Настоящее изобретение относится к способу получения генетически модифицированных микроорганизмов, обладающих способностью препятствовать ядерному распаду стероидов, к использованию таких микроорганизмов для накопления стероидов, а также к накоплению таких модифицированных микроорганизмов.
Способность разрушать фитостерины широко распространена среди нокардиоформных актиномицет и требует набора ферментов, разрушающих боковую цепь и структуру стероидного ядра.
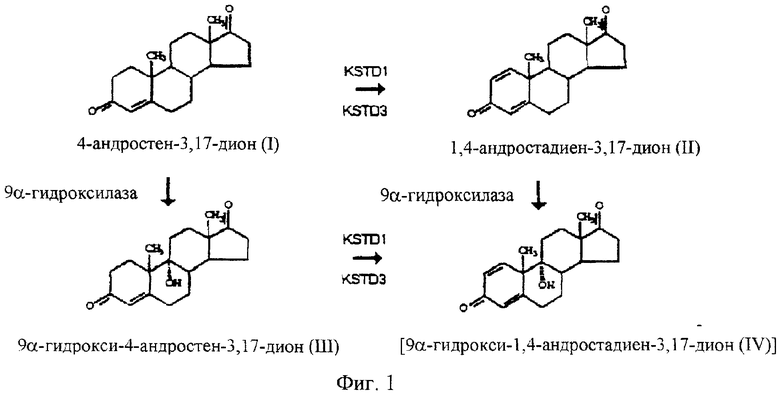
Фермент 3-кетостероид Δ1-дегидрогеназа (KSTD) [4-ен-3-оксостероид: (акцептор)-1-ен-оксидоредуктаза, ЕС 1.3.99.4] участвует в разрыве кольца В стероидного ядра в результате введения двойной связи в положении С1-С2. Более точно, данный фермент участвует в превращении 4-андростен-3,17-диона в 1,4-андростадиен-3,17-дион и 9α-гидрокси-4-андростен-3,17-диона в 9α-гидрокси-1,4-андростадиен-3,17-дион (см. Фигуру 1). Данный фермент идентифицировали у нескольких бактерий: Arthrobacter simplex (Penasse and Peyre, 1968 Rhodococcus. Crit. Rev. Biotech. 14:29-73), Pseudomonas (Levy and Talalay, 1959 J.Biol. Chem. 234:2009-20013; 1959 J. Biol. Chem. 234:2014-2021), Nocardia restrictus (Sih and Bennet, 1962 Biochem. Biophys. Acta 56:587-592), Nocardia corallina (Itagaki с соавт., 1990 Biochim. Biophys. Acta 1038:60-67), Nocardia opaca (Drobnič с соавт., 1993 Biochim, Biophys. Res. Comm. 190:509-515), Mycobacterium fortuitum (Wovcha с соавт., 1979 Biochim. Biophys. Acta 574:471-479) и Rhodococcus erythropolis IMET7030 (Kaufmann с соавт., 1992 J.Steroid. Biochem. Molec. Biol. 43:297-301). KSTD N. opaca охарактеризована в качестве флавопротеина (Lestrovaja с соавт., 1978 Z.Allg. Microbiol. 18:189-196). Полностью охарактеризованы лишь гены, кодирующие KSTD у A.simplex, Comamonas testosteroni и Rhodococcus rhodochrous (Plesiat с соавт., 1991 J.Bacteriol. 173:7219-7227; Molnár с соавт., 1995 Mol. Microbiol. 15:895-905; Morii с соавт., 1998 J.Biochem. 124:1026-1032).
Исключительное ингибирование стероидной 1,2-дегидрогеназы обусловливает накопление 9α-гидрокси-4-андростен-3,17-диона, наилучшего исходного материала для кортикоидного синтеза (Kieslich К., 1985 J.Basic Microbiol. 25: 461-474). 9α-Гидроксиандрогены являются промышленно важными в качестве антиандрогенов, антиэстрогенов и противозачаточных средств. 9α-гидроксигруппа легко дегидратируется до 9(11)-дегидросистемы и является исходной структурой для образования 9α-галогенкортикоидов.
Виды Rhodococcus широко известны своим большим катаболическим потенциалом (Warhurst and Fewson, 1994 Rhodococcus. Crit. Rev. Biotech. 14:29-73; Bell с соавт., 1998 J.Appl. Microbiol. 85:195-210). Некоторые виды Rhodococcus способны расщеплять природные фитостерины, которые являются недорогим исходным материалом для получения биоактивных стероидов (Kieslich К., 1986 Drug Res. 36:888-892). Штаммы Rhodococcus и Mycobacterium, обработанные мутагенами и/или выращенные вместе с ферментными ингибиторами, превращают стерины в 4-андростен-3,17-дион и 1,4-андростадиен-3,17-дион (Martin, 1977 Adv. Appl. Microbiol. 22:29-58).
Хотя клонирование kstQ и экспрессия неактивного белка KSTD R. erythropolis IMET7030 в Escherichia coli уже описаны (Wagner с соавт., 1992 J.Basic Microbiol. 32:65-71, 1992 J.Basic Microbiol. 32:269-277), а нуклеотидная последовательность N.ораса ( с соавт., 1993 Biochem. Biophys. Res. Comm. 190:509-515) (синоним R.erythropolis IMET7030) известна (DDBJ/EMBL/GenBank U59422), о молекулярной характеристике данного гена не сообщалось. Активность KSTD существенна для ядерного распада стероидов, а инактивации гена kstD необходима для накопления промежуточных стероидных соединений. В соответствии с одним аспектом настоящего изобретения создана нуклеотидная последовательность гена kstD R.erythropolis. Белок KSTD1 кодируется нуклеотидами 820-2329 из SEQ ID NO:1.
с соавт., 1993 Biochem. Biophys. Res. Comm. 190:509-515) (синоним R.erythropolis IMET7030) известна (DDBJ/EMBL/GenBank U59422), о молекулярной характеристике данного гена не сообщалось. Активность KSTD существенна для ядерного распада стероидов, а инактивации гена kstD необходима для накопления промежуточных стероидных соединений. В соответствии с одним аспектом настоящего изобретения создана нуклеотидная последовательность гена kstD R.erythropolis. Белок KSTD1 кодируется нуклеотидами 820-2329 из SEQ ID NO:1.
Инактивация генов является мощным инструментом для анализа генной функции и для введения блоков метаболизма. Разрушение гена с помощью нерепликативного вектора, несущего селективный маркер, является обычно используемым способом для генной инактивации. Конструирование штаммов с нужными свойствами достигается через создание метаболического пути, однако может возникнуть необходимость в постепенной инактивации или замене нескольких генов. Это возможно лишь при наличии подходящего метода по введению немаркированных генных делеций или генных замен, что позволяет технологически создавать бесконечные варианты метаболизма без участия множества маркеров. В соответствии со вторым аспектом настоящего изобретения разработана схема постадийной инактивации генов, предпочтительно дегидрогеназных генов, вовлеченных в разрушение стероидов. В частности, настоящее изобретение применимо для инактивации генов, участвующих в накоплении 9α-гидрокси-4-андростен-3,17-диона, вследствие роста микроорганизмов на 4-андростен-3,17-дионе. Предпочтительно инактивируется, по меньшей мере, ген kstD1.
Неожиданно было обнаружено, что разрушение гена kstD1, кодирующего у R.erythropolis SQ1 3-кетостероид Δ1-дегидрогеназу, не приводит к инактивации ядерного распада стероидов. Остаточная активность проявляется из-за присутствия второго фермента. В настоящее время установлено, что для получения штамма с полностью блокированным ядерным распадом стероидов необходимо инактивировать более чем один ген. Предпочтительно второй фермент представляет собой дегидрогеназу, более предпочтительно изоэнзим KSTD. С целью получения возможности разрушения или удаления нескольких генов, предпочтительно может использоваться метод сайт-направленного мутагенеза. Метод по введению немаркированных генных делеций предпочтителен для постадийной инактивации генов KSTD. Полученные генетически модифицированные штаммы не должны содержать гетерологичную ДНК.
В соответствии со вторым предпочтительным вариантом осуществления настоящего изобретения инактивируется, по меньшей мере, ген kstD2. Более предпочтительно, чтобы были инактивированы оба гена kstD1 и kstD2. Еще одним аспектом настоящего изобретения является нуклеотидная последовательность гена kstD2 из R.erythropolis. Белок KSTD2 кодируется нуклеотидами 1-1678 из SEQ ID NO:5.
О методах по введению немаркированных генных делеций в клетки рода Rhodococcus не сообщается. Вместе с тем, методы генного делетирования или генного замещения описаны в отношении некоторых других представителей актиномицет, а именно Streptomyces (Hillemann с соавт., 1991 Nucleic Acid Res. 19:727-731; Hosted and Baltz, 1997 J.Bacteriol. 179:180-186), Corynebacterium (Schäfer с соавт., 1994 Gene 145:69-73) и Mycobacterium (Marklund с соавт., 1995 J.Bacteriol. 177:6100-6105; Norman с соавт., 1995 Mol. Microbiol. 16:755-760; Sander с соавт., 1995 Mol. Microbioi. 16:991-1000; Pelicic с соавт., 1996 Mol. Microbiol. 20:919-125; Knipfer с соавт., 1997 Plasmid 37:129-140). Контрселективные маркеры могут использоваться для скрининга редкого второго рекомбинационного события, приводящего к генной делеции или генной замене. В этом отношении и sacB и rpsL оказались полезными репортерными генами (Hosted and Baltz, 1997 J.Bacteriol. 179: 180-186; Schäfer с соавт., 1994 J.Bacteriol. 172: 1663-1666; Sander с соавт., 1995 Mol. Microbiol. 16: 991-1000; Pelicic с соавт., 1996 Mol. Microbiol. 20: 919-125; Jäger с соавт., 1992 J. Bacteriol. 174: 5462-5465), но могут также использоваться и другие подходящие маркеры. Об использовании rspL у Rhodococcus не сообщалось, но sacB (кодирующий левансахаразу Bacillus subtilis) представляет для этого биологического рода сильный позитивный селективный маркер (Jäger с соавт., 1995 FEMS Microbiol. Lett. 126:1-6; Denise-Larose с соавт., 1998 Appl. Environ. Microbiol. 64:4363-4367).
Левансахараза В.subtilis, кодируемая геном sacB, катализирует гидролиз сахаров и синтез леванов (высокомолекулярные фруктозные полимеры). Экспрессия sacB в присутствии сахарозы является летальной для Rhodococcus. Биохимическая основа токсичности левансахаразного действия на сахарозу до сих пор неизвестна. Летальность (т.е. присутствие или отсутствие сахарозы), обусловленная геном sacB, может, следовательно, использоваться в качестве контрселективного маркера. Контрселекция в данном контексте означает, что экспрессия данного маркера является летальной, а не повышающей резистентность, как, например, в случае селективных маркеров (например, резистентные маркеры).
Контрселекция нужна для отбора таких мутантов, которые подвергаются второму рекомбинационному событию, теряя при этом маркер sacB и вводя требуемую мутацию. Преимущество данной системы заключается в том, что в течение отбора будут выживать лишь наиболее подходящие мутанты. По сравнению с системой, в которой используется только один селективный маркер, контрселекция позволяет избежать трудоемкого процесса скрининга, так как утрачивается маркер резистентности, который необходим для системы с одним селективным маркером.
Преимущество немаркированной мутации заключается в том, что она допускает повторное введение мутаций в тот же самый штамм. Чужеродная ДНК (векторная ДНК) удаляется в процессе введения данной мутации. Вновь введенная векторная ДНК, предназначенная для введения второй мутации, не может по этой причине интегрироваться в сайт предыдущей мутации (путем гомологичной рекомбинации между векторными ДНК). Интеграция несомненно произойдет, если векторная ДНК все еще присутствует в данной хромосоме и даст начало большому числу ложнопозитивных продуктов интеграции. Данная система позволяет использовать отдельный ген антибиотика для введения бесконечного множества мутаций. Немаркированная мутация делает возможным легко использовать ее в промышленности, поскольку отсутствие гетерологичной ДНК позволяет легко удалять ферментационный бульон.
Инактивация гена в результате делеции гена позволяет создавать устойчивые, неревертирующие мутанты. Преимущественно небольшие гены (<500 п.н.) легче инактивируются с помощью генной делеции по сравнению с разрушением гена в результате одиночной рекомбинационной интеграции. Мутагенез, осуществляемый с помощью генной делеции, может также применяться для инактивации кластера в несколько генов из генома. Способ мутагенеза путем генного делетирования может также применяться для замены гена (например, замена гена дикого типа на мутантный ген).
Предпочтительным штаммом для мутагенеза катаболических генов стероидных дегидрогеназ является Rhodococcus erythropolis. Вместе с тем и у других видов возможно возникновение немаркированной делеции гена kstD1 и/или kstD2, осуществляемой с помощью генетической конъюгации, если молекулярная организация у них та же (или подобная), что и у R.erythropolis SQ1. Данные виды предпочтительно относятся к роду Rhodococcus, но могут использоваться также и такие близкородственные виды как Nocardia, Mycobacterium и Arthrobacter.
Генная инактивация у Rhodococcus затруднена, когда происходят нелогичные рекомбинационные события, приводящие к случайной геномной интеграции данного мутагенного вектора (Desomer с соавт., 1991 Mol. Microbiol. 5:2115-2124; Barnes с соавт., 1997 J.Bacteriol. 179:6145-6153), феномен, с которым авторы изобретения столкнулись при попытке разрушить ген kstD1 у R.erythropolis SQ1. Нелогичная рекомбинация также является хорошо известным феноменом у некоторых медленно растущих видов Mycobacterium (McFadden, 1996 Mol. Microbiol. 121:205-211). Показано, что конъюгативный перенос плазмиды из Е.coli S17-1 в Rhodococcus минимизирует случайную интеграцию (Powell and Archer, 1998 Antinie van Leeuwenhoek 74:175-188). Доказана возможность кокъюгативной мобилизации плазмид из штамма S17-1 Е.coli во многие различные штаммы коринеформных бактерий и в Rhodococcus fascians DSM20131 (Schäfer с соавт., 1990 J.Bacteriol. 172:1663-1666; Jäger с соавт., 1995 FEMS Microbiol. Lett. 126:1-6). В соответствии с настоящим изобретением конъюгативный перенос мутагенного вектора, несущего ген sacB в качестве контрселективного маркера, поэтому применяли для введения немаркированной генной делеции в стероидный катаболизм у R.erythropolis SQ1.
В качестве дополнительного варианта осуществления настоящего изобретения введение второго генного инактивационного события выполняют с использованием тех же методов, которые рассмотрены в Примерах для гена kstD2. Для дальнейшей генной инактивации могут опять использоваться эти же методы или же УФ-облучение или такие химические средства, как, например, нитрогуанидин или диэпоксиэтан. Методы введения генных мутаций, осуществляемые таким образом, хорошо известны в данной области.
Хорошо также известны и методы по созданию носителей, используемых в протоколе мутагенеза (Sambrook с соавт., Molecular Cloning: a Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, новейшее издание). Кроме того, методики по сайт-направленному мутагенезу, лигированию добавочных последовательностей, ПЦР, секвенированию ДНК и созданию подходящих экспрессионных систем в настоящее время также хорошо известны в данной области техники. Части или всю ДНК, кодирующие нужный белок, можно создать в результате синтеза с использованием стандартных твердофазных методик, преимущественно для включения сайтов рестрикции, облегчающих лигирование.
Модификации и варианты данного способа по введению разрушенных генных мутаций или немаркированной генной делеции, также как трансформирование и конъюгирование, будут понятны специалистам в данной области из предшествующего подробного описания настоящего изобретения. Подразумевается, что такие модификации и варианты находятся в рамках настоящей заявки.
В соответствии с другим аспектом настоящего изобретения микроорганизмы, обладающие множеством инактивированных генов, могут использоваться для накопления стероидных промежуточных соединений. Накопленный продукт преимущественно представляет собой 9α-гидрокси-4-андростен-3,17-дион. Данное исходное вещество может зависеть от наличия ферментных генов, которые инактивируются. Подходящими исходными веществами являются, например, фитостерины или 4-андростен-3,17-дион. Предпочтительным исходным веществом является 4-андростен-3,17-дион.
Преимущество представленного способа состоит в том, что можно обеспечить высокоэффективное превращение исходного стероида в искомый накапливаемый продукт. Его выход может превысить 80%, предпочтительно более чем 90% и часто достигает величины почти 100%.
Еще один аспект настоящего изобретения относится к генетически модифицированным микроорганизмам с множеством инактивированных генов, которые участвуют в стероидном разложении. В частности, данные гены являются дегидрогеназами. Предпочтительно инактивируются, по меньшей мере, ген kstD1 или kstD2. Более предпочтительной является инактивация обоих генов kstD1 и kstD2. Предпочтительными являются микроорганизмы, относящиеся к роду Rhodococcus. Наиболее предпочтительным является штамм Rhodococcus erythropolis RG1-UV29.
Штаммы микроорганизмов Rhodococcus erythropolis RG1-UV29 и Rhodococcus erythropolis RG1 были депонированы в Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSMZ), Mascheroder Weg 1b, D-38124 Braunschweig, Германия под каталожными номерами соответственно DSM 13157 и DSM 13156. Данное депонирование осуществляли в соответствии с Будапештским Договором.
Специалистам должно быть понятно, каким образом использовать методы и вещества для создания микроорганизмов, утрачивающих способность к разрушению стероидного ядра, описанные в настоящем документе и на которые дана ссылка. Подобным образом можно инактивировать множество генов, кодирующих несколько других ферментов, разрушающих стероидное ядро.
Нижеследующие примеры являются иллюстративными для настоящего изобретения и никоим образом не должны интерпретироваться в качестве ограничивающих рамки настоящего изобретения.
Описание чертежей
Фигура 1
Схематическое изображение ядерного распада стероидов у R.erythropolis SQ1. Указано расположение изоэнзимов KSTD1 и KSTD2 фермента 3-кетостероид Δ1-дегидрогеназы (KSTD).
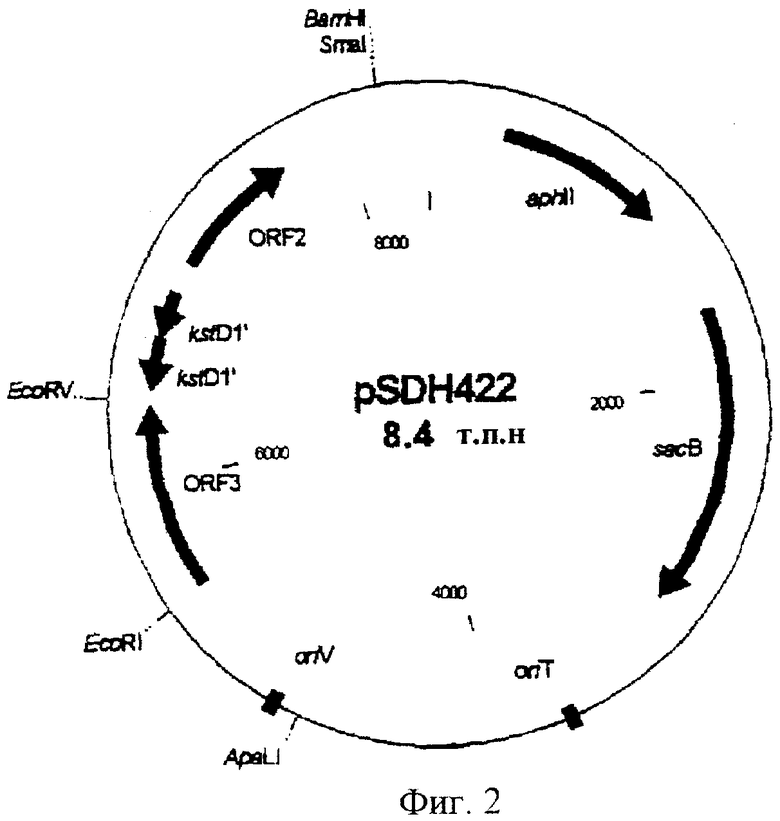
Фигура 2
Схематическое изображение мутагенного вектора pSDH422 вместе с контрселективным маркером sacB, используемого для создания штамма Rhodococcus erythropolis RG1, с 1062 п.н. немаркированной делецией гена kstD1. ORF2 и ORF3 представляют собой фланкирующие гены гена kstD1 R.erythropolis SQ1.
Фигура 3
Схематически представленная общая молекулярная организация гена kstD1 у штамма дикого типа R.erythropolis SQ1 и после включения pSDH422 при помощи однократного кроссинговера соответственно по правому (штамм SDH422-3) и левому локусу-мишени (штамм SDH422-4) kstD1. Окно-вставка: Саузерн-анализ с использованием в качестве зонда kstD1, где хромосомная ДНК R.erythropolis дикого типа (дорожка 1) расцеплена BamHI, штамма SDH422-3 (дорожка 2), SDH422-4 (дорожка 3) и двух отдельных делеционных мутантов kstD1 (дорожки 4 и 5).
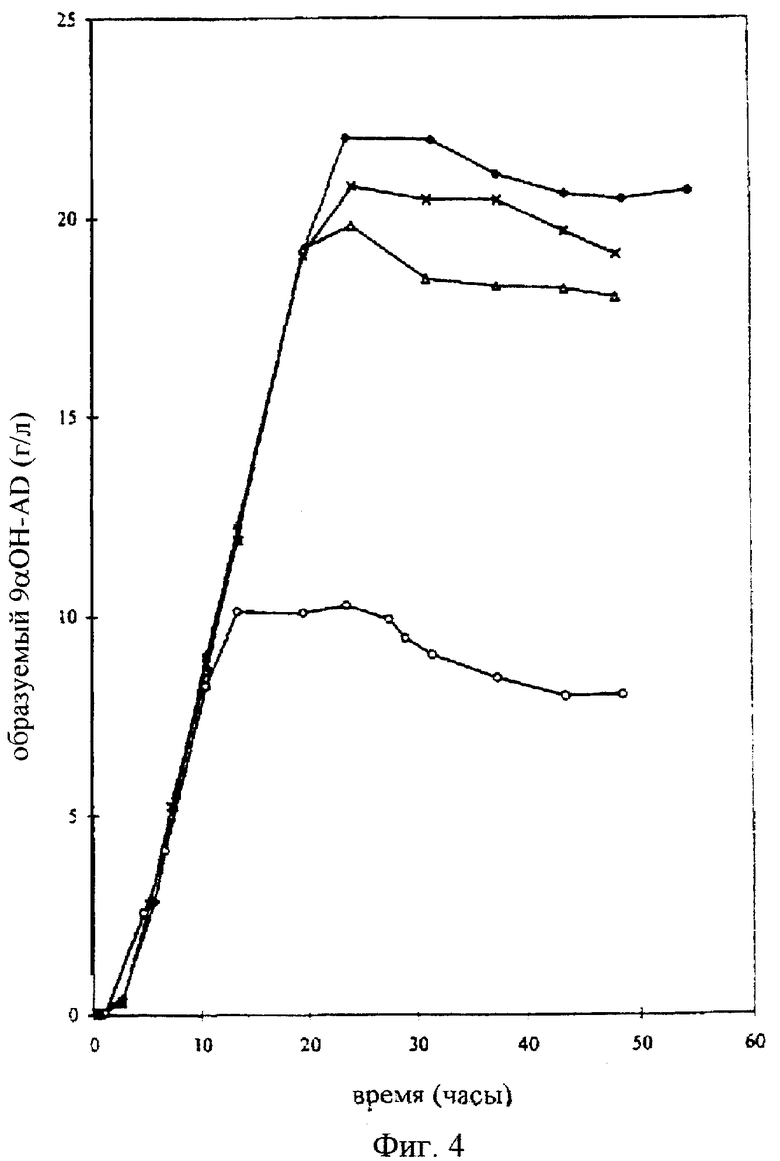
Фигура 4
Биоконверсия 4-андростен-3,17-диона в 9α-гидрокси-4-андростен-3,17-дион в 6-литровой культуре Rhodococcus erythropolis SQ1 UV29. 10 г/л AD (-O-) и 20 г/л AD (-Δ-, , -Х-).
, -Х-).
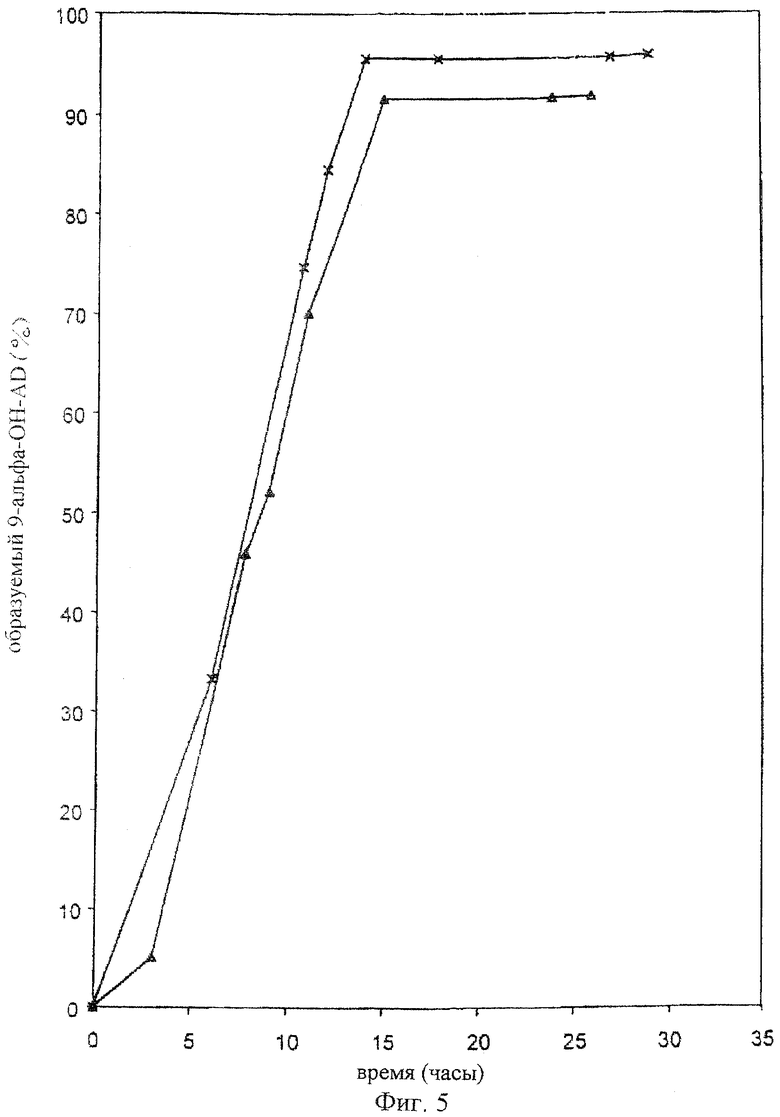
Фигура 5
Биоконверсия 4-андростен-3,17-диона в 9α-гидрокси-4-андростен-3,17-дион в 6-литровой культуре Rhodococcus erythropolis RG8. 10 г/л AD(-Δ-, -X-).
Примеры
Пример 1
Характеристика kstD1
Вырожденный олигонуклеотидный зонд kstD [5' ttcgg (c/g)gg (c/g)ac(c/g)tc(c/g)gc(c/g)tactc(c/g)gg(c/g)gc(c/g)tc(c/g) atctgg] (SEQ ID NO: 2) создавали из выровненных N-концевых частей известных последовательностей белка KSTD A.simplex, С. testosteroni и N.ораса. После расщепления общей ДНК R.erythropolis SQ1 рестриктазой BglII определяли размер фрагментов с помощью центрифугирования в градиенте сахарозы. Саузерн-анализ при 68°С (отмывка в жестких условиях с помощью 2×SSC в течение 2×15 мин и 0,1×SSC в течение 2×10 мин) полученных фракций дает 6 т.п.н. фрагмент ДНК, гибридизующий с меченным диоксигенином олигонуклеотидным зондом kstD. Данный фрагмент лигируют в BglII-сайт Rhodococcus-E.coli челночного вектора pDA71 (Dabs с соавт., 1995 Development of improved Rhodococcus plasmid vectors and their use in cloning genes of potential commercial and medical importance, p.129-135. В: Proceedings of the Ninth Symposium on the Actinomycetes, Москва, Россия) и субклонируют в расщепленную с помощью BamHI плазмиду pBluescript II KS (Stratagene) (pSDH200).
По результатам анализа рестрикционного картирования авторы изобретения заключили, что лишь один EcoRV-сайт присутствует в указанном 6 т.п.н. фрагменте, делящий его на одинаковые по размеру фрагменты, приблизительно 3 т.п.н. Саузерн-анализ свидетельствует, что EcoRV-фрагмент pSDH200 размером около 2,9 т.п.н. содержит последовательности, гомологичные kstD-олинуклеотиду. Нуклеотидное секвенирование обнаруживает открытую рамку считывания в 1,533 н. (kstD1, смотрите SEQ ID NO:1), кодирующую KSTD1, что демонстрируется при помощи гетерологичной экспрессии у Escherichia. Дальнейшее нуклеотидное секвенирование позволяет обнаружить две ORF - 1,533 н. (ORF1) и 627 н. (ORF2), кодирующие предполагаемые белки соответственно из 510 аминокислот и 208 аминокислот.
Пример 2
Штамм с делецией kstD1.
Был сконструирован мутагенный вектор, который содержит фрагмент хромосомной ДНК R.erythropolis SQ1 с делецией kstD1. 1062 п.н. BsmI-фрагмент pSDH200, кодирующий большую внутреннюю часть KSTD1, делетируют для создания pSDH200)BsmI. Для того чтобы создать мутагенный вектор 2724 п.н. SmaI/EcoRI-фрагмент из pSDH200)BsmI, заякоривающий 468 п.н. остаток kstD1 и его фланкирующие области, клонируют в SmaI/EcoRI-сайт pK18mobsacB (pSDH422, см. Фигуру 2). Вектор pSDH422, кодирующий устойчивость к канамицину, отбирают для интеграции данного мутагенного вектора в хромосому, а заякоривающий ген sacB из В.subtills, предназначенный для контрселекции, вводят в E.coli S17-1 и мобилизуют в R.erythropolis SQ1 с помощью конъюгации следующим образом. Клетки реципиентного штамма R.erythropolis SQ1 распределяют на LBP-агар с добавлением 30 мкг·м-1 налидиксовой кислоты и растят их в течение 5 дней. Мутагенный вектор pSDH422 вначале вводят в E.coli S17-1 посредством трансформации.
Трансформанты (приблизительно 1000 на чашку), выращенные в течение ночи на селективной среде (25 мкг·мл-1 канамицина), инкубируют при комнатной температуре в течение последующих 24 ч. Колонии обоих штаммов, Rhodococcus и E.coli, ресуспендируют в конечном объеме 1,5 мл LBP (1% бактопептон (Difco), 0,5% дрожжевой экстракт (BBL) и 1% NaCl). Смешивают аликвотные пробы по 750 мкл от каждого штамма и мягко осаждают центрифугированием. Полученный осадок клеток ресуспендируют в 1 мл LBP, а клетки распределяют на неселективный LBP-агар в виде 250 мкл-овых аликвот. После выращивания в течение ночи при 30°С полученный совместно выращенный материал ресуспендируют в 2 мл LBP-среды и 100 мкл-овые аликвоты распределяют на LBP-агар с добавлением канамицина (200 мкг·мл-1) и налидиксовой кислоты (30 мкг·мл-1). Через 3 дня появляются трансконъюгаты R.erythropolis SQ1. Все полученные трансконъюгаты Rhodococcus, устойчивые к канамицину (kanr), были чувствительны к сахарозе (sucs); рост после получения реплики на LBPS-агаре (1% бактопептон, 0,5% дрожжевой экстракт, 1% NaCl, 10% сахарозы) с добавлением 200 мкг·мл-1 канамицина не происходил.
Саузерн-анализ (Фиг.3) микроорганизмов дикого типа (дорожка 1: одиночная полоса приблизительно 4500 п.н.) и двух транскокъюгатов, SDH422-3 (дорожка 2: две полосы приблизительно 2900 и 10100 п.к.) и SDH422-4 (дорожка 3: две полосы приблизительно 4000 и 9000 п.н.) обнаруживает, что оба типа штаммов имеют одну копию pSDH422, включенную в локус-мишень благодаря единственному рекомбинационному событию. Генная делеция kstD1, полученная в штамме SDH422-3 R.erythropolis после ночного роста в неселективных условиях, становится видимой после пассирования культуры клеток на селективной среде, т.е. на LBSP-агаре.
Делетирование гена kstD1 достигается после ночного роста в неселективных условиях и последующего пассирования культуры клеток на селективной среде, т.е. на LBPS-агаре. ПЦР колоний с использованием kstD1-праймеров (прямой праймер [5' gcgcatatgcaggactggaccagcgagtgc] (SEQ ID NO:3); обратный праймер [5' gcgggatccgcgttacttcgccatgtcctg] (SEQ ID NO:4)) на 9 sucr/kans-колонияx дает 6 ПЦР-продуктов с размером фрагментов 468 п.н., включающих делетированный ген kstD1. Делецию гена подтверждали в Саузерн-анализе при 60°С (отмывка в жестких условиях с помощью 2×SSC в течение 2×5 мин и 0,1 SSC в течение 2×5 мин) с использованием в качестве зонда гена kstD1, случайным образом меченного диоксигенином. 4,5 т.п.н. kstD1-фрагмента ДНК дикого типа, полученный после расщепления хромосомной ДНК BamHI, уменьшался до 3,4 т.п.н. у мутантов с генной делецией, свидетельствуя о предполагаемой делеции фрагмента kstD1-ДНК размером 1062 п.н. Этот полученный штамм назвали R.erythropolis RG1.
Пример 3
Инактивация стероидного Δ1-дегидрогенирования в УФ-мутагенезе.
Клетки (2·108 КОЕ·мл-1) R.erythropolis RG1 поздней экспоненциальной фазы, выращенные в 10 мМ глюкозоминеральной среды (К2НРО4 4,65 г·л-1, NaH2PO4·H2O 1,5 г·л-1, NH4Cl 3 г·л-1, MgSO4·7H2O г·л-1, Vishniac-микроэлементы, рН 7,2), подвергают ультразвуковой обработке в течение короткого периода времени для получения одиночных клеток. Разбавленные (104) образцы распределяют на глюкозоминеральную агаровую среду и облучают в течение 15-20 сек с помощью УФ-лампы (Philips TAW 15W) на расстоянии 27 см, что в среднем приводит к 95% гибели клеток. Через 4 дня инкубирования колонии помещают на минеральную агаровую среду, содержащую 4-андростен-3,17-дион (0,5 г·л-1, растворенного в DMSO (50 мг·мл-1)). Через 3-4 дня отбирают дефектных по росту на стероиде мутантов для дальнейшей характеристики. Для отбора штаммов, блокированных по Δ1-дегидрогенированию, полученную мутантную популяцию скринируют на наличие мутантов с дефектным ростом на 4-андростен-3,17-дионе, способных расти на минеральной агаровой среде, содержащей 1,4-андростадиен-3,17-дион (0,25 г·л-1). На этом основании сделано заключение, что соответствующий ген инактивирован. Этот ген назвали kstD3 (см. Фигуру 1).
Пример 4
Микробиологическое 9α-гидроксилирование 4-андростен-3,17-диона при помощи УФ-мутантов Rhodococcus erythropolis UV-29.
Rhodococcus erythropolis SQ1 UV-29 представляет собой УФ-мутант, который способен превращать 4-андростен-3,17-дион (AD) в 9α-гидрокси-4-андростен-3,17-дион (9αOH-AD) с концентрацией от 10 до 20 г/л.
Данную конверсию осуществляют с использованием нижеследующего способа.
В 250 мл-овой колбе Эрленмейера, содержащей 75 мл стерильной среды (1,5% дрожжевой экстракт, 1,5% глюкоза; рН 7,0), на роторной качалке (180 об./мин) в течение 24 часов выращивают при 28°С Rhodococcus erythropolis SQ1 UV-29 (прекультура). В 10-литровый ферментер, с 6 литрами в нем стерильной культуральной жидкости (1,5% дрожжевой экстракт, 1,5% глюкоза, 0,01% пеногасящего агента пропиленгликоля; рН 7,5) инокулируют прекультуру (1%) и инкубируют при 28°С в течение 16 часов при барботировании стерильным воздухом, при этом культуру перемешивают для стимуляции глубинного роста. Затем вводят суспензию из 60 граммов 4-андростен-3,17-диона в 300 мл полисорбата (0,1%). После чего продолжают аэробную инкубацию с перемешиванием при 28°С в течение 24 часов. Полученные культуральные образцы экстрагируют метанолом (70%) и перед определением стероидного состава с помощью ЖХВД фильтруют через 0,45 мкм-ный глухой фильтр. Эту же операцию осуществляют in triplo (трижды) с удвоенной концентрацией AD в количестве 20 г/л, добавляя 120 г AD вместо 60 г.
Как показано на фигуре 4, за 24 часа 10-20 г/л 4-андростен-3,17-диона почти полностью превращаются в 9α-гидрокси-4-андростен-3,17-дион (приблизительно 93% от общего количества 4-андростен-3,17-диона).
Пример 5
Характеристика kstD2.
Генную библиотеку R.erythropolis RG1 вводят посредством электротрансформации в компетентный штамм R.erythropolis RG1-UV29. Колонии получают репликой на минеральной агаровой среде, содержащей 4-андростен-3,17-дион (0,5 г/л) в качестве единственного источника углерода и энергии. Комплементацию фенотипа штамма RG1-UV29 оценивают через три дня инкубирования при 30°С. Колонии, растущие на минеральной агаровой среде с 4-андростен-3,17-дионом, культивируют в LBP-среде для выделения плазмидной ДНК, которую в дальнейшем повторно вводят в штамм RG1-UV29 для контроля истинной комплементации. Плазмиду pKSD101, выделяемую из трансформанта, который демонстрирует восстановление роста на минеральной среде с 4-андростен-3,17-дионом, вводят в Е.coli DH5α для последующего анализа. В pKSD101 идентифицируют вставку, приблизительно 6,5 т.п.н. ДНК фототрофных бактерий, и подвергают аналитическому рестрикционному картированию, субклонированию и последующим экспериментам по комплементации. 3,6 т.п.н. фрагмент ДНК EcoRI все еще сохраняет способность восстанавливать фенотип штамма RG1-UV29 и поэтому его субклонируют в pBluescript(II)KS (pKSD105) для нуклеотидного секвенирования. Анализ последовательности нуклеотидов обнаруживает присутствие большой открытой рамки считывания (ORF) размером 1,698 н., кодирующей предлагаемый белок из 565 аминокислот с расчетной молекулярной массой 60,2 кДа. Данную ORF обозначили kstD2 (SEQ ID NO:5) (которая идентична ранее описанной kstD3 - см. Пример 3). Выведенная аминокислотная последовательность kstD2 демонстрирует высокое сходство с известной 3-кетостероид Δ1-дегидрогеназой (KSTD), указывая, что kstD2 кодирует второй фермент KSTD у R.erythropolis RG1.
Пример 6
Штаммы с делецией kstD2.
Штамм RG7 R.erythropolis является мутантным штаммом, полученным из дикого типа штамма SQ1 R.erythropolis, содержащего единственную генную делецию kstD2. Штамм RG8 R.erythropolis создали в результате последовательной делеции в штамме дикого типа SQ1 R.erythropolis двух генов, кодирующих активность 3-кетостероид Δ1-дегидрогеназы, т.е. kstD1 и kstD2. Штамм RG8 получают в результате делеции гена kstD2 из генома, мутантного по делеции kstD1 штамма RG1 R.erythropolis. Метод, используемый для делеции гена kstD2, аналогичен методу, описанному для делетирования гена kstD1 в примере 2, за тем исключением, что использовали разные мутагенные векторы (pKSD201 вместо pSDH422).
Мутагенный вектор pKSD201 создают следующим образом. Делетирование внутреннего 1,093 п.н. фрагмента ДНК из гена kstD2 расщеплением MluI и последующее самолигирование pKSD105 позволяет создать pKSD200. 2,4 т.п.н. фрагмент EcoRI из pKSD200, содержащий мутировавший ген kstD2, лигируют в EcoRI-обработанную pK18mobsacB, создавая тем самым pKSD201. Плазмиду pKSD201 вводят в Е.coli S17-1 и мобилизуют путем конъюгации в штамм SQ1 R.erythropolis (чтобы создать штамм RG7), или - в штамм RG1 (чтобы создать штамм RG8). Трансконъюгаты (sucs kanr), полученные в результате целевого включения pKSD201 в данный геном, появляются через 3 дня роста при 30°С. Делецию kstD2 осуществляют в течение ночного роста одного выбранного трансконъюгата (sucs kanr) в неселективных условиях (т.е. на LBP-среде) и последующего помещения на селективную агаровую среду LBPS. ПЦР колоний осуществляют на 15 колониях sucr/kans с помощью kstD2-праймеров (прямой праймер [5' gcgcatatggctaagaatcaggcaccc] (SEQ ID NO:6); обратный праймер [5' gcgggatccctacttctctgctgcgtgatg] (SEQ ID NO:7)), получая 4 ПЦР-продукта с размером фрагмента 0,6 т.п.н., включающих оставшуюся часть гена kstD2. В Саузерн-анализе, с использованием в качестве зонда dig-меченного гена kstD2, хромосомная ДНК дикого типа, обработанная с помощью Asp718, и эти 4 полученных мутанта подтверждают делецию kstD2: 2,4 т.п.н. фрагмент Asp718 ДНК дикого типа уменьшается до 1,3 т.п.н. у мутантных штаммов.
Пример 7
Микробиологическое 9α-гидроксилирование 4-андростен-3,17-диона с помощью штамма RG8 R.erythropolis.
Rhodococcus erythropolis RG8 представляет собой двойной мутант по делеции kstD1 и kstD2, который способен превращать 4-андростен-3,17-дион (AD) в 9α-гидрокси-4-андростен-3,17-дион (9αOH-AD) с концентрацией 10 г/л.
Данное превращение осуществляют с использованием нижеследующего способа:
В 250 мл-овой колбе Эрленмейера, содержащей 75 мл стерильной среды (1,5% дрожжевой экстракт, 1,5% глюкоза; рН 7,0), выращивают в течение 24 часов на роторной качалке при 28°С Rhodococcus erythropolis RG8 (прекультура). В 10-литровый ферментер, с 6 литрами в нем стерильной культуральной жидкости (1,5% дрожжевой экстракт, 1,5% глюкоза, 0,01% пеногасящего агента пропиленгликоля; рН 7,5), инокулируют прекультуру (1%) и инкубируют при 28°С в течение 16 часов при барботировании стерильным воздухом, при этом культуру перемешивают для стимуляции глубинного роста. Затем вводят суспензию из 60 граммов 4-андростен-3,17-диона в 300 мл полисорбата (0,1%). После чего продолжают аэробную инкубацию с перемешиванием при 28°С в течение 24 часов. Образцы отбирают в течение данного процесса. Данные образцы экстрагируют метанолом (70%) и перед определением стероидного состава с помощью ЖХВД фильтруют через 0,45 мкм-ный глухой фильтр. Данный процесс осуществляют дважды.
Как показано на фигуре 5, за 15 часов 10 г/л 4-андростен-3,17-диона почти полностью превращаются в 9α-гидрокси-4-андростен-3,17-дион (приблизительно 92-96% от общего количества 4-андростен-3,17-диона).
| название | год | авторы | номер документа |
|---|---|---|---|
| ШТАММ Rhodococcus erythropolis ВКПМ Ac-1740 ДЛЯ ПОЛУЧЕНИЯ 9 АЛЬФА-ГИДРОКСИСТЕРОИДОВ | 2007 |
|
RU2351645C1 |
| РЕКОМБИНАНТНЫЕ МИКОБАКТЕРИАЛЬНЫЕ ШТАММЫ-ПРОДУЦЕНТЫ И ИХ ИСПОЛЬЗОВАНИЕ ДЛЯ ПОЛУЧЕНИЯ 22-ФУНКЦИОНАЛИЗИРОВАННЫХ СТЕРОИДОВ | 2019 |
|
RU2767610C2 |
| ПРИМЕНЕНИЕ БАКТЕРИЙ, ОТНОСЯЩИХСЯ К ГРУППЕ НОКАРДИОФОРМНЫХ АКТИНОМИЦЕТОВ, ДЛЯ ПОЛУЧЕНИЯ ФАРМАЦЕВТИЧЕСКОЙ КОМПОЗИЦИИ И СПОСОБ ИСПОЛЬЗОВАНИЯ ТАКОЙ ФАРМАЦЕВТИЧЕСКОЙ КОМПОЗИЦИИ | 2010 |
|
RU2543663C2 |
| МИКРОБИОЛОГИЧЕСКИЙ СПОСОБ ПОЛУЧЕНИЯ 1,2-ДЕГИДРИРОВАННЫХ ПРОИЗВОДНЫХ Δ-3-КЕТОСТЕРОИДОВ РЯДА АНДРОСТАНА В ВОДНО-ОРГАНИЧЕСКИХ СРЕДАХ | 2010 |
|
RU2447154C1 |
| БИОТЕХНОЛОГИЧЕСКАЯ ОПТИМИЗАЦИЯ МИКРООРГАНИЗМОВ | 2020 |
|
RU2832723C2 |
| СПОСОБ ПОЛУЧЕНИЯ АНДРОСТА-1,4-ДИЕН-3,17-ДИОНА ИЗ СТЕРИНОВ РАСТИТЕЛЬНОГО И ЖИВОТНОГО ПРОИСХОЖДЕНИЯ | 2005 |
|
RU2297455C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ 1-БУТАНОЛА | 2006 |
|
RU2429295C2 |
| СПОСОБ ПОЛУЧЕНИЯ СТЕРОИДНЫХ ПРОИЗВОДНЫХ ВОССТАНОВЛЕНИЕМ ОКСОСТЕРОИДНЫХ СОЕДИНЕНИЙ ИЛИ ОКИСЛЕНИЕМ ГИДРОКСИСТЕРОИДНЫХ СОЕДИНЕНИЙ С ИСПОЛЬЗОВАНИЕМ ГИДРОКСИСТЕРОИДНОЙ ДЕГИДРОГЕНАЗЫ | 2007 |
|
RU2426791C2 |
| СПОСОБ ПОЛУЧЕНИЯ СТИГМАСТ-4-ЕН-3-ОНА ИЗ БЕТА-СИТОСТЕРОЛА | 2011 |
|
RU2472857C1 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ЧЕТЫРЕХУГЛЕРОДНЫХ СПИРТОВ | 2006 |
|
RU2394913C2 |

Изобретение относится к биотехнологии. Предложен способ конструирования генетически модифицированных штаммов микроорганизмов, разрушающих стероиды, в котором инактивируются множество генов, таких, например, как гены стероидных дегидрогеназ, вовлеченных в разрушение стероидного ядра. Примером таких генов может быть ген kstD1. Штаммы с множеством инактивируемых генов, кодирующих ферменты, разрушающие стероиды, позволяют более эффективно накапливать промежуточные стероидные соединения. Предпочтительным продуктом накопления является 9α-гидрокси-4-андростен-3,17-дион. 7 з.п. ф-лы, 5 ил.
| US 5004695 А, 02.04.1991 | |||
| СПОСОБ ПОЛУЧЕНИЯ АНДРОСТА-1,4-ДИЕН-3,17-ДИОНА | 1993 |
|
RU2039824C1 |

