., (2000) Cell Immunol. 204(1):64-74.
[204] Патенты США 4689338, 4929624, 5238944, 5266575, 5268376, 5346905, 5352784, 5389640, 5395937, 5482936, 5494916, 5525612, 6083505, 6440992, 6627640, 6656938, 6660735, 6660747, 6664260, 6664264, 6664265, 6667312, 6670372, 6677347, 6677348, 6677349, 6683088, 6703402, 6743920, 6800624, 6809203, 6888000 и 6924293.
[205] Jones (2003) Curr Opin Investig Drugs 4:214-218.
[206] WO2004/060308.
[207] WO2004/064759.
[208] US 6924271.
[209] US 2005/0070556.
[210] US 5658731.
[211] Патент США 5011828.
[212] WO2004/87153.
[213] US 6605617.
[214] WO02/18383.
[215] WO2004/018455.
[216] WO03/082272.
[217] WO2006/002422.
[218] Johnson et al., (1999) Bioorg Med Chem Lett 9:2273-2278.
[219] Evans et al., (2003) Expert Rev Vaccines 2:219-229.
[220] Andrianov et al., (1998) Biomaterials 19:109-115.
[221] Payne et al., (1998) Adv Drug Delivery Review 31:185-196.
[222] US 5057540.
[223] WO96/33739.
[224] EP-A-0109942.
[225] WO96/11711.
[226] WO00/07621.
[227] Barr et al., (1998) Advanced Drug Delivery Reviews 32:247-271.
[228] Sjolanderet et al., (1998) Advanced Drug Delivery Reviews 32:321-338.
[229] Pizza et al., (2000) Int J Med Microbiol 290:455-461.
[230] WO95/17211.
[231] WO98/42375.
[232] Singh et al., (2001) J Cont Release 70:267-276.
[233] WO99/27960.
[234] US 6090406
[235] US 5916588
[236] EP-A-0626169.
[237] WO99/52549.
[238] WO01/21207.
[239] WO01/21152.
[240] Signorelli & Hadden (2003) Int Immunopharmacol 3(8): 1177-86.
[241] WO2004/064715.
[242] Cooper (1995) Pharm Biotechnol 6:559-80.
[243] WO03/011223.
[244] Meraldi et al., (2003) Vaccine 21:2485-2491.
[245] Pajak et al., (2003) Vaccine 21:836-842.
[246] US-6586409.
[247] Wong et al., (2003) J Clin Pharmacol 43(7):735-42.
[248] US2005/0215517.
[249] Vaccine Design: The Subunit and Adjuvant Approach (eds. Powell & Newman) Plenum Press 1995 (ISBN 0-306-44867-X).
[250] Vaccine Adjuvants: Preparation Methods and Research Protocols (Volume 42 of Methods in Molecular Medicine series). ISBN: 1-59259-083-7. Ed. O'Ηagan.
[251] WO98/20734.
[252] WO01/30390
[253] Galli et al., PNAS, 2007, 104(1): 3984-3989
[254] ClaassenE et al., Res Immunol, 1992, 143(5):478-83
[255] Vollmer J et al., Eur J Immunol, 2004, 34(l):251-62
[256] Lindblad, E.B., Immunol. Cell Biol, 2004, 82:497-505
[257] Podda A, et al., Expert Rev Vaccines, 2003 2(2): 197-203
[258] Bernasconi et al., Science, 2002, 298(5601):2199-202
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАКЦИНА НА ОСНОВЕ В-КЛЕТОК, НАГРУЖЕННЫХ ЛИГАНДОМ Т-КЛЕТОК-ПРИРОДНЫХ КИЛЛЕРОВ И АНТИГЕНОМ | 2006 |
|
RU2420311C2 |
| УЛУЧШЕННАЯ ВАКЦИНАЦИЯ ПРОТИВ ГРИППА | 2013 |
|
RU2599496C2 |
| ВАКЦИНАЦИЯ МЕНИНГОКОККОВЫМИ КОНЪЮГАТАМИ | 2005 |
|
RU2379052C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2005 |
|
RU2351363C2 |
| ВАКЦИНАЦИЯ МЕНИНГОКОККОВЫМИ КОНЪЮГАТАМИ | 2006 |
|
RU2429011C2 |
| РЕКОМБИНАНТНАЯ ТРЕХВАЛЕНТНАЯ ВАКЦИНА ОТ ГРИППА ЧЕЛОВЕКА | 2012 |
|
RU2485973C1 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ И СПОСОБЫ | 2008 |
|
RU2468034C2 |
| Экспрессионный вектор на основе аденовируса человека 5 серотипа индуцирующий кросс-протективный иммунитет к вирусам гриппа А субтипа H3 и фармацевтическая композиция на его основе. | 2023 |
|
RU2814189C1 |
| РЕКОМБИНАНТНАЯ ПРОТИВОГРИППОЗНАЯ ВАКЦИНА С ШИРОКИМ СПЕКТРОМ ЗАЩИТЫ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2017 |
|
RU2757013C2 |
| РЕКОМБИНАНТНЫЕ ВИРУСЫ ГРИППА А | 2001 |
|
RU2280690C2 |
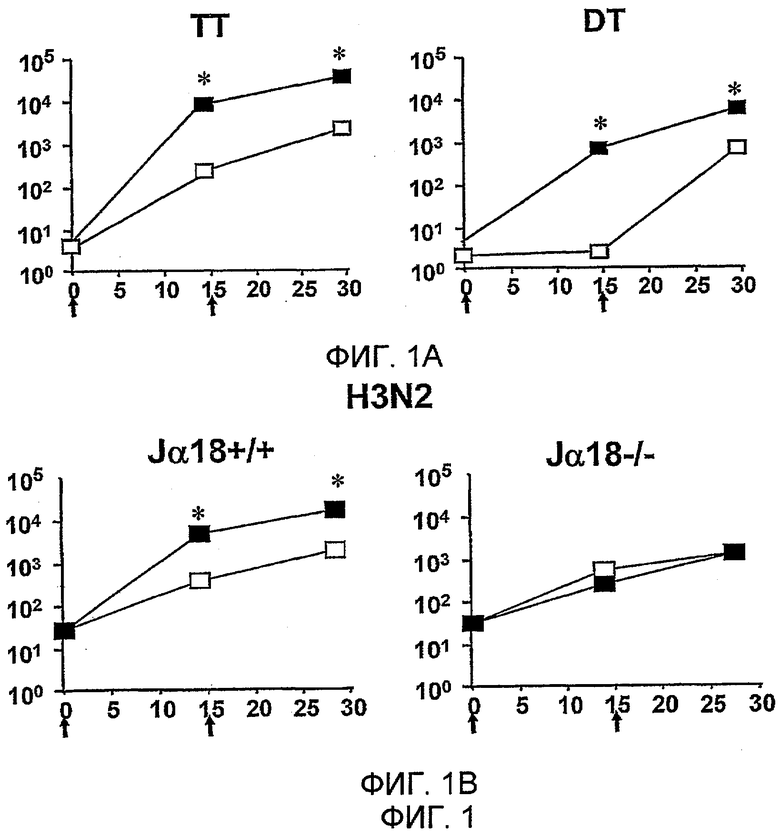
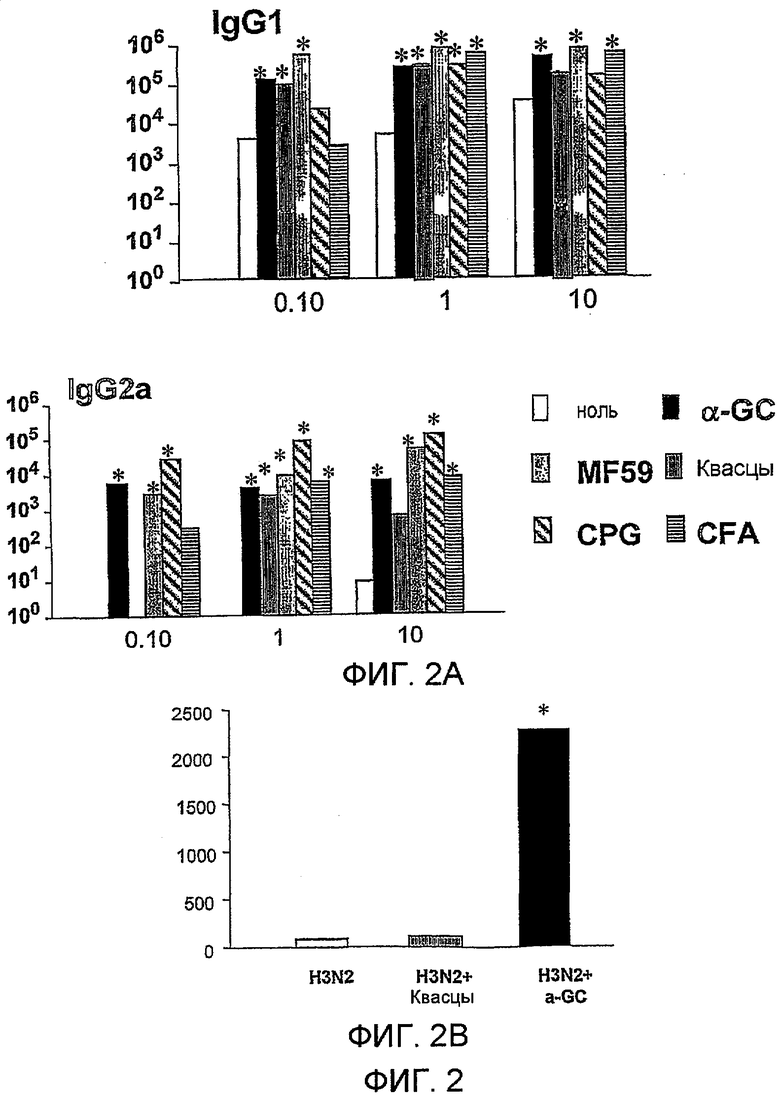
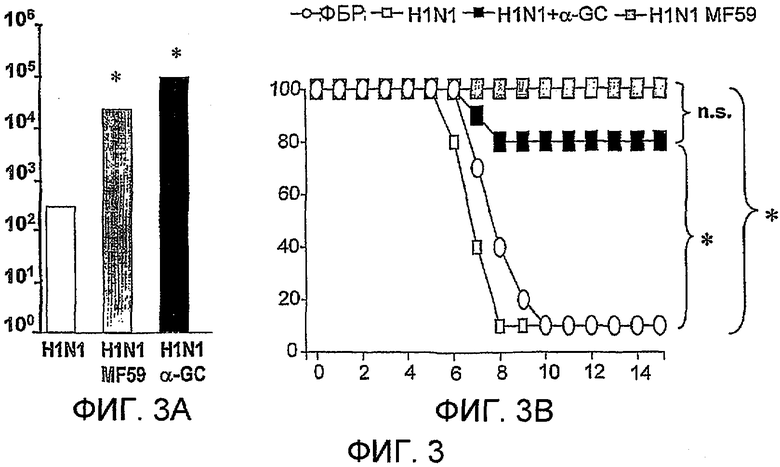
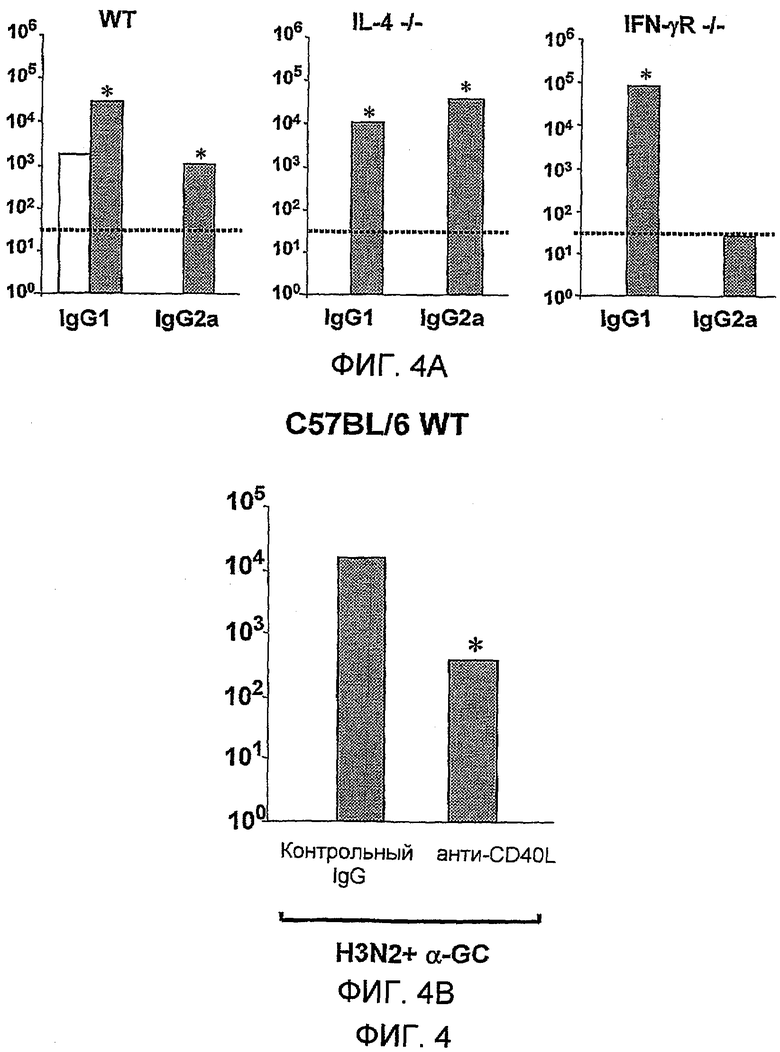
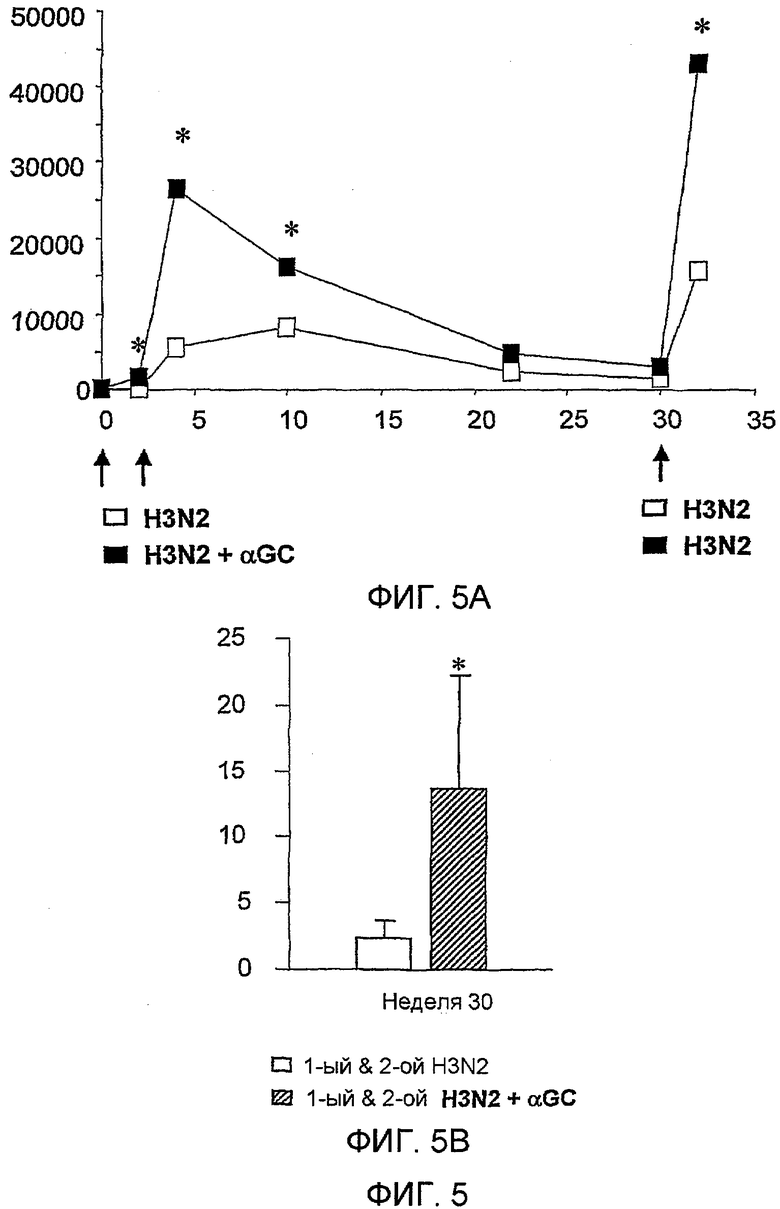
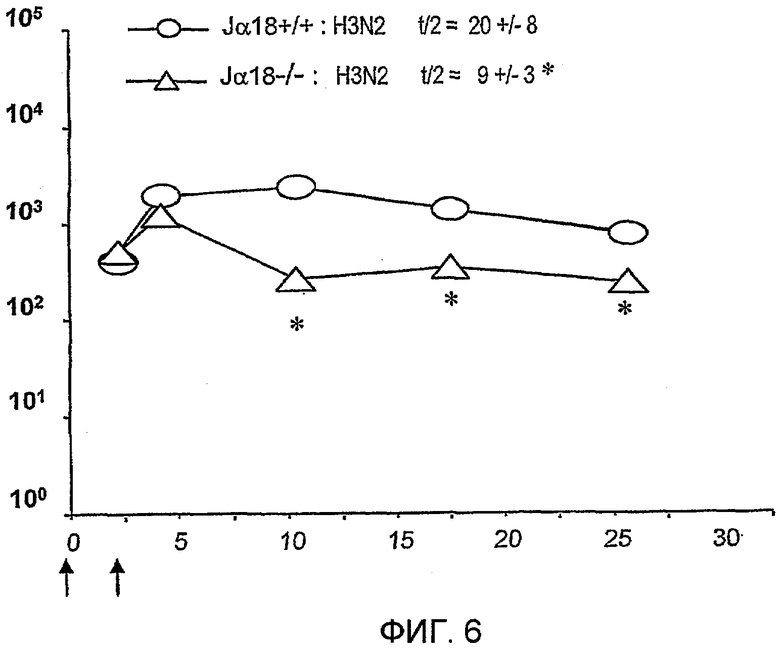
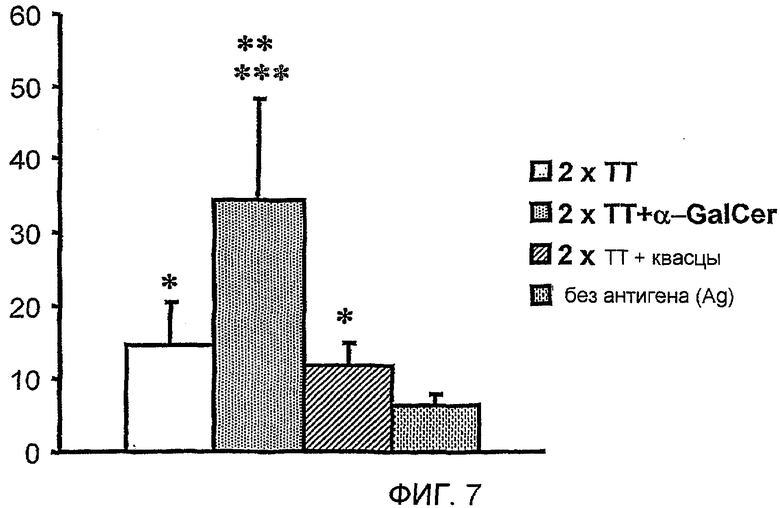
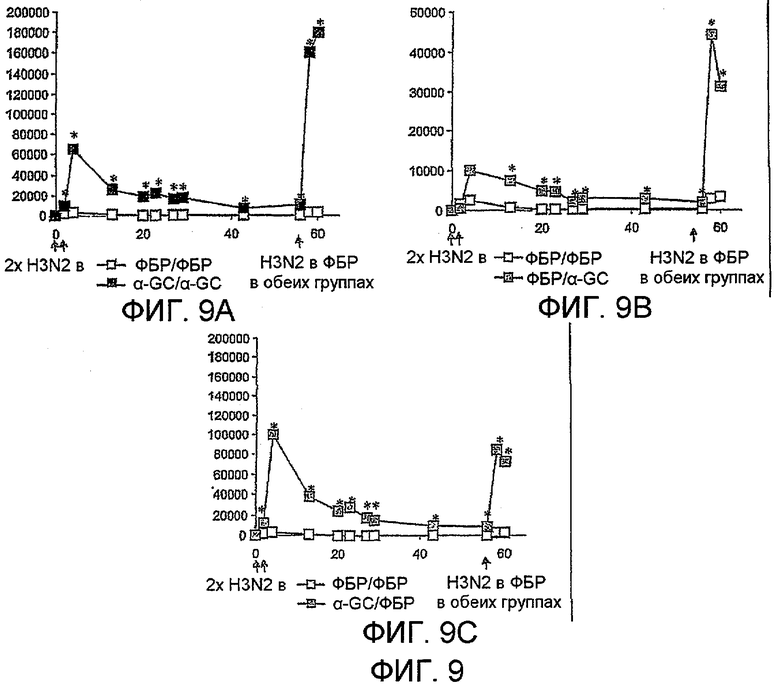
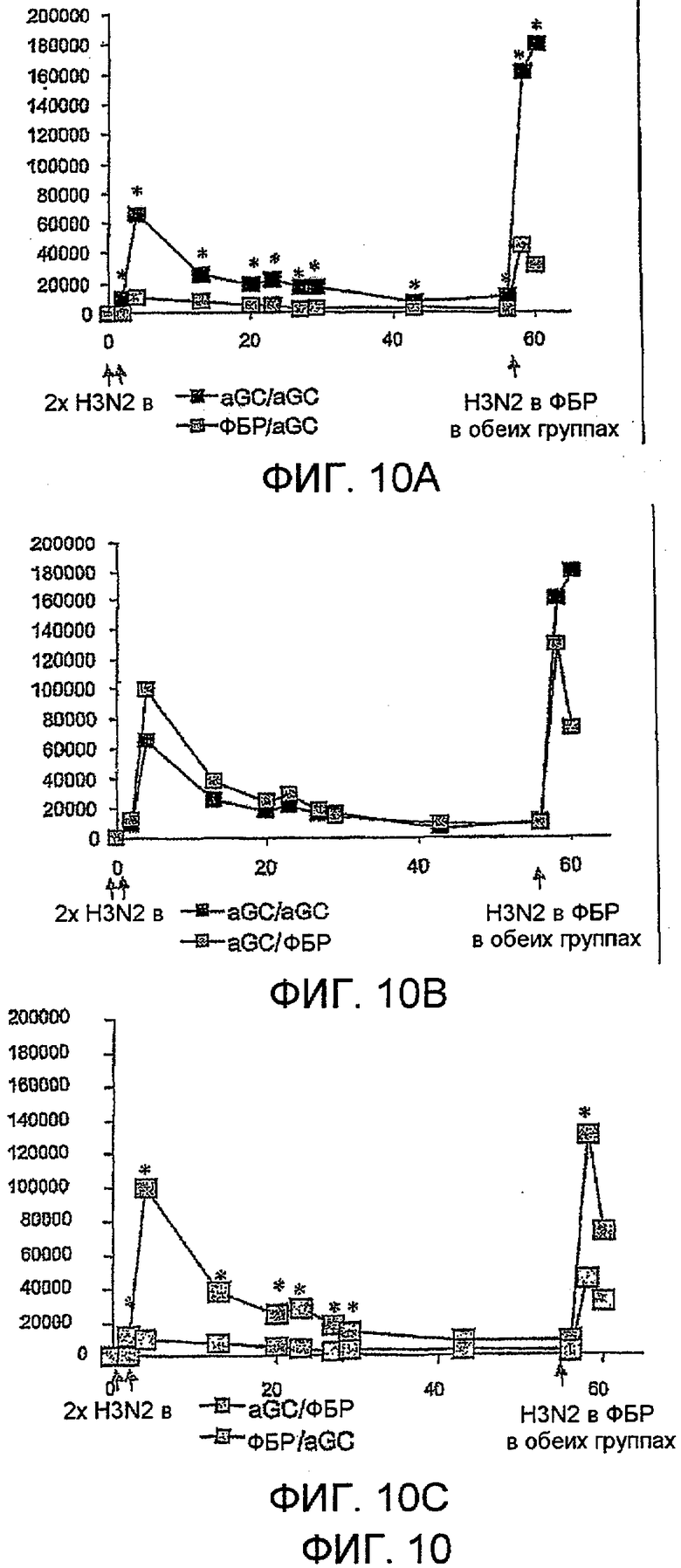
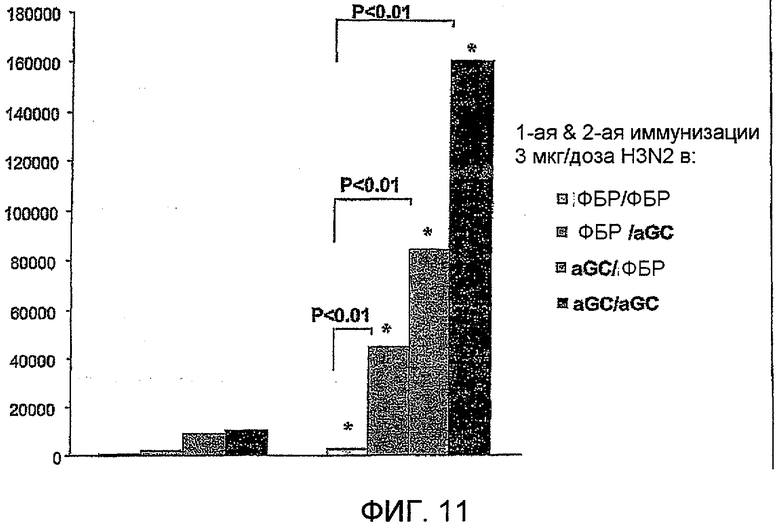
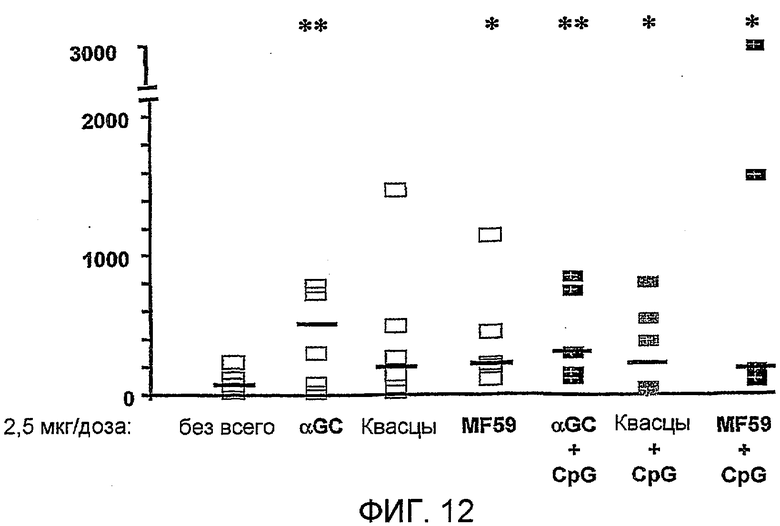
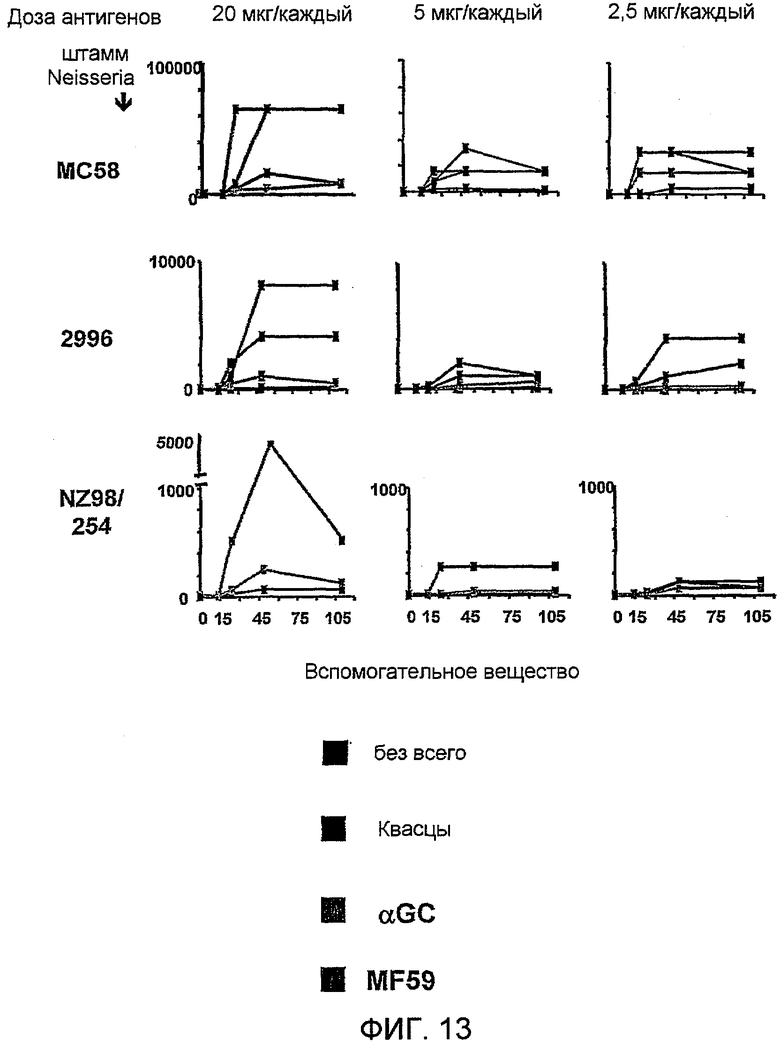
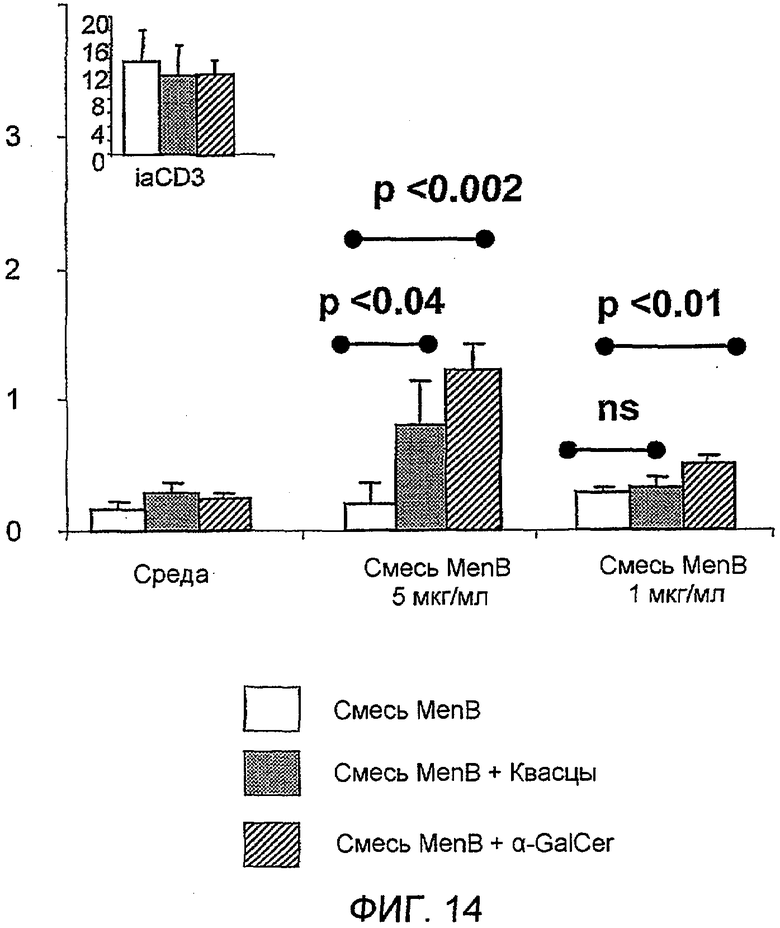

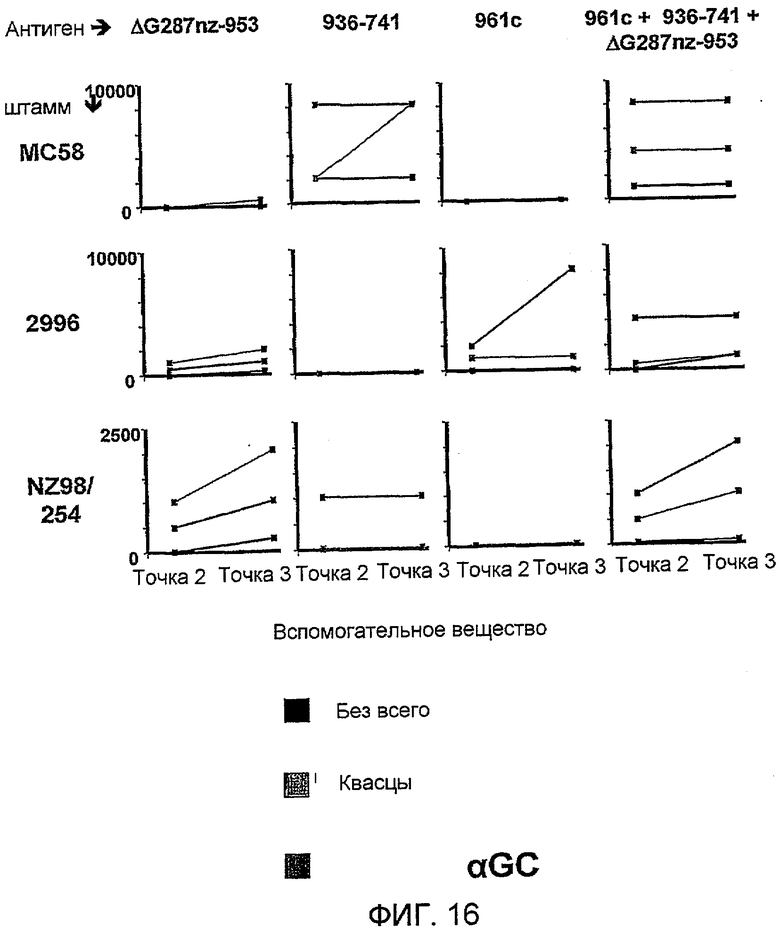
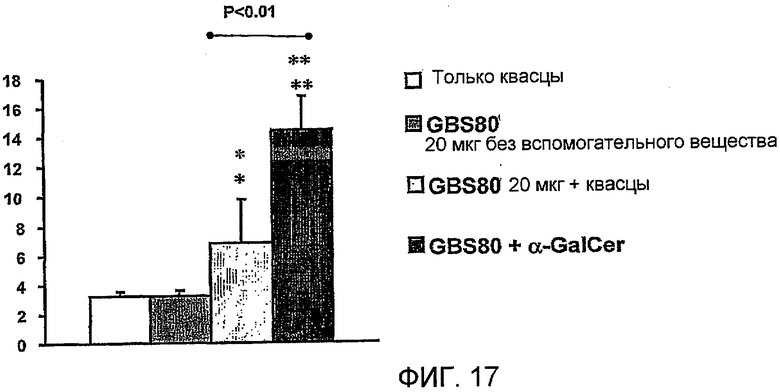
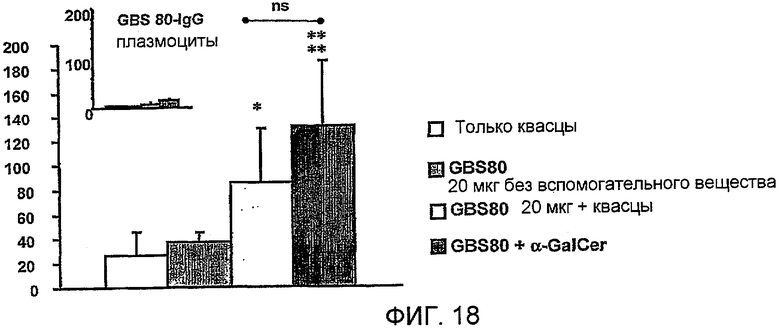
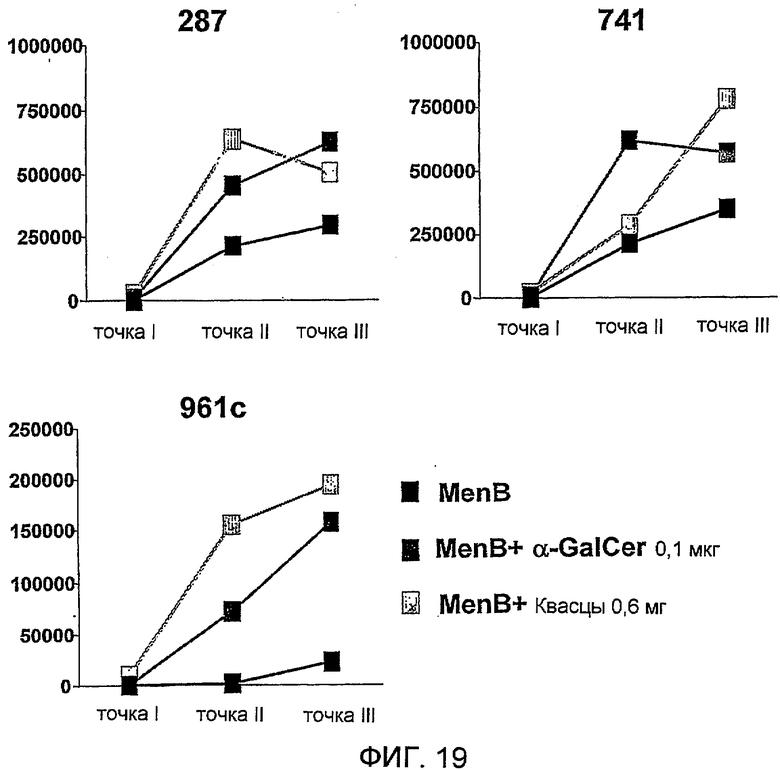
Группа изобретений относится к иммуногенным композициям, содержащим α-галактозилцерамид и белковый антиген стрептококка группы B. Группа изобретений также относится к применению α-галактозилцерамида и белкового антигена стрептококка группы B для индуцирования иммунного ответа. Применение заявленной группы изобретений приводит к вызыванию долговременной иммунологической памяти в отсутствие повторных доз антигена и/или в отсутствие множественных возбуждающих доз. 12 н. и 15 з.п. ф-лы, 19 ил., 2 табл., 4 пр.
1. Иммуногенная композиция, содержащая: а) α-галактозилцерамид и b) белковый антиген из стрептококка группы В.
2. Способ индуцирования иммунного ответа у пациента, включающий введение пациенту α-галактозилцерамида и белкового антигена из стрептококка группы В.
3. Способ по п.2, где антиген и α-галактозилцерамид вводят одновременно, последовательно или раздельно.
4. Применение белкового антигена из стрептококка группы В для индуцирования иммунного ответа у пациента, где указанный антиген вводят с α-галактозилцерамидом.
5. Применение α-галактозилцерамида для индуцирования иммунного ответа у пациента, в котором указанный α-галактозилцерамид вводят с белковым антигеном из стрептококка группы В.
6. Применение а) белкового антигена из стрептококка группы B и b) α-галактозилцерамида для индуцирования иммунного ответа у пациента.
7. Применение белкового антигена из стрептококка группы В для индуцирования иммунного ответа у пациента, где пациент был предварительно обработан α-галактозилцерамидом.
8. Применение по любому из пп.4-7, где количество α-галактозилцерамида, вводимое указанному пациенту, меньше 10 мкг/кг массы тела пациента.
9. Набор для индуцирования иммунного ответа у пациента, содержащий: (а) белковый антиген из стрептококка группы B и (b) α-галактозилцерамид.
10. Способ индуцирования иммунного ответа на белковый антиген из стрептококка группы B у пациента, который в этом нуждается, включающий введение указанному пациенту композиции, содержащей:
a) белковый антиген из стрептококка группы B и
b) α-галактозилцерамид,
таким образом, что число и/или частота доз указанной композиции, необходимые для того, чтобы указанный пациент был способен на иммунный ответ при последующем подвергании указанному антигену, понижены по сравнению с введением указанного антигена в отсутствие α-галактозилцерамида.
11. Способ по п.10, где число и/или частота доз указанной композиции, необходимые для того, чтобы указанный пациент был способен на защитный иммунный ответ при последующем подвергании указанному антигену, понижены по сравнению с введением указанного антигена в отсутствие α-галактозилцерамида.
12. Способ по п.11, в котором число доз указанной композиции, необходимое для того, чтобы указанный пациент был способен на защитный иммунный ответ при последующем подвергании указанному антигену, понижено по сравнению с введением указанного антигена в отсутствие α-галактозилцерамида.
13. Способ по п.12, где число доз, требуемое для индуцирования защитного иммунного ответа, понижено до однократной возбуждающей дозы.
14. Способ по п.11, где частота повторных доз указанной композиции, необходимая, чтобы указанный пациент был способен на защитный иммунный ответ при последующем подвергании указанному антигену, понижена по сравнению с введением указанного антигена в отсутствие α-галактозилцерамида.
15. Способ по п.14, где повторные дозы вводят с интервалами более одного года.
16. Способ по п.15, где необходимость в повторных дозах полностью устранена.
17. Способ вызывания иммунного ответа на белковый антиген из стрептококка группы B у пациента, включающий введение указанному пациенту:
a) белкового антигена из стрептококка группы B и
b) α-галактозилцерамида,
где указанные антиген и α-галактозилцерамид также вводили указанному пациенту предварительно более чем за один год.
18. Применение белкового антигена из стрептококка группы В и α-галактозилцерамида для вызывания иммунного ответа у пациента, где указанные антиген и α-галактозилцерамид также вводили указанному пациенту предварительно более чем за 1 год.
19. Применение по п.18, где иммунный ответ представляет собой защитный иммунный ответ.
20. Применение по п.18 или 19, где антиген и α-галактозилцерамид вводят одновременно, последовательно или раздельно.
21. Применение по п.18 или 19, где количество α-галактозилцерамида, вводимое указанному пациенту, меньше 10 мкг/кг массы тела пациента.
22. Способ вызывания иммунного ответа на белковый антиген из стрептококка группы B у пациента, включающий введение указанному пациенту:
a) белкового антигена из стрептококка группы B и
b) α-галактозилцерамида,
где количество α-галактозилцерамида, включенное в композицию, меньше 10 мкг/кг массы тела пациента.
23. Применение белкового антигена из стрептококка группы В и α-галактозилцерамида для вызывания иммунного ответа у пациента, где количество α-галактозилцерамида меньше 10 мкг/кг массы тела пациента.
24. Применение по п.23, где иммунный ответ представляет собой защитный иммунный ответ.
25. Применение по п.24, где α-галактозилцерамид и антиген вводят одновременно, последовательно или раздельно.
26. Применение по п.23, где α-галактозилцерамид активирует инвариантные NKT-клетки.
27. Применение по п.23, где α-галактозилцерамид увеличивает уровни IFN-γ, IL-4 и IL-13, секретируемых инвариантными NKT-клетками, по сравнению с уровнями IFN-γ, IL-4 и IL-13, секретируемых инвариантными NKT-клетками, в отсутствие α-галактозилцерамида.
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| US2005192248 A1, 01.09.2005 | |||
| WO2005000348 А2, 01.06.2005 | |||
| WO2006026389 A2, 09.03.2006 | |||
| KO S-Y ET AL | |||
| Alpha-galactosylceramide can act as a nasal vaccine adjuvant inducing protective immune responses against viral infection and tumor | |||
| JOURNAL OF IMMUNOLOGY, WILLIAMS & WILKINS CO, US, vol | |||
| Ручной прибор для загибания кромок листового металла | 1921 |
|
SU175A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |

