ЗАЯВЛЕНИЕ О ПРИОРИТЕТЕ
Настоящая заявка подтверждает полезность, в соответствии с п.35U.S.C. § 119(e), предварительной заявки на патент США, серия No. 61/220278, зарегистрированной 25 июня, 2009 г., полное содержание которой включено в настоящую заявку в качестве ссылки.
ЗАЯВЛЕНИЕ ОТНОСИТЕЛЬНО ПРАВИТЕЛЬСТВЕННОЙ ПОДДЕРЖКИ ИССЛЕДОВАНИЙ
Настоящее изобретение было сделано при правительственной поддержке в рамках гранта номер 5-P01-HL06350, выданного Национальному Институту Здоровья. Соответственно, правительство США имеет определенные права на настоящее изобретение.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к химерным полипептидам человеческого фактора свертывания крови VII (FVII), обладающим повышенной свертывающей активностью и меньшими тромботическим осложнениями, чем доступные в настоящее время полипептиды фактора VII, а также к полинуклеотидным конструкциям, кодирующим такие полипептиды, к векторам и клеткам-хозяевам, включающим и экспрессирующим указанные полинуклеотиды, к фармацевтическим композициям, их применению и способам лечения.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Свертывание крови представляет собой процесс, состоящий из целого ряда сложных взаимодействий различных компонентов (или факторов) крови, которые в итоге приводят к образованию фибринового сгустка. В основном, компоненты крови, которые участвуют в том, что называется коагуляционный "каскад", представляют собой энзиматически неактивные белки (пронзимы или зимогены), которые превращаются в протеолитические ферменты под действием активатора (который сам представляет собой активированный фактор свертывания крови).
Факторы свертывания крови, которые подвергаются таким превращениям, в целом называются "активными факторами", и на письме такие факторы обозначают с добавлением буквы "a" к наименованию соответствующего фактора свертывания крови (например, активированный фактор VII будет обозначаться как фактор VIIa или FVIIa).
В норме, инициация гемостатического процесса опосредована образованием комплекса между тканевым фактором и фактором VIIa. Этот комплекс затем превращает факторы IX (FIX) и X (FX) в их активные формы. Фактор Xa (FXa) превращает ограниченные количества протромбина в тромбин на клетках, содержащих тканевой фактор. Тромбин активирует тромбоциты и факторы V (FV) и VIII (FVIII) в факторы Va (FVa) и VIIIa (FVIIIa), где оба выполняют функцию кофакторов в дальнейшем процессе, ведущем к полной тромбиновой активации. Этот процесс включает образование фактора Xa под действием фактора IXa (FIXa) (в комплексе с фактором VIIIa) и происходит на поверхности активированных тромбоцитов. В конечном счете, тромбин превращает фибриноген в фибрин, что приводит к образованию фибринового сгустка. Исследования последних лет показали, что фактор VII и тканевой фактор являются основными инициаторами процесса свертывания крови.
Фактор VII представляет собой гликопротеин плазмы, который циркулирует в крови в виде одноцепочечного зимогена. Зимоген обладает слабой каталитической активностью. Одноцепочечный фактор VII может быть превращен в двухцепочечный фактор VIIa под действием фактора Xa, фактора XIIa, фактора IXa, фактора VIIa или тромбина in vitro. Считается, что фактор Xa является основным физиологическим активатором фактора VII. Как и в случае некоторых других белков, вовлекаемых в гемостаз, активность фактора VII зависит от витамина K, который необходим для гамма-карбоксилирования множества остатков глютаминовой кислоты, которые объединяются в кластеры вблизи амино-конца белка. Эти гамма-карбоксилированные глютаминовые кислоты необходимы для индуцированного ионом металла взаимодействия фактора VII с фосфолипидами. Превращение зимогена фактора VII в активированную двухцепочечную молекулу происходит путем расщепления пептидной связи во внутреннем пептиде Arg152-Ile153. Кроме того, хорошо известно, что высокие концентрации фактора VII ведут к аутоактивации in vitro. В присутствии тканевого фактора, фосфолипидов и ионов кальция, двухцепочечный фактор VIIa быстро активирует фактор X или фактор IX путем ограниченного протеолиза.
Ген, кодирующий FVII (hFVII), был картирован на хромосоме 13 в положении q34-qter 9 (de Grouchy et al., Hum Genet 1984; 66:230-233). Он содержит девять экзонов и охватывает 12,8 тыс. н.п. (O'Hara et al., Proc Natl Acad Sci USA 1987; 84:5158-5162). Организация гена и структура белка FVII аналогичны таковым для К-зависимых белков с прокоагулянтной активностью, где экзоны 1a и 1b кодируют сигнальную последовательность; экзон 2 - пропептид и домен GLA; экзон 3 - короткий гидрофобный участок; экзоны 4 и 5 - домены GLA, подобные таковым для эпидермального фактора роста; и экзоны 6-8 - каталитический домен GLA сериновой протеазы (Yoshitake et al., Biochemistry 1985; 24: 3736-3750).
Фактор IX (кристмас-фактор) представляет собой зимоген сериновой протеазы, активный в нормальном процессе гемостаза, и в этом случае для проявления ферментативной активности требуется карбоксилирование специфических остатков глютаминовой кислоты. Факторы IX, X, VII и белок C являются близко родственными паралогами одного семейства сериновых протеаз, с высокой степенью идентичности по аминокислотной последовательности и соответствующей интрон-экзоновой организацией генов, кодирующих эти белки. Указанные близко родственные белки характеризуются сходной структурой функциональных доменов GLA от амино- до карбокси-конца и включают домен GLA γ-карбоксиглютаминовой кислоты (GLA), два домена GLA фактора, подобного эпидермальному фактору роста (EGF), пептид активации и каталитический домен. Белок S включает 666 аминокислот, витамин K-зависимый белок с доменом GLA, 4 EGF-подобных домена, чувствительный к тромбину участок и 2 домена ламинина.
Зависимые от витамина K белки свертывания плазмы крови содержат домен GLA, который функционирует как сайт прикрепления белка к мембранам, где указанный домен GLA характеризуется высокой консервативностью среди различных коагулирующих белков. Несмотря на сходство, домены GLA демонстрируют широко варьирующую аффинность в отношении фосфолипидов, при этом домен GLA белка S обладает наивысшей аффинностью для фосфолипидов. (Ellison et al., Biochemistry, 1998; 37:7997-8003), (McDonald et al., Biochemistry 1997; 36:5120-27).
Зачастую бывает желательно стимулировать или усиливать коагулирующий каскад в системе свертывания крови у субъекта. Фактор VIIa применялся для контроля расстройств, сопровождающихся кровотечением, которые имеют несколько причин, таких как недостаточность фактора свертывания крови (например, гемофилия A и B или недостаточность факторов свертывания крови XI или VII) или присутствие ингибиторов факторов свертывания. Фактор VIIa применялся также для контроля сильного кровотечения у субъектов с нормально функционирующей системой свертывания крови (в отсутствие недостаточности фактора свертывания крови или ингибиторов каких-либо факторов свертывания крови). Такое кровотечение может быть, например, вызвано дефектом в тромбоцитарной функции, тромбоцитопенией или болезнью Виллебранда.
Кровотечение является одной из важнейших проблем хирургии и в случае других форм травмы. Например, фактор VII широко применялся для лечения раненых солдат в Ираке и Афганистане. (Perkins JG, et al. The Journal of Trauma. 2007; 62: 1095-9; discussion 9-101). Его использование позволило спасти много жизней, но во многих случаях медицинского применения, после лечения этим средством наблюдаются побочные эффекты, такие как инсульт или другие тромботические явления. Однако, согласно общему мнению врачей, использующих FVIIa, его применение спасло значительно больше жизней, чем потеряло. Возможно, самой хорошей иллюстрацией этого является то, что в предшествующих войнах примерно 30 процентов раненых умирало от полученных повреждений, тогда как во время военных действий в Персидском заливе это количество сократилось примерно до 10 процентов. (Gawande A, et al., N Engl J Med. 2004;351:2471-5).
Исследование гемофилии B у трансгенных мышей, экспрессирующих фактор VIIa, показало, что постоянная экспрессия фактора VIIa на низких уровнях (ниже 1,5 мкг/мл) восстанавливает свертывающую активность у мышей с геомофилией В. Однако, уровни фактора VIIa у мышей дикого типа или у мышей с гемофилией В свыше 2 мкг/мл ведут к тромбозам в сердце и легких; при этом и сердце, и легкие являются сайтами высокой экспрессии тканевого фактора. Эти данные позволяют полагать, что высокие уровни фактора VIIa в кровотоке индуцируют тромбоз в том случае, если они контактируют с тканевым фактором, который становится доступным после повреждения сосудов в сердце и легких. (Margaritas et al., J. Clin. Invest. 2004; 113:1025-31). Кроме того, эти исследования показали, что осуществляемый через вектор перенос от собаки гена фактора VIIa собакам с гемофилией является и безопасным, и эффективным подходом в краткосрочной и среднесрочной перспективе. (Margaritas et al., Gene Therapy 2009; 113:3682-3689).
Ввиду тех опасностей, с которыми сопряжено лечение фактором VII, этот продукт, как и многие другие продукты, должен применяться по разрешению и в соответствии с рекомендациями регламентирующих органов. Например, по данным Европейского Медицинского Агентства (The European Medicines Agency), Подразделение по оценке препаратов для применения человеком (Human Medicines Evaluation Unit), проводимая в настоящее время терапия фактором VII сопряжена с риском тромбоза и развитием синдрома диссеминированного внутрисосудистого свертывания, особенно в тех ситуациях, когда фактор VII вводится пациентам с наличием в анамнезе ишемической болезни сердца или болезни печени, пациентам после проведения операций, новорожденным и лицам, имеющим риск тромбоза и диссеминированной внутрисосудистой коагуляции. См., например, Core SPC for Human Plasma Derived Coagulation Factor VII Products (CPMP/BPWG/2048/01), July, 2004.
Ранее было показано, что домен EGF-1 фактора VIIa играет решающую роль в определении аффинности фактора VIIa для тканевого фактора. С использованием синтетического субстрата и фактора X, в обоих случаях при наличии и в отсутствие тканевого фактора, было показано, что полипептиды фактора VIIa с доменом EGF-1 фактора IX обладают меньшей каталитической активностью, чем фактор VIIa дикого типа. (Jin et al., Biochemistry, 1999, 28:1185-92). С первого взгляда, может сложиться впечатление, что это позволяет обходиться без использования химерных конструкций для лечения кровотечений; однако, в соответствии с предположением Monroe (British Journal of Haematology 1997; 99:542-549), механизм действия FVIIa при лечении гемофилии и кровотечения не зависит от тканевого фактора. При этом, общее мнение специалистов в данной области разделилось в вопросе об активности фактора VIIa, независимого от тканевого фактора.
Коммерческие препараты рекомбинантного человеческого FVIIa продаются под торговой маркой NovoSeven® и NovoSeven® RT. Указанные препараты NovoSeven® и NovoSeven® RT показаны для лечения эпизодов кровотечения у пациентов с гемофилией A или B, и на фармацевтическом рынке доступен лишь препарат rFVIIa для лечения эпизодов кровотечения. В последнее время было показано, что NovoSeven® может связываться с регидратированными лиофилизированными тромбоцитами, которые могут вводиться в сочетании для локализации фактора VII в сайте повреждения. (Fischer et al., Platelets, 2008; 19:182-91). Дополнительно, было показано, что селективное ПЭГилирование фактора VII может повышать его период полувыведения из плазмы (Stennicke et al., Thromb. Haemost, 2008; 100:920-28) и что рекомбинантный человеческий фактор VII с 3 аминокислотными замещениями имеет повышенную активность на поверхности тромбоцитов. (Moss et al., J. Thromb. Haemost., 2009; 7:299-305). Ожидается, что ПЭГилирование химерных молекул фактора VIIa по настоящему изобретению будет действовать в том же направлении, приводя к повышению периода полувыведения из плазмы. Аналогично, можно полагать, что другие модификации белков, известные в данной области, такие как ковалентное присоединение не полипептидных фрагментов с образованием конъюгатов, например, гликозилирование, будут действовать аналогично молекулам фактора VIIa по настоящему изобретению, т.е., можно ожидать, что свойства, придаваемые белку за счет ковалентного присоединения не полипептидного фрагмента будут переданы также химерным молекулам фактора VII.
В этой связи, имеется потребность в вариантах фактора VIIa, обладающих высокой свертывающей способностью, которые могут вводиться в относительно низких дозах, и в вариантах, которые создают меньше нежелательных побочных эффектов, таких как тромботические осложнения, ассоциированные с проводимой в настоящее время терапией.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В настоящей заявке описываются химерные молекулы фактора FVIIa, в частности, химерные молекулы hFVIIa, включающие домены hFVIIa и домены из одного или нескольких белков системы свертывания крови, обеспечивающих одно или несколько желательных качеств. В этой связи, химерные молекулы FVIIa по настоящему изобретению, характеризуются одним или несколькими улучшенными свойствами, в сравнении с коммерчески доступным rFVIIa, включая такие показатели, как более высокая свертывающая активность и/или возможность их введения в относительно низких дозах и/или снижение возникающих при этом неблагоприятных побочных эффектов. Следовательно, медицинское лечение химерными молекулами по настоящему изобретению дает преимущества в сравнении с доступным в настоящее время соединением rFVIIa, такие как потенциально сниженные дозы и/или ослабленные нежелательные побочные эффекты.
Репрезентативные химерные полипептиды FVIIa по настоящему изобретению включают химерный FVIIa, включающий EGF-2 и каталитические домены FVII и домен GLA витамин K-зависимого белка коагуляции и домен EGF-1 витамин K-зависимого белка коагуляции. В конкретных вариантах осуществления настоящего изобретения, химерные полипептиды FVIIa по настоящему изобретению включают 1) химерный FVIIa, включающий домены GLA и EGF-1 из FIX и домен EGF-2 и каталитический домен FVII; 2) химерный FVIIa, включающий домен EGF-1 из FIX и домены GLA, EGF-2 и каталитический домен FVII; 3) химерный FVIIA, включающий домен GLA белка S, домен EGF-1 из FIX и домен EGF-2 и каталитический домен FVII; 4) химерный FVIIa, включающий домены GLA и EGF-1 белка S и домен EGF-2 и каталитический домен FVII; 5) химерный FVIIa, включающий домен EGF-1 белка S и домены GLA, EGF-2 и каталитический домен FVII; и 6) химерный FVIIa, включающий домен EGF-1 белка S, домен GLA из FIX и домен EGF-2 и каталитический домен FVII. Репрезентативные химерные полипептиды FVIIa по настоящему изобретению могут также включать FVIIa дикого типа или любой из описанных выше химерных полипептидов FVIIa, которые содержат аминокислотные замещения в домене EGF-1 или в домене GLA. Указанные замещения могут представлять собой консервативные замещения или неконсервативные замещения. Такие замещения могут включать замещение изолейцина по остатку 69 аланином и/или замещение аргинина по остатку 79 аланином из домена EGF-1. Дополнительные замещения могут включать замещение лизина по остатку 5 аргинином, которое придает домену GLA более высокую связывающую аффинность с коллагеном типа IV, но, по всей видимости, не затрагивает связывание с тромбоцитами (Gui et al., J. Thromb Haemost. 2009; 7:1843-1851); замещение метионина по остатку 306 другой аминокислотой, предпочтительно консервативное аминокислотное замещение, более предпочтительно аланином, что также снижает аффинность в отношении тканевого фактора; и замещение валина по остатку 158 аспартатом, замещение глютамата по остатку 296 валином и/или замещение метионина по остатку 298 глютамином, что приводит к повышению удельной активности фактора VIIa.
Настоящее изобретение также относится к способам лечения расстройств, ассоциированных с кровотечением, у субъекта с указанным расстройством путем введения одного или нескольких химерных полипептидов FVIIa по настоящему изобретению. Способ лечения расстройства, ассоциированного с кровотечением, может включать способ введения указанному субъекту молекулы нуклеиновой кислоты, включающей нуклеотидную последовательность, кодирующую полипептид фактора VIIa по настоящему изобретению.
Дополнительно, настоящее изобретении относится к способу лечения расстройства, ассоциированного с кровотечением, у субъекта, имеющего такое расстройство, путем введения белка, включающего домен GLA, где указанный белок направлен и/или концентрируется вблизи сайта появления сгустка в определенной концентрации белка в кровотоке в плазме у субъекта. Это может включать использование доменов, которые связываются с большей аффинностью с тромбоцитами, находящимися на месте образования сгустка или в непосредственной близости от него, чем применяемые в настоящее время полипептиды фактора VII. Примеры таких доменов включают домен GLA разных белков коагуляции, которые связываются с отрицательно заряженными фосфолипидными слоями, локализованными на поверхности тромбоцитов. Неограничивающие примеры таких домен GLA включают домен GLA из FIX, который связывается более прочно, чем домен GLA из FVIIa, с тромбоцитами и фосфолипидами (Melton et al. "Location of the platelet binding site in zymogen coagulation factor IX" Blood Coagul Fibrinolysis 12:237-243 (2001)) и домен GLA белка S, который связывается с фосфолипидами более прочно, чем любой другой известный домен GLA (McDonald et al. "Comparison of naturally occurring vitamin K-dependent proteins: correlation of amino acid sequences and membrane binding properties suggests a membrane contact site" Biochemistry:36:5l20-5127 (1997)). Неограничивающие примеры белка, содержащего домен GLA, включают рекомбинантный белок, включающий домен GLA из FIX или белка S, химерный белок, включающий домен GLA из FIX или белка S, и/или химерный полипептид FVIIa по настоящему изобретению, включающий домен GLA из FIX или белка S.
Настоящее изобретение, в другом варианте своего осуществления, относится к способу лечения субъекта, имеющего расстройство, ассоциированное с кровотечением, путем введения химерного полипептида фактора VIIa по настоящему изобретению, который включает каталитический домен, полученный из полипептида фактора VII, и один или несколько доменов, направленных на тромбоциты. Такие направленные на тромбоциты домены включают домены из белков, которые взаимодействуют или связываются с поверхностью тромбоцита. Такое взаимодействие или связывание с поверхностью тромбоцита может осуществляться через мембранные фосфолипиды тромбоцита или через белки и/или рецепторы клеточной поверхности тромбоцита. Неограничивающие примеры таких доменов включают Al домен фактора фон Виллебранда, который является основным сайтом связывания с тромбоцитарным гликопротеином lb (Emsley et al., JBC, 273:10396-10401 (1998)), и Fab фрагмент (домен) из антител, которые связываются с мембранными белками и/или рецепторами тромбоцита, таких как антитело, которое связывается с мембранными фосфолипидами тромбоцита (Out et al., Blood, 77:2655-2659(1991)).
Такое нацеливание коагулирующих белков на сайт сгустка с использованием домена, который связывается с тромбоцитами и имеет сниженную аффинность к тканевому фактору, обладает дополнительным и неожиданным преимуществом, а именно: снижением риска осложнений, ассоциированных с применяемой в настоящее время терапией фактором VII, такого как тромбогенность. Указанный подход может также использоваться для нацеливания других терапевтически полезных полипептидов или других молекул на тромбоциты или на сайт сгустка. Такие терапевтически полезные молекулы могут включать антикоагулянты и т.п.
Настоящее изобретение также относится к способу лечения расстройства, ассоциированного с кровотечением, у субъекта, имеющего такое расстройство, путем введения такому субъекту полипептида со сниженной тромбогенностью, в сравнении с тромбогенностью фактора VII (например, сниженной по меньшей мере примерно на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 90%, и т.п. относительно тромбогенности нативного фактора VII). Указанная "сниженная тромбогенность" может быть определена разными способами по известным методикам, включающим, без ограничения, определение на основании выявления сниженного числа сгустков, сгустков меньшего размера, большего периода времени, необходимого для образования сгустков (тест in vivo или in vitro), по меньшему числу летальных исходов от образования тромбов и/или по более длительному периоду выживания, в сравнении с контролем. Полипептид, обладающий сниженной тромбогенностью в сравнении с фактором VII, может представлять собой, например, химерный полипептид фактора VII по настоящему изобретению или его активный фрагмент. Неограничивающие примеры таких полипептидов включают химерный FVIIa, включающий домены GLA и EGF-1 из FIX и EGF-2 и каталитические домены из FVII, и химерный FVIIa, включающий домен GLA белка S, домен EGF-1 из FIX и EGF-2 и каталитические домены из FVII.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 приведен схематический вид химерных молекул FVIIa, используемых в настоящем изобретении. FVII: Фактор VII; FIX: Фактор IX
На фиг. 2 проиллюстрировано образование тромбина (нМ) в нормальных условиях и в гемофильных условиях (с удаленным фактором IX и фактором VIII) в модельной клеточной системе гемофилии при разных концентрациях фактора VIIa дикого типа.
На фиг. 3 проиллюстрировано образование тромбина (нМ) в нормальных условиях и в гемофильных условиях (с удаленным фактором IX и фактором VIII) в модельной клеточной системе гемофилии при введении 50 нМ фактора VIIa дикого типа или 10 нМ химерного фактора VIIa.
На фиг. 4 показан пик тромбина, относительно контрольного значения, образуемого в модельной клеточной системе гемофилии при введении 50 нМ фактора VIIa дикого типа или 10 нМ химерного фактора VIIa.
На фиг. 5 проиллюстрировано отклонение показателя «остановка кровотечения» в анализе коагулирующей активности у мышей с гемофилией B без лечения, у мышей с гемофилией B при введении 2 мг/кг NovoSeven®, у мышей с гемофилией B при введении 2 мг/кг химерной молекулы FVIIa и у мышей дикого типа без лечения.
На фиг.6 показана последовательность репрезентативного химерного фактора VII по настоящему изобретению, включающая сигнальный участок, пропептид, домены GLA и EGF1 фактора IX (подчеркнуты) (SEQ ID NO: 1).
На фиг.7 показана последовательность репрезентативного химерного фактора VII по настоящему изобретению, включающая сигнальный участок, пропептид и EGF1 домен фактора IX (подчеркнуты), и домен GLA белка S (жирный шрифт) (SEQ ID NO: 2).
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Ниже приводится более подробное описание настоящего изобретения. При этом, настоящее изобретение может быть представлено в разных формах, однако, не ограничивается приведенными вариантами. Скорее, приведенные варианты даны для более полного представления настоящего изобретения и дополнительно иллюстрируют его для лучшего понимания специалистами в данной области.
Терминология, используемая в настоящем описании, дана для целей описания только конкретных вариантов и ее не следует трактовать в плане какого-либо ограничения настоящего изобретения. В тексте настоящего описания и в прилагаемой формуле изобретения, единственная форма артиклей ("a," "an" и "the") включает также множественную форму, если из контекста явно не следует обратное.
Если особо не указано иное, все используемые в настоящем описании термины (включая технические и научные термины) имеют те значения, которые являются стандартными для специалистов в той области, к которой относится настоящее изобретение. Следует также понимать, что термины, такие как термины, определяемые в обычно используемых словарях, следует рассматривать в контексте настоящей заявки согласно значению, которое соответствует данному контексту и релевантной области, и которое не следует трактовать в идеализированном или чересчур формализованном смысле, если особо не указано иное. Терминология, используемая в тексте настоящего изобретения, дана для целей описания только конкретных вариантов его осуществления и ее не следует трактовать в плане ограничения настоящего изобретения. Все публикации, патентные заявки, патенты и другие ссылки, процитированные в настоящем описании, включены в него полностью в качестве ссылок.
Кроме того, в тексте настоящего описания, форма «и/или» относится ко всем возможным сочетаниям соответствующих, одного или нескольких терминов, и включает эти сочетания, а когда их следует рассматривать в альтернативном смысле, обозначает отсутствие таких сочетаний («или»).
Если из контекста не следует иное, то следует особо отметить, что различные элементы настоящего изобретения, приведенные в настоящем описании, могут использоваться в любом сочетании.
Кроме того, согласно настоящему изобретению, в некоторых вариантах осуществления настоящего изобретения, любой элемент или сочетание элементов, приведенное в данном тексте, может быть исключен/исключено или пропущен/пропущено. Для пояснения этого положения, следует отметить, что если в описании указано, что комплекс включает компоненты A, B и C, этим конкретно указывается, что любой из компонентов A, B или C, или их сочетание может быть пропущено и отклонено, в единственном виде или в любом сочетании.
В контексте настоящего описания, такую фразу, как "состоящий по существу из" (и ее грамматические варианты) следует интерпретировать как включающую все процитированные материалы и стадии, а также такие, которые явно не влияют на базовые и новые характеристики, одну или несколько, заявленного изобретения. См., In re Herz, 537 F.2d 549, 551-52, 190 U.S.P.Q. 461, 463 (CCPA 1976); см. также MPEP § 2111.03. Таким образом, фразу "состоящий по существу из" в тексте настоящего описания не следует понимать как эквивалентную термину "включающий".
Термин "примерно", используемый в тексте настоящего описания применительно к измеряемому значению, такому как количество или концентрация и т.п., следует понимать как включающий все вариации в 20%, 10%, 5%, 1%, 0,5% или даже 0,1% от указанного количества.
Термин "активность" в тексте настоящего описания обозначает способность полипептида фактора VII превращать свой субстрат, фактор X в активный фактор Xa.
Термин "собственная активность" также включает способность образовывать тромбин на поверхности активированных тромбоцитов в отсутствие тканевого фактора.
Термин "N-концевой домен GLA" включает аминокислотную последовательность от примерно 61 аминокислотного остатка до примерно 105 остатка в аминокислотной последовательности SEQ ID NO: 18), аминокислотную последовательность от примерно 47 аминокислотного остатка до примерно 92 аминокислотного остатка в аминокислотной последовательности SEQ ID NO: 19, любые пост-трансляционные модификации идентифицированных выше аминокислотных последовательностей, любые консервативные замещения аминокислот в идентифицированных аминокислотных последовательностях, добавление аминокислотных остатков к идентифицированным аминокислотным последовательностям, делеции аминокислотных остатков из идентифицированных аминокислотных последовательностей или любую другую аминокислотную последовательность из белка коагулирующей системы, который связывается с фосфолипидными мембранами.
Термин "EGF-1" обозначает участок из 30-40 аминокислот, содержащий шесть цистеинов, обнаруженных первоначально в EGF (эпидермальный фактор роста), а также в совокупности белков, вовлекаемых в клеточную сигнальную функцию, и коагулирующих белков, где практически все известные EGF-подобные домены содержат дисульфидные связи 1-3, 2-4 и 5-6. Домен EGF-1 в факторе VII охватывает участок от примерно 106 аминокислотного остатка до примерно 142 остатка в аминокислотной последовательности SEQ ID NO: 18. Домен EGF-1 в факторе IX охватывает участок от примерно 93 аминокислотного остатка до примерно 129 аминокислотного остатка в аминокислотной последовательности SEQ ID NO: 19. Домен EGF-1 в белке S охватывает участок от примерно 117 аминокислотного остатка до примерно 155 остатка в аминокислотной последовательности SEQ ID NO: 20. Указанные аминокислотные последовательности могут включать любые пост-трансляционные модификации идентифицированных выше аминокислотных последовательностей, любые консервативные замещения аминокислот в идентифицированных аминокислотных последовательностях, любое добавление аминокислотных остатков к идентифицированным аминокислотным последовательностям и/или любые делеции аминокислотных остатков из идентифицированных аминокислотных последовательностей.
Термин "EGF-2" обозначает второй EGF-подобный домен в серии (из двух или более EGF-подобных доменов). Домен EGF-2 в факторе VII охватывает участок от примерно 147 аминокислотного остатка до примерно 188 остатка в аминокислотной последовательности SEQ ID NO: 18. Домен EGF-2 в факторе IX охватывает участок от примерно 130 аминокислотного остатка до примерно 171 остатка в аминокислотной последовательности SEQ ED NO: 19. Домен EGF-2 в белке S охватывает участок от примерно 157 аминокислотного остатка до примерно 200 остатка в аминокислотной последовательности SEQ ID NO: 20. Указанные аминокислотные последовательности могут включать любые пост-трансляционные модификации идентифицированных выше аминокислотных последовательностей, любые консервативные замещения аминокислот в идентифицированных аминокислотных последовательностях, любое добавление аминокислотных остатков к идентифицированным аминокислотным последовательностям и/или любые делеции аминокислотных остатков из идентифицированных аминокислотных последовательностей.
Термин "каталитический домен" в тексте настоящего описания обозначает домен в белке, который участвует в расщеплении пептидных связей. Каталитический домен в факторе VII охватывает участок от примерно 213 аминокислотного остатка до примерно 452 аминокислотного остатка в аминокислотной последовательности SEQ ID NO: 18. Каталитический домен в факторе IX охватывает участок от примерно 227 аминокислотного остатка до примерно 459 аминокислотного остатка в аминокислотной последовательности SEQ ID NO: 19. Указанные аминокислотные последовательности могут включать любые пост-трансляционные модификации идентифицированных выше аминокислотных последовательностей, любые консервативные замещения аминокислот в идентифицированных аминокислотных последовательностях, любое добавление аминокислотных остатков к идентифицированным аминокислотным последовательностям и/или любые делеции аминокислотных остатков из идентифицированных аминокислотных последовательностей.
Трехбуквенная аббревиатура "GLA" обозначает 4-карбоксиглютаминовую кислоту (γ- карбоксиглютамат).
Термин "протеазный домен" обозначает домен в белке, который вовлекается в расщепление пептидных связей и который, как считается, в основном локализован на участке от примерно 213 аминокислотного остатка до карбокси-концевого аминокислотного остатка в SEQ ID NO: 18. (тяжелая цепь фактора VIIa). Термин "полипептид фактора VII" в контексте настоящего описания обозначает любой белок, включающий аминокислотную последовательность 61-466 из нативного человеческого фактора VII (SEQ ID NO: 18) или ее варианты или фрагменты. Указанный термин включает, без ограничения, человеческий фактор VII, человеческий фактор VIIa и их варианты. Термин "фактор VII" в контексте настоящего описания включает молекулу неактивного одноцепочечного зимогена фактора VII, а также молекулу активированного двуцепочечного фактора VII (фактор VIIa). Этот термин включает белки с аминокислотной последовательностью 61-466 (SEQ ID NO: 18) из нативного человеческого фактора VII или фактора VIIa. Для любого специалиста в данной области очевидно, что минорные изменения в данной последовательности будут, как ожидается, действовать аналогичным образом и что вовлекаемые в процесс домены, полипептиды и фактор VII будут несколько сокращаться в размере или удлиняться или будут содержать изменения, не влияющие на суть настоящего изобретения. Таким образом, данное определение также охватывает белки и пептиды с несколько модифицированной аминокислотной последовательностью, например, с модифицированной N-частью, где указанные модификации включают делеции или добавления N-концевых аминокислот, так что, в некоторых вариантах осуществления настоящего изобретения, рассматриваемые в нем белки по существу сохраняют активность фактора VIIa (например, сохраняют примерно 50%, 60%, 70%, 80%, 90%, 95% и т.п. от активности нативного фактора VIIa). Термин "фактор VIIa" или "FVIIa" в контексте настоящего описания обозначает продукт, состоящий из активированной формы (фактор VIIa). Термин "фактор VII" или "фактор VIIa" в указанном выше определении также включает природные аллельные вариации, которые могут существовать и которые встречаются от одного индивидуума к другому.
Кроме того, степень и локализация гликозилирования или других пост-трансляционных модификаций могут варьировать, в зависимости от экспрессии в выбранных клетках-хозяевах, ткани или виде животного и от природы организма-хозяина или от тканевой среды.
Термин "домен", в контексте настоящего описания, обозначает часть белковой последовательности и структуру, которая при этом может возникать, функционировать и существовать независимо от остальной части белковой цепи. Домен способен формировать компактную трехмерную структуру и зачастую она может, независимо от остальной части, демонстрировать стабильность и создавать складчатую организацию. Один домен может появляться в широком множестве эволюционно родственных белков. Домены могут варьировать по длине от примерно 25 аминокислот до примерно 500 аминокислот. Термин "домен" может также включать домен белка дикого типа, где указанный белок дикого типа содержал, один или несколько аминокислотных остатков, которые были изменены путем консервативного замещения. Поскольку они обладают автономной стабильностью в имеющейся белковой среде, то с использованием методов генетической инженерии такие домены могут быть «замещены» на участке между одним и другим белком с образованием химерных белков.
Термин "вариант" или "варианты" в контексте настоящего описания используется для обозначения фактора VII с аминокислотной последовательностью 61-466 (SEQ ID NO: 18) из нативного фактора VII или фактора VIIa, где одна или несколько аминокислот из родительского белка были замещены другой аминокислотой и/или где одна или несколько аминокислот из родительского белка были делетированы и/или где одна или несколько аминокислот были встроены в белок и/или где одна или несколько аминокислот были добавлены к родительскому белку и/или где домен GLA был замещен доменом GLA, взятым из другого белка (например, доменом GLA, который связывается с мембранами тромбоцитов или с фосфолипидными мембранами), и/или где домен GLA был замещен связывающимся с тромбоцитами доменом, взятым из другого белка (например, Al доменом фактора Виллебранда, который является главным сайтом связывания тромбоцитарного гликопротеина lb (Emsley et al., JBC, 273:10396-10401 (1998)), и Fab фрагментом (доменом) антител, который связывается с белками и/или рецепторами тромбоцитарной мембраны, как в случае антитела, которое связывается с фосфолипидами тромбоцитарной мембраны (Out et al., Blood, 77:2655-2659 (1991)), и/или где домен EGF-1 фактора VII был замещен доменом EGF-1 из другого белка (например, доменом EGF-1, который связывается с меньшей аффинностью с тканевым фактором, таким как домен EGF-1 фактора IX). Такое добавление может происходить либо на N-концевом участке, либо на C-концевом участке родительского белка, или на обоих указанных участках белка, а также на его внутренней части.
Таким образом, в некоторых вариантах осуществления настоящего изобретения, рассматриваемый/ые в настоящем изобретении "вариант" или "варианты" может/могут все еще обладать коагулирующей активностью FVII, свойственной его активированной форме. Однако, в некоторых вариантах осуществления настоящего изобретения, рассматриваемый/ые в настоящем изобретении "вариант" или "варианты" может/могут не обладать коагулирующей активностью.
Таким образом, в некоторых вариантах осуществления настоящего изобретения, рассматриваемый в нем вариант по меньшей мере на 40%, 50%, 60% или 70% идентичен аминокислотной последовательности 61-466 (SEQ ID NO: 18) из нативного человеческого фактора VII или фактора VIIa. Например, домен EGF-1 из FVII характеризуется идентичностью на уровне 65,7% с доменом EGF-1 из FIX, и домен GLA из FVII характеризуется идентичностью на уровне 58,6% с доменом GLA из FIX и идентичностью на уровне 51% с доменом GLA белка S. В одном варианте осуществления настоящего изобретения, рассматриваемый в нем вариант по меньшей мере на 80% идентичен аминокислотной последовательности 61-466 (SEQ ID NO: 18) из нативного человеческого фактора VII или фактора VIIa. В другом варианте осуществления настоящего изобретения, рассматриваемый в нем вариант по меньшей мере на 90% идентичен аминокислотной последовательности 61-466 (SEQ ID NO: 18) из нативного человеческого фактора VII или фактора VIIa. В еще одном варианте осуществления настоящего изобретения, рассматриваемый в нем вариант по меньшей мере на 95% идентичен аминокислотной последовательности 61-466 (SEQ ID NO.18) из нативного человеческого фактора VII или фактора VIIa.
Термин "любая другая аминокислота", в контексте настоящего описания, обозначает аминокислоту, отличающуюся от той аминокислоты, которая присутствует в данном положении в естественном состоянии. Указанный термин включает, без ограничения, аминокислоты, которые могут кодироваться полинуклеотидом. Предпочтительно, такая другая аминокислота является природной аминокислотой в своей естественной L-форме и может кодироваться полинуклеотидом. Соответствующим конкретным примером является L-цистеин (Cys).
В контексте настоящего описания, термин "оперативно связанный" (и его грамматические формы) относится к ковалентному соединению двух или более аминокислотных последовательностей путем ферментативного лигирования или другим способом, с образованием такой конфигурации, относительно друг друга, объединяемых последовательностей, которая позволяет осуществлять их нормальное функционирование. Например, нуклеотидная последовательность, кодирующая пред-последовательность или секреторный лидер, оперативно связана с нуклеотидной последовательностью, кодирующей полипептид, если он экспрессируется как пред-белок, который участвует в секреции полипептида: промотор или энхансер оперативно связан с кодирующей последовательностью, если он влияет на транскрипцию данной последовательности; сайт связывания рибосомы оперативно связан с кодирующей последовательностью, если он расположен таким образом, что способствует трансляции данного белка или пептида. В основном, термин "оперативно связанный" означает, что объединяемые нуклеотидные последовательности являются непрерывными, и в случае секреторного лидера, непрерывными и находящимися в рамке считывания. Связывание легче всего осуществляется путем лигирования в подходящих для этого сайтах рестрикции. Если таких сайтов нет, то используют синтетические олигонуклеотидные адапторы или линкеры, в сочетании со стандартными методами рекомбинантных ДНК.
Термин "вектор", в контексте настоящего описания, обозначает любой элемент нуклеиновой кислоты, способный к амплификации в клетке-хозяине. Соответственно, указанный вектор может представлять собой автономно реплицирующийся вектор, т.е. вектор, который существует в виде внехромосомного элемента и репликация которого не зависит от хромосомной репликации, например, плазмиды. Альтернативно, указанный вектор может представлять собой такой вектор, который, при введении его в клетку-хозяина, интегрируется в геном клетки-хозяина и реплицируется вместе с хромосомой(ами), в которую(ые) он был интегрирован. Выбор вектора часто зависит от клетки-хозяина, в которую его предстоит ввести. Векторы включают, без ограничения, плазмидные векторы, фаговые векторы, вирусы или космидные векторы. Векторы обычно содержат ориджин репликации и по меньшей мере один селектируемый ген, т.е. ген, кодирующий продукт, который легко детектируется или присутствие которого необходимо для роста клеток.
Термин "клетка-хозяин", в контексте настоящего описания, обозначает любую клетку, включая гибридные клетки, в которых может экспрессироваться гетерологичная ДНК. Типичные клетки-хозяева включают, без ограничения, клетки насекомых, дрожжевые клетки, клетки млекопитающих, включая человеческие клетки, такие как BHK, CHO, HEK и COS клетки. В практике осуществления настоящего изобретения, культивируемые клетки-хозяева представляют собой предпочтительно клетки млекопитающих, более предпочтительно, установленную линию клеток млекопитающих, включающую, без ограничения, клеточные линии CHO (например, ATCC CCL 61), COS-1 (например, ATCC CRL 1650), почки детеныша хомяка (BHK) и HEK293 (например, ATCC CRL 1573; Graham et al., J. Gen. Virol. 36:59-72, 1977). Подходящая клеточная линия BHK представляет собой клеточную линию tk-tsl3 BHK (Waechter and Baserga, Proc. Natl. Acad. Sci. USA 79:1106-1110, 1982), которая далее будет обозначаться как линия клеток BHK 570. Клеточная линия BHK 570 доступна от Американской Коллекции типовых культур (American Type Culture Collection, 10801 University, Boulevard, Manassas, Va. 20110), с номером доступа в ATCC CRL 10314. Клеточная линия tk-ts13 BHK также доступна от ATCC с номером доступа CRL 1632. Другие подходящие клеточные линии включают, без ограничения, клеточные линии Rat Hep I (гепатома крысы; ATCC CRL 1600), Rat Hep II (гепатома крысы; ATCC CRL 1548), TCMK (ATCC CCL 139), линия клеток легкого человека (ATCC HB 8065), NCTC 1469 (ATCC CCL 9.1) и клетки DUKX (Urlaub and Chasin, Proc. Natl. Acad. Sci. USA 77:4216-4220, 1980). Кроме того, могут использоваться клетки 3T3, клетки Namalwa, миеломы и продукты слияния миелом с другими клетками.
В контексте настоящего описания, термин "подходящая ростовая среда" обозначает среду, содержащую питательные элементы и другие компоненты, необходимые для роста клеток и экспрессии последовательности нуклеиновой кислоты, кодирующей полипептид фактора VII по настоящему изобретению.
Термин "конъюгат" (или используемый взаимозаменяемо термин "конъюгированный полипептид") означает в контексте настоящего описания гетерогенную (такую как композитную или химерную) молекулу, образуемую путем ковалентного присоединения одного или нескольких полипептидов к одному или нескольким не полипептидным фрагментам, таким как полимерные молекулы, липофильные соединения, сахарные фрагменты или органические дериватизирующие агенты. Предпочтительно, указанный конъюгат растворим при релевантных концентрациях и в соответствующих условиях, т.е. растворим в физиологических жидкостях, таких как кровь. Примеры конъюгированных полипептидов по настоящему изобретению включают гликозилированные и/или ПЭГилированные полипептиды.
Термин "ковалентное присоединение" означает, что полипептид и не полипептидный фрагмент либо напрямую были ковалентно соединены друг с другом, либо каким-то образом опосредованно были ковалентно соединены друг с другом через один или несколько промежуточных фрагментов, таких как мостиковые, спейсерные или связующие фрагменты, один или несколько. Термин "неконъюгированный полипептид" может использоваться применительно к полипептидной части конъюгата.
При использовании в контексте настоящего описания, термин "не полипептидный фрагмент" обозначает молекулу, которая способна конъюгировать с присоединяемой группой полипептида по настоящему изобретению. Подходящие примеры таких молекул включают полимерные молекулы, сахарные фрагменты, липофильные соединения или органические дериватизирующие агенты. Термин «конъюгат», используемый в описании настоящего изобретения, следует понимать таким образом, что не полипептидный фрагмент соединен с полипептидной частью конъюгата через группу присоединения, имеющуюся в полипептиде. Как указывалось выше, такой не полипептидный фрагмент может быть непосредственно ковалентно присоединен к группе присоединения или он может опосредованно соединен с группой присоединения через один или несколько вклинивающихся фрагментов, таких как мостиковые, спейсерные или связующие фрагменты, один или несколько.
Термин "полимерная молекула" обозначает молекулу, образуемую путем ковалентного связывания двух или более мономеров, где ни один из мономеров не представляет собой аминокислотный остаток, за исключением тех случаев, когда указанный полимер представляет собой человеческий альбумин или другой широко представленный в плазме белок. Термин "полимер" может использоваться взаимозаменяемо с термином "полимерная молекула". Указанный термин включает углеводные фрагменты, присоединяемые путем гликозилирования in vitro, т.е., синтетического гликозилирования, обычно осуществляемого in vitro путем ковалентного связывания молекулы углевода с группой присоединения на полипептиде, необязательно с использованием сшивающего агента.
Углеводная молекула, присоединяемая путем гликозилирования in vivo, такого как N- или O-гликозилирование (подробно описанное ниже), в настоящем описании обозначается как "сахарный фрагмент". За исключением тех случав, где число не полипептидных фрагментов, таких как полимерные молекулы, одна или несколько, или сахарные фрагменты в конъюгате явно указаны, любая ссылка на "не полипептидный фрагмент", содержащийся в конъюгате, или иным образом используемый в настоящем изобретении, будет представлять собой ссылку на один или несколько не полипептидных фрагментов, таких как полимерные молекулы, одна или несколько, или сахарные фрагменты.
Термин "группа присоединения" в контексте настоящего описания обозначает функциональную группу полипептида, в частности, его аминокислотный остаток, или углеводный фрагмент, способный присоединять не полипептидный фрагмент, такой как полимерная молекула, липофильная молекула, сахарный фрагмент или органический дериватизирующий агент.
Применительно к N-гликозилированию in vivo, термин "группа присоединения" используется в нестандартном варианте для указания аминокислотных остатков, состоящих из сайта N-гликозилирования (с последовательностью N-X-S/T/C, где X обозначает любой аминокислотный остаток, за исключением пролина, N обозначает аспарагин и S/T/C обозначает либо серин, треонин, либо цистеин, предпочтительно серин или треонин, и наиболее предпочтительно треонин). При том, что аспарагиновый остаток в сайте N-гликозилирования обозначает такой остаток, к которому присоединяется сахарный фрагмент в процессе гликозилирования, такое присоединение не может быть достигнуто, если не присутствуют другие аминокислотные остатки в сайте N-гликозилирования.
Соответственно, в том случае, когда не полипептидный фрагмент обозначает сахарный фрагмент и конъюгация осуществляется через N-гликозилирование, фразу "аминокислотный остаток, включающий группу присоединения для не полипептидного фрагмента", используемую применительно к изменениям в аминокислотной последовательности рассматриваемого полипептида, следует понимать как указание на то, что один или несколько аминокислотных остатков, составляющих сайт N-гликозилирования, изменяют таким образом, что функциональный сайт N-гликозилирования либо вводят в аминокислотную последовательность, либо удаляют из указанной последовательности.
В контексте настоящего описания, термин "лечить" или "лечение" включает и контроль, и/или профилактику ожидаемого кровотечения, такие как хирургия, и регулирование уже имеющегося кровотечения, такого как в случае травмы, или при необходимости лечения больных гемофилией, с целью подавления или минимизирования кровотечения. Соответственно, в данный термин "лечение" включается вариант профилактического введения полипептида фактора VIIa по настоящему изобретению.
Термин "эпизод кровотечения" в контексте настоящего описания включает неконтролируемое и избыточное кровотечение. Эпизоды кровотечения могут представлять собой основную проблему, как в случае хирургии, так и других форм повреждения тканей. Неконтролируемое и избыточное кровотечение может возникать у субъектов как с нормально функционирующей системой свертывания крови, так и у субъектов, имеющих расстройства в системе свертывания крови или расстройства, ассоциированные с кровотечением.
В контексте настоящего описания, термин "расстройство, ассоциированное с кровотечением" отражает любой дефект, врожденный, приобретенный или индуцированный, который может быть клеточного, физиологического или молекулярного происхождения и который проявляется в виде кровотечений. Соответствующие примеры включают недостаточность по фактору свертывания крови (например, гемофилия A и B или недостаточность факторов свертывания крови XI или VII), наличие ингибиторов факторов свертывания крови, дефектную функциию тромбоцитов, тромбоцитопению, болезнь Виллебранда или кровотечение, вызванное хирургическим вмешательством или травмой.
Избыточные кровотечения могут возникать у субъектов с нормально функционирующей системой свертывания (в отсутствие недостаточности факторов свертывания крови или ингибиторов какого-либо из факторов свертывания крови) и могут быть вызваны дефектом в тромбоцитарной функции, тромбоцитопенией или болезнью Виллебранда. В таких случаях, имеющиеся кровотечения могут напоминать кровотечения, вызываемые гемофилией, поскольку гемостатическая система, как и в случае гемофилии, не содержит или содержит аномальные "соединения", обязательные для функционирования системы коагуляции (такие как тромбоциты или белковый фактор фон Виллебранда), что вызывает обильные кровотечения. В случае субъектов, которые имели обширное повреждение ткани в результате хирургического вмешательства или травмы, нормальный гемостатический механизм может быть сдвинут из-за потребности в срочном гемостазе, так что у них может развиваться кровотечение, несмотря на нормальный гемостатический механизм.
Достижение удовлетворительного состояния гемостаза также становится проблемой при возникновении кровотечения в органах, таких как головной мозг, область внутреннего уха и глаза, где ограничена возможность хирургического вмешательства с целью остановки кровотечения. Такая же проблема может возникнуть при отборе образцов для биопсии из разных органов (печень, легкое, опухолевая ткань, желудочно-кишечный тракт), а также при лапароскопии. Все эти ситуации объединяет то, что весьма сложно остановить кровотечение с помощью хирургических методов (швы, гемостатические клипсы и т.п.), что имеет место также при диффузном кровотечении (геморрагический гастрит и профузное маточное кровотечение). Острые и профузные кровотечения могут также возникать у субъектов при проведении антикоагулянтной терапии, у которых дефектный гемостаз может быть индуцирован проводимой терапией. Такие субъекты нуждаются в хирургическом вмешательстве, в том случае, если требуется срочное противодействие антикоагулянтному эффекту.
Радикальная ретролобковая аденомэктомия представляет собой стандартную процедуру, проводимую в случае субъектов с локализованным раком предстательной железы. Эта операция часто осложняется заметной и иногда даже массивной потерей крови. Значительная потеря крови в ходе простатэктомии в основном связана со сложной анатомической ситуацией, при которой отмечается васкуляризованные сайты разной плотности, которые не легко доступны при хирургическом гемостазе и которые могут возникать при диффузном кровотечении на большой площади. Кроме того, интрацеребральная геморрагия представляет собой труднее всего поддающуюся лечению форму инсульта, которая ассоциирована с высокой смертностью и ростом гематомы в первые несколько часов после возникновения интрацеребральной геморрагии. Лечение с использованием rFVIIa может ограничивать рост гематомы, снизить смертность и улучшить функциональные результаты к 90 дням. Однако, при использовании доступного в настоящее время rFVIIa, имеется риск развития тромбоэмболических побочных реакций. Молекулы химерного фактора VII по настоящему изобретению, с характерной для них сниженной тромбогенностью, могут дать возможность преодолеть эту проблему при лечении инсульта. Другая ситуация, которая может вызывать проблемы в случае неудовлетворительного гемостаза, возникает у субъектов с нормальным механизмом гемостаза, когда им проводят антикоагулянтную терапию с целью предупреждения тромбоэмболической болезни. Такая терапия может включать гепарин, другие виды протеогликанов, варфарин или другие формы антагонистов витамина K, а также аспирин и другие ингибиторы агрегации тромбоцитов.
В одном варианте осуществления настоящего изобретения, кровотечение ассоциировано с гемофилией. В другом варианте осуществления настоящего изобретения, кровотечение ассоциировано с гемофилией, определяемой наличием приобретенных ингибиторов. В другом варианте осуществления настоящего изобретения, кровотечение ассоциировано с тромбоцитопенией. В другом варианте осуществления настоящего изобретения, кровотечение ассоциировано с болезнью Виллебранда. В другом варианте осуществления настоящего изобретения, кровотечение ассоциировано с серьезным повреждением ткани. В другом варианте осуществления настоящего изобретения, кровотечение ассоциировано с тяжелой травмой. В другом варианте осуществления настоящего изобретения, кровотечение ассоциировано с хирургией. В другом варианте осуществления настоящего изобретения, кровотечение ассоциировано с лапароскопией. В другом варианте осуществления настоящего изобретения, кровотечение ассоциировано с геморрагическим гастритом. В другом варианте осуществления настоящего изобретения, указанное кровотечение представляет собой профузное маточное кровотечение. В другом варианте осуществления настоящего изобретения, кровотечение происходит в органах с ограниченной возможностью остановки в них кровотечения с помощью механических способов. В другом варианте осуществления настоящего изобретения, кровотечение происходит в головном мозге, внутренней области уха или глазах. В другом варианте осуществления настоящего изобретения, кровотечение ассоциировано с процессом отбора образцов для биопсии. В другом варианте осуществления настоящего изобретения, кровотечение ассоциировано с антикоагулянтной терапией.
Термин "субъект" в контексте настоящего описания обозначает любое животное, в частности млекопитающих, таких как люди, и может, где это приемлемо, использоваться взаимозаменяемо с термином "пациент".
Термин "усиление нормальной гемостатической системы" обозначает улучшение способности данной системы создавать тромбин или образовывать функциональный сгусток крови.
Термин "генная терапия" относится к способу изменения экспрессии эндогенного гена за счет экзогенного введения гена. В контексте настоящего описания, термин "генная терапия" также относится к замене дефектного гена, кодирующего дефектный белок, или к замене недостающего гена путем введения функционального гена, соответствующего дефектному или утраченному гену, в соматические или стволовые клетки индивидуума, при наличии такой необходимости. Генная терапия может проводиться по методам, осуществляемым ex vivo, в соответствии с которыми дифференцированные или соматические стволовые клетки отбирают из организма индивидуума и затем вводят нормальную копию дефектного гена в отобранные клетки с использованием вирусного вектора, такого как носитель для генной доставки. Кроме того, существующие технологии непосредственного введения гена in vivo позволяют осуществлять перенос гена в клетки индивидуума in situ большого числа вирусных векторов, липосом, комплексов ДНК-белок или оголенной ДНК для достижения терапевтического результата. Термин "генная терапия" также относится к замене дефектного гена, кодирующего дефектный белок, путем введения полинуклеотида, который функционирует по существу так же, как и дефектный ген, или как функционировал бы белок, при условии, что они не являются дефектными, в соматические или стволовые клетки индивидуума, при наличии такой необходимости.
В настоящем изобретении, генная терапия используется для введения субъекту молекулы нуклеиновой кислоты, включающей нуклеотидную последовательность, кодирующую химерный FVII по настоящему изобретению. Соответственно, настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, включающей нуклеотидную последовательность, кодирующую химерный белок FVII по настоящему изобретению. Такая молекула нуклеиновой кислоты может присутствовать в конструкции нуклеиновой кислоты (например, в векторе или плазмиде). Такая конструкция нуклеиновой кислоты может присутствовать в клетке. Настоящее изобретение также относится к способу доставки химерного белка FVII по настоящему изобретению в клетку, включающему введение в клетку молекулы нуклеиновой кислоты, включающей нуклеотидную последовательность, кодирующую химерный белок FVII, в условиях, при которых указанная нуклеотидная последовательность экспрессируется с образованием в клетке химерного белка FVII. Указанная клетка может представлять собой клетку, которая вводится субъекту, и/или клетку, которая уже имеется у субъекта.
Получение химерного фактора VII
Полипептиды химерного фактора VII по настоящему изобретению могут быть получены с использованием технологии рекомбинантных нуклеиновых кислот. В основном, процедура состоит в том, что модифицируют клонированную последовательность нуклеиновой кислоты фактора VII дикого типа, с тем чтобы она кодировала желательный белок. Затем, указанную модифицированную последовательность встраивают в вектор экспрессии, который, в свою очередь, трансформируют или трансфицируют в клетки-хозяева. В качестве клеток-хозяев могут применяться клетки высших эукариотов, в частности, культивируемые клетки млекопитающих. В настоящее время известны полноразмерные нуклеотидная и аминокислотная последовательности для человеческого фактора VII (патент США No. 4784950, где описаны клонирование и экспрессия рекомбинантного человеческого фактора VII, а также приведены номера доступа в GenBank® No. J02933 и AAA51983 и номер доступа в SwissProt No. P08709-1) и описана аминокислотная последовательность SEQ ID NO: 18. Последовательность бычьего фактора VII описана в работе Takeya et al., J. Biol. Chem. 263:14868-14872 (1988)). Полноразмерные нуклеотидная и аминокислотная последовательности фактора IX известны (см. Davie et al.,Proc. Natl. Acad. Sci. U.S.A 1982; 79:6461-6464; Jaye et al., Nucleic Acids Res. 1983; 11:2325-2335; и McGraw et al., Proc. Natl. Acad. Sci. U.S.A. 1985; 82:2847-2851, описаны также номера доступа в GenBank® NoNo. J00136 и AAA98726 и номер доступа в SwissProt No.P00740), приведена аминокислотная последовательность SEQ ID NO: 19; а также полноразмерные нуклеотидная и аминокислотная последовательности для белка S (см., Hoskins et al., Proc. Natl.Acad. Sci. U.S.A. 1987; 84:349-353, номера доступа в GenBank® NoNo. Ml5036 и AAA36479, номер доступа в SwissProt No. P07225), а также описана аминокислотная последовательность SEQ ID NO: 20. Полноразмерные нуклеотидные и аминокислотные последовательности для других коагулирующих белков известны и могут быть просмотрены с использованием SwissProt или на вэб-сайте NCBI.
Изменения в аминокислотной последовательности могут быть достигнуты с помощью множества методик. Модификация последовательности нуклеиновой кислоты может быть выполнена с использованием сайт-специфического мутагенеза. Методы проведения сайт-специфического мутагенеза известны в данной области и описаны, например, в работах Zoller and Smith (DAM 3:479-488, 1984) или в работе «Splicing by extension overlap», Horton et al., Gene 77, 1989, pp. 61-68. Таким образом, с использованием нуклеотидной и аминокислотной последовательностей фактора VII можно ввести одно или несколько нужных изменений. Аналогично, для специалистов в данной области хорошо известны процедуры получения конструкций ДНК с использованием полимеразно-цепьевой реакции и специфических праймеров (см., PCR Protocols, 1990, Academic Press, San Diego, Calif,USA). Кроме того, полипептиды химерного фактора VII по настоящему изобретению могут быть получены при введении уникальных сайтов рестрикции в нуклеотидные последовательности, кодирующие различные полипептиды, которые могут использоваться для выделения фрагментов нуклеотидных последовательностей, кодирующих полные домены различных полипептидов. Далее, такие различные нуклеотидные последовательности могут быть подвергнуты рекомбинации с получением различных химерных полипептидов фактора VII по настоящему изобретению.
Например, в нуклеотидные последовательности, кодирующие различные коагулирующие белки, могут быть встроены уникальные сайты рестрикции, так что в этом случае домены GLA, EGF-1, EGF-2 и каталитический домен могут легко обмениваться между различными коагулирующими белками. Домен EGF-1 может свободно вращаться вокруг домена EGF-2 (Pike et al. Proc Natl. Acad. Sci. 1999; 96:8925-8930), тогда как домены GLA и EGF-1 могут свободно вращаться вокруг друг друга, когда присутствует кальций (Sunnerhagen et al. Nature Structural Biology 1995; 2:504-509). Согласно данному варианту осуществления настоящего изобретения, домены GLA и EGF-1 могут быть получены из одного и того же коагулирующего белка, но из белка, отличного от EGF-2 и каталитический домены. Химерные полипептиды фактора VIIa по настоящему изобретению могут, в этой связи, включать домены из любого коагулирующего белка, при условии, что по меньшей мере 50% одного домена представляет собой домен фактора VII. Кроме того, химерные полипептиды фактора VIIa по настоящему изобретению могут включать домены GLA и EGF-1 из одного и того же белка или из разных белков.
Конструкция нуклеиновой кислоты, кодирующая химерные полипептиды фактора VII по настоящему изобретению, может быть получена из генома или из кДНК, например, путем создания геномной библиотеки или библиотеки кДНК с последующим скринингом последовательностей ДНК, кодирующих полностью или части различных полипептидов, путем гибридизации с использованием синтетических олигонуклеотидных зондов, в соответствии с процедурами стандартных методик (см., Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd. Ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y., 1989).
Конструкция нуклеиновой кислоты, кодирующая химерные полипептиды фактора VII, может быть также получена путем синтеза с помощью известных стандартных методик, например, по способу с использованием фосфоамидита, описанному в работе Beaucage and Caruthers, Tetrahedron Letters 22 (1981), 1859-1869, или по способу, описанному Matthes et al., EMBO Journal 3 (1984), 801-805. Согласно методу, включающему использование фосфоамидита, синтезируют олигонуклеотиды, например, в автоматическом синтезаторе ДНК, далее проводят очистку, отжиг, лигирование и клонируют в подходящих векторах.
Кроме того, указанная конструкция нуклеиновой кислоты может представлять собой смесь синтетических и геномных нуклеиновых кислот, смесь синтетической нуклеиновой кислоты и кДНК или смесь геномной ДНК и кДНК, полученную путем лигирования фрагментов синтетических последовательностей, геномных последовательностей или последовательностей кДНК (где это приемлемо), фрагментов, соответствующих различным частям полной конструкции нуклеиновой кислоты, в соответствии со стандартными методиками.
Последовательности ДНК, используемые при получении химерных полипептидов фактора VII по настоящему изобретению, в типичном случае кодируют пре-про-полипептид на амино-конце фактора VII, так чтобы достигались соответствующий пост-трансляционный процессинг (например, гамма-карбоксилирование остатков глютаминовой кислоты) и секреция из клетки-хозяина. Указанный пре-про-полипептид может быть получен из фактора VII или другого плазматического белка, зависимого от витамина К, такого как фактор IX, фактор X, протромбин, белок С или белок S. Как известно специалистам в данной области, в аминокислотные последовательности химерных полипептидов могут быть введены дополнительные модификации фактора VII, таким образом, что такие модификации в итоге не будут в существенной мере ухудшать способность белка действовать в качестве коагулянта.
Векторы экспрессии, используемые для целей экспрессии вариантов фактора VIIa, будут включать промоторы, способные направлять транскрипцию клонированного гена или кДНК. Подходящие промоторы, которые могут использоваться в культивируемых клетках млекопитающих, включают вирусные промоторы и клеточные промоторы. Вирусные промоторы включают промотор SV40 (Subramani et al., Mol. Cell. Biol.1:854-864, 1981) и промотор CMV (Boshart et al., Cell 41:521-530, 1985). Особенно подходящим вирусным промотором является главный поздний промотор из аденовируса 2 (Kaufman and Sharp, Mol. Cell. Biol. 2:1304-1319, 1982). Другой особенно предпочтительный вирусный промотор представляет собой промотор фактора элонгации-1-альфа (CHEF1) из китайского хомяка. Клеточные промоторы включают промотор мышиного каппа-гена (Bergman et al., Proc. Natl. Acad. Sci. USA81:7041-7045, 1983) и VH промотор мыши (Loh et al., Cell 33:85-93, 1983). Особенно подходящим клеточным промотором является промотор мышиного металлотионеина-1 (Palmiter et.al., Science 222:809-814, 1983). Векторы экспрессии могут также включать набор сайтов сплайсинга РНК, расположенных по направлению считывания информации промотора и против направления считывания информации от сайта встраивания самой последовательности химерного фактора VII. Подходящие сайты сплайсинга РНК могут быть получены из генов аденовируса и/или иммуноглобулина. В векторах экспрессии также содержится сигнал полиаденилирования, расположенный по направлению считывания информации от сайта встраивания. Особенно подходящие сигналы полиаденилирования включают ранний или поздний сигнал полиаденилирования из SV40 (Kaufman and Sharp), сигнал полиаденилирования из 5 Elb области аденовируса, терминатор из гена гормона роста человека (DeNoto et al. Nucl. Acids Res. 9:3719-3730, 1981) или сигнал полиаденилирования из гена фактора VII человека или гена бычьего фактора VII. Векторы экспрессии могут также включать некодирующую лидерную последовательность вируса, такую как тройной лидер аденовируса 2, расположенный между промотором и сайтами сплайсинга РНК; а также энхансерные последовательности, такие как энхансер SV40.
Клонированные последовательности ДНК вводят в культивируемые клетки млекопитающих, например, путем трансфекции с использованием фосфата кальция (Wigler et al., Cell 14:725-732, 1978; Corsaro and Pearson, Somatic Cell Genetics 7:603-616, 1981; Graham and Van der Eb, Virology 52d:456-467, 1973) или посредством электропорации (Neumann et al., EMBO J. 1:841-845, 1982). Для идентификации и отбора клеток, которые экспрессируют экзогенную ДНК, ген, который придает селектируемый фенотип (селектируемый маркер) в основном вводят в клетки вмести с геном или кДНК, представляющими интерес. Подходящие селектируемые маркеры включают гены, которые придают резистентность к лекарственным средствам, таким как неомицин, гигромицин и метотрексат. Селектируемый маркер может представлять собой амплифицируемый селектируемый маркер. Подходящий амплифицируемый селектируемый маркер представляет собой последовательность дигидрофолятредуктазы (DHFR). Другим подходящим селектируемым маркером является гистидинол. Селектируемые маркеры описаны в обзоре Thilly (Mammalian Cell Technology. Butterworth Publishers, Stoneham, Mass., который включен в настоящее описание в качестве ссылки). Любой специалист со средним уровнем знаний в данной области может легко выбрать подходящие селектируемые маркеры.
Селектируемые маркеры могут вводиться в клетку на отдельной плазмиде одновременно с геном, который представляет интерес, или они могут вводиться на одной плазмиде. Если они вводятся на одной плазмиде, то селектируемый маркер и представляющий интерес ген могут находиться под контролем разных промоторов или одного и того же промотора, где указанный промотор организован таким образом, чтобы создавать дицистронную матрицу. Конструкции такого типа известны в данной области (например, Levinson and Simonsen, патент США No. 4713339). К смеси, которая вводится в клетки, может быть полезно вводить дополнительную ДНК, которая выполняет функцию, известную как «ДНК-носитель».
После того, как клетки захватывают ДНК, их растят в течение 1-2 дней, до начала экспрессии гена, представляющего интерес. Среда, которая используется для культивирования клеток, может представлять собой стандартную среду, подходящую для роста клеток-хозяев, такую как минимальная или комплексная среда, содержащая нужные добавочные компоненты. Подходящие среды доступны от коммерческих поставщиков или могут быть получены по известным рецептам (см, например, каталоги Американской Коллекции типовых культур (American Type Culture Collection)). Среды получают с использованием процедуры, известной в данной области (см., например, ссылки на бактерии и дрожжи; Bennett, J. W. and LaSure, L., editors, More Gene Manipulations in Fungi, Academic Press, CA, 1991). Ростовая среда в основном включает источник углерода, источник азота, незаменимые аминокислоты, необходимые сахара, витамины, соли, фосфолипиды, белки и факторы роста. Для целей продукции гамма-карбоксилированных химерных полипептидов фактора VII, используемая среда будет содержать витамин К, предпочтительно, в концентрации примерно от 0,1 нг/мл до примерно 20 мкг/мл. Далее проводится селекция по лекарственному препарату для отбора растущих клеток, которые стабильно экспрессируют селектируемый маркер. В случае клеток, которые были трансфицированы амплифицируемым селектируемым маркером, концентрация лекарственного средства может быть повышена, с тем чтобы можно было отобрать большее число копий клонированных последовательностей, что повышает уровень экспрессии. Клоны стабильно трансфицированных клеток далее скринировали для идентификации экспрессии желательного химерного полипептида фактора VII.
Подходящие линии клеток млекопитающих включают CHO (ATCC CCL 61), COS-1 (ATCC CRL 1650), клетки почки детеныша хомяка (BHK) и клетки 293 (ATCC CRL 1573; Graham et al., J. Gen.Virol. 36:59-72, 1977). Подходящие клеточные линии BHK представляют собой tk.sup.-tsl3 BHK клеточную линию (Waechter and Baserga, Proc. Natl. Acad. Sci. USA 79:1106-1110, 1982), которая далее будет называться как клетки BHK 570. Клеточная линия BHK 570 доступна от Американской Коллекции типовых культур (American Type Culture Collection, 12301 Parklawn Dr., Manassas, Va. 20852, с номером доступа ATCC CRL 10314. Клеточная линия tk-ts13 BHK также доступна от ATCC с номером доступа CRL 1632. Кроме того, может использоваться множество других клеточных линий, включающих клетки Rat Hep I (линия клеток гепатома крысы; ATCC CRL 1600), Rat Hep II (линия клеток гепатомы крысы; ATCC CRL 1548), TCMK (ATCC CCL 139), линию клеток легкого человека (ATCC HB 8065), NCTC 1469 (ATCC CCL 9.1) и клетки DUKX (Urlaub and Chasin, Proc. Natl. Acad. Sci. USA 77:4216-4220, 1980).
Химерные полипептиды фактора VII по настоящему изобретению могут быть конъюгированы для пролонгирования периода полувыведения полипептида in vivo. Такая конъюгация может снижать почечный клиренс и повышать количество фактора VII, присутствующего in vivo, что в свою очередь, снижает частоту введения химерных полипептидов фактора VII. Подходящие конъюгаты и способы получения конъюгированных полипептидов фактора VII описаны в патенте США No. 7442524, который включен в настоящее описание полностью в качестве ссылки.
Трансгенные животные
Для получения химерных полипептидов фактора VII по настоящему изобретению могут использоваться трансгенные животные в рамках соответствующей технологии. Предпочтительно, чтобы указанные белки образовывались в молочной железе организма самки млекопитающего-хозяина. Экспрессия в молочной железе и последующая секреция в молоко белка, представляющего интерес, позволяет преодолеть многие трудности, возникающие при выделении белков из других источников. Молоко легко собирается, доступно в больших количествах и биохимически хорошо охарактеризовано. Кроме того, основная часть молочных белков присутствуют в молоке в высоких концентрациях (в типичном случае, в количестве примерно от 1 до 15 г/л). Химерные полипептиды фактора VII по настоящему изобретению могут также продуцироваться в моче организма-хозяина животного, что имеет преимущество в сравнении с получением их в молоке, поскольку моча образуется и у самцов, и у самок животных, и, кроме того, в моче содержится меньше белков, чем в молоке, что важно с точки зрения выделения.
С коммерческой точки зрения, в случае использования молока в качестве источника химерных полипептидов фактора VII по настоящему изобретению, очевидна предпочтительность использования в качестве организма-хозяина животного, которое продуцирует большое количество молока. Хотя для этой цели могут использоваться мелкие животные, такие как мыши и крысы (и предпочтительно на соответствующей стадии), предпочтительно использовать домашних млекопитающих, включающих, без ограничения, свиней, коз, овец и крупный рогатый скот. Овцы особенно хорошо подходят, ввиду таких показателей, как фактор трансгенности в генезе данного вида, выход молока, стоимость и легкая доступность оборудования для сбора овечьего молока (см., например, PCT публикацию No. WO 88/00239, где дается сравнительное описание факторов, влияющих на выбор вида организма-хозяина). В основном, в качестве организма-хозяина, желательно выбирать породу животного, которая выводилась для использования в качестве производителя молока, такую как восточная фрисландская овца, или для выведения породы производителей молока на основе трансгенной линии. В любом случае, должны использоваться только здоровые животные.
Для достижения экспрессии в молочной железе, используют промотор транскрипции из гена молочного белка. Гены молочного белка включают гены, кодирующие казеины (см. патент США No. 5304489), бета-лактоглобулин, альфа-лактальбумин и кислый сывороточный белок. В этом случае, приемлем промотор бета-лактоглобулина (BLG). В случае овечьего гена бета-лактоглобулина, в основном используют область, отстоящую проксимально по меньшей мере на 406 п.н. от 5'-фланкирующей последовательности гена, хотя приемлемы более крупные части 5'-фланкирующей последовательности, вплоть до примерно 5 тысяч п.н., такие как сегмент ДНК размером примерно 4,25 тысяч п.н., охватывающий 5'-фланкирующий промотор и некодирующую часть гена бета-лактоглобулина (см., Whitelaw et al., Biochem. J. 286: 31-39 (1992)). Приемлемы также аналогичные фрагменты промотора ДНК из других видов.
Для достижения экспрессии в моче, используют промотор, специфичный для уротелия. Например, может использоваться промотор, который осуществляет экспрессию уроплакин-родственных генов (см., например, патент США No. 6001646, который включен в настоящее описание полностью в качестве ссылки).
Другие области гена бета-лактоглобулина также могут включаться в указанные конструкции, как и геномные участки экспрессируемого гена. В основном, считается, что конструкции без интронов, например, плохо экспрессируются по сравнению с конструкциями, которые содержат такие последовательности ДНК (см., Brinster et al., Proc. Natl. Acad. Sci. USA 85: 836-840 (1988); Palmiter et al., Proc. Natl. Acad. Sci. USA 88: 478-482 (1991); Whitelaw et al., Transgenic Res. 1: 3-13 (1991); PCT публикация No. WO 89/01343; и PCT публикация No. WO 91/02318, где каждая из работ включена в настоящее описание полностью в качестве ссылки). В этом отношении, предпочтительно в основном, где это возможно, использовать геномные последовательности, содержащие полностью или некоторые из нативных интронов гена, кодирующего белок или полипептид, представляющий интерес, так что предпочтительно также включение по меньшей мере некоторых интронов, например, из гена бета-лактоглобулина. Один такой участок представляет собой сегмент ДНК, который обеспечивает сплайсинг интрона и полиаденилирования РНК от 3'-некодинующего участка овечьего гена бета-лактоглобулина. В случае замещения природными 3'-некодирующими последовательностями гена, указанный сегмент овечьего бета-лактоглобулина может усиливать и стабилизировать уровни экспрессии белка или полипептида, представляющего интерес. В других вариантах осуществления настоящего изобретения, участок, окружающий инициирующий кодон ATG в химерной последовательности фактора VII, замещают соответствующими последовательностями из гена, специфичного для молочного белка. Такое замещение обеспечивает возможность предположительной тканеспецифичной инициации для усиления экспрессии. Полезно проводить замещение полных химерных пре-пропоследовательностей фактора VII и 5'-некодирующих последовательностей последовательностями, например, из BLG гена, хотя могут замещаться и более мелкие участки.
Для экспрессии химерных полипептидов фактора VIIa в трансгенных животных, сегмент ДНК, кодирующий химерный фактор VIIa, оперативно связывают с дополнительными сегментами ДНК, необходимыми для экспрессии, для целей получения единиц экспрессии. Такие дополнительные сегменты включают указанный выше промотор, а также последовательность, которая обеспечивает терминацию транскрипции и полиаденилирование мРНК. Единицы экспрессии будут также включать сегмент ДНК, кодирующий последовательность секреторного сигнала, оперативно связанную с сегментом, кодирующим химерный фактор VIIa. Последовательность секреторного сигнала может представлять собой нативную последовательность секреторного сигнала фактора VII или может представлять собой другой белок, такой как белок молока (см. von Heijne, Nucl. Acids Res. 14: 4683-4690 (1986); и Meade et al., патент США No. 4873316, которые включены в настоящее описание в качестве ссылки).
Конструирование единиц экспрессии для использования в трансгенных животных может в удобном варианте проводиться путем встраивания химерной последовательности фактора VII в плазмидный или фаговый вектор, содержащий дополнительные сегменты ДНК, хотя единица экспрессии может быть сконструирована по существу в рамках любой последовательности процедур лигирования. Особенно удобно использовать вектор, содержащий сегмент ДНК, кодирующий молочный белок, и замещать кодирующую последовательность для молочного белка химерным полипептидом фактора VII, что приводит к созданию гена слияния, который включает контрольные последовательности экспрессии из гена молочного белка. В любом случае, клонирование единиц экспрессии в плазмидах или других векторах облегчает амплификацию химерной последовательности фактора VII. Амплификация может проводиться, в удобном варианте, в бактериальных клетках-хозяевах (например, в E. coli), при этом, такие векторы в типичном случае включают ориджин репликации и селектируемый маркер, функционирующий в бактериальных клетках-хозяевах. Затем единицы экспрессии вводят в оплодотворенные яйца (включающие эмбрионы на ранней стадии) выбранного вида организма-хозяина. Введение гетерологичной ДНК может быть осуществлено любым из известных способов, включающих микроинъекцию (см., например, патент США No. 4873191), инфекцию ретровирусом (Jaenisch, Science 240: 1468-1474 (1988)) или сайт-направленную интеграцию с использованием эмбриональных стволовых клеток (ES) (см. обзор Bradley et al., Bio/Technology 10: 534-539 (1992)). Затем яйца имплантируют в яйцеводы или матку псевдобеременных самок и оставляют развиваться до рождения. Потомство, несущее введенную ДНК в зародышевой линии, может передавать ДНК своему потомству по нормальному менделевскому правилу, что приводит к развитию трансгенных животных. Основные процедуры получения трансгенных животных известны в данной области (см., например, Hogan et al., Manipulating the Mouse Embryo: A Laboratory Manual. Cold Spring Harbor Laboratory,1986; Simons et al., Bio/Technology 6: 179-183 (1988); Wall et al., Biol. Reprod. 32: 645-651 (1985); Buhler et al., Bio/Technology 8: 140-143 (1990); Ebert et al., Bio/Technology 9: 835-838 (1991); Krimpenfort et al., Bio/Technology 9: 844-847 (1991); Wall et al., J. Cell. Biochem. 49: 113-120 (1992); патент США No. 4873191; патент США No. 4873316; PCT публикация No. WO 88/00239, PCT публикация No. WO 90/05188, PCT публикация No. WO 92/11757; и публикация по патенту Великобритании No. GB 87/00458). Методы введения последовательностей чужеродных ДНК в млекопитающих и манипуляции с их зародышевыми клетками первоначально были разработаны для мышей (см., например, Gordon et al., Proc. Natl. Acad. Sci. USA 77: 7380-7384 (1980); Gordon and Ruddle, Science 214: 1244-1246 (1981); Palmiter and Brinster, Cell 41: 343-345 (1985); Brinster et al., Proc. Natl. Acad. Sci. USA 82: 4438-4442 (1985); and Hogan et al.). Эти методики были впоследствии адаптированы для использования на более крупных животных, включая домашних животных (см., например, PCT публикации NoNo. WO 88/00239, WO 90/05188 и WO 92/11757; и Simons et al., Bio/Technology 6: 179-183 (1988)). В основном, наиболее эффективный используемый в настоящее время способ создания трансгенных мышей или видов домашних животных включает инъекцию нескольких сотен линейных молекул ДНК, представляющих интерес, в одно из про-ядер оплодотворенного яйца, по стандартным методам. Может также проводиться инъекция ДНК в цитоплазму зиготы.
Может также использоваться продукция в трансгенных растениях. При этом, экспрессия может быть расширена или направлена на конкретный орган, такой как клубень (см, Hiatt, Nature 344:469-479 (1990); Edelbaum et al., J. Interferon Res. 12:449-453 (1992); Sijmons et al., Bio/Technology 8:217-221 (1990); и EP 0255378).
Продукция химерных полипептидов фактора VII по настоящему изобретению может быть также достигнута при трансляции in vitro, такой как, например, с использованием лизата ретикулоцитов кролика с использованием экстракта зародышей пшеницы и бесклеточной системы из E. coli. Указанная трансляция in vitro может быть объединенной или сопряженной. Объединенная in vitro трансляция основана на транскрипции с использованием полимеразы бактериофага с последующей трансляцией в лизате ретикулоцитов кролика или в лизате зародышей пшеницы. Сопряженная трансляция in vitro основана на бесклеточном лизате из E. coli.
Восстановление и активация
Химерные полипептиды фактора VII по настоящему изобретению выделяют из культуральной среды культивируемых клеток или из молока. Химерные полипептиды фактора VII по настоящему изобретению могут быть очищены с использованием множества процедур, известных в данной области, которые включают, без ограничения, хроматографию (например, ионообменную, аффинную, гидрофобную, хроматографию. хроматофокусирование и вытеснительную хроматографию), процедуру электрофореза (например, препаративное изоэлектрофокусирование (ИЭФ), процедуру, основанную на дифференциальной растворимости (например, осаждение сульфатом аммония) или экстракции (например, ProteinPurification. J.-C. Janson and Lars Ryden, editors, VCH Publishers, New York, 1989). Дополнительно, очистка может быть проведена с использованием стандартных химических способов очистки, таких как высокоэффективная жидкостная хроматография. Другие способы очистки, включающие осаждение цитратом бария, известные в данной области, также могут быть применимы для очистки новых химерных полипептидов фактора VII, приведенных в настоящем описании (см., например, Scopes, R, Protein Purification. Springer-Verlag, N.Y., 1982).
Для терапевтических целей предпочтительно, чтобы полипептиды химерного фактора VII по настоящему изобретению были по существу чистыми. Соответственно, в одном варианте осуществления настоящего изобретения, варианты фактора VII по настоящему изобретению очищали по меньшей мере до уровня 90-95% чистоты, предпочтительно до уровня по меньшей мере примерно 98% чистоты. Степень чистоты можно оценить, например, с помощью гель-электрофореза, секвенирования по амино-концевой кислоте и при использовании ВЭЖХ с обращением фазы.
Химерный полипептид фактора VII расщепляли по сайту его активации, с тем чтобы пробразовать его в двухцепочечную форму. Активация может быть осуществлена по процедурам, известным в данной области, таким как процедуры, описанные в работах Osterud, et al., Biochemistry 11:2853-2857 (1972); Thomas, U.S. Pat. No. 4456591; Hedner and Kisiel, J. Clin. Invest. 71:1836-1841 (1983); или Kisiel and Fujikawa, Behring Inst. Mitt. 73:29-42 (1983). Альтернативно, химерный полипептид фактора VII может быть активирован путем концентрирования химерного полипептида фактора VII и приведения его в контакт с положительно заряженной поверхностью или смолой, например, как описано в работе Bjoern et al. (Research Disclosure, 269 September 1986, pp. 564-565). Фактор VII может быть также активирован при пропускании его через колонку для ионообменной хроматографии, такую как Mono Q® (Pharmacia fine Chemicals) или т.п. Химерный полипептид фактора VII может быть активирован в растворе путем получения раствора, включающего по существу чистый препарат химерного полипептида фактора VII, с последующим добавлением к раствору аминосоединения, Ca2+ до конечной концентрации примерно 5 мМ - 50 мМ (например, такой как от примерно 10 мМ до примерно 30 мМ) и далее при корректировании конечного pH раствора до уровня примерно 7,2-8,6 (такого как от примерно 7,6 до примерно 8,2); с последующим инкубированием полученной активирующей смеси при температуре от примерно 2°C до примерно 25°C в течение периода времени, достаточного для превращения 90% химерного полипептида FVII в химерный полипептид FVIIa и, необязательно, с последующим выделением FVIIa из активирующей смеси. Полученный активированный химерный полипептид фактора VII может быть затем получен в виде композиции и использован для введения, как описано ниже.
Генная терапия
Химерные полипептиды фактора VII по настоящему изобретению могут быть также использованы в способах лечения расстройств, ассоциированных с кровотечением, в рамках генной терапии. В этом варианте осуществления настоящего изобретения, химерные полипептиды фактора VII по настоящему изобретению кодируются молекулами нуклеиновой кислоты, которые могут быть введены в клетки субъекта путем транспорта ex vivo или транспорта in vivo.
Любой из способов генной терапии, применяемый в данной области, может быть использован в настоящем изобретении. Ниже описаны репрезентативные способы. Общий обзор способов осуществления генной терапии приведен в работах Goldspiel et al., Clinical Pharmacy 1993, 12:488-505; Wu and Wu, Biotherapy 1991, 3:87-95; Tolstoshev, Ann. Rev. Pharmacol. Toxicol. 1993, 32:573-596; Mulligan, Science 1993, 260:926-932; и Morgan and Anderson, Ann. Rev. Biochem. 1993; 62 :191-217; May, TIBTECH 1993, 11:155-215. В настоящем изобретении могут использоваться широко известные в области применения технологии рекомбинантных ДНК способы, описанные в работах: Ausubel et al., (eds.), 1993, Current Protocols in Molecular Biology, John Wiley & Sons, NY; Kriegler, 1990, Gene Transfer and Expression, A Laboratory Manual Stockton Press, NY; and in Chapters 12 and 13, Dracopoli et al., (eds.), 1994, Current Protocols in Human Genetics, John Wiley & Sons, NY; Colosimo et al., Biotechniques 2000;29(2):314-8, 320-2, 324.
Полинуклеотиды, кодирующие химерные полипептиды фактора VII, могут быть встроены в соответствующий вектор клонирования. Векторы, подходящие для генной терапии, включают вирусы, такие как аденовирусы, адено-ассоциированный вирус (AAV), вирус осповакцины, вирус герпеса, бакуловирусы и ретровирусы, парвовирусы, лентивирусы, бактериофаги, космиды, плазмиды, грибные векторы и другие рекомбинантные носители, в типичном случае используемые в данной области, которые были описаны как подходящие для экспрессии во многих эукариотических и прокариотических организмах-хозяевах и которые могут использоваться для генной терапии, а также просто для экспрессии гена.
В одном варианте осуществления настоящего изобретения, указанный вектор представляет собой вирусный вектор. Вирусные векторы, в особенности аденовирусные векторы, могут быть включены в состав комплекса с катионным амфифильным соединением, таким как катионный липид, поли-L-лизин (PLL) и диэтиламиноэтилдекстран (DELAE-декстран), что повышает эффективность вирусной инфекции целевых клеток (см., например, РСТ публикацию по патенту No. PCT/US97/21496, зарегистрированному 20 ноября 1997 года, которая включена в настоящее описание в качестве ссылки). Подходящие вирусные векторы, которые могут использоваться в настоящем изобретении, включают векторы, полученные из вируса осповакцины, вирусы герпеса, AAV и ретровирусов. Обзор вирусов, применяемых при генной терапии, приведен, например, в работах Mah et al., Clin. Pharmacokinet. 2002; 41(12):901-11; Scott et al., Neuromuscul. Disord. 2002;12 Suppl LS23-9.
В одном варианте осуществления настоящего изобретения, используют вектор, в котором кодирующие последовательности и любые другие желательные последовательности фланкированы участками, которые способствуют гомологичной рекомбинации по желательному сайту в геноме, что обеспечивает экспрессию конструкции на основе молекулы нуклеиновой кислоты, которая была интегрирована в геном (Koller and Smithies, Proc. Natl. Acad. Sci. USA 1989, 86:8932-8935; Zijlstra ct al., Nature 1989, 342:435-438; U.S. Pat. No. 6244113 to Zarling et al.; и патент США No. 6200812 to Pati et al., где каждая из работ включена полностью в настоящее описание в качестве ссылки).
Доставка вектора в организм пациентов может быть либо прямой, и в этом случае пациент непосредственно подвергается воздействию вектора или доставляемого комплекса, или опосредованной, в этом случае клетки вначале трансформируют вектором in vitro и затем трансплантируют в организм пациента. Оба указанных подхода известны, соответственно, как генная терапия in vivo и генная терапия ex vivo.
В одном варианте осуществления настоящего изобретения, указанный вектор непосредственно вводится in vivo, после чего он поступает в клетки субъекта и опосредует экспрессию гена. Это может быть осуществлено с использованием множества различных методов, известных в данной области, которые обсуждались выше, например, путем конструирования его в качестве части соответствующего вектора экспрессии и введения таким образом, чтобы он стал внутриклеточным, например, путем инфекции с использованием дефектного или аттенуированного ретровирусного или другого вирусного вектора (см. патент США No. 4980286) или путем непосредственной инъекции оголенной ДНК, или путем бомбардировки микрочастицами (например, с использованием генной пушки Biolistic, Dupont); или путем нанесения покрывающего слоя, включающего липиды, или рецепторы клеточной поверхности, или трансфицирующие агенты, путем инкапсулирования в биополимеры (например, с использованием полисахарида поли-β-l-64-N-ацетилглюкозамина; см., патент США No. 5635493), путем инкапсулирования в липосомы, микрочастицы или микрокапсулы; путем введения в сочетании с пептидом или другим лигандом, который, по известным данным, поступает в ядро; или путем введения в сочетании с лигандом, который подвергается эндоцитозу, зависимому от рецептора (см., например, Wu and Wu, J. Biol. Chem. 1987, 62:4429-4432), и т.п. В другом варианте, может быть получен комплекс нуклеиновая кислота-лиганд, в котором лиганд включает фузогенный вирусный пептид для разрушения эндосом, что позволяет избежать лизосомной деградации нуклеиновой кислоты, или катионные 12-членные пептиды, например, полученные из сложного генного локуса Antennapedia, который может использоваться для транспортировки терапевтической ДНК в клетки (Mi et al., Mol. Therapy 2000, 2:339-47). В еще одном варианте, нуклеиновая кислота может быть направлена in vivo для специфического поглощения клетками и экспрессии, путем нацеливания на специфический рецептор (см., например, публикации PCT NoNo. WO 92/06180, WO 92/22635, WO 92/20316 и WO 93/14188). Дополнительно, может также использоваться методика, известная как магнитофекция, для доставки векторов в организм млекопитающих. Эта методика объединяет векторы с наночистицами-суперпарамагнетиками для доставки в условиях воздействия магнитных полей. Такое применение снижает время доставки и повышает эффективность действия вектора (Schcrer et. al., Gene Therapy 2002; 9:102-9).
В одном варианте осуществления настоящего изобретения, указанная нуклеиновая кислота может быть введена с использованием липидного носителя. Липидные носители могут быть ассоциированы с оголенными нуклеиновыми кислотами (например, с плазмидной ДНК) для облегчения прохождения через клеточные мембраны. Для этой цели могут использоваться катионные, анионные или нейтральные липиды. Однако, применяют катионные липиды, поскольку, как было показано, они лучше ассоциируются с ДНК, которая в основном имеет отрицательный заряд. Было также показано, что катионные липиды опосредуют внутриклеточную доставку плазмидной ДНК (Feigner and Ringold, Nature 1989; 337:387). Показано также, что внутривенная инъекция мышам комплексов катионный липид - плазмида приводит к экспрессии ДНК в легких (Brigham et al., Am. J. Med. Sci. 1989; 298:278). См. также Osaka et al., J. Pharm. Sci. 1996; 85(6):612-618; San et al., Human Gene Therapy 1993; 4:781-788; Senior et al., Biochemica et Biophysica Acta 1991; 1070:173-179); Kabanov and Kabanov, Bioconjugate Chem. 1995; 6:7-20; Liu et al., Pharmaceut. Res. 1996; 13; Remy et. al, Bioconjugate Chem. 1994; 5:647-654; Behr, J-P., Bioconjugate Chem. 1994; 5:382-389; Wyman et al, Biochem. 1997; 36:3008-3017; патент США No. 5939401 to Marshall et al; патент США No. 6331524 to Scheule et al.
Репрезентативные катионные липиды включают такие липиды, которые были описаны, например, в патенте США No. 5283185 и, в частности, в патенте США No. 5767099, описание которых включено в настоящую заявку в качестве ссылки. В одном варианте осуществления настоящего изобретения, катионный липид представляет собой N4-сперминхолестерилкарбамат (GL-67), описанный в патенте США No. 5767099. Дополнительные, применимые для этой цели липиды включают N4-спермидинхолестерилкарбамат (GL-53) и 1-(N4-спермин)-2,3-дилаурилглицеролкарбамат (GL-89).
Для введения in vivo вирусных векторов, может использоваться подходящее иммуносупрессорное лечение в сочетании с вирусным вектором, например, с аденовирусным вектором, для того, чтобы избежать иммунной дезактивации вирусного вектора и трансфицированных клеток. Например, могут вводиться иммуносупрессорные цитокины, такие как интерлейкин-12 (IL-12), интерферон-гамма (IFN-гамма) или анти-CD4 антитело, которые блокируют гуморальные или клеточные иммунные ответы на вирусные векторы. В этой связи, полезно использовать вирусный вектор, который разработан таким образом, чтобы экспрессировалось минимальное число антигенов.
Соматические клетки могут быть созданы ex vivo таким образом, чтобы они имели конструкцию, кодирующую химерный полипептид фактора VII по настоящему изобретению, с использованием любого из описанных способов, и затем имплантированы в организм индивидуума. Этот способ в основном описан в PCT публикации No. WO 93/09222 (Selden et al.) Кроме того, эта методика была использована в запатентованной технологии ImPACT для клеточной доставки, описанной в работе Payumo et al., Clin. Orthopaed. and Related Res. 2002; 403S: S228-S242. В такого рода системе для генной терапии, соматические клетки (например, фибробласты, гепатоциты или эндотелиальные клетки) отбирают от пациента, культивируют in vitro, трансфицируют вместе с одним или несколькими терапевтически значимыми генами, фиксируют основные характеристики и вновь вводят в организм пациента. Могут использоваться и первичные клетки (полученные от индивидуума или ткани и сконструированные определенным образом перед пассажами), и вторичные клетки (пассированные in vitro перед введением in vivo), так же как и иммортализированные клеточные линии, известные в данной области. Соматические клетки, применимые для их использования в способах по настоящему изобретению, включают, без ограничения, соматические клетки, такие как фибробласты, кератиноциты, эпителиальные клетки, эндотелиальные клетки, гепатоциты, сформированные элементы крови, клетки мышц, другие соматические клетки, которые могут культивироваться, и предшественники соматических клеток. В одном варианте, указанные клетки представляют собой гепатоциты.
Конструкции, которые включают полинуклеотид, кодирующий химерные полипептиды FVIIa по настоящему изобретению, и, необязательно, нуклеиновые кислоты, кодирующие селектируемый маркер, в сочетании с дополнительными последовательностями, необходимыми для экспрессии химерного FVIIa в первичных или вторичных клетках реципиента, используются для трансфекции первичных или вторичных клеток, в которых должен продуцироваться кодируемый продукт. Такие конструкции включают, без ограничения, векторы на основе инфекционных вирусов, такие как ретровирусные векторы, векторы на основе вируса герпеса, аденовирусные векторы, векторы на основе аденоассоциированного вируса, векторы на основе вируса эпидемического паротита и полиовирусные векторы.
Для опосредованной транспортировки с использованием клеточных типов эпидермиса, включающих кератиноциты, меланоциты и дендритные клетки, особенно хорошо подходит трансдермальная доставка (Pfutzner et al, Expert Opin. Investig. Drugs 2000; 9:2069-83).
В одном варианте осуществления настоящего изобретения, используемый для генной терапии вектор представляет собой одноцепочечный самокомплементарный аденоассоциированный вирус. Дополнительно, указанный самокомплементарный аденоассоциированный вирус может содержать малый трансиретиновый промотор-энхансер, интроны из малого вируса мышей и сигнал полиаденилирования из бычьего гормона роста. Указанный вектор может также содержать последовательность RKRRKR, описанную в работе Margaritis et al., которая вызывает секрецию фактора VII в его активной форме, то есть фактора VIIa (Margaritis et al., J. Clin. Invest., 2004; 113:1025-31).
Введение и фармацевтические композиции
Химерные полипептиды фактора VII по настоящему изобретению могут использоваться для контроля ассоциированным с кровотечением расстройств, которые вызваны рядом причин, таких как недостаточность фактора свертывания крови (например, гемофилия A и B или недостаточность коагулирующих факторов XI или VII) или наличие ингибиторов фактора свертывания крови, или они могут использоваться для контроля избыточного кровотечения у субъектов с нормально функционирующей системой свертывания крови (без недостаточности по фактору свертывания или наличия ингибиторов против любого из коагулирующих факторов). Кровотечения могут быть вызваны дефектной функцией тромбоцитов, тромбоцитопенией или болезнью Виллебранда. Такие кровотечения могут также наблюдаться у субъектов, имеющих повышенную фибринолитическую активность, которая была индуцирована различными стимулами.
У субъектов с обширным повреждением тканей, связанным с хирургией или травмой, гемостатический механизм может быть перегружен в связи с потребностью в срочном гемостазе, и у них может возникнуть кровотечение, несмотря на нормально функционирующий гемостатический механизм. Достижение удовлетворительного гемостаза также является проблемой при кровотечении в органах, таких как головной мозг, область внутреннего уха и глаза, и может также представлять собой проблему в случае диффузного кровотечения (геморрагический гастрит и профузное маточное кровотечение), когда трудно идентифицировать источник. Такая же проблемы может возникнуть в процессе отбора образцов биопсии из разных органов (печени, легкого, опухолевой ткани, желудочно-кишечного тракта), а также при лапароскопический хирургии. Во всех этих ситуациях, трудно обеспечить гемостаз хирургическими способами (швы, зажимы и т.п.). Острые профузные кровотечения могут возникать у субъектов при проведении антикоагулянтной терапии, у которых дефектный гемостаз вызывается проводимой терапией. Такие субъекты могут нуждаться в хирургическом вмешательстве, если антикоагулянтный эффект был достигнут слишком быстро. Другая ситуация, которая может вызвать проблемы в случае неудовлетворительного гомеостаза, может возникнуть, когда субъекту с нормально функционирующим механизмом гемостаза проводят антикоагулянтную терапию для предупреждения тромбоэмболической болезни. Такая терапия может включать введение гепарина, других форм протеогликанов, варфарина или других форм антагониста витамина К, а также аспирина и других ингибиторов агрегации тромбоцитов.
Для лечения, в сочетании с осторожным инвазивным вмешательством, химерные полипептиды фактора VII по настоящему изобретению в типичном случае могут вводиться в течение примерно 24 часов до проведения такого инвазивного вмешательства и до 7 дней или более, после него. Введение в качестве коагулянта может осуществляться множеством способов, приведенных в настоящем описании. Доза химерных полипептидов фактора VII варьирует от примерно 0,05 мг/день до примерно 500 мг/день, предпочтительно, от примерно 1 мг/день до примерно 200 мг/день, и более предпочтительно, от примерно 10 мг/день до примерно 175 мг/день для субъекта весом 70 кг, в качестве первичной и поддерживающей доз, в зависимости от веса субъекта и тяжести состояния. Любой специалист в данной области может легко определить оптимальную дозу для данного субъекта и данного состояния.
Фармацевтические композиции в основном предназначены для использования их с целью парентерального введения в режиме профилактического и/или терапевтического лечения. Предпочтительно, фармацевтические композиции вводят парентерально, то есть внутривенно, подкожно или внутримышечно, или могут вводиться непрерывной или пульсирующей инфузией. Альтернативно, фармацевтические композиции могут быть изготовлены для последующего их введения различными способами, которые включают, без ограничения, пероральный, внутривенный, интрацеребральный, интраназальный, трансдермальный, внутрибрюшинный, внутримышечный, внутрилегочный, вагинальный, ректальный, внутриглазной или любой другой подходящий способ.
Композиции для парентерального введения включают химерный полипептид фактора VII по настоящему изобретению в сочетании с фармацевтически приемлемым носителем, предпочтительно растворенным, причем предпочтительно в водном носителе. Может использоваться множество водных носителей, таких как вода, забуференная вода, 0,4% солевой раствор, 0,3% глицин и т.п. Химерные полипептиды фактора VII по настоящему изобретению могут быть также изготовлены в виде композиций, которые пролонгируют стабильность и срок хранения, например, в сочетании с метионином и сахарозой. Химерные полипептиды фактора VII по настоящему изобретению могут быть также изготовлены с получением липосомных препаратов для доставки или нацеливания на сайты повреждения. Липосомные препараты в основном описаны, например, в патентах США NoNo. 4837028, 4501728 и 4975282. Композиции могут быть простерилизованы с использованием стандартных, хорошо известных методов стерилизации. Полученные водные растворы могут быть упакованы для последующего использования или могут быть профильтрованы в асептических условиях и лиофилизированы, где лиофилизированный препарат объединяют со стерильным водным раствором перед введением. Указанные композиции могут содержать фармацевтически приемлемые вспомогательные вещества, способствующие поддержанию физиологических условий, таких как корректоры pH и забуферивающие агенты, средства для корректировки тонических свойств и т.п., например, такие как ацетат натрия, лактат натрия, хлорид калия, хлорид кальция и т.п. Указанные композиции могут также содержать консерванты, изотонирующие добавки, неионные поверхностно-активные вещества или детергенты, антиоксиданты и/или другие дополнительные добавки.
Концентрация химерного полипептида фактора VII в этой композиции может широко варьировать, то есть, от количества менее, чем примерно 0,5% вес.%, обычно от примерно 1 вес.% или по меньшей мере примерно 1 вес.%, до примерно 15-20 вес.% и предпочтительно изготавливаются с учетом объемов жидкости, вязкости и т.п., в соответствии с конкретным выбранным способом введения. Таким образом, типичная фармацевтическая композиция для внутривенной инфузии может быть изготовлена с содержанием 250 мл стерильного раствора Рингера и 10 мг химерного полипептида фактора VII. Фактические способы получения парентерально вводимых композиций хорошо известны или очевидны специалистам в данной области и описаны более подробно, например, в работе Remington's Pharmaceutical Sciences, 18th ed., Mack Publishing Company, Easton, Pa. (1990).
Композиции, содержащие химерный полипептид фактора VII по настоящему изобретению, могут вводиться в режиме профилактического и/или терапевтического лечения. При терапевтическом применении, композиции вводят субъекту, который уже имеет описанное выше заболевание, в количествах, достаточных для излечивания, облегчения или частичной остановки заболевания или его осложнений. Количество, которое является адекватным для достижения этой цели, определяется как «терапевтически эффективное количество». Как известно специалистам в данной области, количество, эффективное для достижения данной цели, будет зависеть от тяжести заболевания или повреждения, а также от веса и общего состояния субъекта. В основном, однако, эффективное количество варьирует от примерно 0,05 мг до примерно 500 мг химерного полипептида фактора VII в день для субъекта весом 70 кг, при этом, чаще всего используются дозировки от примерно 1,0 мг до примерно 200 мг химерного полипептид фактора VII в день.
Следует иметь виду, что средства по настоящему изобретению в основном могут применяться при серьезных заболеваниях или повреждениях, то есть в опасных для жизни или потенциально опасных для жизни ситуациях. В таких случаях, лечащий врач, ввиду минимального количества чужеродных веществ и в целом отсутствия иммуногенности человеческого фактора VII для людей, может, учитывая эти факторы, и даже это будет желательно, вводить значительный избыток композиций с таким химерным полипептидом фактора VII.
При профилактическом применении, композиции, содержащие химерный полипептид фактора VII по настоящему изобретению, вводят субъекту, который является чувствительным или иным образом имеет риск заболевания или повреждения, которое может повлиять на коагулирующую систему данного субъекта. Такое количество будет определяться как «профилактически эффективная доза». В случае профилактического применения, точное количество также зависит от состояния здоровья субъекта и его веса, и доза в этом случае варьирует в основном от примерно 0,05 мг до примерно 500 мг в день для субъекта весом 70 кг, и чаще всего используется доза от примерно 1,0 мг до примерно 200 мг в день для субъекта весом 70 кг.
Однократное или множественное введение композиции могут осуществляться с использованием доз и режима введения, выбранных лечащим врачом. Для субъектов, проходящих амбулаторное лечение, в случае которых требуется введение ежедневных поддерживающих доз, химерные полипептиды фактора VII могут вводиться путем непрерывной инфузии с использованием, например, переносной насосной системы.
Химерные полипептиды фактора VII по настоящему изобретению могут быть также изготовлены в виде пролонгированных композиций или композиций с длительным высвобождением. Способы изготовления пролонгированных композиций или композиций с длительным высвобождением известны в данной области и включают, без ограничения, использование полупроницаемых матриц из твердых гидрофобных частиц, содержащих полипептид.
Локальная доставка химерных полипептидов фактора VII по настоящему изобретению, например, в случае местного введения, может осуществляться, например, с помощью распыления, перфузии, катетеров с двойным баллоном, стента, встроенного в сосудистые трансплантаты или стенты, с использованием гидрогелей, наносимых на баллонные катетеры, и в рамках других хорошо известных способы. В любом случае, фармацевтические композиции должны обеспечивать количество варианта фактора VII, достаточное для эффективного лечения субъекта.
Приведенные ниже примеры были включены для иллюстрации настоящего изобретения. Некоторые аспекты приведенных ниже примеров описаны применительно к методикам и процедурам, которые авторы настоящего изобретения обнаружили или рассматривают как подходящие в практике осуществления настоящего изобретения. В свете настоящего изобретения и с учетом основного состояния уровня знаний в данной области, следует понимать, что приведенные примеры даны лишь с пояснительной целью и возможны различные изменения, модификации и перестановки без отступления от основных принципов настоящего изобретения.
ПРИМЕРЫ
Пример I - Конструирование и экспрессия химерных полипептидов фактора VII
кДНК человеческого фактора VII и человеческго фактора IX получали и вводили следующие уникальные сайты рестрикции для облегчения доменного обмена:
Сайт BstEII находится на 5'-конце домена EGF-1 (остатки 47-49 фактора VII; остатки 48-50 фактора IX);
Сайт SacI находится на соединении доменов EGF-1 и EGF-2 (остатки 82-83 фактора VII; остатки 83-84 фактора IX);
Сайт NotI находится на 3'-конце домена EGF-2 (остатки 135-137 фактора VII; остатки 132-134 фактора IX).
Различные химерные молекулы были сконструированы с использованием этих сайтов рестрикции для обмена доменами между фактором VII и фактором IX, и это проиллюстрировано на фиг. 1. Такие же или аналогичные сайты рестрикции могут быть использованы для обмена доменами между фактором VII и белком S.
Аминокислотная последовательность полученных химерных соединений представлена в виде SEQ ID NO: 1 (сигнал фактора IX, пропептид, домены GLA и EGF1, с EGF2 и каталитическим доменами фактора VII) и в виде SEQ ID NO: 2 (сигнальный и пропептидный домены фактора IX, домен GLA белка S, EGF1 домен фактора IX, с EGF2 и каталитическим доменами фактора VII). Выбранные варианты из этих рекомбинантных ДНК были субклонированы в векторе экспрессии pCMV5 для трансфекции клеток млекопитающих.
Полученные рекомбинантные конструкции затем были подвергнуты контрансфекции с pSV2neo и pCMVhGC в клеточную линию почки человека 293 (ATCC CRL 1573). Клон, экспрессирующий высокий уровень каждой конструкции, был отобран и подвергнут скринингу, как было ранее описано (Toomey et al., 1991, J. Biol. Chem., 266:19198-19202). Каждый скринированный клон размножали в круглодонных колбах площадью 900 см2 для крупномасштабной продукции, и рекомбинантные белки очищали по методу псевдоафинной хроматографии с использованием быстротекучей Q-Sepharose и элюции в градиенте кальция с последующим проведением элюции в градиенте NaCl.
Пример II - Получение тромбина с использованием полипептидов фактора VIIa
Образование тромбина при использовании фактора VIIa определяли в рамках in vitro модели гемофилии. Моноциты использовали в качестве источника тканевого фактора и объединяли с неактивированными тромбоцитами и синтетической плазмой, содержащей плазменные концентрации факторов V, VII, IX, VIII и XI и плазменные концентрации антитромбина и TFPI. Условия гемофилии были созданы путем удаления факторов VIII и IX.
Моноциты получали при отборе 4 мл крови от здоровых индивидуумов с последующим помещением их в пробирку с цитратом натрия. Кровь осторожно наслаивали на верхнюю часть разделяющей среды для лимфоцитов Accu-Prep™ в количестве 3 мл (Accurate Chemicals, NY, USA), в конической пробирке на 15 мл, и центрифугировали со скоростью 1500 об/мин в течение 30 минут. Далее отбирали мононуклеарный слой и добавляли его к равному объему Versene (Lineberger Tissue Culture) при температуре 4°C и центрифугировали со скоростью 800 об/мин в течение 10 минут. Полученный осадок ресуспендировали в 5 мл Versene при температуре 4°C и центрифугировали со скоростью 800 об/мин в течение 10 минут. Полученный осадок ресуспендировали в 4 мл макрофагальной среды SFM (Life Technologies, CA, USA) с добавкой 500 нг/мл липополисахарида (LPS) (Sigma-Aldrich, MO, USA), и клетки вносили в лунки 96-луночного микротитрационного планшета в количестве 200 мкл/лунку. Планшет инкубировали при температуре 37°C, с содержанием 5% CO2 в течение двух часов, затем промывали три раза макрофагальной средой SFM и инкубировали в течение ночи.
Тромбоциты получали при отборе 4 мл крови от здорового индивидуума и помещали в пробирку с цитратом натрия. Кровь осторожно наслаивали на верхнюю часть разделяющей среды для лимфоцитов Accu-Prep™ в количестве 5 мл (Accurate Chemicals, NY, USA), в конической пробирке на 15 мл, и центрифугировали со скоростью 1500 об/мин в течение 30 минут. Затем отбирали тромбоцитарный слой и добавляли его к равному объему смеси цитрат-глюкоза-солевой раствор с содержанием 10 мкг/мл простагландина E1 (PGE1) (Sigma-Aldrich, MO, USA) и центрифугировали со скоростью 800 об/мин в течение 10 минут. Полученный осадок отбрасывали и тромбоциты отделяли от плазменных белков гель-фильтрацией на сефарозе в бескальциевом буфере Tyrodes с добавкой 1 мг/мл овальбумина. Восстановленные тромбоциты хранили при температуре 37°C.
Синтетическую плазму для теста in vitro, в котором имитировалась гемофилия, получали с использованием ингибитора C-1-эстеразы фактора XI, в концентрации 5 мкг/мл, с добавлением через один час инкубации с предварительно полученными моноцитами 200 мкг/мл антитромбина, 0,07 мкг/мл TFPI, 100 мг/мл протромбина, 8 мкг/мл фактора X и 0,5 мкг/мл фактора VII. После ночной инкубации добавляли фактор V в концентрации 7 мкг/мл. Для подержания нормальных условий, фактор IX добавляли через 1 час инкубации в концентрации 4 мкг/мл и после ночной инкубации добавляли фактор VIII в дозе 1 Ед/мл.
260 мкл тромбоцитов добавляли к указанным выше растворам синтетической плазмы, и полученную суспензию добавляли к моноцитам для запуска реакции. В определенные временные интервалы отбирали 10 мкл образцов и добавляли к 90 мкл буфера для тестирования тромбина, содержащего 1 мМ ЭДТА и 0,5 мМ Gly-Pro-Arg-pNA (Centerchem Inc., CT, USA). Добавляли 100 мкл 50% уксусной кислоты для остановки расщепления синтетического субстрата и измеряли показатель ОП при длине волны 405 нм. Концентрацию тромбина определяли по следующей формуле:
[тромбин] = коэффициент разбавления × ((A405-фоновое значение)/(значение в точке стоп - значение в точке старт))/(коэффициент преобразования),
где коэффициент преобразования представляет собой 1 нМ тромбина, которому соответствует показатель ОП/мин 0,0117 при длине волны 405 нм.
Образование тромбина в нормальных условиях (при включении факторов VIII и IX) демонстрирует пик продукции тромбина с короткой лаг-фазой (фиг. 2). Повышение концентраций фактора VIIa дикого типа способствовало продукции тромбина в условиях гемофилии (фиг. 2).
Пример III - Образование тромбина с использованием химерных полипептидов фактора VIIa
Образование тромбина с использованием химерных белков, полученных в примере 1, определяли в рамках in vitro модели гемофилии, описанной в примере 2. При добавлении 10 нМ химерного фактора VIIa, содержащего домен EGF-2 и каталитический домен фактора VII и GLA и домен EGF-1 фактора IX, или химерного фактора VIIa, содержащего GLA, EGF-2 и каталитический домен фактора VII и домен EGF-1 фактора IX, отмечалась активность, аналогичная той, которую демонстрировал фактор VIIa дикого типа в концентрации 50 нМ (фиг. 3).
Если сравнивать полученный пик продукции тромбина, в сравнении с контролем, то видно, что ни фактор VIIa дикого типа, ни исследованные химерные полипептиды фактора VIIa не восстанавливали пик продукции тромбина до уровня контроля. Однако, использование 10 нМ химерного фактора VIIa, содержащего EGF-2 и каталитический домен фактора VII и GLA и домен EGF-1 фактора IX, или химерного фактора VIIa, содержащего GLA, EGF-2 и каталитический домен фактора VII и домен EGF-1 фактора IX, восстанавливало пик продукции тромбина до уровня аналогичного тому, который демонстрировал фактор VIIa дикого типа в концентрации 50 нМ (фиг. 4).
Пример IV - Анализ химерных полипептидов фактора VIIa на свертывающую активность
Свертывающую активность химерных полипептидов фактора VIIa оценивали in vivo в тесте на свертываемость крови, описанном в работе Buyue, Y., et al., 2008, Blood, 112:3234-3241. В общих чертах, процедура состоит в том, что фактор VII вводили мыши с гемофилией B (2 мг/кг NovoSeven®, 2 мг/кг химерного фактора VIIa, содержащего GLA и домен EGF-1 фактора IX и EGF-2 и каталитический домен фактора VIIa, или 1,4 мг/кг химерного фактора VIIa, содержащего домен GLA белка S, домен EGF-1 фактора IX и EGF-2 и каталитический домен фактора VIIa), через катетер, вставленный в вену конечности. Мышь дикого типа использовали в качестве контроля. Мышей анестезировали и разрезали подкожную вену другой конечности путем прокалывания иглой малого размера. Дистальную часть затем вырезали, вставляя кончик ножниц в вену и делали небольшой надрез. Далее, формировался сгусток и записывали время до остановки кровотечения. Затем сгусток удаляли иглой 30 размера и фиксировали время до остановки кровотечения. Этот процесс повторяли в течение 30 минут. Регистрировали количество сгустков как число повреждений. Зафиксированные временные показатели усредняли и записывали в качестве среднего времени. Эксперимент проводили на одной мыши в качестве мыши дикого типа с использованием химерного фактора VIIa, содержащего домен GLA белка S, домен EGF-1 фактора IX и EGF-2 и каталитический домен фактора VIIa. Для химерного фактора VIIa, содержащего GLA и домен EGF-1 фактора IX и EGF-2 и каталитический домен фактора VIIa, эксперимент проводили на двух мышах с гемофилией и получали те же результаты. В случае мыши с гемофилией B, которой не вводили никакого фактора VII, было зарегистрировано 0,25 повреждений, как видно из данных таблицы 1, и это указывает на то, что только у одной из 4 мышей не было кровотечения без первоначального повреждения.
Эксперимент повторяли, оценивая одну мышь с гемофилией B в варианте без обработки, одну мышь с гемофилией B, которой вводили 2 мг/кг NovoSeven®, одну мышь с гемофилией B, которой вводили 2 мг/кг химерного фактора VIIa, содержащего GLA и домен EGF-1 фактора IX и EGF-2 и каталитический домен фактора VIIa, и одну мышь дикого типа в варианте без обработки. Для мыши с гемофилией B, которой вводили 2 мг/кг химерного фактора VIIa, содержащего GLA и домен EGF-1 фактора IX и EGF-2 и каталитический домен фактора VIIa, отмечалось снижение среднего времени до остановки кровотечения, в сравнении с мышью с гемофилией B, которой вводили 2 мг/кг NovoSeven®, как и с мышью дикого типа (фиг. 5).
Пример V - Тромбогенность химерных полипепидов фактора VIIa
Тромбогенность химерных полипептидов фактора VIIa по настоящему изобретению может быть проанализирована на модели мышей, которые экспрессируют высокие уровни этих белков. В последнее время было показано, что 50% мышей, экспрессирующих более чем 2 мг/мл рекомбинантного фактора VIIa из мыши дикого типа, умирали от тромбоза в течение 16 месяцев. У мышей с гемофилией B, которые экспрессировали аналогичные уровни фактора VIIa, отмечался тот же уровень смертности, а у мышей штамма C57BL/6J, в случае обратного скрещивания, тромбозы происходили намного раньше (<4 месяцев) (Aljamali MN, et al J Clin Invest. 2008; 118:1825-1834).
Может быть получена мышь с гемофилией B (штамм C57BL/6J), которая экспрессирует 2-10 мкг или более химерных полипептидов, при этом, если экспрессия химерных полипептидов фактора Factor VII вызывает тромбоз, то эти мыши должны вырабатывать их рано (<4 месяцев).
Указанный штамм мышей может быть получен при использовании нового самокомплементарного аденоассоциированного вируса (AAV), разработанного Др. Paul Monahan из Центра генной терапии (Gene Therapy Center UNC). (Wu Z, et al. Mol Ther. 2008;16:280-289). Этот вектор содержит малый промотор-энхансер трансиретина (TTR промотор), интроны из малого вируса мышей, MVM и pA сигнал из бычьего гормона роста. Было показано, что инъекция 1×1011 векторных геномов, основанных на этом векторе, в портальную вену приводила к длительной экспрессии фактора IX на уровне 7 мкг на мл в течение 8 недель после инфекции (Wu Z, et al. Mol Ther. 2008;16:280-289).
кДНК для мышиного и человеческого фактора VIIa, а также для химерных полипептидов фактора VII по настоящему изобретению может быть использована для оценки тромбогенности. Margaritis et al. описали последовательность RKRRKR (SEQ ID NO: 21), которая вызывает секрецию фактора VII в активной форме, то есть в виде фактора VIIa. (Margaritis et al., J. Clin. Invest., 2004; 113:1025-31), которая также может быть использована.
Для оценки этой гипотезы получали полипептиды мышиного фактора VII дикого типа и химерные полипептиды мышиного фактора VII. Ранее было показано, что мышиный тканевой фактор не реагирует с человеческим фактором VII, и, в этой связи, мышиные полипептиды были использованы для демонстрации того, что тканевой фактор не оказывает нужного эффекта, в случае введения фактора VIIa при травме, но ассоциирован с тромбозом. Мышиные полипептиды, которые могут быть использованы для оценки этой гипотезы, включают мышиный фактор VII дикого типа (SEQ ID NO: 3), мышиный фактор VII дикого типа с последовательностью RKRRKR, которая вызывает секрецию полипептида в двуцепочечной активной форме (FVIIa) (SEQ ID NO: 4), сигнал для мышиного фактора IX, пропептид, домены GLA и EGF1, с EGF2 и каталитическим доменами фактора VII и последовательностью RKRRKR (SEQ ID NO: 5) и сигнал для мышиного фактора IX и пропептидные домены, домен GLA белка S, EGF1 домен фактора IX, с EGF2 и каталитическим доменами фактора VII и последовательностью RKRRKR (SEQ ID NO: 6). Химерные полипептиды мышиного фактора VII (SEQ ID NONO: 5 и 6) имели сниженную афинность для тканевого фактора (возможно, в 100 раз сниженную, в сравнении с молекулой FVII дикого типа), и этого количества должно быть достаточно для предупреждения нежелательных тромбозов.
Химерные полипептиды фактора VII могут быть также дополнительно модифицированы для дальнейшего снижения аффинности к тканевому фактору. Примером такой мутации являются изменение метионина по остатку 306 на аланин. Могут быть также проведены другие модификации химерных полипептидов фактора VII, которые приведут к повышению удельной активности фактора VIIa. Примеры таких модификаций включают указанные ниже эквиваленты остатков 158, 296 и 298 в человеческой последовательности: V158D, E296V и M298Q.
Полипептиды дикого типа и химерные полипептиды мышиного фактора VII, описанные выше, могут быть сконструированы с использованием мышиной кДНК для фактора VII в качестве матрицы и с использованием олигонуклеотидных праймеров для амплификации различных доменов, с последующим встраиванием конструкций в вектор pSC-TTR-mvm, и конструкция такого типа описана в работе Wu et al., Molecular Therapy, 2008; 16:280-289. Конкретно, фактор VII мыши дикого типа может быть амплифицирован на основе мышиной кДНК для фактор VII с использованием следующих праймеров:
5'-TGAGGATCCCCACCATGGTTCCACAGGCGCATGGGCT-3' (SEQ ID NO: 7) и 5'-TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC-3' (SEQ ID NO: 8). Амплифицированный фрагмент может быть затем расщеплен с использованием BamHI/SphI и встроен в вектор pSC-TTR-mvm между сайтами BgIII/SphI. Далее, последовательность поладенилирования BGH может быть встроена в полученный вектор в сайте SphI.
Конструкция мышиного фактора VII дикого типа с последовательностью RKRRKR может быть получена аналогичным способом. Конкретно, фактор VII мыши дикого типа с последовательностью RKRRKR может быть амплифицирован на основе мышиной кДНК для фактора VII с использованием следующих праймеров:
5'-TGAGGATCCCCACCATGGTTCCACAGGCGCATGGGCT-3' (SEQ ID NO: 7) и 5'-ACAATGCGTTTTCGCCGCTTACGGCGGCCTTGGCGGCTGCTGGAGT-3' (SEQ ID NO: 9), с получением первого фрагмента; и
5'-TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC-3' (SEQ ID NO: 8) и 5'-GCCGCCGTAAGCGGCGAAAACGCATTGTGGGAGGCAACGTGTGCCC-3' (SEQ ID NO: 10), с получением второго фрагмента. Затем проводят перекрывающуюся ПЦР с использованием двух фрагментов в качестве матрицы и указанных ниже праймеров:
5'-TGAGGATCCCCACCATGGTTCCACAGGCGCATGGGCT-3'(SEQ ID NO: 7) и 5'-TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC-3' (SEQ ID NO: 8). Далее, полученный амплифицированный фрагмент может быть расщеплен с использованием BamHI/SphI ивстроен в вектор pSC-TTR-mvm между сайтами Bglll/Sphl. Впоследствии, последовательность полиаденилирования BGH может быть встроена в полученный вектор в сайте SphI.
Конструкция, содержащая сигнал для мышиного фактора IX, пропептид, домены GLA и EGF1, с EGF2 и каталитическим доменами фактора VII и последовательностью RKRRKR (SEQ ID NO: 5), конструируется аналогичным образом. Конкретно, получают первый фрагмент с использованием мышиной кДНК для фактора IX в качестве матрицы и указанных ниже праймеров:
5'-AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC-3' (SEQ ID NO: 11) и
5'-GATCAGCTGCTCATTCTTGCTTTTTTCACAGTTCCTTCCTTCAAATC-3'(SEQ ID NO: 12). Второй фрагмент создают с использованием мышиного фактора VII в качестве матрицы и следующих праймеров:
5'-GATTTGAAGGAAGGAACTGTGAAAAAAGCA AGAATGAGC-AGCTGATC-3' (SEQ ID NO: 13) и 5'-ACAATGCGTTTTCGCCGCTTACGGCGGCCTTGGCGGCTGCTGGAGT-3'(SEQ ID NO: 9).
Третий фрагмент может быть создан с использованием мышиного фактора VII в качестве матрицы и следующих праймеров:
5'-TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC-3'(SEQ ID NO: 8) и
5'-GCCGCCGTAAGCGGCGAAAACGCATTGTGGGAGGCAACGTGTGCCC-3' (SEQ ED NO: 10). Затем может быть проведена перекрывающаяся ПЦР с использованием этих трех фрагментов и праймеров:
5'-AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC-3' (SEQ ID NO: 11) и 5'-TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC-3' (SEQ ID NO: 8). Полученный амплифицированный фрагмент затем расщепляют с использованием BamHI/SphI и встраивают в вектор pSC-TTR-mvm между сайтами BglII/SphI. Затем встраивают сигнал полиаденилирования BGH в полученный вектор в SphI сайте.
Аналогичный способ может быть использован для создания конструкции, содержащей сигнал мышиного для фактора IX и пропептидные домены, домены GLA белка S, EGF1 домен фактора IX с EGF2 и каталитическим доменами фактора VII и последовательностью RKRRKR (SEQ ID NO: 6). Конкретно, может быть получен первый фрагмент с использованием кДНК для фактора IX и следующи[ праймеров:
5'-AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC-3' (SEQ ID NO: 11) и
5'-GTTTCTTCGAACAAGGTATTTGCTCTCTTTGGACGGGTAAGAATTTTG-3' (SEQ ID NO: 14). Второй фрагмент может быть получен с использованием мышиного белка S в качестве матрицы и следующих праймеров:
5'-CAAAATTCTTACCCGTCCAAAGAGAGCAAATACCTTGTTCGAAGAAAC-3' (SEQ ID NO: 15) и
5'-GATCTCCATCAACATACTGCTTATAAAAATAATCCGTCTCGGGATT-3' (SEQ ID NO: 16). Третий фрагмент может быть получен с использованием вектора с сигналом для мышиного фактора VI, пропептида, доменов GLA и EGF1, с EGF2 и каталитическим доменами фактора VII и последовательностью RKRRKR, в качестве матрицы, и следующих праймеров:
5'-AATCCCGAGACGGATTATTTTTATAAGCAGTATGTTGATGGAGATC-3'(SEQ ID NO: 17) и 5'-TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC-3' (SEQ ID NO: 8). Затем проводят перекрывающуюся ПЦР с использованием этих трех фрагментов в качестве матрицы и следующих праймеров:
5'- AGGCCTGAAGATCTCCACCATGAAGCACCTGAACACCGTC-3' (SEQ ID NO: 11) и 5'-TTCCCCAGCATGCCTACAGTAGTGGGAGTCGGAAAAC-3' (SEQ ID NO: 8). Полученный амплифицированный фрагмент затем расщепляют с использованием BamHI/SphI и встраивают в вектор pSC-TTR-mvm между сайтами BglII/SphI. Впоследствии, может быть встроена последовательность полиаденилирования BGH в полученный вектор в SphI сайте.
Векторные конструкции с мышиным фактором VII дикого типа (SEQ ID NO: 3) и мышиным фактором VII дикого типа, содержащим последовательность RKRRKR (SEQ ID NO: 4), каждую, инъецировали двум самцам мышей с гемофилией B определенного возраста (5X1011 векторных геномов на мышь). Векторные конструкции, содержащие сигнал для мышиного фактора IX, пропептид, домены GLA и EGF1, с EGF2 и каталитическим доменами фактора VII и последовательностью RKRRKR (SEQ ID NO: 5) и сигнал для мышиного фактора IX и пропептидные домены, домена GLA белка S, домен EGF1 фактора IX, с EGF2 и каталитическим доменами фактора VII и последовательностью RKRRKR (SEQ ID NO: 6) были, каждый, инъецированы трем самцам мышей с гемофилией определенного возраста (5X1011 векторных геномов на мышь). Примерно через три недели после инъекции все мыши, которым инъецировали либо мышиный фактор VII дикого типа (SEQ ID NO: 3), либо мышиный фактор дикого типа с последовательностью VII RKRRKR (SEQ ID NO: 4), умерли. Мыши, которым инъецировали либо сигнал для фактора IX, пропептид, домены GLA и EGF1, с EGF2 и каталитическим доменами фактора IX мыши и последовательностью RKRRKR (SEQ ID NO: 5), либо мышиный сигнал для фактора IX и пропептидные домены, домен GLA белка S, домен EGF1 фактора IX, с EGF2 и каталитическим доменами фактора VII и последовательностью RKRRKR (SEQ ID NO: 6), были живы и не демонстрировали неблагоприятных симптомов примерно через пять недель после инъекции. Кровь отбирали для анализа у мышей один раз в неделю и через 6-8 недель после инъекции определяли уровень полипептида мышиного фактора VII. Кроме того, может быть опосредованно оценен уровень антигенов против полипептидов мышиного фактора VII, при измерении свертывающей активности in vitro в образцах крови, взятых у мышей после инъекции (Margaritas et al., Gene Therapy 2009; 113:3682-3689).
Приведенное выше описание является иллюстрацией настоящего изобретения, и его не следует трактовать в ограничительном смысле. Настоящее изобретение определяется прилагаемой формулой изобретения со всеми включенными в нее эквивалентами. Все публикации, патентные заявки, патенты, патентные публикации, последовательности, идентифицированные по номерам доступа в GenBank® и/или SNP, а также другие процитированные в тексте заявки материалы включены в него полностью в качестве ссылок, применительно к каждому предложению и/или абзацу, к которому эта ссылка относится.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЙ КОАГУЛИРУЮЩИЙ ФАКТОР VIIa С ПРОДЛЕННЫМ ВРЕМЕНЕМ ПОЛУЖИЗНИ | 2007 |
|
RU2466141C2 |
| ПОЛИПЕПТИД ФАКТОРА КОАГУЛЯЦИИ VII ЧЕЛОВЕКА, ЕГО ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2002 |
|
RU2325401C2 |
| МОДИФИЦИРОВАННЫЕ ВИТАМИН К-ЗАВИСИМЫЕ ПОЛИПЕПТИДЫ | 2005 |
|
RU2396347C2 |
| ПОЛИПЕПТИД ФАКТОРА VII СВЕРТЫВАНИЯ КРОВИ, ЕГО ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2001 |
|
RU2326126C2 |
| ПОЛИПЕПТИДЫ НА ОСНОВЕ МОДИФИЦИРОВАННОГО ФАКТОРА VII И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2571931C2 |
| ПРОИЗВОДНЫЕ ФАКТОРА VII СВЕРТЫВАНИЯ КРОВИ | 2002 |
|
RU2338752C2 |
| ВАРИАНТЫ ДОМЕНА GLA ФАКТОРА VII ИЛИ VIIa | 2004 |
|
RU2373282C2 |
| ПРОИЗВОДНЫЕ ФАКТОРОВ СВЕРТЫВАНИЯ КРОВИ VII И VIIa, КОНЪЮГАТЫ И КОМПЛЕКСЫ, СОДЕРЖАЩИЕ ИХ, И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2620072C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВИТАМИН К-ЗАВИСИМЫХ БЕЛКОВ | 2001 |
|
RU2321633C2 |
| GLA ДОМЕНЫ В КАЧЕСТВЕ ТЕРАПЕВТИЧЕСКИХ АГЕНТОВ | 2014 |
|
RU2705786C2 |





















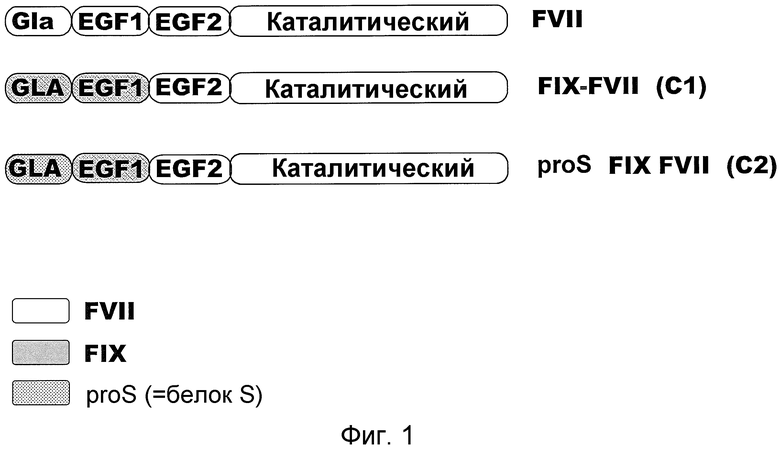
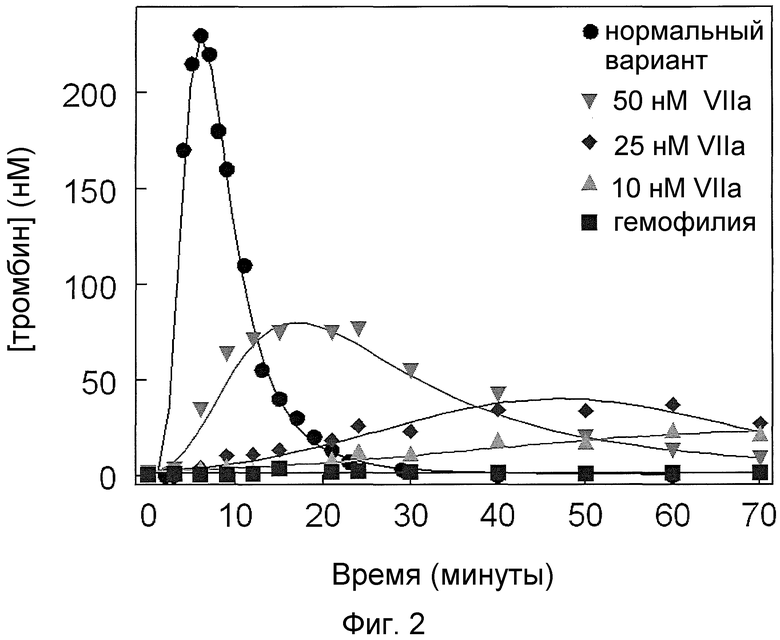
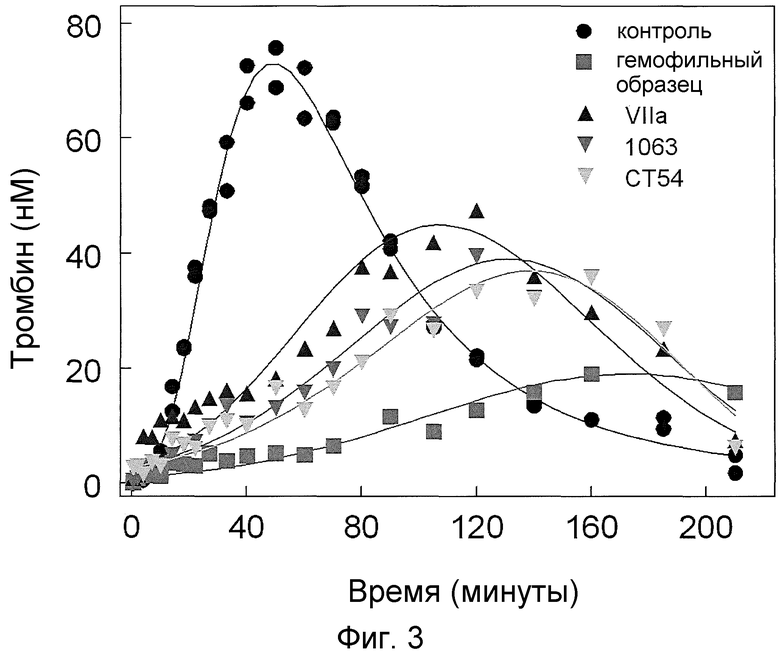
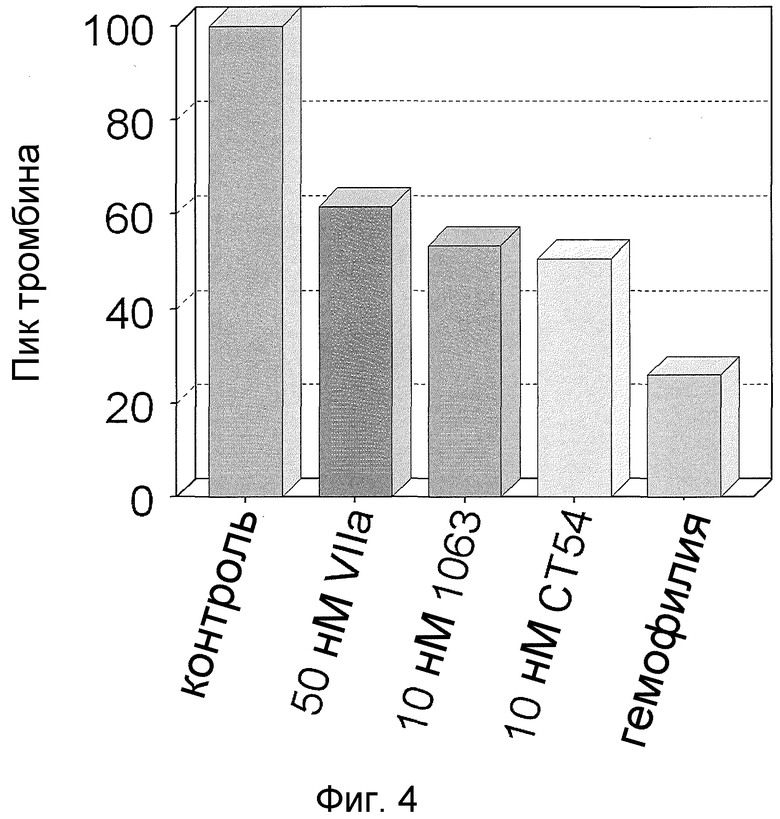
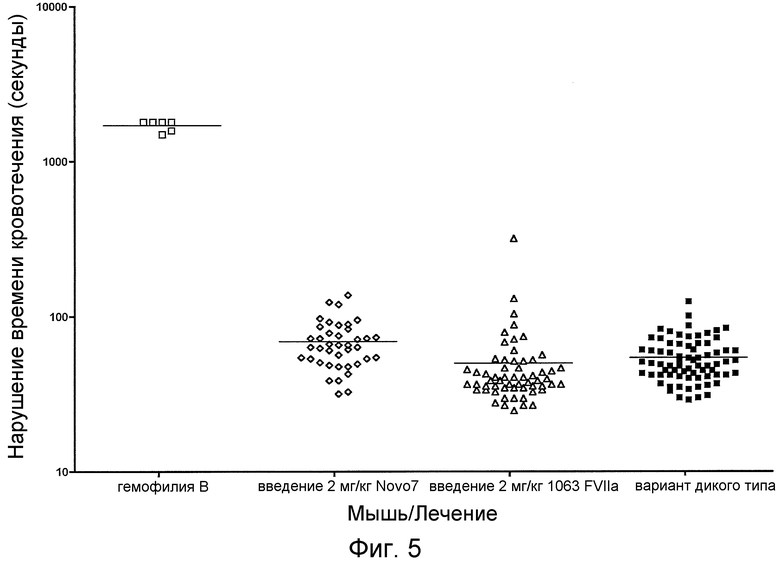


Группа изобретений относится к медицине, а именно к гематологии и может быть использована при лечении расстройств, ассоциированных с кровотечением. Способ по изобретению включает введение субъекту химерного полипептида фактора VIIa, где указанный химерный полипептид фактора VIIa включает домен EGF-2 и каталитический домен фактора VII; домен GLA, выбранный из группы, состоящей из домена GLA фактора VII, домена GLA фактора IX и домена GLA белка S; и домен EGF-1, выбранный из группы, состоящей из домена EGF-1 фактора IX и домена EGF-1 белка S. Химерный полипептид фактора VIIa по изобретению включает домен EGF-2 и каталитический домен фактора VII; домен GLA белка S;и домен EGF-1, выбранный из группы, состоящей из домена EGF-1 белка S и домена EGF-1 фактора IX. Молекула нуклеиновой кислоты по изобретению кодирует химерный полипептид фактора VIIa. Также изобретения касаются вектора, включающего молекулу нуклеиновой кислоты, клетки, включающей химерный полипептид фактора VIIa и клетки, включающей вектор. Использование изобретений позволяет снизить дозы вводимого белка и ослабить нежелательные побочные эффекты. 6 н.з. и 5 з.п. ф-лы, 7 ил., 5 пр.
1. Способ лечения расстройства, ассоциированного с кровотечением, у субъекта, имеющего расстройство кровотечения, путем введения химерного полипептида фактора VIIa, где указанный химерный полипептид фактора VIIa включает домен EGF-2 и каталитический домен фактора VII; домен GLA, выбранный из группы, состоящей из домена GLA фактора VII, домена GLA фактора IX и домена GLA белка S; и домен EGF-1, выбранный из группы, состоящей из домена EGF-1 фактора IX и домена EGF-1 белка S.
2. Способ по п.1, где расстройство, ассоциированное с кровотечением, выбирают из группы, состоящей из: недостаточности по фактору свертывания крови; дефектной функции тромбоцитов; тромбоцитопении; болезни Виллебранда; наличия ингибирования фактора свертывания крови; кровотечения, вызванного хирургическим вмешательством; и кровотечения, вызванного травмой.
3. Способ по п.2, где расстройство, ассоциированное с кровотечением, представляет собой недостаточность по фактору свертывания крови.
4. Способ по п.3, где недостаточность по фактору свертывания крови представляет собой гемофилию.
5. Способ по п.1, где лечение расстройства, ассоциированного с кровотечением, включает введение субъекту молекулы нуклеиновой кислоты, включающей нуклеотидную последовательность, кодирующую химерный полипептид фактора VIIa.
6. Способ по п.1, где домен GLA представляет собой домен GLA фактора IX, и указанный домен GLA фактора IX содержит замену лизина по остатку 5 аргинином.
7. Химерный полипептид фактора VIIa, где указанный химерный полипептид фактора VIIa включает:
домен EGF-2 и каталитический домен фактора VII;
домен GLA белка S;и
домен EGF-1, выбранный из группы, состоящей из домена EGF-1 белка S и домена EGF-1 фактора IX.
8. Молекула нуклеиновой кислоты, кодирующая химерный полипептид фактора VIIa по п.7.
9. Вектор, включающий молекулу нуклеиновой кислоты по п.8.
10. Клетка, включающая химерный полипептид фактора VIIa по п.7.
11. Клетка, включающая вектор по п.9.
| WO 2008127702 A2 (CATALYST BIOSCIENAES, INC.), 23.10.2008 |

