Область техники, к которой относится изобретение
Настоящее изобретение относится к веществу, используемому в качестве терапевтического и/или профилактического средства против патологического метаболизма костной ткани, и к способу лечения и/или профилактики патологического метаболизма костной ткани.
Предпосылки изобретения
Известно, что кость является динамическим органом с постоянной перестройкой путем повторяющегося формирования и резорбции, в связи с чем изменяется ее морфология и поддерживаются уровни кальция в крови. В здоровой кости поддерживается равновесие между формированием кости остеобластами и резорбцией кости остеокластами, и костная масса поддерживается постоянной. В отличие от этого, при потере равновесия между формированием кости и резорбцией кости, происходит патологический метаболизм костной ткани, такой как остеопороз (WO 07/093042 и Endocrinological Review, (1992) 13, pp. 66-80).
В качестве факторов, регулирующих метаболизм костной ткани, опубликовано множество системных гормонов и местных цитокинов, и эти факторы взаимодействуют друг с другом для формирования и поддержания кости (WO 07/093042 и Endocrinological Review, (1996) 17, pp. 308-332). Изменение костной ткани при старении и возникновение в связи с этим остеопороза, хорошо известно, но механизм его возникновения включает различные факторы, такие как уменьшение секреции половых гормонов и аномалии рецепторов гормонов; изменение местной экспрессии цитокинов в кости; экспрессия связанных со старением генов; и невозможность дифференцировки или дисфункция остеокластов или остеобластов, и таким образом, остеопороз трудно рассматривать как просто связанный с возрастом физиологический феномен. Первичный остеопороз в основном разделяют на постменопаузальный остеопороз, связанный с уменьшением секреции эстрогенов, и старческий остеопороз, связанный со старением, но прогрессирование фундаментальных исследований механизмов регуляции формирования кости и резорбции кости является необходимым для выяснения механизма его возникновения и для поиска лекарственных средств против него.
Остеокласты представляют собой многоядерные клетки, происходящие из гематопоэтических стволовых клеток, и посредством высвобождения ионов хлорида и ионов водорода на поверхность кости, на которой происходит адгезия остеокластов, остеокласты окисляют промежуток между поверхностью кости и остеокластами, а также секретируют катепсин К, который представляет собой кислую протеазу, или тому подобное. (American Journal of Physiology, (1991) 260, C1315-C1324). Это вызывает деградацию фосфата кальция, активацию кислых протеаз и деградацию белков костного матрикса, что приводит к резорбции кости.
Обнаружено, что клетки-предшественники остеокластов дифференцируются до остеокластов путем стимуляции RANKL (лигандом активатора рецептора NF-кВ), экспрессированным на клеточной мембране остеобластов/стромальных клеток, присутствующих на поверхности кости (Proceedings of the National Academy of Science of the United States of America, (1998) 95, pp. 3597-3602, и Cell, (1998) 93, pp. 165-176). Выявлено, что RANKL представляет собой мембранный белок, продуцируемый остеобластами/стромальными клетками, его экспрессию регулирует фактор резорбции кости, RANKL индуцирует дифференцировку клеток-предшественников остеокластов в зрелые многоядерные остеокласты, и тому подобное. (Proceedings of the National Academy of Science of the United States of America, (1998) 95, pp. 3597-3602, и Journal of Bone and Mineral Research, (1998) 23, S222). Кроме того, обнаружено, что у мышей с нокаутом, лишенных RANKL, развивается подобное остеопетрозу заболевание, и таким образом, доказано, что RANKL является физиологическим фактором, индуцирующим дифференцировку остеокластов (Nature, (1999) 397, pp. 315-323).
В качестве фармацевтических препаратов для лечения заболеваний метаболизма костной ткани или для сокращения времени лечения используют бисфосфонаты, активный витамин D3, кальцитонин и его производные, гормоны, такие как эстрадиол, SERM (избирательные модуляторы рецепторов эстрогенов), иприфлавон, витамин K2 (менатетренон), PTH, препараты кальция и тому подобное. Однако эти лекарственные средства не всегда являются удовлетворительными в отношении терапевтического эффекта, и необходима разработка лекарственного средства с более сильным терапевтическим эффектом.
Клеточные мембраны иммуноцитов покрыты плотным покрытием из различных гликанов, таких как сиалированные гликаны, которые узнают различные гликан-связывающие белки. Связывающие сиаловую кислоту иммуноглобулиноподобные лектины (далее в настоящем документе обозначенные как «siglec»), представляют собой семейство мембранных белков типа I, которые узнают сиалированные гликаны и связывают их. Многие siglec экспрессированы на клеточных мембранах иммуноцитов и узнают сиаловую кислоту, сходным образом присутствующую на клеточных мембранах иммуноцитов, и регулируют взаимодействие клеток или функцию клеток, и считают, что они вовлечены в иммунные ответы (Nature Reviews Immunology, (2007) 7, pp. 255-266). Однако, существует также множество молекул siglec, физиологические функции которых еще не выяснены. Siglec-15 (связывающий сиаловую кислоту иммуноглобулиноподобный лектин 15) представляет собой молекулу, которая, как недавно опубликовано, принадлежит к Siglec (смотри, например, Непатентный документ 10) и является идентичной молекуле, называемой CD33L3 (подобная CD33 молекула 3). Эта молекула является высоко эволюционно консервативной от рыб до человека, и обнаружена ее сильная экспрессия в дендритных клетках и/или макрофагах селезенки и лимфатических узлов человека. Кроме того, в результате тестирования связывания с использованием пробы сиаловой кислоты, обнаружено также, что Siglec-15 человека связывает Neu5Acα2-6GalNAc, и что Siglec-15 мыши дополнительно связывает Neu5Acα2-3Galβ1-4Glc и тому подобное (смотри, например, Glycobiology, (2007) 17, pp. 838-846). До недавнего времени физиологическая роль Siglec-15 не была выявлена, однако, опубликовано, что экспрессия Siglec-15 увеличивается при дифференцировке и созревании остеокластов, и дифференцировку остеокластов ингибируют уменьшением экспрессии Siglec-15 посредством РНК-интерференции (смотри, например, WO 07/093042). Кроме того, эффект антитела против Siglec-15 на дифференцировку остеокластов впервые выявили в WO 09/48072 (опубликованной 16 апреля 2009 г.), однако, последовательность антитела, которое можно вводить человеку, еще не выяснена.
Краткое изложение сущности изобретения
Проблемы, решаемые настоящим изобретением
Целью изобретения является ген, специфически экспрессирующийся при различных формах патологического метаболизма костной ткани, который наблюдают при остеопорозе, ревматоидном артрите, метастазировании злокачественной опухоли в кость или тому подобное, вещества, ингибирующего дифференцировку и созревание остеокластов и их активность, и терапевтического и/или профилактического средства против патологического метаболизма костной ткани.
Средства для решения проблем
Авторы настоящего изобретения проводили исследования для выяснения механизма дифференцировки, созревания и активации остеокластов, с целью обнаружения вещества, оказывающего терапевтический и/или профилактический эффект при патологическом метаболизме костной ткани. В результате авторы настоящего изобретения обнаружили, что экспрессия гена Siglec-15 увеличивается при дифференцировке и созревании остеокластов, и обнаружили также, что дифференцировку остеокластов ингибирует антитело, специфически связывающееся с Siglec-15. Кроме того, авторы настоящего изобретения гуманизировали полученное антитело крысы против Siglec-15 мыши, и таким образом, было создано настоящее изобретение.
Таким образом, изобретение относится к следующим изобретениям.
(1) Антитело, связывающее полипептид, содержащий остатки аминокислот 39-165 аминокислотной последовательности SEQ ID NO:2, и ингибирующее образование остеокластов и/или резорбцию кости остеокластами, или функциональный фрагмент антитела.
(2) Антитело или функциональный фрагмент антитела по п.(1), отличающиеся тем, что:
последовательность тяжелой цепи содержит вариабельную область с CDRH1, CDRH2 и CDRH3, и CDRH1 содержит аминокислотную последовательность SEQ ID NO:44, CDRH2 содержит любую из аминокислотных последовательностей SEQ ID NO:45 и SEQ ID NO:97, и CDRH3 содержит аминокислотную последовательность SEQ ID NO:46; и
последовательность легкой цепи содержит вариабельную область с CDRL1, CDRL2, и CDRL3, и CDRL1 содержит аминокислотную последовательность SEQ ID NO:47, CDRL2 содержит аминокислотную последовательность SEQ ID NO:48, и CDRL3 содержит аминокислотную последовательность SEQ ID NO:49.
(3) Антитело или функциональный фрагмент антитела по п.(2), отличающиеся тем, что содержат последовательность вариабельной области тяжелой цепи, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:41, и последовательность вариабельной области легкой цепи, содержащую остатки аминокислот 21-132 аминокислотной последовательности SEQ ID NO:43.
(4) Функциональный фрагмент антитела по любому из пп.(1)-(3), выбранный из группы, состоящей из Fab, F(ab')2, Fab' и Fv.
(5) Антитело по любому из пп.(1)-(3), отличающееся тем, что представляет собой scFv.
(6) Антитело или функциональный фрагмент антитела по любому из пп.(1)-(4), отличающийся тем, что антитело представляет собой химерное антитело.
(7) Антитело или функциональный фрагмент антитела по п.(6), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-470 аминокислотной последовательности SEQ ID NO:41 и последовательность легкой цепи, содержащую остатки аминокислот 21-237 аминокислотной последовательности SEQ ID NO:43.
(8) Антитело или функциональный фрагмент антитела по любому из пп.(1)-(4) и (6), отличающийся тем, что антитело является гуманизированным.
(9) Антитело по пп.(7) или (8), где тяжелая цепь обладает константной областью тяжелой цепи иммуноглобулина G2 человека, и легкая цепь обладает константной областью легкой цепи иммуноглобулина κ человека.
(10) Антитело, ингибирующее образование остеокластов и/или резорбцию кости остеокластами, или функциональный фрагмент антитела, где антитело содержит:
(a) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих аминокислотных последовательностей:
a1) аминокислотная последовательность, содержащая остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:51;
a2) аминокислотная последовательность, содержащая остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:53;
a3) аминокислотная последовательность, содержащая остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:55;
a4) аминокислотная последовательность, содержащая остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:57;
a5) аминокислотная последовательность, содержащая остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:59;
а6) аминокислотная последовательность, содержащая остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:99;
a7) аминокислотная последовательность, содержащая остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:101;
a8) аминокислотная последовательность, обладающая гомологией по меньшей мере 95% с любой из аминокислотных последовательностей, выбранных из a1)-a7);
a9) аминокислотная последовательность, обладающая гомологией по меньшей мере 99% с любой из аминокислотных последовательностей, выбранных из a1)-a7); и
a10) аминокислотная последовательность, включающая замену, делецию или добавление от одного до нескольких остатков аминокислот в любой из аминокислотных последовательностей, выбранных из a1)-a7); и
(b) вариабельную область легкой цепи, выбранную из группы, состоящей из следующих аминокислотных последовательностей:
b1) аминокислотная последовательность, содержащая остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:61;
b2) аминокислотная последовательность, содержащая остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:63;
b3) аминокислотная последовательность, содержащая остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:65;
b4) аминокислотная последовательность, содержащая остатки аминокислот 21-132 аминокислотной последовательности SEQ ID NO:67;
b5) аминокислотная последовательность, содержащая остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:69;
b6) аминокислотная последовательность, содержащая остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:71;
b7) аминокислотная последовательность, содержащая остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:103;
b8) аминокислотная последовательность, содержащая остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:105;
b9) аминокислотная последовательность, обладающая гомологией по меньшей мере 95% с любой из аминокислотных последовательностей, выбранных из b1)-a8);
b10) аминокислотная последовательность, обладающая гомологией по меньшей мере 99% с любой из аминокислотных последовательностей, выбранных из b1)-b8); и
b11) аминокислотная последовательность, включающая замену, делецию или добавление от одного до нескольких остатков аминокислот в любой из аминокислотных последовательностей, выбранных из b1)-b8).
(11) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:51 и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:61.
(12) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:53, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:63.
(13) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:55, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:65.
(14) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:55, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:67.
(15) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:57, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:69.
(16) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:59, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:71.
(17) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:69.
(18) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:101, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:69.
(19) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:103.
(20) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 аминокислотной последовательности SEQ ID NO:105.
(21) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-470 аминокислотной последовательности SEQ ID NO:51, и последовательность легкой цепи, содержащую остатки аминокислот 21-238 аминокислотной последовательности SEQ ID NO:61.
(22) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-470 аминокислотной последовательности SEQ ID NO:53, и последовательность легкой цепи, содержащую остатки аминокислот 21-238 аминокислотной последовательности SEQ ID NO:63.
(23) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-470 аминокислотной последовательности SEQ ID NO:55, и последовательность легкой цепи, содержащую остатки аминокислот 21-238 аминокислотной последовательности SEQ ID NO:65.
(24) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-470 аминокислотной последовательности SEQ ID NO:55, и последовательность легкой цепи, содержащую остатки аминокислот 21-237 аминокислотной последовательности SEQ ID NO:67.
(25) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-470 аминокислотной последовательности SEQ ID NO:57, и последовательность легкой цепи, содержащую остатки аминокислот 21-238 аминокислотной последовательности SEQ ID NO:69.
(26) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-470 аминокислотной последовательности SEQ ID NO:59, и последовательность легкой цепи, содержащую остатки аминокислот 21-238 аминокислотной последовательности SEQ ID NO:71.
(27) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-466 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую остатки аминокислот 21-238 аминокислотной последовательности SEQ ID NO:69.
(28) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-466 аминокислотной последовательности SEQ ID NO:101, и последовательность легкой цепи, содержащую остатки аминокислот 21-238 аминокислотной последовательности SEQ ID NO:69.
(29) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-466 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую остатки аминокислот остатки 21-238 аминокислотной последовательности SEQ ID NO:103.
(30) Антитело или функциональный фрагмент антитела по п.(10), отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую остатки аминокислот 20-466 аминокислотной последовательности SEQ ID NO:99 и последовательность легкой цепи, содержащую остатки аминокислот 21-238 аминокислотной последовательности SEQ ID NO:105.
(31) Фармацевтическая композиция, отличающаяся тем, что содержит по меньшей мере одно из антител или функциональных фрагментов антител по пп.(1)-(30).
(32) Фармацевтическая композиция по п.(31), отличающаяся тем, что является терапевтическим и/или профилактическим средством против патологического метаболизма костной ткани.
(33) Фармацевтическая композиция для лечения и/или профилактики патологического метаболизма костной ткани, отличающаяся тем, что содержит по меньшей мере одно из антител или функциональных фрагментов антител по пп.(1)-(30) и по меньшей мере один член, выбранный из группы, состоящей из бисфосфонатов, активного витамина D3, кальцитонина и его производных, гормонов, таких как эстрадиол, SERM (избирательных модуляторов рецепторов эстрогенов), иприфлавона, витамина K2 (менатетренона), препаратов кальция, PTH (паратиреоидного гормона), нестероидных противовоспалительных средств, растворимых рецепторов TNF, антител против TNF-α или функциональных фрагментов антител, антител против PTHrP (родственного паратиреоидному гормону белка) или функциональных фрагментов антител, антагонистов рецептора IL-1, антител против рецептора IL-6 или функциональных фрагментов антител, антител против RANKL или функциональных фрагментов антител и OCIF (фактора ингибирования остеокластогенеза).
(34) Фармацевтическая композиция по п.(32) или (33), где патологический метаболизм костной ткани выбран из группы, состоящей из остеопороза, разрушения кости, сопровождающего ревматоидный артрит, злокачественной гиперкальциемии, разрушения кости, сопровождающего множественную миелому или метастазирование злокачественной опухоли в кость, гигантоклеточной опухоли, остеопении, потери зубов из-за периодонтита, остеолиза около протеза сустава, разрушения кости при хроническом остеомиелите, болезни Педжета кости, нефрогенной остеодистрофии и несовершенного остеогенеза.
(35) Фармацевтическая композиция по п.(34), отличающаяся тем, что патологический метаболизм костной ткани представляет собой остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость.
(36) Фармацевтическая композиция по п.(35), отличающаяся тем, что патологический метаболизм костной ткани представляет собой остеопороз.
(37) Фармацевтическая композиция по п.(36), отличающаяся тем, что остеопороз представляет собой постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит.
(38) Способ лечения и/или профилактики патологического метаболизма костной ткани, отличающийся введением по меньшей мере одного из антител или функциональных фрагментов антител по пп.(1)-(30).
(39) Способ лечения и/или профилактики патологического метаболизма костной ткани, отличающийся введением одновременно или последовательно по меньшей мере одного из антител или функциональных фрагментов антител по пп.(1)-(30) и по меньшей мере одного члена, выбранного из группы, состоящей из бисфосфонатов, активного витамина D3, кальцитонина и его производных, гормонов, таких как эстрадиол, SERM (избирательных модуляторов рецепторов эстрогенов), иприфлавона, витамина K2 (менатетренона), препаратов кальция, PTH (паратиреоидного гормона), нестероидных противовоспалительных средств, растворимых рецепторов TNF, антител против TNF-α или функциональных фрагментов антител, антител против PTHrP (родственного паратиреоидному гормону белка) или функциональных фрагментов антител, антагонистов рецептора IL-1, антител против рецептора IL-6 или функциональных фрагментов антител, антител против RANKL или функциональных фрагментов антител и OCIF (фактора ингибирования остеокластогенеза).
(40) Способ лечения и/или профилактики по п.(38) или (39), отличающийся тем, что патологический метаболизм костной ткани представляет собой остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость.
(41) Способ лечения и/или профилактики по п.(40), отличающийся тем, что патологический метаболизм костной ткани представляет собой остеопороз.
(42) Способ лечения и/или профилактики по п.(41), отличающийся тем, что остеопороз представляет собой постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит.
(43) Полинуклеотид, кодирующий антитело по любому из пп.(3), (7) и (10)-(30).
(44) Полинуклеотид по п.(43), отличающийся тем, что содержит нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:40, и нуклеотидную последовательность, содержащую нуклеотиды 61-396 нуклеотидной последовательности SEQ ID NO:42.
(45) Полинуклеотид по п.(44), отличающийся тем, что содержит нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:40, и нуклеотидную последовательность, содержащую нуклеотиды 61-711 нуклеотидной последовательности SEQ ID NO:42.
(46) Полинуклеотид по п.(43), отличающийся тем, что содержит:
(a) полинуклеотид, выбранный из группы, состоящей из следующих нуклеотидных последовательностей:
a1) нуклеотидная последовательность, содержащая нуклеотиды 58-420 нуклеотидной последовательности SEQ ID N0: 50;
a2) нуклеотидная последовательность, содержащая нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:52;
a3) нуклеотидная последовательность, содержащая нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:54;
a4) нуклеотидная последовательность, содержащая нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:56;
a5) нуклеотидная последовательность, содержащая нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:58;
a6) нуклеотидная последовательность, содержащая нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:98;
a7) нуклеотидная последовательность, содержащая нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:100;
a8) нуклеотидная последовательность полинуклеотида, который гибридизуется с полинуклеотидом, содержащим нуклеотидную последовательность, комплементарную любой из нуклеотидных последовательностей, выбранных из a1)-a7), в жестких условиях; и
a9) нуклеотидная последовательность, включающая замену, делецию или добавление от одного до нескольких остатков аминокислот в любой из нуклеотидных последовательностей, выбранных из a1)-a7); и
(b) полинуклеотид, выбранный из группы, состоящей из следующих нуклеотидных последовательностей:
b1) нуклеотидная последовательность, содержащая нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:60;
b2) нуклеотидная последовательность, содержащая нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:62;
b3) нуклеотидная последовательность, содержащая нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:64;
b4) нуклеотидная последовательность, содержащая нуклеотиды 61-396 нуклеотидной последовательности SEQ ID NO:66;
b5) нуклеотидная последовательность, содержащая нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:68;
b6) нуклеотидная последовательность, содержащая нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:70;
b7) нуклеотидная последовательность, содержащая нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:102;
b8) нуклеотидная последовательность, содержащая нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:104;
b9) нуклеотидная последовательность полинуклеотида, который гибридизуется с полинуклеотидом, содержащим нуклеотидную последовательность, комплементарную любой из нуклеотидных последовательностей, выбранных из b1) - b8), в жестких условиях; и
b10) нуклеотидная последовательность, включающая замену, делецию или добавление от одного до нескольких нуклеотидов в любой из нуклеотидных последовательностей, выбранных из b1) - b8).
(47) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:50, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:60.
(48) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:52, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:62.
(49) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:54, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:64.
(50) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:54, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:66.
(51) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:56, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:68.
(52) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:58, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:70.
(53) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:68.
(54) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:100, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:68.
(55) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:102.
(56) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:104.
(57) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:50, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:60.
(58) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:52, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:62.
(59) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:54, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:64.
(60) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:54, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-711 нуклеотидной последовательности SEQ ID NO:66.
(61) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:56, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:68.
(62) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:58, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:70.
(63) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-1398 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:68.
(64) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-1398 нуклеотидной последовательности SEQ ID NO:100, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:68.
(65) Полинуклеотид по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 58-1398 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:102.
(66) Полинуклеотид, по п.(46), отличающийся тем, что содержит полинуклеотид, содержащий нуклеотидную, последовательность, содержащую нуклеотиды 58-1398 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:104.
(67) Вектор, содержащий любой из полинуклеотидов по пп.(43)-(66).
(68) Трансформированная клетка-хозяин, содержащая любой из полинуклеотидов по пп.(43)-(66).
(69) Трансформированная клетка-хозяин, содержащая вектор по п.(67).
(70) Способ получения антитела по любому из пп.(3), (7) и (10)-(30), включающий культивирование клетки-хозяина по п.(68) или (69) и очистку антитела из полученного продукта культивирования.
Преимущества изобретения
В соответствии с изобретением можно получить терапевтическое и/или профилактическое средство против патологического метаболизма костной ткани, механизмом действия которого является ингибирование дифференцировки и созревания остеокластов и их активности резорбции кости.
Краткое описание рисунков
[Фиг. 1] На фиг. 1 показаны результаты оценки чистоты Siglec-15-His мыши, очищенного хроматографией на колонке HisTrap HP и хроматографией на колонке Resource Q, посредством SDS-полиакриламидного электрофореза и окрашивания серебром.
[Фиг. 2] На фиг. 2 показаны результаты детекции характеристик Siglec-15-His мыши, очищенного хроматографией на колонке HisTrap HP и хроматографией на колонке Resource Q, посредством SDS-полиакриламидного электрофореза и Вестерн-блоттинга с использованием антитела против V5-HRP.
[Фиг. 3] На фиг. 3 показаны результаты оценки чистоты Siglec-15-Fc мыши, очищенного хроматографией на колонке HiTrap с белком A, посредством SDS-полиакриламидного электрофореза и окрашивания серебром.
[Фиг. 4] На фиг. 4 показаны результаты тестирования связывания моноклонального антитела крысы против Siglec-15 мыши в планшете с иммобилизованным на нем Siglec-15-Fc мыши, способом ELISA. Символ (♦) обозначает антитело #1A1, символ (■) обозначает антитело #3A1, символ (▲) обозначает антитело #8A1, символ (×) обозначает антитело #24A1, символ (•) обозначает антитело #32A1, символ (O) обозначает антитело #34A1, символ (+) обозначает антитело #39A1, символ (-) обозначает антитело #40A1, символ (-) обозначает антитело #41B1, символ (◊) обозначает антитело #61A1, и символ (□) обозначает контрольное IgG.
[Фиг. 5] На фиг. 5 показаны результаты тестирования эффекта добавления моноклонального антитела крысы против Siglec-15 (#3A1, #8A1, или #32A1) на дифференцировку остеокластов (стимуляция RANKL) из неадгерентных клеток костного мозга мыши. К тому же, контрольное IgG крысы на фигуре представляет собой отрицательный контроль, общий для фиг. 5 и 6.
[Фиг. 6] На фиг. 6 показаны результаты тестирования эффекта добавления моноклонального антитела крысы против Siglec-15 (#34A1, #39A1, или #40A1) на дифференцировку остеокластов (стимуляция RANKL) из неадгерентных клеток костного мозга мыши. К тому же, поликлональное антитело No. 3 кролика против Siglec-15 мыши на фигуре представляет собой положительный контроль, общий для фиг. 5 и 6.
[Фиг. 7] На фиг. 7 показаны результаты оценки чистоты Siglec-15-His человека, очищенного хроматографией на колонке HisTrap HP и хроматографией на колонке Resource Q, посредством SDS-полиакриламидного электрофореза и окрашивания серебром.
[Фиг. 8] На фиг. 8 показаны результаты оценки чистоты Siglec-15-Fc человека, очищенного хроматографией на колонке с белком A, посредством SDS-полиакриламидного электрофореза.
[Фиг. 9] На фиг. 9 показаны результаты тестирования связывания моноклонального антитела крысы против Siglec-15 мыши в планшете с иммобилизованным на нем Siglec-15-Fc человека, способом ELISA. Символ (♦) обозначает антитело #1A1, символ (■) обозначает антитело #3A1, символ (▲) обозначает антитело #8Al, символ (×) обозначает антитело #24A1, символ (•)обозначает антитело #32A1, символ (O) обозначает антитело #34A1, символ (+) обозначает антитело #39A1, символ (-) обозначает антитело #40A1, символ (-) обозначает антитело #41B1, символ (◊) обозначает антитело #61A1, и символ (□) обозначает контрольное IgG.
[Фиг. 10] На фиг. 10 показаны микрофотографии, изображающие, посредством окрашивания TRAP, ингибирование образования гигантских остеокластов из нормальных клеток-предшественников остеокластов человека посредством добавления моноклонального антитела крысы против Siglec-15 мыши.
[Фиг. 11] На фиг. 11 показаны микрофотографии, изображающие, посредством окрашивания TRAP, ингибирование образования гигантских остеокластов из нормальных клеток-предшественников остеокластов человека посредством добавления моноклонального антитела крысы против Siglec-15 мыши (антитело #32A1).
[Фиг. 12] Фиг. 12 представляет собой график, показывающий ингибирование активности резорбции кости нормальных остеокластов человека посредством добавления моноклонального антитела крысы против Siglec-15 мыши (антитело #32A1) (N=6).
[Фиг. 13] Фиг. 13A представляет собой график, показывающий эффект увеличения минеральной плотности костной ткани поясничного отдела позвоночника при введении моноклонального антитела крысы против Siglec-15 мыши крысам после овариэктомии в течение 4 недель; и фиг. 13B представляет собой график, показывающий эффект уменьшения выделения дезоксипиридинолина с мочой при введении моноклонального антитела крысы против Siglec-15 мыши крысам после овариэктомии в течение 4 недель.
[Фиг. 14] Фиг. 14 представляет собой график, показывающий, что моноклональное антитело крысы против Siglec-15 мыши #32A1 связывает домен V-set Siglec-15 человека, посредством конкурентного ELISA.
[Фиг. 15] Фиг. 15 представляет собой график, показывающий, что антитело крысы #32A1 и полученное из него химерное антитело человека обладают по существу одинаковой аффинностью для Siglec-15-Fc мыши по конкурентному ELISA.
[Фиг. 16] На фиг. 16 показан график, изображающий, по ферментативной активности TRAP, ингибирование образования остеокластов мыши посредством добавления моноклонального антитела крысы против Siglec-15 мыши (#32A1 антитело) и полученного из него химерного антитела (N=3).
[Фиг. 17] На фиг. 17 показаны микрофотографии, изображающие, посредством окрашивания TRAP, ингибирование образования гигантских остеокластов из нормальных клеток-предшественников остеокластов человека посредством добавления химерного антитела человека, полученного из моноклонального антитела крысы против Siglec-15 мыши #32A1.
[Фиг. 18] Фиг. 18 представляет собой изображение, на котором подтверждено, что 6 типов гуманизированных антител крысы против Siglec-15 мыши связывают белок Siglec-15 мыши зависимым от концентрации антител образом, способом ELISA с использованием планшета с иммобилизованным на нем Siglec-15-Fc мыши.
[Фиг. 19] Фиг. 19 представляет собой изображение, на котором подтверждено, что 6 типов гуманизированных антител крысы против Siglec-15 мыши связывают белок Siglec-15 человека зависимым от концентрации антител образом, способом ELISA с использованием планшета с иммобилизованным на нем Siglec-15-Fc человека.
[Фиг. 20] На фиг. 20 показаны микрофотографии, изображающие, посредством окрашивания TRAP, ингибирование образования гигантских остеокластов мыши при стимуляции TNFα посредством добавления моноклонального антитела крысы против Siglec-15 мыши (антитело #32A1) или полученного из него химерного антитела.
[Фиг. 21] На фиг. 21 показан график, изображающий, по ферментативной активности TRAP, ингибирование образования остеокластов мыши при стимуляции TNFα посредством добавления моноклонального антитела крысы против Siglec-15 мыши (антитело #32A1) или полученного из него химерного антитела (N=3).
[Фиг. 22] На фиг. 22 показан график, изображающий, по ферментативной активности TRAP, ингибирование образования остеокластов крысы посредством добавления моноклонального антитела крысы против Siglec-15 мыши (антитело #8A1 или #32A1) (N=3).
[Фиг. 23] На фиг. 23 показана клонированная нуклеотидная последовательность тяжелой цепи #32A1 крысы и ее аминокислотная последовательность.
[Фиг. 24] На фиг. 24 показана клонированная нуклеотидная последовательность легкой цепи #32A1 крысы и ее аминокислотная последовательность.
[Фиг. 25] На фиг. 25 показана нуклеотидная последовательность тяжелой цепи химерного антитела человека #32A1 и ее аминокислотная последовательность.
[Фиг. 26] На фиг. 26 показана нуклеотидная последовательность легкой цепи химерного антитела человека #32A1 и ее аминокислотная последовательность.
[Фиг. 27] На фиг. 27 показана нуклеотидная последовательность h#32A1-T1H и его аминокислотная последовательность.
[Фиг. 28] На фиг. 28 показана нуклеотидная последовательность h#32A1-T2H и его аминокислотная последовательность.
[Фиг. 29] На фиг. 29 показана нуклеотидная последовательность h#32A1-T3H и его аминокислотная последовательность.
[Фиг. 30] На фиг. 30 показана нуклеотидная последовательность h#32A1-T5H и его аминокислотная последовательность.
[Фиг. 31] На фиг. 31 показана нуклеотидная последовательность h#32A1-T6H и его аминокислотная последовательность.
[Фиг. 32] На фиг. 32 показана нуклеотидная последовательность h#32A1-T1L и его аминокислотная последовательность.
[Фиг. 33] На фиг. 33 показана нуклеотидная последовательность h#32A1-T2L и его аминокислотная последовательность.
[Фиг. 34] На фиг. 34 показана нуклеотидная последовательность h#32A1-T3L и его аминокислотная последовательность.
[Фиг. 35] На фиг. 35 показана нуклеотидная последовательность h#32A1-T4L и его аминокислотная последовательность.
[Фиг. 36] На фиг. 36 показана нуклеотидная последовательность h#32A1-T5L и его аминокислотная последовательность.
[Фиг. 37] На фиг. 37 показана нуклеотидная последовательность h#32A1-T6L и его аминокислотная последовательность.
[Фиг. 38] На фиг. 38 показаны аминокислотные последовательности h#32A1-T7H, h#32A1-T8H и h#32A1-T9H.
[Фиг. 39] На фиг. 39 показаны аминокислотные последовательности h#32A1-T10H, h#32A1-T11H и h#32A1-T12H.
[Фиг. 40] На фиг. 40 показаны аминокислотные последовательности h#32A1-T7L, h#32A1-T8L, h#32A1-T9L, h#32A1-T10L и h#32A1-T11L.
[Фиг. 41] На фиг. 41 показаны аминокислотные последовательности h#32A1-T12L, h#32A1-T13L, h#32A1-T14L, h#32A1-T15L и h#32A1-T16L.
[Фиг. 42] На фиг. 42 показаны аминокислотные последовательности h#32A1-T17L, h#32A1-T18L, h#32A1-T19L, h#32A1-T20L и h#32A1-T21L.
[Фиг. 43] На фиг. 43 показаны аминокислотные последовательности h#32A1-T22L, h#32A1-T23L, h#32A1-T24L и h#32A1-T25L.
[Фиг. 44] На фиг. 44 показан график, изображающий, по ферментативной активности TRAP, ингибирование образования остеокластов мыши посредством добавления гуманизированного антитела крысы против Siglec-15 мыши (h#32A1-T1, h#32A1-T2 или h#32A1-T3). К тому же, антитело крысы #32A1 на фигуре представляет собой положительный контроль, общий для фиг. 44 и 45.
[Фиг. 45] На фиг. 45 показан график, изображающий, по ферментативной активности TRAP, ингибирование образования остеокластов мыши посредством добавления гуманизированного антитела крысы против Siglec-15 мыши (h#32A1-T4, h#32A1-T5 или h#32A1-T6).
[Фиг. 46] На фиг. 46 показан график, изображающий, по ферментативной активности TRAP, ингибирование образования остеокластов мыши посредством добавления гуманизированного антитела крысы против Siglec-15 мыши (h#32A1-H1-1/L5 или h#32A1-H5/L5). К тому же, антитело #32A1 крысы и химерное антитело человека #32A1 на фигуре представляют собой положительный контроль, общий для фиг. 46 и 47.
[Фиг. 47] На фиг. 47 показан график, изображающий, по ферментативной активности TRAP, ингибирование образования остеокластов мыши посредством добавления гуманизированного антитела крысы против Siglec-15 мыши (h#32A1-H1-1/L2-16 или h#32A1-H-1/L2-15).
[Фиг. 48] На фиг. 48 показан график, изображающий ингибирование активности резорбции кости нормальных остеокластов человека посредством добавления гуманизированного антитела крысы против Siglec-15 мыши (h#32A1-T1, h#32A1-T2, h#32A1-T3, h#32A1-T4, h#32A1-T5 или h#32A1-T6). К тому же, антитело крысы #32A1 и химерное антитело человека #32A1 на фигуре представляют собой положительный контроль, общий для фиг. 48 и 49.
[Фиг. 49] На фиг. 49 показан график, изображающий ингибирование активности резорбции кости нормальных остеокластов человека посредством добавления гуманизированного антитела крысы против Siglec-15 мыши (h#32A1-H1-1/L5, h#32A1-H5/L5, h#32A1-H1-1/L2-16 или h#32A1-H1-1/L12-5).
[Фиг. 50] Фиг. 50 представляет собой термограмму, полученную для определения термостабильности антитела h#32A1-H5/L5.
[Фиг. 51] Фиг. 51 представляет собой термограмму, полученную для определения термостабильности антитела h#32A1-H1-1/L5.
[Фиг. 52] Фиг. 52 представляет собой термограмму, полученную для определения термостабильности антитела h#32A1-H1-1/L12-5.
[Фиг. 53] Фиг. 53 представляет собой термограмму, полученную для определения термостабильности антитела h#32A1-H1-1/L2-16.
[Фиг. 54] На фиг. 54 показана нуклеотидная последовательность h#32A1-H1-1 и его аминокислотная последовательность.
[Фиг. 55] На фиг. 55 показана нуклеотидная последовательность h#32A1-H5 и его аминокислотная последовательность.
[Фиг. 56] На фиг. 56 показана нуклеотидная последовательность h#32A1-L2-15 и его аминокислотная последовательность.
[Фиг. 57] На фиг. 57 показана нуклеотидная последовательность h#32A1-L2-16 и его аминокислотная последовательность.
Способ осуществления изобретения
Термин «ген», как используют в настоящем документе, включает не только ДНК, но также мРНК, кДНК и кРНК.
Термин «полинуклеотид», как используют в настоящем документе, используют с таким же значением, как «нуклеиновая кислота», и он также включает ДНК, РНК, зонды, олигонуклеотиды и праймеры.
Термины «полипептид» и «белок», как используют в настоящем документе, используют взаимозаменяемо.
Термин «фракция РНК», как используют в настоящем документе, относится к фракции, содержащей РНК.
Термин «клетка», как используют в настоящем документе, включает также клетки в животном-индивидууме и культивируемые клетки.
Термин «Siglec-15», как используют в настоящем документе, используют с таким же значением, как «белок Siglec-15».
Термин «образование остеокластов», как используют в настоящем документе, используют с таким же значением, как «дифференцировка остеокластов» или «созревание остеокластов».
Термин «функциональный фрагмент антитела», как используют в настоящем документе, относится к частичному фрагменту антитела, обладающему активностью связывания антигена, и включает Fab, F(ab')2, Fv, scFv, диатела, линейные антитела, полиспецифические антитела, сформированные из фрагментов антител, и тому подобное. Термин включает также Fab', который представляет собой моновалентный фрагмент в вариабельной области антитела, полученный обработкой F(ab')2 в восстанавливающих условиях. Однако, термин не является ограниченным этими молекулами, при условии, что фрагмент обладает аффинностью связывания антигена. Кроме того, эти функциональные фрагменты включают не только фрагмент, полученный обработкой полноразмерной молекулы белка антитела подходящим ферментом, но также белок, продуцированный в подходящей клетке-хозяине с использованием генетически модифицированного гена антитела.
Термин «эпитоп», как используют в настоящем документе, относится к частичному пептиду или к частичной третичной структуре Siglec-15, с которыми связывается специфическое антитело против Siglec-15. Вышеупомянутый эпитоп, представляющий собой частичный пептид Siglec-15, можно определить способом, хорошо известным специалистам в данной области, таким как иммуноанализ. Однако, можно применять, например, следующий способ. Получены различные частичные структуры Siglec-15. При получении частичных структур можно использовать известный способ синтеза олигопептидов. Например, серию полипептидов с соответствующим образом уменьшенной длиной, полученной последовательным укорачиванием Siglec-15 с С-конца или N-конца, получают с использованием способа генетической рекомбинации, известной специалистам в данной области. Затем исследуют реактивность антитела против этих полипептидов и приблизительно определяют участок узнавания. Затем синтезируют пептиды, обладающие самой короткой длиной, и определяют реактивность по отношению к этим пептидам, при помощи чего можно определить эпитоп. Кроме того, эпитоп, представляющий собой частичную третичную структуру Siglec-15, связываемую специфическим антителом против Siglec-15, посредством определения остатков аминокислот Siglec-15, соседних с антителом, посредством рентгеноструктурного анализа. Если второе антитело против Siglec-15 связывается с частичным пептидом или частичной третичной структурой, с которыми связывается первое антитело против Siglec-15, можно определить, что первое антитело и второе антитело разделяют один и тот же эпитоп. Кроме того, посредством подтверждения, что второе антитело против Siglec-15 конкурирует с первым антителом против Siglec-15 за связывание с Siglec-15 (то есть, второе антитело ингибирует связывание между Siglec-15 и первым антителом), можно определить, что первое антитело и второе антитело разделяют один и тот же эпитоп, даже если последовательность или структура специфического эпитопа не определена. Кроме того, когда первое антитело и второе антитело связывают один и тот же эпитоп, а также первое антитело оказывает конкретный эффект, такой как активность нейтрализации антигена, можно ожидать, что второе антитело обладает такой же активностью.
Известно, что каждая из тяжелой и легкой цепи молекулы антитела обладает тремя определяющими комплементарность областями (CDR). Определяющую комплементарность область называют также гипервариабельным доменом, и она присутствует в вариабельной области каждой тяжелой и легкой цепи антитела. Она представляет собой участок, обладающий необычайно высокой изменчивостью первичной структуры, и существуют три отдельных CDR в первичной структуре каждой тяжелой и легкой полипептидной цепи. В этом описании, в отношении определяющих комплементарность областей антитела, определяющие комплементарность области тяжелой цепи представлены CDRH1, CDRH2 и CDRH3 со стороны N-конца аминокислотной последовательности тяжелой цепи, и определяющие комплементарность области легкой цепи представлены CDRL1, CDRL2 и CDRL3 со стороны N-конца аминокислотной последовательности легкой цепи. Эти участки расположены близко друг к другу в третичной структуре и определяют специфичность для антигена, с которым связывается антитело.
Фраза «гибридизацию проводят в жестких условиях», как используют в настоящем документе, относится к гибридизации, которую проводят в условиях, при которых идентификации можно достигать проведением гибридизации при 68°C в коммерчески доступном растворе для гибридизации ExpressHyb (изготовленном в Clontech, Inc.) или проведением гибридизации при 68°C в присутствии 0,7-1,0 М NaCl с использованием фильтра с иммобилизованной на нем ДНК, с последующим проведением промывки при 68°C с использованием 0,1-2 х раствора SSC (1 x раствор SSC состоит из 150 мМ NaCl и 15 мМ цитрата натрия) или в эквивалентных этим условиях.
1. Siglec-15
Авторы настоящего изобретения обнаружили, что ген Siglec-15 специфически экспрессируется в гигантоклеточных опухолях, и обнаружили также, что уровень экспрессии гена Siglec-15 увеличивается, когда происходящая из моноцитов линия клеток дифференцирует в остеокласты.
В отношении использования Siglec-15 по изобретению, Siglec-15 очищают непосредственно из моноцитов или клеток костного мозга человека, не относящегося к человеку млекопитающего (такого как морская свинка, крыса, мышь, кролик, свинья, овца, крупный рогатый скот или обезьяна) или курицы, и используют, или получают фракцию клеточной мембраны из вышеупомянутых клеток, и ее можно использовать. Кроме того, Siglec-15 можно получать его синтезом in vitro или его продукцией в клетке-хозяине посредством генной инженерии. Конкретно, при генно-инженерной продукции, кДНК Siglec-15 встраивают в вектор, способный экспрессировать кДНК Siglec-15, и Siglec-15 синтезируют в растворе, содержащем фермент, субстрат и вещество, дающее энергию, необходимые для транскрипции и трансляции, или другую прокариотическую или эукариотическую клетку-хозяина трансформируют для экспрессии Siglec-15, в результате чего можно получить белок.
Нуклеотидная последовательность кДНК Siglec-15 человека задепонирована в GenBank под инвентарным номером NM_213602 и представлена как SEQ ID NO:1 в разделе “Список последовательностей”, и его аминокислотная последовательность представлена как SEQ ID NO:2 в разделе “Список последовательностей”. Нуклеотидная последовательность кДНК Siglec-15 мыши задепонирована в GenBank под инвентарным номером XM_884636 и представлена как SEQ ID NO:3 в разделе “Список последовательностей”, и его аминокислотная последовательность представлена как SEQ ID NO:4 в разделе “Список последовательностей”. Зрелый Siglec-15 человека, из которого удалена сигнальная последовательность, соответствует аминокислотной последовательности, содержащей остатки аминокислот 21-328 аминокислотной последовательности SEQ ID NO:2. Кроме того, Siglec-15 мыши, из которого удалена сигнальная последовательность, соответствует аминокислотной последовательности, содержащей остатки аминокислот 21-341 аминокислотной последовательности SEQ ID NO:4. К тому же, Siglec-15 иногда называют подобным антигену CD33 3, подобным молекуле CD33 3, CD33-подобным 3 или CD33L3, и все они представляют собой одну и ту же молекулу.
кДНК Siglec-15 можно получать, например, так называемым способом ПЦР, в котором полимеразную цепную реакцию (далее в настоящем документе обозначенную как «ПЦР») проводят с использованием библиотеки кДНК, экспрессирующей кДНК Siglec-15, в качестве матрицы и праймеров для специфической амплификации кДНК Siglec-15 (Saiki, R. K., et al., Science, (1988) 239, 487-49).
Кроме того, полинуклеотид, гибридизующийся с полинуклеотидом, содержащим нуклеотидную последовательность, комплементарную по меньшей мере одной нуклеотидной последовательности, выбранной из нуклеотидных последовательностей, представленных как SEQ ID NO:1 и 3 в разделе “Список последовательностей”, в жестких условиях и кодирующий белок, обладающий биологической активностью, сравнимой с активностью Siglec-15, также рассматривают как кДНК Siglec-15. Кроме того, полинуклеотид, представляющий собой вариант сплайсинга, транскрибируемый с локуса Siglec-15 человека или мыши, или полинуклеотид, гибридизующийся с последовательностью, комплементарной ему, в жестких условиях и кодирующий белок, обладающий биологической активностью, сравнимой с активностью Siglec-15, также рассматривают как кДНК Siglec-15.
Кроме того, белок, содержащий аминокислотную последовательность, содержащую замену, делецию или добавление одной или нескольких аминокислот по меньшей мере в одной из аминокислотных последовательностей, выбранных из аминокислотных последовательностей SEQ ID NO:2 и 4 в разделе “Список последовательностей”, или аминокислотную последовательность, полученную удалением сигнальной последовательности из любой из этих последовательностей, и обладающий биологической активностью, сравнимой с активностью Siglec-15, также рассматривают как Siglec-15. Кроме того, белок, содержащий аминокислотную последовательность, кодируемую вариантом сплайсинга, транскрибируемым с локуса Siglec-15 человека или мыши, или аминокислотную последовательность, содержащую замену, делецию или добавление одной или нескольких аминокислот в ней, и обладающий биологической активностью, сравнимой с активностью Siglec-15, также рассматривают как Siglec-15.
2. Детекция патологического метаболизма костной ткани
Анализ уровня экспрессии гена Siglec-15 в группе тестируемых образцов из различных костных тканей человека показал, что уровень экспрессии гена значительно повышен в гигантоклеточной опухоли (GCT), которая представляет собой опухоль костной ткани с большим количеством возникающих подобных остеокластам многоядерных гигантских клеток, и характеризуется клинической картиной с остеолитическим разрушением костной ткани (Bullough et al., Atlas of Orthopedic Pathology 2nd edition, pp. 17.6-17.8, Lippincott Williams & Wilkins Publishers (1992)).
Обнаружено также, что уровень экспрессии гена Siglec-15 увеличивается, когда происходящая из моноцитов линия клеток дифференцирует в остеокласты.
Соответственно, считают, что Siglec-15 связан с патологиями человека, такими как GCT, при которых увеличивается резорбция кости. Другими словами, измерение уровня экспрессии гена Siglec-15 и/или Siglec-15 в каждой клетке и/или в каждой ткани позволяет определение состояния патологического метаболизма костной ткани, сопровождающееся сверхэкспрессией Siglec-15. Патологический метаболизм костной ткани, как используют в настоящем документе, относится к нарушению, отличающемуся общей потерей костной массы, и оно включает в качестве конкретных неограничивающих примеров, остеопороз (постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит), разрушение кости, сопровождающее ревматоидный артрит, злокачественную гиперкальциемию, разрушение кости, сопровождающее множественную миелому или метастазирование злокачественной опухоли в кость, гигантоклеточную опухоль, остеопению, потерю зубов из-за периодонтита, остеолиз около протеза сустава, разрушение кости при хроническом остеомиелите, болезнь Педжета кости, нефрогенную остеодистрофию и несовершенный остеогенез.
По изобретению, «тестируемый образец» для использования для исследования уровня экспрессии гена Siglec-15 и/или Siglec-15 относится к образцу из ткани костного мозга, кости, предстательной железы, яичек, полового члена, мочевого пузыря, почки, полости рта, глотки, губы, языка, десны, носоглотки, пищевода, желудка, тонкого кишечника, толстого кишечника, ободочной кишки, печени, желчного пузыря, поджелудочной железы, носа, легких, мягких тканей, кожи, молочной железы, матки, яичника, головного мозга, щитовидной железы, лимфатического узла, мышцы, жировой ткани или тому подобное, или крови, жидкости организма, продуктов выделения или тому подобное, полученных от тестируемого субъекта, клинического материала и т.д., однако, по изобретению, кровь или костный мозг являются более предпочтительными.
Что касается RANKL, который, как известно, ассоциирован с дифференцировкой остеокластов, получена мышь с нокаутом, и проанализирован фенотип при потере функции RANKL (Young-Yun Kong, et. al., Nature (1999) 397, pp. 315-323). Посредством получения мыши с нокаутом, лишенной Siglec-15, таким же способом, как выше, можно анализировать фенотип, при котором утрачена функция Siglec-15.
3. Получение антитела против Siglec-15
Антитело по изобретению, нацеленное против Siglec-15, может быть получено посредством иммунизации животного с помощью Siglec-15 или произвольного полипептида, выбранного из аминокислотной последовательности Siglec-15, и сбора и очистки продуцированного in vivo антитела общепринятым способом. Биологический вид Siglec-15, предназначенного для использования в качестве антигена, не является ограниченным человеком, и животного можно иммунизировать с помощью Siglec-15, полученного из животного, отличного от человека, такого как мышь или крыса. В этом случае, посредством исследования перекрестной реактивности между связыванием антитела с полученным гетерологичным Siglec-15 и Siglec-15 человека, можно выбрать антитело, применимое в случае заболевания человека.
Кроме того, моноклональное антитело может быть получено путем слияния продуцирующих антитела клеток, продуцирующих антитело против Siglec-15, с клетками миеломы для получения гибридомы известным способом (например, Kohler and Milstein, Nature, (1975) 256, pp. 495-497; Kennet, R. ed., Monoclonal Antibodies, pp. 365-367, Plenum Press, N.Y. (1980)).
К тому же, Siglec-15, предназначенный для использования в качестве антигена, можно получать посредством генной инженерии, чтобы заставить клетку-хозяина продуцировать ген Siglec-15.
Конкретно, получают вектор, способный экспрессировать ген Siglec-15, и полученным вектором трансфицируют клетку-хозяина для экспрессии гена, и затем очищают экспрессированный Siglec-15. Далее в настоящем документе конкретно описан способ получения антитела против Siglec-15.
(1) Получение антигена
Примеры антигена, предназначенного для использования для получения антитела против Siglec-15, включают Siglec-15, полипептид, содержащий частичную аминокислотную последовательность, содержащую по меньшей мере 6 последовательных аминокислот из Siglec-15, и производное, полученное добавлением к нему данной аминокислотной последовательности или носителя.
Siglec-15 можно очищать непосредственно из тканей опухолей или клеток опухолей человека и использовать. Кроме того, Siglec-15 можно получать его синтезом in vitro или вызывая его продукцию в клетке-хозяине посредством генной инженерии.
Что касается генной инженерии, конкретно, кДНК Siglec-15 встраивают в вектор, способный экспрессировать кДНК Siglec-15, и Siglec-15 синтезируют в растворе, содержащем фермент, субстрат и вещество, дающее энергию, необходимые для транскрипции и трансляции, или другую прокариотическую или эукариотическую клетку-хозяина трансформируют для экспрессии Siglec-15, в результате чего можно получить антиген.
Кроме того, антиген можно получать также в форме секреторного белка посредством экспрессии слитого белка, полученного лигированием внеклеточного домена Siglec-15, который представляет собой мембранный белок, с константной областью антитела в подходящей системе хозяин-вектор.
кДНК Siglec-15 можно получать, например, так называемым способом ПЦР, при котором полимеразную цепную реакцию (далее в настоящем документе обозначенную как «ПЦР») проводят с использованием библиотеки кДНК, экспрессирующей кДНК Siglec-15, в качестве матрицы и праймеров для специфической амплификации кДНК Siglec-15 (смотри Saiki, R. K., et al., Science, (1988) 239, pp. 487-489).
В отношении синтеза полипептида in vitro, в качестве неограничивающего примера Rapid Translation System (RTS), изготовленную в Roche Diagnostics, Inc.
Примеры прокариотического хозяина включают Escherichia coli и Bacillus subtilis. Для трансформации клетки-хозяина намеченным геном, клетку-хозяина трансформируют с использованием плазмидного вектора, содержащего репликон, т.е., точку начала репликации, происходящую из вида, совместимого с хозяином, и регуляторную последовательность. Кроме того, вектор предпочтительно обладает последовательностью, способной задавать фенотипическую избирательность трансформированной клетки.
Примеры эукариотической клетки-хозяина включают клетки позвоночных, клетки насекомых и клетки дрожжей. Что касается клеток позвоночных, в качестве неограничивающих примеров, часто используют дефицитные по дигидрофолатредуктазе линии (Urlaub, G. and Chasin, L. A., Proc. Natl. Acad. Sci. USA (1980) 77, pp. 4126-4220) клеток обезьяны COS (Gluzman, Y., Cell, (1981) 23, pp. 175-182, ATCC CRL-1650), фибробласты мыши NIH3T3 (ATCC No. CRL-1658) и клетки яичника китайского хомяка (клетки CHO; ATCC: CCL-61); и тому подобное.
Полученный таким образом трансформант можно культивировать общепринятым способом, и при культивировании трансформанта намеченный полипептид продуцируется внутри клеток или вне клеток.
Подходящую среду, предназначенную для культивирования, можно выбирать из различных общепринятых культуральных сред, в зависимости от применяемой клетки-хозяина. Если применяют Escherichia coli, можно использовать, например, среду LB, дополненную антибиотиком, таким как ампициллин или IPMG, по необходимости.
Рекомбинантный белок, продуцируемый внутри клетки или вне клетки таким трансформантом посредством такого культивирования, можно отделять и очищать любым из различных известных способов разделения с использованием физических или химических свойств белка.
Конкретные примеры способов включают обработку общепринятым осаждающим белок средством, ультрафильтрацию, различные типы жидкостной хроматографии, такие как хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография и аффинная хроматография, диализ и их сочетание.
Кроме того, посредством присоединения метки из шести остатков гистидина к рекомбинантному белку, подлежащему экспрессии, белок можно эффективно очищать на никель-аффинной колонке. Альтернативно, посредством присоединения Fc-области IgG к рекомбинантному белку, подлежащему экспрессии, белок можно эффективно очищать с помощью колонки с белком A.
Посредством сочетания вышеупомянутых способов, большое количество намеченного полипептида можно легко получать с высоким выходом и высокой чистотой.
(2) Получение моноклонального антитела против Siglec-15
Примеры антитела, специфически связывающего Siglec-15, включают моноклональное антитело, специфически связывающее Siglec-15, и способ получения антитела является таким, как описано ниже.
Для получения моноклонального антитела, как правило, необходимы следующие функциональные стадии:
(a) очистка биополимера, подлежащего использованию в качестве антигена;
(b) получение продуцирующих антитело клеток посредством иммунизации животного посредством инъекции антигена, сбора крови, анализа титра антитела в ней для определения, когда следует вырезать селезенку;
(c) получение клеток миеломы (далее в настоящем документе обозначенных как «миелома»);
(d) слияние продуцирующих антитело клеток с миеломой;
(e) скрининг группы гибридом, продуцирующих намеченное антитело;
(f) разделение гибридом на клоны отдельных клеток (клонирование);
(g) необязательно, культивирование гибридомы или выращивание животного с имплантированной гибридомой для продукции большого количества моноклонального антитела;
(h) исследование полученного таким образом моноклонального антитела по биологической активности и специфичности связывания, или анализ того же самого по свойствам в качестве меченного реагента; и тому подобное.
Далее в настоящем документе способ получения моноклонального антитела подробно описан, следуя вышеуказанным стадиям, однако, способ не является ограниченным этим, и, например, можно использовать продуцирующие антитело клетки и миелому, отличные от клеток селезенки.
(a) Очистка антигена
В качестве антигена можно использовать Siglec-15, полученный способом, как описано выше, или его частичный пептид.
Кроме того, фракцию мембран, полученную из рекомбинантных клеток, экспрессирующих Siglec-15, или сами рекомбинантные клетки, экспрессирующие Siglec-15, а также частичный пептид из белка по изобретению, химически синтезированный способом, известным специалистам в данной области, также можно использовать в качестве антигена.
(b) Получение продуцирующих антитело клеток
Антиген, полученный на стадии (a), смешивают с адъювантом, таким как полный или неполный адъювант Фрейнда, или сульфат алюминия-калия, и полученную смесь используют в качестве иммуногена для иммунизации экспериментального животного. В качестве экспериментального животного можно легко использовать любого животного, используемого в известном способе получения гибридомы. Конкретно, можно использовать, например, мышь, крысу, козу, овцу, крупный рогатый скот, лошадь или тому подобное. Однако, с точки зрения простоты доступности клеток миеломы, подлежащих слиянию с выделенными продуцирующими антитело клетками, мышь или крысу предпочтительно используют в качестве животного, подлежащего иммунизации.
Кроме того, линия мышей или крыс, подлежащая использованию, не является конкретно ограниченной, и в случае мыши можно использовать, например, различные линии, такие как A, AKR, BALB/c, BDP, BA, СЕ, C3H, 57BL, C57BL, C57L, DBA, FL, HTH, HT1, LP, NZB, NZW, RF, R III, SJL, SWR, WB и 129, а в случае крысы можно использовать, например, Wistar, Low, Lewis, Sprague, Dawley, ACI, BN, Fischer и тому подобное.
Эти мыши и крысы являются коммерчески доступными от заводчиков/распространителей экспериментальных животных, например, CLEA Japan, Inc. и Charles River Laboratories Japan, Inc.
Среди них, принимая во внимание совместимость при слиянии с клетками миеломы, как описано ниже, в случае мыши, линия BALB/c, и в случае крысы, линии Wistar и Low являются особенно предпочтительными в качестве животного, подлежащего иммунизации.
Кроме того, принимая во внимание антигенную гомологию между человеком и мышью, является также предпочтительным использовать мышь, обладающую сниженной биологической функцией удаления аутоантител, то есть, мышь с аутоиммунным заболеванием.
Возраст мыши или крысы на время иммунизации предпочтительно составляет 5-12 недель, более предпочтительно, 6-8 недель.
Для иммунизации животного Siglec-15 или его рекомбинантным белком, можно использовать, например, известный способ, подробно описанный, например, в Weir, D. M., Handbook of Experimental Immunology Vol. I. II. III., Blackwell Scientific Publications, Oxford (1987), Kabat, E. A. and Mayer, M. M., Experimental Immunochemistry, Charles С Thomas Publisher Springfield, Illinois (1964) или тому подобное.
Среди этих способов иммунизации, предпочтительный конкретный способ по изобретению представляет собой, например, следующий.
То есть, сначала, фракция мембранных белков, служащую антигеном, или клетки, в которых вызвана экспрессия антигена является/являются внутрикожно или внутрибрюшинно вводимыми животному.
Однако, сочетание обоих способов введения является предпочтительным для увеличения эффективности иммунизации, и когда внутрикожное введение проводят в первой половине, а внутрибрюшинное введение проводят во второй половине или только при последнем дозировании, эффективность иммунизации можно особенно увеличить.
Расписание введения антигена меняется в зависимости от типа животного, подлежащего иммунизации, индивидуальных отличий, или тому подобное. Однако, как правило, расписание введения, при котором частота введения антигена составляет 3-6 раз, и интервал дозирования составляет 2-6 недель, является предпочтительным, и расписание введения, при котором частота введения антигена составляет 3-4 раза, и интервал дозирования составляет 2-4 недели, является более предпочтительным.
Кроме того, доза антигена меняется в зависимости от типа животного, подлежащего иммунизации, индивидуальных отличий, или тому подобное, однако, как правило, устанавливают дозу 0,05-5 мг, предпочтительно, приблизительно 0,1-0,5 мг.
Бустер-иммунизацию проводят через 1-6 недель, предпочтительно, 2-4 недели, более предпочтительно, 2-3 недели после введения антигена, как описано выше.
Доза антигена на время проведения бустер-иммунизации меняется в зависимости от типа или размера животного или тому подобное, однако, в случае мыши, доза, как правило, составляет 0,05-5 мг, предпочтительно, 0,1-0,5 мг, более предпочтительно, приблизительно 0,1-0,2 мг.
Клетки селезенки или лимфоциты, включая продуцирующие антитело клетки, в стерильных условиях удаляют у иммунизированного животного через 1-10 суток, предпочтительно, 2-5 суток, более предпочтительно, 2-3 суток после бустер-иммунизации.
В это время измеряют титр антитела, и если животного, обладающего значительно увеличенным титром антитела, используют в качестве источника продуцирующих антитело клеток, последующую процедуру можно выполнять более эффективно.
Примеры способа измерения титра антител, подлежащего использованию по изобретению, включают способ RIA и способ ELISA, но способ не является ограниченным ими.
Например, если применяют способ ELISA, измерение титра антител по изобретению можно выполнять способами, как описано ниже.
Сначала очищенный или частично очищенный антиген адсорбируют на поверхности твердой фазы, такой как 96-луночный планшет для ELISA, и поверхность твердой фазы без адсорбированного на ней антигена, покрывают белком, неродственным антигену, таким как бычий сывороточный альбумин (далее в настоящем документе обозначенный как «BSA»). После промывки поверхности, поверхность приводят в контакт с серийно разведенным образцом (например, сыворотки мыши) в качестве первичного антитела, чтобы позволить антителу в образце связываться с антигеном.
Затем, в качестве вторичного антитела, антитело, меченное ферментом, против антитела мыши, добавляют и позволяют связываться с антителом мыши. После промывки добавляют субстрат для фермента и изменение поглощения, происходящее из-за развития окраски, индуцированного деградацией субстрата или тому подобное, измеряют и рассчитывают титр антитела на основании измерения.
Отделение продуцирующих антитело клеток от клеток селезенки или лимфоцитов можно осуществлять известным способом (например, Kohler et al., Nature (1975), 256, p. 495; Kohler et al., Eur. J. Immunol. (1977), 6, p. 511; Milstein et al., Nature (1977), 266, p. 550; Walsh, Nature (1977), 266, p. 495).
Например, в случае клеток селезенки, можно применять общий способ, в котором продуцирующие антитело клетки отделяют гомогенизацией селезенки для получения клеток фильтрацией через сито из нержавеющей стали и суспендированием клеток в минимальной эссенциальной среде Игла (MEM).
(c) Получение клеток миеломы (далее в настоящем документе обозначенных как «миелома»)
Клетки миеломы, подлежащие использованию для слияния клеток, не являются конкретно ограниченными, и подходящие клетки можно выбирать из известных линий клеток. Однако, из соображений удобства, когда гибридому отбирают из слитых клеток, является предпочтительным использование линии, дефицитной по HGPRT (гипоксантин-гуанин фосфорибозилтрансфераза), способ отбора для которой разработан.
Более конкретно, примеры HGPRT-дефицитного штамма включают X63-Ag8(X63), NS1-ANS/1(NS1), P3X63-Ag8.U1(P3U1), X63-Ag8.653(X63.653), SP2/0-Ag14(SP2/0), MPC11-45.6TG1.7(45.6TG), F0, S149/5XXO и BU.1, полученные из мышей; 210.RSY3.Ag.1.2.3(Y3), полученная из крыс; и U266AR(SKO-007), GM1500-GTG-A12(GM1500), UC729-6, LICR-LOW-HMy2(HMy2) и 8226AR/NIP4-1(NP41), полученные из человека.
Эти HGPRT-дефицитные линии доступны, например, из Американской коллекции типовых культур (ATCC) или тому подобное.
Эти линии клеток субкультивировали в подходящей среде, такой как среда с 8-азагуанином [среда, полученная добавлением 8-азагуанина к среде RPMI-1640, дополненной глутамином, 2-меркаптоэтанолом, гентамицином и эмбриональной телячьей сывороткой (далее в настоящем документе обозначенной как «FCS»)]; среда Дульбекко, модифицированная по способу Исков (далее в настоящем документе обозначенная как «IMDM»), или модифицированная по способу Дульбекко среда Игла (далее в настоящем документе, обозначенный как «DMEM»). В этом случае, за 3-4 суток до выполнения слияния клеток, клетки субкультивировали в нормальной среде [например, среде ASF104 (изготовленной в Ajinomoto Co., Ltd.), содержащую 10% FCS] для получения не менее 2×107 клеток на сутки слияния клеток.
(d) Слияние клеток
Слияние между продуцирующими антитело клетками и клетками миеломы можно соответствующим образом проводить известным способом (Weir, D. M. Handbook of Experimental Immunology Vol. I. II. III., Blackwell Scientific Publications, Oxford (1987), Kabat, E. A. and Mayer, M. M., Experimental Immunochemistry, Charles С Thomas Publisher, Springfield, Illinois (1964), etc.), в таких условиях, что выживаемость клеток не чрезмерно снижена.
В качестве такого способа можно использовать химический способ, в котором продуцирующие антитело клетки и клетки миеломы смешивают в растворе, содержащем полимер, такой как полиэтиленгликоль, в высокой концентрации, физический способ с использованием электростимуляции или тому подобное.
Среди этих способов, конкретный пример химического способа является таким, как описано ниже.
То есть, в случае, когда полиэтиленгликоль используют в растворе, содержащем полимер в высокой концентрации, продуцирующие антитело клетки и клетки миеломы смешивают в растворе полиэтиленгликоля с молекулярной массой 1500-6000, более предпочтительно, 2000-4000, при температуре от 30 до 40°C, предпочтительно, от 35 до 38°C, в течение 1-10 минут, предпочтительно, 5-8 минут.
(e) Отбор группы гибридом
Способ отбора гибридом, полученных вышеупомянутым слиянием клеток, не является конкретно ограниченным. Как правило, применяют способ отбора с HAT (гипоксантин, аминоптерин, тимидин) (Kohler et al., Nature (1975), 256, p. 495; Milstein et al., Nature (1977), 266, p. 550).
Этот способ является эффективным, когда гибридомы получены с использованием клеток миеломы из HGPRT-дефицитной линии, которые не могут выживать в присутствии аминоптерина.
Таким образом, при культивировании неслитых клеток и гибридом в среде HAT, только гибридомам, устойчивым к аминоптерину, избирательно позволяют выживать и пролиферировать.
(f) Разделение на клоны отдельных клеток (клонирование)
В качестве способа клонирования гибридом можно использовать известный способ, такой как способ с метилцеллюлозой, способ с мягкой агарозой, или способ предельных разведений (смотри, например, Barbara, В. M. and Stanley, M. S.: Selected Methods in Cellular Immunology, W. H. Freeman and Company, San Francisco (1980)). Среди этих способов, в частности, способ предельных разведений является предпочтительным.
По этому способу, линию клеток фибробластов, полученную из зародыша крысы, или питающие клетки, такие как нормальные клетки селезенки мыши, клетки тимуса, или клетки асцита, рассевают в микропланшет.
При этом гибридомы разводят в среде для получения плотности клеток 0,2-0,5 клеток на 0,2 мл. Аликвоту 0,1 мл разведенной суспензии гибридом добавляют в каждую лунку и культивирование продолжают в течение приблизительно 2 недель, в то же время заменяя приблизительно 1/3 культурального раствора свежей средой в предопределенные интервалы времени (например, каждые 3 суток), в результате чего клоны гибридомы могут пролиферировать.
Гибридомы в лунках, в которых подтвержден титр антитела, подвергали, например, клонированию способом предельных разведений с повтором 2-4 раза. Гибридому, для которой подтвердили стабильный титр антител, выбирают в качестве линии гибридомы, продуцирующей моноклональное антитело против Siglec-15.
Примеры клонированной таким образом линии гибридомы, включают гибридому #32A1, описанную в WO 09/48072. Гибридома #32A1 депонирована в Международном Патентном Депозитарии Организмов Национального Института Развития Прикладной Науки и Технологий (расположенном в Central 6, 1-1-1 Higashi, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan) 28 августа 2008 г., и ей присвоен инвентарный номер FERM BP-10999 с наименованием anti-Siglec-15 Hybridoma #32A1. К тому же, в этом описании, антитело, продуцируемое гибридомой #32A1, обозначено как «антитело #32A1» или просто «#32A1». Кроме того, антитела, полученные в Примерах из этого описания, отличные от антитела #32A1, также представлены наименованиями антител таким же образом. Частичный фрагмент, содержащий вариабельную область тяжелой цепи антитела #32A1, обладает аминокислотной последовательностью, содержащей остатки аминокислот 20-167 из SEQ ID NO:28 в разделе “Список последовательностей”. Кроме того, частичный фрагмент, содержащий последовательность вариабельной области легкой цепи антитела #32A1, обладает аминокислотной последовательностью, содержащей остатки аминокислот 21-139 из SEQ ID NO:30 в разделе “Список последовательностей”.
(g) Получение моноклонального антитела культивированием гибридомы
Посредством культивирования отобранной таким образом гибридомы, можно эффективно получать моноклональное антитело. Однако, перед культивированием, является предпочтительным проводить скрининг гибридомы, продуцирующей немеченое моноклональное антитело.
В таком скрининге можно применять известный способ.
Измерение титра антитела по изобретению можно осуществлять, например, способом ELISA, объясненным в пункте (b), описанном выше.
Гибридому, полученную способом, описанным выше, можно сохранять в замороженном состоянии в жидком азоте или в морозильнике при -80°C или ниже.
После завершения клонирования среду меняют со среды HT на нормальную среду, и культивируют гибридому.
Крупномасштабное культивирование проводят посредством ротационного культивирования с использованием большого культурального сосуда или посредством культивирования с постоянным перемешиванием.
Из супернатанта, полученного при крупномасштабном культивировании, моноклональное антитело, специфически связывающееся с белком по изобретению, можно получать посредством очистки с использованием способа, известного специалистам в данной области, такого как гель-фильтрация.
Кроме того, гибридому инъецируют в брюшную полость мыши той же самой линии, что и гибридома (например, вышеупомянутой BALB/c), или мыши Nu/Nu для пролиферации гибридомы, посредством чего можно получать асциты, содержащие большое количество моноклонального антитела по изобретению.
В случае, когда гибридому вводят в брюшную полость, если минеральное масло, такое как 2,6,10,14-тетраметилпентадекан (пристан), вводят за 3-7 суток до этого, можно получать большее количество асцитов.
Например, иммунодепрессант предварительно инъецируют в брюшную полость мыши той же самой линии, что и гибридома, для инактивации T-клеток. Через 20 суток после этого, 106-107 клеток клона гибридомы суспендируют в бессывороточной среде (0,5 мл), и суспензию инъецируют в брюшную полость мыши. Как правило, когда брюшная полость расширяется и заполняется асцитами, асциты собирают у мыши.
По этому способу, моноклональное антитело можно получить в концентрации, приблизительно в 100 раз или более выше, чем в культуральном растворе.
Моноклональное антитело, полученное вышеупомянутым способом, можно очищать способом, описанных, например, в Weir, D. M. : Handbook of Experimental Immunology Vol. I, II, III, Blackwell Scientific Publications, Oxford (1978).
Таким образом, примеры способа включают способ осаждения сульфатом аммония, гель-фильтрацию, ионообменную хроматографию и аффинную хроматографию.
В качестве простого способа очистки, можно использовать также коммерчески доступный набор для очистки моноклональных антител (например, набор MAbTrap GII, изготовленный в Pharmacia, Inc.) или тому подобное.
Полученное таким образом моноклональное антитело обладает высокой антигенной специфичностью для Siglec-15.
(h) Анализ моноклонального антитела
Изотип и подкласс полученного таким образом моноклонального антитела, можно определить следующим образом.
Во-первых, примеры способа идентификации включают способ Оухтерлони, способ ELISA и способ RIA.
Способ Оухтерлони является простым, но при низкой концентрации моноклонального антитела необходима операция концентрирования.
С другой стороны, при использовании способа ELISA или способа RIA, посредством прямой реакции культурального супернатанта с твердой фазой с адсорбированным антигеном и с использованием антител, соответствующих различным типам изотипов и подклассов иммуноглобулинов, в качестве вторичных антител, можно идентифицировать изотип и подкласс моноклонального антитела.
Кроме того, в качестве более простого способа, можно использовать также коммерчески доступный набор для идентификации (например, набор Mouse Typer, изготовленный в Bio-Rad Laboratories, Inc.) или тому подобное.
Кроме того, количественное определение белка можно осуществлять способом Фолина-Лоури и способом расчета, основанным на оптической плотности при 280 нм [1,4 (OD 280) = 1 мг/мл иммуноглобулина].
(3) Другие антитела
Антитело по изобретению включает не только вышеупомянутое моноклональное антитело против Siglec-15, но также рекомбинантное антитело, полученное искусственной модификацией с целью уменьшения гетерологичной антигенности для человека, такое как химерное антитело, гуманизированное антитело и антитело человека. Эти антитела можно получать с использованием известного способа.
В качестве химерного антитела, можно привести пример антитела, в котором вариабельные и константные области антитела получены из различных видов, например, химерное антитело, в котором происходящая из мыши или крысы вариабельная область антитела соединена с константной областью человеческого происхождения (смотри Proc Natl. Acad. Sci. USA, 81, 6851-6855, (1984)). В качестве одного примера химерного антитела, полученного из антитела крысы против белка мыши #32A1, можно привести антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-470 из SEQ ID NO:41 в разделе “Список последовательностей”, и легкую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 21-237 из SEQ ID NO:43 в разделе “Список последовательностей”.
В качестве примера гуманизированного антитела можно привести антитело, полученное вставкой только определяющей комплементарность области (CDR) в антитело человеческого происхождения (смотри Nature (1986) 321, pp. 522-525), и антитело, полученное прививкой части остатков аминокислот каркаса, так же как последовательности CDR, в антитело человека, способом CDR-прививки (WO 90/07861). В качестве примера гуманизированного антитела из антитела #32A1 крысы, можно привести произвольное сочетание тяжелой цепи, содержащей вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 из SEQ ID NO:51 в разделе “Список последовательностей”, остатки аминокислот 20-140 SEQ ID NO:53, остатки аминокислот 20-140 SEQ ID NO:55, остатки аминокислот 20-140 из SEQ ID NO:57, остатки аминокислот 20-140 SEQ ID NO:59, остатки аминокислот 20-140 SEQ ID NO:99, остатки аминокислот 20-140 SEQ ID NO:101, или остатки аминокислот 1-121 любой из аминокислотных последовательностей, представленных на SEQ ID NO:72-77, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 SEQ ID NO:61, остатки аминокислот 21-133 SEQ ID NO:63, остатки аминокислот 21-133 SEQ ID NO:65, остатки аминокислот 21-132 из SEQ ID NO:67, остатки аминокислот 21-133 SEQ ID NO:69, остатки аминокислот 21-133 SEQ ID NO:71, остатки аминокислот 21-133 SEQ ID NO:103, остатки аминокислот 21-133 SEQ ID NO:105 или остатки аминокислот 1-113 любой из аминокислотных последовательностей, представленных на SEQ ID NO:78-96.
В качестве примера предпочтительного сочетания можно привести антитело, содержащее тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 SEQ ID NO:51, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 SEQ ID NO:61, антитело, содержащее тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 SEQ ID NO:53, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 SEQ ID NO:63, антитело, содержащее тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 SEQ ID NO:55, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 SEQ ID NO:65, антитело, содержащее тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 SEQ ID NO:55, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 SEQ ID NO:67, антитело, содержащее тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 SEQ ID NO:57, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 SEQ ID NO:69, антитело, содержащее тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 SEQ ID NO:59, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 SEQ ID NO:71, антитело, содержащее тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 из SEQ ID NO:99, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 из SEQ ID NO:69, антитело, содержащее тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 из SEQ ID NO:101, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 из SEQ ID NO:69, антитело, содержащее тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 из SEQ ID NO:99, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 из SEQ ID NO:103, или антитело, содержащее тяжелую цепь, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 20-140 из SEQ ID NO:99, и легкую цепь, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую остатки аминокислот 21-133 из SEQ ID NO:105.
В качестве примера более предпочтительного сочетания, можно привести антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-470 из SEQ ID NO:51, и легкую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 21-238 из SEQ ID NO:61, антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-470 из SEQ ID NO:53, и легкой цепью, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 21-238 из SEQ ID NO:63, антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-470 из SEQ ID NO:55, и легкую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 21-238 из SEQ ID NO:65, антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-470 из SEQ ID NO:55, и легкую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 21-237 из SEQ ID NO:67, антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-470 из SEQ ID NO:57, и легкую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 21-238 из SEQ ID NO:69, антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-470 из SEQ ID NO:59, и легкой цепью, обладающей аминокислотной последовательностью, содержащей остатки аминокислот 21-238 из SEQ ID NO:71, антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-466 из SEQ ID NO:99, и легкой цепью, обладающей аминокислотной последовательностью, содержащей остатки аминокислот 21-238 из SEQ ID NO:69, антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-466 из SEQ ID NO:101, и легкую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 21-238 из SEQ ID NO:69, антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-466 из SEQ ID NO:99, и легкую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 21-238 из SEQ ID NO:103, или антитело, содержащее тяжелую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 20-466 из SEQ ID NO:99, и легкую цепь, обладающую аминокислотной последовательностью, содержащей остатки аминокислот 21-238 из SEQ ID NO:105.
Однако, гуманизированное антитело, полученное из антитела #32A1, не является ограниченным вышеупомянутыми гуманизированными антителами, при условии, что гуманизированное антитело обладает всеми 6 типами последовательностей CDR из #32A1 и обладает активностью ингибирования образования остеокластов. К тому же, вариабельная область тяжелой цепи из антитела #32A1 обладает CDRH1 (DYFMN), содержащей аминокислотную последовательность SEQ ID NO:44, или одной из CDRH2 (QIRNKIYTYATFYAESLEG), содержащей аминокислотную последовательность SEQ ID NO:45, и CDRH2 (QIRNKIYTYATFYA) SEQ ID NO:97, и CDRH3 (SLTGGDYFDY), содержащей аминокислотную последовательность SEQ ID NO:46. CDRH2 SEQ ID NO:45, находится в соответствии с определением Kabat (SEQUENCES OF PROTEINS OF IMMUNOLOGICAL INTEREST VOL. I, FIFTH EDTION (1991)). CDRH2, представленная на SEQ ID NO:97, сделана короче на пять остатков на С-конце, чем по определению Kabat. В последовательности тяжелой цепи, содержащей эту CDRH2, последовательность CDR, полученная из крысы, сделана короче и включено больше из каркасной последовательности человека, и таким образом, при введении человеку, она намного менее вероятно будет узнаваемой как гетерологичный антиген. Кроме того, вариабельная область легкой цепи антитела #32A1 обладает CDRL1 (RASQSVTISGYSFIH), содержащей аминокислотную последовательность SEQ ID NO:47, CDRL2 (RASNLAS), содержащей аминокислотную последовательность SEQ ID NO:48, и CDRL3 (QQSRKSPWT), содержащей аминокислотную последовательность SEQ ID NO:49.
Кроме того, антитело по изобретению включает антитело человека. Антитело человека против Siglec-15 относится к антителу человека, обладающего только последовательностью гена антитела, полученной из хромосомы человека. Антитело человека против Siglec-15 можно получать способом с использованием продуцирующей антитело человека мыши, обладающей фрагментом хромосомы человека, содержащим гены тяжелой и легкой цепи антитела человека (смотри Tomizuka, K. et al., Nature Genetics (1997) 16, pp. 133-143; Kuroiwa, У. et al., Nucl. Acids Res. (1998) 26, pp. 3447-3448; Yoshida, H. et al., Animal Cell Technology: Basic and Applied Aspects vol. 10, pp. 69-73 (Kitagawa, Y., Matuda, T. and Iijima, S. eds.), Kluwer Academic Publishers, 1999; Tomizuka, K. et al., Proc Natl. Acad. Sci. USA (2000) 97, pp. 722-727 и т.д.).
Такую мышь, продуцирующую антитело человека, можно получать конкретно следующим образом. Генетически модифицированное животное, у которого эндогенные локусы генов тяжелых и легких цепей иммуноглобулинов нарушены, и вместо этого, локусы генов тяжелых и легких цепей иммуноглобулинов введены посредством вектора на основе искусственной хромосомы дрожжей (YAC) или тому подобное, получают посредством получения животного с нокаутом и трансгенного животного, и скрещивания этих животных.
Кроме того, способом генной инженерии, с использованием кДНК, кодирующей также тяжелую цепь и легкую цепь антитела человека, соответственно, и предпочтительно, вектора, содержащего кДНК, трансформируют эукариотические клетки, и трансформант, продуцирующий рекомбинантное моноклональное антитело человека, культивируют, в результате чего антитело также можно получать из культурального супернатанта.
По настоящему изобретению, в качестве хозяина можно использовать, например, эукариотические клетки, предпочтительно, клетки млекопитающих, такие как клетки CHO, лимфоциты или клетки миеломы.
Кроме того, известен также способ получения происходящего из фагового дисплея антитела человека, отобранного скринингом библиотеки антител человека (смотри Wormstone, I. M. et al., Investigative Ophthalmology & Visual Science. (2002) 43 (7), pp. 2301-2308; Carmen, S. et al., Briefings in Functional Genomics and Proteomics (2002), 1 (2), pp. 189-203; Siriwardena, D. et al., Opthalmology (2002) 109 (3), pp. 427-431 и т.д.).
Например, можно использовать способ фагового дисплея, в котором вариабельная область антитела человека экспрессирована на поверхности фага в форме одноцепочечного антитела (scFv), и отбирать фаг, который связывается с антигеном (Nature Biotechnology (2005), 23, (9), pp. 1105-1116).
Посредством анализа гена фага, отобранного на основании связывания с антигеном, можно определить последовательность ДНК, кодирующую вариабельную область антитела человека, связывающегося с антигеном.
Если определена последовательность ДНК scFv, которое связывается с антигеном, можно получать антитело человека посредством получения вектора экспрессии, обладающего этой последовательностью, и введения вектора в подходящего хозяина для его экспрессии (WO 92/01047, WO 92/20791, WO 93/06213, WO 93/11236, WO 93/19172, WO 95/01438, WO 95/15388, Annu. Rev. Immunol (1994) 12, pp. 433-455, Nature Biotechnology (2005) 23 (9), pp. 1105-1116).
Антитела, полученные вышеуказанным способом, оценивают по свойству связывания с антигеном способом, описанным в примере 25, или тому подобное, и можно выбирать предпочтительное антитело. В качестве одного из примеров другого показателя при сравнении свойств антител, можно привести стабильность антител. Дифференциальная сканирующая калориметрия (DSC), показанная в примере 33, представляет собой инструмент, способный быстро и точно измерять среднюю точку температуры термической денатурации (Tm), которая представляет собой удобный показатель относительной структурной стабильности белков. Посредством измерения значений Tm с использованием DSC и сравнения значений, можно сравнивать разницу в термостабильности. Известно, что для стабильности антител при хранении показана некоторая корреляция с термостабильностью антител (Lori Burton, et. al., Pharmaceutical Development and Technology (2007) 12, pp. 265-273), и предпочтительное антитело можно выбирать с использованием термостабильности в качестве показателя. Примеры других показателей для выбора антител включают следующие: выход в подходящей клетке-хозяине является высоким и способность к агрегации в водном растворе является низкой. Например, для антитела, для которого показан наивысший выход, не всегда показывают высокую термостабильность, и таким образом, необходимо выбирать антитело, наиболее подходящее для введения человеку, посредством проведения сравнительной оценки на основании вышеупомянутых показателей.
Кроме того, известен также способ, в котором последовательности полноразмерной тяжелой и легкой цепи антитела лигируют с использованием подходящего линкера, в результате чего получают одноцепочечный иммуноглобулин (Lee, H-S, et. al., Molecular Immunology (1999) 36, pp. 61-71; Shirrmann, T. et. al., mAbs (2010), 2, (1) pp. 1-4). При димеризации такого одноцепочечного иммуноглобулина, полученный димер может обладать структурой и активностью, сходными со структурой и активностью антитела, которое само по себе является тетрамером. Кроме того, антитело по изобретению может представлять собой антитело, обладающее вариабельной областью одной тяжелой цепи и не обладающее последовательностью легкой цепи. Такое антитело называют однодоменным антителом (sdAb) или нанотелом, и фактически, его наблюдали у верблюдов и лам, и опубликовано, что оно обладает аффинностью связывания антигена (Muyldemans S. et. al., Protein Eng. (1994) 7 (9), 1129-35, Hamers-Casterman C. et. al., Nature (1993) 363 (6428) 446-8). Понятно, что вышеупомянутые антитела представляют собой типы функциональных фрагментов антитела по изобретению.
Кроме того, посредством контроля гликозилирования, при котором гликан связывают с антителом по изобретению, можно усиливать зависимую от антитела клеточную цитотоксическую активность. Что касается способов гликозилирования антител, известны WO 99/54342, WO 00/61739, WO 02/31140 и т.д. Однако, способы не являются ограниченными этим.
В случаях, когда антитело получают, сначала выделяя ген антитела, и затем вводя ген в подходящего хозяина, можно использовать сочетание подходящего хозяина и подходящего вектора экспрессии. Конкретные примеры гена антитела включают сочетание гена, кодирующего последовательность тяжелой цепи антитела, описанного в этом описании, и гена, кодирующего последовательность его легкой цепи. При трансформации клетки-хозяина, можно встраивать последовательность гена тяжелой цепи и последовательность гена легкой цепи в один и тот же вектор экспрессии, а также в различные векторы экспрессии по отдельности. В случаях, когда эукариотические клетки используют в качестве хозяина, можно использовать клетки животных, клетки растений и эукариотические микроорганизмы. В качестве примеров клеток животных, можно привести (1) клетки млекопитающих, например, дефицитные по дигидрофолатредуктазе линии (Urlaub, G. and Chasin, L. A., Proc. Natl. Acad. Sci. USA (1980) 77, pp. 4126-4220) клеток обезьяны COS (Gluzman, Y., Cell, (1981) 23, pp. 175-182, ATCC CRL-1650), фибробласты мыши NIH3T3 (ATCC No. CRL-1658) и клетки яичника китайского хомяка (клетки CHO; ATCC: CCL-61). Кроме того, в случаях, когда используют прокариотические клетки, в качестве примеров можно привести Escherichia coli и Bacillus subtilis. Посредством введения гена намеченного антитела в эти клетки посредством трансформации и культивирования трансформированных таким образом клеток in vitro, можно получать антитело. В вышеупомянутом способе культивирования выход иногда может меняться в зависимости от последовательности антитела, и таким образом, можно выбрать антитело, которое легко получать как лекарственное средство, с использованием выхода в качестве показателя, среди антител, обладающих сравнимой активностью связывания.
Не существует ограничений по изотипу антитела по изобретению, и его примеры включают IgG (IgG1, IgG2, IgG3, IgG4), IgM, IgA (IgA1, IgA2) IgD, и IgE, и его предпочтительные примеры включают IgG и IgM, и далее, его более предпочтительные примеры включают IgG1 и IgG2.
Кроме того, антитело по изобретению может представлять собой функциональный фрагмент антитела, обладающий антигенсвязывающим участком антитела, или его модифицированный фрагмент. Фрагмент антитела можно получать обработкой антитела протеазой, такой как папаин или пепсин, или модификацией гена антитела способом генной инженерии и экспрессией модифицированного гена в подходящих культивируемых клетках. Среди этих фрагментов антител, фрагмент, обладающий всеми или частью функций полноразмерной молекулы антитела, можно назвать функциональным фрагментом антитела. В качестве примеров функций антитела, в основном можно привести активность связывания антигена, активность нейтрализации активности антигена, активность увеличения активности антигена, зависимую от антитела цитотоксическую активность, комплемент-зависимую цитотоксическую активность и комплемент-зависимую клеточную цитотоксическую активность. Функцией функционального фрагмента антитела по изобретению является активность связывания с Siglec-15, предпочтительно, активность ингибирования образования остеокластов, более предпочтительно, активность ингибирования процесса слияния клеток остеокластов.
Примеры фрагмента антитела включают Fab, F(ab')2, Fv, одноцепочечное Fv (scFv), в котором молекулы Fv тяжелой цепи и легкой цепи лигированы через подходящий линкер, диатело (диатела), линейное антитело и полиспецифическое антитело, состоящее из фрагментов антител. Кроме того, Fab', который представляет собой моновалентный фрагмент в вариабельной области антитела, полученный обработкой F(ab')2 в восстанавливающих условиях, также рассматривают как фрагмент антитела.
Кроме того, антитело по изобретению может представлять собой полиспецифическое антитело со специфичностью по меньшей мере для двух различных антигенов. Как правило, такая молекула связывается с двумя антигенами (то есть, биспецифическое антитело), однако, «полиспецифическое антитело», как используют в настоящем документе, включает антитело, обладающее специфичностью для двух или более (например, трех) антигенов.
Полиспецифическое антитело по изобретению может представлять собой полноразмерное антитело или фрагмент такого антитела (например, биспецифическое антитело F(ab')2). Биспецифическое антитело можно получать лигированием тяжелых и легких цепей (пар HL) из двух типов антител, или можно также получать слиянием гибридом, продуцирующих различные моноклональные антитела, для получения слитых клеток, продуцирующих биспецифические антитела (Millstein et.al., Nature (1983) 305, pp. 537-539).
Антитело по изобретению может представлять собой одноцепочечное антитело (обозначаемое также scFv). Одноцепочечное антитело можно получать лигированием вариабельной области тяжелой цепи и вариабельной области легкой цепи антитела через полипептидный линкер (Pluckthun, The Pharmacology of monoclonal Antibodies, 113 (edited by Rosenburg and Moore, Springer Verlag, New York, pp. 269-315 (1994)), Nature Biotechnology (2005), 23, pp. 1126-1136). Кроме того, фрагмент BiscFv, полученный лигированием двух молекул scFv через полипептидный линкер, также можно использовать в качестве биспецифического антитела.
Этот способ получения одноцепочечного антитела известен в данной области (смотри, например, Патенты США No. 4946778, 5260203, 5091513, 5455030 и т.д.). В этих scFv вариабельная область тяжелой цепи и вариабельная область легкой цепи антитела лигированы через линкер, который не формирует конъюгат, предпочтительно, через полипептидный линкер (Huston, J. S. et. al., Proc. Natl. Acad. Sci. USA (1988), 85, pp. 5879-5883). В scFv вариабельная область тяжелой цепи и вариабельная область легкой цепи могут происходить из одного и того же антитела или из различных антител.
В качестве полипептидного линкера, подлежащего использованию для лигирования вариабельных областей, используют, например, данный одноцепочечный пептид, состоящий из 12-19 остатков.
ДНК, кодирующую scFv, можно получать проведением амплификации с использованием ДНК, кодирующей полную аминокислотную последовательность или желательную частичную аминокислотную последовательность, выбранную из тяжелой цепи или вариабельной области тяжелой цепи из вышеупомянутого антитела и легкой цепи или вариабельной области легкой цепи из него, в качестве матрицы, способом ПЦР с использованием пары праймеров, определяющей оба ее конца, и дополнительным проведением амплификации посредством объединения ДНК, кодирующей часть полипептидного линкера, и пары праймеров, определяющей оба ее конца, так чтобы лигировать оба конца тяжелой цепи и легкой цепи, соответственно.
Далее, после получения ДНК, кодирующей scFv, вектор экспрессии, содержащий эту ДНК, и хозяина, трансформированного вектором экспрессии, можно получать общепринятыми способами. Далее, с использованием полученного хозяина, scFv можно получать общепринятыми способами. Фрагмент антитела из него можно продуцировать в хозяине посредством получения гена и экспрессии гена таким же образом, как описано выше.
Антитело по изобретению можно подвергать мультимеризации для увеличения его аффинности для антигена. Антитело, подлежащее мультимеризации, может представлять собой один тип антитела или множество антител, которые узнают множество эпитопов одного и того же антигена. В качестве примера способа мультимеризации антитела, можно привести связывание домена CH3 IgG с двумя молекулами scFv, связывание со стрептавидином, введение мотива спираль-петля-спираль и тому подобное.
Антитело по изобретению может представлять собой поликлональное антитело, которое представляет собой смесь множества типов антител против Siglec-15, обладающих различными аминокислотными последовательностями. В качестве одного примера поликлональных антител, можно привести смесь множества типов антител, обладающих различными CDR. В качестве такого поликлонального антитела, культивируют смесь клеток, продуцирующих различные антитела, и можно использовать антитело, очищенное из полученной культуры (смотри WO 2004/061104).
В качестве модифицированного антитела, можно использовать также антитело, связанное с любым из различных типов молекул, таких как полиэтиленгликоль (PEG).
Кроме того, антитело по изобретению может присутствовать в форме конъюгата, сформированного между любым из этих антител и другим лекарственным средством (иммуноконъюгата). Примеры такого антитела включают те, в которых антитело является конъюгированным с радиоактивным материалом или соединением, оказывающим фармакологическое действие (Nature Biotechnology (2005) 23, pp. 1137-1146).
Полученное антитело можно очищать до гомогенности. Выделение и очистку антитела можно проводить с применением общепринятого способа выделения и очистки белка. Например, антитело можно выделять и очищать посредством выбора и комбинации соответствующим образом хроматографии на колонке, фильтрации на фильтрах, ультрафильтрации, осаждения солью, диализа, препаративного электрофореза в полиакриламидном геле, электрофореза с изоэлектрической фокусировкой, и тому подобное (Strategies for Protein Purification and Characterization: A Laboratory Course Manual, Daniel R. Marshak et al. eds. Cold Spring Harbor Laboratory Press (1996); Antibodies: A Laboratory Manual. Ed Harlow and David Lane, Cold Spring Harbor Laboratory (1988)), но способ не является ограниченным этим.
Примеры такой хроматографии включают аффинную хроматографию, ионообменную хроматографию, гидрофобную хроматографию, гель-фильтрационную хроматографию, обращеннофазовую хроматография и адсорбционную хроматографию.
Такую хроматографию можно проводить с применением жидкостной хроматографии, такой как HPLC или FPLC.
В качестве примера колонки для применения в аффинной хроматографии, можно привести колонку с белком A и колонку с белком G.
Например, в качестве примера колонки с использованием белка A, можно привести Hyper D, POROS, Сефарозу FF (Pharmacia) и тому подобное.
Кроме того, с использованием носителя с иммобилизованным на нем антигеном, антитело можно очищать также с использованием свойства связывания антитела с антигеном.
4. Лекарственное средство, содержащее антитело против Siglec-15
Из антител против Siglec-15, полученных способом, описанным в вышеуказанном пункте «3. Получение антитела против Siglec-15», можно получить антитело, нейтрализующее биологическую активность Siglec-15. Такое антитело, нейтрализующее биологическую активность Siglec-15, ингибирует биологическую активность Siglec-15 in vivo, т.е., дифференцировку и/или созревание остеокластов, и таким образом, его можно использовать в качестве терапевтического и/или профилактического средства при патологическом метаболизме костной ткани, вызванном патологической дифференцировкой и/или созреванием остеокластов, в виде лекарственного средства. Патологический метаболизм костной ткани может представлять собой любое нарушение, характеризующееся общей потерей костной массы (остеопению или остеолиз). Как правило, лечение и/или профилактика с помощью антитела против Siglec-15 является/являются применимыми в случае, когда необходимо ингибирование резорбции кости. Примеры патологического метаболизма костной ткани, который можно лечить и/или предотвращать с помощью антитела против Siglec-15, включают остеопороз (постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит), разрушение кости, сопровождающее ревматоидный артрит, злокачественную гиперкальциемию, разрушение кости, сопровождающее множественную миелому или метастазирование злокачественной опухоли в кость, гигантоклеточную опухоль, остеопению, потерю зубов из-за периодонтита, остеолиз около протеза сустава, разрушение кости при хроническом остеомиелите, болезнь Педжета кости, нефрогенную остеодистрофию и несовершенный остеогенез, однако, патологический метаболизм костной ткани не является ограниченным этим, при условии, что заболевание сопровождается общей потерей костной массы, вызванной остеокластами. Примеры антитела против Siglec-15, подлежащего применению в качестве вышеупомянутого лекарственного средства, включают химерное антитело и гуманизированное антитело, полученное из антитела #32A1 способом, описанным в пункте 3. «(3) Другие антитела». Кроме того, химерное антитело, гуманизированное антитело и антитело человека, разделяющие тот же самый эпитоп, что и антитело #32A1, можно использовать в качестве лекарственного средства. Разделяет ли конкретное антитело против Siglec-15 тот же самый эпитоп, что и антитело #32A1, можно подтвердить наблюдением, связывают или нет эти антитела один и тот же специфический частичный пептид Siglec-15. Кроме того, можно определять также, что если конкретное антитело против Siglec-15 конкурирует с антителом #32A1 за связывание с Siglec-15, эти антитела разделяют один и тот же эпитоп.
Активность in vitro антитела против Siglec-15 нейтрализации биологической активности Siglec-15 можно определять, например, по активности ингибирования дифференцировки клеток со сверхэкспрессией Siglec-15 в остеокласты. Например, антитело против Siglec-15 добавляют к клеткам RAW 264.7 или RAW 264, которые представляют собой линию клеток, происходящих из моноцитов мыши, в различных концентрациях, и можно определять активность ингибирования дифференцировки в остеокласты при стимуляции RANKL или TNF-α. Кроме того, антитело против Siglec-15 добавляют к первичной культуре клеток, происходящих из костного мозга, в различных концентрациях, и можно определять активность ингибирования дифференцировки в остеокласты при стимуляции RANKL, TNF-α, или активным витамином D3. Кроме того, антитело против Siglec-15 добавляют к нормальным клеткам-предшественникам остеокластов человека (Normal Human Natural Osteoclast Precursor Cells, доступным из Sanko Junyaku Co., Ltd., Кат. No. 2T-110) в различных концентрациях, и можно определять активность ингибирования дифференцировки в остеокласты при стимуляции RANKL или M-CSF. Такой ингибирующий эффект на дифференцировку остеокластов можно определять с использованием ингибирования активности устойчивой к тартрату кислой фосфатазы (TRAP) остеокластов в качестве показателя. Кроме того, ингибирующий эффект на дифференцировку остеокластов можно определять также с использованием ингибирования образования TRAP-положительных многоядерных остеокластов, т.е., ингибирования слияния клеток остеокластов в качестве показателя. Кроме того, при экспериментальном использовании пит-анализа (Takada et al., Bone and Mineral, (1992) 17, 347-359) с использованием клеток, полученных из бедренной кости и/или большой берцовой кости, активность in vitro ингибирования резорбции кости остеокластами можно определять добавлением антитела против Siglec-15 к клеткам, полученным из бедренной кости и/или большой берцовой кости в различных концентрациях, и наблюдением формирования пит-клеток на слое дентина. В качестве системы для определения in vitro активности ингибирования резорбции кости остеокластами, возможно также использование планшета, покрытого коллагеном человека, конъюгированным с европием. Терапевтический или профилактический эффект in vivo антитела против Siglec-15 на метаболизм костной ткани с использованием экспериментального животного можно подтверждать введением антитела против Siglec-15 в модели остеопороза на животных или трансгенному животному со сверхэкспрессией Siglec-15 и измерением каких-либо изменений в остеокластах.
Полученное таким образом антитело, нейтрализующее биологическую активность Siglec-15 является применимым в качестве лекарственного средства, в частности, в качестве фармацевтической композиции для лечения или профилактики патологического метаболизма костной ткани, такого как остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость, или в качестве антитела для иммунологической диагностики такого заболевания.
При лечении ревматоидного артрита (RA) главной проблемой является потеря костной массы, сопровождающая возникновение заболевания. Опубликовано, что в этой потере костной массы, сопровождающей RA, остеокласты играют первичную роль. Цитокины считают наиболее важными для индукции (дифференцировки и созревания) и активации остеокластов, и причиной разрушения кости при RA являются RANKL и TNF-α (Romas E. et al., Bone 30, pp. 340-346, 2002). OCIF/OPG, который представляет собой рецептор-ловушку для RANKL, может ингибировать образование остеокластов, индуцированное RANKL, но не ингибирует образование остеокластов индуцированное TNF-α. С другой стороны, антитело против Siglec-15 по изобретению эффективно ингибирует образование остеокластов, индуцированное как RANKL, так и TNF-α. Таким образом, ожидают, что антитело против Siglec-15 по изобретению может ингибировать потерю костной массы и разрушение кости, индуцированное TNF-α при RA или тому подобное, более сильно, чем блокатор RANKL (OCIF/OPG, антитело против RANKL, или тому подобное).
В качестве одного примера, для лечения или профилактики патологического метаболизма костной ткани, антитело против Siglec-15 можно вводить отдельно или в сочетании по меньшей мере с одним другим лекарственным средством против относящегося к костной ткани заболевания. В качестве другого примера, антитело против Siglec-15 можно вводить в сочетании с терапевтически эффективным количеством лекарственного средства против патологического метаболизма костной ткани. Лекарственные средства, которые можно вводить в сочетании с антителом против Siglec-15, включают в качестве неограничивающих примеров: бисфосфонаты (например, алендронат, этидронат, ибандронат, инкадронат, памидронат, ризедронат и золедронат), активный витамин D3, кальцитонин и его производные, гормоны, такие как эстрадиол, SERM (избирательные модуляторы рецепторов эстрогенов), иприфлавон, витамин K2 (менатетренон), препараты кальция, PTH (паратиреоидный гормон), нестероидные противовоспалительные средства (например, целекоксиб и рофекоксиб), растворимые рецепторы TNF (например, этанерцепт), антитела против TNF-α или функциональные фрагменты антител (например, инфликсимаб), антитела против PTHrP (родственного паратиреоидному гормону белка) или функциональные фрагменты антител, антагонисты рецептора IL-1 (например, анакинра), антитела против рецептора IL-6 или функциональные фрагменты антител (например, тоцилизумаб), антитела против RANKL или функциональные фрагменты антител (например, деносумаб) и OCIF (фактор ингибирования остеокластогенеза). В зависимости от состояния патологического метаболизма костной ткани или предназначенной степени лечения и/или профилактики, можно вводить два или три, или более типов лекарственных средств, и эти лекарственные средства можно поставлять все вместе посредством включения их в один и тот же препарат. Эти лекарственные средства и антитело против Siglec-15 можно поставлять все вместе посредством включения их в один и тот же препарат. Кроме того, лекарственные средства можно поставлять все вместе посредством включения их в один и тот же набор, предназначенный для использования для лечения и/или профилактики. Кроме того, эти лекарственные средства и антитело против Siglec-15 можно поставлять по отдельности. В случае введения при генотерапии, ген белкового лекарственного средства против заболевания костной ткани и ген антитела против Siglec-15 можно встраивать ниже одной и той же промоторной области или различных промоторных областей, и их можно вводить в один и тот же вектор или различные векторы.
Конъюгацией лекарственного средства против заболевания костной ткани с антителом против Siglec-15 или его фрагментом, можно получить нацеленный конъюгат лекарственного средства, как описано в M. C. Garnet «Targeted drug conjugates: principles and progress», Advanced Drug Delivery Reviews, (2001) 53, 171-216. Для достижения этой цели можно применять любой фрагмент антитела, отличный от молекулы антитела, при условии, что он полностью не теряет способности узнавать остеокласты, и примеры этого включают такие фрагменты, как Fab, F(ab')2 и Fv. По изобретению, антитело и фрагмент можно применять одинаковым способом. Конъюгат, сформированный антителом против Siglec-15 или его фрагментом и лекарственным средством против заболевания костной ткани, может представлять собой любую из различных форм, описанных в M. C. Garnet «Targeted drug conjugates: principles and progress». Advanced Drug Delivery Reviews, (2001) 53, 171-216, G. T. Hermanson «Bioconjugate Techniques» Academic Press, California (1996), Putnam and J. Kopecek 44 Polymer Conjugates with Anticancer Activity» Advances in Polymer Science (1995) 122, 55-123 и тому подобное. То есть, в качестве примера можно привести конъюгат, в котором антитело против Siglec-15 и лекарственное средство против заболевания костной ткани конъюгированы друг с другом химически и напрямую или через спейсер, такой как олигопептид, и конъюгат, сформированный посредством подходящего носителя для лекарственного средства. Примеры носителя для лекарственного средства включают липосому и водорастворимый полимер. Более конкретные примеры конъюгата, сформированного посредством такого носителя для лекарственного средства, включают конъюгат, в котором антитело и лекарственное средство против заболевания костной ткани включены в липосому, и липосома и антитело конъюгированы друг с другом, и конъюгат, в котором лекарственное средство против заболевания костной ткани конъюгировано с водорастворимым полимером (соединением с молекулярной массой от приблизительно 1000 до 100000) химически и напрямую или через спейсер, такой как олигопептид, и антитело конъюгировано с водорастворимым полимером. Конъюгацию антитела (или его фрагмента) с лекарственным средством против заболевания костной ткани или с носителем для лекарственного средства, таким как липосома или водорастворимый полимер, можно осуществлять способом, известным специалистам в данной области, таким как способ, описанный в G. T. Hermanson «Bioconjugate Techniques» Academic Press. California (1996), Putnam and J. Kopecek «Polymer Conjugates with Anticancer Activity» Advances in Polymer Science (1995) 122, 55-123. Включение лекарственного средства против заболевания костной ткани в липосому можно осуществлять способом, известным специалистам в данной области, таким как способ, описанный в D. D. Lasic «Liposomes: From Physics to Applications» Elsevier Science Publishers В. V., Amsterdam (1993) или тому подобное. Конъюгацию лекарственного средства против заболевания костной ткани с водорастворимым полимером можно осуществлять способом, известным специалистам в данной области, таким как способ, описанный в D. Putnam and J. Kopecek «Polymer Conjugates with Anticancer Activity» Advances in Polymer Science (1995) 122, 55-123. Конъюгат антитела (или его фрагмента) и белкового лекарственного средства против заболевания костной ткани (или его фрагмента) можно получать способами, известными специалистам в данной области, посредством генной инженерии, отличной от вышеупомянутого способа.
Изобретение относится также к фармацевтической композиции, содержащей терапевтически и/или профилактически эффективное количество антитела против Siglec-15 и фармацевтически приемлемый разбавитель, носитель, солюбилизирующее средство, эмульгатор, консервант и/или адъювант.
Изобретение относится также к фармацевтической композиции, содержащей терапевтически и/или профилактически эффективное количество антитела против Siglec-15, терапевтически и/или профилактически эффективное количество по меньшей мере одного лекарственного средства против заболевания костной ткани и фармацевтически приемлемые разбавитель, носитель, солюбилизирующее средство, эмульгатор, консервант, и/или адъювант. Лекарственные средства против заболевания костной ткани включают в качестве неограничивающих примеров, бисфосфонаты (например, алендронат, этидронат, ибандронат, инкадронат, памидронат, ризедронат, и золедронат), активный витамин D3, кальцитонин и его производные, гормоны, такие как эстрадиол, SERM (избирательные модуляторы рецепторов эстрогенов), иприфлавон, витамин K2 (менатетренон), препараты кальция, PTH (паратиреоидный гормон), нестероидные противовоспалительные средства (например, целекоксиб и рофекоксиб), растворимые рецепторы TNF (например, этанерцепт), антитела против TNF-α или функциональные фрагменты антител (например, инфликсимаб), антитела против PTHrP (родственного паратиреоидному гормону белка) или функциональные фрагменты антител, антагонисты рецептора IL-1 (например, анакинра), антитела против рецептора IL-6 или функциональные фрагменты антител (например, тоцилизумаб), антитела против RANKL или функциональные фрагменты антител (например, деносумаб) и OCIF (фактор ингибирования остеокластогенеза).
Вещество, подлежащее использованию в препарате, пригодном для фармацевтической композиции по изобретению, предпочтительно, является нетоксичным для индивидуума, которому вводят фармацевтическую композицию, в отношении дозы и концентрации.
Фармацевтическая композиция по изобретению может содержать вещество для фармацевтического применения, способное изменять или поддерживать ее pH, осмотическое давление, вязкость, прозрачность, цвет, изотоничность, асептические условия, стабильность, растворимость, скорость высвобождения, скорость всасывания и проходимость. Такие вещества для фармацевтического применения включают в качестве неограничивающих примеров, аминокислоты, такие как глицин, аланин, глутамин, аспарагин, аргинин и лизин; противомикробные средства; антиоксиданты, такие как аскорбиновая кислота, сульфат натрия, и гидросульфит натрия; буферы, такие как фосфатные, цитратные, боратные буферы, гидрокарбонат натрия и растворы Tris-HCl; наполнители, такие как маннит и глицин; хелатирующие средства, такие как этилендиаминтетраацетат (ЭДТА); комплексообразующие средства, такие как кофеин, поливинилпирролидин, β-циклодекстрин и гидроксипропил-β-циклодекстрин; расширители, такие как глюкоза, манноза и декстрин; другие углеводы, такие как моносахариды и дисахариды; окрашивающие средства; вкусовые вещества; разбавители; эмульгаторы; гидрофильные полимеры, такие как поливинилпирролидин; консерванты, такие как полипептиды с низкой молекулярной массой, солеобразующие противоионы, хлорид бензалкония, бензойная кислота, салициловая кислота, тимеросал, фенетиловый спирт, метилпарабен, пропилпарабен, хлоргексидин, сорбиновая кислота и пероксид водорода; растворители, такие как глицерин, пропиленгликоль и полиэтиленгликоль; сахарные спирты такие как маннит и сорбит; суспендирующие вещества; поверхностно-активные вещества, такие как сложный эфир сорбитана, полисорбаты, включая полисорбат 20 и полисорбат 80, Тритон, трометамин, лецитин и холестерин; увеличивающие стабильность средства, такие как сахароза и сорбит; увеличивающие эластичность средства, такие как хлорид натрия, хлорид калия, и маннит и сорбит; транспортные средства; наполнители; и/или фармацевтические адъюванты. Количество этих веществ, добавляемых для фармацевтического применения, предпочтительно, от 0,01 до 100 раз превышает, особенно предпочтительно, от 0,1 до 10 раз превышает массу антитела против Siglec-15. Специалисты в данной области могут соответствующим образом определять предпочтительный состав фармацевтической композиции в препарате в зависимости от заболевания, для которого применяют композицию, способа введения, подлежащего применению, или тому подобное.
Наполнитель или носитель в фармацевтической композиции может присутствовать в форме жидкости или твердого вещества. Подходящий наполнитель или носитель может представлять собой пригодную для инъекций воду, физиологический солевой раствор, искусственную спинномозговую жидкость или другое вещество, общепринятое для парентерального введения. Кроме того, нейтральный физиологический солевой раствор или физиологический солевой раствор, содержащий сывороточный альбумин, также можно использовать в качестве носителя. Фармацевтическая композиция может содержать Tris-буфер с pH 7,0-8,5, ацетатный буфер с pH 4,0-5,5 или цитратный буфер с pH 3,0-6,2. Кроме того, такой буфер можно дополнять сорбитом или другим соединением. Примеры фармацевтической композиции по изобретению включают фармацевтическую композицию, содержащую антитело против Siglec-15, и фармацевтическую композицию, содержащую антитело против Siglec-15 и по меньшей мере одно лекарственное средство против заболевания костной ткани. Фармацевтическую композицию по изобретению получают в форме лиофилизированного продукта или жидкости в качестве лекарственного средства, обладающего выбранным составом и требуемой чистотой. Фармацевтическую композицию, содержащую антитело против Siglec-15, и фармацевтическую композицию, содержащую антитело против Siglec-15 и по меньшей мере одно лекарственное средство против патологического метаболизма костной ткани, можно составлять также в лиофилизированный продукт с использованием подходящего наполнителя, такого как сахароза.
Фармацевтическую композицию по изобретению можно получать для парентерального введения или для желудочно-кишечной абсорбции посредством перорального введения. Состав и концентрацию препарата можно определять в зависимости от способа введения. Чем выше аффинность антитела против Siglec-15, содержащегося в фармацевтической композиции по изобретению, составляет для Siglec-15, то есть, чем ниже его константа диссоциации (значение Kd) для Siglec-15, тем больше антитело против Siglec-15 может проявлять свою эффективность в качестве лекарственного средства, даже при уменьшении дозы для человека. Таким образом, дозу фармацевтической композиции по изобретению для человека также можно определять на основании этих соображений. Что касается дозы, в случае, когда антитело человека против Siglec-15 вводят человеку, антитело можно вводить в дозе от приблизительно 0,1 до 100 мг/кг один раз в одни - 180 суток.
Примеры лекарственной формы фармацевтической композиции по изобретению включают инъекции, включая инфузии, суппозитории, средства для трансназального введения, средства для подъязычного введения и абсорбенты для чрескожного введения.
Примеры
Далее в настоящем документе изобретение описано более конкретно по отношению к примерам, однако, изобретение не является ограниченным ими. Следует отметить, что соответствующие операции, относящиеся к генным манипуляциям, в следующих примерах проводили способами, описанными в «Molecular Cloning» (написанном Sambrook, J., Fritsch, E. F. и Maniatis, Т., опубликованном в Cold Spring Harbor Laboratory Press в 1989), или, в случае использования коммерчески доступных реагентов или наборов, их используют согласно приложенным к ним протоколам, если нет иных указаний.
Пример 1
Получение конструкции, экспрессирующей растворимый белок Siglec-15 мыши
Частичная нуклеотидная последовательность, кодирующая внеклеточный домен белка Siglec-15 мыши, представлена как SEQ ID NO:5 в разделе “Список последовательностей”, и его аминокислотная последовательность представлена как SEQ ID NO:6 в разделе “Список последовательностей”. С применением такой частичной последовательности растворимый белок Siglec-15 мыши можно получать в культуральном супернатанте клеток животных или тому подобное.
a) Амплификация гена растворимого Siglec-15 мыши посредством ПЦР
В качестве праймеров для амплификации кДНК внеклеточного домена Siglec-15 мыши посредством ПЦР, олигонуклеотид, обладающий последовательностью 5'-ggggacaagt ttgtacaaaa aagcaggctt caccATGGAG GGGTCCCTCC AACTC-3' (mSiglec-15-ECD-F: SEQ ID NO:7 в разделе “Список последовательностей”); и олигонуклеотид, обладающий последовательностью 5'-ggggaccact ttgtacaaga aagctgggtc TCCGGGGGCG CCGTGGAAGC GGAAC-3' (mSiglec-15-ECD-R: SEQ ID NO:8 в разделе “Список последовательностей”), синтезировали общепринятым способом. К тому же, эти праймеры сконструированы как праймеры для амплификации для получения исходного клона gateway, так что последовательность attB1 добавлена к mSiglec-15-ECD-F, и последовательность attB2 добавлена к mSiglec-15-ECD-R. ПЦР проводили с использованием этого сочетания праймеров и полинуклеотида, содержащего последовательность открытой рамки считывания Siglec-15 мыши в качестве матрицы, общепринятым способом. Условия для термоциклера устанавливали следующим образом: после нагревания при 94°C в течение 5 минут, температурный цикл «94°С в течение 0,5 минуты, 55°C в течение 0,5 минуты и 68°C в течение 1,5 минут» повторяли 15 раз, с последующим нагреванием при 68°C в течение 5 минут и инкубацией при 4°C.
b) Получение исходного клона реакцией Gateway BP
Исходный клон, в который кДНК внеклеточного домена Siglec-15 мыши встраивали посредством способа Gateway (Invitrogen, Inc.) с использованием системы сайт-специфической рекомбинации фага лямбда, получали следующим способом. Сначала проводили реакцию BP с использованием BP Clonase между продуктом ПЦР, обладающим последовательностью attB на обоих концах, полученным в a), и pDNOR221 (изготовленной в Invitrogen, Inc.), которая представляет собой донорный вектор, обладающий последовательностью attP. С использованием этой реакционной смеси трансформировали Escherichia coli DH10B, проводили ПЦР колоний для поиска устойчивых к лекарственному средству клонов, и подтверждали размер вставок. Затем для клона, в котором подтверждено наличие вставки правильного размера, проводили анализ секвенированием всей интересующей последовательности ДНК. В результате получен исходный клон, полностью идентичный намеченной нуклеотидной последовательности (SEQ ID NO:5 в разделе “Список последовательностей”), кодирующей внеклеточный домен белка Siglec-15 мыши.
c) Получение экспрессирующего клона реакцией Gateway LR
Экспрессирующий клон, в который кДНК внеклеточного домена Siglec-15 мыши встраивали посредством способа Gateway (Invitrogen, Inc.) с использованием системы сайт-специфической рекомбинации фага лямбда, получали следующим способом. Исходный клон, полученный в b), содержит вставку, обладающую последовательностью attL на обоих концах. Проводили реакцию LR с использованием LR Clonase между этим исходным клоном и двумя типами векторов назначения, обладающих последовательностью attR. К тому же, в качестве векторов назначения использовали два типа векторов назначения: pDONM, сконструированный так, что метка эпитопа V5 и метка 6×His добавлены к С-концу вставки; и phIgFc, сконструированный так, что метка Fc человека добавлена к С-концу вставки. С использованием реакционной смеси, полученной посредством реакции LR, трансформировали Escherichia coli DH10B, и проводили ПЦР колоний для получения устойчивых к лекарственному средству клонов, и подтверждали размер вставок. Затем, для клона, в котором подтверждено наличие вставки правильного размера, проводили анализ секвенированием обоих концов от стороны вставки до стороны вектора.
Последовательности праймеров для анализа секвенированием:
5'-tgcgtgaagg tgcagggcag-3' (mSiglec-15-ECD-seq-upstm: SEQ ID NO:9 в разделе “Список последовательностей”) и
5'-cctcgcctgg tcgggtc-3' (mSiglec-15-ECD-seq-dnstm: SEQ ID NO:10 в разделе “Список последовательностей”)
В результате анализа секвенированием получены экспрессирующие клоны (растворимый Siglec-15 мыши/pDONM и растворимый Siglec-15 мыши/phIgFc), в которых произошла правильная рекомбинация как для pDONM, так и для phIgFc, соответственно. Посредством трансфекции клеток животных или тому подобное растворимым Siglec-15 мыши/pDONM, мРНК, обладающую последовательностью оснований SEQ ID NO:11 в разделе “Список последовательностей”, транскрибируют и транслируют в белок (Siglec-15-His мыши), обладающий аминокислотной последовательностью SEQ ID NO:12 в разделе “Список последовательностей”. Кроме того, посредством трансфекции клеток животных или тому подобное растворимым Siglec-15 мыши/phIgFc, мРНК, обладающую последовательностью оснований SEQ ID NO:13 в разделе “Список последовательностей”, транскрибируют и транслируют в белок (Siglec-15-Fc мыши), обладающий аминокислотной последовательностью SEQ ID NO:14 в разделе “Список последовательностей”.
Пример 2
Крупномасштабное получение культурального раствора, содержащего растворимый белок Siglec-15 мыши, с использованием клеток 293-F
Два типа векторов экспрессии (растворимый Siglec-15 мыши/pDONM и растворимый Siglec-15 мыши/phIgFc), полученные в примере 1, получали в количестве приблизительно 5 мг, соответственно. К тому же, при очистке плазмид из Escherichia coli после крупномасштабного культивирования использовали набор для выделения плазмид в гига-количествах Invitrogen PureLink HiPure (изготовленный в Invitrogen, Inc.). Полученную таким образом плазмиду смешивали с Opti-MEM (изготовленной в Invitrogen, Inc.), полученную смесь стерилизовали фильтрацией, в нее добавляли 10 мл реагента для трансфекции 293 фектина (изготовленного в Invitrogen, Inc.), и полученную смесь инкубировали при комнатной температуре в течение 20 минут. Эту смесь добавляли к клеткам Freestyle 293-F (изготовленным в Invitrogen, Inc.), культивированным в колбах Эрленмейера, так чтобы плотность клеток достигла 1,1×106 клеток/мл ×5 л (1 л/колбу ×5 колб) в среде для экспрессии FreeStyle 293 (изготовленной в Invitrogen, Inc.). После того, как клетки подвергали культивированию с встряхиванием (125 оборотов/мин) при концентрации CO2 8,0% в течение 96 часов (4 суток) при 37°C, культуральный раствор собирали и центрифугировали для получения культурального супернатанта. Считают, что в полученном таким образом культуральном супернатанте экспрессируются белок, в котором метка эпитоп V5 и метка 6х His добавлены к C-концу внеклеточного домена Siglec-15 мыши (Siglec-15-His мыши) и белок, в котором метка Fc человека добавлена к C-концу внеклеточного домена Siglec-15 мыши (Siglec-15-Fc мыши), соответственно.
Пример 3
Очистка Siglec-15-His мыши
a) Хроматография на колонке HisTrap HP
К 2 л культурального раствора экспрессирующих Siglec-15-His мыши клеток 293F, полученных в примере 2, добавляли 225 мл 10 × буфера (500 мМ Tris, 1,5 M NaCl, 200 мМ имидазол, pH 8,0), и полученную смесь хорошо перемешивали и фильтровали через фильтр Sterivex-GV (изготовленный в Millipore Co., Ltd.). Этот культуральный раствор наносили со скоростью потока 2 мл/мин на колонку, содержащую три 5 мл колонки HisTrap HP (изготовленные в Amersham Biosciences. Inc.), соединенные последовательно и предварительно обработанные удаляющим пирогены средством PyroCLEAN (изготовленным в ALerCHEK, Inc.) и промывали дистиллированной водой для инъекций. После промывки колонки с помощью 60 мл 50 мМ Tris-HCl буфера (pH 8,0), содержащего 300 мМ NaCl при скорости потока 1 мл/мин, белок, адсорбированный на колонке, элюировали с помощью 50 мл 50 мМ Tris-HCl буфера (pH 8,0), содержащего 300 мМ NaCl и 500 мМ имидазол, при скорости потока 1 мл/мин. Элюат фракционировали по 1 мл на фракцию в пробирки mini-sorp (изготовленные в Nunc, Inc.), в которые предварительно добавляли по 10 мкл 10% Tween 20. После этого приблизительно 20 мл раствора, полученного объединением фракций (фракции 14-20) и содержащего белок, концентрировали до 2,5 мл с помощью центрифугируемого мембранного концентратора Amicon Ultra-15 (изготовленного в Millipore Co., Ltd.), концентрат наносили на обессоливающую колонку PD-10 (изготовленную в Amersham Biosciences, Inc.), которую предварительно уравновешивали фосфатно-солевым буфером, содержащим 0,01% Tween 20 (T-PBS), с последующей элюцией T-PBS, в результате чего получали 3,5 мл образца, растворитель в котором заменен на T-PBS.
b) Хроматография на колонке Resource Q
К 3,5 мл образца, который очищали хроматографией на колонке HisTrap HP, и растворитель в котором заменяли на T-PBS, добавляли 22,5 мл 50 мМ Tris-HCl буфера (pH 7,5), содержащего 0,1% CHAPS, и полученную смесь перемешивали. Затем смесь центрифугировали при 4°C в течение 30 минут при 3000 об./мин и преципитат удаляли. После фильтрации полученного супернатанта через фильтр Millex-GV (изготовленный в Millipore Co., Ltd.), фильтрат наносили при скорости потока 1 мл/мин на колонку Resource Q 6 мл (изготовленную в Amersham Biosciences, Inc.), которую предварительно уравновешивали 50 мМ Tris-HCl буфером (pH 7,5), содержащим 0,1% CHAPS. После этого колонку промывали этим буфером при скорости потока 1 мл/мин и собирали фракцию белка, не адсорбированного на колонке. Белок, адсорбированный на колонке, элюировали буфером 50 мМ Tris-HCl (pH 7,5), содержащим 0,1% CHAPS и 1 M NaCl, при скорости потока 1 мл/мин. Затем 26,5 мл из фракции, не адсорбированной на колонке, концентрировали до 2,0 мл с помощью центрифугируемого мембранного концентратора Amicon Ultra-15 (изготовленного в Millipore Co., Ltd.), концентрат центрифугировали при 4°C в течение 10 минут при 3000 об./мин, и преципитат удаляли. Супернатант после центрифугирования подвергали криоконсервации при -80°C до использования. Вышеупомянутую процедуру очистки (хроматографию на колонке HisTrap HP и хроматографию на колонке Resource Q) проводили, повторяя ее дважды.
с) Детекция и анализ чистоты очищенного Siglec-15-His мыши
С использованием образца, полученного посредством вышеупомянутой процедуры очистки (хроматографии на колонке HisTrap HP и хроматографии на колонке Resource Q), проводили SDS-полиакриламидный электрофорез в восстанавливающих условиях и окрашивание серебром. То есть, к 5 мкл из каждого образца, очищенного на соответствующих стадиях очистки, добавляли равное количество раствора для обработки с SDS, и полученную смесь термически обрабатывали при 95°C в течение 10 минут. По 0,3 мкл из каждого из термически обработанных образцов использовали для SDS-полиакриламидного электрофореза. В качестве геля для электрофореза использовали 8-25% полиакриламидный градиентный гель (изготовленный в Amersham Biosciences, Inc.), и электрофорез проводили с использованием PhastSystem (изготовленной в Amersham Biosciences, Inc.). Кроме того, в качестве маркеров молекулярной массы использовали маркеры молекулярной массы Rainbow (изготовленные в Amersham Biosciences, Inc.). После завершения электрофореза проводили окрашивание серебром с использованием набора PhastGel Silver (изготовленного в Amersham Biosciences, Inc.) и PhastSystem. Результаты показаны на фиг. 1. Показано, что белок, обладающий молекулярной массой приблизительно 35 кДа (Siglec-15-His мыши), являлся эффективно очищенным и концентрированным во фракции белка, не адсорбированной на колонке Resource Q.
Электрофорез проводили в таких же условиях, за исключением того, что маркеры для Вестерн-блоттинга ECL DualVue (изготовленные в Amersham Biosciences, Inc.) использовали в качестве маркеров молекулярной массы, и белок из геля переносили (проводили блоттинг) на PVDF мембрану (Hybond-P, изготовленный в Amersham Biosciences, Inc.) с использованием набора для полусухого переноса PhastTransfer (изготовленного в Amersham Biosciences, Inc.) и PhastSystem. Эту PVDF мембрану осторожно встряхивали в 10 мл блокирующего средства (BlockAce, изготовленного в Snow Brand Milk Products, Co., Ltd.), содержащего 0,1% Tween 20, при комнатной температуре в течение 1 часа. К этому блокирующему раствору добавляли 10 мкл HRP с S-белком (изготовленной в Amersham Biosciences, Inc.) и добавляли 10 мкл антитела против V5-HRP (моноклонального антитела против Pk-TAG-HRP, изготовленного в Acris Antibodies), и мембрану в растворе осторожно встряхивали при комнатной температуре в течение дополнительного 1 часа. PVDF мембрану промывали 4 раза посредством осторожного встряхивания в 50 мл фосфатно-солевого буфера (PBS), содержащего 0,01% Tween 20, в течение 5 минут. После промывки PVDF мембрану обрабатывали согласно протоколу, прилагаемому к набору для детекции ECL (изготовленному в Amersham Biosciences, Inc.) для проявления окраски в полосе белка, и появившуюся окраску детектировали с использованием мини-камеры для ECL (изготовленной в Amersham Biosciences, Inc.) и пленки Polaroid (Polapan 3200B, изготовленной в Polaroid, Inc.). Результаты показаны на фиг. 2. Также по этим результатам можно подтвердить, что белок, обладающий молекулярной массой приблизительно 35 кДа (Siglec-15-His мыши) и узнаваемый антителом против V5-HRP, являлся эффективно очищенным и концентрированным во фракции белка, не адсорбированной на колонке Resource Q.
d) Измерение концентрации белка очищенного Siglec-15-His мыши
Для очищенного Siglec-15-His мыши (фракции белка, не адсорбированной на колонке Resource Q), концентрацию белка измеряли с помощью набора для анализа белка DC (изготовленного в Bio-Rad Laboratories, Inc.) с использованием бычьего сывороточного альбумина в качестве стандартного образца. Как показано в таблице 1, всего 1,66 мг очищенного белка Siglec-15-His мыши получили посредством проведения процедуры очистки дважды.
Пример 4
Очистка Siglec-15-Fc мыши
a) Хроматография на колонке HiTrap c белком A
1,8 л культурального раствора экспрессирующих Siglec-15-Fc мыши клеток 293F, полученных в примере 2, фильтровали через фильтр Sterivex-GV (изготовленный в Millipore Co., Ltd.), и затем фильтрат наносили на 5 мл колонку HiTrap c белком A (изготовленную в Amersham Biosciences, Inc.), предварительно уравновешенную с помощью PBS Дульбекко (D-PBS, изготовленного в Invitrogen, Inc.) при скорости потока 5 мл/мин. После промывки колонки с помощью D-PBS при скорости потока 5 мл/мин, белок, адсорбированный на колонке, элюировали с помощью 50 мл буфера 0,1 M цитрата натрия (pH 3,0) при скорости потока 5 мл/мин. Элюат фракционировали по 5 мл на фракцию в пробирки mini-sorp (изготовленные в Nunc, Inc.), и немедленно после этого, в них добавляли 1,3 мл 1 M Tris для нейтрализации элюата. После этого раствор, полученный объединением фракций (фракции 1 и 2), в которых детектировали белок, концентрировали до 2,5 мл с помощью центрифугируемого мембранного концентратора Amicon Ultra-15 (изготовленного в Millipore Co., Ltd.), концентрат наносили на обессоливающую колонку PD-10 (изготовленную в Amersham Biosciences, Inc.), предварительно уравновешенную физиологическим солевым раствором для инъекций Otsuka (TO-SS, изготовленным в Otsuka Pharmaceutical Co., Ltd.), содержащим 0,01% Tween 20, с последующей элюцией TO-SS, в результате чего получали 3,5 мл образца, растворитель в котором заменен на TO-SS. Этот образец подвергали криоконсервации при -80°C до использования. С использованием 2,9 л культурального раствора клеток 293F, такую же процедуру очистки проводили, повторяя ее еще раз.
b) Детекция и анализ чистоты очищенного Siglec-15-Fc мыши
С использованием образца, полученного посредством вышеупомянутой процедуры очистки, проводили SDS-полиакриламидный электрофорез в восстанавливающих условиях и окрашивание серебром. То есть, к 5 мкл из каждого образца, очищенного на соответствующих стадиях очистки, добавляли равное количество раствора для обработки с SDS и полученную смесь термически обрабатывали при 95°C в течение 10 минут. По 0,3 мкл из образца, полученного разведением каждого из термически обработанных образцов 1/300 или 1/900 с помощью разведенного до половины концентрации раствора для обработки с SDS, использовали для SDS-полиакриламидного электрофореза. Электрофорез и окрашивание серебром проводили таким же способом, как анализ чистоты Siglec-15-His мыши, описанный в c) из примера 3. Результаты показаны на фиг. 3 вместе с результатами оценки предварительных условий очистки в малом масштабе (pH применяемого культурального раствора составлял 8,9 или 7,0). Показано, что белок, обладающий молекулярной массой приблизительно 55 кДа (Siglec-15-Fc мыши) являлся эффективно очищенным и концентрированным во фракции белка, элюированной с колонки HiTrap с белком A.
с) Измерение концентрации белка очищенного Siglec-15-Fc мыши
Для очищенного Siglec-15-Fc мыши (фракции белка, элюированной с обессоливающей колонки PD-10), концентрацию белка измеряли с помощью набора для анализа белка DC (изготовленного в Bio-Rad Laboratories, Inc.) с использованием бычьего сывороточного альбумина в качестве стандартного образца. Как показано в таблице 2, всего 92 мг очищенного белка Siglec-15-Fc мыши получили посредством проведения процедуры очистки дважды.
Пример 5
Получение гибридомы крысы, продуцирующей моноклональное антитело крысы против Siglec-15 мыши
a) Получение антигена
Белок Siglec-15-His мыши, полученный в примере 3, получали при 100 мкг/0,5 мл, и к нему добавляли равное количество адъюванта, и получали эмульсию с использованием стеклянного шприца. В качестве адъюванта полный адъювант Фрейнда (FCA, изготовленный в Difco Laboratories, Inc.) использовали только для первой иммунизации, и неполный адъювант Фрейнда (FICA, изготовленный в Difco Laboratories, Inc.) использовали для второй и последующих иммунизаций.
b) Иммунизация крыс
Четырех крыс (Wistar, самки, в возрасте 6 недель, закуплены из CLEA Japan, Inc.) использовали в качестве иммунизируемых животных. Эмульсию, полученную в a), инъецировали подкожно и внутрикожно с использованием иглы для инъекций 27 G, так что количество антигена составляло 50 мкг на крысу. Иммунизацию проводили всего 4 раза каждые 7 суток после первой иммунизации. Небольшое количество (200 мкл) крови отбирали из хвостовой вены через 7 суток после даты 4-й иммунизации, и получали антисыворотку. Чтобы подтвердить титр антитела в антисыворотке, проводили ELISA с использованием белка Siglec-15-His мыши, использованного в качестве антигена, белка Siglec-15-Fc мыши, полученного в примере 4, или бычьего сывороточного альбумина (BSA), каждый из которых являлся иммобилизованным. В результате, реактивность с белком Siglec-15-His мыши и белком Siglec-15-Fc мыши наблюдали у всех четырех крыс (крысы No. 1-4) С другой стороны, реактивности с BSA не наблюдали. По этим результатам подтвердили, что титр антитела в сыворотке каждой из иммунизированных крыс увеличивался, и таким образом, крысу No. 2, для которой показали самый высокий титр антител, подвергали процедуре слияния клеток.
c) Слияние клеток
Слияние клеток проводили общепринятым способом слияния клеток селезенки мыши (крысы) с клетками миеломы. Цельную кровь отбирали из сердца крысы под эфирной анестезией, и крысу подвергали эвтаназии, и затем вырезали селезенку. Собранные клетки селезенки и клетки P3X63Ag8.653 (ATCC CRL 1580), представляющие собой клетки миеломы мыши, подвергали слиянию клеток с использованием полиэтиленгликоля (PEG). Полученные клетки рассевали в 96-луночный планшет, и к ним добавляли среду, содержащую гипоксантин (H), аминоптерин (A) и тимидин (T) (селективная среда HAT), и затем клетки культивировали в течение 7-10 суток. Культуральный супернатант собирали из 61 лунки, в которых подтвердили выживание гибридом, полученных слиянием клеток. Затем титр антитела оценивали посредством ELISA с использованием белка Siglec-15-His мыши, использованного в качестве антигена, белка Siglec-15-Fc мыши, полученного в примере 4, или BSA, каждый из которых являлся иммобилизованным, и проводили скрининг гибридом, продуцирующих моноклональное антитело против Siglec-15 мыши. По результатам скрининга выбрали 12 лунок, для которых показали высокий титр антител, и гибридомы, содержащиеся в лунках, подвергали процедуре клонирования.
d) Клонирование гибридом
Для отобранных таким образом гибридом первую стадию клонирования проводили способом предельного разведения. После предельных разведений гибридомы культивировали в течение 2 недель, и титр антитела в культуральном супернатанте подтверждали ELISA с использованием белка Siglec-15-Fc мыши, полученный в примере 4 или BSA, каждый из которых являлся иммобилизованным. Для 11 клонов, для которых подтвердили, что они являются положительными клонами, проводили вторую стадию клонирования (таким же способом, как и первую стадию клонирования), в результате всего в конце получено 10 клонов гибридомы, продуцирующих моноклональное антитело против Siglec-15 мыши (#1A1, #3A1, #8A1, #24A1, #32A1, #34A1, #39A1, #40A1, #41B1, #61A1). К тому же, гибридома #32A1 депонирована в Международном Патентном Депозитарии Организмов Национального Института Развития Прикладной Науки и Технологий (расположенном в Central 6, 1-1-1 Higashi, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan) 28 августа 2008 г., и ей присвоен инвентарный номер FERM BP-10999 с наименованием anti-Siglec-15 Hybridoma #32A1.
Пример 6.
Получение моноклонального антитела крысы против Siglec-15 мыши
а) Получение асцитов “голых” мышей
мышей
Гибридомы, полученные в примере 5, культивировали с использованием среды TIL I (изготовленной в Immuno-biological Laboratories Co., Ltd.), среды, содержащей 10% FCS. Субкультивирование клеток проводили посредством способа, в котором культуральный раствор разводили приблизительно до одной четвертой каждые двое - трое суток с использованием временной точки, когда клетки вырастали до приблизительно 5×105 клеток/мл в качестве показателя. Каждую из культивированных таким образом гибридом внутрибрюшинно имплантировали “голой” мыши, которым предварительно внутрибрюшинно вводили пристан (0,2 мл/мышь), 1×107 клеток на мышь. При имплантации трех “голых” мышей использовали для каждого из 10 клонов гибридомы. После имплантации асциты собирали, когда наблюдали достаточное накопление асцитов, и объединяли с асцитами, собранными от двух других мышей после имплантации той же самой гибридомы, измеряли количество объединенных таким образом асцитов, и асциты подвергали криоконсервации до очистки антитела. Количества собранных асцитов для соответствующих гибридом обобщены в таблице 3.
b) Очистка антитела
Общее количество собранных асцитов подвергали очистке IgG с использованием 20 мл колонки с белком G (изготовленной в GE Healthcare, Co., Ltd.). Чистоту очищенных IgG анализировали анализом гель-фильтрации (хроматография на колонке Superdex 200), и некоторые из антител подвергали концентрированию центрифугированием через мембрану. То есть, среди очищенных моноклональных антител против Siglec-15 мыши, 9 типов антител, за исключением антитела #24A1, концентрировали до приблизительно одной шестой - одной восьмой исходного объема посредством центрифугирования антител при 3000 об./мин в течение 30-60 минут при 4°C с использованием центрифугируемого мембранного концентратора Amicon Ultra-15 (изготовленного в Millipore Co., Ltd.). Затем, для антитела #24A1 и других концентрированных 9 типов антител, концентрацию белка измеряли с помощью набора для анализа белка DC (изготовленного в Bio-Rad Laboratories, Inc.) с использованием бычьего сывороточного альбумина (BSA) в качестве стандартного образца. Посредством вышеупомянутого способа получали моноклональное антитело против Siglec-15 мыши.
Пример 7
Оценка свойства связывания моноклонального антитела крысы против Siglec-15 мыши с белком Siglec-15 мыши
Свойство связывания моноклонального антитела крысы против Siglec-15 мыши с белком Siglec-15 мыши оценивали способом ELISA. Белок Siglec-15-Fc мыши, полученный в примере 4, разводили до 5 мкг/мл буфером 0,1 M карбонатом натрия (pH 9,5), и полученный раствор добавляли в 96-луночный планшет (изготовленный в Nalge Nunc International, Inc., Кат. No. 430341) по 100 мкл/лунку. Затем планшет оставляли при комнатной температуре на 1 час, раствор удаляли, и буфер для промывки (фосфатно-солевой буфер, содержащий 0,05% Tween 20) добавляли по 300 мкл/лунку и затем удаляли. Затем эту процедуру промывки проводили еще раз, фосфатно-солевой буфер, содержащий 25% BlockAce (изготовленный в Dainippon Sumitomo Pharma Co., Ltd.) добавляли по 200 мкл/лунку, и планшет оставляли при комнатной температуре на 1 час, таким образом проводили блокирование. Жидкость удаляли, и планшет промывали дважды по 300 мкл/лунку буфера для промывки. Затем каждое из моноклональных антител крысы против Siglec-15 мыши, полученных в примере 6, или контрольное IgG крысы (изготовленное в R&D Systems, Inc.) разводили до конечной концентрации от 1,28 до 20000 нг/мл (серии 5-кратного разведения) буфером для ELISA (фосфатно-солевой буфер, содержащий 12,5% BlockAce и 0,05% Tween 20), и полученный раствор разведенного антитела добавляли в планшет по 100 мкл/лунку. Затем планшет оставляли при комнатной температуре на 1 час, жидкость удаляли, и планшет промывали 3 раза по 300 мкл/лунку буфера для промывки. Затем меченное HRP (пероксидазой хрена) антитело козы против IgG крысы (изготовленное в Beckman Coulter, Inc.), разведенное в 1000 раз буфером для ELISA, добавляли по 100 мкл/лунку, и планшет оставляли при комнатной температуре на 1 час. Жидкость удаляли, и планшет промывали 3 раза по 300 мкл/лунку буфера для промывки, и затем, с использованием набора для проявления окрашивания для пероксидазы (изготовленного в Sumitomo Bakelite Co., Ltd.), окрашивание проявляли согласно протоколу, сопровождающему набор. После проявления окрашивания оптическую плотность при 492 нм измеряли с использованием считывателя для микропланшетов (изготовленного в Nihon Molecular Devices Corporation). В результате подтвердили, что все из оцениваемых 10 тестируемых образцов моноклональных антител крысы против Siglec-15 мыши связывают белок Siglec-15 мыши зависимым от концентрации антител образом (Фиг. 4). С другой стороны, в случае контрольного IgG крысы, не наблюдали связывания с белком Siglec-15 мыши.
Пример 8
Получение неадгерентных клеток костного мозга мыши
Бедренную кость и большую берцовую кость вырезали у самца мыши ddY в возрасте 5-8 недель, и удаляли мягкие ткани. Оба конца бедренной кости или большой берцовой кости отрезали, и D-PBS инъецировали с использованием шприца с иглой 25 калибра для инъекций, чтобы вытолкнуть клетки костного мозга, которые собирали в пробирку для центрифугирования. Центрифугирование проводили при комнатной температуре в течение 5 минут при 100 g, и супернатант удаляли. К полученному осадку клеток добавляли 1 мл гемолитического буфера (Red Blood Cell Lysing Buffer, изготовленного в Sigma Co., Ltd.), чтобы суспендировать его, и полученную суспензию оставляли при комнатной температуре на 5 минут. К ней добавляли 20 мл D-PBS, и суспензию центрифугировали при комнатной температуре в течение 5 минут при 100 g, и супернатант удаляли. К полученному осадку клеток добавляли 10 мл среды MEM-α (изготовленной в Invitrogen, Inc.), содержащей 5 нг/мл M-CSF (изготовленной в R&D Systems, Inc.), и 10% эмбриональную бычью сыворотку (FBS), чтобы суспендировать его. Затем полученную суспензию пропускали через клеточное сито (40 мкм Nylon, изготовленный в BD Falcon) для удаления агрегатов. Полученные клетки переносили в 75 см2 T-флакон (для прикрепления адгерентных клеток) и культивировали в течение ночи в CO2 инкубаторе. После культивирования в течение ночи клетки, у которых не произошла адгезия к T-флакону, выделяли и использовали в качестве неадгерентных клеток костного мозга мыши.
Пример 9
Оценка биологической активности моноклонального антитела крысы против Siglec-15 мыши на основании теста образования остеокластов мыши
С использованием всех 10 тестируемых образцов моноклональных антител против Siglec-15 мыши, полученных в примере 6, оценивали эффект на дифференцировку остеокластов из неадгерентных клеток костного мозга мыши. Неадгерентные клетки костного мозга мыши, полученные способом из примера 8, получали при 1,5×105 клеток/мл в среде α-MEM, содержащей 10% FBS и 10 нг/мл M-CSF (изготовленный в R&D Systems, Inc.), и полученный препарат клеток рассевали в каждую лунку 96-луночного планшета в количестве 200 мкл, и клетки культивировали в течение 2 суток в CO2 инкубаторе. Старый культуральный раствор из 96-луночного планшета удаляли, и 100 мкл среды MEM-α добавляли в каждую лунку, 100 мкл среды MEM-α, содержащей 10% FBS, в которую добавляли RANKL человека (RANKL, изготовленный в Peprotech, Inc.) и M-CSF, добавляли для получения конечных концентраций 20 нг/мл и 10 нг/мл, соответственно. К культуральному раствору клеток каждое из моноклональных антител крысы против Siglec-15 мыши, полученных в примере 6, образец, полученный удалением азида натрия из коммерчески доступного контрольного IgG крысы (очищенный IgG крысы, изготовленный в R&D Systems, Inc.), или отдельно полученное поликлональное антитело кролика против Siglec-15 мыши (No. 3) добавляли в концентрации от 32 до 1000 нг/мл, и клетки культивировали в течение дополнительных 3 суток в CO2 инкубаторе. К тому же, поликлональное антитело (No. 3) представляет собой антитело, для которого уже подтверждено ингибирование образования остеокластов в экспериментальной системе, описанной в этом примере. После завершения культивирования активность устойчивой к тартрату кислой фосфатазы (TRAP) образованных остеокластов измеряли способом, описанным ниже. Культуральный раствор в каждой лунке 96-луночного планшета удаляли отсасыванием, и 50 мкл буфера 50 мМ цитрата натрия (pH 6,1), содержащего 1% Тритон X-100, добавляли в каждую лунку. Затем планшет встряхивали в течение 5 минут на встряхивателе для планшетов для лизиса клеток. В каждую лунку добавляли 50 мкл раствора субстрата (50 мМ буфера цитрата натрия (pH 6,1), содержащего 5 мг/мл п-нитрофенилфосфата и 0,46% тартрата натрия), и планшет инкубировали при комнатной температуре в течение 5 минут. После инкубации 50 мкл 1 н раствора гидроксида натрия добавляли в каждую лунку 96-луночного планшета для остановки ферментативной реакции. После остановки ферментативной реакции измеряли оптическую плотность в каждой лунке при 405 нм, и полученное измерение использовали в качестве показателя активности TRAP. Результаты показаны на фиг. 5 и 6. Значительного ингибирования активности TRAP не наблюдали в случае коммерчески доступного контрольного IgG крысы. С другой стороны, значительное ингибирование активности TRAP наблюдали в случаях, когда антитело #32A1 добавляли в диапазоне от 32 нг/мл до 1000 нг/мл, и антитело #8A1 и аффинно очищенное поликлональное антитело кролика No. 3 добавляли в диапазоне от 63 нг/мл до 1000 нг/мл. Также в случаях добавления антитела #3A1, антитела #34A1 и антитела #39A1, зависимое от дозы ингибирование активности TRAP наблюдали при относительно высокой концентрации 500 нг/мл или выше. Ингибирования образования остеокластов мыши другими антителами не наблюдали. По приведенным выше результатам антитела, которые сильно ингибировали образование остеокластов мыши (дифференцировку и созревание остеокластов), обнаружены среди полученных моноклональных антител крысы против Siglec-15 мыши. Кроме того, в качестве примера свойства, общего для антитела #3A1, антитела #8A1, антитела #32A1, антитела #34A1 и антитела #39A1, можно привести активность ингибирования образования остеокластов в концентрации 1000 нг/мл, т.е., 1 мкг/мл или меньше.
Пример 10
Получение конструкции, экспрессирующей растворимый белок Siglec-15 человека
Частичная нуклеотидная последовательность, кодирующая внеклеточный домен белка Siglec-15 человека, представлена как SEQ ID NO:15 в разделе “Список последовательностей”, и его аминокислотная последовательность представлена как SEQ ID NO:16 в разделе “Список последовательностей”. С применением такой частичной последовательности растворимый белок Siglec-15 человека можно получать в культуральном супернатанте клеток животных или тому подобное.
a) Амплификация гена растворимого Siglec-15 человека посредством ПЦР
В качестве праймеров для амплификации кДНК внеклеточного домена Siglec-15 человека посредством ПЦР, олигонуклеотид, обладающий последовательностью 5'-ggggacaagt ttgtacaaaa aagcaggctt caccATGGAA AAGTCCATCT GGCTGC-3' (hSiglec-15-ECD-F: SEQ ID NO:17 в разделе “Список последовательностей”); олигонуклеотид, обладающий последовательностью 5'-ggggaccact ttgtacaaga aagctgggtc CCCGCTGGCG CCATGGAAGC GG-3' (hSiglec-15-ECD-R: SEQ ID NO:18 в разделе “Список последовательностей”), синтезировали общепринятым способом. К тому же, эти праймеры сконструированы как праймеры для амплификации для получения исходного клона gateway, такого, что последовательность attB1 добавлена к hSiglec-15-ECD-F, и последовательность attB2 добавлена к hSiglec-15-ECD-R. ПЦР проводили с использованием этого сочетания праймеров и полинуклеотида, содержащего последовательность открытой рамки считывания Siglec-15 человека в качестве матрицы, общепринятым способом. Полученную реакционную смесь после ПЦР очищали с использованием набора для очистки продуктов ПЦР PureLink (изготовленного в Invitrogen, Inc.).
b) Получение исходного клона реакцией Gateway BP
Исходный клон, в который кДНК внеклеточного домена Siglec-15 человека встраивали посредством способа Gateway (Invitrogen, Inc.) с использованием системы сайт-специфической рекомбинации фага лямбда, получали следующим способом. Сначала проводили реакцию BP с использованием BP Clonase проводили между продуктом ПЦР, обладающим последовательностью attB на обоих концах, полученным в a), и pDNOR221 (изготовленной в Invitrogen, Inc.), которая представляет собой донорный вектор, обладающий последовательностью attP. С использованием этой реакционной смеси трансформировали Escherichia coli TOP10, проводили ПЦР колоний для поиска устойчивых к лекарственному средству клонов, и подтверждали размер вставок. Затем для клона, в котором подтверждено наличие вставки правильного размера, проводили анализ секвенированием всей интересующей последовательности ДНК. В результате, получен исходный клон, полностью идентичный намеченной нуклеотидной последовательности (SEQ ID NO:15 в разделе “Список последовательностей”), кодирующей внеклеточный домен белка Siglec-15 человека.
c) Получение экспрессирующего клона реакцией Gateway LR
Экспрессирующий клон, в который кДНК внеклеточного домена Siglec-15 человека встраивали посредством способа Gateway (Invitrogen, Inc.) с использованием системы сайт-специфической рекомбинации фага лямбда, получали следующим способом. Исходный клон, полученный в b), содержит вставку, обладающую последовательностью attL на обоих концах. Проводили реакцию LR с использованием LR Clonase между этим исходным клоном и двумя типами векторов назначения, обладающих последовательностью attR. К тому же, в качестве векторов назначения, использовали два типа векторов назначения: pDONM сконструированный так, что метка эпитопа V5 и метка 6х His добавлены к С-концу вставки; и phIgFc, сконструированный так, что метка Fc человека добавлена к С-концу вставки. С использованием реакционной смеси, полученной посредством реакции LR, трансформировали Escherichia coli TOP10, и проводили анализ секвенированием для полученных устойчивых к лекарственному средству клонов для подтверждения того, что произошла правильная рекомбинация.
В результате анализа секвенированием, получены экспрессирующие клоны (растворимый Siglec-15 человека/pDONM и растворимый Siglec-15/phIgFc), в которых произошла правильная рекомбинация как для pDONM, так и для phIgFc, соответственно. Посредством трансфекции клеток животных или тому подобное растворимым Siglec-15 человека/pDONM, мРНК, обладающую последовательностью оснований SEQ ID NO:19 в разделе “Список последовательностей”, транскрибируют и транслируют в белок (Siglec-15 человека-His), обладающий аминокислотной последовательностью SEQ ID NO:20 в разделе “Список последовательностей”. Кроме того, посредством трансфекции клеток животных или тому подобное растворимым Siglec-15 человека/phIgFc, мРНК, обладающую последовательностью оснований SEQ ID NO:21 в разделе “Список последовательностей”, транскрибируют и транслируют в белок (Siglec-15-Fc человека), обладающий аминокислотной последовательностью SEQ ID NO:22 в разделе “Список последовательностей”.
Пример 11
Крупномасштабное получение культурального раствора, содержащего растворимый белок Siglec-15 человека, с использованием клеток 293-F
a) Получение культурального раствора, содержащего Siglec-15-His человека
Растворимый Siglec-15 человека/pDONM, полученный в примере 10, получали в количестве приблизительно 25 мг. К тому же, при очистке плазмид из Escherichia coli после крупномасштабного культивирования использовали набор для выделения плазмид в гига-количествах Invitrogen PureLink HiPure (изготовленный в Invitrogen, Inc.). Полученную таким образом плазмиду смешивали с Opti-MEM (изготовленной в Invitrogen, Inc.), в нее добавляли 50 мл реагента для трансфекции 293 фектина (изготовленного в Invitrogen, Inc.), и полученную смесь инкубировали при комнатной температуре в течение 20 минут. Эту смесь добавляли к клеткам 293-F (изготовленным в Invitrogen, Inc.), культивированным в среде для экспрессии Freestyle 293 (изготовленной в Invitrogen, Inc.), содержащей 1% пенициллин-стрептомицин, так чтобы плотность клеток достигла 1,0-3,4×106 клеток/мл, с использованием 25 л аппарата-биопроцессора для культивирования (WAVE Bioreactor). Затем клетки подвергали культивированию с встряхиванием (30 оборотов/мин) при концентрации CO2 от 6 до 12% в течение 96 часов (4 суток) при 37°C, культуральный раствор собирали и центрифугировали для получения культурального супернатанта. Наблюдали, что в полученном таким образом культуральном супернатанте экспрессируются белок, в котором метка эпитоп V5 и метка 6х His добавлены к C-концу внеклеточного домена Siglec-15 человека (Siglec-15-His человека).
b) Получение культурального раствора, содержащего Siglec-15-Fc человека
Растворимый Siglec-15 человека/phIgFc, полученный в примере 10, получали в количестве приблизительно 5 мг. К тому же, при очистке плазмид из Escherichia coli после крупномасштабного культивирования использовали набор для выделения плазмид в гига-количествах Invitrogen PureLink HiPure (изготовленный в Invitrogen, Inc.). Полученную таким образом плазмиду смешивали с Opti-MEM (изготовленной в Invitrogen, Inc.) с последующей стерилизацией фильтрацией. Затем в нее добавляли 10 мл реагента для трансфекции 293 фектина (изготовленного в Invitrogen, Inc.), и полученную смесь инкубировали при комнатной температуре в течение 20 минут. Эту смесь добавляли к клеткам Freestyle 293-F (изготовленным в Invitrogen, Inc.), культивировали в колбах Эрленмейера, так чтобы плотность клеток достигла 1,0-3,0×106 клеток/мл × 5 л (1 л/флакон × 5 флаконов) в среде для экспрессии Freestyle 293 (изготовленной в Invitrogen, Inc.). Затем клетки подвергали культивированию с встряхиванием (125 оборотов/мин) при концентрации CO2 8,0% в течение 96 часов (4 суток) при 37°C, культуральный раствор собирали и центрифугировали для получения культурального супернатанта. Наблюдали, что в полученном таким образом культуральном супернатанте экспрессируется белок, в котором метка Fc человека добавлены к C-концу внеклеточного домена Siglec-15 человека (Siglec-15-Fc человека).
Пример 12
Очистка растворимого белка Siglec-15 человека
a) Очистка растворимого Siglec-15-His человека
a-i) хроматография на колонке HisTrap HP
К 12 л культурального раствора экспрессирующих Siglec-15-His человека клеток 293F, полученных в примере 11, добавляли 1350 мл 10×буфера (500 мМ Tris, 1,5 M NaCl, 200 мМ имидазол, pH 8,0) и полученную смесь хорошо перемешивали и фильтровали через фильтр MilliPak-60 (изготовленный в Millipore Co., Ltd.). Этот культуральный раствор наносили при скорости потока 10 мл/мин на колонку с Ni-Сефарозой HP (изготовленной в Amersham Biosciences, Inc.) 100 мл, предварительно промытую чистой водой (водой Milli-Q). Затем колонку промывали 400 мл буфера 50 мМ Tris-HCl (pH 8,0), содержащего 300 мМ NaCl, при скорости потока 8 мл/мин, белок, адсорбированный на колонке, элюировали с помощью 200 мл буфера 50 мМ Tris-HCl (pH 8,0), содержащего 300 мМ NaCl и 500 мМ имидазол, при скорости потока 2,5 мл/мин, и элюат фракционировали в пробирки mini-sorp (изготовленные в Nunc, Inc.). Чтобы предотвратить преципитацию белка, 8 мл раствора 5 M NaCl добавляли к приблизительно 40 мл фракции, содержащей элюированный белок, с последующим перемешиванием, и затем полученную смесь концентрировали до приблизительно 20 мл с помощью центрифугируемого мембранного концентратора Amicon Ultra-15 (изготовленного в Millipore Co., Ltd.). Нерастворимое вещество, полученное во время концентрирования, удаляли центрифугированием при 3000 об./мин в течение 30 минут при 4°C, и 2,5 мл полученного супернатанта наносили на обессоливающую колонку PD-10 (изготовленную в Amersham Biosciences, Inc.) предварительно уравновешенную фосфатно-солевым буфером, содержащим 1 M NaCl (N-PBS), с последующей элюцией N-PBS, в результате чего получали 3,5 мл образца, растворитель в котором заменен на N-PBS. Эту процедуру проводили посредством повторения ее еще 7 раз, и получали приблизительно 28 мл раствора частично очищенного Siglec-15-His человека.
a-ii) хроматография на колонке Resource Q
12 мл образца, который очищали хроматографией на колонке с Ni-Сефарозой HP, и растворитель в котором заменяли на N-PBS, подвергали диализу в течение ночи при 4°C против буфера 50 мМ Tris-HCl (pH 7,5), содержащего 0,1% CHAPS (1 л, 3 раза), и полученный диализат центрифугировали при 3000 об./мин в течение 30 минут при 4°C, и преципитат удаляли. Затем полученный супернатант фильтровали через фильтр Millex-GV (изготовленный в Millipore Co., Ltd.), фильтрат наносили при скорости потока 1 мл/мин на колонку Resource Q 6 мл (изготовленную в Amersham Biosciences, Inc.), предварительно уравновешенную буфером 50 мМ Tris-HCl (pH 7,5), содержащим 0,1% CHAPS. Затем колонку промывали этим буфером при скорости потока 1 мл/мин и собирали фракцию белка, не адсорбированного на колонке. Белок, адсорбированный на колонке, элюировали буфером 50 мМ Tris-HCl (pH 7,5), содержащим 0,1% CHAPS и 1 M NaCl, при скорости потока 1 мл/мин. Затем 26,5 мл из фракции, не адсорбированной на колонке, концентрировали до 3,0 мл с помощью центрифугируемого мембранного концентратора Amicon Ultra-15 (изготовленного в Millipore Co., Ltd.), концентрат центрифугировали при 3000 об./мин в течение 10 минут при 4°C и преципитат удаляли. 2,5 мл полученного супернатанта наносили на обессоливающую колонку PD-10 (изготовленную в Amersham Biosciences, Inc.), предварительно уравновешенную фосфатно-солевым буфером, содержащим 50 мМ гидрохлорид аргинина (pH 7,0, A-PBS), с последующей элюцией A-PBS, в результате чего получали 3,5 мл образца, растворитель в котором заменен на A-PBS. Гидрохлорид аргинина в растворитель полученного образца добавляли, чтобы предотвратить преципитацию растворимого Siglec-15-His человека. Супернатант после центрифугирования подвергали криоконсервации при -80°C до использования. Вышеупомянутую процедуру очистки (хроматографию на колонке Resource Q) проводили, повторяя ее дважды.
a-iii) Детекция и анализ чистоты очищенного Siglec-15-His человека
С использованием образца, полученного посредством вышеупомянутой процедуры очистки (хроматографии на колонке с Ni-Сефарозой HP и хроматографии на колонке Resource Q), проводили SDS-полиакриламидный электрофорез в восстанавливающих условиях и окрашивание серебром. То есть, к 5 мкл из каждого образца, очищенного на соответствующих стадиях очистки, добавляли равное количество раствора для обработки с SDS, и полученную смесь термически обрабатывали при 95°C в течение 10 минут. По 0,3 мкл из каждого из термически обработанных образцов использовали для SDS-полиакриламидного электрофореза. Процедуру электрофореза проводили таким же способом, как в примере 3, за исключением того, что в качестве маркеров молекулярной массы использовали маркеры молекулярной массы Rainbow (изготовленные в Amersham Biosciences, Inc.). После завершения электрофореза проводили окрашивание серебром с использованием набора PhastGel Silver (изготовленного в Amersham Biosciences, Inc.) и PhastSystem. Результаты показаны на фиг. 7. Показано, что белок, обладающий молекулярной массой приблизительно 35 кДа (Siglec-15-His человека), являлся эффективно очищенным и концентрированным во фракции белка, не адсорбированной на колонке Resource Q.
a-iv) Измерение концентрации белка очищенного Siglec-15-His человека
Для очищенного Siglec-15-His человека (фракции белка, не адсорбированной на колонке Resource Q), концентрацию белка измеряли с помощью набора для анализа белка DC (изготовленного в Bio-Rad Laboratories, Inc.) с использованием бычьего сывороточного альбумина в качестве стандартного образца. Посредством проведения процедуры очистки дважды всего получили 1,66 мг очищенного Siglec-15-His человека.
b) Очистка растворимого Siglec-15-Fc человека
b-i) хроматография на колонке HiTrap с белком A
1,5 л культурального раствора экспрессирующих Siglec-15-Fc человека клеток 293F, полученных в b) из примера 11, фильтровали через фильтр Sterivex-GV (изготовленный в Millipore Co., Ltd.), и затем фильтрат наносили при скорости потока 5 мл/мин на 5 мл колонку HiTrap с белком A (изготовленную в Amersham Biosciences, Inc.), предварительно уравновешенную PBS Дульбекко (D-PBS, изготовленным в Invitrogen, Inc.). Затем колонку промывали 70 мл D-PBS при скорости потока 5 мл/мин, белок, адсорбированный на колонке, элюировали с помощью 24 мл буфера 0,1 M цитрата натрия (pH 3,0) при скорости потока 1,2 мл/мин. Элюат фракционировали по 1,2 мл в пробирки mini-sorp (изготовленные в Nunc, Inc.), и немедленно после этого, туда добавляли 0,31 мл 1 M Tris для нейтрализации элюата. Аликвоту 2,5 мл из раствора (приблизительно 7,5 мл), полученного объединением фракций элюированного белка (фракции 5-9), наносили на обессоливающую колонку PD-10 (изготовленную в Amersham Biosciences, Inc.), предварительно уравновешенную фосфатно-солевым буфером, содержащим 50 мМ гидрохлорид аргинина (pH 7,0, A-PBS), с последующей элюцией A-PBS, в результате чего получили 3,5 мл образца, растворитель в котором заменен на A-PBS. Эту процедуру проводили посредством повторения ее дважды. Гидрохлорид аргинина в растворитель добавляли, чтобы предотвратить преципитацию растворимого Siglec-15-Fc человека. 2,5 мл оставшегося раствора фракций элюированного белка (фракции 5-9) наносили на обессоливающую колонку PD-10 (изготовленную в Amersham Biosciences, Inc.), предварительно уравновешенную фосфатно-солевым буфером, содержащим 1 M NaCl (pH 6,7, N-PBS), с последующей элюцией N-PBS, в результате чего получали 3,5 мл образца, растворитель в котором заменен на N-PBS. NaCl в растворитель в полученном образце добавляли, чтобы предотвратить преципитацию растворимого Siglec-15-Fc человека без добавления содержащего аминогруппу соединения, такого как аргинин. Образцы, полученные вышеупомянутым способом, подвергали криоконсервации при -80°C до использования.
b-ii) Детекция и анализ чистоты очищенного Siglec-15-Fc человека
С использованием образцов, полученных посредством вышеупомянутой процедуры очистки, проводили SDS-полиакриламидный электрофорез в восстанавливающих условиях и окрашивание серебром. То есть, к 5 мкл из каждого образца, очищенного на соответствующих стадиях очистки, добавляли равное количество раствора для обработки с SDS, и полученную смесь термически обрабатывали при 95°C в течение 10 минут. По 0,3 мкл образцов, полученных разведением каждого из термически обработанных образцов 1/100 или 1/300 с помощью половины концентрации раствора для обработки с SDS, использовали для SDS-полиакриламидного электрофореза. Электрофорез и окрашивание серебром проводили таким же способом, как в анализе чистоты Siglec-15-His, описанном в a-iii). Результаты показаны на фиг. 8. Показано, что белок, обладающий молекулярной массой приблизительно 55 кДа (Siglec-15-Fc человека), являлся эффективно очищенным и концентрированным во фракции белка, элюированного с колонки HiTrap с белком A.
b-iii) Измерение концентрации белка очищенного Siglec-15-Fc человека
Для очищенного Siglec-15-Fc человека (фракции белка, элюированной с обессоливающей колонки PD-10) концентрацию белка измеряли с помощью набора для анализа белка DC (изготовленного в Bio-Rad Laboratories, Inc.) с использованием бычьего сывороточного альбумина в качестве стандартного образца. Как показано в таблице 4, всего 25,2 мг очищенного Siglec-15-Fc человека получили посредством проведения процедуры очистки дважды.
Пример 13
Оценка свойства связывания моноклонального антитела крысы против Siglec-15 мыши с белком Siglec-15 человека
Свойство связывания моноклонального антитела крысы против Siglec-15 мыши с белком Siglec-15 человека оценивали способом ELISA. Белок Siglec-15-Fc человека (подвергнутый замене буфера на A-PBS), полученный в b) из примера 12, разводили до 5 мкг/мл буфером 0,1 M карбонатом натрия (pH 9,5), и полученный раствор добавляли в 96-луночный планшет (изготовленный в Nalge Nunc International, Inc., Кат. No. 430341) по 100 мкл/лунку. Затем планшет оставляли при комнатной температуре на 1 час, раствор удаляли, и буфер для промывки (фосфатно-солевой буфер, содержащий 0,05% Tween 20) добавляли по 300 мкл/лунку и затем удаляли. Затем эту процедуру промывки проводили еще раз, фосфатно-солевой буфер, содержащий 25% BlockAce (изготовленный в Dainippon Sumitomo Pharma Co., Ltd.) добавляли по 200 мкл/лунку, и планшет оставляли при комнатной температуре на 1 час, таким образом проводили блокирование. Жидкость удаляли, и планшет промывали дважды по 300 мкл/лунку буфера для промывки. Затем каждое из моноклональных антител крысы против Siglec-15 мыши, полученных в примере 6, или контрольное IgG крысы (изготовленное в R&D Systems, Inc.) разводили до конечной концентрации от 1,28 до 20000 нг/мл (серии 5-кратного разведения) буфером для ELISA (фосфатно-солевой буфер, содержащий 12,5% BlockAce и 0,05% Tween 20), и полученный раствор разведенного антитела добавляли в планшет по 100 мкл/лунку. Затем планшет оставляли при комнатной температуре на 1 час, жидкость удаляли, и планшет промывали 3 раза по 300 мкл/лунку буфера для промывки. Затем меченное HRP (пероксидазой хрена) антитело козы против IgG крысы (изготовленное в Beckman Coulter, Inc.), разведенное в 1000 раз буфером для ELISA, добавляли по 100 мкл/лунку, и планшет оставляли при комнатной температуре на 1 час. Жидкость удаляли и планшет промывали 3 раза по 300 мкл/лунку буфера для промывки, и затем, с использованием набора для проявления окрашивания для пероксидазы (изготовленного в Sumitomo Bakelite Co., Ltd.), окрашивание проявляли согласно протоколу, сопровождающему набор. После проявления окрашивания оптическую плотность при 492 нм измеряли с использованием считывателя для микропланшетов (изготовленного в Nihon Molecular Devices Corporation). В результате подтвердили, что все из оцениваемых 10 тестируемых образцов моноклональных антител крысы против Siglec-15 мыши связывают белок Siglec-15 человека зависимым от концентрации антител образом (Фиг. 9). В частности, активность связывания 5 тестируемых образцов, а именно, #1A1, #3A1, #24A1, #32A1 и #61A1, являлась высокой, и активность связывания 3 тестируемых образцов, а именно, #8A1, #34A1, и #39A1, являлась относительно низкой. С другой стороны, в случае контрольного IgG крысы, не наблюдали связывания с белком Siglec-15. По вышеуказанным результатам, показано, что моноклональные антитела крысы против Siglec-15 мыши, полученные в примере 6, связывают не только Siglec-15 мыши, но также Siglec-15 человека, и более того, обнаружено, что некоторые антитела сильно связывают Siglec-15 человека.
Пример 14
Эффект добавления моноклонального антитела крысы против Siglec-15 мыши на слияние клеток и активность резорбции кости нормальных клеток-предшественников остеокластов человека (оценка биологической активности in vitro)
Поскольку подтвердили, что моноклональные антитела крысы против Siglec-15 мыши связывают также Siglec-15 человека в примере 13, исследовали эффекты этих антител на образование остеокластов человека и активность резорбции кости.
a) Эффект добавления моноклонального антитела крысы против Siglec-15 мыши на слияние клеток остеокластов из нормальных клеток-предшественников остеокластов человека (Окрашивание TRAP)
Нормальные клетки-предшественники остеокластов человека (Normal Human Natural Osteoclast Precursor Cells, закупленные из Sanko Junyaku Co., Ltd., Кат. No. 2T-110), рассевали в 96-луночный планшет по 1×104 клеток/лунку согласно протоколу, сопровождающему клетки. В качестве среды использовали минимальную поддерживающую среду для клеток-предшественников остеокластов (OPBM, закупленную из Sanko Junyaku Co., Ltd., Кат. No. PT-8201), дополненную набором добавок OPGM (закупленным из Sanko Junyaku Co., Ltd., Кат. No. PT-9501), содержащую эмбриональную бычью сыворотку (конечная концентрация: 10%), RANKL человека (конечная концентрация: 66 нг/мл), M-CSF человека (конечная концентрация: 33 нг/мл) и тому подобное. К полученному культуральному супернатанту добавляли каждое из моноклональных антител крысы против Siglec-15 мыши, полученных в примере 6, или контрольное IgG крысы (изготовленное в R&D Systems, Inc.) для получения конечной концентрации 30 мкг/мл, и клетки культивировали в течение 4 суток в CO2 инкубаторе. После культивирования супернатант удаляли, и 10% нейтральный формалин добавляли для фиксации клеток. После фиксации клеток клетки дважды промывали дистиллированной водой, окрашивающий раствор TRAP (0,27 мМ нафтол AS-MX фосфат (изготовленный в Sigma Co., Ltd.), 1,6 мМ LB соль быстрого красно-фиолетового (изготовленную в Sigma Co., Ltd.), 1% диметилформамид, 50 мМ тартрат натрия, 0,1 M буфер ацетат натрия (pH 5,0)) добавляли по 100 мкл/лунку, и реакции позволяли проходить при комнатной температуре в течение 5 минут. После реакции клетки дважды промывали дистиллированной водой, и затем наблюдали под микроскопом (Фиг. 10). В результате образование гигантских остеокластов, происходящее из-за высокой степени слияния клеток, являлось почти полностью ингибированным добавлением антитела #32A1. Кроме того, в случае антитела #41B1, образование гигантских остеокластов, происходящее из-за высокой степени слияния клеток, также являлось значительно ингибированным. С другой стороны, в случае других моноклональных антител крысы против Siglec-15 мыши (антитела #1A1 и других) и контрольного IgG крысы, такого значительно ингибирования слияния клеток остеокластов не наблюдали. Этим способом выявили, что многоядерность и слияние клеток TRAP-положительных остеокластов из нормальных клеток-предшественников остеокластов человека являлись ингибированными посредством моноклональных антител, специфически связывающих белок Siglec-15.
b) Эффект добавления моноклонального антитела крысы против Siglec-15 мыши (#32A1) на слияние клеток остеокластов из нормальных клеток-предшественников остеокластов человека (Окрашивание TRAP)
Нормальные клетки-предшественники остеокластов человека (Normal Human Natural Osteoclast Precursor Cells, закупленные из Sanko Junyaku Co., Ltd., Кат. No. 2T-110) рассевали в 96-луночный планшет по 1×104 клеток/лунку согласно протоколу, сопровождающему клетки. В качестве среды использовали минимальную поддерживающую среду для клеток-предшественников остеокластов (OPBM, закупленную из Sanko Junyaku Co., Ltd., Кат. No. PT-8201), дополненную набором добавок OPGM (закупленным из Sanko Junyaku Co., Ltd., Кат. No. PT-9501), содержащую эмбриональную бычью сыворотку (конечная концентрация: 10%), RANKL человека (конечная концентрация: 68,4 нг/мл), M-CSF человека (конечная концентрация: 33 нг/мл) и тому подобное. К полученному культуральному супернатанту добавляли моноклональное антитело крысы против Siglec-15 мыши (антитело #32A1), полученное в примере 6, для получения конечной концентрации 0,1, 0,3, 1 или 3 мкг/мл, и клетки культивировали в течение 3 суток в CO2 инкубаторе. После культивирования супернатант удаляли, и 10% нейтральный формалин добавляли для фиксации клеток. После фиксации клеток клетки дважды промывали дистиллированной водой, и окрашивающий раствор TRAP (0,27 мМ нафтол AS-MX фосфат (изготовленный в Sigma Co., Ltd.), 1,6 мМ LB соль быстрого красно-фиолетового (изготовленная в Sigma Co., Ltd.), 1% диметилформамид, 50 мМ тартрат натрия, 0,1 M буфер ацетат натрия (pH 5,0)) добавляли по 100 мкл/лунку, и реакции позволяли проходить при комнатной температуре в течение 5 минут. После реакции клетки дважды промывали дистиллированной водой, и затем наблюдали под микроскопом (Фиг. 11). В результате образование TRAP-положительных многоядерных остеокластов являлось ингибированным зависимым от концентрации антитела #32A1 образом в диапазоне от 0,3 мкг/мл до 3 мкг/мл.
c) Эффект добавления моноклонального антитела крысы против Siglec-15 мыши (#32A1) на активность резорбции кости нормальных клеток-предшественников остеокластов человека (Оценка с использованием покрытого коллагеном планшета)
Известно, что остеокласты высвобождают протеазу, такую как катепсин К, и разрушают коллаген I типа, являющийся структурным компонентом костной ткани. В наборе для анализа OsteoLyse (изготовленном в Lonza, Inc., Кат. No. PA-1500), предоставлен 96-луночный планшет, покрытый коллагеном человека, конъюгированным с европием (96-луночный планшет для культивирования клеток OsteoLyse), и можно оценивать активность резорбции кости остеокластов in vitro посредством измерения количества флуоресцентных фрагментов коллагена, высвобождаемых в супернатант, когда остеокласты культивируют в планшете. Нормальные клетки-предшественники остеокластов человека (Normal Human Natural Osteoclast Precursor Cells остеокластов, закупленные из Sanko Junyaku Co., Ltd., Кат. No. 2T-110), рассевали в 96-луночный планшет для культивирования клеток OsteoLyse по 1×104 клеток/лунку согласно протоколу, сопровождающему клетки. В качестве среды использовали минимальную поддерживающую среду для клеток-предшественников остеокластов (OPBM, закупленную из Sanko Junyaku Co., Ltd., Кат. No. PT-8201), дополненную набором добавок OPGM (закупленным из Sanko Junyaku Co., Ltd., Кат. No. PT-9501), содержащую эмбриональную бычью сыворотку (конечная концентрация: 10%), RANKL человека (конечная концентрация: 68,4 нг/мл), M-CSF человека (конечная концентрация: 33 нг/мл) и тому подобное. К полученному культуральному супернатанту добавляли моноклональное антитело крысы против Siglec-15 мыши (антитело #32A1), полученное в примере 6, для получения конечной концентрации 0,1, 0,3, 1 или 3 мкг/мл, и клетки культивировали в течение 3 суток в CO2 инкубаторе. Собирали аликвоту 10 мкл культурального супернатанта и к ней добавляли 200 мкл высвобождающего флуорофор реагента, включенного в набор для анализа OsteoLyse, и измеряли интенсивность флуоресценции (Возбуждение: 340 нм, Излучение: 615 нм) с использованием флуоресцентного считывателя для планшетов (ARVO MX, изготовленного в Perkin Elmer Inc.), таким образом определяли количество свободных флуоресцентных фрагментов коллагена, высвобожденных в культуральный супернатант (Фиг. 12). В результате, количество флуоресцентных фрагментов коллагена, увеличенное при добавлении RANKL, являлось уменьшенным посредством антитела #32A1 зависимым от концентрации образом в диапазоне от 0,3 мкг/мл до 3 мкг/мл. По этим результатам выявили, что активность резорбции кости остеокластов человека являлась ингибированной посредством исследуемого моноклонального антитела, специфически связывающего белок Siglec-15.
Пример 15
Оценка биологической активности моноклонального антитела крысы против Siglec-15 мыши с использованием крысы после овариэктомии
a) Способ эксперимента с животными
Яичники с обеих сторон удаляли у самок крыс F344 (полученных из Charles River Laboratories Japan, Inc.) в возрасте 12 недель, и крыс разделяли на три группы: группу с введением носителя; группу с введением моноклонального антитела крысы против Siglec-15 мыши #8A1; и группу с введением моноклонального антитела крысы против Siglec-15 мыши #32A1. Кроме того, одну группу подготавливали в качестве группы с ложной операцией. В группах с введением антитела моноклональное антитело крысы против Siglec-15 мыши #8A1 или моноклональное антитело крысы против Siglec-15 мыши #32A1, полученные в примере 6, внутрибрюшинно вводили в дозе 1 мг/кг три раза в неделю с повторением в течение 4 недель начиная со следующих суток после операции. В группе с введением носителя и в группе с ложной операцией, PBS, содержащий 0,01% Tween 20, внутрибрюшинно вводили в качестве носителя. Через 4 недели после начала введения, мочу собирали в течение 24 часов в условиях голодания, и образцы мочи сохраняли при -80°C до измерения. После завершения сбора мочи, крыс подвергали эвтаназии, и поясничный отдел позвоночника вырезали у каждой крысы.
b) Измерение минеральной плотности костной ткани поясничного отдела позвоночника
Мягкие ткани, присоединенные к вырезанному поясничному отделу позвоночника, удаляли, и выделяли поясничные позвонки от 4-го до 6-го. Вырезанные поясничные позвонки обезжиривали и дегидратировали встряхиванием в этаноле и затем высушивали на воздухе, и минеральную плотность костной ткани измеряли с использованием костного денситометра (DCS-600EX, изготовленный в Aloka Co., Ltd.). Результаты показаны на фиг. 13(A). Значительное уменьшение минеральной плотности костной ткани в поясничный отдел позвоночника наблюдали в группе после овариэктомии по сравнению с группой после ложной операции, однако, в группах с введением антител #8A1 и #32A1 уменьшение минеральной плотности костной ткани из-за овариэктомии являлось значительно ингибированным.
c) Измерение выделения дезоксипиридинолина с мочой
Множество сшитых метаболитов коллагена I типа явно отражают метаболическое ремоделирование кости, в частности, резорбцию кости. Прежде всего, дезоксипиридинолин локализован главным образом в коллагене костной ткани, и таким образом, его считают высоко надежным в качестве показателя резорбции кости.
Криоконсервированные образцы мочи оттаивали, и нерастворимое вещество осаждали способом центрифугирования, в результате чего получали супернатант. Количество дезоксипиридинолина, содержащегося в этом супернатанте, измеряли с использованием Osteolinks «DPD» (изготовленного в DS Pharma Biomedical Co., Ltd.). Кроме того, с использованием теста на креатинин Wako (изготовленного в Wako Pure Chemical Industries, Ltd.), содержание креатинина в супернатанте также измеряли, и рассчитывали количество дезоксипиридинолина, скорректированное по креатинину. Результаты показаны на фиг. 13(B). Выделение дезоксипиридинолина с мочой являлось значительно увеличенным в группе после овариэктомии по сравнению с группой после ложной операции, и таким образом, это указывает на то, что у крыс после овариэктомии резорбция кости остеокластами является увеличенной. С другой стороны, в группах с введением антител #8A1 и #32A1, увеличение выделения дезоксипиридинолина из-за овариэктомии являлось ингибированным, так что уровень выделения дезоксипиридинолина являлся сравнимым с уровнем в группе после ложной операции. По этому результату также подтвердили в моделях на животных, что исследуемые моноклональные антитела, специфически связывающие Siglec-15, ингибируют резорбцию кости остеокластами, и это явно показывает, что из-за ингибирующего эффекта на резорбцию кости, уменьшение минеральной плотности костной ткани в поясничном отделе позвоночника у крыс после овариэктомии являлось ингибированным.
Пример 16
Определение участка связывания (эпитопа) моноклонального антитела крысы против Siglec-15 мыши
a) Экспрессия и очистка V-set домена Siglec-15 человека
ДНК, кодирующую белок, в котором метка His и последовательность узнавания тромбина присоединены к N-концу V-set домена Siglec-15 человека (полипептид, содержащий остатки аминокислот 39-165 аминокислотной последовательности в Базе данных белков NCBI с инвентарным номером NP_998767 или аминокислотная последовательность, представленная на SEQ ID NO:2 в разделе “Список последовательностей”), встраивали в вектор pDEST14 (Invitrogen, Inc., Кат. No. 11801-016). С использованием этой плазмиды трансформировали Escherichia coli Rosetta-gamiB (DE3) (Novagen, Inc., Кат. No. 71136-4) и культивировали в среде ТВ (Invitrogen, Inc., Кат. No. 22711-022). После культивирования бактериальные клетки гомогенизировали ультразвуком, полученный гомогенат центрифугировали, и супернатант очищали на колонке HisTrap HP (GE Healthcare, Co., Ltd., Кат. No. 17-5247-01). Затем метку His отщепляли тромбином, и затем V-set домен Siglec-15 человека очищали с использованием колонки Mono S5/50 GL (GE Healthcare, Co., Ltd., Кат. No. 17-5168-01) и колонки Superdex 75 10/300 (GE Healthcare, Co., Ltd., Кат. No. 17-5174-01), пока не получали отдельную полосу с молекулярной массой 14 кДа при электрофорезе.
b) Очистка растворимого Siglec-15-Fc человека
Растворимый Siglec-15-Fc человека очищали способом, описанным в примере 12.
c) Конкурентный ELISA V-set домена человека и моноклонального антитела крысы против Siglec-15 мыши #32A1
В 96-луночный планшет maxi-sorp (изготовленный в Nunc, Inc., номер модели: 442404), 100 мкл антитела козы против Fc человека (Jackson ImmunoResearch, Inc., номер модели: 109-005-098) (1,25 мкг/мл) добавляли в каждую лунку и иммобилизовали в течение ночи при 4°C. Затем 96-луночный планшет maxi-sorp дважды промывали PBS, 100 мкл растворимого Siglec-15-Fc человека (1 мкг/мл) добавляли в каждую лунку и иммобилизовали при комнатной температуре в течение 1 часа. Затем 300 мкл раствора 5% обезжиренного молока/PBS добавляли в каждую лунку, и блокирование проводили при комнатной температуре в течение 3 часов. Одновременно, 2 мкг/мл моноклонального антитела крысы против Siglec-15 мыши #32A1 смешивали с равным количеством 0, 0,032, 0,16, 0,8, 4 или 20 мкг/мл V-set домена Siglec-15 человека, и реакции позволяли проходить при комнатной температуре в течение 1,5 часов. Затем 96-луночный планшет maxi-sorp дважды промывали PBS, в него добавляли 100 мкл смешанного раствора моноклонального антитела крысы против Siglec-15 мыши #32A1 и V-set домена Siglec-15 человека, и реакции позволяли проходить при комнатной температуре в течение 1 часа. Затем 96-луночный планшет maxi-sorp промывали 5 раз с помощью раствора 0,05% Tween 20 (Bio-Rad Laboratories, Inc., Кат. No. 170-6531)/PBS (далее в настоящем документе обозначенного как «раствор PBST»), добавляли 100 мкл HRP-меченного антитела козы против IgG крысы (Beckman Coulter, Inc., Кат. No. 732664, разведенного в 2000 раз), и реакции позволяли проходить при комнатной температуре в течение 1 часа. Затем планшет промывали 5 раз раствором PBST, туда добавляли 100 мкл жидкости для проявления окраски из набора субстрата ELISA POD ABTS (Nacalai Tesque Co., Ltd., Кат. No. 14351-80), и реакции позволяли проходить в течение 30 минут. Затем туда добавляли 100 мкл раствора для остановки реакции, и измеряли оптическую плотность при 405 нм. Результаты конкурентного ELISA показывают, что связывание моноклонального антитела крысы против Siglec-15 мыши #32A1 с иммобилизованным Siglec-15 человека является ингибированным зависимым от концентрации V-set домена человека образом. Соответственно, показано, что эпитоп для моноклонального антитела крысы против Siglec-15 мыши #32A1 представляет собой V-set домен Siglec-15 человека (домен, содержащий остатки аминокислот 39-165 аминокислотной последовательности в Базе данных белков NCBI с инвентарным номером NP_998767 или аминокислотная последовательность, представленная как SEQ ID NO:2 в разделе “Список последовательностей”) (Фиг. 14).
Пример 17
Амплификация кДНК, кодирующей вариабельную область моноклонального антитела крысы против Siglec-15 мыши #32A1, и ее анализ секвенированием
a) Получение мРНК
Гибридому #32A1 культивировали по a) из примера 6. Из 4×107 клеток гибридомы #32A1, приблизительно 65 мкг мРНК получали с использованием набора для выделения мРНК QuickPrep (GE Healthcare, Co., Ltd.).
b) Синтез кДНК (5'-RACE-Ready кДНК)
Синтез кДНК (5'-RACE-Ready кДНК) проводили с использованием 0,3 мкг мРНК, полученной в a) с помощью обратной транскриптазы PrimeScript (TaKaRa Bio, Inc.) и набора для амплификации кДНК SMART RACE (Clontech Co., Ltd.).
c) Амплификация кДНК из гена вариабельной области тяжелой цепи #32A1 посредством 5'-RACE ПЦР
В качестве праймеров для амплификации кДНК из гена вариабельной области тяжелой цепи #32A1 посредством ПЦР использовали UPM (Универсальную смесь праймеров A, приложенную к набору для амплификации кДНК SMART RACE) и олигонуклеотид, обладающий последовательностью 5'-GGCCGGGTGGGCTACGTTGCAGGTGACGGTCTG-3' (RG2AR2: SEQ ID NO:23 в разделе “Список последовательностей”). В качестве UPM использовали UPM, приложенную к набору для амплификации кДНК SMART RACE (Clontech Co., Ltd.), и RG2AR2 сконструировали из последовательности константной области тяжелой цепи крысы (IgG2a) в базе данных и синтезировали общепринятым способом.
кДНК из гена вариабельной области тяжелой цепи #32A1 амплифицировали посредством 5'-RACE ПЦР с использованием этого сочетания праймеров и кДНК, синтезированной в b) (5'-RACE-Ready кДНК) в качестве матрицы. ПЦР проводили с использованием набора для ПЦР Advantage 2 (Clontech Co., Ltd.), и условия для термоциклера устанавливали следующим образом: после нагревания при 94°C в течение 1 минуты, температурный цикл «94°C в течение 0,5 минуты и 72°C в течение 3 минут» повторяли 5 раз, затем температурный цикл «94°C в течение 0,5 минуты, 70°C в течение 0,5 минуты, и 72°C в течение 3 минут» повторяли 5 раз, и затем температурный цикл «94°C в течение 0,5 минуты, 68°C в течение 0,5 минуты и 72°C в течение 3 минут» повторяли 20 раз с последующей инкубацией при 4°C.
d) Амплификация кДНК из гена вариабельной области легкой цепи #32A1 посредством 5'-RACE ПЦР
В качестве праймеров для амплификации кДНК из гена вариабельной области легкой цепи #32A1 посредством ПЦР использовали UPM (Универсальную смесь праймеров A, приложенную к набору для амплификации кДНК SMART RACE) и олигонуклеотид, обладающий последовательностью 5'-CATGCTGTACGTGCTGTCTTTGCTGTCCTGATCAG-3' (RKR2: SEQ ID NO:24 в разделе “Список последовательностей”). В качестве UPM использовали UPM, приложенную к набору для амплификации кДНК SMART RACE (Clontech Co., Ltd.), и RKR2 сконструировали из последовательности константной области легкой цепи крысы (κ цепь) в базе данных и синтезировали общепринятым способом.
кДНК из гена вариабельной области легкой цепи #32A1 амплифицировали посредством 5'-RACE ПЦР с использованием этого сочетания праймеров и кДНК, синтезированной в b) (5'-RACE-Ready кДНК) в качестве матрицы. ПЦР проводили с использованием набора для ПЦР Advantage 2 (Clontech Co., Ltd.), и условия для термоциклера устанавливали следующим образом: после нагревания при 94°C в течение 1 минуты, температурный цикл «94°C в течение 0,5 минуты и 72°C в течение 3 минут» повторяли 5 раз, затем температурный цикл «94°C в течение 0,5 минуты, 70°C в течение 0,5 минуты и 72°C в течение 3 минут» повторяли 5 раз, и затем температурный цикл «94°C в течение 0,5 минуты, 68°C в течение 0,5 минуты и 72°C в течение 3 минут» повторяли 20 раз с последующей инкубацией при 4°C
e) Определение последовательностей оснований кДНК вариабельных областей тяжелой и легкой цепи
кДНК вариабельной области тяжелой цепи, амплифицированную в c), очищали с использованием набора для очистки продуктов ПЦР MinElute (QIAGEN Inc.), и затем проводили анализ секвенированием последовательности ДНК. В качестве праймера для секвенирования олигонуклеотид, обладающий последовательностью 5'-CTCCAGAGTTCCAGGTCACGGTGACTGGC-3' (RG2AR3: SEQ ID NO:25 в разделе “Список последовательностей”) сконструировали из последовательности константной области тяжелой цепи крысы (IgG2a) в базе данных и синтезировали общепринятым способом.
кДНК вариабельной области легкой цепи, амплифицированную в d) очищали с использованием набора для очистки продуктов ПЦР MinElute (QIAGEN Inc.), и затем проводили анализ секвенированием последовательности ДНК. В качестве праймера для секвенирования олигонуклеотид, обладающий последовательностью 5'-TCCAGTTGCTAACTGTTCCG-3' (sqRK: SEQ ID NO:26 в разделе “Список последовательностей”) сконструировали из последовательности константной области легкой цепи крысы (κ цепь) в базе данных и синтезировали общепринятым способом.
кДНК, содержащая вариабельную область тяжелой цепи, полученная анализом секвенированием, обладает последовательностью оснований SEQ ID NO:27 в разделе “Список последовательностей”, и кодирует аминокислотную последовательность SEQ ID NO:28 в разделе “Список последовательностей”. Аминокислотная последовательность, представленная аминокислотами под номерами 1-19 из SEQ ID NO:28, соответствует сигнальной последовательности для секреции, аминокислотная последовательность, представленная ее аминокислотами под номерами 20-140, соответствует вариабельной области тяжелой цепи, и аминокислотная последовательность, представленная ее аминокислотами под номерами 141-167, соответствует константной области тяжелой цепи (частичной). Вышеупомянутая вариабельная область тяжелой цепи содержит CDRH1 (DYFMN), содержащую аминокислотную последовательность SEQ ID NO:44, CDRH2 (QIRNKIYTYATFYAESLEG), содержащую аминокислотную последовательность SEQ ID NO:45, и CDRH3 (SLTGGDYFDY), содержащую аминокислотную последовательность SEQ ID NO:46. Кроме того, кДНК, содержащая вариабельную область легкой цепи, обладает последовательностью оснований SEQ ID NO:29 в разделе “Список последовательностей”, и кодирует аминокислотную последовательность SEQ ID NO:30 в разделе “Список последовательностей”. Аминокислотная последовательность, представленная аминокислотами под номерами 1-20 из SEQ ID NO:30, соответствует сигнальной последовательности для секреции, аминокислотная последовательность, представленная ее аминокислотами под номерами 21-132, соответствует вариабельной области легкой цепи, и аминокислотная последовательность, представленная ее аминокислотами под номерами 133-139, соответствует константной области легкой цепи (частичной). Вышеупомянутая вариабельная область легкой цепи содержит CDRL1 (RASQSVTISGYSFIH), содержащую аминокислотную последовательность SEQ ID NO:47, CDRL2 (RASNLAS), содержащую аминокислотную последовательность SEQ ID NO:48, и CDRL3 (QQSRKSPWT), содержащую аминокислотную последовательность SEQ ID NO:49.
Пример 18
Получение конструкции, экспрессирующей ген химерного антитела человека из моноклонального антитела крысы против Siglec-15 мыши #32A1
a) Получение универсальных векторов экспрессии pEF1/FCCU-1 и pEF6KCL
a)-i) Конструирование экспрессирующего легкую цепь человека вектора pEF6KCL
Проведением ПЦР с использованием плазмиды pEF6/V5-HisB (Invitrogen) в качестве матрицы, а также с использованием следующих праймеров, получали фрагмент ДНК от непосредственно 3' от BGHpA (2174) до SmaI (2958) (фрагмент ДНК, содержащий точку начала репликации f1, и промотор и точку начала репликации SV40, далее в настоящем документе обозначенный как «фрагмент A»).
5'-CCACGCGCCCTGTAGCGGCGCATTAAGC-3' (праймер EFF1: SEQ ID NO:31 в разделе “Список последовательностей”)
5'-AAACCCGGGAGCTTTTTGCAAAAGCCTAGG-3' (праймер EFsmaR: SEQ ID NO:32)
Полученный фрагмент A и фрагмент ДНК (SEQ ID NO:33, далее в настоящем документе обозначенный как «фрагмент B»), содержащий последовательность ДНК, кодирующую сигнальную последовательность для секреции κ цепи человека, константную область κ цепи человека и человеческий сигнал добавления поли-A, лигировали посредством ПЦР с перекрыванием фрагментов. Полученный таким образом фрагмент ДНК, в котором фрагмент A и фрагмент В являлись лигированными (далее в настоящем документе обозначенный как «фрагмент A+B»), расщепленный рестрикционными ферментами KpnI и SmaI, лигировали с плазмидой pEF6/V5-HisB (Invitrogen), расщепленной рестрикционными ферментами KpnI и SmaI, в результате чего сконструировали экспрессирующий легкую цепь человека вектор pEF6KCL, обладающий сигнальной последовательностью, участком клонирования, константной областью κ цепи человека и сигнальной последовательностью человека для добавления поли-A, на 3'-конце от промотора EF1.
a)-ii) Конструирование pEF1/KCL
Фрагмент ДНК, полученный расщеплением pEF6KCL, полученной вышеупомянутым способом, рестрикционными ферментами KpnI и SmaI, лигировали с pEF1/myc-HisB (Invitrogen, Inc.), расщепленной KpnI и SmaI, посредством чего сконструировали плазмиду pEF1/KCL.
a)-iii) Конструирование экспрессирующего тяжелую цепь человека вектора pEF1/FCCU-1
Фрагмент ДНК (SEQ ID NO:34 в разделе “Список последовательностей”), содержащий последовательность ДНК, кодирующую сигнальную последовательность IgG1 и аминокислотную последовательность константной области, расщепляли рестрикционными ферментами NheI и PmeI, и лигировали с плазмидой pEF1/KCL, расщепленной NheI и PmeI, в результате чего сконструировали экспрессирующий тяжелую цепь человека вектор pEF1/FCCU-1, обладающий сигнальной последовательностью, участком клонирования, константной областью тяжелой цепи человека и сигнальной последовательностью человека для добавления поли-A, на 3'-конце от промотора EF1.
b) Получение экспрессирующей тяжелую цепь химерного антитела человека #32A1 конструкции
В качестве праймеров для амплификации кДНК вариабельной области тяжелой цепи #32A1 посредством ПЦР олигонуклеотид, обладающий последовательностью 5'-aaagctgagcGAGGTGCAAATTTTGGAGACTGGAGGAGGC-3' (32A1HF: SEQ ID NO:35 в разделе “Список последовательностей”) и олигонуклеотид, обладающий последовательностью 5'-aaagctgagctGACTGTGACCATGACTCCTTGGCCCCAG-3' (32A1HR: SEQ ID NO:36 в разделе “Список последовательностей”), синтезировали общепринятым способом.
К тому же, для вставки продукта ПЦР в pEF1/FCCU-1, эти праймеры сконструированы так, что добавлена последовательность узнавания рестрикционного фермента BlpI. ПЦР проводили общепринятым способом с использованием этого сочетания праймеров и кДНК вариабельной области тяжелой цепи, очищенной в e) из примера 17, в качестве матрицы. Полученный продукт ПЦР очищали с использованием набора для очистки продуктов ПЦР MinElute (QIAGEN, Inc.), и затем фрагмент ДНК, полученный расщеплением продукта ПЦР рестрикционным ферментом BlpI (NEW ENGLAND BIOLABS, Inc.), встраивали в pEF1/FCCU-1 в участок расщепления рестрикционного фермента BlpI, в результате чего сконструировали конструкцию, экспрессирующую тяжелую цепь химерного антитела человека #32A1. Последовательность вставки подтверждали анализом секвенированием. В качестве праймера для секвенирования олигонуклеотид, обладающий последовательностью 5'-TAATACGACTCACTATAGGG-3' (F11: SEQ ID NO:37 в разделе “Список последовательностей”) синтезировали общепринятым способом. Вектор экспрессии, в который можно правильно встраивать вставку, назван «32A1H/pEF1/FCCU».
c) Получение экспрессирующей легкую цепь химерного антитела человека #32A1 конструкции
В качестве праймеров для амплификации кДНК вариабельной области легкой цепи #32A1 посредством ПЦР олигонуклеотид, обладающий последовательностью 5'-aaacatatggcGACATTGTCTTGACCCAGTCTCCTGCTTTGG-3' (32A1LF: SEQ ID NO:38 в разделе “Список последовательностей”) и олигонуклеотид, обладающий последовательностью 5'-aaacgtacgTCTCAATTCCAGCTTGGTGCCTCCAGCG-3' (32A1LR SEQ ID NO:39 в разделе “Список последовательностей”) синтезировали общепринятым способом.
К тому же, для вставки продукта ПЦР в pEF6KCL, эти праймеры сконструированы так, что последовательность узнавания рестрикционного фермента NdeI добавлена к 32A1LF, и последовательность узнавания рестрикционного фермента BsiWI добавлена к 32A1LR. ПЦР проводили общепринятым способом с использованием этого сочетания праймеров и кДНК вариабельной области легкой цепи, очищенной в e) из примера 17, в качестве матрицы. Полученный продукт ПЦР очищали с использованием набора для очистки продуктов ПЦР MinElute (QIAGEN, Inc.), и затем фрагмент ДНК, полученный расщеплением продукта ПЦР рестрикционными ферментами NdeI (TaKaRa Bio, Inc.) и BsiWI (NEW ENGLAND BIOLABS, Inc.), встраивали в pEF6KCL в участок расщепления рестрикционными ферментами NdeI (TaKaRa Bio, Inc.) и BsiWI (NEW ENGLAND BIOLABS, Inc.), в результате чего сконструировали конструкцию, экспрессирующую легкую цепь химерного антитела человека #32A1. Последовательность вставки подтверждали анализом секвенированием. В качестве праймера для секвенирования использовали праймер F11, представленный как SEQ ID NO:37 в разделе “Список последовательностей”. Вектор экспрессии, в который можно правильно встраивать вставку, назван «32A1L/pEF6KCL».
Ген тяжелой цепи химерного антитела человека #32A1, полученный в b), обладает нуклеотидной последовательностью SEQ ID NO:40 в разделе “Список последовательностей” и кодирует аминокислотную последовательность SEQ ID NO:41 в разделе “Список последовательностей”. Аминокислотная последовательность, представленная аминокислотами под номерами 1-19 из SEQ ID NO:41, соответствует сигнальной последовательности для секреции, аминокислотная последовательность, представленная ее аминокислотами под номерами 20-140, соответствует вариабельной области тяжелой цепи, и аминокислотная последовательность, представленная ее аминокислотами под номерами 141-470, соответствует константной области тяжелой цепи. Кроме того, ген легкой цепи химерного антитела человека #32A1, полученный в c), обладает нуклеотидной последовательностью SEQ ID NO:42 в разделе “Список последовательностей” и кодирует аминокислотную последовательность SEQ ID NO:43 в разделе “Список последовательностей”. Аминокислотная последовательность, представленная аминокислотами под номерами 1-20 из SEQ ID NO:43, соответствует сигнальной последовательности для секреции, аминокислотная последовательность, представленная ее аминокислотами под номерами 21-132, соответствует вариабельной области легкой цепи, и аминокислотная последовательность, представленная ее аминокислотами под номерами 133-237, соответствует константной области легкой цепи.
Пример 19
Получение химерного антитела человека из моноклонального антитела крысы против Siglec-15 мыши #32A1
a) Получение химерного антитела человека
3×107 клеток из клеток 293 Freestyle в логарифмической фазе роста рассевали в 30 мл свежей среды для экспрессии для Freestyle 293 (Invitrogen, Inc.) (получали четыре партии, в которых 30-мл культуры брали в качестве одной партии) и подвергали культивированию с встряхиванием (125 об./мин.) при 37°C в инкубаторе с 8% CO2. 300 мкг полиэтиленимина (изготовленного в Polyscience, Inc. #24765) растворяли в 1 мл среды Opti-Pro SFM (изготовленной в Invitrogen, Inc.), и полученный раствор оставляли при комнатной температуре на 5 минут. Экспрессирующий тяжелую цепь химерного антитела человека #32A1 вектор 32A1H/pEF1/FCCU и экспрессирующий легкую цепь химерного антитела человека #32A1 вектор 32A1L/pEF6KCL, полученные в примере 18, подготавливали с использованием набора для очистки плазмид PureLink HiPure (Invitrogen, Inc.). 32A1H/pEF1/FCCU (15 мкг) и 32A1L/pEF6KCL (45 мкг), суспендировали в 1 мл среды Opti-Pro SFM (Invitrogen, Inc.), и полученную суспензию добавляли к 1 мл смешанной жидкости полиэтиленимин/Opti-Pro SFM, которую оставляли при комнатной температуре на 5 минут, и полученную смесь оставляли при комнатной температуре на дополнительные 5 минут. Затем 2 мл смешанной жидкости полиэтиленимин/векторы экспрессии/Opti-Pro SFM добавляли к каждой партии суспензии клеток 293 Freestyle, и продолжали культивирование с встряхиванием. После культивирования клеток в течение 7 суток при 37°C в 8% CO2, культуральный супернатант собирали из каждой партии.
b) Очистка химерного антитела человека
90 мл культурального супернатанта (для трех партий), полученного выше, фильтровали через фильтр (изготовленный в NALGENE, Inc., #295-4545), и 0,5 мл MabSelect SuRe (изготовленного в GE Healthcare Bio-science Co., Ltd., #17-5438-01), уравновешенного PBS, добавляли к фильтрату, и полученную смесь встряхивали в течение ночи при 80 об./мин. и 10°C. На следующие сутки, носитель собирали и промывали PBS, и затем антитело элюировали раствором 1 M аргинина (pH 4,0). Элюат наносили на колонку PD-10 (изготовленную в GE Healthcare Bio-science Co., Ltd., #17-0851-01) для замены жидкости на PBS, и затем концентрировали с помощью Amicon Ultra-4 (изготовленного в Millipore Co., Ltd., #UFC805008), в результате чего получили 1,2 мл химерного антитела человека (0,98 мг/мл). Концентрацию антитела рассчитывали по измерению при 280 нм, полученному с использованием Биофотометра Hitachi Diode Array U-0080D.
Пример 20
Оценка свойства связывания химерного антитела человека из моноклонального антитела крысы против Siglec-15 мыши #32A1 с белком Siglec-15 мыши
Конкурентное ингибирование антитела крысы #32A1 против очищенного химерного антитела человека, полученного в примере 19, определяли следующим способом. Siglec-15-Fc мыши, очищенный в примере 4, разводили до 1 мкг/мл с помощью PBS, разведенный раствор распределяли по 100 мкл/лунку в иммунопланшете (изготовленном в Nunc, Inc., #437111), и планшет оставляли стоять в течение ночи при 4°C, посредством чего белок адсорбировали на планшете. На следующие сутки каждую лунку промывали дважды раствором PBS-T (PBS, 0,05% (об./об.) Tween 20), раствор, полученный разведением обезжиренного молока (изготовленного в Morinaga Milk Industry Co., Ltd.) до 5% с помощью PBS, распределяли по 350 мкл/лунку, и планшет оставляли стоять при комнатной температуре в течение 2 часов. Жидкость из каждой лунки удаляли, и каждую лунку промывали 3 раза раствором PBS-T. Затем смешанный раствор (раствор PBS, содержащий обезжиренное молоко в конечной концентрации 0,5%), содержащий химерное антитело человека при 0,25 мкг/мл и антитело #32A1 или контрольное антитело IgG крысы (изготовленное в R&D Systems, Inc., #6-001-A) в различных концентрациях (0 мкг/мл, 0,016 мкг/мл, 0,05 мкг/мл, 0,16 мкг/мл, 0,5 мкг/мл, 1,66 мкг/мл или 5 мкг/мл) распределяли по 100 мкл/лунку, и планшет оставляли стоять при комнатной температуре в течение 2 часов. Затем каждую лунку промывали 3 раза раствором PBS-T, конъюгированное с щелочной фосфатазой антитело козы против IgG человека AffiniPure (изготовленное в Jackson ImmunoResearch, Inc., #109-055-097), разведенное в 2500 раз раствором TBS-T (TBS, 0,05% (об./об.) Tween 20), добавляли по 100 мкл/лунку, и планшет оставляли стоять при комнатной температуре в течение 1 часа. Жидкость из каждой лунки удаляли, и каждую лунку промывали 5 раз раствором TBS-T. Затем раствор флуоресцентного субстрата (изготовленный в Roche Co., Ltd., #11681982001) добавляли по 100 мкл/лунку, и позволяли проходить флуоресцентной реакции. Через 10 минут после добавления раствора флуоресцентного субстрата интенсивность флуоресценции измеряли с использованием считывателя для планшетов. В результате, показано, что антитело #32A1 ингибирует связывание химерного антитела человека с Siglec-15-Fc мыши зависимым от концентрации образом. Затем, когда антитело #32A1 и химерное антитело человека смешивали в такой же концентрации (1:1), показали приблизительно 40% конкурентное ингибирование. Таким образом, считают, что антитело #32A1 и химерное антитело человека обладают по существу одинаковой аффинностью для Siglec-15-Fc мыши. С другой стороны, для контрольного IgG крысы не показали конкурентного ингибирования (Фиг. 15).
Пример 21
Оценка биологической активности химерного антитела человека из моноклонального антитела крысы против Siglec-15 мыши #32A1 на основании теста образования остеокластов мыши и теста образования остеокластов человека
a) Тест образования остеокластов мыши
С использованием химерного антитела человека из моноклонального антитела крысы против Siglec-15 мыши #32A1, полученного в примере 19, исследовали эффект на дифференцировку остеокластов из неадгерентных клеток костного мозга мыши. Неадгерентные клетки костного мозга мыши, полученные способом из примера 8, получали при 1,5×105 клеток/мл в среде α-MEM, содержащей 10% FBS и 10 нг/мл M-CSF (изготовленный в R&D Systems, Inc.), полученный препарат клеток рассевали в каждую лунку 96-луночного планшета в количестве 200 мкл, и клетки культивировали в течение 2 суток в CO2 инкубаторе. Старый культуральный раствор из 96-луночного планшета удаляли, и в каждую лунку добавляли 100 мкл среды MEM-α, 100 мкл среды MEM-α, содержащей 10% FBS, к которой RANKL человека (RANKL, изготовленный в Peprotech, Inc.) и M-CSF добавляли для получения конечных концентраций 20 нг/мл и 10 нг/мл, соответственно. К культуральному раствору клеток моноклональное антитело крысы против Siglec-15 мыши #32A1, полученное в примере 6, или химерное антитело человека из моноклонального антитела крысы против Siglec-15 мыши #32A1, полученное в примере 19, добавляли в концентрации от 3 до 100 нг/мл, и клетки культивировали в течение дополнительных 3 суток в CO2 инкубаторе. После завершения культивирования активность устойчивой к тартрату кислой фосфатазы (TRAP) образованных остеокластов измеряли способом, описанным в примере 9. После остановки ферментативной реакции измеряли оптическую плотность в каждой лунке при 405 нм, и полученное измерение использовали в качестве показателя активности TRAP. Результаты показаны на фиг. 16. Зависимое от дозы ингибирование активности TRAP наблюдали при концентрации 50 нг/мл или выше в случае антитела #32A1 и химерного антитела человека из моноклонального антитела крысы против Siglec-15 мыши #32A1. По этому результату показали, что химерное антитело человека из моноклонального антитела крысы против Siglec-15 мыши #32A1 обладало активностью ингибирования образования остеокластов (дифференцировки и созревания остеокластов) по существу, сравнимой с активностью антитела крысы #32A1.
b) Оценка с использованием нормальных клеток-предшественников остеокластов человека (Окрашивание TRAP)
Нормальные клетки-предшественники остеокластов человека (Normal Human Natural Osteoclast Precursor Cells, закупленные из Sanko Junyaku Co., Ltd., Кат. No. 2T-110) рассевали в 96-луночный планшет по 1×104 клеток/лунку согласно протоколу, сопровождающему клетки. В качестве среды использовали минимальную поддерживающую среду для клеток-предшественников остеокластов (OPBM, закупленную из Sanko Junyaku Co., Ltd., Кат. No. PT-8201), дополненную набором добавок OPGM (закупленным из Sanko Junyaku Co., Ltd., Кат. No. PT-9501), содержащую эмбриональную бычью сыворотку (конечная концентрация: 10%), RANKL человека (конечная концентрация: 66 нг/мл), M-CSF человека (конечная концентрация: 33 нг/мл) и тому подобное. К полученному культуральному супернатанту добавляли химерное антитело человека из моноклонального антитела крысы против Siglec-15 мыши #32A1, полученное в примере 19, при конечной концентрации 0,3, 1, 3 или 10 мкг/мл, и клетки культивировали в течение 3 суток в CO2 инкубаторе. После культивирования супернатант удаляли, и 10% нейтральный формалин добавляли для фиксации клеток. После фиксации клеток клетки дважды промывали дистиллированной водой, окрашивающий раствор TRAP (0,27 мМ нафтол AS-MX фосфат (изготовленный в Sigma Co., Ltd.), 1,6 мМ LB соль быстрого красно-фиолетового (изготовленная в Sigma Co., Ltd.), 1% диметилформамид, 50 мМ тартрат натрия, 0,1 M буфер ацетат натрия (pH 5,0)) добавляли по 100 мкл/лунку, и реакции позволяли проходить при комнатной температуре в течение 5 минут. После реакции клетки дважды промывали дистиллированной водой, и затем наблюдали под микроскопом (Фиг. 17). В результате образование TRAP-положительных многоядерных остеокластов являлось почти полностью ингибированным добавлением химерного антитела человека #32A1 в диапазоне от 0,3 мкг/мл до 10 мкг/мл.
Пример 22
Конструирование гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1 (1)
a) Конструирование гуманизированного варианта #32A1
a)-i) Молекулярное моделирование вариабельной области #32A1
Молекулярное моделирование вариабельной области #32A1 проводили способом, общеизвестным как моделирование по гомологии (Methods in Enzymology, 203, 121-153, (1991)). Первичные последовательности (доступны трехмерные структуры, выведенные из рентгеновских кристаллических структур) вариабельных областей иммуноглобулина человека, задепонированные в Банке данных белка (Nuc. Acid Res. 35, D301-D303 (2007)), сравнивали с вариабельной областью #32A1, определенной выше. В результате 1LK3 выбрано как обладающее наибольшей гомологией последовательности с вариабельной областью легкой цепи #32A1 среди антител, обладающих сходной делецией в каркасной области. Кроме того, 1AD0 выбрано как обладающее наибольшей гомологией последовательности с вариабельной областью тяжелой цепи #32A1. Трехмерную структуру каркасной области составляли получением «модели каркаса» посредством сочетания координат 1LK3 и 1AD0, соответствующих легким и тяжелым цепям #32A1. CDR #32A1 классифицировали согласно классификации Thornton et al. (J. Mol. Biol., 263, 800-815, (1996)) следующим образом: CDRL1, CDRL2, CDRL3, CDRH1 и CDRH2 приписаны к 15A, 7A, 9A, 10A и 12B, соответственно. CDRH3 классифицированы как k(8)С согласно H3-правилам (FEBS letter 399, 1-8 (1996)). Затем репрезентативные конформации соответствующих CDR интегрировали в модель каркаса.
Наконец, для получения возможной молекулярной модели вариабельной области #32A1 в отношении энергии, проводили расчет энергии для исключения неблагоприятных межатомных контактов. Вышеуказанную процедуру проводили с использованием коммерчески доступной программы предсказания трехмерной структуры белка Prime и программы поиска координат MacroModel (Schrodinger, LLC).
a)-ii) Конструирование аминокислотной последовательности гуманизированного #32A1
Гуманизированное антитело #32A1 конструировали способом, общеизвестным как прививка CDR (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)). Акцепторное антитело выбирали двумя способами на основании гомологии аминокислот внутри каркасной области. Последовательность каркасной области #32A1 сравнивали с последовательностями всех каркасных областей человека в Базе данных Kabat (Nuc. Acid Res. 29, 205-206 (2001)), содержащей аминокислотные последовательности антител. В результате антитело M37GO37'CL выбрано в качестве акцептора на основании 73% гомологии последовательности в каркасной области. Остатки аминокислот в каркасной области M37GO37'CL выравнивали с остатками аминокислот #32A1 и идентифицировали положения, где использованы различные аминокислоты. Положения этих остатков анализировали с использованием трехмерной модели #32A1, сконструированной выше. Затем донорные остатки для вставки в акцептор выбирали согласно критериям, представленным в Queen et al. (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)).
Последовательность каркасной области #32A1 сравнивали с последовательностями всех каркасных областей человека в IgBLAST (Nuc Acid Res.36, D25-D30 (2007)). В результате, для L-цепи, AAB34430 выбрано в качестве акцептора на основании 79% гомологии последовательности в каркасной области. В отношении H-цепи, CAF31288 выбрано в качестве акцептора на основании 78% гомологии последовательности в каркасной области. В отношении L-цепи, остатки аминокислот в каркасной области AAB34430, и в отношении H-цепи, остатки аминокислот в каркасной области CAF31288, выравнивали с остатками аминокислот #32A1 и идентифицировали положения, где использованы различные аминокислоты. Положения этих остатков анализировали с использованием трехмерной модели #32A1, сконструированной выше. Затем донорные остатки для прививки в акцептор выбирали согласно критериям, представленным в Queen et al. (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)). В этих способах, посредством переноса некоторых выбранных донорных остатков в акцепторное антитело, последовательности гуманизированного #32A1 сконструированы, как описано в следующих примерах.
b) Гуманизация тяжелой цепи #32A1
b)-i) тяжелая цепь h#32A1-T1H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены аминокислот под номерами 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 43 (треонин), 61 (глутаминовая кислота), 89 (валин), 95 (аспарагиновая кислота), 96 (серин), 97 (глутаминовая кислота), 98 (серин), 100 (валин), 104 (валин), 105 (серин), 114 (изолейцин), 118 (треонин), 134 (валин), 135 (метионин), и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на лейцин, валин, серин, глутамин, аланин, глицин, фенилаланин, аспарагин, аланин, лизин, аспарагин, лейцин, метионин, аспарагин, валин, аланин, треонин, лейцин, и серин, соответственно, названа «тяжелая цепь h#32A1-T1H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T1H-типа представлена как SEQ ID NO:51 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-19 аминокислотной последовательности SEQ ID NO:51, последовательность, содержащая остатки аминокислот 20-140 из нее, и последовательность, содержащая остатки аминокислот 141-470 из нее, соответствуют сигнальной последовательности, вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:51, представлена как SEQ ID NO:50 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-57 нуклеотидной последовательности SEQ ID NO:50, последовательность, содержащая нуклеотиды 58-420 из нее, и последовательность, содержащая нуклеотиды 421-1410 из нее, кодируют сигнальную последовательность, последовательность вариабельной области тяжелой цепи и последовательность константной области тяжелой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:50 и аминокислотная последовательность SEQ ID NO:51 показаны также на фиг. 27.
b)-ii) тяжелая цепь h#32A1-T2H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены аминокислот под номерами 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 43 (треонин), 95 (аспарагиновая кислота), 96 (серин), 97 (глутаминовая кислота), 98 (серин), 100 (валин), 104 (валин), 114 (изолейцин), 118 (треонин), 134 (валин), 135 (метионин) и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на лейцин, валин, серин, глутамин, аланин, аспарагин, аланин, лизин, аспарагин, лейцин, метионин, валин, аланин, треонин, лейцин, и серин, соответственно, названа «тяжелая цепь h#32A1-T2H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T2H- типа представлена как SEQ ID NO:53 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-19 аминокислотной последовательности SEQ ID NO:53, последовательность, содержащая остатки аминокислот 20-140 из нее, и последовательность, содержащая остатки аминокислот 141-470 из нее, соответствуют сигнальной последовательности, вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:53, представлена как SEQ ID NO:52 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-57 из нуклеотидной последовательности SEQ ID NO:52, последовательность, содержащая нуклеотиды 58-420 из нее, и последовательность, содержащая нуклеотиды 421-1410 из нее, кодируют сигнальную последовательность, последовательность вариабельной области тяжелой цепи и последовательность константной области тяжелой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:52 и аминокислотная последовательность SEQ ID NO:53 показаны также на фиг. 28.
b)-iii) тяжелая цепь h#32A1-T3H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 26 (треонин), 32 (лизин), 104 (валин), 114 (изолейцин), 134 (валин), 135 (метионин), и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на валин, серин, глутамин, метионин, валин, треонин, лейцин, и серин, соответственно, названа «тяжелая цепь h#32A1-T3H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T3H-типа представлена как SEQ ID NO:55 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-19 аминокислотной последовательности SEQ ID NO:55, последовательность, содержащая остатки аминокислот 20-140 из нее, и последовательность, содержащая остатки аминокислот 141-470 из нее, соответствуют сигнальной последовательности, вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:55, представлена как SEQ ID NO:54 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-57 нуклеотидной последовательности SEQ ID NO:54, последовательность, содержащая нуклеотиды 58-420 из нее, и последовательность, содержащая нуклеотиды 421-1410 из нее, кодируют сигнальную последовательность, последовательность вариабельной области тяжелой цепи и последовательность константной области тяжелой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:54 и аминокислотная последовательность SEQ ID NO:55 показаны также на фиг. 29.
b)-iv) тяжелая цепь h#32A1-T5H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены аминокислот под номерами 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 61 (глутаминовая кислота), 89 (валин), 95 (аспарагиновая кислота), 97 (глутаминовая кислота), 99 (серин), 104 (валин), 105 (серин), 114 (изолейцин), 134 (валин), 135 (метионин) и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на валин, валин, серин, глутамин, глицин, фенилаланин, аспарагин, лизин, треонин, метионин, аспарагин, валин, треонин, лейцин и серин, соответственно, названа «тяжелая цепь h#32A1-T5H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T5H-типа представлена как SEQ ID NO:57 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-19 аминокислотной последовательности SEQ ID NO:57, последовательность, содержащая остатки аминокислот 20-140 из нее, и последовательность, содержащая остатки аминокислот 141-470 из нее, соответствуют сигнальной последовательности, вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:57, представлена как SEQ ID NO:56 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-57 нуклеотидной последовательности SEQ ID NO:56, последовательность, содержащая нуклеотиды 58-420 из нее, и последовательность, содержащая нуклеотиды 421-1410 из нее, кодируют сигнальную последовательность, последовательность вариабельной области тяжелой цепи и последовательность константной области тяжелой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:56 и аминокислотная последовательность SEQ ID NO:57 показаны также на фиг. 30.
b)-v) тяжелая цепь h#32A1-T6H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены аминокислот под номерами 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 95 (аспарагиновая кислота), 97 (глутаминовая кислота), 99 (серин), 104 (валин), 114 (изолейцин), 134 (валин), 135 (метионин) и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на валин, валин, серин, глутамин, аспарагин, лизин, треонин, метионин, валин, треонин, лейцин, и серин, соответственно, названа «тяжелая цепь h#32A1-T6H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T6H-типа представлена как SEQ ID NO:59 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-19 аминокислотной последовательности SEQ ID NO:59, последовательность, содержащая остатки аминокислот 20-140 из нее, и последовательность, содержащая остатки аминокислот 141-470 из нее, соответствуют сигнальной последовательности, вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:59, представлена как SEQ ID NO:58 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-57 нуклеотидной последовательности SEQ ID NO:58, последовательность, содержащая нуклеотиды 58-420 из нее, и последовательность, содержащая нуклеотиды 421-1410 из нее, кодируют сигнальную последовательность, последовательность вариабельной области тяжелой цепи и последовательность константной области тяжелой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:58 и аминокислотная последовательность SEQ ID NO:59 показаны также на фиг. 31.
b)-vi) тяжелая цепь h#32A1-T7H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены аминокислот под номерами 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 43 (треонин), 89 (валин), 95 (аспарагиновая кислота), 96 (серин), 97 (глутаминовая кислота), 98 (серин), 100 (валин), 104 (валин), 105 (серин), 114 (изолейцин), 118 (треонин), 134 (валин), 135 (метионин), и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на лейцин, валин, серин, глутамин, аланин, фенилаланин, аспарагин, аланин, лизин, аспарагин, лейцин, метионин, аспарагин, валин, аланин, треонин, лейцин, и серин, соответственно, названа «тяжелая цепь h#32A1-T7H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T7H-типа представлена как SEQ ID NO:72 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-121 аминокислотной последовательности SEQ ID NO:72 и последовательность, содержащая остатки аминокислот 122-451 из нее, соответствуют вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:72 показана также на фиг. 38.
b)-vii) тяжелая цепь h#32A1-T8H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены аминокислот под номерами 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 43 (треонин), 61 (глутаминовая кислота), 95 (аспарагиновая кислота), 96 (серин), 97 (глутаминовая кислота), 98 (серин), 100 (валин), 104 (валин), 105 (серин), 114 (изолейцин), 118 (треонин), 134 (валин), 135 (метионин) и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на лейцин, валин, серин, глутамин, аланин, глицин, аспарагин, аланин, лизин, аспарагин, лейцин, метионин, аспарагин, валин, аланин, треонин, лейцин и серин, соответственно, названа «тяжелая цепь h#32A1-T8H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T8H-типа представлена как SEQ ID NO:73 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-121 аминокислотной последовательности SEQ ID NO:73, и последовательность, содержащая остатки аминокислот 122-451 из нее, соответствуют вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:73 показана также на фиг. 38.
b-viii) тяжелая цепь h#32A1-T9H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены аминокислот под номерами 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 43 (треонин), 61 (глутаминовая кислота), 89 (валин), 95 (аспарагиновая кислота), 96 (серин), 97 (глутаминовая кислота), 98 (серин), 100 (валин), 104 (валин), 114 (изолейцин), 118 (треонин), 134 (валин), 135 (метионин) и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на лейцин, валин, серин, глутамин, аланин, глицин, фенилаланин, аспарагин, аланин, лизин, аспарагин, лейцин, метионин, валин, аланин, треонин, лейцин и серин, соответственно, названа «тяжелая цепь h#32A1-T9H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T9H-типа представлена как SEQ ID NO:74 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-121 аминокислотной последовательности SEQ ID NO:74, и последовательность, содержащая остатки аминокислот 122-451 из нее, соответствуют вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:74 показана также на фиг. 38.
b)-ix) тяжелая цепь h#32A1-T10H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены аминокислот под номерами 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 43 (треонин), 95 (аспарагиновая кислота), 96 (серин), 97 (глутаминовая кислота), 98 (серин), 100 (валин), 104 (валин), 105 (серин), 114 (изолейцин), 118 (треонин), 134 (валин), 135 (метионин), и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на лейцин, валин, серин, глутамин, аланин, аспарагин, аланин, лизин, аспарагин, лейцин, метионин, аспарагин, валин, аланин, треонин, лейцин, и серин, соответственно, названа «тяжелая цепь h#32A1-T10H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T10H-типа представлена как SEQ ID NO:75 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-121 аминокислотной последовательности SEQ ID NO:75 и последовательность, содержащая остатки аминокислот 122-451 из нее, соответствуют вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:75 показана также на фиг. 39.
b)-x) тяжелая цепь h#32A1-T11H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены аминокислот под номерами 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 43 (треонин), 89 (валин), 95 (аспарагиновая кислота), 96 (серин), 97 (глутаминовая кислота), 98 (серин), 100 (валин), 104 (валин), 114 (изолейцин), 118 (треонин), 134 (валин), 135 (метионин) и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на лейцин, валин, серин, глутамин, аланин, фенилаланин, аспарагин, аланин, лизин, аспарагин, лейцин, метионин, валин, аланин, треонин, лейцин, и серин, соответственно, названа «тяжелая цепь h#32A1-T11H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T11H-типа представлена как SEQ ID NO:76 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-121 аминокислотной последовательности SEQ ID NO:76, и последовательность, содержащая остатки аминокислот 122-451 из нее, соответствуют вариабельной области тяжелой цепи и константной области тяжелой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:76 показана также на фиг. 39.
b)-xi) тяжелая цепь h#32A1-T12H-типа:
Гуманизированная тяжелая цепь #32A1, сконструированная посредством замены остатков аминокислот 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 43 (треонин), 61 (глутаминовая кислота), 95 (аспарагиновая кислота), 96 (серин), 97 (глутаминовая кислота), 98 (серин), 100 (валин), 104 (валин), 114 (изолейцин), 118 (треонин), 134 (валин), 135 (метионин) и 140 (лейцин) из тяжелой цепи #32A1 SEQ ID NO:28 в разделе “Список последовательностей”, на лейцин, валин, серин, глутамин, аланин, глицин, аспарагин, аланин, лизин, аспарагин, лейцин, метионин, валин, аланин, треонин, лейцин и серин, соответственно, названа «тяжелая цепь h#32A1-T12H-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-T12H-типа представлена как SEQ ID NO:77 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-121 аминокислотной последовательности SEQ ID NO:77, и последовательность, содержащая остатки аминокислот 122-451 из нее, соответствуют вариабельной области тяжелой цепи и константной области тяжелой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:77 показана также на фиг. 39.
с) Гуманизация легкой цепи #32A1
c)-i) легкая цепь h#32A1-T1L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 41 (серин), 66 (глутамин), 68 (аргинин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 122 (аланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, аспарагин, пролин, лизин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, тирозин, глицин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T1L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T1L-типа представлена как SEQ ID NO:61 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-20 аминокислотной последовательности SEQ ID NO:61, последовательность, содержащая остатки аминокислот 21-133 из нее, и последовательность, содержащая остатки аминокислот 134-238 из нее, соответствуют сигнальной последовательности, вариабельной области легкой цепи, и константной области легкой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:61, представлена как SEQ ID NO:60 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-60 нуклеотидной последовательности SEQ ID NO:60, последовательность, содержащая нуклеотиды 61-399 из нее, и последовательность, содержащая нуклеотиды 400-714 из нее, кодируют сигнальную последовательность, последовательность вариабельной области легкой цепи, и последовательность константной области легкой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:60 и аминокислотная последовательность SEQ ID NO:61 показаны также на фиг. 32.
c)-ii) легкая цепь h#32A1-T2L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 66 (глутамин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 1101 (валин), 104 (aspartic acid), 106 (изолейцин), 108 (треонин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, пролин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, валин, изолейцин, лизин, и треонин, соответственно, названа «легкая цепь h#32A1-T2L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T2L-типа представлена как SEQ ID NO:63 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-20 аминокислотной последовательности SEQ ID NO:63, последовательность, содержащая остатки аминокислот 21-133 из нее, и последовательность, содержащая остатки аминокислот 134-238 из нее, соответствуют сигнальной последовательности, вариабельной области легкой цепи, и константной области легкой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:63, представлена как SEQ ID NO:62 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-60 нуклеотидной последовательности SEQ ID NO:62, последовательность, содержащая нуклеотиды 61-399 из нее, и последовательность, содержащая нуклеотиды 400-714 из нее, кодируют сигнальную последовательность, последовательность вариабельной области легкой цепи, и последовательность константной области легкой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:62 и аминокислотная последовательность SEQ ID NO:63 показаны также на фиг. 33.
c)-iii) легкая цепь h#32A1-T3L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 127 (лейцин), 129 (лейцин), 130 (аргинин), и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на аспарагиновую кислоту, серин, глутаминовую кислоту, серин, серин, лейцин, глутаминовую кислоту, валин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T3L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T3L-типа представлена как SEQ ID NO:65 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-20 аминокислотной последовательности SEQ ID NO:65, последовательность, содержащая остатки аминокислот 21-133 из нее, и последовательность, содержащая остатки аминокислот 134-238 из нее, соответствуют сигнальной последовательности, вариабельной области легкой цепи и константной области легкой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:65 представлена как SEQ ID NO:64 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-60 нуклеотидной последовательности SEQ ID NO:64, последовательность, содержащая нуклеотиды 61-399 из нее, и последовательность, содержащая нуклеотиды 400-714 из нее, кодируют сигнальную последовательность, последовательность вариабельной области легкой цепи, и последовательность константной области легкой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:64 и аминокислотная последовательность SEQ ID NO:65 показаны также на фиг. 34.
c)-iv) легкая цепь h#32A1-T4L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 36 (глутамин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 129 (лейцин), 130 (аргинин), и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на глутаминовую кислоту, серин, серин, лейцин, глутаминовую кислоту, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T4L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T4L-типа представлена как SEQ ID NO:67 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-20 аминокислотной последовательности SEQ ID NO:67, последовательность, содержащая остатки аминокислот 21-132 из нее, и последовательность, содержащая остатки аминокислот 133-237 из нее, соответствуют сигнальной последовательности, вариабельной области легкой цепи и константной области легкой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:67, представлена как SEQ ID NO:66 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-60 нуклеотидной последовательности SEQ ID NO:66, последовательность, содержащая нуклеотиды 61-396 из нее, и последовательность, содержащая нуклеотиды 400-711 из нее, кодируют сигнальную последовательность, последовательность вариабельной области легкой цепи, и последовательность константной области легкой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:66 и аминокислотная последовательность SEQ ID NO:67 показаны также на фиг. 35.
c)-v) легкая цепь h#32A1-T5L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 41 (серин), 66 (глутамин), 68 (аргинин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 110 (фенилаланин), 122 (аланин), 123 (глицин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на аспарагиновую кислоту, серин, глутаминовую кислоту, аспарагин, пролин, лизин, серин, серин, лейцин, глутаминовую кислоту, валин, тирозин, глицин, глутамин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T5L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T5L-типа представлена как SEQ ID NO:69 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-20 аминокислотной последовательности SEQ ID NO:69, последовательность, содержащая остатки аминокислот 21-133 из нее, и последовательность, содержащая остатки аминокислот 134-238 из нее, соответствуют сигнальной последовательности, вариабельной области легкой цепи и константной области легкой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:69, представлена как SEQ ID NO:68 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-60 нуклеотидной последовательности SEQ ID NO:68, последовательность, содержащая нуклеотиды 61-399 из нее, и последовательность, содержащая нуклеотиды 400-714 из нее, кодируют сигнальную последовательность, последовательность вариабельной области легкой цепи и последовательность константной области легкой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:68 и аминокислотная последовательность SEQ ID NO:69 показаны также на фиг. 36.
c)-vi) легкая цепь h#32A1-T6L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 66 (глутамин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на аспарагиновую кислоту, серин, глутаминовую кислоту, пролин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T6L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T6L-типа представлена как SEQ ID NO:71 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-20 аминокислотной последовательности SEQ ID NO:71, последовательность, содержащая остатки аминокислот 21-133 из нее, и последовательность, содержащая остатки аминокислот 134-238 из нее, соответствуют сигнальной последовательности, вариабельной области легкой цепи и константной области легкой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:71 представлена как SEQ ID NO:70 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-60 нуклеотидной последовательности SEQ ID NO:70, последовательность, содержащая нуклеотиды 61-399 из нее, и последовательность, содержащая нуклеотиды 400-714 из нее, кодируют сигнальную последовательность, последовательность вариабельной области легкой цепи и последовательность константной области легкой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:70 и аминокислотная последовательность SEQ ID NO:71 показаны также на фиг. 37.
c)-vii) легкая цепь h#32A1-T7L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 66 (глутамин), 68 (аргинин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 122 (аланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, пролин, лизин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, тирозин, глицин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T7L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T7L-типа представлена как SEQ ID NO:78 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:78, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:78 показана также на фиг. 40.
c)-viii) легкая цепь h#32A1-T8L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 41 (серин), 66 (глутамин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 122 (аланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, аспарагин, пролин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, тирозин, глицин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T8L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T8L-типа представлена как SEQ ID NO:79 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:79, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:79 показана также на фиг. 40.
c)-ix) легкая цепь h#32A1-T9L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 41 (серин), 66 (глутамин), 68 (аргинин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 122 (аланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, аспарагин, пролин, лизин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, глицин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T9L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T9L-типа представлена как SEQ ID NO:80 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ 10 NO:80, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:80 показана также на фиг. 40.
c)-x) легкая цепь h#32A1-T10L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 41 (серин), 66 (глутамин), 68 (аргинин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислота, серин, глутаминовую кислоту, аспарагин, пролин, лизин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, тирозин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T10L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T10L-типа представлена как SEQ ID NO:81 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:81, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:81 показана также на фиг. 40.
c)-xi) легкая цепь h#32A1-T11L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 66 (глутамин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 122 (аланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, пролин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, тирозин, глицин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T11L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T11L-типа представлена как SEQ ID NO:82 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:82, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:82 показана также на фиг. 40.
c)-xii) легкая цепь h#32A1-T12L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 66 (глутамин), 68 (аргинин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 122 (аланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, пролин, лизин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, глицин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T12L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T12L-типа представлена как SEQ ID NO:83 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:83, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:83 показана также на фиг. 41.
c)-xiii) легкая цепь h#32A1-T13L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 66 (глутамин), 68 (аргинин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, пролин, лизин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, тирозин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T13L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T13L-типа представлена как SEQ ID NO:84 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:84, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:84 показана также на фиг. 41.
c)-xiv) легкая цепь h#32A1-T14L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 41 (серин), 66 (глутамин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 122 (аланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, аспарагин, пролин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, глицин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T14L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T14L-типа представлена как SEQ ID NO:85 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:85 и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:85 показана также на фиг. 41.
c)-xv) легкая цепь h#32A1-T15L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 41 (серин), 66 (глутамин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 127 (лейцин), 129 (лейцин), 130 (аргинин), и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовая кислота, аспарагин, пролин, валин, серин, серин, лейцин, глутаминовая кислота, валин, валин, тирозин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T15L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T15L-типа представлена как SEQ ID NO:86 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:86 и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:86 показана также на фиг. 41.
c)-xvi) легкая цепь h#32A1-T16L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 41 (серин), 66 (глутамин), 68 (аргинин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 127 (лейцин), 129 (лейцин), 130 (аргинин), и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, аспарагин, пролин, лизин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T16L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T16L-типа представлена как SEQ ID NO:87 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:87, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:87 показана также на фиг. 41.
c)-xvii) легкая цепь h#32A1-T17L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 66 (глутамин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 122 (аланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, пролин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, глицин, валин, изолейцин, лизин и треонин, соответственно, названа «h#32A1-T17L-типа легкая цепь».
Аминокислотная последовательность легкой цепи h#32A1-T17L-типа представлена как SEQ ID NO:88 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:88, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:88 показана также на фиг. 42.
c)-xviii) легкая цепь h#32A1-T18L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 66 (глутамин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, пролин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, тирозин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T18L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T18L-типа представлена как SEQ ID NO:89 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:89 и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:89 показана также на фиг. 42.
c)-xix) легкая цепь h#32A1-T19L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 66 (глутамин), 68 (аргинин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, пролин, лизин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T19L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T19L-типа представлена как SEQ ID NO:90 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:90 и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:90 показана также на фиг. 42.
c)-xx) легкая цепь h#32A1-T20L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), 29 (аланин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 36 (глутамин), 41 (серин), 66 (глутамин), 81 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, аспарагиновую кислоту, серин, глутаминовую кислоту, аспарагин, пролин, валин, серин, серин, лейцин, глутаминовую кислоту, валин, валин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T20L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T20L-типа представлена как SEQ ID NO:91 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:91, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:91 показана также на фиг. 42.
c)-xxi) легкая цепь h#32A1-T21L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 21 (аспарагиновая кислота), 24 (лейцин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 31 (аланин), 32 (валин), 34 (лейцин), 36 (глутамин), 40 (изолейцин), 66 (глутамин), 99 (аспарагин), 100 (пролин), 101 (валин), 102 (глутамин), 103 (аланин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 122 (аланин), 123 (глицин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на глутаминовую кислоту, метионин, треонин, серин, лейцин, пролин, глутаминовую кислоту, лейцин, аланин, серин, серин, лейцин, глутаминовую кислоту, пролин, глутаминовую кислоту, фенилаланин, лейцин, тирозин, глицин, глутамин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T21L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T21L-типа представлена как SEQ ID NO:92 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:92 и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:92 показана также на фиг. 42.
c)-xxii) легкая цепь h#32A1-T22L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 21 (аспарагиновая кислота), 24 (лейцин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 31 (аланин), 32 (валин), 34 (лейцин), 36 (глутамин), 40 (изолейцин), 66 (глутамин), 99 (аспарагин), 100 (пролин), 101 (валин), 102 (глутамин), 103 (аланин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 123 (глицин), 127 (лейцин), 129 (лейцин), 130 (аргинин), и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на глутаминовую кислоту, метионин, треонин, серин, лейцин, пролин, глутаминовую кислоту, лейцин, аланин, серин, серин, лейцин, глутаминовую кислоту, пролин, глутаминовую кислоту, фенилаланин, лейцин, глутамин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T22L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T22L-типа представлена как SEQ ID NO:93 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:93, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:93 показана также на фиг. 43.
c)-xxiii) легкая цепь h#32A1-T23L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 24 (лейцин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 31 (аланин), 32 (валин), 34 (лейцин), 36 (глутамин), 40 (изолейцин), 66 (глутамин), 99 (аспарагин), 100 (пролин), 101 (валин), 102 (глутамин), 103, (аланин), 104 (aspartic acid), 106 (изолейцин), 108 (треонин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на метионин, треонин, серин, лейцин, пролин, глутаминовую кислоту, лейцин, аланин, серин, серин, лейцин, глутаминовую кислоту, пролин, глутаминовую кислоту, фенилаланин, лейцин, валин, изолейцин, лизин, и треонин, соответственно, названа «легкая цепь h#32A1-T23L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T23L-типа представлена как SEQ ID NO:94 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:94, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:94 показана также на фиг. 43.
c)-xxiv) легкая цепь h#32A1-T24L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами 21 (аспарагиновая кислота), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 31 (аланин), 32 (валин), 34 (лейцин), 36 (глутамин), 40 (изолейцин), 66 (глутамин), 99 (аспарагин), 100 (пролин), 101 (валин), 102 (глутамин), 103 (аланин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 122 (аланин), 123 (глицин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на глутаминовую кислоту, треонин, серин, лейцин, пролин, глутаминовую кислоту, лейцин, аланин, серин, серин, лейцин, глутаминовую кислоту, пролин, глутаминовую кислоту, фенилаланин, лейцин, тирозин, глицин, глутамин, валин, изолейцин, лизин и треонин, соответственно, названа «легкая цепь h#32A1-T24L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T24L-типа представлена как SEQ ID NO:95 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:95, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:95 показана также на фиг. 43.
c)-xxv) легкая цепь h#32A1-T25L-типа:
Гуманизированная легкая цепь #32A1, сконструированная посредством замены аминокислот под номерами между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 31 (аланин), 32 (валин), 34 (лейцин), 36 (глутамин), 40 (изолейцин), 99 (аспарагин), 100 (пролин), 101 (валин), 102 (глутамин), 103 (аланин), 104 (аспарагиновая кислота), 106 (изолейцин), 110 (фенилаланин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из легкой цепи #32A1 SEQ ID NO:30 в разделе “Список последовательностей”, на треонин, серин, лейцин, пролин, глутаминовую кислоту, лейцин, серин, серин, лейцин, глутаминовую кислоту, пролин, глутаминовую кислоту, фенилаланин, тирозин, валин, изолейцин, лизин, и треонин, соответственно, названа «легкая цепь h#32A1-T25L-типа».
Аминокислотная последовательность легкой цепи h#32A1-T25L-типа представлена как SEQ ID NO:96 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-113 аминокислотной последовательности SEQ ID NO:96, и последовательность, содержащая остатки аминокислот 114-218 из нее, соответствуют вариабельной области легкой цепи и константной области легкой цепи, соответственно. Аминокислотная последовательность SEQ ID NO:96 показана также на фиг. 43.
Пример 23
Получение гена гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1
a) Конструирование векторов, экспрессирующих легкие цепи h#32A1-T1L, h#32A1-T2L, h#32A1-T3L, h#32A1-T4L, h#32A1-T5L и h#32A1-T6L-типа
ДНК, каждая из которых содержит ген, кодирующий вариабельную область легкой цепи h#32A1-T1L, h#32A1-T2L, h#32A1-T3L, h#32A1-T4L, h#32A1-T5L или h#32A1-T6L-типа, слитый с секреторной сигнальной последовательностью, представленной: аминокислотами под номерами 1-133 из SEQ ID NO:61 в разделе “Список последовательностей”; аминокислотами под номерами 1-133 из SEQ ID NO:63; аминокислотами под номерами 1-133 из SEQ ID NO:65; аминокислотами под номерами 1-132 из SEQ ID NO:67; аминокислотами под номерами 1-133 из SEQ ID NO:69; или аминокислотами под номерами 1-133 из SEQ ID NO:71, синтезировали (Medical & Biological Laboratories Co., Ltd., Artificial Gene Synthesis Service). Затем, каждый из фрагментов ДНК, полученный расщеплением синтезированных ДНК с помощью рестрикционных ферментов NheI и BsiWI, встраивали в универсальный вектор для экспрессии легкой цепи гуманизированного антитела (pEF6KCL) в участок, расщепленный рестрикционными ферментами NheI и BsiWI, посредством чего конструировали векторы для экспрессии легкой цепи h#32A1-T1L, h#32A1-T2L, h#32A1-T3L, h#32A1-T4L, h#32A1-T5L, и h#32A1-T6L-типа. Полученные таким образом векторы экспрессии названы «pEF6KCL/h#32A1-T1L», «pEF6KCL/h#32A1-T2L», «pEF6KCL/h#32A1-T3L», «pEF6KCL/h#32A1-T4L», «pEF6KCL/h#32A1-T5L» и «pEF6KCL/h#32A1-T6L», соответственно.
b) Конструирование векторов, экспрессирующих тяжелую цепь h#32A1-T1H, h#32A1-T2H, h#32A1-T3H, h#32A1-T5H и h#32A1-T6H
ДНК, каждая из которых содержит ген, кодирующий вариабельную область тяжелой цепи h#32A1-T1H, h#32A1-T2H, h#32A1-T3H, h#32A1-T5H или h#32A1-T6H-типа, слитый с секреторной сигнальной последовательностью, представленной аминокислотами под номерами 1-140 из SEQ ID NO:51, 53, 55, 57 или 59 в разделе “Список последовательностей”, синтезировали (Medical & Biological Laboratories Co., Ltd., Artificial Gene Synthesis Service). Затем, каждый из фрагментов ДНК, полученный расщеплением синтезированных ДНК с помощью рестрикционным ферментом BlpI, встраивали в универсальный вектор для экспрессии тяжелой цепи гуманизированного антитела (pEF1/FCCU-1) в участок, расщепленный рестрикционным ферментом BlpI, посредством чего конструировали векторы для экспрессии тяжелой цепи h#32A1-T1H, h#32A1-T2H, h#32A1-T3H, h#32A1-T5H и h#32A1-T6H. Полученные таким образом векторы экспрессии названы «pEF1/FCCU/h#32A1-T1H», «pEF1/FCCU/h#32A1-T2H», «pEF1/FCCU/h#32A1-T3H», «pEF1/FCCU/h#32A1-T5H» и «pEF1/FCCU/h#32A1-T6H», соответственно.
Пример 24
Получение гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1
a) Получение гуманизированного антитела
1,5×108 клеток из клеток 293 Freestyle в логарифмической фазе роста рассевали в 100 мл свежей среды для экспрессии для Freestyle 293 (Invitrogen, Inc.) и подвергали культивированию с встряхиванием (125 об./мин.) при 37°C в инкубаторе с 8% CO2. 1 мг полиэтиленимина (изготовленного в Polyscience, Inc. #24765) растворяли в 4 мл среды Opti-Pro SFM (изготовленной в Invitrogen, Inc.), и полученный раствор оставляли при комнатной температуре на 5 минут. Плазмиду, экспрессирующую тяжелую цепь (0,05 мг), и плазмиду, экспрессирующую легкую цепь (0,15 мг), полученные с использованием набора PureLink HiPure (Invitrogen, Inc.), суспендировали в 4 мл среды Opti-Pro SFM (Invitrogen, Inc.). Затем 4 мл смешанной жидкости полученные экспрессирующие плазмиды/Opti-Pro SFM добавляли к 4 мл смешанной жидкости полиэтиленимин/Opti-Pro SFM, которую оставляли при комнатной температуре на 5 минут, и полученную смесь оставляли при комнатной температуре на дополнительные 5 минут. Затем 8 мл смешанной жидкости полиэтиленимин/экспрессирующие плазмиды/Opti-Pro SFM добавляли к суспензии клеток 293 Freestyle, и продолжали культивирование с встряхиванием. После культивирования клеток в течение 7 суток при 37°C в 8% CO2, культуральный супернатант собирали.
b) Очистка гуманизированного антитела
Культуральный супернатант, полученный в a) выше, очищали аффинной хроматографией с белком A. 100 мл культурального супернатанта помещали в 500-мл флакон с пробкой, и к этому добавляли 1 мл суспензии (50% взвесь) MabSelect SuRe (изготовленного в GE Healthcare Bio-science Co., Ltd.), уравновешенного PBS. Полученную смесь перемешивали в течение ночи при 100 об./мин. в инкубаторе при 10°C. Затем культуральный супернатант 293F/суспензию MabSelect SuRe наносили в пустую 5 мл центрифугируемую колонку Zeba (PIERCE, Inc.). После того, как всю смолу выливали в колонку, колонку промывали 10 мл 1 M NaCl. Затем, 1 мл раствора 1 M аргинина (pH 4,0) наносили на колонку, и собирали фракцию, содержащую антитело. Фракцию наносили в центрифугируемое фильтрующее устройство (Amicon Ultra-4, молекулярная масса фракции: 50 K, Millipore Co., Ltd.), и проводили замену жидкости на цитратный буфер и концентрирование. Конечный объем доводили до 200 мкл, которые использовали в качестве очищенного образца.
Гуманизированное антитело из #32A1, полученное посредством комбинации pEF6KCL/h#32A1-T1L и pEF1/FCCU/h#32A1-T1H, названо «h#32A1-T1»; гуманизированное антитело из #32A1, полученное посредством комбинации pEF6KCL/h#32A1-T2L и pEF1/FCCU/h#32A1-T2H, названо «h#32A1-T2»; гуманизированное антитело из #32A1, полученное посредством комбинации pEF6KCL/h#32A1-T3L и pEF1/FCCU/h#32A1-T3H, названо «h#32A1-T3»; гуманизированное антитело из #32A1, полученное посредством комбинации pEF6KCL/h#32A1-T4L и pEF1/FCCU/h#32A1-T3H, названо «h#32A1-T4»; гуманизированное антитело из #32A1, полученное посредством комбинации pEF6KCL/h#32A1-T5L и pEF1/FCCU/h#32A1-T5H, названо «h#32A1-T5»; и гуманизированное антитело из #32A1, полученное посредством комбинации pEF6KCL/h#32A1-T6L и pEF1/FCCU/h#32A1-T6H, названо «h#32A1-T6».
Пример 25
Оценка свойства связывания гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1 с белком Siglec-15 мыши
Свойство связывания гуманизированных антител из антитела крысы против Siglec-15 мыши #32A1 T1-T6 с Siglec-15-Fc мыши и Siglec-15-Fc человека оценивали следующим способом. Siglec-15-Fc мыши, очищенный в примере 4, или Siglec-15-Fc человека, очищенный в примере 12, разводили до 1 мкг/мл с помощью PBS, и разведенный раствор распределяли по 100 мкл/лунку в иммунопланшете (изготовленном в Nunc, Inc., #437111), и планшет оставляли стоять в течение ночи при 4°C, посредством чего каждый белок адсорбировали на планшете. На следующие сутки каждую лунку промывали 5 раз раствором PBS-T (PBS, 0,05% (об./об.) Tween 20), и раствор, полученный разведением обезжиренного молока (изготовленного в Morinaga Milk Industry Co., Ltd.) до 5% с помощью PBS, распределяли по 350 мкл/лунку, и планшет оставляли стоять при комнатной температуре в течение 2 часов. Жидкость из каждой лунки удаляли, и каждую лунку промывали 5 раз раствором PBS-T. Затем каждое из очищенных гуманизированных антител из антител крысы против Siglec-15 мыши #32A1 T1-T6, полученных в примере 24, или химерное антитело человека, полученное в примере 19, разводили до конечной концентрации от 1 до 0,004 мкг/мл (серии разведения в 4 раза) раствором PBS, содержащим 0,5% обезжиренное молоко, разведенный раствор распределяли по 100 мкл/лунку, и планшет оставляли стоять при комнатной температуре на 2 часа. В это время концентрацию каждого антитела определяли проведением измерения при 280 нм с использованием спектрофотометра DU-7400 (изготовленного в Beckman Coulter, Inc.) и проведением расчетов на основе молекулярного молярного коэффициента поглощения каждого антитела. Затем каждую лунку промывали 5 раз раствором PBS-T, конъюгированное с щелочной фосфатазой антитело козы против IgG человека (изготовленное в Jackson ImmunoResearch, Inc., #109-055-097), разведенное в 2500 раз раствором TBS-T (TBS, 0,05% (об./об.) Tween 20) добавляли по 100 мкл/лунку, и планшет оставляли стоять при комнатной температуре в течение 1 часа. Жидкость из каждой лунки удаляли, и каждую лунку промывали TBS-T. Затем раствор флуоресцентного субстрата (изготовленный в Roche Co., Ltd., #11681982001) добавляли по 100 мкл/лунку, и позволяли проходить флуоресцентной реакции. Через 10 минут после добавления раствора флуоресцентного субстрата в случае планшета с адсорбированным на нем Siglec-15-Fc мыши, и через 15 минут в случае планшета с адсорбированным на нем Siglec-15-Fc человека, интенсивность флуоресценции измеряли с использованием Spectra Max M5 (изготовленного в Molecular Devices, Inc.). В результате подтвердили, что все из исследуемых шести гуманизированных антител из антитела крысы против Siglec-15 мыши связывают белок Siglec-15 мыши (Фиг. 18) и белок Siglec-15 человека (Фиг. 19) зависимым от концентрации антител образом.
Пример 26
Эффект добавления химерного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1 на дифференцировку остеокластов из адгерентных клеток костного мозга мыши (стимуляция TNFα)
С использованием моноклонального антитела крысы против Siglec-15 мыши #32A1, полученного в примере 6, и химерного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1, полученного в примере 19, исследовали эффект на дифференцировку остеокластов из неадгерентных клеток костного мозга мыши при стимуляции TNFα. Неадгерентные клетки костного мозга мыши, полученные способом из примера 8, получали при 1,5×105 клеток/мл в среде α-MEM, содержащей 10% эмбриональную бычью сыворотку (FBS), 10 нг/мл M-CSF, и 2 нг/мл TGF-β (изготовленного в R&D Systems, Inc.), полученный препарат клеток рассевали в каждую лунку 96-луночного планшета в количестве 200 мкл, и клетки культивировали в течение 2 суток в CO2 инкубаторе. Старый культуральный раствор из 96-луночного планшета удаляли, и в каждую лунку добавляли 100 мкл среды MEM-α, 100 мкл среды MEM-α, содержащей 10% FBS, к которой рекомбинантный TNFα человека (изготовленный в R&D Systems, Inc.) и M-CSF добавляли для получения конечных концентраций 30 нг/мл и 10 нг/мл, соответственно. К культуральному раствору клеток моноклональное антитело крысы против Siglec-15 мыши #32A1, полученное в примере 9, или химерное антитело из моноклонального антитела крысы против Siglec-15 мыши #32A1, полученное в примере 19, добавляли в концентрации от 0,125 до 4 мкг/мл, и клетки культивировали в течение дополнительных 3 суток в CO2 инкубаторе. В то же самое время подготавливали также лунку, в которой клетки культивировали при добавлении рекомбинантного OCIF/OPG человека, полученного способом, описанным в описании Публикации Патентной заявки No. WO 96/26217, в концентрации 100 нг/мл. После завершения культивирования окрашивание TRAP проводили с использованием набора с кислой фосфатазой лейкоцитов (изготовленного в Sigma Co., Ltd.) согласно протоколу, сопровождающему набор, и наблюдали образование TRAP-положительных многоядерных остеокластов. В результате образование TRAP-положительных гигантских многоядерных остеокластов являлось ингибированным посредством добавления поликлонального антитела против Siglec-15 мыши (Фиг. 20). Хотя малые одноядерные остеокласты образовывались даже в случае, когда добавляли моноклональное антитело крысы против Siglec-15 мыши #32A1 или полученное из него химерное антитело, образование больших многоядерных остеокластов, индуцированное TNFα, являлось почти полностью ингибированным. С другой стороны, даже при добавлении достаточного количества OCIF/OPG, который является блокатором RANKL, образование больших многоядерных остеокластов являлось только немного ингибированным. Кроме того, таким же способом, активность устойчивой к тартрату кислой фосфатазы (TRAP) образованных остеокластов измеряли способом, описанным в примере 9. После остановки ферментативной реакции, измеряли оптическую плотность в каждой лунке при 405 нм, и полученное измерение использовали в качестве показателя активности TRAP. Результаты показаны на фиг. 21. Значительное ингибирование активности TRAP наблюдали в случае антитела #32A1 и химерного антитела #32A1 в диапазоне от 125 нг/мл до 4 мкг/мл. По этим результатам показано, что моноклональное антитело крысы против Siglec-15 мыши и полученное из него химерное антитело сильно ингибируют процесс слияния клеток при дифференцировке и созревании остеокластов, индуцированных TNFα.
Пример 27
Оценка биологической активности моноклональных антител крысы против Siglec-15 мыши #8A1 и #32A1 на основании теста образования остеокластов крысы
С использованием моноклональных антител крысы против Siglec-15 мыши #8A1 и #32A1, полученных в примере 6, оценивали эффект на дифференцировку остеокластов из неадгерентных клеток костного мозга крысы. Неадгерентные клетки костного мозга крысы получали посредством модификации способа из примера 8. То есть, бедренную кость вырезали из обеих ног самки крысы F344 в возрасте 12 недель и удаляли мягкие ткани. Оба конца бедренной кости отрезали, и среду MEM-α инъецировали с использованием шприца с иглой 23 калибра для инъекций, чтобы вытолкнуть клетки костного мозга, которые собирали в пробирку для центрифугирования. Центрифугирование проводили при комнатной температуре в течение 5 минут при 100 g, и супернатант удаляли. К полученному осадку клеток добавляли 1 мл гемолитического буфера (Red Blood Cell Lysing Buffer, изготовленного в Sigma Co., Ltd.), чтобы суспендировать его, и полученную суспензию оставляли при комнатной температуре в течение 5 минут. К ней добавляли 20 мл D-PBS, суспензию центрифугировали при комнатной температуре в течение 5 минут при 100 g, и супернатант удаляли. К полученному осадку клеток добавляли 10 мл среды MEM-α (изготовленной в Invitrogen, Inc.), содержащей 5 нг/мл M-CSF (изготовленной в R&D Systems, Inc.) и 10% эмбриональную бычью сыворотку (FBS) добавляли, чтобы суспендировать его. Затем полученную суспензию пропускали через клеточное сито (40 мкм Nylon, изготовленный в BD Falcon) для удаления агрегатов. Полученные клетки переносили в 75 см2 T-флакон (для прикрепления адгерентных клеток) и культивировали в течение ночи в CO2 инкубаторе. После культивирования в течение ночи клетки, у которых не произошла адгезия к T-флакону, выделяли и использовали в качестве неадгерентных клеток костного мозга крысы.
Неадгерентные клетки костного мозга крысы, полученные этим способом, культивировали и образования остеокластов крысы проводили способом из примера 9. То есть, полученные таким образом неадгерентные клетки костного мозга крысы получали при 1,5×105 клеток/мл в среде α-MEM, содержащей 10% FBS и 10 нг/мл M-CSF (изготовленной в R&D Systems, Inc.), и полученный препарат клеток рассевали в каждую лунку 96-луночного планшета в количестве 200 мкл, и клетки культивировали в течение 2 суток в CO2 инкубаторе. Старый культуральный раствор из 96-луночного планшета удаляли, и 100 мкл среды MEM-α добавляли в каждую лунку, 100 мкл среды MEM-α, содержащей 10% FBS, в которую добавляли RANKL человека (RANKL, изготовленный в Peprotech, Inc.) и M-CSF для получения конечных концентраций 20 нг/мл и 10 нг/мл, соответственно. К культуральному раствору клеток моноклональное антитело крысы против Siglec-15 мыши #8A1 или #32A1, полученное в примере 6, добавляли в концентрации от 31 до 1000 нг/мл, и клетки культивировали в течение дополнительных 3 суток в CO2 инкубаторе. После завершения культивирования активность устойчивой к тартрату кислой фосфатазы (TRAP) образованных остеокластов измеряли способом, описанным в примере 9. После остановки ферментативной реакции измеряли оптическую плотность в каждой лунке при 405 нм, и полученное измерение использовали в качестве показателя активности TRAP. Результаты показаны на фиг. 22. В случае антитела #8A1, зависимое от концентрации ингибирование активности TRAP наблюдали при концентрации 63 нг/мл или выше, и в случае антитела #32A1, зависимое от концентрации ингибирование активности TRAP наблюдали при концентрации 31 нг/мл или выше. По этим результатам показали, что моноклональные антитела крысы против Siglec-15 мыши #8A1 и #32A1 сильно ингибируют образование остеокластов крысы таким же образом, как и образование остеокластов мыши, и активность in vitro ингибирования образования остеокластов крысы антитела #32A1 является более высокой, чем активность антитела #8A1.
Пример 28
Конструирование гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1 (2)
a) Конструирование гуманизированного варианта #32A1
a)-i) Конструирование аминокислотной последовательности гуманизированного #32A1
Гуманизированное антитело #32A1 конструировали способом, общеизвестным как прививка CDR (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)). В отношении определения областей CDR, определения Kabat (SEQUENCES OF PROTEINS OF IMMUNOLOGICAL INTEREST VOL. I, FIFTH EDTION (1991)) использовали для всех областей, за исключением CDRH2. В отношении CDRH2, ее последовательность сделана короче на пять остатков с С-конца, чем по определению Kabat, с использованием собственного определения. В последовательности тяжелой цепи, содержащей определенную таким образом CDRH2, последовательность CDR, происходящая из крысы, сделана короче, и включено больше последовательностей из каркасной области человека, и таким образом, при ее введении человеку, она с намного меньшей вероятностью будет узнана как гетерологичный антиген. Акцепторное антитело выбрано на основании гомологии аминокислот с каркасной областью. Последовательность каркасной области #32A1 сравнивали с последовательностями всех каркасных областей человека в Базе данных Kabat (Nuc. Acid Res. 29, 205-206 (2001)), включающей аминокислотные последовательности антител. В результате, для L-цепи, GF4/1.1'CL выбрано в качестве акцептора на основании 71% гомологии последовательности в каркасной области. В отношении H-цепи, M37GO37'CL выбрано в качестве акцептора на основании 73% гомологии последовательности в каркасной области. В отношении L-цепи, остатки аминокислот в каркасной области GF4/1.1'CL, и в отношении H-цепи, остатки аминокислот в каркасной области M37GO37'CL выравнивали с остатками аминокислот #32A1, и идентифицировали положения, где использованы различные аминокислоты. Положения этих остатков анализировали с использованием трехмерной модели #32A1, сконструированной в a)-i) из примера 22. Затем донорные остатки для прививки в акцептор выбирали согласно критериям, представленным в Queen et al. (Proc Natl. Acad. Sci. USA 86, 10029-10033 (1989)). Посредством переноса некоторых избранных донорных остатков в акцепторное антитело, последовательности гуманизированного #32A1 сконструированы, как описано в следующих примерах.
a)-ii) Гуманизация тяжелой цепи #32A1
a)-ii)-i) тяжелая цепь h#32A1-H1-1-типа:
Гуманизированная тяжелая цепь 32A1, сконструированная посредством замены аминокислот под номерами 23 (изолейцин), 24 (лейцин), 26 (треонин), 32 (лизин), 43 (треонин), 61 (глутаминовая кислота), 83 (глутаминовая кислота), 85 (лейцин), 86 (глутаминовая кислота), 89 (валин), 95 (аспарагиновая кислота), 96 (серин), 97 (глутаминовая кислота), 98 (серин), 100 (валин), 104 (валин), 105 (серин), 114 (изолейцин), 118 (треонин), 134 (валин), 135 (метионин) и 140 (лейцин) из 32A1, представленного на SEQ ID NO:28 в разделе “Список последовательностей”, на лейцин, валин, серин, глутамин, аланин, глицин, аланин, валин, лизин, фенилаланин, аспарагин, аланин, лизин, аспарагин, лейцин, метионин, аспарагин, валин, аланин, треонин, лейцин, и серин, соответственно, названа «тяжелая цепь h#32A1-H1-1-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-H1-1-типа представлена как SEQ ID NO:99 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-19 аминокислотной последовательности SEQ ID NO:99, последовательность, содержащая остатки аминокислот 20-140 из нее, и последовательность, содержащая остатки аминокислот 141-466 из нее, соответствуют сигнальной последовательности, вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:99 представлена как SEQ ID NO:98 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-57 нуклеотидной последовательности SEQ ID NO:98, последовательность, содержащая нуклеотиды 58-420 из нее, и последовательность, содержащая нуклеотиды 421-1398 из нее, кодируют сигнальную последовательность, последовательность вариабельной области тяжелой цепи и последовательность константной области тяжелой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:98 и аминокислотная последовательность SEQ ID NO:99 показаны также на фиг. 54.
a)-iii) Гуманизация легкой цепи #32A1
a)-iii)-i) легкая цепь h#32A1-L2-15-типа:
Гуманизированная легкая цепь 32A1, сконструированная посредством замены аминокислот под номерами 21 (аспарагиновая кислота), 23 (валин), 24 (лейцин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 31 (аланин), 32 (валин), 34 (лейцин), 36 (глутамин), 40 (изолейцин), 66 (глутамин), 99 (аспарагин), 100 (пролин), 101 (валин), 102 (глутамин), 103 (аланин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 110 (фенилаланин), 122 (аланин), 123 (глицин), 127 (лейцин), 129 (лейцин), 130 (аргинин), и 132 (аланин) из 32A1, представленного на SEQ ID NO:30 в разделе “Список последовательностей”, на глутаминовую кислоту, лейцин, метионин, треонин, серин, лейцин, пролин, глутаминовую кислоту, лейцин, аланин, серин, серин, лейцин, глутаминовую кислоту, пролин, глутаминовую кислоту, фенилаланин, лейцин, тирозин, глицин, глутамин, валин, изолейцин, лизин, и треонин, соответственно, названа «легкая цепь h#32A1-L2-15-типа».
Аминокислотная последовательность легкой цепи h#32A1-L2-15-типа представлена как SEQ ID NO:103 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-20 аминокислотной последовательности SEQ ID NO:103, последовательность, содержащая остатки аминокислот 21-133 из нее, и последовательность, содержащая остатки аминокислот 134-238 из нее, соответствуют сигнальной последовательности, вариабельной области легкой цепи, и константной области легкой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:103, представлена как SEQ ID NO:102 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-60 нуклеотидной последовательности SEQ ID NO:102, последовательность, содержащая нуклеотиды 61-399 из нее, и последовательность, содержащая нуклеотиды 400-714 из нее, кодируют сигнальную последовательность, последовательность вариабельной области легкой цепи, и последовательность константной области легкой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:102 и аминокислотная последовательность SEQ ID NO:103 показаны также на фиг. 56.
a)-iii)-ii) легкая цепь h#32A1-L2-16-типа:
Гуманизированная легкая цепь 32A1, сконструированная посредством замены аминокислот под номерами 21 (аспарагиновая кислота), 23 (валин), 24 (лейцин), между 29 и 30 (т.е. вставки остатка, отсутствующего в каркасной области крысы), 31 (аланин), 32 (валин), 34 (лейцин), 36 (глутамин), 40 (изолейцин), 66 (глутамин), 99 (аспарагин), 100 (пролин), 101 (валин), 102 (глутамин), 103 (аланин), 104 (аспарагиновая кислота), 106 (изолейцин), 108 (треонин), 123 (глицин), 127 (лейцин), 129 (лейцин), 130 (аргинин) и 132 (аланин) из 32A1, представленного на SEQ ID NO:30 в разделе “Список последовательностей”, на глутаминовую кислоту, лейцин, метионин, треонин, серин, лейцин, пролин, глутаминовую кислоту, лейцин, аланин, серин, серин, лейцин, глутаминовую кислоту, пролин, глутаминовую кислоту, фенилаланин, лейцин, глутамин, валин, изолейцин, лизин, и треонин, соответственно, названа «легкая цепь h#32A1-L2-16-типа».
Аминокислотная последовательность легкой цепи h#32A1-L2-16-типа представлена как SEQ ID NO:105 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-20 аминокислотной последовательности SEQ ID NO:105, последовательность, содержащая остатки аминокислот 21-133 из нее, и последовательность, содержащая остатки аминокислот 134-238 из нее, соответствуют сигнальной последовательности, вариабельной области легкой цепи и константной области легкой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:105, представлена как SEQ ID NO:104 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-60 нуклеотидной последовательности SEQ ID NO:104, последовательность, содержащая нуклеотиды 61-399 из нее, и последовательность, содержащая нуклеотиды 400-714 из нее, кодируют сигнальную последовательность, последовательность вариабельной области легкой цепи, и последовательность константной области легкой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:104 и аминокислотная последовательность SEQ ID NO:105 показаны также на фиг. 57.
b) Получение гена гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1
b)-i) Конструирование универсального вектора pEG2, экспрессирующего тяжелую цепь человека.
Фрагмент ДНК, содержащий последовательность ДНК, кодирующую аминокислоты из сигнальной последовательности и константной области IgG2 человека SEQ ID NO:106, синтезировали (Medical & Biological Laboratories Co., Ltd., Artificial Gene Synthesis Service), и расщепляли рестрикционными ферментами NheI и PmeI, посредством чего получали фрагмент ДНК приблизительно 1100 п.о. Этот фрагмент ДНК лигировали с фрагментом ДНК приблизительно 6200 п.о., полученный расщеплением pEF1/FCCU-1 с помощью NheI и PmeI и удалением фрагмента ДНК приблизительно 1100 п.о., посредством чего конструировали универсальный вектор pEG2, экспрессирующий тяжелую цепь гуманизированного антитела (hIgG2-типа).
b)-ii) Конструирование вектора, экспрессирующего тяжелую цепь h#32A1-H1-1
Фрагмент ДНК, полученный синтезом ДНК, содержащей ген, кодирующий вариабельную область тяжелой цепи h#32A1-H1-1, слитую с сигнальной последовательностью для секреции, представленной аминокислотами под номерами 1-140 из SEQ ID NO:99 в разделе “Список последовательностей” (Invitrogen), и расщеплением ДНК ресткриционным ферментом BlpI, встраивали в универсальный вектор (pEG2), экспрессирующий тяжелую цепь гуманизированного антитела (hIgG2-типа), в участок расщепления рестрикционного фермента BlpI, посредством чего конструировали универсальный вектор, экспрессирующий тяжелую цепь h#32A1-H1-1. Полученный таким образом вектор экспрессии назван «pEG2/h#32A1-H1-1».
b)-iii) Конструирование вектора, экспрессирующего тяжелую цепь h#32A1-H5
Гуманизированная тяжелая цепь 32A1, в которой сигнальная последовательность и константная область тяжелой цепи h#32A1-T5H-типа, сконструированной в b)-iv) из примера 22, заменены на последовательности из IgG2 типа, названа «тяжелая цепь h#32A1-H5-типа».
Аминокислотная последовательность тяжелой цепи h#32A1-H5-типа представлена как SEQ ID NO:101 в разделе “Список последовательностей”. Последовательность, содержащая остатки аминокислот 1-19 аминокислотной последовательности SEQ ID NO:101, последовательность, содержащая остатки аминокислот 20-140 из нее, и последовательность, содержащая остатки аминокислот 141-466 из нее, соответствуют сигнальной последовательности, вариабельной области тяжелой цепи, и константной области тяжелой цепи, соответственно. Нуклеотидная последовательность, кодирующая аминокислотную последовательность SEQ ID NO:101, представлена как SEQ ID NO:100 в разделе “Список последовательностей”. Последовательность, содержащая нуклеотиды 1-57 нуклеотидной последовательности SEQ ID NO:100, последовательность, содержащая нуклеотиды 58-420 из нее, и последовательность, содержащая нуклеотиды 421-1398 из нее, кодируют сигнальную последовательность, последовательность вариабельной области тяжелой цепи и последовательность константной области тяжелой цепи, соответственно. Нуклеотидная последовательность SEQ ID NO:100 и аминокислотная последовательность SEQ ID NO:101 показаны также на фиг. 55.
Фрагмент ДНК, полученный расщеплением pEF1/FCCU/h#32A1-T5H, полученной в примере 23, с помощью рестрикционного фермента BlpI, встраивали в универсальный вектор (pEG2), экспрессирующий тяжелую цепь гуманизированного антитела (hIgG2 типа) в участок расщепления рестрикционным ферментом BlpI, посредством чего сконструировали вектор, экспрессирующий тяжелую цепь h#32A1-H5. Полученный таким образом вектор экспрессии назван «pEG2/h#32A1-H5».
b)-iv) Конструирование векторов, экспрессирующих легкие цепи h#32A1-L2-15 и h#32A1-L2-16-типа
ДНК, каждая из которых содержит ген, кодирующий вариабельную область легкой цепи h#32A1-L2-15 или h#32A1-L2-16-типа, слитый с секреторной сигнальной последовательностью, представленной аминокислотами под номерами 1-133 SEQ ID NO:103 в разделе “Список последовательностей” или аминокислотами под номерами 1-133 из SEQ ID NO:105, синтезировали (Invitrogen), и векторы, экспрессирующие легкие цепи h#32A1-L2-15 и h#32A1-L2-16-типа, конструировали таким же способом, как в примере 23. Полученные таким образом векторы экспрессии названы «pEF6KCL/h#32A1-L2-15» и «pEF6KCL/h#32A1-L2-16», соответственно.
c) Получение гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1
c)-i) Получение гуманизированного антитела
1,2×109 клеток из клеток Freestyle 293F (Invitrogen) в логарифмической фазе роста рассевали в 1,2 л свежей среды для экспрессии для Freestyle 293 (Invitrogen) и подвергали культивированию с встряхиванием при 37°C в инкубаторе с 8% CO2 при 90 об./мин. в течение 1 часа. 3,6 мг полиэтиленимина (Polyscience, #24765) растворяли в 20 мл среды Opti-Pro SFM (Invitrogen). Затем плазмиду, экспрессирующую H-цепь, (0,4 мг) и плазмиду, экспрессирующую L-цепь (0,8 мг), полученные с использованием набора для очистки плазмид PureLink HiPure (Invitrogen, Inc.) суспендировали в 20 мл среды Opti-Pro SFM. Затем 20 мл полученной смешанной жидкости экспрессирующие плазмиды/Opti-Pro SFM добавляли к 20 мл смешанной жидкости полиэтиленимин/Opti-Pro SFM, и полученную смесь осторожно перемешивали и затем оставляли на 5 минут. Затем смесь добавляли к клеткам Freestyle 293F, и культивирование с встряхиванием проводили в течение 7 суток при 37 °C в инкубаторе с 8% CO2 при 90 об./мин. Полученный культуральный супернатант фильтровали через одноразовый капсульный фильтр (Advantec #CCS-045-E1H).
Гуманизированное антитело из #32A1, полученное посредством комбинации pEF6KCL/h#32A1-T5L и pEG2/h#32A1-H5, названо «h#32A1-H5/L5»; гуманизированное антитело из #32A1, полученное посредством комбинации pEF6KCL/h#32A1-T5L и pEG2/h#32A1-H1-1, названо «h#32A1-H1-1/L5»; гуманизированное антитело из #32A1, полученное посредством комбинации pEF6KCL/h#32A1-L2-15 и pEG2/h#32A1-H1-1, названо «h#32A1-H1-1/L2-15»; и гуманизированное антитело из #32A1, полученное посредством комбинации pEF6KCL/h#32A1-L2-16 и pEG2/h#32A1-H1-1, названо «h#32A1-H1-1/L2-16».
c)-ii) Очистка гуманизированного антитела
Культуральный супернатант, полученный в c)-i) выше, очищали двухстадийным способом, включающим аффинную хроматографию с р-белком A (при 4-6°C) и керамическим гидроксиапатитом (при комнатной температуре). Стадию замены буфера после аффинной хроматографии с р-белком A и после очистки посредством керамического гидроксиапатита проводили при комнатной температуре. Сначала 1100-1200 мл культурального супернатанта наносили на MabSelect SuRe (изготовленный в GE Healthcare Bioscience Co., Ltd., 2×1 мл колонки HiTrap, соединенные последовательно), уравновешенные PBS. Затем культуральный раствор наливали в колонку, колонку промывали 15-30 мл PBS. Затем проводили элюцию раствором 2 M гидрохлорида аргинина (pH 4,0), и собирали фракцию, содержащую антитело. Фракцию наносили на обессоливающую колонку (изготовленную в GE Healthcare Bioscience Co., Ltd., 2×5 мл обессоливающие колонки HiTrap, соединенные последовательно), и жидкость заменяли буфером, содержащим 5 мМ фосфат натрия, 50 мМ MES, и 20 мМ NaCl при pH 6,5. Затем раствор антитела, подвергнутый замене буфера, наносили на колонку с керамическим гидроксиапатитом (Japan Bio-Rad Laboratories, Inc., колонка с гидроксиапатитом Bio-Scale CHT2-1 (объем 2 мл)), уравновешенную буфером, содержащим 5 мМ NaPi, 50 мМ MES и 20 мМ NaCl при pH 6,5. Затем проводили элюцию линейным градиентом концентраций хлорида натрия, и собирали фракцию, содержащую антитело. Фракцию наносили на обессоливающую колонку (изготовленную в GE Healthcare Bioscience Co., Ltd., 2×5 мл обессоливающие колонки HiTrap, соединенные последовательно) и жидкость заменяли на CBS (10 мМ цитратный буфер, содержащий 140 мМ хлорид натрия, pH 6,0). Наконец, полученный раствор концентрировали с использованием центрифугируемого устройства для фильтрации UF VIVASPIN 20 (фракционируемая молекулярная масса: 30 K, Sartorius Co., Ltd., при 4°C), концентрацию IgG доводили до 1,0 мг/мл или более, и полученный таким образом раствор использовали в качестве очищенного образца. Объемы соответствующих очищенных образцов являлись следующими: h#32A1-H1-1/L5: 2,2 мл, h#32A1-H5/L5: 1,8 мл, h#32A1-H1-1/L2-16: 6,0 мл, и h#32A1-H1-1/L2-15: 2,2 мл.
Пример 29
Оценка биологической активности гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1 на основании теста образования остеокластов мыши
Ингибирующий эффект на образование остеокластов мыши, показанный для гуманизированных антител, полученных в примерах 24 и 28, оценивали посредством частичной модификации способа, описанного в примере 9. Неадгерентные клетки костного мозга мыши, полученные способом из примера 8, получали при 1,5×105 клеток/мл в среде α-MEM, содержащей 10% FBS и 10 нг/мл M-CSF (изготовленной в R&D Systems, Inc. ), полученный препарат клеток рассевали в каждую лунку 96-луночного планшета в количестве 200 мкл, и клетки культивировали в течение 2 суток в CO2 инкубаторе. Старый культуральный раствор из 96-луночного планшета удаляли, и 100 мкл среды MEM-α добавляли в каждую лунку, 100 мкл среды MEM-α, содержащей 10% FBS, в которую добавляли RANKL человека (RANKL, изготовленный в Peprotech, Inc.) и M-CSF для получения конечных концентраций 20 нг/мл и 10 нг/мл, соответственно. К культуральному раствору клеток моноклональное антитело крысы против Siglec-15 мыши (антитело #32A1), полученное в примере 6, или каждое из 6 типов гуманизированных антител #32A1 (h#32A1-T1, h#32A1-T2, h#32A1-T3, h#32A1-T4, h#32A1-T5 и h#32A1-T6), полученных в примере 24, добавляли в концентрации от 3 до 100 нг/мл, и клетки культивировали в течение дополнительных 3 суток в CO2 инкубаторе. После завершения культивирования активность устойчивой к тартрату кислой фосфатазы (TRAP) образованных остеокластов измеряли следующим способом. Культуральный раствор из каждой лунки 96-луночного планшета удаляли отсасыванием, и 50 мкл буфера 50 мМ цитрата натрия (pH 6,1), содержащего 1% Тритон X-100, добавляли в каждую лунку. Затем планшет встряхивали в течение 5 минут в встряхивателе для планшетов для лизиса клеток. В каждую лунку добавляли 50 мкл раствора субстрата (буфер 50 мМ цитрат натрия (pH 6,1), содержащий 5 мг/мл п-нитрофенилфосфата и 0,46% тартрат натрия), и планшет инкубировали при комнатной температуре в течение 10 минут. После инкубации 50 мкл раствора 1 н гидроксида натрия добавляли в каждую лунку 96-луночного планшета для остановки ферментативной реакции. После остановки ферментативной реакции в каждой лунке измеряли поглощение при 405 нм, и полученное измерение использовали в качестве показателя активности TRAP. Результаты показаны на Фиг. 44 и 45. Кроме того, для моноклонального антитела крысы против Siglec-15 мыши (антитело #32A1), полученного в примере 6, химерного антитела человека из антитела #32A1, полученного в примере 19, и 4 типов гуманизированных антител #32A1 (h#32A1-H1-1/L5, h#32A1-H5/L5, h#32A1-H1-1/L2-16 и h#32A1-H1-1/L2-15), полученных в примере 28, биологическую активность оценивали таким же способом, как в этом тесте, и результаты показаны на фиг. 46 и 47. Во всех случаях 10 типов гуманизированных антител #32A1 (h#32A1-T1, h#32A1-T2, h#32A1-T3, h#32A1-T4, h#32A1-T5, h#32A1-T6, h#32A1-H1-1/L5, h#32A1-H5/L5, h#32A1-H1-1/L2-16, и h#32A1-H1-1/L2-15), для которых оценивали активность, наблюдали сильную активность ингибирования образования остеокластов, по существу, сравнимую с активностью моноклонального антитела крысы против Siglec-15 мыши (антитело #32A1) или химерного антитела человека из антитела #32A1.
Пример 30
Эффект добавления гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1 на активность резорбции кости нормальных клеток-предшественников остеокластов человека (Оценка с использованием покрытого коллагеном планшета)
Нормальные клетки-предшественники остеокластов человека (Normal Human Natural Osteoclast Precursor Cells, закупленные из Sanko Junyaku Co., Ltd., Кат. No. 2T-110) рассевали в 96- луночный планшет для культивирования OsteoLyse (закупленный из Sanko Junyaku Co., Ltd., Кат. No. PA-1500) при 1×104 клеток/лунку согласно протоколу, сопровождающему клетки. В качестве среды использовали минимальную поддерживающую среду для клеток-предшественников остеокластов (OPBM, закупленную из Sanko Junyaku Co., Ltd., Кат. No. PT-8201), дополненную набором добавок OPGM (закупленным из Sanko Junyaku Co., Ltd., Кат. No. PT-9501), содержащую эмбриональную бычью сыворотку (конечная концентрация: 10%), RANKL человека (конечная концентрация: 63,8 нг/мл), M-CSF человека (конечная концентрация: 33 нг/мл) и тому подобное. К полученному культуральному супернатанту добавляли каждое из моноклонального антитела крысы против Siglec-15 мыши (антитело #32A1), полученное в примере 6, химерное антитело человека из антитела #32A1, полученного в примере 19, и 10 типов гуманизированных антител #32A1 (h#32A1-T1, h#32A1-T2, h#32A1-T3, h#32A1-T4, h#32A1-T5, h#32A1-T6, h#32A1-H1-1/L5, h#32A1-H5/L5, h#32A1-H1-1/L2-16 и h#32A1-H1-1/L2-15), полученные в примерах 24 и 28, в конечной концентрации 4, 20, 100 или 500 нг/мл, и клетки культивировали в течение 5 суток в CO2 инкубаторе. Собирали аликвоту 10 мкл культурального супернатанта, к ней добавляли 200 мкл высвобождающего флуорофор реагента, включенного в набор для анализа OsteoLyse, и с использованием флуоресцентного считывателя для планшетов (ARVO MX, изготовленного в Perkin Elmer Inc.) проводили флуориметрию с временным разрешением (флуориметр для флуориметрии с временным разрешением) (Возбуждение: 340 нм, Излучение: 615 нм), подходящую для измерения флуоресценции с использованием европия, посредством чего определяли количество свободных флуоресцентных фрагментов коллагена, высвобожденных в культуральный супернатант (Фиг. 48 и 49). В результате, количество флуоресцентных фрагментов коллагена, увеличенное при добавлении RANKL, являлось уменьшенным посредством антитела крысы #32A1 или химерного антитела человека #32A1 зависимым от концентрации образом. Таким же способом, также посредством всех из исследованных 10 типов гуманизированных антител #32A1, наблюдали зависимое от концентрации ингибирование. По этим результатам выявили, что активность резорбции кости остеокластов человека являлась ингибированной посредством гуманизированных антител #32A1, специфически связывающих белок Siglec-15.
Пример 31
Биологическая оценка гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1 с использованием крыс после овариэктомии
Ингибирующий эффект на уменьшение минеральной плотности костной ткани у крыс после овариэктомии, наблюдаемый для гуманизированных антител, полученных в примерах 24 и 28, можно оценивать способом, описанным в примере 15.
Пример 32
Оценка биологической активности гуманизированного антитела из моноклонального антитела крысы против Siglec-15 мыши #32A1 на основании теста образования остеокластов мыши при стимуляции TNFα
Ингибирующий эффект на образование остеокластов мыши при стимуляции TNFα, показанный для гуманизированных антител, полученных в примерах 24 и 28, можно оценивать способом, описанным в примере 26.
Пример 33
Определение термостабильности гуманизированного антитела с использованием дифференциальной сканирующей калориметрии (DSC)
Для h#32A1-H5/L5, h#32A1-H1-1/L5, h#32A1-H1-1/L2-15 и h#32A1-H1-1/L2-16 определяли термостабильность. Каждый из этих образцов получали в концентрации 0,5 мг/мл (в растворе CBS), и аликвоты по 400 мкл из них использовали в качестве раствора образца для измерения DSC. Условия для измерения DSC устанавливали следующим образом. А именно, начальную температуру устанавливали на 20°C, конечную температуру устанавливали на 100°C, скорость увеличения температуры устанавливали на 60°C/час, и время на фильтре устанавливали на 10 секунд. В качестве сравнительного раствора использовали CBS. В качестве аппарата для измерения DSC использовали платформу VP-Capillary DSC, изготовленную в MicroCal Inc. USA. Фон (кривую сканирования, полученную при заполнении сравнительным раствором также ячейки для образца), вычитали из кривой сканирования, полученной для каждого раствора образца, посредством чего проводили поправку на фон. На фиг. 50 показана термограмма для h#32A1-H5/L5, На фиг. 51 показана термограмма для h#32A1-H1-1/L5, на фиг. 52 показана термограмма для h#32A1-H1-1/L2-15, и на фиг. 53 показана термограмма для h#32A1-H1-1/L2-16. Когда величину вершины пика в каждой термограмме принимали в качестве средней точки температуры термической денатурации (Tm) в области Fab, величина Tm h#32A1-H5/L5 составляла 81,2°C, величина Tm h#32A1-H1-1/L5 составляла 84,1°C, величина Tm h#32A1-H1-1/L2-15 составляла 80,0°C, и величина Tm h#32A1-1-1/L2-16 (IgG2) составляла 77,9°C.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
Химерное или гуманизированное антитело против Siglec-15 по изобретению обладает способностью ингибировать дифференцировку остеокластов или активность резорбции кости, и фармацевтическая композиция, содержащая антитело против Siglec-15, может быть терапевтическим или профилактическим средством против заболевания с патологическим метаболизмом костной ткани
Инвентарный номер
FERM BP-10999
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-В7-Н3-АНТИТЕЛО | 2012 |
|
RU2668170C2 |
| НОВОЕ АНТИТЕЛО ПРОТИВ DR5 | 2011 |
|
RU2590711C2 |
| НОВОЕ АНТИТЕЛО ПРОТИВ DR5 | 2011 |
|
RU2644678C1 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ AXL | 2010 |
|
RU2571224C2 |
| АНТИТЕЛО, НАПРАВЛЕННОЕ НА БЕЛОК SIGLEC-15, СВЯЗАННЫЙ С ОСТЕОКЛАСТАМИ | 2008 |
|
RU2475499C2 |
| АНТИТЕЛО К EPHA2 | 2008 |
|
RU2525133C2 |
| АНТИТЕЛО ПРОТИВ GPR20 И КОНЪЮГАТЫ АНТИТЕЛО ПРОТИВ GPR20-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2018 |
|
RU2772804C2 |
| АНТИТЕЛО ПРОТИВ GARP | 2016 |
|
RU2769379C2 |
| АНТИ-СD3-АНТИТЕЛО И МОЛЕКУЛА, СОДЕРЖАЩАЯ ЭТО АНТИТЕЛО | 2017 |
|
RU2790326C2 |
| КОНЪЮГАТ АНТИ-TROP2 АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2014 |
|
RU2705367C2 |
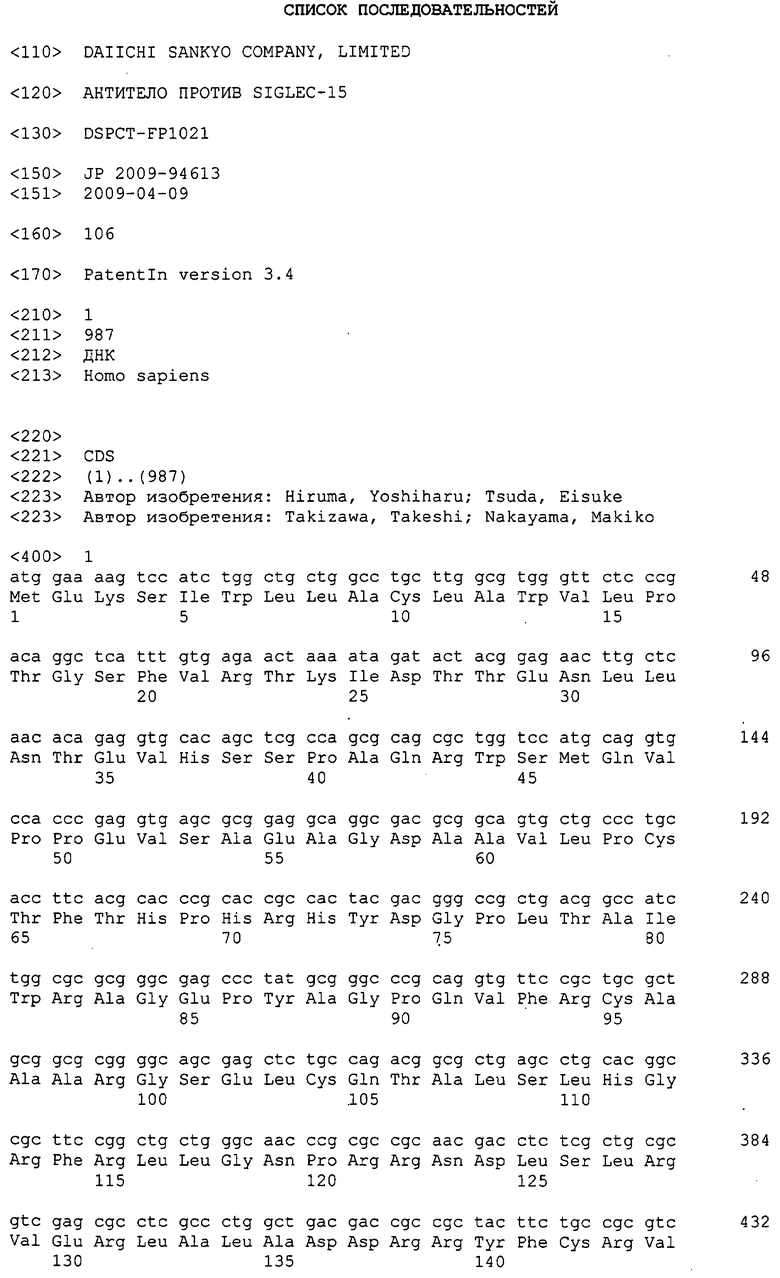








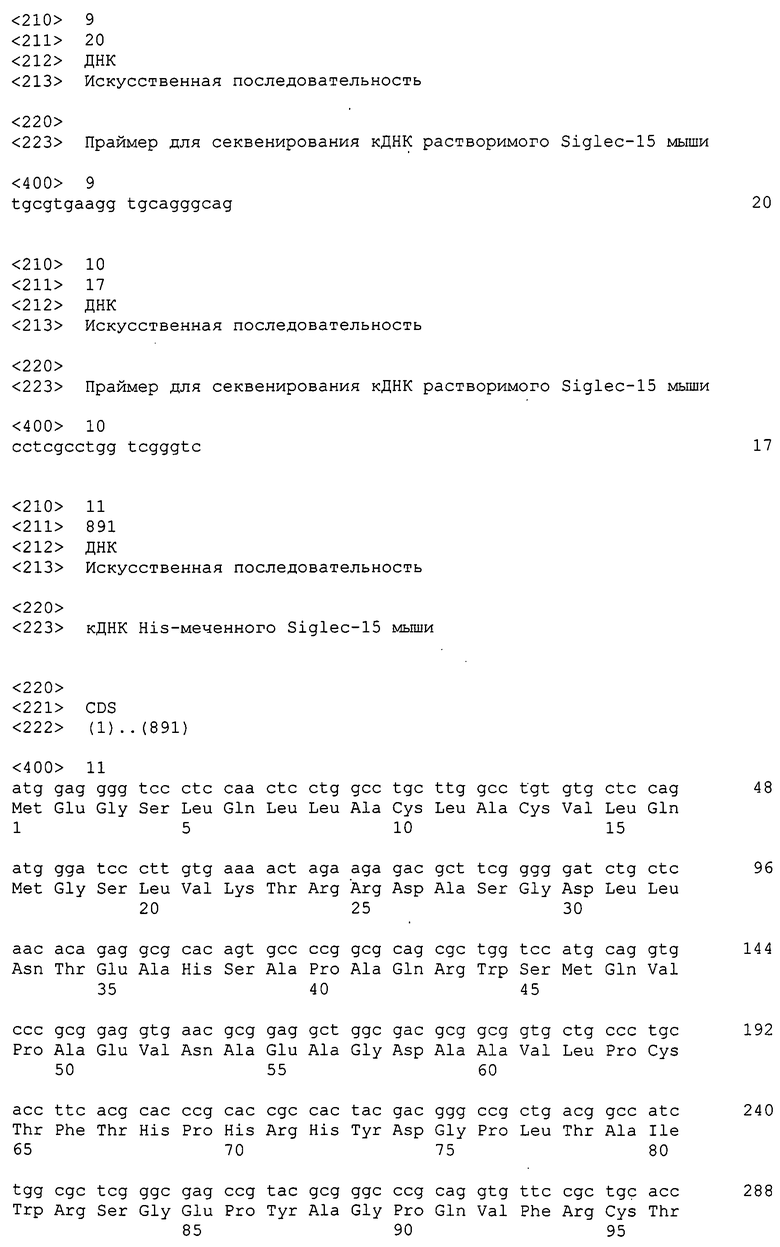































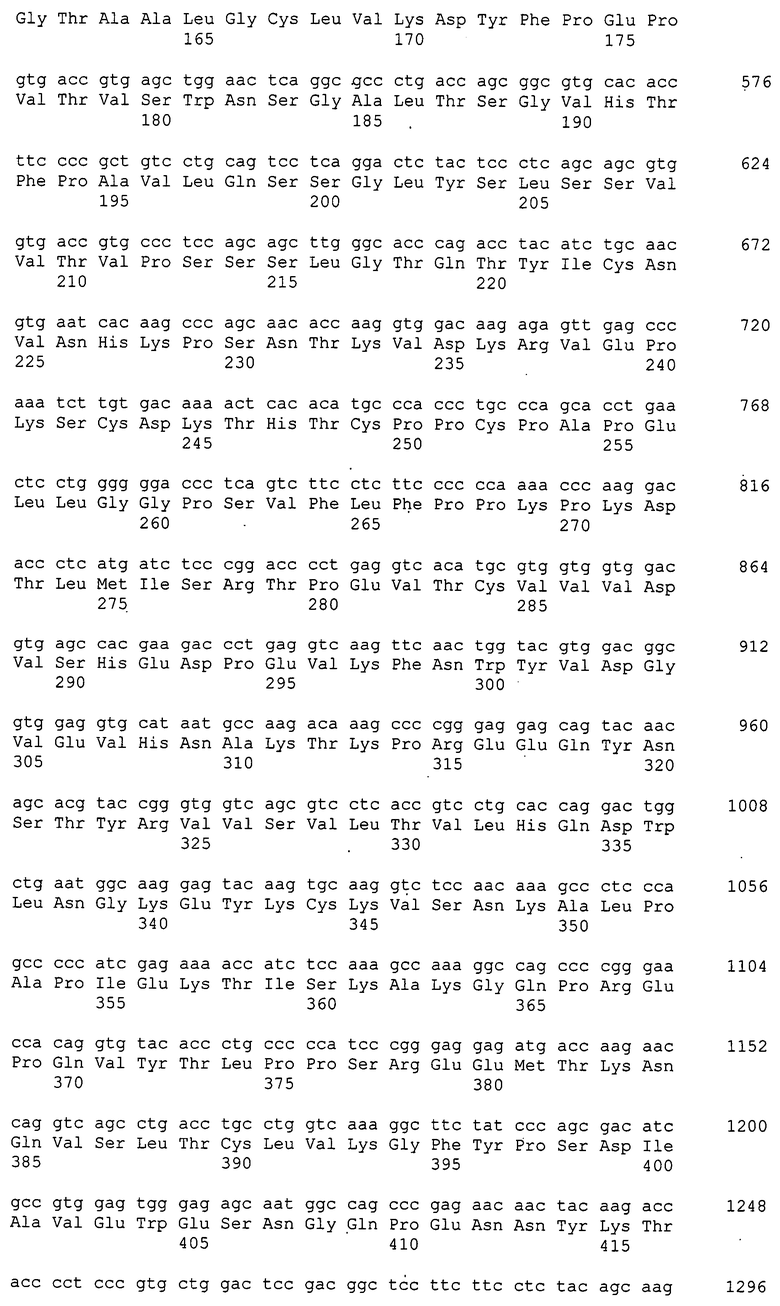

































































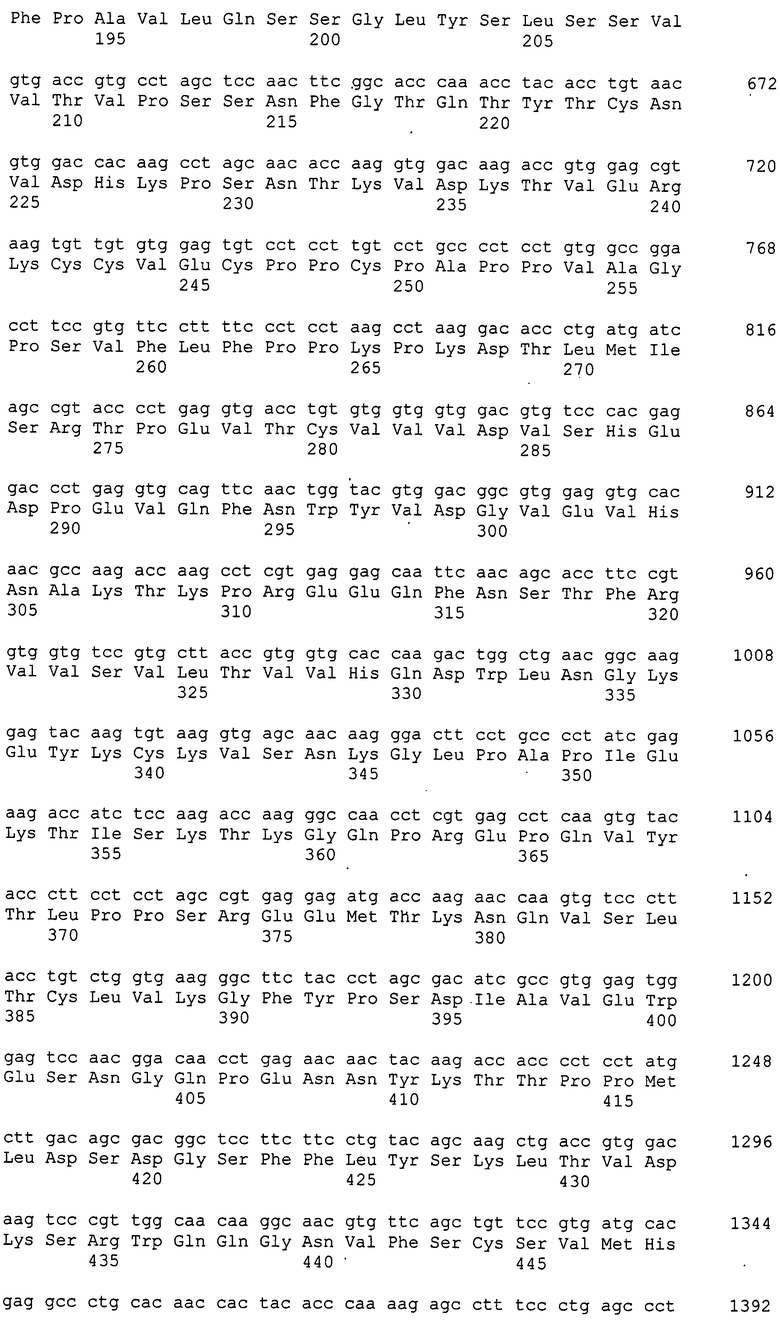












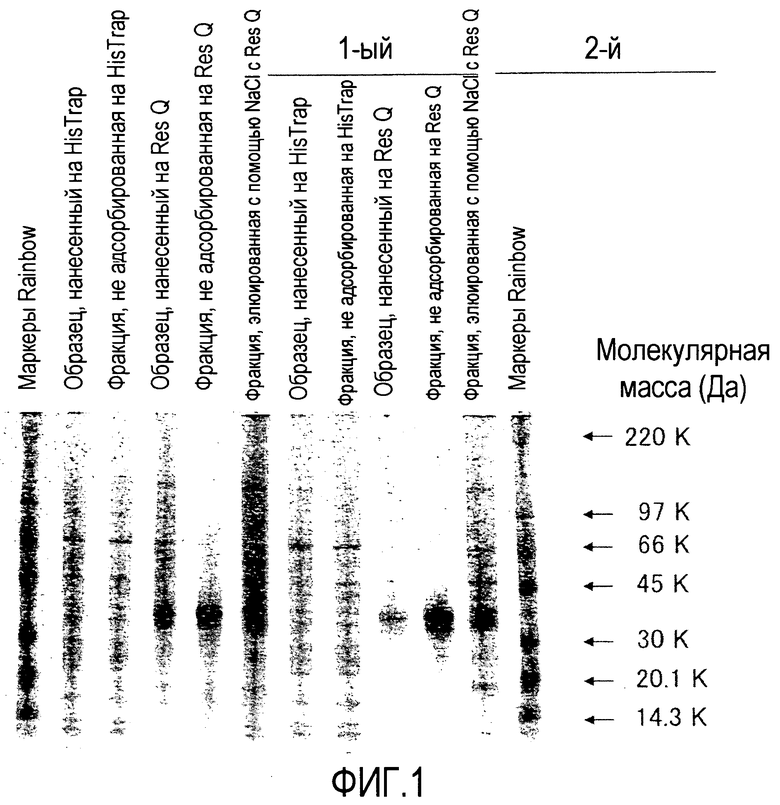
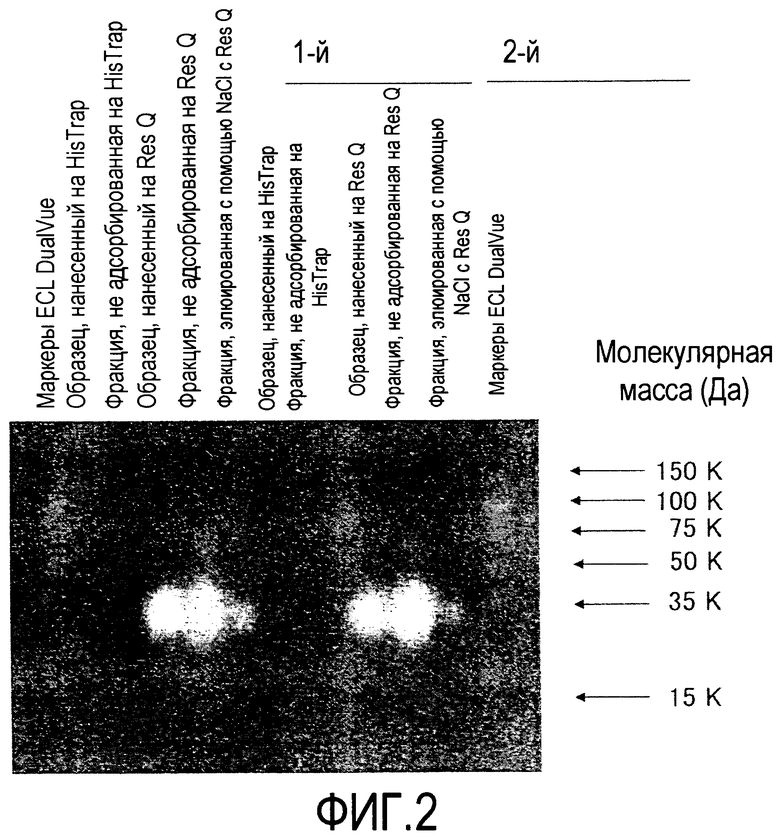
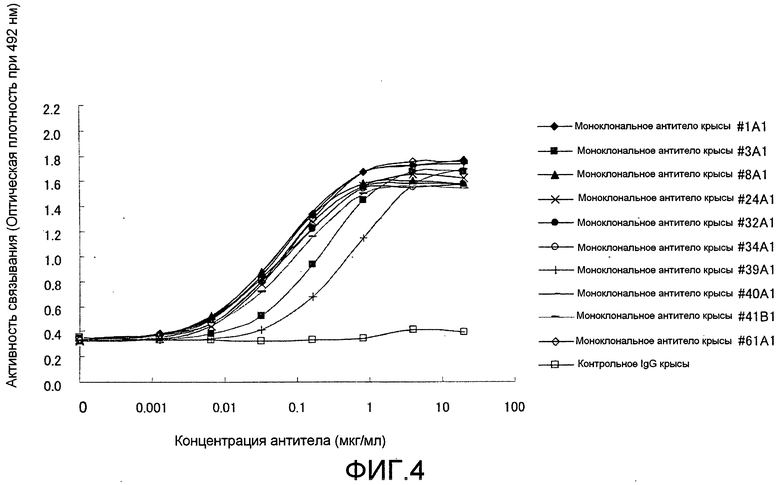
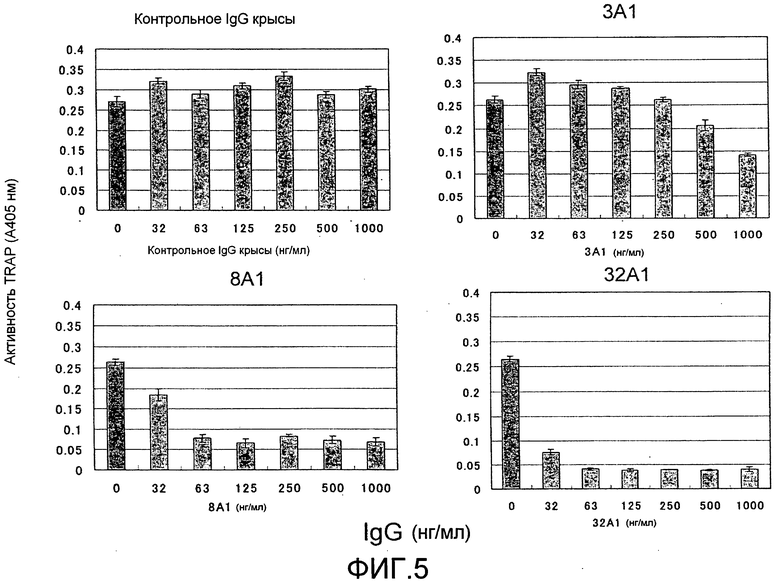
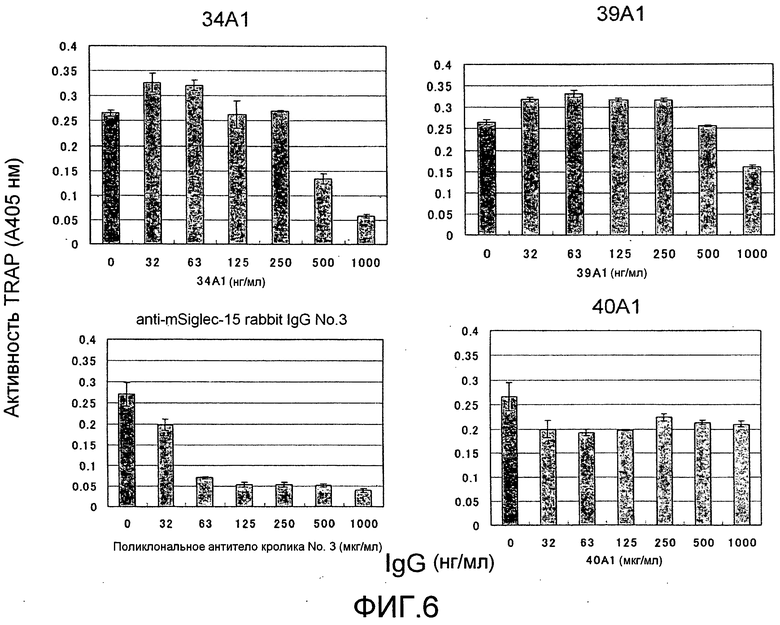
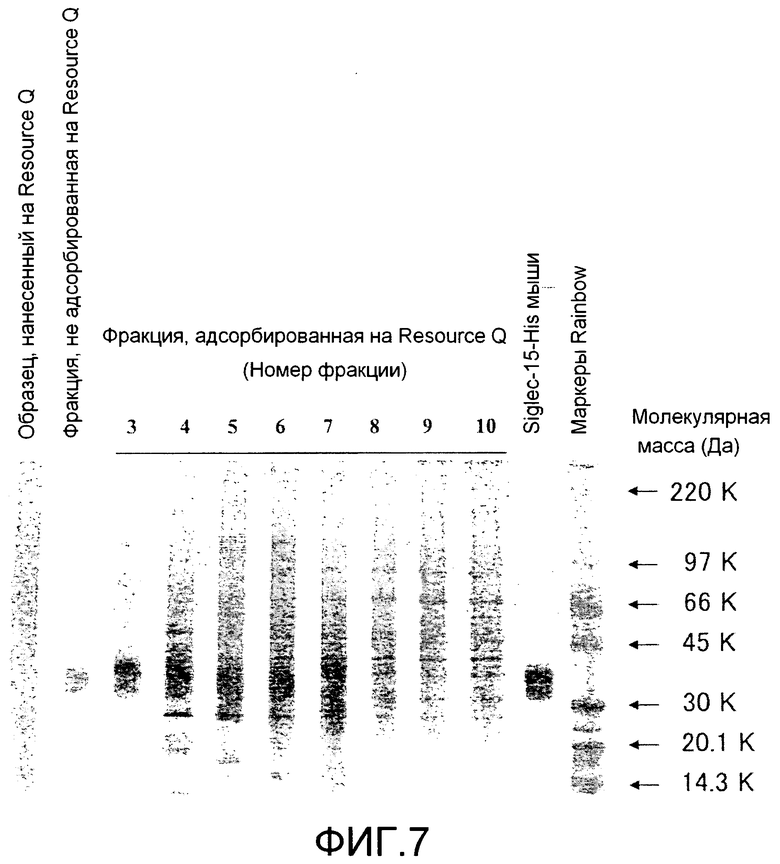
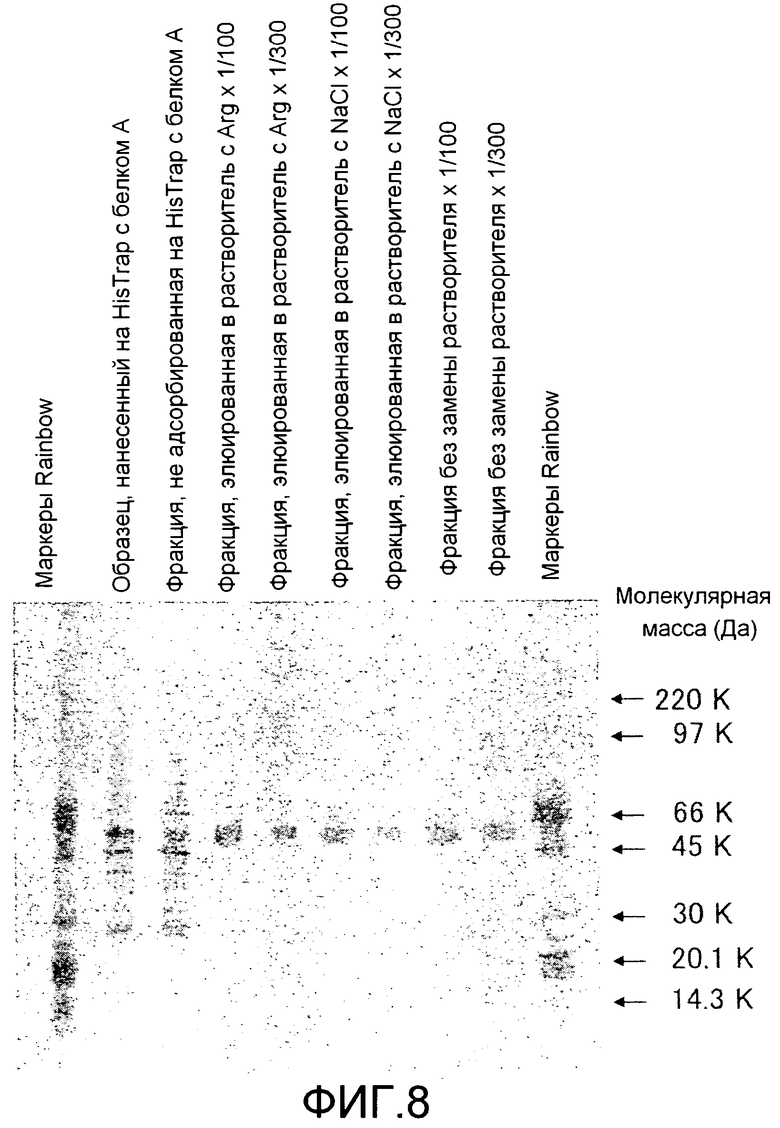
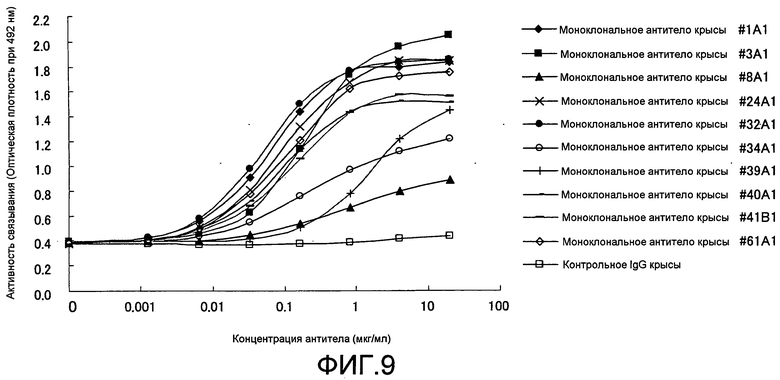
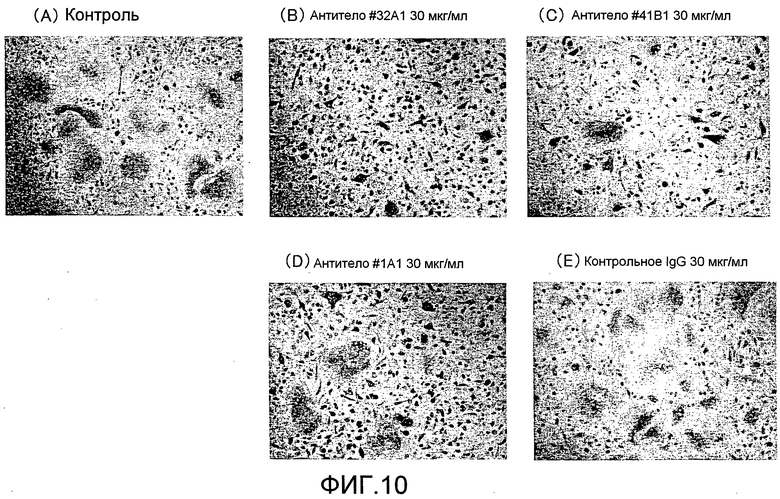
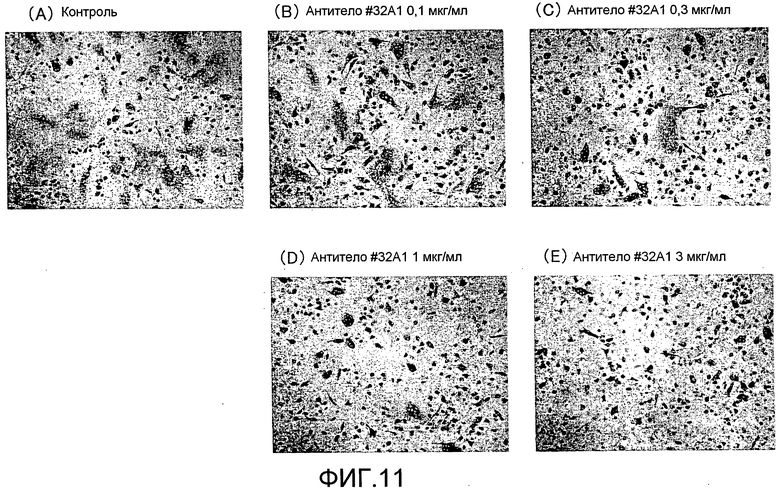

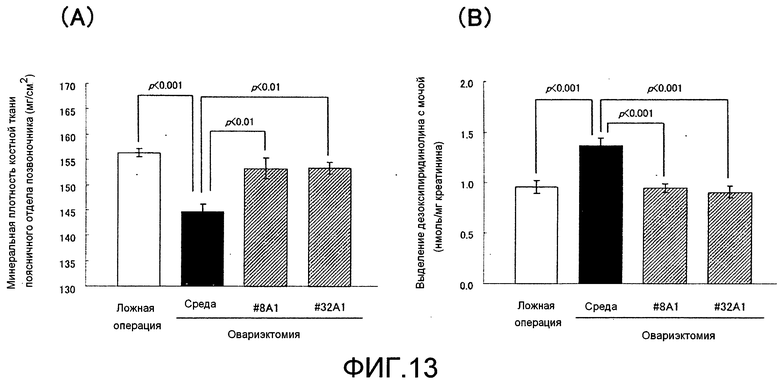
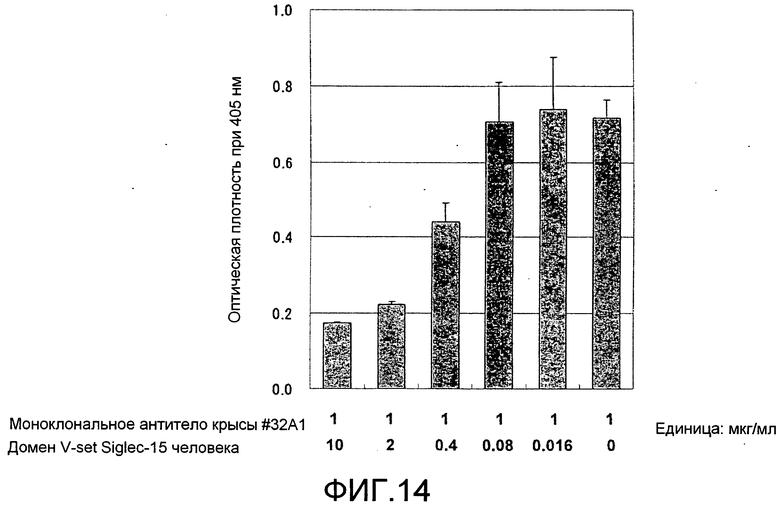
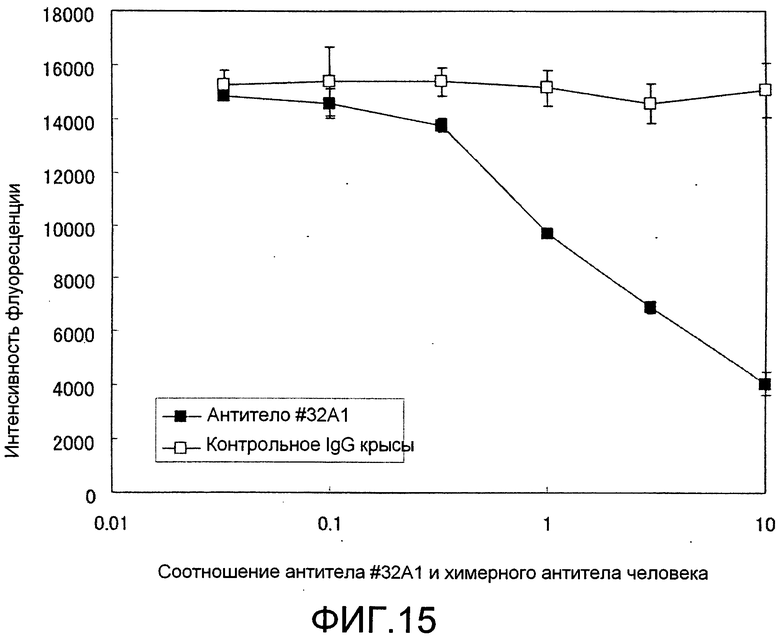
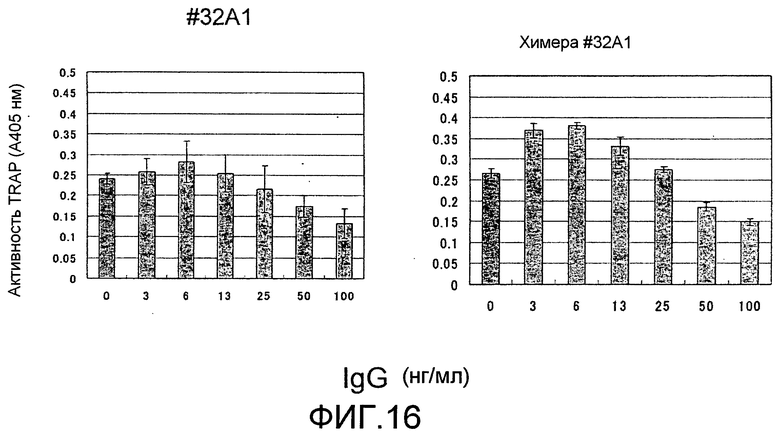
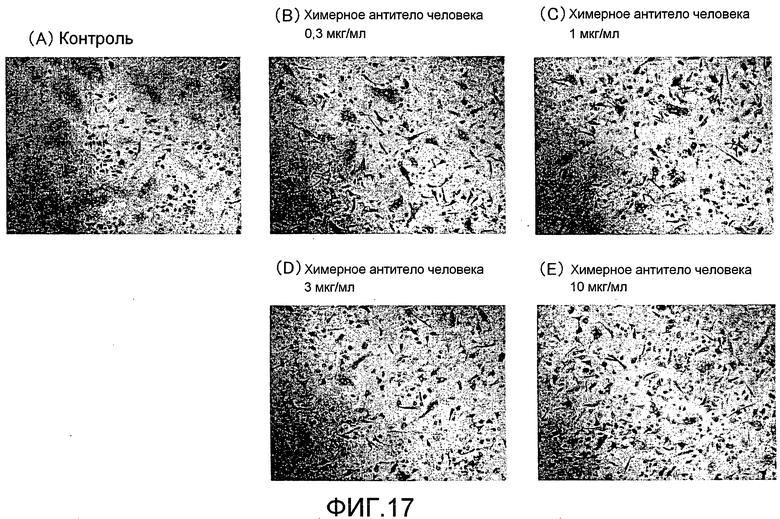
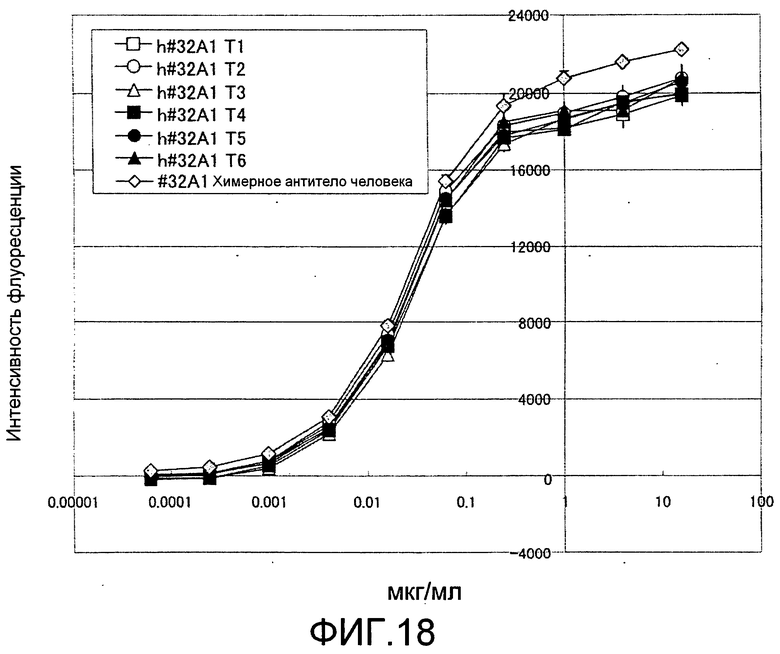
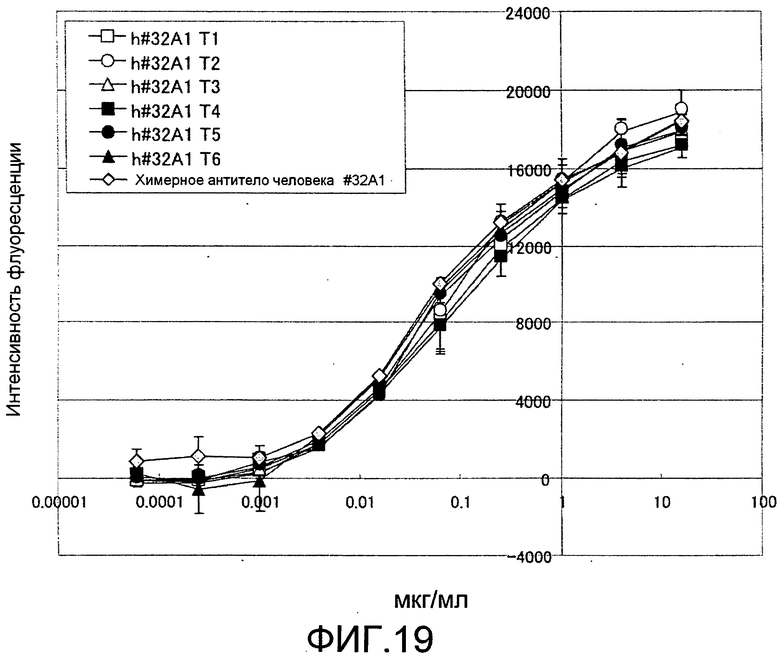

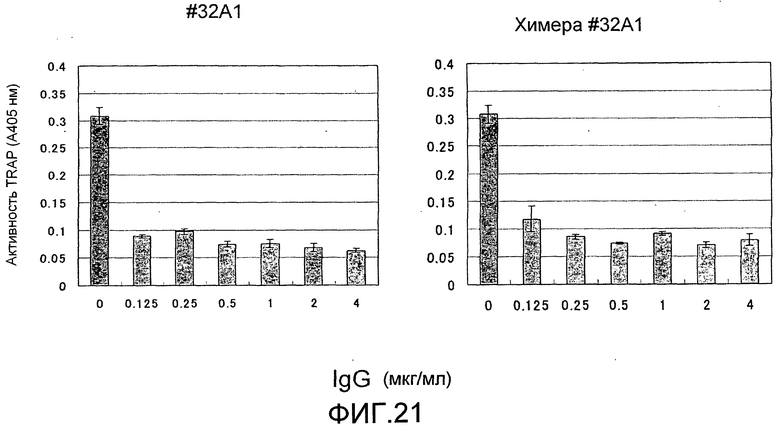
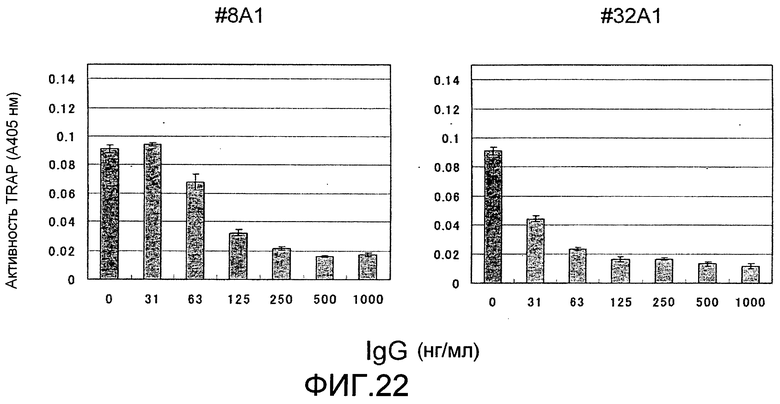
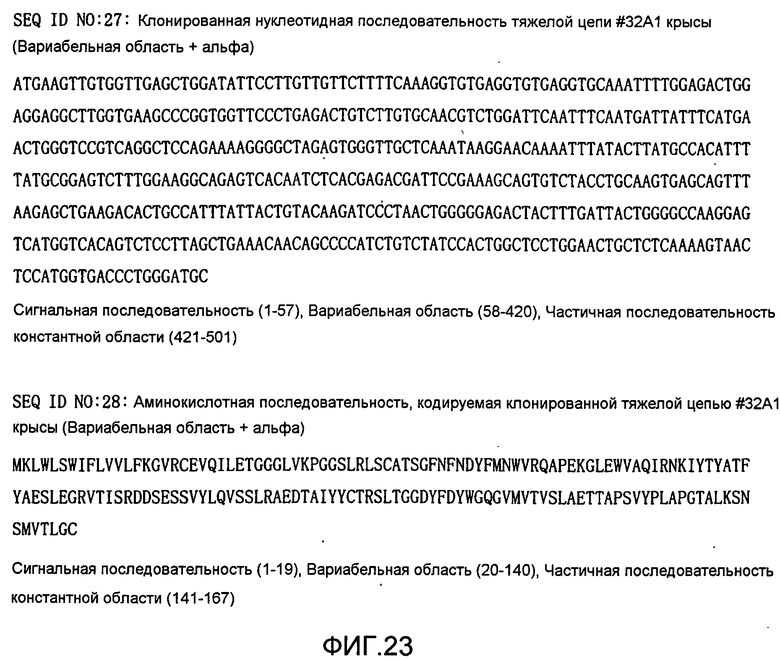









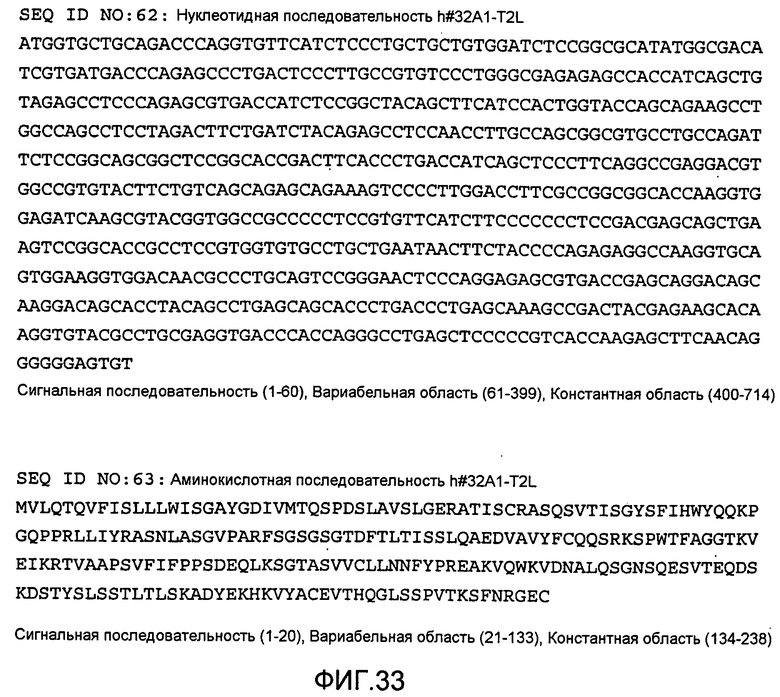



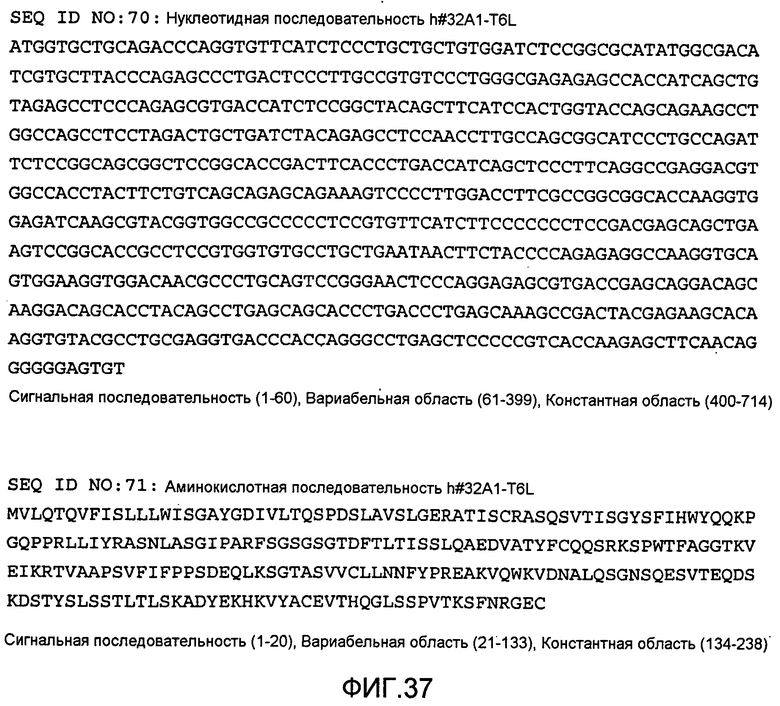






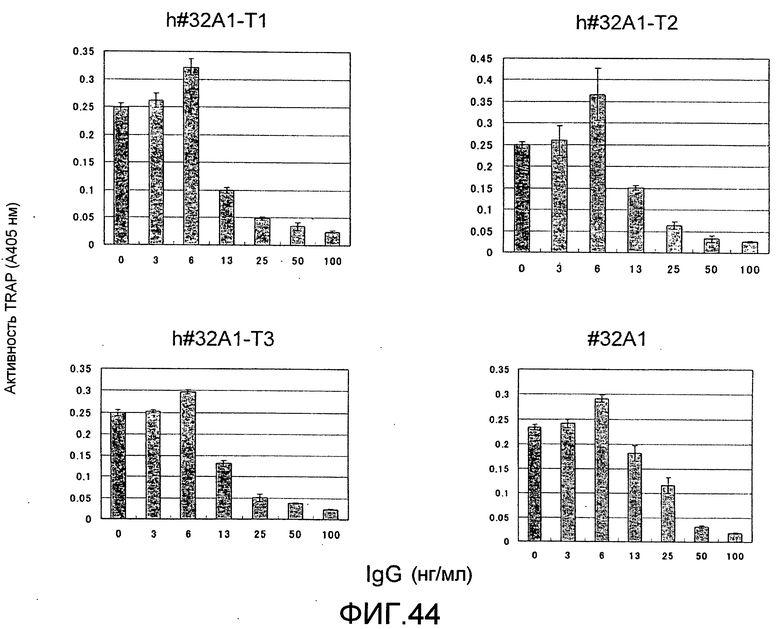
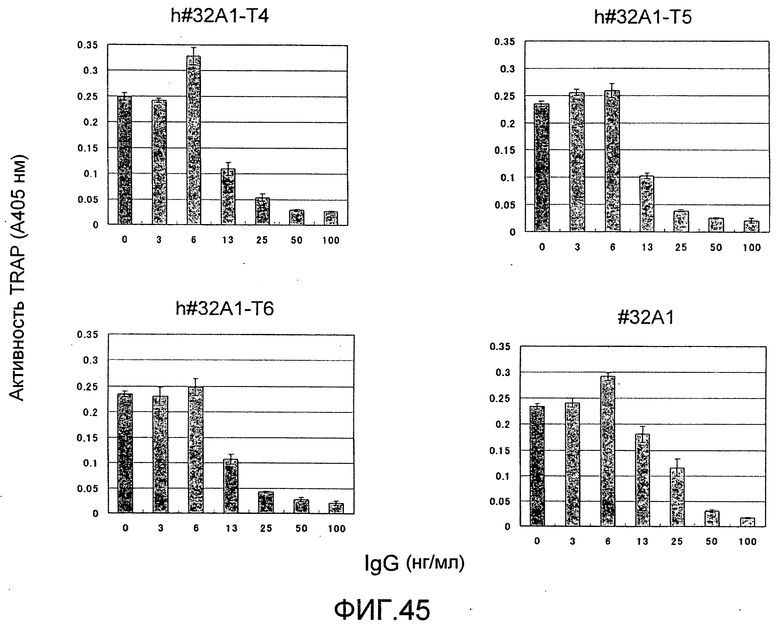
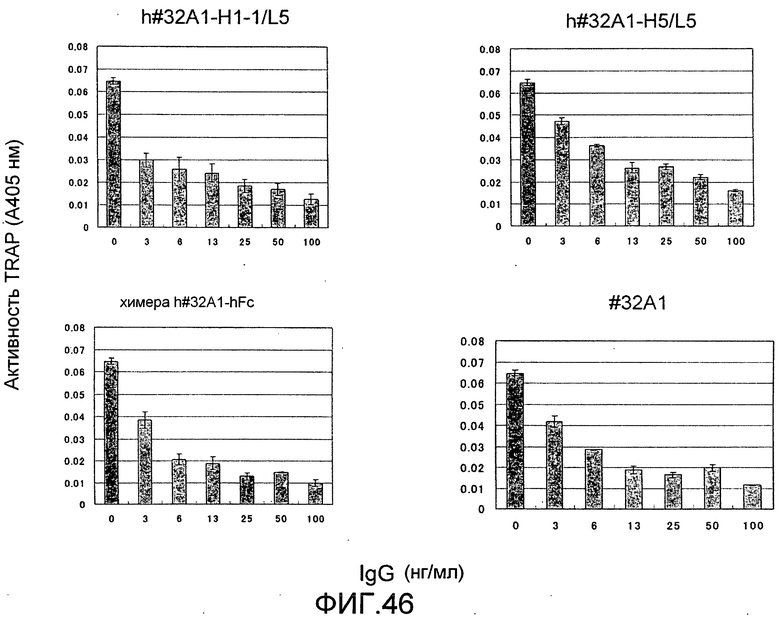
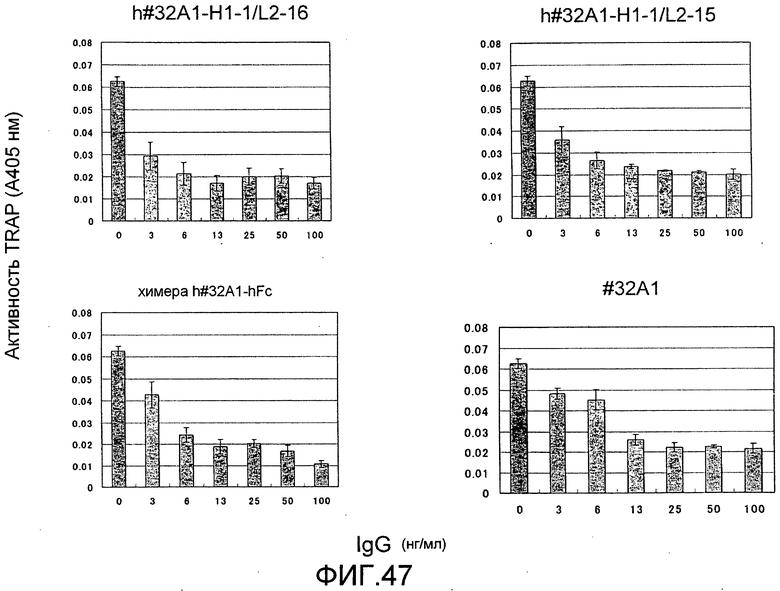
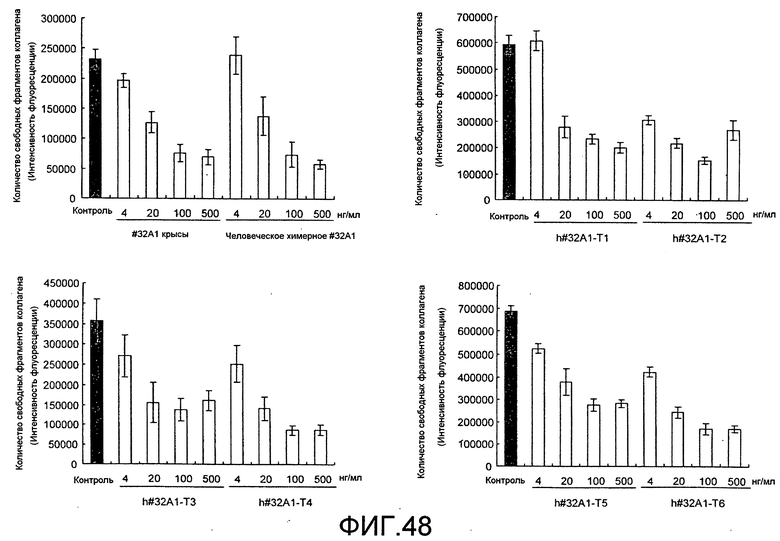
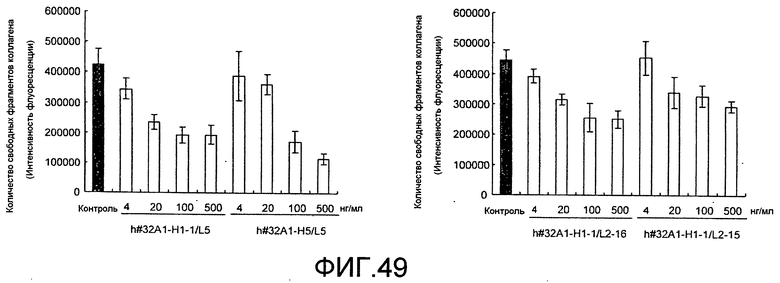
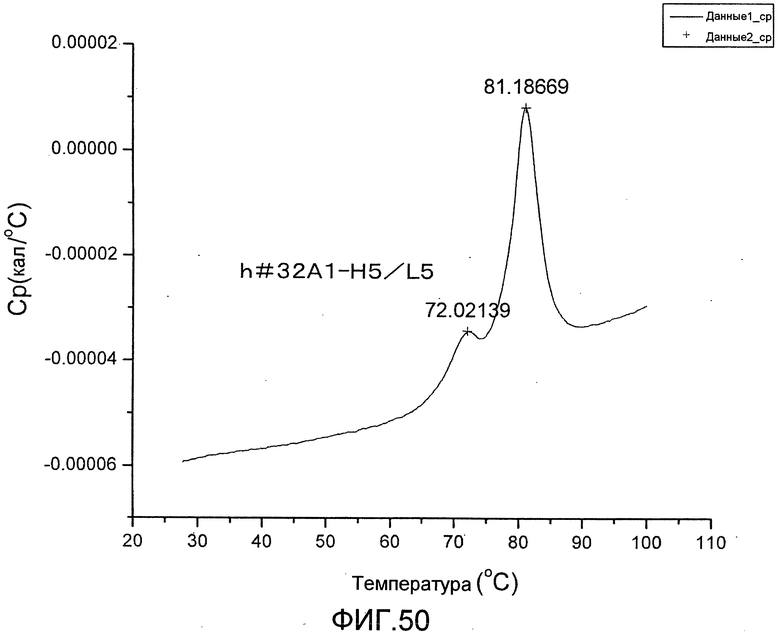
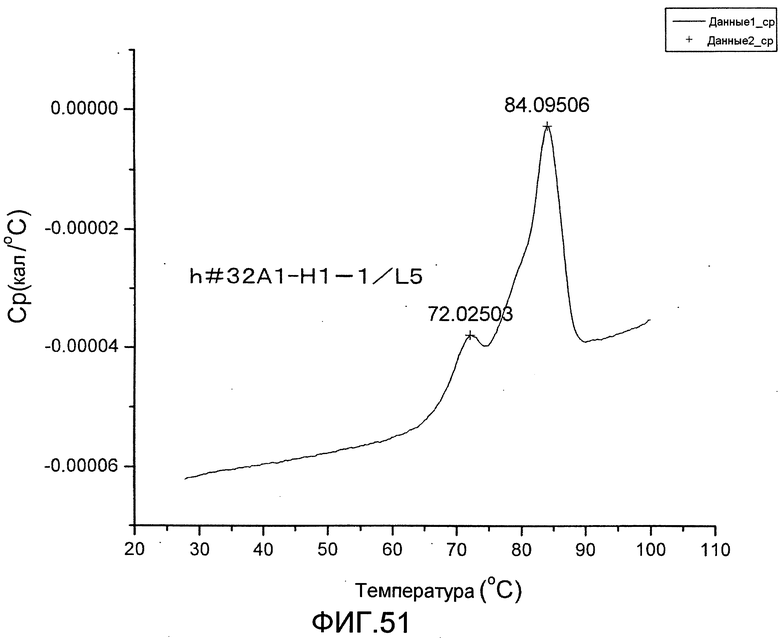
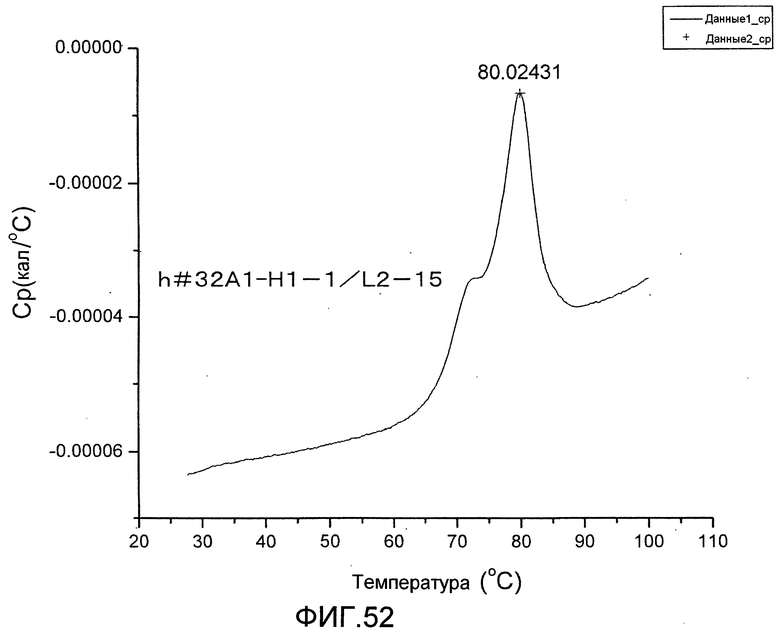
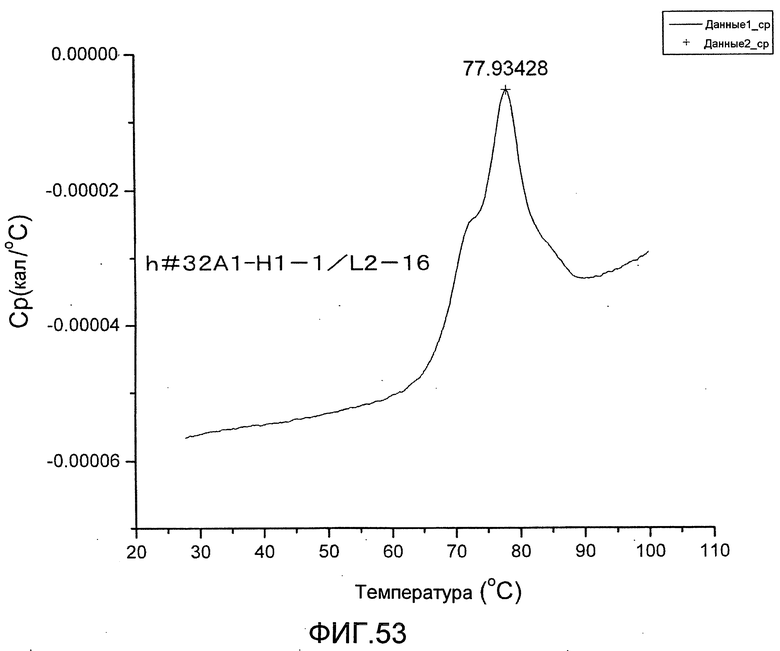




Изобретение относится к области биотехнологии и иммунологии. Описано антитело и его функциональный фрагмент, специфически узнающее Siglec-15 человека и обладающее активностью ингибирования образования остеокластов. Также описана фармацевтическая композиция для лечения и/или профилактики патологического метаболизма костной ткани, которая содержит такое антитело. Изобретение может быть использовано в медицине. 14 н. и 59 з.п. ф-лы, 57 ил., 4 табл., 33 пр.
1. Антитело или функциональный фрагмент антитела, связывающие полипептид, состоящий из аминокислотных остатков 39-165 аминокислотной последовательности SEQ ID NO:2, и ингибирующие образование остеокластов и, необязательно, остеокластную резорбцию кости, где антитело ингибирует in vitro образование остеокластов при концентрации, составляющей 1 мкг/мл или менее, и где антитело содержит:
последовательность тяжелой цепи, содержащую вариабельную область с CDRH1, CDRH2 и CDRH3, где CDRH1 содержит аминокислотную последовательность SEQ ID NO:44, CDRH2 содержит любую из аминокислотных последовательностей SEQ ID NO:45 или SEQ ID NO:97 и CDRH3 содержит аминокислотную последовательность SEQ ID NO:46; и
последовательность легкой цепи, содержащую вариабельную область с CDRL1, CDRL2 и CDRL3, где CDRL1 содержит аминокислотную последовательность SEQ ID NO:47, CDRL2 содержит аминокислотную последовательность SEQ ID NO:48 и CDRL3 содержит аминокислотную последовательность SEQ ID NO:49.
2. Антитело или функциональный фрагмент антитела по п.1, отличающиеся тем, что содержат последовательность вариабельной области тяжелой цепи, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:41, и последовательность вариабельной области легкой цепи, содержащую аминокислотные остатки 21-132 аминокислотной последовательности SEQ ID NO:43.
3. Функциональный фрагмент антитела по п.1, выбранный из группы, состоящей из Fab, F(ab′)2, Fab′ и Fv.
4. Антитело по п.1, отличающееся тем, что представляет собой scFv.
5. Антитело или функциональный фрагмент антитела по п.1, отличающиеся тем, что антитело представляет собой химерное антитело.
6. Антитело или функциональный фрагмент антитела по п.5, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-470 аминокислотной последовательности SEQ ID NO:41, и последовательность легкой цепи, содержащую аминокислотные остатки 21-237 аминокислотной последовательности SEQ ID NO:43.
7. Антитело или функциональный фрагмент антитела по п.1, отличающиеся тем, что антитело является гуманизированным.
8. Антитело по п.6, где тяжелая цепь обладает константной областью тяжелой цепи иммуноглобулина G2 человека и легкая цепь обладает константной областью легкой цепи κ иммуноглобулина человека.
9. Антитело или функциональный фрагмент антитела по п.1, где антитело содержит:
(а) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих аминокислотных последовательностей:
a1) аминокислотной последовательности, содержащей аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:51;
а2) аминокислотной последовательности, содержащей аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:53;
а3) аминокислотной последовательности, содержащей аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:55;
а4) аминокислотной последовательности, содержащей аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:57;
а5) аминокислотной последовательности, содержащей аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:59;
а6) аминокислотной последовательности, содержащей аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:99;
а7) аминокислотной последовательности, содержащей аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:101;
а8) аминокислотной последовательности, которая по меньшей мере на 95% гомологична любой аминокислотной последовательности, выбранной из a1)-а7);
а9) аминокислотной последовательности, которая по меньшей мере на 99% гомологична любой аминокислотной последовательности, выбранной из a1)-a7); и
а10) аминокислотной последовательности, включающей замену, делецию или добавление от одного до нескольких аминокислотных остатков в любой аминокислотной последовательности, выбранной из a1)-а7); и
(b) вариабельную область легкой цепи, выбранную из группы, состоящей из следующих аминокислотных последовательностей:
b1) аминокислотной последовательности, содержащей аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:61;
b2) аминокислотной последовательности, содержащей аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:63;
b3) аминокислотной последовательности, содержащей аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:65;
b4) аминокислотной последовательности, содержащей аминокислотные остатки 21-132 аминокислотной последовательности SEQ ID NO:67;
b5) аминокислотной последовательности, содержащей аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:69;
b6) аминокислотной последовательности, содержащей аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:71;
b7) аминокислотной последовательности, содержащей аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:103;
b8) аминокислотной последовательности, содержащей аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:105;
b9) аминокислотной последовательности, которая по меньшей мере на 95% гомологична любой аминокислотной последовательности, выбранной из b1)-b8);
b10) аминокислотной последовательности, которая по меньшей мере на 99% гомологична любой аминокислотной последовательности, выбранной из b1)-b8); и
b11) аминокислотной последовательности, включающей замену, делецию или добавление от одного до нескольких аминокислотных остатков в любой аминокислотной последовательности, выбранной из b1)-b8).
10. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:51, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:61.
11. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:53, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:63.
12. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:55, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:65.
13. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:55, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 21-132 аминокислотной последовательности SEQ ID NO:67.
14. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:57, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:69.
15. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:59, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:71.
16. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:69.
17. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:101, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:69.
18. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:103.
19. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 20-140 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую вариабельную область легкой цепи, содержащую аминокислотную последовательность, содержащую аминокислотные остатки 21-133 аминокислотной последовательности SEQ ID NO:105.
20. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-470 аминокислотной последовательности SEQ ID NO:51, и последовательность легкой цепи, содержащую аминокислотные остатки 21-238 аминокислотной последовательности SEQ ID NO:61.
21. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-470 аминокислотной последовательности SEQ ID NO:53, и последовательность легкой цепи, содержащую аминокислотные остатки 21-238 аминокислотной последовательности SEQ ID NO:63.
22. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-470 аминокислотной последовательности SEQ ID NO:55, и последовательность легкой цепи, содержащую аминокислотные остатки 21-238 аминокислотной последовательности SEQ ID NO:65.
23. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-470 аминокислотной последовательности SEQ ID NO:55, и последовательность легкой цепи, содержащую аминокислотные остатки 21-237 аминокислотной последовательности SEQ ID NO:67.
24. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-470 аминокислотной последовательности SEQ ID NO:57, и последовательность легкой цепи, содержащую аминокислотные остатки 21-238 аминокислотной последовательности SEQ ID NO:69.
25. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-470 аминокислотной последовательности SEQ ID NO:59, и последовательность легкой цепи, содержащую аминокислотные остатки 21-238 аминокислотной последовательности SEQ ID NO:71.
26. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-466 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую аминокислотные остатки 21-238 аминокислотной последовательности SEQ ID NO:69.
27. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-466 аминокислотной последовательности SEQ ID NO:101, и последовательность легкой цепи, содержащую аминокислотные остатки 21-238 аминокислотной последовательности SEQ ID NO:69.
28. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-466 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую аминокислотные остатки 21-238 аминокислотной последовательности SEQ ID NO:103.
29. Антитело или функциональный фрагмент антитела по п.9, отличающиеся тем, что содержат последовательность тяжелой цепи, содержащую аминокислотные остатки 20-466 аминокислотной последовательности SEQ ID NO:99, и последовательность легкой цепи, содержащую аминокислотные остатки 21-238 аминокислотной последовательности SEQ ID NO:105.
30. Фармацевтическая композиция для лечения и/или профилактики патологического метаболизма костной ткани, отличающаяся тем, что содержит по меньшей мере одно из антител или функциональных фрагментов антител по пп.1-29 в дозе от 0,1 до 100 мг/кг от одного раза в сутки до одного раза в 180 суток и по меньшей мере один представитель, выбранный из группы, состоящей из бисфосфонатов, активного витамина D3, кальцитонина и его производных, гормонов, таких как эстрадиол, SERM (избирательных модуляторов рецепторов эстрогенов), иприфлавона, витамина K2 (менатетренона), препаратов кальция, РТН (паратиреоидного гормона), нестероидных противовоспалительных средств, растворимых рецепторов TNF, антител против TNF-α или функциональных фрагментов антител, антител против PTHrP (родственного паратиреоидному гормону белка) или функциональных фрагментов антител, антагонистов рецептора IL-1, антител против рецептора IL-6 или функциональных фрагментов антител, антител против RANKL или функциональных фрагментов антител и OCIF (фактора ингибирования остеокластогенеза).
31. Фармацевтическая композиция по п.30, где патологический метаболизм костной ткани выбран из группы, состоящей из остеопороза, разрушения кости, сопровождающего ревматоидный артрит, злокачественной гиперкальциемии, разрушения кости, сопровождающего множественную миелому или метастазирование злокачественной опухоли в кость, гигантоклеточной опухоли, остеопении, потери зубов из-за периодонтита, остеолиза около протеза сустава, разрушения кости при хроническом остеомиелите, болезни Педжета кости, нефрогенной остеодистрофии и несовершенного остеогенеза.
32. Фармацевтическая композиция по п.31, отличающаяся тем, что патологический метаболизм костной ткани представляет собой остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость.
33. Фармацевтическая композиция по п.32, отличающаяся тем, что патологический метаболизм костной ткани представляет собой остеопороз.
34. Фармацевтическая композиция по п.33, отличающаяся тем, что остеопороз представляет собой постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит.
35. Фармацевтическая композиция для лечения и/или профилактики патологического метаболизма костной ткани, отличающаяся тем, что содержит по меньшей мере одно из антител или функциональных фрагментов антител по пп.1-29 в дозе от 0,1 до 100 мг/кг от одного раза в сутки до одного раза в 180 суток.
36. Фармацевтическая композиция по п.35, отличающаяся тем, что является терапевтическим и/или профилактическим средством против патологического метаболизма костной ткани.
37. Фармацевтическая композиция по п.36, где патологический метаболизм костной ткани выбран из группы, состоящей из остеопороза, разрушения кости, сопровождающего ревматоидный артрит, злокачественной гиперкальциемии, разрушения кости, сопровождающего множественную миелому или метастазирование злокачественной опухоли в кость, гигантоклеточной опухоли, остеопении, потери зубов из-за периодонтита, остеолиза около протеза сустава, разрушения кости при хроническом остеомиелите, болезни Педжета кости, нефрогенной остеодистрофии и несовершенного остеогенеза.
38. Фармацевтическая композиция по п.37, отличающаяся тем, что патологический метаболизм костной ткани представляет собой остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость.
39. Фармацевтическая композиция по п.38, отличающаяся тем, что патологический метаболизм костной ткани представляет собой остеопороз.
40. Фармацевтическая композиция по п.39, отличающаяся тем, что остеопороз представляет собой постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит.
41. Применение антитела по пп.1-29 или его функционального фрагмента для получения лекарственного средства для лечения и/или профилактики патологического метаболизма костной ткани, отличающееся введением одновременно или последовательно антитела или функционального фрагмента и по меньшей мере одного члена, выбранного из группы, состоящей из бисфосфонатов, активного витамина D3, кальцитонина и его производных, гормонов, таких как эстрадиол, SERM (избирательных модуляторов рецепторов эстрогенов), иприфлавона, витамина K2 (менатетренона), препаратов кальция, РТН (паратиреоидного гормона), нестероидных противовоспалительных средств, растворимых рецепторов TNF, антител против TNF-α или функциональных фрагментов антител, антител против PTHrP (родственного паратиреоидному гормону белка) или функциональных фрагментов антител, антагонистов рецептора IL-1, антител против рецептора IL-6 или функциональных фрагментов антител, антител против RANKL или функциональных фрагментов антител и OCIF (фактора ингибирования остеокластогенеза).
42. Применение по п.41, отличающееся тем, что патологический метаболизм костной ткани представляет собой остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость.
43. Применение по п.42, отличающееся тем, что патологический метаболизм костной ткани представляет собой остеопороз.
44. Применение по п.43, отличающееся тем, что остеопороз представляет собой постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит.
45. Применение антитела по пп.1-29 или его функционального фрагмента для получения лекарственного средства для лечения и/или профилактики патологического метаболизма костной ткани.
46. Применение по п.45, отличающееся тем, что патологический метаболизм костной ткани представляет собой остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость.
47. Применение по п.46, отличающееся тем, что патологический метаболизм костной ткани представляет собой остеопороз.
48. Применение по п.47, отличающееся тем, что остеопороз представляет собой постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит.
49. Способ лечения патологического метаболизма костной ткани, отличающийся одновременным или последовательным введением эффективного количества по меньшей мере одного из антител или функциональных фрагментов антител по пп.1-29 и по меньшей мере одного представителя, выбранного из группы, состоящей из бисфосфонатов, активного витамина D3, кальцитонина и его производных, гормонов, таких как эстрадиол, SERM (избирательных модуляторов рецепторов эстрогенов), иприфлавона, витамина K2 (менатетренона), препаратов кальция, РТН (паратиреоидного гормона), нестероидных противовоспалительных средств, растворимых рецепторов TNF, антител против TNF-α или функциональных фрагментов антител, антител против PTHrP (родственного паратиреоидному гормону белка) или функциональных фрагментов антител, антагонистов рецептора IL-1, антител против рецептора IL-6 или функциональных фрагментов антител, антител против RANKL или функциональных фрагментов антител и OCIF (фактора ингибирования остеокластогенеза).
50. Способ лечения по п.49, отличающийся тем, что патологический метаболизм костной ткани представляет собой остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость.
51. Способ лечения по п.50, отличающийся тем, что патологический метаболизм костной ткани представляет собой остеопороз.
52. Способ лечения по п.51, отличающийся тем, что остеопороз представляет собой постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит.
53. Способ профилактики патологического метаболизма костной ткани, отличающийся одновременным или последовательным введением эффективного количества по меньшей мере одного из антител или функциональных фрагментов антител по пп.1-29 и по меньшей мере одного представителя, выбранного из группы, состоящей из бисфосфонатов, активного витамина D3, кальцитонина и его производных, гормонов, таких как эстрадиол, SERM (избирательных модуляторов рецепторов эстрогенов), иприфлавона, витамина K2 (менатетренона), препаратов кальция, РТН (паратиреоидного гормона), нестероидных противовоспалительных средств, растворимых рецепторов TNF, антител против TNF-α или функциональных фрагментов антител, антител против PTHrP (родственного паратиреоидному гормону белка) или функциональных фрагментов антител, антагонистов рецептора IL-1, антител против рецептора IL-6 или функциональных фрагментов антител, антител против RANKL или функциональных фрагментов антител и OCIF (фактора ингибирования остеокластогенеза).
54. Способ профилактики по п.53, отличающийся тем, что патологический метаболизм костной ткани представляет собой остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость.
55. Способ профилактики по п.54, отличающийся тем, что патологический метаболизм костной ткани представляет собой остеопороз.
56. Способ профилактики по п.55, отличающийся тем, что остеопороз представляет собой постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит.
57. Способ лечения патологического метаболизма костной ткани, отличающийся введением эффективного количества по меньшей мере одного из антител или функциональных фрагментов антител по пп.1-29.
58. Способ лечения по п.57, отличающийся тем, что патологический метаболизм костной ткани представляет собой остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость.
59. Способ лечения по п.58, отличающийся тем, что патологический метаболизм костной ткани представляет собой остеопороз.
60. Способ лечения по п.59, отличающийся тем, что остеопороз представляет собой постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит.
61. Способ профилактики патологического метаболизма костной ткани, отличающийся введением эффективного количества по меньшей мере одного из антител или функциональных фрагментов антител по пп.1-29.
62. Способ профилактики по п.61, отличающийся тем, что патологический метаболизм костной ткани представляет собой остеопороз, разрушение кости, сопровождающее ревматоидный артрит, или разрушение кости, сопровождающее метастазирование злокачественной опухоли в кость.
63. Способ профилактики по п.62, отличающийся тем, что патологический метаболизм костной ткани представляет собой остеопороз.
64. Способ профилактики по п.63, отличающийся тем, что остеопороз представляет собой постменопаузальный остеопороз, старческий остеопороз, вторичный остеопороз из-за применения лекарственного средства, такого как стероид или иммунодепрессант, или остеопороз, сопровождающий ревматоидный артрит.
65. Полинуклеотид, кодирующий антитело по любому из пп.2, 6 и 9-29.
66. Полинуклеотид по п.65, отличающийся тем, что содержит нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:40, и нуклеотидную последовательность, содержащую нуклеотиды 61-396 нуклеотидной последовательности SEQ ID NO:42.
67. Полинуклеотид по п.66, отличающийся тем, что содержит нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:40, и нуклеотидную последовательность, содержащую нуклеотиды 61-711 нуклеотидной последовательности SEQ ID NO:42.
68. Полинуклеотид по п.66, отличающийся тем, что содержит:
(а) полинуклеотид, выбранный из группы, состоящей из следующих нуклеотидных последовательностей:
a1) нуклеотидной последовательности, содержащей нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:50;
а2) нуклеотидной последовательности, содержащей нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:52;
а3) нуклеотидной последовательности, содержащей нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:54;
а4) нуклеотидной последовательности, содержащей нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:56;
а5) нуклеотидной последовательности, содержащей нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:58;
а6) нуклеотидной последовательности, содержащей нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:98;
а7) нуклеотидной последовательности, содержащей нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:100;
а8) нуклеотидной последовательности полинуклеотида, который гибридизуется с полинуклеотидом, содержащим нуклеотидную последовательность, комплементарную любой из нуклеотидных последовательностей, выбранных из a1)-a7), в жестких условиях; и
а9) нуклеотидной последовательности, включающей замену, делецию или добавление от одного до нескольких нуклеотидов в любой нуклеотидной последовательности, выбранной из a1)-a7); и
(b) полинуклеотид, выбранный из группы, состоящей из следующих нуклеотидных последовательностей:
b1) нуклеотидной последовательности, содержащей нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:60;
b2) нуклеотидной последовательности, содержащей нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:62;
b3) нуклеотидной последовательности, содержащей нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:64;
b4) нуклеотидной последовательности, содержащей нуклеотиды 61-396 нуклеотидной последовательности SEQ ID NO:66;
b5) нуклеотидной последовательности, содержащей нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:68;
b6) нуклеотидной последовательности, содержащей нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:70;
b7) нуклеотидной последовательности, содержащей нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:102;
b8) нуклеотидной последовательности, содержащей нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:104;
b9) нуклеотидной последовательности полинуклеотида, который гибридизуется с полинуклеотидом, содержащим нуклеотидную последовательность, комплементарную любой нуклеотидной последовательности, выбранной из b1)-b8), в жестких условиях; и
b10) нуклеотидной последовательности, включающей замену, делецию или добавление от одного до нескольких нуклеотидов в любой нуклеотидной последовательности, выбранной из b1)-b8).
69. Полинуклеотид по п.68, отличающийся тем, что содержит полинуклеотид, содержащий:
нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:50, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:60; или
нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:52, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:62; или
нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:54, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:64; или
нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:54, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-396 нуклеотидной последовательности SEQ ID NO:66; или
нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:56, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:68; или
нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:58, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:70; или
нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:68; или
нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:100, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:68; или
нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:102; или
нуклеотидную последовательность, содержащую нуклеотиды 58-420 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-399 нуклеотидной последовательности SEQ ID NO:104; или
нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:50, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:60; или
нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:52, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:62; или
нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:54, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:64; или
нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:54, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-711 нуклеотидной последовательности SEQ ID NO:66; или
нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности, SEQ ID NO:56, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:68; или
нуклеотидную последовательность, содержащую нуклеотиды 58-1410 нуклеотидной последовательности SEQ ID NO:58, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:70; или
нуклеотидную последовательность, содержащую нуклеотиды 58-1398 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:68; или
нуклеотидную последовательность, содержащую нуклеотиды 58-1398 нуклеотидной последовательности SEQ ID NO:100, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:68; или
нуклеотидную последовательность, содержащую нуклеотиды 58-1398 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:102; или
нуклеотидную последовательность, содержащую нуклеотиды 58-1398 нуклеотидной последовательности SEQ ID NO:98, и полинуклеотид, содержащий нуклеотидную последовательность, содержащую нуклеотиды 61-714 нуклеотидной последовательности SEQ ID NO:104.
70. Вектор экспрессии, содержащий полинуклеотид по пп.65-69.
71. Трансформированная клетка-хозяин, содержащая полинуклеотид по пп.65-69, использующаяся для экспрессии антитела по любому из пп.2, 6, 9-29.
72. Трансформированная клетка-хозяин, содержащая вектор по п.70, использующаяся для экспрессии антитела по любому из пп.2, 6, 9-29.
73. Способ получения антитела по любому из пп.2, 6 и 9-29, включающий культивирование клетки-хозяина по п.71 или 72 и очистку антитела из полученного продукта культивирования.

