Область техники, к которой относится изобретение
Настоящее изобретение относится к области биотехнологии, в частности к генетической инженерии, и может быть использовано в различных областях биотехнологии и научных исследованиях (тонкий органический синтез, селективное определение обычных и N-замещенных D-аминокислот и глицина и др.). Для достижения указанных целей сконструирована рекомбинантна плазмидная ДНК pNhaDAAO, содержащая модифицированный ген nhadaao, кодирующий оксидазу D-аминокислот архей Natrarchaeobius halalkaliphilus AArcht4 (NhaDAAO). Полученная плазмида обеспечивает экспрессию. включенного в ее нуклеотидную последовательность гена nhadaao в клетках Escherichia coli. В результате трансформации штамма E.coli BL21(DE3) предложенной плазмидой получен новый рекомбинантный штамм-продуцент E.coli BL21(DE3)/pNhaDAAO ВКПМ В-14556, обеспечивающий экспрессию NhaDAAO в активной форме. Для обеспечения эффективного контроля экспрессии в целевом штамме-продуценте в модифицированном гене nhadaao gjckt стоп кодона TAA в исходном гене в качестве добавлен второй стоп-кодон TGA. В результате трансформации штамма E.coli BL21(DE3) предложенной плазмидой получен новый рекомбинантный штамм-продуцент E.coli BL21(DE3))/pNhaDAAO ВКПМ В-14556, обеспечивающий экспрессию NhaDAAO в активной форме. Оксидазы D-аминокислот находят применение в самых различных областях науки и техники - фармацевтическая промышленность и тонкий органический синтез, определение D-аминокислот как маркеров различных заболеваний человека (включая нейродегенеративные), для определения обсемененности пищи бактериями, как потенциальный антимикробный агент и др.
Уровень техники
Оксидаза D-аминокислот (DAAO, КФ 1.2.1.2) играет важную роль в функционировании как прокариот, так низших (дрожжи и грибы) и высших эукариот (млекопитающие). В бактериях, дрожжах и микроскопических грибах основная роль этого фермента сводится к утилизации экзогенных D-аминокислот (в первую очередь D-Ala) [Tishkov, V. I. & Khoronenkova, S. V. (2005). D-Amino acid oxidase: structure, catalytic mechanism, and practical application. Biochemistry (Mosc.) 70, 40-54, doi:BCM70010051; Pollegioni, L., Piubelli, L., Sacchi, S., Pilone, M. S., & Molla, G. (2007a). Physiological functions of D-amino acid oxidases: from yeast to humans. Cell Mol. Life Sci. 64, 1373-1394, doi:10.1007/s00018-007-6558-4; Takahashi, S., Abe, K., & Kera, Y. (2015). Bacterial d-amino acid oxidases: Recent findings and future perspectives. Bioengineered. 6, 237-241, doi:10.1080/21655979.2015.10529172]. У высших эукариот - позвоночных и особенно млекопитающих, основная роль DAAO заключается в поддержании определенного уровня D-аминокислот, которые являются регуляторами важнейших процессов, в первую очередь нервной деятельности. Например, снижение уровня D-Ser в спинномозговой жидкости за счет повышенной активности DAAO приводит к шизофрении [Chumakov, I., Blumenfeld, M., Guerassimenko, O., et al. (2002). Genetic and physiological data implicating the new human gene G72 and the gene for D-amino acid oxidase in schizophrenia. Proc. Natl. Acad. Sci. U. S. A 99, 13675-13680, doi:10.1073/pnas.182412499; Cheng, Y. J., Lin, C. H., & Lane, H. Y. (2021). D-Amino Acids and pLG72 in Alzheimer's Disease and Schizophrenia. Int. J. Mol. Sci. 22, doi: 10.3390/ijms222010917]. Протекание болезней Альцгеймера и Паркинсона сопровождается повышением уровня D-Ala [Cheng, Y. J., Lin, C. H., & Lane, H. Y. (2021). D-Amino Acids and pLG72 in Alzheimer's Disease and Schizophrenia. Int. J. Mol. Sci. 22, doi: 10.3390/ijms222010917; Pernot, P., Mothet, J. P., Schuvailo, O., Soldatkin, A., Pollegioni, L., Pilone, M., Adeline, M. T., Cespuglio, R., & Marinesco, S. (2008). Characterization of a yeast D-amino acid oxidase microbiosensor for D-serine detection in the central nervous system. Anal. Chem. 80, 1589-1597, doi:10.1021/ac702230w4,5]. Оксидаза D-аминокислот также широко используется на практике [Khoronenkova, S. V. & Tishkov, V. I. (2008). D-amino acid oxidase: physiological role and applications. Biochemistry (Mosc.) 73, 1511-1518, doi: 10.1134/s0006297908130105; Pollegioni, L., Molla, G., Sacchi, S., Rosini, E., Verga, R., & Pilone, M. S. (2008). Properties and applications of microbial D-amino acid oxidases: current state and perspectives. Appl. Microbiol. Biotechnol. 78, 1-16, doi:10.1007/s00253-007-1282-4; Pollegioni, L. & Molla, G. (2011). New biotech applications from evolved D-amino acid oxidases. Trends Biotechnol. 29, 276-283, doi:;10.1016/j.tibtech.2011.01.010]. Например, DAAO из дрожжей Trigonopsis variabilis используется в двустадийном биокаталитическом процессе получения 7-аминоцефалоспорановой кислоты (7-АЦК) из цефалоспорина С [Barber MS, Giesecke U, Reichert A, Minas W. 2004. Industrial enzymatic production of cephalosporin-based beta-lactams. Adv. Biochem. Eng Biotechnol. 88:179-215.10; Atroshenko, D. L., Shelomov, M. D., Zarubina, S. A., Negru, N. Y., Golubev, I. V., Savin, S. S., & Tishkov, V. I. (2019). Multipoint TvDAAO Mutants for Cephalosporin C Bioconversion. Int. J. Mol. Sci. 20, doi: 10.3390/ijms20184412]. 7-АЦК является исходным синтоном для производства полусинтетических цефалоспоринов различных поколений. По сравнению с ранее используемым чисто химическим такой процесс позволяет сократить расход органических растворителей в 400 раз. Объем производства 7-АЦК составляет более 10 тыс. тонн в год. Описан биокаталитический способ получения природного гербицида широкого спектра действия-L-глюфозината, в котором на первой стадии используется DAAO [Green B.M., Gradley M.L. Methods For Making L-Glufosinate. US 2017 / 0253897 A1 Published 07.09.2017. filed 28.02.2017]. DAAO используется в фармацевтической промышленности, например, при получении ингибитора ангиотензинпревращаюего фермент Омапатрилата [Patel, R. N. (2001). Enzymatic synthesis of chiral intermediates for Omapatrilat, an antihypertensive drug. Biomol. Eng 17, 167-182.].
Отдельной областью современной аналитической биотехнологии является разработке методов селективного определения различных D-аминокислот в сложных биологических системах. Например, D-Ala является сшивающим агентом в муреиновом слое клеток грам-положительных и грам-отрицательных бактерий. Поэтому наличие в образце D-Ala позволяет определить бактериальное заражение объекта даже если бактерии уже неживые. Однако в первую очередь особое внимание уделяется определению D-аминокислот в тканях и жидкостях человека, поскольку они являются регуляторами важнейших процессов, в первую очередь нервной деятельности. Поскольку основная часть аминокислот в организме находится в виде L-изомера необходимы стереоселективные методы детекции именно D-аминокислот и такая селективность как раз может быть достигнута с помощью оксидаз D-аминокислот [Molla, G., Piubelli, L., Volonte, F., & Pilone, M. S. (2012). Enzymatic detection of D-amino acids. Methods Mol. Biol. 794, 273-289, doi:10.1007/978-1-61779-331-8_18]. Для экспресс-анализа и его проведения in situ были разработаны биосенсоры для определения ряда D-аминокислот [Sacchi, S., Rosini, E., Caldinelli, L., & Pollegioni, L. (2012). Biosensors for D-amino acid detection. Methods Mol. Biol. 794, 313-324, doi:10.1007/978-1-61779-331-8_21]. Однако разработка новых методов и биосенсоров на D-аминокислоты ограничена как свойствами известных оксидаз D-аминокислот, так и их малым выбором. В настоящее время получены и охарактеризовано небольшое количество DAAO - из почек свиньи [Fukui, K., Watanabe, F., Shibata, T., & Miyake, Y. (1987). Molecular cloning and sequence analysis of cDNAs encoding porcine kidney D-amino acid oxidase. Biochemistry 26, 3612-3618, doi:10.1021/bi00386a054], мыши [Tada, M., Fukui, K., Momoi, K., & Miyake, Y. (1990)] Cloning and expression of a cDNA encoding mouse kidney D-amino acid oxidase. Gene 90, 293-297, doi: 10.1016/0378-1119(90)90193-u], человека [Fukui, K. & Miyake, Y. (1992). Molecular cloning and chromosomal localization of a human gene encoding D-amino-acid oxidase. J. Biol. Chem. 267, 18631-18638, doi:S0021-9258(19)37007-3], карпа [Sarower, M. G., Okada, S., & Abe, H. (2003). Molecular characterization of D-amino acid oxidase from common carp Cyprinus carpio and its induction with exogenous free D-alanine. Arch. Biochem. Biophys. 420, 121-129, doi: 10.1016/j.abb.2003.09.035], дрожжей и микроскопических грибов Fusarium solani (FsoDAAO) [Isogai, T., Ono, H., Ishitani, Y., Kojo, H., Ueda, Y., & Kohsaka, M. (1990). Structure and expression of cDNA for D-amino acid oxidase active against cephalosporin C from Fusarium solani. J. Biochem. 108, 1063-1069, doi:10.1093/oxfordjournals.jbchem.a123306], Trigonopsis variabilis (TvaDAAO) [Gonzalez, F. J., Montes, J., Martin, F., Lopez, M. C., Ferminan, E., Catalan, J., Galan, M. A., & Dominguez, A. (1997). Molecular cloning of TvDAO1, a gene encoding a D-amino acid oxidase from Trigonopsis variabilis and its expression in Saccharomyces cerevisiae and Kluyveromyces lactis. Yeast 13, 1399-1408, doi: doi:10.1002/(SICI)1097-0061(199712)13:15<1399], Rhodosporidium toruloides (предыдущее название Rhodotorula gracilis) (RtoDAAO) [Pollegioni, L., Molla, G., Campaner, S., Martegani, E., & Pilone, M. S. (1997). Cloning, sequencing and expression in E. coli of a D-amino acid oxidase cDNA from Rhodotorula gracilis active on cephalosporin C. J. Biotechnol. 58, 115-123, doi: 10.1016/s0168-1656(97)00142-9]. Pichia pastoris (PpaDAAO) [Klompmaker, S. H., Kilic, A., Baerends, R. J., Veenhuis, M., & van der Klei, I. J. (2010). Activation of a peroxisomal Pichia pastoris D-amino acid oxidase, which uses d-alanine as a preferred substrate, depends on pyruvate carboxylase. FEMS Yeast Res. 10, 708-716, doi:10.1111/j.1567-1364.2010.00647.x], Candida boidinii (CboDAAO) [Yurimoto, H., Hasegawa, T., Sakai, Y., & Kato, N. (2001). Characterization and high-level production of D-amino acid oxidase in Candida boidinii. Biosci. Biotechnol. Biochem. 65, 627-633], Rasamsonia emersonii strain YA (RemDAAO) [Shimekake, Y., Furuichi, T., Abe, K., Kera, Y., & Takahashi, S. (2019). A novel thermostable D-amino acid oxidase of the thermophilic fungus Rasamsonia emersonii strain YA. Sci. Rep. 9, 11948, doi:10.1038/s41598-019-48480-y [doi];10.1038/s41598-019-48480-y. ] и бактерий из Rubrobacter xylanophilus (RxyDAAO) [Takahashi, S., Furukawara, M., Omae, K., Tadokoro, N., Saito, Y., Abe, K., & Kera, Y. (2014). A Highly Stable D-Amino Acid Oxidase of the Thermophilic Bacterium Rubrobacter xylanophilus. Appl. Environ. Microbiol. 80, 7219-7229, doi: 10.1128/AEM.02193-14], Streptomyces coelicolor (ScoDAAO) [Saito, Y., Takahashi, S., Kobayashi, M., Abe, K., & Kera, Y. (2014). D-Amino acid oxidase of Streptomyces coelicolor and the effect of D-amino acids on the bacterium. Annals of Microbiology 64, 1167-1177. doi.org/10.1007/s13213-013-0756-0] и Arthrobacter protophormiae (AprDAAO) [Geueke, B., Weckbecker, A., & Hummel, W. (2007). Overproduction and characterization of a recombinant D-amino acid oxidase from Arthrobacter protophormiae. Appl. Microbiol. Biotechnol. 74, 1240-1247, doi:10.1007/s00253-006-0776-9]. Однако, все описанные в литературе DAAO, используемые для детекции D-аминокислот, имеют широкий спектр субстратной специфичности, т.е. проявляют активность со многими D-аминокислотами, что не позволяет обеспечить селективность анализа к определенной аминокислоте. В некоторых случаях, условия анализа позволяют достигнуть определенной селективности за счет выбора фермента с определенной субстратной специфичностью или исходя из особенности условий проведения анализа. Например, DAAO из дрожжей R. gracilis имеет одинаковые каталитические характеристики с D-Ser и D-Ala и селективность определения D-Ser в спинномозговой жидкости достигается только за счет того, что физиологическая концентрация D-Ala в 10 раз ниже, чем концентрация D-Ser [Pernot, P., Mothet, J. P., Schuvailo, O., Soldatkin, A., Pollegioni, L., Pilone, M., Adeline, M. T., Cespuglio, R., & Marinesco, S. (2008). Characterization of a yeast D-amino acid oxidase microbiosensor for D-serine detection in the central nervous system. Anal. Chem. 80, 1589-1597, doi:10.1021/ac702230w]. Однако данный подход покрывает небольшую область необходимых анализов. Проведение экспериментов по изменению спектра субстратной специфичность не позволило решить данную проблему [Sacchi, S., Lorenzi, S., Molla, G., Pilone, M. S., Rossetti, C., & Pollegioni, L. (2002). Engineering the substrate specificity of D-amino-acid oxidase. J. Biol. Chem. 277, 27510-27516, doi:10.1074/jbc.M203946200; Setoyama, C., Nishina, Y., Mizutani, H., Miyahara, I., Hirotsu, K., Kamiya, N., Shiga, K., & Miura, R. (2006). Engineering the substrate specificity of porcine kidney D-amino acid oxidase by mutagenesis of the "active-site lid". J. Biochem. 139, 873-879, doi: 10.1093/jb/mvj094; Komarova, N.V., Golubev, I.V., Khoronenkova, S.V., Chubar', T.A., & Tishkov, V.I. (2012). Engineering of substrate specificity of D-amino acid oxidase from the yeast Trigonopsis variabilis: directed mutagenesis of Phe258 residue. Biochemistry (Mosc.) 77, 1181-1189, doi: 10.1134/S0006297912100100]. Из уровня техники известен рекомбинантный штамм E.coli BL21(DE3)/pNhyDAAO. - продуцент оксидазы D-аминокислот, высокоспецифичной к D-Phe [RU2803295, опубл. 12.09.2023].
Кроме обычных D-аминокислот в организме образуются их различные производные, в первую очередь варианты, в которых алкилирована аминогруппа. Наиболее известными метаболитами являются саркозин и бетаин (N-метилглицин и N-триметилглицин соответственно). В литературе появляются все больщее число работ, в которых обсуждается роль таких модифицированных аминокислот при возникновении различных заболеваний, например болезни Паркинсона [Socha E, Koba M, Kośliński P. Amino acid profiling as a method of discovering biomarkers for diagnosis of neurodegenerative diseases. Amino Acids. 2019;51(3):367-371. doi: 10.1007/s00726-019-02705-6]. В этом случае требуется определение не одной аминокислоты, а их набора, причем необходимо анализировать как свободные, так и модифицированные аминокислоты. Такой набор можно определять с помощью различных инструментальных методов (LC-MS, ВЭЖХ, капиллярный электрофорез и т.д.). Однако эти методы очень трудоемки, требуют дорогого оборудования и расходных материалов и могут быть реализованы только в больших аналитических диагностических лабораториях. Альтернативой являются биосенсоры на основе ферментов. В случае оксидаз это могут быть электрохимические биосенсоры, которые детектируют реакцию по образованию пероксида водорода (например, анализ глюкозы). К сожалению, ферменты с широким спектром субстратной специфичности, способные окислять как свободные, так и модифицированные аминокислоты, в настоящее время неизвестны. Например, саркозиноксидаза окисляет субстрат до глицина и формальдегида, но уже не способна окислять глицин и тем более другие аминокислоты. Оксидазы, способные окислять бетаин, в литературе не описаны.
Решением данной проблемы является создание рекомбинантных штаммов -продуцентов новых оксидаз D-аминокислот с необходимой субстратной специфичностью, способных окислять как свободные, так и модифицированные аминокислоты. Для выбора пути и способа решения проблемы необходимо учитывать следующие факторы:
1. Фермент должен обладать необходимым спектром субстратной специфичности, т.е должен обеспечивать возможность широкого набора свободных аминокислот, глицина и их модифицированных производных. Прототипы таких оксидаз в литературе не описаны
2. Создание рекомбинантного штамма - продуцента DAAO. Природные штаммы микроорганизмов не могут быть использованы для получения фермента, поскольку уровень экспрессии DAAO в нативных штаммах очень низок. Например, в случае Arthrobacter protophormiae удельная активность в бесклеточном экстракте составляет всего 0,003 ед. на мг белка, что соответствует уровню экспрессии 0,01%, в то время как в рекомбинантных штаммах уровень экспрессии составляет до 30-40% от растворимого белка клетки. Поэтому для обеспечения возможности получения достаточных количеств целевого фермента необходимо использовать методы генной инженерии и создавать рекомбинантный штамм-продуцент. В литературе описано получение рекомбинантных DAAO в различных штаммах-продуцентах. В случае DAAO дрожжей и микроскопических грибов используются как бактерии E.coli [Molla, G., Vegezzi, C., Pilone, M. S., & Pollegioni, L. (1998). Overexpression in Escherichia coli of a recombinant chimeric Rhodotorula gracilis d-amino acid oxidase. Protein Expr. Purif. 14, 289-294, doi: 10.1006/prep.1998.0956; Dib, I., Stanzer, D., & Nidetzky, B. (2007). Trigonopsis variabilis D-amino acid oxidase: control of protein quality and opportunities for biocatalysis through production in Escherichia coli. Appl. Environ. Microbiol. 73, 331-333, doi:10.1128/AEM.01569-06; RU2310687, опубл. 20.11.2007], так и различные типы дрожжей [Dominguez, A. (1997). Molecular cloning of TvDAO1, a gene encoding a D-amino acid oxidase from Trigonopsis variabilis and its expression in Saccharomyces cerevisiae and Kluyveromyces lactis. Yeast 13, 1399-1408; Abad, S., Nahalka, J., Winkler, M., Bergler, G., Speight, R., Glieder, A., & Nidetzky, B. (2011). High-level expression of Rhodotorula gracilis D-amino acid oxidase in Pichia pastoris. Biotechnol. Lett. 33, 557-563, doi:10.1007/s10529-010-0456-9; Isoai, A., Kimura, H., Reichert, A., Schorgendorfer, K., Nikaido, K., Tohda, H., Giga-Hama, Y., Mutoh, N., & Kumagai, H. (2002). Production of D-amino acid oxidase (DAO) of Trigonopsis variabilis in Schizosaccharomyces pombe and the characterization of biocatalysts prepared with recombinant cells. Biotechnol. Bioeng. 80, 22-32, doi:10.1002/bit.10345; Yurimoto, H., Hasegawa, T., Sakai, Y., & Kato, N. (2001). Characterization and high-level production of D-amino acid oxidase in Candida boidinii. Biosci. Biotechnol. Biochem. 65, 627-633]. В случае бактериальных DAAO в качестве штамма-продуцента используются только клетки E.coli [Takahashi, S., Furukawara, M., Omae, K., Tadokoro, N., Saito, Y., Abe, K., & Kera, Y. (2014). A Highly Stable D-Amino Acid Oxidase of the Thermophilic Bacterium Rubrobacter xylanophilus. Appl. Environ. Microbiol. 80, 7219-7229, doi: 10.1128/AEM.02193-14; Saito, Y., Takahashi, S., Kobayashi, M., Abe, K., & Kera, Y. (2014). D-Amino acid oxidase of Streptomyces coelicolor and the effect of D-amino acids on the bacterium. Ann. Microbiol. 64, 1167-1177. doi.org/10.1007/s13213-013-0756-0; Geueke, B., Weckbecker, A., & Hummel, W. (2007). Overproduction and characterization of a recombinant D-amino acid oxidase from Arthrobacter protophormiae. Appl. Microbiol. Biotechnol. 74, 1240-1247, doi:10.1007/s00253-006-0776-9].
Исходя из вышеизложенного следует констатировать, что в настоящее время отсутствуют прототипы и аналоги DAAO, окисляющих как свободные аминокислоты, так и их N-модифицированные варианты.
Таким образом, техническая проблема, решаемая посредством заявляемого изобретения, заключается в необходимости создания рекомбинантного штамма E-coli - продуцента оксидазы D-аминокислот, которая имеет широкий спектр субстратной специфичности и способна окислять как свободные, так и модифицированные по аминогруппе глицин и различные D-аминокислоты.
Раскрытие изобретения
Техническим результатом заявляемой группы изобретений является разработка плазмиды и штамма, модифицированного данной плазмидой, которая обеспечивает синтез целевого фермента NhaDAAO в клетках E.coli, которая имеет широкий спектр субстратной специфичности и способна окислять как свободные, так и модифицированные по аминогруппе глицин и различные D аминокислоты. За счет наличия в плазмиде гена белка-репрессора laqI осуществляется строгий контроль процесса экспрессии. Отсутствие преждевременной гибели клеток свидетельствует о нетоксичности экспрессируемого фермента. Высокий уровень экспрессии достигается за счет использования высокоэффективного промотора РНК-полимеразы фага Т7.
Штамм продуцент E.coli BL21(DE3)/pNhaDAAO содержит на хромосоме ген РНК-полимеразы фага Т7, что позволяет использовать полученную плазмиду pNhaDAAO для экспрессии целевого белка. Кроме того, исходный штамм E.coli BL21(DE3), который использовали для получения целевого штамма-продуцента мутантен по основной протеазе клеток E.coli (lonA-), что значительно предотвращает деградацию целевого фермента в процессе культивирования штамма-продуцента. Заявляемый штамм продуцент E.coli BL21(DE3)/pNhaDAAO продуцирует целевой фермент в активной форме.
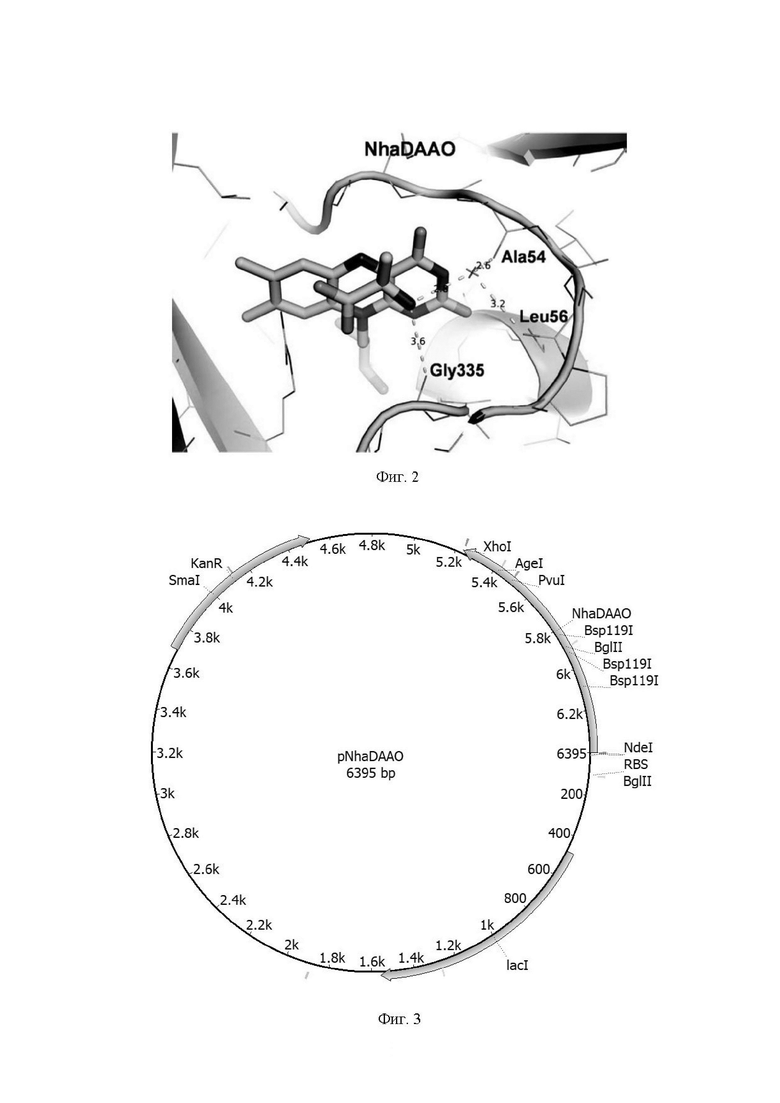
Технический результат достигается рекомбинантной плазмидой pNhaDAAO размером 6395 п.о., содержащей модифицированный ген nhadaao оксидазы D-аминокислот из архей N. halalkaliphilus AArcht4 под контролем T7 промотора. обеспечивающей синтез оксидазы D-аминокислот из архей N. halalkaliphilus AArcht4 (NhaDAAO) в клетках E. coli. Плазмида pNhaDAAO получена путем клонирования по сайтам NdeI/Xho фрагмента размером 1165 п.о. (последовательность SEQ ID №3) с геном, кодирующим NhaDAAO (1158 п.о.) в плазмиду pET24a(+), расщепленную рестриктазами NdeI и Xho.
Также технический результат достигается рекомбинантным штаммом Escherichia coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 - продуцентом оксидазы D-аминокислот NhaDAAO, полученным путем трансформации штамма Escherichia. coli BL21(DE3) заявляемой плазмидной ДНК, задепонированный во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под номером В-14556.
Для обеспечения эффективной экспрессии новой DAAO в клетках E.coli ген фермента клонируется в плазмидный вектор под контроль сильного промотора РНК-полимеразы фага Т7. В используемом для клонирования векторе pET24a(+) имеется ген Kan, который в качестве селективного маркера обеспечивает устойчивость к канамицину. Кроме того, исходная плазмида pET24a(+) содержит ген белка-репрессора lac-промотора lacI, который предотвращает неспецифическую базовую экспрессию целевого белка. Для обеспечения строгого контроля терминации трансляции в нуклеотидную последовательность гена nhadaao в дополнение к собственному стоп-кодону ТАА добавлен второй стоп-кодон TGA.
Краткое описание чертежей
Изобретение поясняется следующими чертежами.
Фиг.1 Выравнивание аминокислотных последовательностей потенциальных DAAO, отобранных после поиска по гомологии в геномах бактерий в базах UniProt (NCBI) и коллекции микроорганизмов МГУ имени М.В. Ломоносова и ФИЦ Биотехнологии РАН. Выравнивание выполнено с помощью программы Clustal X 1.83.
Сокращения.
Бактерии: RxyDAAO - Rubrobacter xylanophilus; ScoDAAO - Streptomyces coelicolor; AprDAAO - Arthrobacter protophormiae; NhyDAAO - Natronosporangium hydrolyticum ACPA39; GthDAAO - Gandjariella thermophila; CthDAAO - Chloracidobacterium thermophilum B; RtaDAAO - Rubrobacter taiwanensis; RraDAAO - Rubrobacter radiotolerans DSM 5868. Археи: NhaDAAO - Natrarchaeobius halalkaliphilus AArcht4
Фиг. 2. Модельная структура активного центра оксидазы D-аминокислот из N. halalkaliphilus AArcht4. Структура получена моделированием с использованием алгоритма AjphaFold2. Показаны остатки Ala54, Leu56 и Gly335, обеспечивающие дополнительное пространство в активном центре для связывания аминокислот с модифицированной аминогруппой.
Фиг. 3. Физическая и генетическая карта плазмидного вектора pNhaDAAO. Обозначены положения индикаторных сайтов рестрикции, рибосом-связывающий участок, ген устойчивости к канамицину (Kan) и ген lac-репрессора (lacI).
Фиг. 4. Представлена последовательность фрагмента NdeI-XhoI с последовательностью SEQ ID № 3.
Осуществление изобретения
С учетом требуемой субстратной специфичности новой DAAO и необходимости создания системы экспрессии (комбинации "экспрессирующий вектор"/"штамм-реципиент") этого фермента достижение поставленных целей было реализовано за счет того, что
1) в результате анализа генов потенциальных DAAO в геномах бактерий и архей была отобрана оксидаза D-аминокислот, которая по данным биоинформационно-структурного анализа должна была обладать требуемой субстратной специфичностью.
2) сконструирован экспрессионный вектор, содержащий ген отобранного фермента и который должен обеспечивать экспрессию этого гена в клетках E.coli. В качестве базового вектора для клонирования использовали плазмиду pET24a(+), которая обеспечивает длительное сохранение экспрессионного вектора в клетке в процессе культивирования за счет наличия гена устойчивости к канамицину и которая не допускает неспецифической базовой экспрессии целевого гена за счет наличия в плазмидном векторе гена белка- ингибитора lac- промотора lacI.
3) путем трансформации полученным вектором получается рекомбинантный штамм E.coli - продуцент выбранной DAAO. Проводится культивирование полученного штамма-продуцента для проверки его способности экспрессировать активный рекомбинантный фермент
4) изучается профиль субстратная специфичность новой DAAO.
I. Выбор целевого фермента проводится в три этапа:
1) первый этап включает выбор гена на основе биоинформативного анализа аминокислотных последовательностей потенциальных DAAO бактерий и архей. Отбирают последовательность, которая демонстрирует отличие от последовательностей «классических» DAAO;
2) вторым этапом является сравнение модельной структуры отобранного фермента с экспериментальными и модельными структурами DAAO с известными свойствами. Выбирают фермент с величиной RMSD в граничном диапазоне для DAAO и за пределами RMSD с глициноксидазами;
3) далее был проведен анализ структуры активного центра, на основании которого был сделан выбор в пользу фермента из архей Natrarchaeobius halalkaliphilus AArcht4 (NhaDAAO).
II. Конструирование экспрессирующего вектора включает три этапа:
1) анализ нуклеотидной последовательности гена, кодирующего NhaDAAO, Для обеспечения эффективной терминации трансляции гена nhadaao в клетках E.coli вслед имеющимся в гене стоп-кодоне TAA был добавлен другой стоп-кодон TAG, Такой тандем стоп-кодонов эффективен к клетках E.coli;
2) клонирование гена mhadaao проводили с помощью полимеразной цепной реакции с использованием прямого и обратного праймеров (SEQ ID №1 и SEQ ID №2). Для клонирования в экспрессирующий вектор в прямой и обратный праймеры были введены соответственно сайты рестрикции NdeI и XhoI;
3) экспрессирующий вектор pNhaDAAO был получен путем лигирования продукта ПЦР с геном nhadaao в плазмидный в вектор pET24a(+) после их обработки рестриктазами Ndei и XhoI. Клонированная последовательность указана как SEQ ID №3 (фиг. 4). Сконструированный вектор pNhaDAAO (Фиг 3) содержит ген устойчивости к канамицину и ген белка-репрессора lac-промотора.lacI.
Ш. Конструирование рекомбинантного штамма E.coli - продуцента NhaDAAO
Рекомбинантный штамм E. coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 - продуцент NhaDAAO был получен трансформацией компетентных клеток E.coli BL21(DE3) экспрессирующей плазмидой pNhaDAAO. Сконструированный штамм Escherichia coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 продуцировал рекомбинантную NhaDAAO в активной форме.
Штамм E.coli BL21(DE3)/pNhaDAAO депонирован во Всероссийской коллекции промышленных микроорганизмов как E. coli BL21(DE3)/pNhaDAAO ВКПМ В-14556.
Заявляемый штамм E. coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 имеет следующие морфологические и физиолого-биохимические характеристики:
Морфологические признаки
Клетки прямые, палочковидные, подвижные, грамотрицателиные, неспорообразующие. При выращивании в течение 24-48 час при температуре 30-37°С на агаризованных средах (LB, 2YT), содержащих канамицин (25 мкг/мл), колонии гладкие, круглые, блестящие, край ровный.
Состав среды LB, мас.%:
Бакто-триптон - 1
дрожжевой экстракт - 0,5
NaCl - 1,
вода - остальное.
Состав среды 2YT, мас.%:
Бакто-триптон - 1,6
дрожжевой экстракт - 1,0
NaCl - 0,5
вода - остальное.
Физиолого-биохимические признаки
Штамм растет при температуре от 15 до 37°С (оптимум 30-37°С). В качестве источника азота использует органический азот в виде триптона, аминокислот.
Штамм E. coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 синтезирует оксидазу D-аминокислот NhaDAAO после нндукции 0,1 мМ ИПТГ (изопропил-β-D-тиогалактозидом).
Генотипические признаки
Штамм E. coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 устойчив к канамицину.
Штамм E.coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 содержит на плазмиде pNhaDAAO ген оксидазы D-аминокислот nhadaao из N. halalkaliphilus AArcht4 с дополнительным стоп-кодоном TAG под контролем T7 промотора.
Условия хранения:
1. В ночную культуру, выращенную на жидкой среде 2YT с канамицином (25 мгк/мл), добавляют стерильный глицерин (до 15%), расфасовывают по 1,5 мл в пластиковые микропробирки и хранят в замороженном виде при -70°С.
2. Хранят в лиофилизированном виде в ампулах при +4°C.
3. Штамм может поддерживаться регулярными пересевами (1 раз в 3-4 недели) на агаризованной среде LB или 2YT, содержащей канамицин 25 мкг/мл.
IV. Определение спектра субстратной специфичности
Для определения спектра субстратной специфичности используют препараты бесклеточного экстракта с новой NhaDAAO. Для получения препаратов NhaDAAO штамм E.coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 культивировали при 30°С на среде 2YT в присутствии канамицина (25 мкг/мл). Индукцию биосинтеза проводили с помощью ИПТГ (0,1 мМ) по достижении поглощения культуральной среды A600 1 ед. поглощения. Культивирование продолжали при 20°С в течение 48 часов. Клетки осаждали центрифугированием при 5000 об/мин при 4°С. Далее клетки ресуспендировали в 0,1 мМ фосфатном буфере, pH 8,0 и разрушали на ультразвуковом дезинтеграторе при 4°С. Клеточный дебрис осаждали центрифугированием при 14000 об/мин, 30 мин, 4°С. Супернатант использовали для определения активности с набором обычных и модифицированных по аминогруппе D-аминокислот (10 мМ) в 0,1 М буфере при рН 8,0. Активность определяли пероксидазным методом с о-дианизидином в качестве субстрата пероксидазы.
Таким образом, были получены:
- рекомбинантная плазмида pNhaDAAO размером 6395 п.о., содержащая модифицированный ген nhadaao оксидазы D-аминокислот из N. halalkaliphilus AArcht4 под контролем T7 промотора. обеспечивающая синтез оксидазы D-аминокислот из N. halalkaliphilus AArcht4 (NhaDAAO) в клетках E. coli. Плазмида pNhaDAAO получена путем клонирования по сайтам NdeI/Xho фрагмента размером 1165 п.о. (последовательность SEQ ID №3) с геном, кодирующим NhaDAAO (1158 п.о.) в плазмиду pET24a(+), расщепленную рестриктазами NdeI и Xho;
- рекомбинантный штамм Escherichia coli BL21(DE3)/pNhaDAAO - продуцент оксидазы D-аминокислот NhaDAAO, полученный путем трансформации штамма Escherichia. coli BL21(DE3) плазмидной ДНК.
Изобретение поясняется следующими примерами.
Пример 1. Выбор источника новой DAAO на основе биоинформацинно-структурного анализа.
Разработанный нами биоинформационно-структурный подход поиска генов потенциальных DAAO в геномах микроорганизмов [Atroshenko D.L., Golovina D.I., Sergeev E.P., Shelomov M.D., Elcheninov A.G., Kublanov I.V., Chubar T.A., Pometun A.A., Savin S.S., Tishkov V.I. Bioinformatics-structural approach to the search for new D-amino acid oxidases. Acta Naturae 2022, Vol. 14 № 4 (55), p. 57-68. DOI: 10.32607/actanaturae.11812] позволил отобрать 9 последовательностей из бактерий и одну из архей (фиг. 1).
Анализ результатов множественного выравнивания (Фиг. 1) свидетельствует, что бактериальные ферменты (первые 9 последовательностей) имеют практическую одинаковую длину (отличие только на N-конце). На фиг. 1 хорошо видно, что последовательности DAAO из бактерий хорошо совпадают по длине, в то время как фермент из архей (NhaDAAO, нижняя строчка) имеет более длинную последовательность, что приводит к появлению разрывов в выравнивании ферментов. Всего в выравнивании наблюдается 9 четко локализованных разрывов, а также на С-конце NhaDAAO имеется более длинная последовательность. Наличие дополнительных участков в последовательности NhaDAAO должно влиять на каталитические свойства фермента и поэтому уже на этапе сравнения аминокислотных последовательностей можно высказать предположение, что NhaDAAO должна иметь субстратную специфичность несколько отличную от ферментов из бактерий.
Дополнительным аргументом в пользу предположения об индивидуальности субстратной специфичности NhaDAAO служат данные по сравнению трехмерных экспериментальных и модельных структур DAAO и глициноксидаз. Из таблицы 1 следует, что NhaDAAO однозначно не относится к глициноксидазам - среднеквадратичное отклонение составляет более 9 ангстрем, в то время как величина RMSD при сравнении с оксидазами D-аминокислот довольно значительно - 2-3 ангстрема, при этом между «классическими» DAAO такие различия составляет 2 ангстрема и менее. Интересным является факт, что наименьшее отличие наблюдается между структурами NhaDAAO и DAAO человека (1,96 ангстрема).
Таблица 1.
Среднеквадратичное отклонение между структурами оксидаз D-аминокислот и глициноксидаз.
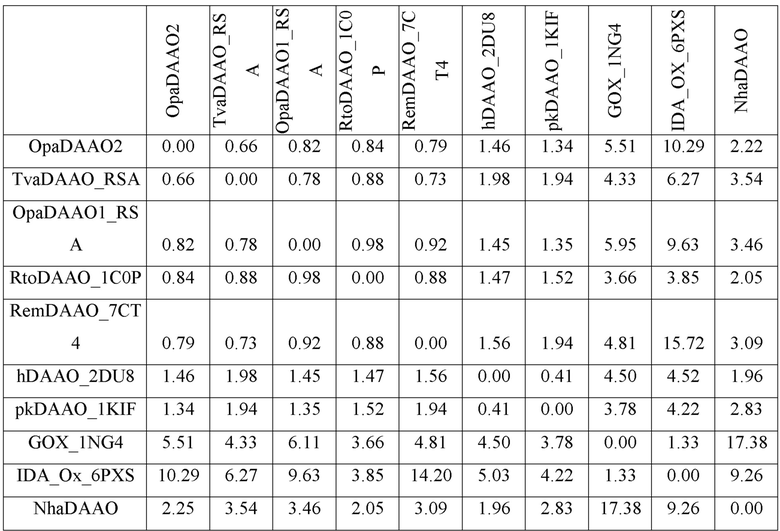
Анализ модельного активного центра NhaDAAO (фиг. 2) указывает на его существенное отличие в связывании субстрата в активном центре от активного центра других оксидаз D-аминокислот. Обычно в DAAO аминогруппа D-аминокислоты связывается водородными связями с кислородом полиппетидной цепи близлежащей альфа спирали и через молекулу воды удерживается двумя остатками аспарагина/глутамина. В случае NhaDAAO на месте высококонсервативных остатков аспарагина/глутамина находятся остатки Ala54 и Leu56 (боковой радикал последнего направлен в противоположную сторону от субстрата), а в 335 положении вместо остатка Ser расположен остаток Gly.
Таким образом, на основании биоинформационно-структурного анализа для создания штамма-продуцента новой DAAO была выбрана NhaDAAO.
Пример 2. Конструирование экспрессионного вектора pNhaDAAO
Экспрессионный вектор pNhaDAAO получали клонированием в плазмиду pET24a(+) фрагмента ДНК с геном nhadaao. Фрагмент ДНК с геном nhadaao и дополнительным стоп-кодоном получали с помощью полимеразной цепной реакции (ПЦР). В качестве матрицы использовали геномную ДНК Natrarchaeobius halalkaliphilus AArcht4, которую выделяли из клеток указанных архей с помощью набора для выделения геномной ДНК фирмы ThermoFisher Scientific «GeneJET Genomic DNA Purification Kit» согласно протоколу фирмы производителя. Для проведения ПЦР использовали прямой AArcht4_DAAO_F_NdeI
5'-GCAGGTTCAATACATATGCAACCAGACATAACTGTCCTCG-3'
и обратный праймеры
AArcht4_DAAO_R_XhoI
5'-CGTACACTCGAGCTATTATTCGCATATATGTTTGGTTAGTGTTCG-3'
В прямой праймер был введен сайт рестрикции NdeI CATATG (указан полужирным шрифтом). В обратном праймере для клонирования был введен сайт рестрикции XhoI CTCGAG, а вместо стоп-кодона TAA введен тандем стоп-кодонов TAATAG (выделено подчеркиванием).
ПЦР проводили в тонкостенной пробирке объемом 0,5 мл. Состав реакционной смеси приведен в таблице 2.
Таблица 2.
Состав реакционной смеси для проведения полимеразной цепной реакции
Перед началом ПЦР реакционную смесь без ДНК-полимеразы инкубировали 3 мин при 98°С, затем в пробирку добавляли фермент и проводили ПЦР по следующей программе 1-я стадия - 95°С, 15 с; 2-стадия - 56°С, 20 с и 3-я стадия - 72°С, 2 мин, всего -35 циклов. После этого реакционную смесь выдерживали еще 5 мин при 72°С. Продукт ПЦР очищали с помощью электрофореза в 1% агарозном геле. Вырезали фрагмент размером 1000 п.о и выделяли ДНК из геля с помощью набора для выделения и очистки ДНК из агарозного геля «PureLink Quick Gel Extraction Kit» (ThermoFisher Scientific).
Очищенный ПЦР фрагмент ДНК и плазмиду pET24a(+) обрабатывают рестриктазами NdeI и XhoI. Условия проведения реакций представлены в таблице 3.
Таблица 3
Условия расщепления продукта ПЦР и плазмиды pET24a(+)ю
Реакцию проводят в пробирке объемом 0,5 мл в течение 1,5 часов при 37°С. Продукты обработки рестриктазами очищали с помощью электрофореза в 1% агарозном геле. Затем ДНК экстрагировали из геля и очищали с помощью набора для выделения и очистки ДНК из агарозного геля «PureLink Quick Gel Extraction Kit» (ThermoFisher Scientific).
Очищенные продукты рестрикции лигировали при следующих условиях (Таблица 4).
Таблица 4
Условия лигирования продуктов рестрикции ПЦР и плазмиды pET24a(+).
Реакцию лигирования проводили в течение 3 часов при 22°С. Затем по стандартной методике [Sambrook J, Maniatis T, Fritsch EF. 1989. Molecular cloning a laboratory manual Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory] продуктом реакции лигирования трансформировали компетентные клетки E.coli Dh5α (50 мкл компетентных клеток + 7 мкл лигазной смеси), После теплового шока клетки инкубировали в 1 мл среды LB при 37°С и затем высевали на агаризованную среду LB, содержащую 25 мкг/мкл канамицина, и инкубировали в течение ночи при 37°С.
Для выделения плазмид с чашки брали единичную колонию, инкубировали в 4 мл среды LB, содержащую 25 мкг/мкл канамицина, и инкубировали в течение ночи при 37°С. Полученные клетки осаждали на центрифуге при 5000 об/мин, 4°С 5 мин и из них выделяли плазмидную ДНК с помощью набора для выделения плазмидной ДНК «PureLink™ HiPure Plasmid Miniprep Kit». Выделение плазмид проводили в трех повторах.
Секвенирование полученных плазмид с помощью стандартных секвенирующих праймеров T7for и T7rev (Novagen) показало, что во всех плазмидах клонирована последовательность ДНК, соответствующая последовательности SEQ ID №3.
Полученная плазмида получила название pNhaDAAO (Фиг. 3).
Пример 3. Получение рекомбинантного штамма E.coli - продуцента NhaDAAO. Полученной плазмидой pNhaDAAO трансформировали штамм E. coli BL21(DE3)]F-ompTgaldcmlonhsdSB(rB-mB-)λ(DE3[lacIlacUV5-T7p07ind1sam7nin5]) [malB+]K-12(λS). Отбор клонов проводили на агаризованной среде LB, содержащую 25 мкг/мкл канамицина.
Для проверки эффективности продуцирования NhaDAAO полученным штаммом с чашки отбирали отдельные колонии и выращивали в 4 мл среды 2YT, содержащей 25 мкг/мкл канамицина, в течение ночи при 30°С. Затем отбирали 1 мл культуральной жидкости и переносили в колбу с отбойниками объемом 1 л со 100 мл среды 2YT, содержащей 25 мкг/мкл канамицина. Клетки культивировали при 30°С до поглощения на 600 нм А600 1 ед. поглощения, добавляли индуктор ИПТ до концентрации 0,1 мМ, снижали температуру до 20°С и культивировали в течение 48 часов. По окончании клетки осаждали центрифугированием при 6000 об/мин в течение 10 мин при 4°С. Супернатант удаляли, а биомассу ресуспендировали в 0,1 М фосфатном буфере рН 8.0 (10 мас %). Суспензию клеток 20 мл подвергали заморозке - разморозке и затем обрабатывали ультразвуком при постоянном охлаждении 4°С - 4 цикла 1 мин озвучивания + 4 минуты охлаждения. Клеточный дебрис осаждали центрифугированием при 15000 об/мин в течение 20 мин при 4°С и определяли активность в бесклеточном экстракте. Активность определяли в 0,1 М фосфатном буфере, рН 8,0, при 30°С пероксидазным методом в присутствии 10 мМ субстрата (см. пример 4). В качестве субстрата пероксидазы использовали о-дианизидин (10 мг/л). Культивирование проводили в трех повторах. Выход биомассы составил 15±2 г/л среды. Активность в бесклеточном экстракте с саркозином составила 0,32 ед. на л среды. Таким образом, полученный рекомбинантный штамм E.coli BL21(DE3)/pNhaDAAO экспрессирует NhaDAAO в активной форме. Полученный рекомбинантный штамм Escherichia coli BL21(DE3)/pNhaDAAO зарегистрирован во Всероссийской коллекции промышленных микроорганизмов как Escherichia coli BL21(DE3)/pNhaDAAO ВКПМ В-14556.
Пример 4. Субстратная специфичность NhaDAAO.
Были проведены тесты ферментативной активности NhaDAAO с различными субстратами. Активность определяли в 0,05 М фосфатном буфере, рН 8,0, при комнатной температуре пероксидазным методом в присутствии 10 мМ соответствующей аминокислоты. В качестве субстрата пероксидазы использовали о-дианизидин (10 мг/л).
Таблица 5
Субстратная специфичность NhaDAAO
*Приведенные результаты являются средним из трех измерений.
Из таблицы 5 видно, что NhaDAAO проявляет ферментативную активность со всеми изученными аминокислотами, однако наибольшую активность он проявляет с глицином и серином. Особо следует отметить, что фермент проявляет ферментативную активность как со свободными, так и и с N-замещенными формами аминокислот, что является уникальным случаем для оксидаз D-аминокислот. Подобные ферменты до настоящего времени в литературе не описаны.
Таким образом можно сделать вывод, что созданный нами рекомбинантный штамм Escherichia coli BL21(DE3)/pNhaDAAO ВКПМ В-14556 - продуцент NhaDAAO, позволяет получить фермент, который имеет большой биотехнологический и биоаналитический потенциал. Фактически NhaDAAO представляет универсальный биокатализатор, который можно использовать с большим количеством субстратов, в первую очередь с N-замещенными аминокислотами. Созданный штамм-продуцент - первый штамм, который экспрессирует DAAO из архей.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Плазмида
pNhaDAAO.xml" softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2023-12-27">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText></ApplicationNumberText>
<FilingDate></FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>107-2023</ApplicantFileReference>
<ApplicantName languageCode="ru">Федеральное государственное
бюджетное образовательное учреждение высшего образования
"Московский государственный университет имени
М.В.Ломоносова" (МГУ)</ApplicantName>
<ApplicantNameLatin>Federal state budgetary education institution of
higher education "Lomonosov Moscow State University"
(MSU)</ApplicantNameLatin>
<InventionTitle languageCode="ru">РЕКОМБИНАНТНАЯ ПЛАЗМИДА pNhaDAAO,
ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ОКСИДАЗЫ ОБЫЧНЫХ И N-ЗАМЕЩЕННЫХ D-АМИНОКИСЛОТ
(NhaDAAO) АРХЕЙ Natrarchaeobius halalkaliphilus AArcht4 (NhaDAAO) В
КЛЕТКАХ Escherichia coli И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli
BL21(DE3)/pNhaDAAO ВКПМ В-14556 - ПРОДУЦЕНТ NhaDAAO</InventionTitle>
<SequenceTotalQuantity>3</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>40</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..40</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gcaggttcaatacatatgcaaccagacataactgtcctcg</INSDSeq
_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>45</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..45</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cgtacactcgagctattattcgcatatatgtttggttagtgttcg</IN
SDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>1165</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1165</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tatgcaaccagacataactgtcctcggtgccggggtcaatggcacctcc
accgcgcttgcgcttacactcctcggatacaacacccagatagttgcagatcagttcgcctatgaggagc
aacatcgagaccctcgtttttcctccgcgtacggtgcaggtgccattcttccatactcggtcggcatgga
cagcctggatgaaaccttcgaggagagtcagacggtttttcggctgttagaggagttgagcgttcttggc
gtccggaagaacgaccactactggatcgacgaggaagggcgtggtccttcgaacgccccgaagtacctcg
acggattccgaaaagtcgaatcgccgaacgcccttcctcgacggacgggttcagaacacgtgacggactt
tactgccgagatgcttttcgccgatatgccgacgtacatggaagggttgtacaggttgtatcaggcggct
ggagggagtattcgaaagcgaaccgtgacacgcacagatctaaccgaaatgcccggactactgatcaact
gcactgggctacggagtccggaattgttcgaagacagcgcaccgtaccacgcggcacgtggccatctcgt
caccgcacagcgtgcaccgattccgaaactcgatggaacgatgatttcgtactcctactccgtcgcagga
gatcgaaaaggggtctactgcttcccccgaatggatgggatagtgatgggaggaacacatcagtcagtcc
cctaccggccggacgagaagccgtgtttcccacccctgaacgaaccgaccacgaaaatcggggggacgga
ggttccgaaacgcatcgtagaattgaacgccgaactcctcgctcaactgggtgtagacgtagagcgatct
gacctggaagtctcgatcggctaccgaccgctccgtgaccccgagggagacggcgtacgcgtcgaactgg
ccgaagaatcgtgtggaccggtcgtccacaactacggccacggtggtgcgggtatctcggtatcctgggg
gtgtgcgataagcgtagcacggctcgtccgtgaagcaattggtgacagggaagtcccccccgctccagtt
tcacgacctgcaccgctgcttccactctatcgaacactaaccaaacatatatgcgaataatagctc</IN
SDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---


Изобретение относится к области биотехнологии, в частности к генетической инженерии, и представляет собой рекомбинантную плазмидную ДНК pNhaDAAO, содержащую модифицированный ген Nhadaao, кодирующий оксидазу D-аминокислот архей Natrarchaeobius halalkaliphilus AArcht4 (NhaDAAO). Полученная плазмида обеспечивает экспрессию включенного в ее нуклеотидную последовательность модифицированного гена в клетках Escherichia coli. В результате трансформации штамма E. coli BL21(DE3) предложенной плазмидой получен новый рекомбинантный штамм-продуцент E. coli BL21(DE3))/pNhaDAAO ВКПМ В-14556, обеспечивающий экспрессию NhaDAAO в активной форме. Изобретение позволяет получать NhaDAAO в активной форме с высокой степенью эффективности. 2 н.п. ф-лы, 4 ил., 5 табл., 4 пр.
1. Рекомбинантный вектор pNhaDAAO для экспрессии оксидазы D-аминокислот архей Natrarchaeobius halalkaliphilus AArcht4 (NhaDAAO), содержащий NdeI-XhoI фрагмент размером 5230 п.о. вектора pET24a(+) и NdeI-XhoI фрагмент размером 1165 п.о., представляющий собой нуклеотидную последовательность SEQ ID № 3.
2. Рекомбинантный штамм Escherichia coli BL21(DE3)/pNhaDAAO – продуцент NhaDAAO, полученный путем трансформации штамма Escherichia. coli BL21(DE3) плазмидной ДНК по п.1, задепонированный во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под номером В-14556.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pNhyDAAO, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ОКСИДАЗЫ D-АМИНОКИСЛОТ БАКТЕРИЙ Natronosporangium hydrolyticum ACPA39 (NhyDAAO) В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli BL21(DE3)/pNhyDAAO ВКПМ В-14337 - ПРОДУЦЕНТ NhyDAAO | 2022 |
|
RU2803295C1 |
| МУТАНТНАЯ ОКСИДАЗА D-АМИНОКИСЛОТ | 2009 |
|
RU2412998C1 |
| МУТАНТНАЯ ОКСИДАЗА D-АМИНОКИСЛОТ (ВАРИАНТЫ) | 2012 |
|
RU2507262C1 |
| РЕКОМБИНАНТНАЯ ДНК, КОДИРУЮЩАЯ ФУНКЦИОНАЛЬНО АКТИВНЫЙ ГИБРИДНЫЙ БЕЛОК ОКСИДАЗЫ D-АМИНОКИСЛОТ С ХИТИНСВЯЗЫВАЮЩИМ ДОМЕНОМ (DAOcbd) РЕКОМБИНАНТНАЯ ПЛАЗМИДА pVR1, ОБЕСПЕЧИВАЮЩАЯ ЕГО СИНТЕЗ В КЛЕТКАХ Escherichia coli, И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli С 41(DE3)/pVR1 - ПРОДУЦЕНТ DAOcbd | 2006 |
|
RU2310688C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pETTvDAO2, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ОКСИДАЗЫ D-АМИНОКИСЛОТ (DAO) ДРОЖЖЕЙ Trigonopsis variabilis В КЛЕТКАХ Escherichia coli И РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli C41(DE3)/pETTvDAO2 - ПРОДУЦЕНТ DAO | 2006 |
|
RU2310687C1 |
| Tishkov VI, Khoronenkova SV | |||
| D-Amino acid oxidase: structure, catalytic mechanism, and practical application | |||
| Biochemistry (Mosc) | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| PMID: 15701048. | |||

