По данной заявке запрашивается приоритет в соответствии с 35 U.C.S.§119(е) согласно предварительной заявке США No. 60/928,884, поданной 11 мая 2007 года, и предварительной заявке No. 61/005,463, поданной 5 декабря 2007 года. Обе заявки включены в данное описание посредством ссылки.
СУТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к композициям и методам доставки терапевтических веществ в организме, в частности в центральную нервную систему (ЦНС).
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Гематоэнцефалический барьер является одним из наименее проницаемых в биологических системах. Множество факторов обеспечивают его непроницаемость. С помощью электронной микроскопии было показано, что плотные соединения между эндотелиальными клетками кровеносных сосудов мозга, а также другие особенности клеток эндотелия (ограниченный пиноцитоз, надостаток межклеточного пространства) предотвращают образование плазматического ультрафильтрата. Ферментативная активность в гематоэнцефалическом барьере также ограничивает проникновение некоторых веществ, в особенности моноаминов и небольших пептидов (Baranczyk-Kuzma and Audus (1987) J. Cereb. Blood Flow Metab., 7:801-805; Hardebo and Owman (1990) Pathophysiology of the BBB, pp.41-55 (Johansson et al., Eds.) Elsevier, Amsterdam; Miller et al. (1994) J. Cell. Physiol., 161:333-341; Brownson et al. (1994) J. Pharmacol. Exp. Ther., 270:675-680; Brownlees and Williams (1993) J. Neurochem., 60:793-803). Более того, система выброса веществ из мозга в кровь, частью которой является Р-гликопротеин, в свою очередь также препятствует накоплению низкомолекулярных соединений и жирорастворимых веществ (Taylor, E.M. (2002) Clin. Pharmacokinet., 41:81-92; Schinkel et al. (1996) J. Clin. Invest., 97:2517-2524). Набор второстепенных факторов, таких как связывание белков рецепторами, ферментативная деградация, клиренс и сиквестрация тканями, также ограничивает способность веществ проникать через гематоэнцефалический барьер, благодаря ограничению их представления; эти факторы играют особенно важную роль в распределении экзогенно введенных веществ (Banks and Kastin (1993) Proceedings of the International Symposium on Blood Binding and Drug Transfer, pp.223-242 (Tillement et al., Eds.) Fort and Clair, Paris).
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В соответствии с настоящим изобретением предложены методы лечения нейродегенеративных заболеваний. Данные методы подразумевают введение терапевтически эффективной дозы композиции, содержащей а) по меньшей мере один комплекс терапевтического полипептида и синтетического полимера, содержащего по меньшей мере один заряд, противоположный заряду терапевтического полипептида, и б) по меньшей мере один фармацевтически допустимый носитель. В конкретном варианте осуществления настоящего изобретения, синтетический полимер содержит по меньшей мере один неионный сегмент и по меньшей мере один полиионный сегмент. В другом варианте осуществления настоящего изобретения вводимый комплекс может пересекать гематоэнцефалический барьер.
В другом аспекте данного изобретения методы лечения нейродегенеративных заболеваний у пациентов включают введение терапевтических доз композиции, содержащей изолированные клетки, содержащие по крайней мере один комплекс, содержащий терапевтический полипептид и синтетический полимер, содержащий по меньшей мере один заряд, противоположный заряду терапевтического полипептида, а также по крайней мере один фармацевтически допустимый носитель. В конкретном варианте осуществления настоящего изобретения синтетический полимер содержит по меньшей мере один неионный сегмент и по меньшей мере один полиионный сегмент. В другом варианте осуществления настоящего изобретения клетки после введения смогут пройти через гематоэнцефалический барьер. Клетки могут быть взяты у пациента, подвергающегося лечению. В конкретном варианте осуществления настоящего изобретения клетки являются иммунными клетками, такими как моноциты, макрофаги, моноциты костного мозга, дендритные клетки, лимфоциты, Т-клетки, нейтрофилы, эозинофилы или базофилы.
В соответствии с еще одним аспектом настоящего изобретения, представленные выделенные клетки содержат по крайней мере один комплекс, содержащий по меньшей мере один интересующий белок и синтетический полимер, содержащий по меньшей мере один заряд, противоположный заряду интересующего белка. Композиции, содержащие клетки, также представлены.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Рисунок 1А - схематичное представление структуры полипептид-полиионного комплекса (может также называться нанозимом). Рисунок 1Б - гель-ретардация фермент-полиионного комплекса при различных значениях Z. Образцы подвергались электрофорезу в ПААГ (7.5%) в неденатурирующих условиях (без ДСН). Дорожка 1: свободный фермент; дорожки 2-4: комплексы фермента с ПЭИ-ПЭГ при прогрессивно увеличивающемся Z (0.5, 2,4). Рисунки 1В-Д - кривые зависимости изменения среднего диаметра (1В-Д) и зета-потенциала комплексов каталазы с полиионами при различных условиях: Рисунок 1В: Z в растворе ФСБ; Рисунок 1Г: ионная сила (Z=1, рН 7.4); Рисунок 1Д: рН (Z=1, [NaCl]=0.15 М). Рисунок 1Е - электронная микрофотография комплекса каталазы с полиионом. Шкала 100 нм. Рисунок 1Ж - зависимость ферментативной активности каталазы в комплексе с полиионом (Z=1). Активность каталазы при различных значениях Z измерялась по скорости распада перекиси водорода. Данные представлены как средние значения ±СОС (n=4). Статистическая достоверность различий значений ферментативной активности комплекса каталазы с полиионом по сравнению с активностью каталазы обозначена звездочкой: (*) р<0,05. Ферментативная активность каталазы не изменялась в широком диапазоне концентрации блок сополимера, значительно уменьшаясь только при Z=50.
Рисунок 2А - ПААГ электрофорез комплексов чБХЭ/ПЛЛ-г-ПЭО(2). Номера дорожек соответствуют номерам образцов в таблице 1. Рисунок 2Б - ПААГ электрофорез свободной лБХЭ и комплексов лБХЭ/ПЭИ-г-ПЭО(2) различного состава. Номера дорожек соответствуют номерам образцов в таблице 2.
Рисунок 3 - зависимость диаметра частиц, образованных в смесях лБХЭ/ПЭИ-г-ПЭО(2) и (■) чБХЭ/ПЭИ-г-ПЭО(2) при различных значениях Z+/-. Концентрация БХЭ 0,15 мг/мл, 23°С в 10 мМ фосфатном буфере, рН 7,4.
Рисунок 4А - ПААГ электрофорез (А) свободной лБХЭ и (Б) комплекса лБХЭ/ПЭИ-г-ПЭО (7) (Z+/-=10,3) при различных разбавлениях. Исходная концентрация лБХЭ 0,167 мг/мл. Рисунок 4Б - ПААГ электрофорез (А) свободной чБХЭ; (Б) несшитого комплекса чБХЭ/ПЭИ-г-ПЭО (2) (Z+/-=1.2); и (В) сшитого комплекса чБХЭ/ПЭИ-г-ПЭО(2) (Z+/-=1.2; степень сшивания 85%) при разных разбавлениях (1:1000, 1:5000, и 1:250). Начальная концентрация чБХЭ 0,15 мг/мл.
Рисунки 5А-В - ПААГ электрофорез свободной чБХЭ (дорожка А); несшитого комплекса чБХЭ/ПЭИ-г-ПЭО (2) (дорожка Б) (Z+/-=1.2); и сшитого комплекса чБХЭ/ПЭИ-г-ПЭО (2) (дорожка В) (Z+/-=1.2) при различных разбавлениях: 1000, 500 и 250. Степень сшивания 85%, 40% и 20% на рисунках 5А, 5Б и 5В соответственно. Начальная концентрация чБХЭ 0,15 мг/мл.
Рисунок 6 - ПААГ электрофорез сшитых комплексов чБХЭ/ПЭИ-г-ПЭО(2) (Z+/-=1.2) с разной степенью сшивания при разбавлении в 500 раз. Конечная концентрация чБХЭ 0,15 мг/мл.
Рисунок 7 - IVIS изображения мышей, которым внутривенно ввели комплекс CuZnСОД с полиионом. Флуоресценция Alexa 680 регистрировалась системой IVIS 200 в различные моменты времени после внутривенной инъекции (в хвостовую вену) комплекса CuZnСОД с полиионом, меченным Alexa 680.
Рисунки 8А и 8Б - ПААГ электрофорез комплексов чБХЭ/ПЭИ-г-ПЭО(2) и лБХЭ/ПЭИ-г-ПЭО(2) соответственно. Номера дорожек соответствуют номерам образцов в таблице 9. Концентрация чБХЭ и лБХЭ 0,15 мг/мл.
Рисунки 9А и 9Б - ПААГ электрофорез сшитых комплексов чБХЭ/ПЭИ-г-ПЭО(2) и лБХЭ/ПЭИ-г-ПЭО(2) при Z+/-=1.0 или Z+/-=2.0 соответственно. Степень сшивания 40%. Дорожка А - свободная чБХЭ; дорожка Б - несшитый комплекс чБХЭ/ПЭИ-г-ПЭО; дорожка В - сшитый комплекс чБХЭ/ПЭИ-г-ПЭО; дорожка Г - свободная чБХЭ; дорожка Д - несшитый комплекс лБХЭ/ПЭИ-г-ПЭО; и дорожка Е - сшитый комплекс лБХЭ/ПЭИ-г-ПЭО. Конечная концентрация БХЭ 0,0003 мг/мл.
Рисунок 10 - цитотоксичность полипептид-полиионного комплекса (Z=1) или соответствующих концентраций ПЭИ-ПЭГ в МККМ. Клетки были инкубированы в течение 24 часов с различными концентрациями полипептид-полиионного комплекса или блок сополимера, отмыты и затем инкубированы 48 часов при 37°С. Выживаемость клеток была определена с помощью сульфородмаина-Б. Абсорбция была измерена при 490 нм, используя Microkinetics ридер ВТ2000. Полученные значения были выражены как процентное отношение контрольных значений абсорбции клеток, к которым полипептид-полиионный комплекс не был добавлен. Все измерения были повторены 8 раз. Цитотоксичный эффект свободной каталазы или полиионного комплекса каталазы с ПЭИ-ПЭГ в МККМ не был показан.
Рисунок 11А - кинетика аккумуляции свободной каталазы и каталазы в комплексе с полиионом (Z=1) в моноцитах. Клетки были инкубированы с Alexa 594-меченной каталазой или фермент-полиионным комплексом в течение различного времени. Затем клетки были лизированы, и флуоресценция клеточных лизатов была измерена (λвозб=580 нм, λисп=617 нм). Данные представлены как средние значения ± СОС (n=4). Рисунок 11Б - аккумуляция полиионного комплекса каталазы в МККМ при различных значениях Z. Рисунок 11В - внутриклеточная локализация полиионного комплекса РИТЦ-меченой каталазы в МККМ. Клетки растили на предметных стеклах, после чего инкубировали с комплексом каталазы с ПЭИ-ПЭГ (Z=1) в течение 24 часов. После инкубации клетки зафиксировали и окрасили F-актинспецифичным фаллоидином, меченным Орегон Зеленым 488. Конфокальные микрофотографии были получены с помощью системы ACAS-570.
Рисунок 12А - профиль выделения комплекса каталазы с полиионом из МККМ. Клетки были нагружены комплексом каталазы с ПЭИ-ПЭГ (Z=1) в течение одного часа, отмыты ФСБ и инкубированы в среде без каталазы на протяжении различного времени. Количество выделенной каталазы, а также удержанной клетками было определено с помощью флуоресцентной спектрофотометрии. Данные представлены как средние значения ±СОС (n=4). Рисунок 12Б - профиль инициированного выделения каталазы из МККМ в среду. Зрелые МККМ были нагружены сначала Alexa 594 меченным комплексом каталазы с полиионом (Z=1) в течение одного часа, отмыты ФСБ и затем инкубированы в среде без каталазы, содержащей или несодержащей 10 µМ форбол миристат ацетата (ФМА) в течение различного времени. Количество выделенной в среду каталазы было определено с помощью флуоресцентной спектрофотометрии. Данные представлены как средние значения ±СОС (n=4). Добавление ФМА в среду привело к увеличению выделения фермента на 50%.
Рисунки 13А и 13Б показывают сохранение ферментативной активности каталазы внутри МККМ. На рисунке 13А МККМ были нагружены либо свободной каталазой, либо полиионным комплексом каталазы (Z=1), затем клетки отмыли и инкубировали в среде без каталазы в течение различного времени. Активность каталазы, выделенной из МККМ, была определена спектрофотометрически. На рисунке 13Б полиионные комплексы каталазы различного состава (Z) были загружены в клетки, которые затем были инкубированы в среде без каталазы в течение 2 часов. Активность каталазы определяли в собранной среде спектрофотометрически. Данные представлены как средние значения ±СОС (n=4). Статистическая достоверность различий значений ферментативной активности полиионного комплекса каталазы по сравнению с активностью свободной каталазы обозначена звездочкой: (*) р<0,05.
Рисунок 14А - схема модуляции активных форм кислорода полиионным комплексом каталазы, выделенным из МККМ. МККМ были нагружены блок сополимером (2 мг/мл; рисунок 14В), свободной каталазой или полиионным комплексом каталазы (Z=1) (Рисунки 14Б и 14Г). Затем клетки были отмыты и инкубированы 2 часа в буфере Рингера-Кребса. В то же время мышиные микроглиальные клетки были стимулированы добавлением либо 200 нг/мл ФНО-α (48 часов), либо 0,5 µг/мл Н-α-синуклеина (Рисунок 14Г). Затем к супернатантам, содержащим выделенный фермент, собранным от МККМ, были добавлены растворы Amplex Red и пероксидазы корня хрена, после чего они были добавлены к активированным клеткам микроглии. В качестве контроля использовались клетки микроглии, инкубированные в свежей среде (Рисунок 14Б). Количество пероксида водорода, выделенного клетками микроглии и расщепленного каталазой, было определено спектрофотометрически.
Данные представлены как средние значения ±СОС (n=6). Статистическая достоверность различий количеств расщепленной перекиси водорода выделенным полиионным комплексом каталазы или свободной каталазой по сравнению с активированной микроглией (контроль) обозначена звездочкой: (*) р<0,05, (**) р<0.005.
Рисунок 15 - биораспределение 125I-меченого полиионного комплекса каталазы в мышах, подвергшихся лечению МФТП. Мышам ввели МККМ (106 клеток/мышь), нагруженных полиионным комплексом каталазы (Z=1, 50 µCi/мышь), или только полиионный комплекс (контрольная группа). Спустя 24 часа мыши были умерщвлены, и радиоактивность разных органов была измерена. Данные представлены как средние значения ±СОС (n=4). Статистическая достоверность различий транспорта МККМ, нагруженных полиионным комплексом каталазы по сравнению со свободным комплексом обозначена двумя звездочками: (**) р<0,005.
Рисунок 16 - биораспределение Alexa 680 меченого полипептид-полиионного комплекса, нагруженного в МККМ. Клетки внутривенно ввели мышам, подвергшимся действию МФТП.
Рисунок 17 демонстрирует нейрозащитный эффект МККМ, содержащих полипептид-полиионный комплекс, нагруженный каталазой, против потери дофаминэргических нейронов, вызванной действием МФТП. Уровень N-ацетил-аспартата был значительно понижен в контрольной группе мышей и немного увеличен у мышей, подвергшихся лечению МККМ, нагруженных полиионным комплексом каталазы (n=4).
Рисунок 18 - периферически введенный CuZnСОД-полиионный комплекс ингибирует увеличение кровяного давления, вызванное интрацеребровентрикулярной (ИЦВ) инъекцией ангиотензина II. Максимальное изменение среднего артериального давления (САД) после ИЦВ ангиотензина II было измерено через 0, 1, 2 и 5 дней после введения через сонную артерию свободной CuZnСОД или CuZnСОД-полиионного комплекса.
Рисунок 19 - нейрозащитный эффект МККМ, нагруженных полиионным комплексом каталазы, против потери дофаминэргических нейронов, вызванной МФТП.
Рисунок 10 - ПААГ электрофорез полиионного комплекса каталазы с различными сшивками. Образцы были подвержены ПААГ электрофорезу (10% гель) при денатурирующих условиях (ДСН). Дорожки: 1 - маркеры молекулярного веса; 2 - каталаза; и полиионные комплексы, сшитые 3 - ЭДК, 4 - ГА, 5 - БСЗ.
Рисунок 21 - ПААГ электрофорез СОД-полиионного комплекса с различными сшивками. Образцы были подвержены ПААГ электрофорезу (10% гель) при денатурирующих условиях (ДСН). Дорожки: 1 - маркеры молекулярного веса; 2 - СОД; 3 - несшитый полиионный комплекс; и полиионные комплексы с различными сшивками: 4 -ЭДК, 4 - ГА, 6 - БСЗ.
Рисунок 22А - ПААГ электрофорез полиионного комплекса каталазы и СОД с различными сшивками. Образцы были подвержены ПААГ электрофорезу (10% гель) при денатурирующих условиях (ДСН). Дорожки: 1 - несшитый комплекс; полиионные комплексы с различными сшивками: 2 - ГА; 3 - ЭДК; 4 - БСЗ и 5 - ЭДК-S-NHS. Окрашивание произведено антителами к каталазе. Рисунок 22Б - ПААГ электрофорез полиионных комплексов каталазы и СОД с различными сшивками. Образцы были подвержены ПААГ электрофорезу (10% гель) при денатурирующих условиях (ДСН). Дорожки: 1 - несшитый комплекс; полиионные комплексы с различными сшивками: 2 - ГА; 3 - ЭДК; 4 - БСЗ и 5 - ЭДК-S-NHS. Окрашивание произведено антителами к СОД.
Рисунок 23 - биораспределение Li-COR, меченных МККМ, нагруженных полиионным комплексом каталазы. МККМ были выделены из BALB/C мышей, выращены до созревания (12 дней) и помечены Li-COR. Затем их нагружали полиионным комплексом каталазы в течение 2 часов. Нагруженные МККМ внутривенно ввели обритым EALB/C мышам (50 млн/мышь). Мышей держали на жидкой диете в течение 24 часов.
Детальное описание изобретения
По данной заявке предложены композиции и методы для сайт-специфической и/или продолжительной доставки интересующего белка/полипептида. В частности, композиции, содержащие полиионный комплекс интересующего полипептида с синтетическим полимером, имеющим суммарный заряд, противоположный суммарному заряду интересующего белка.
В предпочтительном варианте осуществления настоящего изобретения синтетические полимеры, входящие в состав комплексов, являются блок сополимерами. В частности, данные синтетические полимеры представляют собой блок сополимеры, включающие по меньшей мере один полиионный сегмент и по меньшей мере один неионный водорастворимый полимерный сегмент. Наиболее просто блок сополимеры можно определить как конъюгаты по крайней мере двух разных полимерных сегментов (Tirrel, М. In: Interactions of Surfactants with Polymers and Proteins. Goddard E.D. и Ananthapadmanabhan, K.P. (eds.), CRC Press, Boca Raton, Ann Arbor, London, Tokyo, pp.59-122, 1992). Наиболее простая структура блок сополимера включает два сегмента, соединенных концами, образуя диблок типа А-В. Последующая конъюгация концами более чем двух сегментов образует триблок типа А-В-А, мультиблок А-Б-А-Б или мультисегмент типа А-В-С-. Если в структуре блок сополимера можно выделить главную цепь, в которой одна или несколько повторяющихся единиц соединяются с разными сегментами полимера, тогда сополимер имеет привитую (графт) архитектуру, например типа А(В)n. Более сложными структурами являются, например, звездообразные сополимеры типа (АВ)n или АnВm, которые имеют более двух полимерных сегментов, соединенных в одном центре. Типичный блок сополимер в настоящем изобретении имеет структуру А-В или В-А, где А это полиионный блок, а В - неионный водорастворимый полимерный блок. Блоки блок сополимера могут иметь от 2 до 1000 повторяющихся мономеров.
Размер комплексов от приблизительно 5 до 500 нм является предпочтительным, более предпочтительным от приблизительно 5 до 250 нм, более предпочтительным от приблизительно 10 до 140 нм и еще более предпочтительным от приблизительно 20 до 100 нм. Комплексы не агрегируются и остаются в пределах предпочитаемого диапазона размеров на протяжении по меньшей мере одного часа после разведения в водном растворе при физиологических значениях рН и ионной силы, например в фосфатно-солевом буфере (ФСБ) при рН 7,4. Размер комплексов может быть измерен как средний диаметр с помощью динамического светорассеяния (см. Batrakova et al. (2007) Bioconjugate Chem., 18:1498-1506). Предпочтительно, чтобы после растворения в водном растворе комплексы оставались стабильны, то есть не агрегировались и/или оседали по крайней мере в течение 2 часов, предпочтительнее в течение 12 часов, еще предпочтительнее в течение 24 часов.
Полиионный блок сополимера имеет суммарный заряд, противоположный заряду интересующего белка. Например, если интересующий белок имеет суммарный отрицательный заряд, тогда полиионный блок будет иметь суммарный положительный заряд при соответствующем рН. Полиионный блок может быть поликатионом (то есть полимером, имеющим суммарный положительный заряд при соответствующем рН) или полианионом (то есть полимером, имеющим суммарный отрицательный заряд при соответствующем рН). В конкретном варианте осуществления настоящего изобретения, полиионный блок имеет по меньшей мере три заряда, предпочтительно по меньшей мере 10 зарядов, более предпочтительно по меньшей мере 15 зарядов. В предпочтительном варианте осуществления настоящего изобретения заряды располагаются достаточно близко друг к другу. Действительно, опуская теоретические подробности, известно, что когда расстояние между зарядами на полиэлетролите меньше определенного критического значения, малые противоионы, содержащиеся в растворе, могут конденсироваться вокруг цепи полиэлектролита. Например, «длина Бьеррума» в водном растворе полиэлектролитов равна приблизительно 7 Å (см. Manning (1980) Biopolymers, 19:37-59). Такие противоионы могут высвобождаться во внешний раствор при реакции полиэлектролитов с противоположно заряженными полиионами, таким образом обеспечивая «движущую силу» для образования полиэлектролитных комплексов (Kabanov et al. (2002) Structure, dispersion stability and dynamics of DNA and polycation complexes. In Pharmaceutical Perspectives of Nucleic Acid-Based Therapeutics (S.W. Kim, R. Mahato, Eds.) Taylor & Francis, London, New York, pp.164-189).
Степень полимеризации полиионных блоков обычно находится в пределах между 10 и примерно 100000. Более предпочтительна степень полимеризации между примерно 20 и примерно 10000, еще более предпочтительна степень полимеризации между примерно 10 и 1000, еще более предпочтительна степень полимеризации между примерно 10 и 200.
Полиионный блок включает поликатионные и полианионные сегменты. Примеры поликатионных сегментов включают, но не ограничиваются, полимеры и сополимеры и их соли, содержащие блоки, состоящие из одного или более мономеров, включая без ограничения, первичные, вторичные и/или третичные амины, каждый из которых может быть частично или полностью кватернизирован, таким образом, образуя четвертичные соли аммония. Примерами таких мономеров являются катионные аминокислоты (например, лизин, аргинин, гистидин, орнитин и тому подобные), алкиленимины (например, этиленимин, пропиленимин, бутиленимин, пентиленимин, гексиленимин, спермин и т.п.), виниловые мономеры (например, винилкапролактам, винилпиридин и т.п., акрилаты и метакрилаты (например, N,N-диметиламиноэтил акрилат, N,N-диметиламиноэтил метакрилат, N,N-диэтиламиноэтил акрилат, N,N-диэтиламиноэтил метакрилат, t-бутиламиноэтил метакрилат, акрилоксиэтилтриметил галид аммония, акрилоксиэтил-диметибензил галид аммония, метакриламидопрорилтриметил галид аммония и т.п.), аллильные мономеры (например, диметил-диаллил хлорид аммония), алифатические, гетероциклические или ароматические неионы.
Поликатионы и фрагменты поликатионов могут быть получены путем полимеризации мономеров, которые сами по себе могут не быть катионами, таких как, например, 4-винилпиридин, которые затем переходят в поликатионную форму путем различных химических реакций мономеров, например алкилирования, что приводит к возникновению ионизируемых групп. Преобразование мономеров может быть неполным. В результате часть мономеров в сополимере не имеет ионизируемых групп, как например, в сополимере винилпиридина и галида N-алкилвинилпиридиния.
Сополимеры, входящие в состав поликатионных участков, могут содержать более одного типа мономерных единиц, включая комбинации катионных мономерных единиц с по меньшей мере одним другим типом мономерных единиц, включая, например, другие катионы, анионы, цвиттерионы, гидрофильные неионные и/или гидрофобные мономерные единицы. Такие поликатионные участки могут быть получены путем сополимеризации более чем одного типа химически различных мономеров. В таком сополимере заряженные группы должны быть расположены достаточно близко друг к другу, чтобы позволить образование комплекса при реакции с другими компонентами. В предпочтительном варианте осуществления настоящего изобретения, количество некатионных единиц должно быть достаточно мало, чтобы полимер или полимерный блок оставался в целом положительно заряженным. Полимер, содержащий поликатионный участок, может состоять из смеси двух или более полимеров различной структуры, таких как полимеры с разной степенью полимеризации, структурой скелета и/или функциональными группами.
Примеры полианионных блоков включают, но не ограничиваются, полимеры и их соли, содержащие единицы, полученные от одного или более мономеров, включая: ненасыщенные этиленовые монокарбоновые кислоты, ненасыщенные этиленовые дикарбоновые кислоты, этиленовые мономеры, включающие сульфоновую кислотную группу, их соли щелочных металлов и аммониевые соли. Набор примеров таких мономеров включает акриловую кислоту, метакриловую кислоту, аспарагиновую кислоту, альфа-акриламидометилпропансульфоновую кислоту, 2-акриламидо-2-метилпропансульфоновую кислоту, диоксипиридинкарбоновую кислоту, цитраконовую кислоту, транскоричную кислоту, глутаминовую кислоту, трансглутаконовую кислоту, 4-гидрокси коричную кислоту, итаконовую кислоту, фумаровую кислоту, линоленовую кислоту, линолевую кислоту, малеиновую кислоту, нуклевые кислоты, транс-бета-гидромуконовую кислоту, транс-транс-муконовую кислоту, олеиновую кислоту, 1,4-фенилендиакриловую кислоту, фосфат 2-пропен-1-сульфоновую кислоту, рицинолевую кислоту, 4-стиролсульфоновую кислоту, стиролсульфоновую кислоту, 2-сульфоэтил метакрилат, винилсульфоновую кислоту, винилбензилсульфоновую кислоту, винил фосфорную кислоту, винилбензойную кислоту и винилгликолевую кислоту и т.п., а также карбоксилированный декстран, сульфурированный декстран, геперин и т.п. Набор примеров полианионов включает, но не ограничивается, полималеиновую кислоту, полиаминокислоты (например, полиаспартат, полиглутамат и их сополимеры), полиакриловую кислоту и т.п.
Полианионы и участки полианионов могут быть получены путем полимеризации мономеров, которые сами по себе могут не быть анионными или гидрофильными, как, например, трет-бутил метакрилат или цитраконовый ангидрид, которые затем переходят в полианионную форму посредством различных химических реакций мономерных единиц, например гидролиза, что приводит к образованию ионизированных групп. Преобразование мономерных единиц может быть неполным. В результате часть мономеров в сополимере не имеет ионизируемых групп, как например, в сополимере трет-бутил метакрилата и метакриловой кислоты.
Сополимеры, входящие в состав полианионных участков, могут содержать более одного типа мономерных единиц, включая комбинации анионных единиц с по меньшей мере одним типом единиц, включая, например, другие анионы, катионы, цвиттерионы, гидрофильные неионные и/или гидрофобные единицы. Такие полианионные участки могут быть получены путем сополимеризации более чем одного типа химически различных мономеров. В таком сополимере заряженные группы должны быть расположены достаточно близко друг к другу, чтобы позволить образование комплекса при реакции с другими компонентами. В предпочтительном варианте осуществления настоящего изобретения, количество неанионных единиц должно быть достаточно мало, чтобы полимер или полимерный блок оставался в целом положительно заряженным. Полимер, содержащий полианионный участок, может состоять из смеси двух или более полимеров различной структуры, таких как полимеры с разной степенью полимеризации, структурой скелета и/или функциональными группами.
В предпочтительном варианте осуществления настоящего изобретения, полиионный сегмент представляет собой полипептид, принадлежащий к группе полимеров или сополимеров лизина, гистидина, аргинина, орнитина, аспартата и/или глутамата и их солей. Набор примеров подобных синтетических полиионов включает полилизин, полигистидин, полиаргинин, полиарнитин, полиаспартат, полиглутамат и их соли. В другом предпочтительном варианте полиионый сегмент принадлежит к группе, состоящей из полиакриловой кислоты, полиалкилен акриловой кислоты, полиалкиленимина, полиэтиленимина, полифосфатов и их солей.
Неионный водорастворимый полимерный сегмент может быть выбран из группы, состоящей из полиэтилен оксида, сополимера этиленоксида и пропиленоксида, полисахарида, полиакриламида, полиглицерола, поливинилалкоголя, поливинилпирролидона, поливинилпиридин N-оксида, сополимера винилпиридин N-оксида и винилпиридина, полиоксазолина и полиакроилморфолина или их производных. Неионные полимерные сегменты предпочтительно должны быть нетоксичными и неиммуногенными. В конкретном варианте осуществления настоящего изобретения водорастворимыми полимерами являются полиэтиленоксид (ПЭО), полиэтиленгликоль (ПЭГ) или сополимер этиленоксида и пропиленоксида. Если неионным водорастворимым полимерным сегментом является полиэтиленоксид, тогда предпочтительный молекулярный вес этого полимера находится между примерно 300 и примерно 20000, более предпочтительно между примерно 1500 и примерно 15000, еще более предпочтительно между примерно 2000 и 10000 и еще более предпочтительно между примерно 4000 и 10000.
Полиионный и неионный водорастворимый полимерные сегменты могут содержать разные концевые группы. Например, метод синтеза может привести к включению различных концевых групп.
Комплексы данного изобретения могут спонтанно самоорганизовываться в частицы наноразмера. Не вдаваясь в теорию, считается, что сформированные частицы имеют структуру типа ядро-оболочка (core-shell). Ядро частиц включает белок-полиионный комплекс, а гидрофильная оболочка включает неионный водорастворимый сегмент сополимера. Действительно, нейтрализация заряда полииона приводит к образованию гидрофобных доменов, которые склонны к сегрегации в водной среде. Несмотря на это, водорастворимые неионные сегменты предотвращают агрегацию и макроскопическое разделение фаз. В результате комплексы самоорганизуются в частицы наноразмера и образуют стабильные водные дисперсии.
Для образования защитного наноконтейнера для интересующего полипептида или белка, блок сополимеры синтезируются путем конъюгации полиионного сегмента (например, полиэтиленимина (ПЭИ, 2000 Да)) и неионного водорастворимого сегмента (например, полиэтиленоксида (ПЭО) 10000 Да)). (Vinogradov et al. (1999) Bioconjug. Chem., 10:851-60). Комплексы могут быть получены путем добавления раствора интересующего белка (например, каталазы (1 мг/мл)) к раствору блок сополимера (например, ПЭИ-ПЭГ (2 мг/мл)) в буфере (например, в фосфатно-солевом буфере (рН 7.4)), образуя молочно-белые дисперсии.
В конкретном варианте осуществления настоящего изобретения, частицы достигают клеток тела в изотоническом растворе при рН 7.4. Однако комплексы могут быть приготовлены до введения частиц при больших или меньших значениях рН. Известно, что многие полипептиды, представляющие интерес для данного изобретения, являются полиамфолитами, содержащими как положительно, так и отрицательно заряженные группы. Баланс положительно и отрицательно заряженных групп в таких полипептидах зависит как от их химической структуры, так и от рН внешней среды. При значениях рН ниже изоэлектрической точки (рI) полипептиды могут быть положительно заряжены, в то время как при значениях рН выше рI комплексы могут быть отрицательно заряжены. Таким образом, в соответствии с данным изобретением, комплексы могут быть получены при реакции полипептидов с полианионом при рН ниже pI. Данные комплексы могут быть также получены при реакции полипептидов с полиионами при рН выше pI. После образования комплексов pH раствора может быть изменен до желаемого значения, например до, 7.4, для дальнейшего применения. В некоторых случаях полипептиды могут содержать участки или домены с множественными положительно или отрицательно заряженными группами, расположенными близко друг к другу. Подобные полипептиды могут образовывать комплексы с противоположно заряженными полиионами (например, поликатионами в случае, если участок полипептида содержит множественные отрицательно заряженные группы, или полианионами, если участок полипептида содержит множественные положительно заряженные группы) при рН как выше, так и ниже pI.
Ядро комплексов может быть сшитым. Сшивки могут химически связывать функциональные группы полипептида, полииона или полипептида и полииона, включая сшивки между полипептидом и полиионом. Сшивки могут быть расщепляемыми или разлагаемыми и могут расщепляться внутри тела или внутри клетки. Известно много различных методов сшивки, которые могут быть использованы в данном случае (G. Hermanson, Bioconjugate Techniques, Elsevier, 1996, 785 p.). Примерами сшивок являются, без ограничения, 1-этил-3-(3-диметиламинопропил)карбодиимин (ДЭК), глутаровый альдегид (ГА), формальдегид, дивинилсульфон, полиангидрид, эпихлоргидрин, диглицидиловый эфир этиленгликоля, диглицидиловый эфир бутандиола, полиглицидиловый эфир полиглицерола, полиэтиленгликоль, диглицидиловый эфир полипропиленгликоля, бис- или полиэпокси сшивки (например, 1,2,3,4,-диэпоксибутан или 1,2,7,8-диэпоксиоктан), а также перечисленные в G. Hermanson (Bioconjugate Techniques, Elsevier, 1996). В конкретном варианте осуществления настоящего изобретения степень сшивания полипептид-полиионного комплекса варьируется от примерно 40% до примерно 70%, предпочтительно от примерно 40% до примерно 60%, более предпочтительно от примерно 40% до примерно 50%. Избыток блок сополимера в полипептид-полиионном комплексе может уменьшить степень сшивки, необходимую для стабильности комплекса.
Полипептид-полиионные комплексы настоящего изобретения могут вводиться млекопитающим, в частности людям. Ниже в данном документе показано, что полипептид-полиионные комплексы настоящего изобретения способны пересекать гематоэнцефалический барьер и доставлять интересующие полипептиды в ЦНС, в частности, если пациент имеет нейродегенеративное или нейровоспалительное заболевание или нарушение. Не вдаваясь в теорию, частицы полипептид-полиионных комплексов после введения в организм млекопитающих могут быть поглощены циркулирующими клетками, способными достигать мозг. Таким образом, часть полипептида будет доставлена в мозг этими клетками. В частности, этими клетками могут быть циркулирующие клетки иммунной системы, например, моноциты или макрофаги, предпочтительно моноциты, генерированные из клеток костного мозга (МККМ), макрофаги, дендритные клетки, лимфоциты, предпочтительно Т-клетки, нейтрофилы, эозинофилы, базофилы и их комбинации.
Более того, предполагается, что комплексы настоящего изобретения могут защитить полипептид внутри клетки. В то же время, благодаря особой структуре ядро-оболочка, комплексы нетоксичны для клетки-хозяина и не нарушают ее функциональные свойства. В частности, комплексы не нарушают способность клеток достигать очага заболевания.
Не вдаваясь в теорию, также предполагается, что время циркуляции свободных или захваченных циркулирующими клетками комплекса может быть увеличено. В результате время представления комплексов гематоэнцефалическому барьеру увеличивается, и возрастает процент введенной дозы полипептида, доставленного в мозг.
Более того, не вдаваясь в теорию, также предполагается, что комплексы могут связываться и проникать в нервные клетки и/или в периферические нервные окончания, после чего они транспортируются в мозг путем ретроградного транспорта (Zweifel et al. (2005) Nat. Rev. Neurosci., 6:615-625; U.S. Patent Application Publication 2003/0083299) или схожего механизма. Уникальная структура комплексов настоящего изобретения, в частности, комбинация ионных и неионных полимерных цепей в сополимерах, обеспечивает защиту полипептидов, минимизирует повреждения клеток и тканей, а также способствует свободной миграции комплексов в мозг.
Полипептид-полиионные комплексы настоящего изобретения могут вводиться парентерально, включая, но не ограничиваясь, подкожный, внутривенный и интраперитониальный методы введения. Кроме того, возможно введение полипептид-полиионных комплексов непосредственно в нервную систему, в частности в полость позвоночного канала, внутрицеребрально или эпидурально. Полипептид-полиионные комплексы также могут вводиться внутримышечно, внутрикожно или через сонную артерию. Возможно использование комбинации различных методов введения.
В соответствии с другим вариантом осуществления настоящего изобретения, клетки нагружаются полипептид-полиионным комплексом и затем вводятся пациенту, как терапевтический препарат. Конкретнее, клетки являются циркулирующими клетками крови, в частности клетками иммунной системы. Иммунные клетки включают без ограничения моноциты, макрофаги, моноциты, генерированные из клеток костного мозга, дендритные клетки, лимфоциты, Т-клетки, нейтрофилы, эозинофилы, базофилы и/или их комбинации. Нагруженные клетки способны проникать через гематоэнцефалический барьер и доставлять интересующий полипептид, в частности, если пациент имеет нейродегенеративное или нейровоспалительное заболевание или нарушение. Клетки могут быть выделены из млекопитающих, используя доступные методы выделения и сортировки. Как описано ниже, клетки могут быть нагружены полипептид-полиионным комплексом путем инкубации с полипептид-полиионным комплексом. Нагруженные клетки могут быть введены парентерально, включая, но не ограничиваясь, подкожным, внутривенным и интраперитониальным методами введения. Кроме того, возможно введение полипептид-полиионных комплексов непосредственно в нервную систему, в частности в полость позвоночного канала, внутрицеребрально или эпидурально. Полипептид-полиионные комплексы также могут вводиться внутримышечно, внутрикожно или через сонную артерию. Возможно использование комбинации различных методов введения.
Нейровоспаление, возникшее в результате активации одноядерных фагоцитов мозга (периваскулярных и паренхимальных макрофагов и микроглии), а также астроцитов и эндотелиальных клеток, может посредством паракринных механизмов усилить нервные повреждения в широком спектре болезней, таких как болезнь Альцгеймера, Паркинсона, Хантингтона, ВИЧ-ассоциированные нейрокогнитивные расстройства, губчатый энцефалопатит и инсульт. Воспалительные инфильтраты ЦНС при этих расстройствах очень комплексны и многогранны. На первых стадиях одноядерные фагоциты начинают каскад, который позже приводит к активации и рекрутингу искусственного иммунитета и, в конце концов, нейродегенерации. Микроглия - это первые одноядерные фагоциты в ЦНС, которые отвечают на повреждение и чьей основной функцией является защита мозга. Активированная микроглия участвует в воспалительном процессе, в результате которого образуются нейротоксичные факторы, включающие хинолиновую кислоту, супероксидные анионы, матричные металлопротеазы, оксид азота, арахидоновую кислоту и ее метаболиты, хемокины, воспалительные цитокины и экситотоксины, включая глутамат, которые приводят к нейродегенерации. С другой стороны нейрозащитные функции микроглии могут быть контролированы через ее способность выделять нейротрофины, а также связывать и удалять экситотоксины из межклеточного пространства. Действительно известно, что микроглия положительно влияет на выживаемость нейронов после повреждений мозга. Не вдаваясь в теорию, предполагается, что схожесть механизмов нейродегенерации может быть использована для достижения терапевтического эффекта с помощью иммунных клеток, нагруженных полипептид-полиионными комплексами. В предпочтительном варианте осуществления настоящего изобретения будут использоваться одноядерные фагоциты, обладающие замечательной способностью пересекать гематоэнцефалический барьер, благодаря миграции и способности к экстравазации.
Показательный метод вышеописанного варианта осуществления настоящего изобретения включает: выделение интересующих клеток из пациента, инкубирование выделенных клеток с полипептид-полиионным комплексом и введение их обратно пациенту. Не вдаваясь в теорию, предполагается, что данный подход возможен благодаря способности полипептид-полиионных комплексов защищать их груз от агрессивного протеолиза в лизосомах фагоцитов. Также предполагается, что структура полипептид-полиионных комплексов типа ядро-оболочка не влияет на способность клеток проходить через гематоэнцефалический барьер и переносить свой груз в мозг.
I. Определения
Следующие определения предоставлены, чтобы улучшить понимание настоящего изобретения
Использованный здесь термин «полимер» означает молекулы, образованные при химическом соединении двух или более повторяющихся единиц или мономеров. Термин «блок сополимер» в наиболее простом случае относится к конъюгатам по меньшей мере двух разных полимерных сегментов, из которых каждый полимерный сегмент включает две или более расположенные рядом единицы одного типа.
Термин «выделенный белок» или «выделенный и очищенный белок» также несколько раз использован здесь. Этот термин в первую очередь относится к белку, полученному путем экспрессии отдельной молекулы нуклеиновой кислоты. Альтернативно, этот термин может относиться к белку, который был в достаточной степени отделен от других белков, с которыми он естественно связан, и таким образом переведен в «достаточно чистую» форму. Понятие «выделенный» не исключает искусственные или синтетические смеси с другими компонентами или материалами, а также примеси, которые не влияют на основную активность и могут присутствовать, например, из-за неполной очистки или добавления стабилизаторов.
«Полипептид» или «белок» иногда использованы здесь как взаимозаменяемые понятия и означают молекулярную цепь, состоящую из аминокислот. Термин «полипептид» включает пептиды, олигопептиды и белки. Эти термины также означают постэкспрессионные модификации полипептидов, например гликозилирование, ацетилирование, фосфорилирование и т.п. Вдобавок, бекловые фрагменты, аналоги, мутированные или вариантные белки, сшитые белки и т.п. также попадают под значение полипептид.
Термин «изолированный» может означать белок, нуклеиновую кислоту, вещество или клетки, которые были достаточно отделены от окружения, с которым они естественно были бы связаны таким образом, что присутствуют в «достаточно чистой» форме. «Изолированный» не обязательно исключает искусственные или синтетические смеси с другими компонентами или материалами, а также примеси, которые не влияют на основную активность и могут присутствовать, например, из-за неполной очистки.
«Фармацевтически приемлемый» означает одобренный управляющим органом федерального или местного правительства или перечисленный в Фармакопеи США или другой общепризнанной фармакопеи для использования в животных, в частности в людях.
«Носитель» относится к, например, растворителю, адъюванту, консерванту (например, тимерсол, бензил алкоголь), антиоксиданту (например, аскорбиновая кислота, метабисульфат натрия), солюбилизатору (например, Твин 80, Полисорбат 80), эмульгатору, буферу (например, Трис-HCl, ацетатный или фосфатный), воде, водным растворам, маслам, веществам, увеличивающим объем (например, лактоза, маннитол), наполнителям, вспомогательным веществам или средам, с которыми вводится, активный агент настоящего изобретения. Подходящие фармацевтические носители описаны в книге Э.В.Мартина "Remington's Pharmaceutical Sciences" (Mack Publishing Co., Easton, PA); Gennaro, A.R., Remington: The Science and Practice of Pharmacy, 20th Edition (Lippincott, Williams and Wilkins), 2000; Liberman, et al., Eds., Pharmaceutical Dosage Forms, Marcel Decker, New York, N.Y., 1980; and Kibbe, et al., Eds., Handbook of Pharmaceutical Excipients (3rd Ed.), American Pharmaceutical Association, Washington, 1999.
II. Терапевтический препарат
В то время как в предпочтительном варианте осуществления настоящего изобретения белки заключены внутри полимерного комплекса, в область настоящего изобретения также попадает инкапсуляция в полимерные комплексы других терапевтических агентов или интересующих веществ. Такие агенты или вещества включают без ограничения полипептиды, пептиды, нуклеиновые кислоты или синтетические и натуральные лекарства. В предпочтительном варианте осуществления настоящего изобретения, терапевтический агент является полипептидом или белком. В то время как на протяжении всего описания настоящего изобретения упоминаются полипептид-полиионные комплексы, использование белков также предполагается в настоящем изобретении. Здесь термины полипептид и белок часто используются взаимозаменяемо.
В предпочтительном варианте осуществления настоящего изобретения интересующий белок в полимерном комплексе является терапевтическим белком, то есть он влияет на снижение проявления симптомов и/или лечит заболевание, расстройство, патологию и/или симптомы, с ним связанные. Белки могут иметь терапевтическое действие против неврологических расстройств (в частности, ЦНС), включая без ограничений, нейродегенеративные расстройства, болезни Альцгеймера, Паркинсона, Хантингтона, инсульт, травмы, инфекции, менингит, энцефалит, глиомы, рак (включая метастазы в мозгу), ВИЧ-1-ассоциированную деменцию, ВИЧ-ассоциированные нейрокогнитивные расстройства, паралич, амиотропный латеральный склероз (АЛС или болезнь Лу Герига), множественный склероз (МС), ЦНС-ассоциированную кардиоваскулярную болезнь, прионную болезнь, ожирение, расстройства обмена веществ, воспалительное заболевание и лизосомные болезни накопления (ЛБН, такие как, без ограничения, болезнь Гаучера, Помпа, Нимана Пика, синдром Гунтера, мукополисахараидоз I (МПС I), ганглиозидоз типа II, синдром Санфилиппо, болезнь Тай-Саха, болезнь Сандхоффа, болезнь Краббе, метахроматическая лейкодистрофия и болезнь Фабри). Терапевтически активные белки включают, но не ограничиваются, ферментами, антителами, гормонами, факторами роста, другими полипептидами, введение которых в мозг может влиять на снижение проявления симптомов и/или лечить заболевание, расстройство, патологию и/или симптомы, с ним связанные. Нейроактивные полипептиды, пригодные для настоящего изобретения включают, но не ограничиваются, эндокринные факторы, факторы роста, факторы, выделяемые гипоталамусом, нейротрофические факторы, паракринные факторы, нейротрансмиттерные полипептиды, антитела и фрагменты антител, которые связываются с любыми из вышеперечисленных полипептидов, антителами и фрагментами антител, которые связываются с рецепторами этих полипептидов, цитокинами, эндорфинами, нейтрализующими полипептидами, агонистами рецепторов экспрессированных в ЦНС, полипептидами, вовлеченными в лизосомальные болезни накопления и т.п. В данном варианте осуществления настоящего изобретения терапевтический белок оказывает действие на ЦНС. В другом варианте осуществления настоящего изобретения терапевтический белок не пересекает гематоэнцефалический барьер.
Примеры специфичных белков включают без ограничения каталазу, теломеразу, супероксид дисмутазу (СОД), глутатионпероксидазу, глутаминазу, цитокины, эндорфины (например, энкефалин), факторы роста (например, эпидермальный фактор роста (ЭФР), основной и кислотный факторы роста фибробластов (aFGF и bFGF), инсулиноподобный фактор роста I (IGF-I), нейротрофический фактор головного мозга (BDNF), глиальный нейротрофический фактор (GDNF), нейротрофический фактор тромбоцитов (PDNF), фактор роста сосудов (VGF), фактор роста нервов (NGF), инсулиноподобный фактор роста (IGF-II), фактор некроза опухоли (TNF), фактор ингибирования лейкемии (LIF), различные интерлейкины и т.п.), антиапоптозные белки (Blc-2, PI3 киназа и т.п.), вещества, связывающие амилоид бета (например, антитела), модуляторы α-, β- и/или γ-секретаз, вазоактивный интестинальный пептид, лептин, кислую альфа-гликозилазу (GAA), кислую сфингомиелазу, N-ацтилгалактозамин-4-сульфатазу А и нейротрансмиттеры (см. e.g., Schapira, A.H. (2003) Neurology 61:S56-63; Ferrari et al. (1990) Adv Exp Med Biol. 265:93-99; Ferrari et al. (1991) J Neurosci Res. 30:493-497; Koliatsos et al. (1991) Ann Neurol. 30:831-840; Dogrukol-Ak et al. (2003) Peptides 24:437-444; Amalfitano et al. (2001) Genet Med. 3:132-138; Simonaro et al. (2002) Am J Hum Genet. 71:1413-1419; Muenzer et al. (2002) Acta Paediatr Suppl. 91:98-99; Wraith et al. (2004) J Pediatr. 144:581-588; Wicklow et al. (2004) Am J Med Genet. 127A:158-166; Grabowski (2004) J Pediatr. 144:815-19; Auclair et al. (2003) Mol Genet Metab. 78:163-174; Przybylska et al. (2004) J Gene Med. 6:85-92). Лизосомальные болезни накопления - это наследственные генетические дефекты, которые приводят к дефициту ферментов и нарушают естественный метаболический цикл (Enns and Huhn, (2008) Neurosurg. Focus 24:E12). Это приводит к различным прогрессирующим физическим и/или умственным нарушениям, и считается, что доставка этих дефицитных ферментов в мозг может вылечить эти заболевания. Различные ферменты, вовлеченные в лизосомальные болезни накопления или ферменты, которые могут восполнить функцию дефицитных белков, могут быть доставлены с помощью методов, предложенных в настоящем изобретении.
В одном варианте осуществления, настоящее изобретение может быть использовано как лечение острой нервной токсичности вредных агентов, основываясь на доставке в мозг бутирилхолинэстеразы (БХЭ) или ацетилхолинэстеразы, реактиваторов холинэстеразы (например, оксидов), веществ, связывающих органофосфаты, и ингибиторов карбаматного типа. Так как бутирилхолинэстераза также гидролизует множество лекарств, содержащих сложные эфиры, такие как кокаин и сукцинилхолин, БХЭ в составе комплексов настоящего изобретения имеет терапевтический потенциал против кокаиновой зависимости и токсичности (e.g., Carmona et.al. (1999) Drug Metab. Dispos., 28:367-371; Carmona (2005) Eur. J. Pharmacol., 517:186-190).
Методы настоящего изобретения подразумевают использование полипептидных комплексов, содержащих один или несколько эффективных полипептидов, или использование нескольких комплексов, содержащих различные полипептиды, которые могут вводиться самостоятельно или с клетками, одновременно или отдельно друг от друга. Комплексы могут находиться в одной или разных композициях.
III. Методы введения препарата
Описанные здесь полипептид-полиионные комплексы и клетки, их содержащие, могут вводиться пациентам как фармацевтический препарат. Термин «пациент» в данном контексте означает человеческую или животную особь. Полипептид-полиионные комплексы, а также клетки, их содержащие, могут быть использованы в терапевтических целях под наблюдением врача.
Фармацевтическая композиция, содержащая полипептид-полиионный комплекс и/или клетки, нагруженные полипептид-полиионным комплексом настоящего изобретения, может быть легко сформулирована с любым подходящим фармацевтическим носителем. Например, комплексы и клетки могут быть сформулированы с подходящей средой, например водой, фосфатным солевым буфером, этанолом, многоатомными спиртами (например, глицеролом, пропиленгликолем, жидким полиэтиленгликолем и т.п.), диметилсульфоксидом (DMSO), маслами, детергентами, суспендирующими веществами или их подходящей смесью. Концентрация полипептид-полиионного комплекса и/или клеток в выбранной среде может варьироваться, и выбор среды может зависеть от метода введения фармацевтического препарата. Кроме случаев, когда использование любой из обозначенных выше сред несовместимо с полипептид-полиионными комплексами или клетками, предполагается их использование как фармацевтического препарата.
Согласно настоящему изобретению подходящие для каждого пациента доза и схема приема полипептид-полиионных комплексов и/или клеток могут быть определены врачом, принимая во внимание возраст, пол, вес, общее физическое состояние и конкретное состояние пациента и его заболевания, для лечения которого используются полипептид-полиионные комплексы или клетки. Врач также может принимать во внимание способ введения, фармацевтический носитель и активность полипептид-полиионного комплекса или клеток.
Выбор подходящего фармацевтического препарата будет также зависеть от способа его введения. Например, полипептид-полиионный комплекс или клетки, содержащие полипептид-полиионный комплекс настоящего изобретения, может вводиться путем прямой инъекции в область в непосредственной близости гематоэнцефалического барьера. В данном случае фармацевтический препарат содержит полипептид-полиионный комплекс или клетки, диспергированные в среде, подходящей для участка инъекции.
Полипептид-полиионные комплексы или клетки настоящего изобретения могут вводиться любым методом, таким как внутривенно в кровяное русло, перорально, подкожно, внутримышечно или интраперитониально. В данной области уже существуют фармацевтические препараты для инъекций. Если инъекция выбрана как метод введения полипептид-полиионного комплекса или клеток, необходимо принять меры, чтобы обеспечить доставку достаточного количества молекул или клеток к клеткам-мишеням для проявления биологического эффекта.
Фармацевтические композиции, содержащие комплекс или клетки настоящего изобретения в качестве активного ингредиента в смеси с фармацевтически приемлемым носителем, могут быть приготовлены в соответствии с общепринятыми фармацевтическими методами приготовления лекарственных смесей. Носитель может иметь большое разнообразие форм в зависимости от того, какой способ приготовления желателен для введения, например, внутривенного, перорального, прямой инъекции, интракраниального и интравитриального.
Фармацевтический препарат в данном изобретении может быть приготовлен в виде единицы дозирования для облегчения введения и обеспечения постоянства дозы. Под единицей дозирования здесь подразумевается физически изолированная единица фармацевтического препарата, подходящая для пациента, находящегося на лечении. Каждая единица дозирования должна содержать необходимое количество активного компонента в смеси с фармацевтическим носителем, чтобы вызвать желаемый эффект. Методы для определения подходящей единицы дозирования хорошо известны и отработаны.
Единица дозирования может быть пропорционально увеличена или уменьшена в зависимости от веса пациента. Концентрации, достаточные для частичного снятия симптомов конкретного патологического состояния, могут быть определены из расчетов кривой дозовой зависимости концентрации, как принято в данной области.
По данной заявке, подходящая единица дозирования для введения полипептид-полиионных комплексов или клеток, содержащих полипептид-полиионные комплексы, может быть определена по оценке токсичности данных молекул или клеток в животных моделях. Минимальные и максимальные дозы полипептид-полиионных комплексов или клеток, входящих в состав фармацевтического препарата, могут быть определены, основываясь на анализе целебных и побочных эффектов лечения различными дозами препарата.
Подходящая единица дозирования может также быть определена по анализу, эффективности действия полипептид-полиионных комплексов или клеток в комбинации с другими стандартными лекарствами. Единицы дозирования полипептид-полиионных комплексов могут быть определены индивидуально или в комбинации с любым другим методом лечения в зависимости от наблюдаемого эффекта.
Введение фармацевтического препарата, содержащего полипептид-полиионные комплексы или клетки, может производиться через определенные промежутки времени, например, по меньшей мере дважды или более в день до тех пор, пока проявление патологических симптомов достаточно снизится, после чего дозировка может быть уменьшена до уровня, достаточного для поддержания состояния. Подходящие временные промежутки между введением препарата будут зависеть от состояния пациента.
Нижеперечисленные примеры описывают наглядные методы применения настоящего изобретения и никоим образом не ограничивают сферу его применения.
Пример 1
Существует острая необходимость доставки терапевтических полипептидов в пораженные участки мозга при болезнях Альцгеймера и Паркинсона (AD и PD) (Brinton, R.D. (1999) Int. J. Fertil. Womens Med., 44:174-85; Gozes, I. (2001) Trends Neurosci., 24:700-5; Kroll et al. (1998) Neurosurgery 42:1083-100), инфекциях (менингит, энцефалит, прионная болезнь и ВИЧ-ассоциированная деменция), (Bachis et al. (2005) Ann. N. Y. Acad. Sci., 1053:247-57; Wang et al. (2003) Virology 305:66-76), инсульте Koliatsos et al. (1991) Ann. Neurol., 30:831-40; Dogrukol-Ak et al. (2003) Peptides 24:437-44), лизосомальных болезнях накопления Desnick et al. (2002) Nat. Rev. Genet., 3:954-66; Urayama et al. (2004). Proc. Natl. Acad. Sci., 101:12658-63), ожирении (Banks, W. (2003) Curr. Pharm. Des., 9:801-809; Banks et al. (2002) J. Drug Target., 10:297-308) и других метаболических и воспалительных заболеваний ЦНС, которую невозможно переоценить.
Воспаление играет важную роль в метаболических и дегенеративных заболеваниях нервной системы (Perry et al. (1995) Curr. Opin. Neurobiol., 5:636-41). Сильные воспалительные процессы приводят к увеличенному выделению активных форм кислорода (АФК), что в свою очередь вызывает клеточную смерть и нейродегенерацию. Уровень АФК и других нейротоксинов может быть снижен путем воздействия на нейровоспалительные процессы при заболевании направленными антиоксидантами или лекарствами, которые ингибируют выделение или образование провоспалительных цитокинов и экозаноидов, что в итоге приведет к улучшению исхода заболевания (Prasad, et al. (1999) Curr. Opin. Neurol., 12:761-70).
Тем не менее, подобные методы ограничиваются тем, что лекарства должны не только пройти через гематоэнцефалический барьер, но также достигнуть достаточной концентрации, чтобы оказать воздействие на механизмы прогрессирующего заболевания. Кроме того, поскольку воспалительные процессы чаще всего происходят на ранних стадиях заболевания, терапевтические препараты должны применяться рано и достаточно часто. Ограниченная доставка лекарств является основным препятствием для разработки новых принципов лечения расстройств нервной системы.
Одним из таких заболеваний является болезнь Паркинсона, второе по распространенности нейродегенеративное расстройство у людей старше 65 лет. Это заболевание характеризуется недостатком нейротрансмиттера дофамина вследствие потери дофаминэргических нейронов в черной субстанции и их иннервации в стриатуме. Нейропатология болезни Паркинсона включает воспаление в мозгу, активацию микроглии и последующее выделение нейротоксинов, включая АФК, которые играют ключевую роль в повреждении и смерти клеток (McGeer et al. (1988) Neurology 38:1285-91; Busciglio et al. (1995) Nature 378:776-9; Ebadi et al. (1996) Prog. Neurobiol., 48:1-19; Wu et al. (2003) Proc. Natl. Acad. Sci., 100:6145-50). Мозг больного болезнью Паркинсона имеет пониженные уровни антиоксидантов и противоокислительных ферментов (Ambani et al. (1975) Arch. Neurol., 32:114-8; Riederer et al. (1989) J. Neurochem., 52:515-20; Abraham et al. (2005) Indian J. Med. Res., 121:111-5), что приводит к пониженной способности противостоять окислительному стрессу и нейродегенерации. Существует множество доказательств того, что антиоксиданты способны ингибировать воспалительные процессы и таким образом защищать дофаминэргические нейроны в лабораторных и животных моделях болезни Паркинсона (Wu et al. (2002) J. Neurosci., 22:1763-71; Du et al. (2001) Proc. Natl. Acad. Sci., 98:14669-74; Kurkowska-Jastrzebska et al. (2002) Int. Immunopharmacol., 2:1213-8; Teismann et al..(2001) Synapse 39:167-74; Ferger et al. (1999) Naunyn Schmiedebergs Arch. Pharmacol., 360:256-61; Ferger et al. (1998) Naunyn Schmiedebergs Arch. Pharmacol., 358:351-9; Peng et al. (2005) J. Biol. Chem., 280:29194-8). Реакция превращения перекиси водорода, известной АФК, до воды и молекулярного кислорода, катализируемая каталазой, известна как одна из самых быстрых для всех известных ферментов. Убедительные доказательства свидетельствуют, что антиоксиданты способны ингибировать воспалительные процессы и защищать до 90% дофаминэргических нейронов in vitro и in vivo (Wu et al. (2002) J. Neurosci., 22:1763-71; Du et al. (2001) Proc. Natl. Acad. Sci., 98:14669-74; Kurkowska-Jastrzebska et al. (2002) Int. Immunopharmacol., 2:1213-8; Teismann et al. (2001) Synapse 39:167-74; Ferger et al. (1999) Naunyn. Schmiedebergs Arch. Pharmacol., 360:256-61; Ferger et al. (1998) Naunyn. Schmiedebergs Arch. Pharmacol., 358:351-9; Peng et al. (2005) J. Biol. Chem., 280:29194-8). В in vitro модели болезни Паркинсона на первичной культуре церебральных тучных клеток было показано, что каталаза способна защитить клетки от токсичного действия АФК (Prasad et al. (1999) Curr. Opin. Neurol., 12:761-70; Gonzalez-Polo et al. (2004) Cell Biol. Int., 28:373-80). Более того, низкомолекулярный активатор каталазы, разагилин, оказывал нейрозащитное действие в мышиной модели болезни Паркинсона (Maruyama et al. (2002) Neurotoxicol. Teratol., 24:675-82). Было проведено несколько клинических испытаний низкомолекулярных антиоксидантов, среди которых R-токоферол и депренил наиболее активно снижали скорость прогрессирования болезни Паркинсона (Group, T.P.S. (1993) N. Engl. J., 328:176-183). Тем не менее, как говорилось выше, большинство испытаний не показали значительных улучшений из-за ограниченного транспорта R-токоферола через гематоэнцефалический барьер (Pappert et al. (1996) Neurology, 47:1037-42).
Материалы и методы
Материалы. Каталаза печени быка, полиэтиленимин (ПЭИ) (2К, разветвленный, 50% водный раствор), сульфородамин-В, додецил сульфат натрия, Сефадекс G-25 и Тритон Х-100 были приобретены в Sigma-Aldrich (St-Louis, МО). Эпоксид метоксиполиэтиленгликоля был приобретен в Shearwater Polymer Inc., Huntsville, AL.
МФТП. Интоксикация C57BL/6 мышей 1-метил-4-фенил-1,2,3,6-тетрагидропиридином (МФТП) проводилась, как описано ранее (Benner et al. (2004) Proc. Natl. Acad. Sci., 101:9435-40). Через 12 часов после инъекции МФТП мышам внутривенно ввели 50 µCi/мышь полипептид-полиионного комплекса, меченного 125I. Спустя 24 часа мышей умертвили и определили количество радиоактивности в основных органах (мозг, селезенка, печень, легкие и почки), используя гамма-счетчик 1480 Wizard 3 (Perkin-Elmer Life Sciences, Shelton, CT). Количество доставленного фермента было выражено как процентное отношение введенной дозы для целого органа.
Конъюгаты ПЭИ-ПЭГ. Сополимер был синтезирован согласно модифицированному протоколу (Nguyen et al. (2000) Gene Ther., 7:126-38) путем конъюгации ПЭИ и эпоксида метоксиполиэтиленгликоля. Коротко, к 5% водному раствору ПЭИ был добавлен водный раствор эпоксид метоксиполиэтиленгликоля. Смесь инкубировали в течение ночи при комнатной температуре. Полученные конъюгаты очистили от избытка ПЭИ (а также от низкомолекулярных примесей) путем диализа в течение 48 часов против воды (вода менялась дважды, используя диализные мембраны Spectra/For с границей пропускания 6000-8000 Да). Затем конъюгаты были сконцентрированы in vacuo. Для конечной очистки конъюгаты были растворены в 20 мл 100% метанола и полученный раствор покапельно был добавлен к 400 мл эфира. Преципитат был осажден центрифугированием (400 g, 5 минут), дважды отмыт и высушен в эксикаторе. Детальная характеристика полученного продукта осуществлялась спектрофотометрически и масс-спектрометрически, как описано ранее (Nguyen et al. (2000) Gene Ther., 7:126-38).
Блок иономерные комплексы. Определенные количества каталазы (1 мг/мл) и блок сополимера (2 мг/мл) были независимо растворены в фосфатно-солевом буфере при комнатной температуре. Раствор фермента добавляли покапельно к раствору блок сополимера при постоянном помешивании. Отношение +/- зарядов (Z) было рассчитано путем деления количества протонированных аминогрупп ПЭИ-ПЭГ при рН 7.4 (Vinogradov et al. (1998) Bioconjugate Chem., 9:805-812) на общее количество глутамина и аспарагина в каталазе. Состав, размер, дисперсионная стабильность, морфология, форма и структура полученных комплексов были охарактеризованы с помощью ряда физико-химических методов (ПААГ электрофорез, динамическое светорассеяние, просвечивающая электронная микроскопия), как было описано ранее (Vinogradov et al. (1999) Bioconjugate Chem., 10:851-60; Lemieux et al. (2000) J. Drug Target 8:91-105; Vinogradov et al. (2004) J. Drug Target., 12:517-26; Vinogradov et al. (2005) J. Controlled Release, 107:143-57).
ПЛАТ электрофорез. Образование комплексов было проверено с помощью анализа сдвига в полиакриламидном геле. Комплексы фермента с различным значением Z были загружены в 7,5% ПААГ в присутствии 5 мМ Триса, 50 мМ глицина и рН 8,3 при неденатурирующих условиях (без ДСН), чтобы сохранить комплекс. Белковые полосы были визуализированы, используя первичные поликлональные кроличьи антитела против каталазы (Ab 1877, Abeam Inc, Cambridge, MA; 1:6000) и вторичные антикроличьи антитела, меченные пероксидазой корня хрена (Amersham Life Sciences, Cleveland, ОН; 1:1500). Белковые полосы были визуализированы, используя хемилюминесцентный кит (Pierce, Рокфорд, Rockford, IL).
Динамическое светорассеяние. Фактический гидродинамический диаметр и зета-потенциал полипептид-полиионных комплексов измерялись с помощью спектроскопии корреляции фотонов на ′ZetaPlus′ анализаторе зета-потенциала (Brookhaven Instruments, Санта Барбара, шт. Калифорния), как было описано ранее (Bronich et al. (2000) J. Am. Chem. Soc., 122:8339-8343; Vinogradov et al. (1999) Colloids Surf. B-Biointerfaces 16:291-304).
Просвечивающая электронная микроскопия. Капля дисперсии комплекса каталазы и ПЭИ-ПЭГ (Z=1) в фосфатно-солевом буфере была помещена на медную сетку, покрытую формваром (меш 150, Ted Pella Inc., Redding, CA). Высушенная сетка с полипептид-полиионным комплексом была окрашена сульфатом ванадия и визуализирована на просвечивающем электронном микроскопе Philips 201 (Philips/FEI Inc., Briarcliff Manor, NY).
Каталаза и активность каталазы. Активность фермента в полимерных наночастицах была измерена по изменению скорости разложения перекиси водорода каталазой при различных отношениях зарядов и наблюдалась по изменению абсорбции при 240 нм (коэффициент экстинкции перекиси водорода 44×106 М-1 cm1).
Мечение полиионного комплекса каталазы 125I. 125I-меченые полиионные комплексы каталазы были получены путем инкубации раствора белка в ФСБ (1 мг/мл) с Na125I (I мCi) в течение 15 минут в присутствии йодирующего реагента IODO-BEADS (Pierce, Pierce, Rockford, IL) и последующей очистки от несвязавшейся метки хроматографией с высаливанием (Pierce, Rockford, IL). 125I-меченая каталаза (400 µСi/мл, 0.7 мг/мл) была смешана с ПЭИ-ПЭГ блок сополимером (Z=1).
Статистический анализ. Для всех экспериментов данные представлены как средние значения ±СОС. Тесты статистической значимости отклонения между различными группами были сделаны, используя односторонний ANOVA-анализ с множественными сравнениями (попарное сравнение по критерию Фишера), используя GraphPad Prism 4.0_(GraphPad software, San Diego, CA). Минимальное значение р, равное 0,05, было определено как уровень значимости для всех тестов.
Результаты
Блок иономерные комплексы способны спонтанно образовываться при смешивании блок иономеров с противоположно заряженными поверхностно активными веществами или полиэлектролитами (Harada et al. (2001) J. Controlled Release 72:85-91; Kabanov et al. (1995) Bioconjugate Chem., 6:639-643; Harada et al. (1995) Macromolecules 28:5294-5299; Bronich et al. (1997) Macromolecules 30:3519-3525). Нейтрализация зарядов на полиионе приводит к образованию гидрофобных доменов, которые сегрегируют в водных растворах в ядра мицелл полиионных комплексов. Водорастворимые неионные сегменты блок иономеров (например, ПЭГ) предотвращают агрегацию и макроскопическое разделение фаз. В результате эти комплексы самоорганизуются в частицы наноразмера и формируют стабильные водные дисперсии (Рисунок 1А). Каталаза имеет суммарный отрицательный заряд при физиологических условиях. Таким образом, полиионные комплексы были получены в фосфатном буфере (рН 7,4) путем смешивания фермента (1 мг/мл) и положительно заряженного ПЭИ-ПЭГ (2 мг/мл).
Каталаза и комплексы ПЭИ-ПЭГ были сформированы при различных значениях Z (от 0 до 4). Они были подвержены электрофорезу в ПААГ в неденатурирующих условиях и впоследствии перенесены на нитроцеллюлозную мембрану. Белковые полосы были визуализированы антителами против каталазы (Рисунок 1Б). Интенсивность полос уменьшалась с увеличением количества сополимера. Это означает, что размер образующихся комплексов не позволял им войти в гель, что было подтверждено динамическим светорассеянием. Добавление ПЭИ-ПЭГ к раствору каталазы (1 мг/мл) привело к образованию частиц наноразмера с относительно небольшим индексом полидисперсности (примерно 0,1-0,2), тогда как свободная каталаза не образовывала частиц.
Размер частиц зависел от зарядного отношения, ионной силы и рН (Рисунок 1В, 1Г и 1Д). В ФСБ средний диаметр частиц увеличивался с увеличением зарядного отношения и стабилизировался на уровне 90-100 нм при зарядном отношении, равном 1 и выше (Рисунок 1В). Зета-потенциал увеличивался с увеличением количества блок сополимера (Рисунок 1В). При постоянном зарядном отношении (Z=1) и отсутствии солей наблюдалось образование агрегатов размером больше 600 нм (Рисунок 1Г). Добавление соли привело к уменьшению размера частиц, который стабилизировался на 90-100 нм, когда концентрация NaCl достигла 0,15 М. Вероятно образование больших неравновесных агрегатов полиэлектролитных комплексов при смешивании растворов каталазы и ПЭИ-ПЭГ. В бессолевых условиях эти агрегаты не могут уравновеситься и остаются «замороженными» в этом состоянии из-за слабого обмена полиионами (Kabanov, V. (1994) Polym. Sci., 36:143-156; Kabanov, V. (2003) Fundamentals of Polyelectrolyte Complexes in Solution and the Bulk. In Multilayer Thin Films (Decher, G., and Schlenoff, J., Eds.) pp 47-86, Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim). При добавлении соли обмен полиионами ускоряется, что приводит к образованию маленьких равновесных частиц. Эти частицы стабильны в диапазоне рН от 7,4 до 11,5, но необратимо агрегируют при более высоких или низких значениях рН (Рисунок 1Д). Внутри этого диапазона каталаза и ПЭИ-ПЭГ были противоположно заряжены. Агрегация комплексов происходила из-за протонации и инверсии заряда каталазы (рI 6,5) при низких значениях рН или депротонации ПЭИ при высоких значениях рН. Таким образом, частицы полипептид-полиионного комплекса были стабильны при физиологическом рН и ионной силе. При этих условиях по форме частицы были близки к сферическим (Рисунок 1E). Никаких изменений в ферментативной активности каталазы не было обнаружено при зарядовых отношениях, использованных для последующей загрузки клеток, доставки и выделения частиц клетками (Рисунок 1Ж).
Чтобы определить, могут ли полипептид-полиионные комплексы достигать воспаленных участков мозга, отражающих состояние, как при болезни Паркинсона, была использована модель с применением МФТП. МФТП вызывает у людей и не принадлежащих к человеческому роду приматов серьезные и необратимые повреждения мозга, как при болезни Паркинсона (Langston et al. (1986) Clin. Neuropharmacol. 9:485-507). МФТП начинает самоподдерживающийся процесс нигростриальной нейродегенерации (Langston et al. (1999) Ann. Neurol. 46:598-605). У мышей МФТП воспроизводит практически все биохимические и патологические симптомы болезни Паркинсона, включая селективную дегенерацию дофаминэргических нейронов в черной субстанции и стриатуме (Schmidt et al. (2001) J. Neural. Transm. 108:1263-82) and glial inflammation (Gao et al. (2003) Trends Pharmacol. Sci., 24:395-401).
С57В1/6 мышам, интоксицированным МФТП, внутривенно ввели полипептид-полиионные комплексы, содержащие 125I-меченую каталазу. Спустя 24 часа радиоактивность можно было обнаружить в мозгу, а также в других органах.
Пример 2
Для приготовления комплексов бутирилхолинэстеразы (БХЭ) с поли-L-лизин-графт-поли(этиленоксидом), БХЭ/ПЛЛ-г-ПЭО, использовался катионный графт блок иономер ПЛЛ-г-ПЭО (2), содержащий приблизительно 1,4 цепи ПЭО, привитые на основную цепь ПЛЛ. Согласно 1Н-ЯМР анализу молекулярный вес ПЛЛ-г-ПЭО (2) составлял приблизительно 24000 г/моль. В данном исследовании использовались человеческая (чБХЭ) и лошадиная (лБХЭ) бутирилхолинэстеразы.
Комплексы чБХЭ с ПЛЛ-г-ПЭО(2) были приготовлены путем простого смешения растворов блок иономера и белка в буфере (фосфатный буфер, 10 мМ, рН 7,4). Состав смесей при зарядовом соотношении между компонентами, близком к стехиометрическому, был исследован и представлен в таблице 1. Состав смеси был определен как молярное отношение концентраций остатков сиаловой кислоты, входящей в состав БХЭ и аминогрупп ПЛЛ-г-ПЭО(2). (СК/ Л). Состав смесей БХЭ/ПЛЛ-г-ПЭО(2) был также определен через общее количество карбоксильных групп (глутамата, аспартата и сиаловой кислоты) в белке и рассчитан как отношение концентраций аминогрупп в ПЛЛ-г-ПЭО(2) и карбоксильных групп в белке (Z+/-).
Степень включения чБХЭ в блок иономерные комплексы была анализирована с помощью гель-электрофореза в ПААГ в неденатурирующих условиях. На рисунке 2А показан ПААГ различных смесей чБХЭ и ПЛЛ-г-ПЭО(2). Интенсивность полос чБХЭ значительно уменьшалась с увеличением доли сополимера в смеси. Это свидетельствует о связывании сополимера ПЛЛ-г-ПЭО (2) и БХЭ, что приводит к нейтрализации заряда сополимера. При Z+/- = 1,0 наблюдалась практически полная остановка миграции комплекса в геле.
Комплексы лБХЭ и ПЛЛ-г-ПЭО(2) были приготовлены подобным образом. Составы смесей представлены в таблице 2. Степень включения лБХЭ в блок иономерные комплексы была анализирована с помощью гель-электрофореза в ПААГ в неденатурирующих условиях. На рисунке 2Б показан ПААГ различных смесей лБХЭ и ПЛЛ-г-ПЭО (2). Полное включение лБХЭ наблюдалось при использовании избытка блок иономера в смеси (Z+/- = 6.2). Подобные результаты были получены для комплексов лБХЭ и ПЛЛ-г-ПЭО с плотностью графтирования 6,6 цепей ПЭО на одну цепь ПЛЛ (ПЛЛ-г-ПЭО (7)).
Далее комплексы обоих типов БХЭ с ПЛЛ-г-ПЭО(2) были охарактеризованы с помощью динамического светорассеяния. Данные для всех комплексов приведены на рисунке 3. Частицы по размеру немного больше свободного белка были обнаружены во всех смесях блок иономеров и БХЭ.
Молекулярные массы (Mw) комплексов чБХЭ/ПЛЛ-г-ПЭО были измерены с помощью равновесного седиментационного анализа. Все измерения были произведены при 20°С при скорости вращения ротора 4000 оборотов в минуту в течение 24 часов. Полученный профиль равновесного осаждения регистрировался по УФ-абсорбции. Для расчета молекулярного веса по данным равновесной седиментации использованное значение среднего молекулярного объема белка равнялось 0.73 см3/г. Рассчитанные значения молекулярных масс представлены в таблице 3.
Эти данные свидетельствуют о том, что образованные комплексы чБХЭ и ПЛЛ-г-ПЭО включают одну молекулу белка. Наблюдаемое увеличение молекулярной массы по сравнению со свободным белком указывают на то, что на один тетрамер белка в комплексе приходится 2-3 цепи сополимера ПЛЛ-г-ПЭО.
Активность чБХЭ в комплексе была определена с помощью метода, основанного на гидролизе йодида бутирилтиохолина, и представлена в таблице 4. Изменений ферментативной активности BChE в составе комплекса не наблюдалось даже в присутствии избытка блок иономера. Так как для определения ферментативной активности БХЭ используются очень низкие концентрации комплекса, было необходимо показать, что при таких разведениях комплексы сохраняют свою целостность. Различные разбавления комплексов были анализированы ПААГ электрофорезом с последующей окраской по Карновскому-Рутсе (Karnovsky and L. Roots (1964) J. Histochem, Cytochem, 12:219-221). Этот метод «прямой окраски» тиохолина является невероятно чувствительным при очень низких концентрациях БХЭ. Типичный ПААГ представлен на Рисунке 4А. Данные свидетельствуют, что комплексы диссоциируют при больших разбавлениях.
Мультимолекулярная структура типа ядро-оболочка блок иономерных комплексов может быть стабилизирована сшивками между полимерными цепями. Полученные сшитые комплексы являются, по сути, отдельными молекулами наноразмера, которые стабильны при разбавлении и могут выдержать изменения во внешней среде, такие как изменения рН, ионной силы, состава растворителя и силы сдвига, без изменения структуры. Таким образом, для увеличения стабильности БХЭ/блок иономерных комплексов были использованы сшиватели, а именно глутаровый альдегид (ГА), амино-реактивный гомофункциональный сшиватель. Сшивание происходит благодаря образованию иминов (шиффовых оснований) между альдегидными группами ГА и первичными аминогруппами как белка, так и полилизиновых сегментов блок иономера.
Сшивка БХЭ/ПЛЛ-г-ПЭО(2) комплексов (Z+/-=1,2, 0,15 мг/мл в расчете на BChE) осуществлялась в 10 мМ фосфатном буфере (рН 7,4) при добавлении 0,25% водного раствора ГА. Количество ГА было рассчитано, основываясь на желаемой степени сшивки (85%), определяемой как отношение общего количества альдегидных групп в растворе ГА к общему количеству остатков лизина в ПЛЛ-г-ПЭО сополимере. Растворы сшиваемых комплексов инкубировали в течение 5 часов при комнатной температуре. Стабильность сшитых комплексов определялась по методу Карновского-Рутсе. Смеси комплексов были разбавлены соответственно в 1000, 500 и 250 раз. Свободная чБХЭ и исходный несшитый комплекс были разбавлены в той же степени и использовались в качестве контроля. На рисунке 4Б показаны белки и комплексы после разделения электрофорезом в ПААГ. Полосы свободной чБХЭ не были видны в соответствующих дорожках сшитых чБХЭ/ПЛЛ-г-ПЭО комплексов при разбавлении до 1000 раз. В отличие от этого, в разбавленных исходных комплексах наблюдалась полная диссоциация и высвобождение свободной чБХЭ. Полученные результаты свидетельствуют о том, что стабильность блок иономерных комплексов, содержащих чБХЭ в ядре, может быть значительно увеличена путем их сшивания.
Ферментативная активность чБХЭ в составе сшитых комплексов была определена используя йодид бутирилтиохолина в качестве субстрата. Результаты представлены в таблице 5. Данные показывают, что сшивка комплексов чБХЭ/ПЛЛ-г-ПЭО приводит к потере ферментативной активности чБХЭ (активность чБХЭ в сшитом комплексе уменьшилась на 75% по сравнению со свободной чБХЭ). В целом, сшивка ядра комплексов чБХЭ/ПЛЛ-г-ПЭО приводит к значительной устойчивости сшитых комплексов к разбавлению.
Для получения комплексов с различной степенью сшивки к чБХЭ/РЬЬ-д-РЕО(2) комплексам (Z+/-=1.2, 0.15 мг/мл в расчете на BChE) в 10 мМ фосфатном буфере добавляли различные концентрации ГА. 3 мкл раствора ГА различной концентрации добавляли к 120 мкл раствора комплекса, как показано в таблице 6. Количество ГА было рассчитано, исходя из желаемой степени сшивки, определяемой как отношение общего количества альдегидных групп в растворе ГА к общему количеству остатков лизина в ПЛЛ-г-ПЭО сополимере. Стоит заметить, что степень сшивки является теоретической величиной и отображает максимально возможное значение, а не реальное, которое должно быть меньше. Целевая степень сшивки варьировалась от 10% до 100%. Смеси держали при комнатной температуре в течение 5 часов.
Стабильность сшитых комплексов определялась по методу Карновского-Рутсе. Смеси комплексов были разбавлены соответственно в 1000, 5000 и 250 раз. Свободная чБХЭ и исходный несшитый комплекс были разбавлены в той же степени и использовались в качестве контроля. На рисунках 5А и 5Б показаны показательные гели после электрофореза комплексов с различными степенями сшивки (85%, 40% и 20%). Комплексы, приготовленные с целевой степенью сшивки 85% и 40%, были стабильны и не диссоциировали при разбавлении до 1000 раз. Полосы свободной чБХЭ не были видны на дорожках, соответствующих сшитым комплексам чБХЭ/ПЛЛ-г-ПЭО(2) с целевой степенью сшивки 85% (рисунок 5А) и 40% (рисунок 5Б). Разбавление комплексов-предшественников привело к полной диссоциации и высвобождению свободной чБХЭ (дорожки Б). Комплексы, приготовленные с целевой степенью сшивки 20%, частично диссоциировали при разбавлении (Рисунок 5 В). Действительно, при разбавлении в 250 раз на дорожке была видна полоса, соответствующая свободной чБХЭ.
На рисунке 6 представлен ПААГ после электрофореза комплексов чБХЭ/PLL-g-PEO(2) Z+/-=1,2, приготовленных с разными степенями сшивки и разбавленными в 500 раз. Полоса свободной БХЭ была видна на дорожках сшитых комплексов со степенью сшивки 30% и меньше. Данные показывают, что предпочтительная степень сшивки чБХЭ/ПЛЛ-г-ПЭО(2) комплексов должна быть 40% и более, чтобы предотвратить деградацию комплексов при разбавлении.
Молекулярная масса (Mw) сшитых комплексов чБХЭ /ПЛЛ-г-ПЭО(2) была измерена методом равновесного седиментационного анализа. Все имерения были произведены при 20°С при скорости ротора 6000 оборотов в минуту в течение 24 часов. Полученный профиль равновесного осаждения регистрировался по УФ-абсорбции. Для расчета молекулярного веса по данным равновесной седиментации использованное значение среднего молекулярного объема белка равнялось 0.73 см3/г. Рассчитанные значения молекулярных масс представлены в таблице 7. Рассчитанные молекулярные массы сшитых комплексов сравнимы с молекулярными массами комплексов-предшественников. Полученные данные показывают, что реакции сшивки происходили внутри отдельных комплексов и не привели к агрегации и сшивке частиц между собой.
Ферментативная активность чБХЭ в составе сшитых комплексов была определена, используя йодид бутирилтиохолина в качестве субстрата. Результаты представлены в таблице 8. Данные показывают, что сшивка комплексов чБХЭ /ПЛЛ-г-ПЭО влияет на ферментативную активность чБХЭ. Увеличение степени сшивки приводит к снижению ферментативной активности. Например, активность чБХЭ в сшитом комплексе при степени сшивки 85% уменьшилась на 75% по сравнению со свободной чБХЭ, а при 100% сшивании наблюдалась полная потеря активности. С другой стороны при степени сшивки 40% активность уменьшилась всего на 20%. Таким образом, с помощью химической сшивки ядра чБХЭ/ПЛЛ-г-ПЭО можно контролировать стабильность комплексов при разбавлении и при этом сохранять ферментативную активность белка, встроенного в ионное ядро комплекса.
(ед./мг)
Миграция и локализация чБХЭ, доставленной с помощью полимерных комплексов, in vivo была исследована в нуль-зиготных по бутирилхолинэстеразе (ВСhЕ-/-) мышах с помощью оптической визуализации. (ВСhЕ-/-) нокаутные мыши были получены путем направленной делеции части гена чБХЭ (accession number M99492; Li et. al. (2008) J. Pharm. Exp. Ther., 324:1146-1154). лБХЭ была мечена IRDye®800CW красителем (Li-cor, Lincoln, NE), флуоресцирующим в ближней инфракрасной области. По рассчитанной степени мечения на один тетрамер белка приходилась одна молекула красителя. Комплексы, содержащие меченую лБХЭ, были приготовлены путем смешивания 16 мкл раствора лБХЭ/IRDye с 57 мкл раствора ПЛЛ-г-ПЭО(2) (10 мг/мл) и 8 мкл 10-кратного ФСБ (0,1 М фосфатный буфер, 1,4 М NaCI, pH 7,4). Полученные комплексы были затем сшиты глутаровым альдегидом. Количество добавленного ГА было рассчитано исходя из целевой степени сшивки 40%. Смесь инкубировалась при комнатной температуре в течение 5 часов. Сшитый БХЭ/IRDye/ПЛЛ-г-ПЭО(2) комплекс был стабилен при разбавлении, как было показано методом Карновского-Рутсе. Ферментативная активность лБХЭ/IRDye, включенной в сшитый комплекс, уменьшилась на 35%.
Перед визуализацией шерсть животного на дорзальной и вентральной сторонах удалили с помощью крема Nair. Мышей держали на специальной жидкой диете, чтобы уменьшить флуоресценцию из желудка и кишечника, наблюдаемую от стандартного животного корма. Комплексы вводились либо интратекально (ИТ), либо внутримышечно (ВМ). После анестезии мышам вводили меченую свободную лБХЭ либо лБХЭ/IRDye в составе сшитого комплекса. Флуоресценция лБХЭ/ IRDye in vivo измерялась в течение 48 часов с помощью системы IVIS 200. Аккумуляцию лБХЭ/IRDye в мозгу в составе сшитого комплекса можно было наблюдать спустя 2,5 часа после интратекальной инъекции. Флуоресценцию лБХЭ/IRDye в мозге также можно было детектировать спустя 48 часов после ВМ введения комплекса.
Для определения активности лБХЭ, доставленной в мозг, ткани мозга извлекли из эйтанизированных мышей для анализа. Активность лБХЭ была определена используя метод Эллмана (Duysen, et al. (2001) J. Pharm. Exp.Ther. 299:528-535). Единицы активности выражались как количество микромоль бутирилтиохолина, гидролизированных в минуту при pH 7,0 и 25°С. Результаты представлены в таблице 9.
ткани)
Результаты свидетельствуют о том, что БХЭ, доставленная в мозг внутри полимерного комплекса, аккумулировалась и задерживалась в ткани мозга тестированных животных.
Пример 3
Биораспределение CuZnСОД-полиионного комплекса в животных исследовалось, как описано ниже.
Мечение белка. CuZn супероксид дисмутазу (CuZnСОД; 2 мг) растворили в 1 мл ФСБ (0,1 М фосфат калия, 1,5 М хлорид натрия, рН 7,4) при комнатной температуре. рН был увеличен до 8,5 путем добавления 100 мкл 1М фосфатного буфера (K2H2PO4). Полученный раствор добавили к раствору активного красителя, Alexa 680 (Molecular Probes, Inc., Eugene, OR, cat # A-20172), и инкубировали 1 час при перемешивании при комнатной температуре.
Очистка меченой CuZnСОД. Реакционная смесь (1 мл) была нанесена на колонку Sephadex G25 (0.5×26 см), используя фосфатный буфер (10 мМ, рН 7,4), как элюирующий буфер. Меченый белок и несвязавшийся краситель представлены двумя окрашенными полосками. Первая окрашенная полоска (светло-голубая) была собрана через 30 минут в 8 фракций (150 мкл каждая). Концентрация белка, определенная по методу бицинхониновой кислоты, была 0,75 мг/мл. Раствор меченого белка был лиофилизирован и хранился при -20°С.
Приготовление белок-содержащих полиионных комплексов. Чтобы получить CuZnСОД-полиионный комплекс с зарядовым отношением (Z)=2:1, 500 мкл раствора Alexa 680-меченой СОД в физиологическом буфере были добавлены покапельно к 830 мкл раствора блок сополимера полиэтиленимина (ПЭИ) и поли(этиленгликоля) (ПЭИ-ПЭГ, 2 мг/мл) при постоянном перемешивании. Зарядовое отношение (Z) было рассчитано путем деления количества аминогрупп ПЭИ-ПЭГ, протонированных при pH 7,4, на общее количество Gln и Asp в CuZnСОД. Полученный раствор СuZnСОД-полиионного комплекса инкубировали по крайней мере 1 час перед дальнейшим использованием.
Визуализация биораспределения CuZnСОД-полиионного комплекса в мышах. Перед экспериментом мышей анестезировали пентобарбиталом i.p. инъекцией 30-40 мг/кг массы тела, обрили и депилировали (чтобы уменьшить блокирование флуоресценции шерстью). Мышей держали на специальной жидкой диете на протяжении 72 часов (чтобы исключить автофлуоресценцию из желудка и кишечника от твердой еды). Мышам ввели Alexa 680-меченый полиионный комплекс СuZnСОД через хвостовую вену. Затем мыши были анестезированы смесью 1,5% изофлурана, 66% оксида азота и остаточного кислорода и помещены в камеру для визуализации. Биораспределение полиионных комплексов CuZnСОД определялось путем измерения in vivo флуоресценции Alexa 680 с помощью системы IVIS 200. Alexa 680-меченые полиионные комплексы CuZnСОД начинали накапливаться в мозге через час после i.v. инъекции и оставались там, по крайней мере в течение 24 часов (Рисунок 7). Данные свидетельствуют о том, что периферически введенные полиионные комплексы CuZnСОД достигли мозга.
Пример 4
Для получения блок сополимерных комплексов БХЭ были использованы ПЛЛ-ПЭО блок сополимеры. Поли-L-лизин-графт-полиэтиленоксид (ПЛЛ-б-ПЭО) был синтезирован, как описано ранее (см., e.g., Harada et al. (1995) Macromolecules 28:5294). α-Метокси-ω-амино-полиэтиленгликоль с молекулярной массой 5600 г/моль и достаточно узким распределением молекулярного веса (1,27) (Biotech GmbH, Германия) был использован в качестве макроинициатора для синтеза блок сополимера. ПЛЛ-6-ПЭО были охарактеризованы с помощью 1H ЯМР спектроскопии на спектрометра Varian 500 МГц, используя D2O в качестве растворителя. Длина ПЛЛ сегмента была рассчитана равной 25. Рассчитанная молекулярная масса ПЛЛ-г-ПЭО равнялась примерно 24000 г/моль. Этот полимер был определен как ПЛЛ-б-ПЭО. Отношение интенсивности пиков от метиленовых протонов ПЭО (ОСН2СН2:δ=3.62 ppm) и ε-метиленовых протонов ПЛЛ ((СН2)3СН2NH3:δ=2.9 ppm) было измерено для расчета степени полимеризации для ППЛ сегмента и было равно 36. Рассчитанная молекулярная масса ПЛЛ-6-ПЭО приблизительно равнялась 10200 г/моль. Этот полимер был определен как ПЛЛ-б-ПЭО.
Концентрация аминогрупп в растворе ПЛЛ-б-ПЭО была определена методом обратного титрования. Рассчитанная концентрация аминогрупп в растворе 5 мг/мл ПЛЛ-б-ПЭО была равна 6,1 мМ.
Для приготовления комплексов с ПЛЛ-б-ПЭО использовались чБХЭ и лБХЭ. Комплексы были приготовлены путем простого смешивания буферных растворов (10 мМ фосфатный буфер, рН 7,4) блок сополимеров и белка в различных соотношениях и представлены в таблице 10. Состав смесей БХЭ/ПЛЛ-б-ПЭО был выражен в пересчете на общее количество карбоксильных групп (Glu, Asp и сиаловая кислота) в белке и рассчитан как отношение концентраций аминогрупп в ПЛЛ-6-ПЭО и карбоксильных групп в белке (Z+/-).
Степень включения БХЭ в блок иономерные комплексы анализировалась с помощью гель-электрофореза в ПААГ в неденатурирующих условиях. На рисунках 8А и 8Б показана картина гель-электрофореза смесей чБХЭ/ПЛЛ-б-ПЭО и лБХЭ/ПЛЛ-б-ПЭО соответственно. В обоих случаях интенсивность полос БХЭ уменьшалась с увеличением концентрации блок сополимера в смеси. Это свидетельствует о том, что ПЛЛ-б-ПЭО блок сополимер связывался с БХЭ и нейтрализовывал ее заряд. Миграция комплексов обеих смесей была практически полностью остановлена при значениях Z+/- в районе 2,0. Следует заметить, что для образования блок иономерных комплексов БХЭ из лошадиной сыворотки, используя графт-сополимеры (ПЛЛ-г-ПЭО(2) или ПЛЛ-г-ПЭО(7), требовался избыток сополимера в смеси (Z+/-=6.2).
Затем комплексы БХЭ обоих типов и ПЛЛ-б-ПЭО были охарактеризованы с помощью динамического светорассеяния. Данные для всех типов комплексов представлены в таблице 11. Частицы по размеру немного больше свободного белка были обнаружены во всех смесях блок иономеров и БХЭ.
Далее был изучен эффект сшивки ядра комплексов БХЭ/ПЛЛ-б-ПЭО на их стабильность. Для этого был использован глутаровый альдегид (ГА), амино-реактивный гомофункциональный сшиватель. К растворам комплексов чБХЭ/ПЛЛ-б-ПЭО и лБХЭ/ПЛЛ-б-ПЭО (Z+/-=1,0, 0,15 мг/мл в расчете на БХЭ) в 10 мМ фосфатном буфере был добавлен 0,008% водный раствор ГА. Количество ГА было рассчитано, исходя из целевой степени сшивания (40%), определяемой как отношение общего количества альдегидных групп в растворе ГА к общему количеству остатков Lys в ПЛЛ-б-ПЭО сополимере. Растворы комплексов со сшивателем были инкубированы 5 часов при комнатной температуре. Стабильность сшитых комплексов определялась по методу Карновского-Рутсе. Сшитые комплексы были разбавлены в 500 раз. Свободная чБХЭ и исходный несшитый комплекс, разбавленные в той же степени, использовались в качестве контроля. Картина гель-электрофореза представлена на рисунке 9А. Результаты показывают, что комплексы БХЭ/ПЛЛ-б-ПЭО, приготовленные при Z+/-=1,0 и при целевой степени сшивания 40%, не были стабильны при разбавлении, что привело к их диссоциации. Белковая полоса, соответствующая свободной БХЭ, была видна на всех дорожках, соответствующих сшитым комплексам (соответственно дорожки В и Е на рисунке 9А), и их несшитых предшественников (соответственно дорожки Б и Д на рисунке 9А).
В другой группе экспериментов, к комплексам чБХЭ/ПЛЛ-б-ПЭО, приготовленным при Z+/-=2,0, был добавлен 0,016% раствор ГА, чтобы получить 40% сшивания. Сшитые комплексы были разбавлены в 500 раз. Свободная БХЭ и исходные несшитые комплексы использовались в качестве контроля. Картина гель-электрофореза представлена на рисунке 9Б. Как показано на рисунке 9Б, белковая полоса БХЭ не видна на дорожках В и Е, соответствующих сшитым чБХЭ/ПЛЛ-б-ПЭО и лБХЭ/ПЛЛ-б-ПЭО при Z+/-=2,0. В отличие от этого, разбавление комплексов-предшественников привело к полной диссоциации и высвобождению свободной БХЭ (дорожки Б и Д на рисунке 9Б). Таким образом, для успешного сшивания ядра БХЭ/ПЛЛ-б-ПЭО комплекса необходим небольшой избыток блок сополимера.
Ферментативная активность БХЭ в составе несшитых и сшитых БХЭ/ПЛЛ-б-ПЭО комплексов была определена, используя бутирилтиохолин в качестве субстрата. Данные представлены в таблице 12. Ферментативная активность лБХЭ в сшитом комплексе лБХЭ/ПЛЛ-б-ПЭО при Z+/-=2,0 оставалась практически неизменной. Более того, ферментативная активность чБХЭ в сшитом чБХЭ/ПЛЛ-б-ПЭО комплексе не уменьшилась по сравнению с активностью чБХЭ в несшитом комплексе чБХЭ/ПЛЛ-б-ПЭО.
Пример 5
Проникновение в мозг происходит вследствие установления градиента хемокинов, вызванного нейровоспалительными процессами (Kadiu et al. (2005) Neurotox. Res., 8:25-50; Gorantla et al. (2006) J. Leukocyte Biol., 80:1165-1174). Таким образом была разработана модель болезни Паркинсона для тестирования доставки лекарств клетками. Во-первых, с помощью различных воспалительных факторов была вызвана продукция АФК клетками микроглии. Одним из факторов являлся нитрированный α-синуклеин, который считается, что выделяется в межклеточное пространство при болезни Паркинсона и стимулирует активацию микроглии (Gendelman, Н. (2006) Neurotoxicology 27:1162; Mosley et al. (2006) Clin. Neurosci. Res., 6:261-281; El-Agnaf et al. (2003) FASEB J., 17:1945-7). Во-вторых, воспаление, вызванное 1-метил-4-фенил-1,2,3,6-тетрагидропиридином (МФТП), служило направляющей для миграции в мозг макрофагов, генерированных из клеток костного мозга (МККМ). Хорошо известно, что воспалительные факторы вызывают миграцию лейкоцитов в мозг путем диапедеза по хемотаксису (Anthony et al. (1997) Brain 120:435-44; Anthony et al. (2001) Prog. Brain Res., 132:507-24; Blamire et al. (2000) J. Neurosci., 20:8153-9; Persidsky et al. (1999) Am. J. Pathol., 155:1599-611; Kuby, J. (1994) Immunology; Freeman, WH. and Co., New York). Моноциты могут мигрировать по межклеточным пространствам в мозге и могут проходить через межклеточные контакты клеток эндотелия мозга (Pawlowski et al. (1988) J. Exp. Med., 168:1865-82; Lossinsky et al. (2004) Histol. Histopathol., 19:535-64). Они могут поглощать чужеродные частицы и затем высвобождать их путем экзоцитоза. Таким образом, эти свойства позволяют использовать макрофаги в качестве носителей для лечения нейровоспалительных процессов (Daleke et al. (1990) Biochim. Biophys. Acta 1024:352-66; Lee et al. (1992) Biochim. Biophys. Acta 1103:185-97; Nishikawa et al. (1990) J. Biol. Chem., 265:5226-31; Fujiwara et al. (1996) Biochim. Biophys. Acta 1278:59-67).
В данном случае МККМ использовались как носители для доставки фармацевтических концентраций каталазы в мозг. Главной проблемой этого подхода является то, что макрофаги эффективно разрушают захваченные честицы (Fujiwara et al. (1996) Biochim. Biophys. Acta 1278:59-67). Следовательно, активность фермента внутри клетки-носителя необходимо защитить. Этого можно достичь путем включения белка в полимерные наноносители (наносферы, липосомы, мицеллы, наночастицы) (Aoki et al. (2004) Int. J. Hypertherm., 20:595-605; Calvo et al. (2001) Pharm. Res., 18:1157-1166; Gref et al.; (1994) Science 263:1600-1603; Harada et al. (1999) Science 283:65-7; Jaturanpinyo (2004) Bioconjugate Chem., 15:344-8; Kabanov et al. (2002) J. Controlled Release 82:189-212; Kwon, G.S. (2003) Crit. ReV. Ther. Drug Carrier Syst., 20:357-403; Mora et al. (2002) Pharm. Res., 19:1430-8; Rousseau et al. (1999) Exp.Brain Res., 125:255-64; Torchilin, V.P. (2000) Eur. J. Pharm. Sci., 11:S81-91; Vinogradov et al. (2004) Bioconjugate Chem., 15:50-60). Ранее было показано, что полиэлектролитные комплексы могут связывать ферменты (Kabanov et al. (1977) Mol. Biol. (Russian), 11:582-596; Kabanov, V. (1994) Polym. Sci., 36:183-197; Kabanov et al. (2004) J. Phys. Chem. B, 108:1485-1490). Полиэлектролитные комплексы ферментов наноразмера могут быть получены путем самоорганизации ферментов с противоположно заряженными блок полиэлектролитами, содержащими ионный и неионный водорастворимый блоки (Harada et al. (2001) J. Controlled Release 72:85-91; Harada et al. (2003) J. Am. Chem. Soc., 125:15306-7). Полученные наночастицы содержат ядро, состоящее из комплекса белка и полиэлектролита, которое окружено короной из водорастворимого неионного полимера, например полиэтиленгликоля (ПЭГ). В настоящей работе комплексы каталазы были получены путем реакции катионного блок сополимера, полиэтиленимина-полиэтиленгликоля (ПЭИ-ПЭГ), который ранее использовали для доставки полинуклеотидов (Vinogradov et al. (1998) Bioconjugate Chem., 9:805-812).
Полученные блок иономерные комплексы каталазы могут быть поглощены МККМ. Доказательством того, что подобная модификация может защитить каталазу от деградации, служит то, что впоследствии МККМ высвобождают эти комплексы во внешнюю среду на протяжении по меньшей мере 4-5 дней и что они могут переносить каталазу в воспаленные участки мозга, как например, в МФТП модели болезни Паркинсона.
Материалы и методы
Материалы, как в примере 1.
МККМ. Клетки костного мозга были выделены из бедренной кости самок C57BL/6 мышей, как описано ранее (Dou et al. (2006) Blood 108:2827-35). Клетки культивировали 10 дней в среде, содержащей 1000 ед./мл фактора, стимулирующего колонии макрофагов (ФСКМ) (Wyeth Pharmaceutical, Cambridge, MA). Чистоту культуры макрофагов определяли с помощью проточной цитометрии на FACSCalibur (BD Biosciences, San Jose, CA).
Микроглия. Мозги, выделенные из неонатальных C57BL/6 мышей (1-3 дней от роду), отмыли холодным сбалансированным солевым раствором Хенкса (ССРХ) и измельчили. Супернатант заменили 2,5% раствором трипсина и ДНКазы (1 мг/мл) и инкубировали 30 минут при 37°С и затем добавили 10 мл 10% термоактивированной эмбриональной телячьей сыворотки в ССБХ. Затем смесь осадили центрифугированием (5 минут, 1500 оборотов в минуту, 4°С) и перевели в обогащенную среду культивирования с добавлением ФСКМ. Клетки культивировали до созревания (обычно 10 дней).
МФТП, как в примере 1.
ПЭИ-ПЭГ конъюгаты, как в примере 1.
Блок иономерные комплексы, как в примере 1.
ПААГ электрофорез, как в примере 1.
Динамическое светорассеяние, как в примере 1.
Просвечивающая электронная микроскопия, как в примере 1.
Каталаза и ее активность, как в примере 1.
Мечение каталазы Alexa Finer 594 и изотиоционатом родамина (РИТЦ). Для изучения загрузки и выделения каталазы она была помечена с помощью Alexa Fluor 594 кита для мечения белков (А10239, Molecular probes, Inc., Eugene, OR) согласно протоколу производителя. Для конфокальной микроскопии каталаза была помечена РИТЦ. Коротко, к раствору каталазы (1 мг/мл) в 0,1М натрий-фосфатном буфере, рН 8,5 добавили раствор РИТЦ (10 мг/мл) в ДМСО и инкубировали 2 часа при комнатной температуре. Меченую каталазу очистили от низкомолекулярных примесей путем гель-фильтрации на колонке Sephadex G-25 (1×20 см) в ФСБ при скорости элюации 0,5 мл/мин и затем лиофильно высушили.
Загрузка и выделение полипептид-полиионных комплексов в МККМ. МККМ растили на 24-луночных плашках (2.5×106 клеток/лунку) (Batrakova et al. (1998) Pharm. Res., 15:1525-1532; Batrakova et al. (2005) Bioconjugate Chem., 16:793-802). Затем их преинкубировали в экспериментальном буфере (122 мМ NaCl, 25 мМ NаНСО3, 10 мМ глюкозы, 3 мМ КСl, 1.2 мМ MgSO4, 0.4 мМ K2HPO4, 1.4 мМ CaCl2, and 10 мМ HEPES) в течение 20 минут. После преинкубации к клеткам добавили свободную Alexa-Fluor 594 меченую каталазу (0,7 мг/мл) в экспериментальном буфере или полипептид-полиионный комплекс в различные моменты времени. Затем клетки отмыли 3 раза холодным ФСБ и растворили в 1% растворе Тритона Х 100. Для измерения количества выделенного из МККМ полипептид-полиионного комплекса нагруженные МККМ инкубировали в свежей среде в течение различных периодов времени. Флуоресценция в каждом образце измерялась на флуоресцентном спектрофотометре Shimadzu RF5000 (λвозб=580 nm, λисп=617 nm). Количество полипептид-полиионных комплексов было нормализовано на общее количество белка и выражено в мкг фермента на мг белка в экспериментах по загрузке и в мкг фермента в мл среды в экспериментах с выделением комплексов. Данные представлены как средние значения ± СОС (n=4).
Внутриклеточная локализация полипептид-полиионных комплексов. Моноциты были выращены на предметных стеклах для конфокальной микроскопии (Kabanov et al. (1995) Bioconjugate Chem., 6:639-643) и инкубированы с РИТЦ-мечеными полипептид-полиионными комплексами (Z=1) в течение 2.4 часов при 37°С. Затем клетки зафиксировали в 4% растворе параформальдегида и покрасили F-актин-специфичным Oregon Green 488-меченым фаллоидином и ядерным красителем ToPro-3 (Molecular Probes, Inc., Eugene, OR).
Измерения антиокислительной активности. Зрелые мышиные МККМ нагружали либо свободным ферментом, либо фермент-полиионным комплексом (Z=1) в течение одного часа, отмыли ФСБ и добавили свежую среду. Через различные промежутки времени среду собирали и измеряли в ней антиокислительную активность фермента, выделившегося из МККМ, по скорости распада перекиси водорода.
Флуоресцентный метод с использованием красителя Ampex Red. Мышиные микроглиальные клетки были выращены в 96-лучных плашках (0.1×106 клеток/лунку). Клетки стимулировали либо фактором некроза опухоли альфа (TNF-α) (200 нг/мл) в течение 48 часов, либо нитрированным альфа-синуклеином (N-α-syn) (0,5 мкМ), чтобы вызвать продукцию АФК. Параллельно, МККМ, выращенные в 24-лучных плашках, нагружали либо свободной каталазой (1 мг/мл), либо полиионным комплексом каталазы в течение одного часа и затем 2 часа ингибировали в буфере Кребса-Рингера (145 мМ NaCl, 4,86 мМ КСl, 5,5 мМ глюкозы, 5,7 мМ NaH2PO4, 0,54 мМ CaCl2, 1,22 мМ MgCl2, pH 7,4), чтобы собрать выделившуюся во внешнюю среду каталазу. После инкубации к супернатантам от клеток, нагруженных свободной каталазой или полиионным комплексом каталазы, был добавлен стоковый раствор красителя Ampex Red (10 U/mL HRP, 10 mM Ampex Red). В случае, когда микроглия была стимулирована N-α-syn, в супернатанты было также добавлено 0,5 мкМ агрегированного N-α-syn. Полученные растворы были добавлены к активированной микроглии, после чего разложение АФК свободной каталазой или каталазой в полиионном комплексе измерялось по флуоресценции при λвозб=563 nm, λисп=587 nm. Эффект действия супернатантов, собранных от ненагруженных клеток или нагруженных только ПЭИ-ПЭГ на распад АФК, сравнивали с контрольными экспериментами.
Мечение полипептид-полиионного комплекса каталазы 125I. Как в примере 1. 125I-меченую каталазу (400 µСi/мл, 0,7 мг/мл) смешали с ПЭИ-ПЭО блок сополимером (Z=1). Полученными комплексами нагружали зрелые моноциты (80×106 МККМ в 1 мл среды) в течение 2 часов при 37°С. После инкубации нагруженные моноциты отмыли 3 раза холодным ФСБ.
Статистический анализ, как в примере 1.
Результаты
Получение полипептид-полиионных комплексов описано выше в примере 1. Исследования клеточной выживаемости с помощью сульфородамина В (SRB) показали, что полипептид-полиионные комплексы (так же как и свободные каталаза и сополимер) не вызывали цитотоксичность в МККМ в широком диапазоне концентраций (от 0,03 до 1000 мкг/мл каталазы; Рисунок 10). Кинетика аккумуляции свидетельствует о быстром захвате клетками (МККМ) как свободной каталазы, так и полипептид-полиионного комплекса (Рисунок 11А). Следует заметить, что свободный фермент захватывался макрофагами почти в 2 раза быстрее по сравнению с полипептид-полиионным комплексом. Через час инкубации примерно 30 мкг каталазы в составе полипептид-полиионного комплекса/106 клеток было захвачено макрофагами. Захват полипептид-полиионного комплекса за 60 минут уменьшился с увеличением зарядового отношения (Рисунок 11Б), что может быть следствием эффекта короны ПЭГ. Конфокальные микрофотографии показывают везикулярную или цитоплазматическую локализацию РИТЦ-меченой каталазы, добавленной к МККМ в виде полипептид-полиионных комплексов (Рисунок 11В).
Зрелые МККМ сначала нагрузили Alexa Fluor 594-меченым полиионным комплексом каталазы (в течение 60 минут) и затем культивировали в свежей среде на протяжении различного времени. Нагруженные МККМ высвобождали каталазу во внешнюю среду в течение по меньшей мере 4-5 дней (Рисунок 12А). В течение того же времени количество фермента, связанного с клетками, пропорционально уменьшалось. Действие 10 мкМ форбол миристат ацетата (ФМА), сильного активатора сигнального пути протеин киназы С и стимулятора продукции АФК (Chang et al. (1993) Immunology 80:360-366) на МККМ, нагруженные полипептид-полиионным комплексом, вызывало усиление выделения фермента в среду на примерно 50% (Рисунок 12Б). Это свидетельствует о том, что выделение полипептид-полиионного комплекса МККМ может зависеть от клеточной активации.
МККМ, нагруженные свободной каталазой или полиионным комплексом каталазы, поместили в свежую среду, после чего через различные промежутки времени определяли активность каталазы в среде. Противоположно случаю, когда МККМ были нагружены свободной каталазой, которая полностью теряла активность после выделения в среду, МККМ, нагруженные полипептид-полиионным комплексом, выделяли активный фермент в течение по меньшей мере 24 часов (Рисунок 13А). Максимальная активность выделившегося фермента наблюдалась, когда МККМ были нагружены полипептид-полиионным комплексом, приготовленным в стехиометрическом соотношении (Z=1) (Рисунок 13Б). Таким образом, данные показывают, что включение каталазы в блок иономерный комплекс с ПЭИ-ПЭГ предохраняет фермент, который впоследствии выделяется из клеток в активной форме.
Антиокислительную емкость наноформуляций каталазы оценивали по подавлению выработки АФК клетками микроглии. Для этого МККМ, нагруженные либо свободной каталазой, либо полипептид-полиионным комплексом, инкубировали 2 часа в буфере Кребса-Рингера, после чего полученный супернатант добавили к клеткам микроглии, активированным TNF-α (200 нг/мл). Таким образом, было показано, что каталаза, содержащаяся в супернатантах, собранных от МККМ, нагруженных свободной каталазой или полиионным комплексом каталазы, способна разлагать пероксид водорода, выделяемый микроглиальными клетками (Рисунок 14А). Больший эффект был показан для полиионного комплекса каталазы, что согласуется со способностью комплекса предохранять фермент внутри клеток-носителей. При этом супернатанты, собранные от ненагруженных МККМ (Рисунок 14Б) или от нагруженных только ПЭИ-ПЭГ (Рисунок 14В), практически не имели никакого эффекта на уровне перекиси водорода. Чтобы определить, будут ли эти эффекты воспроизводиться в микроглии, активированной факторами, типичными для болезни Паркинсона, клетки стимулировали 0,5 мкМ N-α-syn. При болезни Паркинсона агрегированный N-α-syn представляет собой цитоплазматические включения, которые выделяются после смерти дофаминэргических нейронов и являются основным компонентом телец Леви (Zhang et al. (2005) FASEB J., 19:533-42). Считается, что эти агрегированные белки служат стимулом для активации микроглии (Gendelman, H. (2006) Neurotoxicology 27:1162; Thomas et al. (2007) J. Neurochem. 100:503-19). В данном случае концентрация пероксида водорода также была значительно снижена при добавлении супернатантов МККМ, нагруженных полиионными комплексами каталазы (Рисунок 14Г). Таким образом, настоящее исследование показывает, что полиионные комплексы каталазы, выделившиеся из МККМ, могут смягчать окислительный стресс, вызванный активацией микроглии. Действительно, полиионные комплексы каталазы, выделившиеся из МККМ, способны снижать количество перекиси водорода гораздо более эффективно, чем свободная каталаза, что подтверждает, что полиионный комплекс эффективно предохраняет ферментативную активность каталазы в МККМ.
Для того чтобы определить, могут ли МККМ, нагруженные полиионными комплексами каталазы, достигать воспаленные участки мозга, как в болезни Паркинсона, была использована МФТП модель. Двум группам С57В1/6 мышей, интоксицированных МФТП, внутривенно вводили либо только полипептид-полиионный комплекс, содержащий 125I-меченую каталазу, либо МККМ, нагруженные полиионным комплексом каталазы. Спустя 24 часа наблюдалось значительное увеличение радиоактивности в селезенке, печени, легких, почках и мозге в группе мышей, которым ввели МККМ, содержащие полипептид-полиионный комплекс по сравнению с мышами, которым ввели только полипептид-полиионный комплекс (Рисунок 15). Стоит заметить, что после адоптивного переноса полипептид-полиионного комплекса макрофагами примерно 0,6% введенной дозы было обнаружено в мозге, что было в два раза больше, чем у мышей, которым внутривенно вводили свободный полипептид-полиионный комплекс. Таким образом, полученные данные свидетельствуют о том, что адоптивно пересаженные МККМ, нагруженные полипептид-полиионным комплексом, увеличивают доставку фермента в мозг, а также в другие периферические ткани, которые являются местами миграции макрофагов.
Эффективный транспорт терапевтических полипептидов в мозг необходим для успешного лечения нейровоспалительных заболеваний. Для этого было исследовано, могут ли МККМ быть использованы как носители для доставки каталазы, мощного антиокислителя. Давно известно, что макрофаги и микроглия, а также другие мононуклеарные фагоциты, могут захватывать коллоидные наночастицы, такие как липосомы или наносуспензии, и впоследствии переносит и выделять препарат в области повреждения ткани, воспаления или заболевания (Dou et al. (2006) Blood 108:2827-35; Dou et al. (2007) Virology 358:148-158; Gorantla et al. (2006) J. Leukocyte Biol., 80:1165-1174; Daleke et al. (1990) Biochim. Biophys. Acta 1024:352-66; Jain et al. (2003) Int. J. Pharm., 261:43-55).
Более того, была также изучена способность МККМ пересекать гематоэнцефалический барьер (Lawson et al. (1992) Neuroscience 48:405-15; Sirtard et al. (2004) FASEB J., 18:998-1000; Male et al. (2001) Prog. Brain Res., 132:81-93; Streit et al. (1999) Prog. Neurobiol., 57:563-81; Kokovay et al. (2005) Neurobiol. Dis., 19:471-8; Kurkowska-Jastrzebska et al. (1999) Acta Neurobiol. Exp.(Wars) 59:1-8; Kurkowska-Jastrzebska et al. (1999) Exp. Neurol., 156:50-61; Simard et al. (2006) Mol. Psychiatry 11:327-35). В частности, было показано, что моноциты проникают в мозге в МФТП модели болезни Паркинсона (Kokovay et al. (2005) Neurobiol. Dis., 19:471-8; Kurkowska-Jastrzebska et al. (1999) Acta Neurobiol. Exp.(Wars) 59:1-8; Kurkowska-Jastrzebska et al. (1999) Exp. Neurol., 156:50-61). МФТП вызывает временное повсеместное увеличение скорости инфильтрации моноцитов в средний мозг, страту, септу и гиппокамп. В этих исследованиях наибольшая аккумуляция моноцитов-макрофагов наблюдалась на следующий день после введения МФТП. Основываясь на этом, можно предположить, что адоптивно пересаженные моноциты, нагруженные каталазой, могут доставлять фермент в наиболее поврежденные участки мозга при болезни Паркинсона, включая черную субстанцию и стриатум в мышах, подвергшихся действию МФТП.
Чтобы защитить каталазу от деградации внутри МККМ, белок иммобилизировали в блок иономерный комплекс с катионным блок сополимером, ПЭИ-ПЭГ. Полученные наночастицы имели размер 60-100 нм и были стабильны при физиологических условиях (рН, ионная сила). Чтобы достигнуть наибольшей загрузки в МККМ и сохранить ферментативную активность каталазы, состав и структуру полиионных комплексов каталазы можно варьировать. Захват чужеродных частиц, так же как их последующее высвобождение, является одной из основных функций макрофагов (Stout et al. (1997) Front. Biosci., 2:dl97-206). Здесь было показано, что МККМ могут захватывать значительные количества полипептид-полиионного комплекса (приблизительно 30 мкг каталазы/106 клеток) в относительно короткий промежуток времени (40-60 минут), после чего высвобождать его во внешнюю среду на протяжении 4-5 дней. Это позволяет сделать предположение, что клетки, нагруженные полипептид-полиионным комплексом, после адоптивной пересадки могут иметь достаточно времени, чтобы достигнуть мозга и высвободить каталазу. Более того, было показано (Schorlemmer et al. (1977) Clin. Exp. Immunol., 27:198-207; Allison et al. (1974) Symp. Soc. Exp. Biol., 419-46; Cardella et al. (1974) Nature 247:46-8), что экзоцитоз можно стимулировать активацией моноцитов и макрофагов. Вышеописанные эксперименты показывают, что выделение полипептид-полиионного комплекса клетками может быть усилено при стимуляции ФМА. Также было показано, что блок иономерный комплекс предохраняет ферментативную активность каталазы внутри клетки-хозяина. Причем клетки, нагруженные полиионным комплексом фермента, выделяли активный фермент в среду на протяжении по меньшей мере 24 часов. Более того, культуральные супернатанты, собранные от клеток, нагруженных полипептид-полиионными комплексами, имели сильный антиокислительный эффект на АФК, выделенные микроглией, которая была активирована либо N-α-syn, либо TNF-α. Таким образом, на моделях клеточных культур было показано, что МККМ, нагруженные полипептид-полиионным комплексом, могут ослаблять окислительный стресс, связанный с нейродегенеративными процессами. Наконец, in vivo было показано, что адоптивная пересадка МККМ, нагруженных полипептид-полиионными комплексами, может увеличить количество доставленного меченого фермента в ткани, включая двоекратное увеличение количества доставленного фермента в мозгу мышей, подвергшихся действию МФТП. Интересно, что значительное количество меченого фермента было обнаружено в мозгу после инъекции свободного полипептид-полиионного комплекса. Возможно, полипептид-полиионный комплекс захватывается циркулирующими моноцитами, которые могут доставить фермент в мозг.
Пример 6
In vivo визуализация с помощью системы визуализации in vivo (IVIS). BALB/C мыши получили инъекцию МФТП (чтобы вызвать нейровоспаление, как в болезни Паркинсона), после чего их обрили (чтобы уменьшить блокировку флуоресценции шерстью). МККМ, нагруженные Alexa Fluor 680-меченым полипептид-полиионным комплексом (ПЭИ-ПЭГ, Z=1), были введены внутривенно мышам, подвергшимся действию МФТП (50 млн/мышь). Мыши были визуализированы с помощью IVIS на протяжении различного времени (Рисунок 16). Значительное количество полипептид-полиионного комплекса было обнаружено в интоксицированном МФТП мозге. Стоит заметить, что в контрольной группе мышей, которым не вводили МФТП, не было зафиксировано флуоресценции в мозге, что указывает на то, что МККМ усиливали доставку полипептид-полиионного комплекса через гематоэнцефалический барьер в воспаленные участки мозга.
Гистопатологическая оценка in vivo токсичности МККМ, нагруженных полипептид-полиионным комплексом. Здоровым C57BL/6 мышам ввели либо МККМ, нагруженные полипептид-полиионным комплексом (10 млн/мышь), либо ФСБ (контроль). Спустя 48 часов мозг, печень, селезенка и почки были взяты для некропсии. Органы были окрашены гематоксилином и эозином и анализированы на световом микроскопе. При этом не было обнаружено признаков апоптоза, нарушения гематоэнцефалического барьера, нейровоспалительного ответа в нейронах; макровезикулярного стеатоза и некроза в гепатоцитах; признаков холестаза в печени; или признаков острого трубчатого некроза в почках.
Нейрозащитный эффект МККМ, нагруженных полипептид-полиионным комплексом, против вызванной МФТП потери дофаминэргических нейронов в мышах. Чтобы оценить нейрозащитный эффект полипептид-полиионного комплекса, интоксицированным МФТП мышам внутривенно ввели МККМ, нагруженные полипептид-полиионным комплексом. На 7 день после инъекции были измерены уровни N-ацетил аспартата (NAA) (метаболита нейронов) в черной субстанции и стриатуме (наиболее пораженные области мозга при болезни Паркинсона). Инъекция МФТП вызвала значительное снижение уровня NAA в черной субстанции и стриатуме контрольной группы мышей (Рисунок 17). В противоположность этому, снижение уровня NAA не было обнаружено в интоксицированных МФТП мышах, которым ввели МККМ, нагруженные полипептид-полиионным комплексом. В дополнительных экспериментах было показано снижение уровня воспаления, измеренного как отношение уровня астроцитоза в экспериментальных мышах к уровню астроцитоза в контроле через 2 дня. Данные указывают на то, что полиионный комплекс каталазы обладает нейрозащитным действием при нейродегенерации, вызванной МФТП.
Пример 7
Периферическое введение CuZnСОД-полиионного комплекса ингибирует острый ответ кровяного давления при центральном введении AngII. Чтобы показать эффективность периферического введения CuZnСОД-полиионного комплекса для модуляции сигнального пути AngII в мозге, был использован CuZnСОД-полиионный комплекс, описанный в примере 3. В частности, был исследован эффект периферического введения (через сонную артерию) CuZnСОД-полиионного комплекса на острое увеличение кровяного давления, вызванное AngII (100 нг), введенного интрацеребровентрикулярно (ICV). Вызванные таким образом изменения среднего артериального давления (САД) были регистрированы в кроликах через 0, 1, 2 и 5 дней после ICV введения CuZnСОД-полиионного комплекса или свободной CuZnСОД. Изменения САД после ICV введения AngII были значительно снижены через 1 и 2 дня после лечения СuZnСОД-полиионным комплексом по сравнению с ответом в день 0 (Рисунок 18). В противоположность этому, лечение свободной CuZnСОД, которая является активной, но не может пересечь гематоэнцефалический барьер, не имело эффекта на изменение кровяного давления, вызванное ICV введенным AngII (Рисунок 18). Данные показывают, что периферически введенный CuZnСОД-полиионный комплекс способен проникать в AngII-чувствительные нейроны в ЦНС и модулировать AngII-опосредованные кардиоваскулярные ответы. Действительно, в конкретном варианте осуществления настоящего изобретения представлены методы лечения повышенного кровяного давления у пациентов, которые включают введение смеси, содержащей: а) по меньшей мере один комплекс, содержащий CuZnСОД и синтетический полимер, содержащий по меньшей мере один заряд, противоположный заряду CuZnСОД, и б) по меньшей мере один фармацевтически приемлемый носитель. В конкретном варианте осуществления настоящего изобретения комплекс, содержащий СuZnСОД и синтетический полимер, содержащий по меньшей мере один заряд, противоположный заряду СuZnСОД, находится внутри клеток, которые вводятся пациенту.
Пример 8
Нейротрофический фактор головного мозга (BDNF) является основным нейротрофическим белком с молекулярным весом 27,3 кДа с изоэлектрической точкой 10,23. BDNF имеет суммарный положительный заряд (+10,23) при нейтральном рН (Philo et. al. (1994) J. Biol. Chem., 269:27840-27846). Поэтому для включения BDNF в полиионный комплекс был использован анионный блок сополимер, ПЭО-6-поли(метакрилат натрия) (ПЭО-г-ПМА) (рК карбоксильной группы 5,2). Комплексы были приготовлены путем простого смешения забуференных водных растворов блок сополимерного и белкового компонентов. Отношение полимер/белок в смесях было рассчитано путем деления суммарной рассчитанной концентрации карбоксильных групп в ПЭО-6-ПМА на суммарную концентрацию остатков Lys и Arg в белке. При смешивании растворы оставались прозрачными и выпадение осадка не наблюдалось.
Герцептин (трастузумаб) является гуманизированными античеловеческими моноклональными антителами против рецептора эпидермального фактора роста 2 (HER2/c-erbB2). Было показано, что герсептин эффективен против первичных и экстракраниальных метастатических раков молочной железы, которые сверхэкспрессируют HER2. Тем не менее, у пациентов с метастазами в мозге, гематоэнцефалический барьер ограничивает его использование (Kinoshita et. al. (2006) PNAS, 103:11719-11723).
Герсептин является основным белком с молекулярной массой 145,5 кДа и изоэлектрической точкой 8,45. Герсептин имеет суммарный положительный заряд (+12) при нейтральном рН. Для инкорпорации герсептина в полиионный комплекс был использован анионный блок сополимер, ПЭО-б-поли(метакрилат натрия) (ПЭО-6-ПМА). Комплексы были приготовлены путем простого смешения забуференных водных растворов блок сополимерного и белкового компонентов. Отношение полимер/белок в смесях было рассчитано путем деления суммарной рассчитанной концентрации карбоксильных групп в ПЭО-б-ПМА на суммарную концентрацию остатков Lys и Arg в белке. При смешивании растворы оставались прозрачными и выпадение осадка не наблюдалось.
Лептин является белковым гормоном с молекулярной массой 18,7 кДа, который играет ключевую роль в регуляции поглощения энергии, включая регуляцию (снижение) аппетита и (усиление) метаболизма. Лептин имеет изоэлектрическую точку 5,85 и суммарный отрицательный заряд (приблизительно -2) при физиологическом рН. Для получения лептин-полиионного комплекса был использован катионный блок иономер с графт-архитектурой, поли-L-лизин-графт-полиэтилегоксид, ПЛЛ-г-ПЭО(2), содержащий приблизительно 1,4 цепей ПЭО, привитых на основную цепь полииона. Комплексы были приготовлены путем простого смешения забуференных водных растворов графт-сополимерного и белкового компонентов. Отношение полимер/белок в смесях было рассчитано путем деления суммарной рассчитанной концентрации карбоксильных групп в ПЛЛ-г-ПЭО(2) на суммарную концентрацию остатков Asp и Glu в белке. При смешивании растворы оставались прозрачными и выпадение осадка не наблюдалось.
Пример 9
Предотвращение воспаления в МФТП-интоксицированных мышах моноцитами, нагруженными полиионными комплексами каталазы
Чтобы вызвать патологические изменения, характерные для болезни Паркинсона, самцам C7BL/6 мышей-реципиентов было введено 18 мг МФТП/кг веса тела в ФСБ посредством 4 интраперитониальных инъекций, производимых каждые 2 часа (МФТП (Sigma Chemical Co., St. Louis, МО)). Контрольной группе мышей внутривенно вводили физраствор спустя 18 часов, половине МФТП-интоксицированных мышей внутривенно ввели моноциты, нагруженные полиионным комплексом каталазы (10 млн/мышь).
Другой половине мышей внутривенно вводили физраствор. Активная фаза смерти нейронов и пик нейровоспалительной активности наблюдались примерно через 2 дня после инъекции МФТП. Таким образом, два дня спустя области среднего мозга были выделены из контрольных мышей, интоксицированных МФТП, и из мышей, которых после инъекции МФТП лечили моноцитами, нагруженными каталазой. Мозги были мгновенно заморожены и погружены в среду для окситоциновой пробы. Иммуногистохимический анализ проводился на интактных срезах толщиной 30 мкм, фиксированных в 4% параформальдегиде в течение 24 часов и постфиксированных в сахарозе в течение 48 часов при 4°С. Срезы ткани хранили в 0,01% растворе азида натрия в ФСБ. Перед визуализацией срезы отмыли 3 раза ФСБ. Затем срезы блокировали 7% нормальной козьей сывороткой (НКС) в течение 1 часа.
Для визуализации активированной микроглии (окраска Мас-1) срезы тканей были иммуноокрашены крысиными CD11b первичными антителами (AbD Serotec, Raleigh, NC), разведенными 1:200 в 7% НКС в течение ночи при 4°С. Образцы были инкубированы со вторичными козьими антикрысиными антителами, меченными Alexa Fluor 594 (Invitrogen Corporation, Carlsbad, CA), разбавленными 1:200 в 7% НКС в течение 45 минут при комнатной температуре.
Для визуализации астроцитов:,, срезы ткани были пермеабилизированы 1% Тритоном Х100 в 5% НКС в ФСБ в течение 10 минут и блокированы в течение часа 5% НКС. Затем срезы были инкубированы с кроличьими первичными антителами к антиглиальному фибриллярному кислому белку, разбавленными 1:1000 в 5% НКС в течение 18 часов при 4°С. Образцы затем были инкубированы с козьими антикроличьими вторичными антителами, меченными Alexa Fluor 488 (Molecular Probes), разбавленными 1:200, в течение 45 минут при комнатной температуре. Срезы были заключены в среду Aquamount. Иммунореактивность была анализирована путем флуоресцентного анализа. Интенсивность флуоресценции была рассчитана при помощи программного обеспечения ImageJ (National Institute of Health; NIH). Площадь была измерена как функция уровня экспрессии CD11b, используя ImageJ (National Institute of Health; NIH).
Данные, представленные в таблице 13, отчетливо показывают, что инъекции МФТП вызывают сильные воспалительные процессы в черной субстанции и приводят к активации микроглии и астроцитозу. В противоположность этому, лечение МФТП интоксицированных мышей моноцитами, нагруженными каталазой, предотвращало нейровоспаление на уровне здоровых животных.
Нейрозащитный эффект моноцитов, нагруженных полиионным комплексом каталазы, против потери дофаминэргических нейронов, вызванной МФТП в мышах
Чтобы количественно и неинвазивно определить нейрозащитный эффект каталазы в черной субстанции и стриатуме после интоксикации мышей МФТП, которая привела к прогрессии болезни Паркинсона, была использована новая технология нейровизуализации, которая позволяет оценивать уровни N-ацетил аспартата (NAA) в нейронах с помощью магнитной резонансной спектроскопической визуализации (magnetic resonance spectroscopic imaging (MRSI)).
Для этого мыши сначала были сканированы до введения МФТП. Потом половине мышей ввели нагруженные полиионным комплексом каталазы МККМ (25 млн МККМ/100 мкл/мышь). Другой половине ввели ФСБ и использовали в качестве контроля для определения максимума нейродегенерации. Уровень метаболита нейронов мозга, N-ацетил аспартата (NAA), в черной субстанции и стриатуме был измерен с помощью MSRI на седьмой день после лечения. МРТ и MRSI осуществлялись на системе Bruker Avance 7T/21 см, работающей при 300,41 МГц, используя активно разобщенную 72 мм объемную передающую катушку и сделанную в лаборатории 1,25×1,5 см принимающую поверхностную катушку. Магнитно-резонансные изображения были получены при следующих параметрах: размер поля сканирования - 20 мм, количество сечений - 25, толщина сечений - 0,5 мм в чередующемся порядке, размер матрицы - 128×128. Построение Т2 томограммы и ко-регистрацию с гистологией проводили по 8 изображениям от 8 отраженных сигналов (интервал между импульсами 12 мс), рефокусированных при помощи последовательности КПМГ импульсов РЧ. Спектроскопические изображения были получены при помощи оптимизированного цифрового бинормального рефокусированного возбуждения, используя три ортогональных рефокусированных импульса, избирательные для определенного сечения (биномиальное возбуждение с избирательным рефокусированием по объему). Спектроскопические изображения интересующего объема с измерениями 8×2,4×1,5 мм были получены, используя пространственное кодирование 24×24 в 20-мм поле сканирования с усреднением 4-х измерений в сечении, содержащем черную субстанцию, имея номинальный размер элемента объема 1 мкл. Общее время экспозиции - 80 минут. Обработка MRSI изображений. Спектроскопические изображения были преобразованы с помощью Фурье преобразований в объеме фазового кодирования и реформатированы с помощью MatLab Mathworks Inc, Nantick, MA.
Спектры были согласованы, используя AMARES в пакете программного обеспечения JMRUI. Параметры моделирования и константы были генерированы, используя спектры фантомов.
Спектроскопические изображения свободной воды были получены при идентичных параметрах, за исключением: TR=1 с, NA=1 и коэффициент усиления приемника = 1000. Свободная вода использовалась в качестве внутреннего стандарта для каждого элемента объема, чтобы количественно определить концентрацию метаболита из подавленных MRSI данных.
Техник, осуществлявший аппроксимацию данных, не знал их источника. Калибровка отношения амплитуд сигналов метаболита и воды при соответствующих коэффициентах усиления приемника была измерена на фантомах. Расчеты были произведены в MatLab (The Mathworks Inc, Nantick, MA), и концентрации метаболита были выведены в формате ASCII (для базы данных) и в двоичном формате (для совмещения с МРТ).
Как видно из Рисунка 19, инъекции МФТП вызвали потерю значительного количества NAA в черной субстанции и стриатуме мышей контрольной группы. В противоположность этому, снижения уровня NAA не наблюдалось у мышей, интоксицированных МФТП и подвергшихся лечению МККМ, нагруженных полипептид-полиионным комплексом. Данные свидетельствуют о том, что нагруженные клетки могут достигать поврежденные участки мозга в достаточном количестве, и высвобождать активную каталазу, и таким образом оказывать нейрозащитное действие в мышиной модели болезни Паркинсона.
Пример 10
Аккумуляция полиионного комплекса каталазы в различных типах клеток-носителей
Кроме МККМ, было показано, что другие клетки-носители, такие как дендритные клетки (ДК) или Т-лимфоциты, которые также могут проникать в воспаленные участки мозга, могут быть использованы для доставки полиионных комплексов каталазы.
Нагрузка клеток производилась так же, как в случае МККМ. Коротко, ДК или Т-лимфоциты посадили в 96-луночные плашки при плотности 104 клеток/лунку и инкубировали с Alexa Fluor 594-меченым полиионным комплексом каталазы (Z=1) в течение различных промежутков времени. Затем клетки отмыли и лизировали 1% раствором Тритона Х100. Интенсивность флуоресценции клеточных лизатов была измерена и нормализирована на количество клеток (Таблица 14).
Таким образом, было показано, что, как и в случае МККМ, оба типа клеток быстро аккумулировали значительное количество наночастиц каталазы (соответственно 112 мкг, 21 мкг и 30 мкг на миллион ДК, Т-лимфоцитов и МККМ). Это позволяет обеспечить успешную доставку полиионных комплексов каталазы в мозг, используя различные типы клеток.
Пример 11
Сшивка полиионного комплекса каталазы
Для стабилизации комплекса путем сшивки блок сополимера и белка были использованы различные агенты.
Глутаровый альдегид
Для получения полиионного комплекса, 0,5 мл раствора каталазы (0,5 мг/мл) в 60 мМ фосфатном буфере, рН 7,4 было смешано с 0,5 мл раствора блок сополимера. Затем 4 мкл (100-кратный избыток в пересчете на количество NH2-групп) глутарового альдегида (Fluka, # 49632, 25% водный раствор) было добавлено к смеси при постоянном помешивании. Смесь инкубировали в течение 2 часов при комнатной температуре. Затем было добавлено 7,5 мкл раствора боргидрида натрия (5×10-2M) в 1М растворе NaOH. Боргидрид натрия добавляли в 2 приема с разницей в 20 минут. Затем смесь инкубировали в течение одного часа при комнатной температуре и очистили при помощи гель-фильтрации на колонке Sephadex G25.
N-этил-N′-(3-диметиламинопропил)-карбодиимид (ЭДК)
Полиионный комплекс каталазы был получен, как описано выше. Затем было добавлено 1,5 мг ЭДК (30-кратный избыток в пересчете на количество карбоксильных групп) при постоянном помешивании. После инкубации смесь была очищена путем гель-фильтрации на колонке Sephadex G25.
Бис-(сульфосукцинимидил)суберат, натриевая соль (БС3)
Полиионный комплекс каталазы был получен, как описано выше. Затем к смеси добавили 2 мг БС3 (7-кратный избыток) при постоянном помешивании. Смесь инкубировали в течение 3 часов при комнатной температуре. После инкубации смесь была очищена путем гель-фильтрации на колонке Sephadex G25.
Сшивку полиионных комплексов каталазы подтвердили вестерн-блотом. Образцы подвергли ПААГ электрофорезу в 10% геле в денатурирующих условиях (в присутствии ДСН), что разрушает несшитые комплексы. Затем гели были блоттированы и белковые полосы были визуализированы первичными антителами к каталазе (Abeam, ab1877). На рисунке 20 представлены изображения гелей полиионных комплексов каталазы, сшитых различными сшивками. Дорожка 1: стандарты молекулярных масс; дорожка 2: свободная каталаза; дорожка 3: полиионный комплекс каталазы, сшитый ЭДК; дорожка 4: полиионный комплекс каталазы, сшитый ГА; дорожка 5: полиионный комплекс каталазы, сшитый БС3.
Как показано на рисунке 20, полная конъюгация была достигнута при использовании ГА, что следует из отсутствия полосы каталазы, так как большие комплексы не могли войти в гель (дорожка 4). Использование ЭДК в качестве сшивки (дорожка 3) также привело к сшиванию, хотя полной конъюгации достигнуть не удалось при данных условиях, на что указывает присутствие полосы свободной каталазы. Сшивка БС3 привела к образованию меньших комплексов, которые смогли войти в гель, но с большим замедлением по сравнению со свободной каталазой (дорожка 2).
Сшивка полиионных комплексов супероксид дисмутазы (СОД)
Были получены схожие сшитые комплексы СОД и блок сополимера.
ГА
Для получения полиионного комплекса СОД было смешано 0,5 мл раствора СОД (1 мг/мл) в 60 мМ фосфатном буфере, рН 7,4 и 0,5 мл раствора блок сополимера (0,25 мг/мл) в таком же буфере. Затем к смеси добавили 10 мкл (1000-кратный избыток в пересчете на количество NH2-групп) ГА при постоянном помешивании. Смесь инкубировали в течение 2 часов при комнатной температуре. Затем было добавлено 5 мкл раствора боргидрида натрия (5×10-2 М) в 1 М NaOH. Боргидрид натрия добавляли в 2 приема с интервалом в 20 минут. После этого смесь инкубировали в течение еще одного часа при комнатной температуре и очистили путем гель-фильтрации на колонке Sephadex G25.
ЭДК
Полиионные комплексы СОД были получены, как описано выше. Затем к смеси было добавлено 1,5 мг ЭДК (12-кратный избыток в пересчете на количество СОО-групп) при постоянном перемешивании. Смесь инкубировали в течение 2 часов при комнатной температуре. После инкубации смесь очистили путем гель-фильтрации на колонке Sephadex G25.
БС3
Полиионные комплексы СОД были получены, как описано выше. Затем к смеси было добавлено 1,7 мг БС3 (в пересчете на остатки Lys) при постоянном перемешивании. Затем смесь инкубировали в течение 3-х часов при комнатной температуре. После инкубации смесь очистили путем гель-фильтрации на колонке Sephadex G25.
Сшивку полиионных комплексов СОД подтвердили вестерн-блотом. Образцы подвергли ПААГ электрофорезу в 10% геле в денатурирующих условиях (в присутствии ДСН), что разрушает несшитые комплексы. Затем гели были блоттированы и белковые полосы были визуализированы первичными антителами к СОД (Calbiochaem, # 574597). На рисунке 21 представлены изображения гелей полиионных комплексов СОД, сшитых различными сшивками. Дорожка 1: стандарты молекулярных масс; дорожка 2: свободная СОД; дорожка 3: несшитый полиионный комплекс; 4: полиионный комплекс СОД, сшитый ЭДК; дорожка 5: полиионный комплекс СОД, сшитый ГА; дорожка 6: полиионный комплекс СОД, сшитый БС3.
Как показано на рисунке 21, с помощью ЭДК (дорожка 4) не была достигнута полная конъюгация при данных конкретных условиях, так как видна полоса свободной СОД. В противоположность этому при использовании ГА (дорожка 5) и БС3 (дорожка 6) конъюгация была полной, на что указывает отсутствие полосы СОД на вестерн-блоте, так как образовавшиеся большие комплексы не могли войти в гель.
Сшивка полиионных комплексов каталазы/СОД
В целом для получения смешенных полиионных комплексов каталазы/СОД сначала смешали каталазу и СОД при рН 6,8 (при данном рН каталаза заряжена отрицательно (рI 7.28), а СОД - положительно (pI 6.32)). Затем добавили раствор блок сополимера. Для конъюгации блок сополимера с белками были использованы разные сшивки, как описано выше.
ГА
Для получения полиионного комплекса каталазы/СОД 1 мг каталазы и 1,33 мг СОД растворили в 60 мМ фосфатном буфере, рН 6,8. После этого к смеси добавили 1,3 мг блок сополимера и инкубировали 10 минут при комнатной температуре. Затем к смеси добавили 5 мкл (9-кратный избыток (в пересчете на количество NH2-групп)) ГА при постоянном помешивании. Смесь инкубировали в течение ночи (8 часов) при 4°С. После инкубации к смеси добавили 6,5 мкл раствора боргидрида натрия (5×10-2 М) в 1 М NaOH. Боргидрид натрия добавляли в 2 приема с интервалом в 20 минут. Смесь инкубировали в течение еще одного часа при комнатной температуре и очистили путем гель-фильтрации на колонке Sephadex G25.
ЭДК
Полиионный комплекс каталазы/СОД был получен, как описано выше. Затем к смеси добавили 10 мг ЭДК (20-кратный избыток (в пересчете на количество СОО-групп), при постоянном помешивании. Смесь инкубировали в течение ночи (8 часов) при 4°С. После инкубации смесь очистили путем гель-фильтрации на колонке Sephadex G25.
БС3
Полиионный комплекс каталазы/СОД был получен, как описано выше. Затем к смеси добавили 8,6 мг БС3 (10-кратный избыток в пересчете на остатки Lys) при постоянном перемешивании. После инкубации смесь очистили путем гель-фильтрации на колонке Sephadex G25.
ЭДК-сульфо-NHS
Чтобы стабилизировать промежуточный ЭДК комплекс, был использован гидроксисукцинимид (сульфо-NHS). Для этого полиионный комплекс каталазы/СОД был синтезирован, как описано ранее. Затем к смеси сначала было добавлено 10 мг ЭДК (20-кратный избыток (в пересчете на количество СОО-групп)) при постоянном перемешивании, а затем 2 мг сульфо-NHS, и реакционную смесь инкубировали в течение 3 часов при комнатной температуре. После инкубации смесь очистили путем гель-фильтрации на колонке Sephadex G25.
Сшивку полиионных комплексов каталазы/СОД подтвердили вестерн-блотом. Образцы подвергли ПААГ электрофорезу в 10% геле при денатурирующих условиях (в присутствии ДСН). Затем гели были блоттированы и белковые полосы были визуализированы первичными антителами к СОД и каталазе. На рисунке 22A представлены изображения гелей полиионных комплексов каталазы/СОД, сшитых различными сшивками и окрашенных антителами к каталазе. Несшитый полиионный комплекс каталазы/СОД; полиионный комплекс каталазы/СОД (дорожка 1); сшитый ГА (дорожка 2); ЭДК (дорожка 3); БС3 (дорожка 4); ЭДК-S-NHS (дорожка 5). Как показано на рисунке 22А, использование ГА привело к полной конъюгации (дорожка 2); конъюгация ЭДК с другой стороны не была полной (дорожка 3), о чем свидетельствует небольшая полоса каталазы. Стабилизация промежуточного комплекса ЭДК сульфо-N-гидроксисукцинимидом (дорожка 5) привела к значительному улучшению сшивки по сравнению с только ЭДК (дорожка 3). На рисунке 22Б представлены изображения гелей полиионных комплексов каталазы/СОД, сшитых различными сшивками и окрашенных антителами к СОД. Несшитый полиионный комплекс каталазы/СОД (дорожка 1); полиионный комплекс каталазы/СОД, сшитый ГА (дорожка 2); ЭДК (дорожка 3); БС3 (дорожка 4); ЭДК-S-NHS (дорожка 5).
Эти результаты подтверждают данные при окраске геля антителами к каталазе. Полная конъюгация была получена при использовании ГА (дорожка 2); сшивка ЭДК (дорожка 3) привела к неполной конъюгации (так как присутствует значительное окрашивание свободной СОД). Использование БС3 (дорожка 4) и сульфо-N-гидроксисукцинимида вместе с ЭДК (дорожка 5) привело к практически полной конъюгации.
Пример 12
Визуализация биораспределения МККМ в мышах, интоксицированных МФТП
Перед началом эксперимента самок BALB/C мышей анестезировали инъекцией пентобарбитала i.p. (30-40 мг/кг массы тела), обрили и депелировали (чтобы уменьшить блокировку флуоресценции шерстью). Мышей держали на жидкой диете в течение 72 часов (чтобы избавиться от автофлуоресценции в желудке и кишечнике от твердой еды). Мышам ввели 18 мг свободного основания МФТП/кг массы тела в ФСБ путем 4 интреперитониальных инъекций каждые 2 часа (МФТП (Sigma Chemical Co., St. Louis, МО)). Спустя 18 часов мышам ввели Li-COR-меченые МККМ, нагруженные полиионным комплексом каталазы, путем инъекции через хвостовую вену (50 млн/мышь). Затем, мыши были анестезированы смесью 1,5% изофлурана, 66% оксида азота и остаточным кислородом и помещены в камеру для визуализации. Биораспределение меченых МККМ, нагруженных полиионным комплексом каталазы, измерялось по интенсивности флуоресценции Li-COR-меченных МККМ in vivo, используя систему визуализации с газовой анестезией серии IVIS 200. Li-COR-меченные МККМ, нагруженные полиионным комплексом каталазы, начали накапливаться в мозге через 2 часа после iv инъекции. Концентрация МККМ достигала максимума через 4-7 часов и оставалась повышенной по меньшей мере в течение 48 часов после инъекции (Рисунок 23). Данные показывают, что периферически введенные МККМ, нагруженные полиионным комплексом каталазы, способны достигать и накапливаться в мозге в значительных количествах.
Все публикации и патенты, процитированные в данном описании, включены в него во всей своей полноте путем ссылки. Хотя вышеизложенное изобретение было детально проиллюстрировано с помощью примеров для ясности понимания, специалистам в данной области будет очевидно, что могут быть сделаны многочисленные изменения и модификации, не отступая от сути изобретения, которое ограничено исключительно объемом прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИМЕРНЫЕ НАНОЧАСТИЦЫ СОСТАВА ФЕРМЕНТ-ПОЛИКАТИОН-ПОЛИАНИОН, СОДЕРЖАЩИЕ АНТИОКСИДАНТНЫЙ ФЕРМЕНТ, ДЛЯ ПРИМЕНЕНИЯ В МЕДИЦИНЕ И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2013 |
|
RU2575836C2 |
| СПОСОБ И КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА | 2011 |
|
RU2541100C2 |
| ЭРИТРОФЕРРОН И ERFE-ПОЛИПЕПТИДЫ И СПОСОБЫ РЕГУЛЯЦИИ МЕТАБОЛИЗМА ЖЕЛЕЗА | 2013 |
|
RU2684216C2 |
| ФАКТОРЫ, ДЕЙСТВУЮЩИЕ НА АКТИВНОСТЬ ФЕРМЕНТА, ВЫСВОБОЖДАЮЩЕГО РЕЦЕПТОР ФАКТОРА НЕКРОЗА ОПУХОЛЕЙ | 1999 |
|
RU2279478C2 |
| ВАРИАНТЫ ГЕМАГГЛЮТИНИНА И НЕЙРАМИНИДАЗЫ ГРИППА | 2010 |
|
RU2535970C2 |
| ХИМЕРНЫЕ МОЛЕКУЛЫ ФАКТОРА VII | 2010 |
|
RU2563231C2 |
| СПОСОБ ЛЕЧЕНИЯ БУЦИНДОЛОЛОМ, ОСНОВАННЫЙ НА ОПРЕДЕЛЕНИИ ГЕНОТИПА | 2005 |
|
RU2389798C2 |
| ПЕПТИДЫ НАПРАВЛЕННОГО ДЕЙСТВИЯ НА VEGFR-1/NRP-1 | 2008 |
|
RU2488592C2 |
| АНТАГОНИСТЫ НЕУРЕГУЛИНА И ПРИМЕНЕНИЕ ИХ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ | 2011 |
|
RU2587619C2 |
| ОНКОЛИТИЧЕСКИЙ HSV-ВЕКТОР | 2014 |
|
RU2719190C2 |
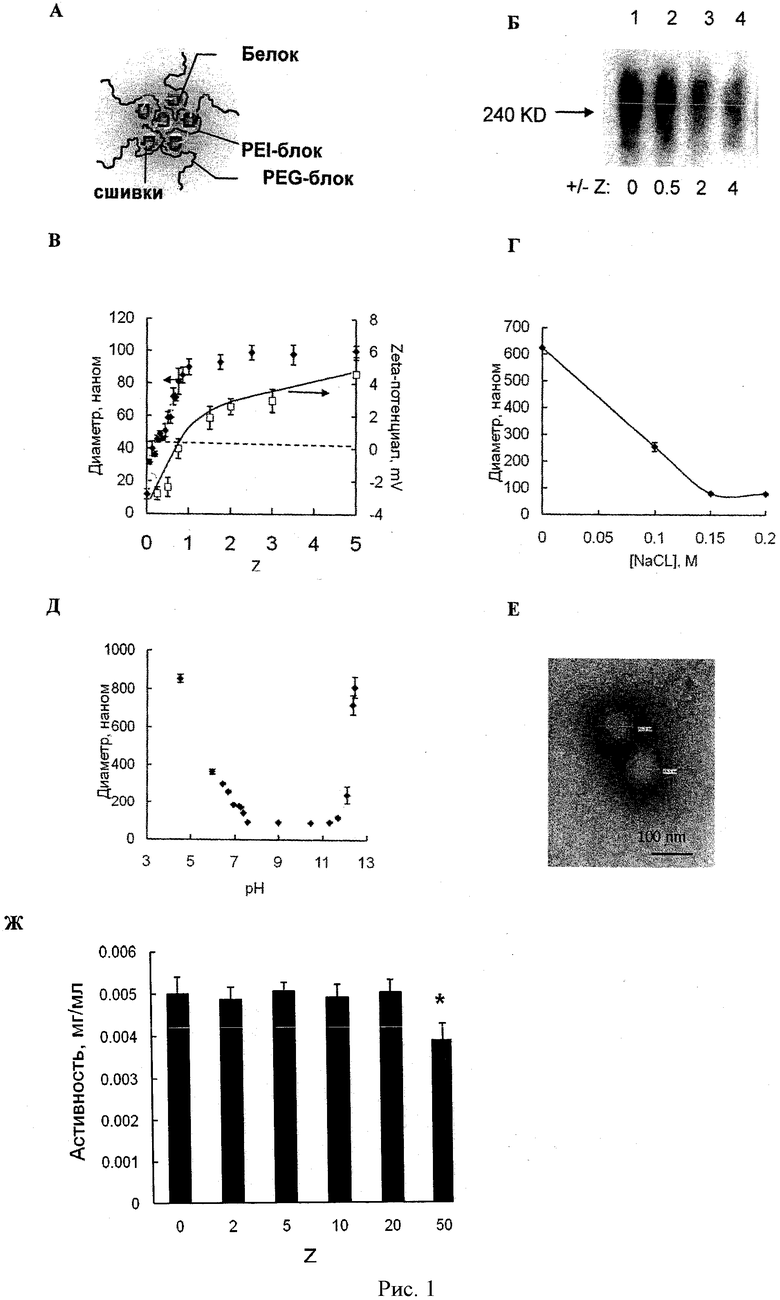
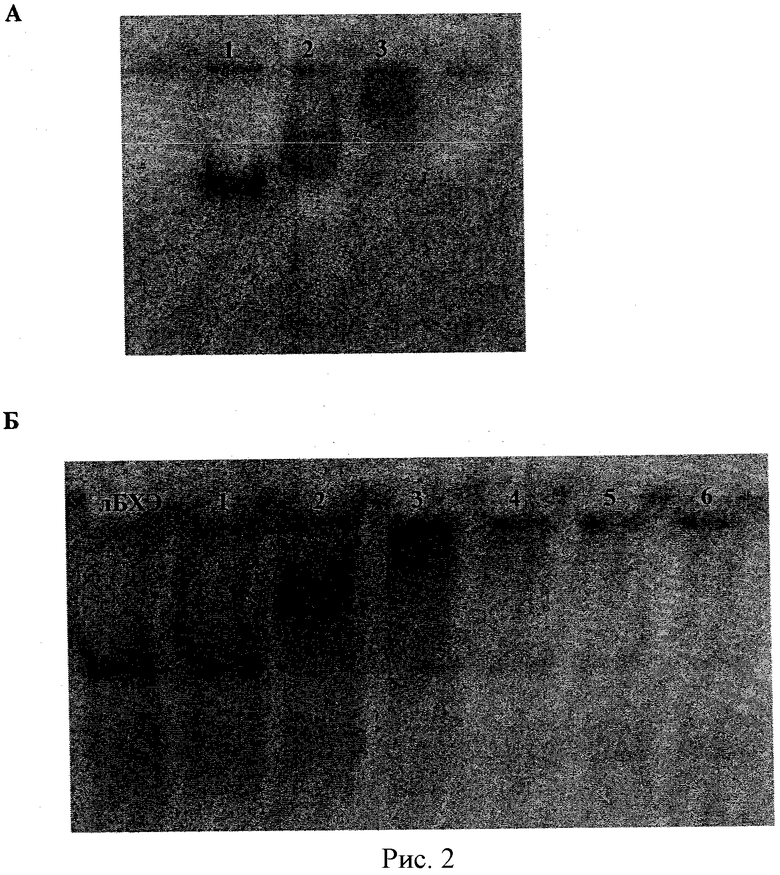
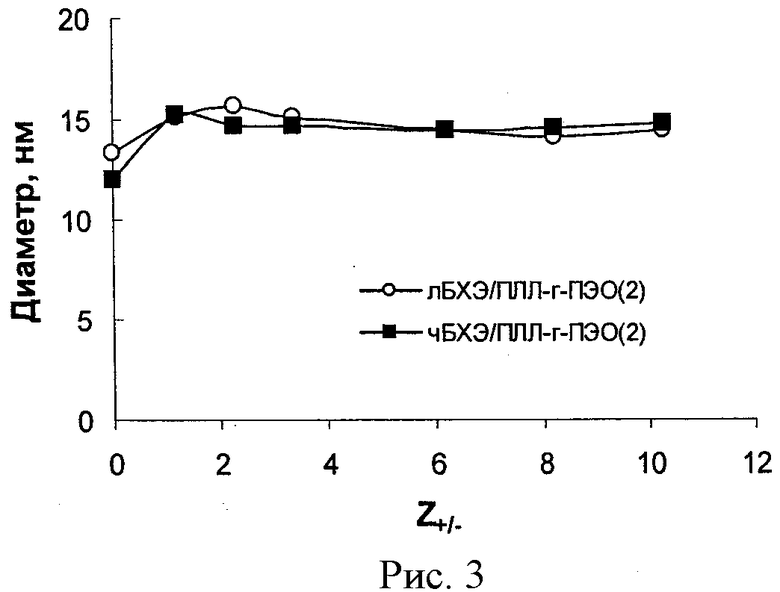
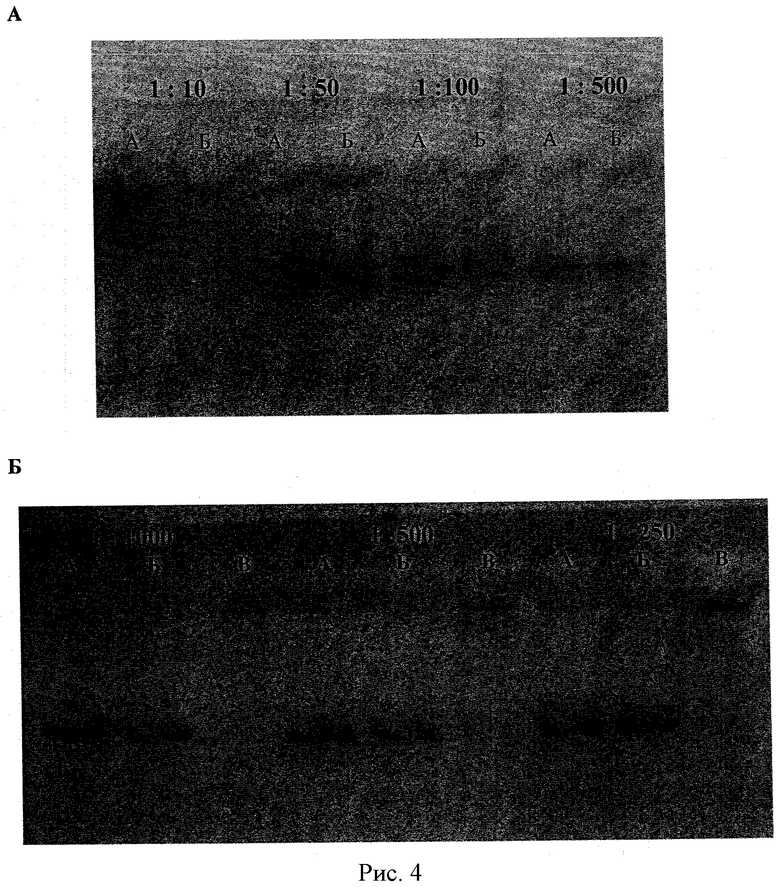
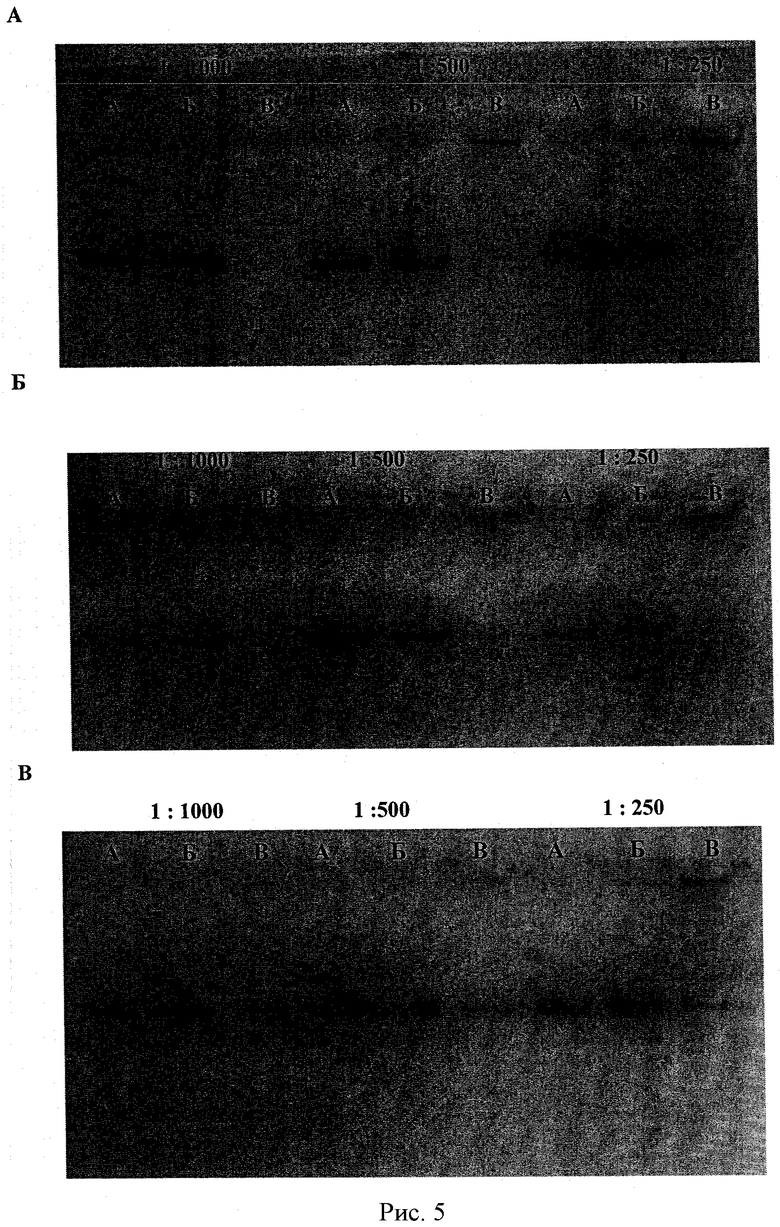
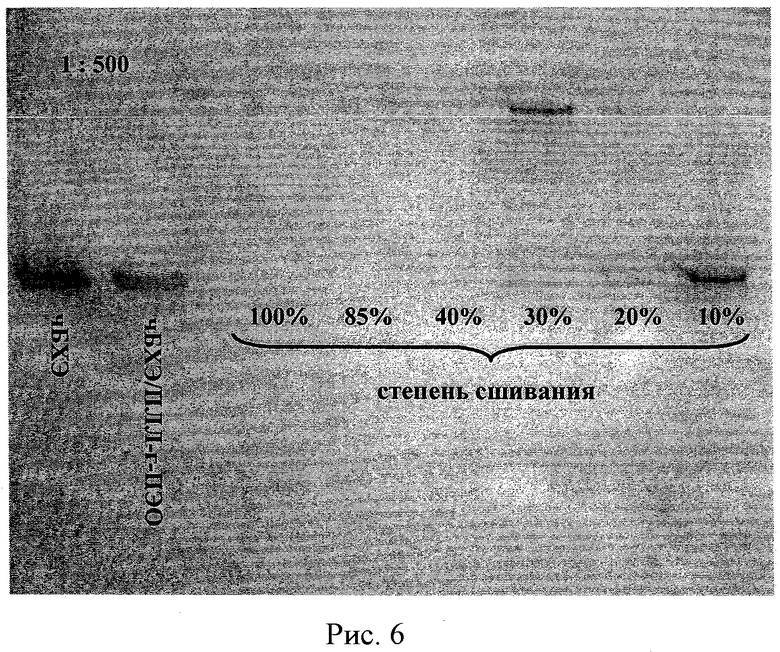
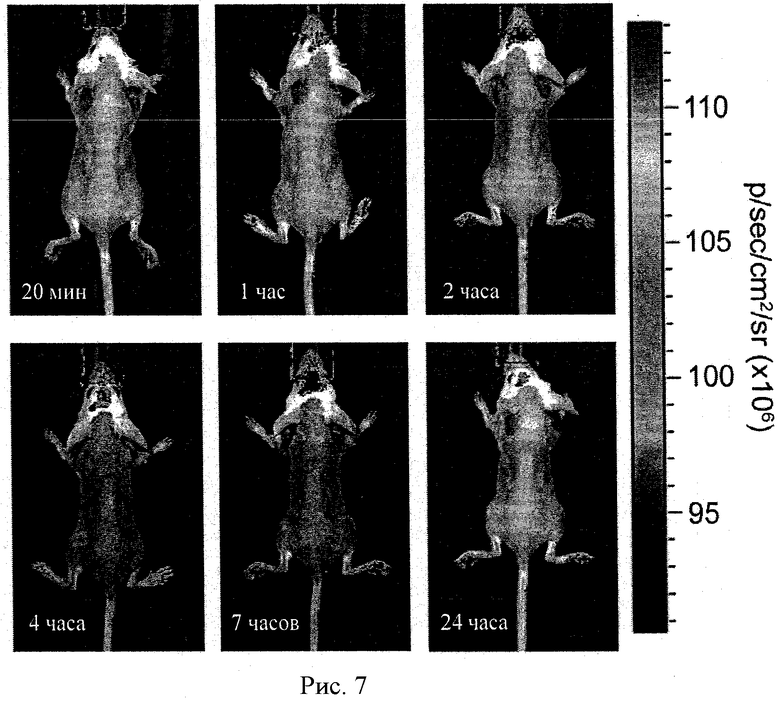


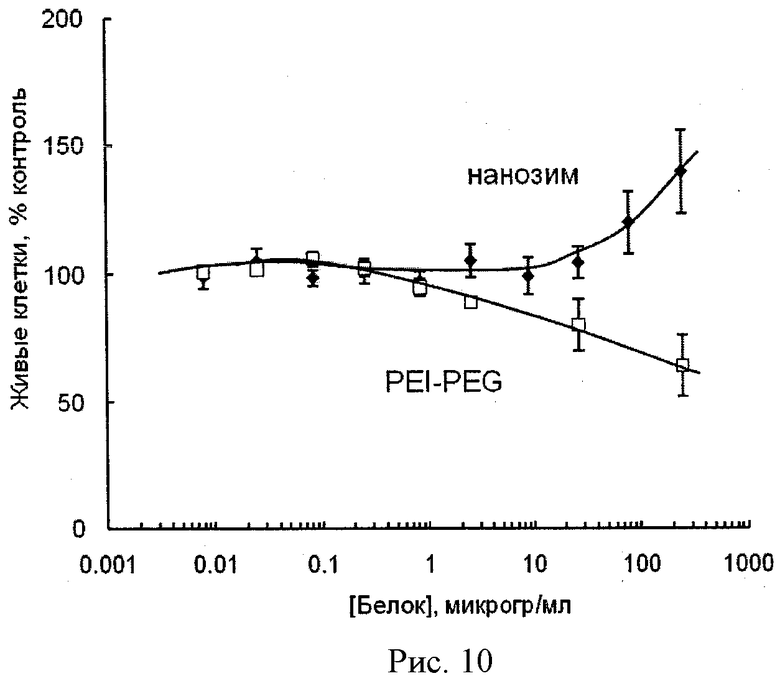
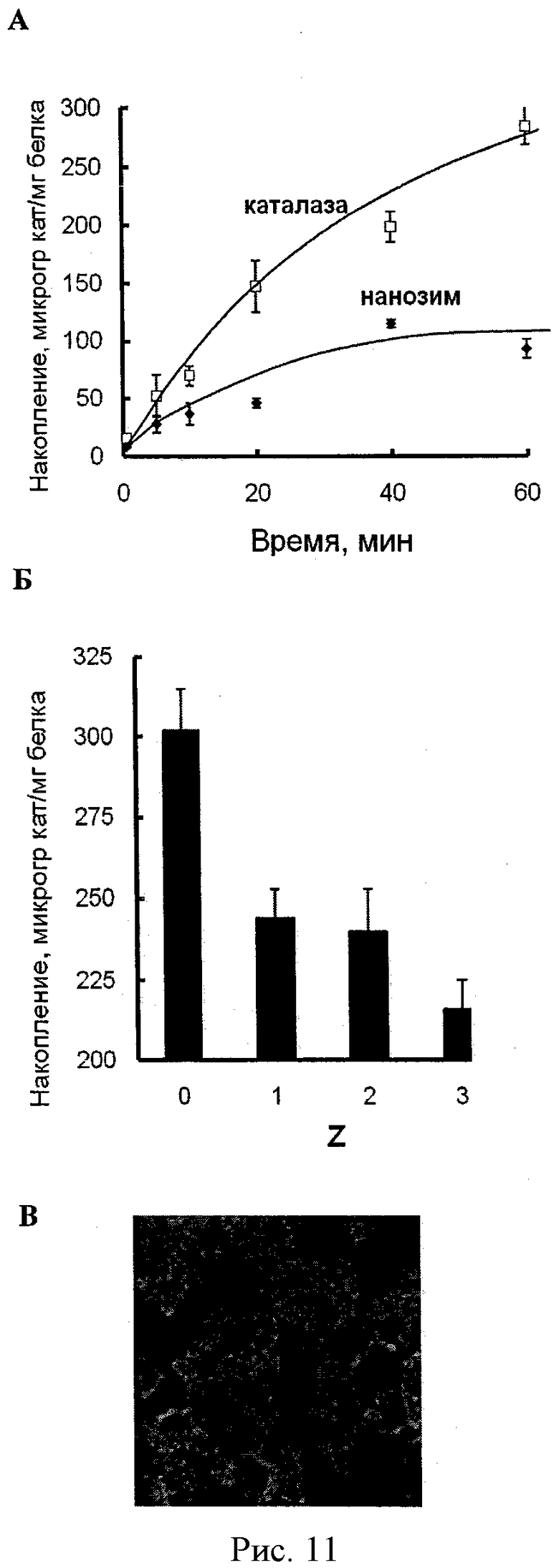
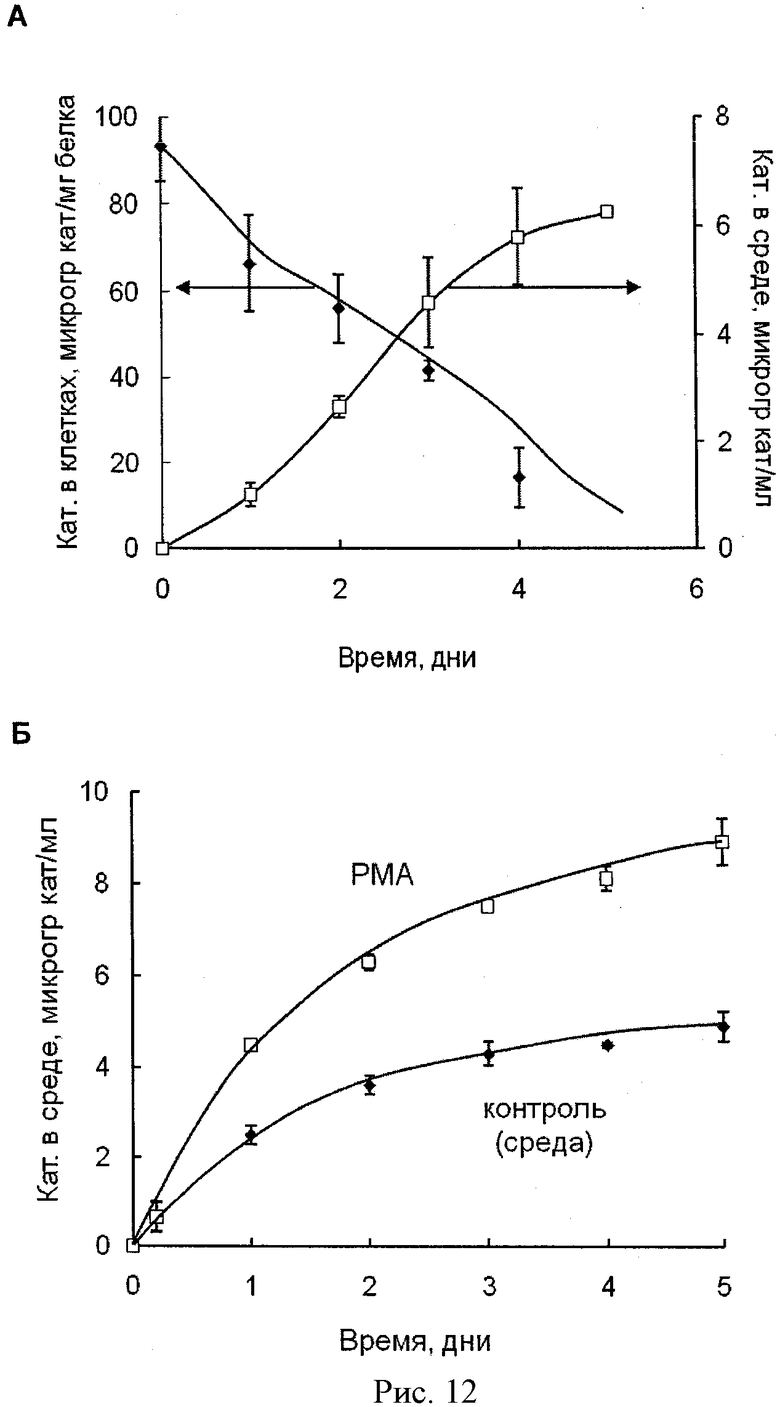
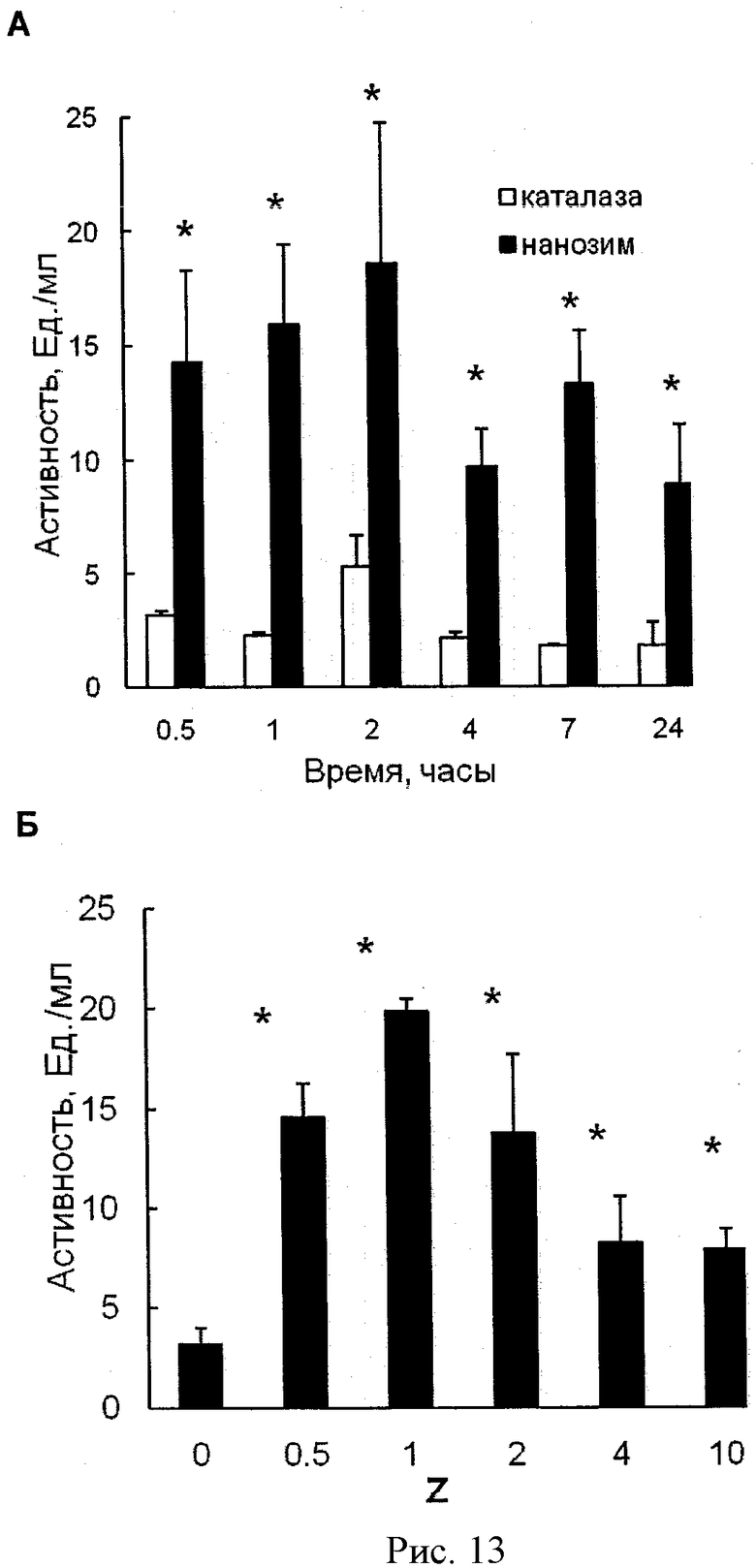
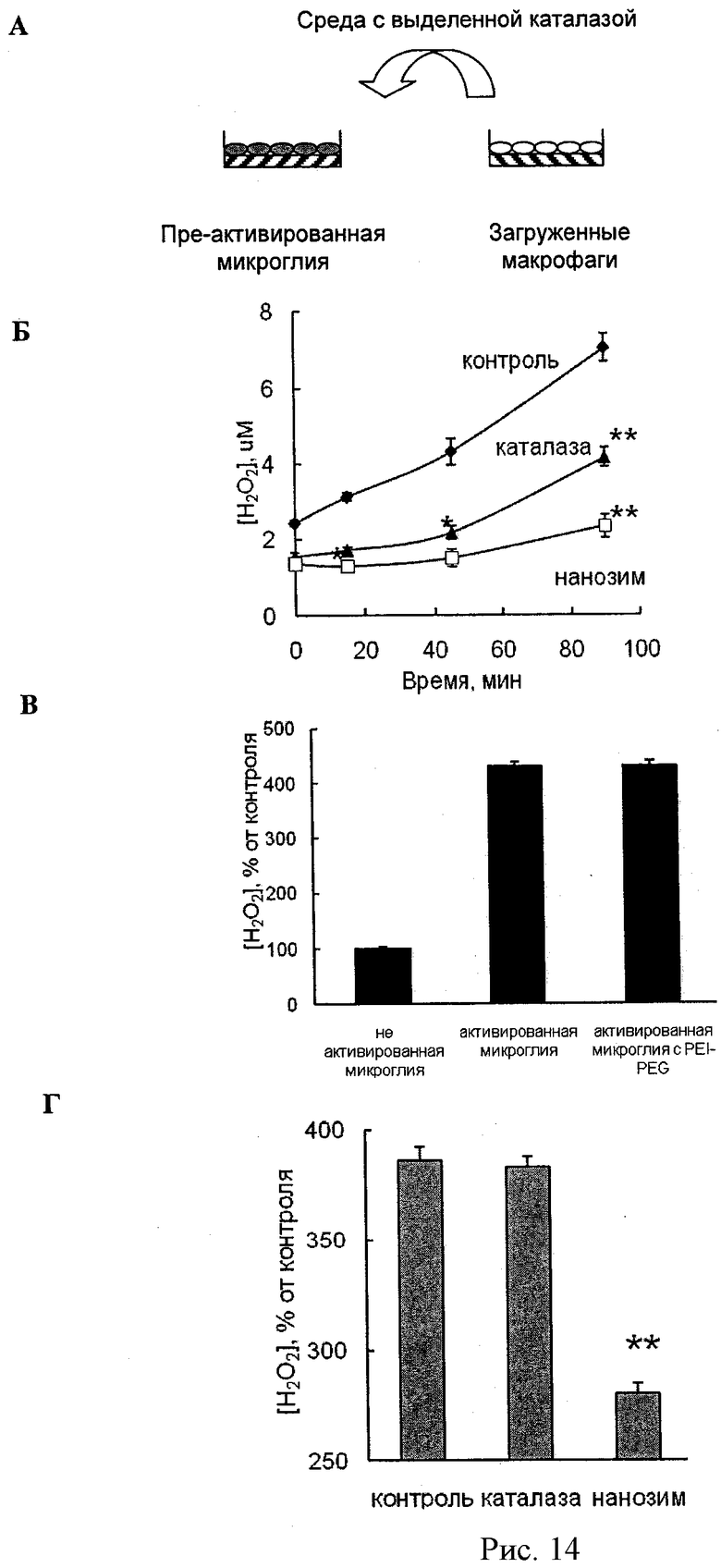
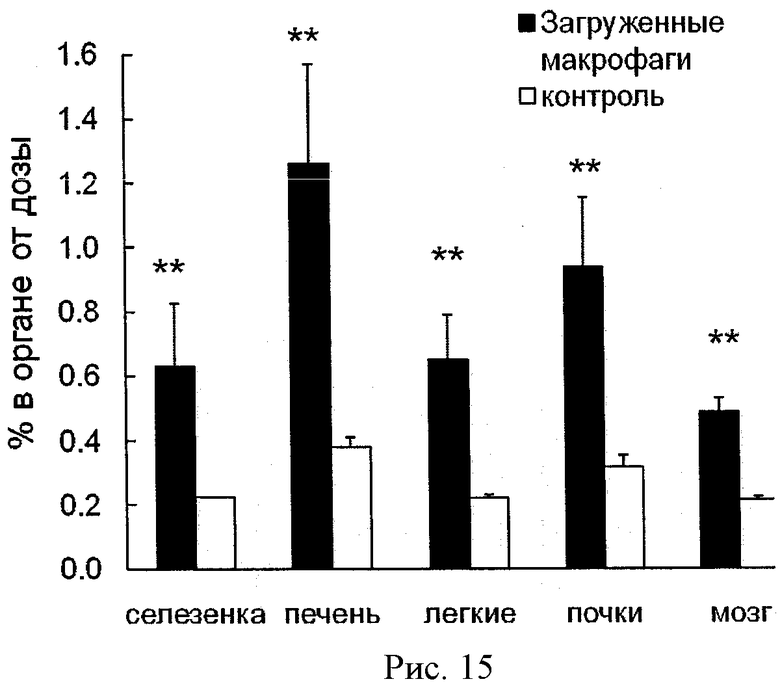
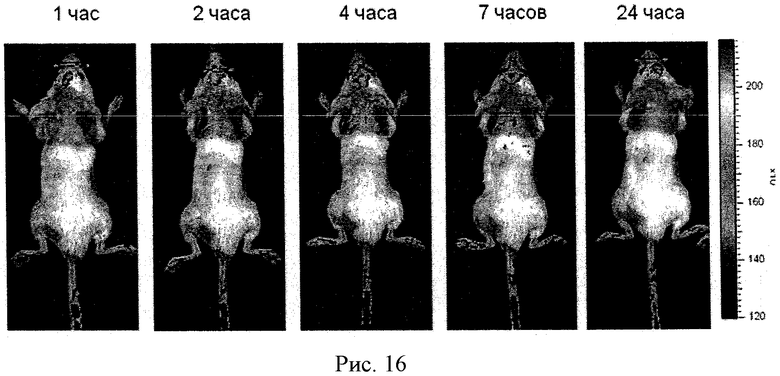
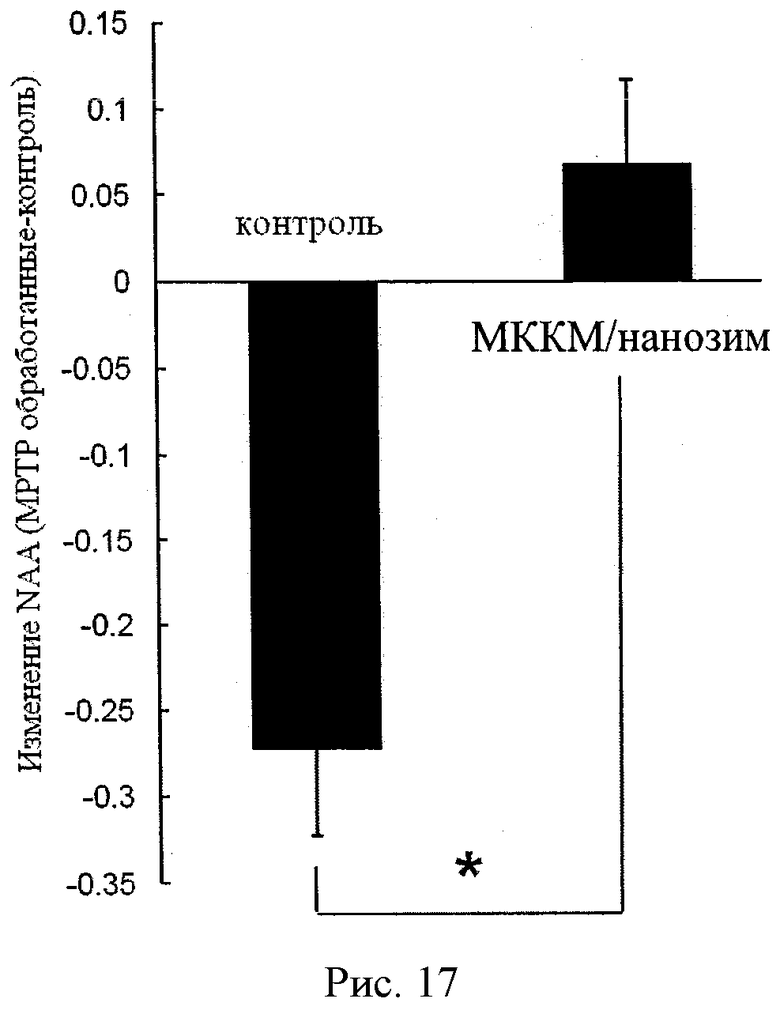


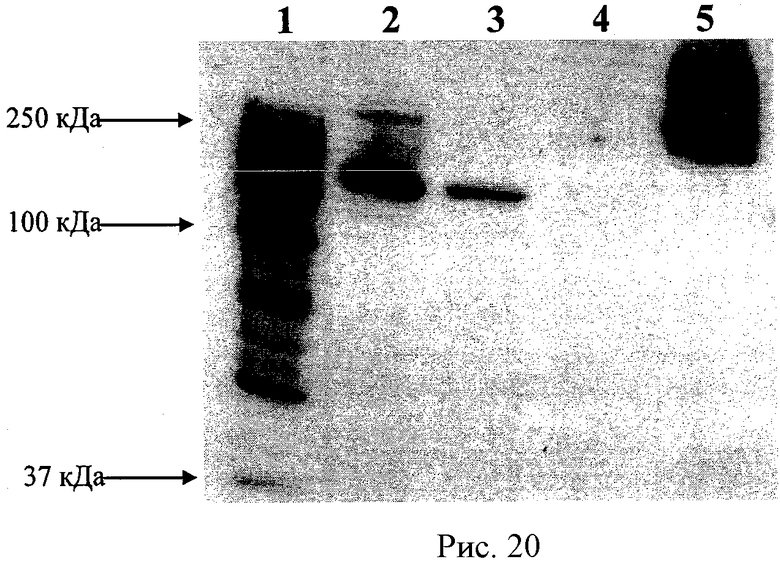
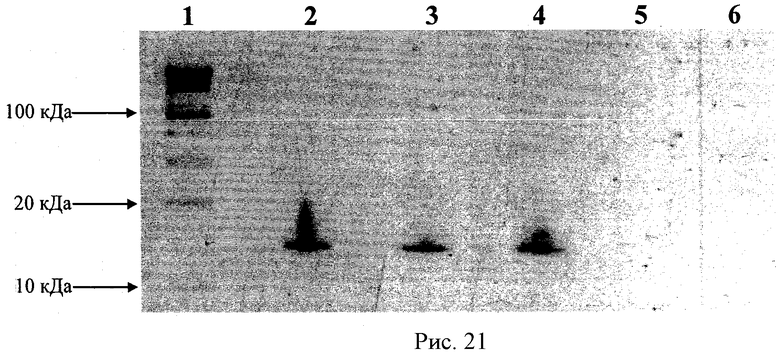
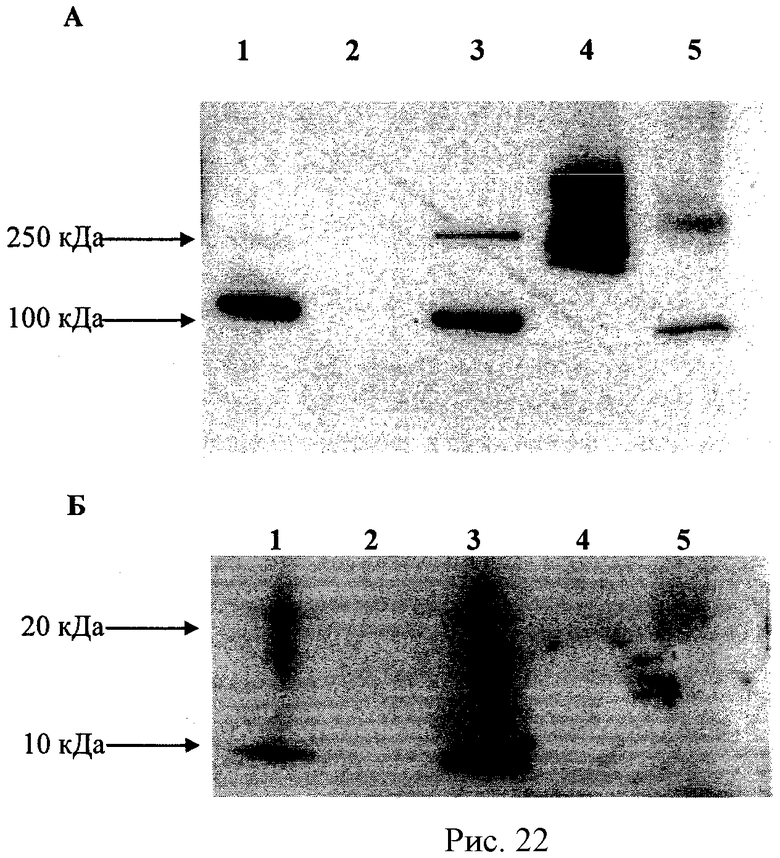
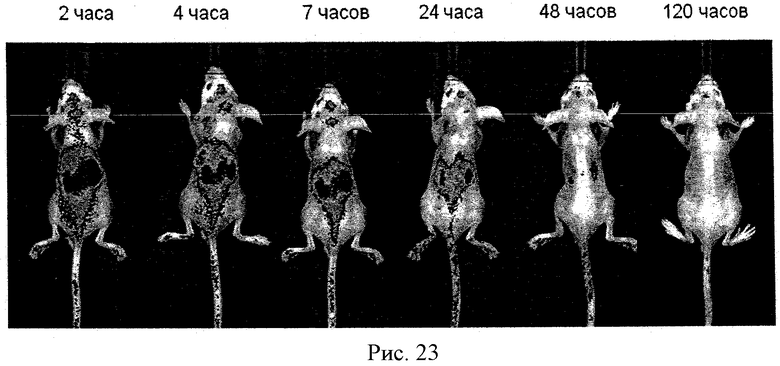
Группа изобретений относится к лечению нейрологических заболеваний центральной нервной системы. Способ лечения нейрологического нарушения центральной нервной системы у пациента включает введение терапевтически активных количеств композиции, включающей: а) по меньшей мере один комплекс, включающий терапевтический полипептид и блок сополимер, где указанный блок сополимер включает по меньшей мере один растворимый в воде неионный сегмент и по меньшей мере один полиионный сегмент, где указанный полиионный сегмент включает по меньшей мере один заряд, противоположный заряду терапевтического полипептида, где указанный комплекс имеет морфологию ядро-оболочка, где ядро содержит терапевтический полипептид и полиионный сегмент блок сополимера и где оболочка содержит водорастворимый неионный сегмент блок сополимера, и б) по меньшей мере один фармацевтически приемлемый носитель, где указанный по меньшей мере один комплекс преодолевает гематоэнцефалический барьер. Способ лечения нейрологического нарушения центральной нервной системы у пациента включает введение терапевтически активных количеств композиции, включающей: а) выделенные клетки, содержащие по меньшей мере один комплекс, включающий терапевтический полипептид и блок сополимер, где указанный блок сополимер включает по меньшей мере один растворимый в воде неионный сегмент и по меньшей мере один полиионный сегмент, где указанный полиионный сегмент включает по меньшей мере один заряд, противоположный заряду терапевтического полипептида, где указанный комплекс имеет морфологию ядро-оболочка, где ядро содержит терапевтический полипептид и полиионный сегмент блок сополимера и где оболочка содержит водорастворимый неионный сегмент блок сополимера, и б) по меньшей мере один фармацевтически приемлемый носитель, где указанные клетки преодолевают гематоэнцефалический барьер. Выделенная клетка, которая содержит по меньшей мере один комплекс, содержащий по меньшей мере один белок и блок сополимер, где указанный сополимер включает по меньшей мере один растворимый в воде неионный сегмент и по меньшей мере один полиионный сегмент, где указанный полиионный сегмент содержит по меньшей мере один заряд, противоположный заряду указанного белка, где указанный комплекс имеет морфологию ядро-оболочка, где ядро содержит терапевтический полипептид и полиионный сегмент блок сополимера и где оболочка содержит водорастворимый неионный сегмент блок сополимера. Композиция, содержащая указанные выделенные клетки и по меньшей мере один фармацевтически приемлемый носитель. Способ доставки белка через гематоэнцефалический барьер в центральную нервную систему, включающий введение субъекту указанной композиции. Способ доставки белка через гематоэнцефалический барьер в центральную нервную систему, включающий введение субъекту композиции, содержащей: а) по меньшей мере один комплекс, включающий терапевтический полипептид и блок сополимер, где указанный блок сополимер включает по меньшей мере один растворимый в воде неионный сегмент и по меньшей мере один полиионный сегмент, где указанный полиионный сегмент включает по меньшей мере один заряд, противоположный заряду терапевтического полипептида, где указанный комплекс имеет морфологию ядро-оболочка, где ядро содержит терапевтический полипептид и полиионный сегмент блок сополимера и где оболочка содержит водорастворимый неионный сегмент блок сополимера, и б) по меньшей мере один фармацевтически приемлемый носитель. Группа изобретений обеспечивает повышение эффективности лечения нейрологических заболеваний. 6 н. и 33 з.п. ф-лы, 23 ил., 14 табл., 12 пр.
1. Способ лечения нейрологического нарушения центральной нервной системы у пациента, нуждающегося в этом, включающий введение терапевтически активных количеств композиции, включающей:
а) по меньшей мере один комплекс, включающий терапевтический полипептид и блок сополимер, где указанный блок сополимер включает по меньшей мере один растворимый в воде неионный сегмент и по меньшей мере один полиионный сегмент, где указанный полиионный сегмент включает по меньшей мере один заряд, противоположный заряду терапевтического полипептида, где указанный комплекс имеет морфологию ядро-оболочка, где ядро содержит терапевтический полипептид и полиионный сегмент блок сополимера и где оболочка содержит водорастворимый неионный сегмент блок сополимера, и
б) по меньшей мере один фармацевтически приемлемый носитель, где указанный по меньшей мере один комплекс преодолевает гематоэнцефалический барьер.
2. Способ по п.1, в котором указанный полиионный сегмент отрицательно заряжен, а указанный терапевтический полипептид имеет суммарный положительный заряд при рН 7,4.
3. Способ по п.1, в котором указанный полиионный сегмент положительно заряжен, а указанный терапевтический полипептид имеет суммарный отрицательный заряд при рН 7,4.
4. Способ по п.1, в котором указанный полиионный сегмент выбран из группы, состоящей из полиалкиленимина, полилизина, полиаргинина, полиаспартата, полиглутамата, полиакрилата, полиалкилен акрилового полимера и их сополимеров.
5. Способ по п.1, где указанный водорастворимый неионный сегмент является полиэтиленоксидом.
6. Способ по п.1, где указанный терапевтический полипептид выбран из группы, состоящей из ферментов, антител, гормонов и факторов роста.
7. Способ по п.1, где указанный терапевтический полипептид обладает терапевтическим действием в центральной нервной системе.
8. Способ по п.1, в котором указанный терапевтический полипептид и указанный полиионный сегмент блок сополимера химически сшиты.
9. Способ по п.1, в котором указанный терапевтический полипептид выбран из группы, состоящей из эндокринных факторов, факторов роста, высвобождающих факторов гипоталамуса, нейротрофических факторов, нейротрансмиттеров, паракринных факторов, антител, фрагментов антител, цитокинов, эндорфинов, антагонистов полипептидов, агонистов рецепторов, экспрессируемых клетками ЦНС, полипептидов болезней лизосомального накопления и антиапоптозных белков.
10. Способ по п.1, в котором указанный терапевтический полипептид выбран из группы, состоящей из каталазы, супероксид дисмутазы и глутатионпероксидазы.
11. Способ по п.1, в котором указанный терапевтический полипептид выбран из группы, состоящей из бутирилхолинэстеразы, ацетилхолинэстеразы, реактиваторов холинэстеразы, скавенджеров органофосфатов и ингибиторов карбаматного типа.
12. Способ по п.1, где указанное нейрологическое нарушение представляет собой нейровоспалительное заболевание или нарушение.
13. Способ по п.12, где указанное нейровоспалительное заболевание или нарушение представляет собой болезнь Паркинсона.
14. Способ по п.1, где указанная композиция вводится внутривенно.
15. Способ лечения нейрологического нарушения центральной нервной системы у пациента, нуждающегося в этом, включающий введение терапевтически активных количеств композиции, включающей:
а) выделенные клетки, содержащие по меньшей мере один комплекс, включающий терапевтический полипептид и блок сополимер, где указанный блок сополимер включает по меньшей мере один растворимый в воде неионный сегмент и по меньшей мере один полиионный сегмент, где указанный полиионный сегмент включает по меньшей мере один заряд, противоположный заряду терапевтического полипептида, где указанный комплекс имеет морфологию ядро-оболочка, где ядро содержит терапевтический полипептид и полиионный сегмент блок сополимера и где оболочка содержит водорастворимый неионный сегмент блок сополимера, и
б) по меньшей мере один фармацевтически приемлемый носитель, где указанные клетки преодолевают гематоэнцефалический барьер.
16. Способ по п.15, в котором указанный полиионный сегмент отрицательно заряжен, а указанный терапевтический полипептид имеет суммарный положительный заряд при pH 7,4.
17. Способ по п.15, в котором указанный полиионный сегмент положительно заряжен, а указанный терапевтический полипептид имеет суммарный отрицательный заряд при pH 7,4.
18. Способ по п.15, в котором указанный полиионный сегмент выбран из группы, состоящей из полиалкиленимина, полилизина, полиаргинина, полиаспартата, полиглутамата, полиакрилата, полиалкилен акрилового полимера и их сополимеров.
19. Способ по п.15, где указанный водорастворимый неионный сегмент является полиэтиленоксидом.
20. Способ по п.15, в котором указанный терапевтический полипептид и указанный полиионный сегмент блок сополимера химически сшиты.
21. Способ по п.15, в котором указанный терапевтический полипептид выбран из группы, состоящей из каталазы, супероксид дисмутазы и глутатионпероксидазы.
22. Способ по п.15, в котором указанный терапевтический полипептид выбран из группы, состоящей из бутирилхолинэстеразы, ацетилхолинэстеразы, реактиваторов холинэстеразы, скавенджеров органофосфатов и ингибиторов карбаматного типа.
23. Способ по п.15, где указанное нейрологическое нарушение представляет собой нейровоспалительное заболевание или нарушение.
24. Способ по п.23, где указанное нейровоспалительное заболевание или нарушение представляет собой болезнь Паркинсона.
25. Способ по п.15, где указанная композиция вводится внутривенно.
26. Способ по п.15, в котором указанные клетки выделены из пациента, подвергающегося лечению.
27. Способ по п.15, в котором указанные клетки являются иммунными клетками.
28. Способ по п.27, в котором указанные иммунные клетки включают по меньшей мере одну клетку, выбранную из группы, состоящей из моноцитов, макрофагов, моноцитов, генерированных из клеток костного мозга, дендритных клеток, лимфоцитов, Т-клеток, нейтрофилов, эозинофилов и базофилов.
29. Способ по п.28, в котором указанные иммунные клетки включают по меньшей мере одну клетку, выбранную из группы, состоящей из моноцитов и макрофагов.
30. Способ по п.29, в котором указанные иммунные клетки являются моноцитами, генерированным из клеток костного мозга.
31. Выделенная клетка, которая содержит по меньшей мере один комплекс, содержащий по меньшей мере один белок и блок сополимер, где указанный сополимер включает по меньшей мере один растворимый в воде неионный сегмент и по меньшей мере один полиионный сегмент, где указанный полиионный сегмент содержит по меньшей мере один заряд, противоположный заряду указанного белка, где указанный комплекс имеет морфологию ядро-оболочка, где ядро содержит терапевтический полипептид и полиионный сегмент блок сополимера и где оболочка содержит водорастворимый неионный сегмент блок сополимера.
32. Выделенная клетка по п.31, где указанная клетка является иммунной клеткой.
33. Выделенная клетка по п.32, где указанная иммунная клетка выбрана из группы, состоящей из моноцитов, макрофагов, моноцитов, генерированных из клеток костного мозга, дендритных клеток, Т-клеток, нейтрофилов, эозинофилов и базофилов.
34. Выделенная клетка по п.33, где указанная иммунная клетка выбрана из группы, состоящей из моноцитов и макрофагов.
35. Выделенная клетка по п.34, где указанная иммунная клетка является моноцитом, генерированным из клеток костного мозга.
36. Выделенная клетка по п.31, где указанный белок обладает терапевтическим действием при нейрологическом нарушении.
37. Композиция, содержащая выделенные клетки по п.31 и по меньшей мере один фармацевтически приемлемый носитель.
38. Способ доставки белка через гематоэнцефалический барьер в центральную нервную систему, где указанный способ включает введение субъекту композиции по п.37.
39. Способ доставки белка через гематоэнцефалический барьер в центральную нервную систему, где указанный способ включает введение субъекту композиции, содержащей
а) по меньшей мере один комплекс, включающий терапевтический полипептид и блок сополимер, где указанный блок сополимер включает по меньшей мере один растворимый в воде неионный сегмент и по меньшей мере один полиионный сегмент, где указанный полиионный сегмент включает по меньшей мере один заряд, противоположный заряду терапевтического полипептида, где указанный комплекс имеет морфологию ядро-оболочка, где ядро содержит терапевтический полипептид и полиионный сегмент блок сополимера и где оболочка содержит водорастворимый неионный сегмент блок сополимера, и
б) по меньшей мере один фармацевтически приемлемый носитель.
| VINOGRADOV S.V | |||
| ET AL.: 'Nanosized Cationic Hydrogels for Drug Delivery: Preparation, Properties and Interaction with Cells' ADVANCED DRUG DELIVERY REVIEWS vol | |||
| Видоизменение прибора для получения стереоскопических впечатлений от двух изображений различного масштаба | 1919 |
|
SU54A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| WU J | |||
| ET AL.: 'Quantitative Evaluation of Monocyte Transmigration into the Brain Following Chemical Opening of the Blood-Brain Barrier in Mice' | |||

