Область техники, к которой относится изобретение
Изобретение относится к молекулярной биологии, фармацевтике и медицинской химии, в частности к разработке, созданию и валидации новых биомишеней и тест-систем с целью поиска химических соединений-лидеров, которые могут являться потенциальными кандидатами в лекарства для лечения заболеваний человека.
Уровень техники
Одна из основных современных задач фармацевтики состоит в обнаружении высокоспецифичных терапевтических мишеней, позволяющих проводить эффективную разработку новых лекарственных веществ. Важнейшими среди таковых мишеней являются протеинкиназы - белки, осуществляющие передачу внутриклеточных сигналов в клетках. Серин-треониновые протеинкиназы (СТПК) - одни из наиболее универсальных регуляторов жизнеспособности про- и эукариотических клеток. Этим энзимам принадлежат ключевая роль в контроле таких процессов, как апоптоз, пролиферация и дифференцировка клеток, транспорт веществ из клетки и др [Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, Т.; Sudarsanam, S. The protein kinase complement of the human genome. //Science, 2002, 298, 1912-1934; Cohen, P. Protein kinases - the major drug targets of the twentyfirst century? //Nat. Rev. Drug Discov., 2002, 1, 309-315; Danilenko V.N., Osolodkin D.I., Lakatosh S.A., Preobrazhenskaya M.N., Shtil A.A. Bacterial Eukaryotic Type Serine-Threonine Protein Kinases: Tools for Targeted Anti-Infective Drug Design. / Current Topics in Medicinal Chemistry, 2011, 11, 1352-1369; Anizon F, Shtil A.A., Danilenko V.N., Moreau P. Fighting Tumor Cell Survival: Advances in the Design and Evaluation of Pim Inhibitors. // Curr Med Chem. 2010; 17 (34): 4114-33].
Нарушение функционирования протеинкиназ приводит ко многим заболеваниям человека. [D'Alessandris С, Lauro R, Presta I, Sesti G. C-reactive protein induces phosphorylation of insulin receptor substrate-1 on Ser307 and Ser612 in L6 myocytes, thereby impairing the insulin signalling pathway that promotes glucose transport. // Diabetologia. 2007 Apr; 50 (4): 840-9; Shirai H., Autieri M., Eguchi S. Small GTP-binding proteins and mitogen-activated protein kinases as promising therapeutic targets of vascular remodeling. //Curr Opin Nephrol Hypertens. 2007, Mar; 16 (2): P.111-115].
Киназа гликогенсинтаза - 3 (GSK3) Homo sapiens представляет собой серинтреониновую протеинкиназу, существующую в двух изоформах (GSK3α и GSK3β), каждая из которых кодируется отдельным геном [Coghlan MP, Culbert AA, Cross DA, Corcoran SL, Yates JW, Pearce NJ, Rausch OL, Murphy GJ, Carter PS, Roxbee Cox L, Mills D, Brown MJ, Haigh D, Ward RW, Smith DG, Murray KJ, Reith AD, Holder JC. Selective small molecule inhibitors of glycogen synthase kinase-3 modulate glycogen metabolism and gene transcription. // Chem Biol. 2000 Oct; 7 (10): 793-803; Kim L, Kimmel AR. GSK3, a master switch regulating cell-fate specification and tumorigenesis. //Curr Opin Genet Dev. 2000 Oct; 10 (5): 508-14].
Киназа гликогенсинтаза - 3β (GSK3β) играет ключевую роль в различных сигнальных путях, связанных с рецепторами тирозинкиназ и G-белками. [Doble B.W., Woodgett J.R. GSK-3: tricks of the trade for a multi-tasking kinase. // J Cell Sci. 2003 Apr 1; 116 (Pt 7): 1175-86]. Нарушения в регуляции этих сигнальных путей являются причинами развития заболеваний в организме человека, таких как диабет типа II [Kaidanovich О, Eldar-Finkelman H. The role of glycogen synthase kinase-3 in insulin resistance and type 2 diabetes. // Expert Opin Ther Targets. 2002 Oct; 6 (5): 555-61], болезнь Альцгеймера [Grimes C.A., Jope R.S. The multi-faceted roles of glycogen synthase kinase-3β in cellular signaling. // ProgNeurobiol 2001; 65: 391-426], нарушения ЦНС, в том числе маниакально-депрессивное расстройство и нейродегенеративные заболевания, хронические воспалительные заболевания [Hoeflich KP, Luo J, Rubie EA, Tsao MS, Jin O, Woodgett JR. Requirement for glycogen synthase kinase-3beta in cell survival and NF-kappaB activation. // Nature. 2000 Jul 6; 406 (6791): 86-90].
Лечение многих заболеваний (диабет; состояния, ассоциированные с диабетом; хронические нейродегенеративные заболевания, включая болезнь Альцгеймера, маниакально-депрессивный психоз; расстройства настроения; шизофрения; нейротравматические нарушения, такие как острый шок; выпадение волос и рак) также ассоциированны с необходимостью ингибирования GSK3β [Патент WO 00/38675, 2000 г., Патент WO 00/21927, 2000 г.]. В качестве ингибиторов GSK3β также могут быть использованы производные 2,4-диаминотиазола [Патент WO 01/56567, 2001 г.] и пептидные ингибиторы [Патент WO 01/49709, 2001 г.]. Для ингибирования циклинзависимыхкиназ, GSK3β и казеин-киназы 1 для лечения нейродегенеративных заболеваний, таких как болезнь Альцгеймера, диабет, воспалительные патологии и рак, используют гимениальдизин или его производные [Патент WO 01/41768, 2001 г.]. Для получения лекарственных средств также используются индирубиновые производные, ингибирующие GSK3β [Патент WO 01/37819,2001 г.].
По литературным данным, селективными ингибиторами протеинкиназы GSK3β являются: SB 216763 (3-(2,4-Dichlorophenyl)-4-(1-methyl-1H-indol-3-yl)-1H-pyrrole-2,5-dione), SB 415286 (3-(3-chloro-4-hydroxyphenylamino)-4-(4-nitrophenyl)-1H-pyrrole-2,5-dione), alsterpaullone [Coghlan MP, Culbert AA, Cross DA, Corcoran SL, Yates JW, Pearce NJ, Rausch OL, Murphy GJ, Carter PS, Roxbee Cox L, Mills D, Brown MJ, Haigh D, Ward RW, Smith DG, Murray KJ, Reith AD, Holder JC. Selective small molecule inhibitors of glycogen synthase kinase-3 modulate glycogen metabolism and gene transcription. //Chem Biol. 2000 Oct; 7 (10): 793-803; Braeuning A, Buchmann A. The glycogen synthase kinase inhibitor 3-(2,4-dichlorophenyl)-4-(1-methyl-1H-indol-3-yl)-1H-pyrrole-2,5-dione (SB216763) is a partial agonist of the aryl hydrocarbon receptor. //Drug Metab Dispos. 2009 Aug; 37 (8): 1576-80; Leost M, Schultz C, LinkA, WuYZ, BiematJ, MandelkowEM, BibbJA, SnyderGL, GreengardP, ZaharevitzDW. Paullones are potent inhibitors of glycogensynthase kinase-3 beta and cyclin-dependent kinase 5/p25. // EurJBiochem 2000, 267: 5983-5994].
Таким образом, целенаправленный отбор ингибиторов протеинкиназы GSK3β является перспективным направлением в лекарственной терапии вышеописанных заболеваний. Поэтому актуальной является задача создания тест-систем для эффективного скрининга ингибиторов СТПК.
В настоящее время отбор ингибиторов протеинкиназы GSK3β проводится in silico с помощью докинга с 3D-структурами с последующей проверкой их действия in vitro на очищенном ферменте [Wang F, Liu M, Liu J. In silico prediction of inhibitory effects of pyrazol-5-one and indazole derivatives on GSK3β kinase enzyme. //Journal of Molecular Structure, 1024, p.94-103, Sep 2012]. Однако не все ингибиторы, отобранные in silico и in vitro проявляют активность в клеточных системах.
Ранее разработана тест-система на основе штамма Streptomyces lividans TK24(66) для прескрининга ингибиторов СТПК [Danilenko V.N., Simonov A.Y., Lakatosh S.A., Kubbutat M.H., Totzke F., Schächtele C., Elizarov S.M., Bekker O.B., Printsevskaya S.S., Luzikov Y.N., Reznikova M.I., Shtil A.A., Preobrazhenskaya M.N. Search for inhibitors of bacterial and human protein kinases among derivatives of diazepines[1,4]annelated with maleimide and indole cycles. // J Med Chem. 2008 Dec 25; 51 (24): 7731-6]. Ключевым элементом разработанной тест-систем является фермент аминогликозид-3′-фосфотрансфераза тип VIII (AphVIII) штамма Streptomyces rimosus ATCC 10970, обеспечивающий устойчивость актинобактерий к аминогликозидным антибиотикам. Ранее нами был изолирован и секвенирован ген aphVIII и проведено клонирование его в штамм E.coli BL21(DE3) в составе экспрессионного вектора рЕТ16b под контроль транскрипционного и трансляционного сигналов бактериофага Т7 в рамке считывания с АТG экспрессионного участков. [Сизова И.А., Хегеманн П., Фурманн M., Даниленко В.Н. Аминогликозид-3'-фосфотрансфераза VIII из Streptomyces rimosus: сравнение с аминогликозид-3'-фосфотрансферазами из штаммов-продуцентов аминогликозидов и с эукариотическими протеинкиназами. // Молекулярная биология, 2002, т.36, №1, с.27-36]. Важной особенностью APHVIII S. rimosus является зависимость активности фермента от уровня его фосфорилирования серин-треониновыми протеинкиназами (СТПК). [Елизаров С.М., Сергиенко О.В., Сизова И.А., Даниленко В.Н. Зависимость активности аминогликозид-3'-фосфотрансферазы типа VIII от серин-треонин-протеинкиназ у Streptomyces rimosus 11 Молекулярная биология, 2005. т.39. №2. с.1-19]. Методами направленного мутагенеза и молекулярного моделирования установлено, что ключевым для модуляции активности APHVIII является Ca2+-зависимое фосфорилирование по остатку Ser-146 в активационной петле фермента. Сравнительный анализ канамицинкиназной активности нефосфорилированной и фосфорилированной форм исходного и мутантных вариантов APHVIII белка показал, что модификация Ser-146 в APHVIII приводит к 6-7-кратному увеличению канамицинкиназной активности APHVIII [Елизаров С.М., Алексеева М.Г., Новиков Ф.Н., Чилов Г.Г., Маслов Д.А., Даниленко В.Н. Идентификация сайтов фосфорилирования аминогликозидфосфотрансферазы VIII Streptomyces rimosus. // Биохимия, 2012, том 77, вып.11, с.1504-1512].
Известно, что штаммы Е.coli не содержат СТПК эукариотического типа. Это дает возможность сконструировать клеточную систему для тестирования ингибиторов протеинкиназы GSK3β, основанную на фосфорилировании AphVIII. Сконструирована модельная тест-система на основе штамма Escherichia coli APHVIII/Pk25. Разработанная тест-система может быть использована для первичного отбора (скрининга) ингибиторов протеинкиназы Pk25 штамма Streptomyces coelicolor и ее структурных. [Беккер, О.Б., Алексеева, М.Г., Осолодкин, Д.И., Палюлин, В.А., Елизаров, С.М. и Даниленко, В.Н. Новая тест-система для скрининга ингибиторов серин-треониновых протеинкиназ: Escherichia coli APHVIII/Pk25. ActaNaturae: 2010, с. 114-127].
Однако для отбора ингибиторов протеинкиназы GSK3β и других протеинкиназ, участвующих в патогенезе заболеваний человека до настоящего времени не сконструирована бактериальная тест-система эффективного скрининга.
Раскрытие изобретения
Задачей настоящего изобретения является создание новой тест-системы E.coli/gsk3β/aphVIII для скрининга ингибиторов GSK3β.
Принцип тест системы: фосфорилирование aphVIII no Ser146 посредством GSK3β приводит к устойчивости штамма Е.coli BL21(DE3)/gsk3β/aphVIII к канамицину; добавление ингибитора препятствует фосфорилированию aphVIII и снижает устойчивость к канамицину.
Промышленной применимостью новой тест-системы E.coli/gsk3β/aphVIII является селективный скрининг ингибиторов протеинкиназы GSK3β, в результате которого могут быть отобраны вещества, которые могут являться потенциальными лекарственными препаратами для лечения заболеваний человека (таких как диабет, хронические нейродегенеративные заболевания, включая болезнь Альцгеймера, маниакально-депрессивный психоз, нейротравматические нарушения, выпадение волос и рак).
1. Конструирование тест-системы E.coli/gsk3β/aphVIII.
Для конструирования тест-системы проведено клонирование генов каталитического домена протеинкиназы gsk3β и аминогликозидфосфотрансферезы aphVIII в E.coli, затем получены конструкции, содержащие гены аминогликозидфосфотрансферазы aphVIII и протеинкиназы gsk3β в одном экспрессионном векторе.
1.1. Клонирование генов каталитического домена протеинкиназы gsk3β и аминогликозидфосфотрансферезы aphVIII.
На первом этапе конструирования тест-системы проведено клонирование гена каталитического домена протеинкиназы gsk3β (915 п.н.) в Е.coli в составе экспрессионного вектора рЕТ32а под контроль транскрипционного и трансляционного сигналов бактериофага Т7 в рамке считывания с ATG экспрессионного участка.
Для повышения уровня экспрессии гена каталитического домена gsk3β для клонирования проведен синтез данного гена с адаптацией кодонов для Е.coli (http://www.kazusa.or.jp/codon/).
На последовательности 1 (см. перечень последовательностей) представлена нуклеотидная последовательность синтезированного гена. На последовательности 2 представлен сравнительный анализ синтезированной и исходной последовательностей генов gsk3β, отличия выделены подчеркиванием. Гомология нуклеотидных последовательностей составляет от 75,6%.
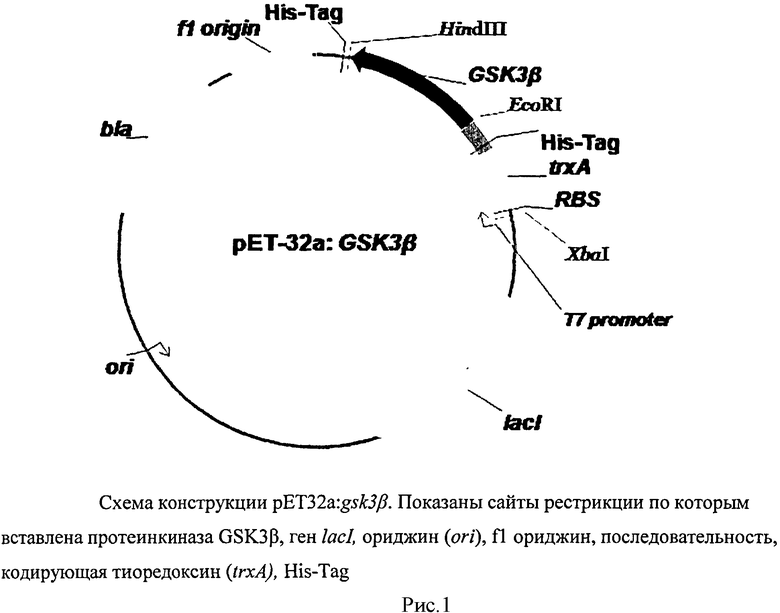
Для выбора сайтов клонирования проведен рестрикционный анализ синтезированного гена gsk3β по программе NEBcutter V2.0 (http://tools.neb.com/NEBcutter2/index.php). По результатам этого анализа клонирование в вектор рЕТ32а производилось по сайтам EcoRI-HindIII. Полученная плазмида обозначена pET32a:GSK3β.
Схема конструкции полученного вектора представлена на рисунке 1.
Ген aphVIII из штамма S.rimosus был ранее клонирован в экспрессионный вектор рЕТ16b по сайтам эндонуклеаз рестрикции XhoI и BamHI. Полученная плазмида обозначена pET16b:aphVIII. Последовательность гена aphVIII депонирована в банке генов NCBI/GenBank (код доступа AAG11411). (Сизова И.А., Хегеманн П., Фурманн М., Даниленко В.Н. Аминогликозид-3'-фосфотрансфераза VIII из Streptomyces rimosus: сравнение с аминогликозид-3'-фосфотрансферазами из штаммов-продуцентов аминогликозидов и с эукариотическими протеинкиназами. // Молекулярная биология, 2002, т.36, №1, с.27-36).
1.2. Получение целевых конструкций, содержащих гены аминогликозидфосфотрансферазы aphVIII и протеинкиназы gsk3β.
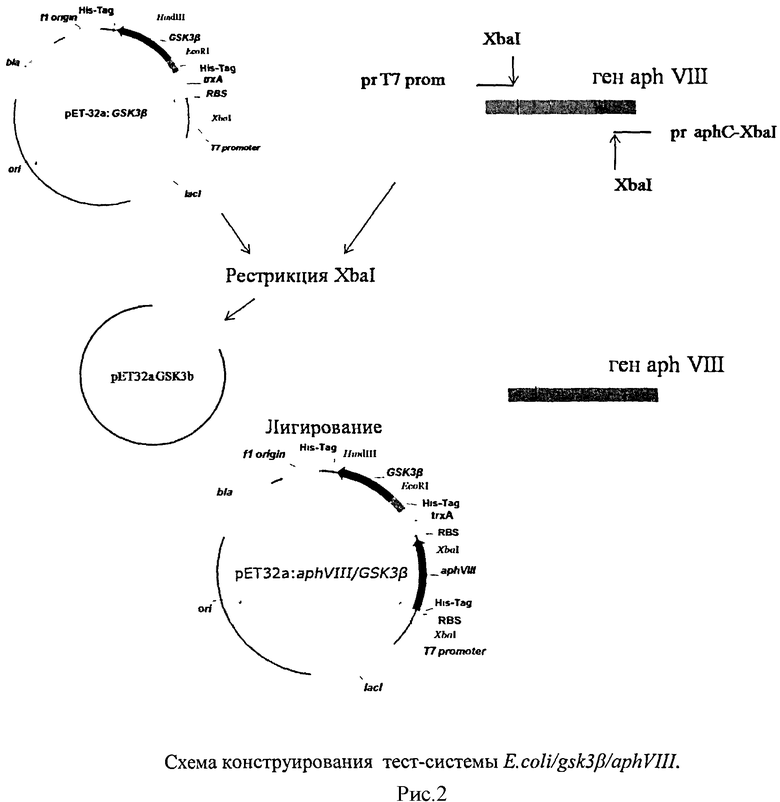
С целью получения целевых конструкций проведено клонирование гена aphVIII в плазмиду pET32a:GSK3β по сайту эндонуклеазы рестрикции XbaI. При таком клонировании гены aphVIII и gsk3β находятся под одним промотером, но имеют свои энхансеры, сайты связывания с рибосомой (RBS), инициирующие кодоны и терминаторы.
Полученная плазмида обозначена pET32a:aphVIII/GSK3β.
Схема получения целевой конструкции представлена на рисунке 2.
Для контроля уровня фосфорилирования AphVIII по уровню устойчивости к канамицину проведено клонирование гена aphVIII в плазмиду рЕТ32а по сайту эндонуклеазы рестрикции XbaI. Полученная плазмида обозначена рЕТ32а:aphVIII.
2. Модификация области S-146 аминогликозидфосфотрансферазы AphVIII для фосфорилирования киназой GSK3β.
Для повышения селективности тест-системы необходимо, чтобы протеинкиназа GSK3β фосфорилировала AphVIII, а для этого необходимо модифицировать потенциальный сайт фосфорилирования Ser-146, то есть приблизить его по своей структуре к каноническому сайту для GSK3β - S-X-X-X-S/T-P. (Dajani R, Fraser E, Roe SM, Young N, Good V, Dale TC, Pearl LH. Crystal structure of glycogen synthase kinase 3 beta: structural basis for phosphate-primed substrate speciflcity and autoinhibition // Cell. 2001. Jun 15; 105 (6): 721-32).
Получено 2 модификации сайта фосфорилирования Ser-146, предусматривающие аминокислотные замены, выделенные подчеркиванием.
AphVIII-1: AVAEGS146VDLED→ASAEGS146VDLSD;
AphVIII-2: AVAEGS146VDLED→ASPEGS146VDLSD.
Для получения мутантного варианта AphVIII-1 проведены аминокислотные замены: V142→S142, E150→S150. Для получения мутантного варианта AphVIII-2 проведены аминокислотные замены: V142→S142, А143→Р143 и Е150→S150.
С этой целью проведен сайт-направленный мутагенез гена aphVIII по методу точковых мутаций. Для проведения мутагенеза сконструированы олигонуклеотиды, предусматривающие необходимые замены: для получения варианта AphVIII-1 - GSK1(+) и GSK1(-); для получения варианта AphVIII-2 - GSK2(+) и GSK2(-) (таблица 1).
Мутантные гены aphVIII-1 и aphVIII-2 клонировали в плазмиду рЕТ16b по сайтам эндонуклеаз рестрикции XhoI и BamHI, а затем в плазмиды рЕТ32а и рЕТ32a:GSK3β по сайту XbaI. Полученные плазмиды обозначены рЕТ32а:aphVIII-1, pET32a:aphVIII-2, pET32a:aphVIII-1/GSK3β, pET32a:aphVIII-2/GSK3β соответственно.
Таким образом:
1) Сконструированная бактериальная тест-система E.coli/gsk3β/aphVIII-1 включает каталитический домен протеинкиназы GSK3β и субстрат для фосфорилирования - белок AphVIII, гены которых клонированы в одном экспрессионном векторе рЕТ32а.
2) Селективность разработанной тест-системы обусловлена адаптацией потенциального сайта фосфорилирования Ser-146 AphVIII для протеинкиназы GSK3β.
3) Назначением разработанной тест-системы E.coli/gsk3β/aphVIII-1 является селективный скрининг ингибиторов протеинкиназы GSK3β - потенциальных лекарственных препаратов для лечения заболеваний человека.
4) Подобные бактериальные тест-системы могут быть сконструированы для скининга ингибиторов других СТПК, участвующих в патогенезе заболеваний человека.
Осуществление изобретения
Синтез гена каталитического домена gsk3β с адаптацией кодонов для Е.coli проводят в компании «Евроген» (http://www.evrogen.ru/about/contact-us.shtml).
Для клонирования гена каталитического домена протеинкиназы gsk3β 5 мкг плазмидной ДНК рЕТ32а инкубируют с 50 ед. FastDigest®EcoRI и 50 ед. FastDigest®HindIII ферментов в присутствии 10Х FastDigest буфера фирмы «Fermentas» (Литва) в течение 1 ч при 37°C. Параллельно 3 мкг синтезированного гена каталитического домена gsk3β инкубируют с 30 ед. FastDigest®EcoRI и 30 ед. FastDigest®HindIII ферментов в тех же условиях. Затем осаждают ДНК добавлением 1/10 объема KAс с 2,5 объема этанола, растворяют в воде и проводят электрофорез в 1% агарозном геле для определения концентрации полученных фрагментов. Полученные фрагменты обрабатывают ДНК-лигазой фага Т4 (30 ед./мкл) фирмы «Fermentas» (Литва) из расчета 1 мкл фермента на 0,5 мкг ДНК в присутствии 10Х лигазного буфера. Инкубацию проводят в течение 18 часов при 12°C. Полученной лигазной смесью трансформируют компетентные клетки штамма E.coli DH5a, полученные хлоркальциевым методом. Трансформированные клетки подращивают в течение 1,5 часа и высевают на агаризованную среду, содержащую L-бульон с ампициллином (150 мкг/мл).
Скрининг колоний, выросших через 18 ч при 37°C, проводят методом ПЦР-скрининга с использованием набора «Амплификация» фирмы «Dialat Ltd» (Россия) на приборе «Терцик» («ДНК-технология»). Для этого выросшие колонии скалывают и ресуспендируют в 15 мкл стерильной воды. Готовят смесь для ПЦР (на 100 мкл): 10 мкл 10×ПЦР буфера; 10 мкл смеси 2,5 mM ΣdNTPs; 5 мкл DMSO; 4 мкл 50 mМ MgCl2 олигонуклеотиды на вектор рЕТ32а - S.Tag и T7term в концентрации 20 пмоль; 0,8 мкл фермента Taq-полимеразы. Смесь аликвотят по 23 мкл в микропробирки для ПЦР и добавляют по 3 мкл полученной суспензии клеток. Параметры реакции ПЦР: лизис клеток и денатурация геномной ДНК при 95°C в течение 5 мин; затем 30 циклов амплификации: 94°C в течение 1 мин (денатурация), при 52°C в течение 1 мин (отжиг олигонуклеотидов), 72°C в течение 2 мин (достройка (элонгация) цепи); финальная элонгация фрагментов при 72°C в течение 10 мин. Результаты амплификации учитывают путем анализа исследуемых образцов с помощью электрофореза в 1% агарозном геле. Размер амплифицированных фрагментов определяют с использованием маркера λ DNA/EcoRI+HindIII («Fermentas»). Положительными являются клоны, дающие при ПЦР фрагмент длиной 1416 п.н. (длина клонируемого гена+длина линкера плазмиды рЕТ32а, содержащего ген тиоредоксина). Из отобранных клонов выделяют плазмидные ДНК, в которых проводят рестрикционный анализ наличия вставки и ресеквенируют синтезированный ген каталитического домена gsk3β. В результате проведенного клонирования получена плазмида pET32a:GSK3β.
Для проведения мутагенеза на первом этапе проводят амплификации с плазмиды рЕТ16b:aphVIII: в отдельных пробирках амплифицируют левую (5'-концевую) часть гена, с использованием пар праймеров AphN+GSK1(-) или GSK2(-), и правую (3'-концевую) часть гена, с использованием пар праймеров GSK1(+) или GSK2(+)+AphC, часть гена. В результате образуется два фрагмента ДНК, перекрывающихся на участках, которые содержат включенные праймеры, предусматривающие необходимые нуклеотидные замены и полностью комплементарные друг другу. На второй стадии ПЦР полученные фрагменты смешивают в эквимолярном соотношении и после цикла тепловой денатурации и ренатурации используют в качестве матрицы в другой ПЦР с внешними праймерами AphN и AphC.
Полученные в результате амплификации фрагменты ДНК препаративно выделяют из агарозного геля, секвенируют для подтверждения наличия необходимых замен и клонируют в плазмиду рЕТ16b по сайтам эндонуклеаз рестрикции XhoI и BamHI. Скрининг рекомбинантных клонов осуществляют при помощи ПЦР в описанных выше условиях.
Для получения целевых конструкций проводят клонирование гена aphVIII в плазмиду pET32a:GSK3β. Фрагмент, содержащий ген aphVIII, амплифицируют с плазмидных ДНК pET16b/aphVIII с использованием олигонуклеотидов Т7prom и AphC-XbaI (гомологичен C-концевой области гена и содержит сайт для узнавания рестриктазой XbaI). Все олигонуклеотиды, используемые для конструирования тест-системы, приведены в таблице 1. Состав смеси для ПЦР и параметры реакции приведены выше. Затем 5 мкг плазмидной ДНК pET32a:GSK3β инкубируют с 50 ед. FastDigest®XbaI в присутствии 10Х FastDigest буфера фирмы «Fermentas» (Литва) в течение 1 ч при 37°C. Параллельно 3 мкг амплифицированного фрагмента aphVIII инкубируют с 30 ед. FastDigest®XbaI в тех же условиях. Осаждают ДНК добавлением 1/10 объема KAс с 2,5 объема этанола и растворяют в воде. Затем 1 мкг рестрицированной плазмидной ДНК обрабатывают антарктической фосфатазой (NEB) в присутствии 10Х NEB-буффера в течение 10 мин при 37°C, инактивируют фермент в течение 10 мин при 65°C и обрабатывают ДНК-лигазой фага Т4 вместе с рестрицированным фрагментом, содержащим ген aphVIII. Трансформированные клетки E.coli DH5a высевают на агаризованную среду, содержащую L-бульон с ампициллином. Скрининг колоний проводят методом ПЦР-скрининга с использованием олигонуклеотидов Т7prom и AphC-XbaI, что позволяет сразу отбирать клоны, содержащие ген aphVIII в прямой ориентации.
Для изучения экспрессии клонируемых генов полученными плазмидами трансформируют штамм Е.coli BL21(DE3), содержащий индуцибельный ген РНК-полимеразы бактериофага Т7. Клетки E.coli BL2I(DE3), содержащие сконструированные плазмиды, выращивают в колбах в жидкой среде (L-бульон), содержащей ампициллин, в аэрируемых условиях 250 об/мин, при 37°C до оптической плотности 0,6 (~2 часа), затем индуцируют экспрессию добавлением ИПТГ до финальной концентрации 1,0 mM/мл среды. Далее проводят культивирование при 28°C в течение 5 часов, после чего отбирают биомассу и ресуспендируют в буфере следующего состава: 62,5 mМ Трис-HCl pH 6,8; 5% глицерин; 2% меркаптоэтанол; 0,1% SDS; бромфеноловый синий. Затем клетки разрушают кипячением в течение 10 мин и анализируют фракцию белков с помощью электрофореза в полиакриламидном геле.
Для определения активности ингибиторов используют метод бумажных дисков. Методика заключается в определении величины зоны подавления роста штамма Е.coli BL21(DE3)/gsk3β/aphVIII, засеянного газоном на агаризованной среде, вокруг бумажных дисков, содержащих антибиотик или антибиотик в сочетании с ингибиторами протеинкиназы GSK3β. Тестирование исследуемых веществ осуществляют в субингибирующих концентрациях. Концентрацию канамицина подбирют экспериментально.
Бактерии, выращенные на агаризованной среде LB с ампициллином, засевают в жидкую среду LB и выращивают в термостатируемом в шейкере в течение 18 часов при 37°C при 250 об/мин. Затем бактериальную суспензию смешивают с расплавленной агаризованной средой М9, содержащей ампициллин (150 мкг/мл) и индуктор IPTG (50 мкг/мл), в соотношении 1:1 (v/v) и заливают полученной смесью чашки Петри с агаризованной средой М9 с ампициллином и IPTG. После застывания на поверхность агара накладывают бумажные диски, содержащие антибиотик канамицин или канамицин и ингибитор протеинкиназ. Культуру инкубируют в течение 16 часов при +37°C.
Примеры валидации и использования тест-системы E.coli/gsk3β/aphVIII по настоящему изобретению.
Пример 1
Исследование белков AphVIII-1 и AphVIII-1, фосфорилированного GSK3β, методом изоэлектрического фокусирования
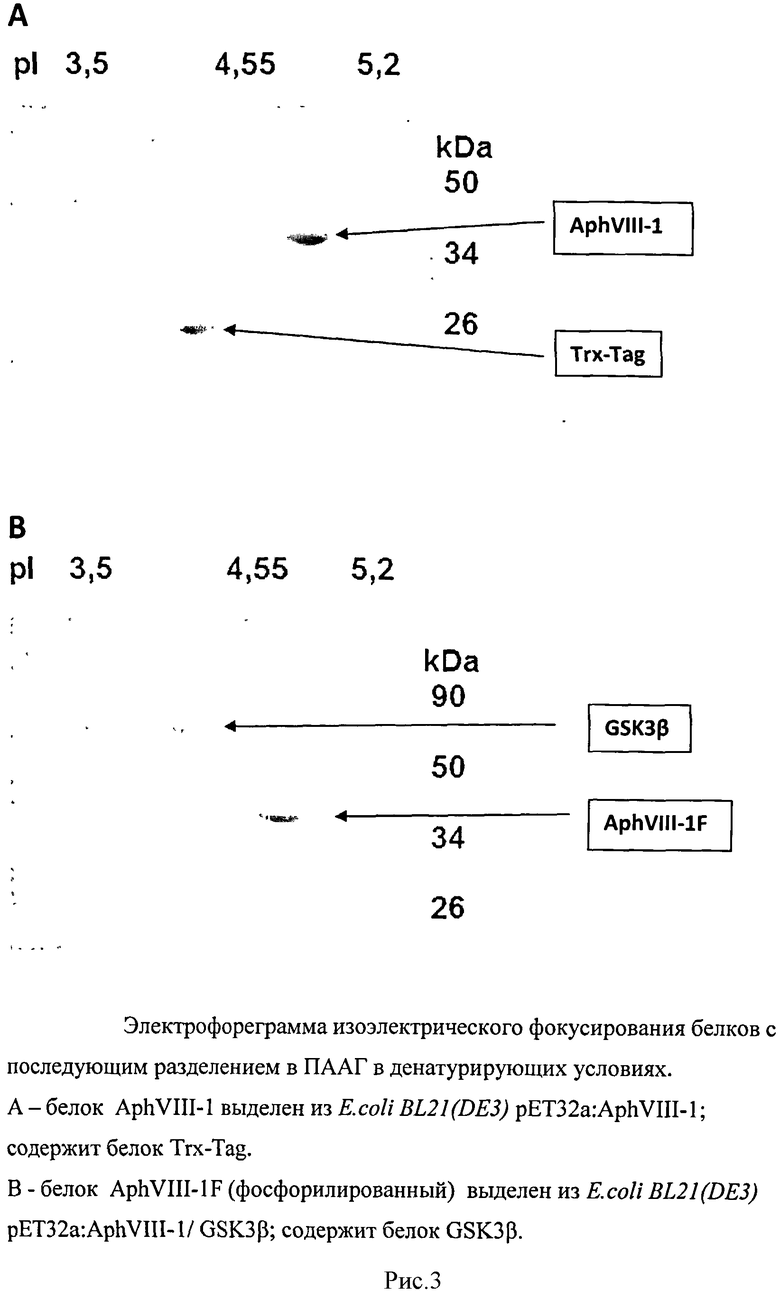
Для проверки пост-трансляционной модификации, а именно селективного фосфорилирования белка AphVIII протеинкиназой GSK3β в конструкции E.coli/gsk3β/aphVIII-1 на первом этапе выделяли белки AphVIII-1 и AphVIII-1/GSK3β с помощью набора Ni-NTA Spin Kit Handbook («Qiagen») в денатурирующих условиях. Затем проводили двумерный электрофорез выделенных белков в полиакриламидном геле.
Исследуемые белки вносили в полиакриламидный гель в концентрации 8 мкг/мл и проводили электрофорез в двух направлениях. Результаты электрофоретического разделения исследуемых белков представлены на рисунке 3.
Для дальнейших исследований белков AphVIII-1 и AphVIII-1/GSK3β фрагменты гелей, содержащие исследуемые белки, были вырезаны и переданы в НИИ Биомедицинской Химии им. В.Н. Ореховича РАМН для проведения масс-спектрометрического анализа. Масс-спектрометрический анализ показал, что фрагмент геля, содержащего белок AphVIII-1/GSK3β является посттрансляционной модификацией AphVIII-1, связанной с фосфорилированием его протеинкиназой GSK3β.
Пример 2
Выбор и валидация оптимальной тест-системы E.coli/gsk3β/aphVIII
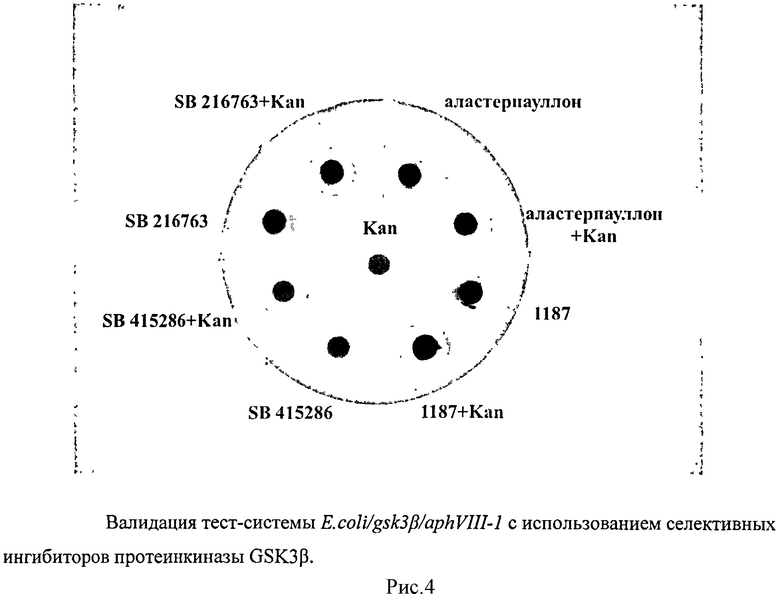
Все созданные конструкции были исследованы на устойчивость к аминогликозидному антибиотику канамицину. Штамм E.coli BL21 (DE3), содержащий плазмиду с исходным вариантом гена aphVIII, был устойчив к канамицину в концентрации 325 мкг/мл. Замены, произведенные в aphVIII-1, снижали уровень устойчивости на 38%, а в случае гена aphVIII-2 приводили к существенному снижению уровня устойчивости. В случае совмещения aphVIII и gsk3β в одном векторе для исходного фермента AphVIII и его варианта AphVIII-2 уровень устойчивости к канамицину не изменялся, а для конструкции aphVIII-1/gsk3β возрастал на 20%. Поэтому для последующих исследований была выбрана конструкция E.coli/gsk3β/aphVIII-1.
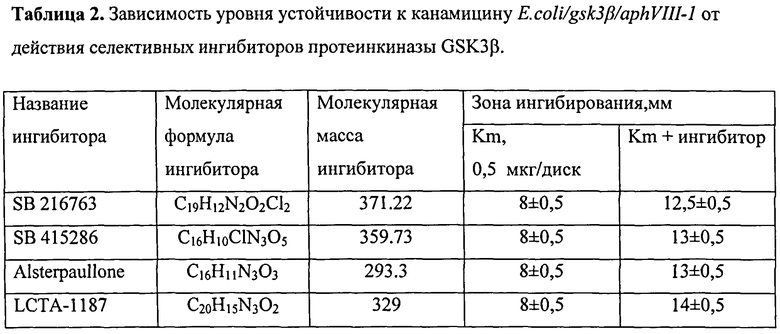
Для валидации тест-системы было исследовано влияние селективных ингибиторов протеинкиназы GSK3β - SB 216763; SB 415286; Alsterpaullone и LCTA-1187. Все ингибиторы вносили в концентрации 100 мкМ/диск. Исследования по валидации показали, что все 4 ингибитора увеличивают зону подавления роста. Это свидетельствует о том, что сконструированная тест-система E.coli/gsk3β/aphVIII-1 работает и может быть использована для отбора ингибиторов протеинкиназы GSK3β (рисунок 4, таблица 2).
Пример 3
Скрининг ингибиторов СТПК в валидированной тест-системе E.coli/gsk3β/aphVIII-1
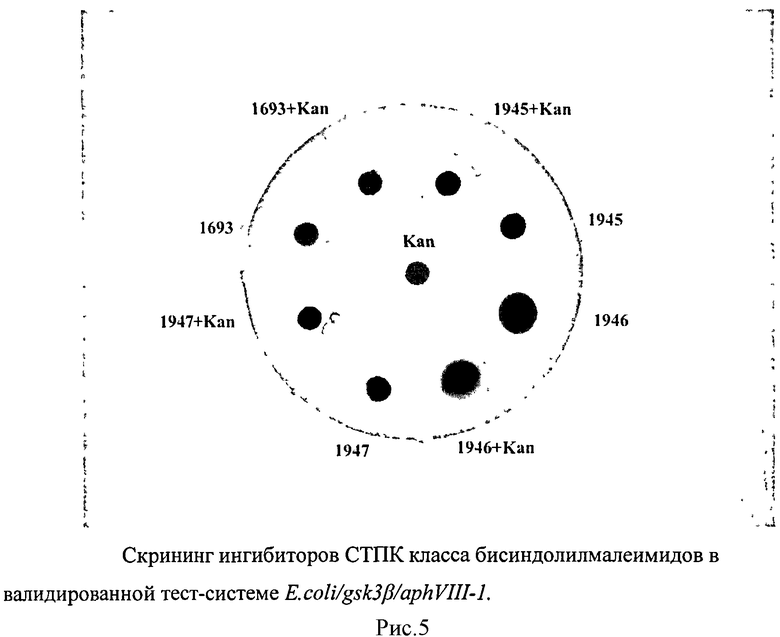
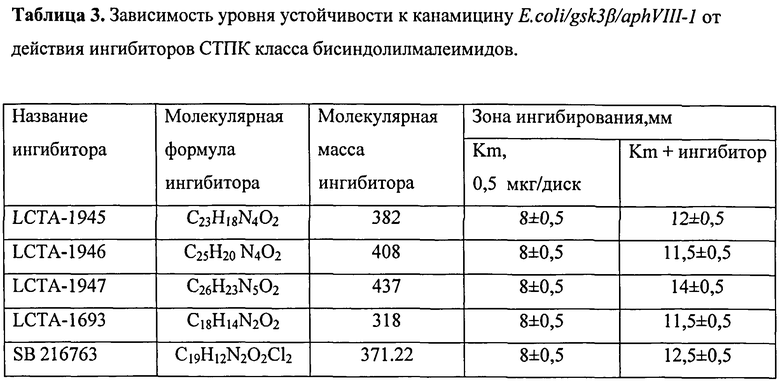
В валидированной тест-системе E.coli/gsk3β/aphVIII-1 проводился скрининг низкомолекулярных веществ класса бисиндолилмалеимидов, синтезированных в НИИНА им. Г.Ф. Гаузе РАМЫ и показавших активность в качестве ингибиторов СТПК на других тест-системах, созданных в лаборатории (таблица 6). Исследования проводили в 3-5 повторностях. Для выявления наиболее активного из исследуемых веществ, проводился анализ действия ингибиторов LCTA-1945, LCTA-1946, LCTA-1947, LCTA-1693 и LCTA-1210 в сравнении с селективным ингибитором протеинкиназы GSK3β - SB216763.
Вещество LCTA-1947 при совместном действии с канамицином, дает увеличение зоны ингибирования. При этом зона ингибирования превышает зону, получаемую при действии стандартного ингибитора фермента GSK3p SB216763, что говорит о повышенной активности вещества LCTA-1947 в отношении ингибирования протеинкиназы GSK3β. Все остальные вещества дают зону ингибирования роста бактерий, сопоставимую с зоной ингибирования SB 216763 (Рисунок 5, таблица 3).
Таким образом, отобрано наиболее активное вещество класса бисиндолилмалеимидов - LCTA-1947, которое может являться хит-соединением.




Изобретение относится к области молекулярной биологии и касается тест-системы для скрининга ингибиторов протеинкиназы GSK3β. Охарактеризованное изобретение представляет собой генетическую конструкцию, представленную на рис.2, полученную на основе экспрессионного вектора pET32a, и включающую ген каталитического домена протеинкиназы GSK3β, клонированного по сайтам эндонуклеаз рестрикции EcoRI и HindIII, и ген субстрата - аминогликозидфосфотрансферазы aphVIII из Streptomyces rimosus, клонированный по сайту эндонуклеазы рестрикции XbaI, в котором модифицирован сайт фосфорилирования AphVIII Ser-146 (AVAEGS146VDLED→ ASAEGS146VDLSD). Тест-система позволяет проводить прескрининг in situ ингибиторов GSK3β - низкомолекулярных соединений различных химических классов, способных проникать в клетки E.coli и грамположительных бактерий и может быть использовано в фармацевтике и медицинской химии, в частности в разработке, создании и валидации новых биомишеней и тест-систем с целью поиска химических соединений-лидеров, которые могут являться потенциальными кандидатами в лекарства для лечения заболеваний человека. 5 ил., 3 табл., 3 пр.
Тест-система E.coli/gsk3β/aphVIII для скрининга ингибиторов протеинкиназы GSK3β человека, представляющая собой генетическую конструкцию, представленную на рис.2, полученную на основе экспрессионного вектора pET32a и включающую ген каталитического домена протеинкиназы GSK3β, клонированного по сайтам эндонуклеаз рестрикции EcoRI и HindIII, и ген субстрата - аминогликозидфосфотрансферазы aphVIII из Streptomyces rimosus, клонированный по сайту эндонуклеазы рестрикции XbaI, в котором модифицирован сайт фосфорилирования AphVIII Ser-146 (AVAEGS146VDLED→ ASAEGS146VDLSD).
| БЕККЕР ОЛЬГА БОРИСОВНА, СОЗДАНИЕ БАКТЕРИАЛЬНОЙ ТЕСТ-СИСТЕМЫ ДЛЯ СКРИНИНГА ИНГИБИТОРОВ ПРОТЕИНКИНАЗ НА ОСНОВЕ ГЕНОВ ФОСФОТРАНСФЕРАЗ, Автореферат | |||
| дисс., Москва 2011, стр.1-24 | |||
| Jaakko Mattila et al., RNAi screening for kinases and phosphatases identifies FoxO regulators, Proc Natl Acad Sci U S A., 2008 September 30, Vol.105, N39, p.p.14873"14878 |

