Настоящее изобретение относится к фармацевтической композиции для защиты животного от расстройства, возникающего в результате инфекции бактерией, которая относится к группе нокардиоформных актиномицетов. Изобретение также относится к применению живых ослабленных бактерий данной группы для получения указанной композиции и к способу лечения животного данной композицией.
Среди бактерий класса Actinobacteria имеется отряд бактерий, называемый Actinomycetales, обычно именуемый актиномицетами. Бактерии, которые относятся к этому отряду, представляют собой жгутиковые грамположительные бактерии (однако несколько видов имеют сложные структуры клеточной стенки, что делает классическое окрашивание по Граму менее подходящим или даже неподходящим, что имеет место, например, со многими видами, которые относятся к семейству Actinomycetales Mycobacteriaceae) с высоким содержанием G+C (гуанина+цитозина). Они лучше всего известны как обитающие в почве организмы, хотя различные штаммы населяют растения и животных, включая людей. Они образуют устойчивые споры, которые часто прикрепляются к воздушному мицелию или гифам.
Актиномицеты играют роль в разрушении органического материала. Несколько видов используются в промышленности и фармацевтических исследованиях ввиду их типичных свойств.
Большинство актиномицетов являются непатогенными для животных (термин «животные», используемый в связи с настоящим изобретением, включает людей). Однако внутри многих подотрядов актиномицетов (наряду с другими Streptosporangineae, Micrococcineae, Streptomycineae и Frankineae) имеется один подотряд, а именно Corynebacterineae, который включает наряду с большим количеством непатогенных бактерий существенное число патогенов животных. Представляется, что данные патогены относятся к филогенетической группе, известной как нокардиоформные актиномицеты, которая включает семейства Mycobacteriaceae, Nocardiaceae и Corynebacteriaceae (см. наряду с другими источниками главу 11, озаглавленную: Rhodococus equi: Pathogenesis and Replication in Macrophages, in «Opportunistic Intracellular Bacteria and Immunity», by Lois J. Paradise et al. (eds.), New York, 1999). Примечательно, против большей части заболеваний, связанных с инфекцией указанными патогенными микроорганизмами, едва ли существует какое-либо адекватное профилактическое лечение (т.е. лечение перед или по существу одновременно с вызывающим заболевание патогенным микроорганизмом, которое может быть способно или предотвратить возникновение заболевания, или, по меньшей мере, смягчить воздействия заболевания). В течение последних лет было подтверждено признание того, что семейства Mycobacteriaceae, Nocardiaceae и Corynebacteriaceae филогенной группы нокардиоформных актиномицетов представляют собой очень тесно связанные семейства внутри подотряда Corynebacterineae (см. также источник University of California, San Diego, Outline of Senior Project, Marelle L. Yehuda, June 2, 2005). Стало также ясно, что, в частности, патогенные бактерии в данной группе, по меньшей мере, те, по поводу инфекции которыми нет адекватного профилактического лечения (таких, как, например, Mycobacterium uberculosis, Nocardia seriolae и Rhodococcus equi), имеют важное общее свойство: инфекция обычно происходит через кожу или слизистую оболочку с последующей диссеминацией бактерий внутри макрофагов и репликацией внутри указанных макрофагов (см. наряду с другими источниками Microbes and Infection 7, 2005, 1352-1363; Proceedings of the National Academy of Sciences, June 7, 2005, Vol. 102, no 23, pp 8327 - 8332; Nature Medicine 13, 282-284, 2007; Transplantation Proceedings, Volume 36, Issue 5, June 2004, pp 1415-1418). Действительно, макрофаги находятся на передней линии иммунной защиты организма от микробных инфекций, но в отличие от бактерий, которые зависят от избегания фагоцитоза для выживания в организме хозяина, предусматриваемые в настоящее время патогенные бактерии внутри данной группы нацелены на макрофаги для выживания и даже реплицируются в организме хозяина. Настоящее изобретение относится к данным бактериям, которые обладают способностью выживать внутри макрофагов животного и в связи с настоящим изобретением будут именоваться нокардиоформными актиномицетами, выживающими в макрофагах.
Очевидно, нокардиоформные актиномицеты, выживающие в макрофагах, эволюционировали для избегания основных функций защиты животного от микробов. В частности, Mycobacterium tuberculosis, микроб, вызывающий туберкулез, представляет собой вид, который успешно эксплуатировал макрофаги в качестве своей первичной ниши in vivo, но другие бактериальные виды, которые относятся к группе нокардиоформных актиномицетов, включая Mycobacteriaceae, Nocardiaceae и Corynebacteriaceae, приняли такую же стратегию. Они представляют собой, например, Mycobacterium ulcerans, которая вызывает язву Бурули, Mycobacterium avium paratuberculosis, которая вызывает болезнь Джона у крупного рогатого скота и которая связана с болезнью Крона у людей, Mycobacterium bovis, которая вызывает бычий туберкулез, Mycobacterium avium, которая связана с оппортунистической инфекцией индивидов с нарушенным иммунитетом, таких как пациенты со СПИДом, Nocardia seriolae и Nocardia farcinia, которые вызывают нокардиоз у рыб, Nocardia asteroids, которая вызывает инфекцию у реципиентов почечного трансплантата, Rhodococcus equi (ранее известный как Corynebacterium), который вызывает пневмонию у жеребят и который также связан с оппортунистическими инфекциями у индивидов с нарушенным иммунитетом, Corynebacterium pseudotuberculosis, которая вызывает абсцессы наряду с другими органами в легких у овец, коз, лошадей и иногда также у людей и т.д. Все эти бактериальные виды имеют общую способность выживать внутри макрофагов, инфицировать их и реплицироваться внутри указанного типа клетки-хозяина.
Данное типичное свойство серьезно препятствует лечению расстройств (в настоящем описании термин «расстройство» используется в качестве эквивалента «заболевания»), возникающих в результате инфекции бактерией, которая относится к группе выживающих в макрофагах нокардиоформных актиномицетов. Во многих случаях, когда клинические признаки действительно присутствуют, проводится лечение антибиотиками. Однако это обременительно, поскольку внутри макрофагов присутствует значительное количество бактерий и они труднодостижимы антибиотиками. Поэтому лечение антибиотиками часто занимает много времени и дает неоднозначный эффект. По поводу таких заболеваний, как туберкулез у людей, нокардиоз у рыб и пневмония у жеребят, было бы предпочтительно профилактическое лечение. Такое профилактическое лечение обычно основано на применение вакцины, содержащей убитые или живые ослабленные бактерии, происходящие из бактерий дикого типа. Оказалось, что в отношении выживающих в макрофагах нокардиоформных актиномицетов убитые (инактивированные) вакцины (т.е. вакцины, содержащие убитые бактерии или их одну или более субъединиц в качестве терапевтического средства) неэффективны. В настоящее время в целом считается, что для успешного профилактического лечения против выживающих в макрофагах нокардиоформных актиномицетов необходимы живые бактерии, поскольку только они способны достичь макрофагов и имитировать бактерии дикого типа в степени, достаточной для запуска адекватного иммунного ответа. Действительно, для лечения туберкулеза имеется живая вакцина (БЦЖ, бацилла Кальметта-Герена), основанная на виде Mycobacterium bovis, который тесно связан с видом Mycobacterium tuberculosis. Однако защитный эффект был незначительным. В отношении вида Nocardiaceae, таких как Rhodococcus equi и Nocardia seriolae, в настоящее время нет выпускаемых промышленностью вакцин. В отношении вида Corynebacterium pseudotuberculosis была предпринята попытка борьбы с использованием аутогенных вакцин, однако это имело неоднозначный эффект (RUMA Guidelines, National Office of Animal health, Hertfordshire, United Kingdom, 2006).
Существует очевидная потребность в адекватном профилактическом лечении для защиты животного от расстройства, возникающего в результате инфекции выживающими в макрофагах нокардиоформными актиномицетами. Лечение в этом смысле означает стимуляцию иммунной системы животного-мишени в степени, достаточной, по меньшей мере, для уменьшения негативных эффектов заражения микроорганизмами дикого типа. Целью является то, что это ведет к защите от расстройства, т.е. предотвращается возникновение расстройства или, по меньшей мере, уменьшается уровень инфекции или клинические признаки заболевания у животного и, следовательно, также уменьшается тяжесть заболевания. Тот факт, что выживающие в макрофагах нокардиоформные актиномицеты приняли такую же стратегию выживания в организме хозяина, приводит к идее общей стратегии для профилактического лечения против инфекции указанными бактериями.
В этом отношении отмечается описание в литературе решающей роли, которую играет метаболизм холестерина в выживании нокардиоформных актиномицетов в макрофагах, который может быть важным фактором вирулентности (Proceedings of the National Academy of Science, February 6, 2007, vol. 104, no. 6, pp 1947-1952). Предполагалось также, что указанный метаболизм обеспечивает логические мишени для новых терапевтических средств для борьбы с вызывающими заболевания штаммами, т.е. лекарственных средств для лечения после возникновения инфекции. Действительно, при ретроспективном анализе можно найти другое подтверждающее доказательство установленного факта, что для всех выживающих в макрофагах нокардиоформных актиномицетов метаболизм холестерина играет роль в выживании и персистенции бактерий в макрофагах хозяина. Например, из главы 11 (озаглавленной: «Rhodococus equi: Pathogenesis and Replication in Macrophages (Патогенез и репликация в макрофагах) ») в монографии «Opportunistic Intracellular Bacteria and Immunity (Оппортунистические внутриклеточные бактерии и иммунитет)», под редакцией Lois J. Paradise et al., New York, 1999) известно, что существуют большие сходства в клинической симптоматологии между инфекциями, вызванными несколькими нокардиоформными актиномицетами, и было определено, что холестеролоксидаза является ферментным компонентом или фактором вирулентности. В руководстве «Veterinary Microbiology (Ветеринарная микробиология)», том 56, выпуск 3-4, июнь 1997, 269-276, показано, что Corynebacterium pseudotuberculosis вовлечена в процесс холестеролоксидазы вместе с Rhodococcus equi.
Поэтому на первый взгляд представляется привлекательной разработка фармацевтической композиции для защиты животного против расстройства, возникающего в результате инфекции выживающими в макрофагах нокардиоформными актиномицетами (такая композиция может также именоваться вакциной) с использованием признания того, что метаболизм холестерина играет решающую роль в выживании этих бактерий в макрофагах. Однако, понимая, что предложения, высказанные в приведенной выше в настоящем описании статье PNAS (статье 2007), относительно лекарственного средства и таким образом нацеленные на полное уничтожение бактерий вмешательством в их метаболизм холестерина, преследующие такую же стратегию, представляются неподходящими для живой вакцины: если предпринимается попытка ослабления бактерии прекращением ее выживания в существенном сайте репликации, то бактерия не будет реплицироваться и выживать у животного-хозяина. Действительно, для лечения лекарственными средствами это является идеальной ситуацией. Однако для живой вакцины, если полностью блокируется выживание бактерии, то ожидается имитация вакцины, содержащей убитые бактерии. Оказалось, что такие вакцины неэффективны для лечения инфекции выживающими в макрофагах нокардиоформными актиномицетами. Все же предпринимались попытки оценки применения живых бактерий, у которых нарушен метаболизм холестерина, в фармацевтической композиции для защиты животного от заражения болезнетворными нокардиоформными актиномицетами дикого типа. Примером таких попыток является живая вакцина на основе мутанта холестеролоксидазы (ChoE) штамма 103+ Rhodococcus equi дикого типа (Prescott in Veterinary Microbiology 118, 2006, pp 240-246). Данная попытка была безуспешной. Однако не потому, что она не вызывала защиты, что следовало бы ожидать на основании технических положений приведенной выше в настоящем описании в качестве ссылки статьи PNAS (PNAS February 6, 2007, vol. 104, no. 6, pp 1947-1952), а потому, что мутантный штамм был все еще слишком вирулентным. Мутант еще был способен выживать и размножаться в макрофагах на уровне, сравнимом с R. Equi дикого типа. Также представляется, что антигенная нагрузка этого аномального в отношении холестерина мутанта сравнима с антигенной нагрузкой организма дикого типа. Поэтому мутант все же был способен вызывать заболевание. Действительно, тем временем было также установлено, что живая мутантная Rhodococcus equi, которая вообще неспособна захватывать холестерин (мутант пермеазы захвата стерина supAB, как представлено в публикациях Van der Geize et al. at the 4th Havemeyer Workshop on Rhodococcus equi, Edinburgh, 13-16 July, 2008; и Van der Geize et al.: «A novel method to generate unmarked gene deletions in the intracellular pathogen Rhodococcus equi using 5-fluorocytosine conditional lethality» в монографии Nucleic Acids Research 2008; doi: 10.1093/nar/gkn811, далее также приводимые в качестве ссылки как «Van der Geize et al., 2008»), что означает полную блокаду метаболизма холестерина (по меньшей мере, когда холестерин используется в качестве исходного соединения), все же способна выживать и продолжает оставаться в макрофагах (Van der Geize et al., 2008) и таким образом является еще слишком вирулентной. Представляется, что для ослабления живой Rhodococcus equi требуется дополнительная мутация, оказывающая эффект вне метаболизма холестерина (Prescott: Veterinary Microbiology 125, 2007, 100-110). На основании данных результатов был сделан вывод, что сам метаболизм холестерина не является важным фактором вирулентности и не может использоваться для достаточного ослабления данных бактерий. Очевидно, бактерии, имеющие мутации их метаболизма холестерина, имеют такую же или, по меньшей мере, сравнимую антигенную нагрузку, как организм дикого типа, и таким образом, хотя они способны обеспечить адекватную защиту (то есть, если обсуждаемое животное выживает заражение мутированными бактериями), они еще слишком вирулентны для использования в фармацевтической композиции. Таким образом считалось, что нацеливание на фармацевтическую композицию для профилактического лечения, причем на композицию, содержащую живые бактерии, которые ослаблены инактивацией генов, участвующих в метаболизме холестерина, является тупиковым.
Однако, к удивлению, заявитель обнаружил, что для защиты животного от расстройства, возникающего в результате инфекции выживающей в макрофаге нокардиоформной актиномицетой, можно применять фармацевтическую композицию, содержащую живые бактерии вида нокардиоформных актиномицетов (обычно являющиеся таким же видом, как инфицирующая бактерия, или, альтернативно, являющиеся очень близко родственным видом, таким образом, имеющие много общих эпитопов T-клеток, как в случае с Mycobacterium tuberculosis в сравнении с Mycobacterium bovis), причем живые бактерии ослаблены инактивацией гена, который кодирует протеин, участвующий в разрушении метилгексагидроиндандиона пропионата, и фармацевтически приемлемый носитель для переноса бактерий.
«Ослабленные» в данном смысле означает неспособные вызвать полный набор симптомов заболевания, который обычно связан с вирулентным (часто дикого типа) патогенным аналогом ослабленной бактерии.
«Инактивация» в контексте настоящего изобретения означает, что ген, например, хотя и является частью оперона (т.е. набора генов, необходимого для действительной экспрессии протеина на функциональном уровне), или полностью удаляется из генома, или заменяется (любой известной или даже еще подлежащей разработке в будущем методикой; см., например, публикацию Introduction to Biotechnology and Genetic Engineering, A.J. Nair, INFINITY SCIENCE PRESS LLC, 2008, главу 13 "Genetic Techniques", pp 476-496 и главу 15 "Recombinant DNA Technology", pp 563-612) с тем, чтобы он больше не кодировал соответствующий протеин дикого типа или был больше недоступен для полной транскрипции, или любым другим изменением в геноме с тем, чтобы протеин дикого типа не был получен ослабленной бактерией in vivo, по меньшей мере, на уровне, подходящем для поддержания нормального катаболизма метилгексагидроиндандиона пропионата по сравнению с ситуацией, при которой ген (или оперон, если это применимо) представлен в форме, подходящей для поддержания нормального метаболизма.
«Кодирующий протеин» в контексте настоящего изобретения значит, что ген (например, хотя и являющийся частью оперона) непосредственно кодирует протеин или субъединицу протеина (множественные субъединицы, вместе образующие ферментативно активный протеин), или кодирует одно или более промежуточных соединений, которые превращаются или непосредственно, или через множество стадий в протеин или его субъединицу (множественные субъединицы, вместе образующие ферментативно активный протеин).
«Фармацевтически приемлемый носитель» может представлять собой растворитель, дисперсионную среду, покрытие, противобактериальное и противогрибковое средство, изотонический или задерживающий всасывание агент и тому подобные, которые физиологически совместимы с животным-мишенью и приемлемы для него, например, наряду с другими моментами, изготовлены в стерильном виде. Некоторыми примерами таких несущих сред являются вода, солевой раствор, солевой раствор с фосфатным буфером, жидкость для бактериальной культуры, декстроза, глицерин, этанол и тому подобные, а также их комбинации. Они могут обеспечить получение жидкой, полутвердой и твердой лекарственной форм в зависимости от предполагаемого способа введения. Общеизвестно, что присутствие несущей среды несущественно для эффективности вакцины, но она может значительно упростить дозирование и введение антигена. Кроме носителя и антигена, фармацевтическая композиция может содержать другие вещества, такие как адъюванты, стабилизаторы, модификаторы вязкости или другие компоненты, добавляемые в зависимости от предполагаемого применения или требуемых свойств композиции.
В фармацевтической композиции по настоящему изобретению присутствуют живые бактерии, причем указанные бактерии мутируют с тем, чтобы инактивировался ген, который кодирует протеин, участвующий в разрушении метилгексагидроиндандиона пропионата. Общеизвестно, что метилгексагидроиндандион пропионат (также известный как HIP или 3aα-H-4α(3'-пропионовая кислота)-7αβ-метилгексагидро-1,5-индандион) и 5-гидрокси-метилгексагидроинданон пропионат (также известный как HIL или 3aα-H-4α(3'-пропионовая кислота)-5α-гидрокси-7aβ-метилгексагидро-1-инданон-δ-лактон) образуются во время разрушения холестерина актинобактериями, включая выживающие в макрофагах нокардиоформные актиномицеты. Недавно в бактериальных видах, которые относятся к подотряду Corynebacterineae, был идентифицирован оперон (называемый ipdAB: разрушение инданона пропионата Альфа+Бета), кодирующий α и β субъединицу трансферазы, которая участвует в разрушении HIP (см. одновременно рассматриваемую международную патентную заявку PCT/EP2008/060844, поданную 19 августа 2008 г, основанную на первичной заявке на патент США, поданной 21 августа 2007 г.). Известный мутант с нокаутом трансферазы больше неспособен разрушать HIP и HIL (см. фиг.3 патентной заявки, приведенной выше в настоящем описании в качестве ссылки) и он не растет на HIP, HIL или 4-андростен-3,17-дионе. В любом случае «участвует в разрушении HIP» значит, что нокаутный мутант больше неспособен расти на HIP в качестве единственного источника углерода и энергии или, по меньшей мере, неспособен расти на HIP на уровне, который может быть получен немутантной бактерией. В настоящем неясно, катаболизирует ли трансфераза разрушение HIP сама, или реакция катализируется, отчего зависит разрушение HIP. Однако разрушение HIP происходит на относительно поздней стадии метаболизма холестерина и представляет собой очень специфическую стадию в пути разрушения холестерина. На основании общедоступных представлений о мутациях в метаболизме холестерина (указанные выше в настоящем описании ссылки на работы Prescott) следовало ожидать, что эта мутация привела бы к тому, что живая бактерия, хотя и обеспечивает защитный эффект, была бы слишком вирулентной. Однако к удивлению заявителя оказывается, что такой мутант адекватно ослаблен, что связано со значимо сниженной выживаемостью мутанта внутри макрофагов. Причина того, почему ген, который участвует в разрушении HIP, играет такую важную роль в выживании внутри макрофагов, неясна, это даже представляется противоречащим результатам предшествующего уровня техники, которые показывают, что даже полная блокада метаболизма холестерина оказывает недостаточный ослабляющий эффект, по-видимому, поскольку нокардиоформные актиномицеты не оказывают эффекта на выживаемость макрофагов. Поэтому было достаточно удивительным обнаружение того, что настоящая мутация, которая воздействует на второстепенную стадию катаболизма холестерина, создает серьезные препятствия выживанию патогенных нокардиоформных актиномицетов в макрофагах. В частности, было обнаружено, что мутантные живые бактерии все же способны поступать в макрофаги и оставаться в них (следовательно, обеспечивая стимул защитного иммунного ответа), но на очень низком уровне, который, как представляется, значительно снижает их вирулентность, что в свою очередь делает их приемлемыми для профилактического лечения.
В одном варианте осуществления инактивируется множество генов в одном опероне. Путем инактивации множества генов уменьшается вероятность изменения бактерии в фенотип, напоминающий дикий тип. В частности, инактивируются гены ipdA и ipdB. Путем инактивации указанных генов может быть обеспечено эффективное и безопасное ослабление. В предпочтительном варианте осуществления инактивация достигается делецией, по меньшей мере, одного гена делецией немаркированного гена. Преимущество немаркированной мутации состоит в том, что она обеспечивает возможность повторного введения мутаций в тот же штамм. Инородная ДНК (векторная ДНК) удаляется в процессе введения мутации. Поэтому вновь введенная векторная ДНК для введения второй мутации не может интегрироваться в сайте предыдущей мутации (путем гомологичной рекомбинации между векторными ДНК). Интеграция определенно произойдет, если векторная ДНК еще присутствует в хромосоме, и вызовет появление большого числа ложноположительных интегрантов. Система обеспечивает возможность использования одного антибиотического гена для введения бесконечного числа мутаций. Немаркированная мутация также обеспечивает возможность легкого использования в промышленности ввиду отсутствия гетерогенной ДНК, обеспечивающей возможность легкого удаления ферментационного бульона. Инактивация генов делецией генов обеспечивает возможность конструирования устойчивых ревертирующих мутантов. Особенно маленькие гены (<500 пар оснований) легче инактивируются делецией генов по сравнению с разрывом генов интеграцией одной рекомбинации. Генный делеционный мутагенез может также применяться для инактивации кластера нескольких генов из генома. Стратегия генного делеционного мутагенеза может также применяться для замещения генов (например, изменения дикого типа в мутантный ген).
В одном варианте осуществления бактерии относятся к семейству Nocardiaceae или Mycobacteriaceae. Предпочтительно бактерии относятся к родам Rhodococcus, Nocardia или Mycobacterium и в частности относятся к любому из видов Rhodococcus equi, Nocardia seriolae, Mycobacterium tubercolosis, Mycobacterium ulcerans, Mycobacterium bovis или Mycobacterium avium paratubercolosis. В отношении этих видов до настоящего времени промышленностью не выпускаются адекватные вакцины. Настоящее изобретение обеспечивает возможность получения фармацевтических композиций, которые могут применяться в качестве вакцин для борьбы с этими бактериями и поэтому смягчают течение соответствующих заболеваний, которые они вызывают у животного.
В одном варианте осуществления фармацевтическая композиция представлена в форме, подходящей для перорального введения. Кроме того, это очень удобный путь введения, в частности, стало ясно, что данный путь введения является безопасным. Парентеральное введение может вызвать развитие абсцессов. Предпочтительно, живые бактерии присутствуют в концентрации от 1×104 до 1×1010 CFU (колониеобразующих единиц) на дозу.
Настоящее изобретение также относится к бактериям Rhodococcus equi, происходящим из штамма, депонированного в Национальной Коллекции Культур Микроорганизмов Института Пастера в Париже, Франция, под № CNCM 1-4108 или № CNCM 1-4109, и к бактериям, которые относятся к данному штамму.
Настоящее изобретение также относится к использованию живых бактерий, которые относятся к группе нокардиоформных актиномицетов, способных выживать внутри макрофагов животного, причем живые бактерии ослабляются инактивацией гена, который кодирует протеин, участвующий в разрушении метилгексагидроиндандиона пропионата, для получения фармацевтической композиции для защиты животного от расстройства, возникающего в результате инфекции соответствующей бактерией дикого типа.
Изобретение также относится к способу лечения животного для защиты его от расстройства, возникающего в результате инфекции бактерией, относящейся к группе нокардиоформных актиномицетов, обладающих способностью выживать внутри макрофагов животного, включающему введение животному фармацевтической композиции, содержащей живые бактерии вида нокардиоформных актиномицетов, причем живые бактерии ослаблены инактивацией гена, который кодирует протеин, участвующий в разрушении метилгексагидроиндандиона пропионата.
Изобретение будет далее объяснено с использованием следующих примеров, описывающих определенные варианты осуществления настоящего изобретения, причем указанные варианты осуществления распространяются на три части:
Часть A: Идентификация и конструирование штаммов
Часть B: Выживание в макрофагах как модель для ослабления in vivo
Часть C: Эффективность мутантных бактерий в защите против инфекции диким типом
ЧАСТЬ A: ИДЕНТИФИКАЦИЯ И КОНСТРУИРОВАНИЕ ШТАММОВ
A1 Культуральные среды и условия роста
Штаммы R. equi выращивали при 30°C (200 об/мин) в среде Luria-Bertani (LB), состоящей из бактотриптона (BD), Дрожжевого экстракта (BD) и 1% NaCl (Merck) или минеральной среде с ацетатом. M. smegmatis mc2155 (Snapper et al., 1990, Mol. Microbiol. 4:1911-1919) выращивали при 37°C (200 об/мин) в триптическом соевом бульоне BBL (TSB; BD) с добавкой 0,05% Твин 80. Минеральная среда с ацетатом (MM-Ac) содержала K2HPO4 (4,65 г/л), NaH2PO4-H2O (1,5 г/л), Na-ацетат (2 г/л), NH4Cl (3 г/л), MgSO4·7H2O (1 г/л), тиамин (40 г/л, стерилизованный фильтрацией; Sigma) и основной раствор Vishniac (1 мл/л). Основной раствор Vishniac получали следующим образом (модифицированным способом Vishniac and Santer, 1957, Rev. 21: 195-213): EDTA (этилендиаминтетрауксусную кислоту) (10 г/л) и ZnSO4·7H2O (4,4 г/л) растворяли в дистиллированной воде (pH 8 с использованием 2 M KOH). Затем добавляли CaCl2·2 H2O (1,47 г/л), MnCl2·7H2O (1 г/л), FeSO4·7H2O (1 г/л), (NH4)6 Mo7O24·4 H2O (0,22 г/л), CuSO4·5 H2O (0,315 г/л) и CoCl2·6 H2O (0,32 г/л) в указанном порядке при pH 6 и, наконец, хранили при pH 4.
Для роста на твердых средах добавляли бактоагар (15 г/л; BD). Основной раствор 5-фторцитозина (Sigma- Aldrich) (10 мг/мл) получали в дистиллированной воде, растворяли нагреванием до 50°C, стерилизовали фильтрацией и добавляли к автоклавированной среде.
Штамм INS436 Nocardia seriola обычным образом выращивали при 26°С (200 об/мин) в бульоне Eugon (BD) с добавлением Твин 80 (0,05%). Для роста на твердых средах добавляли бактоагар (15 г/л; BD). К агаровой среде добавляли сахарозу (2%) для sacB-зависимого сахарозного отбора.
A2 Идентификация ipdA, ipdB и fadE30 в штамме 103+ R. equi и ipdAB в штамме INS436 N. seriolae
Как указано выше в настоящем описании, обнаружено, что гены ipdA и ipdB Rhodococcus участвуют в разрушении метилгексагидроиндандиона пропионата (HIP; 3aα-H-4α(3'-пропионовая кислота)-7aβ-метилгексагидро-1,5-индандиона) и 5-гидрокси-метилгексагидроиндандиона пропионата (HIL; 3aα-H-4α(3'- пропионовая кислота)-5α-гидрокси-7aβ-метилгексагидро-1-инданон-δ-лактона). Биоинформационные анализы протеиновых последовательностей IpdA и IpdB штамма SQ1 R. erythropolis в геномных базах данных выявили, что гены, кодирующие IpdA и IpdB, и их видимая оперонная организация сохранились в геноме R. equi 103+ (штамма дикого типа, полученного от J. F. Prescott, Ontario, Canada; как приведено в качестве ссылки в руководстве Veterinary Microbiology 118 (2006) 240-246). Геномная последовательность R. equi 103+ была определена группой секвенирования R. equi в Sanger Institute, Hinxton, Cambridge, UK (геном, опубликованный как «R. equi 103S»). Анализ генома, кроме того, выявил, что R. equi 103+ содержит дополнительные паралогичные гены ipdA и ipdB, обозначенные соответственно как ipdA2 и ipdB2. Данные гены локализуются снаружи от кластера катаболических генов холестерина. Аминокислотные последовательности IpdA, IpdB, IpdA2 и IpdB2 изображены в прилагаемых идентичностях SEQ ID соответственно под №№ 1, 2, 3 и 4. Идентичности аминокислотных последовательностей протеинов IpdA и IpdB R. equi 103+ данными паралогичными и несколькими другими актинобактериальными ортологами приведены в таблице 6. Эта таблица представляет обзор генов, идентифицированных в других геномах нокардиоформных актиномицетов, кодирующих ортологи IpdA и IpdB Rhodococcus equi 103+. В связи с настоящим изобретением данные и другие ортологи называются IpdA и IpdB. Идентичность протеинов указывает процентную долю идентичности аминокислотных последовательностей полной длины с IpdA и IpdB R. equi 103S. Актинобактериальные геномные последовательности были получены из геномного сервера BLAST для микробных геномов Национального Центра Биотехнологической Информации (NCBI). Данные о последовательности R. equi 103+ были получены группой секвенирования R. в Sanger Institute. Геном штамма 103+ (известный как 103S) использовали для этих целей идентификации. Практическая работа с Rhodococcus equi, как иллюстрируется ниже в настоящем описании, проводилась со штаммом RE1 R. equi (выделенным у жеребенка, страдающего грануломатозной пневмонией, вызванной инфекцией Rhodococcus equi).
Второй ген, участвующий в разрушении метилгексагидроиндандиона пропионата, представляет собой fadE30. Рост мутанта ΔfadE30 значительно нарушен на HIL и HIP по существу без проявления роста после 24 часов инкубации. Ген fadE30 Rhodococcus equi был идентифицирован исследованием сходства протеиновых последовательностей, выполненным на геноме R. equi 103+, имеющемся в институте Sanger (http://www.sanger.ac.uk). Аннотированную протеиновую последовательность FadE30 штамма RHA1 Rhodococcus jostii (Ro4596, номер доступа в Генном банке ABG96382) использовали в качестве матрицы протеиновой последовательности (McLeod et al., 2006, in Proc. Natl. Acad. Sci. U.S.A. 103:15582-15587; и Van der Geize et al., 2007, в Proc. Natl. Acad. Sci. U.S.A. 104:1947-1952). Исследование базы данных сходства с использованием Ro04596 выявило ген R. equi 103S, кодирующий протеин, который проявил идентичность аминокислотной последовательности 73% с Ro04596. Данный протеин был аннотирован как FadE30 R. equi 103S (SEQ ID No 43) и его соответствующий ген был назван fadE30. Ортологичные гены, кодирующие FadE30 у других актинобактерий, могут быть идентифицированы аналогичным образом. Их отбор представлен в таблице 8.
Геномный локус ipdAB N. seriola амплифицировали PCR (ПЦР, полимеразной цепной реакцией) в трех частях с использованием олигонуклеотидных праймеров, разработанных на высококонсервативных нуклеотидных последовательностях актинобактериального локуса ipdAB. Данные консервативные области были идентифицированы совмещением нуклеотидных последовательностей нескольких известных актинобактериальных последовательностей генов ipdAB. Нуклеотидную геномную последовательность области ipdAB Nocardia farcinica (nfa05080- nfa05090) (DDBJ номер доступа AP006618) использовали в качестве первичной матрицы для разработки олигонуклеотидных ПЦР-праймеров. Используемые олигонуклеотидные праймерные последовательности перечислены в таблице 5. Хромосомную ДНК N. seriola INS436 использовали в качестве матрицы для ПЦР. Гены ipdAB N. seriola амплифицировали с использованием праймеров ipdA-actino-F и ipdB-actino-R (ПЦР 22), и находящуюся выше по ходу транскрипции область ipdAB амплифицировали с использованием ipd-актино-F2 и ipdA-актино-R (ПЦР 23), а находящуюся ниже по ходу транскрипции область амплифицировали с использованием ipdB-актино-F и ipd-актино-R (ПЦР 21). Продукты ПЦР клонировали в вектор клонирования pGEM-T и определяли нуклеотидные последовательности вставок. В последующем были разработаны другие пары праймеров на полученной последовательности ДНК и использованы для реклонирования и ресеквенирования локуса ipd. Это привело к получению полной нуклеотидной последовательности локуса ipdAB N. Seriola, охватывающей 4139 пар оснований. Секвенированный фрагмент ДНК содержал гены ipdA и ipdB N. Seriola и их соседние гены. Выведенные протеиновые последовательности IpdA и IpdB N. seriola INS436 показаны соответственно в SEQ ID NO 58 и SEQ ID NO 59.
A3 Клонирование, ПЦР и выделение геномной ДНК
DH5α Escherichia coli использовали в качестве хозяина для всех процедур клонирования. Рестрикционные ферменты были получены у компании Fermentas GmbH. Хромосомную ДНК клеточных культур выделяли с использованием набора бактериальной геномной ДНК GenElute Bacterial Genomic DNA Kit (Sigma-Aldrich) в соответствии с инструкциями производителя.
ПЦР выполняли в реакционной смеси (25 мкл), состоящей из Трис-HCl (10 мМ, pH 8), 1x стандартного полимеразного буфера, dNTP (деоксинуклеотид трифосфат) (0,2 мМ), DMSO (диметилсульфоксид) (2%), ПЦР-праймеры (каждого по 10 нг/мкл, таблица 5) и высокоспецифичной ДНК-полимеразы (Fermentas) или ДНК-полимеразы Pwo (Roche Applied Science). Для ПЦР колоний клеточный материал смешивали со 100 мкл хлороформа и 100 мкл 10 мМ Tris-HCl при pH 8, энергично перемешивали вихревой мешалкой и центрифугировали (2 мин, 14000×g). Образец верхней водной фазы (1 мкл) в последующем использовали в качестве матрицы для ПЦР. Стандартная ПЦР включала 5-минутную стадию плавления ДНК при 95°C с последующими 30 циклами по 45 сек денатурации при 95°C, 45 сек отжига при 60°C и 1-3 мин удлинения при 72°C. Используемое время удлинения зависело от длины ожидаемого ПЦР ампликона, принимая 1,5 мин/1 тысячу оснований в качестве общего правила.
A4 Электротрансформация R. equi, M. smegmatis и N. seriolae
Клетки штаммов R. equi трансформировали электропорообразованием, по существу, как описано (Van der Geize et al., в принятой к публикации в NAR статье, приведенной выше в качестве ссылки; Navas et al., 2001, J. Bacteriol. 183: 4796-4805). Вкратце, клеточные культуры выращивали в 50 мл среды LB при 30°C до тех пор, пока OD600 (оптическая плотность при длине волн 600 нм) не достигала 0,8-1,0. Клетки подвергали пеллетированию (20 мин при 4500×g) и дважды промывали 10% ледяным глицерином. Подвергнутые пеллетированию клетки ресуспендировали в 0,5-1 мл ледяного 10% глицерина и делили на аликвоты по 200 мкл.
Клетки M. smegmatis mc2155 трансформировали электропорообразованием, по существу, как описано в литературе (Jacobs et al., 1991, Methods Enzymol. 204:537-555). Вкратце, клеточные культуры (250 мл) выращивали при 37°C в среде TSB+0,05% Твин 80 до тех пор, пока OD600 достигала 0,8, клали на лед на полтора часа и центрифугировали (10 мин при 5000×g) для пеллетирования клеток. Клеточные пеллеты дважды промывали дистиллированной водой и ресуспендировали в конечном объеме 1 мл 10% глицерина и делили на аликвоты по 200 мкл.
Элюированную MilliQ плазмидную ДНК (5-10 мкл; набор GenElute Plasmid Miniprep Kit, Sigma-Aldrich) добавляли к 200 мкл клеток в кюветы с зазорами 2 мм. Электропорообразование выполняли одиночным импульсом 12,5 кВ/см, 1000 Ом и 25 мкфарад. Подвергнутые электропорообразованию клетки осторожно смешивали с 1 мл среды LB (R. equi) или 1 мл TSB+0,05% Твин 80 (M. smegmatis) и давали возможность восстановиться в течение 2 ч (R. equi) или 5 ч (M. smegmatis) при 37°C и 200 об/мин. Аликвоты (200 мкл) восстановленных клеток высевали на селективную агаровую среду. Трансформанты R. equi отбирали на агаре LB, содержащем апрамицин (50 мкг/мл) и появлялись после 2-3 дней инкубации при 30°C. Трансформанты M. smegmatis отбирали на агаровой среде с TSB+0,05% Твин 80, содержащей канамицин (10 мкл/мл), и они появлялись после 4-5 дней инкубации при 37°C.
Для N. seriolae прекультуру (20 мл) штамма INS436 выращивали в течение 5 дней при 26°C (200 об/мин) в бульоне Eugon+0,05% Твин 80 (OD6oonm=6) и использовали для инокуляции 100 мл свежего бульона Eugon+0,05% среды Твин 80 (1:100). Первичную культуру выращивали в течение ночи при 26°C в течение 20 ч до OD600нм=1,3.
Клетки подвергали пеллетированию (20 мин, 4000 g) при 4°C и дважды промывали 50 мл ледяного 10% глицерина. Подвергнутые пеллетированию клетки ресуспендировали в 500 мкл 10% глицерина, делили на аликвоты по 200 мкл и немедленно использовали для электротрансформации. Подвергнутую элюированию MilliQ плазмидную ДНК (5-10 мкл; набор GenElute Plasmid Miniprep Kit, Sigma-Aldrich) добавляли к 200 мкл клеток в кювету с зазором 2 мм, смешивали и оставляли на 1 мин на льду. Электропорообразование выполняли одиночным импульсом 1,75 кВ/см, 200 Ом и 50 мкфарад (приблизительная продолжительность импульса 9,3 мс). Подвергнутые порообразованию клетки осторожно смешивали с 1 мл бульонной среды Eugon с добавлением 0,05% Твин 80 и давали возможность восстановиться в течение 3,5 ч при 26°C и 220 об/мин. Аликвоты по 50 и 100 мкл восстановленных клеток высевали на селективный агар Eugon с добавлением 0,05% Твин 80 и канамицина (20 мкг/мл). Трансформанты появлялись после 7 дней инкубации при 26°C.
A5 Делеция немаркированных генов в штаммах R. equi выбора с использованием 5-фторцитозина (5-FC)
Мутанты R. equi получали делецией немаркированных генов, по существу, как описано в литературе (Van der Geize et al., 2008). Трансформанты R. equi, полученные в результате электропорообразования клеток дикого типа или мутантных клеток, штрихами высевали на агаровую среду LB с добавлением апрамицина для подтверждения устойчивости к апрамицину (ApraR). Четыре трансформанта ApraR на одну трансформацию выращивали в течение ночи (20-24 ч) при 30°C и 200 об/мин в 25 мл среды LB и высевали в 101-103-кратных разведениях в среде MM-Ac на чашки с агаром MM-Ac с добавлением 5-FC (100 мкг/мл) в аликвотах по 100 мкл. Устойчивые к 5-FC колонии, появляющиеся после 3 дней инкубации при 30°C, высевали штрихами перепечатыванием с чашки на чашку с агаром LB и агаром LB с добавлением апрамицина (50 мкг/мл) для отбора чувствительных к апрамицину (Apras) и устойчивых к 5-FC (5-FCR) колоний. Apras/5-FCR колонии проверяли на присутствие желательной делеции генов ПЦР колоний с использованием праймеров, амплифицирующих локус делеции генов (таблица 5). Геномную ДНК выделяли из потенциальных мутантов с делецией генов и использовали для подтверждения делеции гена с использованием праймеров, амплифицирующих локус гена ipdAB или ipdAB2, а также находящихся выше и ниже по ходу транскрипции областей указанных локусов праймерами, как представлено в таблице 5.
A6 Конструкция плазмид для делеции генов ipdAB и ipdAB2 в R. equi
Плазмиду pSelAct-ipd1 для генерирования делеции немаркированного гена оперона ipdAB в R. equi RE1 конструировали следующим образом. Находящиеся выше (1368 пар оснований; праймеры ipdABequiUP-F и ipdABequiUP-R) и ниже (1396 пар оснований; праймеры ipdABequiDOWN-F и ipdABequiDOWN-R) по ходу транскрипции фланкирующие области генов ipdAB амплифицировали ПЦР (Таблица 5, PCR1 и PCR2). Полученные ампликоны лигировали в переваренный EcoRV pBluescript(ll)KS, предоставляя плазмиды pEqui14 и pEqui16 соответственно для находящейся выше и ниже по ходу транскрипции области. Spel/EcoRV (1,4 т.п.о.), являющийся фрагментом pEqui14, лигировали в Spel/EcoRV, переваренный pEqui16, генерируя pEqui18. EcoRI/HindIII (2,9 т.п.о.), являющийся фрагментом pEqui18, содержащего делецию гена ipdAB, и его фланкирующие области обрабатывали фрагментом Klenow и лигировали в переваренный SmaI, суицидальный вектор pSelAct (Van der Geize et al., 2008). Полученная плазмида была обозначена pSelAct-ipd1 для конструирования мутанта с делецией гена ipdAB R. equi Δipd AB (также именуемого RG1341). Этот мутант был депонирован в Национальной Коллекции Культур Микроорганизмов Института Пастера в Париже, Франция, под № CNCM 1-4108.
Мутант R. equi с двойной делецией генов ΔipdABΔipdAB2 (также именуемый RG2837) получали делецией немаркированного гена оперона в мутантном штамме R. Equi ΔipdAB с использованием плазмиды pSelAct-ΔipdAB2. Плазмиду pSelAct-ΔipdAb2 конструировали следующим образом. Находящиеся выше (1444 пар оснований; праймеры ipdAB2equiUP-F и ipdAB2equiUP-R) и ниже по ходу транскрипции (1387 пар оснований; ipdAB2equiDOWN-F, ipdAB2equiDOWN-R) области ipdAB2 амплифицировали ПЦР с использованием геномной ДНК в качестве матрицы (таблица 5, PCR6 и PCR7). Ампликоны лигировали в SmaI, переваренный pSelAct, получая в результате соответственно плазмиды pSelAct-ipdAB2equiUP и pSelAct-ipdAB2equiDOWN. После переваривания обеих плазмид BgIII/SpeI фрагмент pSelAct-ipdAB2equiDOWN (1381 пар оснований) лигировали в pSelAct-ipdAB2equiUP, приводя к получению pSelAct-ΔipdAB2, используемой для конструирования делеции гена ΔipdAB2. Полученный в результате мутант R. equi ΔipdABΔipdAB2 был депонирован в Национальной Коллекции Культур Микроорганизмов Института Пастера в Париже, Франция, под № CNCM 1-4109.
A7 Конструирование плазмиды для делеции гена ipdAB в M. smegmatis mc 2 155
Плазмиду pK18-ipdABsmeg, используемую для делеции немаркированного гена генов ipdAB в M. smegmatis mc2155, конструировали следующим образом.
Находящиеся выше (1502 пар оснований; праймеры ipdABsmegUP-F и ipdABsmegUP-R) и ниже (1431 пар оснований; праймеры ipdABsmegDOWN-F и ipdABsmegDOWN-R) по ходу транскрипции фланкирующие области генов ipdAB амплифицировали ПЦР с использованием геномной ДНК M. smegmatis mc2155 в качестве матрицы (таблица 5, PCR12 и PCR13). Полученные ампликоны лигировали в переваренную SmaI pK18mobsacB (Schafer et al., 1994, Gene 145:69-73), в результате приводя к получению соответственно pK18-ipdABsmegUP и pK18-ipdABsmegDOWN. Фрагмент ДНК 1,5 т.п.о., полученный из переваренной BamHI/SpeI pK18-ipdABsmegUP, в последующем лигировали в pK18-ipdABsmegUP линеаризированную SamHI/Spel, приводя в результате к конструированию pK18-ipdABsmeg, используемой для делеции гена ipdAB.
A8 Конструирование плазмиды для делеции гена fadE30 в R. equi
Плазмиду pSelAct-fadE30 для генерирования делеции немаркированного гена fadE30 в R. equi RE1 конструировали следующим образом. Находящиеся выше (1511 пар оснований; праймеры fadE30equiUP-F и fadE30equiUP-R) и ниже (1449 пар оснований; праймеры fadE30equiDOWN-F и fadE30equiDOWN-R) по ходу транскрипции фланкирующие геномные области fadE30 амплифицировали стандартной ПЦР, используя высокоспецифичную ДНК-полимеразу (Fermentas GmbH) (таблица 5; PCR 15 и PCR 16). Полученные ампликоны лигировали в клонирующий pGEM-T вектор (Promega Benelux), предоставляя pGEMT-fadE30UP и pGEMT- fadE30DOWN. Фрагмент ДНК BcuI/Bg/II (1,4 т.п.о.) отрезали от pGEMT-fadE30DOWN и лигировали в BcuI/Bg/II, линеаризованную pGEMT-fadE30UP, приводя в результате к получению pGEMT-fadE30. Для конструирования pSelAct-fadE30, pGEMT-fadE30 переваривали Ncol и BcuI и обрабатывали фрагментом Klenow. Тупоконечный фрагмент ДНК (2,9 т.п.о.), несущий делецию гена fadE30, лигировали в переваренную SmaI pSelAct (van der Geize et al., 2008). Полученная плазмида была обозначена pSelAct-fadE30 и использовалась для конструирования мутантного штамма R. Equi ΔfadE30.
A9 Конструирование плазмиды для делеции гена ipdAB в N. seriola INS436
Плазмиду pK18ipdABNser, используемую для делеции немаркированного гена генов ipdAB в N. seriola INS436, конструировали следующим образом. Находящиеся выше (1487 пар оснований; праймеры ipdABNserUP-F и ipdABNserUP-R; PCR 24) и ниже (1049 пар оснований; праймеры ipdABNserDOWN-F и ipdABNserDOWN-R; PCR 25) по ходу транскрипции фланкирующие области генов ipdAB амплифицировали ПЦР, используя геномную ДНК N. seriola INS436 в качестве ДНК матрицы (таблица 5). Полученные ампликоны лигировали в переваренную SmaI pK18mobsacB (Schafer ef al., 1994, in Gene 145: 69-73)), приводя к получению соответственно pK18-ipdABNserUP и pK18-ipdABNserDOWN. Фрагмент ДНК 1,07 т.п.о., полученный из переваренной SmaI/PstI pK18-ipdABNserDOWN, был в последующем лигирован в pK18-ipdABNserUP, которая была линеаризована SmaI/PstI, приводя в результате к конструированию плазмиды pK18-ipdABNser, которая использовалась для делеции гена ipdAB.
A10 Конструирование мутантных штаммов R. equi ΔipdAB и R. equi ΔipdABΔipdAB2
Мутанты R. equi с делецией немаркированных генов ipdAB (RG1341) и ipdABipdAB2 (RG2837) конструировали, используя стратегию двухэтапной гомологичной рекомбинации контрселекцией с использованием 5-фторцитозина, разработанной для R. equi (Van der Geize et al., 2008). Для конструирования ΔipdAB-мутантного штамма RG1341 R. equi нерепликативная плазмида pSelAct-ipd1 была мобилизована в штамм RE1 R. equi электротрансформацией. 4 трансформанта ApraR, полученные в результате гомологичной рекомбинации между плазмидой pSelAct-ipd1 и геномом RE1 в последующем подвергали селекции с использованием 5-FC для отбора по возникновению второго редкого явления гомологичной рекомбинации, приводящей к делеции гена. 18 случайно выбранных колоний Apras/5FCR подвергали ПЦР колонии, и 3 колонии FCR/Apras дали ампликон ожидаемого размера (296 пар оснований, таблица 5, PCR5). Геномную ДНК выделяли из указанных трех мутантов ΔipdAB и подвергали анализу ПЦР локуса ipdAB и его фланкирующих областей выше и ниже по ходу транскрипции (таблица 5, PCR3 и PCR4). Данный анализ подтвердил присутствие подлинной делеции гена ipdAB в двух из трех случаев и не выявил аберрантных геномных перегруппировок в локусе ipdAB. Присутствие vapA в качестве маркера плазмиды вирулентности было подтверждено ПЦР (таблица 5, PCR11). Был выбран один ipdAB-мутантный штамм, обозначенный R. equi RG1341, и использовался для дальнейшей работы.
Мутантный штамм с двойной делецией генов был сконструирован из штамма RG2837 с использованием плазмиды pSelAct-ΔipdAB2, по существу, как описано для выделения одного мутанта ΔipdAB. 4 трансформанта ApraR, полученные в результате электропорообразования клеток штамма RG1341 с использованием pSelAct-ΔipdAB2, подвергали селекции 5-FC для выбора колоний Apras/5-FCR. Последующий ПЦР-анализ 18 колоний Apras/5-FCR подтвердил, что 2 колонии содержали делецию гена ΔipdAB2, о чем свидетельствует полученный ампликон размером 123 пары оснований с использованием олигонуклеотида, полученного для амплификации оперона ipdAB2 (таблица 5, PCR10). Дальнейший анализ ПЦР находящихся выше и ниже локуса ipdAB2 по ходу транскрипции областей подтвердил присутствие делеции гена ipdAB2 и не выявил аберрантных геномных перегруппировок (таблица 5, PCR8 и PCR9). Также присутствие гена вирулентности vapA было подтверждено ПЦР (таблица 5, PCR11). Один мутантный штамм RG2837 с двойной делецией гена ΔipdABΔipdAB2 был выбран для дальнейшей работы.
A11 Конструирование мутантного штамма ΔipdAB M. smegmatis
Мутант M. smegmatis mc2155 с делецией немаркированного гена ipdAB конструировали с использованием системы контрселекции sacB (Pelicic et al., 1996, MoI. Microbiol. 20:919-925; Van der Geize et al., 2001, FEMS Microbiol Lett. 205:197-202) следующим образом. Для конструирования мутанта ΔipdAB штамма M. smegmatis mc2155 нерепликативную плазмиду pK18-ipdABsmeg мобилизовали в M. smegmatis электропорообразованием. Были получены несколько трансформантов. Один устойчивый к канамицину трансформант выращивали в течение 2 дней при 37°C неселективно в среде TSB, содержащей 0,05% Твин 80, и в последующем высевали на чашки с TSB агаром, содержащим 2% сахарозу, для отбора чувствительных к канамицину (Kms) и устойчивых к сахарозе (SucR) двойных рекомбинантов контрселекцией sacB. Колонии, появлявшиеся после 3 дней инкубации, штриховыми отпечатками высевали на TSB агар и TSB агар с добавлением канамицина (10 мкг/мл) для отбора колоний Kms/SucR. Истинные колонии Kms/SucR далее проверяли ПЦР колоний для выявления присутствия делеции гена ipdAB (таблица 5, PCR14). Геномную ДНК выделяли из трех потенциальных мутантов ipdAB. ПЦР-анализ подтвердил присутствие делеции гена ipdAB, и один мутантный штамм ipdAB был выбран для дальнейшей работы и обозначен как M. smegmatis ΔipdAB.
A12 Конструирование мутантного штамма R. equi ΔfadE30
Делецию немаркированного гена fadE30 в R. equi RE1 генерировали с использованием стратегии двухэтапной гомологичной рекомбинации контрселекцией 5-фторурацилом, разработанной для R. equi (Van der Geize et al., 2008). Для конструирования мутантного штамма ΔfadE30 нерепликативную плазмиду pSelAct-fadE30 мобилизовали в штамм RE1 R. equi электротрансформацией. Два трансформанта ApraR, полученные в результате гомологичной рекомбинации между плазмидой pSelAct-fadE30 и геномом RE1, в последующем подвергали селекции 5-FC для селекции по возникновению явления второй редкой гомологичной рекомбинации в результате делеции гена fadE30. 18 случайно взятых колоний Apras/5FCR подвергали ПЦР колоний с использованием праймеров fadE30cont-F и fadE30cont-R (таблица 5; PCR 19) и 13 колоний FCR/Apras дали ампликон ожидаемого размера (428 пар оснований). Геномную ДНК выделяли из двух потенциальных мутантов ΔfadE30 и подвергали ПЦР-анализу. ПЦР-анализ локуса fadE30 с использованием олигонуклеотидных праймеров fadE30contr-F и fadE30contr-R (таблица 5) подтвердил присутствие делеции гена fadE30 и отсутствие гена fadE30 дикого типа. ПЦР-анализ фланкирующих областей выше и ниже по ходу транскрипции показал получение ожидаемых продуктов, имеющих длину соответственно 1,86 т.п.о. и 1,76 т.п.о. Анализ подтвердил присутствие истинной делеции гена fadE30 в обоих случаях и не выявил аберрантных геномных перегруппировок в локусе fadE30. Присутствие vapA как маркера плазмиды вирулентности было подтверждено ПЦР. Один мутантный штамм fadE30 был обозначен R. equi ΔfadE30 и использован для дальнейшей работы.
A13 Конструирование мутантного штамма N. seriola ΔipdAB
Мутант N. seriola INS436 с делецией немаркированного гена ipdAB конструировали с использованием системы контрселекции sacB (Pelicic et al., 1996, in MoI. Microbiol. 20:919-925; Van der Geize et al., 2001, in FEMS Microbiol Lett. 205:197-202) следующим образом. Нерепликативную плазмиду pK18-ipdABNser мобилизовали в N. seriola INS436 электротрансформацией. Затем несколько устойчивых к канамицину трансформантов выращивали неселективно в течение 7 дней при 26°С в бульонной среде Eugon, содержащей 0,05% Твин 80. Отбор чувствительных к канамицину (Kms) и устойчивых к сахарозе (SucR) двойных рекомбинантов контрселекцией sacB в последующем выполняли высеванием на чашки с агаром Eugon, содержащим 2% сахарозу. Колонии, появившиеся после 7 дней инкубации при 26°C, штриховыми отпечатками высевали на чашки с агаром Eugon и чашки с агаром Eugon с добавлением канамицина (20 мкг/мл) для отбора колоний Kms/SucR. Истинные колонии Kms/SucR далее проверяли ПЦР колоний для выявления присутствия гена ipdAB с использованием праймеров ipdABNser-F и ipdABNser-R (таблица 5). Геномную ДНК выделяли из трех потенциальных мутантов ipdAB. ПЦР-анализ с использованием пары праймеров ipdABNser-F и ipdABNser-R (PCR 23) привел к выявлению продукта ПЦР размером 222 пары оснований и отсутствия продукта ПЦР дикого типа размером 1624 пары оснований, подтверждая присутствие делеции гена ipdAB. Пару праймеров IpdABNser-F2 и IpdABNserDOWN-Contr-R (PCR 26) использовали для дополнительного подтверждения делеции гена ipdAB. Данная пара праймеров привела к получению ожидаемого продукта размером 1148 пар оснований для мутанта ipdAB, тогда как продукт ПЦР размером 2560 пар оснований был получен только для штамма дикого типа. Один мутантный штамм был обозначен N. seriola ΔipdAB и выбран для дальнейшей работы.
ЧАСТЬ B: ВЫЖИВАНИЕ В МАКРОФАГАХ КАК МОДЕЛЬ ДЛЯ ОСЛАБЛЕНИЯ IN VIVO
B1 Использованные штаммы
Вирулентные штаммы
Штамм RE1: материнский штамм дикого типа. Данный штамм растет на холестерине.
Штамм RE1ΔsupAB: делеция гена supAB. Данный штамм неспособен расти на холестерине.
Невирулентный штамм
Штамм 103-: не содержит плазмиды вирулентности размером от 80 до 90 т.п.о. и, как известно, является апатогенным у лошадей (Takai et al. 2000, Infect. Immun. 68: 6840-6847). Данный штамм растет на холестерине.
Штаммы в соответствии с изобретением
Штамм RE1ΔipdAB (RG1341): ген ipdAB делетируется в кластере катаболизма холестерина. Данный штамм не растет на 4-андростен-3,17-дионе (AD).
Штамм RE1ΔipdAB-AD+: бактерии штамма RE1ΔipdAB, адаптированные для роста на AD (если штамм RE1ΔipdAB высевается в высокой концентрации (более чем 106 КОЕ/мл) при AD как единственном источнике углерода, затем могут возникнуть несколько колоний для роста на AD).
Штамм RE1ΔipdABipdAB2 (RG2837): второй набор генов ipdAB (не являющийся частью кластера катаболизма холестерина) также делетируется. Данный штамм не растет на AD.
Штамм ΔfadE30 R. Equi. Рост данного штамма резко нарушен на HIP/HIL и AD.
B2 Культуры Rhodococcus equi для инфекции макрофагов
Различные штаммы Rhodococcus equi, подлежащие тестированию в анализе выживания в макрофагах, выращивали в течение ночи (17 ч) при 37°C и 100 об/мин в питательной бульоне (Difco) для достижения конечной концентрации 1-2×108 КОЕ/мл. Использовали только свежеполученные культуры. После инокуляции макрофагов проводили определение количества живых микроорганизмов (чашечный подсчет) для подтверждения титра инфекционности.
B3 Культуры Rhodococcus для контрольного заражения жеребят
Штаммы RE1 Rhodococcus equi, RE1ΔipdAB и RE1ΔipdAB-AD+, высевали на кровяной агар и инкубировали в течение 24 часов при 37°C. Бактерии собирали 4 мл стерильного изотонического PBS на чашку. Бактериальную суспензию разбавляли стерильным изотоническим PBS для достижения конечной концентрации 4×104 КОЕ/мл. Транспортировку проводили при окружающей температуре; разбавленные культуры использовали в пределах 4 часов после получения. После контрольного заражения проводили определение количества живых бактерий (чашечный подсчет) для подтверждения титра инфекционности. Количества живых суспензий составили соответственно 4,35×104 КОЕ/мл для RE1, 7,1×104 КОЕ/мл для REΔipdAB и 5,8×104 КОЕ/мл для RE1ΔipdAB-AD+.
B4 Системы тестирования
Клеточная линия макрофагов
Линию клеток U937 (моноцитов человека) использовали для тестирования выживания штаммов Rhodococcus equi. Моноциты выращивали в среде RPMI 1640+NaHCO3+NAPYR+глюкоза (среда RPMI 1640), забуференной 10 мМ HEPES и с добавлением 200 МЕ/мл пенициллина и стрептомицина и 10% фетальной телячьей сыворотки (FBS). Клетки выращивали в суспензии при 37°C и 5% CO2.
Жеребята
Использовали 8 жеребят: 7 жеребят в возрасте от 3 до 5 недель и одного жеребенка в возрасте 7 недель (все с кобылами). Жеребят распределяли на 3 группы по 3, 3 и 2 жеребенка, соблюдая равномерное возрастное распределение по группам. Животных содержали в отдельных помещениях. Во время эксперимента жеребята вскармливались материнским молоком и кобыл кормили в соответствии со стандартными процедурами. Доступ к питьевой воде был неограничен.
Во время T=0 всех жеребят подвергали контрольному заражению интратрахеально 100 мл культуры для контрольного заражения штамма RE1, RE1ΔipdAB или RE1ΔipdAB-AD+ (см. выше в данном описании под заголовком «Культуры Rhodococcus для контрольного заражения жеребят») с использованием шприца с иглой, так называемой транстрахеальной инжекции.
B5 Экспериментальные процедуры и параметры
Тест выживания в макрофагах
Для анализа выживаемости в макрофагах моноциты выращивали в течение нескольких дней, как описано выше в настоящей заявке. Культуральную среду замещали свежей культуральной средой и клетки активировали в течение ночи 60 нг/мл форбол-12-миристат- 13-ацетатом (PMA) для индукции их дифференциации в макрофаги.
Дифференцированные клетки осаждали центрифугированием (5 мин при 200×g) и клеточный осадок после центрифугирования ресуспендировали в свежей не содержащей антибиотики среде RPMI 1640 с 10% FBS. Для каждого подлежащего тестированию штамма пробирку, содержащую 10 мл клеточной суспензии приблизительно с 106 клеток/мл, инокулировали Rhodococcus equi при множественности инфекции (MOI) приблизительно 10 бактерий на макрофаг.
Бактерии инкубировали с макрофагами в течение 1 часа при 37°C и 5% CO2. Среду замещали 10 мл среды RPMI 1640 с добавлением 10% FBS и 100 мкг/мл гентамицина и снова инкубировали в течение 1 часа для уничтожения любых внеклеточных бактерий. Макрофаги (с интернализованными R. equi) осаждали центрифугированием (5 мин при 200×g) и клеточный осадок после центрифугирования ресуспендировали в 40 мл среды RPMI 1640, забуференной 10 мМ HEPES, и с добавлением 10% FBS и 10 мкг/мл гентамицина. Данную суспензию делили по 4 культуральным флаконам (по 10 мл в каждый) и инкубировали при 37°C и 5% CO2. После 4, 28, 52 и 76 часов инкубации макрофаги (один культуральный флакон на штамм) осаждали центрифугированием (5 мин при 200×g) и клеточный осадок после центрифугирования дважды промывали в 1 мл не содержащей антибиотик среде RPMI 1640. Наконец, клеточный осадок лизировали 1% Triton X-100 в 0,01 M солевом растворе с фосфатным буфером (PBS) с последующим определением количества живых бактерий (чашечный подсчет).
Контрольное заражение жеребят
1 - Ректальная температура: измерялась за 1 день до контрольного заражения, в день контрольного заражения (непосредственно перед контрольным заражением) и затем один раз в день после контрольного заражения до вскрытия.
2 - Клиническое обследование: в течение 3 недель после контрольного заражения лошадей ежедневно обследовали для выявления клинических признаков.
3 - Патологоанатомическое исследование и бактериология: на 21-й день после контрольного заражения жеребят взвешивали и затем умерщвляли анестезией ксилазином (100 мг/100 кг) и кетамином (500 мг/100 кг) с последующим кровопусканием до смерти. Легкие взвешивали для расчета отношения массы легких к массе тела. Полное патологоанатомическое исследование выполняли с обращением особого внимания на легкие и связанные лимфоузлы. В случае отклонений от нормы патологоанатом, если считал необходимым, брал образцы для гистологического исследования.
Образцы ткани (1 см3) иссекали из семи стандартных участков, представляющих доли каждой половины легких (3 участка на половину легких + добавочная доля); для каждого участка предпочтительно выбиралась пораженная ткань, если она присутствовала. Образцы с зеркальным отражением (два образца эквивалентной доли на каждой половине) объединяли для получения образцов на жеребенка + образец добавочной доли. Каждый (объединенный) образец гомогенизировали, серийно разбавляли и инокулировали на чашки с кровяным агаром и затем инкубировали при 37°C в течение 16-24 часов. Колонии Rhodococcus подсчитывали и выражали в виде КОЕ/мл гомогената. Дополнительные мазки брали из всех участков с патологическими изменениями. Мазки штрихами высевали на чашки с кровяным агаром и затем инкубировали при 37°C в течение 16-24 часов. Rhodococcus equi первоначально идентифицировали по его типичной негемолитической мукоидной морфологии колонии. Дальнейшую идентификацию проводили окрашиванием по Граму, API/Phoenix и/или ПЦР.
B6 Результаты
Выживание в макрофагах
Результаты двух отдельных экспериментов показаны в таблице 1 (в сочетании с фиг.1) и 2 (в сочетании с фиг.2). Результаты показывают, что мутант RE1ΔsupAB способен выживать в макрофагах таким же образом, как материнский штамм дикого типа RE1, указывая на то, что метаболизм холестерина не является существенным для выживания в макрофагах. Это согласуется с признанием того, что метаболизм холестерина, как таковой, не имеет значения для вирулентности. Напротив, выживание в макрофагах штамма RE1ΔipdAB, штамма RE1ΔipdABipdAB2, штамма RE1ΔipdAB-AD+ и штамма 103- было явно снижено. Следует, однако, отметить, что бактерии все же способны выживать в макрофагах, но на значительно сниженном уровне (обычно в концентрации в 100-1000 раз ниже уровня дикого типа). Штамм 103- не содержит плазмиду вирулентности размером от 80 до 90 т.п.о. и, как известно, является авирулентным у лошадей (Takai et al.). Данный штамм 103- не подходит в качестве вакцинного штамма, поскольку он не вызывает защитный иммунный ответ, вероятно, ввиду того, что он не содержит плазмиду вирулентности. В таблице 9 (в сочетании с фиг.4) показаны результаты для мутантного штамма R. equi ΔfadE30, в сравнении со штаммом дикого типа RE1 и штаммом RE1 ΔipdAB. Ясно, что мутант fadE30 имеет характеристику выживания, сравнимую со штаммом ΔipdAB, а также меньше способен к выживанию в макрофагах.
Результаты со штаммами, которые имеют мутацию в опероне, кодирующем протеин, участвующий в активности разрушения метилгексагидроиндандиона пропионата (т.е. штаммы RE1ΔipdAB, RE1ΔipdABipdAβ2, RE1ΔipdAB-AD+ и R. equiΔfadE30), показывают, что данные штаммы менее способны к выживанию в макрофагах. В частности, их способности к выживанию сравнимы со способностями к выживанию апатогенного штамма 103-. Это уже является хорошим показателем адекватного ослабления. Штамм RE1ΔipdAB-AD+ продемонстрировал такой же фенотип макрофагов, как штамм RE1ΔipdAB, указывая на то, что существенным для выживания в макрофагах на уровне дикого типа скорее является интактный оперон ipdAB, а не интактный метаболизм холестерина. Одиночные делеции гена ipdAB (в генном кластере катаболизма холестерина) привели к сниженному выживанию в макрофагах. Дополнительная делеция в копии указанных генов (ipdA и ipdB вне кластера катаболизма холестерина, называемых ipdA2 и ipdB2) не оказывала дополнительного ослабляющего эффекта в тесте с макрофагами.
Учитывая данные результаты, штаммы RE1ΔipdAB и RE1ΔipdAB-AD+ (=штамму RE1ΔipdAB, адаптированному для роста на AD) вводили интратрахеально жеребятам (обычная процедура контрольного заражения) и сравнивали с материнским штаммом дикого типа RE1 для тестирования ослабления in vivo.
Ректальная температура
В таблице 3 (в сочетании с фиг.3) представлены результаты измерения ректальной температуры. Группа 1 представляет собой группу, которая получала штамм RE1 дикого типа. Группы 2 и 3 получали соответственно RE1ΔipdAB и RE1ΔipdAB-AD+. Величины температуры с 3 по 10 день не показаны, поскольку они не выявили никаких значимых изменений от нормальной ректальной температуры. Аномальные величины температуры указаны жирным шрифтом. У двух из трех жеребят (№ 18 и 19), подвергнутых контрольному заражению материнским штаммом RE1 дикого типа, не проявилось явное увеличение значений ректальной температуры с 14 дня после контрольного заражения и далее. Увеличение значений ректальной температуры после 14 дней совпадало с развитием клинических признаков (см. ниже). У жеребенка № 21 (зараженного штаммом RE1ΔipdAB) проявилось незначительное повышение температуры (39,1°C) в 1-й день после контрольного заражения, что, вероятнее всего, не связано с инфекцией Rhodococcus (время инкубации при данной модели контрольного заражения составляет > 7 дней). В последующем у жеребят, подвергнутых контрольному заражению RE1ΔipdAB или RE1 ΔipdAB-AD+, не обнаруживалась повышенная температура.
Клинические признаки после контрольного заражения
Действительно, клинические балльные оценки с 7 по 21-й день показали, что у жеребят № 18 и 19, подвергнутых контрольному заражению материнским штаммом RE1 дикого типа, развились признаки респираторного заболевания с 13-го дня после контрольного заражения и далее. У жеребца № 17 (также из группы контрольного заражения RE1) после контрольного заражения проявились лишь слабые клинические признаки. У жеребят, которые были подвергнуты контрольному заражению мутантным штаммом RE1ΔipdAB и RE1ΔipdAB-AD+, не проявились отчетливые признаки респираторного заболевания. Клинические эффекты последних двух групп были главным образом основаны на несколько увеличенной частоте сердечных сокращений, что также (частично) могло быть вызвано стрессом, связанным с обращением с животными, поскольку она также присутствовала перед контрольным заражением.
Патологоанатомическое исследование
Данные патологоанатомического исследования легких показаны в таблице 4. После контрольного заражения материнским штаммом RE1 дикого типа у жеребят развились признаки респираторного заболевания (в частности, у жеребят № 18 и 19). Жеребята, подвергнутые контрольному заражению мутантными штаммами, оставались здоровыми. Через 21 день после контрольного заражения жеребят умерщвляли и проводили их вскрытие. При вскрытии у всех жеребят, подвергнутых контрольному заражению штаммом дикого типа, оказалось наличие типичной пиогрануломатозной пневмонии, из очага которой был повторно выделен R. equi в виде чистой культуры, и идентичность штамма дикого типа была подтверждена ПЦР. У жеребенка № 18, R. equi дикого типа был также выделен из увеличенного медиастинального лимфоузла.
В легких жеребят, подвергнутых контрольному заражению мутантными штаммами, не проявлялись пневмонические области и Rhodococcus не был выделен за исключением несколько увеличенного бронхиального лимфоузла жеребенка № 20 и здоровой легочной ткани жеребенка № 24. Идентичность данных изолятов была подтверждена как RE1ΔipdAB и RE1ΔipdAB-AD+ соответственно ПЦР и ростом на агаре AD.
B7. Заключение по ЧАСТИ B экспериментов
У штаммов RE1ΔipdAB и RE1ΔipdAB-AD+ отчетливо нарушено выживание в макрофагах и они ослаблены у жеребят. Представляется, что нокаут второй копии гена ipdAB (приводящий к получению RE1ΔipdABipdAB2) не оказывает дополнительного эффекта на тест выживания в макрофагах и, вероятно, также и у жеребят. Комбинированные результаты in vivo и in vitro, полученные со штаммами RE1, RE1supAB, 103-, RE1ΔipdAB и RE1ΔipdAB-AD+, указывают на хорошую корреляцию между уровнем выживания в макрофагах и вирулентностью in vivo для бактерии, относящейся к нокардиоформным актиномицетам. В частности, представляется, что, когда мутантный штамм имеет значительно сниженную способность выживания в макрофагах, обычно примерно от 2 до 3 логарифмов относительно вирулентного материнского штамма, то мутантный штамм значительно ослаблен относительно материнского штамма.
На основании общеизвестного факта, что путь инфекции и факторы вирулентности разделяются среди нокардиоформных актиномицетов, а следовательно, того факта, что Rhodococcus equi обычно используется в качестве модели для исследования Mycobacteriaceae, в частности, в отношении факторов вирулентности, связанных с выживанием в макрофагах и персистенцией в них (см. наряду с другими источниками PNAS, February 6, 2007, vol. 104, no. 6, pp 1947-1952), понятно, что сниженная выживаемость бактерий в результате инактивации гена, который кодирует протеин, участвующий в разрушении метилгексагидроиндандиона пропионата, является генерической для ослабления нокардиоформных актиномицетов.
ЧАСТЬ C: ЭФФЕКТИВНОСТЬ МУТАНТНОЙ БАКТЕРИИ В ЗАЩИТЕ ПРОТИВ ИНФЕКЦИИ ДИКИМ ТИПОМ
C1. Введение
Считается, что фармацевтической композиции, содержащей живые мутантные бактерии в соответствии с настоящим изобретением, присуща антиинфекционная эффективность. Это можно понять, признав, что общеизвестные представления о мутациях метаболизма холестерина отчетливо указывают на то, что бактерии, имеющие мутации метаболизма холестерина, все же имеют антигенную нагрузку, сравнимую с таковой организмов дикого типа. В комбинации с представленными в настоящем изобретении последними данными о том, что мутантные живые бактерии еще способны поступать в макрофаги и выживать в них, не оставляет сомнения то, что сохраняется стимул иммунного ответа. Тем не менее, заявитель провел эксперименты для подтверждения данного мнения. Для этого использовали вакцину, содержащую живого Rhodococcus equi. Пока не существует вакцины, как таковой, против данной бактерии, что делает настоящие эксперименты релевантными. Но важнее то, что данная бактерия была общепризнанна в качестве хорошей модели для других нокардиоформных актиномицетов, в частности Mycobaterium tuberculosis. Другие подтверждающие эксперименты (для получения подтверждающих результатов с охватом полного диапазона нокардиоформных актиномицетов) могут, например, проводиться с Nocardia seriolae (см. параграф C6). Они представляют собой типичных патогенов рыб, против которых нет адекватной вакцины, по тем же причинам, что нет адекватной вакцины против других нокардиоформных актиномицетов, выживающих в макрофагах.
C2. Структура эксперимента
Для данного исследования использовали 16 жеребят в возрасте от 2 до 4 недель. Жеребят делили на 4 группы по 4 жеребенка и вакцинировали перорально 1 мл вакцины, содержащей различные дозы RE1ΔipdAB (составленной в стерильном солевом растворе с фосфатным буфером). Группу 1 вакцинировали 5×109 КОЕ, группу 2 вакцинировали 5×108 КОЕ; группу 3 вакцинировали 5×107 КОЕ и группу 4 оставляли в качестве не вакцинированного контроля. Вакцинацию проводили во время T=O и во время T=2 недели. Во время T=4 недели всех жеребят подвергали контрольному заражению 100 мл культуры вирулентного штамма 85F Rhodococcus equi (содержащего 5×106 КОЕ на дозу 100 мл).
В течение 3-недельного периода после контрольного заражения проводилась клиническая оценка состояния лошадей. Жеребят взвешивали в день первой вакцинации, в день контрольного заражения и в день вскрытия. Через 3 недели после контрольного заражения (или ранее в случае тяжелых клинических признаков) жеребят взвешивали и подвергали эвтаназии и выполняли полное патологоанатомическое исследование с обращением особого внимания на легкие и респираторные лимфоузлы. Легкие взвешивали для расчета отношения массы легких к массе тела. Для исследования брали образцы ткани из всех легочных долей.
C3. Материалы и методы
Тестируемые изделия
Вакцину, содержащую живой Rhodococcus штамма REIΔipdAB, составляли в солевом растворе с фосфатным буфером (PBS). Культуру для контрольного заражения получали следующим образом: штамм 85F Rhodococcus equi высевали на кровяной агар и инкубировали в течение 24 часов при 37°C. Бактерии собирали в 4 мл стерильного изотонического PBS на чашку. Бактериальную суспензию разбавляли стерильным изотоническим PBS для достижения конечной концентрации 4×104 бактерий/л. Транспортировку проводили при окружающей температуре; разбавленную культуру использовали в пределах 4 часов после получения. После контрольного заражения определяли количество живых бактерий для подтверждения титра инфекционности. Титр составил 5,3×104 КОЕ/мл.
Экспериментальные процедуры и параметры
Повторное выделение бактерий выполняли взятием ректальных мазков непосредственно перед каждой вакцинацией и в 0, 1, 2, 3, 6, 10, 14, 15, 16, 17, 20 и 24-й день (после вакцинации). Мазки носовой полости брали у жеребят во время T=O (непосредственно перед первой вакцинацией). Полученные в виде мазков образцы серийно разбавляли в солевом физиологическом растворе и высевали на кровяной агар и инкубировали при 37°C в течение 16-24 часов. Колонии Rhodococcus первоначально идентифицировали типичной морфологией негемолитической мукоидной колонии, подсчитывали и выражали в виде КОЕ/мл. В каждый день выделения случайным методом выбирали три повторных изолята (от трех различных лошадей, если они присутствовали) и использовали для подтверждения идентичности: окрашивание по Граму, API/Phoenix и ПЦР на генах ipdAB.
В течение исследования лошадей ежедневно наблюдал биотехнолог для выявления каких-либо отклонений от нормы в отношении общего состояния здоровья и/или поведения. Начиная за один день до контрольного заражения, лошадей ежедневно обследовали (до вскрытия) с целью выявления клинических признаков. Взвешивание проводили непосредственно перед первой вакцинацией, непосредственно перед первым контрольным заражением и непосредственно перед вскрытием. Таким образом можно было рассчитать прибавку массы тела и отношение массы легких к массе тела.
На 14-20-й день после контрольного заражения (или раньше в случае тяжелых клинических признаков) жеребят умерщвляли анестезией ксилазином (100 мг/100 кг) и кетамином (500 мг/100 кг) и последующим кровопусканием до смерти. Легкие взвешивали для расчета отношения массы легких к массе тела. Выполняли полное патологоанатомическое исследование. Образцы ткани (1 см3) иссекали из семи стандартных участков, репрезентативных для долей каждой половины легких (3 участка на половину легких + добавочная доля); для каждого участка предпочтительно выбирали пораженную ткань, если она присутствовала. Образцы, являющиеся зеркальным изображением (два образца эквивалентной доли на каждой половине), объединяли для получения 3 образцов на жеребенка + образца добавочной доли. Каждый (объединенный) образец гомогенизировали, серийно разводили и инокулировали на чашки с кровяным агаром и затем инкубировали при 37°C в течение 16-24 часов. Колонии Rhodococcus подсчитывали и выражали в виде КОЕ/мл гомогената. Балльную оценку поражения легких пневмонией для каждого животного получали установлением процентной доли консолидации для апикальных (левых+правых), каудальных (левых+правых) и добавочных легочных узлов. Для каждого животного указанные процентные доли складывали для получения цифры балльной оценки одного легкого. Таким образом получается легочная балльная оценка от 0 до 500.
C4. Результаты
Результаты после контрольного заражения суммированы в таблице 7. По суммированным результатам ясно, что у всех четырех контролей развились тяжелые признаки пиогрануломатозной пневмонии, вызванной R. equi в качестве единственного возбудителя. У двух вакцинатов № 4 и 5 имелись признаки, сравнимые с контролями, но у всех других вакцинатов были гораздо более легкие признаки или по существу не было признаков пневмонии. Процент массы легких (объективный количественный показатель пневмонии) подтверждает частичную защиту у большинства вакцинатов.
У вакцинированного жеребенка № 2 имелась пневмония, вызванная неспецифическими бактериями. В действительности, данного жеребенка можно рассматривать как защищенного, поскольку Rhodococcus не выделялся, несмотря на массивное контрольное заражение. Кроме того, от трех вакцинированных жеребят с пневмонией были выделены смешанные инфицирующие возбудители. Данные смешанные инфекции, вероятно, оказывали отрицательное влияние на различные параметры защиты. Хотя защитный эффект вакцины очевиден, эффект зависимости реакции от дозы не наблюдался. В действительности оказалось, что самая низкая доза обеспечивала наилучшие результаты. На основании этого в сочетании с тем фактом, что жеребята имеют незрелую желудочно-кишечную флору, считается, что оптимальная доза составляет от 1×104 до 1×1010 КОЕ.
C5. Заключение по части C экспериментов
Оказалось, что все три пероральные дозы вакцины безопасны для молодых жеребят и вызывали существенную защиту от тяжелого интратрахеального контрольного заражения. В данных экспериментах и гены ipdA, и ipdB оперона ipdAB были удалены из генома бактерии в вакцине. Однако ясно, что другие мутации, вовлекающие один и тот же оперон, могут быть равным образом эффективны. Мутация, которая, например, воздействует только на один из генов ipdA или ipdB, сама по себе может быть в равной мере эффективной. В любом из последних случаев участвующая трансфераза не может быть получена и таким образом достигается получение такого же фенотипа. Действительно, это может быть также выведено из данных Rengarajan (PNAS, June 7, 2005, Vol. 102, No. 23, pp 8327-8332). В данной ссылке представлено релевантное доказательство для другой нокардиоформной актиномицеты, а именно Mycobacterium tuberculosis: инактивация любого одного из ортологичных генов ipd (называемых соответственно rv3551 и rv3552 в M. tuberculosis) приводит к такому же фенотипу.
C6. Подтверждение эффективности в качестве вакцины мутанта Nocardia seriolae
Описанные выше эксперименты для Rhodococcuse equi повторяли для другой актиномицеты, патогена рыб Nocardia seriolae, который вызывает нокардиоз у рыб. Для этого эксперимента использовали штамм дикого типа INS436 и штамм ΔipdAB, как указано выше в настоящем описании в параграфе A13.
Группам из 20 желтохвостов (Seriola quinqueradiata) внутрибрюшинно инъецировали или штамм дикого типа, или мутантный штамм (в различных концентрациях), и одну группу из 20 рыб оставляли в качестве контроля. Рыб наблюдали в течение 2-3 недель для выявления смертности и других клинических реакций с целью оценки ослабления мутантного штамма. В конце указанного периода наблюдения проводили контрольное заражение выживших рыб фиксированной дозой штамма дикого типа для оценки эффективности мутантного штамма в качестве вакцины. Рыб наблюдали в течение еще 2 недель.
Для штамма дикого типа инокулят получали из 2,25×108 кое/мл. Для мутантного штамма он составил 7,85×108 кое/мл (примерно в 3 раза выше). Из этих сходных инокулятов получали двукратные, 20-кратные, 200-кратные и 2000-кратные разведения для использования в исследовании ослабления. Материал для внутрибрюшинного контрольного заражения содержал 1,2×106 кое/мл.
Результаты эксперимента ослабления представлены в таблице 10 (в которой показана смертность в конце периода наблюдения). Из этого становится ясно, что мутантный штамм ослаблен относительно штамма дикого типа. Хотя сходный инокулят для мутантного штамма был в 3 раза выше по кое/мл, смертность значительно ниже в группе, которая получала мутантный штамм. Результаты экспериментов по определению эффективности представлены в таблице 11. Хотя получить полную защиту было невозможно, ясно, что мутант обеспечивает значительную защиту (с учетом того факта, что в контрольной группе 60% рыб погибли в течение периода наблюдения). Даже в группе рыб, которые получали разбавление в 2000 раз, гораздо меньше рыб погибло из-за контрольного заражения Nocardia seriolae дикого типа.
Было показано, что инактивация гена, который кодирует протеин, участвующий в разрушении метилгексагидроиндандиона пропионата, ведет к получению ослабленного и защитного вакцинного штамма во второй нокардиоформной актиномицете. Это является дополнительным подтверждением того, что данное изобретение применимо по широкому диапазону нокардиоформных актиномицетов.



















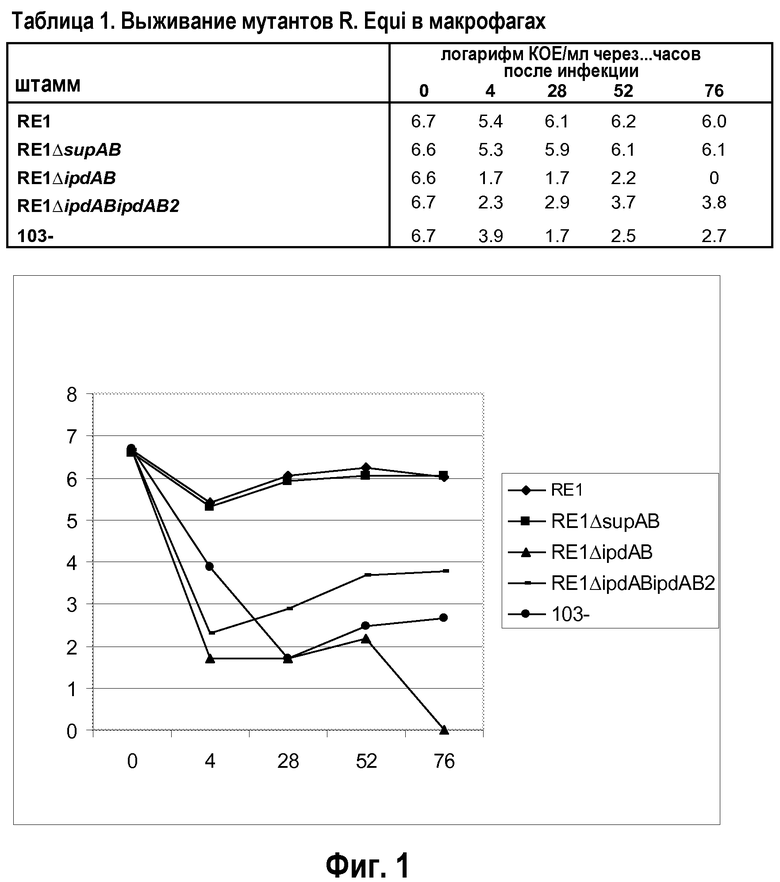
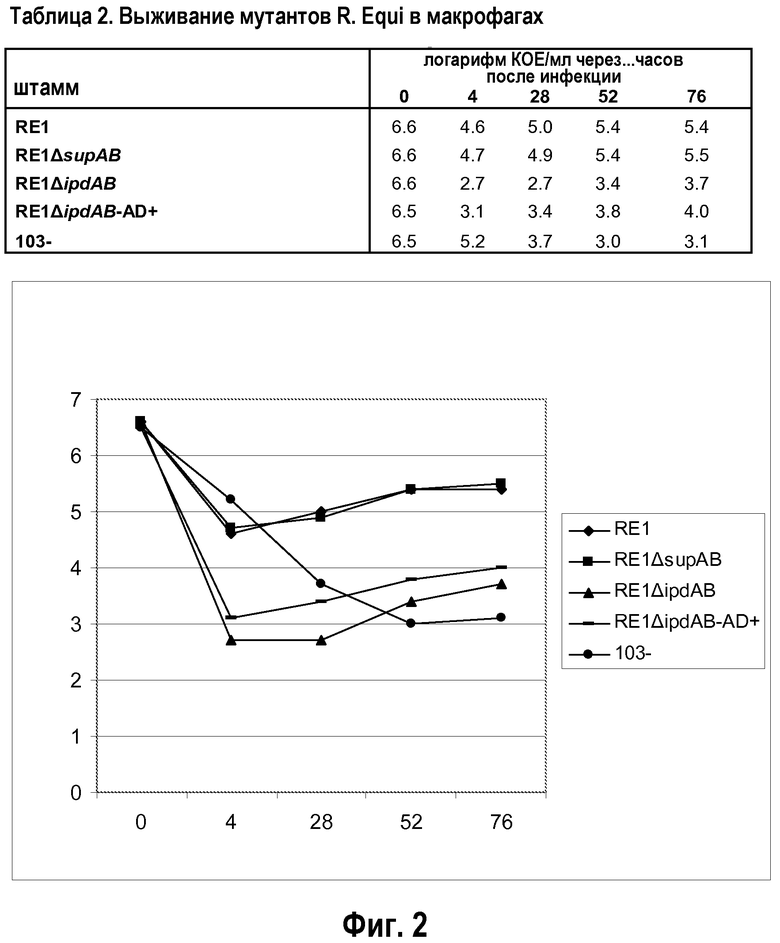

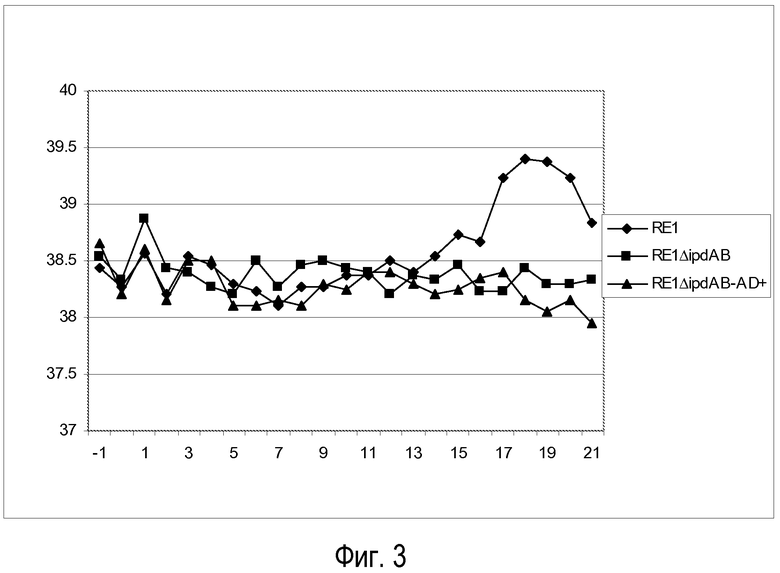

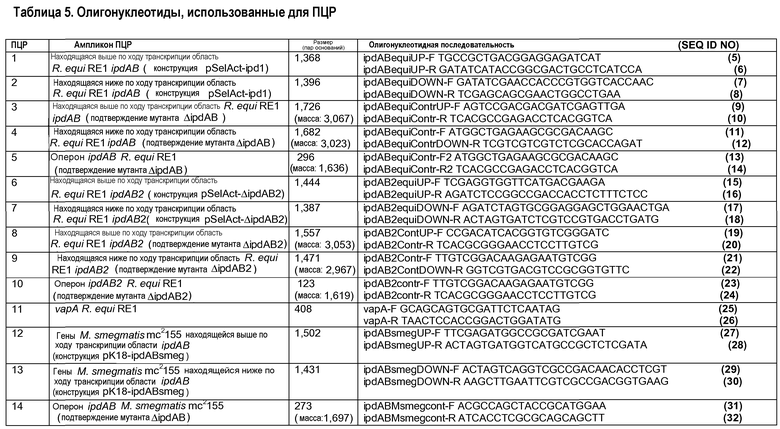
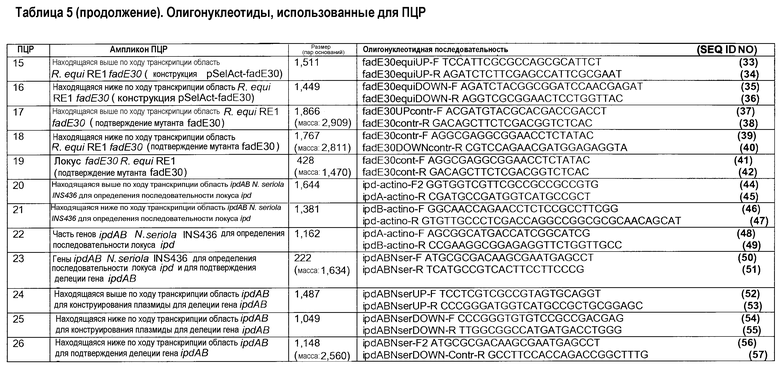
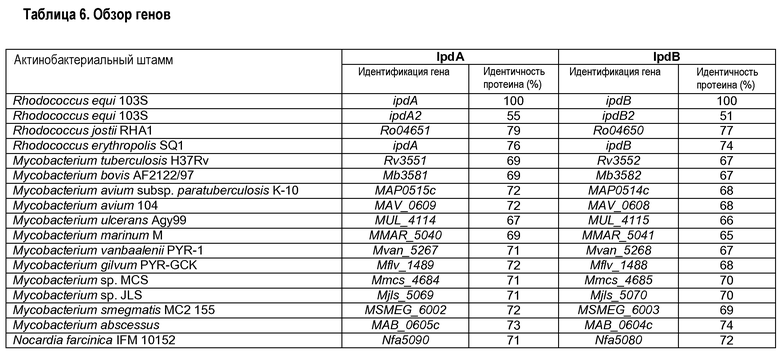
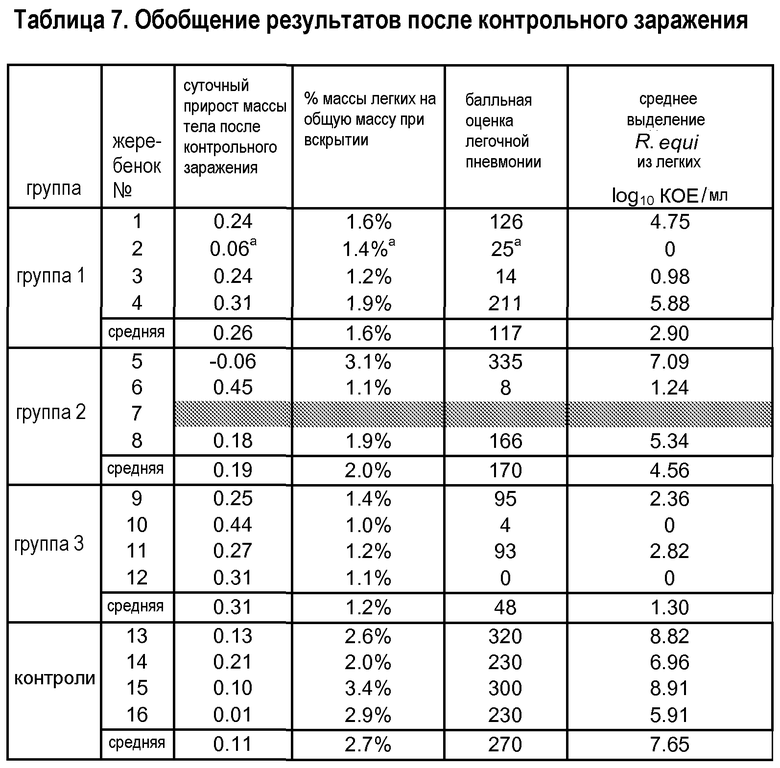
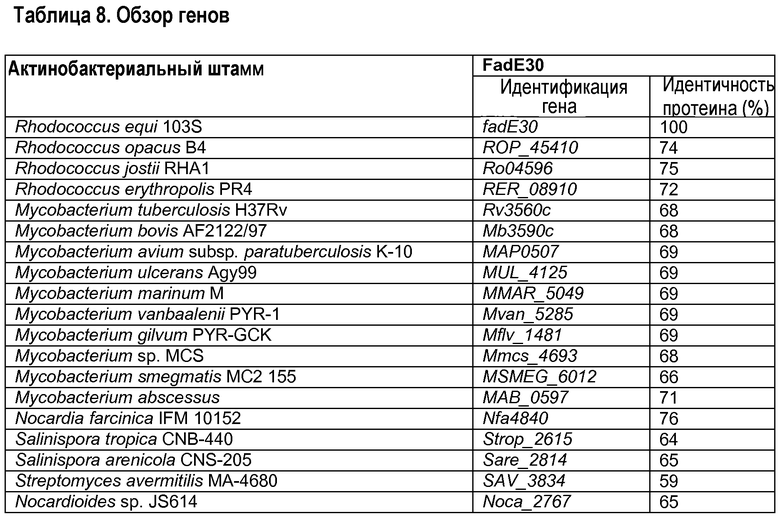
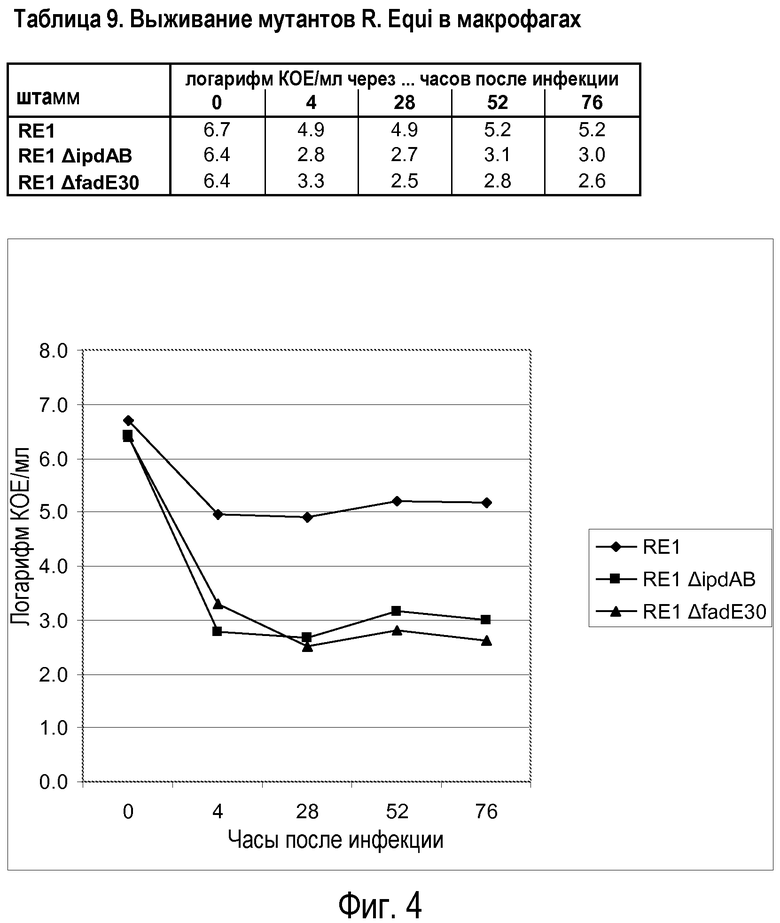

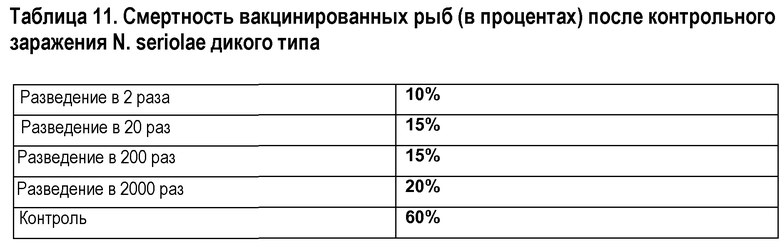
Группа изобретений касается применения живых бактерий, которые относятся к группе нокардиоформных актиномицетов для получения фармацевтической композиции и способа использования такой композиции. Охарактеризовано применение нокардиоформных актиномицетов, обладающих способностью выживать внутри макрофагов животного, причем живые бактерии ослаблены инактивацией гена, который кодирует протеин, участвующий в разрушении метилгексагидроиндандиона пропионата, для получения фармацевтической композиции. Представленный способ обработки животного для защиты его от расстройства, возникающего в результате инфекции бактерией, которая относится к группе нокардиоформных актиномицетов, включает введение животному фармацевтической композиции, содержащей эффективное количество таких бактерий. Предложенные изобретения могут быть использованы для профилактики инфекций, вызываемых нокардиоформными актиномицетами. 3 н. и 9 з.п. ф-лы, 4 ил., 11 табл.
1. Фармацевтическая композиция для защиты животного от расстройства, возникающего в результате инфекции бактерией, которая относится к группе нокардиоформных актиномицетов, обладающих способностью выживать внутри макрофагов животного, содержащая эффективное количество живых бактерий вида нокардиоформных актиномицетов, причем живые бактерии ослаблены инактивацией гена, который кодирует протеин, участвующий в разрушении метилгексагидроиндандиона пропионата, и фармацевтически приемлемый носитель для указанных живых бактерий.
2. Фармацевтическая композиция по п.1, отличающаяся тем, что инактивированы множественные гены в опероне.
3. Фармацевтическая композиция по п.2, отличающаяся тем, что инактивированы гены ipdA и ipdB.
4. Фармацевтическая композиция по п.1, где инактивация осуществляется делецией по меньшей мере одного гена путем делеции немаркированного гена.
5. Фармацевтическая композиция по п.1, отличающаяся тем, что бактерии относятся к семейству Nocardiaceae или Mycobacteriaceae.
6. Фармацевтическая композиция по п.5, отличающаяся тем, что бактерии относятся к родам Rhodococcus, Nocardia или Mycobacterium.
7. Фармацевтическая композиция по п.6, отличающаяся тем, что бактерии относятся к видам Rhodococcus equi, Nocardia seriolae, Mycobacterium tuberculosis, Mycobacterium ulcerans, Mycobacterium bovis или Mycobacterium avium paratuberculosis.
8. Фармацевтическая композиция по п.1, отличающаяся тем, что она представлена в форме, подходящей для перорального введения.
9. Фармацевтическая композиция по п.1, отличающаяся тем, что живые бактерии присутствуют в концентрации от 1×104 до 1×1010 КОЕ на дозу.
10. Фармацевтическая композиция по п.7, отличающаяся тем, что указанные живые бактерии представляют собой бактерии Rhodococcus equi штамма, депонированного в Национальной Коллекции Культур Микроорганизмов Института Пастера в Париже, Франция, под №CNCM I-4108 или №CNCM 1-4109.
11. Применение живых бактерий, которые относятся к группе нокардиоформных актиномицетов, обладающих способностью выживать внутри макрофагов животного, причем живые бактерии ослаблены инактивацией гена, который кодирует протеин, участвующий в разрушении метилгексагидроиндандиона пропионата, для получения фармацевтической композиции для защиты животного от расстройства, возникающего в результате инфекции соответствующей бактерией дикого типа.
12. Способ обработки животного для защиты его от расстройства, возникающего в результате инфекции бактерией, которая относится к группе нокардиоформных актиномицетов, обладающих способностью выживать внутри макрофагов животного, включающий введение животному фармацевтической композиции, содержащей живые бактерии нокардиоформных актиномицетов, причем живые бактерии ослаблены инактивацией гена, который кодирует протеин, участвующий в разрушении метилгексагидроиндандиона пропионата.
| US 7297339 B2, 20.11.2007 | |||
| ПРЕПАРАТ ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ, ВЫЗЫВАЕМЫХ МИКОБАКТЕРИЯМИ, АКТИНОМИЦЕТАМИ, ПРЕЖДЕ ВСЕГО NOCARDIA SP., И ВОЗБУДИТЕЛЯМИ ТОКСОПЛАЗМОЗА | 1994 |
|
RU2136272C1 |

