Изобретение относится к биологии и медицине, а именно к способам исследования влияния окружающей и внутренней среды организма на здоровье человека и животных, и может быть использовано для определения у них биологического возраста, скорости старения, прогнозирования долголетия индивидов при различных состояниях организма и управления этими показателями жизнедеятельности.
Известно, что живые системы являются открытыми термодинамическими системами и характеризуются сложной упорядоченной структурой. Уровни их организации значительно выше, чем в неживой природе. Для сохранения и увеличения своей высокой упорядоченности живые системы в меру свойственной им открытости (в том числе на организменном уровне) непрерывно обмениваются энергией, веществом и информацией с внешней средой и при этом выполняют работу по уменьшению энтропии (рассеивания энергии в окружающую среду), неизбежно нарастающей за счет потерь в связи с теплообменом, броуновским движением и старением молекул, и т.д. [Николис Г., Пригожий И. Познание сложного. М., 1990. - С.293]. Процесс этого обмена называется метаболизмом. Известно, что предпочтительным является метаболизм с минимальным уровнем энтропии, так как именно он обеспечивает работу системы с максимальной экономией потерь и устойчивостью во внешней среде [Пригожий И. От существующего к возникающему. - М., 1985. - 113 с.; Пригожий И. Введение в термодинамику необратимых процессов. Пер. с англ. М., 1960; Франк Г.М., Кузин A.M. О сущности жизни. - М., 1964. - 350 с.]. На этой основе нами выдвигается гипотеза о том, что чем выше уровень метаболизма в живой системе, то есть, чем более интенсивно она обменивается энергией, веществом и информацией с внешней средой, тем эта система вынуждена совершать большую работу по поддержанию гомеостаза для сохранения минимального уровня энтропии, нести в связи с этим более значительные потери, стать более открытой к окружающей среде, а следовательно, и уязвимой к ее неблагоприятным воздействиям. Следуя этой гипотезе, уровень открытости живой системы можно рассматривать как показатель качества ее физиологического состояния, имеющий обратную связь с характеристиками этого качества - здоровьем, работоспособностью, продолжительностью жизни. Следует отметить, что другие авторы [Фролов В.А., Моисеева Т.Ю. Живой организм как информационно-термодинамическая система. - Вестник РУДН, 1999, №1. - С.6-14] тоже рассматривают открытость живой системы в связи с продолжительностью ее жизни на этапе эволюции к закрытой термодинамической системе. Таким образом, метаболизм, энтропия, открытость живой системы к окружающей воздушной среде могут не только характеризовать качество происходящих в этой системе процессов жизнеобеспечения, но и быть его первопричиной. Самому понятию открытости живой системы к окружающей среде можно дать следующее определение: открытость живой системы - это присущая ей развитость универсального свойства целесообразно жизнеобеспечивающего взаимодействия с окружающей средой.
В связи с вышеизложенным нами поставлена задача по разработке способа определения энтропии в организме человека или животного для получения возможности управлять процессами жизнеобеспечения.
Энтропия в организме человека или животного может характеризоваться кинетикой O2 на этапах его перемещения из атмосферы в организм, которая зависит от содержания О2 во вдыхаемом воздухе и в воздухе, содержащемся в альвеолах легких (альвеолярном), времени полного насыщения эритроцита кислородом в легких, времени, предоставленного эритроциту для отдачи полученного в легких О2 клеткам организма, и силы связи эритроцитарного гемоглобина с О2.
Известно, что содержание О2 во вдыхаемом воздухе зависит от его содержания в зоне дыхания. Естественное содержание O2 в воздухе открытых пространств выше, чем в замкнутых объемах и равно в среднем 20,9%. Содержание O2 в альвеолярном воздухе относится к числу индивидуальных гомеостатических констант и (при прочих равных условиях: возраста, устойчивости к недостатку кислорода и др.) находится во взаимодействии с показателями работоспособности и общего состояния здоровья организма [Сиротинин Н.Н., 1971; Евгеньева Л.Я., 1974; Карпман В.Л., Любина Б.Г., 1982; Меерсон Ф.З., 1981, и др.].
Известно, что продолжительность пребывания эритроцитов в легочных капиллярах зависит от скорости легочного кровотока и составляет 0,25-0,75 с. Этого времени достаточно для оксигенации крови, поскольку в норме эритроцит полностью насыщается O2 за 0,25 с [Зайко Н.Н., Быць Ю.В., Атаман А.В. и др. Патологическая физиология (Учебник для студентов мед. вузов). - К "Логос", 1996]. Таким образом, время полного насыщения эритроцита кислородом в легких, равное 0,25 с, характеризует период или фазу эффективного (прямого или открытого) контакта эритроцита с O2 альвеолярного воздуха. Известно, что время отдачи эритроцитом полученного в легких кислорода клеткам организма до следующего прохода эритроцита через легкие для насыщения кислородом характеризует период или фазу неэффективного (непрямого или закрытого) контакта эритроцита циркулирующей крови с O2 альвеолярного воздуха. Длительность этого периода (фазы) значительно превышает длительность прямого контакта эритроцита циркулирующей крови с O2 альвеолярного воздуха и зависит от скорости кровообращения или времени (Т) полного оборота циркулирующей крови в организме, на который (при прочих равных условиях) влияет частота сердечных сокращений (ЧСС) [Бабский Е.Б., Зубков А.А., Косицкий Г.И., Ходоров Б.И. Физиология человека. - М.: Медицина, 1966. - С.117]. Например, у взрослого человека в норме при ЧСС 75 уд./мин (состояние мышечного покоя) Т равен в среднем 21,5 с. С учетом известных возрастных, половых и межвидовых различий в величине отношения массы сердца к массе тела [Жеденов В.Н. Легкие и сердце животных и человека. 2 изд. М., 1961. - 478 с.] величину Т при различных ЧСС у животных и человека можно определить по следующему математическому выражению:
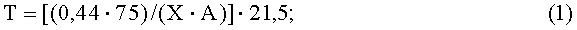
Т - время полного оборота эритроцита с током циркулирующей крови в организме (время полного оборота циркулирующей крови у исследуемого животного и человека, за которое циркулирующая кровь совершает полный оборот в сумме малого и большого кругов кровообращения), с;
0,44 - среднестатистическая относительная масса сердца человека (по отношению к общей массе тела), для которой характерно время полного оборота крови за 21,5 с при частоте сердечных сокращений 75 уд./мин, %;
75 - частота сердечных сокращений (ЧСС), при которой время полного оборота циркулирующей крови у человека совершается в среднем за 21,5 с, уд./мин;
21,5 - время полного оборота циркулирующей крови у человека при ЧСС 75 уд./мин, с;
Х - фактическая или (при невозможности измерения) характерная для человека и исследуемого вида животного среднестатистическая относительная масса сердца, %; (по данным [Жеденов В.Н. Легкие и сердце животных и человека. 2 изд. М., 1961. - 478 с.] масса сердца от общей массы тела в среднем равна 1/215 у мужчин и 1/250 - у женщин);
А - ЧСС фактическая, измеренная в момент исследования индивида, уд/мин.
Известно [Эккерт Р., Рэнделл Д., Огастин Дж. Физиология животных. Т.2. М., 1992], что сила связи эритроцитарного гемоглобина с O2 или устойчивость оксигемоглобина к диссоциации при прочих равных условиях зависит от водородного показателя (рН) крови, который, например, с ростом в ней напряжения CO2 снижается и, тем самым, уменьшает силу связи гемоглобина с O2 (сродство гемоглобина к O2), что способствует высвобождению O2 в плазму крови и поступлению оттуда в окружающие ткани. Известно также, что между изменением концентраций CO2 и O2 в организме имеет место реципрокная (взаимно обратная) связь. Поэтому, если содержание CO2 в любой части организма закономерно влияет на силу связи гемоглобина с O2, то влияние этой силы на дальнейшее продвижение O2 в структуры организма можно учитывать величиной концентрации альвеолярного O2.
Однако взятые в отдельности, указанные физиологические показатели, влияющие на взаимодействие атмосферного O2 со структурами организма (фазы прямого и непрямого контактов эритроцита циркулирующей крови с альвеолярным O2 в легких и его концентрация), не могут в полной мере характеризовать его энтропию, поскольку в этом случае не учитывается их сочетанное влияние на метаболические процессы.
Задачей изобретения является определение энтропии в организме человека или животного по взаимодействию фаз прямого и непрямого контактов эритроцита циркулирующей крови с альвеолярным O2 в легких и его концентрацией.
Указанная задача решается в заявляемом способе определения энтропии в организме человека или животного, заключающемся в учете времени прямого контакта эритроцита циркулирующей крови с альвеолярным O2, равного 0,25 с, определении времени полного оборота эритроцита с током циркулирующей крови в организме при фактическом числе ударов сердца за минуту по величине отношения произведения выраженной в процентах среднестатистической относительной массы сердца человека, равной 0,44, на выраженное в ударах сердца за минуту число 75 к произведению выраженной в процентах относительной массы сердца исследуемого индивида на число имеющихся у него в момент исследования фактических ударов сердца за минуту, умноженного на выраженное в секундах время полного оборота эритроцита с током циркулирующей крови, равное числу 21,5 при 75 сокращениях сердца за минуту, измерении выраженного в процентах содержания O2 в альвеолярном воздухе, и отличающемся тем, что энтропию в организме человека или животного определяют по величине, получаемой от произведения отношения времени прямого контакта эритроцита циркулирующей крови с альвеолярным O2 к времени полного оборота эритроцита с током циркулирующей крови в организме при фактическом числе ударов сердца за минуту на выраженное в процентах содержание O2 в альвеолярном воздухе.

где α - энтропия в организме человека или животного, %;
0,25 - число, соответствующее времени полного насыщения эритроцита циркулирующей крови в организме кислородом, с;
Т - время полного оборота эритроцита с током циркулирующей крови в организме, с;
Co2 - содержание кислорода в альвеолярном воздухе легких, %.
Предлагаемый способ определения энтропии в организме человека или животного основан на учете того, что с ростом частоты сердечных сокращений (ЧСС) суммарная (за определенное время) длительность прямых контактов эритроцита циркулирующей крови с кислородом альвеолярного воздуха увеличивается, а непрямых контактов - уменьшается, что сопровождается увеличением обмена веществ в организме и приростом необратимого рассеяния свободной энергии в окружающую среду. Так у человека (например, за 10 минут) суммарная длительность прямых контактов эритроцита с О2 альвеолярного воздуха при ЧСС 75 уд./мин (Т=21,5 с) составляет 7 с (то есть 600 с/21,5 с=27,9 оборотов циркулирующей крови; 27,9·0,25 с≈7 с), при ЧСС 100 уд./мин (Т=16,1 с) - 9,3 с, а при ЧСС 180 уд./мин (Т=8,96 с) - 16,7 с. Вместе с тем, за это же время суммарная длительность непрямых контактов эритроцита циркулирующей крови с кислородом альвеолярного воздуха при ЧСС 75 уд./мин составляет 593 с [то есть 600 с/21,5 с=27,9 оборотов циркулирующей крови; 27,9·(21,5 с-0,25 с)=593 с], при ЧСС 100 уд./мин - 591 с, а при ЧСС 180 уд./мин - 583 с. Таким образом, в предлагаемом способе открытость организма атмосфере, метаболизм и энтропия увеличиваются с ростом ЧСС за счет увеличения фазы прямого контакта эритроцита с атмосферой (альвеолярный воздух-атмосфера) в единицу времени и сокращения противоположной фазы без газообмена с атмосферой.
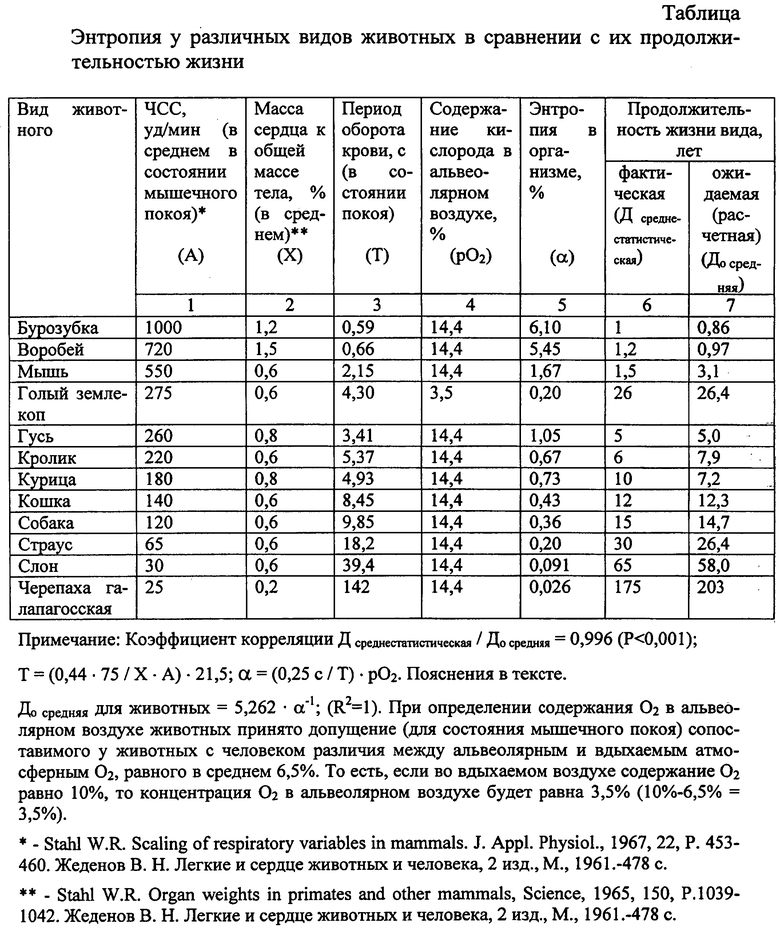
В таблице приведены примеры определения энтропии (α) у 12 различных видов животных, которая сопоставлялась с имеющимися в литературе сведениями о среднестатистической продолжительности жизни (Дсреднестатистическая) вида этих животных. На основе приведенных данных получено следующее уравнение степенной регрессии, характеризующее зависимость между α и среднестатистической продолжительностью жизни (Дсреднестатистическая):
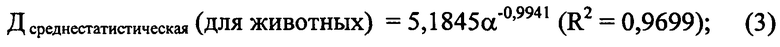
где 5,1845 - эмпирический коэффициент;
R2 - величина достоверности аппроксимации между Дсреднестатистическая и α.
В целях упрощения математического выражения 3 нами разработана формула 4 с коэффициентом корреляции r Дсреднестатистическая/До средняя=0,996; Р<0,001:
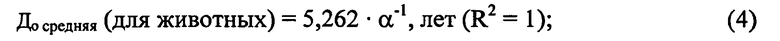
где До средняя - ожидаемая средняя продолжительность жизни;
5,262 - эмпирический коэффициент;
R2 - величина достоверности аппроксимации между До средняя и α.
Полученная зависимость продолжительности жизни вида животного от энтропии в организме позволяет объяснить считающееся парадоксальным долгожительство грызуна «Голый землекоп» (Heterocephalus glaber) исключительно обитанием этого млекопитающего в трудно проветриваемых подземных условиях (туннели диаметром 2-4 см, глубиной до 2 м, длиной до 5 км) с предельно низким содержанием O2 во вдыхаемом воздухе от 8 до 12% (в среднем 10%) и смертельной для многих других животных концентрацией CO2 (10%). Имеются данные о содержании на поверхности кожи и слизистых оболочках у этих грызунов высоких концентраций углекислоты [Шиндер А. Животное, не чувствующее боли // Еженедельник 2000. - 27.06-03.07.2008. №26 (420)], которые у других видов животных не наблюдаются. Указанные условия существования голого землекопа приводят к крайне низким концентрациям O2 в альвеолах легких (3,5%) и по представленным в таблице данным более чем в 8 раз уменьшают энтропию в сравнении с другими равными по массе грызунами, что, по-видимому, и приводит к существенному (более чем в 15 раз) увеличению длительности жизни особей этого их вида. В доступной нам литературе указанный феномен долгожительства Heterocephalus glaber объясняется с позиций генетики приобретенным особым свойством его организма, но это еще не характеризует саму первопричину (внешнюю причину) образования и закрепления этого свойства у данного вида грызуна. Из полученных результатов следует, что (при прочих равных условиях) продолжительность жизни организма есть, скорее всего, средневзвешенная величина, определяемая длительностью его состояний в процессе онтогенеза, характеризуемых интенсивностью взаимодействия эритроцитов циркулирующей крови с атмосферным кислородом.
Однако на основании анализа литературы (Гаврилов Л.А., Гаврилова Н.С. Биология продолжительности жизни М.: Наука. 1991. - 280 с.) следует считать неверным перенесение закономерностей животного мира на понимание проблем человеческого долголетия, детерминируемого, прежде всего, социально-экономическими факторами (уровнем медицинского обеспечения, безопасностью труда и эффективностью отдыха, материальной обеспеченностью и духовным комфортом). Поскольку социально-экономические условия жизни Homo sapiens существенно изменились за время его эволюции, то измерение ожидаемой продолжительности жизни современного человека с применением выявленной и отраженной в формуле 4 закономерности нуждается в дополнении, учитывающем влияние этих условий на долголетие.
Средняя продолжительность жизни человека в Палеолите (2,6 миллиона лет назад), когда условия его жизни мало отличались от животных, была равна 31 году [Бужилова А.П. К вопросу о семантике коллективных захоронений в эпоху палеолита. В кн.: Этиология человека и смежные дисциплины. Современные методы исследований. Под ред. Бутовской, М.: Ин-т этиологии и антропологии, 2004. С.21-35], что соответствует результату, получаемому для человекообразных обезьян, например для мужской особи гориллы:
α (для гориллы)=(0,25 с/21,5 с)·14,4%=0,167%;
До средняя=5,262·0,167-1=31,5 года.
Принимая во внимание расчеты Б.Ц. Урланиса [Урланис Б.Ц. Увеличение продолжительности жизни в СССР // Социальные исследования: Сб. - М.: Наука, 1965. - С.151, 153; Урланис Б.Ц. Этюд о возрасте // Неделя. - 1966. - №40], в которых он на примере наиболее передовых и благополучных стран статистически доказывает, что видовая или характерная для человека, как одного из видов живых существ, биологическая продолжительность жизни (обозначенная автором как нормальная) должна составлять 90 лет, мы скорректировали формулу 4, преобразовав ее в формулу 5, учитывающую дополнительные 58 лет, которые, по-нашему мнению, должны проживать мужчины и женщины в нормальных социально-экономических условиях труда и быта. Так, например, если учесть, что у взрослого человека концентрация O2 в альвеолярном воздухе в норме равна 14,4% [Бабский Е.Б., Зубков А.А., Косицкий Г.И., Ходоров Б.И. Физиология человека. - М.: Медицина, 1966. - С.117, 143], то (при характерной для мужчин в состоянии мышечного покоя средней ЧСС 72 уд./мин и массе сердца 1/215 от общей массы тела) период полного оборота циркулирующей крови в организме равен 21,4 с, α и До средняя составляют:
α=(0,25 с/21,4 с)·14,4%=0,168%;
До средняя=5,262·0,168-1=31,3 года.
В результате, вклад нормальных социально-экономических условий в ожидаемую продолжительность жизни для мужчин составляет: 90 лет - 31,3 года=58,7 лет.
При характерной для женщин в состоянии мышечного покоя средней ЧСС 78 уд./мин и массе сердца 1/250 от общей массы тела период полного оборота циркулирующей крови в организме равен 22,7 с, α и До средняя составляют:
α=(0,25 с/22,7 с)·14,4%=0,158%;
До средняя=5,262·0,158-1=33,3 лет.
В результате вклад нормальных социально-экономических условий в ожидаемую продолжительность жизни для женщин составляет: 90 лет - 33,3 года=56,7 лет.
На основе этих полученных данных нами, как отмечено выше, принята усредненная для мужчин и женщин величина вклада нормальных социально-экономических условий в ожидаемую продолжительность жизни, равная 58 лет.
Известно, что в отличие от нормальных социально-экономических условий, обеспечивающих человеку видовую (нормальную) продолжительность жизни, относящиеся к исследуемым региону и временному периоду проживания реальные социально-экономические условия формируют среднюю продолжительность жизни. Например, если средняя продолжительность жизни в России в 2011 году (по данным Росстата) составляла 64,3 лет для мужчин и 76,1 лет для женщин, то обусловленный в этот период времени вклад имеющихся (в 2011 году) социально-экономических условий в ожидаемую продолжительность жизни россиянина составлял:
64,3 лет-31,3 лет=33,0 лет (для мужчин);
76,1 лет-33,3 лет=42,8 лет (для женщин).
В формулировках нормальная и средняя ожидаемые продолжительности жизни смысловое содержание выражений «нормальная и средняя» учитывает, прежде всего, социально-экономические условия жизни (нормальные - характеризуют условия, близкие к идеальным, в наибольшей мере способствующим достижению видовой, биологической продолжительности жизни, средние - отражают реальные условия в регионе в данный период времени проживания). С учетом вышесказанного, ожидаемая продолжительность предстоящей жизни человека (До) должна рассчитываться с применением следующего математического выражения:

где А - ожидаемое количество лет прожития за счет социально-экономических условий (при условиях, близких к идеальным, обозначаемых нормальными, - 58 лет; при других условиях - количество лет, получаемых в результате вычитания из известных статистических данных о средней продолжительности жизни в регионе в данный период времени проживания 31,3 лет для мужчин и 33,3 лет для женщин). Обозначение остальных символов приведено выше.
Выдающийся современный геронтолог академик Д.Ф. Чеботарев указывает на то, что видовая продолжительность жизни должна служить реальным ориентиром для увеличения средней продолжительности жизни. Разница между этими величинами представляет резерв, который вполне может быть освоен за счет улучшения условий и образа жизни. Тактической задачей геронтологии он считает борьбу с преждевременным старением и хотя бы частичное освоение тех резервов, которые, безусловно, есть у человека и которые определяются неиспользованным периодом между современной средней и видовой продолжительностью жизни, сохранение практического здоровья в течение всего периода так называемого третьего возраста (от 60 до 90 лет). Стратегической задачей он считает продление активного долголетия сверх сроков видовой длительности жизни человека [Чеботарев Д.Ф. Физиологические механизмы старения. Л.: Наука, 1982. - 228 с.]. В формуле, определяющей конечные цели геронтологии «Добавлять не только годы к жизни, но и жизнь к годам», - воплощены и тактические, и стратегические задачи этой науки, объединены и медицинские, и социальные проблемы старения. Поэтому разработку средств, позволяющих оценивать развитие таких резервов организма, которые работают на достижение активного долголетия с преодолением нормальной продолжительности жизни, следует рассматривать как одну из важных первичных ступеней на пути решения комплексной проблемы старения. В этой связи считаем, что разработанный нами способ определения открытости организмов человека и животных атмосфере является важным средством для успешного разрешения указанной проблемы, поскольку дает возможность, например, выявлять и априорно оценивать развитие резерва долголетия организма на этапах онтогенеза и при различных функциональных состояниях выявлять сходство и различие в формировании этого резерва у человека и животных.
Приведем примеры использования заявляемого способа на людях и некоторых животных, находящихся в различных функциональных состояниях (мышечный покой, физическая нагрузка, нарушение сердечно-сосудистой и дыхательной систем, период новорожденности и грудной возраст постнатального онтогенеза).
У мужчины при выполнении работы средней тяжести ЧСС равна 100 уд./мин, концентрация O2 в альвеолярном воздухе, измеренная газоанализатором ПГА-12 в последних порциях выдыхаемого воздуха, поддерживается на уровне 14,4%. Следовательно энтропия в организме человека при выполнении работы средней тяжести составляет:
α=(0,25 с/15,4 с)·14,4%=0,23%.
При такой величине энтропии нормальная и средняя ожидаемые в 2011 году продолжительности жизни могут составлять:
До нормальная=(5,262·0,23-1)+58 лет=80,9 лет;
До средняя=(5,262·0,23-1)+33,0 лет=55,9 лет.
У мужчины с нарушением сердечно-сосудистой и дыхательной систем ЧСС в состоянии мышечного покоя равна 95 уд./мин, при выполнении работы средней тяжести - 130 уд./мин, концентрация O2 в альвеолярном воздухе, измеренная газоанализатором ПГА-12 в указанных состояниях, равна 16,1%. Следовательно энтропия в организме будет составлять:
- (в состоянии мышечного покоя) α1=0,25 с/16,2 с·16,1%=0,25%;
- (в состоянии выполнения работы средней тяжести) α2=0,25 с/11,9 с·16,1%=0,34%.
Нормальная и средняя ожидаемые продолжительности жизни мужчины с нарушениями сердечно-сосудистой и дыхательной систем будет составлять:
До1=(5,262·0,25-1)+58 лет=79,0 лет (нормальная в состоянии мышечного покоя);
До2=(5,262·0,34-1)+58 лет=73,5 лет (нормальная в состоянии выполнения работы средней тяжести);
До1=(5,262·0,25-1)+33,0 лет=54,0 лет (средняя в состоянии мышечного покоя);
До2=(5,262·0,34-1)+33,0 лет=48,5 лет (средняя в состоянии выполнения работы средней тяжести).
У новорожденного мальчика ЧСС равна 150 уд./мин, масса сердца в общей массе тела равна 0,89%, концентрация O2 в альвеолярном воздухе составляет 17,8%. Через 1/2 года и через год ЧСС и содержание O2 в альвеолярном воздухе ребенка уменьшились до 130 и 120 уд./мин, 17,3 и 17,2% соответственно. Следовательно, энтропия в организме составляет:
- у новорожденного α=0,25 с/5,31 с·17,8%=0,84%,
- через 1/2 года после рождения α=0,25 с/6,13 с·17,3%=0,70%,
- через год после рождения α=0,25 с/6,64 с·17,2%=0,65%.
Нормальная ожидаемая продолжительность жизни, измеренная при указанных функциональных состояниях организма, будет равна:
- у новорожденного До=(5,262·0,84-1)+58 лет=64,3 года
- через 1/2 года после рождения До=(5,262·0,70-1)+58 лет=65,5 лет
- через год после рождения До=(5,262·0,65-1)+58 лет=66,1 лет.
Средняя ожидаемая продолжительность жизни будет равна:
- у новорожденного До=(5,262·0,84-1)+33,0 лет=39,3 лет
- через 1/2 года после рождения До=(5,262·0,70-1)+33,0 лет=40,5 лет
- через год после рождения До=(5,262·0,65-1)+33,0 лет=41,1 лет.
Выявленные различия в величине энтропии в организме при указанных его состояниях согласуются с риском нарушения здоровья, которому в большей мере подвергаются новорожденные, видимо, в связи с недостаточно сформированными механизмами обмена веществ. В частности, в пересчете на массу тела младенцы и дети младшего возраста пьют больше воды, потребляют больше пищи и вдыхают больше воздуха, чем взрослые [Дьяченко В.Г., Рзянкина М.Ф., Солохина Л.В. Руководство по социальной педиатрии: учебное пособие / В.Г. Дьяченко, М.Ф Рзянкина, Л.В. Солохина / Под ред. В.Г. Дьяченко. - Хабаровск: Изд-во Дальневосточ. гос. мед. ун-та. - 2012. - 322 с.]. Указанные результаты апробации заявляемого способа согласуются с данными литературы о том, что биологический возраст организма не есть величина постоянная, он изменяется при различных состояниях, обусловленных возрастом, физической активностью, здоровьем, психоэмоциональным стрессом и др. факторами [Позднякова Н.М., Прощаев К.И., Ильницкий А.Н., Павлова Т.В., Башук В.В. Современные взгляды на возможности оценки биологического возраста в клинической практике // Фундаментальные исследования. - 2011. - №2 - С.17-22].
У домового воробья ЧСС в состоянии мышечного покоя равна 460 уд./мин, а в полете - 950 уд./мин (этот вид животных имеет среднюю продолжительность жизни 1,2 лет и относительную массу сердца 1,5%; [Жеденов В.Н. Легкие и сердце животных и человека. 2 изд. М., 1961. - 478 с.]), концентрация О2 в альвеолярном воздухе 14,4%. Следовательно, энтропия в организме домового воробья при этих состояниях будет равна:
- (в состоянии мышечного покоя) α1=(0,25 с/1,03 с)·14,4%=3,49%;
- (при полете) α2=(0,25 с/0,50 с)·14,4%=7,20%.
Средняя ожидаемая продолжительность жизни этого воробья будет равна:
- (в состоянии мышечного покоя) До=(5,262·3,49-1)=1,5 лет;
- (при полете) До=(5,262·7,20-1)=0,73 лет.
Из примеров использования заявляемого способа следует, что с ростом энтропии в организме человека или животного нормальная и средняя ожидаемые продолжительности жизни индивидов сокращаются и наоборот. Полученные результаты применения заявляемого способа согласуются с известными результатами физиологических исследований [Маршак М.Е. Физиологическое значение углекислоты. - М.: Медицина, 1969. - 145 с.; Агаджанян Н.А., Елфимов А.И. Функции организма в условиях гипоксии и гиперкапнии. М.: Медицина, 1986. - 272 с.; Агаджанян Н.А., Катков А.Ю. Резервы нашего организма. М.: Знание, 1990. - 240 с.], в которых установлено влияние тренировок организма к недостатку O2 и избытку CO2 на укрепление здоровья, повышение работоспособности и увеличение продолжительности жизни. Поскольку в исследованиях этих авторов достоверно установлено, что тренировки к недостатку O2 и избытку CO2 снижают ЧСС, частоту и глубину легочного дыхания, содержание O2 в альвеолярном воздухе, то указанное благотворное влияние таких тренировок на организм можно объяснить достигнутым уменьшением его открытости атмосфере и необратимого рассеяния свободной энергии в окружающую среду.
Так, при систематических тренировках с волевыми задержками легочного дыхания и вдыханием гипоксических-гиперкапнических воздушных смесей с содержанием O2 15-9% и CO2 5-11% альвеолярный воздух содержит O2 8,5; 7,5%. В результате (при ЧСС, например, 50 уд./мин) Т=32,25 с; α=0,0659%; 0,0581%. Тогда нормальная ожидаемая продолжительность жизни будет равна:
До=(5,262·0,0659-1)+58 лет=138 лет;
До1=(5,262·0,0581-1)+58 лет=149 лет.
Средняя ожидаемая продолжительность жизни для мужчин будет равна:
До=(5,262·0,0659-1)+33,0 лет=113 лет;
До1=(5,262·0,0581-1)+33,0 лет=124 лет.
Таким образом, в заявляемом способе определения энтропии в организме человека или животного решена задача изобретения: энтропия в организме человека или животного определяется по взаимодействию фаз контакта эритроцита циркулирующей крови с альвеолярным O2 в легких и его концентрацией.
ЛИТЕРАТУРА
1. Агаджанян Н.А., Елфимов А.И. Функции организма в условиях гипоксии и гиперкапнии. М.: Медицина, 1986. - 272 с.
2. Агаджанян Н.А., Катков А.Ю. Резервы нашего организма. М.: Знание, 1990. - 240 с.
3. Бабский Е.Б., Зубков А.А., Косицкий Г.И., Ходоров Б.И. Физиология человека. - М.: Медицина, 1966. - С.117, 143.
4. Бужилова А.П. К вопросу о семантике коллективных захоронений в эпоху палеолита. В кн.: Этиология человека и смежные дисциплины. Современные методы исследований. Под ред. Бутовской, М.: Ин-т этиологии и антропологии, 2004. - С.21-35.
5. Гаврилов Л.А., Гаврилова Н.С. Биология продолжительности жизни. М.: Наука, 1991. - 280 с.
6. Дьяченко В.Г., Рзянкина М.Ф., Солохина Л.В. Руководство по социальной педиатрии: учебное пособие / В.Г. Дьяченко, М.Ф Рзянкина, Л.В. Солохина / Под ред. В.Г. Дьяченко. - Хабаровск: Изд-во Дальнево-сточ. гос. мед. ун-та, 2012. - 322 с.
7. Евгеньева Л.Я. Дыхание спортсмена.- Киев, Здоров'я, 1974. - 101 с.
8. Жеденов В.Н. Легкие и сердце животных и человека. 2 изд. М., 1961. - 478 с.
9. Зайко Н.Н., Быць Ю.В., Атаман А.В. и др. Патологическая физиология (Учебник для студентов мед. вузов). - К "Логос", 1996.
10. Карпман В.Л., Любина Б.Г. Динамика кровообращения у спортсменов. М.: Физкультура и спорт, 1982. - 135 с.
11. Маршак М.Е. Физиологическое значение углекислоты. - М.: Медицина, 1969. - 145 с.
12. Меерсон Ф.З. Адаптация, стресс и профилактика. М., 1981.
13. Николис Г., Пригожий И. Познание сложного. М., 1990. - С.293.
14. Позднякова Н.М., Прощаев К.И., Ильницкий А.Н., Павлова Т.В., Башук В.В. Современные взгляды на возможности оценки биологического возраста в клинической практике // Фундаментальные исследования, 2011. - №2 - С.17-22.
15. Пригожий И.Р. Введение в термодинамику необратимых процессов. Пер. с англ. М., 1960.
16. Пригожий И. От существующего к возникающему. - М., 1985. - 113 с.
17. Сиротинин Н.Н. Регуляция дыхания и физиологическое приспособление дыхательной функции при гипоксии // Физиол. жив. СССР, 1971. - Т.7. - №12.
18. Урланис Б.Ц. Увеличение продолжительности жизни в СССР // Социальные исследования: Сб. - М.: Наука, 1965. - С.151, 153.
19. Урланис Б.Ц. Этюд о возрасте // Неделя, 1966. - №40.
20. Франк Г.М., Кузин A.M. О сущности жизни. - М.,1964. - 350 с.
21. Чеботарев Д.Ф. Физиологические механизмы старения. Л.: Наука, 1982. - 228 с.
22. Шиндер А. Животное, не чувствующее боли // Еженедельник 2000.-27.06-03.07.2008. №26 (420).
23. Эккерт Р., Рэнделл Д., Огастин Дж. Физиология животных. Т.2. М., 1992.
24. Stahl W.R. Organ weights in primates and other mammals, Science, 1965, 150, P.1039-1042.
25. Stahl W.R. Scaling of respiratory variables in mammals. J. Appl. Physiol., 1967, 22, P.453-460.
Изобретение относится к биологии и медицине, а именно к изучению влияния окружающей и внутренней среды организма на здоровье человека или животного. Способ касается исследования энтропии в организме. Для этого определяют относительную по отношению к массе тела массу сердца в % (X), число сердечных сокращений (А) и содержание кислорода в альвеолярном воздухе легких в % (Со2). Расчет проводят по формуле: α=(0,25/Т)·Co2, где α - энтропия в %, Т - время полного оборота эритроцита с током циркулирующей крови в сек, при этом Т=[(0,44·75)/(X·А)]·21,5. Способ позволяет измерять основную объединяющую живые системы характеристику организма, что может быть использовано для определения биологического возраста, состояния здоровья, для изучения влияния различных средств профилактики нарушений здоровья и продления жизни. 1 табл.
Способ определения энтропии в организме человека или животного, отличающийся тем, что определяют относительную по отношению к массе тела массу сердца в % (X), число сердечных сокращений (А) и содержание кислорода в альвеолярном воздухе легких в % (Со2) и расчет проводят по формуле: α=(0,25/Т)·Co2, где α - энтропия в %, Т - время полного оборота эритроцита с током циркулирующей крови в сек, при этом Т=[(0,44·75)/(X·А)]·21,5.
| СПОСОБ ИССЛЕДОВАНИЯ АКТИВНОСТИ МЕТАБОЛИЧЕСКИХ ПРОЦЕССОВ В КЛЕТКЕ | 1992 |
|
RU2050001C1 |
| СПОСОБ ДИАГНОСТИКИ НАРУШЕНИЙ МЕТАБОЛИЗМА В ОРГАНИЗМЕ В УСЛОВИЯХ ОКИСЛИТЕЛЬНОГО СТРЕССА | 2010 |
|
RU2436101C1 |
| СПОСОБ ОЦЕНКИ МЕТАБОЛИЗМА И СИСТЕМЫ ТРАНСПОРТА КИСЛОРОДА К ГОЛОВНОМУ МОЗГУ У БОЛЬНЫХ С ТЯЖЕЛОЙ ЧЕРЕПНО-МОЗГОВОЙ ТРАВМОЙ | 2004 |
|
RU2264162C2 |
| JP 4829496 B2, 07.12.2011 | |||
| УСТРОЙСТВО ДЛЯ УСПОКОЕНИЯ КОЛЕБАНИЙ УПРУГОГО ЭЛЕМЕНТА ПЕРЕМЕННОЙ ЖЕСТКОСТИ | 2003 |
|
RU2241251C1 |
| СКОБЕЛЕВ Е | |||
| И., Функциональная активность системы транспорта кислорода и гемодилюция | |||
| IY Всесоюзный съезд анестезиологов и реаниматологов, Одесса, 1989, C | |||
| Катодный усилитель | 1923 |
|
SU492A1 |
| ТИЩЕНКО И | |||
| М | |||
| Роль функциональной активности | |||

