Изобретение относится к биотехнологии и медицине и касается аттенуированных бактерий В.pertussis, содержащих нокаутную мутацию в гене dnt и продуцирующих нетоксическую, иммунологически активную, токсоидную форму коклюшного токсина, их применению в качестве живой и инактивированной вакцин и вектора для поливалентной вакцины. Интраназальная иммунизация мышей живыми и инактивированными аттенуированными бактериями B.pertussis обеспечивает эффективную и пролонгированную защиту от заражения вирулентным штаммом возбудителя коклюша.
Уровень техники
Вакцинация против коклюша в течение многих лет проводится коклюшной корпускулярной вакциной (ККВ). ККВ обеспечивает защиту от заболевания более 85% привитых детей. Тем не менее, из-за возникающих разнообразных побочных реакций после вакцинации в ряде развитых стран, а также у нас в стране, наблюдалось снижение охвата детского населения прививками. Как следствие, увеличилась заболеваемость не только новорожденных детей, но и детей школьного возраста и взрослых, особенно, находящихся в контакте с детьми в организованных коллективах [Centers for Disease Control and Prevention. Pertussis United States, 1997-2000. Morb. Mortal. Wkly. Rep. 2002, 51:73-76].
Детальное изучение иммунобиологических свойств возбудителя коклюша позволило установить роль конкретных факторов вирулентности в патогенезе заболевания, формировании иммунного ответа и разработать менее реактогенные бесклеточные коклюшные вакцины (БКВ). Показано, что ведущим фактором, определяющим патогенез, клинику заболевания и обеспечивающим формирование иммунитета, является коклюшный токсин (КТ).
Используемые в настоящее время бесклеточные KB значительно различаются по составу антигенов коклюшного микроба, методам их очистки и обезвреживания KT [Mooi, F.R., I.H. van Loo, A.J.King. Emerg. Infect. Dis. 2001, 7:.526-28]. При производстве БКВ используются трудоемкие и дорогостоящие технологические операции, а их хранение требует специальных условий. Несмотря на то, что бесклеточные KB, по сравнению с ККВ, обладают более низкой реактогенностью, отмечены случаи неврологических осложнений и смертей новорожденных, связанные с вакцинацией БКВ [Чупринина Р.П., Озерецковский Н.А., Алексеева И.А., 2001, 3:19-22]. Эффективность поликомпонентных БКВ и высокий стабильный уровень защитной активности ККВ подтверждает важную роль в защите против коклюша белков, связанных с наружной мембраной клетки, в частности филаментозного гемагглютинина, пертактина, агглютиногенов 2 и 3.
Опыт применения моно- и поликомпонентных БКВ показал, что их низкая реактогенность обусловлена, в частности, практически полным отсутствием в препаратах термолабильного или дермонекротического эндотоксина (ДНТ). Этот факт позволяет предположить, что бактерии, не продуцирующие эндотоксин, наиболее перспективны для производства безвредных вакцин, интраназальное введение которых будет нивелировать и отрицательный эффект липополисахарида. Наблюдения за профилактической эффективностью противококлюшных вакцин в течение последних 20 лет свидетельствуют о том, что иммунитет после вакцинации БКВ сохраняется не более 4-х лет, после иммунизации ККВ 5-6 лет, а после перенесенного заболевания продолжительность напряженного иммунитета достигает 10 лет [Guiso.N, Njamkepo Е., Vié Ie Sage.F, et al. Vaccine. 2007, 25:1390-13907; Mattoo S., James D. Clinical Microbiology Reviews. 2005, 18:326-382; Purdy K..W, Hay J..W, Botteman M..F, Ward J..I. Clin Infect Dis. 2004, 39:29-30].
Таким образом, максимальная длительность и напряженность иммунитета, скорее всего, могут быть обеспечены в результате использования для массовой вакцинации (ревакцинации) детей и взрослых живой вакцины интраназального применения, созданной на основе полноценных аттенуированных бактерий В.pertussis. Возможность разработки такой вакцины продемонстрирована в ряде исследований [Mielcarek N., Debrie A.S., Raze D. et al. PLoS Pathogens. 2006, 2:0662-70; Stevenson A, Roberts M. FEMS Immunol Med Microbiol. 2003, 37:121-128]. Показано также, что бактерии В.pertussis могут быть использованы в качестве вектора для конструирования поливалентных живых вакцин, предназначенных для интраназальной иммунизации. Интраназальная иммунизация животных бактериями, продуцирующими гетерологичные антигены, сопровождается выработкой IgG и IgA как против филаментозного гемагглютинина B.pertussis, так и против слитых с ним гетерологичных антигенов. При этом антитела обнаруживаются не только в сыворотке крови, но и в выделениях генитального тракта мышей [Mielcarek N., Debrie A.S., Raze D. et al. PLoS Pathogens. 2006, 2:0662-70; Stevenson A., Roberts M. FEMS Immunol. Med.Microbiol. 2003, 37:121-128].
Направленная аттенуация бактерий B.pertussis путем конструирования мутантов B.pertussis, продуцирующих токсоид КТ, описана ранее [Синяшина Л.Н., Нечаева Е.В., Амелина И.П., Каратаев Г.И., ЖМЭИ, 2009]. Однако одним из существенных недостатков указанного аттенуированного штамма является способность продуцировать дермонекротический токсин, кодируемый геном dnt, являющийся одним из факторов патогенности бактерий рода Bordetella, определяющим реактогенность вакцины. Как отмечено выше, дермонекротический токсин, кодируемый последовательностью хромосомы dnt, является второй важной мишенью для снижения токсичности бактерий рода Bordetella. Гены dnt присутствуют в хромосомах практически всех представителей рода Bordetella и почти не отличаются друг от друга [Kashimoto Т., Katahira J., Cornejo W.R. et al. Infect Immun. 1999, 67:3727-32; Parkhill J., Sebaihia M., Preston A. et al. Nat. Genet. 2003, 35:32-4; Pullinger G.D., Adams Т.Е., Mullan P.B. et al. Infect Immun. 1996, 64:4163-71; Schmidt G., U.M. Goehring, J. Schirmer, et al. J. Bio Chem. 1999, 274:31875-3188]. Исключение составляют некоторые штаммы, так называемого комплекса IV В. bronchiseptica, выделенные от людей и утратившие ген dnt [Diavatopoulos D.A., Craig A. et al. PLOS Pathogens. 2005, 1:373-383]. Синтез ДНТ находится под контролем bvg-локуса. Bvg- клетки B.pertussis не продуцируют ДНТ [Stibitz S., Miller J.F. In V.L.Miller, J.B.Kaper, D.A.Portney, R.R.Isberg (ed.), Molecular genetics of bacterial pathogenesis. American Society for Microbiology, Washington, D.C., 1994, 407-422]. Нет данных, указывающих на участие ДНТ в формировании иммунного ответа. Дермонекротический, термолабильный или летальный мышиный токсин представляет собой белковую молекулу, состоящую из одной полипептидной цепи с молекулярной массой 140 кДа. ДНТ является эндотоксином, локализован в цитоплазме и клеточной стенке бактерий, инактивируется нагреванием при 56°С [Horiguchi Y, Nakai Т, Kume К.. Infect. Immun. 1991, 59: 1112-1116; Horiguchi Y., Senda Т., Sugimoto N. et al. J.Cell Sci. 1995, 108:3243-51; Kashimoto Т, Katahira J, Cornejo W.R. et al. Infect Immun. 1999, 67:3727-32; Pullinger G.D., Adams Т.Е., Mullan P.B. et al. Infect Immun. 1996, 64:4163-71]. При внутрикожном введении животным ДНТ вызывает некрозы. После внутривенного и внутрибрюшинного введения животным ДНТ наблюдаются значительные некрозы, кровоизлияния, отеки и дегенеративные изменения в печени, почках, селезенке. Сообщалось, что ДНТ ингибирует активность щелочной фосфатазы и уменьшает накопление коллагена 1 типа в остеобласто-подобных клетках. Предполагается, что ДНТ может влиять на способность клеток к дифференциации, стимулирует синтез ДНК и белка, что является следствием модификации GTF- связывающего белка RhoA эукариотических клеток [Horiguchi Y., Nakai Т., Kume К. Infect. Immun. 1991, 59:1112-1116; Horiguchi Y., Senda Т., Sugimoto N. et al. J. Cell Sci. 1995, 108:3243-51]. Картирована область ферментативного центра и участка, ответственного за связывание ДНТ с рецептором эукариотической клетки [13, 20]. Роль ДНТ в патогенезе коклюша изучена недостаточно. Предполагается, что ДНТ принимает участие в приживлении и колонизации коклюшного микроба [Diavatopoulos D.A., Craig A. et al. PLOS Pathogens. 2005, 1:373-383; Horiguchi Y., Senda Т., Sugimoto N. et al. J. Cell Sci. 1995, 108:3243-51].
Известен аттенуированный штамм Bordetella pertussis, содержащий мутацию в гене ptx, делецию гена dnt и гетерологичный ген ampG, продукт которого значительно снижает количество трахеального цитотоксина (WO/2007/104451). В экспериментах на новорожденных мышах показано, что штамм способен индуцировать иммунный ответ у лабораторных животных. Однако защитные свойства аттенуированного тройного мутанта Bordetella pertussis оценивались по способности бактерий вирулентного штамма размножаться в носоглотке и легких мышей после их интраназальной вакцинации аттенуированными бактериями, что не соответствует тесту, рекомендованному Всемирной Организацией здравоохранения и используемому в большинстве стран, включая Россию, для оценки протективности коклюшной вакцины. Несоответствие условий анализа протективности международным стандартам (в значительной степени заниженные дозы заражения бактериями вирулентного штамма, способ заражения и метод оценки протективности) не позволяют оценить пригодность известного аттенуированного тройного мутанта Bordetella в качестве кандидата живой вакцины.
Раскрытие изобретения
Авторами настоящего изобретения впервые предложен штамм аттенуированных бактерий B.pertussis, не проявляющих активности дермонекротического токсина и продуцирующих генетически измененную иммунологически активную нетоксичную, токсоидную форму коклюшного токсина, способный обеспечивать при интраназальном способе вакцинации эффективный иммунитет против вирулентного штамма бактерий B.pertussis.
Другой аспект изобретения касается живой вакцины против возбудителя коклюша B.pertusis на основе аттенуированного двойного мутанта B.pertusis KS. Еще один аспект изобретения касается инактивированной цельноклеточной (корпускулярной) вакцины на основе аттенуированного штамма, содержащего мутантные гены ptx и dnt.
Еще один аспект изобретения касается использования аттенуированного штамма Bordetella в качестве вектора при создании поливалентной вакцины для профилактики или лечения инфекций, вызываемых различными патогенами, в частности возбудителем туберкулеза (Mycobacterium tuberculosis).
В еще одном аспекте изобретение обеспечивает штамм аттенуированных бактерий В.pertussis, содержащих мутантные гены ptx и dnt, деропонированный в Коллекции штаммов бактерий Федерального государственного бюджетного учреждения «Научно-исследовательский институт эпидемиологии и микробиологии имени почетного академика Н.Ф.Гамалеи Министерства здравоохранения и социального развития Российской Федерации» № 171/331 B.p KS-4.
Краткое описание чертежей
Фиг.1а. Схема конструирования и структура рекомбинантной плазмиды pKS1.
Стрелками обозначены последовательности генов в составе рекомбинантных плазмид: cat - серая, dnt - черная, bla - незакрашенная, tet - косая штриховка. Ломаными линиями обозначены положения праймеров Дф (Cga tat tcc agt cgt tcg tg), Др (Tcg ttc aac аса tcc aag gc), использованных для амплификации и клонирования фрагмента последовательности гена dnt и идентификации последовательности гена cad в составе рекомбинантных плазмид pKS2 и pKS3 и хромосомы рекомбинанта KS B.pertussis, содержащей мутантный ген dnt.
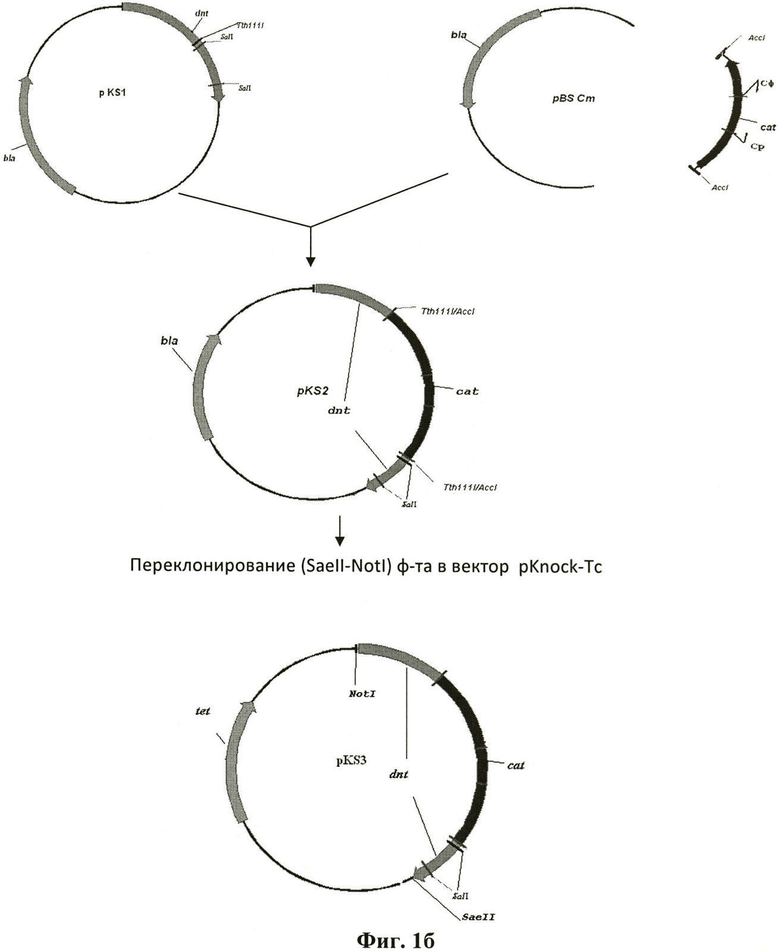
Фиг.1б. Схема конструирования и структура рекомбинантных плазмид pKS2 и pKS3.
Стрелками обозначены последовательности генов в составе рекомбинантных плазмид: cat - серая, dnt - черная, bla - незакрашенная, tet - косая штриховка. Ломаными линиями обозначены положения праймеров Дф (Cga tat tcc agt cgt tcg tg), Др (Tcg ttc aac аса tcc aag gc), использованных для амплификации и клонирования фрагмента последовательности гена dnt и идентификации последовательности гена cad в составе рекомбинантных плазмид pKS2 и pKS3 и хромосомы рекомбинанта KS B.pertussis, содержащей мутантный ген dnt.
Фиг.2. Схема аллельного обмена между копиями фрагментов мутантных (плазмида и pKS3) и нативных (хромосома B.pertussis 5) последовательностей гена dnt. Структура фрагмента хромосомы KS B.pertussis, содержащего мутантный ген dnt.
Ломаными линиями обозначены положения праймеров  использованных для амплификации и клонирования фрагмента последовательности гена dnt и идентификации последовательности гена cad в составе рекомбинантных плазмид pKS2 и pKS3 и хромосомы рекомбинанта KS B.pertussis, содержащей мутантный ген dnt. Длинные линии на верхней части рисунка фланкируют участки последовательности гена dnt, участвующие в гомологичной рекомбинации в процессе аллельного обмена.
использованных для амплификации и клонирования фрагмента последовательности гена dnt и идентификации последовательности гена cad в составе рекомбинантных плазмид pKS2 и pKS3 и хромосомы рекомбинанта KS B.pertussis, содержащей мутантный ген dnt. Длинные линии на верхней части рисунка фланкируют участки последовательности гена dnt, участвующие в гомологичной рекомбинации в процессе аллельного обмена.
Фиг.3. Электрофорез продуктов ПЦР ДНК B.pertussis KS.

Осуществление изобретения
Настоящее изобретение впервые обеспечивает аттенуированные бактерии B.pertussis, не проявляющие активности дермонекротического токсина, коклюшного токсина и общей токсической активности, но сохранивших способность связывания с рецепторами эукариотической клетки (мутацией не затронута область гена dnt, кодирующая участок связывания токсина с рецептором эукариотической клетки) и продуцирующих генетически измененную иммунологически активную нетоксичную, токсоидную форму коклюшного токсина (оперон ptx), причем аттенуированные двойные мутанты B.pertussis являются нереактогененными и обеспечивает эффективную защиту лабораторных животных, вакцинированных интраназально, против интрацеребральной инфекции вирулентным штаммом B.pertussis.
В одном из аспектов изобретение направлено на аттенуированные бактерии В.pertussis KS, несущие нокаутную мутацию в гене dnt, сконструированные на основе описанного ранее мутанта В.pertussis 5, продуцирующего токсоид КТ (Синяшина Л.Н., Нечаева Е.В., Амелина И.П., Каратаев Г.И., ЖМЭИ, 2009). Согласно современной классификации бактерии исходного вирулентного штамма В.pertussis 173 и мутанта В.pertussis 5 относятся к генотипу ptxS1A4, соответствующему генотипу большинства штаммов бактерий B.pertussis, используемых на протяжении последних 60 лет для производства противококлюшных вакцин в РФ и ряде зарубежных стран [Borisova. О; Kombarova S.Y.; Zakharova, N.S. et al. Clin vaccine Immunol. 2007; 14:234-238; Audrey J King, Tamara van Gorkom, Jeroen L.A.; Penningsye at al. BMC Genomics. 2010; 11:196]. В качестве бактерий используемых для конструирования двойных мутантов, содержащих описанные мутации в опероне ptx и гене dnt, могут быть использованы не только бактерии B.pertusis 173 и B.pertusis 5, имеющие генотип ptxS1A4, но и бактерии с другими генотипами, в частности ptxS1 A1, выделяемые от больных коклюшем в настоящее время.
Согласно предложенному изобретению для мутагенеза была выбрана область гена dnt, кодирующая токсический центр белка. В отличие от описанной ранее делеции [Kashimoto Т., Katahira J. Cornejo W.R. et al. Infect Immun. 1999, 67:3727-32; Pullinger G.D., Adams Т.Е., Mullan P.B. et al. Infect Immun. 1996, 64:4163-71] используемая согласно изобретению инсерция последовательности гена cad в Tth1111 сайт гена dnt не затрагивает область гена, кодирующего участок связывания ДНТ с рецептором эукариотической клетки.
Конъюгативная плазмида pKS3, несущая фрагмент последовательности dnt, содержащий инсерцию гена cad и не реплицирующаяся в клетках микроорганизмов рода Bordetella, получена для конструирования аттенуированных бактерий B.pertussis, содержащих мутантные гены ptx и dnt, согласно изобретению (Фиг.1б, пример 2). Бактерии B.pertussis, содержащие мутации ptxS1 и нокаутную мутацию dnt в хромосоме, сконструированы в результате аллельного обмена между природной копией оперона dnt в составе хромосомы реципиента и мутантной копией в составе плазмиды pKS3 (фиг.2, пример 3). Плазмиду pKS3 передавали в бактерии B.pertussis в результате скрещивания S17 Е.coli/pKS3XKmRRifR и В. pertussis 5. Локализация инсерции последовательности в гене dnt подтверждена путем проведения ПЦР на ДНК хромосомы трансконъюганта с праймерами Дфв (Gct gct ggg caa tgt gtt cg) - Cp (Ctc aat gta cct ata acc aga. ccg ttc) (фиг.3, пример 3).
Таким образом, в соответствии с изобретением сконструированы бактерии B.pertussis KS, содержащие мутантный оперон ptx, инсерцию гена kan, расположенную вблизи мутантного оперона в участке, не оказывающем влияния на экспрессию оперона ptx или транспорт субъединиц рекомбинантного белка через бактериальную стенку, и инактивированную инсерцией гена cad последовательность гена dnt.
В еще одном аспекте изобретение обеспечивает штамм аттенуированных бактерий B.pertussis, содержащих мутантные гены ptx и dnt, деропонированный в Коллекции штаммов бактерий Федерального государственного бюджетного учреждения «Научно-исследовательский институт эпидемиологии и микробиологии имени почетного академика Н.Ф.Гамалеи Министерства здравоохранения и социального развития Российской Федерации» под № 171/331 B.p KS-4.
Специфическая токсическая активность КТ, синтезируемого аттенуированными бактериями B.pertussis согласно изобретению, была оценена в экспериментах по определению степени лейкоцитоза и гистаминсенсибилизации у мышей и эффекта образования кластеров клеток СНО, результаты представлены в табл.1.
Из данных, представленных в табл.1, видно, что количество КТ, продуцируемого рекомбинантными бактериями B.pertussis KS, практически не отличается от количества КТ, продуцируемого бактериями вирулентного штамма B.pertussis 173. Специфическая токсическая активность КТ, синтезируемого рекомбинантными бактериями B.pertussis KS согласно изобретению, напротив, значительно отличается от специфической токсической активности КТ, синтезируемого бактериями вирулентного штамма B.pertussis 173. Аттенуированные бактерии B.pertussis, синтезирующие мутантный КТ, вызывают более низкий лейкоцитоз (ЛСА 21×103 против 63×103), гистаминсенсибилизацию у мышей (ГСД50 2.4×109 против 6,9×103) и эффект образования кластеров клеток СНО при значительно большей концентрации токсоида, чем КТ, продуцируемый природными вирулентными бактериями (одинаковый кластер-эффект при концентрации токсина меньшей в 104 раз, чем токсоида).
Определение токсической активности ДНТ рекомбинантных клеток B.pertussis KS определяли в опытах на животных (морских свинках и мышах), результаты представлены в табл.2.
Данные, представленные в табл.2, демонстрируют, что аттенуированные рекомбинантные бактерии B.pertussis KS, в отличие от бактерий исходного вирулентного штамма 173 и бактерий B.pertussis 5, не вызывают гибели мышей при внутрибрюшинном введении препаратов и некрозов в кожной пробе на морских свинках и кроликах.
Общую токсичность аттенуированных бактерий B.pertussis KS согласно изобретению, характеризующую действие на организм мышей всех токсинов возбудителя коклюша, в том числе и цитотоксина, определяли в соответствии с требованиями РФ после введения инактивированной прогреванием бактериальной суспензии лабораторным мышам внутрибрюшинно [Инструкция по отбору, проверке и хранению штаммов B.pertussis для изготовления коклюшной вакцины и коклюшного компонента ассоциированных вакцин. 1987, Москва]. Результаты теста изменения массы тела мышей (табл.3) показали, что в отличие от инактивированных бактерий вирулентного штамма B.pertussis 173, живые аттенуированные бактерии B.pertussis KS при внутрибрюшинном введении 10 МОЕ (Международный оптический стандарт микробных клеток, 1 МОЕ соответствует 1 миллиарду микробных клеток) в 0,5 мл 0,85% раствора хлорида натрия проявляли низкую токсичность.
Защитные свойства бактерий определяли по проценту выживаемости мышей при иммунизации их определенным количеством микробных клеток в соответствии с международными стандартами, принятыми в РФ [Инструкция по отбору, проверке и хранению штаммов В.pertussis для изготовления коклюшной вакцины и коклюшного компонента ассоциированных вакцин. 1987, Москва], результаты представлены в табл.4.
При интраназальной иммунизации мышам вводили от 4 до 500 млн живых микробных клеток в 25 мкл, однократно. Через 14 дней после иммунизации мышей заражали интрацеребрально высоковирулентным штаммом B.pertussis 18323. В качестве положительного контроля использовали препарат ОСО-3 (Отраслевой стандартный образец корпускулярной коклюшной вакцины, ГИСК им.Л.А.Тарасевича), который вводили мышам внутрибрюшинно в количестве 10 МОЕ. После интрацеребрального заражения мышей высоковирулентным штаммом 18323 наблюдали защиту от гибели от 80 до 100% в зависимости от количества бактерий, использованных для вакцинации.
Еще один аспект изобретения касается использования аттенуированных двойных мутантов B.pertusis KS в качестве живой вакцины против возбудителя коклюша B.pertusis. Вакцина согласно изобретению может содержать фармацевтически приемлемый носитель, растворитель или наполнитель, а также адъюванты, например двуокись алюминия. Фармацевтически приемлемые носители включают, но не ограничиваются, фосфатным буфером, дистиллированной водой, эмульсиями "масло в воде" и т.п. Предпочтительно вакцина согласно изобретению вводится интраназально в одной дозе.
Еще один аспект изобретение касается инактивированной цельноклеточной вакцины на основе аттенуированных бактерий, содержащих мутантные гены ptx и dnt. Инактивация живых бактерий аттенуированного двойного мутанта B.pertusis KS производится с помощью обработки гексаметилентетрамином (ГМТА) в соответствии с описанным ранее методом (Авт. свид. №1497802). Токсическую активность определяли в опытах на мышах, морских свинках и кроликах, результаты экспериментов приведены в табл.4 и 5 (Пример 5). Данные, представленные в таблицах 4 и 5, демонстрируют, что корпускулярная вакцина, содержащая инактивированные аттенуированные рекомбинантные бактерии B.pertussis KS, не имеет специфической активности коклюшного и дермонекротического токсинов (уровень ЛСА и ГСА ниже, чем у ОСО-3), не вызывает гибели мышей при внутрибрюшинном введении препаратов и некрозов в кожной пробе на морских свинках и кроликах.
Защитные свойства корпускулярной вакцины на основе инактивированных аттенуированных бактерий B.pertussis KS определяли при интрацеребральном заражении высоковирулентным штаммом B.pertussis 18323 (табл.6, пример 5). В качестве положительного контроля использовали препарат ОСО-3, который вводили мышам внутрибрюшинно в количестве, соответствующем опытным образцам - 0,5; 0,1; 0,02×109 микробных клеток. После внутрибрюшинного введения животным инактивированных бактерий B.pertussis KS, в зависимости от количества бактерий, использованных для вакцинации, наблюдали защиту мышей от 80 до 100% от последующего интрацеребрального заражения вирулентными бактериями B.pertusis 18323.
Инактивированная цельноклеточная вакцина согласно изобретению может содержать перечисленные выше фармацевтически приемлемый носитель, растворитель или наполнитель, используемые для живой вакцины, а также адъюванты, например двуокись алюминия. Инактивированная цельноклеточная вакцина согласно изобретению вводится подкожно.
В еще одном из аспектов изобретение обеспечивает поливалентную вакцину на основе аттенуированных двойных мутантов Bordetella pertussis в качестве вектора. Поливалентный вакцинный штамм В. pertussis продуцирует антигены возбудителя туберкулеза ESAT-6, CFP-10 и Ag85A (заявки на патент РФ 2010123879, 2010123883, 2010123865, соответственно) в составе слитого комплекса с адгезином FGA или/и автотранспортным белком BrkA возбудителя коклюша. Белки ESAT-6, CFP10 и Ag85A экспонированы на С-конце химерного белка, что обеспечивает их представление на поверхности бактерии-вектора и сохранение их иммуногенных свойств, и, как следствие, индукцию выработки специфических защитных антител против возбудителя коклюша и основных протективных антигенов возбудителя туберкулеза при интраназальном введении человеку.
Изобретение иллюстрируется следующими примерами, представленными для подтверждения, но не ограничения объема притязаний.
Пример 1. Бактериальные штаммы E.coli, В.pertussis и условия их культивирования
Штаммы В. Pertussis 173 [Rifr, Серовар 1.2.3.7.13., ЛСА (40-50)], полученный из коллекция штаммов НИИЭМ им.Н.Ф.Гамалеи, и B.pertussis 5 [Rifr, Kmr, Серовар 1.2.0.7.13; ЛСА 17-20 т., ptx(M)], а также бактерии B.pertussis KS [Rifr, Kmr, Cmr, DNT], полученные согласно изобретению, выращивались на среде Борде-Жангу с добавлением 10% дефибринированной крови барана и жидкой питательной среде типа Коен-Уиллер.
Штаммы E.coli S17-1 λpir [TprSmr recA, thi, pro, hsdR-M+ RP4:2-Tc:Mu::Km Tn7 λpir], E.coli Dh5 α [F, Ф80dА (lacZ)ml5, recAl, endAl, gyrA96, thi-1 hsdR17(rk--mk+), supE44, relAl, glnV44, deoR, Δ(lacZYA-argF)U169], E.coli Dh5 α CmrApr/pBScad, полученные из коллекции штаммов НИИЭМ им. Н.Ф.Гамалеи, и штаммы E.coli Dh5α/pKS1, E.coli Dh5α/pKS2, E.coliS17-1 λpir)/pKnock-Tc и E.coli S17-1 λpir/pKS3, полученные согласно изобретению, выращивались в жидкой или на агаризованной среде «L», казеиновоугольном агаре (КУА). Антибиотики добавляли в концентрациях (мкг/мл): ампициллин - 100; канамицин - 25, рифампицин - 100, хлорамфеникол - 25, тетрациклин - 20.
Пример 2. Конструирование плазмиды для рекомбинационного обмена между копиями нативного и мутантного генов dnt
Для конструирования плазмиды с целью рекомбинационного обмена между копиями нативного и мутантного генов dnt был использован конъюгативный вектор pKnock - Тс [Tet ori-R6K mob RP4], не способный реплицироваться в бактериях рода Bordetella [Синяшина Л.Н., Нечаева Е.В., Амелина И.П., Каратаев Г.И., ЖМЭИ, 2009]. Схема конструирования плазмиды pKS3 и ее структура представлены на фиг.1б. Плазмида pKSl [pGEM (dnt)], содержащая последовательность гена dnt, инактивированную инсерцией гена cad (Фиг.1а), получена в результате клонирования продукта ПЦР ДНК B.pertusis-5 с праймерами Дф (Cga tat tcc agt cgt tcg tg) и, Дp (Tcg ttc aac аса tcc aag gc) в вектор pGem (PromegaUSA) и последующей интеграции гена cad (фрагмента Acc1 плазмиды pBScad) в TthlllI сайт гена dnt (плазмида pKS2). Структура рекомбинантной плазмиды pKS2 представлена на Фиг.1б. Плазмида pKS3 получена в результате переклонирования (SaeII-NotI) фрагмента плазмиды pKS2 в соответствующие сайты плазмиды pKnock-Tc (фиг.1б). Структура рекомбинантных плазмид подтверждена с помощью рестрикционного анализа и ПЦР с использованием различных сочетаний праймеров 
Рестрикционный анализ, лигирование, электрофорез ДНК, клонирование последовательностей ДНК проводили в соответствии с методическим руководством Маниатиса с соавт. [Маниатис Т., Э.Фрич, Дж.Сэмбрук. Молекулярное клонирование. Мир, Москва, 1984]. Выделение ДНК плазмид, хромосомы и продуктов ПЦР из геля проводили с помощью систем Wizard Plus SV MiniPrepsDNA purification system (Promega США), Wizard Plus MiniPrepsDNA purification system (Promega США) и Wizard SV Gel and PCR Clean-up System (Promega США). Полимеразную цепную реакцию (ПЦР) проводили на приборе «Терцик» (Россия) при использовании праймеров, синтезированных АО "Синтол".
Пример 3. Конструирование бактерий В.pertussis SK, содержащих мутантный ген dnt в составе хромосомы
Трансформацию клеток E.coli ДНК плазмид и лигированной смесью проводили в соответствии с методическим руководством Маниатиса с соавт. [Маниатис Т., Э.Фрич, Дж.Сэмбрук. Молекулярное клонирование. Мир, Москва, 1984]. Скрещивание между бактериями E.coli S17 и В.pertussis проводили в соответствии с методическим руководством Миллера [Миллер Дж. В. Эксперименты в молекулярной генетике. Мир, Москва, 1984. 436]. В качестве реципиента для конструирования аттенуированных бактерий использовался охарактеризованный ранее KmRRifR штамм В.pertussis 5, содержащий мутацию в гене ptx.
Бактерии В.pertussis, содержащие мутации ptxS1 и нокаутную мутацию dnt в хромосоме, сконструированы в результате аллельного обмена между природной копией оперона dnt в составе хромосомы реципиента и мутантной копией в составе плазмиды pKS3 (фиг.2). Плазмиду pKS3 передавали в бактерии В.pertussis в результате скрещивания S17 E.coli/pKS3XKmRRifR и В.pertussis 5. После трехкратной расчистки на среде КУА с хлорамфениколом бактерии проверяли по маркерам антибиотикоустойчивости, наличию плазмидной ДНК и последовательностей генов kan, cad и dnt в ПЦР с праймерами  Все проверенные бактериальные клоны трансконъюгантов имели фенотип СmR KmR RifR, не содержали плазмидного маркера устойчивости к тетрациклину и плазмидной ДНК. В ПЦР ДНК хромосомы, выделенной из бактерий трансконъюгантов, с праймерами Сф-Ср, и Кф-Кр показано наличие последовательностей, кодирующих устойчивости к канамицину и хлорамфениколу. В результате ПЦР ДНК хромосомы трансконъюгантов с праймерами Дфв-Дрв выявлены фрагмент размером 2380 п.о. и фрагмент размером 870 п.о., что соответствует размеру фрагмента dnt, содержащего вставку гена cat (фиг.3).
Все проверенные бактериальные клоны трансконъюгантов имели фенотип СmR KmR RifR, не содержали плазмидного маркера устойчивости к тетрациклину и плазмидной ДНК. В ПЦР ДНК хромосомы, выделенной из бактерий трансконъюгантов, с праймерами Сф-Ср, и Кф-Кр показано наличие последовательностей, кодирующих устойчивости к канамицину и хлорамфениколу. В результате ПЦР ДНК хромосомы трансконъюгантов с праймерами Дфв-Дрв выявлены фрагмент размером 2380 п.о. и фрагмент размером 870 п.о., что соответствует размеру фрагмента dnt, содержащего вставку гена cat (фиг.3).
Для подтверждения локализации инсерции последовательности в гене dnt проведена ПЦР на ДНК хромосомы трансконъюганта с праймерами Дфв-Ср (фиг.3). Так как праймер Дфв (Ccg caa tgg gcc gag ctg tt) расположен за пределами фрагмента dnt, клонированного в составе плазмиды pKS3, то в результате ПЦР с праймерами Дфв-Ср образуется специфический продукт размером 870 п.о. только при наличии инсерции последовательности cad в сайт TthlllI последовательности dnt. Специфичность полученных продуктов подтверждена результатами рестрикции эндонуклеазами АсcI и SalGI. Определение последовательности гeнa ptxS1 в составе хромосомы аттенуированных бактерий B.pertussis KS подтвердило наличие замен Arg9-Lys9 и Glul29-Glyl29 в области, кодирующей ферментативный центр КТ.
Таким образом, в соответствии с изобретением сконструированы бактерии B.pertussis KS, содержащие мутантный оперон ptx, инсерцию гена kan, расположенную вблизи мутантного оперона в участке, не оказывающем влияния на экспрессию оперона ptx или транспорт субъединиц рекомбинантного белка через бактериальную стенку [Синяшина Л.Н., Нечаева Е.В., Амелина И.П., Каратаев Г.И., ЖМЭИ, 2009], и инактивированную инсерцией гена cad последовательность гена dnt.
Пример 4. Иммунобиологические свойства аттенуированных бактерий B.pertussis KS
Лейкоцитозстимулирующую (ЛСА) и гистаминсенсибилизирующую (ГСА) активности КТ, а также количество КТ определяли в соответствии с известными методами [Инструкция по отбору, проверке и хранению штаммов B.pertussis для изготовления коклюшной вакцины и коклюшного компонента ассоциированных вакцин. 1987, Москва; Синяшина Л.Н., Нечаева Е.В., Амелина И.П., Каратаев Г.И., ЖМЭИ, 2009]. Из данных, представленных в табл.1 видно, что количество КТ, продуцируемого рекомбинантными бактериями B.pertussis KS, практически не отличается от количества КТ, продуцируемого исходными вирулентными бактериями. Специфическая токсическая активность КТ, синтезируемого аттенуированными рекомбинантными бактериями B.pertussis и бактериями вирулентного штамма, напротив, значительно отличается. Аттенуированные бактерии B.pertussis, синтезирующие мутантный КТ, вызывают более низкий лейкоцитоз, гистаминсенсибилизацию у мышей и эффект образования кластеров клеток СНО при значительно большей концентрации токсоида, чем природного КТ. В таблице 1 снижение токсичности КТ приведено в процентах. Эти цифры отображают отношение концентрации природного токсина к мутантному, при добавлении которых к культуральной среде наблюдается одинаковый кластер - эффект монослоя клеток СНО. Значения измеренных параметров практически не отличаются у одиночных и двойных мутантов и близки к специфической активности токсоида, определенной ранее [Синяшина Л.Н., Нечаева Е.В., Амелина И.П., Каратаев Г.И., ЖМЭИ, 2009; Pizza M., Covacci A., Bartoloni A., et al. Science, 1989, 246: 497-500].
Токсическую активность ДНТ рекомбинантных клеток определяли в опытах на животных (кроликах, морских свинках и мышах). Кроликам весом 1,5-2,0 кг и морским свинкам весом 300-350 г вводили внутрикожно 2.0 МОЕ, 1 МОЕ и 0,5 МОЕ живых микробных клеток в логарифмической фазе в объеме 0,2 мл. Контроль - ОСО-5 (отраслевой стандартный образец. Тот же, что и ОСО-3, только живые бактерии). Некроз учитывали через 24 и 48 ч.
При определении активности ДНТ в опытах на мышах определяли 50% летальную дозу [Инструкция по отбору, проверке и хранению штаммов B.pertussis для изготовления коклюшной вакцины и коклюшного компонента ассоциированных вакцин. 1987, Москва]. Животным весом 10-12 г вводили живые бактерии внутрибрюшинно в соответствующих разведениях. Результаты учитывали в течение 48 ч.
Для оценки остаточной токсичности аттенуированные бактерии B.pertussis KS выращивали на среде КУА в течение 36-48 ч, смывали 0.85% раствором хлорида натрия, концентрацию бактериальных клеток доводили до 50-60 МОЕ в 1 мл. Каждой мыши вводили внутрибрюшинно 10 МОЕ в объеме 0,5 мл. Контрольным животным вводили 0.5 мл 0.85% раствора хлорида натрия. За животными наблюдали в течение 7 суток. Массу мышей определяли через 72 ч и через 7 суток после введения. Абсолютный прирост массы тела мышей, которым ввели аттенуированные бактерии, на седьмые сутки был 100% и выше. Относительный прирост массы мышей по сравнению с контрольными мышами, которым вводили 0,85% раствор хлорида натрия, был более 80%. Через 72 ч масса мышей, которым вводили аттенуированные бактерии, была не ниже их массы до введения препарата и составляла 20% относительного прироста массы контрольных мышей.
Защитные свойства бактерий определяли по проценту выживаемости мышей при иммунизации их определенным количеством микробных клеток в соответствии с международными стандартами, принятыми в РФ [Инструкция по отбору, проверке и хранению штаммов B.pertussis для изготовления коклюшной вакцины и коклюшного компонента ассоциированных вакцин. 1987, Москва]. При интраназальной иммунизации мышам вводили от 4 до 500 млн живых микробных клеток в 25 мкл, однократно. Через 14 дней после иммунизации мышей заражали интрацеребрально высоковирулентным штаммом B.pertussis 18323. В качестве положительного контроля использовали препарат ОСО 3, который вводили мышам внутрибрюшинно в количестве, соответствующем 1000 млн микробных клеток. После внутрибрюшинного и интраназального введения животным живых бактерий B.pertussis 5 и KS, в зависимости от количества бактерий, использованных для вакцинации, наблюдали от 80 до 100% их защиту от интрацеребрального заражения вирулентными бактериями B.pertusis 18323.
Стабильность структуры генома и свойств рекомбинантных бактерий B.pertussis KS определяли после 15 пассажей на среде КУА, лиофильного высушивания в сахарозо-желатинозной среде и хранения при 4°С (срок наблюдения более 1 года), а также после высева бактерий из легочной ткани мышей, инфицированных живыми аттенуированными бактериями. Для изучения приживаемости штаммов и развития инфекционного процесса суспензии микробных клеток в концентрации 1×109 в казеиновом гидролизате рН 7.0 вводили в каждую ноздрю мыши в объеме 0.3 мл, параллельно в том же объеме вводили внутривенно. Через 72 ч проверяли ЛСА и проводили посевы из легких. В посевах из легких мышей на среде КУА с кровью через 3 и 6 дней после их заражения не регистрировали бактериальных колоний. В группе мышей, зараженных внутривенно вирулентным штаммом B.pertussis 173, наблюдали высокий лейкоцитоз (около 100000), тогда как у мышей, зараженных аттенуированными бактериями, значение ЛСА находилось в пределах 17.000-23.000 лейкоцитов/мл, что соответствует ЛСА при иммунизации мышей коммерческой ККВ. ЛСА исходных и мутантных бактерий B.pertussis при интраназальном введении микробных клеток не превышал 10-15000. К 8-му дню после интраназального введения микробной суспензии исходного штамма и мутантных бактерий B.pertussis KS и 5 число выросших колоний при посеве гомогената легких мышей достигало 150-200, в то время как после внутривенного введения мышам бактерий B.pertussis KS и 5 высевалось не более 40 колоний. При внутривенном заражении бактериями исходного штамма и посеве на среду КУА гомогената из легких мышей роста отдельных колоний не наблюдали. К 14-му дню после заражения количество колоний при посеве гомогената легких мышей, зараженных интраназально бактериями B.pertussis 173 и мутантами B.pertussis 5 и B.pertussis KS, достигало 3×103, 104 и 3×104, соответственно, и продолжало сохраняться в таком количестве до конца третьей недели после заражения. Различия между числом бактерий дикого штамма и двойного мутанта B.pertussis KS - статистически значимы (Р=0,95). К концу четвертой недели после интраназального и внутривенного заражения животных исходными и аттенурованными бактериями B.pertussis в высевах гомогената легких мышей не выявили бактериальных колоний на индикаторных чашках. Отдельные колонии аттенуированных бактерий B.pertussis KS, выросших на индикаторных чашках после посева гомогенатов легких мышей на четвертой неделе инфекции, перепечатывали на селективные чашки, содержащие антибиотики канамицин и хлорамфеникол и определяли стабильность маркеров в процессе размножения бактерий в организме животных. Все 100 проверенных колоний сохраняли оба маркера устойчивости к антибиотикам. Три отдельные колонии были размножены на среде КУА и использованы для проверки ЛСА и дермонекротической активности. Аналогичные исследования выполнены с бактериями B.pertussis KS после 15 пассажей на среде КУА и лиофильного высушивания штаммов в сахарозо-желатинозной среде и хранении при 4°С в течение не менее 12 месяцев. Показано, что бактерии, выделенные из легких спустя три недели после инфицирования мышей, бактерии после пассажей на искусственной питательной среде КУА и бактерии, прошедшие процедуру лиофилизации, не имеют дермонекротической активности и проявляют ЛСА в пределах, зарегистрированных у бактерий B.pertussis KS до пассажа в организме животных и лиофильного высушивания.
Параллельно с высевом на селективную среду гомогената легких мышей, зараженных интраназально, препараты ДНК анализировали с помощью ПЦР на праймерах, специфичных для последовательностей генов ptx, kan, cad и инактивированной последовательности гена dnt. Результаты ПЦР не выявили ДНК бактерий дикого типа в легких мышей уже на пятой недели после заражения. Результаты ПЦР подтвердили, что все тестированные последовательности присутствовали в гомогенатах, выделенных от животных, инфицированных аттенуированными бактериями B.pertussis KS и B.pertussis 5 на протяжении 4-х недель, в течение которых удавалось выделять бактерии B.pertussis из легких инфицированных животных, и далее вплоть до десятой недели. Результаты ПЦР разведения ДНК возбудителя коклюша из препаратов, выделенных в конце 10-й недели, выявили значимые различия в количестве ДНК бактерий B.pertussis KS и B.pertussis 5. В легких мышей, зараженных бактериями двойного мутанта B.pertussis KS, обнаружено в 5-10 раз больше ДНК возбудителя коклюша, чем у мышей, зараженных одиночным мутантом В.pertussis 5. Полученные результаты указывают на лучшую переживаемость бактерий B.pertussis KS, чем бактерий В.pertussis 5 и, тем более, бактерий дикого типа. B.pertussis 173, в легких лабораторных животных, зараженных интраназально.
Таким образом, аттенуированные бактерии B.pertussis согласно изобретению не имеют дермонекротической активности и проявляют низкую общую токсическую активность, соответствующую требованиям, предъявляемым к противококлюшным вакцинам, производимым в РФ. Более того, аттенуированные бактерии B.pertussis согласно изобретению продуцируют иммунологически активную, нетоксичную производную коклюшного токсина и сохраняют структуру мутантных генов ptx и dnt после, по крайней мере, 15 пассажей на искусственных питательных средах, хранения и при размножении в легких лабораторных животных. Интраназальная иммунизация лабораторных мышей живыми бактериями аттенуированного штамма B.pertussis KS обеспечивает защиту от внутримозгового заражения мышей высоковирулентным штаммом 18323 возбудителя коклюша, близкую по защитному эффекту после иммунизации отраслевым стандартным образцам коклюшной корпускулярной вакцины (ОСО-3).
Пример 5. Корпускулярная вакцина на основе аттенуированных бактерий B.pertussis KS, инактивированных гексаметилентетрамином (ГМТА)
Препараты готовили согласно описанному ранее методу (Авт. свид. №1497802). Бактерии В. pertussis выращивают на средах КУА или Коен-Уиллер при 35°С 36 часов, смывают 0,85% раствором хлорида натрия (рН 7,0-7,2) до концентрации 50-60 МОЕ/мл. В суспензию добавляют 10% раствор ГМТА. Выдерживают при 35°С 48-96 часов. Оценку токсичности и защитных свойств образца вакцины согласно изобретению определяли в соответствии с инструкцией по отбору, проверке и хранению штаммов коклюшных бактерий для производства коклюшных вакцин и коклюшного компонента АКДС-вакцин (М., 1987). Образцы вакцины на основе аттунуированных бактерий B.pertussis KS проверяли через 3 месяца после приготовления. Токсическую активность определяли в опытах на мышах, морских свинках и кроликах.
* Относительный прирост массы мышей после внутрибрюшинного (i.p) введения 1×109 м.к. измерялся в % по отношению к массе тела контрольных животных, которым вводили 0,85 р-р хлорида натрия.
Данные, представленные в табл.4 и 5, демонстрируют, что корпускулярная вакцина, приготовленная инактивированием аттенуированных рекомбинантных бактерий B.pertussis KS, не имеет специфической активности коклюшного и дермонекротического токсинов (уровень ЛСА и ГСА ниже, чем у ОСО-3, не вызывает гибели мышей при внутрибрюшинном введении препаратов и некрозов в кожной пробе на морских свинках и кроликах). Результаты теста изменения массы мышей (табл.4) показали, что испытуемая корпускулярная вакцина проявляет низкую токсичность в сравнении с ОСО-3 (прирост массы тела мышей через 72 ч составил 20% и через 7 суток составил более 80%) (табл.6).
В качестве положительного контроля использовали препарат ОСО-3, который вводили мышам внутрибрюшинно в количестве, соответствующем опытным образцам - 0,5; 0,1; 0,02×109 микробных клеток. После внутрибрюшинного введения животным инактивированных бактерий B.pertussis KS в зависимости от количества бактерий, использованных для вакцинации, наблюдали защиту мышей от 80 до 100% от последующего интрацеребрального заражения вирулентными бактериями B.pertusis 18323.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ лиофильного высушивания аттенуированных бактерий B. pertussis, аттенуированная бактерия B. pertussis, штамм аттенуированных бактерий B. Pertussis, вакцина, лиофилизированный вакцинный препарат | 2017 |
|
RU2709657C2 |
| СПОСОБ ОЦЕНКИ ЗАЩИТНОЙ АКТИВНОСТИ КОКЛЮШНЫХ ВАКЦИН | 2015 |
|
RU2623314C2 |
| Способы оценки защитной активности и иммуногенности коклюшных вакцин у человека, применение живой коклюшной рекомбинантной вакцины ГамЖВК | 2021 |
|
RU2799018C2 |
| СПОСОБ МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКОГО ТИПИРОВАНИЯ ШТАММОВ Bordetella pertussis | 2012 |
|
RU2495132C1 |
| СПОСОБ ОЦЕНКИ ИММУНОГЕННОСТИ КОКЛЮШНЫХ ВАКЦИН | 2003 |
|
RU2231793C1 |
| СПОСОБ ДИАГНОСТИКИ КОКЛЮША И ОПРЕДЕЛЕНИЯ АВИРУЛЕНТНЫХ МУТАНТОВ ВОЗБУДИТЕЛЯ И ДИАГНОСТИЧЕСКИЙ НАБОР | 2011 |
|
RU2506316C2 |
| ПОЛИПЕПТИД, РЕКОМБИНАНТНЫЙ ПОЛИНУКЛЕОТИД, ВАКЦИНА | 1993 |
|
RU2129611C1 |
| СПОСОБ ПОЛУЧЕНИЯ АТТЕНУИРОВАННОГО ШТАММА БАКТЕРИЙ SALMONELLA И ВАКЦИНА | 1992 |
|
RU2126447C1 |
| СПОСОБ ДИФФЕРЕНЦИАЦИИ ШТАММОВ BORDETELLA PERTUSSIS | 2005 |
|
RU2299908C1 |
| СПОСОБ ПОЛУЧЕНИЯ АТТЕНУИРОВАННОГО ШТАММА БАКТЕРИЙ SALMONELLA | 1988 |
|
RU2114172C1 |

Изобретение раскрывает аттенуированные бактерии В. pertussis, содержащие мутантные гены ptx и dnt, не проявляющие активности дермонекротического токсина, но сохранившие способность связывания с рецептором эукариотической клетки. Бактерии по изобретению продуцируют генетически измененную иммунологически активную нетоксичную токсоидную форму коклюшного токсина. Изобретение касается штамма бактерий В.pertussis, содержащего указанные мутантные гены, который депонирован в Коллекции штаммов НИИЭМ им. почетного академика Н.Ф.Гамалеи под номером 171/331 В.р KS-4. Штамм может быть использован в качестве вакцины. Вакцина против возбудителя коклюша содержит живые аттенуированные бактерии или инактивированные аттенуированные бактерии В.pertussis. Интраназальная иммунизация вакциной, содержащей живые или инактивированные аттенуированные бактерии В.pertussis по изобретению, проведенная в эксперименте in vivo, обеспечивает эффективную и пролонгированную защиту от заражения вирулентным штаммом возбудителя коклюша. 3 н. и 1 з.п. ф-лы, 4 ил., 6 табл., 5 пр.
1. Аттенуированные бактерии Bordetella pertussis, содержащие мутантные гены ptx и dnt, не проявляющие активности дермонекротического токсина, но сохранившие способность связывания с рецептором эукариотической клетки, и продуцирующие генетически измененную иммунологически активную нетоксичную токсоидную форму коклюшного токсина.
2. Штамм аттенуированных бактерий Bordetella pertussis, содержащих мутантные гены ptx и dnt, депонированный в Коллекции штаммов бактерий Федерального государственного бюджетного учреждения «Научно-исследовательский институт эпидемиологии и микробиологии имени почетного академика Н.Ф.Гамалеи Министерства здравоохранения и социального развития российской Федерации» под №171/331 В.р KS-4, для использования в качестве вакцины.
3. Вакцина против возбудителя коклюша, содержащая аттенуированные бактерии B.pertussis по п.1.
4. Вакцина по п.3, отличающаяся тем, что она содержит живые аттенуированные бактерии или инактивированные аттенуированные бактерии B.pertussis.
| WO 2007104451 A1, 20.09.2007 | |||
| RU 2227746 C1, 27.04.2004 | |||
| СПОСОБ ПОЛУЧЕНИЯ АНАТОКСИНА BORDETELLA PERTUSSIS | 1986 |
|
SU1369042A1 |
| WO 2006122586 A1, 23.11.2006. | |||

