Область техники, к которой относится изобретение
Настоящее изобретение относится к веществу, применимому в качестве терапевтического и/или профилактического средства при аномальном метаболизме костной ткани, способу скрининга вещества, применимого в качестве терапевтического и/или профилактического средства при аномальном метаболизме костной ткани, способу выявления аномального метаболизма костной ткани и способу лечения и/или профилактики аномального метаболизма костной ткани.
Предпосылки к созданию изобретения
Известно, что кость является динамичным органом, который непрерывно реконструируется посредством многократного образования и резорбции, для того, чтобы менять свою собственную морфологию и поддерживать уровни кальция в крови. В здоровой кости поддерживается равновесие между образованием посредством остеобластов и резорбцией кости посредством остеокластов, и костная масса поддерживается постоянной. Напротив, в тех случаях, когда равновесие между формированием кости и резорбцией кости теряется, происходит аномальный метаболизм костной ткани, например остеопороз (Endocrinological Review, (1992) 13, pp. 66-80, Principles of Bone Biology, Academic Press, New York, (1996) pp. 87-102).
Сообщалось о многих системных гормонах и локальных цитокинах в качестве факторов, которые регулируют метаболизм костной ткани, и эти факторы взаимодействуют друг с другом для образования и поддержания костной ткани (Endocrinological Review, (1992) 13, pp. 66-80, Endocrinological Review, (1996) 17, pp. 308-332). Появление остеопороза как изменения костной ткани вследствие старения широко распространено, но механизм его появления охватывает различные факторы, такие как снижение секреции половых гормонов и аномалии рецепторов к этим гормонам, изменения экспрессии местных цитокинов в костной ткани, экспрессия генов старения и нарушения или дисфункции дифференцировки остеокластов или остеобластов, и, следовательно, трудно рассматривать это просто как связанный с возрастом физиологический феномен. Первичный остеопороз главным образом делится на постменопаузный остеопороз вследствие снижения секреции эстрогена и сенильный остеопороз вследствие старения, но успех фундаментальных исследований механизмов регуляции формирования костной ткани и резорбции костной ткани является необходимым для выяснения механизма его появления и для разработки для него терапевтического средства.
Остеокласты представляют собой многоядерные клетки, происходящие из гематопоэтических стволовых клеток, и посредством высвобождения хлорид-ионов и ионов водорода на поверхность кости, на которой адгезируются остеокласты, остеокласты повышают кислотность промежутка между поверхностью кости и остеокластами, а также секретируют катепсин К, который является кислой протеазой, или подобное (American Journal of Physiology, (1991) 260, C1315-C1324). Это вызывает разрушение фосфата кальция, активацию кислых протеаз и разрушение белков костного матрикса, приводя в результате к резорбции костной ткани.
Было обнаружено, что клетки-предшественники остеокластов дифференцируются в остеокласты под действием стимуляции RANKL (рецептора активатора лиганда NF-кB), экспрессирующегося на клеточной мембране остеобластов/стромальных клеток, находящихся на поверхности кости (Proceedings of the National Academy of Science of the United States of America, (1998) 95, pp. 3597-3602, Cell, (1998) 93, pp. 165-176). Было обнаружено, что RANKL представляет собой мембранный белок, продуцируемый остеобластами/стромальными клетками, его экспрессия регулируется фактором резорбции кости, RANKL индуцирует дифференцировку клеток-предшественников остеокластов в многоядерные остеокласты, и подобное (Proceedings of the National Academy of Science of the United States of America, (1998) 95, pp. 3597-3602, Journal of Bone and Mineral Research, (1998) 23, S222). Далее, было обнаружено, что у мышей «нокаут», лишенных RANKL, развивается заболевание, подобное остеопорозу, и, таким образом, доказано, что RANKL является физиологическим фактором, индуцирующим дифференцировку остеокластов (Nature, (1999) 397, pp. 315-323).
В качестве лекарственных средств для лечения заболеваний метаболизма костной ткани или сокращения продолжительности лечения используют бисфосфонаты, активный витамин D3, кальцитонин и его производные, гормональные препараты, такие как эстрадиол, SERM (селективные модуляторы рецепторов эстрогена), иприфлавон, витамин K2 (менатетренон), препараты PTH (паратиреоидного гормона), препараты кальция и подобное. Однако эти лекарственные средства не всегда демонстрируют удовлетворительный терапевтический эффект, и требовалась разработка средства с более мощным терапевтическим действием.
Клеточные мембраны иммунных клеток покрыты плотной оболочкой различных гликанов, таких как сиаловая кислота, которые распознаются различными гликан-связывающими белками. Иммуноглобулин-подобные лектины, связывающие сиаловые кислоты (далее называемые «siglec»), представляют собой семейство мембранных белков I типа, которые распознают сиалилированные гликаны и связываются с ними. Многие «siglec» экспрессируются на клеточных мембранах иммунных клеток и распознают сиаловую кислоту, так же находящуюся на клеточных мембранах иммунных клеток, и регулируют клеточное взаимодействие или функцию клетки, и предполагается, что они участвуют в иммунном ответе (Nature Reviews Immunology, (2007) 7, pp. 255-266). Однако также существует множество молекул «siglec», физиологические функции которых еще не выяснены. Siglec-15 (иммуноглобулин-подобный лектин 15, связывающий сиаловые кислоты) представляет собой молекулу, которая, как недавно было описано, принадлежит Siglec (Glycobiology, (2007) 17, pp. 838-846) и идентична молекуле, называемой CD33L3 (молекула 3, подобная CD33). Эта молекула эволюционно крайне консервативна от рыб до людей, и было обнаружено, что она сильно экспрессируется в дендритных клетках и/или макрофагах селезенки человека и лимфатических узлах. Далее, как результат теста связывания, в котором используется зонд сиаловой кислоты, было обнаружено, что Siglec-15 человека связывается с Neu5Acα2-6GalNAc и что Siglec-15 мыши помимо Neu5Acα2-6GalNAc связывается с Neu5Acα2-3Galβ1-4Glc (Glycobiology, (2007) 17, pp. 838-846). До настоящего времени физиологическая роль Siglec-15 не была раскрыта, однако сообщалось о том, что экспрессия Siglec-15 повышается при дифференцировке и созревании остеокластов, и дифференцировка остеокластов ингибируется снижением экспрессии Siglec-15 посредством РНК интерференции (WO 2007/093042). Однако действие антитела против Siglec-15 на дифференцировку остеокластов пока не выяснено.
Описание изобретения
Проблемы, решаемые настоящим изобретением
Задачей настоящего изобретения является получение: гена, который специфично экспрессируется при различных формах аномального метаболизма костной ткани, например деструкции кости, которые наблюдаются при остеопорозе, ревматоидном артрите, злокачественных метастазах в костную ткань, или подобном; вещества, которое ингибирует дифференцировку и созревание остеокластов и их активность; нового способа скрининга терапевтического и/или профилактического средства для лечения аномального метаболизма костной ткани; вещества, которое ингибирует дифференцировку и созревание остеокластов и их активность; и терапевтического и/или профилактического средства для лечения аномального метаболизма костной ткани.
Способы решения этих проблем
Авторы настоящего изобретения проводили исследования для объяснения механизма дифференцировки остеокластов, созревания и активации с тем, чтобы найти вещество, обладающее терапевтическим и/или профилактическим действием в отношении аномального метаболизма костной ткани. В результате авторы изобретения обнаружили, что экспрессия гена Siglec-15 повышается при дифференцировке и созревании остеокластов, а также обнаружили, что дифференцировка остеокластов ингибируется под действием антитела, которое специфически связывается с Siglec-15, и, следовательно, было осуществлено настоящее изобретение.
То есть, настоящее изобретение включает следующие изобретения.
(1) Антитело, которое специфически распознает один или несколько полипептидов, содержащих аминокислотную последовательность, описанную в одном из следующих пунктов (a)-(i), и ингибирует образование остеокластов и/или остеокластную резорбцию кости, или функциональный фрагмент антитела:
(a) аминокислотную последовательность, представленную SEQ ID NO:2 в перечне последовательностей;
(b) аминокислотную последовательность, содержащую аминокислотные остатки 21-328 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(c) аминокислотную последовательность, содержащую аминокислотные остатки 1-260 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(d) аминокислотную последовательность, содержащую аминокислотные остатки 21-260 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(e) аминокислотную последовательность, представленную SEQ ID NO:4 в перечне последовательностей;
(f) аминокислотную последовательность, содержащую аминокислотные остатки 21-341 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей;
(g) аминокислотную последовательность, содержащую аминокислотные остатки 1-258 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей;
(h) аминокислотную последовательность, содержащую аминокислотные остатки 21-258 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей; и
(i) аминокислотную последовательность, включающую замены, делеции или добавления одного или нескольких аминокислотных остатков в аминокислотной последовательности, описанной в (a)-(h).
(2) Антитело, которое специфически распознает один или несколько полипептидов, содержащих аминокислотную последовательность, кодируемую нуклеотидной последовательностью, описанной в любом из следующих пунктов (j)-(n), и ингибирует образование остеокластов и/или остеокластную резорбцию кости, или функциональный фрагмент этого антитела:
(j) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:1;
(k) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:3;
(l) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:19;
(m) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:43; и
(n) нуклеотидной последовательностью полинуклеотида, который гибридизируется с полинуклеотидом, содержащим нуклеотидную последовательность, комплементарную нуклеотидной последовательности, описанной в пунктах (j)-(m) в строгих условиях.
(3) Антитело или функциональный фрагмент этого антитела по пункту (1) или (2), которое ингибирует процесс клеточного слияния остеокластов.
(4) Антитело или функциональный фрагмент этого антитела по любому из пунктов (1)-(3), которое ингибирует образование остеокластов, индуцированное под действием TNF-α.
(5) Антитело или функциональный фрагмент этого антитела по любому из пунктов (1)-(4), которое ингибирует образование остеокластов in vitro в концентрации 30 мкг/мл или менее.
(6) Антитело или функциональный фрагмент этого антитела по пункту (5), которое ингибирует образование остеокластов in vitro в концентрации 3 мкг/мл или менее.
(7) Антитело или функциональный фрагмент этого антитела по пункту (6), которое ингибирует образование остеокластов in vitro в концентрации 1 мкг/мл или менее.
(8) Антитело или функциональный фрагмент этого антитела по пункту (7), которое ингибирует образование остеокластов in vitro в концентрации от 63 нг/мл до 1 мкг/мл.
(9) Антитело или функциональный фрагмент этого антитела по любому из пунктов (1)-(4), которое ингибирует остеокластную резорбцию кости.
(10) Антитело или функциональный фрагмент этого антитела по пункту (9), которое ингибирует остеокластную резорбцию кости in vitro в концентрации 3 мкг/мл или менее.
(11) Антитело или функциональный фрагмент этого антитела по пункту (10), которое ингибирует остеокластную резорбцию кости in vitro в концентрации от 0,3 мкг/мл до 3 мкг/мл.
(12) Антитело или функциональный фрагмент этого антитела по любому из пунктов (1)-(11), которое получено способом, включающим следующие стадии 1) и 2):
1) стадию получения антитела, которое специфически распознает один или несколько полипептидов, содержащих аминокислотную последовательность, описанную в любом из следующих пунктов (a)-(i):
(a) аминокислотную последовательность, представленную SEQ ID NO:2 в перечне последовательностей;
(b) аминокислотную последовательность, содержащую аминокислотные остатки 21-328 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(c) аминокислотную последовательность, содержащую аминокислотные остатки 1-260 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(d) аминокислотную последовательность, содержащую аминокислотные остатки 21-260 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(e) аминокислотную последовательность, представленную SEQ ID NO:4 в перечне последовательностей;
(f) аминокислотную последовательность, содержащую аминокислотные остатки 21-341 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей;
(g) аминокислотную последовательность, содержащую аминокислотные остатки 1-258 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей;
(h) аминокислотную последовательность, содержащую аминокислотные остатки 21-258 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей; и
(i) аминокислотную последовательность, включающую замены, делеции или добавления одного или нескольких аминокислотных остатков в аминокислотной последовательности, описанной в пунктах (a)-(h); и
2) стадию скрининга антитела, которое проявляет ингибирующую активность в отношении образования остеокластов и/или ингибирующую активность в отношении резорбции кости.
(13) Антитело или функциональный фрагмент этого антитела по любому из пунктов (1)-(11), которое получено способом, включающим следующие стадии 1) и 2):
1) стадию получения антитела, которое специфически распознает один или несколько полипептидов, содержащих аминокислотную последовательность, кодируемую нуклеотидной последовательностью, описанной в любом из следующих пунктов (j)-(n):
(j) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:1;
(k) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:3;
(l) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:19;
(m) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:43; и
(n) нуклеотидной последовательностью полинуклеотида, который гибридизуется с полинуклеотидом, содержащим нуклеотидную последовательность, комплементарную нуклеотидной последовательности, описанной в пунктах (j)-(m) в строгих условиях; и
2) стадию скрининга антитела, которое проявляет ингибирующую активность в отношении образования остеокластов и/или ингибирующую активность в отношении резорбции кости.
(14) Антитело или функциональный фрагмент этого антитела по любому из пунктов (1)-(13), отличающееся тем, что антитело представляет собой моноклональное антитело.
(15) Антитело или функциональный фрагмент этого антитела по пункту (14), отличающееся тем, что имеет такую же эпитопную специфичность, что и антитело, продуцируемое гибридомой #32A1 (FERM BP-10999).
(16) Антитело или функциональный фрагмент этого антитела по пункту (14), отличающийся тем, что конкурирует с антителом, продуцируемым гибридомой #32A1 (FERM BP-10999).
(17) Антитело или функциональный фрагмент этого антитела по пункту (14), отличающееся тем, что это антитело представляет собой антитело, продуцируемое гибридомой #32A1 (FERM BP-10999).
(18) Антитело или функциональный фрагмент этого антитела по пункту (14), отличающееся наличием той же эпитопной специфичности, что и антитело, продуцируемое гибридомой #41B1 (FERM BP-11000).
(19) Антитело или функциональный фрагмент этого антитела по пункту (14), отличающееся тем, что конкурирует с антителом, продуцируемым гибридомой #41B1 (FERM BP-11000).
(20) Антитело или функциональный фрагмент этого антитела по пункту (14), отличающееся тем, что это антитело представляет собой антитело, продуцируемое гибридомой #41B1 (FERM BP-11000).
(21) Антитело или функциональный фрагмент этого антитела по любому из пунктов (1)-(20), отличающееся тем, что это антитело представляет собой химерное антитело.
(22) Антитело или функциональный фрагмент этого антитела по любому из пунктов (1)-(21), отличающееся тем, что это антитело является гуманизированным.
(23) Антитело или функциональный фрагмент этого антитела по любому из пунктов (1)-(16), (18) и (19), отличающееся тем, что это антитело представляет собой антитело человека.
(24) Антитело или функциональный фрагмент этого антитела по любому из пунктов (1)-(23), отличающееся тем, что это антитело представляет собой антитело IgG.
(25) Функциональный фрагмент антитела по любому из пунктов (1)-(24), который выбран из группы, состоящей из Fab, F(ab')2, Fab' и Fv.
(26) Антитело по любому из пунктов (1)-(16), (18) и (19), отличающееся тем, что представляет собой scFv.
(27) Фармацевтическая композиция, отличающаяся тем, что содержит по меньшей мере одно из антител или функциональных фрагментов антител по пунктам (1)-(26).
(28) Фармацевтическая композиция по пункту (27), отличающаяся тем, что представляет собой терапевтическое и/или профилактическое средство для лечения аномального метаболизма костной ткани.
(29) Фармацевтическая композиция для лечения и/или профилактики аномального метаболизма костной ткани, отличающаяся тем, что содержит по меньшей мере одно из антител или функциональных фрагментов антител по пунктам (1)-(26) и по меньшей мере один компонент, выбранный из группы, состоящей из бисфосфонатов, активного витамина D3, кальцитонина и его производных, гормональных препаратов, таких как эстрадиол, SERM (селективных модуляторов рецепторов эстрогена), иприфлавона, витамина K2 (менатетренона), препаратов кальция, препаратов PTH (паратиреоидного гормона), нестероидных противовоспалительных средств, препаратов растворимого TNF рецептора, анти-TNF-α антител или функциональных фрагментов антител, анти-PTHrP (белок, родственный паратиреоидному гормону) антител или функциональных фрагментов этих антител, антагонистов рецептора IL-1, антител против рецептора IL-6 или функциональных фрагментов этих антител, анти-RANKL антител или функциональных фрагментов таких антител и OCIF (фактора, ингибирующего остеокластогенез).
(30) Фармацевтическая композиция по пунктам (28) или (29), где аномальный метаболизм костной ткани выбран из группы, состоящей из остеопороза, деструкции кости, сопровождающей ревматоидный артрит, злокачественной гиперкальциемии, деструкции кости, сопровождающий множественную миелому или злокачественное метастазирование в костную ткань, гигантоклеточной остеобластомы, потери зубов вследствие периодонтита, остеолизиса вокруг протезированного сустава, деструкции кости при хроническом остеомиелите, деформирующей остеодистрофии, почечной остеодистрофии и несовершенного остеогенеза.
(31) Фармацевтическая композиция по пункту (30), отличающаяся тем, что аномальным метаболизмом костной ткани является остеопороз, деструкция кости, сопровождающая ревматоидный артрит или деструкция кости, сопровождающая злокачественное метастазирование в костную ткань.
(32) Фармацевтическая композиция по пункту (31), отличающаяся тем, что остеопороз представляет собой постменопаузный остеопороз, сенильный остеопороз, вторичный остеопороз, вследствие применения таких терапевтических средств, как стероиды или иммунодепрессанты, или остеопороз, сопровождающий ревматоидный артрит.
(33) Способ лечения и/или профилактики аномального метаболизма костной ткани, характеризующийся введением по меньшей мере одного из антител или функциональных фрагментов антител по пунктам (1)-(26).
(34) Способ лечения и/или профилактики аномального метаболизма костной ткани, отличающийся одновременным или последовательным введением по меньшей мере одного из антител или функциональных фрагментов антител по пунктам (1)-(26) и по меньшей мере одного компонента, выбранного из группы, состоящей из бисфосфонатов, активного витамина D3, кальцитонина и его производных, гормональных препаратов, таких как эстрадиол, SERM (селективных модуляторов рецепторов эстрогена), иприфлавона, витамина K2 (менатетренона), препаратов кальция, препаратов PTH (паратиреоидного гормона), нестероидных противовоспалительных средств, препаратов растворимого рецептора TNF, антител против TNF-α или функциональных фрагментов этих антител, анти-PTHrP (белка, родственного паратиреоидному гормону) антител или функциональных фрагментов этих антител, антагонистов рецептора IL-1, антител против рецептора IL-6 или функциональных фрагментов таких антител, антител против RANKL или функциональных фрагментов таких антител и OCIF (фактора, ингибирующего остеокластогенез).
(35) Способ лечения и/или профилактики по пункту (33) или (34), отличающийся тем, что аномальный метаболизм костной ткани представляет собой остеопороз, деструкцию кости, сопровождающую ревматоидный артрит или деструкцию кости, сопровождающую злокачественное метастазирование в костную ткань.
(36) Способ лечения и/или профилактики по пункту (35), отличающийся тем, что остеопороз представляет собой постменопаузный остеопороз, сенильный остеопороз, вторичный остеопороз вследствие применения таких терапевтических средств, как стероиды или иммунодепрессанты, или остеопороз, сопровождающий ревматоидный артрит.
(37) Гибридома #32A1 (FERM BP-10999).
(38) Гибридома #41B1 (FERM BP-11000).
Преимущество изобретения
В соответствии с настоящим изобретением может быть получено терапевтическое и/или профилактическое средство для лечения аномального метаболизма костной ткани, механизмом действия которого является ингибирование дифференцировки и созревания остеокластов и их активности.
Краткое описание чертежей
На фиг. 1 показаны графики, демонстрирующие анализ профиля экспрессии для генов RANK, RANKL, катепсина K и TRAP в тканях гигантоклеточной остеобластомы у людей.
На фиг. 2 показан график, демонстрирующий анализ профиля экспрессии для гена Siglec-15 в тканях гигантоклеточной остеобластомы у людей.
На фиг. 3 показан график, демонстрирующий изменение уровня экспрессии гена Siglec-15 в тех случаях, когда дифференцировка остеокластов была индуцирована из клеток RAW 264.7 или клеток костного мозга мышей.
На фиг. 4 показаны графики, демонстрирующие экспрессию генов катепсина K и TRAP, сопровождающую остеокластную дифференцировку клеток RAW 264.7.
На фиг. 5 показан график, демонстрирующий экспрессию гена Siglec-15, сопровождающую остеокластную дифференцировку клеток RAW 264.7.
На фиг. 6 показаны результаты определения изменения экспрессии мышиного Siglec-15-His в клетках 293F в зависимости от времени культивирования с помощью электрофореза в SDS-полиакриламидном геле и Вестерн-блоттинга, с использованием антитела против 6-His-HRP.
На фиг. 7 показаны результаты определения изменения экспрессии мышиного Siglec-15-Fc в клетках 293F в зависимости от времени культивирования с помощью электрофореза в SDS-полиакриламидном геле и Вестерн-блоттинга, с использованием антитела против IgG-Fc-HRP человека.
На фиг. 8 показаны результаты оценки чистоты мышиного Siglec-15-His, очищенного с помощью колоночной хроматографии HisTrap HP и хроматографии на колонке Resource Q, посредством электрофореза в SDS-полиакриламидном геле и окрашивания серебром.
На фиг. 9 показаны результаты определения поведения мышиного Siglec-15-His, очищенного с помощью колоночной хроматографии HisTrap HP и колоночной хроматографии Resource Q, посредством электрофореза в SDS-полиакриламидном геле и Вестерн-блоттинга, с использованием антитела против V5-HRP.
На фиг. 10 показаны результаты оценки чистоты мышиного Siglec-15-Fc, очищенного с помощью хроматографии на колонке HiTrap с Белком A, посредством электрофореза в SDS-полиакриламидном геле и окрашивания серебром.
На фиг. 11 показаны результаты подтверждения того, что очищенное поликлональное антитело против мышиного Siglec-15 связывается не только с Siglec-15-Fc, а также с Siglec-15-His с помощью электрофореза в SDS-полиакриламидном геле и Вестерн блоттинга, с использованием поликлонального антитела против мышиного Siglec-15 и антитела против кроличьего IgG-HRP.
На фиг. 12 показана хроматограмма поликлонального антитела против Siglec-15 мыши, очищенного с помощью аффинной колонки, с иммобилизованным на ней мышиным Siglec-15-Fc.
На фиг. 13 показаны результаты оценки чистоты мышиного Siglec-15-Fc, очищенного с помощью хроматографии, с использованием аффинной колонки с иммобилизованным на ней Siglec-15-Fc мыши.
На фиг. 14 показаны хроматограммы поликлональных антител против Siglec-15 мыши, очищенных с помощью гель-фильтрационной колонки на Сефарозе 6.
На фиг. 15 показаны результаты тестирования действия аффинно-очищенного поликлонального антитела против Siglec-15 мыши на остеокластную дифференцировку (стимуляция действием RANKL) мышиных неадгезивных клеток костного мозга (N=3).
На фиг. 16 показаны результаты тестирования действия добавления очищенного гель-фильтрацией поликлонального антитела против мышиного Siglec-15 на остеокластную дифференцировку (стимуляция действием RANKL) неадгезивных клеток костного мозга мыши, исходя из ферментативной активности TRAP (N=3).
На фиг. 17 показаны результаты тестирования нейтрализации антигеном ингибирования остеокластной дифференцировки (стимуляция действием RANKL) неадгезивных клеток костного мозга мыши путем добавления поликлонального антитела против Siglec-15 мыши, исходя из ферментативной активности TRAP (N=3).
На фиг. 18 показаны результаты тестирования действия добавления поликлонального антитела против Siglec-15 мыши на остеокластную дифференцировку (стимуляция действием TNF-α) неадгезивных клеток костного мозга мыши, исходя из ферментативной активности TRAP (N=3).
На фиг. 19 показаны микрофотографии, используемые для оценки действия добавления поликлонального антитела против Siglec-15 мыши на остеокластную дифференцировку (стимуляция действием TNF-α) мышиных неадгезивных клеток костного мозга путем окрашивания TRAP.
На фиг. 20 показаны графики, демонстрирующие, исходя из ферментативной активности TRAP, ингибирование остеокластной дифференцировки (стимуляция активным витамином D3) из мышиных клеток костного мозга путем добавления поликлонального антитела против Siglec-15 мыши (N=6).
На фиг. 21 показаны микрофотографии, демонстрирующие посредством окрашивания TRAP, ингибирование образования гигантских остеокластов (стимуляция активным витамином D3) из мышиных клеток костного мозга путем добавления поликлонального антитела против Siglec-15 мыши.
На фиг. 22 показаны микрофотографии, демонстрирующие посредством окрашивания TRAP ингибирование образования гигантских остеокластов (стимуляция под действием RANKL человека) из мышиных клеток костного мозга путем добавления поликлонального антитела против Siglec-15 мыши.
На фиг. 23 показаны микрофотографии, демонстрирующие посредством окрашивания TRAP ингибирование образования гигантских остеокластов (стимуляция под действием RANKL человека) из клеток RAW 264.7, путем добавления поликлонального антитела против Siglec-15 мыши и отмену ингибирующего действия под действием растворимого Siglec-15.
На фиг. 24 показаны результаты тестирования связывания крысиного моноклонального антитела против Siglec-15 мыши на планшете с иммобилизованным на нем мышиным Siglec-15-Fc с помощью ELISA. Знак (◆) означает антитело #1A1, знак (■) означает антитело #3A1, знак (▲) означает антитело #8A1, знак (×) означает антитело #24A1, знак (●) означает антитело #32A1, знак (○) означает антитело #34A1, знак (+) означает антитело #39A1, знак (-) означает антитело #40A1, знак (―) означает антитело #41B1, знак (◇) означает антитело #61A1 и знак (□) означает контрольный IgG.
На фиг. 25 показаны результаты тестирования эффекта добавления моноклонального антитела (#3A1, #8A1 или #32A1) против Siglec-15 мыши на остеокластную дифференцировку (стимуляцию под действием RANKL) мышиных неадгезивных клеток костного мозга. Контрольный IgG крысы на этой фигуре является отрицательным контролем, общим для фиг.25 и 26.
На фиг. 26 показаны результаты тестирования эффекта добавления моноклонального антитела (#34A1, #39A1 или #40A1) против Siglec-15 мыши на остеокластную дифференцировку (стимуляция под действием RANKL) мышиных неадгезивных клеток костного мозга. Кроличье поликлональное антитело № 3 против Siglec-15 мыши на этой фигуре является положительным контролем, общим для фиг.25 и 26.
На фиг. 27 показаны графики, демонстрирующие изменение в экспрессии генов катепсина К, TRAP или Siglec-15 в тех случаях, когда остеокластная дифференцировка была индуцирована из нормальных клеток-предшественников остеокластов человека.
На фиг. 28 показаны результаты проверки чистоты белка человека Siglec-15-His, очищенного колоночной хроматографией HisTrap HP и хроматографией на колонке Resource Q, посредством электрофореза в SDS-полиакриламидном геле.
На фиг. 29 показаны результаты проверки чистоты белка человека Siglec-15-Fc, очищенного с помощью колоночной хроматографии на Белке А, посредством электрофореза в SDS-полиакриламидном геле.
На фиг. 30 показаны хроматограммы поликлональных антител против Siglec-15 человека, очищенных с помощью аффинной колонки с иммобилизованным на ней белком человека Siglec-15-Fc.
На фиг. 31 показаны микрофотографии, демонстрирующие посредством окрашивания TRAP ингибирование образования гигантских остеокластов из нормальных клеток-предшественников остеокластов человека путем добавления поликлонального антитела против Siglec-15 человека.
На фиг. 32 показаны результаты оценки действия добавления поликлонального антитела против Siglec-15 человека на формирование многоядерных остеокластов из нормальных клеток-предшественников остеокластов человека путем подсчитывания числа TRAP-позитивных клеток, имеющих 5 или более ядер, с помощью инвертированного микроскопа.
На фиг. 33 показаны результаты тестирования связывания крысиного моноклонального антитела против Siglec-15 мыши на планшете с иммобилизованными на нем белком человека Siglec-15-Fc с помощью ELISA. Знак (◆) означает антитело #1A1, знак (■) означает антитело #3A1, знак (▲) означает антитело #8A1, знак (×) означает антитело #24A1, знак (●) означает антитело #32A1, знак (○) означает антитело #34A1, знак (+) означает антитело #39A1, знак (-) означает антитело #40A1, знак (―) означает антитело #41B1, знак (◇) означает антитело #61A1 и знак (□) означает контрольный IgG.
На фиг. 34 показаны микрофотографии, демонстрирующие путем окрашивания TRAP ингибирование образования гигантских остеокластов из нормальных клеток-предшественников остеокластов человека путем добавления крысиного моноклонального антитела против Siglec-15 мыши.
На фиг. 35 показаны микрофотографии, демонстрирующие посредством окрашивания TRAP ингибирование образования гигантских остеокластов из нормальных клеток-предшественников остеокластов человека путем добавления крысиного моноклонального антитела против Siglec-15 мыши (антитело #32A1).
На фиг. 36 показан график, демонстрирующий ингибирование активности нормальных остеокластов человека в отношении резорбции костной ткани путем добавления крысиного моноклонального антитела против Siglec-15 мыши (антитело #32A1) (N=6).
Наилучший способ осуществления настоящего изобретения
Термин «ген», используемый в контексте настоящего изобретения, включает не только ДНК, а также мРНК, кДНК и кРНК.
Термин «полинуклеотид», используемый в контексте настоящего изобретения, используется в том же значении, что и нуклеиновая кислота, а также включает ДНК, РНК, зонды, олигонуклеотиды и праймеры.
Термины «полипептид» и «белок», используемые в контексте настоящего изобретения, используются без различия.
Термин «фракция РНК», используемый в контексте настоящего изобретения, относится к фракции, содержащей РНК.
Термин «клетка», используемый в контексте настоящего изобретения, также включает клетки отдельных животных и культивированные клетки.
Термин «Siglec-15», используемый в контексте настоящего изобретения, используется в том же значении, что и белок Siglec-15.
Термин «образование остеокластов», используемый в контексте настоящего изобретения, используется в том же значении, что и «остеокластная дифференцировка» или «созревание остеокластов».
Термин «функциональный фрагмент антитела», используемый в контексте настоящего изобретения, относится к неполному фрагменту антитела, обладающему антиген-связывающей активностью, и включает Fab, F(ab')2, scFv и подобные. Этот термин также охватывает Fab', который является одновалентным фрагментом в вариабельной области антитела, полученного путем обработки F(ab')2 в редуцирующих условиях. Однако этот термин не ограничивается этими молекулами, при условии, что этот фрагмент обладает аффинностью связывания в отношении антигена. Далее, эти функциональные фрагменты включают не только фрагмент, полученный обработкой полноразмерной молекулы белка антитела соответствующим ферментом, а также белок, полученный в соответствующей клетке-хозяине, с использованием генетически модифицированного гена антитела.
Термин «эпитоп», используемый в контексте настоящего изобретения, относится к части пептида Siglec-15, с которым связывается специфичное антитело против Siglec-15. Упомянутый выше эпитоп, который представляет собой часть пептида Siglec-15, может быть определен способами, хорошо известными специалистам в данной области, например иммуноанализом. Альтернативно, например, может быть использован следующий способ. Получают различные отдельные структуры Siglec-15. В получении отдельных структур может быть использована известная методика олигопептидного синтеза. Например, серии полипептидов, имеющих соответствующим образом уменьшенную длину, полученных последовательным укорочением Siglec-15 с С-конца или N-конца, получают, используя метод генной рекомбинации, известный специалистам в данной области. После этого проверяют реакционную способность антитела в отношении этих полипептидов и приблизительно определяют сайт узнавания. Затем синтезируют полипептиды более короткой длины и проверяют реакционную способность этих пептидов, тем самым можно выявить эпитоп. В случаях, если второе антитело против Siglec-15 связывается с частью пептида, с которой связывается первое антитело против Siglec-15, можно определить, что первое антитело и второе антитело делят между собой один и тот же эпитоп. Далее, путем подтверждения того, что второе антитело против Siglec-15 конкурирует с первым антителом против Siglec-15 за связывание с Siglec-15 (то есть второе антитело ингибирует связывание между Siglec-15 и первым антителом), можно определить, что первое антитело и второе антитело делят один и тот же эпитоп, даже если специфическая последовательность эпитопа не была определена. Далее, в тех случаях, когда первое антитело и второе антитело связываются с одним и тем же эпитопом, а также первое антитело обладает специфическим действием, например антиген-нейтрализующей активностью, можно предположить, что второе антитело обладает такой же активностью.
Выражение «гибридизацию проводят в строгих условиях», используемое в контексте настоящего изобретения, относится к гибридизации, проводимой в условиях, при которых идентификация может быть выполнена путем проведения гибридизации при 68°C в коммерчески доступном гибридизационном растворе, гибридизационном растворе ExpressHyb Hybridization (производство Clontech, Inc.) или проведения гибридизации при 68°C в присутствии 0,7-1,0 M NaCl с использованием фильтра с иммобилизованной на нем ДНК, с последующим промыванием при 68°C, используя 0,1-2×SSC раствор (1×SSC раствор состоит из 150 мМ NaCl и 15 мМ цитрата натрия), или в условиях, эквивалентных этим.
1. Siglec-15
Авторы настоящего изобретения обнаружили, что ген Siglec-15 специфично экспрессируется в гигантоклеточных остеобластомах, а также обнаружили, что уровень экспрессии гена Siglec-15 повышается в тех случаях, когда моноцитарная клеточная линия дифференцируется в остеокласты.
Что касается Siglec-15, используемого в настоящем изобретении, Siglec-15 непосредственно очищают из моноцитов или клеток костного мозга человека, животных, не являющихся человеком (таких как морская свинка, крыса, мышь, кролик, свиньи, овцы, крупный рогатый скот или обезьяны), или цыпленка и используют или получают и могут использовать фракцию клеточной мембраны упомянутых выше клеток. Далее, Siglec-15 может быть получен путем синтеза in vitro или продукцией в клетке-хозяине методами генной инженерии. В методах генной инженерии, в частности, кДНК Siglec-15, интегрируют в вектор, способный экспрессировать кДНК Siglec-15, и Siglec-15 синтезируют в растворе, содержащем ферменты, субстраты и энергетические вещества, необходимые для транскрипции и трансляции, или другую прокариотическую или эукариотическую клетку-хозяина трасформируют для экспрессии Siglec-15, таким образом может быть получен белок.
Нуклеотидная последовательность кДНК Siglec-15 человека была зарегистрирована в GenBank под регистрационным номером NM_213602 и представлена последовательностью SEQ ID NO:1 в перечне последовательностей, и ее аминокислотная последовательность представлена последовательностью SEQ ID NO:2 в перечне последовательностей. Нуклеотидная последовательность кДНК Siglec-15 мыши была зарегистрирована в GenBank под регистрационным номером XM_884636 и представлена последовательностью SEQ ID NO:3 в перечне последовательностей, и ее аминокислотная последовательность представлена последовательностью SEQ ID NO:4 в перечне последовательностей. Зрелый Siglec-15 человека, у которого была удалена сигнальная последовательность, соответствует аминокислотной последовательности, состоящей из аминокислотных остатков 21-328 аминокислотной последовательности, представленной SEQ ID NO:2. Далее, Siglec-15 мыши, у которого была удалена сигнальная последовательность, соответствует аминокислотной последовательности, состоящей из аминокислотных остатков 21-341 аминокислотной последовательности, представленной SEQ ID NO:4. В этой связи, Siglec-15 иногда называют CD33 антиген-подобной 3, молекулой 3, подобной CD33, CD33-подобной 3 или CD33L3, и все они представляют одну и ту же молекулу.
кДНК Siglec-15 может быть получена, например, с помощью так называемого ПЦР способа, в котором полимеразно-цепную реакцию (здесь и далее называемую «ПЦР») проводят, используя библиотеку кДНК, экспрессирующую кДНК Siglec-15 в качестве матрицы, и праймеры, которые специфически амплифицируют кДНК Siglec-15 (Saiki, R. K., et al., Science, (1988) 239, 487-49).
Далее, полинуклеотид, который гибридизируется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной по меньшей мере одной нуклеотидной последовательности, выбранной из нуклеотидных последовательностей, представленных SEQ ID NO:1 и 3 в перечне последовательностей, в строгих условиях, и которая кодирует белок, обладающий биологической активностью, сравнимой с активностью Siglec-15, также включен в кДНК Siglec-15. Далее, полинуклеотид, который представляет собой вариант сплайсинга, транскрибируемый с локуса Siglec-15 человека или мыши, или полинуклеотид, который гибридизируется с ним в условиях строгости и кодирует белок, обладающий биологической активностью, сравнимой с активностью Siglec-15, также включен в кДНК Siglec-15.
Далее, белок, который состоит из аминокислотной последовательности, включающей замены, делеции или добавления одной или нескольких аминокислот по меньшей мере в одной аминокислотной последовательности, выбранной из аминокислотных последовательностей, представленных последовательностями SEQ ID NO:2 и 4 в перечне последовательностей или аминокислотной последовательности, из которой была удалена сигнальная последовательность, и имеет биологическую активность, сравнимую с активностью Siglec-15, также включен в Siglec-15. Далее, белок, который состоит из аминокислотной последовательности, кодируемой вариантом сплайсинга, транскрибируемым с локуса Siglec-15 человека или мыши или аминокислотной последовательности, включающей замены, делеции или добавления одной или нескольких аминокислот, и обладает биологической активностью, сравнимой с активностью Siglec-15, также включен в Siglec-15.
2. Выявление аномального метаболизма костной ткани
Анализ уровня экспрессии гена Siglec-15 в группе тестируемых образцов из различных костных тканей человека показал, что уровень экспрессии гена значительно повышается в гигантоклеточной остеобластоме (GCT), которая представляет собой опухоль костной ткани с возрастающим огромным числом многоядерных гигантских клеток, подобных остеокластам, и характеризуется клинической картиной остеолитической деструкции кости (Bullough et al., Atlas of Orthopedic Pathology 2nd edition, pp. 17.6-17.8, Lippincott Williams & Wilkins Publishers (1992)).
Также было обнаружено, что уровень экспрессии гена Siglec-15 повышается в тех случаях, когда моноцитарная клеточная линия дифференцируется в остеокласты.
В соответствии с этим предполагается, что Siglec-15 связан с патологическим состоянием человека, таким как GCT, при котором повышается резорбция кости. Другими словами, измерение уровня экспрессии гена Siglec-15 и/или Siglec-15 в каждой клетке и/или каждой ткани дает возможность определить состояние аномального метаболизма костной ткани, сопровождающегося гиперэкспрессией Siglec-15. Примеры аномального метаболизма костной ткани в контексте настоящего изобретения включают, но не только, остеопороз (постменопаузный остеопороз, сенильный остеопороз, вторичный остеопороз вследствие применения терапевтического средства, такого как стероиды или иммунодепрессанты, или остеопороз, сопровождающий ревматоидный артрит), деструкцию костей, сопровождающую ревматоидный артрит, злокачественную гиперкальциемию, деструкцию костей, сопровождающую множественную миелому или злокачественное метастазирование в костную ткань, гигантоклеточную остеобластому, потерю зубов вследствие периодонтита, остеолиз вокруг протезированного сустава, деструкцию костей при хроническом остеомиелите, деформирующую остеодистрофию (болезнь Педжета), почечную остеодистрофию и несовершенный остеогенез.
В настоящем изобретении «тестируемый образец», используемый для исследования уровня экспрессии гена Siglec-15 и/или Siglec-15, относится к образцу из ткани костного мозга, кости, предстательной железы, яичек, пениса, мочевого пузыря, почек, ротовой полости, глотки, губ, языка, десен, носоглотки, пищевода, желудка, тонкой кишки, толстой кишки, ободочной кишки, печени, желчного пузыря, поджелудочной железы, носа, легких, мягких тканей, кожи, груди, матки, яичников, головного мозга, щитовидной железы, лимфатического узла, мышечной, жировой ткани или подобных, или крови, жидкости организма или выделения или подобного, однако в настоящем изобретении более предпочтительными являются кровь или костный мозг.
3. Способ скрининга вещества, которое ингибирует дифференцировку в остеокласты
В качестве одного варианта осуществления изобретения можно привести в пример способ скрининга вещества, которое ингибирует дифференцировку в остеокласты, путем измерения уровня экспрессии гена Siglec-15 и/или Siglec-15.
В качестве другого варианта осуществления настоящего изобретения может служить примером способ скрининга вещества, которое оказывает терапевтическое действие и/или профилактическое действие на аномальный метаболизм костной ткани, путем идентификации вещества, которое ингибирует активность Siglec-15 в отношении индукции дифференцировки в зрелые остеокласты.
«Тестируемое вещество» относится к веществу, используемому для исследования активности ингибирования дифференцировки в остеокласты способом скрининга по настоящему изобретению. Примеры тестируемого вещества включают соединение, метаболит микроорганизма, экстракт из ткани растения или животного, их производные и их смеси. Далее, нуклеиновая кислота, предназначенная для снижения уровня экспрессии Siglec-15, или ее производные (например, антисмысловой олигонуклеотид, рибозим, dsРНК или siРНК) также могут быть использованы в качестве тестируемого вещества. Доза или концентрация тестируемого вещества может быть установлена соответствующим образом, или множество доз может быть установлено, например, путем получения их серийных разведений. Тестируемое вещество можно вводить в соответствующем виде, например твердом или жидком, и оно может быть растворено в соответствующем буфере, или к нему можно добавить стабилизатор или подобное. В случае способа скрининга с использованием культивированных клеток, тестируемое вещество добавляют к среде, и клетки могут быть культивированы в этой среде. В случае, когда тестируемое вещество добавляют в среду, тестируемое вещество может быть добавлено в начале культивирования или в середине культивирования, и количество операций добавления не ограничено одной. Период культивирования в присутствии тестируемого вещества может быть установлено соответствующим образом, однако предпочтительно составляет от 30 минут до 2 недель, более предпочтительно от 30 минут до 72 часов. В том случае, когда тестируемое вещество вводят млекопитающему индивидууму, путь введения, в том числе пероральное введение, внутривенную инъекцию, внутрибрюшинную инъекцию, чрескожное введение, подкожную инъекцию и подобное, соответственно определяют в зависимости от физических свойств тестируемого вещества и подобного. К тому же, может быть выбрано подходящее время до получения тестируемого образца после введения тестируемого вещества.
Культивируемыми клетками, используемыми в способе скрининга по настоящему изобретению, могут быть нормальные клетки, устойчивая клеточная линия или клетки, демонстрирующие аномальный рост, такие как злокачественные клетки, при условии, что они представляют собой клетки млекопитающих, экспрессирующих Siglec-15. Их примеры включают, но не только, нормальные клетки-предшественники остеокластов человека (Нормальные природные клетки-предшественники остеокластов человека, доступные от Sanko Junyaku Co., Ltd., Cat. No. 2T-110), мышиные моноцитарные клетки RAW 264.7 (ATCC Cat. No. TIB-71), клетки RAW 264 (ECACC Cat. No. 85062803) и мышиные первичные культивированные клетки костномозгового происхождения. В качестве видов животных для получения культивированных клеток предпочтительными являются человек, мышь или другие животные (морские свинки, крысы, кролики, свиньи, овцы, крупный рогатый скот, обезьяны и т.д.), цыплята и подобные, но эти виды не ограничены ими. К тому же, в качестве культивированных клеток более предпочтительно использовать клетки млекопитающих, гиперэкспрессирующих Siglec-15, и, например, могут быть проиллюстрированы клетки RAW 264.7, клетки RAW 264, клетки 293, клетки CHO и клетки COS7, модифицированные введением гена Siglec-15 вместе с его промоторной областью для гиперэкспрессии Siglec-15, и подобные.
Способ скрининга по настоящему изобретению также включает способ выявления экспрессии гена Siglec-15 в клетках органа или ткани, извлеченных из млекопитающего индивидуума после введения тестируемого вещества этому млекопитающему индивидууму без использования культивированных клеток. Орган или ткань, используемые для выявления экспрессии гена, могут быть любыми, при условии, что они экспрессируют Siglec-15, однако предпочтительной является ткань, связанная с аномальным метаболизмом костной ткани, и более предпочтительными являются костная ткань и костный мозг. В качестве видов млекопитающих могут быть использованы млекопитающие, не являющиеся человеком, и предпочтительными являются мыши, крысы или хомяки, и более предпочтительными являются мыши или крысы. В качестве животной модели с аномальным метаболизмом костной ткани можно использовать животное с удаленным яичником, животное с удаленными тестикулами, животное со злокачественной опухолью, имеющее опухолевые клетки, имплантированные под кожу, в кожу, левый желудочек, костный мозг, вену, брюшную полость и подобное, и животное, с удаленным седалищным нервом, животную модель адъювантного артрита, животную модель коллаген-индуцированного артрита, и животную модель индуцированного глюкокортикоидами остеопороза, мышей с ускоренным старением (мыши SAM P6, Matsushita et al., Am. J. Pathol. 125, 276-283 (1986)), животное с удаленной щитовидной/паращитовидной железой, животное, получающее непрерывную инфузию пептида, родственного паратиреоидному гормону (PTHrP), или «нокаут» мышей по фактору, ингибирующему остеокластогенез (OCIF) (Mizuno et al., Biochem. Biophys. Res. Commun., (1998) 247, 610-615), животное с введением растворимого RANKL или подобного. Далее, также может быть использована животная модель с потерей зубов вследствие пародонтита или животное, модифицированное для гиперэкспрессии Siglec-15. Далее, тестируемое вещество, выбранное путем скрининга, вводят любому из упомянутых выше животных-моделей и измеряют каждый параметр, который может быть получен путем измерения числа зрелых остеокластов в костной ткани, плотности кости, прочности кости или морфологии кости, параметров метаболизма костной ткани (CTx, NTx и т.д.) в крови и моче, или параметры, которые изменяются вследствие аномального метаболизма костной ткани, такие как уровни кальция в крови, в соответствии с которыми можно оценить терапевтическое действие и/или профилактическое действие тестируемого соединения на аномальный метаболизм костной ткани.
Культивированные клетки, используемые в способе по настоящему изобретению, могут быть культивированы в любых условиях, пока эти условия дают возможность культивированным клеткам экспрессировать Siglec-15 без добавления тестируемого вещества. Например, существуют известные условия культивирования для культивируемых клеток, и в тех случаях, когда эти клетки экспрессируют в этих условиях Siglec-15, клетки могут быть культивированы в этих условиях. Далее, в том случае, когда выявляется экспрессия Siglec-15 в органе или ткани, удаляемой у млекопитающего индивидуума, условия разведения для животного могут быть любыми, при условии, что эти условия дают возможность животному экспрессировать Siglec-15 без добавления тестируемого вещества.
Для изучения действия тестируемого вещества на экспрессию Siglec-15 существует способ измерения уровня экспрессии гена Siglec-15 и способ измерения уровня экспрессии Siglec-15, который является продуктом трансляции гена Siglec-15. Предполагается, что тестируемое вещество, которое ингибирует экспрессию гена Siglec-15 и/или Siglec-15, представляет собой вещество, оказывающее терапевтическое действие и/или профилактическое действие на аномальный метаболизм костной ткани, предпочтительно остеопороз, или деструкцию кости, сопровождающую ревматоидный артрит и/или злокачественное метастазирование в костную ткань.
Измерение уровня экспрессии гена Siglec-15 или Siglec-15 в культивированных клетках можно осуществить с помощью Нозерн анализа, количественного метода ПЦР, метода ELISA или подобных. В том случае, когда используют культивируемые клетки млекопитающих, наряду с тестируемым веществом, при необходимости, к среде добавляют соответствующее количество RANKL, TNF-α, M-CSF, активного витамина D3, или подобного, а также в контроль без добавления тестируемого вещества добавляют соответствующее количество RANKL, TNF-α, M-CSF, активного витамина D3 или подобного.
Далее, конструируют экспериментальную систему для измерения связывающего количества эндогенного лиганда с Siglec-15 и определяют, ингибируется или нет связывание эндогенного лиганда с Siglec-15 путем добавления тестируемого соединения, тем самым можно осуществить скрининг вещества, которое ингибирует дифференцировку в остеокласты.
Соответствующие способы скрининга будут описаны следующими пунктами (1)-(3).
(1) Способ с использованием гена Siglec-15
В качестве способа скрининга по настоящему изобретению, например, существует способ с использованием культивируемых клеток млекопитающего и способ с использованием отдельного млекопитающего, которые будут описаны ниже, соответственно.
(a) Способ с использованием культивируемых клеток млекопитающих
(i) Способ, включающий следующие стадии a)-c):
a) стадию экстракции суммарной РНК из культивируемых клеток млекопитающих, культивируемых в среде с добавлением тестируемого вещества;
b) стадию выявления различия в уровне экспрессии гена Siglec-15 между суммарной РНК, полученной на стадии a), и суммарной РНК, полученной из культивируемых клеток млекопитающих, культивированных без добавления тестируемого вещества; и
c) стадию определения терапевтического и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия в уровне экспрессии гена, описанного на стадии b).
(ii) Способ, включающий следующие стадии a)-d):
a) стадию экстракции суммарной РНК из культивируемых клеток млекопитающих в среде с добавлением тестируемого вещества;
b) стадию экстракции суммарной РНК из культивируемых клеток млекопитающих, культивированных в среде без добавления тестируемого вещества;
c) стадию измерения уровня экспрессии гена Siglec-15 в суммарной РНК, полученной на стадии a) и в суммарной РНК, полученной на стадии b), соответственно; и
d) стадию определения терапевтического и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия в уровне экспрессии гена, определенной на стадии c) между суммарной РНК, полученной на стадии a) и суммарной РНК, полученной на стадии b).
(b) Способ, в котором используют отдельных млекопитающих
(i) Способ, включающий следующие стадии (a)-(c):
a) стадию экстракции суммарной РНК из тестируемого образца, полученного от млекопитающего индивидуума, с введением тестируемого вещества;
b) стадию выявления различия в уровне экспрессии гена Siglec-15 между суммарной РНК, полученной на стадии a), и суммарной РНК, полученной из тестируемого образца, полученного от млекопитающего индивидуума без введения тестируемого вещества; и
c) стадию определения терапевтического и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия в уровне экспрессии гена, описанного на стадии b).
(ii) Способ, включающий следующие стадии a)-d):
a) стадию экстракции суммарной РНК из тестируемого образца, полученного от млекопитающего индивидуума с введением тестируемого вещества;
b) стадию экстракции суммарной РНК из тестируемого образца, полученного от млекопитающего индивидуума без введения тестируемого вещества;
c) стадию измерения уровня экспрессии гена Siglec-15 в суммарной РНК, полученной на стадии a), и в суммарной РНК, полученной на стадии b), соответственно; и
d) стадию определения терапевтического и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия в уровне экспрессии гена, описанного на стадии c).
(2) Способ с использованием Siglec-15
В качестве способа скрининга с использованием измерения уровня экспрессии Siglec-15, существует способ, в котором используются культивированные клетки млекопитающих, и способ, в котором используют индивидуальное животное, каждый из которых включает следующие стадии.
(a) Способ, в котором используются культивированные клетки млекопитающих
(i) Способ, включающий следующие стадии a) и b):
a) стадию измерения уровня экспрессии Siglec-15 в культивированных клетках млекопитающих, культивированных в среде с добавлением тестируемого вещества; и
b) стадию определения терапевтического действия и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия между уровнем экспрессии белка, измеренным на стадии a), и уровнем экспрессии белка в культивированных клетках млекопитающих, культивированных в среде без добавления тестируемого вещества.
(ii) Способ, включающий следующие стадии a)-c):
a) стадию измерения уровня экспрессии Siglec-15 в культивированных клетках млекопитающих в среде с добавлением тестируемого вещества;
b) стадию измерения уровня экспрессии белка, описанного на стадии a), в культивированных клетках млекопитающих, культивируемых в среде без добавления тестируемого вещества; и
c) стадию определения терапевтического действия и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем выявления различия между уровнем экспрессии белка, измеренного на стадии a), и уровнем экспрессии белка, измеренным на стадии b).
(iii) Способ, включающий следующие стадии a)-c):
a) стадию иммобилизации суммарного белка, полученного из культивированных клеток млекопитающих, культивируемых в среде с добавлением тестируемого вещества;
b) стадию измерения уровня экспрессии Siglec-15 в иммобилизованном белке; и
c) стадию определения терапевтического действия и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия между уровнем экспрессии Siglec-15, определенным на стадии b), и уровнем экспрессии белка в суммарном белке, полученном из культивированных клеток млекопитающих, культивируемых в среде без добавления тестируемого вещества.
(iv) Способ, включающий следующие стадии a)-e):
a) стадию иммобилизации суммарного белка, полученного из культивированных клеток млекопитающих, культивируемых в среде с добавлением тестируемого вещества;
b) стадию иммобилизации суммарного белка, полученного из культивированных клеток млекопитающих, культивируемых в среде без добавления тестируемого вещества;
c) стадию измерения уровня экспрессии Siglec-15 в иммобилизованном белке, описанном на стадии a), используя антитело или лиганд, специфически связывающийся с этим белком;
d) стадию измерения уровня экспрессии Siglec-15 в иммобилизованном белке, описанном на стадии b), используя антитело или лиганд, специфически связывающийся с этим белком; и
e) стадию определения терапевтического действия и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия между уровнем экспрессии белка, измеренным на стадии c), и уровнем экспрессии белка, измеренным на стадии d).
(b) Способ, в котором используют отдельных млекопитающих
(i) Способ, включающий следующие стадии a) и b):
a) стадию измерения уровня экспрессии Siglec-15 в тестируемом образце, полученном от млекопитающего индивидуума, которому было введено тестируемое вещество; и
b) стадию определения терапевтического действия и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия между уровнем экспрессии Siglec-15, измеренном на стадии a), и уровнем экспрессии белка в тестируемом образце, полученном от млекопитающего индивидуума без введения тестируемого вещества.
(ii) Способ, включающий следующие стадии a)-c):
a) стадию измерения уровня экспрессии Siglec-15 в тестируемом образце, полученном от млекопитающего индивидуума, которому было введено тестируемое вещество, используя антитело или лиганд, специфически связывающийся с этим белком;
b) стадию измерения уровня экспрессии белка в тестируемом образце, полученном от млекопитающего индивидуума без введения тестируемого вещества; и
c) стадию определения терапевтического действия и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия между уровнем экспрессии Siglec-15, измеренным на стадии a), и уровнем экспрессии белка, измеренным на стадии b).
(iii) Способ, включающий следующие стадии a)-c):
a) стадию иммобилизации суммарного белка в тестируемом образце, полученном от млекопитающего индивидуума, которому было введено тестируемое вещество;
b) стадию измерения уровня экспрессии Siglec-15 в иммобилизованном белке; и
c) стадию определения терапевтического и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия между уровнем экспрессии Siglec-15, измеренным на стадии b), и уровнем экспрессии белка в тестируемом образце, полученном от млекопитающего индивидуума без введения тестируемого вещества.
(iv) Способ, включающий следующие стадии a)-e):
a) стадию иммобилизации суммарного белка в тестируемом образце, полученном от млекопитающего индивидуума, которому было введено тестируемое вещество;
b) стадию иммобилизации суммарного белка в тестируемом образце, полученном от млекопитающего индивидуума без введения тестируемого вещества;
c) стадию определения уровня экспрессии Siglec-15 в иммобилизованном белке, описанном на стадии a), используя антитело или лиганд, специфически связывающиеся с этим белком;
d) стадию определения уровня экспрессии Siglec-15 в иммобилизованном белке, описанном на стадии b), используя антитело или лиганд, специфически связывающиеся с этим белком; и
e) стадию определения терапевтического действия и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем анализа различия между уровнем экспрессии белка, описанным на стадии c), и уровнем экспрессии белка, описанным на стадии d).
(3) Способ скрининга с использованием эндогенного лиганда
Скрининг вещества, которое ингибирует дифференцировку в остеокласты, также можно проводить путем наблюдения, ингибируется ли или нет связывание эндогенного лиганда с Siglec-15 путем добавления тестируемого вещества. Примеры сиалилированных гликанов, служащих в качестве эндогенного лиганда для Siglec-15, включают Neu5Acα2-6GalNAc, связывающийся с Siglec-15 человека и мыши, и Neu5Acα2-3Galβ1-4Glc связывающийся с Siglec-15 мыши. Однако эндогенный лиганд для Siglec-15 не ограничивается этими гликанами, при условии, что он обладает аффинностью связывания в отношении Siglec-15. Эти эндогенные лиганды могут быть помечены соответствующей меткой, радиоизотопом или флуоресцентным веществом с целью изучения их связывания с Siglec-15. Например, биотинилированный полиакриламид, с которым был связан сиалилированный олигосахарид, такой как Neu5Acα2-6GalNAc, может быть использован в скрининге в качестве зонда, связывающегося с Siglec-15. В качестве Siglec-15, с которым связывается эндогенный лиганд, могут быть использованы Siglec-15-экспрессирующие клетки или мембранная фракция, полученная из этих клеток. Далее, Siglec-15 можно подвергать скринингу после выделения Siglec-15 из клеток, экспрессирующих Siglec-15, с последующей очисткой. В качестве клеток, экспрессирующих Siglec-15, могут быть использованы любые из культивированных клеточных линий, экспрессирующих Siglec-15, клеток, у которых вызвана транзиторная или непрерывная экспрессия гена Siglec-15, путем воздействия на соответствующие культивированные клетки методами генной инженерии, и клетки, экспрессирующие Siglec-15, полученные in vivo. Способ скрининга с использованием такого эндогенного лиганда можно проводить в соответствии со стадиями, описанными ниже.
(i) Способ, включающий следующие стадии a) и b):
a) стадию добавления эндогенного лиганда для Siglec-15 и тестируемого вещества к клеткам, экспрессирующим Siglec-15; и
b) стадию определения терапевтического и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем сравнения связывающегося количества эндогенного лиганда с Siglec-15-экспрессирующими клетками между случаем, когда тестируемое вещество было добавлено, и случаем, когда тестируемое вещество не добавляли.
(ii) Способ, включающий следующие стадии a)-c):
a) стадию получения фракции клеточных мембран Siglec-15-экспрессирующих клеток;
b) стадию добавления эндогенного лиганда для Siglec-15 и тестируемого вещества к фракции клеточных мембран; и
c) стадию определения терапевтического и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем сравнения количества связывания эндогенного лиганда с фракцией клеточных мембран между случаем, когда тестируемое вещество добавляли, и случаем, когда тестируемое вещество не добавляли.
(iii) Способ, включающий следующие стадии a)-c):
a) стадию получения Siglec-15;
b) стадию добавления эндогенного лиганда для Siglec-15 и тестируемого вещества к Siglec-15 в a); и
c) стадию определения терапевтического и/или профилактического действия тестируемого вещества на аномальный метаболизм костной ткани путем сравнения количества связывания эндогенного лиганда с Siglec-15 между случаем, когда тестируемое вещество добавляли, и случаем, когда тестируемое вещество не добавляли.
В случае, когда в соответствующих клетках вызвана экспрессия Siglec-15 методами генной инженерии и полученный в результате Siglec-15 очищают и подвергают скринингу, используемый в скрининге Siglec-15 может быть выбран из полипептидов, состоящих из аминокислотных последовательностей, показанных в следующих пунктах (a)-(i):
(a) аминокислотной последовательности, представленной последовательностью SEQ ID NO:2 в перечне последовательностей;
(b) аминокислотной последовательности, состоящей из аминокислотных остатков 21-328 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(c) аминокислотной последовательности, состоящей из аминокислотных остатков 1-260 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(d) аминокислотной последовательности, состоящей из аминокислотных остатков 21-260 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(e) аминокислотной последовательности, представленной последовательностью SEQ ID NO:4 в перечне последовательностей;
(f) аминокислотной последовательности, состоящей из аминокислотных остатков 21-341 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей;
(g) аминокислотной последовательности, состоящей из аминокислотных остатков 1-258 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей;
(h) аминокислотной последовательности, состоящей из аминокислотных остатков 21-258 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей; и
(i) аминокислотной последовательности, включающей замены, делеции или добавления одного или нескольких аминокислотных остатков в аминокислотной последовательности, описанной в пунктах (a)-(h).
Используемый в скрининге Siglec-15 также может быть выбран из полипептидов, состоящих из аминокислотных последовательностей, кодируемых нуклеотидными последовательностями, показанными в следующих пунктах (j)-(n):
(j) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:1;
(k) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:3;
(l) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:19;
(m) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:43; и
(n) нуклеотидной последовательностью полинуклеотида, который гибридизируется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, описанной в пунктах (j)-(m) в строгих условиях.
Далее, также можно использовать полипептид, полученный путем присоединения соответствующей метки к любому из этих полипептидов, или полипептид, слитый с другим растворимым белком, в качестве мишени для скрининга. К тому же, полипептид, состоящий из аминокислотных остатков 1-20 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей, соответствует сигнальному пептиду Siglec-15 человека, и полипептид, состоящий из аминокислотных остатков 21-260, соответствует внеклеточному домену зрелого белка Siglec-15 человека. Далее, полипептид, состоящий из аминокислотных остатков 1-20 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей, соответствует сигнальному пептиду Siglec-15 мыши, и полипептид, состоящий из аминокислотных остатков 21-258, соответствует внеклеточному домену зрелого белка мыши Siglec-15. Далее, нуклеотидная последовательность, представленная последовательностью SEQ ID NO:43, кодирует внеклеточный домен Siglec-15 человека, кодируемый нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:1, и нуклеотидная последовательность, представленная последовательностью SEQ ID NO:19, кодирует внеклеточный домен Siglec-15 мыши, кодируемый нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:3.
Кандидатные вещества для вещества, которое ингибирует дифференцировку в остеокласты, выбранные с помощью любого из способов скрининга (1)-(3), могут быть вторично оценены путем ингибирования активности тартрат-резистентной кислой фосфатазы (TRAP) остеокластов в качестве показателя, как показано в Примерах 17, 19 и 20. Далее, их вторичную оценку также можно проводить путем ингибирования образования TRAP-позитивных многоядерных остеокластов, т.е. ингибирования клеточного слияния остеокластов в качестве показателя, как показано в Примерах 19, 21, 22 и 35.
(4) Другие способы
Частоту возникновения аномального метаболизма костной ткани с течением времени, степень тяжести аномального метаболизма костной ткани и/или частоту выживания и т.д. определяют для того случая, когда тестируемое вещество вводили млекопитающему индивидууму, у которого вызвана гиперэкспрессия Siglec-15, и случая, когда тестируемое вещество ему не вводят. В том случае, когда у млекопитающего, которому вводят тестируемое соединение, частота возникновения аномального метаболизма костной ткани значительно снижается, степень тяжести аномального метаболизма костной ткани значительно ниже, и/или частота выживаемости повышается примерно на 10% или более, предпочтительно примерно на 30% или более, более предпочтительно примерно на 50% или более, тестируемое вещество может быть выбрано в качестве соединения, обладающего терапевтическим и/или профилактическим действием на аномальный метаболизм костной ткани.
4. Получение антитела против Siglec-15
Антитело против Siglec-15 по настоящему изобретению может быть получено путем иммунизации животного Siglec-15 или произвольным полипептидом, выбранным из аминокислотной последовательности Siglec-15, и получения и очистки этого антитела, продуцированного in vivo в соответствии с общепринятым способом. Биологические виды используемого Siglec-15 в качестве антигена не ограничены человеком, и животное может быть иммунизировано Siglec-15, полученным от животного, отличного от человека, такого как мышь или крыса. В этом случае, путем изучения перекрестной реактивности между антителом, связывающимся с полученым гетерологичным Siglec-15 и Siglec-15 человека, может быть выбрано антитело, применимое при заболевании человека.
Далее, моноклональное антитело может быть получено путем слияния антитело-продуцирующих клеток, которые продуцируют антитело против Siglec-15, с миеломными клетками для создания гибридомы в соответствии с известным способом (например, Kohler and Milstein, Nature, (1975) 256, pp. 495-497; Kennet, R. ed., Monoclonal Antibody, pp. 365-367, Prenum Press, N.Y. (1980)).
Siglec-15, используемый в качестве антигена, может быть получен методами генной инженерии для того, чтобы вызвать экспрессию гена Siglec-15 в клетке-хозяине.
В частности, методы генной инженерии можно проводить следующим образом. Получают вектор, способный экспрессировать ген Siglec-15, и полученный в результате вектор трансфицируют в клетку-хозяина для экспрессии этого гена и затем очищают экспрессированный Siglec-15. В дальнейшем способ получения антитела против Siglec-15 будет описан конкретно.
(1) Получение антигена
Примеры антигена, используемого для получения антитела против Siglec-15, включают Siglec-15, полипептид, состоящий из неполной аминокислотной последовательности, состоящей по меньшей мере из 6 последовательных аминокислотных остатков Siglec-15, и производное, полученное путем добавления к ней заданной аминокислотной последовательности или носителя. Далее, в разделе «3. Способ скрининга вещества, которое ингибирует дифференцировку в остеокласты», Siglec-15, приведенный в качестве примера как мишень для скрининга, также может быть приведен в качестве примера как антиген, используемый для получения антитела против Siglec-15.
Siglec-15 может быть использован после очистки непосредственно из опухолевых тканей или опухолевых клеток человека. Далее, Siglec-15 может быть получен путем синтеза его in vitro или вызывая методами генной инженерии его продукцию клеткой-хозяином.
В методах генной инженерии, в частности, кДНК Siglec-15 встраивают в вектор, способный экспрессировать кДНК Siglec-15, и Siglec-15 синтезируют в растворе, содержащем ферменты, субстраты и энергетические вещества, необходимые для транскрипции и трансляции, или другую прокариотическую или эукариотическую клетку-хозяин трансформируют для экспрессии Siglec-15, посредством чего может быть получен антиген.
Далее, антиген также может быть получен в виде секреторного белка путем экспрессии слитого белка, полученного лигированием внеклеточного домена Siglec-15, который является мембранным белком, с константной областью антитела в соответствующей системе хозяин-вектор.
кДНК Siglec-15 может быть получена, например, с помощью так называемого ПЦР метода, в котором полимеразно-цепную реакцию (здесь и далее называемую «ПЦР») проводят, используя библиотеку кДНК, экспрессирующую кДНК Siglec-15 в виде матрицы и праймеры, которые специфически амплифицируют кДНК Siglec-15 (см. Saiki, R. K., et al., Science, (1988) 239, pp. 487-489).
В качестве системы синтеза полипептида in vitro, например, может служить примером система «Rapid Translation System» (RTS), производства Roche Diagnostics, Inc., но не ограничиваться только этим примером.
Примеры прокариотических хозяев включают Escherichia coli и Bacillus subtilis. Для трансформации клетки-хозяина целевым геном клетку-хозяин трансформируют, используя плазмидный вектор, содержащий репликон, т.е. сайт инициации репликации, полученный из видов, совместимых с хозяином, и регуляторную последовательность. Далее, вектор предпочтительно имеет последовательность, способную придавать трансформированным клеткам фенотипическую селективность.
Примеры эукариотических клеток-хозяев включают клетки позвоночных, клетки насекомых и клетки дрожжей. В качестве клеток позвоночных, например, зачастую используют штаммы с недостаточностью дигидрофолатредуктазы (Urlaub, G. and Chasin, L. A., Proc. Natl. Acad. Sci. USA (1980) 77, pp. 4126-4220) of simian COS cells (Gluzman, Y., Cell, (1981) 23, pp. 175-182, ATCC CRL-1650), фибробласты мыши NIH3T3 (ATCC No. CRL-1658), и ооциты китайского хомячка (CHO cells; ATCC: CCL-61); и подобные, однако не ограничиваются ими.
Полученный таким образом трансформант может быть культивирован в соответствии с общепринятой процедурой, и путем культивирования этого трансформанта целевой полипептид продуцируется внутриклеточно или внеклеточно.
Подходящая среда, используемая для культивирования, может быть выбрана из различных обычно используемых культуральных сред в зависимости от используемой клетки-хозяина. В случае использования Escherichia coli можно использовать среду LB, дополненную антибиотиком, таким как ампициллин или IPMG, при необходимости.
Рекомбинантный белок, продуцируемый трансформантом внутриклеточно или внеклеточно, при культивировании может быть отделен и очищен любым из различных известных способов разделения, используя физические или химические свойства этого белка.
Конкретные примеры таких способов включают обработку стандартным осаждающим белок реагентом, ультрафильтрацию, различные типы жидкостной хроматографии, такие как хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография и аффинная хроматография, диализ и их сочетания.
Далее, добавляя шесть остатков гистидина к экспрессируемому рекомбинантному белку, этот белок может быть эффективно очищен с помощью аффинной колонки с никелевым сорбентом. Альтернативно, путем присоединения Fc-области IgG к экспрессируемому рекомбинантному белку белок может быть эффективно очищен с помощью колонки с Белком А.
Путем сочетания приведенных выше способов огромное количество целевого полипептида можно легко получить с высоким выходом и высокой чистоты.
(2) Получение анти-Siglec-15 моноклонального антитела
Примеры антитела, специфически связывающегося с Siglec-15, включают моноклональное антитело, специфически связывающееся с Siglec-15, и способ получения такого антитела описан ниже.
Для получения моноклонального антитела обычно требуются следующие операционные стадии:
(a) очистки биополимера, используемого в качестве антигена;
(b) получения антитело-продуцирующих клеток путем иммунизации животного инъекцией антигена, сбора крови, исследования титра антител и определения, когда селезенка подлежит резекции;
(c) получения миеломных клеток (здесь и далее называемых «миеломой»);
(d) слияния антитело-продуцирующих клеток с миеломой;
(e) скрининга группы гибридом, продуцирующим целевое антитело;
(f) деления гибридом на отдельные клеточные клоны (клонирование);
(g) необязательно, культивирования гибридомы или выращивания животного с имплантированной гибридомой для получения большого количества моноклональных антител;
(h) исследование биологической активности и специфичности связывания полученного таким образом моноклонального антитела, или исследование тех же свойств с меченым реагентом; и подобное.
В дальнейшем, способ получения моноклонального антитела будет описан подробно вслед за приведенными выше стадиями, однако этот способ не ограничивается ими, и, например, могут быть использованы антитело-продуцирующие клетки и миелома, отличные от клеток селезенки.
(a) Очистка антигена
В качестве антигена может быть использован Siglec-15, полученный способом, описанным выше, или его неполный пептид.
Далее, в качестве антигена также может быть использована мембранная фракция, полученная из рекомбинантных клеток, экспрессирующих Siglec-15, или сами рекомбинантные клетки, экспрессирующие Siglec-15, а также неполный пептид белка по изобретению, химически синтезированный способом, известным специалистам в данной области.
(b) Получение антитело-продуцирующих клеток
Антиген, полученный на стадии (a), смешивают с адъювантом, таким как полный или неполный адъювант Фрейнда или калиево-алюминиевые квасцы, и полученную в результате смесь используют в качестве иммуногена для иммунизации экспериментального животного. В качестве экспериментального животного любое животное, используемое в известном способе получения гибридомы, может быть использовано без каких-либо проблем. В частности, например, могут быть использованы мыши, крысы, козы, овцы, крупный рогатый скот, лошади или подобные. Однако, с точки зрения удобства применения миеломных клеток, подлежащих слиянию с экстрагированными антитело-продуцирующими клетками, в качестве иммунизируемого животного предпочтительно используются мыши или крысы.
Далее, фактически используемая линия мышей или крыс, конкретно не ограничена, и в случае мышей, например, могут быть использованы различные линии, такие как A, AKR, BALB/c, BDP, BA, CE, C3H, 57BL, C57BL, C57L, DBA, FL, HTH, HT1, LP, NZB, NZW, RF, R III, SJL, SWR, WB и 129, и в случае крысы, например, могут быть использованы Wistar, Low, Lewis, Sprague Dawley, ACI, BN, Fischer и подобные.
Эти мыши и крысы являются коммерчески доступными от заводчиков/дистрибьютеров экспериментальных животных, например, CLEA Japan, Inc. и Charles River Laboratories Japan, Inc.
Среди них, с учетом совместимости слияния с миеломными клетками, описанными ниже, особенно предпочтительными в случае мышей является линия BALB/c, а в случае крыс линии Wistar и Low.
Далее, с учетом антигенной гомологии между людьми и мышами также предпочтительно использовать мышей со сниженной биологической функцией удаления аутоантител, то есть мышей с аутоиммунным заболеванием.
Возраст мышей или крыс во время иммунизации предпочтительно составляет от 5 до 12 недель, более предпочтительно от 6 до 8 недель.
Известный способ для иммунизации животного Siglec-15 или его рекомбинантной формой, например, описан подробно, например, в Weir, D. M., Handbook of Experimental Immunology Vol. I. II. III., Blackwell Scientific Publications, Oxford (1987), Kabat, E. A. and Mayer, M. M., Experimental Immunochemistry, Charles C Thomas Publisher Spigfield, Illinois (1964), или подобный.
Среди этих способов иммунизации предпочтительным специфическим способом в настоящем изобретении является, например, следующий.
Таким образом, сначала фракцию мембранных белков, служащих антигеном, или клетки, в которых вызывается экспрессия антигена, вводят животному внутрикожно или внутрибрюшинно.
Однако сочетание обоих путей введения является предпочтительным для повышения эффективности иммунизации, и в том случае, когда внутрикожное введение проводят в первой половине, а внутрибрюшинную инъекцию проводят во второй половине или только при последнем введении дозы, эффективность иммунизации может быть особенно повышена.
Схема введения антигена варьирует в зависимости от типа иммунизируемого животного, индивидуальных отличий или подобного. Однако в основном частота введения антигена предпочтительно составляет от 3 до 6 раз, более предпочтительно от 3 до 4 раз, и интервал дозирования предпочтительно составляет от 2 до 6 недель, более предпочтительно от 2 до 4 недель.
Далее, доза антигена варьирует в зависимости от вида иммунизируемого животного, индивидуальных отличий или подобного. Однако дозу в основном устанавливают до 0,05-5 мг, предпочтительно примерно от 0,1 до 0,5 мг.
Повторную иммунизацию проводят через 1-6 недель, предпочтительно через 2-4 недели, более предпочтительно через 2-3 недели, после введения антигена, как описано выше.
Доза антигена во время проведения повторной иммунизации варьирует в зависимости от вида или размера животного, или подобного, однако, в случае мышей, дозу обычно устанавливают на 0,05-5 мг, предпочтительно от 0,1 до 0,5 мг, более предпочтительно примерно от 0,1 до 0,2 мг.
Клетки селезенки или лимфоциты, в том числе антитело-продуцирующие клетки, асептически удаляют из иммунизированного животного через 1-10 дней, предпочтительно через 2-5 дней, более предпочтительно через 2-3 дня, после повторной иммунизации.
В это время измеряют титр антител, и в том случае, если животное, имеющее достаточно повышенный титр антител, используется в качестве источника антитело-продуцирующих клеток, последующую процедуру можно проводить более эффективно.
Способ измерения титра антител, используемый в настоящем изобретении, включает, например, способ RIA и способ ELISA, но не ограничивается ими.
Например, в том случае, если используется способ ELISA, измерение титра антител в настоящем изобретении можно проводить в соответствии с процедурами, описанными ниже.
Сначала очищенный или частично очищенный антиген адсорбируют на поверхности твердой фазы, например 96-луночном планшете для ELISA, а поверхность твердой фазы, не имеющей адсорбированного на ней антигена, покрывают белком, неродственным этому антигену, таким как бычий сывороточный альбумин (в дальнейшем называемым «BSA»). После промывания поверхности эту поверхность приводят в контакт с серийно разведенным образцом (например, сывороткой мыши), в качестве первичного антитела, чтобы дать возможность антителу, содержащемуся в образце, связаться с этим антигеном.
Далее, в качестве вторичного антитела, добавляют антитело, меченное ферментом против антитела мыши, и дают ему возможность связаться с антителом мыши. После промывания добавляют субстрат для фермента и измеряют поглощение, которое происходит вследствие развития окраски, индуцированного разрушением субстрата или подобного, для вычисления титра антител.
Отделение антитело-продуцирующих клеток от клеток селезенки или лимфоцитов можно проводить в соответствии с известным способом (например, Kohler et al., Nature (1975), 256, p. 495; Kohler et al., Eur. J. Immunol. (1977), 6, p. 511; Milstein et al., Nature (1977), 266, p. 550; Walsh, Nature (1977), 266, p. 495).
Например, в случае клеток селезенки можно использовать общий способ, в котором антитело-продуцирующие клетки отделяют путем гомогенизации селезенки для получения клеток путем фильтрования через сито из нержавеющей стали и суспендирования клеток в минимальной эссенциальной среде Игла (MEM).
(c) Получение миеломных клеток (в дальнейшем называемых «миеломой»)
Миеломные клетки, используемые для слияния клеток, конкретно не ограничены, и подходящие клетки могут быть выбраны из известных клеточных линий. Однако, принимая во внимание преимущества, когда гибридома выбрана из слитых клеток, предпочтительно использовать штаммы с недостаточностью HGPRT (гипоксантин-гуанинфосфорибозил трансферазы), процедура отбора которых была установлена.
Более конкретно, примеры штамма с недостаточностью HGPRT включают X63-Ag8(X63), NS1-ANS/1(NS1), P3X63-Ag8.U1(P3U1), X63-Ag8.653(X63.653), SP2/0-Ag14(SP2/0), MPC11-45.6TG1.7(45.6TG), F0, S149/5XXO и BU.1, полученные от мышей, 210.RSY3.Ag.1.2.3(Y3), полученные от крыс; и U266AR(SKO-007), GM1500.GTG-A12(GM1500), UC729-6, LICR-LOW-HMy2(HMy2) и 8226AR/NIP4-1(NP41), полученные от людей.
Эти штаммы с недостаточностью HGPRT доступны, например, из Американской коллекции типовых культур (ATCC) или подобных.
Эти штаммы клеток субкультивируют в соответствующей среде, такой как среда с 8-азагуанином [среда, полученная путем добавления 8-азагуанина к среде RPMI-1640, дополненной глутамином, 2-меркаптоэтанолом, гентамицином и эмбриональной сывороткой теленка (в дальнейшем называемой FCS»)]; среда Дульбекко, модифицированная по способу Исков (в дальнейшем называемая «IMDM»), или модифицированная по Дульбекко среда Игла (в дальнейшем называемая «DMEM»). В этом случае, за 3-4 дня до проведения слияния клеток, клетки субкультивируют в нормальной среде [например, среде ASF104 (изготовленной Ajinomoto Co., Ltd.), содержащей 10% FCS] для получения не менее 2×107 клеток в день проведения слияния клеток.
(d) Слияние клеток
Слияние между антитело-продуцирующими клетками и миеломными клетками соответствующим образом проводят в соответствии с известным способом (Weir, D. M. Handbook of Experimental Immunology Vol. I. II. III., Blackwell Scientific Publications, Oxford (1987), Kabat, E. A. and Mayer, M. M., Experimental Immunochemistry, Charles C Thomas Publisher, Springfield, Illinois (1964), etc.), в таких условиях, чтобы коэффициент выживания клеток чрезмерно не снижался.
Примеры этого способа включают химический способ, в котором антитело-продуцирующие клетки и миеломные клетки перемешивают в полимерном растворе полиэтиленгликоля высокой концентрации или подобном, и физический способ с использованием электрической стимуляции.
Среди этих способов конкретным примером химического способа является описанный ниже.
Таким образом, в том случае, когда полиэтиленгликоль используют для полимерного раствора высокой концентрации, антитело-продуцирующие клетки и миеломные клетки перемешивают в растворе полиэтиленгликоля с молекулярной массой 1500-6000, более предпочтительно 2000-4000 при температуре от 30 до 40°C, предпочтительно от 35 до 38°C в течение от 1 до 10 минут, предпочтительно от 5 до 8 минут.
(e) Выбор группы гибридом
Способ выбора гибридом, полученных упомянутым выше слиянием клеток, конкретным образом не ограничен. Обычно используют способ селекции HAT (гипоксантин, аминоптерин, тимидин) (Kohler et al., Nature (1975), 256, p. 495; Milstein et al., Nature (1977), 266, p. 550).
Этот способ является эффективным, когда гибридомы получают, используя миеломные клетки штамма с недостаточностью HGPRT, неспособные выживать в присутствии аминоптерина.
Таким образом, путем культивирования неслитых клеток и гибридом в среде HAT только гибридомы, резистентные к аминоптерину, имеют селективную возможность для выживания и пролиферации.
(f) Деление на одноклеточный клон (клонирование)
В качестве способа клонирования для гибридом может быть использован известный способ, такой как метилцеллюлозный способ, способ мягкой агарозы или метод предельных разведений (см., например, Barbara, B. M. and Stanley, M. S.: Selected Methods in Cellular Immunology, W. H. Freeman and Company, San Francisco (1980)). Среди этих способов, в частности, предпочтительным является метод предельных разведений.
В этом способе клеточный штамм фибробластов, полученный из эмбриона крысы или клеток-кормилиц, таких как нормальные клетки селезенки мыши, клетки тимуса или клетки асцитической жидкости, высевают на микропланшет.
При этом гибридомы разводят в среде для получения плотности клеток от 0,2 до 0,5 клеток на 0,2 мл. Аликвоту 0,1 мл разведенной гибридомной суспензии помещают в каждую лунку и культивирование продолжают примерно в течение 2 недель, заменяя примерно 1/3 культурального раствора свежей средой в заранее определенные интервалы времени (например, каждые 3 дня), за счет чего гибридомные клоны могут пролиферировать.
Гибридомы в лунках, для которых титр антител был подтвержден, подвергают, например, клонированию методом предельных разведений многократно от 2 до 4 раз. Гибридома, у которой было подтверждено наличие стабильного титра антител, выбирают в качестве гибридомного штамма, продуцирующего моноклональные антитела против Siglec-15.
Примеры гибридомного штамма, клонированного таким образом, включают гибридому #32A1 и гибридому #41B1. Гибридома #32A1 и гибридома #41B1 были депонированы в международном депозитарии «International Patent Organism Depositary» национального института «National Institute of Advanced Industrial Science and Technology» (расположенного в Central 6, 1-1-1 Higashi, Tsukuba-shi, Ibaraki-ken, 305-8566, Япония) 28 августа 2008. Гибридоме #32A1 был присвоен депозитный номер FERM BP-10999 под названием анти-Siglec-15 гибридома #32A1, и гибридоме #41B1 был присвоен депозитный номер FERM BP-11000 под названием анти-Siglec-15 гибридома #41B1. В этом описании антитело, продуцируемое гибридомой #32A1, представлено «#32A1 антителом» или просто «#32A1», и антитело, продуцируемое гибридомой #41B1, представлено «#41B1 антителом» или просто «#41B1». Далее, антитела, полученные в Примерах в этом описании, отличные от гибридомы #32A1 и гибридомы #41B1, также представлены названиями антител аналогичным образом.
(g) Получение моноклональных антител путем культивирования гибридомы
Моноклональные антитела могут быть эффективно получены путем культивирования отобранной таким образом гибридомы. Однако перед культивированием предпочтительно проводить скрининг гибридомы, которая продуцирует целевое моноклональное антитело.
В этом скрининге может быть использован известный способ.
Измерение титра антител в настоящем изобретении можно проводить, например, с помощью ELISA, изложенного в пункте (b), описанном выше.
Гибридому, полученную описанным выше способом, можно хранить в замороженном виде в жидком азоте или в морозильной камере при -80°C или ниже.
После завершения клонирования среду меняют со среды HT на нормальную среду и гибридому культивируют.
Массовое культивирование проводят путем вращения культуры, используя большой культуральный сосуд, или с помощью роллерного метода.
Из супернатанта, полученного путем массового культивирования, моноклональное антитело, которое специфически связывается с белком по изобретению, может быть получено с помощью очистки, используя способ, известный специалистам в данной области, такой как гель-фильтрация.
Далее, гибридому вводят в брюшную полость мыши той же линии, что и гибридома (например, упомянутые выше BALB/c) или мыши Nu/Nu для пролиферации гибридомы, посредством чего может быть получена асцитическая жидкость, содержащая большое количество моноклональных антител по настоящему изобретению.
В том случае, когда гибридому вводят в брюшную полость, при введении минерального масла, такого как 2,6,10,14-тетраметилпентадекан (пристин) за 3-7 дней до этого, может быть получено большее количество асцитической жидкости.
Например, иммуносупрессант предварительно вводят в брюшную полость мыши той же линии, что и гибридома, для инактивации Т-клеток. Через 20 дней после этого 106-107 клеток гибридомного клона суспендируют в бессывороточной среде (0,5 мл) и эту суспензию вводят в брюшную полость мыши. В основном, когда брюшная полость увеличивается в объеме и наполняется асцитической жидкостью, асцитическую жидкость собирают.
С помощью этого способа моноклональные антитела могут быть получены в концентрации примерно в 100 раз выше, чем концентрация культурального раствора.
Моноклональное антитело, полученное упомянутым выше способом, может быть очищено способом, описанным, например, в Weir, D. M.: Handbook of Experimental Immunology Vol. I, II, III, Blackwell Scientific Publications, Oxford (1978).
Таким образом, примеры способа включают метод преципитации сульфатом аммония, гель-фильтрацию, ионообменную хроматографию и аффинную хроматографию.
Также может быть использован простой способ очистки, коммерчески доступный набор для очистки моноклональных антител (например, набор MAbTrap GII, производства Pharmacia, Inc.) или подобный.
Полученное таким образом моноклональное антитело обладает высокой антигенной специфичностью в отношении Siglec-15.
(h) Исследование моноклонального антитела
Изотип и подкласс полученного таким образом моноклонального антитела можно определить следующим образом.
Во-первых, примеры способа идентификации включают способ по Оухтерлони, метод ELISA и метод RIA.
Способ по Оухтерлони является простым, но когда концентрация моноклонального антитела является низкой, требуется процесс конденсации.
С другой стороны, при использовании метода ELISA или метода RIA, путем непосредственного взаимодействия культурального супернатанта с антиген-адсорбированной твердой фазой и с использованием антител, соответствующих различным типам изотиов иммуноглобулинов и подклассам в качестве вторичных антител, можно идентифицировать изотип и подкласс моноклонального антитела.
Кроме того, в качестве более простого способа также может быть использован коммерчески доступный набор для идентификации (например, набор Mouse Typer, производства Bio-Rad Laboratories, Inc.) или подобный.
Далее, количественное определение белка можно проводить по методу Фолина Лоури, и способ вычисления основан на поглощении при 280 нм [1,4 (OD 280) = иммуноглобулин 1 мг/мл].
(3) Другие антитела
Антитело по настоящему изобретению включает не только упомянутое выше моноклональное антитело против Siglec-15, а также рекомбинантное антитело, полученное путем искусственной модификации с целью снижения гетерологичной антигенности для людей, например, химерное антитело, гуманизированное антитело и антитело человека. Эти антитела можно получать, используя известный способ.
В качестве химерного антитела можно привести пример антитела, в котором вариабельные и константные области антитела происходят из различных видов, например, химерное антитело, в котором вариабельная область антитела полученная от мыши, соединена с константной областью, полученной от человека (см. Proc. Natl. Acad. Sci. USA, 81, 6851-6855, (1984)).
В качестве гуманизированного антитела можно привести пример антитела, полученного путем интеграции только области, определяющей комплементарность (CDR) в антитело, полученное от человека (см. Nature (1986) 321, pp. 522-525), и антитела, полученного путем трансплантации части аминокислотных остатков каркасной области, а также CDR последовательности в антитело человека методом CDR-трансплантации (WO 90/07861).
Далее, антитело по настоящему изобретению включает антитело человека. Антитело человека против Siglec-15 относится к антителу человека, имеющему только генную последовательность антитела, происходящую из хромосомы человека. Антитело человека против Siglec-15 может быть получено способом с использованием мышей, продуцирующих антитела человека, имеющих фрагмент хромосомы человека, содержащий гены H-цепи и L-цепи антитела человека (см. Tomizuka, K. et al., Nature Genetics (1997) 16, pp. 133-143; Kuroiwa, Y. et al., Nuc. Acids Res. (1998) 26, pp. 3447-3448; Yoshida, H. et al., Animal Cell Technology: Basic and Applied Aspects vol. 10, pp. 69-73 (Kitagawa, Y., Matuda, T. and Iijima, S. eds.), Kluwer Academic Publishers, 1999; Tomizuka, K. et al., Proc. Natl. Acad. Sci. USA (2000) 97, pp. 722-727, etc.).
Такое трансгенное животное может быть получено, в частности, следующим образом. Генетически модифицированное животное, у которого были разрушены генные локусы тяжелой и легкой цепи эндогенного иммуноглобулина млекопитающего, не являющегося человеком, и вместо этого, были введены генные локусы тяжелой и легкой цепи иммуноглобулина человека, через вектор на основе искусственной хромосомы дрожжей (YAC) или подобного, создают путем получения «нокаут» животного и трансгенного животного и скрещивания этих животных.
Далее, в соответствии с методиками генной инженерии, используя кДНК, кодирующие такую тяжелую цепь и легкую цепь антитела человека, соответственно, предпочтительно, вектор, содержащий кДНК, трансформируют эукариотические клетки, и культивируют трансформант, который продуцирует рекомбинантное моноклональное антитело человека, посредством чего из культурального супернатанта может быть получено это антитело.
В настоящем изобретении в качестве хозяина, например, могут быть использованы эукариотические клетки, предпочтительно клетки млекопитающих, такие как клетки CHO, лимфоциты или миеломные клетки.
Далее, также известен способ получения антитела человека, полученного из фагового дисплея, отобранного из библиотеки антител человека (см. Wormstone, I. M. et al., Investigative Ophthalmology & Visual Science. (2002) 43 (7), pp. 2301-2308; Carmen, S. et al., Briefings in Functional Genomics and Proteomics (2002), 1 (2), pp. 189-203; Siriwardena, D. et al., Opthalmology (2002) 109 (3), pp. 427-431, etc.).
Например, может быть использован метод фагового дисплея, в котором вариабельная область антитела человека экспрессируется на поверхности фага в виде одноцепочечного антитела (scFv), и выбирают фаг, который связывается с антигеном (Nature Biotechnology (2005), 23, (9), pp. 1105-1116).
Путем анализа гена фага, выбранного на основе связывания с антигеном, можно определить последовательность ДНК, кодирующую вариабельную область антитела человека, которое связывается с этим антигеном.
Если последовательность ДНК scFv, который связывается с антигеном, определена, антитело человека может быть получено путем получения вектора экспрессии, имеющего эту последовательность, и введения этого вектора в соответствующую клетку-хозяин для ее экспрессии (WO 92/01047, WO 92/20791, WO 93/06213, WO 93/11236, WO 93/19172, WO 95/01438, WO 95/15388, Annu. Rev. Immunol (1994) 12, pp. 433-455, Nature Biotechnology (2005) 23 (9), pp. 1105-1116).
В том случае, когда антитело получают после выделения гена антитела, а затем введения этого гена в соответствующего хозяина, можно использовать сочетание соответствующего хозяина и вектора экспрессии.
В том случае, когда эукариотические клетки используют в качестве хозяина, могут быть использованы клетки животных, клетки растений и эукариотические микроорганизмы.
В качестве клеток животных, в качестве примера можно привести клетки млекопитающих, например штаммы с недостаточностью дигидрофолатредуктазы (Urlaub, G. and Chasin, L. A., Proc. Natl. Acad. Sci. USA (1980) 77, pp. 4126-4220) обезьяньи клетки COS (Gluzman, Y., Cell, (1981) 23, pp. 175-182, ATCC CRL-1650), фибробласты мыши NIH3T3 (ATCC No. CRL-1658) и ооциты китайского хомячка (клетки CHO; ATCC: CCL-61).
В тех случаях, когда используют прокариотические клетки, например, в качестве примера можно привести Escherichia coli и Bacillus subtilis.
Антитело может быть получено путем введения гена целевого антитела в эти клетки посредством трансформации и культивирования трансформированных таким образом клеток in vitro.
Не существует ограничения по изотипу антитела по изобретению, и примеры изотипов включают IgG (IgG1, IgG2, IgG3, IgG4), IgM, IgA (IgA1, IgA2), IgD и IgE, и предпочтительные примеры изотипов включают IgG и IgM.
Далее, антитело по настоящему изобретению может представлять собой функциональный фрагмент антитела, имеющего антиген-связывающий сайт антитела или его модифицированный фрагмент. Фрагмент этого антитела может быть получен путем обработки антитела протеазой, такой как папаин или пепсин, или модификацией гена антитела в соответствии с методами генной инженерии и экспрессии модифицированного гена в подходящих культивируемых клетках. Среди этих фрагментов антитела фрагмент, обладающий всеми или частью функций полноразмерной молекулы, может быть назван функциональным фрагментом антитела. В качестве функций антитела в основном в качестве примера можно привести антиген-связывающую активность, нейтрализующую активность в отношении активности антигена, активность в отношении повышения активности антигена, антитело-зависимую цитотоксическую активность, комплемент-зависимую цитотоксическую активность и комплемент-зависимую клеточную цитотоксическую активность. Функцией функционального фрагмента антитела по настоящему изобретению предпочтительно является ингибирующая активность образования остеокластов, более предпочтительно ингибирующая активность процесса клеточного слияния остеокластов.
Примеры фрагмента антитела включают Fab, F(ab')2, Fv, одноцепочечный Fv (scFv), в котором Fv молекулы тяжелой цепи и легкой цепи связаны через соответствующий линкер, диатело (диатела), линейное антитело, и полиспецифическое антитело, состоящее из фрагмента антитела. Далее, Fab', который представляет собой моновалентный фрагмент в вариабельной области антитела, полученный путем обработки F(ab')2 в редуцирующих условиях, также включен в фрагмент антитела.
Далее, антитело по изобретению может представлять собой полиспецифичное антитело со специфичностью по меньшей мере в отношении двух различных антигенов.
В основном такая молекула связывается с двумя антигенами (то есть биспецифичное антитело), однако «полиспецифичное антитело» в контексте настоящего изобретения включает антитело, имеющее специфичность в отношении двух или более (например, трех) антигенов.
Антитело по настоящему изобретению может быть полиспецифичным антителом, состоящим из полноразмерного антитела или фрагмента такого антитела (например, F(ab')2 биспецифичное антитело). Биспецифичное антитело может быть получено путем лигирования тяжелой и легкой цепей (HL пары) двух типов антител, или также может быть получено путем слияния гибридом, которые продуцируют различные моноклональные антитела для получения слитых клеток, которые продуцируют биспецифичное антитело (Millstein et al., Nature (1983) 305, pp. 537-539).
Антитело по настоящему изобретению может быть одноцепочечным антителом (также называемым scFv). Одноцепочечное антитело может быть получено путем лигирования V областей тяжелой цепи и легкой цепи антитела через полипептидный линкер (Pluckthun, The Pharmacology of Monoclonal Antibodies, 113 (под редакцией Rosenburg and Moore, Springer Verlag, New York, pp. 269-315 (1994), Nature Biotechnology (2005), 23, pp. 1126-1136). Далее, BiscFv фрагмент, полученный путем лигирования двух молекул scFv через полипептидный линкер, также может быть использован в качестве биспецифичного антитела.
Способ получения одноцепочечного антитела известен в этой области техники (см., например, патенты США №№ 4946778, 5260203, 5091513, 5455030, и т.д.). В этом scFv V области тяжелой цепи и легкой цепи лигируют посредством линкера, который не образует конъюгат, предпочтительно посредством полипептидного линкера (Huston, J. S. et al., Proc. Natl. Acad. Sci. USA (1988), 85, pp. 5879-5883). В scFv V области тяжелой цепи и легкой цепи могут быть получены из того же антитела или из разных антител. В качестве полипептидного линкера, используемого для лигирования V областей, например, используют заданный одноцепочечный полипептид, состоящий из 12-19 остатков.
ДНК, кодирующая scFv, может быть получена путем проведения амплификации с использованием неполной ДНК, кодирующей полную или желаемую аминокислотную последовательность ДНК, выбранную из ДНК, кодирующей тяжелую цепь или V область тяжелой цепи упомянутого выше антитела и ДНК, кодирующей его легкую цепь или V область легкой цепи в качестве матрицы с помощью метода ПЦР, используя пару праймеров, которые распознают оба его конца, и в дальнейшем проводя амплификацию, объединяя ДНК, кодирующую часть полипептидного линкера и пару праймеров, которые распознают оба его конца, так чтобы лигировать оба конца с тяжелой цепью и легкой цепью соответственно.
Далее, после получения ДНК, кодирующей scFv, содержащий ее вектор экспрессии, и хозяин, трансформированный этим вектором экспрессии, может быть получен в соответствии с общепринятым способом. Далее, scFv можно получить в соответствии с общепринятым способом использованием полученного в результате хозяина.
Фрагмент антитела может быть получен в хозяине путем получения гена и экспрессии этого гена таким же образом, как описано выше.
Антитело по настоящему изобретению может быть мультимеризовано для повышения аффинности к антигену. Мультимеризуемое антитело может представлять собой один тип антитела или множество антител, которые распознают множество эпитопов того же антигена. В качестве способа мультимеризации антитела можно привести пример связывания CH3 домена IgG с двумя молекулами scFv, связывание со стрептавидином, введение мотива спираль-поворот-спираль, и подобное.
Антитело по настоящему изобретению может быть поликлональным антителом, которое представляет собой смесь многочисленных типов анти-Siglec-15 антител, имеющих различные аминокислотные последовательности. В качестве одного примера поликлонального антитела можно привести в пример смесь многочисленных типов антител, имеющих различные CDR. В качестве такого поликлонального антитела культивируют смесь клеток, которые продуцируют различные антитела, и может быть использовано антитело, очищенное из полученной в результате культуры (см. WO 2004/061104).
В качестве модифицированного антитела также может быть использовано антитело, связанное с любой молекулой различных типов, например, полиэтиленгликолем (PEG).
Далее, антитело по настоящему изобретению может быть в форме конъюгата, образованного между любыми из этих антител и другим медицинским средством (иммуноконъюгат). Примеры такого антитела включают антитело, в котором это антитело конъюгировано с радиоактивным веществом или соединением, обладающим фармакологическим действием (Nature Biotechnology (2005) 23, pp. 1137-1146).
Полученное антитело может быть очищено до гомогенности. Отделение и очистку антитела можно проводить, используя общепринятый способ разделения и очистки белков.
Например, антитело может быть отделено и очищено с помощью соответственно выбранного и комбинированного применения хроматографической колонки, фильтра, ультрафильтрации, солевой преципитации, диализа, препаративного электрофореза в полиакриламидном геле, электрофореза с изоэлектрофокусированием и подобного (Strategies for Protein Purification and Characterization: A Laboratory Course Manual, Daniel R. Marshak et al. eds., Cold Spring Harbor Laboratory Press (1996); Antibodies: A Laboratory Manual. Ed Harlow and David Lane, Cold Spring Harbor Laboratory (1988)), но этот способ этим не ограничивается.
Примеры хроматографии включают аффинную хроматографию, ионообменную хроматографию, гидрофобную хроматографию, гель-фильтрационную хроматографию, хроматографию с обращенной фазой и адсорбционную хроматографию.
Такую хроматографию можно проводить, используя жидкостную хроматографию, такую как HPLC или FPLC.
В качестве колонки, используемой в аффинной хроматографии, может быть использована колонка с Белком А и колонка с Белком G.
Например, в качестве колонки можно привести в пример колонку с использованием Белка А, Hyper D, POROS, Сефарозы F. F. (Pharmacia) и подобного.
Далее, используя носитель с иммобилизованным на нем антигеном, антитело также может быть очищено, используя свойство связывания антитела с антигеном.
5. Лекарственное средство, содержащее антитело против Siglec-15
Из антител против Siglec-15, полученных способом, описанным в приведенном выше пункте «4. Получение антитела против Siglec-15», может быть получено антитело, которое нейтрализует биологическую активность Siglec-15. Такое антитело, которое нейтрализует биологическую активность Siglec-15, ингибирует биологическую активность Siglec-15 in vivo, т.е. дифференцировку и/или созревание остеокластов, и, следовательно, может быть использовано в качестве терапевтического и/или профилактического средства для лечения аномального метаболизма костной ткани, вызванного аномальной дифференцировкой и/или созреванием остеокластов, в качестве лекарственного средства. Аномальный метаболизм костной ткани может представлять собой любое заболевание, характеризующееся общей потерей костной массы (остеопения или остеолиз). В основном лечение и/или профилактика с помощью антитела против Siglec-15 применяется в том случае, когда требуется ингибирование резорбции кости. Примеры аномального метаболизма костной ткани, который можно лечить и/или предотвратить с помощью антитела против Siglec-15, включают остеопороз (постменопаузный остеопороз, сенильный остеопороз, вторичный остеопороз вследствие применения терапевтического средства, такого как стероиды или иммунодепрессанты, или остеопороз, сопровождающий ревматоидный артрит), деструкцию кости, сопровождающую ревматоидный артрит, злокачественную гиперкальциемию, деструкцию кости, сопровождающую множественную миелому или злокачественные метастазы в кость, гигантоклеточную остеобластому, потерю зубов вследствие пародонтита, остеолиз вокруг протезированного сустава, деструкцию кости при хроническом остеомиелите, деформирующую остеодистрофию Педжета, почечную остеодистрофию и несовершенный остеогенез, однако аномальный метаболизм костной ткани ими не ограничивается, при условии, что это заболевание сопровождается общей потерей костной массы, вызванной остеокластами. Примеры антитела против Siglec-15, используемого в качестве упомянутого выше лекарственного средства, включают химерное антитело и гуманизированное антитело, полученное из антитела #32A1 или антитела #41B1 способом, описанным в пункте 4. (3) «Другие антитела». Далее, химерное антитело, гуманизированное антитело и антитело человека, имеющие общий эпитоп, что и антитело #32A1 или антитело #41B1, также могут быть использованы в качестве лекарственного средства. Имеет ли конкретное антитело против Siglec-15 общий эпитоп с антителом #32A1 или антителом #41B1 можно подтвердить путем наблюдения, связываются ли эти антитела или нет с одной и той же специфической частью пептида Siglec-15. Далее, также можно определить, что если определенное антитело против Siglec-15 конкурирует с антителом #32A1 или антителом #41B1 за связывание с Siglec-15, эти антитела имеют один и тот же эпитоп.
In vitro активность антитела против Siglec-15 по нейтрализации биологической активности Siglec-15 можно определить, например, по активности ингибирования дифференцировки клеток, которые гиперэкспрессируют Siglec-15, в остеокласты. Например, антитело против Siglec-15 добавляют к клеткам RAW 264.7 или клеткам Raw 264, которые представляют собой моноцитарную клеточную линию мыши, в различных концентрациях, и можно определить активность ингибирования дифференцировки в остеокласты, под действием стимуляции RANKL или TNF-α. Далее, антитело против Siglec-15 добавляют к первичным культивируемым клеткам костного мозга в различных концентрациях, и может быть определена активность ингибирования дифференцировки в остеокласты стимуляцией RANKL, TNF-α или активным витамином D3. Далее, антитело против Siglec-15 добавляют к нормальным клеткам-предшественникам остеокластов человека (нормальные природные клетки-предшественники остеокластов человека, доступные от Sanko Junyaku Co., Ltd., Cat. No. 2T-110) в различных концентрациях, и можно определить активность ингибирования дифференцировки в остеокласты под действием стимуляции RANKL или M-CSF. Такое ингибирующее действие на дифференцировку остеокластов можно определить с помощью ингибирования активности тартрат-резистентной кислой фосфатазы (TRAP) остеокластов в качестве показателя, как показано в Примерах 17, 19, 20 и 26. Далее, ингибирующее действие на дифференцировку остеокластов также можно определить, используя ингибирование образования TRAP-позитивных многоядерных остеокластов, т.е. ингибирование клеточного слияния остеокластов в качестве показателя, как показано в Примерах 19, 21, 22 и 35. Антитела по настоящему изобретению проявляли ингибирующее действие на клеточное слияние в концентрации 30 мкг/мл или менее в упомянутой выше тест-системе для дифференцировки остеокластов, и некоторые антитела проявляли ингибирующее действие даже в концентрации 3 мкг/мл или менее или 1 мкг/мл или менее. Далее, в том случае, когда было протестировано действие в еще более низкой концентрации, было обнаружено, что множество антител проявляло ингибирующее действие на дифференцировку остеокластов даже в диапазоне концентраций от 63 нг/мл до 1 мкг/мл. Далее, в эксперименте исследования образования очагов дефекта ткани (Takada et al., Bone and Mineral, (1992) 17, 347-359) с использованием клеток бедренной кости и/или клеток большой берцовой кости in vitro активность ингибирования резорбции кости под действием остеокластов может быть определена добавлением антитела против Siglec-15 к клеткам бедренной кости и/или клеткам большой берцовой кости в различных концентрациях и наблюдением образования ямок на срезе дентина. В качестве системы для определения in vitro активности ингибирования резорбции кости остеокластами, как показано в Примере 37, также возможно использовать планшет, покрытый коллагеном человека, конъюгированным с европием. В указанной выше тест-системе для резорбции кости остеокластами антитело по изобретению ингибировало резорбцию кости в концентрации 3 мкг/мл или менее, то есть в диапазоне концентраций от 0,3 мкг/мл до 3 мкг/мл. Терапевтическое или профилактическое действие in vivo антитела против Siglec-15 на аномальный метаболизм костной ткани с использованием экспериментального животного может быть подтверждено путем введения антитела против Siglec-15 животному, которое является животным-моделью остеопороза, или трансгенному животному с гиперэкспрессией Siglec-15, и определения изменения в остеокластах.
Полученное таким образом антитело, которое нейтрализует биологическую активность Siglec-15, используют в качестве лекарственного средства, в частности в качестве фармацевтической композиции для лечения или профилактики аномального метаболизма костной ткани, такого как остеопороз, деструкция кости, сопровождающая ревматоидный артрит, или деструкция кости, сопровождающая злокачественное метастазирование в костную ткань, или в качестве антитела для иммунологической диагностики такого заболевания.
При лечении ревматоидного артрита (RA) основной проблемой является утрата костной ткани, сопровождающая появление этого заболевания. Сообщалось о том, что при этой утрате костной ткани, сопровождающей RA, остеокласты играют основную роль. Считается, что цитокины являются наиболее важными для индукции остеокластов (дифференцировки и созревания), а активируют и вызывают деструкцию кости при RA RANKL и TNF-α (Romas E. et al., Bone 30, pp. 340-346, 2002). Как показано в примере 19 настоящего изобретения, OCIF/OPG, который является рецептором для RANKL, может ингибировать образование остеокластов, индуцированное действием RANKL, но не ингибирует образование остеокластов, индуцированное под действием TNF-α. С другой стороны, антитело против Siglec-15 в соответствии с настоящим изобретением эффективно ингибировало образование остеокластов, индуцированное как действием RANKL, так и TNF-α. Следовательно, ожидается, что антитело против Siglec-15 по настоящему изобретению может ингибировать утрату костной ткани и деструкцию кости, индуцированную действием TNF-α при RA или подобном, более эффективно, чем блокатор RANKL (OCIF/OPG, антитело против RANKL или подобное).
В качестве одного примера, для лечения или профилактики аномального метаболизма костной ткани антитело против Siglec-15 можно вводить отдельно или вместе по меньшей мере с одним терапевтическим средством для лечения заболевания, связанного с костной тканью. В качестве другого примера, антитело против Siglec-15 можно вводить вместе с терапевтически эффективным количеством терапевтического средства для лечения аномального метаболизма костной ткани. Примеры терапевтического средства, которое можно вводить вместе с антителом против Siglec-15, включают, но не только, бисфосфонаты, активный витамин D3, кальцитонин и его производные, гормональные препараты, такие как эстрадиол, SERM (селективные модуляторы рецепторов эстрогена), иприфлавон, витамин K2 (менатетренон), препараты кальция, препараты РТН (паратиреоидного гормона), нестероидные противовоспалительные средства, препараты растворимого рецептора TNF, анти-TNF-α антитела или функциональные фрагменты таких антител, анти-PTHrP (белка, родственного паратиреоидному гормону) антитела или функциональные фрагменты таких антител, антагонисты рецептора IL-1, антитела против рецептора IL-6 или функциональные фрагменты таких антител, антитела против RANKL или функциональные фрагменты таких антител и OCIF (фактор, ингибирующий остеокластогенез). В зависимости от состояния аномального метаболизма костной ткани или намеченной степени лечения и/или профилактики, можно вводить два или три, или более типов лекарственных средств, и эти лекарственные средства могут быть доставлены все вместе путем инкапсулирования их в одном препарате. Эти лекарственные средства и антитело против Siglec-15 могут быть доставлены все вместе путем инкапсулирования их в одном препарате. Далее, эти лекарственные средства могут быть доставлены все вместе путем инкапсулирования их в виде набора, используемого для лечения и/или профилактики. Далее, эти лекарственные средства и антитело против Siglec-15 могут быть доставлены по отдельности. В случае введения при генной терапии ген белкового терапевтического средства для лечения заболевания костей и ген антитела против Siglec-15 может быть вставлен ниже той же промоторной области или различных промоторных областей и может быть введен в тот же вектор или различные векторы.
Путем конъюгирования терапевтического средства для лечения заболевания костей с антителом против Siglec-15 или его фрагментом может быть получен лекарственный конъюгат направленного действия, как описано в M. C. Garnet “Targeted drug conjugates: principles and progress”, Advanced Drug Delivery Reviews, (2001) 53, 171-216. Для достижения этой цели, помимо молекулы антитела, можно применять любой фрагмент антитела, при условии, что он не полностью утрачивает способность распознавать остеокласты, и примеры такого фрагмента включают такие фрагменты, как Fab, F(ab')2 и Fv. В настоящем изобретении антитело и фрагмент можно использовать аналогичным образом. Конъюгат, образованный антителом против Siglec-15 или его фрагментом и терапевтическим средством для лечения заболевания костей, может представлять собой любую из различных форм, описанных у M. C. Garnet “Targeted drug conjugates: principles and progress”, Advanced Drug Delivery Reviews, (2001) 53, 171-216, G. T. Hermanson “Bioconjugate Techniques” Academic Press, California (1996), Putnam and J. Kopecek “Polymer Conjugates with Anticancer Activity” Advances in Polymer Science (1995) 122, 55-123 и подобных. Таким образом, в качестве примера можно привести форму конъюгата, в которой антитело против Siglec-15 и терапевтическое средство для лечения заболевания костей конъюгированы друг с другом химически и напрямую или через спейсер, такой как олигопептид, и форму конъюгата через соответствующий носитель лекарственного средства. Примеры носителя лекарственного средства включают липосомы и водорастворимые полимеры. Более конкретные примеры формы конъюгата посредством такого носителя лекарственного средства включают форму конъюгата, в которой антитело и терапевтическое средство для лечения заболевания костей включены в липосому, и эта липосома и антитело конъюгированы друг с другом, и форму конъюгата, в которой терапевтическое средство для лечения заболевания костей конъюгировано с водорастворимым полимером (соединением, имеющим молекулярную массу примерно от 1000 до 100000) химически и напрямую или через спейсер, такой как олигопептид, и антитело конъюгировано с водорастворимым полимером. Конъюгирование антитела (или его фрагмента) с терапевтическим средством для лечения заболевания костей или носителем лекарственного средства, таким как липосомы или водорастворимый полимер, можно осуществить способом, известным специалистам в данной области, таким как способ, описанный у G. T. Hermanson в “Bioconjugate Techniques” Academic Press, California (1996), Putnam and J. Kopecek “Polymer Conjugates with Anticancer Activity” Advances in Polymer Science (1995) 122, 55-123. Включение терапевтического средства для лечения заболевания костей в липосому можно осуществить способом, известным специалистам в данной области, таким как способ, описанный у D. D. Lasic в “Liposomes: From Physics to Applications” Elsevier Science Publishers B. V., Amsterdam (1993) или подобным. Конъюгирование терапевтического средства для лечения заболевания костей с водорастворимым полимером можно осуществить способом, известным специалистам в данной области, таким, как способ, описанный у D. Putnam и J. Kopecek в “Polymer Conjugates with Anticancer Activity” Advances in Polymer Science (1995) 122, 55-123. Конъюгат между антителом (или его фрагментом) и белковым терапевтическим средством для лечения заболевания костей (или его фрагментом) может быть получен способом, известным специалистам в данной области с помощью методов генной инженерии, отличных от упомянутого выше способа.
Настоящее изобретение также относится к фармацевтической композиции, содержащей терапевтически и/или профилактически эффективное количество антитела против Siglec-15 и фармацевтически приемлемый разбавитель, носитель, солюбилизирующий агент, эмульгатор, консервант и/или адъювант.
Настоящее изобретение также относится к фармацевтической композиции, содержащей терапевтически и/или профилактически эффективное количество антитела против Siglec-15, терапевтически и/или профилактически эффективное количество по меньшей мере одного терапевтического средства для лечения заболевания костей и фармацевтически приемлемый разбавитель, носитель, солюбилизирующий агент, эмульгатор, консервант и/или адъювант. Примеры терапевтического средства для лечения заболевания костей включают, но не только, бисфосфонаты, активный витамин D3, кальцитонин и его производные, гормональные препараты, такие как эстрадиол, SERM (селективные модуляторы рецепторов эстрогена), иприфлавон, витамин K2 (менатетренон), препараты кальция, препараты РТН (паратиреоидного гормона), нестероидные противовоспалительные средства, препараты растворимого рецептора TNF, анти-TNF-α антитела или функциональные фрагменты таких антител, анти-PTHrP (белка, родственного паратиреоидному гормону) антитела или функциональные фрагменты таких антител, антагонисты рецептора IL-1, антитела против рецептора IL-6 или функциональные фрагменты таких антител, антитела против RANKL или функциональные фрагменты таких антител и OCIF (фактор, ингибирующий остеокластогенез).
Вещество, используемое в препарате, допустимом в фармацевтической композиции в соответствии с настоящим изобретением, предпочтительно является нетоксичным для пациента, которому вводят эту фармацевтическую композицию, исходя из дозы и концентрации.
Фармацевтическая композиция по настоящему изобретению может содержать вещество для фармацевтического применения, которое способно изменять или поддерживать pH, осмотическое давление, вязкость, прозрачность, цвет, изотоничность, асептическое состояние, стабильность, растворимость, скорость высвобождения, скорость адсорбции и проницаемость. Примеры веществ для фармацевтического применения включают, но не только, аминокислоты, такие как глицин, аланин, глутамин, аспарагин, аргинин и лизин; противомикробные средства; антиоксиданты, такие как аскорбиновая кислота, сульфат натрия и гидрогенсульфит натрия; буферы, такие как фосфатный, цитратный, боратные буферы, растворы бикарбоната и Tris-HCl; наполнители, такие как маннит и глицин; хелатирующие агенты, такие как этилендиаминтетраацетат (EDTA); комплексообразующие агенты, такие как кофеин, поливинилпирролидин, β-циклодекстрин и гидроксипропил-β-циклодекстрин; вещества, добавляемые для увеличения объема, такие как глюкоза, манноза и дестрин; другие углеводы, такие как моносахариды и дисахариды; красители; ароматизаторы; разбавители; эмульгаторы; гидрофильные полимеры, такие как поливинилпирролидин; консерванты, такие как полипептиды с низкой молекулярной массой, основания, образующие противоионы, хлорид бензалкония, бензоат, салициловая кислота, тимеросал, фенетиловый спирт, метилпарабен, пропилпарабен, хлоргексидин, сорбиновая кислота, и перекись водорода; растворители, такие как глицерин, пропиленгликоль и полиэтиленгликоль; сахарные спирты, такие как манитол и сорбитол; суспендирующие агенты; поверхностно-активные вещества, такие как сложный эфир сорбитана, полисорбаты, в том числе полисорбат 20 и полисорбат 80, Тритон, трометамин, лецитин и холестерин; вещества, повышающие стабильность, такие как сахароза и сорбитол; вещества, повышающие эластичность, такие как хлорид натрия, хлорид калия и маннитол и сорбитол; среды для переноса веществ; разбавители; эксципиенты; и/или фармацевтические адъюванты. Дополнительное количество этих веществ для фармацевтического использования предпочтительно составляет от 0,01 до 100 раз, в частности предпочтительно от 0,1 до 10 раз от массы антитела против Siglec-15. Специалисты в данной области могут соответствующим образом определить предпочтительный состав фармацевтической композиции в препарате в зависимости от заболевания, при котором ее назначают, применяемого пути введения и подобного.
Эксципиент или носитель в фармацевтической композиции может быть в форме жидкости или твердого вещества. Подходящим эксципиентом или носителем может быть вода для инъекций, физиологический раствор, искусственная спинно-мозговая жидкость или другое вещество, обычно используемое для парентерального введения. Далее, нейтральный физиологический раствор или физиологический раствор, содержащий сывороточный альбумин, также может быть использован в качестве носителя. Фармацевтическая композиция может содержать Tris буфер с pH 7,0-8,5 или ацетатный буфер с pH 4,0-5,5, который может быть дополнен сорбитом или другим соединением. Примеры фармацевтической композиции по изобретению включают фармацевтическую композицию, содержащую антитело против Siglec-15, и фармацевтическую композицию, содержащую антитело против Siglec-15 и по меньшей мере одно терапевтическое средство для лечения заболевания костей. Фармацевтическую композицию по изобретению получают в форме лиофилизированного продукта или жидкости в виде лекарственного средства, имеющего выбранную композицию и требуемую чистоту. Фармацевтическая композиция, содержащая антитело против Siglec-15, и фармацевтическая композиция, содержащая антитело против Siglec-15 и по меньшей мере одно терапевтическое средство для лечения аномального метаболизма костной ткани, также могут быть получены в виде лиофилизированного продукта, используя подходящий эксципиент, такой как сахароза.
Фармацевтическая композиция по настоящему изобретению может быть получена для парентерального введения или для абсорбции в желудочно-кишечном тракте посредством перорального введения. Состав и концентрацию препарата можно определить в зависимости от способа введения. Поскольку аффинность антитела против Siglec-15, содержащегося в фармацевтической композиции по изобретению в отношении Siglec-15, выше, то есть, поскольку константа диссоциации (значение Kd) для Siglec-15 ниже, антитело против Siglec-15 может демонстрировать свою лекарственную эффективность в более низкой дозе для людей, и, следовательно, доза фармацевтической композиции по настоящему изобретению для людей также можно определить, исходя из этого результата. Что касается дозы, в случае, когда антитело человека против Siglec-15 вводят людям, это антитело можно вводить в дозе примерно от 0,1 до 100 мг/кг однократно от одного до 180 дней.
Примеры лекарственной формы фармацевтической композиции по настоящему изобретению включают инъекции, в том числе инфузии, суппозитории, трансназальные средства, сублингвальные средства и чрескожные абсорбенты.
6. Поиск непосредственно взаимодействующего вещества
Другой вариант осуществления настоящего изобретения включает способ лекарственного моделирования, исходя из конформации Siglec-15, для получения вещества, которое ингибирует активность Siglec-15. Такой способ известен как способ рационального моделирования лекарственных средств и применяется для поиска соединения, которое эффективно ингибирует или активирует ферментативную активность или связывание с лигандом, кофактором или ДНК. В качестве примера такого соединения известен ингибитор протеазы, который служит в качестве средства против ВИЧ, который уже размещен на рынке. Также в трехмерном структурном анализе Siglec-15 по настоящему изобретению в основном можно использовать хорошо известный способ, такой как рентгеновский кристаллографический анализ или метод ядерного магнитного резонанса. Далее, при поиске вещества, которое ингибирует функцию Siglec-15, также можно осуществить лекарственное моделирование, используя компьютерное моделирование лекарственных средств (CADD). В качестве примера в этом случае известно соединение низкой молекулярной массы (WO 99/58515), которое ингибирует действие AP-1 и считается новым геномным лекарственным средством для лечения ревматоидного артрита и подобного. С помощью такого способа возможно получить вещество, которое ингибирует действие Siglec-15 путем непосредственного связывания с Siglec-15 или путем ингибирования взаимодействия между Siglec-15 и другими факторами.
Далее, другой вариант осуществления относится к полипептиду, с которым соединяется Siglec-15 по настоящему изобретению, другими словами, белок-партнер для Siglec-15. То есть, настоящее изобретение относится к способу скрининга белка-партнера, который регулирует активность Siglec-15.
Один вариант осуществления этого способа скрининга включает стадию приведения образца тестируемого белка в контакт с Siglec-15 и выбора белка, который связывается с Siglec-15. В качестве такого способа можно привести пример, например, способа, в котором используют очищенный Siglec-15 и проводят аффинную очистку белка, связывающегося с ним. Один конкретный пример этого способа будет описан ниже. Последовательность, состоящая из шести гистидинов в качестве аффинной метки, объединяют с Siglec-15 с получением слитого белка и полученный в результате белок инкубируют при 4°C в течение 12 часов вместе с раствором клеточного экстракта (фракция, пропущенная через никель-агарозную колонку после загрузки колонки клеточным раствором). Затем другой никель-агарозный носитель добавляют в смесь и эту смесь инкубируют при 4°C в течение 1 часа. После достаточного промывания никель-агарозного носителя промывочным буфером в эту смесь добавляют 100 мМ имидазола для элюции и очистки белка, который специфически связывается с Siglec-15 в растворе клеточного экстракта. Затем очищенный белок анализируют для определения его структуры. Таким способом может быть очищен белок, который непосредственно связывается с Siglec-15, и белок, который не обладает активностью связывания с Siglec-15, но опосредованно связывается с Siglec-15, образуя комплекс с белком в виде субъединицы, которая непосредственно связывается Siglec-15. В качестве альтернативного способа также возможно проводить клонирование посредством анализа far-Western блоттинг или анализа двухгибридной системы, используя клетки дрожжей или млекопитающих, однако это не ограничивается этими способами.
Если кДНК белка-партнера, который напрямую или опосредованно взаимодействует с Siglec-15 получают таким способом, кДНК может быть использована в функциональном скрининге вещества, которое ингибирует взаимодействие между Siglec-15 и белком-партнером. В частности, получают слитый белок между Siglec-15 и глутатион-S-трансферазой и связывают с микропланшетом, покрытым антителом против глутатион-S-трансферазы. Затем биотинилированный белок-партнер приводят в контакт со слитым белком и выявляют связывание белка-партнера со слитым белком, используя конъюгированную со стрептавидином щелочную фосфатазу. Когда добавляют биотинилированный белок-партнер, также добавляют тестируемое вещество для отбора вещества, которое активирует или ингибирует связывание слитого белка с белком-партнером. С помощью этого способа может быть получено вещество, которое напрямую действует на слитый белок, или вещество, которое напрямую действует на белок-партнер.
В том случае, когда слитый белок связывается опосредованно с белком-партнером через другой фактор, это исследование проводят, например, в присутствии раствора клеточного экстракта, содержащего этот фактор. В этом случае, существует возможность, что также можно выбрать вещество, которое может действовать на этот фактор.
Далее, в том случае, когда полученный белок-партнер обладает активностью, стимулирующей функции Siglec-15, возможно проводить скрининг кандидатного вещества, применимого в качестве терапевтического и/или профилактического средства для лечения аномального метаболизма костной ткани, например терапевтического и/или профилактического средства для лечения остеопороза в соответствии со способом тестирования, в котором используется вектор экспрессии гена Siglec-15, как описано выше. Далее, в том случае, когда полученный белок-партнер обладает активностью ингибирования функции Siglec-15, возможно использовать полинуклеотид, кодирующий такой ингибитор в генной терапии аномального метаболизма костной ткани.
Такой полинуклеотид может быть получен, например, путем анализа аминокислотной последовательности идентифицированного ингибитора, синтеза олигонуклеотидного зонда, кодирующего аминокислотную последовательность, и проведения скрининга библиотеки кДНК или геномной библиотеки. Далее, в том случае, когда полипептид, обладающий активностью ингибирования функции Siglec-15, получают из синтезированной случайным образом библиотеки искусственных пептидов, химически синтезируют ДНК, состоящую из нуклеотидной последовательности, кодирующей аминокислотную последовательность этого пептида.
В генной терапии полученный таким образом ген, кодирующий ингибитор, встраивают, например, в вирусный вектор и пациента инфицируют (аттенуированным) вирусом, содержащим полученный в результате рекомбинантный вирусный вектор. В организме пациента продуцируется фактор против деструкции кости и функционирует для ингибирования дифференцировки остеокластов и, следовательно, может быть реализовано лечение и/или профилактика аномального метаболизма костной ткани.
В качестве способа введения генного терапевтического средства в клетку может быть использован либо метод генного переноса с использованием вирусного вектора, либо метод невирусного генного переноса.
Примеры метода генного переноса с использованием вирусного вектора включают метод интеграции ДНК, кодирующей ингибитор Siglec-15, или его мутантную форму, в ДНК-вирус или РНК-вирус, такой как ретровирус, аденовирус, адено-ассоциированный вирус, вирус герпеса, вирус коровьей оспы, поксвирус, вирус полиомиелита или вирус Синдбис для осуществления генного переноса. Среди них, способ с использованием ретровируса, аденовируса, адено-ассоциированного вируса, или вируса коровьей оспы, является особенно предпочтительным. Примеры способа невирусного переноса генов включают способ введения экспрессионной плазмиды непосредственного в мышцу (способ ДНК вакцинации), липосомный метод, способ липофекции, способ микроинъекции, метод с использованием фосфата кальция и метод электропорации. В частности, метод ДНК вакцинации и липосомный метод являются предпочтительными.
Далее, для фактического применения генного терапевтического средства в качестве лекарственного средства существует in vivo метод, в котором ДНК непосредственно вводят в организм, и ex vivo метод, в котором определенный тип клетки получают от человека и ДНК вводят в эту клетку ex vivo, и эту клетку возвращают в организм.
Например, в том случае, когда генное терапевтическое средство вводят с помощью in vivo метода, его вводят посредством соответствующего пути введения, например, через вену или артерию, под кожу, в кожу или в мышцу, в соответствии с заболеванием, симптомами или подобным. Далее, в том случае, когда средство вводят с помощью in vivo метода, генное терапевтическое средство в основном получают в виде инъекции, однако при необходимости может быть добавлен обычно используемый носитель. Далее, в том случае, когда средство получают в виде липосомы или мембранной слитой липосомы (например, липосомы вируса Sendai), оно может быть получено в виде липосомного препарата, такого как суспензия, лиофилизированное средство или центрифужно концентрированное и лиофилизированное средство.
Нуклеотидная последовательность, комплементарная полноразмерной или неполной последовательности нуклеотидной последовательности, представленной SEQ ID NO:1 или 3 в перечне последовательностей, может быть использована для так называемой антисмысловой терапии. Антисмысловая молекула может быть использована в качестве ДНК, которая в основном состоит из 15-30 нуклеотидов и является комплементарной частью нуклеотидной последовательности, выбранной из нуклеотидных последовательностей, представленных последовательностями SEQ ID NO: 1 и 3 в перечне последовательностей, или их стабильным ДНК производным, таким как их фосфоротиоатное, метилфосфонатное, или морфолино производное, или стабильное производное РНК, такое как 2'-O-алкил РНК. Такая антисмысловая молекула может быть введена в клетку способом, известным в области техники, к которой относится настоящее изобретение, например инъекцией крайне малого количества антисмысловой молекулы, формированием молекулы в липосомную капсулу или экспрессией молекулы с использованием вектора, имеющего антисмысловую последовательность. Такая антисмысловая терапия применяется для лечения заболевания, вызванного чрезмерным повышением активности белка, кодируемого нуклеотидной последовательностью, представленной последовательностью ID NO:1 или 3 в перечне последовательностей.
Далее, способ с использованием двухспиральной короткой РНК (siРНК) также может быть приведен в качестве примера (Genes and Developments, January 15, 2001, vol. 15, No. 2, pp. 188-200). Например, получают siРНК против гена Siglec-15 и вводят в клетку в соответствии со способом, описанным в этом документе, посредством чего может быть получено терапевтическое средство для лечения метаболического заболевания костной ткани, сопровождаемого гиперэкспрессией Siglec-15.
Примеры
В дальнейшем, настоящее изобретение будет более конкретно описано со ссылкой на Примеры, однако настоящее изобретение ими не ограничивается. Следует отметить, что соответствующие операции, относящиеся к манипуляции с генами в следующих Примерах, проводили в соответствии со способами, описанными в публикации «Molecular Cloning» (написанной Sambrook, J., Fritsch, E. F. и Maniatis, T., опубликовано Cold Spring Harbor Laboratory Press in 1989), или в случае применения коммерчески доступных реактивов или наборов, выполнены в соответствии с протоколами, прилагаемыми к ним.
Пример 1. Экспрессия гена Siglec-15 человека в ткани гигантоклеточной остеобластомы
Гигантоклеточная остеобластома (GCT) гистологически представляет собой опухоль костной ткани с появлением огромного количества подобных остеокластам многоядерных гигантских клеток и характеризуется клиническими проявлениями остеолитической деструкции кости (Bullough et al., Atlas of Orthopedic Pathology 2nd edition, 17.6-17.8, Lippincott Williams & Wilkins Publishers (1992)). Анализ профиля экспрессии проводили в отношении зонда EST (Affymetrix GeneChip HG-U133 зонд 215856_at: производство Affymetrix, Inc.), имеющего нуклеотидную последовательность, частично перекрывающуюся последовательностью гена Siglec-15 человека в тканях GCT, с использованием базы данных (Genesis 2006 Release 3.0), полученной Gene Logic, Inc. Далее, анализ профиля экспрессии также проводили с использованием зондов EST для RANK (Affymetrix GeneChip HG-U133 зонд 207037_at, производство Affymetrix, Inc.) и RANKL (Affymetrix GeneChip HG-U133 зонд 210643_at, производство Affymetrix, Inc.), которые играют ключевую роль в дифференцировке в остеокласты, и для катепсина К (Affymetrix GeneChip HG-U133 зонд 202450_s_at, производство Affymetrix, Inc.) и TRAP (Affymetrix GeneChip HG-U133 зонд 204638_at, производство Affymetrix, Inc.), которые являются маркерами дифференцировки в остеокласты в тканях GCT, аналогичным образом.
Когда уровни экспрессии сравнивали между 13 случаями нормальных костных тканей, 12 случаями тканей GCT и 16 случаями опухолей костной ткани, отличных от GCT, было обнаружено, что транскрипция RANK и RANKL специфически повышалась в тканях GCT по сравнению с нормальными тканями (Фиг. 1-A). С другой стороны, в опухолях костной ткани, отличных от GCT, в которых считается, что повышение резорбции кости вызывается не всегда, транскрипция RANK и RANKL была ниже, чем в GCT, и, следовательно, предположено, что GCT обеспечивает окружение, в котором усилено образование остеокластов и активация. Далее, когда сравнивают уровни экспрессии генов катепсина K и TRAP, гены транскрибировались на высоком уровне в GCT (Фиг. 1-B), и было сделано предположение о том, что появляется огромное количество остеокластов, обладающих активностью в отношении резорбции кости. Аналогичным образом, когда сравнивали уровни транскрипции гена Siglec-15, было обнаружено, что этот ген транскрибировался на высоком уровне, особенно в GCT, аналогичным образом, что и соответствующие гены RANK, RANKL, катепсина K и TRAP (Фиг. 2). Из этих результатов можно предположить, что Siglec-15 связан с патологическим состоянием человека, при котором повышается резорбция кости, как GCT.
Пример 2. Экстракция суммарной РНК из зрелых остеокластов мыши
a) Моноцитарные клетки RAW 264.7 мыши (ATCC Cat. No. TIB-71) получали в концентрации 4,5×104 клеток/мл в среде α-MEM, содержащей 10% эмбриональную сыворотку теленка. Полученный в результате клеточный препарат помещали в 75 см2 флакон по 10 мл/флакон, и туда добавляли RANKL человека (производство PeproTech Inc.) с получением конечной концентрации 40 нг/мл, и эти клетки культивировали в течение 3 дней в CO2 термостате. Далее, культивирование без добавления RANKL человека проводили аналогичным образом.
После завершения культивирования суммарную РНК экстрагировали из RAW 264.7, культивированных в соответствующих условиях, используя реагент для экстракции суммарной РНК (ISOGEN, производство Nippon Gene Co., Ltd.) в соответствии с протоколом, прилагаемым к этому реагенту. Полученную суммарную РНК хранили при -80°C.
b) При культивировании первичных культивируемых клеток костного мозга мыши в присутствии активного витамина D3 появляется большое количество TRAP-позитивных многоядерных остеокластов (Takahashi et al., Endocrinology, (1988) 122, 1373-1382).
Самцов мышей ddY в возрасте 8 недель подвергали эвтаназии путем смещения шейных позвонков под эфирным наркозом и извлекали бедренную и большую берцовую кости. После удаления мягких тканей отсекали оба конца бедренной или большой берцовой кости. Затем среду α-MEM, содержащую 10% эмбриональную сыворотку теленка, вводили в костный мозг, используя цилиндр шприца с инъекционной иглой 25 калибра, и получали клетки костного мозга. После подсчета количества клеток клетки подготавливали с концентрацией 5×106 клеток/мл в среде α-MEM, содержащей 10% эмбриональную сыворотку теленка. Полученный в результате клеточный препарат помещали в 60 лунок 96-луночного планшета по 100 мкл/лунку и туда добавляли активный витамин D3 (производство Sigma Co., Ltd.) с получением конечной концентрации 2×10-8 M, и клетки культивировали в течение 8 дней в CO2 термостате. Далее, культивирование без добавления активного витамина D3 проводили аналогичным образом. К тому же, замену среды и добавление активного витамина D3 проводили на 3 и 6 день.
После этого суммарную РНК экстрагировали из клеток, культивированных в соответствующих условиях, используя реагент для экстракции суммарной РНК (ISOGEN, производство Nippon Gene Co., Ltd.) в соответствии с протоколом, прилагаемым к этому реагенту. Полученную суммарную РНК хранили при -80°C до использования.
Пример 3. Получение последовательности открытой рамки считывания (ORF) мышиного Siglec-15
a) Синтез первой цепи кДНК
К 1 мкг суммарной РНК, полученной в пункте a) Примера 2, добавляли 1 мкл 1 Ед/мкл ДНазы I и 1 мкл 10×ДНазного буфера I (производство Invitrogen, Inc.), а затем конечный объем доводили до 10 мкл с использованием H2O. После того как реакцию оставляли протекать при комнатной температуре в течение 15 минут, туда добавляли 1 мкл 25 мМ EDTA и полученную в результате смесь нагревали при 65°C в течение 10 минут. Из этого раствора брали аликвоту 8 мкл, и добавляли туда 1 мкл 50 мкМ праймера олиго(dT)20 и 1 мкл 10 мМ dNTP, и полученную в результате смесь нагревали при 65°C в течение 5 минут, а затем инкубировали на льду. К этому раствору добавляли 2 мкл 10×RT буфера (производство Invitrogen, Inc.), 4 мкл 25 мМ MgCl2, 2 мкл 0,1 M дитиотреитола, 1 мкл ингибитора РНазы (RNaseOUT, 40 Ед/мкл, производство Invitrogen, Inc.) и 1 мкл обратной транскриптазы Superscript III (200 Ед/мкл, производство Invitrogen, Inc.) и суммарный объем доводили до 20 мкл. После протекания реакции при 50°C в течение 50 минут смесь нагревали при 85°C в течение 5 минут, а затем инкубировали на льду в течение 1 минуты. После это смесь хранили при -20°C.
b) ПЦР реакция
Олигонуклеотиды, имеющие последовательности: 5'-agaattccac cATGGAGGGG TCCCTCCAAC TC-3' (mSiglec-15-EcoRI kozak-F: SEQ ID NO:5 в перечне последовательностей); и 5'-cgccgctcga gTTATTTCTC ATGGTGAATG AC-3' (mSiglec-15-XhoI-R: SEQ ID NO:6 в перечне последовательностей) в качестве праймеров для амплификации ORF кДНК мышиного Siglec-15 с помощью ПЦР синтезировали в соответствии с общепринятой процедурой. ПЦР проводили, используя это сочетание праймеров и кДНК, полученную в пункте a), и полимеразу высокой точности (производство Invitrogen, Inc.) в соответствии с общепринятой процедурой. Условия для термоциклера устанавливали следующие: после нагревания при 94°C в течение 2 минут температуру цикла «94°C в течение 0,5 минуты, 55°C в течение 0,5 минут и 68°C в течение 1,5 минут» повторяли 35 раз с последующим нагреванием при 68°C в течение 5 минут и инкубированием при 4°C.
c) Клонирование в вектор pcDNA3.1(+)
Реакционный раствор ПЦР, полученный в пункте b), и вектор pcDNA3.1(+) (производство Invitrogen, Inc.) обрабатывали ферментами рестрикции (EcoRI, XhoI) с последующей очисткой на колонке, а затем проводили лигазную реакцию в соответствии с общепринятой процедурой. Escherichia coli DH5α-T1 трансформировали полученным в результате вектором и помещали на чашку, содержащую ампициллин. Из полученных таким образом клонов Escherichia coli выделяли трансформированные Escherichia coli содержащие плазмиду Siglec-15 мыши/pcDNA3.1(+).
Полную нуклеотидную последовательность ORF кДНК, встроенной в полученную плазмиду, анализировали, используя ДНК секвенсер, и в результате было обнаружено, что она представляет собой последовательность, представленную SEQ ID NO:3 в перечне последовательностей. Эта нуклеотидная последовательность была такой же, как кодирующая область ORF предсказанной последовательности, зарегистрированной в базе данных NCBI GenBank как «CD33L3 мыши» (регистрационный номер: XM_884636), и далее, аминокислотная последовательность (SEQ ID NO:4 в перечне последовательностей), кодируемая нуклеотидной последовательностью, была на 100% идентична предсказанной аминокислотной последовательности CD33L3 мыши.
Пример 4. Экспрессия мРНК Siglec-15, сопровождающая дифференцировку остеокластов мыши (ПЦР анализ в режиме реального времени)
a) К 1 мкг суммарной РНК, полученной в пункте a) или b) Примера 2, добавляли 1 мкл 1 Ед/мкл ДНазы I и 1 мкл 10×ДНазного буфера I (производство Invitrogen, Inc.), а затем конечный объем доводили до 10 мкл с использованием H2O. После протекания реакции при комнатной температуре в течение 15 минут туда добавляли 1 мкл 25 мМ EDTA и полученную в результате смесь нагревали при 65°C в течение 10 минут. Из этого раствора брали аликвоту 8 мкл и в нее добавляли 1 мкл 50 мкМ праймера олиго(dT)20 и 1 мкл 10 мМ dNTP, и полученную в результате смесь нагревали при 65°C в течение 5 минут, а затем инкубировали на льду. К этому раствору добавляли 2 мкл 10×RT буфера (производство Invitrogen, Inc.), 4 мкл 25 мМ MgCl2, 2 мкл 0,1 M дитиотреитола, 1 мкл ингибитора РНазы (RNaseOUT, 40 Ед/мкл, производство Invitrogen, Inc.) и 1 мкл обратной транскриптазы Superscript III (200 Ед/мкл, производство Invitrogen, Inc.) и общий объем доводили до 20 мкл. После протекания реакции при 50°C в течение 50 минут смесь нагревали при 85°C в течение 5 минут, а затем инкубировали на льду.
Используя полученную таким образом одноцепочечную кДНК, ПЦР в режиме реального времени проводили с использованием сочетания следующих праймеров и флуоресцентно меченных зондов (TaqMan probe, производство Applied Biosystems, Inc.).
Условия для ПЦР в режиме реального времени:
Праймеры для амплификации Siglec-15 мыши:

ПЦР анализ в режиме реального времени проводили, используя систему ПЦР в режиме реального времени (Детектор последовательности ABI Prism 7700, производство Perkin Elmer Japan Applied Biosystems Division) в следующих условиях. В этой реакции использовали общую реакционную смесь для проведения ПЦР TaqMan Universal PCR Master Mix (производство Applied Biosystems, Inc.). Сначала дистиллированную воду добавляли к 25 пмоль каждого праймера, 8 нг одноцепочечной кДНК и 10 пмоль TaqMan зонда для доведения конечного объема до 25 мкл, а затем туда добавляли 25 мкл общей реакционной смеси TaqMan Universal PCR Master Mix, тем самым получая 50 мкл реакционного раствора. Этот реакционный раствор нагревали при 50°C в течение 2 минут, а затем нагревали при 95°C в течение 10 минут, и в дальнейшем подвергали 40 температурным циклам «95°C в течение 0,25 минут и 60°C в течение 1 минуты», тем самым проводили ПЦР анализ в режиме реального времени. К тому же, уровень экспрессии мРНК для Siglec-15 мыши был скорректирован по уровню экспрессии мРНК для рибосомного белка L32.
В результате уровень экспрессии гена Siglec-15 существенно повышался в обоих случаях, когда остеокласты индуцировали добавлением RANKL к RAW 264.7 и когда остеокласты индуцировали добавлением активного витамина D3 к первичным культивируемым клеткам костного мозга мышей (Фиг. 3).
b) RAW 264.7 получали при 4,5×104 клеток/мл в среде α-MEM, содержащей 10% эмбриональную сыворотку теленка, и полученный в результате клеточный препарат помещали в 75 см2 флакон по 10 мл/флакон, а затем в него добавляли RANKL человека (производство PeproTech Inc.) с получением конечной концентрации 40 нг/мл. Клетки культивировали в течение 0, 1, 2 и 3 дней в CO2 термостате. Далее, культивирование без добавления RANKL человека проводили аналогичным образом.
После завершения культивирования суммарную РНК экстрагировали из RAW 264.7, культивированных в соответствующих условиях, используя реагент для экстракции суммарной РНК (ISOGEN, производство Nippon Gene Co., Ltd.) в соответствии с протоколом, прилагаемым к этому реагенту. Полученную суммарную РНК хранили при -80°C.
К 1 мкг полученной таким образом суммарной РНК добавляли 1 мкг 1 Ед/мкл ДНазы I и 1 мкл 10×ДНаза I буфера (производство Invitrogen, Inc.), а затем конечный объем доводили до 10 мкл с использованием H2O. После протекания реакции при комнатной температуре в течение 15 минут добавляли 1 мкл 25 мМ EDTA, полученную в результате смесь нагревали при 65°C в течение 10 минут. Из этого раствора брали аликвоту 8 мкл и добавляли 1 мкл 50 мкМ олиго(dT)20 праймера и 1 мкл 10 мМ dNTP, полученную в результате смесь нагревали при 65°C в течение 5 минут, а затем инкубировали на льду. К этому раствору добавляли 2 мкл 10×RT буфера (производство Invitrogen, Inc.), 4 мкл 25 мМ MgCl2, 2 мкл 0,1 M дитиотреитола, 1 мкл ингибитора РНазы (RNaseOUT, 40 Ед/мкл, производство Invitrogen, Inc.) и 1 мкл обратной транскриптазы Superscript III (200 Ед/мкл, производство Invitrogen, Inc.) и суммарный объем доводили до 20 мкл. После протекания реакции при 50°C в течение 50 минут эту смесь нагревали при 85°C в течение 5 минут, а затем инкубировали на льду.
Используя полученную таким образом одноцепочечную кДНК, ПЦР в режиме реального времени проводили с использованием сочетания следующих праймеров и флуоресцентно меченных зондов (TaqMan зонд, производство Applied Biosystems, Inc.).
Условия для ПЦР в режиме реального времени:



ПЦР анализ в режиме реального времени проводили, используя систему ПЦР в режиме реального времени (Детектор последовательности ABI Prism 7700, производство Perkin Elmer Japan Applied Biosystems Division) в следующих условиях. В этой реакции использовали общую реакционную смесь для проведения ПЦР TaqMan Universal PCR Master Mix (производство Applied Biosystems, Inc.). Сначала добавляли дистиллированную воду к 25 пмоль каждого праймера, 8 нг одноцепочечной кДНК и 10 пмоль зонда TaqMan для доведения конечного объема до 25 мкл, а затем добавляли 25 мкл общей реакционной смеси для проведения ПЦР TaqMan Universal PCR Master Mix, получая, таким образом, 50 мкл реакционного раствора. Этот реакционный раствор нагревали при 50°C в течение 2 минут, а затем нагревали при 95°C в течение 10 минут и впоследствии подвергали 40 температурным циклам «95°C в течение 0,25 минут и 60°C в течение 1 минуты», таким образом, проводили ПЦР анализ в режиме реального времени. К тому же, уровень экспрессии мРНК каждого гена корректировали по уровню экспрессии мРНК рибосомного белка L32.
В результате уровни экспрессии генов катепсина К и TRAP, которые известны как маркерные молекулы остеокластов, значительно повышались со 2 дня к 3 дню после добавления RANKL (Фигуры 4-A, B). Аналогично, уровень экспрессии гена Siglec-15 также значительно повышался со 2 дня к 3 дню после добавления RANKL (Фиг. 5). Из этих результатов было обнаружено, что экспрессия гена Siglec-15 повышает сопровождающую дифференцировку в остеокласты, и, в частности, ген Siglec-15 сильно экспрессируется на поздней стадии дифференцировки.
Пример 5. Получение экспрессионной конструкции растворимого мышиного белка Siglec-15
Неполная последовательность нуклеиновой кислоты, кодирующая внеклеточный домен мышиного белка Siglec-15, представлена последовательностью SEQ ID NO:19 в перечне последовательностей, и ее аминокислотная последовательность представлена последовательностью SEQ ID NO:20 в перечне последовательностей. С использованием такой неполной последовательности может быть получен растворимый мышиный белок Siglec-15 в культуральном супернатанте клеток животных, или подобном.
a) Амплификация гена растворимого мышиного Siglec-15 с помощью ПЦР
Олигонуклеотиды, имеющие последовательности: 5'-ggggacaagt ttgtacaaaa aagcaggctt caccATGGAG GGGTCCCTCC AACTC-3' (mSiglec-15-ECD-F: SEQ ID NO:21 в перечне последовательностей); и 5'-ggggaccact ttgtacaaga aagctgggtc TCCGGGGGCG CCGTGGAAGC GGAAC-3' (mSiglec-15-ECD-R: SEQ ID NO:22 в перечне последовательностей) в качестве праймеров для амплификации кДНК внеклеточного домена мышиного Siglec-15 с помощью ПЦР, синтезировали в соответствии с общепринятой процедурой. К тому же, эти праймеры были сконструированы в качестве амплификационных праймеров для получения входного клона «шлюза» (gateway entry clone), так что последовательность attB1 добавляют к mSiglec-15-ECD-F и последовательность attB2 добавляют к mSiglec-15-ECD-R. ПЦР проводили, используя это сочетание праймеров и плазмиду Siglec-15 мыши/pcDNA3.1(+), полученную в Примере 3, в качестве матрицы в соответствии с общепринятой процедурой. Устанавливали следующие условия для термоциклера: после нагревания при 94°C в течение 2 минут температуру цикла «94°C в течение 0,5 минут, 55°C в течение 0,5 минут и 68°C в течение 1,5 минут» повторяли 15 раз, с последующим нагреванием при 68°C в течение 5 минут и инкубированием при 4°C.
b) Получение «входного клона» (entry clone) с помощью реакции Gateway BP
Входной клон, в который интегрировали кДНК внеклеточного домена мышиного Siglec-15 с помощью технологии Gateway (Invitrogen, Inc.), используя сайт-специфическую систему рекомбинации фага лямбда, получали следующим способом. Сначала проводили BP реакцию, используя BP клоназу, между ПЦР продуктом, имеющим attB последовательность с обоих концов, полученных в пункте a) и pDNOR221 (производство Invitrogen, Inc.), который является донорным вектором, имеющим последовательность attP. Используя этот реакционный раствор, трансформировали Escherichia coli DH10B, и метод молекулярных колоний проводили для клонов с лекарственной устойчивостью, и подтверждали размер вставок. Затем для клона, имеющего подтвержденную вставку правильного размера, проводили анализ последовательности всей ДНК вставки. В результате получали входной клон, который полностью идентичен целевой последовательности нуклеиновой кислоты (SEQ ID NO:19 в перечне последовательностей), кодирующей внеклеточный домен мышиного белка Siglec-15.
c) Получение экспрессионного клона с помощью реакции Gateway LR
Экспрессионный клон, в который интегрировали кДНК внеклеточного домена мышиного Siglec-15 с помощью технологии Gateway (Invitrogen, Inc.), используя сайт-специфичную систему рекомбинации фага лямбда, получали следующим способом. Входной клон, полученный в пункте b), содержит вставку, имеющую последовательность attL с обоих концов. LR реакцию, используя LR клоназу, проводили между этим входным клоном и двумя типами «адресных векторов», имеющих последовательность attR. К тому же, в качестве «адресных векторов» использовали два типа «адресных векторов»: pDONM сконструированный так, чтобы эпитопная метка V5 и метка 6×His добавлялись к С-концу вставки; и phIgFc, сконструированный так, чтобы метка Fc человека добавлялась к С-концу вставки. Используя реакционный раствор, полученный с помощью LR реакции, трансформировали Escherichia coli DH10B, и проводили метод молекулярных колоний для полученных клонов с лекарственной устойчивостью, и подтверждали размер вставок. Затем для клона, у которого подтверждено наличие вставки правильного размера, проводили анализ последовательности обоих концов со стороны вставки к стороне вектора.
Последовательности праймеров для анализа последовательностей

В результате анализа последовательностей были получены экспрессионные клоны (растворимый мышиный Siglec-15/pDONM и растворимый мышиный Siglec-15/phIgFc), в которых наблюдается правильная рекомбинация как для pDONM, так и для phIgFc соответственно. Путем трансфекции растворимого мышиного Siglec-15/pDONM в клетки животного или подобные мРНК, имеющая последовательность оснований, представленную последовательностью SEQ ID NO:25 в перечне последовательностей, транскрибируется и транслируется в белок (мышиный Siglec-15-His), имеющий аминокислотную последовательность, представленную SEQ ID NO:26 в перечне последовательностей. Далее, путем трансфекции растворимого мышиного Siglec-15/phIgFc в клетки животного или подобные мРНК, имеющая последовательность оснований, представленную последовательностью SEQ ID NO:27 в перечне последовательностей, транскрибируется и транслируется в белок (мышиный Siglec-15-Fc), имеющий аминокислотную последовательность, представленную последовательностью SEQ ID NO:28 в перечне последовательностей.
Пример 6. Изучение оптимального времени культивирования для продукции растворимого мышиного белка Siglec-15
a) Экспрессия белка с использованием клеток 293-F
Два типа экспрессионных плазмид (растворимый мышиный Siglec-15/pDONM и растворимый мышиный Siglec-15/phIgFc), полученных в Примере 5, подготавливали в количестве примерно 100 мкг соответственно. 50 мкг каждой подготовленной плазмиды перемешивали с Opti-MEM (производство Invitrogen, Inc.) с последующей стерилизацией фильтрованием. Затем добавляли 64 мкл реагента для трансфекции 293fectin (производство Invitrogen, Inc.) и полученную в результате смесь инкубировали при комнатной температуре в течение 25 минут. Каждую из полученных таким образом смесей добавляли к клеткам FreeStyle 293-F (производство Invitrogen, Inc.), культивированным во встряхиваемой колбе так, чтобы плотность клеток достигала 1,0×106 клеток/мл×50 мл в экспрессионной среде FreeStyle 293 Expression Medium (производство Invitrogen, Inc.), и эти клетки подвергали ротационному культивированию (125 оборотов/мин) при концентрации CO2 8,0% в течение 96 часов (4 дня) при 37°C. Небольшую часть культурального раствора собирали с 24-часовыми интервалами (время культивирования: 0, 24, 48, 72, 96 часов) и центрифугировали для получения культурального супернатанта. Считается, что в полученных таким образом культуральных супернатантах экспрессируется белок, в котором эпитопная метка V5 и метка 6×His были добавлены к С-концевой стороне внеклеточного домена мышиного Siglec-15 (мышиный Siglec-15-His), и белок, в котором метка Fc человека была добавлена к С-концевой стороне внеклеточного домена мышиного Siglec-15 (мышиный Siglec-15-Fc) соответственно.
b) Изменение уровня экспрессии в зависимости от времени культивирования клеток 293F, экспрессирующих мышиный Siglec-15-His
Используя образцы культурального раствора (время культивирования: 0, 24, 48, 72, 96 часов) клеток 293F, экспрессирующих мышиный Siglec-15-His, полученые в пункте a), и коммерчески доступного белка, содержащего метку «His-tag» (гистидиновая метка), рекомбинантного остеопротегрина человека/his (OPG-Fc-His) (производство R&D systems, Inc.), уровни экспрессии анализировали с помощью электрофореза в SDS-полиакриламидном геле в редуцирующих условиях и Вестерн блоттинга. То есть, к 5 мкл образца, полученного путем концентрирования каждого культурального раствора в 20 раз, используя Microcon YM-10 (производство Millipore Co., Ltd.) или 5 мкл раствора OPG-Fc-His, добавляли эквивалентное количество раствора, обработанного SDS (10 мМ Tris-HCl буфер (pH 8,0), содержащего 1 мМ EDTA, 2,5% SDS, 0,1% бромфенолового синего и 5% 2-меркаптоэтанола), и полученную в результате смесь нагревали при 95°C в течение 10 минут. 0,8 мкл каждого термически обработанного образца использовали для электрофореза в SDS-полиакриламидном геле. В качестве геля для электрофореза использовали 8-25% полиакриламидный градиентый гель (производство Amersham Biosciences, Inc.) и электрофорез проводили, используя PhastSystem (производство Amersham Biosciences, Inc.). Далее, в качестве маркеров молекулярной массы использовали маркеры для Вестерн блоттинга ECL DualVue (производство Amersham Biosciences, Inc.). После завершения электрофореза белок в геле переносили (блотировали) на PVDF мембрану (Hybond-P, производство Amersham Biosciences, Inc.), используя набор для переноса «PhastTransfer Semi-dry Transfer Kit» (производство Amersham Biosciences, Inc.) и PhastSystem. Эту PVDF мембрану переносили в 10 мл блокирующего реагента (BlockAce, производство Snow Brand Milk Products, Co., Ltd.), содержащего 0,1% Tween 20, и осторожно встряхивали при комнатной температуре в течение 1 часа. В этот блокирующий раствор добавляли 5 мкл раствора S-белка HRP (ECL DualVue Western Blotting Markers, производство Amersham Biosciences, Inc.) и 2 мкл антитела против 6-His-HRP (PentaHis HRP Conjugate kit, производство Qiagen, Inc.) и мембрану в этом растворе осторожно встряхивали при комнатной температуре в течение еще 1 часа. Эту PVDF мембрану промывали 4 раза путем осторожного встряхивания ее в 50 мл фосфатного буфера (PBS), содержащего 0,01% Tween 20 в течение 5 минут. После промывания PVDF мембрану промывали в соответствии с протоколом, приложенным к набору «ECL detection kit» (ECL Western blotting detection reagents and analysis system, производство Amersham Biosciences, Inc.) для развития окраски полосы белка, содержащего метку «His-tag», и развившуюся окраску определяли, используя мини-камеру ECL (производство Amersham Biosciences, Inc.) и пленку Polaroid (Polapan 3200B, производство Polaroid, Inc.). Результаты показаны на Фиг. 6. Из этих результатов было выбрано время культивирования 96 часов в качестве времени культивирования для клеток 293F, которые давали наиболее высокую концентрацию белка (Siglec-15-His мыши), молекулярная масса которого составляет примерно 35 кДа и который взаимодействует с антителом против 6-His-HRP.
c) Изменение уровня экспрессии в зависимости от времени культивирования клеток 293F, экспрессирующих Siglec-15-Fc мыши
Используя образцы культурального раствора (время культивирования: 0, 24, 48, 72, 96 часов) клеток 293F, экспрессирующих Siglec-15-Fc мыши, полученные в пункте a), и IgG человека (производство Sigma Co., Ltd), анализировали уровни экспрессии с помощью электрофореза в SDS-полиакриламидном геле в нередуцирующих условиях и Вестерн-блоттинг. То есть, к 5 мкл каждого образца культурального раствора или 5 мкл раствора IgG человека добавляли эквивалентное количество рабочего раствора SDS и полученную в результате смесь нагревали при 95°C в течение 10 минут. Способом, аналогичным способу, описанному выше в пункте b), используя 0,8 мкл каждого термически обработанного образца, проводили электрофорез в SDS-полиакриламидном геле, перенос (блоттинг) на PVDF мембрану, и блокировку PVDF мембраны. К этой PVDF мембране после блокировки добавляли 5 мкл раствора S-белка HRP (ECL DualVue Western Blotting Markers, производство Amersham Biosciences, Inc.) и добавляли 2 мкл антитела против IgG-Fc-HRP человека (Конъюгат пероксидазы и антитела против IgG человека (Fc-специфичный), производство Sigma Co., Ltd), и мембрану в этом растворе осторожно встряхивали при комнатной температуре в течение еще 1 часа. После промывания, проведенного образом, аналогичным способу, описанному выше в пункте b), определяли развившуюся окраску полосы Fc-содержащего белка. Эти результаты показаны на Фиг. 7. Из этих результатов было выбрано время культивирования 96 часов, как время культивирования для клеток 293F, которые продуцировали наиболее высокую концентрацию белка (Siglec-15-Fc мыши), молекулярная масса которого составляет примерно 110 кДа и который взаимодействует с антителом против Fc человека.
Пример 7. Получение в большом масштабе культурального раствора, содержащего растворимый белок мыши Siglec-15, с использованием клеток 293-F
Два типа экспрессионных плазмид (растворимый Siglec-15/pDONM мыши и растворимый Siglec-15/phIgFc мыши), полученных в Примере 5, подготавливали в количестве примерно 5 мг соответственно. К тому же, в очистке плазмид от Escherichia coli, культивированных в большом масштабе, использовали набор «Invitrogen PureLink HiPure Plasmid Gigaprep Kit» (производство Invitrogen, Inc.). Полученные таким образом плазмиды перемешивали с Opti-MEM (производство Invitrogen, Inc.) с последующей стерилизацией фильтрованием. Затем добавляли 10 мл реагента для трансфекции 293fectin (производство Invitrogen, Inc.) и полученную в результате смесь инкубировали при комнатной температуре в течение 20 минут. Каждую из полученных таким образом смесей добавляли к клеткам FreeStyle 293-F (производство Invitrogen, Inc.), культивированным в колбах Эрленмейера, так чтобы плотность клеток достигала 1,1×106 клеток/мл×5 л (1 л/колбу×5 колб) в экспрессионной среде «FreeStyle 293 Expression Medium» (производство Invitrogen, Inc.). После этого клетки подвергали ротационному культивированию (125 оборотов/мин) при концентрации CO2 8,0% в течение 96 часов (4 дня) при 37°C, культуральный раствор собирали и центрифугировали для получения культурального супернатанта. Считается, что в полученных таким образом культуральных супернатантах экспрессируется белок, в котором эпитопная метка V5 и метка 6×His были добавлены с С-концевой стороны внеклеточного домена Siglec-15 мыши (Siglec-15-His мыши), и белок, в котором метка «Fc человека» была добавлена с С-концевой стороны внеклеточного домена Siglec-15 мыши (Siglec-15-Fc мыши) соответственно.
Пример 8. Очистка Siglec-15-His мыши
a) Колоночная хроматография HisTrap HP
К 2 л культурального раствора клеток 293F, экспрессирующих Siglec-15-His мыши, полученных в Примере 7, добавляли 225 мл 10×буфера (500 мМ Tris, 1,5 M NaCl, 200 мМ имидазола, pH 8,0), и полученную в результате смесь хорошо перемешивали и фильтровали через фильтр Sterivex-GV (производство Millipore Co., Ltd.). Этот культуральный раствор наносили на колонку, которая содержала три 5 мл колонки HisTrap HP (производство Amersham Biosciences, Inc.), соединенных последовательно, и была предварительно обработана реактивом, удаляющим пирогенные вещества PyroCLEAN (производство ALerCHEK, Inc.), и промыта дистиллированной водой для инъекций со скоростью потока 2 мл/мин. После промывания колонки 60 мл 50 мМ Tris-HCl буфера (pH 8,0), содержащего 300 мМ NaCl, со скоростью потока 1 мл/мл, белок, адсорбированный на колонке, элюировали 50 мл 50 мМ Tris-HCl буфера (pH 8,0), содержащего 300 мМ NaCl, и 500 мМ имидазола со скоростью потока 1 мл/мин. Элюат фракционировали по 1 мл на фракцию в пробирки MiniSorp (производство Nunc, Inc.), в которые предварительно добавили 10 мкл 10% Tween 20. После этого примерно 20 мл раствора, полученного путем объединения фракций (фракции 14-20), содержащих элюированный белок, концентрировали до 2,5 мл с помощью центрифужного мембранного концентратора Amicon Ultra-15 (производство Millipore Co., Ltd.), концентрат наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая была предварительно уравновешена фосфатным буфером, содержащим 0,01% Tween 20 (T-PBS), с последующей элюцией T-PBS, таким образом получали 3,5 мл образца, растворитель которого был заменен на T-PBS.
b) Хроматография на колонке Resource Q
К 3,5 мл образца, который был очищен хроматографией на колонке HisTrap HP и растворитель которого был заменен на TBS-P, добавляли 22,5 мл 50 мМ Tris-HCl буфера (pH 7,5), содержащего 0,1% CHAPS, и полученную в результате смесь перемешивали. Затем смесь центрифугировали при 4°C в течение 30 минут при 3000 об/мин и преципитат удаляли. После этого полученный в результате супернатант фильтровали через фильтр Millex-GV (производство Millipore Co., Ltd.), этот фильтрат наносили на 6 мл колонку Resource Q (производство Amersham Biosciences, Inc.), которую предварительно уравновешивали 50 мМ Tris-HCl буфером (pH 7,5), содержащим 0,1% CHAPS со скоростью потока 1 мл/мин. После этого колонку промывали этим буфером со скоростью потока 1 мл/мин и собирали белковую фракцию, которая не адсорбировалась на колонке. Белок, адсорбированный на колонке, элюировали 50 мМ Tris-HCl буфером (pH 7,5), содержащим 0,1% CHAPS и 1 M NaCl, со скоростью потока 1 мл/мин. После этого 26,5 мл фракции, которая не была адсорбирована на колонке, концентрировали до 2,0 мл с помощью центрифужного мембранного концентратора Amicon Ultra-15 (производство Millipore Co., Ltd.), этот концентрат центрифугировали при 4°C в течение 10 минут при 3000 об/мин и преципитат удаляли. После центрифугирования супернатант криоконсервировали при -80°C до использования. Упомянутую выше процедуру очистки (HisTrap HP колоночная хроматография и Resource Q колоночная хроматография) проводили, дважды повторяя ее.
c) Выявление и анализ чистоты очищенного Siglec-15-His мыши
Используя образец, полученный с помощью упомянутой выше процедуры очистки (HisTrap HP колоночной хроматографии и Resource Q колоночной хроматографии), проводили электрофорез в SDS-полиакриламидной геле в редуцирующих условиях и окрашивание серебром. То есть, к 5 мкл каждого образца, очищенного с помощью соответствующих стадий очистки, добавляли эквивалентное количество рабочего раствора SDS и полученную в результате смесь подвергали термообработке при 95°C в течение 10 минут. 0,3 мкл каждого термически обработанного образца использовали для электрофореза в SDS-полиакриламидном геле. Электрофорез проводили образом, аналогичным упомянутому выше в пункте b) Примера 6, за исключением того, что в качестве маркеров молекулярной массы использовали маркеры молекулярной массы «Rainbow Molecular Weight Markers» (производство Amersham Biosciences, Inc.). После завершения электрофореза проводили окрашивание серебром, используя набор «PhastGel Silver Kit» (производство Amersham Biosciences, Inc.) и PhastSystem. Эти результаты показаны на Фиг. 8. Было показано, что белок с молекулярной массой примерно 35 кДа (Siglec-15-His мыши) был эффективно очищен и концентрирован в белковой фракции, которая не адсорбировалась на колонке Resource Q.
Электрофорез проводили в тех же условиях, за исключением того, что в качестве маркеров молекулярной массы использовали маркеры «ECL DualVue Western Blotting Markers» (производство Amersham Biosciences, Inc.), и белок в геле переносили (блоттировали) на PVDF мембрану (Hybond-P, производство Amersham Biosciences, Inc.), используя набор для переноса «PhastTransfer Semi-dry Transfer Kit» (производство Amersham Biosciences, Inc.) и PhastSystem. Эту PVDF мембрану осторожно встряхивали в 10 мл блокирующего реагента (BlockAce, производство Snow Brand Milk Products, Co., Ltd.), содержащего 0,1% Tween 20, при комнатной температуре в течение 1 часа. К этому блокирующему раствору добавляли 10 мкл S-белок HRP (производство Amersham Biosciences, Inc.) и 10 мкл антитела против V5-HRP (моноклональное антитело к Pk-TAG-HRP, производство Acris Antibodies GmbH) и мембрану в растворе осторожно встряхивали при комнатной температуре в течение еще 1 часа. PVDF мембрану промывали 4 раза путем осторожного встряхивания в 50 мл фосфатного буфера (PBS), содержащего 0,01% Tween 20 в течение 5 минут. После промывания PVDF мембрану обрабатывали в соответствии с протоколом, прилагаемым к набору «ECL detection kit» (производство Amersham Biosciences, Inc.) для развития окраски полосы белка, и развившуюся окраску выявляли, используя мини-камеру ECL (производство Amersham Biosciences, Inc.) и пленку Polaroid (Polapan 3200B, производство Polaroid, Inc.). Результаты показаны на Фиг. 9. Также на основе этих результатов можно было подтвердить, что белок имеет молекулярную массу примерно 35 кДа (Siglec-15-His мыши) и взаимодействует с антителом против V5-HRP, эффективно очищен и концентрирован в белковой фракции, которая не адсорбировалась на колонке Resource Q.
d) Измерение концентрации белка очищенного мышиного Siglec-15-His
Для очищенного мышиного Siglec-15-His (белковая фракция которого не была адсорбирована на колонке Resource Q) концентрацию белка измеряли с использованием набора «DC-Protein Assay kit» (производство Bio-Rad Laboratories, Inc.), используя бычий сывороточный альбумин в качестве стандартного образца. Как показано в Таблице 1, всего 1,66 мг очищенного белка «Siglec-15-His» было получено путем проведения процедуры очистки дважды.
Пример 9. Очистка мышиного Siglec-15-Fc
a) Хроматография на колонке HiTrap с Белком A
1,8 л культурального раствора клеток 293F, экспрессирующих мышиный Siglec-15-Fc, полученного в Примере 7, фильтровали через фильтр Sterivex-GV (производство Millipore Co., Ltd.), а затем этот фильтрат наносили на 5 мл колонку HiTrap с Белком A (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена PBS Дульбекко (D-PBS, производство Invitrogen, Inc.) при скорости потока 5 мл/мин. После того как колонку промывали D-PBS при скорости потока 5 мл/мин, белок, адсорбированный на колонке, элюировали 50 мл 0,1 M цитрат-натриевого буфера (pH 3,0) при скорости потока 5 мл/мин. Элюат фракционировали по 5 мл на фракцию в пробирки MiniSorp (производство Nunc, Inc.) и сразу после этого добавляли 1,3 мл 1 M Tris для нейтрализации элюата. После этого раствор, полученный путем объединения фракций (фракции 1 и 2), в которых был обнаружен элюированный белок, концентрировали до 2,5 мл с помощью центрифужного мембранного концентратора Amicon Ultra-15 (производство Millipore Co., Ltd.), этот концентрат наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена физиологическим раствором для инъекций Otsuka (TO-SS, производство Otsuka Pharmaceutical Co., Ltd.), содержащим 0,01% Tween 20, с последующей элюцией TO-SS, получая, таким образом, 3,5 мл образца, растворитель которого был заменен на TO-SS. Этот образец криоконсервировали при -80°C до использования. Используя 2,9 л культурального раствора клеток 293F, аналогичную процедуру очистки проводили еще раз, повторяя ее.
b) Определение и анализ чистоты очищенного мышиного Siglec-15-Fc
Используя образец, полученный с помощью указанной выше процедуры очистки, проводили электрофорез в SDS-полиакриламидном геле в редуцирующих условиях и окрашивание серебром. То есть, к 5 мкл каждого образца, очищенного с помощью соответствующих стадий очистки, добавляли эквивалентное количество рабочего раствора SDS и полученную в результате смесь нагревали при 95°C в течение 10 минут. 0,3 мкл образца, полученного путем разбавления каждого из термически обработанных образцов до 1/300 или 1/900 половиной концентрации рабочего раствора SDS, использовали для электрофореза в полиакриламидном геле. Электрофорез и окрашивание серебром проводили способом, аналогичным исследованию чистоты мышиного Siglec-15-His, описанного в пункте c) Примера 8. Результаты показаны на Фиг. 10 вместе с результатами исследования предварительных условий очистки в небольшом масштабе (pH применяемого культурального раствора составил 8,9 или 7,0). Было показано, что белок, имеющий молекулярную массу примерно 55 кДа (мышиный Siglec-15-Fc), был эффективно очищен и концентрирован в белковой фракции, которая была элюирована с колонки HiTrap с Белком A.
c) Измерение концентрации белка очищенного белка мыши Siglec-15-Fc
Для очищенного белка мыши Siglec-15-Fc (белковая фракция, элюированная с обессоливающей колонки PD-10), концентрацию белка измеряли, используя набор «DC-Protein Assay kit» (производство Bio-Rad Laboratories, Inc.), используя бычий сывороточный альбумин в качестве стандартного образца. Как показано в Таблице 2, всего было получено 92 мг очищенного белка мыши «Siglec-15-Fc», проводя процедуру очистки дважды.
Пример 10. Получение кроличьего поликлонального антитела против мышиного Siglec-15 (иммунизация кролика)
a) Получение антигена
Белок мыши «Siglec-15-Fc», полученный в Примере 9, подготавливали в концентрации 100 мкг/0,5 мл, и добавляли эквивалентное количество адъюванта и получали эмульсию, используя стеклянный шприц. В качестве адъюванта полный адъювант Фрейнда (FCA, Производство Difco Laboratories, Inc.) использовали только для первой иммунизации и неполный адъювант Фрейнда (FICA, Производство Difco Laboratories, Inc.) использовали для второй и последующих иммунизаций.
b) Иммунизация кроликов
Три кролика (самки японских белых кроликов с массой тела 3 кг) использовали в качестве иммунизированных животных. Кроме того, кровь брали до иммунизации и получали 1 мл пре-иммунной сыворотки от каждого кролика. Эмульсию, полученную в пункте а), вводили подкожно, и внутрикожно, используя иглу для инъекций 27 G, по 1 мл на кролика. Иммунизацию проводили всего 8 раз каждые 14 дней после первой иммунизации. Цельную кровь собирали через 7 дней от даты 8-й иммунизации, и 76-79 мл антисыворотки было получено от каждого кролика. Титры антител в пре-иммунной сыворотке и антисыворотке подтверждали с помощью метода ELISA, используя иммобилизованный антиген. В результате повышение титра антител в антисыворотке подтверждали у всех трех кроликов. Антисыворотку хранили при -20°C до использования.
Пример 11. Очистка поликлонального антитела против мышиного Siglec-15
a) Хроматография на колонке HiTrap с Белком A
К 20 мл каждой из трех серий кроличьих антисывороток, полученных в Примере 10, добавляли 20 мл PBS Дульбекко (D-PBS, производство Invitrogen, Inc.) и перемешивали, и полученную в результате смесь фильтровали через фильтр Sterivex-GV (производство Millipore Co., Ltd.). Затем фильтрат наносили на 5 мл колонку HisTrap с Белком А (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена D-PBS при скорости потока 2 мл/мин. После того как колонку промыли, используя 37,5 мл D-PBS при скорости потока 2,5 мл/мин, белок, адсорбированный на колонке, элюировали 50 мл 0,1 M цитрат-натриевого буфера (pH 3,0) при скорости потока 2,5 мл/мин. Элюат фракционировали по 2,5 мл на фракцию в пробирки MiniSorp (производство Nunc, Inc.) и сразу после этого 0,65 мл 1 M Tris добавляли для нейтрализации элюата. После этого примерно 10 мл раствора, полученного путем объединения фракций (фракции 2-5), содержащих элюированный белок, концентрировали до 2,5 мл с помощью центрифужного мембранного концентратора Amicon Ultra-15 (производство Millipore Co., Ltd.), этот концентрат наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена физиологическим раствором для инъекций Otsuka (TO-SS), содержащим 0,01% Tween 20, с последующей элюцией с использованием TO-SS, получая, таким образом, 3,5 мл образца, растворитель которого был заменен на TO-SS. Полученный таким образом образец криоконсервировали при -80°C до использования.
b) Выявление и анализ очистки поликлонального антитела против мышиного Siglec-15
Используя образцы (три серии, №№ 1, 2 и 3), полученные с помощью процедуры очистки, описанной в приведенном выше пункте a), проводили электрофорез в SDS-полиакриламидном геле в редуцирующих условиях и окрашивание серебром. То есть, к 5 мкл каждого образца, очищенного с помощью соответствующих стадий очистки, добавляли эквивалентное количество рабочего раствора SDS (10 мМ Tris-HCl буфер (pH 8,0), содержащего 1 мМ EDTA, 2,5% SDS, 0,1% бромфенолового синего и 5% 2-меркаптоэтанол), и полученную в результате смесь нагревали при 95°C в течение 10 минут. 0,3 мкл образца, полученного путем разведения каждого из термически обработанных образцов до 1/100, 1/300 или 1/900 половиной концентрации рабочего раствора SDS, использовали для электрофореза в SDS-полиакриламидном геле. Электрофорез и окрашивание серебром проводили способом, аналогичным анализу очистки мышиного Siglec-15-His, описанному в пункте c) Примера 8. Было показано, что белок IgG, состоящий из тяжелой цепи, имеющей молекулярную массу примерно 45 кДа, и легкой цепи, имеющей молекулярную массу примерно 21 кДа, был эффективно очищен и концентрирован в белковой фракции, которую элюировали с обессоливающей колонки PD-10.
c) Измерение концентрации белка очищенного поликлонального антитела против мышиного Siglec-15
Для очищенного поликлонального антитела против мышиного Siglec-15 (белковая фракция, элюированная с обессоливающей колонки PD-10), концентрацию белка измеряли с помощью набора «DC-Protein Assay kit» (производство Bio-Rad Laboratories, Inc.), используя бычий IgG в качестве стандартного образца. Как показано в Таблице 3, от 100 до 170 мг поликлонального антитела против мышиного Siglec-15 можно было очистить в каждой серии №№ 1-3.
d) Изучение реактивности очищенного поликлонального антитела против мышиного Siglec-15 в отношении внеклеточного домена Siglec-15
Проводили испытание для подтверждения того, что поликлональное антитело против мышиного Siglec-15, полученное в приведенном выше пункте a), связывается не только с Fc-меткой, а также с внеклеточным доменом белка Siglec-15. К 5 мкл образца очищенного мышиного Siglec-15-His (Пример 8) или к 5 мкл образца очищенного мышиного Siglec-15-Fc добавляли эквивалентное количество рабочего раствора SDS (с добавлением или без добавления 5% 2-меркаптоэтанола) и полученную в результате смесь нагревали при 95°C в течение 10 минут. 0,3 мкл каждого из термически обработанных образцов использовали для электрофореза в SDS-полиакриламидном геле и электрофорез и перенос (блоттинг) на PVDF мембрану проводили способом, аналогичным способу, описанному в приведенном выше пункте b) Примера 6. Эту PVDF мембрану осторожно встряхивали в 8 мл блокирующего реагента (BlockAce, производство Snow Brand Milk Products, Co., Ltd.), содержащего 0,1% Tween 20 при комнатной температуре в течение 1 часа. К этому блокирующему раствору добавляли 1,6 мкл поликлонального антитела № 1 против Siglec-15 мыши (Таблица 3) и PVDF мембрану в этом растворе осторожно встряхивали при комнатной температуре в течение еще 1 часа. Эту PVDF мембрану промывали 4 раза осторожным встряхиванием ее в 50 мл PBS, содержащего 0,01% Tween 20 в течение 5 минут. Промытую PVDF мембрану погружали в 8 мл блокирующего реагента «Antibody Diluent ECL Advance Blocking Agent» (ECL Advance Western Blotting Detection Kit, производство Amersham Biosciences, Inc.) и добавляли анти-кроличьи IgG-HRP (производство Amersham Biosciences, Inc.) с получением конечной концентрации 1/200000. Затем добавляли 0,8 мкл раствора S-белок HRP (ECL DualVue Western Blotting Markers, производство Amersham Biosciences, Inc.) и мембрану в растворе осторожно встряхивали при комнатной температуре в течение еще 1 часа. Эту PVDF мембрану промывали 4 раза осторожным встряхиванием ее в 50 мл PBS, содержащего 0,01% Tween 20 в течение 5 минут. После промывания PVDF мембрану обрабатывали в соответствии с протоколом, прилагаемым к набору «ECL Advance Western Blotting Detection Kit» (производство Amersham Biosciences, Inc.), и развитие окраски полосы белка определяли, используя мини-камеру ECL (производство Amersham Biosciences, Inc.) и пленку Polaroid (Polapan 3200B, производство Polaroid, Inc.). Результаты показаны на Фиг. 11. На основании этих результатов было показано, что очищенное поликлональное антитело против мышиного Siglec-15 также связывается с мышиным Siglec-15-His, и это могло быть подтверждено тем, что поликлональное антитело против мышиного Siglec-15 связывается не только с Fc-меткой, а также с внеклеточным доменом белка Siglec-15. То же испытание проводили повторно, и было подтверждено, что поликлональные антитела № 2 и № 3 против мышиного Siglec-15 также связываются с мышиным Siglec-15-His.
Пример 12. Очистка пре-иммунного IgG кролика
Кровь предварительно брали у каждого из трех кроликов, используемых в Примере 10, до начала иммунизации мышиным Siglec-15-Fc и получали из нее пре-иммунную сыворотку. После этого аликвоту 0,8 мл каждого из этих образцов сыворотки перемешивали друг с другом, добавляли 2,4 мл PBS Дульбекко (D-PBS, производство Invitrogen, Inc.) и полученную в результате смесь фильтровали через фильтр Millex-GV (производство Millipore Co., Ltd.). Затем полученный в результате образец сыворотки наносили на 5 мл колонку HiTrap с Белком A (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена D-PBS со скоростью потока 1 мл/мин. После того как колонку промыли, используя 50 мл D-PBS при скорости потока 2,5 мл/мин, белок, адсорбированный на колонке, элюировали 50 мл 0,1 M цитрат-натриевого буфера (pH 3,0) при скорости потока 2,5 мл/мин. Элюат фракционировали по 2,5 мл на фракцию в пробирки MiniSorp (производство Nunc, Inc.) и сразу после этого 0,65 мл 1 M Tris добавляли для нейтрализации элюата. После этого раствор, полученный путем объединения фракций (фракции 2-4), содержащих элюированный белок, концентрировали до 2,5 мл с помощью центрифужного мембранного концентратора Amicon Ultra-15 (производство Millipore Co., Ltd.), этот концентрат наносили на PD-10 обессоливающую колонку (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена физиологическим раствором для инъекций Otsuka (TO-SS), содержащим 0,01% Tween 20, с последующей элюцией с использованием TO-SS, получая, таким образом, 3,5 мл образца, растворитель которого был заменен на TO-SS. Очищенный таким путем образец пре-иммунного IgG кролика подвергали электорофорезу в полиакриламидном геле и окрашиванию серебром способом, описанным выше в пункте c) Примера 8 для подтверждения того, что IgG белок был достаточно очищенным, а затем измеряли концентрацию этого белка. Образец, очищенный таким образом, криоконсервировали при -80°C до использования.
Пример 13. Получение аффинной колонки с иммобилизованным на ней мышиным Siglec-15-Fc
После того как 0,54 мл растворителя 18,5 мг/мл раствора очищенного мышиного Siglec-15-Fc, описанного в Примере 9 (всего 10 мг белка), заменяли буфером для связывания (0,2 M NaHCO3, 0,5 M NaCl, pH 8,3), используя обессоливающую колонку PD-10, полученный в результате раствор концентрировали до 1 мл, используя центрифужный мембранный концентратор Amicon Ultra-4 (производство Millipore Co., Ltd.). После замены изопропанола в NHS-активированной HiTrap колонке (1 мл, производство Amersham Biosciences, Inc.) на 1 мМ соляную килоту 1 мл буфера для связывания, содержащего 10 мг/мл мышиного Siglec-15-Fc, вводили в колонку, используя шприц. После протекания реакции при комнатной температуре в течение 30 минут, для инактивации избытка активных групп, 6 мл блокирующего буфера (этаноламинный буфер, содержащий 0,5 M NaCl, pH 8,3), 6 мл промывочного буфера (ацетат-натриевый буфер, содержащий 0,5 M NaCl, pH 4,0) и 6 мл блокирующего буфера вводили последовательно в соответствии с протоколом Amersham Biosciences, Inc., а затем колонку оставляли при комнатной температуре в течение 30 минут. После этого 6 мл промывочного буфера, 6 мл блокирующего буфера и 6 мл промывочного буфера снова последовательно вводили в колонку и, наконец, буфер в колонке заменяли на 50 мМ Tris-HCl буфер (pH 7,0), содержащий 1 M NaCl и 0,01% Tween 20. Эту колонку хранили при 4°C до использования.
Пример 14. Очистка поликлонального антитела против мышиного Siglec-15 с помощью аффинной колонки
a) Аффинная хроматография на колонке
К 2 мл каждого из очищенных поликлональных антител № 1, 2 и 3 против мышиного Siglec-15, полученных в Примере 11, добавляли 8 мл буфера для нанесения (10 мМ Tris-HCl буфер, содержащий 0,15 M NaCl, pH 7,2) и полученную в результате смесь наносили на аффинную колонку (Пример 13), которая предварительно была уравновешена буфером для нанесения при скорости потока 0,25 мл/мин. После того как колонку промыли, используя 5 мл буфера для нанесения, при скорости потока 0,25 мл/мин, сначала белок, адсорбированный на колонке, элюировали 5 мл 0,1 M глицингидрохлоридного буфера (pH 2,7), содержащего 0,5 M NaCl при скорости потока 0,25 мл/мин, и после этого белок, адсорбированный на колонке, элюировали 5 мл 0,1 M цитрат-натриевого буфера (pH 2,0), содержащего 0,5 M NaCl при скорости потока 0,25 мл/мин. Хроматограмма поликлонального антитела № 3 против мышиного Siglec-15, очищенного с помощью аффинной колонки, показана на Фиг. 12. Элюат фракционировали по 0,5 мл на фракцию в пробирки MiniSorp (производство Nunc, Inc.), и сразу после этого 16 мкл 1 M Tris добавляли к 0,5 мл каждой фракции, элюированной глицингидрохлоридным буфером, и 150 мкл 1 M Tris добавляли к 0,5 мл каждой фракции, элюированной цитрат-натриевым буфером для нейтрализации элюата. Большинство поликлональных антител против мышиного Siglec-15 было элюировано с использованием 0,1 M глицингидрохлоридного буфера (pH 2,7), содержащего 0,5 M NaCl. Примерно 2,5 мл раствора, полученного путем объединения фракций (фракции 3-7), в которых IgG белок, элюированный глицингидрохлоридным буфером, выявлялся в каждой серии, наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена физиологическим раствором для инъекций Otsuka (TO-SS), содержащим 0,01% Tween 20, с последующей элюцией с использованием TO-SS, получая, таким образом, 3,5 мл образца, растворитель которого был заменен на TO-SS. В отношении белковых фракций IgG (фракции 16-19), элюированных цитрат-натриевым буфером в количестве примерно 2,5 мл, фракции для всех трех серий объединяли и полученный в результате раствор концентрировали до 2,5 мл с помощью центрифужного мембранного концентратора Amicon Ultra-4 (производство Millipore Co., Ltd.). Затем этот концентрат наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена TO-SS, с последующей элюцией с использованием TO-SS, получая, таким образом, 3,5 мл образца (Цитрат-E), растворитель которого был заменен на TO-SS. Образцы, полученные таким образом, криоконсервировали при -80°C до использования.
b) Измерение концентрации белка афинно-очищенного поликлонального антитела против мышиного Siglec-15
Для образцов очищенного поликлонального антитела против мышиного Siglec-15 (белковая фракция, элюированная с обессоливающей колонки PD-10), концентрацию белка измеряли с помощью набора «DC-Protein Assay kit» (производство Bio-Rad Laboratories, Inc.), используя бычий IgG в качестве стандартного образца. Образцы, в которых измеряли концентрацию белка, подготавливали таким образом, чтобы концентрация антитела составляла 15 или 50 мкг/мл, и подвергали электрофорезу и окрашиванию серебром аналогичным способом, что и в пункте b) Примера 11. Результаты показаны на Фиг. 13. Было показано, что IgG белок, состоящий из тяжелой цепи, имеющей молекулярную массу примерно 45 кДа, и легкой цепи, имеющей молекулярную массу примерно 21 кДа, был эффективно очищен и концентрирован в белковой фракции, элюированной с колонки PD-10. Как показано в Таблице 4, примерно от 2,3 до 7,4 мг аффинного очищенного поликлонального антитела против мышиного Siglec-15 можно было получить в каждой серии № 1-3 или в виде фракции Цитрат-E.
Пример 15. Очистка аффинно-очищенного поликлонального антитела против Siglec-15 мыши с помощью гель-фильтрационной колонки
a) Колоночная хроматография на Сефарозе 6
Для полного удаления эндотоксина и примесей низкой молекулярной массы из аффинно-очищенного поликлонального антитела против мышиного Siglec-15, полученного в Примере 14, очистку дополнительно проводили с использованием гель-фильтрационной колонки. 1 мл каждого из аффинно-очищенных поликлональных антител № 2 и 3 против мышиного Siglec-15 наносили на колонку с Сефарозой 6 HR 10/30 (производство Amersham Biosciences, Inc.), которая предварительно была обработана реагеном, удаляющим пирогенные вещества PyroCLEAN (производство ALerCHEK, Inc.), и уравновешивали PBS Дульбекко (D-PBS, производство Invitrogen, Inc.), содержащим 0,01% Tween 20, с последующей элюцией с использованием D-PBS, содержащего 0,01% Tween 20 при скорости потока 0,4 мл/мин. Их хроматограммы показаны на Фиг. 14. Элюат фракционировали по 0,5 мл на фракцию в пробирки MiniSorp (производство Nunc, Inc.) и получали 2,0 мл образца поликлонального антитела против мышиного Siglec-15, очищенного с помощью гель-фильтрации (фракции 28-31). Полученный таким образом образец криоконсервировали при -80°C до использования.
b) Измерение концентрации кроличьего белка IgG, очищенного с помощью гель-фильтрационной колонки
Концентрацию белка также измеряли для образца поликлонального антитела против мышиного Siglec-15, очищенного гель-фильтрацией (белковая фракция, элюированная с колонки с Сефарозой 6). Как показано в Таблице 5, поликлональное антитело против мышиного Siglec-15, очищенное с помощью гель-фильтрации, в количестве 2,25 мг и 3,34 мг можно было получить в сериях № 2 и 3 соответственно.
Пример 16. Получение неадгезивных клеток костного мозга мыши
Бедренную и большую берцовую кость извлекали у самцов мышей ddY в возрасте от 5 до 8 недель и удаляли мягкие ткани. Оба конца бедренной или большой берцовой кости отсекали и вводили D-PBS, используя шприц с иглой для инъекций 25 калибра для выталкивания клеток костного мозга, которые собирали в центрифужную пробирку. Центрифугирование проводили при комнатной температуре в течение 5 минут при 100 g и супернатант удаляли. К клеточному осадку добавляли 1 мл гемолитического буфера (лизирующий буфер для эритроцитов, производство Sigma Co., Ltd.) для суспендирования его и полученную в результате суспензию оставляли при комнатной температуре на 5 минут. Добавляли 20 мл D-PBS, эту суспензию центрифугировали при комнатной температуре в течение 5 минут при 100 g и супернатант удаляли. К клеточному осадку добавляли 10 мл среды MEM-α (производство Invitrogen, Inc.), содержащей 5 нг/мл M-CSF (производство R&D systems, Inc.) и 10% эмбриональную сыворотку теленка (FBS) для суспендирования осадка. Затем полученную в результате суспензию пропускали через ячеистый фильтр (40 мкм Nylon, производство BD Falcon) для удаления агрегированных веществ. Полученные в результате клетки переносили в 75 см2-T колбы (для использования адгезивных клеток) и культивировали в течение ночи в CO2 термостате. После культивирования в течение ночи клетки, которые не адгезировались на T-колбе, извлекали и использовали в качестве неадгезивных клеток костного мозга мыши.
Пример 17. Действие добавления поликлонального антитела против мышиного Siglec-15 на остеокластную дифференцировку неадгезивных клеток костного мозга мыши (стимуляция действием RANKL)
Используя поликлональные антитела против мышиного Siglec-15, полученные в Примерах 14 и 15, изучали действие на остеокластную дифференцировку неадгезивных клеток костного мозга мыши. Неадгезивные клетки костного мозга мыши, полученные с помощью упомянутого способа в Примере 16, были получены в концентрации 1,5×105 клеток/мл в среде α-MEM, содержащей 10% FBS и 10 нг/мл M-CSF (производство R&D systems, Inc.), и полученный в результате клеточный препарат высевали в каждую лунку 96-луночного планшета в количестве 200 мкл, и клетки культивировали в течение 2 дней в CO2 термостате. Старый культуральный раствор в 96-луночном планшете удаляли и в каждую лунку добавляли 100 мкл среды MEM-α, содержащей 10% FBS, к которой добавляли RANKL человека (RANKL, производство Peprotech, Inc.) и M-CSF с получением конечной концентрации 20 нг/мл и 10 нг/мл соответственно. К раствору клеточной культуры добавляли аффинно-очищенное антитело № 3, антитело Цитрат-E, антитело № 2, очищенное гель-фильтрацией, антитело № 3, очищенное гель-фильтрацией, пре-иммунный кроличий IgG (полученный в Примерах 12, 14 и 15) или коммерчески доступный контрольный IgG кролика (неиммунный кроличий IgG CLRB00, производство Cedarlane Laboratories Ltd.) в концентрации от 30 до 1000 нг/мл и клетки культивировали в течение еще 3 дней в CO2 термостате. После завершения культивирования активность тартрат-резистентной кислой фосфатазы (TRAP) образовавшихся остеокластов измеряли следующим способом. Культуральный раствор в каждой лунке 96-луночного планшета удаляли отсасыванием и 50 мкл 50 мМ цитрат-натриевого буфера (pH 6,1), содержащего 1% Тритон X-100, добавляли в каждую лунку. Затем планшет встряхивали в течение 5 минут на планшетном шейкере для лизиса клеток. В каждую лунку добавляли 50 мкл раствора субстрата (50 мМ цитрат-натриевого буфера (pH 6,1), содержащего 5 мг/мл п-нитрофенилфосфата и 0,46% тартрат натрия), и планшет инкубировали при комнатной температуре в течение 5 минут. После инкубирования 50 мкл 1 н. раствора гидроксида натрия добавляли в каждую лунку 96-луночного планшета для остановки ферментативной реакции. После остановки ферментативной реакции измеряли поглощение в каждой лунке при 405 нм и это измерение использовали как показатель активности TRAP. Результаты показаны на Фиг.15 и 16. Значительное ингибирование активности TRAP не наблюдалось в случаях пре-иммунного кроличьего IgG и коммерчески доступного контрольного кроличьего IgG. С другой стороны, значительное ингибирование активности TRAP наблюдалось в случаях аффинно-очищенного антитела № 3 в концентрации 30 нг/мл или выше и антитела Цитрат-E в концентрации примерно 130 нг/мл или выше (Фиг. 15). Также в случае антитела № 3, очищенного гель-фильтрацией, значительное ингибирование активности TRAP наблюдалось при концентрации 30 нг/мл или выше. Более того, более эффективная ингибирующая активность наблюдалась в случае антитела № 3, очищенного гель-фильтрацией, чем в случае антитела № 2, очищенного гель-фильтрацией (Фиг. 16). Поскольку активность ингибирования образования остеокластов наблюдалась также у антитела, очищенного гель-фильтрацией, было показано, что активность ингибирования образования остеокластов, наблюдаемая у поликлонального антитела против мышиного Siglec-15, не объяснялась эндотоксином или низкомолекулярными примесями, содержащимися в образце антитела, но объясняется активностью самой молекулы антитела. Исходя из представленных выше результатов, было показано, что поликлональное антитело против мышиного Siglec-15 обладает мощным ингибирующим действием на образование остеокластов (дифференцировку остеокластов и созревание).
Пример 18. Нейтрализация антигеном ингибирования остеокластной дифференцировки неадгезивных клеток костного мозга мыши путем добавления поликлонального антитела против мышиного Siglec-15 (стимуляция действием RANKL)
Было подтверждено, что действие поликлонального антитела против мышиного Siglec-15 зависит от связывания его с антигеном путем предварительного добавления антигена к поликлональному антителу против мышиного Siglec-15 для образования иммунного преципитата. К 10 мкг/мл аффинно-очищенного антитела № 3, полученного в Примере 14, добавляли мышиный Siglec-15-His или Siglec-15-Fc, полученный в Примере 8 или 9 в концентрации 10, 30, 100 или 300 мкг/мл, и полученную в результате смесь инкубировали при 37°C в течение 2 часов. После инкубирования смесь центрифугировали в течение 5 минут с использованием Chibitan и полученный в результате супернатант стерилизовали фильтрацией через фильтр Millex-GV (производство Millipore Co., Ltd.). Неадгезивные клетки костного мозга мыши, полученные способом Примера 16, высевали в 96-луночный планшет с концентрацией 200 мкл/лунку и клетки культивировали в течение 2 дней в CO2 термостате. Старый культуральный раствор в 96-луночном планшете удаляли и в каждую лунку добавляли 100 мкл среды MEM-α, содержащей 10% FBS, к которой добавляли RANKL человека (RANKL, производство Peprotech, Inc.) и M-CSF с получением конечной концентрации 20 нг/мл и 10 нг/мл соответственно. К клеточному культуральному раствору добавляли каждый из тестируемых образцов, полученных выше, в концентрации 1/200 (об/об) и клетки культивировали в течение еще 3 дней в CO2 термостате. После завершения культивирования активность тартрат-резистентной кислой фосфатазы (TRAP) образовавшихся остеокластов измеряли способом, описанным в Примере 17. Результаты показаны на Фиг. 17. В обоих случаях, где антитело было нейтрализовано действием Siglec-15-His и где антитело было нейтрализовано действием Siglec-15-Fc, действие антитела было нейтрализовано и утрачено. Эти результаты демонстрируют, что ингибирующее действие поликлонального антитела против мышиного Siglec-15 на образование остеокластов происходит вследствие связывания его с белком Siglec-15 и блокирования его функции.
Пример 19. Действие добавления поликлонального антитела против мышиного Siglec-15 на остеокластную дифференцировку неадгезивных клеток костного мозга мыши (стимуляция действием TNF)
Используя поликлональное антитело против мышиного Siglec-15, изучали действие на остеокластную дифференцировку неадгезивных клеток костного мозга мыши путем стимуляции под действием TNF. Неадгезивные клетки костного мозга мыши, полученные способом Примера 16, подготавливали в концентрации 1,5×105 клеток/мл в среде α-MEM, содержащей 10% эмбриональную сыворотку теленка (FBS), 10 нг/мл M-CSF и 2 нг/мл TGF-β (производство R&D systems, Inc.), и полученный в результате клеточный препарат высевали в каждую лунку 96-луночного планшета в количестве 200 мкл, и клетки культивировали в течение 2 дней в CO2 термостате. Старый культуральный раствор в 96-луночном планшете удаляли и в каждую лунку добавляли 100 мкл среды MEM-α, содержащей 10% FBS, в которую добавляли рекомбинантный TNF-α человека (производство R&D systems, Inc.) и M-CSF с получением конечной концентрации 30 нг/мл и 10 нг/мл соответственно. В каждый клеточный культуральный раствор добавляли пре-иммунный IgG, полученный в Примере 12, или антитело № 3 против мышиного Siglec-15, очищенное гель-фильтрацией, полученное в Примере 15, в концентрации от 30 до 1000 нг/мл, и клетки культивировали в течение еще 3 дней в CO2 термостате. В то же время также была подготовлена лунка, в которой клетки культивировали путем добавления рекомбинантного OCIF/OPG человека, полученного способом, описанным в описании к патентному документу WO 96/26217, в концентрации от 3 до 100 нг/мл. После завершения культивирования активность тартрат-резистентной кислой фосфатазы (TRAP) образовавшихся остеокластов измеряли способом, описанным в Примере 17. Результаты показаны на Фиг. 18. В случаях пре-иммунного IgG и OCIF/OPG не наблюдалось значительного ингибирования активности TRAP. С другой стороны, в случае антитела № 3 против мышиного Siglec-15, очищенного гель-фильтрацией, наблюдалось примерно 50% ингибирование активности TRAP в концентрации 250 нг/мл или выше. Исходя из этих результатов, было видно, что поликлональное антитело против мышиного Siglec-15 может ингибировать также образование остеокластов, индуцированное под действием TNF (дифференцировку и созревание остеокластов), которое не может быть ингибировано под действием OCIF/OPG.
Для каждой лунки 96-луночного планшета, подготовленной путем проведения культивирования способом, аналогичным описанному выше, окрашивание TRAP проводили, используя набор «Leukocyte Acid Phosphatase kit» (производство Sigma Co., Ltd.) в соответствии с протоколом, прилагаемым к этому набору, и наблюдали образование TRAP-позитивных многоядерных остеокластов. В результате образование TRAP-позитивных гигантских многоядерных остеокластов было ингибировано путем добавления поликлонального антитела против мышиного Siglec-15 (Фиг. 19). Поскольку одноядерные остеокласты образовывались даже в случае, когда добавляли поликлональное антитело против мышиного Siglec-15, было показано, что поликлональное антитело против мышиного Siglec-15 сильно ингибирует процесс клеточного слияния при дифференцировке и созревании остеокластов, индуцированной действием TNF.
Пример 20. Действие добавления поликлонального антитела против мышиного Siglec-15 на остеокластную дифференцировку первичных культивированных клеток костного мозга мыши (активность TRAP)
Самца мыши ddY в возрасте 7 недель подвергали эвтаназии путем смещения шейных позвонков под эфирным наркозом и извлекали бедренную и большую берцовую кости. После удаления мягких тканей оба конца бедренной или большой берцовой кости отсекали. Затем среду α-MEM, содержащую 10% эмбриональную сыворотку теленка, вводили в костный мозг, используя цилиндр шприца с инъекционной иглой 25 калибра, и получали клетки костного мозга. После подсчета количества клеток костного мозга клетки подготавливали в концентрации 5×106 клеток/мл в среде α-MEM, содержащей 10% эмбриональную сыворотку теленка. Полученный в результате клеточный препарат помещали в 96-луночный планшет с концентрацией 100 мкл/лунку и добавляли активный витамин D3 (производство Sigma Co., Ltd.) с получением конечной концентрации 2×10-8 M. К этому супернатанту клеточной культуры добавляли аффинно-очищенное антитело № 3 против мышиного Siglec-15, полученное в Примере 14, или пре-иммунный IgG кролика, полученный в Примере 12, с получением конечной концентрации 4,57, 13,7, 41,2, 123, 370, 1111, 3333 или 10000 нг/мл, и клетки культивировали в течение 8 дней в CO2 термостате. Кроме того, замену среды и добавление тестируемого вещества проводили на 3 и 6 день. Культуральный супернатант удаляли на 8 день культивирования и 10% нейтральный формалин добавляли для фиксации клеток. После фиксации клеток клетки промывали дважды дистиллированной водой, и раствор субстрата TRAP (15 мМ п-нитрофенилфосфат, 50 мМ тартрат натрия, 0,1 M ацетат-натриевый буфер (pH 5,0)) добавляли по 100 мкл/лунку, и реакцию оставляли протекать при комнатной температуре в течение 30 минут. Затем добавляли 1 н. NaOH по 50 мкл/лунку для остановки реакции и поглощение при 405 нм измеряли, используя микропланшетный ридер, оценивая, таким образом, активность TRAP в клетках. В результате активность TRAP ингибировалась в зависимости от дозы добавленного поликлонального антитела против мышиного Siglec-15 (Фиг. 20-A). С другой стороны, в случае, когда добавляли пре-иммунный IgG, снижения активности TRAP не наблюдалось (Фиг. 20-B). Аналогичным образом было обнаружено, что образование TRAP-позитивных остеокластов из клеток костного мозга мыши, индуцированное активным витамином D3, ингибируется антителом, специфически связывающимся с Siglec-15.
Пример 21. Действие добавления поликлонального антитела против мышиного Siglec-15 на клеточное слияние остеокластов из первичных культивированных клеток костного мозга мыши (окрашивание TRAP)
Самца мыши ddY в возрасте 7 недель подвергали эвтаназии путем смещения шейных позвонков под эфирным наркозом и извлекали бедренную и большую берцовую кости. После удаления мягких тканей оба конца бедренной или большой берцовой кости отсекали. Затем среду α-MEM, содержащую 10% эмбриональную сыворотку теленка, вводили в костный мозг, используя цилиндр шприца с инъекционной иглой 25 калибра, и получали клетки костного мозга. После подсчета количества клеток клетки подготавливали в концентрации 5×106 клеток/мл в среде α-MEM, содержащей 10% эмбриональную сыворотку теленка. Полученный в результате клеточный препарат помещали в 96-луночный планшет с концентрацией 100 мкл/лунку. Добавляя активный витамин D3 (производство Sigma Co., Ltd.) в конечной концентрации 2×10-8 M или RANKL человека (производство PeproTech Inc.) в конечной концентрации 80 нг/мл к этому культуральному супернатанту в качестве фактора, индуцирующего дифференцировку остеокластов, получали супернатанты клеточных культур. К каждому из этих супернатантов клеточных культур добавляли аффинно-очищенное антитело № 3 против мышиного Siglec-15, полученное в Примере 14, с получением конечной концентрации 370 или 3333 нг/мл, или добавляли пре-иммунный кроличий IgG, полученный в Примере 12, с получением конечной концентрации 3333 нг/мл. В этой культуральной системе индукции остеокластной дифференцировки под действием активного витамина D3 клетки культивировали в течение 8 дней в CO2 термостате и в системе индукции дифференцировки под действием RANKL человека клетки культивировали в течение 6 дней в CO2 термостате. Кроме того, замену среды и добавление тестируемого вещества проводили на 3 и 6 день. После культивирования супернатант удаляли и 10% нейтральный формалин добавляли для фиксации клеток. После фиксации клеток клетки промывали дважды дистиллированной водой и добавляли раствор для окрашивания TRAP (0,27 мМ нафтол AS-MX фосфат (производство Sigma Co., Ltd.), 1,6 мМ прочный красный-фиолетовый LB соль (производство Sigma Co., Ltd.), 1% диметилформамид, 50 мМ тартрат натрия, 0,1 M ацетат-натриевый буфер (pH 5,0)) в концентрации 100 мкл/лунку, и реакцию оставляли протекать при комнатной температуре в течение 5 минут. Затем клетки промывали дважды дистиллированной водой, а затем наблюдали под микроскопом. В результате в случае, когда дифференцировка остеокластов была индуцирована либо активным витамином D3 (Фиг. 21), либо RANKL человека (Фиг. 22), клеточное слияние остеокластов было ингибировано добавлением поликлонального антитела против мышиного Siglec-15 и образования гигантских остеокластов не наблюдалось. С другой стороны, в случае, когда добавляли пре-иммунный IgG, такого ингибирования клеточного слияния остеокластов не наблюдалось. Аналогичным образом, было обнаружено, что многоядерность и клеточное слияние TRAP-позитивных остеокластов из клеток костного мозга мыши, индуцированное под действием активного витамина D3 или RANKL человека, ингибируется антителом, специфически связывающимся с Siglec-15.
Пример 22. Действие добавления поликлонального антитела против мышиного Siglec-15 на клеточное слияние остеокластов из клеток RAW 264.7 (окрашивание TRAP)
a) Получение поликлонального антитела против мышиного Siglec-15, абсорбированного антигенным белком
Пять смешанных растворов (A-E) получали, используя D-PBS (производство Invitrogen, Inc.), содержащий 0,01% Tween 20, так, что относительные концентрации аффинно-очищенного антитела № 3 против мышиного Siglec-15, полученного в Примере 14, белка мыши «Siglec-15-His» и белка мыши «Siglec-15-Fc», полученного в Примерах 8 и 9, показаны в Таблице 6. Эти смешанные растворы инкубировали при 37°C в течение 2 часов и центрифугировали при 20000×g в течение 10 минут. Полученные в результате супернатанты использовали как 20-кратную концентрацию тестируемых образцов.
b) Оценка использования RAW 264.7 с помощью окрашивания TRAP
RAW 264.7 подготавливали в концентрации 2,25×104 клеток/мл в среде α-MEM, содержащей 10% эмбриональную сыворотку теленка, и полученный в результате клеточный препарат помещали в 96-луночный планшет в концентрации 200 мкл/лунку, и добавляли RANKL человека (производство PeproTech Inc.) с получением конечной концентрации 40 нг/мл. К этому супернатанту клеточной культуры каждый из тестируемых образцов A-E, полученных в пункте a), добавляли в конечной концентрации 1/20 и клетки культивировали в течение 3 дней в CO2 термостате. После культивирования супернатант удаляли и 10% нейтральный формалин добавляли для фиксации клеток. После фиксации клеток клетки промывали дважды дистиллированной водой, и раствор для окрашивания TRAP (0,27 мМ нафтол AS-MX фосфат (производство Sigma Co., Ltd.), 1,6 мМ прочного красного фиолетового LB соли (производство Sigma Co., Ltd.), 1% диметилформамида, 50 мМ тартрат натрия, 0,1 M ацетат-натриевого буфера (pH 5,0)) добавляли в концентрации 100 мкл/лунку, и реакцию оставляли протекать при комнатной температуре в течение 5 минут. Затем клетки промывали дважды дистиллированной водой, а затем наблюдали под микроскопом. Как результат, добавлением поликлонального антитела против мышиного Siglec-15 клеточное слияние остеокластов было существенно ингибировано (Фиг. 23-A) по сравнению с контролем (Фиг. 23-0) без добавления тестируемого образца. Однако путем абсорбции поликлонального антитела против мышиного Siglec-15 в белке мыши «Siglec-15-Fc», используемого в качестве иммунизирующего антигена, ингибирующее действие поликлонального антитела против мышиного Siglec-15 на клеточное слияние остеокластов было отменено (Фиг.23-B, C). Далее, также путем абсорбирования поликлонального антитела против мышиного Siglec-15 и белка мыши «Siglec-15-His» ингибирующее действие поликлонального антитела против мышиного Siglec-15 на клеточное слияние остеокластов было отменено аналогичным образом (Фиг.23-D, E). Исходя из этих результатов, было обнаружено, что многоядерность и клеточное слияние TRAP-позитивных остеокластов из клеток RAW 264.7, индуцированных под действием RANKL человека, ингибируется антителом, специфически связывающимся с Siglec-15. К тому же результаты, представленные обозначениями A-E на Фиг. 23, соответствуют тестируемым образцам A-E в Таблице 6 соответственно.
Пример 23. Создание гибридомы, продуцирующей моноклональные антитела крысы против мышиного Siglec-15
a) Получение антигена
Белок мыши «Siglec-15-His», полученный в Примере 8, подготавливали в концентрации 100 мкг/0,5 мл, и добавляли эквивалентное количество адъюванта, и получали эмульсию с помощью стеклянного шприца. В качестве адъюванта полный адъювант Фрейнда (FCA, Производство Difco Laboratories, Inc.) использовали только для первой иммунизации и неполный адъювант Фрейнда (FICA, Производство Difco Laboratories, Inc.) использовали для второй и последующих иммунизаций.
b) Иммунизация крыс
Четыре крысы (Wistar, самки, возраст 6 недель, приобретенные у CLEA Japan, Inc.) использовали в качестве иммунизируемых животных. Эмульсию, полученную в пункте a), вводили подкожно и внутрикожно, используя инъекционную иглу 27 G, так, чтобы количество антигена составляло 50 мкг на крысу. Иммунизацию проводили всего 4 раза каждые 7 дней после первой иммунизации. Небольшое количество (200 мкл) крови брали из хвостовой вены через 7 дней от даты 4-й иммунизации и получали антисыворотку. Для подтверждения титра антител антисыворотки проводили ELISA, используя иммобилизованный белок мыши Siglec-15-His в качестве антигена, белок мыши Siglec-15-Fc, полученный в Примере 9, или бычий сывороточный альбумин (BSA). В результате реакционная способность в отношении белка мыши «Siglec-15-His» и белка мыши «Siglec-15-Fc» наблюдалась у всех четырех крыс (крысы № 1-4). С другой стороны, реакционной способности в отношении BSA не наблюдалось. Исходя из этих результатов, было подтверждено, что титр антител в сыворотке каждой из иммунизированных крыс повышался, и, следовательно, крысу № 2, у которой был показан наиболее высокий титр антител, подвергали процедуре клеточного слияния.
c) Слияние клеток
Слияние клеток проводили в соответствии с общепринятым способом слияния клеток селезенки мыши (крысы) с миеломными клетками. Цельную кровь получали из сердца крысы под эфирным наркозом и крысу подвергали эвтаназии, а затем извлекали селезенку. Собранные клетки селезенки и клетки P3X63Ag8.653 (ATCC CRL 1580), которые представляют собой миеломные клетки мыши, подвергали клеточному слиянию, используя полиэтиленгликоль (PEG). Полученные в результате клетки высевали в 96-луночный планшет и добавляли среду, содержащую гипоксантин (H), аминоптерин (A) и тимидин (T) (селективная среда HAT), а затем клетки культивировали в течение 7-10 дней. Культуральный супернатант собирали из 61 лунки, в которых было подтверждено выживание гибридом, полученным путем слияния клеток. Затем оценивали титр антител с помощью ELISA, используя иммобилизованный белок мыши «Siglec-15-His», белок мыши «Siglec-15-Fc», полученный в Примере 9, или BSA в качестве антигена, и гибридомы, продуцирующие моноклональные антитела против мышиного Siglec-15, подвергали скринингу. Исходя из результатов скрининга, было отобрано 12 лунок, демонстрирующих высокий титр антител, и гибридомы, содержащиеся в этих лунках, подвергали процедуре клонирования.
d) Клонирование гибридомы
В отношении гибридом, отобранных таким образом, первое клонирование проводили методом предельных разведений. После предельного разведения гибридомы культивировали в течение 2 недель и титр антител в культуральном супернатанте подтверждали с помощью ELISA, используя иммобилизованный белок мыши «Siglec-15-Fc», полученный в Примере 9, или BSA. Для 11 клонов, которые были подтверждены как позитивные клоны, проводили второе клонирование (способом, аналогичным первому клонированию), посредством чего в конце было создано 10 клонов гибридом, продуцирующих моноклональные антитела против мышиного Siglec-15 (#1A1, #3A1, #8A1, #24A1, #32A1, #34A1, #39A1, #40A1, #41B1, #61A1). К тому же гибридомы #32A1 и #41B1 были депонированы в международном депозитарии «International Patent Organism Depositary» национального института «National Institute of Advanced Industrial Science and Technology» (расположен в Central 6, 1-1-1 Higashi, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan) 28 августа 2008. Гибридоме #32A1 был присвоен номер депонирования FERM BP-10999 под названием анти-Siglec-15 гибридома #32A1, и гибридоме #41B1 был присвоен номер депонирования FERM BP-11000 под названием анти-Siglec-15 гибридома #41B1.
Пример 24. Получение моноклонального антитела крысы против мышиного Siglec-15
a) Получение асцитической жидкости у мышей «nude»
Гибридомы, созданные в Примере 23, культивировали, используя среду «TIL Media I» (производство Immuno-biological Laboratories Co., Ltd.), содержащую 10% FCS. Субкультивирование клеток осуществляли путем проведения процедуры, в которой культуральный раствор разводили примерно до одной четвертой каждые два - три дня, используя момент времени, когда клетки были выращены примерно до 5×105 клеток/мл, в качестве ориентира. Каждую гибридому, культивированную таким образом, имплантировали внутрибрюшинно мыши «nude», которой предварительно внутрибрюшинно ввели пристан (0,2 мл/мышь) в концентрации 1×107 клеток на мышь. При имплантации использовали трех мышей «nude» для каждого из 10 клонов гибридом. После имплантации асцитическую жидкость собирали в тот момент, когда наблюдалось достаточное накопление асцитической жидкости, которую объединяли с теми, которые были получены от двух других мышей, которым имплантировали ту же гибридому, и измеряли количество асцитической жидкости, объединенной таким образом, и асцитическую жидкость криоконсервировали до очистки антитела. Количества собранной асцитической жидкости для соответствующих гибридом суммированы в Таблице 7.
b) Очистка антитела
Общее количество собранной асцитической жидкости подвергали IgG очистке, используя 20 мл колонку с Белком G (производство GE Healthcare, Co., Ltd.). Чистоту очищенного IgG исследовали с помощью гель-фильтрации (колоночная хроматография на Superdex 200) и некоторые антитела подвергали концентрации на мембранной центрифуге. То есть, 9 типов антител, за исключением антитела #24A1, концентрировали примерно до одной шестой - одной восьмой исходного объема путем центрифугирования антител при 3000 об/мин в течение 30-60 минут при 4°C, используя центрифужный мембранный концентратор Amicon Ultra-15 (производство Millipore Co., Ltd.). После этого для антитела #24A1 и других концентрированных 9 типов антител концентрацию белка измеряли с помощью набора «DC-Protein Assay kit» (производство Bio-Rad Laboratories, Inc.), используя бычий сывороточный альбумин (BSA) в качестве стандартного образца. С помощью упомянутой выше процедуры было получено моноклональное антитело против мышиного Siglec-15.
Пример 25. Оценка связывающего свойства моноклонального антитела крысы против мышиного Siglec-15 в отношении белка Siglec-15 мыши
Свойство связывания моноклонального антитела крысы против мышиного Siglec-15 с мышиным белком Siglec-15 оценивали с помощью метода ELISA. Белок мыши «Siglec-15-Fc», полученный в Примере 9, разбавляли до 5 мкг/мл 0,1 M карбонат-натриевым буфером (pH 9,5) и полученный в результате раствор добавляли в 96-луночный планшет (производство Nalge Nunc International, Inc., Cat. No. 430341) в концентрации 100 мкл/лунку. После того как планшет оставляли при комнатной температуре в течение 1 часа, раствор удаляли, и добавляли промывочный буфер (фосфатно-буферный солевой раствор, содержащий 0,05% Tween 20) в концентрации 300 мкл/лунку, и удаляли. После этого процедуру промывания проводили еще один раз, добавляли фосфатно-буферный солевой раствор, содержащий 25% BlockAce (производство Dainippon Sumitomo Pharma Co., Ltd.) в концентрации 200 мкл/лунку, и планшет оставляли при комнатной температуре в течение 1 часа, таким образом осуществляли блокировку. Жидкость удаляли и планшет промывали дважды 300 мкл/лунку промывочного буфера. Затем каждое из моноклональных антител крысы против мышиного Siglec-15, полученных в Примере 24, или контрольный IgG крысы (производство R&D systems, Inc.) разбавляли до конечной концентрации от 1,28 до 20000 нг/мл (серии 5-кратных разведений) ELISA буфером (фосфатно-буферный солевой раствор, содержащий 12,5% BlockAce и 0,05% Tween 20), и полученный в результате разбавленный раствор антитела добавляли в планшет в концентрации 100 мкл/лунку. После того как планшет оставляли при комнатной температуре в течение 1 часа, жидкость удаляли и планшет промывали три раза 300 мкл/лунку промывочного буфера. Впоследствии козье антитело, меченное HRP (пероксидазой хрена), против IgG крысы (производство Beckman Coulter, Inc.), разбавленное 1000-раз ELISA буфером, добавляли в концентрации 100 мкл/лунку и планшет оставляли при комнатной температуре в течение 1 часа. Жидкость удаляли и планшет промывали три раза 300 мкл/лунку промывочного буфера, а затем, используя набор для развития окраски для пероксидазы (производство Sumitomo Bakelite Co., Ltd.), проводили цветовую реакцию в соответствии с протоколом, прилагаемым к набору. После развития окраски измеряли поглощение при 492 нм, используя микропланшетный ридер (производство Molecular Devices Corporation, Japan). В результате подтвердилось, что все 10 тестируемых веществ исследуемых моноклональных антител крысы против мышиного Siglec-15 связываются с мышиным белком Siglec-15 зависимым от концентрации антитела образом (Фиг. 24). С другой стороны, в случае контрольного IgG крысы связывания с мышиным белком Siglec-15 не наблюдалось.
Пример 26. Оценка биологической активности моноклонального антитела крысы против мышиного Siglec-15 на основании тестирования образования остеокластов мыши
Используя все 10 тестируемых веществ моноклональных антител против мышиного Siglec-15, полученных в Примере 24, изучали действие на остеокластную дифференцировку неадгезивных клеток костного мозга мыши. Неадгезивные клетки костного мозга мыши, полученные способом Примера 16, подготавливали в концентрации 1,5×105 клеток/мл в среде α-MEM, содержащей 10% FBS и 10 нг/мл M-CSF (производство R&D systems, Inc.), и полученный в результате клеточный препарат высевали в каждую лунку 96-луночного планшета в количестве 200 мкл, и клетки культивировали в течение 2 дней в CO2 термостате. Старый культуральный раствор в 96-луночном планшете удаляли и в каждую лунку добавляли 100 мкл среды MEM-α, содержащей 10% FBS, к которой добавляли RANKL человека (RANKL, производство Peprotech, Inc.) и M-CSF с получением конечных концентраций 20 нг/мл и 10 нг/мл соответственно. К этому раствору клеточной культуры добавляли каждое из моноклональных антител крысы против мышиного Siglec-15, полученных в Примере 24, образец, полученный удалением азида натрия из коммерчески доступного контрольного IgG крысы (очищенный IgG крысы, производство R&D systems, Inc.) или кроличье поликлональное антитело против мышиного Siglec-15 (No. 3), полученное в Примере 14, в концентрации от 32 до 1000 нг/мл, и клетки культивировали в течение еще 3 дней в CO2 термостате. После завершения культивирования измеряли активность тартрат-резистентной кислой фосфатазы (TRAP) образовавшихся остеокластов методом, описанным в Примере 17. После остановки ферментативной реакции изменяли поглощение каждой лунки при 405 нм и это измерение использовали в качестве показателя активности TRAP. Результаты показаны на Фиг.25 и 26. Значительного ингибирования активности TRAP не наблюдалось в случае коммерчески доступного контрольного IgG крысы. С другой стороны, значительное ингибирование активности TRAP наблюдалось в случаях антитела #32A1 в интервале от 32 нг/мл до 1000 нг/мл, антитела #8A1 и аффинно-очищенного кроличьего поликлонального антитела № 3 в интервале от 63 нг/мл до 1000 нг/мл. Также в случаях антитела #3A1, антитела #34A1 и антитела #39A1 дозозависимое ингибирование активности TRAP наблюдалось в сравнительно высокой концентрации 500 нг/мл или выше. Ингибирования образования остеокластов мыши под действием других антител не наблюдалось. Из представленных выше результатов антитела, которые сильно ингибируют образование остеокластов мыши (дифференцировку и созревание остеокластов), были обнаружены среди полученных моноклональных антител крысы против мышиного Siglec-15. Далее, в качестве свойства, общего для антитела #3A1, антитела #8A1, антитела #32A1, антитела #34A1 и антитела #39A1, в качестве примера можно привести активность ингибирования образования остеокластов в концентрации 1000 нг/мл, т.е. 1 мкг/мл или менее.
Пример 27. Экстракция суммарной РНК из зрелых остеокластов человека
В тех случаях, когда нормальные клетки-предшественники остеокластов человека (нормальные природные клетки-предшественники остеокластов человека, приобретенные у Sanko Junyaku Co., Ltd., Cat. No. 2T-110) культивируют в минимальной эссенциальной среде для клеток-предшественников остеокластов (OPBM, приобретенной у Sanko Junyaku Co., Ltd., Cat. No. PT-8201), дополненной комплексом дополнительных веществ OPGM (Osteoclast SingleQuotTM Kit, приобретенный у Sanko Junyaku Co., Ltd., Cat. No. PT-9501), содержащей эмбриональную сыворотку теленка (конечная концентрация: 10%), RANKL человека, M-CSF человека и подобными, через 3-7 дней появляется огромное количество TRAP-позитивных многоядерных остеокластов. Используя эту систему культивирования клеток в соответствии с протоколом, прилагаемым к этому набору, получали зрелые остеокласты человека.
a) Нормальные клетки-предшественники остеокластов человека высевали в 60 лунок 96-луночного планшета в концентрации 1×104 клеток/лунку и добавляли RANKL человека с получением конечной концентрации 66 нг/мл, и клетки культивировали в течение 4 дней в CO2 термостате. Затем суммарную РНК экстрагировали из многоядерных остеокластов, используя реагент для экстракции суммарной РНК (ISOGEN, производство Nippon Gene Co., Ltd.) в соответствии с протоколом, прилагаемым к этому реагенту. Полученную суммарную РНК хранили при -80°C до использования.
b) Нормальные клетки-предшественники остеокластов человека высевали в 84 лунки каждого из 96-луночных планшетов при концентрации 1×104 клеток/лунку, и RANKL человека добавляли в каждую лунку одного из планшетов с получением конечной концентрации 53,2 нг/мл, а RANKL человека не добавляли в другой планшет, и клетки культивировали в течение 3 дней в CO2 термостате. Затем суммарную РНК экстрагировали из клеток, используя реагент для экстракции суммарной РНК (ISOGEN, производство Nippon Gene Co., Ltd.) в соответствии с протоколом, прилагаемым к этому реагенту. Полученную суммарную РНК хранили при -80°C до использования.
Пример 28. Установление последовательности открытой рамки считывания (ORF) Siglec-15 человека
a) Синтез первой цепи кДНК
Используя суммарную РНК, полученную в пункте a) Примера 27, в качестве матрицы, синтез кДНК проводили, используя праймер олиго (dT) (производство Invitrogen, Inc.) и обратную транскриптазу Superscript III (производство Invitrogen, Inc.). Эту процедуру проводили в соответствии с протоколом, прилагаемым к этому ферменту.
b) ПЦР реакция
Олигонуклеотиды, имеющие последовательности: 5'-attaagcttc accATGGAAA AGTCCATCTG GCTGC-3' (hSiglec-15-HindIII kozak-F: SEQ ID NO:29 в перечне последовательностей); и 5'-agtggatccT CACGGTGAGC ACATGGTGGC-3' (hSiglec-15-BamHI-R: SEQ ID NO:30 в перечне последовательностей) в качестве праймеров для амплификации ORF кДНК для Siglec-15 человека с помощью ПЦР, синтезировали в соответствии с общепринятым способом. ПЦР проводили, используя такое сочетание праймеров и кДНК, полученной в пункте a), в соответствии с общепринятым способом. Полученный в результате реакционный раствор для ПЦР очищали, используя набор «PureLink PCR Purification Kit» (производство Invitrogen, Inc.).
c) Клонирование в вектор pcDNA3.1(+)
Очищенный реакционный раствор для ПЦР, полученый в пункте b), и вектор pcDNA3.1(+) (производство Invitrogen, Inc.) обрабатывали ферментами рестрикции (BamHI, HindIII) с последующей нарезкой геля и очисткой, а затем проводили лигазную реакцию в соответствии с общепринятым способом. Escherichia coli TOP10 трансформировали и проводили метод молекулярных колоний для получения клонов с лекарственной резистентностью. Полную нуклеотидную последовательность ORF кДНК, встроенную в плазмиду, анализировали, используя ДНК секвенатор, для клона, в котором был получен амплифицированный продукт ожидаемого размера, и в результате было обнаружено, что эта последовательность представлена SEQ ID NO:1 в перечне последовательностей. Эта нуклеотидная последовательность была такой же, как кодирующая область ORF последовательности, зарегистрированной в базе данных NCBI GeneBank как «Siglec-15 человека» (регистрационный номер: NM_213602), и далее аминокислотная последовательность (SEQ ID NO:2 в перечне последовательностей), кодируемая этой нуклеотидной последовательностью, была на 100% идентична аминокислотной последовательности Siglec-15 человека.
Пример 29. Экспрессия мРНК Siglec-15 человека, сопровождающая дифференцировку остеокластов человека (ПЦР анализ в режиме реального времени)
К 1 мкг суммарной РНК, полученной в пункте b) Примера 27, добавляли 1 мкл 1 Ед/мкл ДНазы I и 1 мкл 10×ДНаза I буфера (производство Invitrogen, Inc.), а затем конечный объем доводили до 10 мкл, используя H2O. После протекания реакции при комнатной температуре в течение 15 минут добавляли 1 мкл 25 мМ EDTA и полученную в результате смесь нагревали при 65°C в течение 10 минут. Из этого раствора брали аликвоту 8 мкл, и добавляли 1 мкл 50 мкМ праймера oligo(dT)20 и 1 мкл 10 мМ dNTP, и полученную в результате смесь нагревали при 65°C в течение 5 минут, а затем инкубировали на льду. К этому раствору добавляли 2 мкл 10×RT буфера (производство Invitrogen, Inc.), 4 мкл 25 мМ MgCl2, 2 мкл 0,1 M дитиотреитола, 1 мкл ингибитора РНазы (RNaseOUT, 40 Ед/мкл, производство Invitrogen, Inc.) и 1 мкл обратной транскриптазы Superscript III (200 Ед/мкл, производство Invitrogen, Inc.) и суммарный объем доводили до 20 мкл. После того как реакцию оставляли протекать при 50°C в течение 50 минут, смесь нагревали при 85°C в течение 5 минут, а затем инкубировали на льду.
Используя полученную таким образом одноцепочечную кДНК, ПЦР в режиме реального времени проводили, используя сочетание следующих праймеров и флуоресцентно меченных зондов (зонд TaqMan, производство Applied Biosystems, Inc.).
Условия для ПЦР в режиме реального времени:
Праймеры для амплификации катепсина К человека:



ПЦР анализ в режиме реального времени проводили, используя систему для ПЦР в режиме реального времени (ABI Prism 7700 детектор последовательности, производство Perkin Elmer Japan Applied Biosystems Division) в следующих условиях. В этой реакции использовали общую реакционную смесь для проведения ПЦР «TaqMan Universal PCR Master Mix» (производство Applied Biosystems, Inc.). Сначала добавляли дистиллированную воду к 25 пмоль каждого праймера, 8 нг одноцепочечной кДНК и 10 пмоль зонда TaqMan для доведения конечного объема до 25 мкл, а затем добавляли 25 мкл общей реакционной смеси «TaqMan Universal PCR Master Mix», получая, таким образом, 50 мкл реакционного раствора. Этот реакционный раствор нагревали при 50°C в течение 2 минут, а затем нагревали при 95°C в течение 10 минут и после этого подвергали 40 температурным циклам «95°C в течение 0,25 минут и 60°C в течение 1 минуты», посредством чего проводили ПЦР анализ в режиме реального времени. К тому же уровень экспрессии мРНК для каждого гена корректировали по уровню экспрессии мРНК для рибосомального белка L32.
В результате было обнаружено, что уровень экспрессии гена Siglec-15 значительно повышен в случае, когда дифференцировка остеокластов человека была индуцирована добавлением RANKL аналогичным образом, как и гены катепсина K и TRAP, которые известны как маркерные молекулы остеокластов (Фиг. 27).
Пример 30. Получение экспрессионной конструкции растворимого белка Siglec-15 человека
Часть последовательности нуклеиновой кислоты, кодирующей внеклеточный домен белка Siglec-15 человека, представлена последовательностью SEQ ID NO:43 в перечне последовательностей, ее аминокислотная последовательность представлена SEQ ID NO:44 в перечне последовательностей. Используя такую неполную последовательность, может быть получен растворимый белок Siglec-15 человека в культуральном супернатанте клеток животных или подобном.
a) Амплификация гена растворимого Siglec-15 человека с помощью ПЦР
Олигонуклеотиды, имеющие последовательности: 5'-ggggacaagt ttgtacaaaa aagcaggctt caccATGGAA AAGTCCATCT GGCTGC-3' (hSiglec-15-ECD-F: SEQ ID NO:45 в перечне последовательностей); и 5'-ggggaccact ttgtacaaga aagctgggtc CCCGCTGGCG CCATGGAAGC GG-3' (hSiglec-15-ECD-R: SEQ ID NO:46 в перечне последовательностей) в качестве праймеров для амплификации кДНК внеклеточного домена Siglec-15 человека с помощью ПЦР, синтезировали в соответствии с общепринятым способом. К тому же эти праймеры были сконструированы в качестве амплификационных праймеров для получения «входного клона шлюза» (gateway entry clone), таким образом, что последовательность attB1 добавляется к hSiglec-15-ECD-F, а последовательность attB2 добавляется к hSiglec-15-ECD-R. ПЦР проводили, используя это сочетание праймеров и плазмиду Siglec-15 человека/pcDNA3.1(+), полученную в Примере 28, в качестве матрицы в соответствии с общепринятым способом. Полученный в результате раствор для ПЦР очищали, используя набор для очистки «PureLink PCR Purification Kit» (производство Invitrogen, Inc.).
b) Получение входного клона с помощью реакции Gateway BP
Входной клон, в который была интегрирована кДНК внеклеточного домена Siglec-15 человека посредством технологии Gateway (Invitrogen, Inc.) с использованием сайт-специфичной рекомбинантной системы фага лямбда, получали следующим способом. Сначала BP реакцию с использованием BP Клоназы проводили между ПЦР продуктом, имеющим последовательность attB с обоих концов, полученным в пункте a), и pDNOR221 (производство Invitrogen, Inc.), который является донорным вектором, имеющим последовательность attP. Используя этот реакционный раствор, трансформировали Escherichia coli TOP10, проводили метод молекулярных колоний для клонов с лекарственной резистентностью и подтверждали размер вставок. Затем для клона с подтвержденной вставкой правильного размера проводили анализ последовательности всей последовательности ДНК этой вставки. В результате был получен входной клон, который полностью идентичен целевой последовательности нуклеиновой кислоты (SEQ ID NO:43 в перечне последовательностей), кодирующий внеклеточный домен белка Siglec-15 человека.
c) Получение экспрессирующего клона с помощью реакции Gateway LR
Экспрессирующий клон, в который была интегрирована кДНК внеклеточного домена Siglec-15 человека с помощью технологии Gateway (Invitrogen, Inc.) с использованием сайт-специфичной рекомбинантной системы фага лямбда, был получен следующим способом. Входной клон, полученный на стадии b), содержит вставку, имеющую последовательность attL с обоих концов. LR реакцию, с использованием LR клоназы проводили между этим входным клоном и двумя типами «адресных векторов» (destination vectors), имеющими последовательность attR. К тому же в качестве «адресных векторов» использовали два типа «адресных векторов»: pDONM, сконструированный таким образом, чтобы эпитопная метка V5 и метка 6×His добавлялись к С-концу вставки; и phIgFc сконструированный таким образом, чтобы метка Fc человека добавлялась к С-концу вставки. Используя реакционный раствор, полученный с помощью LR реакции, трансформировали Escherichia coli TOP10 и проводили анализ последовательности для полученных в результате клонов с лекарственной устойчивостью для подтверждения осуществления корректной рекомбинации.
В результате анализа последовательности экспрессирующие клоны (растворимый Siglec-15 человека/pDONM и растворимый Siglec-15 человека/phIgFc), в которых произошла корректная рекомбинация, были получены как для pDONM, так и для phIgFc соответственно. Путем трансфекции растворимого Siglec-15 человека/pDONM в клетки животных или подобные мРНК, имеющая последовательность оснований, представленную последовательностью SEQ ID NO:47 в перечне последовательностей, транскрибируется и транслируется в белок (человека Siglec-15-His), имеющий аминокислотную последовательность, представленную последовательностью SEQ ID NO:48 в перечне последовательностей. Далее, путем трансфекции растворимого Siglec-15 человека/phIgFc в клетки животных или подобные мРНК, имеющая последовательность оснований, представленную последовательностью SEQ ID NO:49 в перечне последовательностей, транскрибируется и транслируется в белок (человека Siglec-15-Fc), имеющий аминокислотную последовательность, представленную последовательностью SEQ ID NO:50 в перечне последовательностей.
Пример 31. Получение в большом масштабе культурального раствора, содержащего растворимый белок Siglec-15 человека, с использованием клеток 293-F
a) Получение культурального раствора, содержащего белок человека Siglec-15-His
Растворимый белок человека Siglec-15/pDONM, полученный в Примере 30, подготавливали в количестве примерно 25 мг. К тому же в очистке плазмид от Escherichia coli, культивируемых в большом масштабе, использовали набор «Invitrogen PureLink HiPure Plasmid Gigaprep Kit» (производство Invitrogen, Inc.). Полученную таким образом плазмиду перемешивали с Opti-MEM (производство Invitrogen, Inc.), и добавляли 50 мл реагента для трансфекции 293fectin (производство Invitrogen, Inc.), и полученную в результате смесь инкубировали при комнатной температуре в течение 20 минут. Эту смесь добавляли к клеткам FreeStyle 293-F (производство Invitrogen, Inc.), культивируемым в экспрессионной среде FreeStyle 293 (производство Invitrogen, Inc.), содержащей 1% пенициллин-стрептомицин, так, чтобы плотность клеток достигала 1,0-3,4×106 клеток/мл, используя 25 л аппарат для биопроцессов в культуре (WAVE Bioreactor). После того как клетки подвергали культивированию с постоянным перемешиванием (30 вращений/мин) при концентрации CO2 от 6 до 12% в течение 96 часов (4 дня) при 37°C, культуральный раствор собирали и центрифугировали для получения культурального супернатанта. Считается, что в полученном таким образом культуральном супернатанте экспрессируется белок, в котором эпитопная метка V5 и метка 6×His была добавлена с С-концевой стороны внеклеточного домена Siglec-15 человека (Siglec-15 человека-His).
b) Получение культурального раствора, содержащего белок человека Siglec-15-Fc
Растворимый Siglec-15 человека/phIgFc, полученный в Примере 30, получали в количестве примерно 5 мг. К тому же в очистке плазмид от Escherichia coli, культивированных в большом масштабе, использовали набор «Invitrogen PureLink HiPure Plasmid Gigaprep Kit» (производство Invitrogen, Inc.). Полученную таким образом плазмиду перемешивали с Opti-MEM (производство Invitrogen, Inc.) с последующей стерилизацией фильтрованием. Затем добавляли 10 мл реагента для трансфекции 293fectin (производство Invitrogen, Inc.) и полученную в результате смесь инкубировали при комнатной температуре в течение 20 минут. Эту смесь добавляли к клеткам FreeStyle 293-F (производство Invitrogen, Inc.), культивируемым в колбах Эрленмейера, таким образом, чтобы плотность клеток достигала 1,0-3,0×106 клеток/мл×5 л (1 л/колбу×5 колб) в экспрессионной среде FreeStyle 293 (производство Invitrogen, Inc.). После этого клетки подвергали ротационному культивированию (125 вращений/мин) с концентрацией CO2 8,0% в течение 96 часов (4 дня) при 37°C, культуральный раствор собирали и центрифугировали для получения культурального супернатанта. Считается, что в полученном таким образом супернатанте экспрессируется белок, в котором метка Fc человека была добавлена с С-концевой стороны внеклеточного домена Siglec-15 человека (белок человека Siglec-15-Fc).
Пример 32. Очистка растворимого белка Siglec-15 человека
a) Очистка растворимого белка человека Siglec-15-His
a-i) Колоночная хроматография HisTrap HP
К 12 л культурального раствора клеток 293F, экспрессирующих белок человека Siglec-15-His, полученного на стадии a) Примера 31, добавляли 1350 мл 10×буфера (500 мМ Tris, 1,5 M NaCl, 200 мМ имидазол, pH 8,0), и полученную в результате смесь хорошо перемешивали и фильтровали через фильтр MilliPak-60 (производство Millipore Co., Ltd.). Этот культуральный раствор наносили на 100 мл колонку с Ni-Sepharose HP (производство Amersham Biosciences, Inc.), которая предварительно была промыта чистой водой (Milli-Q water) при скорости потока 10 мл/мин. После того как колонку промыли, используя 400 мл 50 мМ Tris-HCl буфера (pH 8,0), содержащего 300 мМ NaCl при скорости потока 8 мл/мин, белок, адсорбированный на колонке, элюировали 200 мл 50 мМ Tris-HCl буфера (pH 8,0), содержащего 300 мМ NaCl и 500 мМ имидазола при скорости потока 2,5 мл/мин, и элюат фракционировали в пробирки MiniSorp (производство Nunc, Inc.). Для предотвращения преципитации белка 8 мл 5 M раствора NaCl добавляли примерно к 40 мл фракции, содержащей элюированный белок, с последующим перемешиванием, а затем полученную в результате смесь концентрировали примерно до 20 мл с помощью центрифужного мембранного концентратора Amicon Ultra-15 (производство Millipore Co., Ltd.). Нерастворимое вещество, образованное во время концентрирования, удаляли центрифугированием при 3000 об/мин в течение 30 минут при 4°C и 2,5 мл полученного в результате супернатанта наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена фосфатно-буферным солевым раствором, содержащим 1 M NaCl (N-PBS), с последующей элюцией с использованием N-PBS, посредством чего получали 3,5 мл образца, растворитель которого был заменен на N-PBS. Эту процедуру проводили еще 7 раз, повторяя ее, и получали примерно 28 мл раствора частично очищенного белка человека Siglec-15 -His.
a-ii) Хроматография на колонке Resource Q
12 мл образца, который был очищен с помощью колоночной хроматографии на Ni-Sepharose HP и растворитель которого был заменен на N-PBS, подвергали диализу в течение ночи при 4°C против 50 мМ Tris-HCl буфера (pH 7,5), содержащего 0,1% CHAPS (1 л, три раза), и полученный в результате диализат центрифугировали при 3000 об/мин в течение 30 минут при 4°C, и преципитат удаляли. После того как полученный в результате супернатант профильтровали через фильтр Millex-GV (производство Millipore Co., Ltd.), этот фильтрат наносили на 6 мл колонку Resource Q (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена 50 мМ Tris-HCl буфером (pH 7,5), содержащим 0,1% CHAPS, со скоростью потока 1 мл/мин. После этого колонку промывали этим буфером со скоростью потока 1 мл/мин и собирали белковую фракцию, которая не была адсорбирована на колонке. Белок, адсорбированный на колонке, элюировали 50 мМ Tris-HCl буфером (pH 7,5), содержащим 0,1% CHAPS и 1 M NaCl со скоростью потока 1 мл/мин. После этого 26,5 мл фракции, которая не была адсорбирована на колонке, концентрировали до 3,0 мл с помощью центрифужного мембранного концентратора Amicon Ultra-15 (производство Millipore Co., Ltd.), концентрат центрифугировали при 3000 об/мин в течение 10 минут при 4°C и преципитат удаляли. 2,5 мл полученного в результате супернатанта наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена фосфатно-буферным солевым раствором, содержащим 50 мМ аргинин гидрохлорида (pH 7,0, A-PBS), с последующей элюцией с использованием A-PBS, получая, таким образом, 3,5 мл образца, растворитель которого был заменен на A-PBS. В растворитель полученного образца добавляли аргинина гидрохлорид для предотвращения преципитации растворимого белка человека Siglec-15-His. Супернатант после центрифугирования криоконсервировали при -80°C до использования. Упомянутую выше процедуру очистки (хроматографию на колонке Resource Q) проводили дважды, повторяя ее.
a-iii) Выявление и анализ чистоты очищенного белка человека Siglec-15-His
Используя образец, полученный с помощью упомянутой выше процедуры очистки (Ni-Sepharose HP колоночная хроматография и Resource Q колоночная хроматография), проводили электрофорез в SDS-полиакриламидном геле в редуцирующих условиях и окрашивание серебром. То есть, к 5 мкл каждого образца, очищенного с помощью соответствующих стадий очистки, добавляли эквивалентное количество рабочего раствора SDS и полученную в результате смесь подвергали термической обработке при 95°C в течение 10 минут. 0,3 мкл каждого из термически обработанных образцов использовали для электрофореза в SDS-полиакриламидном геле. Процедуру электорофореза проводили аналогичным образом, как и способ, описанный в Примере 8, за исключением того, что в качестве маркеров молекулярной массы использовали маркеры «Rainbow Molecular Weight Markers» (производство Amersham Biosciences, Inc.). После завершения электрофореза проводили окрашивание серебром, используя набор «PhastGel Silver Kit» (производство Amersham Biosciences, Inc.) и PhastSystem. Результаты показаны на Фиг. 28. Было показано, что белок, имеющий молекулярную массу примерно 35 кДа (белок человека Siglec-15-His), был эффективно очищен и концентрирован в белковой фракции, которая не адсорбировалась на колонке Resource Q.
a-iv) Измерение концентрации белка очищенного Siglec-15 человека-His
Для очищенного белка человека Siglec-15-His (белковая фракция, которая не была адсорбирована на колонке Resource Q), концентрацию белка измеряли с помощью набора «DC-Protein Assay kit» (производство Bio-Rad Laboratories, Inc.), используя бычий сывороточный альбумин в качестве стандартного образца. Путем проведения процедуры очистки дважды всего было получено 1,66 мг очищенного белка человека Siglec-15-His.
b) Очистка растворимого белка человека Siglec-15-Fc
b-i) Хроматография на колонке HiTrap с Белком A
1,5 л культурального раствора клеток 293F, экспрессирующих белок человека Siglec-15-Fc, полученного в пункте b) Примера 31, фильтровали через фильтр Sterivex-GV (производство Millipore Co., Ltd.), а затем фильтрат наносили на 5 мл колонку HiTrap с Белком А (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена PBS Дульбекко (D-PBS, производство Invitrogen, Inc.) при скорости потока 5 мл/мин. После того как колонку промыли, используя 70 мл D-PBS при скорости потока 5 мл/мин, белок, адсорбированный на колонке, элюировали 24 мл 0,1 M цитрат-натриевого буфера (pH 3,0) при скорости потока 1,2 мл/мин. Элюат фракционировали по 1,2 мл на фракцию в пробирки MiniSorp (производство Nunc, Inc.) и сразу после этого 0,31 мл 1 M Tris добавляли для нейтрализации элюата. Аликвоту 2,5 мл раствора (примерно 7,5 мл), полученного путем объединения элюированных белковых фракций (фракции 5-9), наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена фосфатно-буферным солевым раствором, содержащим 50 мМ аргинин гидрохлорида (pH 7,0, A-PBS), с последующей элюцией с использованием A-PBS, получая, таким образом, 3,5 мл образца, растворитель которого был заменен на A-PBS. Эту процедуру проводили дважды, повторяя ее. В растворитель добавляли аргинина гидрохлорид для предотвращения преципитации растворимого белка человека Siglec-15-Fc. 2,5 мл оставшегося раствора элюированной белковой фракции (фракции 5-9) наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена фосфатно-буферным солевым раствором, содержащим 1 M NaCl (pH 6,7, N-PBS), с последующей элюцией с использованием N-PBS, получая, таким образом, 3,5 мл образца, растворитель которого был заменен на N-PBS. NaCl в растворителе в полученном образце добавляли для предотвращения преципитации растворимого белка человека Siglec-15-Fc без добавления соединения, содержащего аминогруппы, такого как аргинин. Образец белка человека Siglec-15-Fc, растворитель которого был заменен на N-PBS, использовали только когда иммобилизованная колонка была получена в следующем пункте c) Примера 34, а во всех других Примерах использовали белок человека Siglec-15-Fc, растворитель которого был заменен на A-PBS. Образцы, полученные с помощью упомянутой выше процедуры, криоконсервировали при -80°C до использования.
b-ii) Выявление и анализ чистоты очищенного белка человека Siglec-15-Fc
Используя образцы, полученные с помощью упомянутой выше процедуры очистки, проводили электрофорез в SDS-полиакриламидном геле в редуцирующих условиях и окрашивание серебром. То есть, к 5 мкл каждого образца, очищенного с помощью соответствующих стадий очистки, добавляли эквивалентное количество рабочего раствора SDS и полученную в результате смесь нагревали при 95°C в течение 10 минут. 0,3 мкл образца, полученного путем разведения каждого из термически обработанных образцов, до 1/100 или 1/300 половиной концентрации рабочего раствора SDS, использовали для электрофореза в SDS-полиакриламидном геле. Электрофорез и окрашивание серебром проводили способом, аналогичным анализу очистки белка человека Siglec-15-His, описанному в пункте a-iii). Результаты показаны на Фиг. 29. Было показано, что белок, имеющий молекулярную массу примерно 55 кДа (белок человека Siglec-15-Fc), был эффективно очищен и концентрирован в белковой фракции, которая была элюирована с колонки HiTrap с Белком А.
b-iii) Измерение концентрации очищенного белка человека Siglec-15-Fc
Для очищенного белка человека Siglec-15-Fc (белковая фракция, элюированная с обессоливающей колонки PD-10), концентрацию белка измеряли с помощью набора «DC-Protein Assay kit» (производство Bio-Rad Laboratories, Inc.), используя бычий сывороточный альбумин в качестве стандартного образца. Как показано в Таблице 8, всего было получено 25,2 мг очищенного белка человека Siglec-15-Fc путем проведения процедуры очистки дважды.
Пример 33. Получение кроличьего поликлонального антитела против Siglec-15 человека (иммунизация кролика)
a) Получение антигена
Белок человека Siglec-15-Fc, полученный в пункте b) Примера 32, подготавливали в концентрации 100 мкг/0,5 мл, и добавляли эквивалентное количество адъюванта, и получали эмульсию, используя стеклянный шприц. В качестве адъюванта полный адъювант Фрейнда (FCA, Производство Difco Laboratories, Inc.) использовали только для первой иммунизации и неполный адъювант Фрейнда (FICA, Производство Difco Laboratories, Inc.) использовали для второй и последующих иммунизаций.
b) Иммунизация кроликов
Три кролика (самки белых японских кроликов с массой тела 3 кг) использовали в качестве иммунизируемых животных. Кроме того, кровь брали до иммунизации, и было получено по 10 мл пре-иммунной сыворотки на кролика. Эмульсию, полученную в пункте a), вводили подкожно и внутрикожно, используя инъекционную иглу 27 G, так, чтобы количество антигена составляло 50 мкг на кролика. Иммунизацию проводили всего 8 раз каждые 14 дней после первой иммунизации. Цельную кровь собирали через 7 дней с даты 8-й иммунизации и получали 74,4-74,9 мл антисыворотки на кролика. Титры антител в пре-иммунной сыворотке и антисыворотке подтверждали методом ELISA, используя иммобилизованный антиген. В результате повышение титра антител в антисыворотке было подтверждено у всех трех кроликов. Антисыворотку хранили при -20°C до использования.
Пример 34. Очистка кроличьего поликлонального антитела против Siglec-15 человека
a) Хроматография на колонке HiTrap с Белком А
К 40 мл каждой из трех серий антисыворотки кроликов, полученной в пункте b) Примера 33, добавляли 40 мл PBS Дульбекко (D-PBS, производство Invitrogen, Inc.) и перемешивали, и полученную в результате смесь фильтровали через фильтр Sterivex-GV (производство Millipore Co., Ltd.). Затем фильтрат наносили на колонку, которая содержала две 5 мл колонки HiTrap с Белком А (две колонки были соединены последовательно, производство Amersham Biosciences, Inc.) и предварительно была уравновешена с использованием D-PBS при скорости потока 2 мл/мин. После того как колонку промыли, используя 35 мл D-PBS со скоростью потока 1 мл/мин, белок, адсорбированный на колонке, элюировали 50 мл 0,1 M цитрат-натриевого буфера (pH 3,0) со скоростью потока 1 мл/мин. Элюат фракционировали по 2,5 мл на фракцию в пробирки MiniSorp (производство Nunc, Inc.) и сразу после этого 0,6 мл 1 M Tris добавляли для нейтрализации элюата. После этого примерно 15,5 мл раствора, полученного путем объединения фракций (фракции 3-7), содержащих элюированный белок, концентрировали до 5 мл с помощью центрифужного мембранного концентратора Amicon Ultra-15 (производство Millipore Co., Ltd.), аликвоту 2,5 мл этого концентрата наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена физиологическим раствором для инъекций Otsuka (TO-SS), содержащим 0,01% Tween 20, с последующей элюцией с использованием TO-SS, получая, таким образом 3,5 мл образца, растворитель которого был заменен на TO-SS. Эту процедуру проводили дважды, повторяя ее. Образцы, полученные таким образом, криоконсервировали при -80°C до использования.
b) Очистка пре-иммунного кроличьего IgG
Кровь предварительно получали от трех кроликов, используемых в Примере 33, до начала иммунизации с использованием белка человека Siglec-15-Fc, и получали пре-иммунную сыворотку. После этого 5 мл аликвоты каждого из этих образцов сыворотки перемешивали друг с другом, добавляли 15 мл D-PBS и полученную в результате смесь фильтровали через фильтр Millex-GV (производство Millipore Co., Ltd.). Затем полученный в результате образец сыворотки наносили на колонку, которая содержала две 5 мл колонки HiTrap с Белком А (производство Amersham Biosciences, Inc.) и предварительно была уравновешена с использованием D-PBS со скоростью потока 1 мл/мин. После того как колонку промыли, используя 35 мл D-PBS со скоростью потока 1 мл/мин, белок, адсорбированный на колонке, элюировали 50 мл 0,1 M цитрат-натриевого буфера (pH 3,0) со скоростью потока 1 мл/мин. Элюат фракционировали по 2,5 мл на фракцию в пробирки MiniSorp (производство Nunc, Inc.) и сразу после этого 0,6 мл 1 M Tris добавляли для нейтрализации элюата. После этого раствор, полученный объединением фракций (фракции 4-6), содержащих элюированный белок, концентрировали до 2,5 мл с помощью центрифужного мембранного концентратора Amicon Ultra-15 (производство Millipore Co., Ltd.), этот концентрат наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена физиологическим раствором для инъекций Otsuka (TO-SS), содержащим 0,01% Tween 20, с последующей элюцией с использованием TO-SS, получая, таким образом 3,5 мл образца, растворитель которого был заменен на TO-SS. Полученный таким образом образец пре-иммунного кроличьего IgG подвергали электрофорезу в полиакриламидном геле и окрашиванию серебром способом, описанным в Примере 8, для подтверждения того, что белок IgG был достаточно чистым, а затем измеряли концентрацию белка. Очищенный таким образом образец пре-иммунного кроличьего IgG криоконсервировали при -80°C до использования.
c) Получение аффинной колонки с иммобилизованным на ней белком человека Siglec-15-Fc
3 мл очищенного белка человека Siglec-15-Fc, растворитель которого был заменен на N-PBS, полученного в пункте b) Примера 32 (всего 7,8 мг белка), концентрировали до 2 мл, используя центрифужный мембранный концентратор Amicon Ultra-4 (производство Millipore Co., Ltd.). К этому концентрату добавляли буфер для связывания (0,2 M NaHCO3, 0,5 M NaCl, pH 8,3) для доведения конечного объема до 2,5 мл и растворитель заменяли на 3,5 мл буфера для связывания, используя обессоливающую колонку PD-10. После этого изопропанол в NHS-активированной HiTrap колонке (1 мл, производство Amersham Biosciences, Inc.) заменяли на 1 мМ соляную кислоту, 3 мл полученного белка человека Siglec-15-Fc вводили в колонку, используя шприц, и жидкость альтернативно вводили туда, используя другой шприц, соединенный с выходным отверстием колонки для осуществления реакции соединения. После того как реакцию оставляли протекать при комнатной температуре в течение 30 минут, для инактивации избытка активных групп, 6 мл блокирующего буфера (этаноламинный буфер, содержащий 0,5 M NaCl, pH 8,3), 6 мл промывочного буфера (ацетат-натриевый буфер, содержащий 0,5 M NaCl, pH 4,0), и 6 мл блокирующего буфера вводили последовательно в соответствии с протоколом компании Amersham Biosciences, Inc., а затем колонку оставляли при комнатной температуре в течение 30 минут. После этого 6 мл промывочного буфера, 6 мл блокирующего буфера и 6 мл промывочного буфера снова последовательно вводили в колонку и, наконец, этот буфер в колонке заменяли на 10 мМ Tris-HCl буфер (pH 7,2), содержащий 0,15 M NaCl. Эту колонку хранили при 4°C до использования.
d) Очистка кроличьего поликлонального антитела против Siglec-15 человека с помощью аффинной колонки
d-i) Аффинная хроматография на колонке
После того как 7 мл каждого из очищенных поликлональных антител против Siglec-15 человека (№ 1, 2 и 3), полученных в пункте a), профильтровали через фильтр Millex-GV (производство Millipore Co., Ltd.), полученный в результате фильтрат наносили на колонку, полученную в пункте c), на которой был иммобилизован белок человека Siglec-15-Fc и которая предварительно была уравновешена буфером для нанесения при скорости потока 0,25 мл/мин. После того как колонку промыли, используя 5 мл буфера для нанесения при скорости потока 0,25 мл/мин, белок, адсорбированный на колонке, элюировали 5 мл 0,1 M глицин гидрохлоридным буфером (pH 2,7), содержащим 0,5 M NaCl при скорости потока 0,25 мл/мин. Хроматограммы поликлональных антител против Siglec-15 человека (№№ 1, 2 и 3), очищенных с помощью аффинной колонки, показаны на фиг. 30. Элюат фракционировали по 0,5 мл на фракцию в пробирки MiniSorp (производство Nunc, Inc.) и сразу после этого 16 мкл 1 M Tris добавляли для нейтрализации элюата. Примерно 2,5 мл раствора, полученного путем объединения белковых фракций IgG (фракции 3-7), элюированных глицин гидрохлоридным буфером, для каждого антитела, наносили на обессоливающую колонку PD-10 (производство Amersham Biosciences, Inc.), которая предварительно была уравновешена фосфатно-буферным солевым раствором Дульбекко, содержащим 0,01% Tween 20 (T-PBS), с последующей элюцией с использованием T-PBS, получая, таким образом, 3,5 мл образца, a растворитель которого был заменен на T-PBS. Образцы, полученные таким образом, криоконсервировали при -80°C до использования.
d-ii) Измерение концентрации белка аффинно-очищенного кроличьего поликлонального антитела против Siglec-15 человека
Для образцов очищенного кроличьего поликлонального антитела против Siglec-15 человека (белковая фракция, элюированная с обессоливающей колонки PD-10) концентрацию белка измеряли с помощью набора «DC-Protein Assay kit» (производство Bio-Rad Laboratories, Inc.), используя бычий IgG в качестве стандартного образца. Как показано в Таблице 9, примерно от 9,1 до 11,9 мг аффинно-очищенного поликлонального антитела против Siglec-15 человека могло быть получено в каждой серии № 1-3.
Пример 35. Действие добавления кроличьего поликлонального антитела против Siglec-15 человека на клеточное слияние нормальных клеток-предшественников остеокластов человека (окрашивание TRAP)
Нормальные клетки-предшественники остеокластов человека (Нормальные природные клетки-предшественники остеокластов человека, приобретенные у Sanko Junyaku Co., Ltd., Cat. № 2T-110) высевали в 96-луночный планшет в концентрации 1×104 клеток/лунку в соответствии с протоколом, прилагаемым к клеткам. В качестве среды использовали минимальную эссенциальную среду для клеток-предшественников остеокластов (OPBM, приобретенные у Sanko Junyaku Co., Ltd., Cat. № PT-8201), дополненную комплексом дополнительных веществ OPGM (Osteoclast SingleQuotTM Kit, приобретенный у Sanko Junyaku Co., Ltd., Cat. № PT-9501), содержащим эмбриональную сыворотку теленка (конечная концентрация: 10%), RANKL человека (конечная концентрация: 69 нг/мл), M-CSF человека (конечная концентрация: 33 нг/мл) и подобное. К полученному в результате культуральному супернатанту добавляли аффинно-очищенное антитело № 2 против Siglec-15 человека, полученное в пункте d) Примера 34, в конечной концентрации 3 или 30 мкг/мл, или пре-иммунный кроличий IgG, полученный в пункте b) Примера 34 в конечной концентрации 30 мкг/мл, и клетки культивировали в течение 5 дней в CO2 термостате. После культивирования супернатант удаляли и 10% нейтральный формалин добавляли для фиксации клеток. После фиксации клеток клетки дважды промывали дистиллированной водой, и добавляли раствор для окрашивания TRAP (0,27 мМ нафтол AS-MX фосфат (производство Sigma Co., Ltd.), 1,6 мМ устойчивого красного фиолетового LB соль (производство Sigma Co., Ltd.), 1% диметилформамид, 50 мМ тартрат натрия, 0,1 M ацетат-натриевый буфер (pH 5,0)) в концентрации 100 мкл/лунку, и реакцию оставляли протекать при комнатной температуре в течение 5 минут. Затем клетки дважды промывали дистиллированной водой, а затем наблюдали под микроскопом (фиг. 31). В результате, образование гигантских остеокластов, происходящее в результате высокой степени слияния клеток, было существенно ингибировано путем добавления поликлонального антитела против Siglec-15 человека. С другой стороны, в том случае, когда добавляли пре-иммунный IgG, такого ингибирования клеточного слияния остеокластов не наблюдалось. Количество TRAP-позитивных многоядерных клеток, в которых количество ядер равно 5 или более, подсчитывали с помощью инвертированного микроскопа (фиг. 32). В результате значительное ингибирование образования многоядерных остеокластов наблюдалось в лунке, в которую добавляли аффинно-очищенное антитело против Siglec-15 человека № 2 в конечной концентрации 30 мкг/мл. Также в том случае, когда пре-иммунный IgG добавляли в концентрации 30 мкг/мл, наблюдалась тенденция ингибирования образования многоядерных остеокластов. Однако, когда было проведено сравнение с такими лунками, было показано, что образование многоядерных остеокластов было существенно ингибировано добавлением антитела № 2 против Siglec-15 человека. Аналогичным способом, было обнаружено, что многоядерность и клеточное слияние TRAP-позитивных остеокластов из нормальных клеток-предшественников остеокластов человека ингибируется под действием антитела, специфически связывающегося с Siglec-15.
Пример 36. Оценка свойства связывания моноклонального антитела крысы против мышиного Siglec-15 с белком Siglec-15 человека
Свойство связывания моноклонального антитела крысы против мышиного Siglec-15 с белком Siglec-15 человека оценивали с помощью метода ELISA. Белок человека «Siglec-15-Fc», полученный в пункте b) Примера 32, разбавляли до 5 мкг/мл 0,1 M карбонат-натриевым буфером (pH 9,5) и полученный в результате раствор добавляли в 96-луночный планшет (производство Nalge Nunc International, Inc., Cat. № 430341) в концентрации 100 мкл/лунку. После того как планшет оставляли при комнатной температуре в течение 1 часа, раствор удаляли, и промывочный буфер (фосфатно-буферный солевой раствор, содержащий 0,05% Tween 20) добавляли в концентрации 300 мкл/лунку и удаляли. После этого процедуру промывания проводили еще один раз, добавляли фосфатно-буферный солевой раствор, содержащий 25% BlockAce (производство Dainippon Sumitomo Pharma Co., Ltd.) в концентрации 200 мкл/лунку и планшет оставляли при комнатной температуре в течение 1 часа, посредством чего осуществляли блокировку. Жидкость удаляли и планшет промывали дважды с использованием 300 мкл/лунку промывочного буфера. Затем каждое из моноклональных антител крысы против мышиного Siglec-15, полученных в Примере 24, или контрольный IgG крысы (производство R&D systems, Inc.) разбавляли до конечной концентрации от 1,28 до 20000 нг/мл (серии 5-кратных разведений) ELISA буфером (фосфатно-буферный солевой раствор, содержащий 12,5% BlockAce и 0,05% Tween 20) и полученный в результате разбавленный раствор антитела добавляли в планшет в концентрации 100 мкл/лунку. После этого планшет оставляли при комнатной температуре на 1 час, жидкость удаляли и планшет промывали три раза с использованием 300 мкл/лунку промывочного буфера. Впоследствии козье антитело, меченное HRP (пероксидазой хрена) против IgG крысы (производство Beckman Coulter, Inc.), разбавленное до 1000-раз ELISA буфером, добавляли в концентрации 100 мкл/лунку и планшет оставляли при комнатной температуре на 1 час. Жидкость удаляли и планшет промывали три раза с использованием 300 мкл/лунку промывочного буфера, а затем, используя набор для развития окраски для пероксидазы (производство Sumitomo Bakelite Co., Ltd.), проводили развитие цветной реакции в соответствии с протоколом, прилагаемым к этому набору. После развития окраски измеряли поглощение при 492 нм, используя микропланшетный ридер (производство Nihon Molecular Devices Corporation). В результате было подтверждено, что все 10 тестируемых веществ, изучаемых моноклональных антител крысы против мышиного Siglec-15, связываются с белком Siglec-15 человека в зависимости от концентрации антитела (фиг. 33). В частности, связывающая активность 5 тестируемых веществ: #1A1, #3A1, #24A1, #32A1 и #61A1, была высокой, и связывающая активность 3 тестируемых веществ: #8A1, #34A1 и #39A1, была низкой. С другой стороны, в случае контрольного IgG крысы связывания с белком Siglec-15 человека не наблюдалось. Исходя из представленных выше результатов, было показано, что моноклональные антитела крысы против мышиного Siglec-15, полученные в Примере 24, связываются не только с мышиным Siglec-15, а также с Siglec-15 человека, и, более того, было обнаружено, что некоторые антитела прочно связываются с Siglec-15 человека.
Пример 37. Действие добавления моноклонального антитела крысы против мышиного Siglec-15 на слияние клеток и активность в отношении резорбции кости нормальных клеток-предшественников остеокластов человека (оценка биологической активности in vitro)
Поскольку в Примере 36 было подтверждено, что моноклональные антитела крысы против мышиного Siglec-15 связываются также с Siglec-15 человека, изучали действия этих антител на образование остеокластов человека и активность резорбции кости.
a) Действие добавления моноклонального антитела крысы против мышиного Siglec-15 на клеточное слияние остеокластов из нормальных клеток-предшественников остеокластов человека (окрашивание TRAP)
Нормальные клетки-предшественники остеокластов человека (нормальные природные клетки-предшественники остеокластов человека, приобретенные у Sanko Junyaku Co., Ltd., Cat. № 2T-110) высевали в 96-луночный планшет с концентрацией 1×104 клеток/лунку в соответствии с протоколом, прилагаемым к клеткам. В качестве среды была использована минимальная эссенциальная среда для клеток-предшественников остеокластов (OPBM, приобретенная у Sanko Junyaku Co., Ltd., Cat. № PT-8201), дополненная комплексом дополнительных веществ OPGM (Osteoclast SingleQuotTM Kit, приобретенным у Sanko Junyaku Co., Ltd., Cat. № PT-9501), содержащая эмбриональную сыворотку теленка (конечная концентрация: 10%), RANKL человека (конечная концентрация: 66 нг/мл), M-CSF человека (конечная концентрация: 33 нг/мл) и подобное. К полученному в результате культуральному супернатанту добавляли каждое из моноклональных антител крысы против мышиного Siglec-15, полученных в Примере 24, или контрольный IgG крысы (производство R&D systems) с получением конечной концентрации 30 мкг/мл и клетки культивировали в течение 4 дней в CO2 термостате. После культивирования супернатант удаляли и добавляли 10% нейтральный формалин для фиксации клеток. После фиксации клеток клетки дважды промывали дистиллированной водой, и добавляли раствор для окрашивания TRAP (0,27 мМ нафтол AS-MX фосфат (производство Sigma Co., Ltd.), 1,6 мМ устойчивого красного фиолетового LB соль (производство Sigma Co., Ltd.), 1% диметилформамид, 50 мМ тартрат натрия, 0,1 M ацетат-натриевый буфер (pH 5,0)) в концентрации 100 мкл/лунку, и реакцию оставляли протекать при комнатной температуре в течение 5 минут. Затем клетки дважды промывали дистиллированной водой, а затем наблюдали под микроскопом (фиг. 34). В результате образование гигантских остеокластов, происходящее в результате высокой степени клеточного слияния, было почти полностью ингибировано добавлением антитела #32A1. Далее, также в случае антитела #41B1, образование гигантских остеокластов, происходящее в результате высокой степени клеточного слияния, было существенно ингибировано. С другой стороны, в случае других моноклональных антител крысы против мышиного Siglec-15 (например, антитела #1A1) и контрольного IgG крысы такого существенного ингибирования клеточного слияния остеокластов не наблюдалось. Аналогичным образом, было обнаружено, что многоядерность и клеточное слияние TRAP-позитивных остеокластов из нормальных клеток-предшественников остеокластов человека ингибируется моноклональным антителом, связывающимся с белком Siglec-15.
b) Действие добавления моноклонального антитела крысы против мышиного Siglec-15 (#32A1) на клеточное слияние остеокластов из нормальных клеток-предшественников остеокластов человека (окрашивание TRAP)
Нормальные клетки-предшественники остеокластов человека (нормальные природные клетки-предшественники остеокластов человека, приобретенные у Sanko Junyaku Co., Ltd., Cat. № 2T-110) высевали в 96-луночный планшет с концентрацией 1×104 клеток/лунку в соответствии с протоколом, прилагаемым к этим клеткам. В качестве среды использовали минимальную эссенциальную среду для клеток-предшественников остеокластов (OPBM, приобретенную у Sanko Junyaku Co., Ltd., Cat. № PT-8201), дополненную комплексом дополнительных веществ OPGM (Osteoclast SingleQuotTM Kit, приобретенным у Sanko Junyaku Co., Ltd., Cat. № PT-9501), содержащую эмбриональную сыворотку теленка (конечная концентрация: 10%), RANKL человека (конечная концентрация: 68,4 нг/мл), M-CSF человека (конечная концентрация: 33 нг/мл) и подобное. К полученному в результате культуральному супернатанту добавляли моноклональное антитело крысы против мышиного Siglec-15 (#32A1), полученное в Примере 24, с получением конечной концентрации 0,1, 0,3, 1 или 3 мкг/мл и клетки культивировали в течение 3 дней в CO2 термостате. После культивирования супернатант удаляли и добавляли 10% нейтральный формалин для фиксации клеток. После фиксации клеток клетки дважды промывали дистиллированной водой, и добавляли раствор для окрашивания TRAP (0,27 мМ нафтол AS-MX фосфат (производство Sigma Co., Ltd.), 1,6 мМ устойчивого красного фиолетового LB соль (производство Sigma Co., Ltd.), 1% диметилформамид, 50 мМ тартрат натрия, 0,1 M ацетат-натриевый буфер (pH 5,0)) в концентрации 100 мкл/лунку, и реакцию оставляли протекать при комнатной температуре в течение 5 минут. Затем клетки дважды промывали дистиллированной водой, а затем наблюдали под микроскопом (фиг. 35). В результате образование TRAP-позитивных многоядерных остеокластов было ингибировано в зависимости от концентрации антитела #32A1 в интервале от 0,3 мкг/мл до 3 мкг/мл.
c) Действие добавления моноклонального антитела крысы против мышиного Siglec-15 (#32A1) на активность резорбции кости нормальных клеток-предшественников остеокластов человека (оценка с использованием планшетов, покрытых коллагеном)
Известно, что остеокласты высвобождают протеазу, такую как катепсин К, и разрушают коллаген I типа, который является конституциональным компонентом костной ткани. Набор «OsteoLyse Assay Kit» (производство Lonza, Inc., Cat. № PA-1500) предоставляет 96-луночный планшет, покрытый коллагеном человека, конъюгированным с европием (96-луночный OsteoLyse планшет для культивирования клеток), и возможно оценить активность резорбции кости остеокластами in vitro путем измерения количества флуоресцентных фрагментов коллагена, высвобожденных в супернатант при культивировании остеокластов в этом планшете.
Нормальные клетки-предшественники остеокластов человека (нормальные природные клетки-предшественники остеокластов человека, приобретенные у Sanko Junyaku Co., Ltd., Cat. № 2T-110) высевали в 96-луночный OsteoLyse планшет для культивирования клеток с концентрацией 1×104 клеток/лунку в соответствии с протоколом, прилагаемым к этим клеткам. В качестве среды использовали минимальную эссенциальную среду для клеток-предшественников остеокластов (OPBM, приобретенную у Sanko Junyaku Co., Ltd., Cat. № PT-8201), дополненную комплексом дополнительных веществ OPGM (Osteoclast SingleQuotTM Kit, приобретенным у Sanko Junyaku Co., Ltd., Cat. № PT-9501), содержащую эмбриональную сыворотку теленка (конечная концентрация: 10%), RANKL человека (конечная концентрация: 68,4 нг/мл), M-CSF человека (конечная концентрация: 33 нг/мл) и подобное. К полученному в результате культуральному супернатанту добавляли моноклональное антитело крысы против мышиного Siglec-15 (антитело #32A1), полученное в Примере 24, с получением конечной концентрации 0,1, 0,3, 1 или 3 мкг/мл, и клетки культивировали в течение 3 дней в CO2 термостате. Получали аликвоту 10 мкл культурального супернатанта, и добавляли 200 мкл реагента, высвобождающего флуорофор, включенного в набор «OsteoLyse Assay Kit», и измеряли интенсивность флуоресценции (Возбуждение: 340 нм, Эмиссия: 615 нм), используя флуоресцентный планшетный ридер (ARVO MX, производство Perkin Elmer Inc.), посредством чего определяли количество свободных флуоресцентных фрагментов коллагена, высвобожденных в культуральный супернатант (фиг. 36). В результате количество флуоресцентных фрагментов коллагена, увеличенное посредством добавления RANKL, уменьшалось под действием антитела #32A1 зависимым от концентрации образом в интервале от 0,3 мкг/мл до 3 мкг/мл. Исходя из этого результата, было обнаружено, что активность остеокластов человека в отношении резорбции кости ингибируется моноклональным антителом, специфически связывающимся с белком Siglec-15.
Пример 38. Получение моноклонального антитела мыши против Siglec-15 человека
Можно получить антитело против Siglec-15 человека с помощью описанных ниже стадий.
(1) Иммунизация
Растворимый белок Siglec-15 человека, полученный в Примерах 30-32, вводят внутрибрюшинно самкам мышей BALB/c в возрасте от 4 до 10 недель. Через 2 недели раствор той же мембранной фракции вводят внутрибрюшинно мышам для повторной иммунизации.
(2) Слияние клеток
Селезенку извлекают у мышей на третий день после повторной иммунизации, помещают в бессывороточную среду и измельчают на сите шпателем. Клеточную суспензию, пропущенную через сито, центрифугируют для осаждения клеток селезенки.
С другой стороны, миеломные клетки NS1 (Американская коллекция типовых культур TIB-18) промывали бессывороточной средой и суспендировали аналогичным образом.
Используя полученные таким образом клетки, экспрессирующие Siglec-15, и миеломные клетки, проводили слияние клеток в соответствии с общепринятой процедурой.
(3) Скрининг
Слитые клетки, продуцирующие антитела против Siglec-15 человека, могут быть подвержены скринингу способом с использованием cell-ELISA и способом с использованием проточного цитометра.
(4) Клонирование
Для группы клеток, подвергнутых скринингу в приведенном выше пункте (3), серии стадий, включающих способ с использованием cell-ELISA и способ с использованием проточного цитометра, описанного в пункте (3), повторяли 5 раз, посредством чего может быть получено несколько гибридомных клонов, способных продуцировать единичное антитело, которое связывается с клетками, экспрессирующими Siglec-15 человека, но не связывается с этими клетками до трансфекции.
(5) Очистка антитела
Культивируют гибридому «мышь-мышь», полученную посредством осуществления стадий (1)-(4), и собирают полученный в результате супернатант. После этого полученный супернатант собирают и диализируют, проводят частичную очистку антитела, используя прибор для высокоэффективной жидкостной хроматографии. Титр антитела против Siglec-15 человека в каждой фракции хроматографии исследуют методом ELISA, используя белок Siglec-15 человека. Фракции, имеющие высокий титр антител, собирают и наносят на аффинную колонку для очистки антител. После промывания внутренней части колонки буфером для уравновешивания колонки антитело элюируют с буфером для элюции с колонки. Сразу после завершения элюции каждый элюат наносят на верхнюю часть центрифужного ультрафильтра и центрифугируют. После этого фильтрат, собранный на дне фильтра, удаляют, промывание проводят 5 раз путем добавления PBS на верхнюю часть. Жидкость, оставшуюся на верхней части фильтра, используют в качестве образца антитела против Siglec-15 человека.
Пример 39. Получение моноклонального антитела крысы против Siglec-15 человека
Моноклональное антитело крысы против Siglec-15 человека может быть получено с использованием растворимого белка Siglec-15 человека, полученного в примерах 30-32, способом, описанным в Примерах 23 и 24.
Пример 40. Действие моноклонального антитела против Siglec-15 человека на дифференцировку остеокластов
Используя моноклональное антитело против Siglec-15 человека, полученное в Примере 38 или 39, может быть протестировано ингибирующее действие этого антитела на образование остеокластов. Для тестирования действия на образование остеокластов мыши можно использовать способ Примера 17, 19, 20, 21, 22 или 26. Для тестирования ингибирующего действия на образование остеокластов человека может быть использован способ Примера 35 или 37.
Промышленная применимость
Антитело против Siglec-15 по настоящему изобретению обладает способностью ингибировать дифференцировку остеокластов или активность резорбции кости, и фармацевтическая композиция, содержащая антитело против Siglec-15, может быть терапевтическим или профилактическим средством для лечения аномального метаболизма костной ткани.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ Siglec-15 | 2010 |
|
RU2539790C2 |
| БЕЛОК, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩИЙСЯ С ИНГИБИРУЮЩИМ ОСТЕОКЛАСТОГЕНЕЗ ФАКТОРОМ (OCIF) (ВАРИАНТЫ), ДНК ЕГО КОДИРУЮЩАЯ (ВАРИАНТЫ), ДНК В КАЧЕСТВЕ ЗОНДА (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ БЕЛКА (ВАРИАНТЫ), СПОСОБ СКРИНИНГА ВЕЩЕСТВА (ВАРИАНТЫ), АНТИТЕЛО (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ПОЛИКЛОНАЛЬНОГО АНТИТЕЛА, ГИБРИДОМА (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ МОНОКЛОНАЛЬНОГО АНТИТЕЛА, СПОСОБ ИЗМЕРЕНИЯ OCIF-СВЯЗЫВАЮЩЕГО БЕЛКА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ) И ЛЕКАРСТВЕННОЕ СРЕДСТВО (ВАРИАНТЫ) | 1998 |
|
RU2238949C2 |
| НОВЫЕ БЕЛКИ И СПОСОБЫ ПОЛУЧЕНИЯ ТАКИХ БЕЛКОВ | 1996 |
|
RU2194714C2 |
| МОЛЕКУЛА АДГЕЗИИ Т-КЛЕТОК И АНТИТЕЛО, НАПРАВЛЕННОЕ ПРОТИВ МОЛЕКУЛЫ | 2006 |
|
RU2396275C2 |
| МОЛЕКУЛЯРНЫЕ МАССИВЫ АНТИГЕНОВ | 2003 |
|
RU2324704C2 |
| НЕ МЫШИНОЕ АНТИ-M-CSF-АНТИТЕЛО (ВАРИАНТЫ), ЕГО ПОЛУЧЕНИЕ И ИСПОЛЬЗОВАНИЕ | 2005 |
|
RU2401277C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ РАКА | 2013 |
|
RU2631804C2 |
| АНТИГЕННЫЕ МАТРИЦЫ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ КОСТЕЙ | 2002 |
|
RU2322258C2 |
| КОМПОЗИЦИЯ И СПОСОБ ДЛЯ ИММУНИЗАЦИИ, СПОСОБ ПРОДУЦИРОВАНИЯ НЕПРИРОДНОГО УПОРЯДОЧЕННОГО И ПОВТОРЯЮЩЕГОСЯ МАССИВА АНТИГЕНОВ | 2002 |
|
RU2438701C2 |
| КОМПОЗИЦИЯ И СПОСОБ ДЛЯ ИММУНИЗАЦИИ, СПОСОБ ПРОДУЦИРОВАНИЯ НЕПРИРОДНОГО, УПОРЯДОЧЕННОГО И ПОВТОРЯЮЩЕГОСЯ МАССИВА АНТИГЕНОВ И ОБОЛОЧЕЧНЫЙ БЕЛОК | 2002 |
|
RU2295973C2 |

























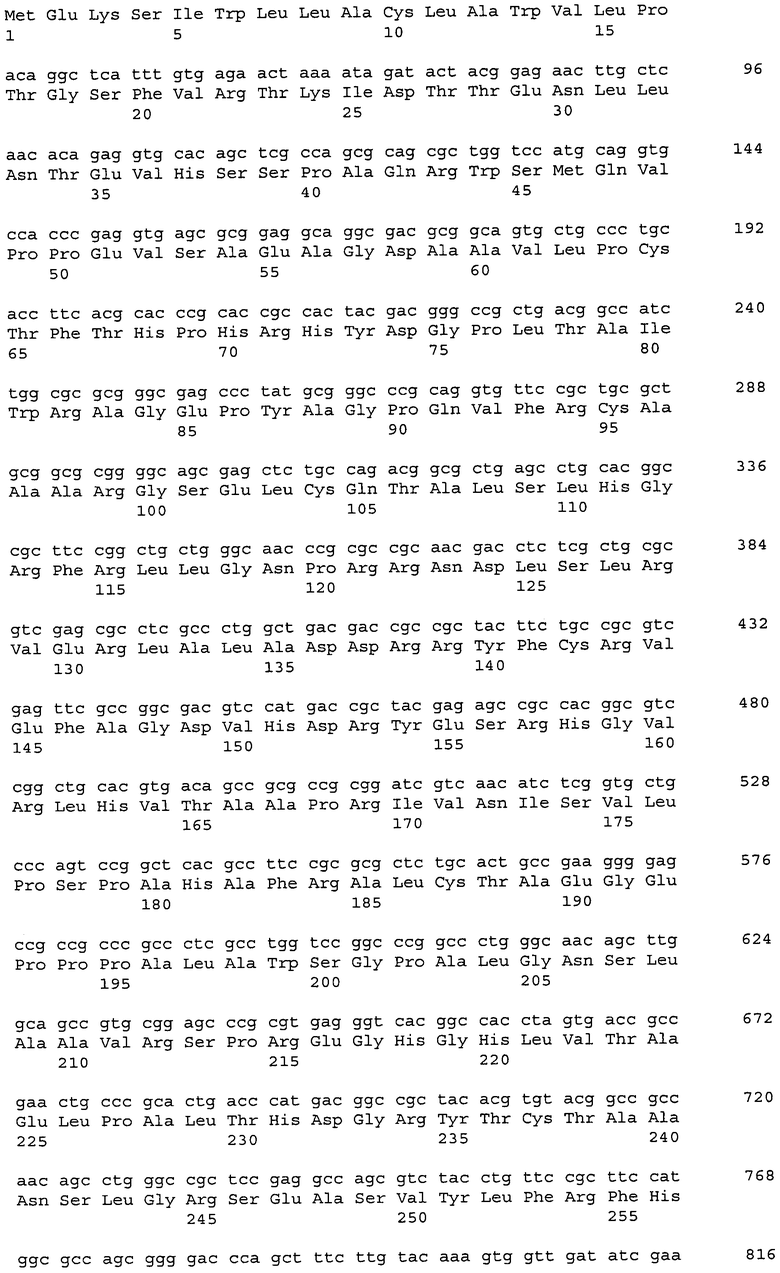






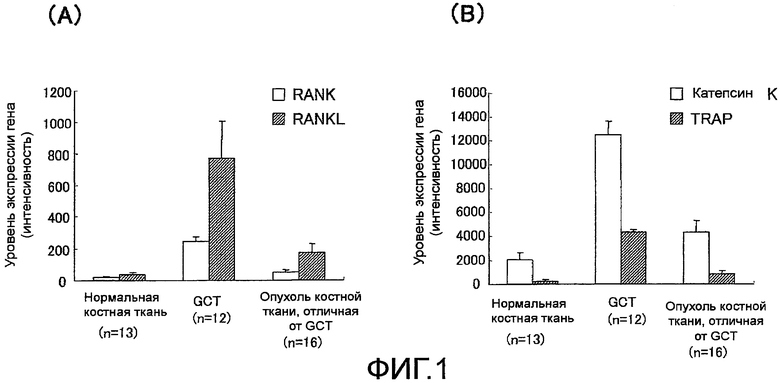
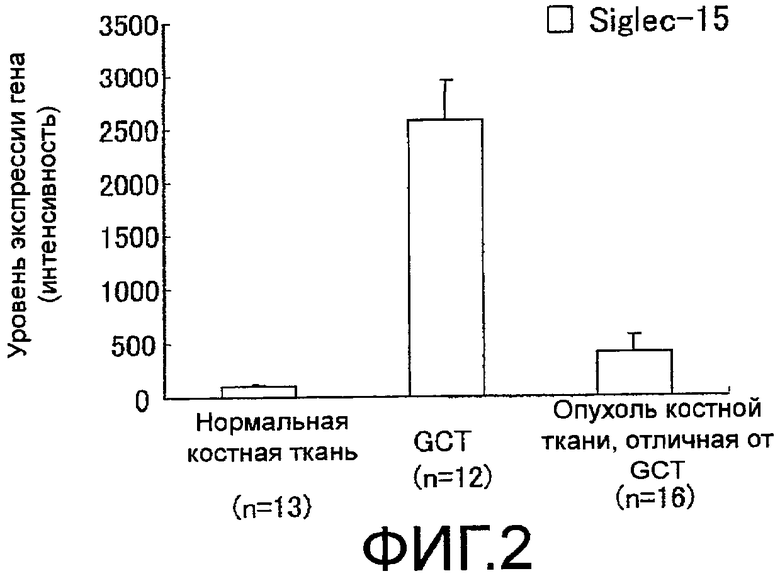
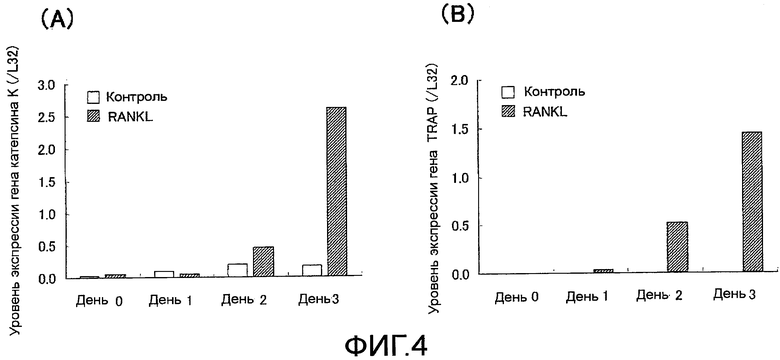
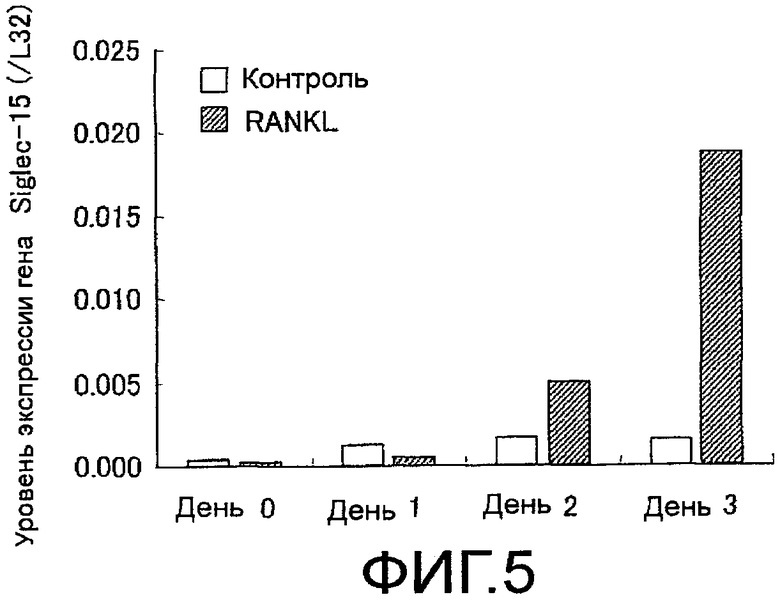
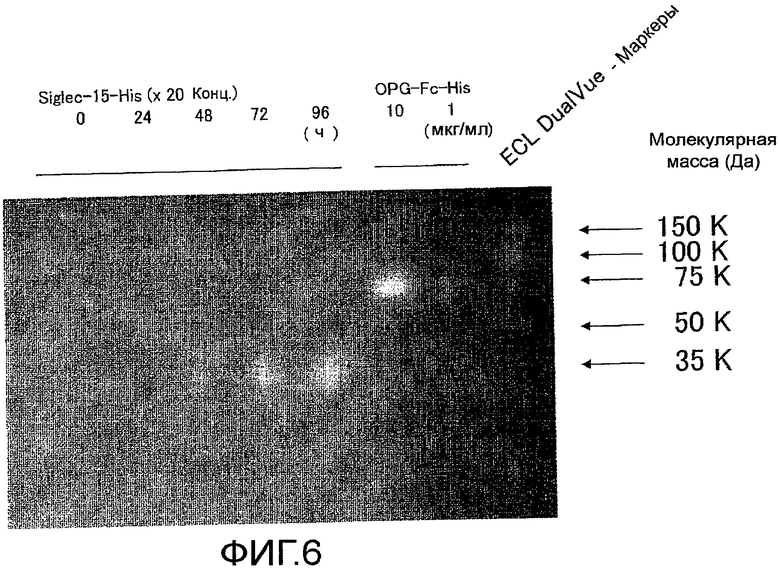
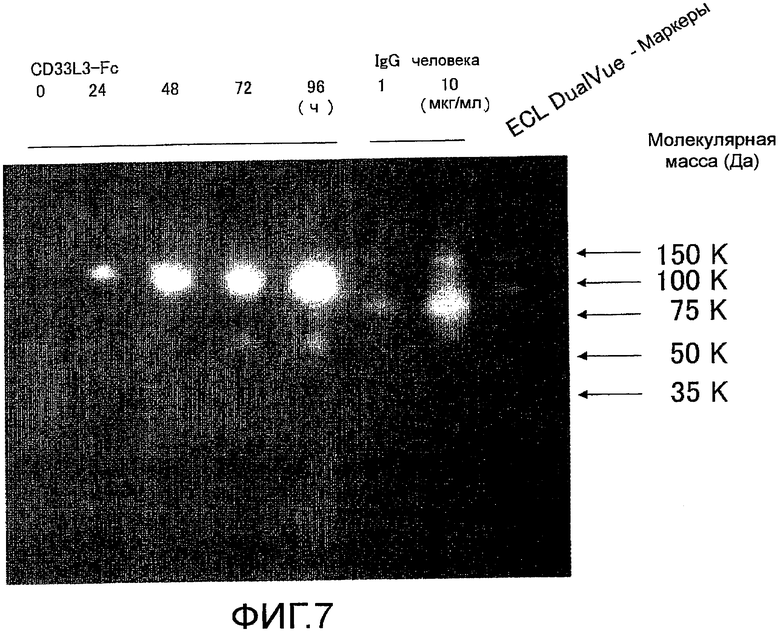
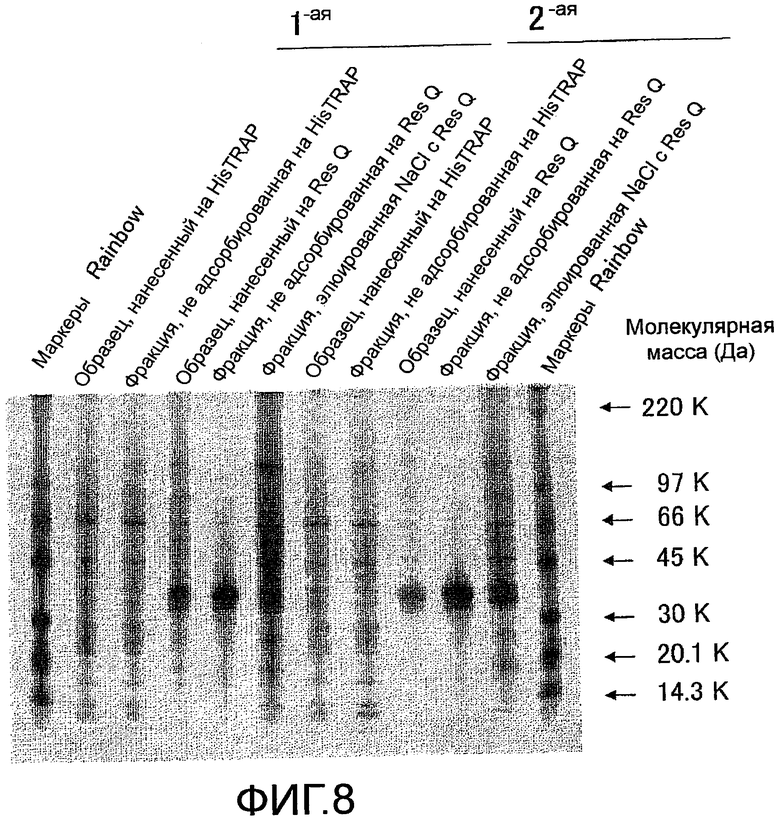
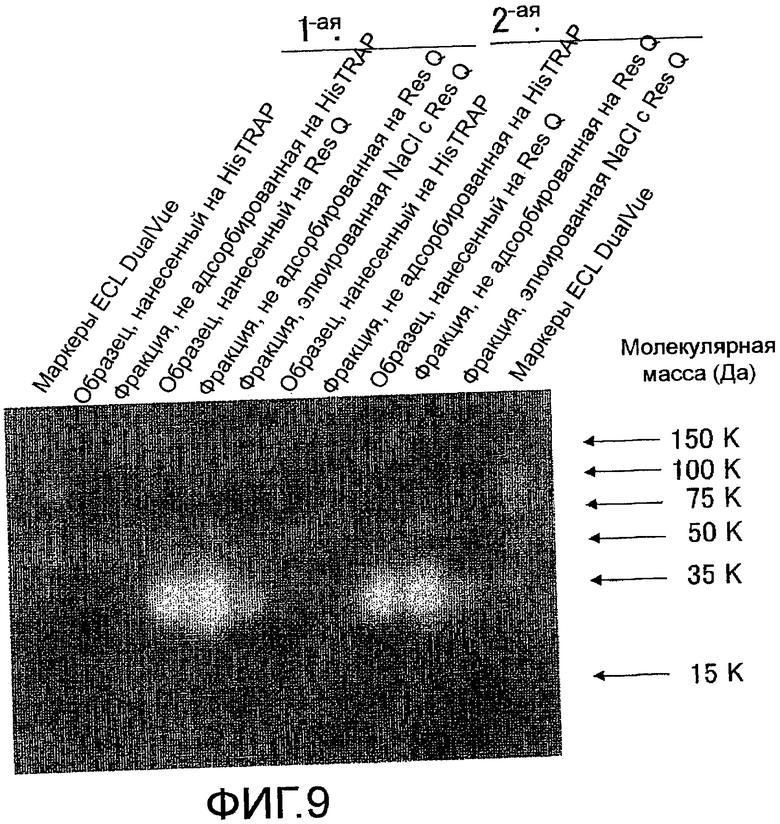
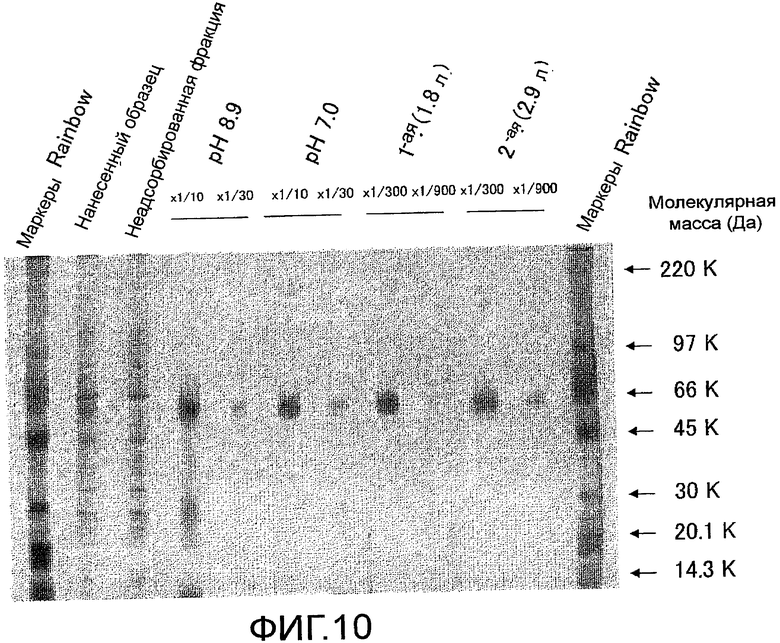
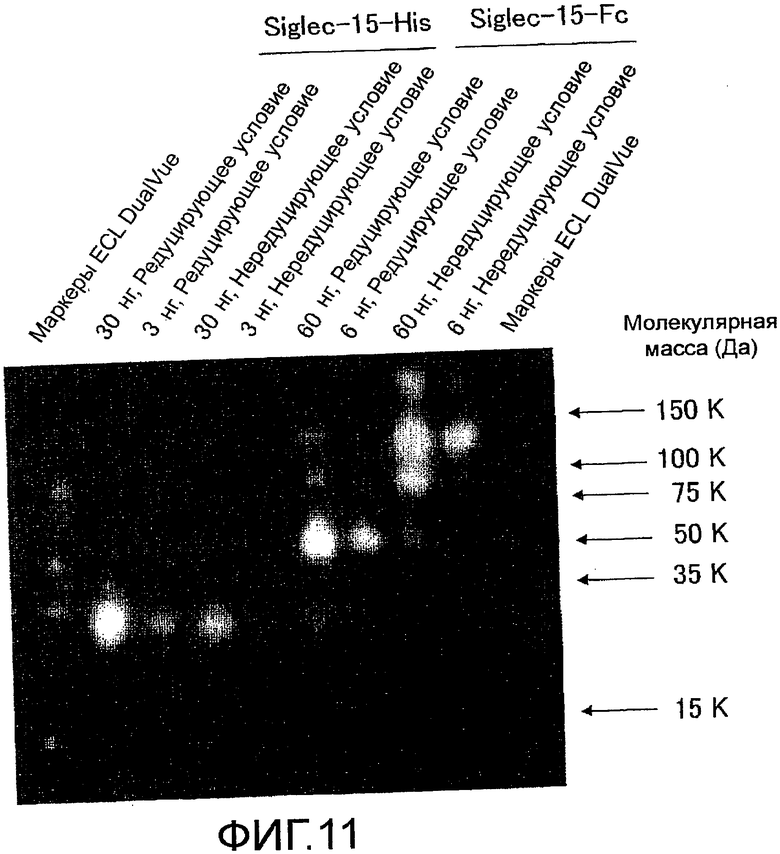
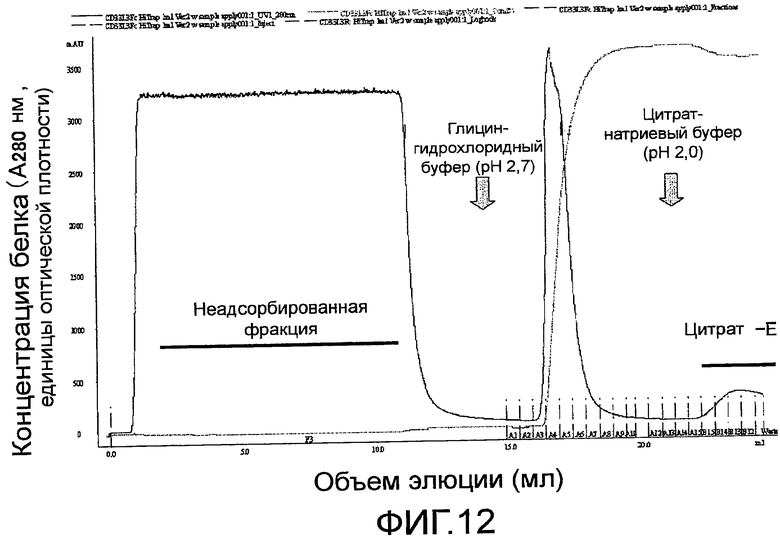
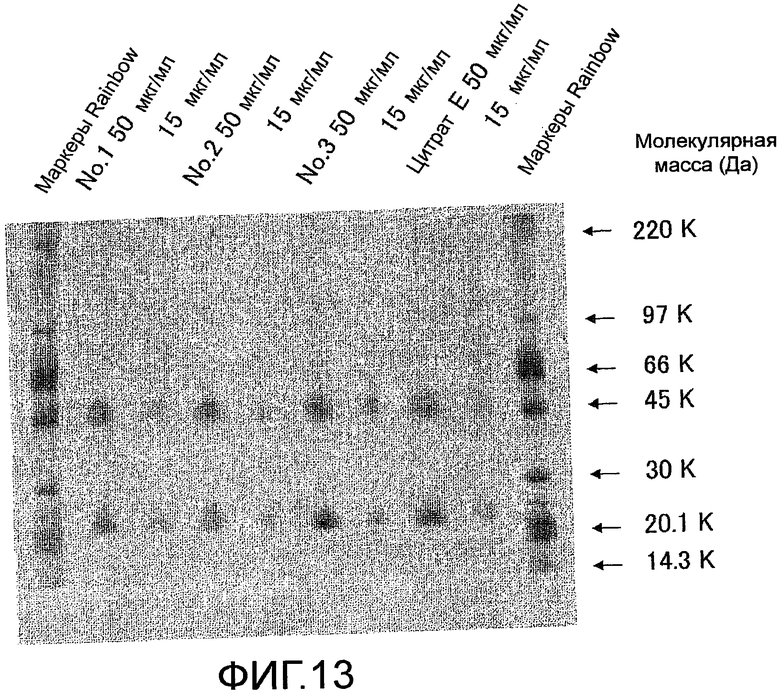
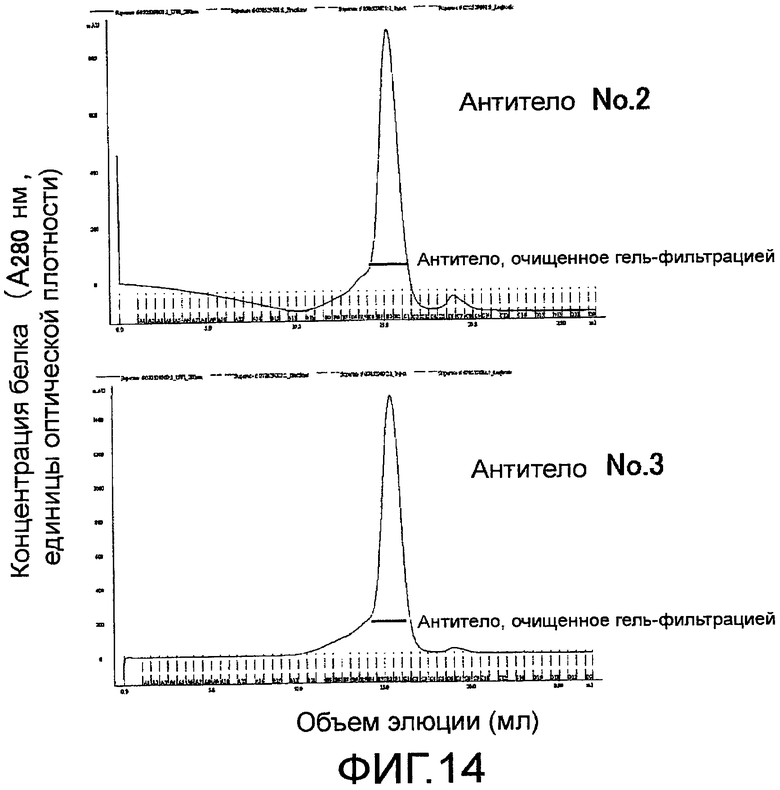
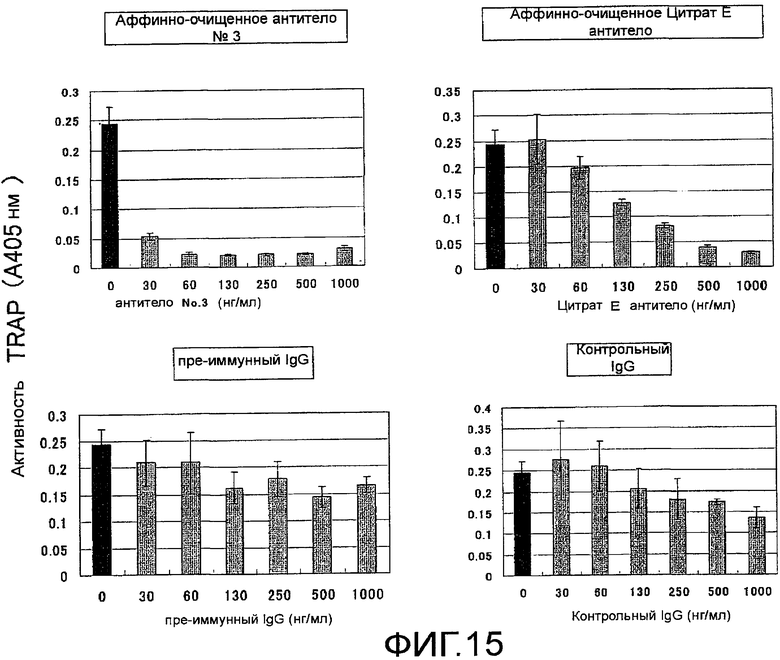
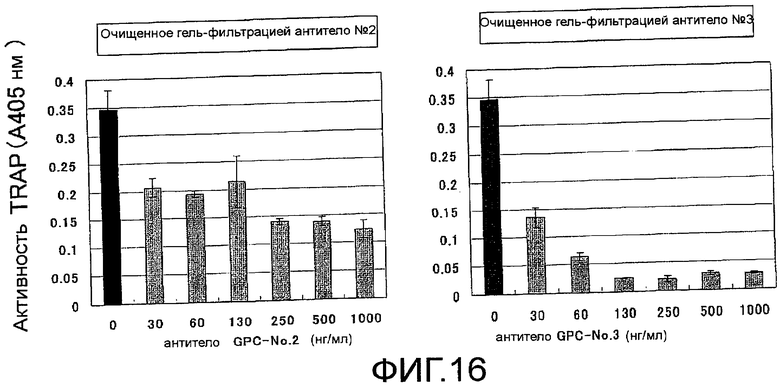
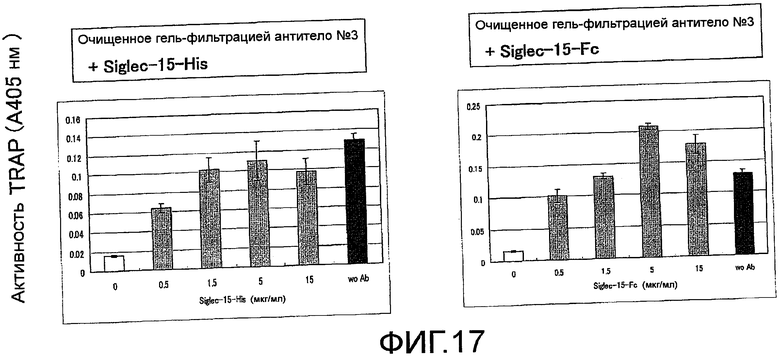
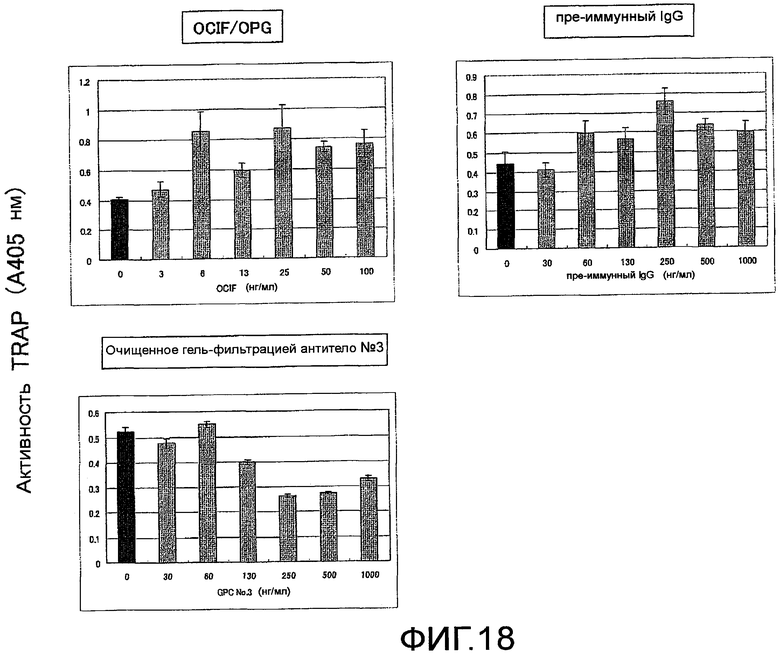
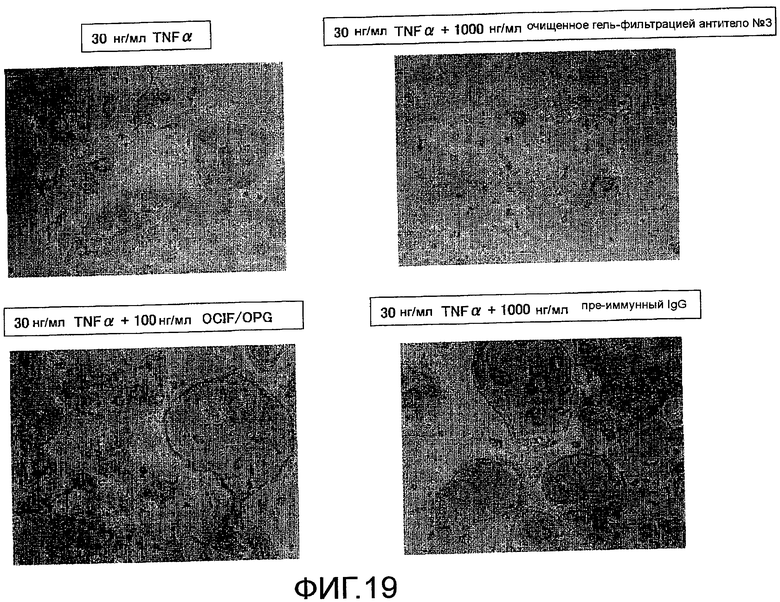
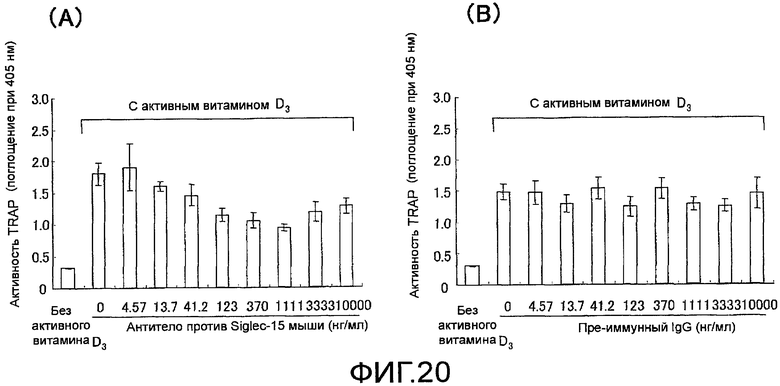
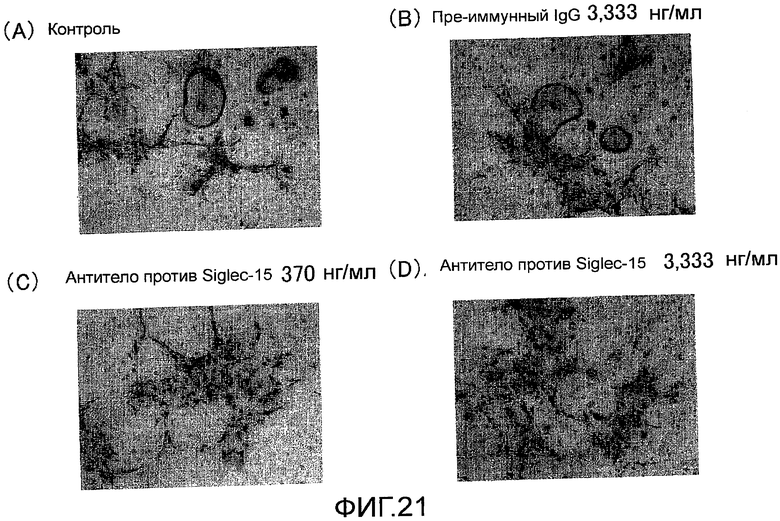


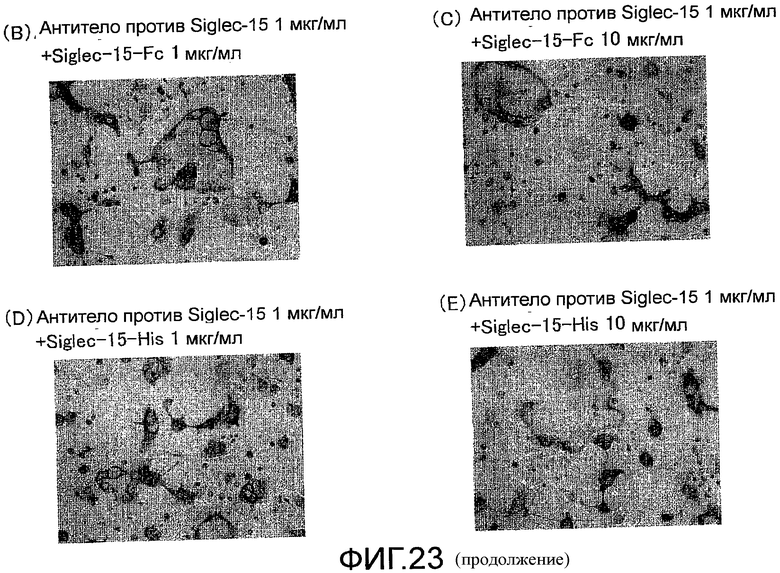
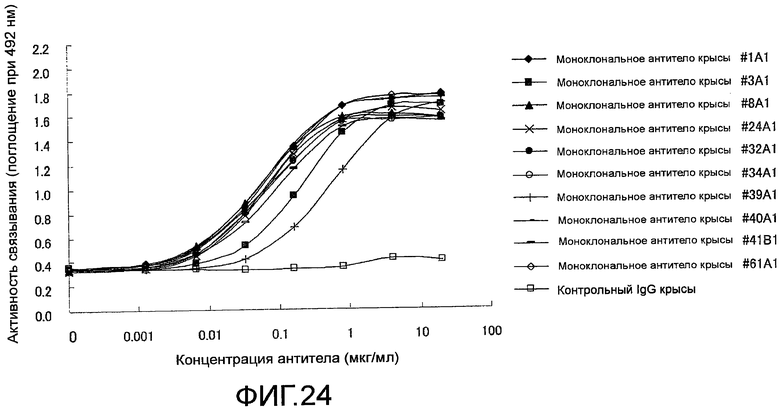
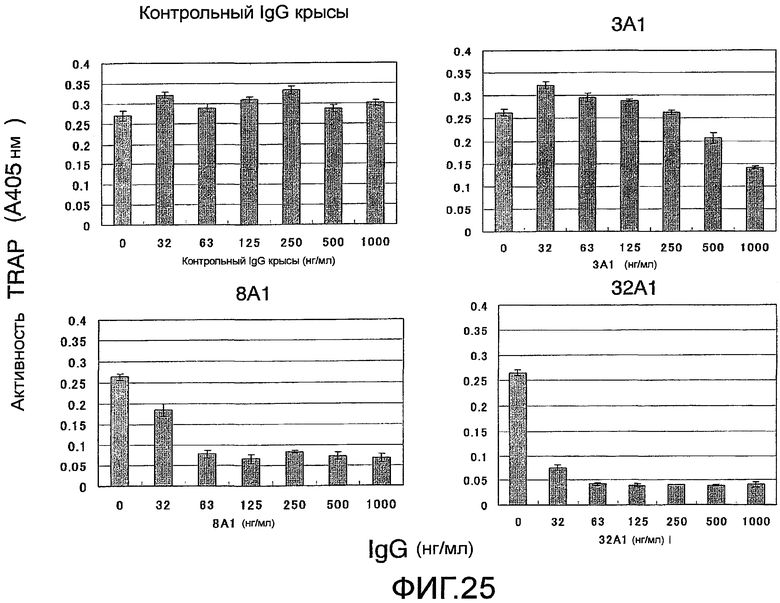
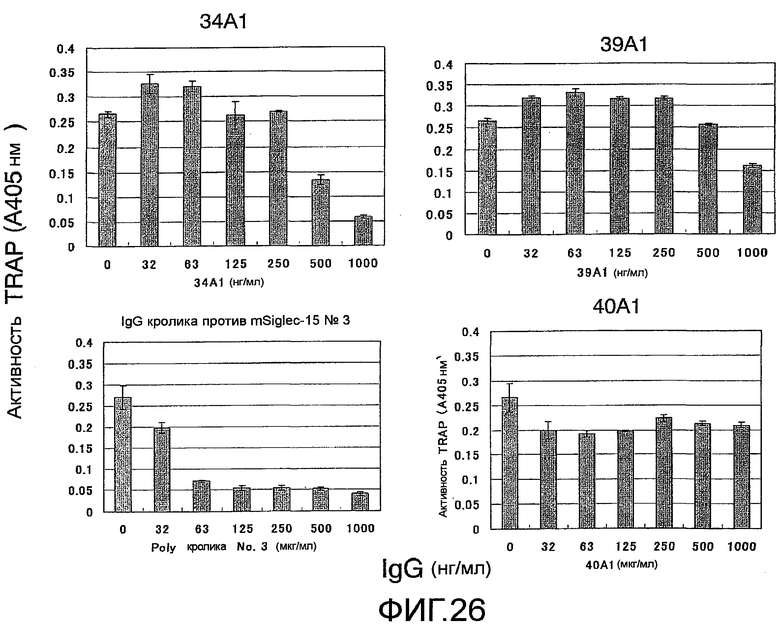

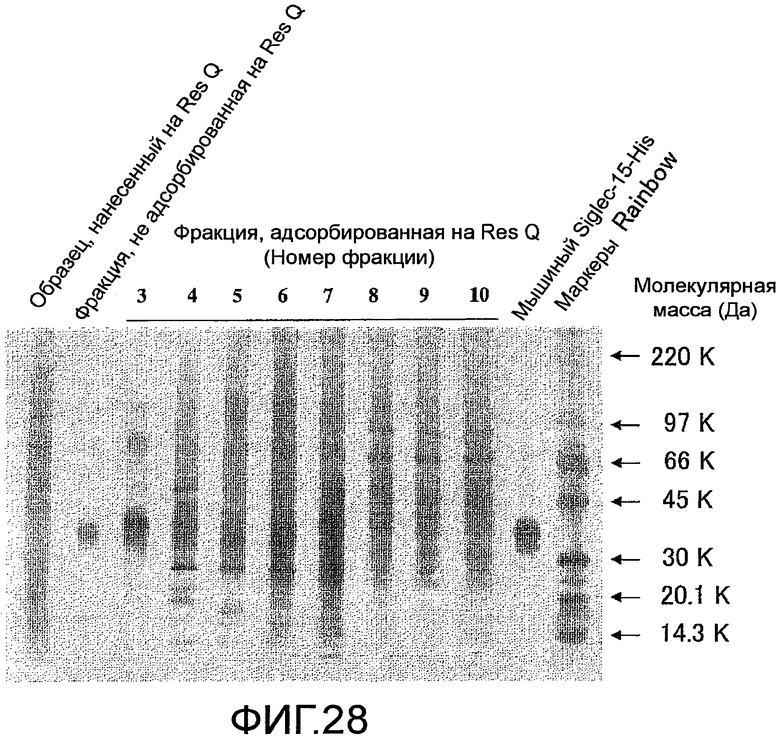
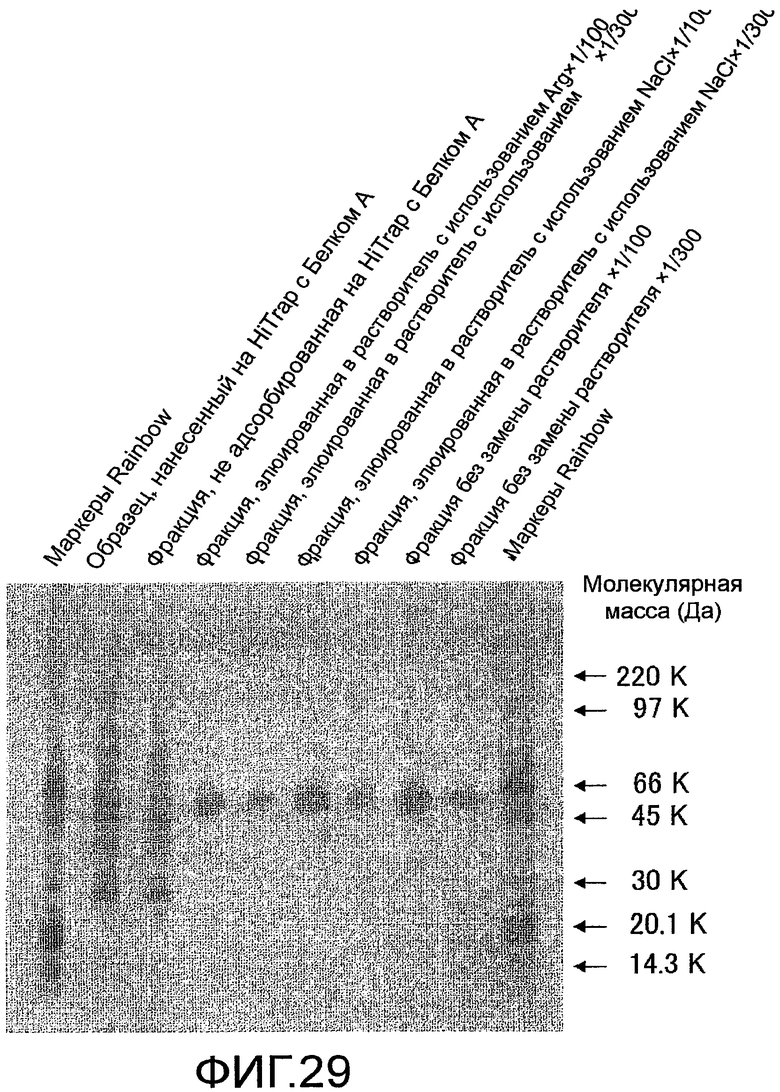
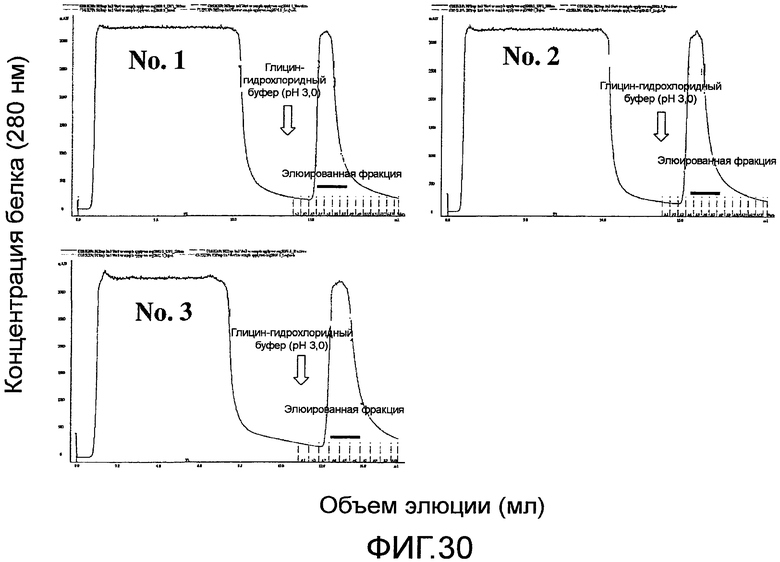
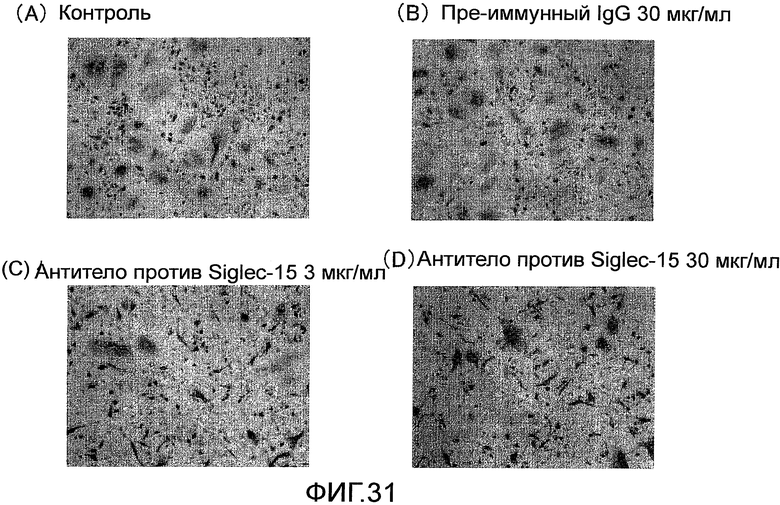
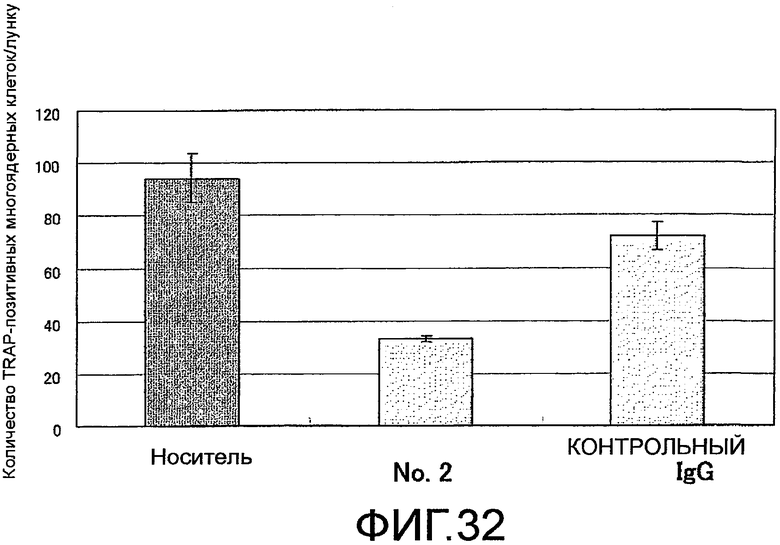
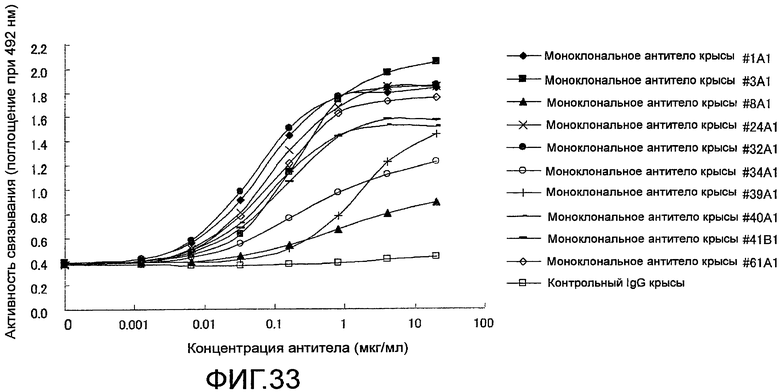
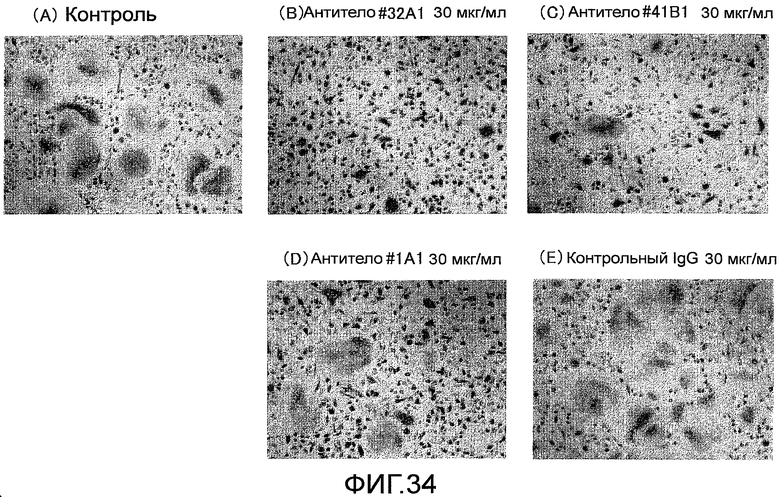
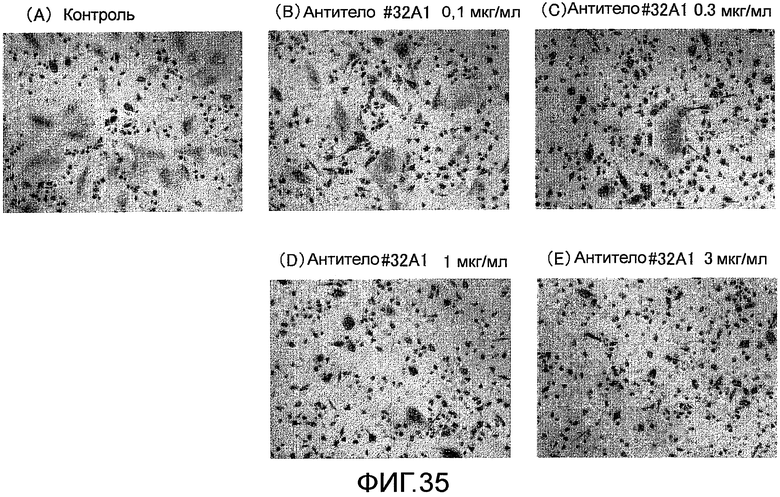
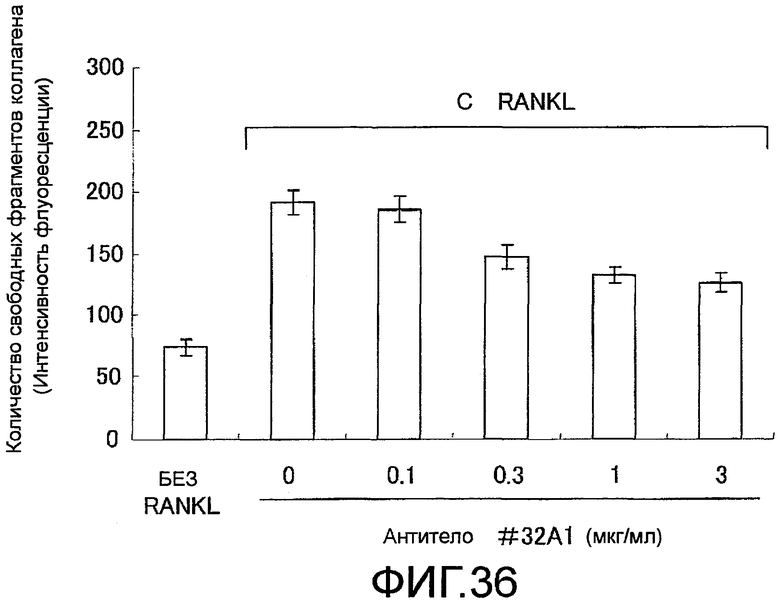
Изобретение раскрывает антитело или его функциональный фрагмент, специфически распознающие полипептид, кодируемый геном в организме человека, и ингибирующие образование остеокластов и, необязательно, остеокластную резорбцию костей. Антитело или функциональный фрагмент можно получить, в частности, гибридомной технологией с использованием гибридом #32A1 (FERM ВР-10999) и #41В1 (FERM ВР-11000), которые раскрыты в изобретении. Описана фармацевтическая композиция на основе антитела или его функционального фрагмента для лечения и/или профилактики патологического метаболизма кости у пациента, а также способы лечения и профилактики аномального метаболизма костной ткани с использованием антител или его функциональных фрагментов. Использование изобретения позволяет с помощью антитела по изобретению специфически распознавать Siglec-15 человека, экспрессируемый геном, и ингибировать образование остеокластов. 9 н. и 30 з.п. ф-лы, 36 ил., 9 табл., 38 пр.
1. Антитело, которое специфически распознает по меньшей мере один полипептид, состоящий из аминокислотной последовательности, описанной в любом из следующих пп.(a)-(i), и ингибирует образование остеокластов и, необязательно, остеокластную резорбцию кости, или функциональный фрагмент этого антитела:
(a) аминокислотную последовательность, представленную последовательностью SEQ ID NO:2 в перечне последовательностей;
(b) аминокислотную последовательность, состоящую из аминокислотных остатков 21-328 аминокислотной последовательности SEQ ID NO:2 в перечне последовательностей;
(c) аминокислотную последовательность, состоящую из аминокислотных остатков 1-260 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(d) аминокислотную последовательность, состоящую из аминокислотных остатков 21-260 аминокислотной последовательности, представленной SEQ ID NO:2 в перечне последовательностей;
(e) аминокислотную последовательность, представленную последовательностью SEQ ID NO:4 в перечне последовательностей;
(f) аминокислотную последовательность, состоящую из аминокислотных остатков 21-341 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей;
(g) аминокислотную последовательность, состоящую из аминокислотных остатков 1-258 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей;
(h) аминокислотную последовательность, состоящую из аминокислотных остатков 21-258 аминокислотной последовательности SEQ ID NO:4 в перечне последовательностей; и
(i) аминокислотную последовательность, включающую замены, делеции или добавления одного аминокислотного остатка в аминокислотной последовательности, описанной в пп.(a)-(h),
где антитело ингибирует in vitro образование TRAP-позитивных многоядерных остеокластов в концентрации 30 мкг/мл или менее.
2. Антитело, которое специфически распознает по меньшей мере один полипептид, состоящий из аминокислотной последовательности, кодируемой нуклеотидной последовательностью, описанной в любом из следующих пп.(j)-(n), и ингибирует образование остеокластов и, необязательно, остеокластную резорбцию кости, или функциональный фрагмент этого антитела:
(j) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:1;
(k) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:3;
(1) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:19;
(m) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:43; и
(n) нуклеотидной последовательностью полинуклеотида, который гибридизируется с полинуклеотидом, содержащим нуклеотидную последовательность, комплементарную нуклеотидной последовательности, описанной в пп.(j)-(m) в строгих условиях,
где антитело ингибирует in vitro образование TRAP-позитивных многоядерных остеокластов в концентрации 30 мкг/мл или менее.
3. Антитело или функциональный фрагмент этого антитела по п.1 или 2, которое ингибирует процесс клеточного слияния остеокластов, индуцированное под действием TNF-α.
4. Антитело или функциональный фрагмент этого антитела по п.1, которое ингибирует образование остеокластов, in vitro в концентрации 3 мкг/мл или менее.
5. Антитело или функциональный фрагмент этого антитела по п.4, которое ингибирует образование остеокластов in vitro в концентрации 1 мкг/мл или менее.
6. Антитело или функциональный фрагмент этого антитела по п.5, которое ингибирует образование остеокластов in vitro в концентрации 63 нг/мл до 1 мкг/мл.
7. Антитело или функциональный фрагмент этого антитела по п.1 или 2, которое ингибирует остеокластную резорбцию кости.
8. Антитело или функциональный фрагмент этого антитела по п.7, которое ингибирует остеокластную резорбцию костей in vitro в концентрации 3 мкг/мл или менее.
9. Антитело или функциональный фрагмент этого антитела по п.8, которое ингибирует остеокластную резорбцию кости in vitro в концентрации от 0,3 мкг/мл до 3 мкг/мл.
10. Антитело или функциональный фрагмент этого антитела по п.1 или 2, которое получено способом, включающим следующие стадии 1) и 2):
1) стадию получения антитела, которое специфически распознает по меньшей мере один полипептид, состоящий из аминокислотной последовательности, описанной в любом из следующих пп.(a)-(i):
(а) аминокислотную последовательность, представленную последовательностью SEQ ID NO:2 в перечне последовательностей;
(b) аминокислотную последовательность, состоящую из аминокислотных остатков 21-328 аминокислотной последовательности SEQ ID NO:2 в перечне последовательностей;
(c) аминокислотную последовательность, состоящую из аминокислотных остатков 1-260 аминокислотной последовательности SEQ ID NO:2 в перечне последовательностей;
(d) аминокислотную последовательность, состоящую из аминокислотных остатков 21-260 аминокислотной последовательности SEQ ID NO:2 в перечне последовательностей;
(e) аминокислотную последовательность, представленную последовательностью SEQ ID NO:4 в перечне последовательностей;
(f) аминокислотную последовательность, состоящую из аминокислотных остатков 21-341 аминокислотной последовательности SEQ ID NO:4 в перечне последовательностей;
(g) аминокислотную последовательность, состоящую из аминокислотных остатков 1-258 аминокислотной последовательности, представленной SEQ ID NO:4 в перечне последовательностей;
(h) аминокислотную последовательность, состоящую из аминокислотных остатков 21-258 аминокислотной последовательности SEQ ID NO:4 в перечне последовательностей; и
(i) аминокислотную последовательность, включающую замены, делеции или добавления одного аминокислотного остатка в аминокислотной последовательности, описанной в пп.(a)-(h); и
2) стадию скрининга антитела, которое демонстрирует ингибирующую активность in vitro в отношении образования TRAP-позитивных многоядерных остеокластов в концентрации 30 мкг/мл или менее.
11. Антитело или функциональный фрагмент этого антитела по п.1 или 2, которое получено способом, включающим следующие стадии 1) и 2):
1) стадию получения антитела, которое специфически распознает по меньшей мере один или полипептид, состоящий из аминокислотной последовательности, кодируемой нуклеотидной последовательностью, описанной в любом из следующих пп.(j)-(n):
(j) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:1;
(k) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:3;
(l) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:19;
(m) нуклеотидной последовательностью, представленной последовательностью SEQ ID NO:43; и
(n) нуклеотидной последовательностью полинуклеотида, который гибридизируется с полинуклеотидом, содержащим нуклеотидную последовательность, комплементарную нуклеотидной последовательности, описанной в пп.(j)-(m) в строгих условиях; и
2) стадию скрининга антитела, которое демонстрирует ингибиторную активность in vitro в отношении образования TRAP-позитивных многоядерных остеокластов в концентрации 30 мкг/мл или менее.
12. Антитело или функциональный фрагмент этого антитела по п.1 или 2, отличающееся тем, что это антитело представляет собой моноклональное антитело.
13. Антитело или функциональный фрагмент этого антитела по п.12, отличающееся тем, что имеет ту же эпитопную специфичность, что и антитело, продуцируемое гибридомой #32А1 (FERM BP-10999).
14. Антитело или функциональный фрагмент этого антитела по п.12, отличающееся тем, что конкурирует с антителом, продуцируемым гибридомой #32A1 (FERM BP-10999).
15. Антитело или функциональный фрагмент этого антитела по п.12, отличающееся тем, что это антитело представляет собой антитело, продуцируемое гибридомой #32А1 (FERM BP-10999).
16. Антитело или функциональный фрагмент этого антитела по п.12, отличающееся тем, что имеет ту же эпитопную специфичность, что и антитело, продуцируемое гибридомой #41В1 (FERM ВР-11000).
17. Антитело или функциональный фрагмент этого антитела по п.12, отличающееся тем, что конкурирует с антителом, продуцируемым гибридомой #41В1 (FERM BP-11000).
18. Антитело или функциональный фрагмент этого антитела по п.12, отличающееся тем, что это антитело представляет собой антитело, продуцируемое гибридомой #41В1 (FERM BP-11000).
19. Антитело или функциональный фрагмент этого антитела по п.1 или 2, отличающееся тем, что это антитело представляет собой химерное антитело.
20. Антитело или функциональный фрагмент этого антитела по п.1 или 2, отличающееся тем, что это антитело является гуманизированным.
21. Антитело или функциональный фрагмент этого антитела по п.1 или 2, отличающееся тем, что это антитело представляет собой антитело человека.
22. Антитело или функциональный фрагмент этого антитела по п.1 или 2, отличающееся тем, что это антитело представляет собой IgG антитело.
23. Функциональный фрагмент антитела по п.1 или 2, который выбран из группы, состоящей из Fab, F(ab')2, Fab' и Fv.
24. Антитело по п.1 или 2, отличающееся тем, что представляет собой scFv.
25. Фармацевтическая композиция для лечения и/или профилактики патологического метаболизма кости у пациента, отличающаяся тем, что содержит по меньшей мере одно из антител или функциональных фрагментов антител по п.1 или 2 в количестве от 0,1 до 100 мг/кг.
26. Фармацевтическая композиция по п.25, отличающаяся тем, что представляет собой терапевтическое и/или профилактическое средство для лечения аномального метаболизма костной ткани.
27. Фармацевтическая композиция для лечения и/или профилактики аномального метаболизма костной ткани у пациента, отличающаяся тем, что содержит по меньшей мере одно из антител или функциональных фрагментов таких антител по п.1 или 2 в количестве от 0,1 до 10 мг/кг и по меньшей мере один компонент, выбранный из группы, состоящей из бисфосфонатов, активного витамина D3, кальцитонина и его производных, гормональных препаратов, таких как эстрадиол, SERM (селективных модуляторов рецепторов эстрогена), иприфлавона, витамина K2 (менатетренона), препаратов кальция, препаратов РТН (паратиреоидного гормона), нестероидных противовоспалительных средств, препаратов растворимого рецептора TNF, анти-TNF-α антител или функциональных фрагментов таких антител, анти-PTHrP (белка, родственного паратиреоидному гормону) антител или функциональных фрагментов таких антител, антагонистов рецептора IL-1, антител против рецептора IL-6 или функциональных фрагментов таких антител, антител против RANKL или функциональных фрагментов таких антител и OCIF (фактора, ингибирующего остеокластогенез).
28. Фармацевтическая композиция по п.26, где аномальный метаболизм костной ткани выбран из группы, состоящей из остеопороза, деструкции костной ткани, сопровождающей ревматоидный артрит, злокачественной гиперкальциемии, деструкции костной ткани, сопровождающей множественную миелому или злокачественное метастазирование в костную ткань, гигантоклеточной остеобластомы, потери зубов вследствие пародонтита, остеолизиса вокруг протезированного сустава, деструкции кости при хроническом остеомиелите, деформирующей остеодистрофии (болезнь Педжета), почечной остеодистрофии и несовершенного остеогенеза.
29. Фармацевтическая композиция по п.28, отличающаяся тем, что аномальный метаболизм костной ткани представляет собой остеопороз, деструкцию костной ткани, сопровождающую ревматоидный артрит или деструкцию костной ткани, сопровождающую злокачественное метастазирование в костную ткань.
30. Фармацевтическая композиция по п.29, отличающаяся тем, что остеопороз представляет собой постменопаузный остеопороз, сенильный остеопороз, вторичный остеопороз вследствие применения таких терапевтических средств как стероиды или иммуносуппрессанты, или остеопороз, сопровождающий ревматоидный артрит.
31. Способ лечения и/или профилактики аномального метаболизма костной ткани, отличающийся тем, что проводят введение по меньшей мере одного из антител или функциональных фрагментов таких антител по п.1 или 2.
32. Применение по меньшей мере одного из антител или функциональных фрагментов таких антител по п.1 или 2 для производства лекарственного средства для лечения и/или профилактики аномального метаболизма костной ткани.
33. Способ лечения и/или профилактики аномального метаболизма костной ткани, отличающийся тем, что проводят одновременное или последовательное введение по меньшей мере одного из антител или функциональных фрагментов таких антител по п.1 или 2, и по меньшей мере одного компонента, выбранного из группы, состоящей из бисфосфонатов, активного витамина D3, кальцитонина и его производных, гормональных препаратов, таких как эстрадиол, SERM (селективных модуляторов рецепторов эстрогена), иприфлавона, витамина K2 (менатетренона), препаратов кальция, препаратов РТН (паратиреоидного гормона), нестероидных противовоспалительных средств, препаратов растворимого рецептора TNF, анти-TNF-α антител или функциональных фрагментов таких антител, анти-PTHrP (белка, родственного паратиреоидному гормону) антител или функциональных фрагментов таких антител, антагонистов рецептора IL-1, антител против рецептора IL-6 или функциональных фрагментов таких антител, антител против RANKL или функциональных фрагментов таких антител и OCIF (фактора, ингибирующего остеокластогенез).
34. Способ лечения и/или профилактики по п.31, отличающийся тем, что аномальный метаболизм костной ткани представляет собой остеопороз, деструкцию костной ткани, сопровождающую ревматоидный артрит или деструкцию костной ткани, сопровождающую злокачественное метастазирование в костную ткань.
35. Способ лечения и/или профилактики по п.34, отличающийся тем, что остеопороз представляет собой постменопаузный остеопороз, сенильный остеопороз, вторичный остеопороз вследствие применения таких терапевтических средств как стероиды или иммуносуппрессанты, или остеопороз, сопровождающий ревматоидный артрит.
36. Способ лечения и/или профилактики по п.33, отличающийся тем, что аномальный метаболизм костной ткани представляет собой остеопороз, деструкцию костной ткани, сопровождающую ревматоидный артрит или деструкцию костной ткани, сопровождающую злокачественное метастазирование в костную ткань.
37. Способ лечения и/или профилактики по п.36, отличающийся тем, что остеопороз представляет собой постменопаузный остеопороз, сенильный остеопороз, вторичный остеопороз вследствие применения таких терапевтических средств как стероиды или иммуносуппрессанты, или остеопороз, сопровождающий ревматоидный артрит.
38. Гибридома #32А1 (FERM ВР-10999) для получения антитела, которое ингибирует образование остеокластов и/или остеокластную резорбцию костей.
39. Гибридома #41В1 (FERM BP-11000) для получения антитела, которое ингибирует образование остеокластов и/или остеокластную резорбцию костей.
| БЕЛОК, СПОСОБНЫЙ ИНГИБИРОВАТЬ ОСТЕОКЛАСТОГЕНЕЗ (OCIF) (ВАРИАНТЫ), КДНК, СПОСОБ ПОЛУЧЕНИЯ БЕЛКА, СПОСОБ ЛЕЧЕНИЯ ОСТЕОПОРОЗА, СПОСОБ ЛЕЧЕНИЯ И/ИЛИ УЛУЧШЕНИЯ ВОСТАНОВЛЕНИЯ МАССЫ КОСТИ, СПОСОБ ЛЕЧЕНИЯ НАРУШЕНИЯ, СВЯЗАННОГО С КОСТНЫМ МЕТАБОЛИЗМОМ, ТРАНСФОРМИРОВАННЫЙ ШТАММ ESCHERICHIA COLI PBK/01F10 И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 1996 |
|
RU2238948C2 |
| WO 2007093042 A1, 23.08.2007 | |||
| ЗАМЕЩЕННЫЕ ПРОИЗВОДНЫЕ ПУРИНА, СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ФАРМАЦЕВТИЧЕСКИЙ ПРЕПАРАТ | 1997 |
|
RU2228335C2 |

