Область техники
Настоящее изобретение относится к антителу, которое связывается с GARP и пригодно в качестве терапевтического противоопухолевого агента, и к способу лечения опухоли с использованием вышеуказанного антитела.
Уровень техники
Регуляторные Т-клетки (Treg) являются основными побудительными клетками, индуцирующими иммунную толерантность, которая наблюдается в опухолевой области онкологических больных. То есть у онкологических больных группы иммунных клеток, которые по существу работают для уничтожения опухолей, оказываются в состоянии иммуносупрессии активированными Treg в опухоли, и это приводит к злокачественной прогрессии опухоли [Непатентная публикация 1].
Преобладающие Повторы Гликопротеина-A (GARP) представляет собой белок с однопроходной трансмембранной структурой [Непатентная публикация 2], и этот белок экспрессируется на клеточной поверхности активированных Treg и образует комплекс с латентным TGF-β (предшественником TGF-β, который является важной молекулой для индуцирования иммунной толерантности) [Непатентная публикация 3].
В результате межклеточных взаимодействий между клетками Treg и клетками-мишенями, к которым Treg индуцируют иммуносупрессию, TGF-β созревает из латентного TGF-β с помощью GARP на клеточной поверхности Treg и секретируется из Treg, а иммуносупрессивные сигналы TGF-β непосредственно передаются клеткам-мишеням [Непатентная публикация 4, 5]. Было продемонстрировано, что связанный с мембраной GARP, экспрессируемый на клеточной поверхности, необходим для такого созревания TGF-β [Непатентная публикация 5]. С другой стороны, было также продемонстрировано, что растворимый GARP, который не имеет трансмембранной области, подавляет пролиферацию CD4-положительных Т-клеток, когда его непосредственно добавляют к культуре клеток [Непатентная публикация 6]. Таким образом, нельзя исключать, что существует иммуносупрессивный механизм GARP, который не требует созревания TGF-β на клеточной мембране.
GARP экспрессируется не только клетками Treg из периферической крови, когда они активированы, но и в клинических условиях инфильтрующими опухоль T-клетками в опухолевых очагах онкологических больных [Непатентная публикация 7], Treg, существующими в асцитах [Непатентная публикация 8], а также Treg, циркулирующими в периферической крови онкологических больных [Непатентная публикация 9].
В сообщении, посвященном изучению влияния ингибирования экспрессии GARP на функцию Treg, GARP, подвергнутый воздействию киРНК, ингибировал иммуносупрессивную функцию Treg в отношении пролиферативных ответов хелперных Т-клеток, но такой ингибирующий эффект был неполным [Непатентная публикация 10].
В другом сообщении антитела против GARP (MHG-8 и LHG-10), которые были получены благодаря их способности ингибировать созревание TGF-β, ингибировали супрессивную функцию клеток A1, которые являются клеточной линией Treg [Непатентная публикация 11], созданной из клеток пациентов с гемохроматозом, в отношении пролиферативных ответов хелперных Т-клеток [Патентная публикация 1 и Непатентная публикация 12]. Однако неизвестно, действительно ли вышеуказанные антитела эффективно проявляют такие ингибирующие эффекты в отношении Treg в микроокружении опухоли, и до настоящего времени не было сообщений об антителах против GARP, обладающих такими эффектами. Также известно антитело, распознающее как GARP, так и TGF-β. [Патентная публикация 2].
Было продемонстрировано, что чрезмерное присутствие и активация Treg у пациентов с малярией и ВИЧ-инфекцией демонстрирует корреляцию с болезненным состоянием [Непатентная публикация 13 и 14], и что удаление Treg приводит к ремиссии болезненного состояния в мышиных моделях [Непатентная публикация 15 и 16].
Список цитирования
Патентные публикации
Патентная публикация 1: WO2015/015003
Патентная публикация 2: WO2016/125017
Непатентные публикации
Непатентная публикация: 1: Int J Cancer. 2010 Aug 15; 127(4): 759-67.
Непатентная публикация: 2: PLoS One. 2008; 3(7): e2705.
Непатентная публикация: 3: Proc Natl Acad Sci USA. 2009; 106(32): 13445-50.
Непатентная публикация: 4: Eur J Immunol. 2009; 39(12): 3315-22.
Непатентная публикация: 5: Mol Biol Cell. 2012; 23(6): 1129-39.
Непатентная публикация: 6: Blood. 2013; 122(7): 1182-91.
Непатентная публикация: 7: Eur J Immunol. 2012 Jul; 42(7): 1876-85.
Непатентная публикация: 8: Clin Immunol. 2013 Oct; 149(1): 97-110.
Непатентная публикация: 9: Cancer Res. 2013; 73: 2435.
Непатентная публикация: 10: Proc Natl Acad Sci USA. 2009 Aug 11; 106(32): 13445-50.
Непатентная публикация: 11: Eur J Immunol. 2009; 39(12): 869-82.
Непатентная публикация: 12: Sci Transl Med. 2015 Apr 22; 7(284)
Непатентная публикация: 13: PLoS One. 2008 Apr 30; 3(4): e2027.
Непатентная публикация: 14: Clin Exp Immunol. 2014 Jun; 176(3): 401-9.
Непатентная публикация: 15: J Immunol. 2012 Jun 1; 188(11): 5467-77.
Непатентная публикация: 16: PLoS Pathog. 2013; 9(12): e1003798.
Сущность изобретения
Техническая Проблема
Целью настоящего изобретения является предложение антитела, которое ингибирует функцию Treg в опухоли и, таким образом, используется в качестве фармацевтического продукта, обладающего терапевтическими эффектами, предложение способа лечения опухоли с использованием вышеуказанного антитела и тому подобного.
Решение проблемы
Авторы настоящего изобретения провели интенсивные исследования, направленные на достижение вышеуказанной цели. В результате авторы изобретения обнаружили антитело, которое специфично связывается с GARP и проявляет активность ингибирования функции Treg через антителозависимую клеточную цитотоксичность, тем самым осуществив настоящее изобретение. Конкретно, настоящее изобретение включает следующие аспекты изобретения.
(1) Антитело, обладающее следующими свойствами:
(а) специфичное связывание с Преобладающими Повторами Гликопротеина-A (GARP);
(b) наличие ингибирующей активности по отношению к иммуносупрессивной функции регуляторных Т-клеток;
(c) наличие активности антителозависимой клеточной цитотоксической (ADCC); и
(d) наличие in vivo противоопухолевой активности.
(2) Антитело согласно (1) выше, где GARP представляет собой молекулу, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 1.
(3) Антитело согласно (1) или (2) выше, которое связывается с:
(а) частями аминокислотной последовательности в положениях аминокислот 366-377, 407-444 и 456-470, представленной в SEQ ID NO: 1,
(b) частями аминокислотной последовательности в положениях аминокислот 54-112 и 366-339, представленной в SEQ ID NO: 1,
(c) частями аминокислотной последовательности в положениях аминокислот 352-339, представленной в SEQ ID NO: 1, или
(d) частями аминокислотной последовательности в положениях аминокислот 18-112, представленной в SEQ ID NO: 1.
(4) Антитело по любому из (1)-(3) выше, которое обладает конкурентной ингибирующей активностью за связывание с GARP против антитела, имеющего:
(а) тяжелую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 2, и легкую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 3,
(b) тяжелую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 4, и легкую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 5,
(c) тяжелую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 25, и легкую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 27, или
(d) тяжелую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 29, и легкую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 31.
(5) Антитело по любому из (1)-(4) выше, где опухоль представляет собой злокачественное новообразование.
(6) Антитело по (5) выше, где злокачественное новообразование представляет собой рак легкого, рак почки, уротелиальный рак, рак толстой кишки, рак предстательной железы, мультиформную глиобластому, рак яичников, рак поджелудочной железы, рак молочной железы, меланому, рак печени, рак мочевого пузыря, рак желудка, рак пищевода или гемобластоз.
(7) Антитело по любому из (1)-(6) выше, которое содержит:
(а) CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 26-35, представленной в SEQ ID NO: 2, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 50-66, представленной в SEQ ID NO: 2, и CDRH3, состоящую из аминокислотной последовательности в положениях аминокислот 99-107, представленной в SEQ ID NO: 2 и CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 23-36, представленной в SEQ ID NO: 3, CDRL2, состоящей из аминокислотной последовательности в положениях аминокислот 52-58, представленной в SEQ ID NO: 3, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 91-101, представленной в SEQ ID NO: 3,
(b) CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 26-35, представленной в SEQ ID NO: 4, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 50-66, представленной в SEQ ID NO: 4, и CDRH3, состоящую из аминокислотной последовательности в положениях аминокислот 99-112, представленной в SEQ ID NO: 4, и CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 23-36, представленной в SEQ ID NO: 5, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 52-58, представленной в SEQ ID NO: 5, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 91-100, представленной в SEQ ID NO: 5,
(c) CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 45-54, представленной в SEQ ID NO: 25, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 69-78, представленной в SEQ ID NO: 25, и CDRH3, состоящую из аминокислотной последовательности в положениях аминокислот 118-125, представленной в SEQ ID NO: 25, и CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 44-44, представленной в SEQ ID NO: 27, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 70-76, представленной в SEQ ID NO: 27, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 109-137, представленной в SEQ ID NO: 27, или
(d) CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 45-54, представленной в SEQ ID NO: 29, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 69-77, представленной в SEQ ID NO: 29, и CDRH3, состоящую из аминокислотной последовательности в положениях аминокислот 117-128, представленной в SEQ ID NO: 29, и CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 44-54, представленной в SEQ ID NO: 31, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 70-76, представленной в SEQ ID NO: 31, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 109-117, представленной в SEQ ID NO: 31.
(8) Антитело по любому из (1)-(7) выше, которое содержит:
(а) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 1-118, представленной в SEQ ID NO: 2, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 1-112, представленной в SEQ ID NO: 3,
(b) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 1-123, представленной в SEQ ID NO: 4, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 1-111, представленной в SEQ ID NO: 5,
(c) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 20-136, представленной в SEQ ID NO: 25, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 21-129, представленной в SEQ ID NO: 27 или
(d) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 20-99, представленной в SEQ ID NO: 29, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 21-129, SEQ ID NO: 31.
(9) Антитело по любому из (1)-(8) выше, где константная область имеет человеческое происхождение.
(10) Антитело по любому из (1)-(9) выше, которое содержит:
(а) тяжелую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 2, и легкую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 3,
(b) тяжелую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 4, и легкую цепь, состоящую из аминокислотной последовательности, представленной в SEQ ID NO: 5,
(c) тяжелую цепь, состоящую из аминокислотной последовательности в положениях аминокислот 20-446, представленной в SEQ ID NO: 25, и легкую цепь, состоящую из аминокислотной последовательности в положениях аминокислот 21-234, представленной в SEQ ID NO: 27, или
(d) тяжелую цепь, состоящую из аминокислотной последовательности в положениях аминокислот 20-449, представленной в SEQ ID NO: 29, и легкую цепь, состоящую из аминокислотной последовательности в положениях аминокислот 21-234, представленной в SEQ ID NO: 31.
(11) Антитело по любому из (1)-(10) выше, которое является гуманизированным.
(12) Антитело по п.11 выше, которое содержит:
вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, выбранной из группы, состоящей из:
(а) аминокислотной последовательности в положениях аминокислот 20-36, представленной в SEQ ID NO: 33,
(b) аминокислотной последовательности в положениях аминокислот 20-136, представленной в SEQ ID NO: 35,
(с) аминокислотной последовательности в положениях аминокислот 20-99, представленной в SEQ ID NO: 41,
(d) аминокислотной последовательности, имеющей по меньшей мере 95% или более гомологии с последовательностью каркасной области, отличной от каждой последовательности CDR в последовательностях (а)-(с), и
(e) аминокислотной последовательности, содержащей делецию, замену или вставку одной или нескольких аминокислот в последовательности каркасной области, отличной от каждой последовательности CDR в последовательностях (a)-(c), и
вариабельную область легкой цепи, состоящую из аминокислотной последовательности, выбранной из группы, состоящей из:
(f) аминокислотной последовательности в положениях аминокислот 21-129, представленной в SEQ ID NO: 37,
(g) аминокислотной последовательности в положениях аминокислот 21-129, представленной в SEQ ID NO: 39,
(h) аминокислотной последовательности в положениях аминокислот 21-129, представленной в SEQ ID NO: 43,
(i) аминокислотной последовательности, имеющей гомологию, составляющую по меньшей мере 95% или более, с последовательностью каркасной области, отличной от каждой последовательности CDR в последовательностях (f)-(h), и
(j) аминокислотной последовательности, содержащей делецию, замену или вставку одной или нескольких аминокислот в последовательности каркасной области, отличной от каждой последовательности CDR в последовательностях (f)-(h).
(13) Антитело по (11) или (12) выше, которое содержит:
(а) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 20-13, представленной в SEQ ID NO: 33, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 21-129, представленной в SEQ ID NO: 37,
(b) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 20-13, представленной в SEQ ID NO: 35, и вариабельную область легкой цепи, состоящей из аминокислотной последовательности в положениях аминокислот 21-129, представленной в SEQ ID NO: 39, или
(с) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 21-99, представленной в SEQ ID NO: 41, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности в положениях аминокислот 21-129, представленной в SEQ ID NO: 43.
(14) Антитело по любому из (11)-(13) выше, которое содержит:
(а) тяжелую цепь, выбранную из группы, состоящей из тяжелой цепи, имеющей аминокислотную последовательность в положениях аминокислот 20-446, представленную в SEQ ID NO: 33, тяжелой цепи, имеющей аминокислотную последовательность в положениях аминокислот 20-446, представленную в SEQ ID NO: 35, и тяжелой цепи, имеющей аминокислотную последовательность в положениях аминокислот 20-449, представленную в SEQ ID NO: 41, и
(b) легкую цепь, выбранную из группы, состоящей из легкой цепи, имеющей аминокислотную последовательность в положениях аминокислот 21-234, представленную в SEQ ID NO: 37, легкой цепи, имеющей аминокислотную последовательность в положениях аминокислот 21-234, представленную в SEQ ID NO: 39, и легкой цепи, имеющей аминокислотную последовательность в положениях аминокислот 21-234, представленную в SEQ ID NO: 43.
(15) Антитело по любому из (11)-(14) выше, которое содержит:
(а) тяжелую цепь, имеющую аминокислотную последовательность в положениях аминокислот 20-466, представленную в SEQ ID NO: 33, и легкую цепь, имеющую аминокислотную последовательность в положениях аминокислот 21-234, представленную в SEQ ID NO: 37,
(b) тяжелую цепь, имеющую аминокислотную последовательность в положениях аминокислот 20-446, представленную в SEQ ID NO: 35, и легкую цепь, имеющую аминокислотную последовательность в положениях аминокислот 21-234, представленную в SEQ ID NO: 39, или
(c) тяжелую цепь, имеющую аминокислотную последовательность в положениях аминокислот 20-449, представленную в SEQ ID NO: 41, и легкую цепь, имеющую аминокислотную последовательность в положениях аминокислот 21-234, представленную в SEQ ID NO: 43.
(16) Полинуклеотид, кодирующий антитело по любому из (1)-(15) выше.
(17) Полинуклеотид по (16) выше, который содержит:
(а) полинуклеотид CDRH1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 76-105, представленной в SEQ ID NO: 6, полинуклеотид CDRH2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 148-198, представленной в SEQ ID NO: 6, и полинуклеотид CDRH3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 295-321, представленной в SEQ ID NO: 6, и полинуклеотид CDRL1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 67-108, представленной в SEQ ID NO: 7, полинуклеотид CDRL2 состоящий из нуклеотидной последовательности в положениях нуклеотидов 154-174, представленной в SEQ ID NO: 7, и полинуклеотид CDRL3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 271-303, представленной в SEQ ID NO: 7,
(b) полинуклеотид, кодирующий CDRH1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 76-105, представленной в SEQ ID NO: 8, полинуклеотид CDRH2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 148-198, представленной в SEQ ID NO: 8, и полинуклеотид CDRH3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 295-336, представленной в SEQ ID NO: 8, и полинуклеотид CDRL1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 67-108, представленной в SEQ ID NO: 9, полинуклеотид CDRL2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 154-174, представленной в SEQ ID NO: 9, и полинуклеотид CDRL3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 271-300, представленной в SEQ ID NO: 9,
(c) полинуклеотид CDRH1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 133-162, представленной в SEQ ID NO: 24, полинуклеотид CDRH2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 205-234, представленной в SEQ ID NO: 24, и полинуклеотид CDRH3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 352-337, представленной в SEQ ID NO: 24, и полинуклеотид CDRL1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 130-162, представленной в SEQ ID NO: 26, полинуклеотид CDRL2 состоящий из нуклеотидной последовательности в положениях нуклеотидов 208-228, представленной в SEQ ID NO: 26, и полинуклеотид CDRL3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 325-351, представленной в SEQ ID NO: 26, или
(d) полинуклеотид CDRH1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 133-162, представленной в SEQ ID NO: 28, полинуклеотид CDRH2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 205-231, представленной в SEQ ID NO: 28, и полинуклеотид CDRH3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 349-338, представленной в SEQ ID NO: 28, и полинуклеотид CDRL1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 130-162, представленной в SEQ ID NO: 30, полинуклеотид CDRL2 состоящий из нуклеотидной последовательности в положениях нуклеотидов 208-228, представленной в SEQ ID NO: 30, и полинуклеотида CDRL3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 325-351, представленной в SEQ ID NO: 30.
(18) Полинуклеотид по (16) или (17) выше, который содержит:
(а) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 1-354, представленной в SEQ ID NO: 6, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 1-336, представленной в SEQ ID NO: 7,
(b) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 1-369, представленной в SEQ ID NO: 8, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 1-333, представленной в SEQ ID NO: 9,
(c) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-408, представленной в SEQ ID NO: 24, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-387, представленной в SEQ ID NO: 26, или
(d) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-417, представленной в SEQ ID NO: 28, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-387, представленной в SEQ ID NO: 30.
(19) Полинуклеотид по любому из (16)-(18) выше, который содержит:
(а) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности, представленной в SEQ ID NO: 6, и полинуклеотид легкой цепи, состоящей из нуклеотидной последовательности, представленной в SEQ ID NO: 7,
(b) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности, представленной в SEQ ID NO: 8, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности, представленной в SEQ ID NO: 9,
(c) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-1398, представленной в SEQ ID NO: 24, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-702, представленной в SEQ ID NO: 26, или
(d) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-1407, представленной в SEQ ID NO: 28, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-702, представленной в SEQ ID NO: 30.
(20) Полинуклеотид по (16) или (17) выше, который содержит:
(а) полинуклеотид вариабельной области тяжелой цепи, выбранный из группы, состоящей из полинуклеотида вариабельной области тяжелой цепи, состоящей из нуклеотидной последовательности в положениях нуклеотидов 58-408, представленной в SEQ ID NO: 32, полинуклеотида вариабельной области тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-408, представленной в SEQ ID NO: 34, и полинуклеотида вариабельной области тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-417, представленной в SEQ ID NO: 40, и
(b) полинуклеотид вариабельной области легкой цепи, выбранный из группы, состоящей из полинуклеотида вариабельной области легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-387, представленной в SEQ ID NO: 36, полинуклеотида вариабельной области легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-387, представленной в SEQ ID NO: 38, и полинуклеотида вариабельной области легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-387, представленной в SEQ ID NO: 42.
(21) Полинуклеотид по (16), (17) или (20) выше, который содержит:
(а) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-408, представленной в SEQ ID NO: 32, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-387, представленной в SEQ ID NO: 36,
(b) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-408, представленной в SEQ ID NO: 34, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-387, представленной в SEQ ID NO: 38, или
(c) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-417, представленной в SEQ ID NO: 40, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-387, представленной в SEQ ID NO: 42.
(22) Полинуклеотид по (16), (17), (20) или (21) выше, который содержит:
(а) полинуклеотид тяжелой цепи, выбранный из группы, состоящей из полинуклеотида тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-1398, представленной в SEQ ID NO: 32, полинуклеотида тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-1398, представленной в SEQ ID NO: 34, и полинуклеотида тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-1407, представленной в SEQ ID NO: 40, и
(b) полинуклеотид легкой цепи, выбранный из группы, состоящей из полинуклеотида легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-702, представленной в SEQ ID NO: 36, полинуклеотида легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-702, представленной в SEQ ID NO: 38, и полинуклеотида легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-702, представленной в SEQ ID NO: 42.
(23) Полинуклеотид по любому из (16), (17) и (20)-(22) выше, который содержит:
(а) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-1398, представленной в SEQ ID NO: 32, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-702, представленной в SEQ ID NO: 36,
(b) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-1398, представленной в SEQ ID NO: 34, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-770, представленной в SEQ ID NO: 38, или
(c) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-1407, представленной в SEQ ID NO: 40, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-702, представленной в SEQ ID NO: 42.
(24) Экспрессирующий вектор, содержащий полинуклеотид по любому из (16)-(23) выше.
(25) Клетки-хозяева, трансформированные экспрессирующим вектором по (24).
(26) Способ получения интересующего антитела или его фрагмента, который включает стадию культивирования клеток-хозяев по (25) выше и стадию сбора интересующего антитела из культуры, полученной с помощью вышеуказанной стадии.
(27) Антитело, полученное способом получения по (26) выше.
(28) Антитело по любому из (1)-(15) и (27) выше, содержащее одну или две или более модификаций, выбранных из группы, состоящей из N-связанного гликозилирования, O-связанного гликозилирования, N-концевого процессирования, C-концевого процессирования, дезамидирования, изомеризации аспарагиновой кислоты, окисления метионина, добавления остатка метионина к N-концу, амидирования остатка пролина и тяжелой цепи, содержащей делецию одной или двух аминокислот на карбоксильном конце.
(29) Антитело по (28) выше, где одна или две аминокислоты делетированы на карбоксильном конце тяжелой цепи.
(30) Антитело по (29) выше, где одна аминокислота делетирована на каждом из карбоксильных концов обеих их тяжелых цепей.
(31) Антитело по любому из (28)-(30) выше, где остаток пролина на карбоксильном конце его тяжелой цепи дополнительно амидирован.
(32) Антитело по любому из (1)-(15) и (27)-(31) выше, где модификация сахарной цепи регулируется для усиления антителозависимой клеточной цитотоксичности.
(33) Фармацевтическая композиция, содержащая по меньшей мере одно из антител по (1)-(15) и (27)-(32) выше.
(34) Фармацевтическая композиция по (33) выше, которая предназначена для применения для противоопухолевой терапии.
(35) Фармацевтическая композиция по (34) выше, где опухоль представляет собой злокачественное новообразование.
(36) Фармацевтическая композиция по (35) выше, где злокачественное новообразование представляет собой рак легких, рак почек, уротелиальный рак, рак толстой кишки, рак предстательной железы, мультиформную глиобластому, рак яичников, рак поджелудочной железы, рак молочной железы, меланому, рак мочевого пузыря, рак желудка, рак пищевода или гемобластоз.
(37) Способ лечения опухоли, который включает введение индивидууму по меньшей мере одного из антител по (1)-(15) и (27)-(32) выше.
(38) Способ лечения по (37) выше, где опухоль представляет собой злокачественное новообразование.
(39) Способ лечения по (38) выше, где злокачественное новообразование представляет собой рак легкого, рак почки, уротелиальный рак, рак толстой кишки, рак предстательной железы, мультиформную глиобластому, рак яичников, рак поджелудочной железы, рак молочной железы, меланому, рак печени, рак мочевого пузыря, рак желудка, рак пищевода или гемобластоз.
Предпочтительные эффекты изобретения
Согласно настоящему изобретению может быть получен терапевтический агент для лечения злокачественного новообразования, содержащий антитело, связывающееся с GARP, и обладающий противоопухолевой активностью, вызванной ADCC-опосредованным ингибированием Treg. Кроме того, чрезмерное присутствие и активация Treg у пациентов с малярией и ВИЧ-инфекцией демонстрирует корреляцию с этим болезненным состоянием, а удаление Treg индуцирует ремиссию болезненного состояния в мышиных моделях заболеваний. Соответственно, можно ожидать, что эффективное ингибирование функции Treg будет оказывать терапевтическое воздействие также на рефрактерные инфекционные заболевания, такие как вызванные малярией и ВИЧ.
Краткое описание чертежей
[Фигура 1]. На Фигуре 1 представлена аминокислотная последовательность (SEQ ID NO: 1) GARP.
[Фигура 2]. На Фигуре 2 представлена аминокислотная последовательность (SEQ ID NO: 2) тяжелой цепи антитела 105F.
[Фигура 3]. На Фигуре 3 представлена аминокислотная последовательность (SEQ ID NO: 3) легкой цепи антитела 105F.
[Фигура 4]. На Фигуре 4 представлена аминокислотная последовательность (SEQ ID NO: 4) тяжелой цепи антитела 110F.
[Фигура 5]. На Фигуре 5 представлена аминокислотная последовательность (SEQ ID NO: 5) легкой цепи антитела 110F.
[Фигура 6]. На Фигуре 6 представлена нуклеотидная последовательность (SEQ ID NO: 6) тяжелой цепи антитела 105F.
[Фигура 7]. На Фигуре 7 представлена нуклеотидная последовательность (SEQ ID NO: 7) легкой цепи антитела 105F.
[Фигура 8]. На Фигуре 8 представлена нуклеотидная последовательность (SEQ ID NO: 8) тяжелой цепи антитела 110F.
[Фигура 9]. На Фигуре 9 представлена показана нуклеотидная последовательность (SEQ ID NO: 9) легкой цепи антитела 110F.
[Фигура 10]. На Фигуре 10 представлено связывание антител с GARP. Антитело 105F и антитело 110F демонстрировали их связывание с GARP в соответствии с методом ИФА.
[Фигура 11]. На Фигуре 11 представлено специфичное связывание антител с GARP. Антитело 105F не связывалось с клетками HEK293T, в которые был введен пустой вектор, и демонстрировало связывание с клетками HEK293T, в которых транзиторно экспрессировался GARP.
[Фигура 12]. На Фигуре 12 представлено специфичное связывание антитела с GARP. Антитело 105F проявляло активность связывания с клетками L428, эндогенно экспрессирующими GARP.
[Фигура 13]. На Фигуре 13 представлено специфичное связывание антител с GARP. Антитело 105F проявляло активность связывания с активированными Treg.
[Фигура 14]. На Фигуре 14 представлена активность ADCC антител. Когда клетки L428, эндогенно экспрессирующие GARP, были подвергнуты направленному воздействию, то наблюдали увеличение активности ADCC в зависимости от концентрации антитела 105F.
[Фигура 15]. На Фигуре 15 представлена ингибирующая активность антитела в отношении функции Treg. Антитело 105F (50 мкг/мл) ингибировало супрессивную функцию Treg в отношении пролиферации против хелперных Т-клеток.
[Фигура 16]. На Фигуре 16 представлена ингибирующая активность антитела в отношении функции Treg. Антитело 105F (10 мкг/мл) ингибировало супрессивную функцию Treg в отношении пролиферации против хелперных Т-клеток. С другой стороны, антитела MHG-8 и LHG-10 не демонстрировали влияния на супрессивную функцию Treg в отношении пролиферации против хелперных Т-клеток.
[Фигура 17]. На Фигуре 17 представлена аминокислотная последовательность (SEQ ID NO: 25) тяжелой цепи антитела c151D.
[Фигура 18]. На Фигуре 18 представлена аминокислотная последовательность (SEQ ID NO: 27) легкой цепи антитела c151D.
[Фигура 19]. На Фигуре 19 представлена аминокислотная последовательность (SEQ ID NO: 29) тяжелой цепи антитела c198D.
[Фигура 20]. На Фигуре 20 представлена аминокислотная последовательность (SEQ ID NO: 31) легкой цепи антитела c198D.
[Фигура 21]. На Фигуре 21 представлена аминокислотная последовательность (SEQ ID NO: 33) тяжелой цепи h151D-H1.
[Фигура 22]. На Фигуре 22 представлена аминокислотная последовательность (SEQ ID NO: 37) легкой цепи h151D-L1.
[Фигура 23]. На Фигуре 23 представлена аминокислотная последовательность (SEQ ID NO: 35) тяжелой цепи h151D-H4.
[Фигура 24]. На Фигуре 24 представлена аминокислотная последовательность (SEQ ID NO: 39) легкой цепи h151D-L4.
[Фигура 25]. На Фигуре 25 представлена аминокислотная последовательность (SEQ ID NO: 41) тяжелой цепи h198D-H3.
[Фигура 26]. На Фигуре 26 представлена аминокислотная последовательность (SEQ ID NO: 43) легкой цепи h198D-L4.
[Фигура 27]. На Фигуре 27 представлена нуклеотидная последовательность (SEQ ID NO: 24) тяжелой цепи антитела c151D.
[Фигура 28]. На Фигуре 28 представлена нуклеотидная последовательность (SEQ ID NO: 26) легкой цепи антитела c151D.
[Фигура 29]. На Фигуре 29 представлена нуклеотидная последовательность (SEQ ID NO: 28) тяжелой цепи антитела c198D.
[Фигура 30]. На Фигуре 30 представлена нуклеотидная последовательность (SEQ ID NO: 30) легкой цепи антитела c198D.
[Фигура 31]. На Фигуре 31 представлена нуклеотидная последовательность (SEQ ID NO: 32) тяжелой цепи антитела h151D-H1.
[Фигура 32]. На Фигуре 32 представлена нуклеотидная последовательность (SEQ ID NO: 36) тяжелой цепи антитела h151D-L1.
[Фигура 33]. На Фигуре 33 представлена нуклеотидная последовательность (SEQ ID NO: 34) тяжелой цепи h151D-H4.
[Фигура 34]. На Фигуре 34 представлена нуклеотидная последовательность (SEQ ID NO: 38) легкой цепи h151D-L4.
[Фигура 35]. На Фигуре 35 представлена нуклеотидная последовательность (SEQ ID NO: 40) тяжелой цепи h198D-H3.
[Фигура 36]. На Фигуре 36 представлена нуклеотидная последовательность (SEQ ID NO: 42) легкой цепи h198D-L4.
[Фигура 37]. На Фигуре 37 представлена активность связывания каждого антитела с GARP-экспрессирующими клетками. h151D-H1L1, h151D-H4L4 и h198D-H3L4 проявляли специфичную активность связывания с GARP.
[Фигура 38]. На Фигуре 38 представлена активность связывания каждого антитела с клетками, ко-экспрессирующими GARP-TGFβ1. Индивидуальные антитела 105F, h151D-H1L1, h151D-H4L4 и h198D-H3L4, связанные как с GARP, так и с GARP-мутантом, которые ко-экспрессировались с TGFβ1, и эти антитела проявляли активность связывания с другой областью в GARP, отличной от случая известных антител MHG8 и LHG10.
[Фигура 39]. На Фигуре 39 представлена активность связывания каждого антитела с клетками L428. Индивидуальные антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4 проявляли активность связывания с эндогенно экспрессированным GARP.
[Фигура 40] На Фигуре 40 представлена активность связывания каждого антитела с Treg. Индивидуальные антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4 проявляли активность связывания с FoxP3-положительными Treg.
[Фигура 41] На Фигуре 41 представлена активность ADCC для каждого антитела. Индивидуальные антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4 проявляли активность ADCC.
[Фигура 42] На Фигуре 42 представлена ингибирующая активность каждого антитела в отношении функции Treg. Индивидуальные антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4 проявляли ингибирующую активность в отношении функции Treg.
[Фигура 43] На Фигуре 43 представлена супрессивная активность Treg в отношении активности лизиса клеток-мишеней клетками CTL.
[Фигура 44] На Фигуре 44 представлено увеличение противоопухолевой активности каждого антитела. Индивидуальные антитела 105F, h151D-H1L1, h151D-H4L4 и h198D-H3L4 ингибировали супрессивную функцию Treg в отношении активности клеточного лизиса клетками CTL.
[Фигура 45] На Фигуре 45 представлена in vivo противоопухолевая активность каждого антитела. Индивидуальные антитела 105F, h151D-H1L1, h151D-H4L4 и h198D-H3L4 проявляли противоопухолевую активность в моделях in vivo.
ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Используемый в настоящем описании термин «злокачественное новообразование» имеет то же значение, что и термин «опухоль».
Используемый в настоящем описании термин «ген» включает не только ДНК, но также ее мРНК и кДНК и ее кРНК.
Используемый в настоящем описании термин «полинуклеотид» имеет то же значение, что и нуклеиновая кислота, и он включает ДНК, РНК, зонд, олигонуклеотид и праймер.
В настоящем описании термин «полипептид» используется так, что он не отличается от термина «белок».
В настоящем описании термин «клетка» включает клетки в индивидуальном животном и культивируемые клетки.
Используемый в настоящем описании термин «GARP» имеет то же значение, что и для белка GARP.
Используемый в настоящем описании термин «цитотоксичность» обозначает, что патологическое изменение вызвано в клетках любым способом. Это означает не только прямое повреждение, но также означает все типы структурных или функциональных повреждений, вызванных в клетках, такие как расщепление ДНК, образование димера основания, хромосомное расщепление, повреждение клеточного митотического аппарата и снижение активностей ферментов различных типов.
Используемый в настоящем описании термин «цитотоксическая активность» обозначает активность, которая вызывает описанную выше цитотоксичность.
Используемый в настоящем описании термин «антителозависимая клеточная цитотоксичность» обозначает «зависимую от антитела клеточную цитотоксическую активность (ADCC)», и эта активность означает опосредованный антителом эффект или активность повреждения клеток-мишеней, таких как опухолевые клетки, NK-клетками.
Используемый в настоящем описании термин «эпитоп» обозначает неполный пептид или неполную трехмерную структуру GARP, с которыми связывается конкретное антитело против GARP. Такой эпитоп, который является неполным пептидом описанного выше GARP, может быть определен способом, хорошо известным специалисту в данной области, таким как иммунологический анализ, например, с помощью следующего способа. Во-первых, получают различные неполные структуры антигена. Что касается получения таких неполных структур, то может быть применен известный способ синтеза олигопептидов. Например, получают ряд пептидов, в которых антиген последовательно укорачивают до соответствующей длины от его С-конца или N-конца с помощью метода генетической рекомбинации, хорошо известного специалисту в данной области, и после этого исследуют реактивность антитела к таким полипептидам, и приблизительно определяют сайты распознавания. После этого синтезируют еще более короткие пептиды и затем исследуют их реактивность с вышеуказанными пептидами, чтобы определить эпитоп. Кроме того, эпитоп, который является неполной трехмерной структурой антигена, который связывается с конкретным антителом, может быть определен путем конкретизации аминокислотных остатков антигена, смежного с вышеописанным антителом, путем рентгеноструктурного анализа.
Используемое в настоящем описании выражение «антитела, связывающиеся с одним и тем же эпитопом», обозначает различные антитела, которые связываются с общим эпитопом. Если второе антитело связывается с неполным пептидом или с неполной трехмерной структурой, с которыми связывается первое антитело, то можно определить, что первое антитело и второе антитело связываются с одним и тем же эпитопом. Кроме того, подтверждая, что второе антитело конкурирует с первым антителом за связывание первого антитела с антигеном (то есть второе антитело препятствует связыванию первого антитела с антигеном), можно определить, что первое антитело и второе антитело связываются с одним и тем же эпитопом, даже если конкретная последовательность или структура эпитопа не определена. Кроме того, когда первое антитело и второе антитело связываются с одним и тем же эпитопом, и к тому же, первое антитело обладает особыми эффектами, такими как противоопухолевая активность, можно ожидать, что второе антитело будет иметь такую же активность, как и у первого антитела. Соответственно, если второе антитело против GARP связывается с неполным пептидом, с которым связывается первое антитело против GARP, можно определить, что первое антитело и второе антитело связываются с одним и тем же эпитопом GARP. Кроме того, подтверждая, что второе антитело против GARP конкурирует с первым антителом против GARP за связывания первого антитела против GARP с GARP, можно определить, что первое антитело и второе антитело являются антителами, связывающимися с одним и тем же эпитопом GARP.
Используемый в настоящем описании термин «CDR» обозначает область, определяющую комплементарность. Известно, что каждая тяжелая цепь и легкая цепь молекулы антитела имеет три CDR. Такая CDR также упоминается как гипервариабельный домен и находится в вариабельной области тяжелой цепи и легкой цепи антитела. Эти области имеют особенно высоко вариабельную первичную структуру и разделены на три участка на первичной структуре полипептидной цепи в каждой из тяжелой цепи и легкой цепи. В настоящем описании, в отношении CDR антитела, CDR тяжелой цепи обозначаются как CDRH1, CDRH2 и CDRH3, соответственно, с N-концевой стороны аминокислотной последовательности тяжелой цепи, тогда как CDR легкой цепи обозначаются как CDRL1, CDRL2 и CDRL3, соответственно, с N-концевой стороны аминокислотной последовательности легкой цепи. Эти сайты расположены близко друг от друга на трехмерной структуре и определяют специфичность антитела к антигену, с которым связывается антитело.
Используемое в настоящем изобретении выражение «гибридизуется в жестких условиях» обозначает, что гибридизацию проводят в коммерчески доступном гибридизационном растворе ExpressHyb Hybridization Solution (производство Clontech) при 68°С или что гибридизацию проводят в условиях с использованием фильтра с иммобилизованной ДНК в присутствии 0,7-1М NaCl при 68°С и после чего промывают при 68°С 0,1-2-кратной концентрацией раствора SSC (где 1xSSC состоит из 150мМ NaCl и 15мМ цитрата натрия) для идентификации или в условиях, эквивалентных этим.
1. GARP
GARP, используемый в настоящем изобретении, может быть непосредственно очищен из GARP-экспрессирующих клеток человека или другого млекопитающего (например, крыса, мышь и т. д.), и затем может быть использован, или может быть получена фракция клеточной мембраны вышеуказанных клеток и может использоваться в качестве GARP. Альтернативно, GARP можно также получить, синтезируя его in vitro или позволив клеткам-хозяевам продуцировать GARP с помощью генетических манипуляций. Согласно такой генетической манипуляции, белок GARP может быть получен, в частности, путем включения кДНК GARP в экспрессирующий вектор, способный экспрессировать кДНК GARP, и затем синтезируя GARP в растворе, содержащем ферменты, субстрат и энергетические материалы, необходимые для транскрипции и трансляции, или путем трансформации клеток-хозяев других прокариот или эукариот, так чтобы они могли экспрессировать GARP.
Аминокислотная последовательность GARP человека представлена в SEQ ID NO: 1 в списке последовательностей. Кроме того, последовательность SEQ ID NO: 1 представлена на Фигуре 1.
Кроме того, белок, который состоит из аминокислотной последовательности, включающей замену, делецию и/или вставку одной или нескольких аминокислот в вышеописанной аминокислотной последовательности GARP, и обладает биологической активностью, эквивалентной биологической активности белка GARP, также включен в понятие GARP.
Зрелый человеческий GARP, из которого удалена сигнальная последовательность, соответствует аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 20-622 в аминокислотной последовательности, представленной в SEQ ID NO: 1.
Кроме того, белок, который состоит из аминокислотной последовательности, содержащей замену, делецию и/или вставку одной или нескольких аминокислот в аминокислотной последовательности, представленной в SEQ ID NO: 1, или из аминокислотной последовательности, представленной в SEQ ID NO: 1, из которой была удалена сигнальная последовательность, и который обладает биологической активностью, эквивалентной активности GARP, также включен в понятие GARP. Кроме того, белок, который состоит из аминокислотной последовательности, кодируемой вариантом сплайсинга, транскрибированным из локуса гена GARP человека, или аминокислотной последовательности, содержащей замену, делецию и/или вставку одной или нескольких аминокислот в вышеуказанной аминокислотной последовательности, и который обладает биологической активностью, эквивалентной биологической активности GARP, также включен в понятие GARP.
2. Получение антител против GARP
Примером антитела против GARP по настоящему изобретению может быть антитело против GARP человека. Антитело против GARP человека означает человеческое антитело, имеющее последовательность генов антитела, полученных исключительно из хромосом человека.
Антитело против GARP человека можно получить с помощью метода с использованием мыши, продуцирующей человеческое антитело, несущей человеческий хромосомный фрагмент, содержащий гены тяжелой цепи и легкой цепи человеческого антитела (см. Tomizuka, K. et al., Nature Genetics (1997) 16, p. 133-143,; Kuroiwa, Y. et al., Nucl.Acids Res. (1998) 26, p. 3447-3448; Yoshida, H. et al., Animal Cell Technology: Basic and Applied Aspects vol. 10, p. 69-73 (Kitagawa, Y., Matsuda, T. and Iijima, S. eds.), Kluwer Academic Publishers, 1999.; Tomizuka, K. et al., Proc. Natl. Acad. Sci. USA (2000) 97, p. 722-727; etc.).
Такая мышь, продуцирующая человеческое антитело, может быть конкретно получена с использованием генетически модифицированного животного, локусы генов тяжелой цепи и легкой цепи эндогенного иммуноглобулина которого были разрушены, и вместо этого были введены локусы генов тяжелой цепи и легкой цепи иммуноглобулина человека с использованием вектора искусственной дрожжевой хромосомы (YAC) или тому подобного, а затем было получено нокаутное животное и трансгенное животное из такого генетически модифицированного животного, после чего скрещивали таких животных друг с другом.
В другом случае антитело против GARP человека можно также получить путем трансформации эукариотических клеток с использованием кДНК, кодирующей каждую из тяжелой цепи и легкой цепи такого человеческого антитела, или предпочтительно с использованием вектора, содержащего кДНК, в соответствии с методами генетической рекомбинации, а затем путем культивирования трансформированных клеток, продуцирующих генетически модифицированное человеческое моноклональное антитело, так что антитело может быть получено из супернатанта культуры. В качестве клеток-хозяев могут использоваться, например, эукариотические клетки и, предпочтительно, клетки млекопитающих, такие как клетки СНО, лимфоциты или миеломы.
Альтернативно, антитело также может быть получено способом получения человеческого антитела с использованием фагового дисплея, где антитело выбирают из библиотеки человеческих антител (см. Wormstone, IM et al., Research Ophthalmology & Visual Science. (2002) 43 (7)), п.2301-2308; Carmen, S. et al., Briefings in Functional Genomics and Proteomics (2002), 1 (2), p. 189-203; Siriwardena, D. et al., Ophthalmology (2002) 109 (3), p. 427-431; etc.). Например, может быть применен метод фагового дисплея, который включает возможность экспрессии вариабельной области человеческого антитела в виде одноцепочечного антитела (scFv) на поверхности фага, а затем отбор фага, связывающегося с антигеном (Nature Biotechnology (2005), 23, (9), стр. 1105-1116).
Анализируя ген фага, который был выбран по его способности связываться с антигеном, можно определить последовательность ДНК, кодирующую вариабельную область человеческого антитела, связывающуюся с антигеном. После определения последовательности ДНК scFv, связывающейся с антигеном, последовательности ДНК константной области антитела дают возможность связываться с ней с получением вектора, экспрессирующего IgG, и содержащего вышеуказанные последовательности, и полученный экспрессирующий вектор затем вводят в подходящие клетки-хозяева и позволяют экспрессироваться в них, получая, таким образом, человеческое антитело (WO92/01047, WO92/20791, WO93/06213, WO93/11236, WO93/19172, WO 95/01438, WO95/15388, Annu. Rev. Immunol (1994) 12, p. 433-455, Nature Biotechnology (2005) 23 (9), p. 1105-1116).
Кроме того, антитело против GARP по настоящему изобретению может быть получено путем иммунизации животного с использованием GARP или любого данного полипептида, выбранного из аминокислотной последовательности GARP, а затем путем сбора и очистки антитела, продуцируемого в живом теле животного. Виды организма GARP, используемого в качестве антигена, не ограничиваются человеком, и, таким образом, животное также может быть иммунизировано GARP, полученным из животного, отличного от человека, такого как мышь или крыса. В этом случае антитело, применимое к заболеванию человека, может быть выбрано путем изучения перекрестной реактивности полученного антитела, связывающегося с гетерологичным GARP, с GARP человека.
Кроме того, клетки, продуцирующие антитела, которые продуцируют антитело против GARP, сливают с клетками миеломы согласно известному методу (например, Kohler and Milstein, Nature (1975) 256, p. 495-497, Kennet, R. ed., Monoclonal Antibodies, p. 365-367, Plenum Press, N.Y. (1980)), чтобы создать гибридомы для получения моноклонального антитела.
Следует отметить, что GARP, используемый в качестве антигена, можно получить, позволив клеткам-хозяевам продуцировать гены GARP в соответствии с генетическими манипуляциями.
В частности, получают вектор, способный экспрессировать ген GARP, и затем вектор вводят в клетки-хозяева, так что ген экспрессируется в них, а затем экспрессированный GARP может быть очищен. В дальнейшем будет конкретно описан способ получения антитела против GARP.
(1) Получение антигена
Примеры антигена, используемого для получения антитела против GARP, могут включать GARP, полипептид, состоящий из по меньшей мере 6 его неполных последовательных аминокислотных последовательностей, и производное, полученное путем добавления любой данной аминокислотной последовательности или носителя к такому GARP или его полипептиду.
GARP может быть непосредственно очищен из опухолевых тканей или опухолевых клеток человека и затем может быть использован. Альтернативно, GARP можно также получить путем его синтеза in vitro или путем разрешения клеткам-хозяевам продуцировать его посредством генетических манипуляций.
Согласно такой генетической манипуляции, антиген может быть получен, в частности, путем включения кДНК GARP в экспрессирующий вектор, способный экспрессировать кДНК GARP, а затем путем синтеза GARP в растворе, содержащем ферменты, субстрат и энергетические материалы, необходимые для транскрипции и трансляции, или путем трансформации клеток-хозяев других прокариот или эукариот, так чтобы они могли экспрессировать GARP.
Также можно получить антиген в виде секреторного белка, позволяя слитому белку, образованному путем лигирования ДНК, кодирующей внеклеточную область GARP, с ДНК, кодирующей константную область антитела, экспрессироваться в виде мембранного белка в подходящем хозяине и/или в векторной системе.
кДНК GARP может быть получена способом, который называется методом ПЦР, который включает в себя осуществление полимеразной цепной реакции (далее именуемой «ПЦР»), например, с использованием библиотеки кДНК, экспрессирующей кДНК GARP в качестве матрицы, а также с использованием праймеров для конкретной амплификации кДНК GARP (см. Saiki, RK, et al.Science (1988) 239, p. 487-489).
Примером in vitro-синтеза полипептида может быть система быстрой трансляции (RTS) производства Roche Diagnostics, но не ограничиваясь этим.
Примеры прокариотических клеток, используемых в качестве клеток-хозяев, могут включать Escherichia coli и Bacillus subtilis. Чтобы трансформировать клетки-хозяева интересующим геном, клетки-хозяева трансформируют плазмидным вектором, содержащим репликон, полученный из видов, совместимых с хозяином, а именно последовательность начала репликации и регуляторную последовательность. В качестве вектора предпочтительным является вектор, имеющий последовательность, способную придавать селективность фенотипа клеткам, подлежащим трансформации.
Примеры эукариотических клеток, используемых в качестве клеток-хозяев, могут включать клетки позвоночных, насекомых и дрожжей. Примеры клеток позвоночных, которые часто могут использоваться, включают клетки COS, которые являются клетками обезьян (Gluzman, Y., Cell (1981) 23, стр. 175-182, ATCC CRL-1650), мышиные фибробласты NIH3T3 (ATCC No. CRL- 1658) и клеточную линию клеток яичников китайского хомячка, дефицитных по дигидрофолатредуктазе (клетки СНО, АТСС CCL-61) (Urlaub, G. and Chasin, LA Proc. Natl. Acad. Sci. USA (1980) 77, p. 4126-4220), но не ограничиваясь ими.
Полученный таким образом трансформант можно культивировать в соответствии с обычными методами, и интересующий полипептид может быть получен внутри или вне клеточной культуры.
В качестве среды, используемой при культивировании, могут быть выбраны различные типы широко используемых сред, если это необходимо, в зависимости от типа выбранных клеток-хозяев. Если клетки-хозяева представляют собой Escherichia coli, то например, антибиотики, такие как ампициллин, или IPMG, могут быть добавлены в среду LB, если необходимо, и затем можно использовать полученную среду.
Рекомбинантный белок, полученный внутри или вне клеток трансформанта в результате описанного выше культивирования, может быть отделен и/или очищен известными методами разделения различных типов с использованием физических или химических свойств белка.
Конкретные примеры способа могут включать обработку с использованием стандартного вещества, осаждающего белок, ультрафильтрацию, различные типы жидкостной хроматографии, такие как молекулярно-ситовая хроматография (гель-фильтрация), абсорбционная хроматография, ионообменная хроматография или аффинная хроматография, метод диализа и их комбинацию.
Кроме того, путем прикрепления гистидиновой метки, состоящей из 6 остатков, к рекомбинантному белку, подлежащему экспрессии, белок может быть эффективно очищен с использованием аффинной никелевой колонки. В другом случае, при связывании Fc-области IgG с подлежащим экспрессии рекомбинантным белком, белок может быть эффективно очищен с использованием колонки с Протеином А.
Комбинируя вышеописанные методы друг с другом, интересующий полипептид может быть получен в крупном масштабе с высоким выходом и с высокой степенью чистоты.
(2) Продуцирование моноклонального антитела против GARP
Примером антитела, специфично связывающегося с GARP, может быть моноклональное антитело, специфично связывающееся с GARP. Способ получения такого моноклонального антитела заключается в следующем.
Для получения моноклонального антитела, как правило, необходимы следующие рабочие стадии.
Конкретно, необходимые рабочие стадии включают:
(а) очистку биополимера, используемого в качестве антигена,
(b) стадию иммунизации животного антигеном путем инъекции, забор крови у животного, изучение титра антител для определения периода удаления селезенки у животного и затем приготовление клеток, продуцирующих антитела,
(c) приготовление клеток миеломы (далее именуемых «миеломы»),
(d) слияние клеток между клетками, продуцирующими антитела, и миеломами,
(e) отбор гибридомной группы, продуцирующей интересующее антитело,
(f) деление на одноклеточные клоны (клонирование),
(g) необязательно, культивирование гибридом для массового продуцирования моноклональных антител или скрещивание животных, которым трансплантировали гибридомы, и
(h) анализ физиологической активности и специфичности связывания полученного таким образом моноклонального антитела или исследование свойств антитела в виде меченого реагента.
Далее будет подробно описан способ получения моноклонального антитела вместе с вышеописанными стадиями. Однако способ получения вышеуказанного антитела не ограничивается этим, и, например, также могут быть использованы клетки, продуцирующие антитела, отличные от клеток селезенки и миелом.
(а) Очистка антигена
В качестве антигена может быть использован GARP, полученный описанным выше способом, или его часть.
Альтернативно, в качестве антигена может быть использована мембранная фракция, полученная из GARP-экспрессирующих рекомбинантных соматических клеток или сами такие GARP-экспрессирующие рекомбинантные соматические клетки или, кроме того, неполный пептид белка по настоящему изобретению, который химически синтезируется согласно способу, хорошо известному специалисту в данной области.
(b) Получение клеток, продуцирующих антитела
Антиген, полученный на стадии (а), смешивают с вспомогательным агентом, таким как полный или неполный адъювант Фрейнда или алюминиевокалиевые квасцы, для получения иммуногена, а затем экспериментальное животное иммунизируют иммуногеном. В качестве такого экспериментального животного может использоваться без каких-либо проблем животное, используемое в известных методах получения гибридом. Конкретные примеры такого животного, которое может быть использовано в настоящем описании, включают мышь, крысу, козу, овцу, быка и лошадь. С точки зрения доступности миеломных клеток, подлежащих слиянию с удаленными клетками, продуцирующими антитела, и т.д., предпочтительно используют мышь или крысу в качестве животного для иммунизации.
Линии фактически используемых мышей и крыс особо не ограничены. В случае мышей примеры линии, которая может быть использована в настоящем описании, включают A, AKR, BALB/c, BDP, BA, CE, C3H, 57BL, C57BL, C57L, DBA, FL, HTH, HT1, LP, NZB, NZW, RF, R III, SJL, SWR, WB и 129. С другой стороны, в случае крыс примеры линии, которая может быть использована в настоящем описании, включают Wistar, Low, Lewis, Sprague Dawley, ACI, BN и Fischer.
Эти мыши и крысы доступны от экспериментальных селекционеров и дистрибьюторов животных, таких как CLEA Japan, Inc. и CHARLES RIVER LABORATORIES JAPAN, INC.
Среди прочего, принимая во внимание совместимость слияния с клетками миеломы, которые обсуждают ниже, линия BALB/c в случае мышей и линии Wistar и Low в случае крыс являются особенно предпочтительными в качестве животных, подлежащих иммунизации.
Кроме того, принимая во внимание гомологию между антигенами человека и мыши, также предпочтительно использовать мышей, чей биологический механизм удаления аутоантител ослаблен, а именно мышей с аутоиммунным заболеванием.
Возраст этих мышей или крыс при иммунизации предпочтительно составляет от 5 до 12 недель и более предпочтительно от 6 до 8 недель.
Чтобы иммунизировать животных с использованием GARP или его рекомбинанта, могут применяться известные способы, которые подробно описаны, например, в Weir, DM, Handbook of Experimental Immunology, Vol. I. II. III, Blackwell Scientific Publications, Oxford (1987), Kabat, EA и Mayer, MM, Experimental Immunochemistry, Charles C Thomas Publisher Springfield, Illinois (1964) и т.д..
Среди этих методов иммунизации метод, предпочтительно применяемый в настоящем изобретении, представляет собой, в качестве конкретного примера следующий способ.
То есть, во-первых, животному вводят внутрикожно или внутрибрюшинно клетки, в которых экспрессируется фракция мембранного белка, используемого в качестве антигена, или антиген.
Для повышения эффективности иммунизации предпочтительным является их комбинированное применение. Эффективность иммунизации может быть особенно усилена, если внутрикожное введение осуществляют в первой половине схемы введения, а внутрибрюшинное введение проводят в ее второй половине или только в последнее введение.
Схема введения антигена различается в зависимости от типа животного, подлежащего иммунизации, индивидуальных различий и т.д.. Вообще предпочтительными являются 3-6 доз антигена и интервал дозирования 2-6 недель и более предпочтительно, 3 или 4 дозы антигена и интервал дозирования 2-4 недели.
Применяемая доза антигена различается в зависимости от типа животного, подлежащего иммунизации, индивидуальных различий и т.д.. Как правило, она составляет 0,05-5 мг и предпочтительно приблизительно 0,1-0,5 мг.
Бустерное введение проводят через 1-6 недель, предпочтительно через 2-4 недели и более предпочтительно через 2-3 недели после описанного выше введения антигена.
Применяемая доза антигена при бустерном введении различается в зависимости от типа животного, его размера и т.д.. В случае мыши, например, применяемая доза антигена составляет, как правило, 0,05-5 мг, предпочтительно 0,1-0,5 мг и более предпочтительно приблизительно 0,1-0,2 мг.
Через 1-10 дней, предпочтительно через 2-5 дня и более предпочтительно через 2 или 3 дня после завершения описанного выше бустерного введения клетки селезенки или лимфоциты, включающие клетки, продуцирующие антитела, асептически удаляют из иммунизированного животного. В это время измеряют титр антител. Животное, в котором титр антитела достаточно высокий, используют в качестве источника клеток, продуцирующих антитела, так что эффективность последующих операций может быть повышена.
Примеры методов измерения титра антител, используемых в настоящем описании, могут включать в себя РИА-метод и метод ИФА, но не ограничиваются ими.
Что касается измерения титра антител в настоящем изобретении, то метод ИФА можно, например, проводить в соответствии со следующими процедурами.
Во-первых, очищенный или частично очищенный антиген адсорбируется на поверхность твердой подложки, такой как 96-луночный планшет для ИФА, а на другую поверхность твердой подложки, на которой такой антиген не адсорбируется, наносят покрытие в виде белка, не относящегося к антигену, такого как бычий сывороточный альбумин (далее именуемый «BSA»). Поверхности промывают и затем дают возможность вступать в контакт с последовательно разбавленным образцом, используемым в качестве первичного антитела (например, мышиная сыворотка), с тем чтобы антитело в образце имело возможность связываться с описанным выше антигеном.
После этого добавляют меченое ферментом антитело против мышиного антитела в качестве вторичного антитела, так что ему дают возможность связаться с мышиным антителом с последующей промывкой. После этого к нему добавляют субстрат фермента и затем измеряют изменение абсорбции с помощью проявления цвета на основе разложения субстрата и т.д., так чтобы рассчитать титр антител.
Клетки, продуцирующие антитело, могут быть отделены от клеток селезенки или лимфоцитов иммунизированного животного в соответствии с известными методами (например, Kohler et al., Nature (1975) 256, p. 495, Kohler et al., Eur.J. Immunol. (1977) 6, p. 511,; Milstein et al., Nature (1977), 266, p. 550,; Walsh, Nature, (1977) 266, p. 495). Например, в случае клеток селезенки может применяться стандартный метод, который включает измельчение селезенки, затем фильтрацию клеток через сито из нержавеющей стали, затем суспендирование фильтрата в минимальной питательной среде Игла (MEM) для отделения клеток, продуцирующих антитела.
(c) Получение миеломных клеток (далее именуемых «миеломы»)
Миеломные клетки, используемые для слияния клеток, особо ничем не ограничены, и клетки могут быть выбраны из известных клеточных линий, если это необходимо, и затем могут быть использованы. Принимая во внимание вопросы удобства при отборе гибридом из слитых клеток, предпочтительно используют клеточные линии, дефицитные по HGPRT (гипоксиантин-GUANINE-фосфорибозилтрансфераза), процедуры отбора которых были установлены.
То есть примеры таких клеточных линий, дефицитных по HGPRT, включают в себя мышиные X63-Ag8 (X63), NS1-ANS/1 (NS1), P3X63-Ag8.U1 (P3U1), X63-Ag8.653 (X63.653), SP2/0-Ag14 (SP2/0), MPC11-45.6TG1.7 (45.6TG), FO, S149/5XXO, и BU. 1; клетки 210 крысиного происхождения. RSY3. Ag. 1. 2,3 (Y3); и клетки U266AR (SKO-007) человеческого происхождения, GM1500⋅GTG-A12 (GM1500), UC729-6, LICR-LOW-HMy2 (HMy2) и 8226AR/NIP4-1 (NP41). Эти клеточные линии, дефицитные по HGPRT, доступны, например, из Американской типированной коллекции клеточных культур (ATCC).
Эти клеточные линии субкультивируют в подходящей среде, такой как 8-азагуаниновая среда [среда, полученная добавлением 8-азагуанина к среде RPMI-1640, содержащей глутамин, 2-меркаптоэтанол, гентамицин и фетальную телячью сыворотку (далее именуемую «FCS»)], среда Дульбекко, модифицированная по способу Исков (далее именуемая «IMDM») или модифицированная Дульбекко среда Игла (далее именуемая «DMEM»). За три или четыре дня до слияния клетки субкультивируют в нормальной среде [например, среда ASF104, содержащая 10% FCS (производства Ajinomoto Co., Inc.)], чтобы обеспечить не менее чем 2×107 клеток в день слияния клеток.
(d) Слияние клеток
Клетки, продуцирующие антитело, могут быть слиты с клетками миеломы, по мере необходимости, в соответствии с известными методами (Weir, D. M., Handbook of Experimental Immunology, Vol. I. II. III., Blackwell Scientific Publications, Oxford (1987), Kabat, E. A. and Mayer, M. M., Experimental Immunochemistry, Charles C Thomas Publisher Springfield, Illinois (1964), и т.д.) в условиях, когда нет чрезмерного снижения выживаемости клеток.
Примеры такого метода, который может быть использован в настоящем описании, включают химический метод, включающий смешивание клеток, продуцирующих антитела, с клетками миеломы в высококонцентрированном растворе полимера, такого как полиэтиленгликоль, и физический метод с использованием электрической стимуляции. Среди этих методов конкретный пример описанного выше химического метода заключается в следующем.
То есть, когда в качестве высококонцентрированного раствора полимера используют полиэтиленгликоль, то клетки, продуцирующие антитела, смешивают с клетками миеломы в растворе полиэтиленгликоля с молекулярной массой 1500-6000, предпочтительно 2000-4000, при температуре 30°С-40°С, предпочтительно 35°С-38°С, и в течение 1-10 минут, предпочтительно 5-8 минут.
(e) Отбор гибридомной группы
Способ отбора гибридом, полученных описанным выше слиянием клеток, ничем особо не ограничивается. В общем, применяется метод отбора с использованием HAT (гипоксантин-аминоптерин-тимидин) (Kohler et al., Nature (1975) 256, p. 495; Milstein et al., Nature (1977) 266, p. 550).
Этот метод эффективен, когда гибридомы получают с использованием миеломных клеток клеточной линии, дефицитной по HGPRT, которые не могут выжить в аминоптерине.
Конкретно, неслитые клетки и гибридомы культивируют в среде HAT, так что остаются и имеют возможность селективного роста только гибридомы, которые устойчивы к аминоптерину.
(f) Разделение на одноклеточные клоны (клонирование)
В качестве методов клонирования гибридом можно применять известные методы, такие как метод метилцеллюлозы, метод мягкой агарозы или метод лимитирующего разведения (см., например, Barbara, B. M. and Stanley, M. S.: Selected Methods in Cellular Immunology, W. H. Freeman and Company, San Francisco (1980)). Среди этих методов особенно предпочтительны методы трехмерного культивирования, такие как метод с использованием метилцеллюлозы. Например, гибридомную группу, образованную слиянием клеток, суспендируют в среде метилцеллюлозы, такой как селективная среда CloneCell-HY Selection Medium D (производства StemCell Technologies, № 03804), и затем культивируют. После этого образовавшиеся гибридомные колонии собирают, так что могут быть получены моноклональные гибридомы. Каждую из собранных колоний гибридом культивируют, и полученный супернатант гибридомной культуры, в котором наблюдается стабильный титр антител, отбирают в качестве гибридомного штамма, продуцирующего моноклональные антитела GARP.
Примеры созданного таким образом гибридомного штамма могут включать гибриды GARP 151D и 198D. В настоящем описании антитело, продуцируемое гибридами GARP 151D и 198D, обозначается как «антитело 151D» или «антитело 198D», или оно просто называется «151D» или «198D».
Вариабельная область тяжелой цепи антитела 151D имеет аминокислотную последовательность, представленную в SEQ ID NO: 15 в списке последовательностей. Кроме того, вариабельная область легкой цепи антитела 151D имеет аминокислотную последовательность, представленную в SEQ ID NO: 17 в списке последовательностей. Следует отметить, что аминокислотная последовательность вариабельной области тяжелой цепи, представленная в SEQ ID NO: 15 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 14 в списке последовательностей. Следует также отметить, что аминокислотная последовательность вариабельной области легкой цепи, представленная в SEQ ID NO: 17 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 16 в списке последовательностей.
Вариабельная область тяжелой цепи антитела 198D имеет аминокислотную последовательность, представленную в SEQ ID NO: 19 в списке последовательностей. Кроме того, вариабельная область легкой цепи антитела 198D имеет аминокислотную последовательность, представленную в SEQ ID NO: 21 в списке последовательностей. Следует отметить, что аминокислотная последовательность вариабельной области тяжелой цепи, представленная в SEQ ID NO: 19 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 18 в списке последовательностей. Следует также отметить, что аминокислотная последовательность вариабельной области легкой цепи, представленная в SEQ ID NO: 21 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 20 в списке последовательностей.
(g) Получение моноклонального антитела путем культивирования гибридом
Отобранные таким образом гибридомы культивируют, так что можно эффективно получать моноклональные антитела. Перед осуществлением культивирования целесообразно провести скрининг гибридом, которые продуцируют интересующее моноклональное антитело.
Для этого скрининга могут применяться известные методы.
Титр антител можно измерить в настоящем изобретении, например, методом ИФА, описанным в пункте (b) выше.
Гибридомы, полученные вышеуказанными способами, могут сохраняться в жидком азоте или в морозильной камере при температуре -80°С или ниже в замороженном состоянии.
После завершения клонирования гибридомы культивируют, заменяя при этом среду НТ на нормальную среду.
Массовое культивирование осуществляют с помощью ротационного культивирования или спин-культивирования, используя большую колбу для культивирования. Супернатант, полученный из этой массовой культуры, очищают в соответствии с методами, хорошо известными специалисту в данной области, такими как гель-фильтрация, с получением моноклонального антитела, специфично связывающегося с белком по настоящему изобретению.
Кроме того, гибридомы вводят в брюшную полость мыши той же линии (например, вышеописанный BALB/c) или мыши Nu/Nu, и гибридомам дают возможность расти в ней, чтобы получить асциты, содержащие большое количество моноклонального антитела по настоящему изобретению.
Когда гибридомы вводят в брюшную полость такой мыши, то может быть получено большее количество асцитов, если ранее мыши вводили минеральное масло, такое как 2,6,10,14-тетраметилпентадекан (пристан), (за 3-7 дней до введения гибридом).
Например, предположим, что иммуносупрессивный агент ранее ввели в брюшную полость мыши той же линии, что и гибридомы, так что Т-клетки дезактивированы. Через 20 дней после инъекции 106-107 гибридом и/или клональные клетки суспендируют в среде, не содержащей сыворотку (0,5 мл), и затем суспензию вводят в брюшную полость. Когда нормальная брюшная полость опухает и собираются асциты, то проводят забор асцитов у мыши. Согласно этому методу можно получить моноклональное антитело в концентрации, которая составляет примерно в 100 раз больше или более, чем в культуральном растворе.
Моноклональное антитело, полученное описанным выше методом, может быть очищено, например, методом, описанным в Weir, DM: Handbook of Experimental Immunology, Vol. I, II, III, Blackwell Scientific Publications, Oxford (1978).
Полученное таким образом моноклональное антитело обладает высокой антигенной специфичностью к GARP.
(h) Анализ моноклонального антитела
Изотип и подтип полученного моноклонального антитела можно определить следующим образом.
Во-первых, примеры метода анализа могут включать метод Оухтерлони, метод ИФА и метод РИА.
Метод Оухтерлони прост, но когда концентрация моноклонального антитела низкая, необходима процедура концентрирования.
С другой стороны, когда используют метод ИФА или метод РИА, супернатант культуры непосредственно реагирует с антигеном, адсорбированным на твердой подложке, а антитело, соответствующее различным изотипам или подклассам иммуноглобулина, используется в качестве вторичного антитела, так что можно идентифицировать изотип и подтип моноклонального антитела.
В качестве более простого метода можно также использовать коммерчески доступный набор для идентификации (например, Mouse Typer Kit, производства BioRad) и т.д..
Кроме того, количественное определение белка может быть осуществлено методом Фолина Лоури и методом вычисления значения на основе поглощения при 280 нм [1,4 (OD 280)=1 мг/мл иммуноглобулина].
Кроме того, также в случае, когда стадии (а)-(h) в вышеуказанном (2) повторяют и моноклональное антитело независимо получают отдельно, причем антитело имеет свойства, эквивалентные свойствам антитела 105F, антитела 110F, антитела, являющегося производным 151D (гуманизированное антитело 151D), и антитела, являющегося производным 198D (гуманизированное антитело 198D). Примером такого антитела может быть антитело, связывающееся с тем же эпитопом, с которым связывается каждое из вышеописанных антител. Антитело 105F распознает области аминокислотной последовательности в положениях аминокислот 366-3377, 407-444 и 456-470 в аминокислотной последовательности (SEQ ID NO: 1) GARP и связывается с ними; антитело 110F распознает области аминокислотной последовательности в положениях аминокислот 54-112 и 366-392 в аминокислотной последовательности (SEQ ID NO: 1) GARP и связывается с ними; антитело, являющееся производным 151D (гуманизированное антитело 151D), распознает аминокислотную последовательность в положениях аминокислот 352-339 в аминокислотной последовательности (SEQ ID NO: 1) GARP и связывается с ней; и антитело, являющееся производным 198D (гуманизированное антитело 198D), распознает аминокислотную последовательность в положениях аминокислот 18-112 в аминокислотной последовательности (SEQ ID NO: 1) GARP и связывается с ней. Соответственно, конкретные примеры вышеуказанного эпитопа могут включать вышеуказанные области в аминокислотной последовательности GARP.
Если вновь полученное моноклональное антитело связывается с неполным пептидом или с неполной трехмерной структурой, с которой связывается вышеописанное антитело 105F и т.д., можно определить, что моноклональное антитело связывается с тем же эпитопом, с которым связывается вышеописанное антитело 105F и т.д.. Кроме того, подтверждая, что моноклональное антитело конкурирует с вышеописанными антителами, такими как антитело 105F, при связывании антител с GARP (т.е. моноклональное антитело препятствует связыванию вышеописанных антител, таких как антитело 105F, с GARP), можно определить, что моноклональное антитело связывается с одним и тем же эпитопом, с которым связывается вышеописанное антитело 105F и т.д., даже если не определена специфичная последовательность или структура эпитопа. Когда подтверждается, что моноклональное антитело связывается с тем же эпитопом, с которым связывается антитело 105F и т.д., то ожидается, что моноклональное антитело должно обладать свойствами, эквивалентными свойствам вышеописанных антител, таких как антитело 105F.
(3) Другие антитела
Антитело по настоящему изобретению также включает генетически рекомбинантные антитела, которые были искусственно модифицированы с целью уменьшения гетерогенной антигенности для человека, такие как химерное антитело, гуманизированное антитело и вышеописанные человеческие антитела, а также вышеописанное моноклональное антитело против GARP. Эти антитела могут быть получены известными методами.
Полученное антитело может быть очищено до гомогенного состояния. Для разделения и очистки антитела могут быть использованы методы разделения и очистки, используемые для обычных белков. Например, колоночная хроматография, фильтрация, ультрафильтрация, высаливание, диализ, препаративный полиакриламидный гель-электрофорез, изоэлектрическое фокусирование и т.д., соответственно, отбираются и объединяются друг с другом, так что антитело можно отделить и очистить (Strategies for Protein Purification and Characterization: A Laboratory Course Manual, Daniel R. Marshak et al. eds., Cold Spring Harbor Laboratory Press (1996); Antibodies: A Laboratory Manual. Ed Harlow and David Lane, Cold Spring Harbor Laboratory (1988)), но примеры методов разделения и очистки не ограничиваются этим.
Примеры хроматографии могут включать аффинную хроматографию, ионообменную хроматографию, гидрофобную хроматографию, гель-фильтрационную хроматографию, хроматографию с обращенной фазой и абсорбционную хроматографию.
Эти хроматографические методы могут быть осуществлены с использованием жидкостной хроматографии, такой как ВЭЖХ или FPLC (жидкостная хроматография быстрого действия).
Примеры колонки, используемой в аффинной хроматографии, могут включать колонку с Протеином А и колонку с Протеином G. Примеры колонки, включающей использование Протеина A, могут включать Hyper D, POROS и Sepharose FF (Pharmacia).
Кроме того, с использованием носителя с иммобилизованным антигеном, антитело можно очистить, используя активность связывания антитела с антигеном.
Полученные антитела оценивают с точки зрения их активности связывания с антигеном в соответствии с методом, описанным в примерах, обсуждаемых ниже, и т.д., так что может быть выбрано предпочтительное антитело.
Стабильность антитела может быть использована в качестве индикатора для сравнения свойств антител. Калориметр дифференциальной сканирующей калориметрии (DSC) представляет собой устройство, способное быстро и точно измерить среднюю точку тепловой денатурации (Tm), что является хорошим показателем относительной структурной стабильности белка. Используя DSC для измерения значений Tm и проводя сравнение относительно полученных значений, можно сравнить различия в термостабильности. Известно, что стабильность сохранения антитела имеет определенную корреляцию с термостабильностью антитела (Lori Burton, et al., Pharmaceutical Development and Technology (2007) 12, pp. 265-273), и, таким образом, предпочтительное антитело может быть выбрано с использованием термостабильности в качестве индикатора. Примеры других показателей для выбора антитела могут включать высокий выход в подходящих клетках-хозяевах и низкую агглютинацию в водном растворе. Например, поскольку антитело с самым высоким выходом не всегда демонстрирует самую высокую термостабильность, необходимо выбрать наиболее подходящее для введения человеку антитело путем его всестороннего исследования на основе вышеуказанных показателей.
Примером человеческого антитела против GARP по настоящему изобретению может быть человеческое антитело против GARP, полученное вышеописанным способом фагового дисплея, и предпочтительные примеры человеческого антитела против GARP по настоящему изобретению могут включать антитело 105F и антитело 110F, каждое из которых имеет следующую структуру.
Тяжелая цепь антитела 105F имеет аминокислотную последовательность, представленную в SEQ ID NO: 2 в списке последовательностей. В аминокислотной последовательности тяжелой цепи, представленной в SEQ ID NO: 2 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-118, представляет собой вариабельную область, тогда как аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 119-448, представляет собой константную область. В SEQ ID NO: 2 в списке последовательностей эта вариабельная область содержит CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 26-35, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 50-66, и CDRH 3, состоящую из аминокислотной последовательности в положениях аминокислот 99-107. Кроме того, последовательность SEQ ID NO: 2 представлена на Фигуре 2.
Легкая цепь антитела 105F имеет аминокислотную последовательность, представленную в SEQ ID NO: 3 в списке последовательностей. В аминокислотной последовательности легкой цепи, представленной в SEQ ID NO: 3 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-112, представляет собой вариабельную область, тогда как аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 113-217, представляет собой константную область. В SEQ ID NO: 3 в списке последовательностей эта вариабельная область содержит CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 23-36, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 52-58, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 91-101. Кроме того, последовательность SEQ ID NO: 3 представлена на Фигуре 3.
Тяжелая цепь антитела 110F имеет аминокислотную последовательность, представленную в SEQ ID NO: 4 в списке последовательностей. В аминокислотной последовательности тяжелой цепи, представленной в SEQ ID NO: 4 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-123, представляет собой вариабельную область, тогда как аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 124-453, представляет собой константную область. В SEQ ID NO: 4 в списке последовательностей эта вариабельная область содержит CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 26-35, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 50-66, и CDRH 3, состоящую из аминокислотной последовательности в положениях аминокислот 99-112. Кроме того, последовательность SEQ ID NO: 4 представлена на Фигуре 4.
Легкая цепь антитела 110F имеет аминокислотную последовательность, представленную в SEQ ID NO: 5 в списке последовательностей. В аминокислотной последовательности легкой цепи, представленной в SEQ ID NO: 5 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-111, представляет собой вариабельную область, тогда как аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 112-216 представляет собой константную область. В SEQ ID NO: 5 в списке последовательностей эта вариабельная область содержит CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 23-36, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 52-58, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 91-100. Кроме того, последовательность SEQ ID NO: 5 представлена на Фигуре 5.
Аминокислотная последовательность тяжелой цепи антитела 105F, представленная в SEQ ID NO: 2 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 6 в списке последовательностей. Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-354 в нуклеотидной последовательности, представленной в SEQ ID NO: 6 в списке последовательностей, кодирует вариабельную область тяжелой цепи антитела 105F, а нуклеотидная последовательность, состоящая из нуклеотидов в положениях 355-1334 кодирует константную область тяжелой цепи антитела 105F. Как представлено в SEQ ID NO: 6, нуклеотидная последовательность, кодирующая вариабельную область, содержит полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 76-105, кодирующий CDRH1, полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 148-198, кодирующий CDRH2, и полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 295-321, кодирующий CDRH3. Кроме того, последовательность SEQ ID NO: 6 представлена на Фигуре 6.
Аминокислотная последовательность легкой цепи антитела 105F, представленная в SEQ ID NO: 3 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 7 в списке последовательностей. Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-336 в нуклеотидной последовательности, представленной в SEQ ID NO: 7 в списке последовательностей, кодирует вариабельную область легкой цепи антитела 105F, и нуклеотидная последовательность, состоящая из нуклеотидов в положениях 337-651, кодирует константную область легкой цепи антитела 105F. Как представлено в SEQ ID NO: 7, нуклеотидная последовательность, кодирующая вариабельную область, содержит полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 67-108, кодирующий CDRL1, полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 154-174, кодирующий CDRL2, и полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 271-303, кодирующий CDRL3. Кроме того, последовательность SEQ ID NO: 7 представлена на Фигуре 7.
Аминокислотная последовательность тяжелой цепи антитела 110F, представленная в SEQ ID NO: 4 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 8 в списке последовательностей. Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-369 в нуклеотидной последовательности, представленной в SEQ ID NO: 8 в списке последовательностей, кодирует вариабельную область тяжелой цепи антитела 110F, и нуклеотидная последовательность, состоящая из нуклеотидов в положениях 370-1359, кодирует константную область тяжелой цепи антитела 110F. Как представлено в SEQ ID NO: 8, нуклеотидная последовательность, кодирующая вариабельную область, содержит полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 76-105, кодирующий CDRH1, полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 148-198, кодирующий CDRH2, и полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 295-336, кодирующий CDRH3. Кроме того, последовательность SEQ ID NO: 8 представлена на Фигуре 8.
Аминокислотная последовательность легкой цепи антитела 110F, представленная в SEQ ID NO: 5 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 9 в списке последовательностей. Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-333 в нуклеотидной последовательности, представленной в SEQ ID NO: 9 в списке последовательностей, кодирует вариабельную область легкой цепи антитела 110F, а нуклеотидная последовательность, состоящая из нуклеотидов в положениях 334-648, кодирует константную область легкой цепи антитела 110F. Как представлено в SEQ ID NO: 9, нуклеотидная последовательность, кодирующая вариабельную область, содержит полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 67-108, кодирующий CDRL1, полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 154-174, кодирующий CDRL2, и полинуклеотид, состоящий из нуклеотидной последовательности в положениях нуклеотидов 271-300, кодирующий CDRL3. Кроме того, последовательность SEQ ID NO: 9 представлена на Фигуре 9.
Что касается антитела по настоящему изобретению, в дополнение к вышеописанному человеческому антителу против GARP, то даже в случае, когда антитело получают независимо, отдельно, в соответствии с методом, отличным от вышеописанного метода получения антител, могут быть получены антитела, имеющие цитотоксичность, эквивалентную цитотоксичности антитела 105F или антитела 110F. Примером такого антитела может быть антитело, связывающееся с тем же эпитопом, с которым связывается антитело 105F или антитело 110F.
Если вновь полученное человеческое антитело связывается с неполным пептидом или с неполной трехмерной структурой, с которой связывается антитело 105F или антитело 110F, можно определить, что полученное антитело связывается с тем же эпитопом, с которым связывается антитело 105F или антитело 110F. Кроме того, подтверждая, что соответствующее антитело конкурирует с антителом 105F или антителом 110F за его связывание с GARP (т.е. соответствующее антитело препятствует связыванию антитела 105F или антитела 110F с GARP), можно определить, что соответствующее антитело связывается с тем же эпитопом, с которым связывается антитело 105F или антитело 110F, даже если специфичная последовательность или структура эпитопа не определена. Если подтверждено, что соответствующее антитело связывается с тем же эпитопом, с которым связывается антитело 105F или антитело 110F, то ожидается, что соответствующее антитело должно иметь цитотоксичность, эквивалентную цитотоксичности антитела 105F или антитела 110F.
Кроме того, антитело по настоящему изобретению включает искусственно модифицированные генетически рекомбинантные антитела. Эти антитела могут быть получены с использованием известных методов. Интересующее антитело предпочтительно представляет собой антитело, содержащее, по меньшей мере, те же самые 6 CDR, что и тяжелая цепь и легкая цепь вышеописанного антитела 105F или 110F, а также имеющее активность ADCC и ингибирующую активность иммуносупрессивной функции Treg. Интересующее антитело не ограничивается конкретным антителом, если оно обладает вышеуказанными свойствами. Более предпочтительно, антитело представляет собой антитело, имеющее вариабельную область тяжелой цепи и вариабельную область легкой цепи вышеописанного антитела 105F или антитела 110F.
Кроме того, путем объединения последовательностей, демонстрирующих высокую гомологию с аминокислотной последовательностью тяжелой цепи и аминокислотной последовательностью легкой цепи антитела 105F или антитела 110F, друг с другом, можно выбрать антитело, обладающее активностью, эквивалентной активности вышеописанного антитела. Такая гомология представляет собой гомологию, составляющую, как правило, 80% или более, предпочтительно 90% или более, более предпочтительно, 95% или более и, наиболее предпочтительно, 99% или более (однако каждая CDR идентична соответствующей области для каждого из вышеописанных антител). Кроме того, также возможно выбрать антитело, обладающее активностью, эквивалентной активности каждого из вышеописанных антител, путем включения аминокислотной последовательности, содержащей замену, делецию или вставку одного или нескольких аминокислотных остатков, в аминокислотную последовательность вышеописанных тяжелой цепи или легкой цепи (исключая каждый сайт CDR).
Кроме того, примеры антител против GARP согласно настоящему изобретению могут включать следующие химерные антитела и гуманизированные антитела.
Пример химерного антитела может включать антитела, в которых вариабельная область и константная область являются гетерологичными друг другу, такие как химерное антитело, образованное путем конъюгации вариабельной области антитела мыши или крысы с константной областью человека (см. Proc. Natl.Acad. Sci. U.S.A., 81, 6851-6855, (1984)).
Химерное антитело, полученное из крысиного антитела 151D против человеческого GARP, представляет собой антитело, состоящее из тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 1-117, представленной в SEQ ID NO: 15, и легкой цепи, содержащей вариабельную область легкой цепи, состоящей из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 1-109, представленной в SEQ ID NO: 17, и это химерное антитело может содержать константную область, полученную от любого данного человека.
Кроме того, химерное антитело, полученное из крысиного антитела 198D против человеческого GARP, представляет собой антитело, состоящее из тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящей из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 1-120, представленной в SEQ ID NO: 19, и легкой цепи, содержащей вариабельную область легкой цепи, состоящей из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 1-109, представленной в SEQ ID NO: 21, и это химерное антитело может содержать константную область, полученную от любого данного человека.
Примеры такого химерного антитела могут включать: антитело, состоящее из тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-446, представленную в SEQ ID NO: 25 в списке последовательностей, и легкой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 21-234, представленную в SEQ ID NO: 27; и антитело, состоящее из тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-449, представленную в SEQ ID NO: 29 в списке последовательностей, и легкой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 21-234, представленную в SEQ ID NO: 31.
Следует отметить, что в последовательности тяжелой цепи, представленной в SEQ ID NO: 25 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-19, представляет собой сигнальную последовательность, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 20-136, представляет собой вариабельную область, и аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 137-446, представляет собой константную область.
Следует также отметить, что в последовательности легкой цепи, представленной в SEQ ID NO: 27 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-20, представляет собой сигнальную последовательность, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 21-129, представляет собой вариабельную область, а аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 130-234, представляет собой константную область.
Кроме того, следует отметить, что в последовательности тяжелой цепи, представленной в SEQ ID NO: 29 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-19, представляет собой сигнальную последовательность, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 20-139, представляет собой вариабельную область, и аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 140-469, представляет собой константную область.
Следует также отметить, что в последовательности легкой цепи, представленной в SEQ ID NO: 31 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-20, представляет собой сигнальную последовательность, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 21-129, представляет собой вариабельную область, а аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 130-234, представляет собой константную область.
Аминокислотная последовательность тяжелой цепи антитела c151D, представленная в SEQ ID NO: 25 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 24 в списке последовательностей. Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-55 в нуклеотидной последовательности, представленной в SEQ ID NO: 24 в списке последовательностей, кодирует сигнальную последовательность тяжелой цепи антитела c151D, нуклеотидная последовательность, состоящая из нуклеотидов в положениях 58-408 в ней, кодирует вариабельную область тяжелой цепи антитела c151D, а нуклеотидная последовательность, состоящая из нуклеотидов в положениях 409-1398, кодирует константную область тяжелой цепи антитела c151D.
Кроме того, аминокислотная последовательность легкой цепи антитела c151D, представленная в SEQ ID NO: 27 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 26 в списке последовательностей. Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-60 в нуклеотидной последовательности, представленной в SEQ ID NO: 26 в списке последовательностей, кодирует сигнальную последовательность легкой цепи антитела c151D, нуклеотидная последовательность, состоящая из нуклеотидов в положениях 61-387 в ней, кодирует вариабельную область легкой цепи антитела c151D, а нуклеотидная последовательность, состоящая из нуклеотидов в положениях 388-702, кодирует константную область легкой цепи антитела c151D.
Кроме того, аминокислотная последовательность тяжелой цепи антитела c198D, представленная в SEQ ID NO: 29 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 28 в списке последовательностей. Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-55 в нуклеотидной последовательности, представленной в SEQ ID NO: 28 в списке последовательностей, кодирует сигнальную последовательность тяжелой цепи антитела c198D, нуклеотидная последовательность, состоящая из нуклеотидов в положениях 58-417 в ней, кодирует вариабельную область тяжелой цепи антитела c198D, а нуклеотидная последовательность, состоящая из нуклеотидов в положениях 418-1407, кодирует константную область тяжелой цепи антитела c198D.
Кроме того, аминокислотная последовательность легкой цепи антитела c198D, представленная в SEQ ID NO: 31 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 30 в списке последовательностей. Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-60 в нуклеотидной последовательности, представленной в SEQ ID NO: 30 в списке последовательностей, кодирует сигнальную последовательность легкой цепи антитела c198D, нуклеотидная последовательность, состоящая из нуклеотидов в положениях 61-387 в ней, кодирует вариабельную область легкой цепи антитела c198D, а нуклеотидная последовательность, состоящая из нуклеотидов в положениях 388-702, кодирует константную область легкой цепи антитела c198D.
Примеры гуманизированного антитела включают гуманизированное антитело, образованное путем включения только области, определяющей комплементарность (CDR), в человеческое антитело (см. Nature (1986) 321, p. 522-525), и гуманизированное антитело, образованное путем трансплантации аминокислотных остатков в некоторые каркасные области, а также последовательности CDR в человеческое антитело в соответствии с методом трансплантации CDR (международная публикация № WO90/07861).
Однако гуманизированное антитело, являющееся производным антитела 151D, не ограничивается конкретным гуманизированным антителом, при условии, что оно сохраняет все 6 CDR-последовательностей антитела 151D и обладает противоопухолевой активностью.
Следует отметить, что вариабельная область тяжелой цепи антитела 151D содержит CDRH1 (GFTFSNYYMA), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 26-35 в SEQ ID NO: 15 в списке последовательностей, CDRH2 (SIGTVGGNTY), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 50-59 в SEQ ID NO: 15, и CDRH3 (EDYGGFPH), состоящей из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 99-106 в SEQ ID NO: 15.
Кроме того, вариабельная область легкой цепи антитела 151D содержит CDRL1 (KASQNVGTNVD), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 24-34 в SEQ ID NO: 17 в списке последовательностей, CDRL2 (GASNRYT), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 50-56 в SEQ ID NO: 17, и CDRL3 (LQYKYNPYT), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 89-97 в SEQ ID NO: 17.
Кроме того, вариабельная область тяжелой цепи антитела 198D содержит CDRH1 (GFSLTSFHVS), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 26-35 в SEQ ID NO: 19 в списке последовательностей, CDRH2 (TISSGGGTY), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 50-58 в SEQ ID NO: 19, и CDRH3 (ISGWGHYYVMDV), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 98-109 в SEQ ID NO: 19.
Кроме того, вариабельная область легкой цепи антитела 198D содержит CDRL1 (QASEDIYSGLA), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 24-34 в SEQ ID NO: 21 в списке последовательностей, CDRL2 (GAGSLQD), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 50-56 в SEQ ID NO: 21, и CDRL3 (QQGLKFPLT), состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 89-97 в SEQ ID NO: 21.
Конкретным примером гуманизированного антитела крысиного антитела 151D может быть любая данная комбинация: тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящей из любой из (1) аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 20-136, представленной в SEQ ID NO: 33 (h151D-H1) или 35 (h151D-H4) в списке последовательностей, (2) аминокислотной последовательности, имеющей гомологию, составляющую по меньшей мере 95% или более, с последовательностью каркасной области, отличной от каждой последовательности CDR в вышеуказанной последовательности (1), и (3) аминокислотной последовательности, содержащей делецию, замену или вставку одной или нескольких аминокислот в последовательность каркасной области, отличной от каждой последовательности CDR в вышеуказанной последовательности (1); и легкой цепи, содержащей вариабельную область легкой цепи, состоящую из любой из (4) аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 21-129, представленной в SEQ ID NO: 37 (h151D-L1) или 39 (h151D- L4), (5) аминокислотной последовательности, имеющую гомологию, составляющую по меньшей мере 95% или более, с последовательностью каркасной области, отличной от каждой последовательности CDR, в вышеуказанной последовательности (4), и (6) аминокислотной последовательности, содержащей делецию, замену или вставку одной или нескольких аминокислот в последовательности каркасной области, отличной от каждой последовательности CDR, в вышеуказанной последовательности (4).
С другой стороны, конкретным примером гуманизированного антитела крысиного антитела 198D может быть любая данная комбинация: тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящей из любой из (1) аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 20-139, представленной в SEQ ID NO: 41 (h198D-H3) в списке последовательностей, (2) аминокислотной последовательности, имеющей гомологию, составляющую по меньшей мере 95% или более, с последовательностью каркасной области, отличной от каждой последовательности CDR в вышеуказанной последовательности (1), и (3) аминокислотной последовательности, содержащей делецию, замену или вставку одной или нескольких аминокислот в последовательности каркасной области, отличной от каждой последовательности CDR в вышеуказанной последовательности (1); и легкой цепи, содержащей вариабельную область легкой цепи, состоящей из любой из (4) аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 21-129, представленной в SEQ ID NO: 43 (h198D-L4) в списке последовательностей, (5) аминокислотной последовательности, имеющей гомологию, составляющую по меньшей мере 95% или более, с последовательностью каркасной области, отличной от каждой последовательности CDR в вышеуказанной последовательности (4), и (6) аминокислотной последовательности, содержащей делецию, замену или вставку одной или нескольких аминокислот в последовательности каркасной области, отличной от каждой последовательности CDR, в вышеуказанной последовательности (4).
Примеры предпочтительной комбинации тяжелой цепи и легкой цепи гуманизированного антитела 151D могут включать: антитело, состоящее из тяжелой цепи, имеющей вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 20-136, представленной в SEQ ID NO: 33, и легкой цепи, имеющей вариабельную область легкой цепи, состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 21-129, представленной в SEQ ID NO: 37; и антитело, состоящее из тяжелой цепи, имеющей вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 20-36, представленной в SEQ ID NO: 35, и легкой цепи, имеющей вариабельную область легкой цепи, состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 21-129, представленной в SEQ ID NO: 39.
Примеры их более предпочтительной комбинации могут включать: антитело (h151D-H1L1), состоящее из тяжелой цепи, имеющей аминокислотную последовательность, представленную в SEQ ID NO: 33, и легкой цепи, имеющей аминокислотную последовательность, представленную в SEQ ID NO: 37; и антитело (h151D-H4L4), состоящее из тяжелой цепи, имеющей аминокислотную последовательность, представленную в SEQ ID NO: 35, и легкой цепи, имеющей аминокислотную последовательность, представленную в SEQ ID NO: 39.
Примером предпочтительной комбинации тяжелой цепи и легкой цепи гуманизированного антитела 198D может быть антитело, состоящее из тяжелой цепи, имеющей вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 20-139, представленной в SEQ ID NO: 41, и легкой цепи, имеющей вариабельную область легкой цепи, состоящую из аминокислотной последовательности, состоящей из аминокислотных остатков в положениях 21-129, представленной в SEQ ID NO: 43.
Примером их более предпочтительной комбинации может быть антитело (h198D-H3L4), состоящее из тяжелой цепи, имеющей аминокислотную последовательность, представленную в SEQ ID NO: 41, и легкой цепи, имеющей аминокислотную последовательность, представленную в SEQ ID NO: 43.
Объединив вместе последовательности, демонстрирующие высокую гомологию с вышеописанными аминокислотными последовательностями тяжелой цепи и аминокислотными последовательностями легкой цепи, можно выбрать антитело, имеющее цитотоксичность, эквивалентную цитотоксичности каждого из вышеописанных антител. Такая гомология представляет собой гомологию, составляющую, как правило, 80% или более, предпочтительно 90% или более, более предпочтительно 95% или более и наиболее предпочтительно 99% или более. Кроме того, также объединив друг с другом аминокислотные последовательности, содержащие замену, делецию или вставку одного или нескольких аминокислотных остатков по отношению к аминокислотной последовательности тяжелой цепи или легкой цепи, можно выбрать антитело, имеющее цитотоксичность, эквивалентную цитотоксичности каждого из вышеописанных антител.
Следует отметить, что термин «несколько» используется в настоящем описании для обозначения 1-10, 1-9, 1-8, 1-7, 1-6, 1-5, 1-4, 1-3, или 1 или 2.
Аминокислотная замена в настоящем описании предпочтительно представляет собой консервативную аминокислотную замену. Консервативная аминокислотная замена представляет собой замену, происходящую в аминокислотной группе, связанной с определенными боковыми цепями аминокислот. Предпочтительными аминокислотными группами являются следующие: кислотная группа=аспарагиновая кислота и глутаминовая кислота; основная группа=лизин, аргинин и гистидин; неполярная группа=аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин и триптофан; и незаряженная полярная групп=глицин, аспарагин, глутамин, цистеин, серин, треонин и тирозин. Другими предпочтительными аминокислотными группами являются следующие: алифатическая гидроксигруппа=серин и треонин; амидосодержащая группа=аспарагин и глутамин; алифатическая группа=аланин, валин, лейцин и изолейцин; и ароматическая группа=фенилаланин, триптофан и тирозин. Такую аминокислотную замену предпочтительно осуществляют так, чтобы свойства вещества, имеющего исходную аминокислотную последовательность, не ухудшались.
Гомология между двумя типами аминокислотных последовательностей может быть определена с использованием параметров по умолчанию алгоритма Blast версии 2.2.2 (Altschul, Stephen F., Thomas L. Madden, Alejandro A. Schaffer, Jinghui Zhang, Zheng Zhang, Webb Miller, and David J. Lipman (1997), «Gapped BLAST and PSI-BLAST: a new generation of protein database search programs,» Nucleic Acids Res. 25: 3389-3402). Алгоритм Blast можно также использовать, обратившись к www.ncbi.nlm.nih.gov/blast через Интернет. Следует отметить, что гомология между нуклеотидной последовательностью антитела по настоящему изобретению и нуклеотидной последовательностью другого антитела также может быть определена с использованием алгоритма Blast.
В аминокислотной последовательности тяжелой цепи гуманизированного антитела 151D, представленной в SEQ ID NO: 33 или 35 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-19, представляет собой сигнальную последовательность, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 20-136, представляет собой вариабельную область, и аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 137-446, представляет собой константную область.
Кроме того, в аминокислотной последовательности легкой цепи гуманизированного антитела 151D, представленной в SEQ ID NO: 37 или 39 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-20, представляет собой сигнальную последовательность, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 21-129, представляет собой вариабельную область, и аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 130-234, представляет собой константную область.
Кроме того, в аминокислотной последовательности тяжелой цепи гуманизированного антитела 198D, представленной в SEQ ID NO: 41 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-19, представляет собой сигнальную последовательность, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 20-139, представляет собой вариабельную область, и аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 140-469, представляет собой константную область.
Кроме того, в аминокислотной последовательности легкой цепи гуманизированного антитела 198D, представленной в SEQ ID NO: 43 в списке последовательностей, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-20, представляет собой сигнальную последовательность, аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 21-129, представляет собой вариабельную область, и аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 130-234, представляет собой константную область.
Аминокислотная последовательность тяжелой цепи гуманизированного антитела 151D, представленная в SEQ ID NO: 33 или 35 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 32 или 34 в списке последовательностей, соответственно. Кроме того, последовательность SEQ ID NO: 33 представлена на Фигуре 21, последовательность SEQ ID NO: 35 представлена на Фигуре 23, последовательность SEQ ID NO: 32 представлена на Фигуре 31, и последовательность SEQ ID NO: ID NO: 34 представлена на Фигуре 33, соответственно.
Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-57, в каждой нуклеотидной последовательности кодирует сигнальную последовательность тяжелой цепи гуманизированного антитела 151D, нуклеотидная последовательность, состоящая из нуклеотидов в положениях 58-408, кодирует вариабельную область тяжелой цепи гуманизированного антитела 151D, и нуклеотидная последовательность, состоящая из нуклеотидов в положениях 409-1398, кодирует константную область тяжелой цепи гуманизированного антитела 151D.
Аминокислотная последовательность тяжелой цепи гуманизированного антитела 198D, представленная в SEQ ID NO: 41 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 40 в списке последовательностей. Кроме того, последовательность SEQ ID NO: 41 представлена на Фигуре 25, а последовательность SEQ ID NO: 40 представлена на Фигуре 35.
Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-55 в вышеуказанной нуклеотидной последовательности, кодирует сигнальную последовательность тяжелой цепи гуманизированного антитела 198D, нуклеотидная последовательность, состоящая из нуклеотидов в положениях 58-417, кодирует вариабельную область тяжелой цепи гуманизированного антитела 198D, а нуклеотидная последовательность, состоящая из нуклеотидов в положениях 418-1407, кодирует константную область тяжелой цепи гуманизированного антитела 198D.
Аминокислотная последовательность легкой цепи гуманизированного антитела 151D, представленная в SEQ ID NO: 37 или 39 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 36 или 38 в списке последовательностей, соответственно. Кроме того, последовательность SEQ ID NO: 37 представлена на Фигуре 22, последовательность SEQ ID NO: 39 представлена на Фигуре 24, последовательность SEQ ID NO: 36 представлена на Фигуре 32, а последовательность SEQ ID NO: ID NO: 38 представлена на Фигуре 34, соответственно.
Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-60, в каждой нуклеотидной последовательности кодирует сигнальную последовательность легкой цепи гуманизированного антитела 151D, нуклеотидная последовательность, состоящая из нуклеотидов в положениях 61-387, кодирует вариабельную область легкой цепи гуманизированного антитела 151D, и нуклеотидная последовательность, состоящая из нуклеотидов в положениях 388-702, кодирует константную область легкой цепи гуманизированного антитела 151D.
Аминокислотная последовательность легкой цепи гуманизированного антитела 198D, представленная в SEQ ID NO: 43 в списке последовательностей, кодируется нуклеотидной последовательностью, представленной в SEQ ID NO: 42 в списке последовательностей. Кроме того, последовательность SEQ ID NO: 43 представлена на Фигуре 26, а последовательность SEQ ID NO: 42 представлена на Фигуре 36.
Нуклеотидная последовательность, состоящая из нуклеотидов в положениях 1-60 в вышеуказанной нуклеотидной последовательности, кодирует сигнальную последовательность легкой цепи гуманизированного антитела 198D, нуклеотидная последовательность, состоящая из нуклеотидов в положениях 61-387, кодирует вариабельную область легкой цепи гуманизированного антитела 198D, а нуклеотидная последовательность, состоящая из нуклеотидов в положениях 388-702, кодирует константную область легкой цепи гуманизированного антитела 198D.
Гомология между этими нуклеотидными последовательностями и нуклеотидными последовательностями других антител также может быть определена с использованием алгоритма Blast.
Еще одним примером антитела по настоящему изобретению может быть человеческое антитело, связывающееся с тем же эпитопом, с которым также связывается гуманизированное антитело 151D или гуманизированное антитело 198D. Антитело против GARP человека означает человеческое антитело, имеющее последовательность генов антитела, полученную только из хромосом человека. Антитело против GARP человека можно получить с помощью вышеуказанного метода.
Если вновь полученное человеческое антитело связывается с неполным пептидом или с неполной трехмерной структурой, с которой связывается гуманизированное антитело 151D или гуманизированное антитело 198D, то можно определить, что человеческое антитело связывается с тем же эпитопом, с которым связывается гуманизированное антитело 151D или гуманизированное антитело 198D. Кроме того, подтверждая, что человеческое антитело конкурирует с гуманизированным антителом 151D или гуманизированным антителом 198D за его связывания с GARP (т.е. человеческое антитело препятствует связыванию гуманизированного антитела 151D или гуманизированного антитела 198D с GARP), можно определить, что человеческое антитело связывается с тем же эпитопом, с которым связывается гуманизированное антитело 151D или гуманизированное антитело 198D, даже если не определена конкретная последовательность или структура эпитопа. Если подтверждено, что интересующее человеческое антитело связывается с тем же эпитопом, с которым связывается гуманизированное антитело 151D или гуманизированное антитело 198D, то ожидается, что человеческое антитело должно иметь цитотоксичность, эквивалентную цитотоксичности гуманизированного антитела 151D или гуманизированного антитела 198D.
Химерное антитело, гуманизированное антитело или человеческое антитело, полученное вышеуказанными методами, оценивают с помощью методов, описанных далее в примерах и т.д., в отношении активности связывания с антигеном, и, таким образом, может быть выбрано предпочтительное антитело.
Настоящее изобретение также включает модификацию антитела. Термин «модификация» используется в настоящем описании для обозначения антитела по настоящему изобретению, которое химически или биологически модифицировано. Примеры такой химической модификации включают связывание химического компонента с аминокислотным скелетом и химическую модификацию N-связанной или O-связанной углеводной цепи. Примеры такой биологической модификации включают антитела, которые подверглись посттрансляционной модификации (например, N-связанную или O-связанную модификацию сахарной цепи, N-концевое или C-концевое процессирование, дезамидирование, изомеризацию аспарагиновой кислоты и окисление метионина), и антитела, к N-концу которых добавляется остаток метионина, в результате чего появляется возможность экспрессии с использованием прокариотических клеток-хозяев. Кроме того, такая модификация также включает меченые антитела для обеспечения возможности обнаружения или выделения антитела по настоящему изобретению или антигена, такого как, например, меченое ферментом антитело, флуоресцентно меченое антитело и антитело, меченное аффинной меткой. Такая модификация антитела по настоящему изобретению применима для улучшения стабильности и удерживания в крови исходного антитела по настоящему изобретению, снижения антигенности, детектирования или выделения такого антитела или антигена и т.д..
Кроме того, регулируя модификацию сахарной цепи (гликозилирование, дефукозилирование и т.д.), которая связывается с антителом по настоящему изобретению, можно повысить антителозависимую клеточную цитотоксичность. В качестве способов регуляции модификации сахарной цепи антитела известны способы, описанные в WO 99/54342, WO 2000/61739, WO2002/31140 и т.д., но не ограничиваясь ими. Антитело по настоящему изобретению также включает антитела, в которых регулируется вышеупомянутая модификация сахарной цепи.
После выделения гена антитела, ген вводят в подходящий хозяин для получения антитела с использованием подходящей комбинации хозяина и экспрессирующего вектора. Конкретным примером гена антитела может быть комбинация гена, кодирующего последовательность тяжелой цепи антитела, описанного в настоящем описании, и гена, кодирующего последовательность легкой цепи описанного антитела. При трансформации клеток-хозяев ген последовательности тяжелой цепи и ген последовательности легкой цепи могут быть вставлены в один экспрессирующий вектор, или, вместо этого, эти гены могут быть вставлены в различные экспрессирующие векторы.
Когда в качестве хозяев используют эукариотические клетки, могут использоваться животные клетки, растительные клетки или эукариотические микроорганизмы. Примеры животных клеток включают клетки млекопитающих, такие как клетки COS, которые являются клетками обезьян (Gluzman, Y., Cell (1981) 23, стр. 175-182, ATCC CRL-1650), мышиные фибробласты NIH3T3 (ATCC No. CRL-1658), и клеточную линию клеток яичника китайского хомячка, дефицитную по дигидрофолатредуктазе (клетки СНО, АТСС CCL-61) (Urlaub, G. and Chasin, LA Proc. Natl. Acad. Sci. USA (1980) 77, p. 4126-4220).
Когда в качестве хозяев используют прокариотические клетки, можно использовать Escherichia coli или Bacillus subtilis.
Интересующий ген антитела вводят в эти клетки для трансформации, и трансформированные клетки затем культивируют in vitro для получения антитела. В вышеуказанной культуре существуют случаи, когда выход различается в зависимости от последовательности антитела, и, следовательно, можно выбрать антитело, которое легко получить в виде лекарственного средства из антител, обладающих эквивалентной активностью связывания, с использованием выхода в качестве индикатора. Соответственно, антитело по настоящему изобретению также включает антитело, полученное описанным выше методом получения антител, который отличается тем, что он включает стадию культивирования трансформированных клеток-хозяев и стадию сбора интересующего антитела из культуры, полученной на вышеуказанной стадии.
Известно, что остаток лизина на карбоксильном конце тяжелой цепи антитела, продуцируемого в культивируемых клетках млекопитающих, удаляется (Journal of Chromatography A, 705: 129-134 (1995)), а также известно, что два аминокислотных остатка на карбоксильном конце тяжелой цепи, глицин и лизин, делетированы, и что остаток пролина, расположенный на карбоксильном конце, вновь амидирован (Analytical Biochemistry, 360: 75-83 (2007)).Однако такая делеция и модификация этих последовательностей тяжелой цепи не влияют на антигенсвязывающую активность и эффекторную функцию (активацию комплемента, антителозависимая клеточная цитотоксичность и т.д.) антитела. Соответственно, настоящее изобретение также включает антитело, которое подверглось вышеуказанной модификации, и конкретные примеры такого антитела включают делеционный мутант, включающий делецию 1 или 2 аминокислот на карбоксильном конце тяжелой цепи, и делеционный мутант, образованный амидированием вышеуказанного делеционного мутанта (например, тяжелая цепь, в которой остаток пролина на карбоксильном конце амидирован). Однако делеционные мутанты, включающие делецию на карбоксильном конце тяжелой цепи антитела в соответствии с настоящим изобретением, не ограничиваются описанными выше делеционными мутантами, если они сохраняют антигенсвязывающую активность и эффекторную функцию. Две тяжелые цепи, составляющие антитело в соответствии с настоящим изобретением, могут представлять собой любой тип тяжелой цепи, выбранный из группы, состоящей из полноразмерного антитела и вышеописанных делеционных мутантов, или комбинацию любых двух типов, выбранных из вышеуказанной группы. Соотношение индивидуальных делеционных мутантов может зависеть от типов культивируемых клеток млекопитающих, которые продуцируют антитело в соответствии с настоящим изобретением, и условий культивирования. Основным ингредиентом антитела в соответствии с настоящим изобретением могут быть антитела, где один аминокислотный остаток делетирован на каждом из карбоксильных концов двух тяжелых цепей.
Примеры изотипа антитела по настоящему изобретению могут включать IgG (IgG1, IgG2, IgG3 и IgG4). Среди прочих предпочтительными являются IgG1 и IgG2.
Примеры общей функции антитела могут включать антигенсвязывающую активность, активность нейтрализации активности антигена, активность повышения активности антигена, активность ADCC, активность антителозависимого клеточного фагоцитоза (ADCP) и активность комплемент-зависимой цитотоксичности (CDC). Функция антитела по настоящему изобретению представляет собой активность связывания с GARP, предпочтительно активность ADCC и более предпочтительно цитотоксичность (противоопухолевая активность), вызванная ADCC-опосредованным ингибированием функции Treg. Кроме того, антитело по настоящему изобретению может иметь активность ADCP и/или активность CDC, а также активность ADCC. В частности, что касается лекарственных средств, содержащих существующие противоопухолевые антитела, то сообщалось, что лекарственные средства непосредственно воздействуют на опухолевые клетки, чтобы блокировать сигналы роста, что они непосредственно воздействуют на опухолевые клетки, чтобы индуцировать сигналы клеточной смерти, что они подавляют ангиогенез, вызывают ADCC через NK-клетки и что они индуцируют активность CDC через комплемент для подавления роста опухолевых клеток (J Clin Oncol 28: 4390-4399. (2010), Clin Cancer Res; 16 (1); 11-20. (2010)). Однако в отношении активности ADCP антитела против GARP по настоящему изобретению, по меньшей мере, авторы настоящего изобретения не знали, что активность ADCP была указана как активность лекарственного средства, содержащего существующее противоопухолевое антитело против GARP.
Антитело по настоящему изобретению может представлять собой антитело, которое было мультимеризовано для усиления аффинности к антигену. Антитело, подлежащее мультимеризации, может быть либо антителом одного типа, либо несколькими антителами, распознающими множественные эпитопы одного антигена. Примеры метода мультимеризации антитела могут включать связывание домена CH3 IgG с двумя scFv (одноцепочечные антитела), связывание антитела со стрептавидином и введение мотива спираль-петля-спираль.
Антитело по настоящему изобретению также может представлять собой поликлональное антитело, которое представляет собой смесь нескольких типов антител против GARP, имеющих различные аминокислотные последовательности. Примером поликлонального антитела может быть смесь нескольких типов антител, содержащих разные CDR. В качестве такого поликлонального антитела можно использовать антитело, полученное путем культивирования смеси клеток, продуцирующих различные антитела, а затем путем очистки полученной культуры (см. WO2004/061104).
В качестве модификации антитела можно использовать антитело, связывающееся с различными типами молекул, такими как полиэтиленгликоль (ПЭГ).
Антитело по настоящему изобретению может дополнительно представлять собой конъюгат, образованный таким антителом и другим лекарственным средством (иммуноконъюгат). Такое антитело может представлять собой, например, антитело, которое связывается с радиоактивным веществом или соединением, имеющим фармакологическое действие (Nature Biotechnology (2005) 23, p. 1137-1146). Примерами такого антитела могут включать Индий (111In) Капромаб пендетид, Технеций (99mTc) Нофетумомаб мерпентан, Индий (111In) Ибритумомаб, Иттрий (90Y) Ибритумомаб и Иод (131I) Тоситумомаб.
3. Лекарственное средство, содержащие антитело против GARP
Так как антитело, полученное методом, описанным в вышеуказанном разделе «2. Получение антител против GARP», проявляет цитотоксичность в отношении Treg, то его можно использовать в качестве лекарственного средства и, в частности, в качестве терапевтического агента для лечения злокачественного новообразования и инфекционных заболеваний (в частности, малярии и ВИЧ-инфекции).
Цитотоксичность, вызванную антителом in vitro, можно измерить на основе активности подавления пролиферативных ответов клеток.
Например, культивируют опухолевую клеточную линию, сверхэкспрессирующую GARP, и в систему культивирования добавляют антитела с разной концентрацией. После этого можно измерить ингибирующую активность антитела в отношении образования очага, образования колонии и роста сфероида.
in vivo терапевтическое действие антитела на злокачественное новообразование экспериментального животного можно измерить, например, путем введения антитела голой мыши, которой была трансплантирована опухолевая клеточная линия, сверхэкспрессирующая GARP, а затем путем измерения изменения в опухолевых клетках.
Примерами типа злокачественного новообразования могут быть рак легкого, рак почек, уротелиальный рак, рак толстой кишки, рак предстательной железы, мультиформная глиобластома, рак яичников, рак поджелудочной железы, рак молочной железы, меланома, рак печени, рак мочевого пузыря, рак желудка, рак пищевода и гемобластоз. Однако тип злокачественного новообразования не ограничивается вышеуказанными примерами, при условии, что опухолевые клетки в качестве терапевтических мишеней экспрессируют GARP.
В качестве вещества, используемого в лекарственном средстве, приемлемом для фармацевтической композиции по настоящему изобретению, предпочтительным является вещество, которое нетоксично при применяемой дозе или применяемой концентрации для пациента, которому следует вводить фармацевтическую композицию.
Фармацевтическая композиция по настоящему изобретению может содержать фармацевтическое вещество для изменения или поддержания рН, осмотического давления, вязкости, прозрачности, цвета, изотоничности, стерильности, стабильности, растворимости, скорости замедленного высвобождения, коэффициента поглощения и проницаемости. Примеры фармацевтического вещества могут включать следующие вещества, но не ограничиваются ими: аминокислоты, такие как глицин, аланин, глутамин, аспарагин, аргинин или лизин; антибактериальные средства; антиоксиданты, такие как аскорбиновая кислота, сульфат натрия или гидросульфит натрия; буферы, такие как фосфатный, цитратный или боратный буфер, гидрокарбонат натрия или раствор Трис-HCl; наполнители, такие как маннит или глицин; хелатирующие агенты, такие как этилендиаминтетрауксусная кислота (ЭДТА); комплексообразователи, такие как кофеин, поливинилпирролидин, β-циклодекстрин или гидроксипропил-β-циклодекстрин; объемообразующие агенты, такие как глюкоза, манноза или декстрин; другие углеводы, такие как моносахариды или дисахариды; краситель; ароматизатор; разбавитель; эмульгатор; гидрофильные полимеры, такие как поливинилпирролидин; низкомолекулярный полипептид; солеобразующие противоионы; антисептики, такие как хлорид бензалкония, бензойная кислота, салициловая кислота, тимеросал, фенэтиловый спирт, метилпарабен, пропилпарабен, хлоргексидин, сорбиновая кислота или перекись водорода; растворители, такие как глицерин, пропиленгликоль или полиэтиленгликоль; сахарные спирты, такие как маннит или сорбит; полисорбаты, такие как суспендирующий агент, эфир сорбитана, полисорбат 20 или полисорбат 80; поверхностно-активные вещества, такие как тритон, трометамин, лецитин или холестерин; усилители устойчивости, такие как сахароза или сорбит; суспендирующий агент; усилители эластичности, такие как хлорид натрия, хлорид калия, маннит или сорбит; транспортные агенты; вспомогательные вещества; и/или фармацевтические адъюванты. Такое фармацевтическое вещество предпочтительно добавляют к антителу против GARP в количестве, которое в 0,001-100 раз, в частности, в 0,1-10 раз выше, чем масса антитела против GARP. Предпочтительный состав фармацевтической композиции в препарате может быть определен специалистом в соответствующей области в зависимости от целевого заболевания, применяемого способа введения и т.д..
Вспомогательным веществом или носителем в фармацевтической композиции может быть жидкость или твердое вещество. Подходящее вспомогательное вещество или носитель может представлять собой воду для инъекций, нормальный физиологический раствор, искусственную спинномозговую жидкость или другие вещества, обычно используемые при парентеральном введении. Нейтральный физиологический раствор или физиологический раствор, содержащий сывороточный альбумин, также можно использовать в качестве носителя. Фармацевтическая композиция может содержать трис-буфер с рН 7-8,5, ацетатный буфер с рН 4-5,5 или цитратный буфер с рН 3-6,2. Кроме того, эти буферы могут также содержать сорбит или другие соединения.
Примеры фармацевтической композиции по настоящему изобретению могут включать фармацевтическую композицию, содержащую антитело против GARP, и фармацевтическую композицию, содержащую антитело против GARP и по меньшей мере одно терапевтическое средство для лечения злокачественного новообразования. Фармацевтическую композицию по настоящему изобретению получают в виде лекарственного средства, имеющего выбранный состав и необходимую чистоту в виде лиофилизированного продукта или жидкости. Такая фармацевтическая композиция, содержащая антитело против GARP, и фармацевтическая композиция, содержащая антитело против GARP и по меньшей мере один терапевтический противоопухолевый агент, также может быть сформована в лиофилизированный продукт, содержащий подходящее вспомогательное вещество, такое как сахароза.
Терапевтический противоопухолевый агент, содержащийся вместе с антителом против GARP в вышеописанной фармацевтической композиции, может вводиться пациенту одновременно, отдельно или непрерывно вместе с антителом против GARP. В другом случае терапевтический противоопухолевый агент и антитело против GARP могут вводиться пациенту с различными интервалами введения. Примеры такого терапевтического противоопухолевого агента могут включать абраксан, карбоплатин, цисплатин, гемцитабин, иринотекан (CPT-11), паклитаксел, пеметрексед, сорафениб, винбластин, лекарственные средства, описанные в международной публикации № WO2003/038043, аналоги LH-RH (лейпрорелин, гозерелин и т.д.), эстрамустин-фосфат, антагонисты эстрогенов (тамоксифен, ралоксифен и т.д.) и ингибиторы ароматазы (анастрозол, летрозол, экземестан и т.д.). Однако примеры терапевтического противоопухолевого агента не ограничиваются вышеописанными лекарственными средствами при условии, что они обладают противоопухолевой активностью.
Пациент, который является объектом для введения, особенно ничем не ограничивается. Он предпочтительно является млекопитающим и более предпочтительно человеком.
Фармацевтическая композиция по настоящему изобретению может быть приготовлена для использования для парентерального введения или для использования для желудочно-кишечного всасывания, включая пероральное введение. Состав и концентрация препарата могут быть определены в зависимости от способа введения. Что касается аффинности антитела против GARP, содержащегося в фармацевтической композиции по настоящему изобретению, по отношению к GARP, а именно что касается константы диссоциации (значение Kd) антитела против GARP и GARP, то по мере увеличения аффинности (то есть низкое значение Kd) фармацевтическая композиция может проявлять лекарственные эффекты, даже если ее применяемая доза для человека снижается. На основании этих результатов можно также определить применяемую дозу фармацевтической композиции по настоящему изобретению для человека. Когда антитело человеческого типа против GARP вводят человеку, антитело можно вводить в дозе примерно от 0,001 до 100 мг/кг один или несколько раз с интервалом от 1 до 180 дней. Примеры формы фармацевтической композиции по настоящему изобретению могут включать инъекцию, включающую капельную инфузию, суппозиторий, трансназальный агент, подъязычный агент и агент трансдермальной абсорбции.
Далее настоящее изобретение будет конкретно описано в следующих примерах. Однако эти примеры не предназначены для ограничения объема настоящего изобретения.
Примеры
В следующих примерах, если не указано иное, отдельные операции, касающиеся генетической манипуляции, осуществляли в соответствии с методом, описанным в «Molecular Cloning» (Sambrook, J., Fritsch, E. F. and Maniatis, T., published by Cold Spring Harbor Laboratory Press in 1989), или другими методами, описанными в экспериментальных руководствах, используемых специалистами в данной области, или когда использовались коммерчески доступные реагенты или наборы, то примеры осуществляли в соответствии с инструкциями, включенными в коммерчески доступные продукты.
Далее настоящее изобретение будет конкретно описано в следующих примерах. Однако эти примеры не предназначены для ограничения объема настоящего изобретения.
Пример 1. Получение антитела
1)-1 Разделение Fab против GARP путем пэннинга в фаговом дисплее
Использовали фаговую библиотеку n-CoDeR Fab (BioInvent) для разделения Fab, связывающегося с GARP. GARP (R & D Systems) биотинилировали, используя NHS-хромогенный-биотиновый-реагент EZ-Link (Thermo Scientific). Для жидкофазного пэннинга биотинилированный GARP в твердофазном состоянии наносили на гранулы со стрептавидином Dynabeads Streptavidin M-280 (Life Technologies) и затем добавляли фаги. Несвязанные фаги удаляли посредством операции промывки с использованием магнита (DynaMag-2, Life Technologies). После этого GARP-связанные фаги собирали путем их обработки трипсином (Sigma-Aldrich) и затем амплифицировали с использованием Escherichia coli. Всего операции пэннинга проводили три раза и фрагмент ДНК, кодирующий Fab, вырезали из поликлональной фагмиды с использованием ферментов рестрикции и затем загружали в экспрессирующий вектор для Escherichia coli. Затем Escherichia coli TOP10F' (Life Technologies) трансформировали экспрессирующим вектором и затем позволяли Fab экспрессироваться в присутствии IPTG (Sigma-Aldrich). Полученный Fab подвергали скринингу с помощью ИФА.
1)-2 Скрининг на GARP-связывающий Fab с помощью ИФА
50 мкл GARP, который разбавляли до 2 мкг/мл с помощью PBS (0,01М фосфатного буферного солевого раствора (рН 7,4), содержащего 0,138М хлорида натрия и 0,0027М хлорида калия Sigma-Aldrich), добавляли в каждую лунку 384-луночного планшета Maxi-Sorp (черный, Nunc), и затем его инкубировали в течение ночи при 4°C для нанесения покрытия на планшет. Альтернативно, 50 мкл NeutrAvidin (Life Technologies), который разбавляли до 1 мкг/мл с помощью PBS, добавляли в такой 384-луночный планшет Maxi-sorp для нанесения покрытия на планшет (путем инкубации в течение ночи при 4°С). Затем, планшет промывали ИФА-буфером (PBS (Sigma-Aldrich), дополненным 0,05% Tween-20 (Bio-RAD)) три раза и затем добавляли туда биотинилированный GARP (1 пмоль/50 мкл PBS /на лунку) с последующей инкубацией при комнатной температуре в течение 1 часа при перемешивании. Планшет три раза промывали ИФА-буфером, затем блокировали с помощью казеинового блокатора (Thermo Scientific) и три раза промывали ИФА-буфером. После этого добавляли супернатант культуры, содержащий Fab, продуцируемый Escherichia coli, и затем планшет инкубировали при комнатной температуре в течение 1 часа при перемешивании. Планшет промывали три раза ИФА-буфером и добавляли 50 мкл разведенного в 2500 раз антитела против человеческого F(ab')2, меченного пероксидазой хрена (R & D Systems). Планшет дополнительно инкубировали при комнатной температуре в течение 1 часа при перемешивании. Реакционную смесь три раза промывали ИФА-буфером, а затем добавляли к лункам хемилюминесцентный субстрат SuperSignal Pico ELISA (Thermo Scientific). Через десять минут хемилюминесценцию измеряли с использованием ридера для планшетов (Envision 2104 Multilabel Reader, Perkin Elmer) и выделяли GARP-связанный Fab.
1)-3. Определение нуклеотидной последовательности ИФА-положительного клона
Вариабельные области тяжелой цепи и легкой цепи ИФА-положительных клонов (105F и 110F) анализировали методом Dye Terminator (BigDye (зарегистрированный товарный знак) Terminator v3.1, Life Technologies).Последовательности основных праймеров, используемых в секвенировании, представлены ниже.
Праймер A: 5'-GAA ACA GCT ATG AAA TAC CTA TTG C-3 '(SEQ ID NO: 10)
Праймер B: 5'-GCC TGA GCA GTG GAA GTC C-3 '(SEQ ID NO: 11)
Праймер C: 5 '-TAG GTA TTT CAT TAT GAC TGT CTC-3' (SEQ ID NO: 12)
Праймер D: 5'-CCC AGT CAC GAC GTT GTA AAA CG-3 '(SEQ ID NO: 13)
В результате описанного выше анализа были определены нуклеотидные последовательности вариабельных областей генов антитела 105F и антитела 110F.
Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 105F представляет собой последовательность, состоящую из нуклеотидов в положениях 1-354 в нуклеотидной последовательности, представленной в SEQ ID NO: 6 в списке последовательностей, и нуклеотидная последовательность вариабельной области легкой цепи антитела 105F представляет собой последовательность, состоящую из нуклеотидов в положениях 1-336 в нуклеотидной последовательности, представленной в SEQ ID NO: 7 в списке последовательностей.
Нуклеотидная последовательность вариабельной области тяжелой цепи антитела 110F представляет собой последовательность, состоящую из нуклеотидов в положениях 1-369 в нуклеотидной последовательности, представленной в SEQ ID NO: 8 в списке последовательностей, и нуклеотидная последовательность вариабельной области легкой цепи антитела 110F представляет собой последовательность, состоящую из нуклеотидов в положениях 1-333 в нуклеотидной последовательности, представленной в SEQ ID NO: 9 в списке последовательностей.
1)-4: Получение полноразмерного IgG, экспрессия и очистка IgG
Полноразмерный IgG ИФА-положительных клонов, включающих 105F и 110F, получали следующим способом.
Определяли нуклеотидную последовательность, кодирующую Fab, и после этого определяли нуклеотидные последовательности, соответствующие вариабельным областям тяжелой цепи и легкой цепи каждого антитела, указанные выше в 1)-3.
Согласно стандартному методу нуклеотидную последовательность вариабельной области вышеописанной тяжелой цепи лигировали с нуклеотидной последовательностью, кодирующей константную область тяжелой цепи человеческого IgG1 (область CH1+Fc: аминокислотная последовательность в положениях аминокислот 119-448 в аминокислотной последовательности, представленной в SEQ ID NO: 2 в списке последовательностей), а также нуклеотидную последовательность вариабельной области вышеописанной легкой цепи лигировали с нуклеотидной последовательностью, кодирующей константную область легкой цепи человеческого IgG1 (CL: аминокислотная последовательность в положениях аминокислот 113-217 в аминокислотной последовательности, представленной в SEQ ID NO: 3 в списке последовательностей). После этого полученный лигат вставляли в экспрессирующий вектор для животных клеток, такой как pcDNA3.3 (Invitrogen), для конструирования вектора, экспрессирующего IgG, для клеток животных.
Нуклеотидную последовательность сконструированного вектора, экспрессирующего IgG, анализировали снова, чтобы подтвердить, что нуклеотидная последовательность полноразмерной тяжелой цепи антитела 105F представляет собой нуклеотидную последовательность, представленную в SEQ ID NO: 6 в списке последовательностей, и что нуклеотидная последовательность полноразмерной легкой цепи антитела 105F представляет собой нуклеотидную последовательность, представленную в SEQ ID NO: 7 в списке последовательностей.
Также было подтверждено, что нуклеотидная последовательность полноразмерной тяжелой цепи антитела 110F представляет собой нуклеотидную последовательность, представленную в SEQ ID NO: 8 в списке последовательностей, и что нуклеотидная последовательность полноразмерной легкой цепи антитела 110F представляет собой нуклеотидную последовательность, представленную в SEQ ID NO: 9 в списке последовательностей.
Кроме того, на основе вышеописанных нуклеотидных последовательностей определяли аминокислотные последовательности полноразмерной тяжелой цепи и полноразмерной легкой цепи антитела 105F, кодированные нуклеотидными последовательностями, и аминокислотные последовательности полноразмерной тяжелой цепи и полноразмерной легкой цепи антитела 110F, кодируемые нуклеотидными последовательностями.
Аминокислотная последовательность тяжелой цепи антитела 105F представляет собой аминокислотную последовательность, представленную в SEQ ID NO: 2 в списке последовательностей, и аминокислотной последовательностью его легкой цепи представляет собой аминокислотную последовательность, представленную в SEQ ID NO: 3 в списке последовательностей.
Аминокислотная последовательность тяжелой цепи антитела 110F представляет собой аминокислотную последовательность, представленную в SEQ ID NO: 4 в списке последовательностей, и аминокислотная последовательностью его легкой цепи представляет собой аминокислотную последовательность, представленную в SEQ ID NO: 5 в списке последовательностей.
IgG антитела 105F или антитела 110F транзиторно экспрессировали путем вставки вышеописанного вектора, экспрессирующего IgG, для клеток животных в клетки FreeStyle 293F (Life Technologies), а затем полученные IgG при необходимости очищали с использованием аффинной колонки с Протеином A (HiTrap Mab Select SuRe, GE Healthcare). После этого буфер, в котором растворяли IgG, заменяли на PBS с использованием Vivaspin 20 (7k MWCO, GE Healthcare), и полученный продукт затем подвергали следующей стадии «1) -5».
1)-5 Подтверждение связывания очищенного IgG с GARP в соответствии с ИФА
100 мкл человеческого GARP (R & D Systems, номер по каталогу: 6055-LR), разведенного до 1 мкг/мл с использованием PBS, добавляли в каждую лунку 96-луночного планшета Maxi-sorp (Black, Nunc), и затем инкубировали в течение ночи при 4°С для нанесения покрытия на планшет.
Планшет три раза промывали ИФА-буфером и затем блокировали казеиновым блокатором при комнатной температуре в течение 1 часа. Планшет промывали три раза ИФА-буфером и добавляли 100 мкл 50 нМ антитела 105F, 50 нМ антитела 110F антитела, 50 нМ человеческого IgG (Jackson Immuno Research), 50 нМ мышиного антитело против GARP (Plato-1, ENZO Life Science) или 50 нМ мышиного IgG (Jackson Immuno Research), и планшет инкубировали при комнатной температуре в течение 1 часа при перемешивании.
Планшет промывали три раза ИФА-буфером. После этого в лунки, обработанные антителом 105F, антителом 110F или человеческим IgG, добавляли 100 мкл меченого HRP антитела против человеческого Fc (R & D Systems), которое было разведено в 5000 раз с помощью PBS. С другой стороны, в лунки, обработанные мышиным антителом против GARP и мышиным IgG, добавляли 100 мкл меченого HRP антитело против мышиного Fc (R & D Systems), которое было разбавлено в 5000 раз с помощью PBS. Планшет инкубировали при комнатной температуре в течение 1 часа при перемешивании.
Планшет промывали ИФА-буфером пять раз и затем к лункам добавляли 0,1 мл хемилюминесцентного субстрата SuperSignal Pico ELISA. Через десять минут хемилюминесценцию измеряли с помощью ридера для планшетов (Envision 2104 Multilabel Reader, Perkin Elmer).
В результате было продемонстрировано, что антитело 105F и антитело 110F связывались с GARP (Фигура 10) также, как связывалось коммерчески доступное антитело против GARP.
Пример 2. Связывание с клетками, экспрессирующими ген антигена
Что касается вектора, экспрессирующего GARP, то был приобретен кДНК-клон человеческого GARP (Origene), и затем его клонировали в вектор pcDNA3.1 (+) (Invitrogen) согласно стандартному методу. После этого была подтверждена его нуклеотидная последовательность.
Вектор, экспрессирующий GARP, и вектор pcDNA3.1, используемый в качестве контроля, каждый трансфецировали в клетки HEK-293T (ATCC: CRL-11268) с использованием Липофектамина 2000 (Invitrogen). Полученные клетки культивировали в среде DMEM (Invitrogen), дополненной 10% фетальной бычьей сывороткой (FBS, Hyclone) в течение ночи в 5% СО2 при 37°С. Затем клетки собирали из планшета методом обработки TrypLE Express (Invitrogen), и клетки дважды промывали буфером MACS (PBS, содержащий 0,5% BSA и 2мМ EDTA, Miltenyi Biotec) и затем суспендировали в том же растворе, как описано выше. Каждое из антител 105F и контрольного человеческого IgG (ENZO Life Science) добавляли к клеточной суспензии, и клетки инкубировали в течение 15 минут при 4°C. Клетки дважды промывали MACS-буфером. Добавляли флуоресцеинизотиоцианат (FITC)-меченное антитело против IgG (Jackson ImmunoResearch Laboratories) и суспендировали, и клетки дополнительно инкубировали при 4°С в течение 15 минут. Клетки промывали с помощью MACS-буфера, затем клетки фиксировали с помощью 1% PFA (полученного из 32%-ного раствора Параформальдегида (ELECTRON MICROSCOPY SCIENCES)) и измеряли с использованием проточного цитометра (FACS Canto II, Becton Dickinson). Данные анализировали с помощью Flowjo (TreeStar). Мертвые клетки удаляли из анализа путем выделения клеток, окрашенных Horizon FVS450 (Becton Dickinson). После этого создавали гистограмму интенсивности флуоресценции FITC живых клеток.
Относительно клеток HEK-293T, трансфецированных только контрольным вектором, гистограмма интенсивности флуоресценции для антитела 105F была аналогична гистограмме для контрольного IgG. С другой стороны, относительно GARP-экспрессирующих клеток HEK-293T было подтверждено, что гистограмма для антитела 105F смещается в сторону сильной интенсивности флуоресценции по сравнению с гистограммой контрольного IgG (Фигура 11). Из вышеуказанных результатов было обнаружено, что антитело 105F специфично связывается с GARP, экспрессируемым клетками HEK-293T.
Пример 3. Связывание с клетками, экспрессирующими эндогенный GARP
3)-1. Анализ проточной цитометрии с использованием клеток L428
Флюоресцентно меченую форму антитела 105F получали с использованием набора для мечения моноклональных антител Alexa Fluor 647 (Invitrogen). Клетки L428 (полученные из DSMZ) дважды промывали MACS-буфером и суспендировали в том же растворе. Меченое антитело 105F добавляли к суспензии клеток, и клетки инкубировали в течение 30 минут при 4°C. Клетки дважды промывали буфером MACS, а затем клетки фиксировали с помощью 1% PFA, и измеряли с помощью проточного цитометра (FACS Canto II, Becton Dickinson). Данные анализировали с использованием FlowJo (TreeStar). Мертвые клетки удаляли путем выделения клеток, окрашенных Horizon FVS450. После этого создавали гистограмму интенсивности флуоресценции FITC живых клеток. По сравнению с гистограммой интенсивности флуоресценции только для клеток L428 гистограмма клеток L428, к которым добавляли антитело 105F, смещается в сторону сильной интенсивности флуоресценции. Таким образом, было подтверждено, что антитело 105F связывалось с GARP, эндогенно экспрессированным клетками (Фигура 12).
3)-2. Проточный цитометрический анализ с использованием человеческих Treg
Мононуклеарные клетки периферической крови (РВМС) от здорового индивидуума разделяли с использованием Ficoll-Paque PLUS (GE Healthcare) и затем разделенные клетки высевали с плотностью 2×106 клеток/мл в среде RPMI1640 (Invitrogen), дополненной 10% FBS (далее называемой «средой RP-F10») в 24-луночный планшет с низкой адгезией (Costar). В лунки добавляли антитело против CD3 (BD Pharmingen) и антитело против CD28 (BD Pharmingen), и клетки культивировали в течение 20 часов. Затем клетки суспендировали в буфере FACS (HBSS (Invitrogen), дополненном 10мМ HEPES (Invitrogen), 2мМ EDTA (Invitrogen) и 2% FBS), и к суспензии добавляли меченое антитело 105F, полученное в стадии 3)-1 выше, и Alexa Fluor 647-меченое антитело против GARP (G14D9, eBioscience). Клетки инкубировали на льду в течение 30 минут. Клетки промывали FACS-буфером и добавляли рабочий раствор Фиксации/Пермеабилизации (eBioscience). Клетки дополнительно инкубировали на льду в течение 30 минут и промывали буфером для пермеабилизации (eBioscience). После этого к клеткам добавляли 2%-ную крысиную сыворотку (eBioscience). Клетки инкубировали при комнатной температуре в течение 15 минут и добавляли PE-меченное антитело против Foxp3 (eBioscience) с последующей инкубацией при комнатной температуре в течение 30 минут. Клетки промывали и фиксировали раствором для тканевой фиксации, который готовили в разбавлением в два раза 4%-ном растворе параформальдегида в фосфатном буфере (Wako Pure Chemical Industries, Ltd.) с D-PBS (Invitrogen) при 4°С в течение 15 минут или более. После того, как клетки промывали буфером FACS, клетки измеряли с использованием проточного цитометра (FACS Canto II, Becton Dickinson) и анализировали с использованием FlowJo (Tree Star). В результате антитело 105F связывалось с FoxP3-положительными Treg также, как коммерчески доступное антитело против GARP, (Фигура 13).
Пример 4. Свойства антител против GARP
4)-1 Активность ADCC
4)-1-1 Получение эффекторных клеток
Замороженные человеческие РВМС от здоровых доноров (Cellular Technology) разделяли путем размораживания в соответствии с протоколами 3)-2 выше. NK-клетки очищали из РВМС, используя набор для выделения NK-клеток (Miltenyi Biotec). Полученные NK-клетки инкубировали в течение ночи в среде RP-F10 (Invitrogen), дополненной 100 МЕ/мл rhIL-2 (Novartis). После этого количество живых клеток подсчитывали с помощью теста исключения с использованием трипанового синего, и клетки затем повторно суспендировали в среде RP-F10 с плотностью клеток 2×105 клеток/мл. Полученные клетки использовали в качестве эффекторных клеток.
4)-1-2. Получение клеток-мишеней
30 мкл (1110 кБк) Хром-51 (51Cr) смешивали с 0,6×106 клеток L428, описанных в примере 3)-1, в среде RPMI1640 (Invitrogen), дополненной 10% FBS, и клетки инкубировали в течение 2 часов в 5% СО2 при 37°С, так чтобы клетки были помечены радиоактивной меткой. Меченые клетки трижды промывали с помощью среды RPMI1640 (Invitrogen), дополненной 10% FBS, и затем клетки снова суспендировали в той же среде с плотностью 4×104 клеток/мл. Полученные клетки использовали в качестве клеток-мишеней.
4)-1-3 Анализ высвобождения 51Cr
Антитело 105F, которое разбавляли с помощью среды RP-F10, так чтобы конечная концентрация составляла 1, 10, 100 или 1000 нг/мл, распределяли в количестве 50 мкл/на лунку в 96-луночный микропланшет с U-образным дном (Costar), и в лунки добавляли клетки-мишени(50 мкл/на лунку). Планшет инкубировали при 4°С в течение 30 минут. Затем к лункам добавляли эффекторные клетки (100 мкл/на лунку) и планшет инкубировали в 5% СО2 при 37°С в течение 4 часов. После этого собирали 50 мкл/на лунку супернатант и наносили на LumaPlate (PerkinElmer), а полученную дозу гамма-излучения измеряли с использованием счетчика гамма-излучения. Степень клеточного лизиса, вызванного активностью ADCC, рассчитывали по следующей формуле.
Степень клеточного лизиса (%)=(AB)/(CB) х 100
A: Количество лунок с образцом
B: Среднее значение (n=3) количества спонтанного высвобождения (лунка без антител и эффекторных клеток). После добавления антитела и при добавлении эффекторных клеток добавляли 50 мкл и 100 мкл среды RP-F10, соответственно. Проводили те же операции, что и для лунок с образцами, кроме указанных выше.
C: среднее значение (n=3) количества максимального высвобождения (лунка, в которой клетки-мишени растворялись с использованием поверхностно-активного вещества). После добавления антитела добавляли 50 мл среды RP-F10. После добавления эффекторных клеток добавляли 100 мкл среды RP-F10, дополненной 2% (об./об.) Triton-X100 (Sigma). Проводили те же операции, что и для лунок с образцами, кроме указанных выше.
Результаты представлены на Фигуре 14. Антитело 105F демонстрировало цитолитическую активность на клетках L428 зависимым от концентрации антитела образом. С другой стороны, контрольный человеческий IgG не демонстрировал такой цитолитической активности. Таким образом, антитело 105F обладало активностью ADCC на клетках L428, экспрессирующих эндогенный GARP. Следует отметить, что человеческие антитела IgG1 против GARP (MHG8 и LHG10), полученные на основе информации о последовательности, описанной в патентной публикации 1, не демонстрировали активности ADCC.
4)-2 Ингибирующая активность в отношении функции Treg
4)-2-1 Получение Treg, Teff (эффекторные Т-клетки: CD4-положительные CD25-отрицательные хелперные Т-клетки) и вспомогательных клеток
CD4-положительные Т-клетки отделяли из РВМС, которые получали таким же образом, как в протоколе 4)-1-1 выше, с использованием набора для выделения CD4 Т-клеток (Miltenyi Biotec), и FITC-меченое антитело против CD4 (Miltenyi Biotec ) и APC-меченое антитело против CD25 (Miltenyi Biotec) добавляли к CD4-положительным Т-клеткам. Клетки инкубировали при 4°С в течение 30 минут. После того как клетки промывали, клетки суспендировали в буфере MACS, а CD4-положительные CD25-отрицательные клетки (Teff) и CD4-положительные CD25-сильноположительные клетки (Treg) отделяли с использованием FACS Aria IIu (Becton Dickinson).
С другой стороны, CD3-положительные клетки удаляли из PBMC с использованием CD3 микросфер (Miltenyi Biotec), и клетки облучали дозой 1 Кл/кг (поглощенная доза: 38,76 Гр/кг (3876 Рад/кг)) с использованием рентгеновского облучателя (Hitachi Medical Corporation) для приготовления вспомогательных клеток.
4)-2-2 Метод совместного культивирования и анализ ингибирующей активности в отношении функции Treg
В качестве культуральной среды использовали среду RPMI1640 (Invitrogen), дополненную пенициллином и стрептомицином (Invitrogen), 1хMEM NEAA (Invitrogen), 1хпируватом натрия (Invitrogen), 5мМ Hepes и 5% человеческой мужской сывороткой AB (Sigma). Teff (2000 клеток/на лунку) и вспомогательные клетки (20000 клеток/на лунку) смешивали и добавляли в каждую лунку 96-луночного планшета с U-образным дном, а Treg дополнительно добавляли и высевали в лунки с плотностью 500 клеток/на лунку. Кроме того, были также подготовлены контрольные лунки без Treg. Антитело против CD3, антитело против CD28 и антитело 105F добавляли в лунки с конечной концентрацией 50 или 10 мкг/мл, и планшет инкубировали в течение 5 дней в 5% СО2 при 37°С. Затем готовили [3H]-тимидин (PerkinElmer) с активностью 18,5 кБк/мл и добавляли в каждую лунку по 20 мкл/на лунку. Клетки дополнительно инкубировали в течение 18 часов. Клетки собирали в Filtermat A (PerkinE lmer) с использованием харвестера клеток (Mach II, Tomtech), а радиоактивность [3H]-тимидина, включенного в клетки, измеряли с помощью сцинтилляционного счетчика (MicroBeta, PerkinElmer). Измеренные данные были выражены в виде скорректированного числа импульсов в минуту (CCPM).
Человеческие антитела IgG1 против GARP (MHG8 и LHG10), полученные на основе информации о последовательности, описанной в патентной публикации 1, также обрабатывали с использованием настоящей экспериментальной системы.
4)-2-3 Расчет ингибирующей активности
Рассчитывали среднее значение для трех лунок при индивидуальных условиях совместного культивирования. Уменьшенное значение пролиферации при совместном культивировании Teff с Treg по сравнению с использованием только Teff определяли как «степень подавления пролиферации Teff, вызванного Treg» (=1 - [CCPM совместной культуры/CCPM только Teff]).
Ингибирующую активность каждого антитела в отношении функции Treg определяли путем вычитания степени подавления пролиферации Teff с помощью Treg в присутствии антитела из того же самого в отсутствии антитела (= [степень подавления при отсутствии добавления антитела] - [степень подавления после добавления каждого антитела]). Следует отметить, что эта ингибирующая активность образца рассчитывается каждый раз в каждом эксперименте.
Результаты ингибирующей активности антитела 105F в отношении функции Treg при 50 мкг/мл (степень ингибирования: 72,6%) представлены на Фигуре 15, а результаты антитела 105F и антител MHG-8 и LHG-10 (10 мкг/мл каждое) представлены на Фигуре 16. Антитела MHG-8 и LHG-10 не проявляли ингибирующей активности в отношении функции Treg (степень ингибирования: 0,8 и 0%, соответственно), тогда как антитело 105F значительно ингибировало функцию Treg (степень ингибирования: 65,8%). Степень ингибирования, вызванная трансдукцией киРНК к GARP в Treg, составляла около 15%, при грубом расчете с использованием значений на Фигуре 5A (CD4+CD25 - (Teff): Treg=4:1) непатентной публикации 10.
Пример 5: Получение крысиного антитела
5)-1 Получение вектора, экспрессирующего GARP
Экспрессирующий вектор, описанный в Примере 2, использовали в качестве вектора, экспрессирующего GARP, а для массового производства использовали набор EndoFree Plasmid Giga Kit (QIAGEN).
5)-2 Иммунизация крыс
Для иммунизации использовали самок крыс WKY/Izm (Япония SLC, Inc.). Во-первых, нижние конечности каждой крысы предварительно обрабатывали гиалуронидазой (SIGMA-ALDRICH), а вектор, экспрессирующий GARP, вводили внутримышечно в те же участки. Впоследствии, используя ECM830 (BTX), осуществляли электропорацию in vivo на тех же участках с использованием двухигольного электрода. Через каждые две недели повторяли ту же самую электропорацию in vivo, и проводили забор лимфатических узлов или селезенки у крысы и использовали для производства гибридом.
5)-3 Получение гибридом
Клетки лимфатических узлов или селезенки сливали с клетками миеломы мыши SP2/0-ag14 (АТСС, №CRL-1581) согласно протоколу слияния клеток под действием электрического тока с использованием LF301 Cell Fusion Unit (BEX), и клетки затем разбавляли в селекционной среде ClonaCell-HY Selection Medium D (StemCell Technologies) и инкубировали. Колонии гибридом, которые появились в культуре, были взяты и отобраны в качестве моноклональных гибридом. Каждую гибридомную колонию культивировали, и супернатант культуры из каждой гибридомы использовали для скрининга гибридом, продуцирующих антитела против GARP.
5)-4 Скрининг антител по методу клеточного ИФА (Cell-ELISA)
5)4-1. Получение клеток, экспрессирующих гены антигенов, для использования в анализе Cell-ELISA
Клетки 293α (стабильная экспрессирующая клеточная линия, полученная из клеток HEK-293 (АТСС: CRL-1573), экспрессирующие интегрин αv и интегрин β3), готовили с плотностью 7,5×105 клеток/мл в среде DMEM (Invitrogen), дополненной 10% FBS. В соответствии с процедурами трансдукции для использования Липофектамина 2000 (Life Technologies) вектор, экспрессирующий GARP, или вектор pcDNA3.1 (+), используемый в качестве отрицательного контроля, трансфецировали в клетки, и клетки распределяли в количестве по 50 мкл каждых в 96-луночный планшет (Corning). После этого клетки культивировали в среде DMEM, дополненной 10% FBS, в течение 24-27 часов в 5% СО2 при 37°С. Полученные трансфецированные клетки использовали для Cell-ELISA в адгезивном состоянии.
5)-4-2 Cell-ELISA
Супернатант культуры клеток 293α, трансфецированных экспрессирующими векторами, полученными в Примере 5)-4-1, удаляли, и супернатант культуры из каждой гибридомы добавляли к клеткам 293α, трансфецированным либо вектором, экспрессирующим GARP, либо вектором pcDNA3.1 (+). Клетки инкубировали при 4°С в течение 1 часа. Клетки в лунках один раз промывали с помощью PBS (+), дополненного 5% FBS, а затем в лунки добавляли антитело против крысиного IgG-пероксидаза, продуцированное в кроликах (SIGMA), которое было разбавлено в 500 раз с помощью PBS (+), дополненного 5% FBS. Клетки инкубировали при 4°С в течение 1 часа. Клетки в лунках три раза промывали с помощью PBS (+), дополненного 5% FBS, и в лунки добавляли OPD-краситель (который был приготовлен путем растворения дигидрохлорида о-фенилендиамина (Wako Pure Chemical Industries, Ltd.) и H2O2 в растворе OPD (0,05 М тринатрийцитрат, 0,1 М динатрий гидрофосфат 12-вода, рН 4,5), так что вещества составляли 0,4 мг/мл и 0,6% (об./об.), соответственно), в количестве 50 мкл/на лунку. Пока планшет инкубировали с перемешиванием в течение некоторого времени, проводили реакцию окрашивания. После этого к планшету добавляли 1М HCl (50 мкл/на лунку) для остановки реакции окрашивания и измеряли оптическую плотность при 490 нм с помощью ридера для планшетов (ENVISION: PerkinElmer). С целью отбора гибридом, которые продуцирует антитело, специфично связывающееся с человеческим GARP, экспрессированным на поверхности клеточной мембраны, в качестве положительных клеток, которые продуцируют антитело против человеческого GARP, отбирали гибридомы, которые продуцировали супернатант культуры, демонстрирующий более высокую поглощательную способность в клетках 293α, трансфецированных вектором, экспрессирующим GARP, чем в клетках, трансфецированных контрольным вектором pcDNA3.1 (+).
5)-5 Скрининг антител в соответствии с проточным цитометрическим методом
5)-5-1 Получение клеток, экспрессирующих гены антигенов, для использования в проточном цитометрическом анализе
Клетки HEK-293T (полученные из АТСС) высевали в 225 см2 колбу (Sumitomo Bakelite Co., Ltd.) с плотностью 5×104 клеток/см2, и затем клетки культивировали в среде DMEM, дополненной 10% FBS, в течение ночи в 5% CO2 при 37°С. На следующий день клетки HEK-293T трансфецировали вектором, экспрессирующим GARP, или вектором pcDNA3.1 (+), используемым в качестве отрицательного контроля, с использованием Липофектамина 2000, и клетки дополнительно инкубировали в течение ночи в 5% СО2 при 37°С. На следующий день трансфецированные клетки HEK-293T обрабатывали TrypLE Express (Life Technologies), промывали средой DMEM, дополненной 10% FBS, и повторно суспендировали в PBS, дополненном 5% FBS. Полученную клеточную суспензию использовали в проточном цитометрическом анализе.
5)-5-2. Проточный цитометрический анализ
Специфичность связывания с человеческим GARP антитела, продуцированного из гибридом, которые были определены как положительные с помощью Cell-ELISA в Примере 5)-4-2, дополнительно подтверждали проточным цитометрическим анализом.
Суспензию транзиторно экспрессирующих клеток HEK-293T, полученных в Примере 5)-5-1, центрифугировали и затем удаляли супернатант. После этого к клеткам добавляли супернатант культуры из каждой гибридомы и суспендировали. Клетки инкубировали при 4°С в течение 1 часа. Клетки дважды промывали с помощью PBS, дополненного 5% FBS, и к клеткам добавляли FITC-конъюгированное антитело к крысиному IgG (SIGMA), которое разбавляли в 500 раз с помощью PBS, дополненного 5% FBS, и суспендировали. Клетки инкубировали при 4°С в течение 1 часа. Клетки дважды промывали с помощью PBS, дополненного 5% FBS, и затем снова суспендировали в PBS, дополненном 5% FBS и 2 мкг/мл 7-аминоактиномицином D (Molecular Probes). Клетки измеряли с использованием проточного цитометра (FC500: производства Beckman Coulter). Данные анализировали с помощью Flowjo (TreeStar). После того, как мертвые клетки были удалены из анализа путем выделения 7-аминоациномицин-D-положительных клеток, создавали гистограмму интенсивности FITC-флуоресценции живых клеток. Гибридомы, продуцирующие антитела, связывающие человеческий GARP (113 клонов), отбирали на основании результатов, где гистограмма для антитела смещалась в сторону сильной интенсивности флуоресценции в клетках HEK-293T, трансфецированных вектором, экспрессирующим GARP, по сравнению с клетками, трансфецированными контрольным вектором pcDNA3.1,
5)-6 Получение моноклонального антитела
5)-6-1 Культивирование гибридом 151D и 198D
Из крысиных гибридом, продуцирующих антитела против человеческого GARP, полученных в 5)-5-2 выше, были отобраны гибридомы 151D и 198D, которые, как предполагается, сильно связываются с человеческим GARP.
Крысиное моноклональное антитело против GARP очищали из супернатанта культуры гибридомы.
Во-первых, объем гибридом, продуцирующих крысиные моноклональные антитела против GARP, был достаточно увеличен с помощью селекционной среды E CloneCell-HY, после чего среду заменяли на Hybridoma SFM (Life Technologies), к которой добавляли 20% FBS с ультранизким содержанием IgG (Life Technologies). После этого гибридомы (8-9×107 клеток) высевали в колбу 1272 см2 (Corning) и затем культивировали в течение 7 дней. Настоящий культуральный супернатант собирали путем центрифугирования и стерилизовали путем пропускания через фильтр 0,8 мкм и через фильтр 0,45 мкм (Corning).
5)-6-2 Очистка моноклонального антитела
Антитело очищали из культурального супернатанта гибридом, полученного в Примере 5)-6-1, в соответствии с процедурой аффинной хроматографии с Протеином G. Антитело адсорбировалось на колонке с Протеином G (GE Healthcare Bioscience), колонку затем промывали с помощью PBS, и затем антитело элюировали водным раствором 0,1 М глицин-HCl (рН 2,7). 1 М Tris-HCl (pH 9) добавляли к элюенту, так что рН доводили до рН 7-7,5. После этого раствор диализовали (Thermo Scientific, кассета диализа Slide-A-Lyzer), так что буфер был заменен на PBS. Используя центробежное фильтрующее устройство UF VIVASPIN20 (отсечка по молекулярной массе: UF30K, Sartorius), антитело концентрировали, так что концентрацию антитела доводили до 0,7 мг/мл или более. Наконец, антитело фильтровали через фильтр Minisart-Plus (Sartorius) с получением очищенного образца.
Пример 6. Клонирование крысиного антитела и продуцирование человеческого химерного антитела
6)-1 Клонирование и секвенирование кДНК крысиного антитела 151D
6)-1-1. Получение суммарной РНК из 151D-продуцирующих гибридом
Для амплификации кДНК, содержащей вариабельную область 151D, из гибридом, продуцирующих 151D, получали суммарную РНК с использованием реагента TRIzol (Ambion).
6)-1-2. Амплификация кДНК, содержащей вариабельную область тяжелой цепи 151D, в соответствии с 5'-RACE ПЦР и ее секвенирование
кДНК, содержащую вариабельную область тяжелой цепи, амплифицировали с использованием приблизительно 1 мкг суммарной РНК, полученной в Примере 6)-1-1, и набора для амплификации кДНК SMARTer RACE (Clontech).
В качестве праймеров, используемых для амплификации кДНК вариабельной области гена тяжелой цепи 151D в соответствии с ПЦР, использовали UPM (Смесь универсального праймера A: входит в комплект для амплификации кДНК SMARTer RACE) и праймеры, сконструированные из последовательностей константных областей известных крысиных тяжелых цепей.
кДНК, содержащую вариабельную область тяжелой цепи, амплифицированную с помощью 5'-RACE ПЦР, клонировали в плазмиду, а затем нуклеотидную последовательность кДНК вариабельной области тяжелой цепи подвергали анализу последовательности.
Определенная нуклеотидная последовательность кДНК, кодирующей вариабельную область тяжелой цепи 151D, представлена в SEQ ID NO: 14, и их аминокислотная последовательность представлена в SEQ ID NO: 15.
6)-1-3 Амплификация кДНК, содержащей вариабельную область легкой цепи 151D в соответствии с 5'-RACE ПЦР, и ее секвенирование
Амплификацию и секвенирование проводили тем же способом, что и в Примере 6) -1-2.Однако в качестве праймеров, используемых для амплификации кДНК вариабельной области гена легкой цепи 151D, в соответствии с процедурой ПЦР использовали UPM (Смесь универсального праймера A: входит в комплект для амплификации кДНК SMARTer RACE) и праймеры, сконструированные из последовательностей константных областей известных крысиных легких цепей.
Определенная нуклеотидная последовательность кДНК, кодирующей вариабельную область легкой цепи 151D, представлена в SEQ ID NO: 16, и ее аминокислотная последовательность представлена в SEQ ID NO: 17.
6)-2 Клонирование и секвенирование кДНК крысиного антитела 198D
Последовательности определяли тем же способом, что и в Примере 6)-1.
Определенная нуклеотидная последовательность кДНК, кодирующей вариабельную область тяжелой цепи 198D, представлена в SEQ ID NO: 18, и ее аминокислотная последовательность представлена в SEQ ID NO: 19. Определенная нуклеотидная последовательность кДНК, кодирующей вариабельную область легкой цепи 198D, представлена в SEQ ID NO: 20, и ее аминокислотная последовательность представлена в SEQ ID NO: 21.
6)-3. Получение вектора, экспрессирующего человеческое химерное антитело
6)-3-1 Конструирование вектора pCMA-LK, экспрессирующего человеческую химерную легкую цепь
Фрагмент приблизительно 5,4-т.п.о., который был получен путем гидролиза плазмиды pcDNA3.3-TOPO/LacZ (Invitrogen) ферментами рестрикции XbaI и PmeI, связывали с фрагментом ДНК, содержащим сигнальную последовательность легкой цепи человека, представленную в SEQ ID NO: 22, и последовательность ДНК, кодирующую константную область κ-цепи человека, с использованием набора для клонирования с помощью ПЦР In-Fusion Advantage (CLONTECH) с получением pcDNA3.3/LK.
Блок экспрессии неомицина удаляли из pcDNA3.3/LK для конструирования pCMA-LK.
6)-3-2. Конструирование вектора pCMA-G1, экспрессирующего тяжелую цепь человеческого химерного антитела IgG1-типа
Фрагмент ДНК, который получали путем гидролиза pCMA-LK с помощью XbaI и PmeI для удаления сигнальной последовательности легкой цепи и константной области его κ-цепи, связывали с фрагментом ДНК, содержащим сигнальную последовательность тяжелой цепи человека, представленную в SEQ ID NO: 23, и последовательность ДНК, кодирующую аминокислоты в константной области человеческого IgG1, с использованием набора для клонирования с использованием ПЦР In-Fusion Advantage (CLONTECH) для конструирования pCMA-G1.
6)-3-3 Конструирование вектора, экспрессирующего тяжелую цепь человеческого химерного 151D
Используя в качестве матрицы кДНК, кодирующую вариабельную область тяжелой цепи крысиного антитела 151D, полученную в Примере 6)-1, проводили ПЦР с праймерами, предназначенными для клонирования In-fusion, с целью амплификации фрагмента ДНК, содержащего кДНК, кодирующую вариабельную область тяжелой цепи. Используя набор для клонирования с использованием ПЦР In-Fusion HD (Clontech), амплифицированный фрагмент ДНК вставляли в сайт pCMA G1, который расщепляли ферментом рестрикции BIpI с тем, чтобы сконструировать вектор, экспрессирующий тяжелую цепь химерного человеческого 151D.
Нуклеотидная последовательность тяжелой цепи человеческого химерного 151D и аминокислотная последовательность этой тяжелой цепи представлены в SEQ ID NO: 24 и SEQ ID NO: 25, соответственно.
6)-3-4 Конструирование вектора, экспрессирующего легкую цепь человеческого химерного 151D
Используя в качестве матрицы кДНК, кодирующую вариабельную область легкой цепи 151D, полученную в Примере 6)-1, проводили ПЦР с праймерами, предназначенными для клонирования In-fusion, с целью амплификации фрагмента ДНК, содержащего кДНК, кодирующую вариабельную область легкой цепи. Используя набор для клонирования с использованием ПЦР In-Fusion HD (Clontech), амплифицированный фрагмент ДНК вставляли в сайт pCMA LK, который расщепляли ферментом рестрикции BsiWI с тем, чтобы сконструировать вектор, экспрессирующий легкую цепь человеческого химерного 151D.
Нуклеотидная последовательность легкой цепи человеческого химерного 151D и аминокислотная последовательность этой легкой цепи представлены в SEQ ID NO: 26 и SEQ ID NO: 27, соответственно.
6)-3-5 Конструирование вектора, экспрессирующего тяжелую цепь человеческого химерного 198D
Используя в качестве матрицы кДНК, кодирующую вариабельную область тяжелой цепи крысиного антитела 198D, полученную в Примере 6)-2, вектор, экспрессирующий тяжелую цепь человеческого химерного 198D, конструировали тем же способом, что и в Примере 6)-3-3.
Нуклеотидная последовательность тяжелой цепи человеческого химерного 198D и аминокислотная последовательность этой тяжелой цепи представлены в SEQ ID NO: 28 и SEQ ID NO: 29, соответственно.
6)-3-6 Конструирование вектора, экспрессирующего легкую цепь человеческого химерного 198D
Используя в качестве матрицы кДНК, кодирующую вариабельную область легкой цепи 198D, полученную в Примере 6)-2, конструировали вектор, экспрессирующий легкую цепь человеческого химерного 198D, тем же способом, что и в Примере 6)-3-4.
Нуклеотидная последовательность легкой цепи химерного человеческого 198D и аминокислотная последовательность этой легкой цепи представлены в SEQ ID NO: 30 и SEQ ID NO: 31, соответственно.
6)-4 Получение человеческого химерного антитела
6)-4-1 Продуцирование человеческого химерного антитела
В соответствии с руководством клетки FreeStyle 293F (Invitrogen) культивировали и пассировали. 1×108 клеток FreeStyle 293F (Invitrogen) в логарифмической фазе роста высевали в 250-мл колбу Фернбах Эрленмейера (CORNING) и затем разбавляли средой для экспрессии FreeStyle293 (Invitrogen) до плотности 2×106 клеток/мл.
Между тем, 20 мкг вектора, экспрессирующего тяжелую цепь, 30 мкг вектора, экспрессирующего легкую цепь, и 150 мкг полиэтиленимина (Polyscience # 24765) добавляли к 5 мл среды Opti-Pro SFM (Invitrogen), и полученную смесь осторожно перемешивали. После инкубации в течение 5 минут смесь добавляли к клеткам FreeStyle 293F.
Клетки инкубировали в инкубаторе (37°С, 8% CO2) со встряхиванием со скоростью 125 об/мин в течение 4 часов, а затем добавляли к культуре 50 мл среды EX-CELL VPRO (SAFC Biosciences), 0,36 мл GlutaMAX I (GIBCO), и 2,5 мл ультрафильтрата Yeastolate (GIBCO). Клетки дополнительно инкубировали в инкубаторе (37°С, 8% СО2) при встряхивании при 125 об./мин. в течение 7 дней. Культуральный супернатант собирали и фильтровали с помощью системы фильтров объемом 250 мл (CORNING, # 431096).
Человеческое химерное антитело 151D, полученное путем объединения вектора, экспрессирующего тяжелую цепь человеческого химерного 151D, с вектором, экспрессирующим легкую цепь человеческого химерного 151D, получило название «c151D», тогда как человеческое химерное антитело 198D, полученное путем объединения вектора, экспрессирующего тяжелую цепь человеческого химерного 198D, с вектором, экспрессирующим легкую цепь человеческого химерного 198D, получило название «c198D».
6)-4-2 Очистка химерного антитела
Культуральный супернатант, полученный в Примере 6)-4-1, очищали одностадийным способом аффинной хроматографии с Протеином A. Культуральный супернатант наносили на колонку (производства GE Healthcare Bioscience), которую набивали смолой MabSelectSuRe, уравновешенной PBS, и колонку затем промывали количеством PBS в два или более раз больше объема колонки. Затем элюирование проводили с использованием 2М раствора гидрохлорида аргинина (рН 4), так чтобы собрать фракцию, содержащую антитело. Эту фракцию подвергали центрифугированию с UF-фильтрующим устройством VIVASPIN20 (отсечка по молекулярной массе: UF30K, Sartorius), так что буфер был заменен на PBS, и антитело было сконцентрировано, тем самым доводя концентрацию антитела до 1 мг/мл или более. Наконец, антитело фильтровали через фильтр Minisart-Plus (Sartorius) с получением очищенного образца.
6)-5 Оценка активности связывания человеческого химерного антитела с человеческим GARP
Константу диссоциации между c151D или c198D, полученными в Примере 6)-4, и человеческим GARP оценивали с использованием Biacore T200 (GE Healthcare Bioscience) в соответствии с методом захвата, который включает в себя захват антитела в качестве лиганда с помощью иммобилизованного Протеина А и затем анализ константы диссоциации с использованием антигена (рекомбинантный человеческий GARP: R & D Systems) в качестве аналита. HBS-EP+(производства GE Healthcare Bioscience) использовали в качестве рабочего буфера, а в качестве сенсорного чипа использовали сенсорный чип с Протеином A (производства GE Healthcare Bioscience).
Человеческое химерное антитело (1 мкг/мл) добавляли на чип со скоростью 10 мкл/мин в течение 20 секунд и затем добавляли раствор с последовательным разведением (от 8 до 128 нМ) антигена при скорости потока 30 мкл/мин в течение 120 секунд. Затем диссоциацию отслеживали в течение 480 секунд. В качестве раствора регенерации добавляли Глицин 1.5 (производства GE Healthcare Bioscience) со скоростью потока 20 мкл/мин в течение 30 секунд.
Модель подбора 1:1 использовали при анализе данных, и рассчитывали константу скорости ассоциации ka, константу скорости диссоциации kd и константу диссоциации (KD; KD=kd/ka).
Результаты представлены Таблице 1.
Таблица 1 Константа диссоциации между c151D или c198D и человеческим GARP
[Таблица 1]
Пример 7. Продуцирование гуманизированного антитела
7)-1 Молекулярное моделирование вариабельной области антитела c151D
Молекулярное моделирование вариабельной области антитела c151D проводили в соответствии со способом, который обычно известен как гомологичное моделирование (Methods in Enzymology, 203, 121-153, (1991)).
Первичную последовательность вариабельной области иммуноглобулина человека, зарегистрированную в белковом банке данных (Nuc.Acid Res.28, 235-242 (2000)) (доступна трехмерная структура, полученная из рентгеновской кристаллической структуры) сравнивали с вариабельной областью антитела c151D.
Трехмерную структуру вариабельной области получали путем объединения друг с другом координат тяжелой цепи и легкой цепи антитела c151D и модели, имеющей высокую гомологию последовательностей с интерфейсами, для того чтобы получить «каркасную модель».
После этого репрезентативную конформацию каждого CDR включали в каркасную модель.
Наконец, чтобы устранить атомарный контакт, который был невыгоден с точки зрения энергии, проводили расчет минимизации энергии. Вышеописанные процедуры проводили с использованием Discovery Studio (Dassault Systemes).
7)-2 Конструирование аминокислотной последовательности гуманизированного антитела 151D
Гуманизированное антитело 151D было сконструировано в соответствии со способом, обычно известным как трансплантация CDR (Proc. Natl.Acad. Sci. USA 86, 10029-10033 (1989)). Акцепторное антитело выбирали на основе гомологии аминокислот в каркасной области.
Последовательность каркасной области антитела c151D сравнивали с каркасной областью консенсусной последовательности подгруппы человека, определенной по KABAT et al.(Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service National Institutes of Health, Bethesda, MD. (1991)). В результате консенсусная последовательность подгруппы 3 γ-цепи человека и подгруппы 1 и 4 κ-цепи человека имела высокую гомологию последовательностей, и на основании этого их выбирали в качестве акцепторов.
Что касается консенсусной последовательности подгруппы 3 γ-цепи человека и консенсусных последовательностей подгруппы 1 κ-цепи человека и подгруппы 4 κ-цепи человека, то аминокислотные остатки в каркасных областях выравнивали с аминокислотными остатками антитела c151D, так чтобы идентифицировать положения, в которых использовались разные аминокислоты. Положения этих остатков анализировали с использованием трехмерной модели антитела c151D, сконструированной в 7)-1 выше, и донорные остатки, которые трансплантировали на акцептор, выбирали на основании критериев, предложенных Queen et al. (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)).
Выбранные таким образом несколько донорных остатков вводили в акцепторное антитело, чтобы сконструировать последовательность гуманизированного антитела h151D таким же образом, как описано в следующих примерах.
7)-3 Конструирование тяжелой цепи h151D-H гуманизированного антитела 151D
7)-3-1 Тяжелая цепь типа h151D-H1
Тяжелая цепь гуманиизированного антитела 151D, сконструированная путем замены остатка аргинина в положении аминокислоты 35 на остаток глицина, остатка лизина в положении аминокислоты 37 на остаток лейцина, остатка лизина в положении аминокислоты 38 на остаток аргинина, остатка серина в положении аминокислоты 42 на остаток аланина, остатка треонина в положении аминокислоты 61 на остаток глицина, остатка глютамина в положении аминокислоты 62 на остаток лизина, остатка аланина в положении аминокислоты 68 на остаток серина, остатка аргинина в положении аминокислоты 80 на остаток аланина, остатка аланина в положении аминокислоты 94 на остаток серина, остатка серина в положении аминокислоты 96 на остаток аспарагина, остатка аспарагиновой кислоты в положении аминокислоты 103 на остаток аспарагина, остатка серина в положении аминокислоты 107 на остаток аланина, остатка треонина в положении аминокислоты 112 на остаток валина, остатка валина в положении аминокислоты 130 на остаток треонина и остатка метионина в положении аминокислоты 131 на остаток лейцина в тяжелой цепи c151D, представленной в SEQ ID NO: 25 в списке последовательностей, получила название тяжелой цепи типа «h151D-H1».
В нуклеотидной последовательности (SEQ ID NO: 32), кодирующей тяжелую цепь типа h151D-H1, зрелая тяжелая цепь, из которой удалена сигнальная последовательность, кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 58-1398, вариабельная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 58-408, а константная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 409-1398. Вышеупомянутая вариабельная область имеет нуклеотидную последовательность, состоящую из нуклеотидов в положениях 133-162, кодирующую CDRH1, нуклеотидную последовательность, состоящую из нуклеотидов в положениях 205-234, кодирующую CDRH2, и нуклеотидную последовательность, состоящую из нуклеотидов в положениях 352-375, кодирующую CDRH3, в SEQ ID NO: 32 в списке последовательностей.
Кроме того, в аминокислотной последовательности (SEQ ID NO: 33) тяжелой цепи типа h151D-H1 зрелая тяжелая цепь, из которой удалена сигнальная последовательность, представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 20-466, вариабельная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 20-136, а константная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 137-446. Вышеуказанная вариабельная область содержит CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 45-54 в SEQ ID NO: 33 в списке последовательностей, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 69-78, и CDRH3, состоящую из аминокислотной последовательности в положениях аминокислот 118-125.
Кроме того, последовательности, представленные в SEQ ID NO: 32 и 33, также представлены на Фигурах 31 и 21, соответственно.
7)-3-2 Тяжелая цепь типа h151D_H4
Тяжелая цепь гуманизированного антитела 151D, сконструированная для замещения остатка аргинина в положении аминокислоты 35 на остаток глицина, остатка лизина в положении аминокислоты 37 на остаток лейцина, остатка лизина в положении аминокислоты 38 на остаток аргинина, остатка серина в положении аминокислоты 42 на остаток аланина, остатка треонина в положении аминокислоты 61 на остаток глицина, остатка глютамина в положении аминокислоты 62 на остаток лизина, остаткка аланина в положении аминокислоты 94 на остаток серина, остатка аспарагиновой кислоты в положении аминокислоты 103 на остаток аспарагина, остатка серина в положении аминокислоты 107 на остаток аланина, остатка треонина в положении аминокислоты 112 на остаток валина, остатка валина в положении аминокислоты 130 на остаток треонина и остатка метионина в положении аминокислоты 131 на остаток лейцина в тяжелой цепи c151D, представленной в SEQ ID NO: 25 в списке последовательностей, получила название «тяжелой цепи типа h151D_H4».
В нуклеотидной последовательности (SEQ ID NO: 34), кодирующей тяжелую цепь типа h151D-H4, зрелая тяжелая цепь, из которой удалена сигнальная последовательность, кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 58-1398, вариабельная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 58-408, а константная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 409-1398. Вышеупомянутая вариабельная область содержит нуклеотидную последовательность, состоящую из нуклеотидов в положениях 133-162, кодирующую CDRH1, нуклеотидную последовательность, состоящую из нуклеотидов в положениях 205-234, кодирующую CDRH2, и нуклеотидную последовательность, состоящую из нуклеотидов в положениях 352, 375, кодирующую CDRH3, в SEQ ID NO: 34 в списке последовательностей.
Кроме того, в аминокислотной последовательности (SEQ ID NO: 35) тяжелой цепи типа h151D-H4 зрелая тяжелая цепь, из которой удалена сигнальная последовательность, представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 20-466, вариабельная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 20-136, а константная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 137-446. Вышеупомянутая вариабельная область содержит CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 45-54 в SEQ ID NO: 35 в списке последовательностей, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 69-78, и CDRH3, состоящую из аминокислотной последовательности в положениях аминокислот 118-125 в ней.
Кроме того, последовательности, представленные в SEQ ID NO: 34 и 35, также представлены на Фигурах 33 и 23, соответственно.
7)-4 Конструирование легкой цепи h151D_L гуманизированного антитела 151D
7)-4-1 Легкая цепь типа h151D-L1
Легкая цепь гуманизированного антитела 151D, сконструированная путем замещения остатка треонина в положении аминокислоты 29 на остаток аспарагиновой кислоты, остатка метионина в положении аминокислоты 31 на остаток лейцина, остатка фенилаланина в положении аминокислоты 32 на остаток аланина, остатка изолейцина в положении аминокислоты 33 на остаток валина, остатка валина в положении аминокислоты 35 на остаток лейцина, остатка аспарагиновой кислоты в положении аминокислоты 37 на остаток глутаминовой кислоты, остатка валина в положении аминокислоты 39 на остаток аланина, остатка метионина в положении аминокислоты 41 на остаток изолейцина, остатка треонина в положении аминокислоты 60 на остаток пролина, остатка треонина в положении аминокислоты 83 на остаток серина, остатка аспарагина в положении аминокислоты 97 на остаток серина, остатка метионина в положении аминокислоты 98 на остаток лейцина, остатка лейцина в положении аминокислоты 103 на остаток валина, остатка треонина в положении аминокислоты 120 на остаток глютамина, остатка лейцина в положении аминокислоты 124 на остаток валина, остатка лейцина в положении аминокислоты 126 на остаток изолейцина, остатка аспарагина в положении аминокислоты 127 на остаток лизина, и остатка аланина в положении аминокислоты 129 на остаток треонина в легкой цепи c151D, представленной в SEQ ID NO: 27 в списке последовательностей, получила название «легкой цепи типа h151D_L1».
В нуклеотидной последовательности (SEQ ID NO: 36), кодирующей легкую цепь типа h151D-L1, зрелая легкая цепь, из которой удалена сигнальная последовательность, кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 61-702, вариабельная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 61-387, а константная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 388-702. Вышеупомянутая вариабельная область содержит нуклеотидную последовательность, состоящую из нуклеотидов в положениях 130-116, кодирующую CDRH1, нуклеотидную последовательность, состоящую из нуклеотидов в положениях 208-228, кодирующую CDRH2, и нуклеотидную последовательность, состоящую из нуклеотидов в положениях 325, 351, кодирующую CDRH3, в SEQ ID NO: 36 в списке последовательностей.
Кроме того, в аминокислотной последовательности (SEQ ID NO: 37) легкой цепи h151D_L1 зрелая легкая цепь, из которой была удалена сигнальная последовательность, представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 21-234, вариабельная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 21-129, а константная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 130-234. Вышеуказанная вариабельная область содержит CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 44-44 в SEQ ID NO: 37 в списке последовательностей, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 70-76, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 109-117 в ней.
Кроме того, последовательности, представленные в SEQ ID NO: 36 и 37, также представлены на Фигурах 32 и 22, соответственно.
7)-4-2 Легкая цепь типа h151D-L4:
Легкая цепь гуманизированного антитела 151D, сконструированная путем замены остатка треонина в положении аминокислоты 29 на остаток серина, остатка метионина в положении аминокислоты 31 на остаток лейцина, остатка фенилаланина в положении аминокислоты 32 на остаток серина, остатка изолейцина в положении аминокислоты 33 на остаток аланина, остатка метионина в положении аминокислоты 41 на остаток изолейцина, остатка треонина в положении аминокислоты 60 на остаток пролина, остатка глутамина в положении аминокислоты 62 на остаток лизина, остатка треонина в положении аминокислоты 83 на остаток серина, остатка аспарагина в положении аминокислоты 97 на остаток серина, остатка метионина в положении аминокислоты 98 на остаток лейцина, остатка аланина в положении аминокислоты 100 на остаток пролина, остатка лейцина в положении аминокислоты 103 на остаток фенилаланина, остатка валина в положении аминокислоты 105 на остаток треонина, остатка треонина в положении аминокислоты 120 на остаток глутамина, остатка лейцина в положении аминокислоты 124 на остаток валина, остатка лейцина в положении аминокислоты 126 на остаток изолейцина, остатка аспарагина в положении аминокислоты 127 на остаток лизина и остатка аланина в положении аминокислоты 129 на остаток треонина в легкой цепи c151D, представленной в SEQ ID NO: 27 в списке последовательностей, получила название «легкой цепи типа h151D-L4».
В нуклеотидной последовательности (SEQ ID NO: 38), кодирующей легкую цепь h151D-L4, зрелая легкая цепь, из которой удалена сигнальная последовательность, кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 61-702, вариабельная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 61-387, а константная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 388-702. Вышеуказанная вариабельная область имеет нуклеотидную последовательность, состоящую из нуклеотидов в положениях 130-162, кодирующую CDRH1CDRL1, нуклеотидную последовательность, состоящую из нуклеотидов в положениях 208-228, кодирующую CDRH2CDRL2, и нуклеотидную последовательность, состоящую из нуклеотидов в положениях 325, 351, кодирующую CDRH3CDRL3, в SEQ ID NO: 38 в списке последовательностей.
Кроме того, в аминокислотной последовательности (SEQ ID NO: 39) легкой цепи h151D_L4 зрелая легкая цепь, из которой была удалена сигнальная последовательность, представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 21-234, вариабельная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 21-129, а константная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 130-234. Вышеуказанная вариабельная область содержит CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 44-54 в SEQ ID NO: 39 в списке последовательностей, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 70-76, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 109-117 в ней.
Кроме того, последовательности, представленные в SEQ ID NO: 38 и 39, также представлены на Фигурах 34 и 24, соответственно.
7)-5 Молекулярное моделирование вариабельной области c198D
Молекулярное моделирование вариабельной области антитела c198D проводили в соответствии со способом, общеизвестным как гомологичное моделирование (Methods in Enzymology, 203, 121-153, (1991)). Первичную последовательность вариабельной области иммуноглобулина человека, зарегистрированную в Белковом Банке Данных (Nuc. Acid Res.28, 235-242 (2000)) (доступна трехмерная структура, полученная из рентгеновской кристаллической структуры) сравнивали с вариабельной областью антитела c198D.
Трехмерную структуру вариабельной области получали путем объединения друг с другом координат тяжелой цепи и легкой цепи антитела c198D и модели, имеющей высокую гомологию последовательностей с их интерфейсами, с тем чтобы получить «каркасную модель».
После этого репрезентативную конформацию каждой CDR включали в каркасную модель.
Наконец, чтобы устранить атомарный контакт, который был невыгоден с точки зрения энергии, проводили расчет минимизации энергии. Вышеописанные процедуры проводили с использованием Discovery Studio (Dassault Systemes).
7)-6 Конструирование аминокислотной последовательности гуманизированного антитела 198D
Гуманизированное антитело 198D конструировали в соответствии со способом, общеизвестным как трансплантация CDR (Proc. Natl.Acad. Sci. USA 86, 10029-10033 (1989)). Акцепторное антитело выбирали на основе гомологии аминокислот в каркасной области.
Последовательность каркасной области антитела c198D сравнивали с каркасной областью консенсусной последовательности подгруппы человека, определенной KABAT et al. (Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service National Institutes of Health, Bethesda, MD. (1991)). В результате консенсусная последовательность подгруппы 2 γ-цепи человека и подгруппы 1 κ-цепи человека имела высокую гомологию последовательности, и на основании этого они были выбраны в качестве акцептора. Кроме того, в акцептор тяжелой цепи вводили несколько остатков в консенсусную последовательность подгруппы 3 γ-цепи человека.
Что касается консенсусной последовательности подгруппы 2 γ-цепи человека, содержащей часть консенсусной последовательности подгруппы 3 γ-цепи человека и консенсусной последовательности подгруппы 1 κ-цепи человека, аминокислотные остатки в каркасных областях выравнивали с аминокислотными остатками антитела c198D, так что были определены положения, в которых использовались различные аминокислоты. Положения этих остатков анализировали с использованием трехмерной модели антитела c198D, сконструированной в 7)-5 выше, и донорные остатки, которые были трансплантированы на акцептор, выбирали на основе критериев, предложенных Queen et al. (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)).
Выбранные таким образом несколько донорных остатков вводили в акцепторное антитело, чтобы сконструировать последовательность гуманизированного антитела h198D, как описано в следующих примерах.
7)-7 Конструирование тяжелой цепи h198D-H гуманизированного антитела 198D
7)-7-1 тяжелая цепь h198D-H3
Тяжелая цепь гуманизированного антитела 198D, сконструированная путем замены остатка глутамина в положении аминокислоты 20 на остаток глутаминовой кислоты, остатка аргинина в положении аминокислоты 24 на остаток валина, остатка пролина в положении аминокислоты 28 на остаток глицина, остатка глутамин в положении аминокислоты 32 на остаток лизина, остатка глутаминовой кислоты в положении аминокислоты 61 на остаток глицина, остатка серина в положении аминокислоты 80 на остаток пролина, остатка аланина в положении аминокислоты 81 на остаток серина, остатка лейцина в положении аминокислоты 86 на остаток валина, остатка серина в положении аминокислоты 87 на остаток треонина, остатка серина в положении аминокислоты 95 на остаток аспарагина, остатка фенилаланина в положении аминокислоты 98 на остаток серина, остатка метионина в положении аминокислоты 101 на остаток лейцина, остатка треонина в положении аминокислоты 103 на остаток серина, остатка лейцина в положении аминокислоты 104 на остаток валина, остатка глутамина в положении аминокислоты 105 на остаток треонина, остатка треонина в положении аминокислоты 106 на остаток аланина, остатка глутаминовой кислоты в положении аминокислоты 107 на остаток аланина, остатка метионина в положении аминокислоты 111 на остаток валина, остатка фенилаланина в положении аминокислоты 113 на остаток тирозина, остатка аланина в положении аминокислоты 133 на остаток треонина и остатка серина в положении аминокислоты 134 на остаток лейцина в тяжелой цепи c198D, представленной в SEQ ID NO: 29 в списке последовательностей, получила название «тяжелой цепи типа h198D_H3».
В нуклеотидной последовательности (SEQ ID NO: 40), кодирующей тяжелую цепь типа h198D-H3, зрелая тяжелая цепь, из которой удалена сигнальная последовательность, кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 58-1407, вариабельная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 58-417, и константная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 418-1407. Вышеуказанная вариабельная область имеет нуклеотидную последовательность, состоящую из нуклеотидов в положениях 130-162, кодирующую CDRH1, нуклеотидную последовательность, состоящую из нуклеотидов в положениях 205-231, кодирующую CDRH2, и нуклеотидную последовательность, состоящую из нуклеотидов в положениях 349, 384, кодирующую CDRH3, в SEQ ID NO: 40 в списке последовательностей.
Кроме того, в аминокислотной последовательности (SEQ ID NO: 41) тяжелой цепи типа h198D-H3 зрелая тяжелая цепь, из которой была удалена сигнальная последовательность, представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 20-469, вариабельная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 20-139, а константная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 140-469.
Кроме того, последовательности, представленные в SEQ ID NO: 40 и 41, также представлены на Фигурах 35 и 25, соответственно.
7)-8 Конструирование легкой цепи h198D-L гуманизированного антитела 198D
7)-8-1 Легкая цепь типа h198D-L4
Легкая цепь гуманизированноо антитела 198D, сконструированная путем замены остатка аланина в положении аминокислоты 29 на остаток серина, остатка глицина в положении аминокислоты 33 на остаток аланина, остатка лейцина в положении аминокислоты 35 на остаток валина, остатка глутаминовой кислотой в положении аминокислоты 37 на остаток аспарагиновой кислоты, остатка треонина в положении аминокислоты 38 на остаток аргинина, остатка глутамина в положении аминокислоты 42 на остаток треонина, остатка глутамина в положении аминокислоты 65 на остаток лизина, остатка глицина в положении аминокислоты 85 на остаток серина, остатка серина в положении аминокислоты 92 на остаток треонина, остатка лизина в положении аминокислоты 94 на остаток треонина, остатка метионина в положении аминокислоты 98 на остаток лейцина, остатка треонина в положении аминокислоты 100 на остаток пролина, остатка глутаминовой кислоты в положении аминокислоты 103 на остаток фенилаланина, остатка глицина в положении аминокислоты 104 на остаток аланина, остатка валина в положении аминокислоты 105 на остаток треонина, остатка серина в положении аминокислоты 120 на остаток глутамина, остатка лейцина в положении аминокислоты 124 на остаток валина и остатка аланина в положении аминокислоты 129 на остаток треонина в легкой цепи c198D, представленной в SEQ ID NO: 31 в списке последовательностей, получила название «легкой цепи типа h198D-L4».
В нуклеотидной последовательности (SEQ ID NO: 42), кодирующей легкую цепь h198D-L4, зрелая легкая цепь, из которой была удалена сигнальная последовательность, кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 61-702, вариабельная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 61-387, а константная область кодируется нуклеотидной последовательностью, состоящей из нуклеотидов в положениях 388-702. Вышеуказанная вариабельная область имеет нуклеотидную последовательность, состоящую из нуклеотидов в положениях 130-162, кодирующую CDRH1CDRL1, нуклеотидную последовательность, состоящую из нуклеотидов в положениях 208-228, кодирующую CDRH2CDRL2, и нуклеотидную последовательность, состоящую из нуклеотидов в положениях 325, 351, кодирующую CDRH3CDRL3, в SEQ ID NO: 42 в списке последовательностей.
Кроме того, в аминокислотной последовательности (SEQ ID NO: 43) легкой цепи h198D_L4 зрелая легкая цепь, из которой была удалена сигнальная последовательность, представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 21-234, вариабельная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 21-129, а константная область представляет собой аминокислотную последовательность, состоящую из аминокислот в положениях 130-234.
Кроме того, последовательности, представленные в SEQ ID NO: 42 и 43, также представлены на Фигурах 36 и 26, соответственно.
7)-9 Конструирование экспрессирующего вектора для гуманизированного антитела
7)-9-1 Конструирование экспрессирующего вектора для гуманизированного антитела h151D-H1L1 против человеческого GARP
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую тяжелую цепь типа h151D-H1, состоящую из нуклеотидов в положениях 58-1398 нуклеотидной последовательности тяжелой цепи типа h151D-H1, представленной в SEQ ID NO: 32 в списке последовательностей (GENEART, служба синтеза искусственных генов). Используя синтезированный фрагмент ДНК, в соответствии с протоколами технологии Cellelligent (R) CHOK1SV BioWa и Lonza был сконструирован экспрессирующий вектор для тяжелой цепи типа h151D-H1. Сконструированный экспрессирующий вектор получил название «GSV-h151D-H1».
Затем синтезировали фрагмент ДНК, содержащий последовательность, кодирующую легкую цепь типпа h151D-L1, состоящую из нуклеотидов в положениях 61-704, 702 нуклеотидной последовательности легкой цепи типа h151D-L1, представленной в SEQ ID NO: 9 36 в списке последовательностей (GENEART, служба синтеза искусственных генов).
Используя синтезированный фрагмент ДНК, в соответствии с протоколами технологии Cellelligent (R) CHOK1SV от BioWa и Lonza был сконструирован экспрессирующий вектор легкой цепи типа h151D-L1. Сконструированный экспрессирующий вектор получил название «GSV-h151D-L1».
Затем конструировали экспрессирующий вектор MACA-1511a из сконструированных таким образом экспрессирующих векторов «GSV-h151D-H1» и «GSV-h151D-L1» в соответствии с протоколами технологии Cellelligent (R) CHOK1SV от BioWa и Lonza. Полученный экспрессирующий вектор получил название «DGV-h151D-H1L1-GS».
7)-9-2 Конструирование экспрессирующего вектора для гуманизированного антитела h151D-H4L4 против человеческого GARP
Как и в случае Примера 7)-9-1, синтезировали фрагмент ДНК, содержащий последовательность, кодирующую тяжелую цепь типа h151D-H4, состоящую из нуклеотидов в положениях 58-1398 нуклеотидной последовательности тяжелой цепи типа h151D-H4, представленной в SEQ ID NO: 34 в списке последовательностей, и фрагмент ДНК, содержащий последовательность, кодирующую легкую цепь типа h151D-L4, состоящую из нуклеотидов в положениях 61-702 нуклеотидной последовательности легкой цепи типа h151D-L4, представленной в SEQ ID NO: 15 38 в списке последовательностей (GENEART, служба синтеза искусственных генов).
Используя синтезированные фрагменты ДНК, в соответствии с протоколами технологии Cellelligent (R) CHOK1SV от BioWa и Lonza был сконструирован экспрессирующий вектор MACA-1514a.Полученный экспрессирующий вектор получил название «DGV-h151D-H4L4-GS».
7)-9-3 Конструирование экспрессирующего вектора для гуманизированного антитела h198D-H3L4 против человеческого GARP
Как и в случае Примера 7)-9-1, синтезировали фрагмент ДНК, содержащий последовательность, кодирующую тяжелую цепь типа h198D-H3, состоящую из нуклеотидов в положениях 58-1407 нуклеотидной последовательности тяжелой цепи типа h198D-H3, представленной в SEQ ID NO: 40 в списке последовательностей, и фрагмент ДНК, содержащий последовательность, кодирующую легкую цепь типа h198D-L4, состоящую из нуклеотидов в положениях 61-702 нуклеотидной последовательности легкой цепи типа h198D-L4, представленной в SEQ ID NO: 42 в списке последовательностей, (GENEART, служба синтеза искусственных генов).
Используя синтезированные фрагменты ДНК, в соответствии с протоколами технологии Cellelligent (R) CHOK1SV от BioWa и Lonza был сконструирован экспрессирующий вектор MACA-1983a. Полученный экспрессирующий вектор получил название «DGV-h198D-H3L4-GS».
7)-10 Получение гуманизированного антитела против человеческого GARP
7)-10-1 Получение клеток, которые продуцируют гуманизированное антитело против человеческого GARP
7)-10-1-1 Получение клеток, которые продуцируют гуманизированное антитело h151D-H1L1 против человеческого GARP
Клетки Potelligent CHOK1SV (BioWa и Lonza) трансфецировали вектором DGV-h151D-H1L1-GS, экспрессирующим гуманизированное антитело h151D-H1L1 против человеческого GARP, который был сконструирован в примере 7)-9-1 в соответствии с протоколами Potelligent (R) CHOK1SV BioWa и Lonza, чтобы сконструировать клеточную линию, продуцирующую гуманизированное антитело H151D-H1L1 против человеческого GARP. Полученная продуцирующая клеточная линия получила название «MAC1-1».
7)-10-1-2 Получение клеток, которые продуцируют гуманизированное антитело h151D-H4L4 против человеческого GARP
Как и в случае Примера 7)-10-1-1, клетки Potelligent CHOK1SV (BioWa и Lonza) трансфецировали вектором DGV-h151D-H4L4-GS, экспрессирующим гуманизированное антитело h151D-H4L4 против человеческого GARP, который был сконструирован в примере 7)-9-2, с тем чтобы сконструировать клеточную линию, продуцирующую гуманизированное антитело h151D-H4L4 против человеческого GARP. Полученная продуцирующая клеточная линия получила название «MAC2-1».
7)-10-1-3 Получение клеток, которые продуцируют гуманизированное антитело h198D-H3L4 против человеческого GARP
Как и в примере 7)-10-1-1, клетки Potelligent CHOK1SV (BioWa и Lonza) трансфецировали вектором DGV-h198D-H3L4-GS, экспрессирующим гуманизированное антитело h198D-H3L4 против человеческого GARP, который был сконструирован в примере 7)-9-3, с тем чтобы сконструировать клеточную линию, продуцирующую гуманизированное антитело h151Dh198D-H4L4H3L4 против человеческого GARP. Полученная клеточная линия получила название «MAC3-1».
7)-10-2 Культивирование клеток, продуцирующих гуманизированное антитело против человеческого GARP
7)-10-2-1 Культивирование клеток, которые продуцируют гуманизированное антитело h151D-H1L1 против человеческого GARP
Полученную в примере 7)-10-1-1 клеточную линию «MAC1-1», продуцирующую гуманизированное антитело h151D-H1L1 против человеческого GARP, культивировали с использованием культурального оборудования в виде реактора Wave (GE Healthcare Japan). Продуцирующую клеточную линию «MAC1-1» оттаивали в среде Dsp04B (JX Energy) и затем культивировали в среде Dsp04B (JX Energy) со скоростью 120 об/мин в инкубаторе (37°C, 5% CO2). Полученный культуральный раствор разбавляли средой C36 (JX Energy) и затем культивировали с наращиванием при 120 об/мин в инкубаторе (37°С, 5% СО2).
Полученный культуральный раствор разбавляли средой С36 при 30×104 клеток/мл и затем переносили в WAVE CELLBAG (GE Healthcare Bioscience) с последующим культивированием при 37°С в 5% СО2, при скорости подачи воздуха 0,3 л/мин, при скорости вращения 18-24 об/мин под углом 6-8° в течение 13 дней.
С 3-го дня после начала культивирования в культуру добавляли среду FM4Ae2 (готовили сами) в количестве 6% от исходного объема культуры в день. Полученный культуральный раствор грубо фильтровали через глубокий фильтр Millistak MC0HC054H1 (Merck Millipore) и затем фильтровали через фильтр 0,22 мкм (Sartorius), прикрепленный к пакетам Flexboy. Этот фильтрат получил название «супернатанта культуры MACA-1511a».
7)-10-2-2 Культивирование клеток, которые продуцируют гуманизированное антитело h151D-H4L4 против человеческого GARP
Так же, как применялось в Примере 7)-10-2-1, полученную в примере 7)-10-1-2 клеточную линию «MAC1-1», продуцирующую гуманизированное антитело h151D-H4L4 против человеческого GARP, культивировали и наращивали, а затем клетки подвергали культивированию с подпиткой с использованием реактора Wave в качестве культурального оборудования (GE Healthcare Japan). Полученную культуру разбавляли средой C36 до 30×104 клеток/мл и затем переносили в WAVE CELLBAG (GE Healthcare Bioscience) с последующим культивированием в течение 13 дней. Полученный культуральный раствор фильтровали, и полученный фильтрат получил название «супернатанта культуры MACA-1514a».
7)-10-2-3 Культивирование клеток, продуцирующих гуманизированное антитело h198D-H3L4 против человеческого GARP
Таким же образом, как и в Примере 7)-10-2-1, полученную в примере 7)-10-1-3 клеточную линию «MAC1-1», продуцирующую гуманизированное антитело h198D-H3L4 против человеческого GARP, культивировали и наращивали, а затем клетки подвергали культивированию с подпиткой с использованием реактора Wave в качестве культурального оборудования (GE Healthcare Japan). Полученную культуру разбавляли средой C36 до 30×104 клеток/мл и затем переносили в WAVE CELLBAG (GE Healthcare Bioscience) с последующим культивированием в течение 13 дней. Полученный культуральный раствор фильтровали, и полученный фильтрат получил название «супернатанта культуры MACA-1983a».
7)-10-3 Очистка гуманизированного антитела против человеческого GARP
7)-10-3-1 Очистка гуманизированного антитела h151D-H1L1 против человеческого GARP
«Супернатант культуры MACA-1511a», полученный в примере 7)-10-2-1, очищали с помощью трехстадийного процесса, а именно аффинной хроматографией с рПротеином A, анионообменной хроматографией и катионообменной хроматографией.
Во-первых, супернатант культуры наносили на аффинную хроматографическую смолу с рПротеином A, которую уравновешивали PBS. После того, как весь культуральный раствор попал в колонку, колонку промывали PBS, буфером, содержащим аргинин, и PBS. Затем оставшееся вещество в колонке элюировали ацетатным буфером и затем собирали пик поглощения при 280 нм. Собранный раствор нейтрализовали трис-буфером и затем грубо фильтровали через стекловолоконный фильтр AP20 (Merck Millipore). Раствор фильтровали через Stericup-GV (Merck Millipore), который представлял собой фильтр 0,22 мкм, и полученный фильтрат определяли как очищенный пул рПротеина A.
Затем очищенный пул рПротеина A наносили на анионообменную хроматографическую смолу, которую уравновешивали PBS. После того, как наносимый раствор весь попадал на колонку, подавали PBS.Собирали проточную фракцию и пик поглощения при 280 нм во время подачи PBS. pH собранного раствора доводили уксусной кислотой, и затем раствор грубо фильтровали через стекловолоконный фильтр AP20 (Merck Millipore). Раствор фильтровали через Stericup-GV (Merck Millipore), который представлял собой фильтр 0,22 мкм, и полученный фильтрат определяли как очищенный пул AEX.
Затем очищенный пул AEX наносили на катионообменную хроматографическую смолу, которую уравновешивали ацетатным буфером. После того как раствор для нанесения весь попадал в колонку, колонку промывали ацетатным буфером. После этого проводили элюирование с использованием ацетатного буфера, содержащего высокую концентрацию NaCl, и собирали пик поглощения при 280 нм. Собранный раствор грубо фильтровали через стекловолоконный фильтр AP20 (Merck Millipore) и затем фильтровали через Stericup-GV (Merck Millipore), который представлял собой фильтр 0,22 мкм. Полученный фильтрат определяли как очищенный пул CEX.
Очищенный пул CEX концентрировали до концентрации антител 25 мг/мл с помощью Pellicon 3 кассеты 30 кДа (Merck Millipore), и буфер затем заменяли на гистидиновый буфер (25мМ гистидин, 5% сорбит, pH 6). Наконец, раствор грубо фильтровали через стекловолоконный фильтр AP20 (Merck Millipore) и затем фильтровали через Stericup-GV (Merck Millipore), который представлял собой фильтр 0,22 мкм, чтобы получить очищенный образец. Этот очищенный образец получил название «h151D-H1L1».
7)-10-3-2 Очистка гуманизированного антитела h151D-H4L4 против человеческого GARP
Таким же образом, как и в примере 7)-10-3-1, «супернатант культуры MACA-1514a», полученный в примере 7)-10-2-2, очищали с помощью трехстадийного процесса, а именно: аффинная хроматография с рПротеином А, анионообменная хроматография и катионообменная хроматография. Очищенный образец получил название «h151D-H4L4».
7)-10-3-3 Очистка гуманизированного антитела h198D-H3L4 против человеческого GARP
Таким же образом, как и в примере 7)-10-3-1, «супернатант культуры MACA-1983a», полученный в примере 7)-10-2-3, очищали с помощью трехстадийного процесса, а именно: аффинная хроматография с рПротеином А, анионообменная хроматография и катионообменная хроматография. Очищенный образец получил название «h198D-H3L4».
7)-11 Оценка активности связывания гуманизированных антител против человеческого GARP с человеческим GARP
Константу диссоциации между каждым из гуманизированных антител против человеческого GARP, H151D-H1L1, h151D-H4L4 и h151Dh198D-H3L4, полученных в примере 7)-10, и GARP оценивали с использованием Biacore T200 (GE Healthcare Bioscience) в соответствии с методом захвата, который включает в себя захват антитела в качестве лиганда иммобилизованным Протеином А, а затем анализ константы диссоциации с использованием антигена в качестве аналита. HBS-EP+(производства GE Healthcare Bioscience) использовали в качестве рабочего буфера, а сенсорный чип с Протеинаом A (производства GE Healthcare Bioscience) использовали в качестве сенсорного чипа.
Человеческое химерное антитело (1 мкг/мл) добавляли на чип со скоростью 10 мкл/мин в течение 20 секунд и добавляли последовательные разведения раствора (от 8 до 128 нМ) антигена при скорости потока 30 мкл/мин в течение 120 секунд. Затем диссоциацию отслеживали в течение 480 секунд. В качестве раствора регенерации добавляли Глицин 1.5 (производства GE Healthcare Bioscience) со скоростью потока 20 мкл/мин в течение 30 секунд.
Модель подбора 1:1 использовали при анализе данных и рассчитывали константу скорости ассоциации ka, константу скорости диссоциации kd и константу диссоциации (KD; KD=kd/ka).
Результаты представлены в Таблице 2.
Таблица 2 Константа диссоциации гуманизированных антител против человеческого GARP
[Таблица 2]
Пример 8: Связывание с клетками, экспрессирующими ген антигена
8)-1 Связывание с GARP
В соответствии со способом, описанным в Примере 2, получали суспензию клеток HEK-293T, в которые трансформировали вектор, экспрессирующий человеческий GARP, или контрольный вектор. h151D-H1L1, h151D-H4L4, h198D-H3L4 и контрольный IgG человека (человеческий IgG: Eureka Therapeutics) добавляли к суспензии клеток, и клетки инкубировали при 4°С в течение 15 минут.
Клетки дважды промывали буфером FACS (PBS (Invitrogen), дополненный 3% FBS) и после этого добавляли антитело против IgG, меченое R-Фикоэритрином (PE) (Jackson ImmunoResearch Laboratories) и Horizon FVS450 (Becton Dickinson) и суспендировали. Клетки дополнительно инкубировали при 4°С в течение 15 минут. Проточный цитометрический анализ проводили, как описано в Примере 2, и генерировали гистограмму интенсивности флуоресценции PE (Фигура 37).
Гистограммы интенсивности флуоресценции для h151D-H1L1, h151D-H4L4 и h198D-H3L4 в клетках HEK-293T, трансфецированных контрольным вектором, были подобны гистограмме для контрольного IgG (на фигуре клетки обозначают как «HEK-293T, трансфецированные пустым вектором»).
С другой стороны, было подтверждено, что гистограммы интенсивности флуоресценции для h151D-H1L1, h151D-H4L4 и h198D_H3L4 смещаются в сторону интенсивной флуоресценции в клетках HEK-293T, экспрессирующих GARP (которые обозначали на фигуре как «hGARP-трансфецированные HEK-293T») по сравнению с гистограммой для контрольного IgG человека.
Из вышеприведенных результатов было обнаружено, что h151D-H1L1, h151D-H4L4 и h198D-H3L4 специфично связывались с GARP.
8)-2 Связывание с GARP-TGF β1
8)-2-1 Конструирование вектора, экспрессирующего мутантный человеческий GARP
Используя вектор, экспрессирующий человеческий GARP (Origene) в качестве матрицы, а также с использованием праймера F (cacggcaacctgctggagcggctgctggggggg) (SEQ ID NO: 44), праймера R (caggctgttcccagacaggtccag) (SEQ ID NO: 45) и набора KOD-Plus-Mutagenesis Kit (SEQ ID NO: 44) Toyobo), YSG в положениях аминокислот 137-139 в аминокислотной последовательности GARP человека (SEQ ID NO: 1) превращали в HGN, чтобы сконструировать вектор, экспрессирующий мутантный человеческий GARP. Затем подтверждали нуклеотидную последовательность этого вектора.
8)-2-2 Ко-экспрессия GARP-TGF β1
Используя Липофектамин 2000 (Invitrogen), клетки HEK-293T трансфецировали вектором, экспрессирующим человеческий TGF β1 (Sino Biological), а также вектором, экспрессирующим человеческий GARP, или вектором, экспрессирующим мутантный человеческий GARP.
Клетки культивировали в среде DMEM (Invitrogen), дополненной 10% FBS, в течение ночи в 5% СО2 при 37°С, и клетки затем собирали из планшета, обрабатывая их TrypLE Express (Invitrogen). Собранные клетки дважды промывали буфером FACS и повторно суспендировали в том же растворе.
Антитела 105F, h151D-H1L1, h151D-H4L4 и h198D-H3L4 по настоящему изобретению, известные антитела (человеческие антитела IgG1 против GARP, MHG8 и LHG10, которые были получены на основе информации о последовательности, описанной в патентной публикации 1), и контрольный человеческий IgG (Eureka Therapeutics) добавляли к суспензии клеток, и клетки инкубировали при 4°С в течение 15 минут.
Клетки дважды промывали FACS-буфером и добавляли PE-меченное антитело против IgG (Jackson ImmunoResearch Laboratories) и Horizon FVS450 (Becton Dickinson) и суспендировали. Клетки дополнительно инкубировали при 4°С в течение 15 минут. Проточный цитометрический анализ проводили, как описано в Примере 2, и генерировали гистограммы интенсивности флуоресценции PE (Фигура 38).
Было подтверждено, что гистограммы для всех антител смещены в сторону интенсивной флуоресценции в клетках HEK-293T, ко-трансфецированных TGF β1 и GARP по сравнению с гистограммами для контрольного IgG (Фигура 38).
С другой стороны, гистограммы для MHG8 и LHG10 не сдвигались и были похожи на гистограммы контрольного IgG в клетках HEK-293T, ко-трансфецированных TGF β1 и мутантным GARP, тогда как гистограммы для антител 105F, h151D -H1L1, h151D-H4L4 и h198D-H3L4 сдвигались в сторону сильной интенсивности флуоресценции в клетках. Таким образом, было продемонстрировано, что антитела MHG8 и LHG10 не связываются с мутантным GARP, как описано в [Непатентной публикации 12].
На основе вышеуказанных результатов было продемонстрировано, что антитела 105F, h151D-H1L1, h151D-H4L4 и h198D-H3L4 связывались как с GARP, так и с GARP-мутантом на клетках, ко-экспрессирующих TGF β1, и было установлено, что эти антитела, связываются с областями, отличными от тех, с которыми связываются антитела MHG8 и LHG10.
Пример 9: Связывание с клетками, экспрессирующими эндогенный GARP
9)-1 Проточный цитометрический анализ с использованием клеток L428
Клетки L428 дважды обрабатывали буфером FACS и суспендировали в том же растворе. После этого, h151D-H1L1, h151D-H4L4, h198D_H3L4, и контролировать человеческий IgG (человеческий IgG: Eureka Therapeutics) добавляли к суспензии, и клетки инкубировали при 4°С в течение 15 минут. Клетки дважды промывали буфером FACS и добавляли и суспендировали PE-меченное антитело против IgG (Jackson ImmunoResearch Laboratories). Клетки инкубировали при 4°С в течение 15 минут. Проточный цитометрический анализ проводили, как описано в Примере 3, и генерировали гистограммы интенсивности флуоресценции PE.
В результате гистограммы для антител h151D-H1L1, h151D-H4L4 и h198D_H3L4 смещались в сторону сильной интенсивности флуоресценции в клетках L428 по сравнению с гистограммами для контрольного IgG. Таким образом, было подтверждено, что h151D-H1L1, h151D-H4L4 и h198D-H3L4 связываются с эндогенно экспрессированным GARP (Фигура 39).
9)-2 Проточный цитометрический анализ с использованием человеческих Treg
Замороженные человеческие РВМС (Cellular Technology) оттаивали в соответствии с протоколами, и РВМС высевали с плотностью 2×106 клеток/мл в 24-луночный планшет (Sumitomo Bakelite Co., Ltd.), используя среду RPMI1640 (Invitrogen), дополненную 10% FBS.
В планшет добавляли магнитные частицы Dynabeads CD3/CD28 для активации человеческих Т-клеток (Life technologies), и клетки культивировали в течение 48 часов. После этого клетки суспендировали в буфере FACS и добавляли антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4 и контролировали человеческий IgG (человеческий IgG: Eureka Therapeutics). APC-меченое антитело против CD4 (Becton Dickinson) также добавляли к суспензии. Клетки инкубировали при 4°С в течение 10 минут.
Клетки промывали буфером FACS, после чего добавляли и суспендировали FITC-меченное антитело против IgG (Jackson ImmunoResearch Laboratories) и Horizon FVS450 (Becton Dickinson). Клетки дополнительно инкубировали при 4°С в течение 15 минут.
Клетки снова промывали буфером FACS и снова суспендировали в растворе с использованием окрашивающего буферного набора FoxP3 (Miltenyi Biotec). После этого к клеткам добавляли PE-меченное антитело против Foxp3 (Miltenyi Biotec), и клетки инкубировали при 4°C в течение 30 минут.
После промывания клеток клетки измеряли с помощью проточного цитометра (FACS Canto II, Becton Dickinson). CD4-положительные клетки анализировали с использованием FlowJo (Tree Star) после того, как мертвые клетки были удалены из анализа путем выделения клеток, окрашенных Horizon FVS450.
Результаты продемонстрировали, что антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4 связываются с FoxP3-положительными Treg (Фигура 40).
Пример 10. Свойства антитела против GARP
10)-1 Активность ADCC
Антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4, известные антитела (человеческие антитела IgG1 против GARP, MHG8 и LHG10, полученные на основе информации о последовательности, описанной в патентной публикации 1), и контрольный человеческий IgG (Sigma) анализировали на предмет их ADCC согласно способу, описанному в Примере 4.
Антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4 проявляли цитолитическую активность на клетках L428 в зависимости от концентрации антител (Фигура 41A).
Напротив, как описано в Примере 4, MHG8 и LHG10 не проявляли такой цитолитической активности таким же образом, как и контрольный человеческий IgG (Фигура 41B).
Из вышеприведенных результатов было продемонстрировано, что антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4 обладают активностью ADCC.
10)-2 Ингибирующая активность в отношении функции Treg
Антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4 анализировали на их ингибирующую активность в отношении функции Treg в соответствии со способом, описанным в Примере 4. Ингибирующая активность h151D-H1L1, h151D-H4L4 и h198D-H3L4 в отношении функции Treg при конечной концентрации 1 мкг/мл представлена на Фигуре 42 (степень ингибирования h151D-H1L1: 81,5%, степень ингибирования h151D -H4L4: 80,4% и степень ингибирования h198D-H3L4: 70,8%).
Было продемонстрировано, что антитела h151D-H1L1, h151D-H4L4 и h198D-H3L4 обладают ингибирующей активностью в отношении функции Treg.
10)-3 Противоопухолевая активность (in vitro)
10)-3-1. Получение цитотоксических Т-лимфоцитов (CTL)
Согласно протоколу из Mie University клетки CTL, имеющие NY-ESO-1-специфический Т-клеточный рецептор (MU28 CD8B35 Клон # 7: полученный из Mie University), инкубировали при плотности клеток 3×105 клеток в колбе 25 см2 (Sumitomo Bakelite Co., Ltd.) в присутствии антитела против CD3 (OKT3: Imgenex), IL-2 (Novartis) и фидерных клеток в среде RPMI1640 (Invitrogen), дополненной 10% человеческой мужской сывороткой AB (Sigma), в течение 7 дней.
Что касается фидерных клеток, то замороженные человеческие РВМС (Cellular Technology) оттаивали и CD8-позитивные клетки удаляли из РВМС с использованием CD8 микросфер (Miltenyi Biotech) для получения CD8-обедненных PBMC (7,5×106 клеток/колба 25 см2), и клетки подвергали рентгеновскому облучению. Кроме того, клетки 103-LCL (полученные от Riken BioResource Center) (1,5×106 клеток/на колбу 25 см2) также подвергали рентгеновскому облучению с использованием рентгеновского облучателя (Hitachi Medical Corporation). Эти клетки использовали в качестве фидерных клеток.
Treg, полученные способом, описанным в Примере 4)-2-1, добавляли после начала культивирования (1,5×105 клеток/колба 25 см2) для оценки подавляющего эффекта Treg в отношении активности клеток CTL. Кроме того, после начала культивирования добавляли Treg (7,5×104 клеток/на колбу 25 см2), полученные вышеуказанным способом, а также антитела 105F, h151D-H1L1, h151D-H4L4, h198D-H3L4 и человеческий IgG1 (Enzo) (10 мкг/мл) для оценки противоопухолевой активности каждого антитела.
После завершения культивирования CD8-положительные клетки очищали и разделяли для приготовления клеток CTL с использованием набора для выделения CD8+T-клеток (Miltenyi Biotech). После этого готовые клетки CTL использовали для оценки активности.
10)-3-2. Получение клеток-мишеней
Клеточную линию клеток меланомы человека, а именно клетки NY-ESO-1, экспрессирующие SK-MEL-52 (полученные от Mie University: Proc Natl Acad Sci U S A. 1980 Jul; 77(7): 4260-4), культивировали с использованием RPMI1640 среды (Invitrogen), дополненной 10% FBS. Мечение клеток с помощью 51Cr проводили, как описано в Примере 4)-1-2, и клетки доводили до концентрации 2×104 клеток/мл. Полученные клетки были определены как клетки-мишени.
10)-3-3 Анализ высвобождения 51Cr
Клетки-мишени распределяли в 96-луночный планшет с U-образным дном (Costar) (50 мкл/на лунку).
Затем в планшет добавляли клетки CTL (100 мкЛ/лунку), так чтобы количество клеток CTL составило в 16, 8, 4 или в 2 раза больше, чем количество клеток-мишеней (клетки CTL: клетки-мишени=16:1, 8:1, 4:1 или 2:1), и клетки инкубировали при 5% СО2 при 37°С в течение 4 часов. После этого клетки обрабатывали в соответствии со способом, описанным в Примере 4)-1-3. Следует отметить, что ингибирующую активность образца рассчитывали каждый раз в каждом эксперименте. Кроме того, было подтверждено, что клетки CTL не проявляют цитолитической активности по отношению к клеткам, которые не экспрессируют NY-ESO-1.
Результаты измерений показаны на Фигурах 43 и 44.
Цитолитическая активность клеток CTL в отношении SK-MEL-52 подавлялась с помощью Treg (Фигура 43).
С другой стороны, скорость клеточного лизиса для клеток CTL против клеток SK-MEL-52 повышалась по мере увеличения числа клеток CTL среди клеток CTL, к которым были добавлены антитела 105F, h151D-H1L1, h151D-H4L4 или h198D-H3L4, и, кроме того, скорость клеточного лизиса была очевидно выше, чем для контрольных клеток CTL, к которым был добавлен контрольный IgG (Фигура 44) при любом отношении мишень-эффектор.
Таким образом, было продемонстрировано, что антитела 105F, h151D-H1L1, h151D-H4L4 и h198D-H3L4 ингибируют супрессивную активность Treg в отношении клеток CTL и улучшают противоопухолевую активность.
10)-4 Противоопухолевая активность (in vivo)
Известно, что противоопухолевые эффекты химерного антитела, обладающего активностью ADCC, могут быть оценены у мышей NOD/Shi-scid, IL-2R (NOG), которым были пересажены клетки L428 и введены PBMC человека (J Immunol. 2009 Oct 1; 183(7): 4782-91).
Клетки L428 (DSMZ), которые были суспендированы в смешанном растворе среды RPMI1640 (Invitrogen) и Matrigel (Becton Dickinson) (1:1) при концентрации клеток 1×107 клеток/мл, трансплантировали в объеме 0,1 мл в подкожный слой подмышечной области мышей NOG (самка, In vivo science). День, когда клетки L428 были трансплантированы, определяли как День 0. На 6-й день мышей делили на группы по величине объема опухоли (n=6 в каждой группе), а группы введения устанавливали следующим образом.
Контроль 1 PBS: вводили в дни 6, 10, 14, 18, 22 и 26, а также человеческие РВМС (Lot: 20140707) вводили в дни 6, 14 и 22
Антитело 105F: вводили в дозе 5 мг/кг в дни 6, 10, 14, 18, 22 и 26, а также человеческие РВМС (Лот: 20140707) вводили в дни 6, 14 и 22
Контроль 2 PBS: вводили в дни 6, 10, 14, 18, 22, а также человеческие РВМС (Лот: 20150924) вводили в дни 6, 14 и 22
Антитело h151D-H1L1: вводили в дозе 1 мг/кг в дни 6, 10, 14, 18 и 22, а также человеческие РВМС (Лот: 20150924) вводили в дни 6, 14 и 22
Антитело h151D-H4L4: вводили в дозе 1 мг/кг в дни 0, 6, 10, 14, 18 и 22, а также человеческие РВМС (Лот: 20150924) вводили в дни 6, 14 и 22
Антитело h198D-H3L4: вводили в дозе 1 мг/кг в дни 0, 6, 10, 14, 18 и 22, а также человеческие РВМС (Лот: 20150924) вводили в дни 6, 14 и 22
Каждое антитело разбавляли с помощью PBS (Invitrogen) и вводили мышам через хвостовую вену (10 мл/кг).
Что касается человеческих РВМС, то замороженные человеческие РВМС (Cellular Technology) оттаивали в соответствии с протоколами и готовили с концентрацией 1×107 клеток/мл. Полученные клетки (0,2 мл) вводили мышам через хвостовую вену.
Длинный диаметр (мм) и короткий диаметр (мм) опухоли измеряли с течением времени с использованием электронного штангенциркуля с цифровой индикацией (Mitutoyo), а затем объем опухоли вычисляли в соответствии со следующим выражением.
Объем опухоли (мм3)=1/2 х [длинный диаметр опухоли] х [короткий диаметр опухоли] х [короткий диаметр опухоли]
Изменение среднего значения±стандартная ошибка (SE) объема опухоли в каждой группе представлено на Фигуре 45.
Антитела 105F, h151D-H1L1, h151D-H4L4 и h198D-H3L4 проявляли противоопухолевую активность к клеткам L428 по сравнению с контрольной группой, которой вводили только PBMC. Таким образом, значительная разница наблюдалась в отношении контрольной группы (105F: t-тест и h151D-H1L1, h151D-H4L4 и h198D-H3L4: множественный сравнительный тест Дуннетта). Результаты теста достоверного отклонения (значения P) в последний день измерения индивидуальных групп (105F: день 31 и h151D-H1L1, h151D-H4L4 и h198D-H3L4: день 25) также показаны на Фигуре.
Следовательно, антитела 105F, h151D-H1L1, h151D-H4L4 и h198D-H3L4 проявляли противоопухолевую активность в моделях in vivo.
Пример 11: Эпитопный анализ антител против GARP
Эпитопы антител против человеческого GARP (105F, 110F, h151D-H1L1 и h198D-H3L4) анализировали с помощью масс-спектрометрии с обменом водорода и дейтерия.
7 мг/мл антитела против человеческого GARP смешивали с 3 мг/мл человеческого GARP (R & D Systems) или с пустым буфером в равных количествах. К полученному раствору добавляли 9 эквивалентов легкой воды или тяжелой воды. Через 30 секунд, 480 секунд или 6000 секунд добавления воды или через одну ночь добавляли к образцу в равных количествах 100мМ фосфорной кислоты, 4 М Gdn-HCl и 150 мМ TCEP (pH 2,5), так что полученную смесь затем подвергали действию дейтерий-содержащей тяжелой воды. Образующийся таким образом образец дейтерий-содержащей тяжелой воды вводили в ВЭЖХ при охлаждении и затем подавали в колонку с иммобилизованным пепсином с помощью 0,1% раствора ТФУ.
Пептидный фрагмент, полученный расщеплением GARP человека в колонке пепсина, сохраняли в колонке- ловушке C18, затем элюировали линейным градиентом воды и ацетонитрила, к которому добавляли 0,1% муравьиную кислоту и 0,025% ТФУ, и затем разделяли на колонке C18. Отделенный фрагмент пептида подвергали масс-спектрометрии с использованием времяпролетного масс-спектрометра.
Скорость замещения дейтерий-содержащей тяжелой воды рассчитывали по массе каждого пептида. Пептидный фрагмент, в котором наблюдалось значительное снижение скорости замещения дейтерий-содержащей тяжелой воды в результате добавления антитела против человеческого GARP, был идентифицирован как эпитопный фрагмент.
В случае 105F в аминокислотных остатках в положениях 366-377, 407-445 и 456-470 человеческого GARP, представленного в SEQ ID NO: 1, было обнаружено подавление скорости замещения дейтерий-содержащей тяжелой воды, и, таким образом, они были идентифицированы как эпитоп.
В случае 110F подавление скорости замещения дейтерий-содержащей тяжелой воды было обнаружено в аминокислотных остатках в положениях 54-112 и 366-392 человеческого GARP, представленного в SEQ ID NO: 1, и, таким образом, они были идентифицированы как эпитоп.
В случае h151D-H1L1 подавление скорости замещения дейтерий-содержащей тяжелой воды было обнаружено в аминокислотных остатках в положениях 352-392 человеческого GARP, представленного в SEQ ID NO: 1, и, таким образом, они были идентифицированы как эпитоп.
В случае h198D-H3L4 подавление скорости замещения дейтерий-содержащей тяжелой воды было обнаружено в аминокислотных остатках в положениях 18-112 человеческого GARP, представленного в SEQ ID NO: 1, и, таким образом, они были идентифицированы как эпитоп.
Промышленная применимость
Антитело против GARP по настоящему изобретению обладает противоопухолевой активностью, вызванной ингибирующей активностью в отношении функции Treg, которая опосредована активностью ADCC, и поэтому фармацевтическая композиция, содержащая антитело против GARP, может быть использована в качестве противоопухолевого агента.
Кроме того, избыточное присутствие Treg и их активация у пациентов с малярией и ВИЧ-инфекцией демонстрирует корреляцию с болезненным состоянием, а удаление Treg вызывает ремиссию каждого заболевания в мышиных моделях заболеваний. Соответственно, можно ожидать, что эффективное ингибирование функции Treg также будет оказывать терапевтическое воздействие на рефракторные инфекции, такие как малярия и ВИЧ.
Список последовательностей Свободный текст
SEQ ID NO: 1 - аминокислотная последовательность GARP
SEQ ID NO: 2 - аминокислотная последовательность тяжелой цепи антитела 105F
SEQ ID NO: 3 - аминокислотная последовательность легкой цепи антитела 105F
SEQ ID NO: 4 - аминокислотная последовательность тяжелой цепи антитела 110F
SEQ ID NO: 5 - аминокислотная последовательность легкой цепи антитела 110F
SEQ ID NO: 6 - нуклеотидная последовательность тяжелой цепи антитела 105F
SEQ ID NO: 7 - нуклеотидная последовательность легкой цепи антитела 105F
SEQ ID NO: 8 - нуклеотидная последовательность тяжелой цепи антитела 110F
SEQ ID NO: 9 - нуклеотидная последовательность легкой цепи антитела 110F
SEQ ID NO: 10 - праймер A
SEQ ID NO: 11 - праймер B
SEQ ID NO: 12 - праймер C
SEQ ID NO: 13 - праймер D
SEQ ID NO: 14 - нуклеотидная последовательность кДНК, кодирующей вариабельную область тяжелой цепи 151D
SEQ ID NO: 15 - аминокислотная последовательность вариабельной области тяжелой цепи 151D
SEQ ID NO: 16 - нуклеотидная последовательность кДНК, кодирующей вариабельную область легкой цепи 151D
SEQ ID NO: 17 - аминокислотная последовательность вариабельной области легкой цепи 151D
SEQ ID NO: 18 - нуклеотидная последовательность кДНК, кодирующей вариабельную область тяжелой цепи 198D
SEQ ID NO: 19 - аминокислотная последовательность вариабельной области тяжелой цепи 198D
SEQ ID NO: 20 - нуклеотидная последовательность кДНК, кодирующей вариабельную область легкой цепи 198D
SEQ ID NO: 21 - аминокислотная последовательность вариабельной области легкой цепи 198D
SEQ ID NO: 22 - нуклеотидная последовательность фрагмента ДНК, содержащего сигнальную последовательность легкой цепи человека, и последовательность, кодирующую аминокислоты в константной области κ-цепи человека
SEQ ID NO: 23 - нуклеотидная последовательность фрагмента ДНК, содержащего сигнальную последовательность тяжелой цепи человека, и последовательность, кодирующую аминокислоты в константной области IgG1 человека
SEQ ID NO: 24 - нуклеотидная последовательность тяжелой цепи человеческого химерного антитела c151D
SEQ ID NO: 25 - аминокислотная последовательность тяжелой цепи человеческого химерного антитела c151D
SEQ ID NO: 26 - нуклеотидная последовательность легкой цепи человеческого химерного антитела c151D
SEQ ID NO: 27 - аминокислотная последовательность легкой цепи человеческого химерного антитела c151D
SEQ ID NO: 28 - нуклеотидная последовательность тяжелой цепи человеческого химерного антитела c198D
SEQ ID NO: 29 - аминокислотная последовательность тяжелой цепи человеческого химерного антитела c198D
SEQ ID NO: 30 - нуклеотидная последовательность легкой цепи человеческого химерного антитела c198D
SEQ ID NO: 31 - аминокислотная последовательность легкой цепи человеческого химерного антитела c198D
SEQ ID NO: 32 - нуклеотидная последовательность гуманизированного антитела h151D-H1
SEQ ID NO: 33 - аминокислотная последовательность гуманизированного антитела h151D-H1
SEQ ID NO: 34 - нуклеотидная последовательность гуманизированного антитела h151D-H4
SEQ ID NO: 35 - аминокислотная последовательность гуманизированного антитела h151D-H4
SEQ ID NO: 36 - нуклеотидная последовательность гуманизированного антитела h151D-L1
SEQ ID NO: 37 - аминокислотная последовательность гуманизированного антитела h151D-L1
SEQ ID NO: 38 - нуклеотидная последовательность гуманизированного антитела h151D-L4
SEQ ID NO: 39 - аминокислотная последовательность гуманизированного антитела h151D-L4
SEQ ID NO: 40 - нуклеотидная последовательность гуманизированного антитела h198D-H3
SEQ ID NO: 41 - аминокислотная последовательность гуманизированного антитела h198D-H3
SEQ ID NO: 42 - нуклеотидная последовательность гуманизированного антитела h198D-L4
SEQ ID NO: 43 - аминокислотная последовательность гуманизированного антитела h198D-L4
SEQ ID NO: 44 - праймер F
SEQ ID NO: 45 - праймер R
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> DAIICHI SANKYO CO. LTD.
<120> АНТИТЕЛО ПРОТИВ GARP
<130> FP1628
<150> JP2015-187488
<151> 2015-09-24
<160> 45
<170> PatentIn version 3.5
<210> 1
<211> 662
<212> БЕЛОК
<213> Человека
<400> 1
Met Arg Pro Gln Ile Leu Leu Leu Leu Ala Leu Leu Thr Leu Gly Leu
1 5 10 15
Ala Ala Gln His Gln Asp Lys Val Pro Cys Lys Met Val Asp Lys Lys
20 25 30
Val Ser Cys Gln Val Leu Gly Leu Leu Gln Val Pro Ser Val Leu Pro
35 40 45
Pro Asp Thr Glu Thr Leu Asp Leu Ser Gly Asn Gln Leu Arg Ser Ile
50 55 60
Leu Ala Ser Pro Leu Gly Phe Tyr Thr Ala Leu Arg His Leu Asp Leu
65 70 75 80
Ser Thr Asn Glu Ile Ser Phe Leu Gln Pro Gly Ala Phe Gln Ala Leu
85 90 95
Thr His Leu Glu His Leu Ser Leu Ala His Asn Arg Leu Ala Met Ala
100 105 110
Thr Ala Leu Ser Ala Gly Gly Leu Gly Pro Leu Pro Arg Val Thr Ser
115 120 125
Leu Asp Leu Ser Gly Asn Ser Leu Tyr Ser Gly Leu Leu Glu Arg Leu
130 135 140
Leu Gly Glu Ala Pro Ser Leu His Thr Leu Ser Leu Ala Glu Asn Ser
145 150 155 160
Leu Thr Arg Leu Thr Arg His Thr Phe Arg Asp Met Pro Ala Leu Glu
165 170 175
Gln Leu Asp Leu His Ser Asn Val Leu Met Asp Ile Glu Asp Gly Ala
180 185 190
Phe Glu Gly Leu Pro Arg Leu Thr His Leu Asn Leu Ser Arg Asn Ser
195 200 205
Leu Thr Cys Ile Ser Asp Phe Ser Leu Gln Gln Leu Arg Val Leu Asp
210 215 220
Leu Ser Cys Asn Ser Ile Glu Ala Phe Gln Thr Ala Ser Gln Pro Gln
225 230 235 240
Ala Glu Phe Gln Leu Thr Trp Leu Asp Leu Arg Glu Asn Lys Leu Leu
245 250 255
His Phe Pro Asp Leu Ala Ala Leu Pro Arg Leu Ile Tyr Leu Asn Leu
260 265 270
Ser Asn Asn Leu Ile Arg Leu Pro Thr Gly Pro Pro Gln Asp Ser Lys
275 280 285
Gly Ile His Ala Pro Ser Glu Gly Trp Ser Ala Leu Pro Leu Ser Ala
290 295 300
Pro Ser Gly Asn Ala Ser Gly Arg Pro Leu Ser Gln Leu Leu Asn Leu
305 310 315 320
Asp Leu Ser Tyr Asn Glu Ile Glu Leu Ile Pro Asp Ser Phe Leu Glu
325 330 335
His Leu Thr Ser Leu Cys Phe Leu Asn Leu Ser Arg Asn Cys Leu Arg
340 345 350
Thr Phe Glu Ala Arg Arg Leu Gly Ser Leu Pro Cys Leu Met Leu Leu
355 360 365
Asp Leu Ser His Asn Ala Leu Glu Thr Leu Glu Leu Gly Ala Arg Ala
370 375 380
Leu Gly Ser Leu Arg Thr Leu Leu Leu Gln Gly Asn Ala Leu Arg Asp
385 390 395 400
Leu Pro Pro Tyr Thr Phe Ala Asn Leu Ala Ser Leu Gln Arg Leu Asn
405 410 415
Leu Gln Gly Asn Arg Val Ser Pro Cys Gly Gly Pro Asp Glu Pro Gly
420 425 430
Pro Ser Gly Cys Val Ala Phe Ser Gly Ile Thr Ser Leu Arg Ser Leu
435 440 445
Ser Leu Val Asp Asn Glu Ile Glu Leu Leu Arg Ala Gly Ala Phe Leu
450 455 460
His Thr Pro Leu Thr Glu Leu Asp Leu Ser Ser Asn Pro Gly Leu Glu
465 470 475 480
Val Ala Thr Gly Ala Leu Gly Gly Leu Glu Ala Ser Leu Glu Val Leu
485 490 495
Ala Leu Gln Gly Asn Gly Leu Met Val Leu Gln Val Asp Leu Pro Cys
500 505 510
Phe Ile Cys Leu Lys Arg Leu Asn Leu Ala Glu Asn Arg Leu Ser His
515 520 525
Leu Pro Ala Trp Thr Gln Ala Val Ser Leu Glu Val Leu Asp Leu Arg
530 535 540
Asn Asn Ser Phe Ser Leu Leu Pro Gly Ser Ala Met Gly Gly Leu Glu
545 550 555 560
Thr Ser Leu Arg Arg Leu Tyr Leu Gln Gly Asn Pro Leu Ser Cys Cys
565 570 575
Gly Asn Gly Trp Leu Ala Ala Gln Leu His Gln Gly Arg Val Asp Val
580 585 590
Asp Ala Thr Gln Asp Leu Ile Cys Arg Phe Ser Ser Gln Glu Glu Val
595 600 605
Ser Leu Ser His Val Arg Pro Glu Asp Cys Glu Lys Gly Gly Leu Lys
610 615 620
Asn Ile Asn Leu Ile Ile Ile Leu Thr Phe Ile Leu Val Ser Ala Ile
625 630 635 640
Leu Leu Thr Thr Leu Ala Ala Cys Cys Cys Val Arg Arg Gln Lys Phe
645 650 655
Asn Gln Gln Tyr Lys Ala
660
<210> 2
<211> 448
<212> БЕЛОК
<213> Человека
<400> 2
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Val Ser Trp Asn Gly Ser Arg Thr His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Arg Gln Leu Ala Glu Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 3
<211> 217
<212> БЕЛОК
<213> Человека
<400> 3
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Val Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Ala Asp Thr Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser
85 90 95
Leu Arg Gly Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 4
<211> 453
<212> БЕЛОК
<213> Человека
<400> 4
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Gly Ile Ser Trp Asn Ser Ala Ile Thr Val Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Ala Gly Gly Arg Tyr Ser Gly Ser Tyr Tyr Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 5
<211> 216
<212> БЕЛОК
<213> Человека
<400> 5
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Arg Ser
85 90 95
Leu Asn Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 6
<211> 1344
<212> ДНК
<213> Человека
<400> 6
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt gactactaca tgagctggat ccgccaggct 120
ccaggcaagg ggctggagtg ggtatcgggt gttagttgga atggcagtag gacgcactat 180
gcagactctg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac actgccgtgt attactgtgc cagacagagg 300
cagctggctg aatttgacta ctggggccaa ggtaccctgg tcaccgtgag ctcagcctcc 360
accaagggcc caagcgtctt ccccctggca ccctcctcca agagcacctc tggcggcaca 420
gccgccctgg gctgcctggt caaggactac ttccccgaac ccgtgaccgt gagctggaac 480
tcaggcgccc tgaccagcgg cgtgcacacc ttccccgctg tcctgcagtc ctcaggactc 540
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 600
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agagagttga gcccaaatct 660
tgtgacaaaa ctcacacatg cccaccctgc ccagcacctg aactcctggg gggaccctca 720
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 780
acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 840
gacggcgtgg aggtgcataa tgccaagaca aagccccggg aggagcagta caacagcacg 900
taccgggtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 960
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1020
aaaggccagc cccgggaacc acaggtgtac accctgcccc catcccggga ggagatgacc 1080
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1140
gagtgggaga gcaatggcca gcccgagaac aactacaaga ccacccctcc cgtgctggac 1200
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1260
ggcaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacccagaag 1320
agcctctccc tgtctcccgg caaa 1344
<210> 7
<211> 651
<212> ДНК
<213> Человека
<400> 7
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacattggg gcgggttatg ttgtacattg gtatcagcag 120
ctcccaggaa cggcccccaa actcctcatc tatgctgaca ccaatcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cagtgggctc 240
cggtccgagg atgaggctga ttattactgc cagtcctatg acagcagcct gagaggttgg 300
gtgttcggcg gaggaaccaa gctgacggtc ctaggtcagc ctaaggctgc ccctagcgtg 360
accctgttcc ctccttccag cgaggagctt caagctaaca aggccaccct ggtgtgtctt 420
atctctgact tctaccctgg cgctgtgacc gtggcctgga aggctgacag ctcccctgtg 480
aaggccggag tggagaccac cacacctagc aagcagtcta acaacaagta cgctgccagc 540
tcctacctga gccttacccc tgagcagtgg aagtctcaca gaagctactc ctgtcaagtg 600
acccacgagg gcagcaccgt ggagaagacc gtggctccta ccgagtgttc c 651
<210> 8
<211> 1359
<212> ДНК
<213> Человека
<400> 8
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatggca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtcgccgga attagttgga acagtgccat cacagtctat 180
gcggactctg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac actgccgtgt attactgtgc aaaagatgcc 300
gggggccggt atagtgggag ctactacttt gactactggg gccaaggtac cctggtcacc 360
gtgagctcag cctccaccaa gggcccaagc gtcttccccc tggcaccctc ctccaagagc 420
acctctggcg gcacagccgc cctgggctgc ctggtcaagg actacttccc cgaacccgtg 480
accgtgagct ggaactcagg cgccctgacc agcggcgtgc acaccttccc cgctgtcctg 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaga 660
gttgagccca aatcttgtga caaaactcac acatgcccac cctgcccagc acctgaactc 720
ctggggggac cctcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 780
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 840
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc ccgggaggag 900
cagtacaaca gcacgtaccg ggtggtcagc gtcctcaccg tcctgcacca ggactggctg 960
aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa 1020
accatctcca aagccaaagg ccagccccgg gaaccacagg tgtacaccct gcccccatcc 1080
cgggaggaga tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 1140
agcgacatcg ccgtggagtg ggagagcaat ggccagcccg agaacaacta caagaccacc 1200
cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggacaag 1260
agcaggtggc agcagggcaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 1320
cactacaccc agaagagcct ctccctgtct cccggcaaa 1359
<210> 9
<211> 648
<212> ДНК
<213> Человека
<400> 9
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gcaggttatg atgtacactg gtatcagcag 120
ctcccaggaa cggcccccaa actcctcatc tatggtaaca gcaatcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cagtgggctc 240
cggtccgagg atgaggctga ttattactgc cagtcctatg acagaagcct gaattgggtg 300
ttcggcggag gaaccaagct gacggtccta ggtcagccta aggctgcccc tagcgtgacc 360
ctgttccctc cttccagcga ggagcttcaa gctaacaagg ccaccctggt gtgtcttatc 420
tctgacttct accctggcgc tgtgaccgtg gcctggaagg ctgacagctc ccctgtgaag 480
gccggagtgg agaccaccac acctagcaag cagtctaaca acaagtacgc tgccagctcc 540
tacctgagcc ttacccctga gcagtggaag tctcacagaa gctactcctg tcaagtgacc 600
cacgagggca gcaccgtgga gaagaccgtg gctcctaccg agtgttcc 648
<210> 10
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер A
<400> 10
gaaacagcta tgaaatacct attg 24
<210> 11
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер B
<400> 11
gcctgagcag tggaagtcc 19
<210> 12
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер C
<400> 12
taggtatttc attatgactg tctc 24
<210> 13
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер D
<400> 13
cccagtcacg acgttgtaaa acg 23
<210> 14
<211> 351
<212> ДНК
<213> Крысы
<400> 14
gaggtgcagc tggtggagtc tgggggaggc ttagtgcagc ctggaaggtc caagaaactc 60
tcctgttcag cctcaggatt cactttcagt aactattaca tggcctgggt ccgccaggct 120
ccaacgcagg gtctggagtg ggtcgcatcc attggtactg ttggtggtaa cacttactat 180
cgagactccg tgaagggccg attcactatc tccagagatg atgcaaaaag caccctatac 240
ctgcaaatgg acagtctgag gtctgaggac acggccactt attactgtgc aagagaggat 300
tacggagggt ttccccactg gggccaagga gtcatggtca cagtctcctc a 351
<210> 15
<211> 117
<212> БЕЛОК
<213> Крысы
<400> 15
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Lys Lys Leu Ser Cys Ser Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Thr Gln Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Gly Thr Val Gly Gly Asn Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Glu Asp Tyr Gly Gly Phe Pro His Trp Gly Gln Gly Val Met
100 105 110
Val Thr Val Ser Ser
115
<210> 16
<211> 327
<212> ДНК
<213> Крысы
<400> 16
aatattgtga tgactcagtc tcccacatcc atgttcatat cagtcggaga cagggtcacc 60
atgaactgta aggccagtca gaatgtggga actaatgtag actggtacca gcagaaaaca 120
gggcagtctc ctaaactgct tatctatggg gcgtccaacc gctacactgg agtccctgat 180
cgcttcacag gcagtggatc tggaacagat ttcactctca ccatcagcaa catgcaggct 240
gaagacctgg ctgtttatga ctgtctacag tataagtaca atccatacac gtttggaact 300
gggaccaagc tggaactgaa ccgggct 327
<210> 17
<211> 109
<212> БЕЛОК
<213> Крысы
<400> 17
Asn Ile Val Met Thr Gln Ser Pro Thr Ser Met Phe Ile Ser Val Gly
1 5 10 15
Asp Arg Val Thr Met Asn Cys Lys Ala Ser Gln Asn Val Gly Thr Asn
20 25 30
Val Asp Trp Tyr Gln Gln Lys Thr Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Met Gln Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Asp Cys Leu Gln Tyr Lys Tyr Asn Pro Tyr
85 90 95
Thr Phe Gly Thr Gly Thr Lys Leu Glu Leu Asn Arg Ala
100 105
<210> 18
<211> 360
<212> ДНК
<213> Крысы
<400> 18
caggtgcagc tgagggagtc aggacctggt ctggtgcagc cctcacagac cctgtccctc 60
acctgcactg tctctgggtt ctcactaacc agctttcatg taagctgggt tcgccagcct 120
ccagagaagg gtctggagtg gattgcaaca atttcaagtg gtggaggtac atattataat 180
tcagctctca aatcccgact gagcatcagc agggacacct ccaagagcca agttttctta 240
aagatgagca ctctgcaaac tgaagacaca gccatgtact tctgtgcccg gatttcgggc 300
tggggccatt actatgttat ggatgtctgg ggtcaaggag cttcagtcac tgtctcctca 360
<210> 19
<211> 120
<212> БЕЛОК
<213> Крысы
<400> 19
Gln Val Gln Leu Arg Glu Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Ser Phe
20 25 30
His Val Ser Trp Val Arg Gln Pro Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Ala Thr Ile Ser Ser Gly Gly Gly Thr Tyr Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Met Ser Thr Leu Gln Thr Glu Asp Thr Ala Met Tyr Phe Cys Ala
85 90 95
Arg Ile Ser Gly Trp Gly His Tyr Tyr Val Met Asp Val Trp Gly Gln
100 105 110
Gly Ala Ser Val Thr Val Ser Ser
115 120
<210> 20
<211> 327
<212> ДНК
<213> Крысы
<400> 20
gacatccaga tgacacagtc tccagcttcc ctgtctggat ctctgggaga aactgtcacc 60
atccaatgtc aagcaagtga ggacatttac agtggtttag cgtggtatca gcagaagcca 120
gggaaatctc ctcagctcct gatctatggt gcaggtagct tacaagacgg cgtcccatca 180
cgattcagtg gcggtggatc tggcacacat tattctctca agatcagcag catgcaaact 240
gaagatgaag gggtttattt ctgtcaacag ggtttaaagt ttccgctcac gttcggttct 300
gggaccaagc tggagatcaa acgggct 327
<210> 21
<211> 109
<212> БЕЛОК
<213> Крысы
<400> 21
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Gly Ser Leu Gly
1 5 10 15
Glu Thr Val Thr Ile Gln Cys Gln Ala Ser Glu Asp Ile Tyr Ser Gly
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Gln Leu Leu Ile
35 40 45
Tyr Gly Ala Gly Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Gly Gly Ser Gly Thr His Tyr Ser Leu Lys Ile Ser Ser Met Gln Thr
65 70 75 80
Glu Asp Glu Gly Val Tyr Phe Cys Gln Gln Gly Leu Lys Phe Pro Leu
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg Ala
100 105
<210> 22
<211> 449
<212> ДНК
<213> Человека
<400> 22
gcctccggac tctagagcca ccatggtgct gcagacccag gtgttcatct ccctgctgct 60
gtggatctcc ggcgcgtacg gcgatatcgt gatgattaaa cgtacggtgg ccgccccctc 120
cgtgttcatc ttccccccct ccgacgagca gctgaagtcc ggcaccgcct ccgtggtgtg 180
cctgctgaat aacttctacc ccagagaggc caaggtgcag tggaaggtgg acaacgccct 240
gcagtccggg aactcccagg agagcgtgac cgagcaggac agcaaggaca gcacctacag 300
cctgagcagc accctgaccc tgagcaaagc cgactacgag aagcacaagg tgtacgcctg 360
cgaggtgacc caccagggcc tgagctcccc cgtcaccaag agcttcaaca ggggggagtg 420
ttaggggccc gtttaaacgg gggaggcta 449
<210> 23
<211> 1132
<212> ДНК
<213> Человека
<400> 23
gcctccggac tctagagcca ccatgaaaca cctgtggttc ttcctcctgc tggtggcagc 60
tcccagatgg gtgctgagcc aggtgcaatt gtgcaggcgg ttagctcagc ctccaccaag 120
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 180
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 240
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 300
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 360
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 420
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 480
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 540
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 600
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 660
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 720
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 780
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 840
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 900
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 960
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1020
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1080
tccctgtctc ccggcaaatg agatatcggg cccgtttaaa cgggggaggc ta 1132
<210> 24
<211> 1398
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Химерное антитело
<400> 24
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctgg gggaggctta gtgcagcctg gaaggtccaa gaaactctcc 120
tgttcagcct caggattcac tttcagtaac tattacatgg cctgggtccg ccaggctcca 180
acgcagggtc tggagtgggt cgcatccatt ggtactgttg gtggtaacac ttactatcga 240
gactccgtga agggccgatt cactatctcc agagatgatg caaaaagcac cctatacctg 300
caaatggaca gtctgaggtc tgaggacacg gccacttatt actgtgcaag agaggattac 360
ggagggtttc cccactgggg ccaaggagtc atggtcacag tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaa 1398
<210> 25
<211> 466
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Химерное антитело
<400> 25
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Lys Lys Leu Ser Cys Ser Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Thr Gln Gly Leu
50 55 60
Glu Trp Val Ala Ser Ile Gly Thr Val Gly Gly Asn Thr Tyr Tyr Arg
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ala Lys Ser
85 90 95
Thr Leu Tyr Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Tyr Gly Gly Phe Pro His Trp Gly Gln
115 120 125
Gly Val Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 26
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Химерное антитело
<400> 26
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
aatattgtga tgactcagtc tcccacatcc atgttcatat cagtcggaga cagggtcacc 120
atgaactgta aggccagtca gaatgtggga actaatgtag actggtacca gcagaaaaca 180
gggcagtctc ctaaactgct tatctatggg gcgtccaacc gctacactgg agtccctgat 240
cgcttcacag gcagtggatc tggaacagat ttcactctca ccatcagcaa catgcaggct 300
gaagacctgg ctgtttatga ctgtctacag tataagtaca atccatacac gtttggaact 360
gggaccaagc tggaactgaa ccgggctgtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 27
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Химерное антитело
<400> 27
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asn Ile Val Met Thr Gln Ser Pro Thr Ser Met Phe
20 25 30
Ile Ser Val Gly Asp Arg Val Thr Met Asn Cys Lys Ala Ser Gln Asn
35 40 45
Val Gly Thr Asn Val Asp Trp Tyr Gln Gln Lys Thr Gly Gln Ser Pro
50 55 60
Lys Leu Leu Ile Tyr Gly Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp
65 70 75 80
Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Asn Met Gln Ala Glu Asp Leu Ala Val Tyr Asp Cys Leu Gln Tyr Lys
100 105 110
Tyr Asn Pro Tyr Thr Phe Gly Thr Gly Thr Lys Leu Glu Leu Asn Arg
115 120 125
Ala Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 28
<211> 1407
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Химерное антитело
<400> 28
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctga gggagtcagg acctggtctg gtgcagccct cacagaccct gtccctcacc 120
tgcactgtct ctgggttctc actaaccagc tttcatgtaa gctgggttcg ccagcctcca 180
gagaagggtc tggagtggat tgcaacaatt tcaagtggtg gaggtacata ttataattca 240
gctctcaaat cccgactgag catcagcagg gacacctcca agagccaagt tttcttaaag 300
atgagcactc tgcaaactga agacacagcc atgtacttct gtgcccggat ttcgggctgg 360
ggccattact atgttatgga tgtctggggt caaggagctt cagtcactgt cagctcagcc 420
tccaccaagg gcccaagcgt cttccccctg gcaccctcct ccaagagcac ctctggcggc 480
acagccgccc tgggctgcct ggtcaaggac tacttccccg aacccgtgac cgtgagctgg 540
aactcaggcg ccctgaccag cggcgtgcac accttccccg ctgtcctgca gtcctcagga 600
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 660
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagagagt tgagcccaaa 720
tcttgtgaca aaactcacac atgcccaccc tgcccagcac ctgaactcct ggggggaccc 780
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 840
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 900
gtggacggcg tggaggtgca taatgccaag acaaagcccc gggaggagca gtacaacagc 960
acgtaccggg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 1020
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1080
gccaaaggcc agccccggga accacaggtg tacaccctgc ccccatcccg ggaggagatg 1140
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1200
gtggagtggg agagcaatgg ccagcccgag aacaactaca agaccacccc tcccgtgctg 1260
gactccgacg gctccttctt cctctacagc aagctcaccg tggacaagag caggtggcag 1320
cagggcaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacccag 1380
aagagcctct ccctgtctcc cggcaaa 1407
<210> 29
<211> 469
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Химерное антитело
<400> 29
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Arg Glu Ser Gly Pro Gly Leu Val Gln
20 25 30
Pro Ser Gln Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu
35 40 45
Thr Ser Phe His Val Ser Trp Val Arg Gln Pro Pro Glu Lys Gly Leu
50 55 60
Glu Trp Ile Ala Thr Ile Ser Ser Gly Gly Gly Thr Tyr Tyr Asn Ser
65 70 75 80
Ala Leu Lys Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln
85 90 95
Val Phe Leu Lys Met Ser Thr Leu Gln Thr Glu Asp Thr Ala Met Tyr
100 105 110
Phe Cys Ala Arg Ile Ser Gly Trp Gly His Tyr Tyr Val Met Asp Val
115 120 125
Trp Gly Gln Gly Ala Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
210 215 220
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys
225 230 235 240
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
245 250 255
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
260 265 270
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
275 280 285
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
290 295 300
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
305 310 315 320
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
325 330 335
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
340 345 350
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
355 360 365
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
370 375 380
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
385 390 395 400
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
405 410 415
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
420 425 430
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
435 440 445
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
450 455 460
Leu Ser Pro Gly Lys
465
<210> 30
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Химерное антитело
<400> 30
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gacatccaga tgacacagtc tccagcttcc ctgtctggat ctctgggaga aactgtcacc 120
atccaatgtc aagcaagtga ggacatttac agtggtttag cgtggtatca gcagaagcca 180
gggaaatctc ctcagctcct gatctatggt gcaggtagct tacaagacgg cgtcccatca 240
cgattcagtg gcggtggatc tggcacacat tattctctca agatcagcag catgcaaact 300
gaagatgaag gggtttattt ctgtcaacag ggtttaaagt ttccgctcac gttcggttct 360
gggaccaagc tggagatcaa acgggctgtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 31
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Химерное антитело
<400> 31
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser
20 25 30
Gly Ser Leu Gly Glu Thr Val Thr Ile Gln Cys Gln Ala Ser Glu Asp
35 40 45
Ile Tyr Ser Gly Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro
50 55 60
Gln Leu Leu Ile Tyr Gly Ala Gly Ser Leu Gln Asp Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Gly Gly Ser Gly Thr His Tyr Ser Leu Lys Ile Ser
85 90 95
Ser Met Gln Thr Glu Asp Glu Gly Val Tyr Phe Cys Gln Gln Gly Leu
100 105 110
Lys Phe Pro Leu Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Ala Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 32
<211> 1398
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 32
atgaagcacc tgtggttctt tctgctgctg gtggccgctc ccagatgggt gctgtctgaa 60
gtgcagctgg tggaatccgg cggaggcctg gtgcagcctg gcggatctct gagactgtct 120
tgtgccgcct ccggcttcac cttctccaac tactacatgg cctgggtgcg acaggcccct 180
ggcaagggac tggaatgggt gtcctctatc ggcaccgtgg gcggcaacac ctactacgcc 240
gattctgtga agggccggtt caccatctcc cgggacgact ccaagaacac cctgtacctg 300
cagatgaact ccctgcgggc cgaggacacc gccgtgtact actgtgccag agaggactac 360
ggcggcttcc ctcattgggg ccagggcaca ctcgtgaccg tgtcctctgc ttccaccaag 420
ggcccctccg tgtttcctct ggccccttcc agcaagtcca cctctggcgg aacagccgct 480
ctgggctgcc tcgtgaagga ctacttcccc gagcccgtga cagtgtcttg gaactctggc 540
gccctgacct ccggcgtgca cacctttcca gctgtgctgc agtcctccgg cctgtactcc 600
ctgtcctccg tcgtgactgt gccctccagc tctctgggca cccagaccta catctgcaac 660
gtgaaccaca agccctccaa caccaaggtg gacaagcggg tggaacccaa gtcctgcgac 720
aagacccaca cctgtccccc ttgtcctgcc cctgaactgc tgggcggacc ttccgtgttc 780
ctgttccccc caaagcctaa ggacaccctg atgatctccc ggacccccga agtgacctgc 840
gtggtggtgg atgtgtccca cgaggaccct gaagtgaagt tcaattggta cgtggacggc 900
gtggaagtgc acaacgccaa gaccaagcct agagaggaac agtacaactc cacctaccgg 960
gtggtgtccg tgctgaccgt gctgcatcag gactggctga acggcaaaga gtacaagtgc 1020
aaggtgtcca acaaggccct gcctgccccc atcgaaaaga ccatctccaa ggccaagggc 1080
cagccccggg aaccccaggt gtacacactg ccccctagcc gggaagagat gaccaagaac 1140
caggtgtccc tgacctgtct cgtgaaaggc ttctacccct ccgatatcgc cgtggaatgg 1200
gagagcaacg gccagcccga gaacaactac aagaccaccc cccctgtgct ggactccgac 1260
ggctcattct tcctgtacag caagctgaca gtggacaagt cccggtggca gcagggcaac 1320
gtgttctcct gctccgtgat gcacgaggcc ctgcacaacc actacaccca gaagtccctg 1380
tccctgagcc ccggcaaa 1398
<210> 33
<211> 466
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 33
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ser Ser Ile Gly Thr Val Gly Gly Asn Thr Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Tyr Gly Gly Phe Pro His Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 34
<211> 1398
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 34
atgaagcacc tgtggttctt tctgctgctg gtggccgctc ccagatgggt gctgtctgaa 60
gtgcagctgg tggaatccgg cggaggcctg gtgcagcctg gcggatctct gagactgtct 120
tgtgccgcct ccggcttcac cttctccaac tactacatgg cctgggtgcg acaggcccct 180
ggcaagggac tggaatgggt ggcctctatc ggcaccgtgg gcggcaacac ctactaccgg 240
gattctgtga agggccggtt caccatctcc cgggacgact ccaagtccac cctgtacctg 300
cagatgaact ccctgcgggc cgaggacacc gccgtgtact actgtgccag agaggactac 360
ggcggcttcc ctcattgggg ccagggcaca ctcgtgaccg tgtcctctgc ttccaccaag 420
ggcccctccg tgtttcctct ggccccttcc agcaagtcta cctccggcgg aacagccgct 480
ctgggctgcc tcgtgaagga ctacttcccc gagcccgtga cagtgtcttg gaactctggc 540
gccctgacca gcggcgtgca cacctttcca gctgtgctgc agtcctccgg cctgtactcc 600
ctgtcctccg tcgtgactgt gccctccagc tctctgggca cccagaccta catctgcaac 660
gtgaaccaca agccctccaa caccaaggtg gacaagcggg tggaacccaa gtcctgcgac 720
aagacccaca cctgtccccc ttgtcctgcc cctgaactgc tgggcggacc ttccgtgttc 780
ctgttccccc caaagcccaa ggacaccctg atgatctccc ggacccccga agtgacctgc 840
gtggtggtgg atgtgtccca cgaggaccct gaagtgaagt tcaattggta cgtggacggc 900
gtggaagtgc acaacgccaa gaccaagcct agagaggaac agtacaactc cacctaccgg 960
gtggtgtccg tgctgaccgt gctgcatcag gactggctga acggcaaaga gtacaagtgc 1020
aaggtgtcca acaaggccct gcctgccccc atcgaaaaga ccatctccaa ggccaagggc 1080
cagccccggg aaccccaggt gtacacactg ccccctagcc gggaagagat gaccaagaac 1140
caggtgtccc tgacctgtct cgtgaaaggc ttctacccct ccgatatcgc cgtggaatgg 1200
gagtccaacg gccagcctga gaacaactac aagaccaccc cccctgtgct ggactccgac 1260
ggctcattct tcctgtacag caagctgaca gtggacaagt cccggtggca gcagggcaac 1320
gtgttctcct gctccgtgat gcacgaggcc ctgcacaacc actacaccca gaagtccctg 1380
tccctgagcc ccggcaaa 1398
<210> 35
<211> 466
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 35
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Ser Ile Gly Thr Val Gly Gly Asn Thr Tyr Tyr Arg
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Asp Tyr Gly Gly Phe Pro His Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 36
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 36
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcctacggc 60
aacatcgtga tgacccagtc ccccgactcc ctggctgtgt ctctgggcga gagagccacc 120
atcaactgca aggcctccca gaacgtgggc accaacgtgg actggtatca gcagaagccc 180
ggccagtccc ctaagctgct gatctacggc gccagcaacc ggtacaccgg cgtgcccgat 240
agattctccg gctctggctc tggcaccgac tttaccctga caatcagctc tctgcaggcc 300
gaggacgtgg ccgtgtacga ctgcctgcag tacaagtaca acccctacac cttcggccag 360
ggcacaaagg tggaaatcaa gcggaccgtg gccgctccct ccgtgtttat cttcccaccc 420
tccgacgagc agctgaagtc cggcacagct tccgtcgtgt gcctgctgaa caacttctac 480
ccccgcgagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg caactcccag 540
gaatccgtga ccgagcagga ctccaaggac agcacctact ccctgtcctc caccctgacc 600
ctgtccaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 660
ctgtctagcc ccgtgaccaa gtctttcaac cggggcgagt gc 702
<210> 37
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 37
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asn Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala
20 25 30
Val Ser Leu Gly Glu Arg Ala Thr Ile Asn Cys Lys Ala Ser Gln Asn
35 40 45
Val Gly Thr Asn Val Asp Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro
50 55 60
Lys Leu Leu Ile Tyr Gly Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Asp Cys Leu Gln Tyr Lys
100 105 110
Tyr Asn Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 38
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 38
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcctacggc 60
aacatcgtga tgacccagtc cccctccagc ctgtctgctt ccgtgggcga cagagtgacc 120
atcaactgca aggcctccca gaacgtgggc accaacgtgg actggtatca gcagaagccc 180
ggcaagtccc ccaagctgct gatctacggc gccagcaaca gatacaccgg cgtgcccgac 240
agattctccg gctctggctc tggcaccgac tttaccctga ccatcagctc cctgcagccc 300
gaggacttcg ccacctacga ctgcctgcag tacaagtaca acccctacac cttcggccag 360
ggcacaaagg tggaaatcaa gcggaccgtg gccgctccct ccgtgtttat cttcccaccc 420
tccgacgagc agctgaagtc cggcacagct tctgtcgtgt gcctgctgaa caacttctac 480
ccccgcgagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg caactcccag 540
gaatccgtga ccgagcagga ctccaaggac agcacctact ccctgtcctc caccctgacc 600
ctgtccaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 660
ctgtctagcc ccgtgaccaa gtctttcaac cggggcgagt gc 702
<210> 39
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 39
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asn Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Asn Cys Lys Ala Ser Gln Asn
35 40 45
Val Gly Thr Asn Val Asp Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro
50 55 60
Lys Leu Leu Ile Tyr Gly Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Asp Cys Leu Gln Tyr Lys
100 105 110
Tyr Asn Pro Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 40
<211> 1407
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 40
atgaagcacc tgtggttctt tctgctgctg gtggccgctc ccagatgggt gctgtctgaa 60
gtgcagctgg tggaatccgg cggaggcctc gtgaagcctt cccagaccct gtctctgacc 120
tgcaccgtgt ccggcttctc cctgacctcc ttccacgtgt catgggtgcg acagcctcca 180
ggcaagggcc tggaatggat cgccaccatc tcctctggcg gcggaaccta ctacaacccc 240
agcctgaagt ccagagtgac catctcccgg gacacctcca agaaccaggt gtccctgaag 300
ctgtcctccg tgaccgccgc tgataccgcc gtgtactact gcgccagaat ctccggctgg 360
ggccactact acgtgatgga cgtgtggggc cagggcaccc tcgtgacagt gtcctctgct 420
tccaccaagg gcccctccgt gtttcctctg gccccttcca gcaagtctac ctccggcgga 480
acagccgctc tgggctgcct cgtgaaagac tacttccccg agcccgtgac cgtgtcttgg 540
aactctggcg ctctgaccag cggcgtgcac acctttccag ctgtgctgca gtcctccggc 600
ctgtactccc tgtccagcgt cgtgactgtg ccctccagct ctctgggcac ccagacctac 660
atctgcaacg tgaaccacaa gccctccaac accaaggtgg acaagcgggt ggaacccaag 720
tcctgcgaca agacccacac ctgtccccct tgtcctgccc ctgaactgct gggcggacct 780
tccgtgttcc tgttcccccc aaagcccaag gacaccctga tgatctcccg gacccccgaa 840
gtgacctgcg tggtggtgga tgtgtcccac gaggaccctg aagtgaagtt caattggtac 900
gtggacggcg tggaagtgca caacgccaag accaagccta gagaggaaca gtacaactcc 960
acctaccggg tggtgtccgt gctgaccgtg ctgcaccagg attggctgaa cggcaaagag 1020
tacaagtgca aggtgtccaa caaggccctg cctgccccca tcgaaaagac catcagcaag 1080
gccaagggcc agccccggga accccaggtg tacacactgc cccctagccg ggaagagatg 1140
acaaaaaatc aggtgtcact gacctgtctc gtgaagggct tctacccctc cgatatcgcc 1200
gtggaatggg agtccaacgg ccagcctgag aacaactaca agaccacccc ccctgtgctg 1260
gactccgacg gctcattctt cctgtacagc aagctgacag tggacaagtc ccggtggcag 1320
cagggcaacg tgttctcctg ctccgtgatg cacgaggccc tgcacaacca ctacacccag 1380
aagtccctgt ccctgagccc cggcaaa 1407
<210> 41
<211> 469
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 41
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys
20 25 30
Pro Ser Gln Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu
35 40 45
Thr Ser Phe His Val Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Ala Thr Ile Ser Ser Gly Gly Gly Thr Tyr Tyr Asn Pro
65 70 75 80
Ser Leu Lys Ser Arg Val Thr Ile Ser Arg Asp Thr Ser Lys Asn Gln
85 90 95
Val Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr
100 105 110
Tyr Cys Ala Arg Ile Ser Gly Trp Gly His Tyr Tyr Val Met Asp Val
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
210 215 220
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys
225 230 235 240
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
245 250 255
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
260 265 270
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
275 280 285
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
290 295 300
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
305 310 315 320
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
325 330 335
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
340 345 350
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
355 360 365
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
370 375 380
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
385 390 395 400
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
405 410 415
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
420 425 430
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
435 440 445
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
450 455 460
Leu Ser Pro Gly Lys
465
<210> 42
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 42
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcctacggc 60
gacatccaga tgacccagag cccttccagc ctgtccgctt ccgtgggcga cagagtgacc 120
atcacctgtc aggcctccga ggacatctac tccggcctgg cctggtatca gcagaagccc 180
ggcaagtccc ccaagctgct gatctacggc gctggatctc tgcaggacgg cgtgccctct 240
agattctccg gctctggatc cggcacccac tacaccctga ccatctccag cctgcagccc 300
gaggacttcg ctacctactt ctgtcagcaa ggcctgaagt tccccctgac cttcggccag 360
ggcaccaagg tggaaatcaa gcggaccgtg gccgctccct ccgtgtttat cttcccaccc 420
tccgacgagc agctgaagtc cggcacagct tctgtcgtgt gcctgctgaa caacttctac 480
ccccgcgagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg caactcccag 540
gaatccgtga ccgagcagga ctccaaggac agcacctact ccctgtcctc taccctgacc 600
ctgtccaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 660
ctgtctagcc ccgtgaccaa gtctttcaac cggggcgagt gc 702
<210> 43
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Гуманизированное антитело
<400> 43
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Glu Asp
35 40 45
Ile Tyr Ser Gly Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro
50 55 60
Lys Leu Leu Ile Tyr Gly Ala Gly Ser Leu Gln Asp Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr His Tyr Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Gly Leu
100 105 110
Lys Phe Pro Leu Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 44
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер F
<400> 44
cacggcaacc tgctggagcg gctgctgggg gagg 34
<210> 45
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер R
<400> 45
caggctgttc ccagacaggt ccag 24
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МУЛЬТИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ОБЛАДАЮЩАЯ ЗАМЕЩАЮЩЕЙ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТЬЮ КОФАКТОРА КОАГУЛИРУЮЩЕГО ФАКТОРА КРОВИ VIII, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ УКАЗАННУЮ МОЛЕКУЛУ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА | 2018 |
|
RU2812909C2 |
| АНТИТЕЛО ПРОТИВ SIRPα | 2019 |
|
RU2791002C2 |
| АНТИТЕЛО ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2781148C2 |
| АНТИ-GARP-TGF-β-АНТИТЕЛА | 2018 |
|
RU2767784C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2021 |
|
RU2840407C1 |
| АНТИ-ORAI1 АНТИТЕЛО | 2015 |
|
RU2724742C2 |
| АНТИТЕЛО ПРОТИВ GPR20 И КОНЪЮГАТЫ АНТИТЕЛО ПРОТИВ GPR20-ЛЕКАРСТВЕННОЕ СРЕДСТВО | 2018 |
|
RU2772804C2 |
| Прогнозирование риска развития нежелательной реакции, связанной с введением антитела к ALK2, и способности отвечать на лечение антителом к ALK2 | 2019 |
|
RU2783762C2 |
| АНТИТЕЛО ПРОТИВ CD147 | 2018 |
|
RU2785293C2 |
| АНТИТЕЛО ПРОТИВ ЧЕЛОВЕЧЕСКОГО TLR7 | 2019 |
|
RU2808123C2 |







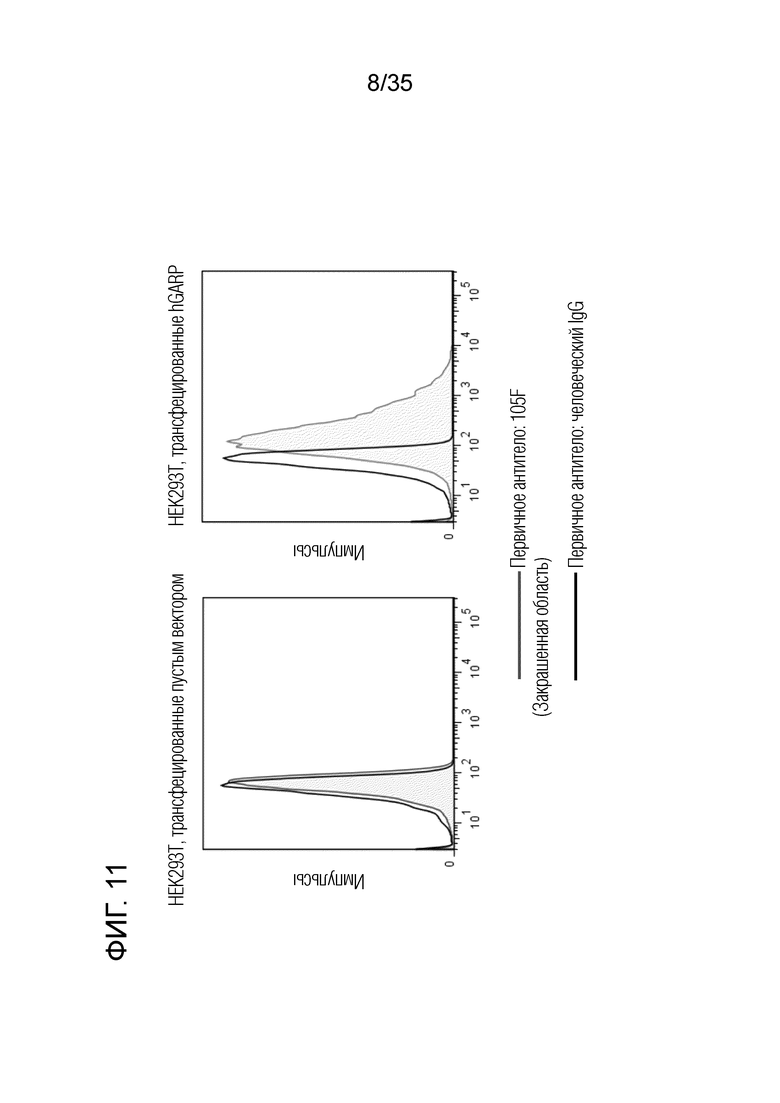


















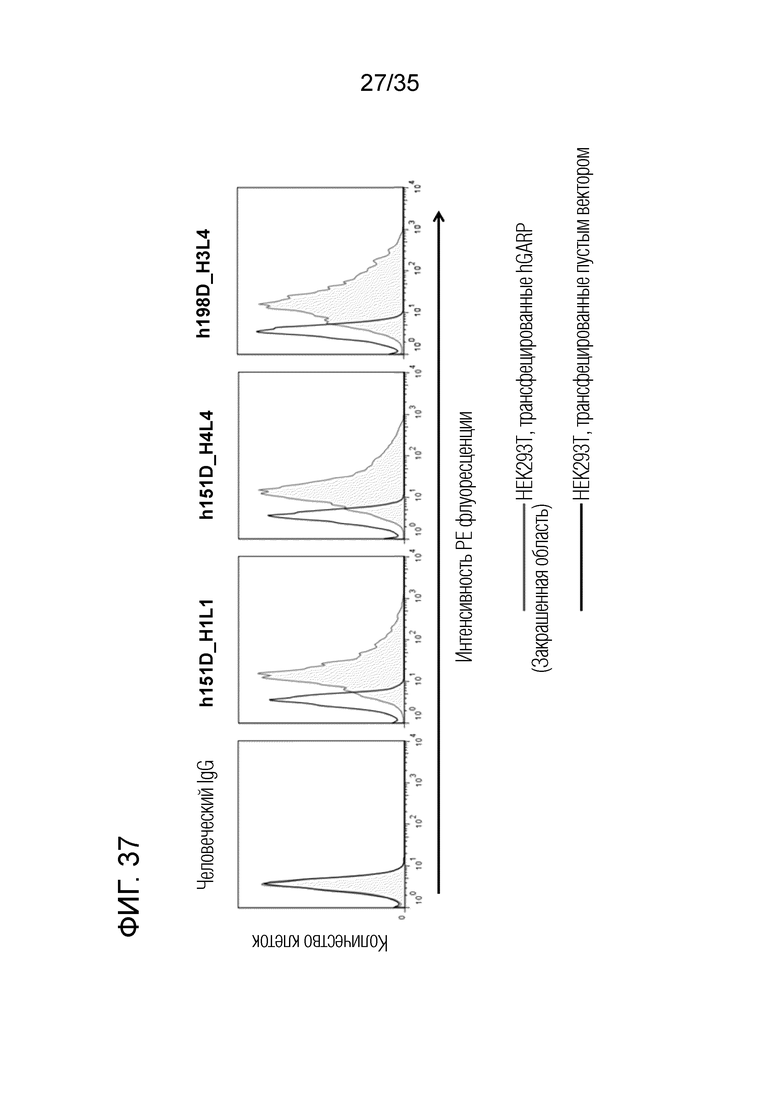

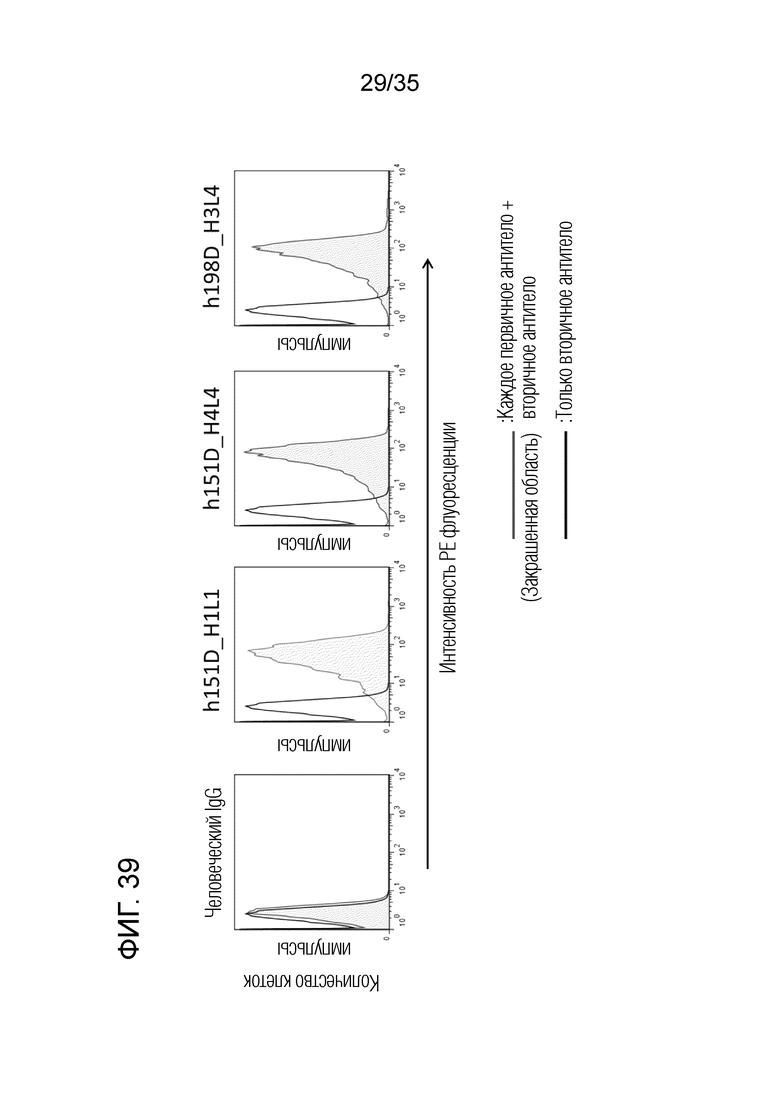



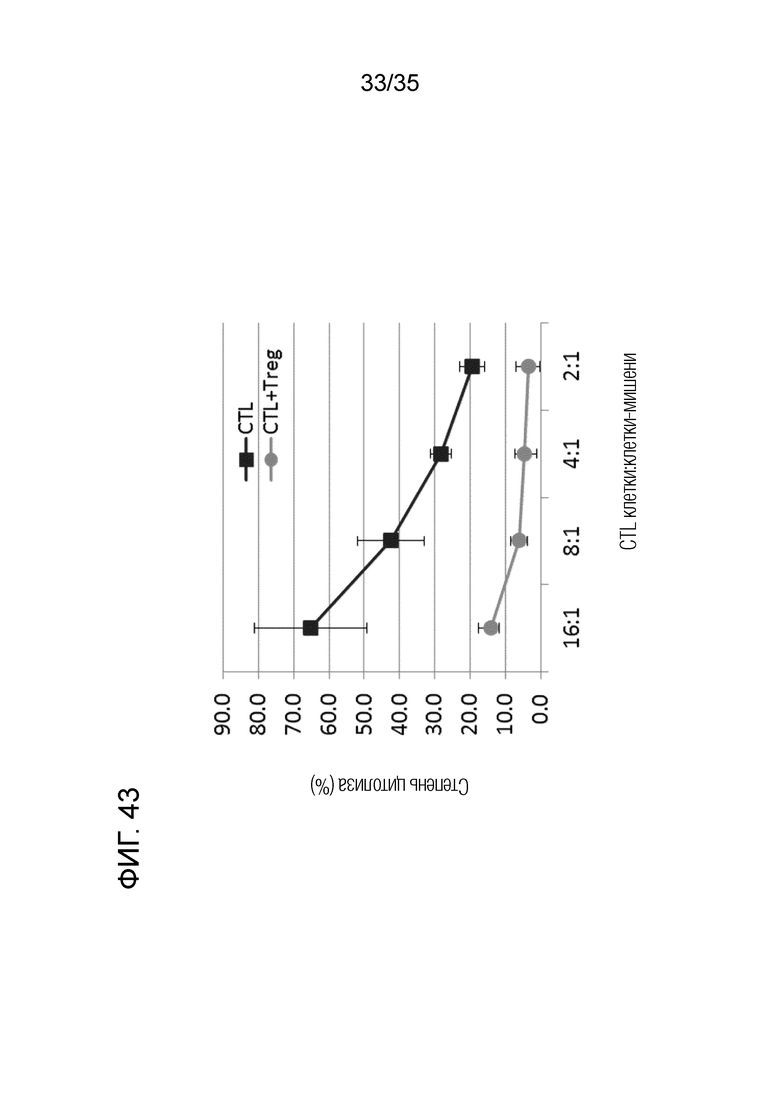
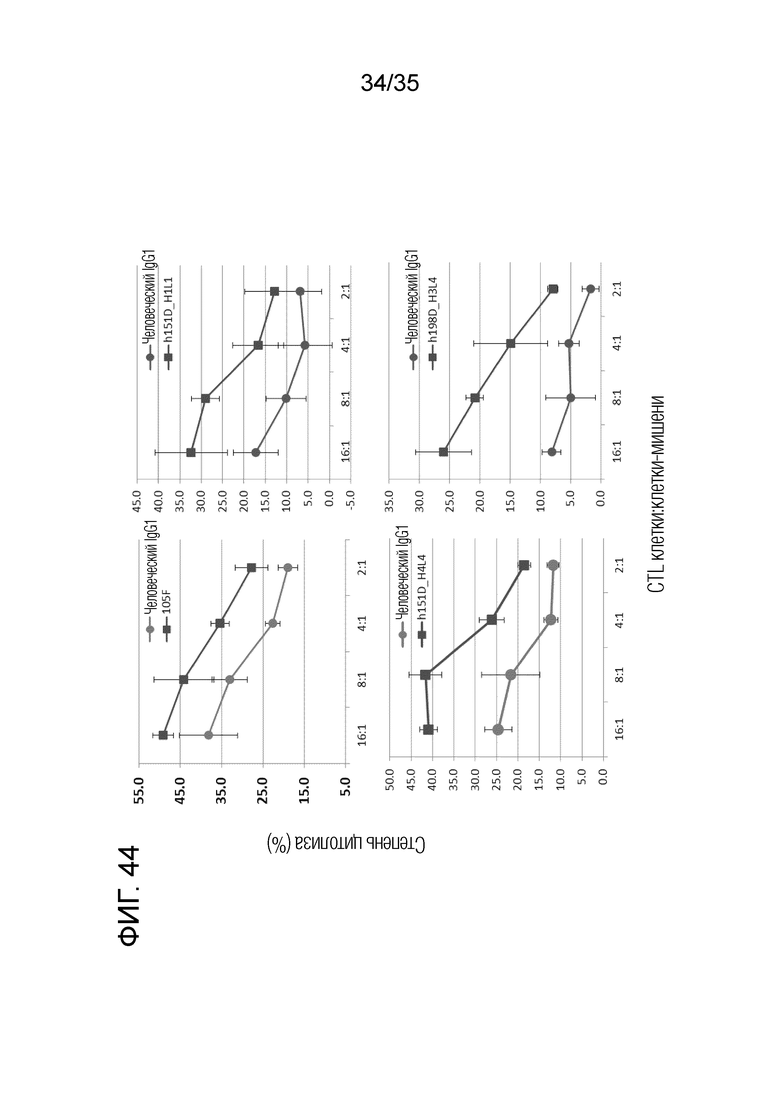
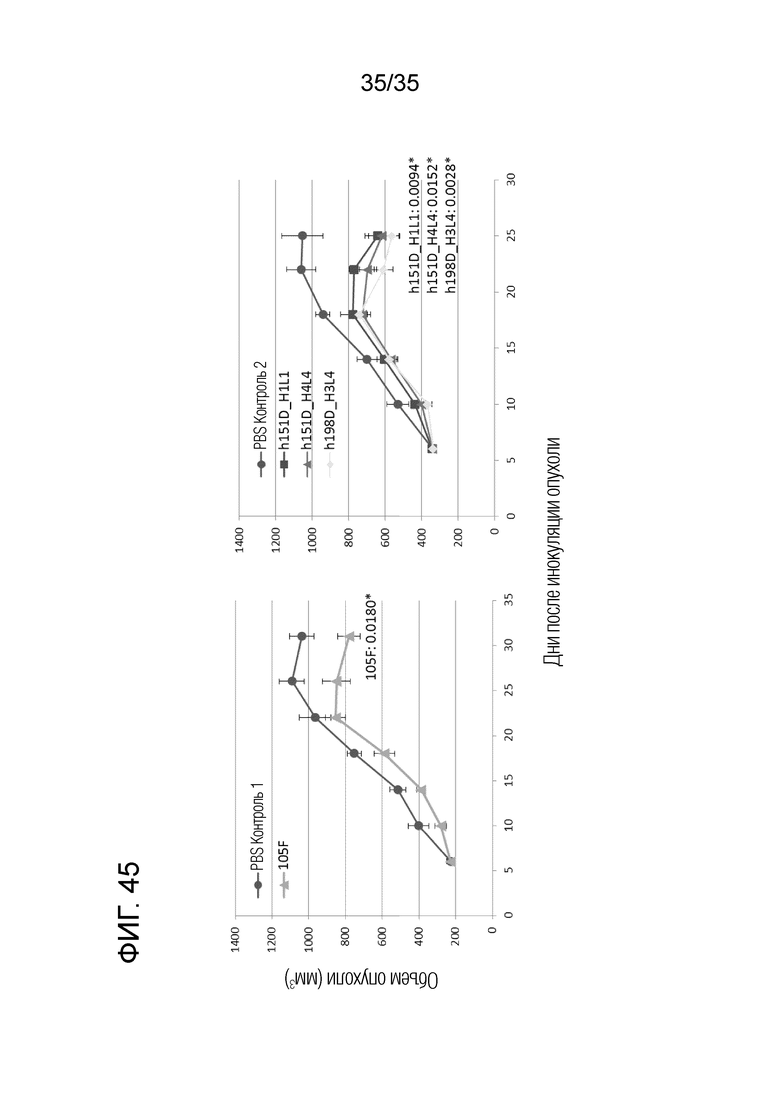
Изобретение относится к области биотехнологии. Предложены варианты антитела, которое специфически связывается с GARP. Также предложены полинуклеотид, кодирующий указанное антитело, включающие указанный полинуклеотид вектор экспрессии и клетка-хозяин, способ получения указанного антитела. Кроме того, изобретение относится к фармацевтической композиции для применения в противоопухолевой терапии и способу лечения опухоли с использованием вышеуказанного антитела. Изобретение обеспечивает получение антитела против GARP, которое проявляет ингибирующую активность в отношении функции Treg, проявляет активность ADCC и может быть использовано в качестве терапевтического противоопухолевого агента. 8 н. и 29 з.п. ф-лы, 45 ил., 2 табл., 11 пр.
1. Антитело, специфически связывающееся с преобладающими повторами гликопротеина-A (GARP), которое имеет:
(1) CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 26-35 последовательности SEQ ID NO: 2, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 50-66 последовательности SEQ ID NO: 2, и CDRH3, состоящую из аминокислотной последовательности в положениях аминокислот 99-107 последовательности SEQ ID NO: 2, и CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 23-36 последовательности SEQ ID NO: 3, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 52-58 последовательности SEQ ID NO: 3, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 91-101 последовательности SEQ ID NO: 3,
(2) CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 26-35 последовательности SEQ ID NO: 4, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 50-66 последовательности SEQ ID NO: 4, и CDRH3, состоящую из аминокислотной последовательности в положениях аминокислот 99-112 последовательности SEQ ID NO: 4, и CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 23-36 последовательности SEQ ID NO: 5, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 52-58 последовательности SEQ ID NO: 5, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 91-100 последовательности SEQ ID NO: 5,
(3) CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 45-54 последовательности SEQ ID NO: 25, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 69-78 последовательности SEQ ID NO: 25, и CDRH3, состоящую из аминокислотной последовательности в положениях аминокислот 118-125 последовательности SEQ ID NO: 25, и CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 44-54 последовательности SEQ ID NO: 27, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 70-76 последовательности SEQ ID NO: 27, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 109-117 последовательности SEQ ID NO: 27, или
(4) CDRH1, состоящую из аминокислотной последовательности в положениях аминокислот 45-54 последовательности SEQ ID NO: 29, CDRH2, состоящую из аминокислотной последовательности в положениях аминокислот 69-77 последовательности SEQ ID NO: 29, и CDRH3, состоящую из аминокислотной последовательности в положениях аминокислот 117-128 последовательности SEQ ID NO: 29, и CDRL1, состоящую из аминокислотной последовательности в положениях аминокислот 44-54 последовательности SEQ ID NO: 31, CDRL2, состоящую из аминокислотной последовательности в положениях аминокислот 70-76 последовательности SEQ ID NO: 31, и CDRL3, состоящую из аминокислотной последовательности в положениях аминокислот 109-117 последовательности SEQ ID NO: 31.
2. Антитело по п.1, которое имеет ингибирующую активность в отношении иммунодепрессивной функции регуляторных Т-клеток.
3. Антитело по п.1 или 2, которое имеет активность антителозависимую клеточную цитотоксическую (ADCC).
4. Антитело по любому из пп.1-3, которое имеет in vivo противоопухолевую активность.
5. Антитело по любому из пп.1-4, где опухоль представляет собой злокачественное новообразование.
6. Антитело по п.5, где злокачественное новообразование представляет собой рак легких, рак почек, уротелиальный рак, рак толстой кишки, рак предстательной железы, мультиформную глиобластому, рак яичников, рак поджелудочной железы, рак молочной железы, меланому, рак печени, рак мочевого пузыря, рак желудка, рак пищевода или рак крови.
7. Антитело по любому из пп.1-6, которое имеет:
(1) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 1-118 SEQ ID NO: 2, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 1-112 SEQ ID NO: 3,
(2) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 1-123 SEQ ID NO: 4, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 1-111 SEQ ID NO: 5,
(3) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 20-136 SEQ ID NO: 25, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 21-129 SEQ ID NO: 27, или
(4) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 20-139 SEQ ID NO: 29, и вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 21-129 SEQ ID NO: 31.
8. Антитело по любому из пп.1-7, где константная область представляет собой константную область человеческого происхождения.
9. Антитело по любому из пп.1-8, которое имеет:
(1) тяжелую цепь, состоящую из аминокислотной последовательности SEQ ID NO: 2, и легкую цепь, состоящую из аминокислотной последовательности SEQ ID NO: 3,
(2) тяжелую цепь, состоящую из аминокислотной последовательности SEQ ID NO: 4, и легкую цепь, состоящую из аминокислотной последовательности SEQ ID NO: 5,
(3) тяжелую цепь, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 20-446 SEQ ID NO: 25, и легкую цепь, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 21-234 SEQ ID NO: 27, или
(4) тяжелую цепь, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 20-469 SEQ ID NO: 29, и легкую цепь, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 21-234 SEQ ID NO: 31.
10. Антитело по любому из пп.1-9, которое является гуманизированным.
11. Антитело по п.10, которое имеет:
вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, выбранной из группы, состоящей из:
(а) аминокислотной последовательности, соответствующей положениям аминокислот 20-136 SEQ ID NO: 33,
(b) аминокислотной последовательности, соответствующей положениям аминокислот 20-136 SEQ ID NO: 35,
(с) аминокислотной последовательности, соответствующей положениям аминокислот 20-139 SEQ ID NO: 41,
(d) аминокислотной последовательности, имеющей гомологию, составляющую по меньшей мере 95% или более, с последовательностью каркасной области, отличающейся от каждой последовательности CDR в последовательностях (а)-(с), и
(e) аминокислотной последовательности, содержащей делецию, замену или вставку одной или нескольких аминокислот в последовательности каркасной области, отличающейся от каждой последовательности CDR в последовательностях (a)-(c), и
вариабельную область легкой цепи, состоящую из аминокислотной последовательности, выбранной из группы, состоящей из:
(f) аминокислотной последовательности, соответствующей положениям аминокислот 21-129 SEQ ID NO: 37,
(g) аминокислотной последовательности, соответствующей положениям аминокислот 21-129 SEQ ID NO: 39,
(h) аминокислотной последовательности, соответствующей положениям аминокислот 21-129 SEQ ID NO: 43,
(i) аминокислотной последовательности, имеющей гомологию по меньшей мере 95% или более, с последовательностью каркасной области, отличающейся от каждой последовательности CDR в последовательностях (f)-(h), и
(j) аминокислотной последовательности, содержащей делецию, замену или вставку одной или нескольких аминокислот в последовательности каркасной области, отличающейся от каждой последовательности CDR в последовательностях (f)-(h).
12. Антитело по п.10 или 11, которое имеет:
(1) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 20-136 SEQ ID NO: 33, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 21-129 SEQ ID NO: 37,
(2) вариабельную области тяжелой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 20-136 SEQ ID NO: 35, и вариабельную область легкой цепи, состоящей из аминокислотной последовательности, соответствующей положениям аминокислот 21-129 SEQ ID NO: 39, или
(3) вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 20-139 SEQ ID NO: 41, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности, соответствующей положениям аминокислот 21-129 SEQ ID NO: 43.
13. Антитело по любому из пп.10-12, которое имеет:
(1) тяжелую цепь, выбранную из группы, состоящей из тяжелой цепи, имеющей аминокислотную последовательность в положениях аминокислот 20-446 SEQ ID NO: 33, тяжелую цепь, имеющую аминокислотную последовательность в положениях аминокислот 20-466 SEQ ID NO: 35, и тяжелую цепь, имеющую аминокислотную последовательность в положениях аминокислот 20-469 SEQ ID NO: 41, и
(2) легкую цепь, выбранную из группы, состоящей из легкой цепи, имеющей аминокислотную последовательность в положениях аминокислот 21-234 SEQ ID NO: 37, легкую цепь, имеющую аминокислотную последовательность в положениях аминокислот 21-234 SEQ ID NO: 39, и легкую цепь, имеющую аминокислотную последовательность в положениях аминокислот 21-234 SEQ ID NO: 43.
14. Антитело по любому из пп.10-13, которое имеет:
(1) тяжелую цепь, имеющую аминокислотную последовательность в положениях аминокислот 20-446 SEQ ID NO: 33, и легкую цепь, имеющую аминокислотную последовательность в положениях аминокислот 21-234 SEQ ID NO: 37,
(2) тяжелую цепь, имеющую аминокислотную последовательность в положениях аминокислот 20-466 SEQ ID NO: 35, и легкую цепь, имеющую аминокислотную последовательность в положениях аминокислот 21-234 SEQ ID NO: 39, или
(3) тяжелую цепь, имеющую аминокислотную последовательность в положениях аминокислот 20-469 SEQ ID NO: 41, и легкую цепь, имеющую аминокислотную последовательность в положениях аминокислот 21-234 SEQ ID NO: 43.
15. Полинуклеотид, кодирующий антитело по любому из пп.1-14.
16. Полинуклеотид по п.15, который имеет:
(1) полинуклеотид CDRH1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 76-105 SEQ ID NO: 6, полинуклеотид CDRH2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 148-198 SEQ ID NO: 6, и полинуклеотид CDRH3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 295-321 SEQ ID NO: 6, и полинуклеотид CDRL1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 67-108 SEQ ID NO: 7, полинуклеотид CDRL2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 154-174 SEQ ID NO: 7, и полинуклеотид CDRL3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 271-303 SEQ ID NO: 7,
(2) полинуклеотид CDRH1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 76-105, представленной в SEQ ID NO: 8, полинуклеотид CDRH2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 148-198 SEQ ID NO: 8, и полинуклеотид CDRH3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 295-336 SEQ ID NO: 8, и полинуклеотид CDRL1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 67-108 SEQ ID NO: 9, полинуклеотид CDRL2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 154-174 SEQ ID NO: 9, и полинуклеотид CDRL3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 271-300 SEQ ID NO: 9,
(3) полинуклеотид CDRH1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 133-162 SEQ ID NO: 24, полинуклеотид CDRH2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 205-234 SEQ ID NO: 24, и полинуклеотид CDRH3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 352-375 SEQ ID NO: 24, и полинуклеотид CDRL1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 130-162 SEQ ID NO: 26, полинуклеотид CDRL2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 208-228 SEQ ID NO: 26, и полинуклеотид CDRL3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 325-351 SEQ ID NO: 26, или
(4) полинуклеотид CDRH1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 133-162 SEQ ID NO: 28, полинуклеотид CDRH2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 205-231 SEQ ID NO: 28, и полинуклеотид CDRH3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 349-384 SEQ ID NO: 28, и полинуклеотид CDRL1, состоящий из нуклеотидной последовательности в положениях нуклеотидов 130-162 SEQ ID NO: 30, полинуклеотид CDRL2, состоящий из нуклеотидной последовательности в положениях нуклеотидов 208-228 SEQ ID NO: 30, и полинуклеотид CDRL3, состоящий из нуклеотидной последовательности в положениях нуклеотидов 325-351 SEQ ID NO: 30.
17. Полинуклеотид по п.15 или 16, который имеет:
(1) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 1-354 SEQ ID NO: 6, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 1-336 SEQ ID NO: 7,
(2) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 1-369 SEQ ID NO: 8, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 1-333 SEQ ID NO: 9,
(3) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-408 SEQ ID NO: 24, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-387, представленной в SEQ ID NO: 26, или
(4) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-417 SEQ ID NO: 28, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-387 SEQ ID NO: 30.
18. Полинуклеотид по любому из пп.15-17, который имеет:
(1) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности SEQ ID NO: 6, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности SEQ ID NO: 7,
(2) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности SEQ ID NO: 8, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности SEQ ID NO: 9,
(3) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-1398 SEQ ID NO: 24, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-702 SEQ ID NO: 26, или
(4) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-1407 SEQ ID NO: 28, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-702 SEQ ID NO: 30.
19. Полинуклеотид по любому из пп.15 или 16, который имеет:
(1) полинуклеотид вариабельной области тяжелой цепи, выбранный из группы, состоящей из полинуклеотида вариабельной области тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-408 SEQ ID NO: 32, полинуклеотида вариабельной области тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-408 SEQ ID NO: 34, и полинуклеотида вариабельной области тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-417 SEQ ID NO: 40, и
(2) полинуклеотид вариабельной области легкой цепи, выбранный из группы, состоящей из полинуклеотида вариабельной области легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-387 SEQ ID NO: 36, полинуклеотида вариабельной области легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-387 SEQ ID NO: 38, и полинуклеотида вариабельной области легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-387 SEQ ID NO: 42.
20. Полинуклеотид по пп.15, 16 или 19, который имеет:
(1) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-408 SEQ ID NO: 32, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-387 SEQ ID NO: 36,
(2) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-408 SEQ ID NO: 34, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-387 SEQ ID NO: 38, или
(3) полинуклеотид вариабельной области тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-417 SEQ ID NO: 40, и полинуклеотид вариабельной области легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-387 SEQ ID NO: 42.
21. Полинуклеотид по пп.15, 16, 19 или 20, который имеет:
(1) полинуклеотид тяжелой цепи, выбранный из группы, состоящей из полинуклеотида тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-1398 SEQ ID NO: 32, полинуклеотида тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-1398 SEQ ID NO: 34, и полинуклеотида тяжелой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 58-1407 SEQ ID NO: 40, и
(2) полинуклеотид легкой цепи, выбранный из группы, состоящей из полинуклеотида легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-702 SEQ ID NO: 36, полинуклеотида легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-702 SEQ ID NO: 38, и полинуклеотида легкой цепи, состоящего из нуклеотидной последовательности в положениях нуклеотидов 61-702 SEQ ID NO: 42.
22. Полинуклеотид по пп.15, 16 и 19-21, который имеет:
(1) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-1398 SEQ ID NO: 32, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-702 SEQ ID NO: 36,
(2) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-1398 SEQ ID NO: 34, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-702 SEQ ID NO: 38, или
(3) полинуклеотид тяжелой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 58-1407 SEQ ID NO: 40, и полинуклеотид легкой цепи, состоящий из нуклеотидной последовательности в положениях нуклеотидов 61-702 SEQ ID NO: 42.
23. Вектор экспрессии, содержащий полинуклеотид по любому из пп.15-22.
24. Клетка-хозяин, трансформированная экспрессирующим вектором по п.23, которая предназначена для получения антитела по пп.1-12.
25. Способ получения интересующего антитела по любому из пп.1-14, который включает стадию культивирования клеток-хозяев по п.24 и стадию сбора интересующего антитела из культуры, полученной на вышеуказанной стадии.
26. Антитело, специфически связывающееся с преобладающими повторами гликопротеина-A (GARP) и полученное способом получения по п.25.
27. Антитело по любому из пп.1-14 и 26, содержащее одну, или две, или более модификаций, выбранных из группы, состоящей из N-связанного гликозилирования, O-связанного гликозилирования, N-концевого процессирования, C-концевого процессирования, дезамидирования, изомеризации аспарагиновой кислоты, окисления метионина, добавления остатка метионина к N-концу, амидирования остатка пролина и тяжелой цепи, содержащей делецию одной или двух аминокислот на карбоксильном конце.
28. Антитело по п.27, где одна или две аминокислоты делетированы на карбоксильном конце его тяжелой цепи.
29. Антитело по п.28, где одна аминокислота делетирована на каждом из карбоксильных концов его обеих тяжелых цепей.
30. Антитело по любому из пп.27-29, где остаток пролина на карбоксильном конце его тяжелой цепи дополнительно амидирован.
31. Антитело по любому из пп.1-14 и 26-30, где модификация сахарной цепи регулируется для усиления антителозависимой клеточной цитотоксичности.
32. Фармацевтическая композиция для применения в противоопухолевой терапии, содержащая по меньшей мере одно из антител по пп.1-14 и 26-31.
33. Фармацевтическая композиция по п.32, где опухоль представляет собой злокачественное новообразование.
34. Фармацевтическая композиция по п.33, где злокачественное новообразование представляет собой рак легких, рак почек, уротелиальный рак, рак толстой кишки, рак предстательной железы, мультиформную глиобластому, рак яичников, рак поджелудочной железы, рак молочной железы, меланому, рак печени, рак мочевого пузыря, рак желудка, рак пищевода или гемобластоз.
35. Способ лечения опухоли, который включает введение индивидууму по меньшей мере одного из антител по пп.1-14 и 26-31.
36. Способ лечения по п.35, где опухоль представляет собой злокачественное новообразование.
37. Способ лечения по п.36, где злокачественное новообразование представляет собой рак легкого, рак почек, уротелиальный рак, рак толстой кишки, рак предстательной железы, мультиформную глиобластому, рак яичников, рак поджелудочной железы, рак молочной железы, меланому, рак печени, рак мочевого пузыря, рак желудка, рак пищевода или рак крови.
| WO 2015015003 A1, 05.02.2015 | |||
| CUENDE J | |||
| et al., Monoclonal antibodies against GARP/TGF-b1 complexes inhibit the immunosuppressive activity of human regulatory T cells in vivo, SCIENCE TRANSLATIONAL MEDICINE, 22 April 2015, Vol.7, Issue 284, 284ra56, стр.1-12 | |||
| US 2013078234 A1, 28.03.2013 | |||
| ЖУЛАЙ Г.А., ОЛЕЙНИК Е.К., Регуляторные Т-лимфоциты |

