Настоящее изобретение относится к удалению загрязняющей ДНК из реакционных смесей для обратной транскрипции, препаратов ДНК-полимеразы с горячим стартом и реакционных смесей для ПЦР с горячим стартом путем применения ДНКазы. Изобретение также относится к предупреждению ложноположительных результатов в реакциях амплификации нуклеиновых кислот путем применения ДНКазы, в частности реакциях амплификации, включая обратную транскрипцию последовательности-мишени, ДНК-полимеразу с горячим стартом и/или барьерную комбинацию для ПЦР с горячим стартом. Изобретение также относится к чрезвычайно термолабильной ДНКазе, подходящей для применения в таких способах.
Способы амплификации нуклеиновых кислот, такие как полимеразные цепные реакции (ПЦР), представляют собой один из наиболее мощных инструментов, имеющихся в биотехнологии, дающих возможность для получения большого количества копий последовательности-мишени из образца, содержащего лишь небольшое количество нуклеиновой кислоты. В случае ПЦР олигонуклеотидные праймеры, комплементарные соответствующим цепям двухцепочечной последовательности-мишени, добавляют к реакционной смеси, содержащей последовательности-мишени и свободные нуклеотиды. Термоциклирование в присутствии ДНК-полимеразы приводит в результате к амплификации последовательности между праймерами. Способность амплифицированных фрагментов, полученных при помощи способа ПЦР, действовать в качестве матриц для последующих циклов ПЦР приводит в результате к быстрому образованию существенного количества последовательности-мишени. Даже единичная копия последовательности-мишени может привести к достаточному количеству нуклеиновой кислоты для того, чтобы обеспечить детекцию, например, путем гибридизации с меченым зондом или включения меченых 32P дезоксинуклеотидтрифосфатов в амплифицированный сегмент.
Лигазная реакция амплификации (ЛРА), также известная как лигазная цепная реакция (ЛЦР), как и ПЦР, использует повторяющиеся циклы и изменение температуры для достижения экспоненциального увеличения во множестве копий последовательности-мишени. В этом способе ДНК-лигаза катализирует присоединение двух олигонуклеотидов, комплементарных соседним областям одной из цепей-мишеней ДНК. Два других олигонуклеотида, комплементарные другой цепи, также могут быть лигированы. После денатурации исходные матричные цепи и две лигированные пары могут действовать в качестве матриц для последующей гибридизации и лигирования.
Амплификация с вытеснением цепи (SDA) использует свойство ферментов, вовлеченных в репарацию ДНК путем вырезания ДНК, замещать имеющую разрезы единичную цепь ДНК в дуплексе ДНК вновь синтезированной цепью. Для получения имеющей разрезы единичной цепи повторно используют фермент эндонуклеазной рестрикции, например Hindi или BsoBI, который лишь разрезает ДНК одной цепи своего сайта распознавания, тогда как противоположная цепь гемифосфоротиолирована. Праймеры, использованные в этом способе, содержат подходящий сайт распознавания, и dATPαS используют в реакции полимеризации.
Амплификация, основанная на последовательности нуклеиновой кислоты (NASBA), также известная как 3SR (самоподдерживающаяся репликация последовательности), по существу представляет собой вариант in vitro естественной ретровирусной транскрипции. 3SR использует повторяющуюся обратную транскрипцию с матрицы РНК с образованием матрицы кДНК. С матрицы кДНК РНК-полимераза продуцирует соответствующую РНК.
Изотермальная петлевая амплификация (ИПА; Notomi, Т., et al., Nuc. Acid Res. 2000 Vol 28 (12) е63) основана на принципе аутоциклического синтеза ДНК с замещением цепи. Используют ДНК-полимеразу, обладающую высокой активностью замещения цепи (например, крупный фрагмент ДНК-полимеразы Bst) со специальным образом разработанными праймерами. Этот процесс включает разделение цепей для выявления новых последовательностей-мишеней без необходимости плавления цепи (таким образом, процесс является изотермическим).
Обратная транскрипция представляет собой процесс, в котором матрица одноцепочечной РНК (оцРНК) транскрибируется в комплементарную одноцепочечную ДНК. Одноцепочечная ДНК может затем использоваться для образования двухцепочечной ДНК (дцДНК). Некоторые ферменты способны продуцировать первую цепь ДНК и синтезировать вторую цепь для образования дцДНК, а другие специфичны только в отношении одной из двух цепей. оцДНК и дцДНК затем могут использоваться во множестве применений молекулярной биологии. Например, они могут использоваться непосредственно в анализах путем детекции с применением зонда (например, Саузерн-блоттинг), экспериментах по секвенированию или в протоколах клонирования. Очень часто кДНК дополнительно амплифицируют в реакции амплификации, такой как ПЦР, ЛЦР, SDA, ИПА или 3SR, например, для получения дополнительного материала для вышеприведенных экспериментов или для количественной оценки количества матрицы РНК, представленной в исходном образце.
Реакции амплификации, связанные с обратной транскрипцией, могут представлять собой "одностадийные" или "двухстадийные" способы. В одностадийном способе компоненты реакции обратной транскрипции и реакции амплификации нуклеиновой кислоты присутствуют в одном реакционном сосуде и типично условия ранней реакции выбраны для того, чтобы обеспечить протекание реакции обратной транскрипции до завершения, и условия реакции затем переключаются на условия, подходящие для того, чтобы обеспечить протекание реакции амплификации нуклеиновой кислоты.
В двухстадийном способе сначала комбинируют компоненты реакции обратной транскрипции и осуществляют реакцию обратной транскрипции. Продукт обратной транскрипции затем комбинируют с компонентами реакции амплификации и подвергают реакции амплификации. В двухстадийном протоколе в "одной пробирке" компоненты реакции амплификации добавляют в один и тот же реакционный сосуд, в котором осуществлялась реакция обратной транскрипции. В двухстадийном протоколе с "двумя пробирками" реакцию амплификации осуществляют в новом реакционном сосуде.
Обратная транскрипция может быть комбинирована с любой из РСА, ИПА, ЛЦР, SDA или BSR в одно- или двухстадийном способе. В случае SDA должен быть выбран термостабильный фермент, замещающий цепь, и термостабильный рестрикционный фермент (например, BsoB1).
Способность этих способов амплификации амплифицировать минимальные количества последовательности-мишени делает их высокочувствительными к загрязнению геномной ДНК в случае РНК последовательностей-мишеней (т.е. реакций амплификации, следующих за обратной транскрипцией или использующих ее) и последовательностями-мишенями в молекулах ДНК с предшествующих реакций амплификации, все из которых могут быть привнесены с реагентами (например, с полимеразой, праймерами, реакционным буфером и т.п), с пилотирующими устройствами, на лабораторных поверхностях, перчатках или путем аэрозолизации. Аэрозоли могут образовываться при нарушении состояния покоя раствора, такого как во время пролития, или даже при нарушении состояния покоя небольшого количества материала на поверхности контейнера, такого как остаток на внутренней поверхности крышки пластиковой пробирки, который может быть аэрозолизирован при открывании пробирки. При исследовании образца нуклеиновой кислоты в целях медицинской диагностики или судебно-медицинских причин влияние ложноположительных результатов, вызванных путем случайного введения в реакционную смесь нуклеиновой кислоты, которая может содержать последовательности-мишени, известного как перенос, может иметь серьезные последствия.
Реакции амплификации, особенно чувствительные к вредным эффектам загрязнения нуклеиновыми кислотами, представляют собой способы количественной ПЦР, поскольку обладают возможностью количественно определять меньше чем 20 копий последовательности ДНК в реакции. Таким образом, даже наименьшие уровни загрязнения нуклеиновыми кислотами могут приводить к ошибочным результатам способов в кПЦР (количественной ПЦР). Дополнительно эти способы требуют детектирования сигналов с амплифицированных нуклеиновых кислот-мишеней, превосходящих неизменный фоновый сигнал. Загрязняющая нуклеиновая кислота может вносить вклад в этот фоновый сигнал и таким образом уменьшать чувствительность способа. Сама по себе минимизация загрязняющей нуклеиновой кислоты максимизирует чувствительность эксперимента количественной ПЦР. В экспериментах, в которых обнаруживают небольшие количества копий нуклеиновых кислот-мишеней, например, при количественной диагностике патогена, основанной на ПЦР, и при количественном определении нагрузки патогеном, наиболее важно, чтобы чувствительность количественной ПЦР была максимизирована, а количество ложноположительных результатов было минимизировано. В области идентификации бактерий и диагностики, где в качестве мишеней способов кПЦР выступают сегменты высококонсервативной бактериальной ДНК (например, гены 16SrРНК или 23SrРНК), загрязнения нуклеиновыми кислотами в результате получения ДНК-полимеразы (которую типично получают из бактерий и систем бактериальной экспрессии), представляют собой значительную проблему. Таким образом, необходимы способы эффективного удаления примесей бактериальной нуклеиновой кислоты из препаратов ДНК-полимеразы. Особенно необходимы способы, которые позволяют достичь этого без вредного влияния на последующие реакции амплификации и без повреждения полимеразы.
Разработано множество способов предупреждения или ограничения влияния переноса. В случае ПЦР они включают гнездовые праймеры, праймеры, которые отжигаются с последовательностями-мишенями внутри границ отжига двух праймеров, используемых для запуска ПЦР (K.B.Mullis et al. Cold Spring Harbour Symposia Vol. LI, p. 263-273, 1986). Более короткий амплифицируемый при помощи ПЦР продукт гнездовых праймеров не может отжигаться с исходными праймерами, таким образом, что является продуктом, претерпевающим перенос, применение исходных праймеров не амплифицирует этот перенос. Тем не менее, перенос не устраняется, и, если те же самые гнездовые праймеры используются в последующей ПЦР, будет амплифицироваться ранее амплифицированный продукт гнездовых праймеров.
Разработаны способы, которые охватывают включение нуклеотиддезоксиуридинтрифосфата (dUTP) в подвергнутую обратной транскрипции/амплифицированную последовательность нуклеиновой кислоты вместо дезокситимидинтрифосфата (dTTP). Поскольку дезоксиуридин (dU) обычно не обнаруживается во встречающейся в природе ДНК, это основание отличает ранее продуцированные ампликоны от новых последовательностей-мишеней. Перед началом осуществления дополнительной обратной транскрипции/реакций амплификации смесь для реакции амплификации может быть обработана ферментом урацил-ДНК-гликозилазой (UNG), который удаляет основание урацил, оставляя сахар-фосфодиэфирный скелет интактным, продуцируя абазический сайт в одноцепочечной (оц) и двухцепочечной (дц) ДНК (US-A-5,418,149). Температуру смеси реакции амплификации увеличивают для отщепления ДНК по абазическим сайтам, что приводит в результате к уменьшению переноса.
Этот способ также имеет недостатки, поскольку введение dUTP в продукт обратной транскрипции/амплификации может иметь отрицательное влияние на последующий анализ продукта, например, путем расщепления рестрикционным ферментом или ПЦР (эффективность полимеризации может быть уменьшена, и возникает препятствие для применения корректно читающих полимераз). Также UNG должен быть необратимо инактивирован, иначе продукты последующих реакций обратной транскрипции/ПЦР будут подвергаться деградации. Повышенная температура представляет собой обычный механизм для инактивации ферментов UNG, но множество из имеющихся в настоящее время в продаже ферментов UNG не подвергаются успешной инактивации даже после воздействия температур реакции ПЦР. Для минимизации влияния остаточной активности UNG температурные стадии, используемые в реакции амплификации, должны быть выше 54°С и реакционный сосуд должен содержаться при высоких температурах или быть немедленно заморожен для предупреждения деградации новых продуктов амплификации, которые также содержат урацил. Недавно описана UNG, полученная из трески, которая может быть полностью необратимо инактивирована при инкубации при 50°С в течение 10 минут, и это делает основанные на UNG подходы более широко применимыми.
Тем не менее, дополнительное ограничение для любой системы UNG заключается в том, что она не дает возможности для очистки реакционной смеси от загрязняющей геномной ДНК, поскольку геномная ДНК не имеет модификации урацила. Соответственно системы UNG не способны устранять загрязнение геномной ДНК вследствие реакций обратной транскрипции.
Также предположили, что индивидуальные реакционные смеси для ПЦР, обработанные перед добавлением ДНК мишени и Taq ДНК-полимеразы ДНКазой I или рестрикционными эндонуклеазами, которые режут внутри относительно последовательности-мишени, таким образом предупреждают амплификацию загрязняющей ДНК (Furrer et al. Nature. Vol.346, p. 324, 1990). Аналогично, реакционные смеси для обратной транскрипции могут быть обработаны таким образом перед добавлением обратной транскриптазы. Для этого способа требуется время удаления загрязнения 30 минут, и для инактивации ДНКазы I или рестрикционной эндонуклеазы после удаления загрязнения реакционную смесь кипятят. Вследствие этой стадии кипячения необходимо добавлять ДНК-полимеразу или обратную транскриптазу после удаления загрязнения. Безусловно, последнее представляет дополнительный риск переноса в преамплификационной смеси/смеси до обратной транскрипции, и предотвращается удаление загрязнения самой ДНК-полимеразы. Концентрации праймера 1 мкМ должны использоваться в этом способе вследствие активности ДНКазы I в отношении одноцепочечной ДНК.
Также описаны ДНКазы, которые являются более термолабильными. В WO 99/007887 раскрыта ДНКаза, выделенная из Pandalus borealis, которая по существу необратимо инактивируется после 2 минут при 94°С. Этот же фермент также по существу необратимо инактивируется после 15 минут при 65°С. Anisimova et al. (Biotechnology Letters; 2009, 31: 251 to 257) описывает произвольным образом мутировавший вариант ДНКазы королевского краба (Камчатский краб, Paralithodes camtschaticus), который инактивируется после инкубации в течение 10 минут при 65°С, хотя инактивация может быть достигнута при температуре 55°С после 10 минут, если используют инактивирующие добавки DTT (1-4 дитиотреитол) и EDTA (этилендиаминтетрауксусная кислота).
EDTA представляет собой хелатообразователь с ионами металлов и таким образом может влиять на действие ферментов, чувствительных к концентрации ионов металлов. Анисимова указывает на то, что на активность ДНКазы королевского краба положительно влияют ионы Mg2+, и, таким образом, EDTA вносит вклад в ее инактивацию путем блокирования этого активатора. ДНК-полимеразы также весьма чувствительны к концентрации ионов металла, и, в частности, содержание Mg2+ в полимеразных реакционных смесях должно тщательно контролироваться. В результате применение EDTA на стадии инактивации ДНКаза имеет потенциал непосредственно ингибировать активность последующих полимеразных реакций. Таким образом, предпочтительно не использовать EDTA на стадиях процессинга, предшествующих полимеразной реакции (например, обратная транскрипция, ПЦР, SDA, 3SR).
Поскольку мутантной ДНКазе королевского краба, предложенной Анисимовой, требуется присутствие EDTA для того, чтобы обеспечить инактивацию при температурах ниже 65°С, эта ДНКаза не подходит для применения на стадии удаления загрязняющей ДНК, предшествующей реакции обратной транскрипции (которую обычно проводят при приблизительно 50°С). Для обеспечения возможности использования этого фермента без риска ингибирования EDTA последующих стадий должна быть стадия инактивации, на которой смесь нагревают до температуры выше 65°С. Поскольку эта температура превышает температуру типичной реакции обратной транскрипции, эта стадия будет отделена и будет дополнительной к стадии обратной транскрипции. Последнее добавляет дополнительную стадию в способ, таким образом увеличивая сложность и трудозатраты в способе и также в области молекулярной биологии, в котором способ может быть повторен множество раз, представляет значительный недостаток с точки зрения затрат энергии и времени использования оборудования. Кроме того, если только обратную траскриптазу не добавляют после обработки ДНКазой и инактивации, ДНКаза будет активной на стадии обратной транскрипции, и поэтому существует риск того, что продукт кДНК будет подвергаться деградации. Более позднее добавление обратной транскриптазы к реакционной смеси может представлять возможность для загрязнения и усложнение способа в целом, вновь с увеличением стоимости.
Авторы настоящего изобретения выяснили, что ДНКаза, которая может быть по существу необратимо инактивирована при температурах, совместимых со стадией обратной транскрипции, и которая по существу специфична в отношении двухцепочечной ДНК, может обеспечивать высокоэффективный и производительный способ. Тем не менее, в настоящее время отсутствует ДНКаза, обладающая этими свойствами. Такая ДНКаза может использоваться для удаления загрязнения полной реакционной смеси для обратной транскрипции (т.е. реакционной смеси, содержащей все основные компоненты, требующиеся для обратной транскрипции молекулы РНК) немедленно перед реакцией обратной транскрипции, и затем после инициации реакции обратной транскрипции (т.е. повышение температуры реакционной смеси до рабочей температуры выбранного для обратной транскрипции фермента, например 50°С или выше) ДНКаза будет по существу необратимо инактивирована в течение реакции обратной транскрипции (при том, что большая часть инактивации идеально осуществляется в течение первых минут реакции). Этот период инактивации является важным, поскольку означает, что вновь образующаяся кДНК не будет деградировать под действием ДНКазы. В отличие от ДНКазы с более высокой температурой инактивации такой ДНКазе не потребуется отдельная стадия инактивации и/или более позднее добавление обратной транскриптазы.
Авторы настоящего изобретения получили фермент, обладающий этими уникальными свойствами. Как все ДНКазы, чрезвычайно термолабильная ДНКаза по изобретению расщепляет ДНК путем расщепления фосфодиэфирных связей сахарофосфатного скелета нуклеиновой кислоты.
Таким образом, в соответствии с настоящим изобретением, предложен способ удаления загрязняющей нуклеиновой кислоты из реакции обратной транскрипции, при котором используют ДНКазу, которая по существу необратимо инактивируется путем нагревания при температуре приблизительно 50°С в течение 5 минут и которая по существу специфична в отношении двухцепочечной ДНК. Предпочтительно этих характеристик инактивации достигают в отсутствие EDTA.
Таким образом, ДНКазу по изобретению используют для деградации загрязняющей двухцепочечной ДНК, присутствующей в реакционной смеси для обратной транскрипции, или представленной в ее индивидуальных компонентах. Таким образом, уровень загрязняющей ДНК в продукте обратной транскрипции (которая может быть амплифицирована и таким образом дать начало для ложноположительных результатов при использовании продукта обратной транскрипции) может быть уменьшен или исключен, и неспецифическая обратная транскрипция также может быть уменьшена или исключена.
В частности, способ включает приведение в контакт реакционной смеси для обратной транскрипции или ее индивидуальных компонентов с ДНКазой по изобретению в условиях, которые дают возможность для расщепления какой-либо двухцепочечной ДНК, и затем нагревание указанной реакционной смеси или ее индивидуальных компонентов для инактивации указанной ДНКазы. Предпочтительно реакционная смесь представляет собой полную реакционную смесь (т.е. включающую праймеры ДНК), и предпочтительно полную реакционную смесь нагревают при температуре, соответствующей рабочей температуре содержащегося в ней фермента для обратной транскрипции.
В еще одном воплощении за реакцией обратной транскрипции следует реакция амплификации нуклеиновой кислоты (например, ПЦР, ЛЦР, SDA, 3SR, ИПА). Предпочтительно ПЦР, ЛЦР или ИПА следуют за реакцией обратной транскрипции. В наиболее предпочтительном воплощении реакция амплификации представляет собой ПЦР.
В еще одном воплощении реакцию обратной транскрипции и реакцию амплификации осуществляют в виде одностадийного способа, т.е. реакционный сосуд имеет все компоненты для реакции обратной транскрипции и реакции амплификации, находящиеся в сосуде одновременно. Тем не менее, также могут быть использованы двухстадийные способы. В таких воплощениях различные компоненты реакционной и частично реакционных смесей могут быть обработаны индивидуально ДНКазой по изобретению.
Альтернативно, в этом аспекте изобретения предложено применение ДНКазы, которая по существу необратимо инактивируется путем нагревания при температуре приблизительно 50°С в течение 5 минут и которая по существу специфична в отношении двухцепочечной ДНК, для удаления загрязняющей нуклеиновой кислоты из реакции обратной транскрипции, предпочтительно где за реакцией обратной транскрипции следует реакция амплификации нуклеиновой кислоты, например обратная транскрипция - ПЦР. Предпочтительно эти характеристики инактивации ДНКазы достигают в отсутствие EDTA.
Как упомянуто выше, изобретение является особенно полезным для предупреждения или ограничения загрязнения и переноса геномной ДНК и, в частности, для предупреждения или уменьшения ложноположительных результатов вследствие переноса и/или загрязнения геномной ДНК.
В еще одном аспекте изобретения также предложен способ предупреждения или уменьшения ложноположительных результатов вследствие загрязнения геномной ДНК и/или переноса в реакциях обратной транскрипции, где в указанном способе используют ДНКазу, которая по существу необратимо инактивируется путем нагревания при температуре приблизительно 50°С в течение 5 минут и которая по существу специфична в отношении двухцепочечной ДНК, для деградации загрязняющей геномной ДНК и/или перенесенной двухцепочечной ДНК, присутствующих в реакционной смеси для обратной транскрипции или ее индивидуальных компонентах. Предпочтительно этих характеристик инактивации ДНКазы достигают в отсутствие EDTA.
ДНКаза по изобретению также подходит для применения в ограничении или уменьшении переноса во всех реакциях амплификации. Последнее происходит в результате того, что чем ниже температура инактивации ДНКазы, тем легче ее инактивировать во время процесса амплификации, и больше степень инактивации, которая может быть достигнута при любой заданной температуре, используемой на стадии инактивации, которая для удобства может представлять собой стадию денатурации ДНК (например, 94°С в течение 5 минут) для протоколов амплификации дцДНК.
В соответствии с настоящим изобретением также предложен способ удаления загрязняющей нуклеиновой кислоты из реакции амплификации нуклеиновой кислоты, при котором используют ДНКазу по изобретению.
Таким образом, ДНКазу по изобретению используют для деградации нецелевой двухцепочечной ДНК, представленной в смеси для реакции амплификации или ее индивидуальных компонентах. Таким образом, неспецифичная амплификация может быть уменьшена или исключена.
В частности, способ включает приведение в контакт смеси для реакции амплификации или ее индивидуальных компонентов, с ДНКазой по изобретению в условиях, которые дают возможность для расщепления какой-либо двухцепочечной ДНК; нагревание указанной реакционной смеси или ее индивидуальных компонентов для инактивации указанной ДНКазы и затем приведение указанной смеси или ее индивидуальных компонентов в контакт с указанной нуклеиновой кислотой-мишенью, которую предполагается амплифицировать.
Альтернативно, в этом аспекте изобретения предложено применение ДНКазы по изобретению для удаления загрязняющей нуклеиновой кислоты из смеси для реакции амплификации.
Как упомянуто выше, изобретение особенно полезно для предупреждения или ограничения переноса в реакциях амплификации нуклеиновых кислот и, в частности, для предупреждения или уменьшения ложноположительных результатов вследствие переноса.
В еще одном аспекте изобретения также предложен способ предупреждения или уменьшения ложноположительных результатов вследствие переноса в реакциях амплификации нуклеиновых кислот, включающий использование ДНКазы по изобретению для деградации перенесенной нецелевой двухцепочечной ДНК, представленной в смеси для реакции амплификации или ее индивидуальных компонентах.
ДНКазу по настоящему изобретению можно также использовать для удаления загрязняющей нуклеиновой кислоты из препаратов ДНК-полимеразы, а также использовать для удаления загрязняющей нуклеиновой кислоты из реакционных смесей для амплификации, содержащих ДНК-полимеразу. Низкая температура активации ДНКазы по настоящему изобретению означает, что инактивация ДНКазы после удаления загрязнения может быть осуществлена без вредного воздействия на полимеразу или при минимальном таком воздействии.
Изобретение особенно подходит для удаления загрязняющей нуклеиновой кислоты из так называемых ДНК-полимераз с горячим стартом. Разработаны многочисленные полимеразы с горячим стартом. Цель, лежащая в основе разработки ДНК-полимераз с горячим стартом, заключалась в модификации полимеразы для предотвращения действия фермента в качестве ДНК-полимеразы (способность удлинять полинуклеотидную последовательность с присоединенным праймером) до тех пор, пока смесь для реакции амплификации не достигнет температуры, близкой к оптимальной каталитической температуре ДНК-полимеразы, или по меньшей мере температуры, при которой отжиг праймера в достаточной степени является специфическим для последовательности, с тем, чтобы избежать или минимизировать неспецифическую амплификацию. Это происходит потому, что при более низких температурах праймеры могут отжигаться неспецифически с образцом нуклеиновой кислоты и приводить к неспецифическим продуктам амплификации, что, в свою очередь, может давать ложные результаты и/или оказывать ингибирующее действие на реакцию. Дополнительно, в некоторых случаях активность полимеразы является менее точной, и ошибки последовательности могут возникать в продуктах амплификации. Эта повышенная специфичность приводит к тому, что полимеразы с горячим стартом особенно подходят для применения для количественной ПЦР.
Один из подходов к разработке ДНК-полимераз с горячим стартом заключается в присоединении к полимеразе термолабильных групп, которые, будучи присоединенными, ингибируют или предупреждают каталитическое действие полимеразы, но которые диссоциируют от полимеразы при температурах, близких к оптимальной каталитической температуре полимеразы, или по меньшей мере температуре, при которой отжиг праймера в достаточной степени является специфическим для последовательности, для того, чтобы избежать или минимизировать неспецифическую амплификацию.
Подходящие термолабильные группы включают специфические в отношении полимеразы антитела и аффитела, другие специфические в отношении полимеразы связывающиеся с ней белки, специфические олигонуклеотидные аптамеры, неспецифические покрытия (например, воск) и ковалентные химические модификации аминокислот полимеразы (например, аминокислот в сайте активности). Удаление загрязнения таких полимераз с использованием ДНКазы, которая инактивируется при температуре, выше температуры активации полимеразы с горячим стартом, может означать, что свойства горячего старта полимеразы с горячим стартом могут быть нарушены. Настоящее изобретение благоприятным образом дает возможность для удаления загрязнений ДНК из препаратов полимераз с горячим стартом ДНКазой и последующей инактивации ДНКазы при температурах, которые ниже температуры активации горячего старта типичных полимераз с горячим стартом, и можно избежать такого негативного воздействия на свойства горячего старта полимераз с горячим стартом.
Обсуждавшиеся выше полимеразы с горячим стартом представляют собой всего лишь один из подходов к проведению ПЦР с горячим стартом. Другие подходы предупреждают контакт одного или более чем одного компонента реакционной смеси для ПЦР с оставшимися компонентами. Типично полимераза или нуклеиновая кислота-мишень изолированы за материалом или в нем (типично липиде, например, воске) с температурой плавления, близкой к температуре оптимальной каталитической температуре полимеразы, или по меньшей мере при температуре, при которой отжиг праймером в значительной степени специфичен для последовательности, для того, чтобы избежать или минимизировать неспецифическую амплификацию. Таким образом, ДНКаза по изобретению также дает возможность для удаления загрязняющей нуклеиновой кислоты этих так называемых "барьерных" комбинаций для ПЦР с горячим стартом без негативного воздействия на этот тип ПЦР с горячим стартом.
Таким образом, в изобретении предложен способ удаления загрязняющей нуклеиновой кислоты из ПЦР с горячим стартом, где указанная реакция представляет собой барьерную комбинацию для ПЦР с горячим стартом и/или использует ДНК-полимеразу с горячим стартом, при этом в указанном способе используют ДНКазы по изобретению.
Таким образом, ДНКазу по изобретению применяют для деградации нецелевой двухцепочечной ДНК, представленной в реакционной комбинации/смеси для ПЦР с горячим стартом. Таким образом, неспецифическая амплификация может быть уменьшена или устранена.
В частности, в способе осуществляют приведение в контакт комбинации/смеси для ПЦР с горячим стартом с ДНКазой по изобретению в условиях, которые дают возможность для расщепления какой-либо двухцепочечной ДНК; нагревание указанной реакционной комбинации/смеси для инактивации указанной ДНКазы и затем приведение указанной амплифицируемой нуклеиновой кислоты-мишени в контакт с оставшимися компонентами реакционной комбинации/смеси.
Альтернативно, в этом аспекте изобретения предложено применение ДНКазы по изобретению для удаления загрязняющей нуклеиновой кислоты из реакции ПЦР с горячим стартом ПЦР, где указанная реакция представляет собой барьерную реакцию с горячим стартом и/или включает полимеразу с горячим стартом.
Изобретение также является особенно полезным для предупреждения или ограничения переноса в реакциях ПЦР с горячим стартом, где указанные реакции представляют собой барьерные реакции с горячим стартом и/или включают ДНК-полимеразу с горячим стартом и, в частности, для предупреждения или уменьшения ложноположительных результатов вследствие переноса.
В еще одном аспекте изобретения также предложен способ предупреждения или уменьшения ложноположительных результатов вследствие переноса в реакциях ПЦР с горячим стартом, где указанные реакции представляют собой барьерные реакции с горячим стартом и/или включают ДНК-полимеразу с горячим стартом, где в указанном способе используют ДНКазу по изобретению для деградации перенесенной нецелевой двухцепочечной ДНК, представленной в комбинации/смеси реакции ПЦР с горячим стартом.
В изобретении также предложен способ удаления загрязняющей нуклеиновой кислоты из препарата ДНК-полимеразы с горячим стартом, при котором используют ДНКазы по изобретению. В этом способе также предложено применение ДНКаз по изобретению.
Таким образом, ДНКазу по изобретению используют для деградации двухцепочечной ДНК, представленной в препарате ДНК-полимеразы с горячим стартом. В частности, в способе осуществляют приведение в контакт препарата ДНК-полимеразы с горячим стартом с ДНКазой по изобретению в условиях, которые дают возможность для расщепления какой-либо двухцепочечной ДНК, представленной в препарате ДНК-полимеразы, и затем нагревают препарат для инактивации указанной ДНКазы.
В настоящем изобретении также предложен способ амплификации in vitro, обратной транскрипции или амплификации ПЦР с горячим стартом нуклеиновой кислоты-мишени, где указанная ПЦР с горячим стартом представляет собой барьерную реакцию с горячим стартом и/или включает ДНК-полимеразу с горячим стартом, отличающийся тем, что в указанном способе осуществляют стадию обработки реакционной смеси, реакционной комбинации или ее индивидуальных компонентов ДНКазой по изобретению перед началом амплификации или реакции обратной транскрипции.
"Обратная транскрипция" представляет собой процесс, в котором РНК-зависимая ДНК-полимераза катализирует образование молекулы ДНК, комплементарной матрице РНК (кДНК). Более конкретно, полимераза катализирует полимеризацию дезоксирибонуклеозидтрифосфатов в последовательность, которая комплементарна (т.е. следует правилам комплементарности оснований Уотсона-Крика) матричной последовательности РНК с праймером.
Идентифицированы многочисленные ферменты, обладающие способностью катализировать эту реакцию, и примеры включают обратную транскриптазу HIV (вируса иммунодефицита человека), обратную транскриптазу AMV (вируса миелобластоза птиц), обратную транскриптазу M-MLV (вируса лейкоза Молони), полимеразу С therm., и полимеразу Tth, но не ограничиваются ими. Эти ферменты имеют диапазон оптимальных рабочих температур. Ферменты, выделенные из организмов, таких как вирусы, которые инфицируют животных-хозяев, имеют оптимальную рабочую температуру приблизительно 37°С. Тем не менее, идентифицированы термостабильные обратные транскриптазы, и их получают путем мутации обратных транскриптаз дикого типа, и именно эти термостабильные ферменты типично используют в реакциях обратной транскрипции в лаборатории. На нижней границе диапазона расположены AMV с рабочим диапазоном от 42 до 60°С, тогда как обратная транскрипционная активность Tth ДНК-полимеразы и C. therm. ДНК-полимеразы имеют рабочие диапазоны от 55 до 70°С и от 60 до 70°С соответственно.
Обычно полная реакционная смесь для обратной транскрипции содержит фермент обратной транскрипции, матрицу РНК, подходящие праймеры, которые могут связываться с матрицей и от которых обратная транскриптаза может начать полимеризацию, dNTP и подходящей буфер. Инкубация смеси при рабочей температуре обратной транскриптазы приводит в результате к продукции кДНК. После завершения реакции обратной транскрипции кДНК можно использовать непосредственно в экспериментах по секвенированию или генотипированию или можно использовать в протоколах по клонированию или детекции.
Под "ферментом обратной транскрипции" понимают любой фермент, который обладает обратной транскрипционной активностью (т.е. способностью катализировать полимеризацию комплементарной цепи ДНК с последовательностью РНК матрицы с праймером или РНК зависимой ДНК-полимеразной активностью). Эта активность может представлять собой единственную активность фермента или, более типично, может представлять собой одну из активностей фермента (например, обратная транскриптаза HIV, обратная транскриптаза M-MLV, обратная транскриптаза AMV, Tth ДНК-полимераза, С. therm, полимераза). Типичные дополнительные активности полимеразы могут включать РНКзу Н, ДНК зависимую ДНК-полимеразу, ДНК-РНК расплетающую активность, Mn2+ зависимую эндонуклеазу. Тем не менее, предпочтительно, чтобы активность РНКзы Н и/или эндонуклеазная активность были минимальными или отсутствовали.
"Препарат фермента обратной транскрипции" представляет собой любой материал, типично раствор, как правило, водный, содержащий фермент обратной транскрипции. В частности, он относится к коммерчески приготовленным препаратам фермента обратной транскрипции, т.е. реагенту-ферменту обратной транскрипции, который может поставляться коммерческим поставщиком лабораторных ферментов, хотя разведенные, скорректированные и/или модифицированные варианты таких препаратов также охвачены этим термином. Препарат фермента обратной транскрипции также может быть приготовлен из фермента обратной транскрипции, который получен из бактериального источника, экспрессирующего фермент обратной транскрипции в природе, и/или который содержит экспрессирующуюся кассету, кодирующую фермент обратной транскрипции. Препарат может быть очищен в такой степени, которая сравнима с исходным препаратом фермента обратной транскрипции, приготовленным непосредственно из бактериального источника.
Термин "реакция амплификации нуклеиновой кислоты" относится к любым способам in vitro увеличения количества копий последовательности-мишени нуклеиновой кислоты. Предпочтительно способы включают "термоциклирование", т.е. включают цитирование при высокой температуре. Способы амплификации включают ПЦР и ее модификации, 3SR, SDA, ЛРА или ЛЦР и ИПА и их модификации, но не ограничиваются ими. ПЦР, ИПА и ЛЦР и их модификации представляют собой способы термоциклирования. Способы могут приводить в результате к линейному или экспоненциальному увеличению количества копий последовательности-мишени. "Модификации" охватывают амплификацию в режиме реального времени, количественную и полуколичественную амплификацию, конкурентную амплификацию и т.д., но не ограничиваются ими.
Нуклеиновая кислота-мишень может представлять собой ДНК или РНК в зависимости от выбранного способа амплификации. Например, для ПЦР мишень представляет собой ДНК, хотя при комбинировании со стадией обратной транскрипции, в качестве мишени можно рассматривать последовательность РНК. 3SR непосредственно амплифицирует последовательности-мишени РНК.
Термин "реакционная смесь для амплификации/обратной транскрипции" относится к любому раствору, как правило, водному, содержащему различные реагенты, используемые для амплификации/обратной транскрипции нуклеиновой кислоты-мишени. Они включают ферменты, водные буферы, соли и нуклеозидтрифосфаты. Термин относится к смесям, которые содержат все необходимые компоненты для осуществления успешной реакции амплификации, и к смесям, которые являются не полными и, таким образом, содержат лишь некоторые (например, по меньшей мере, 2, 3 или 4) из требуемых компонентов. Если реакционная смесь обозначена как "полная", то она содержит все компоненты, необходимые для обратной транскрипции и/или амплификации.
"ДНК-полимераза с горячим стартом" представляет собой ДНК-полимеразу, которая модифицирована, типично путем добавления термолабильных молекулярных единиц, для увеличения температуры, при которой может быть проведена обнаруживаемая полимеризация ДНК полинуклеотида с праймером. Температура, при которой ДНК-полимераза с горячим стартом может приводить к обнаруживаемым уровням ДНК полимеризации, предпочтительно достигает оптимальной каталитической температуры полимеразы.
Термин "препарат ДНК-полимеразы с горячим стартом" относится к любому материалу, типично раствору, как правило, водному, содержащему ДНК-полимеразу с горячим стартом. В частности, он относится к коммерчески приготовленным препаратам ДНК-полимеразы с горячим стартом, т.е. реагенту ДНК-полимеразы с горячим стартом, который может поставляться коммерческим поставщиком лабораторных ферментов, хотя разведенные, скорректированные и/или модифицированные варианты таких препаратов также охвачены этим термином. В качестве препарата ДНК-полимеразы с горячим стартом также может быть использован препарат ДНК-полимеразы с горячим стартом, полученный из бактериального источника, экспрессирующего полимеразу в естественных условиях, и/или который содержит экспрессирующуюся кассету, кодирующую полимеразу. Препарат может быть очищен в такой степени, которая сравнима с исходным препаратом полимеразы, приготовленным непосредственно из бактериального источника. Типично, препарат дополнительно обработан таким образом, чтобы прикрепить к полимеразе блокирующие молекулы, обеспечивающие горячий старт.
"Реакция ПЦР с горячим стартом" представляет собой реакцию амплификации путем ПЦР, в которой обнаруживаемая полимеризация с ДНК полинуклеотида с праймером проявляется лишь при температуре, достигающей оптимальной каталитической температуры полимеразы. Предпочтительные температуры для обнаруживаемой полимеризации следует обсуждать в сочетании с ДНК-полимеразами с горячим стартом.
"Смесь для ПЦР с горячим стартом" представляет собой реакционную смесь для ПЦР, определенную выше, содержащую полимеразу с горячим стартом.
"Барьерная комбинация для ПЦР с горячим стартом" представляет собой реакционный сосуд, содержащий два или более чем два компонента реакционной смеси для ПЦР, где по меньшей мере один компонент изолирован от другого(их) компонента(ов) за или в материале, имеющем температуру плавления, соответствующую температурам обнаруживаемой полимеризации ДНК, как определено выше. Предпочтительно материал представляет собой липид, например воск.
Под "загрязнением" понимают присутствие в реакционной смеси нуклеиновой кислоты, которая может действовать в качестве матрицы для обратной транскрипции и/или амплификации, которая не представляет собой часть совокупности нуклеиновой кислоты, которая рассматривается в качестве мишени для обратной транскрипции/амплификации. Праймеры, используемые в реакционной смеси, не являются примесями.
Предполагается, что термин "удаление загрязняющей нуклеиновой кислоты" охватывает предупреждение и уменьшение загрязнения нуклеиновыми кислотами.
Термин "перенос" используется для описания любой нуклеиновой кислоты, которая случайно или непреднамеренно попадает в реакционную смесь, в частности последовательности-мишени, перенесенные из предшествующих реакций амплификации или обратной транскрипции.
Термин "ложноположительный результат" относится к результату, который, по-видимому, демонстрирует, что исследуемый образец нуклеиновой кислоты содержит последовательности-мишени, но где амплифицированный продукт происходит в результате переноса и/или в случае реакций амплификации, основанных на обратной транскрипции, возможно геномной ДНК. Очевидно, что уменьшение ложноположительных результатов, которое предложено в изобретении, особенно благоприятно в криминалистической и диагностической областях. Способы по изобретению дают возможность для увеличения специфичности амплификации нуклеиновой кислоты.
Термин "ДНКаза" относится к ферменту, который гидролизует фосфодиэфирную связь в скелете ДНК и не является специфичным в отношении нуклеотидной последовательности.
Под термином "по существу необратимо инактивируется" понимают, что после нагревания фермент по меньшей мере на 95% инактивируется, предпочтительно на 98% инактивируется, более предпочтительно фермент на 100% инактивируется. Процентная доля инактивации может быть для удобства измерена путем инкубации образца ДНК (например, продукта ПЦР 500 п.о.) в течение 3 ч с инактивированной ДНКазой или неинактивированной ДНКазой в подходящем буфере (например, Tris, HEPES, PBS (физиологический раствор, забуференный фосфатом)) при 37°С; разделении продуктов реакции на агарозном геле с бромистым этидием путем электрофореза и измерения относительных интенсивностей флуоресценции полос ДНК под УФ (ультрафиолетовым) светом (Пример 2). Альтернативные способы могут быть разработаны специалистами в данной области техники для измерения относительных активностей инактивированной и неинактивированной ДНКазы. Например, можно использовать относительные изменения флуоресценции образцов ДНК, содержащих SYBR зеленый. Дополнительные способы представляют собой анализ Кунитца (Kunitz, М; 1950, S. Gen Physiol, 33:363 и Пример 1 и модифицированный анализ Кунитца, разработанный Yamamoto (Yamamoto, M; 1971, Biochim Biophys Acta, 228:95 и Пример 4).
Даже когда температура реакционной смеси возвращается к комнатной, ДНКаза не восстанавливает свою активность и по существу отсутствует остаточная активность; конкретно менее чем 5%, предпочтительно менее чем 2%, наиболее предпочтительно не остается никакая обнаруживаемая активность ДНКазы.
По существу необратимая инактивация предпочтительно возникает в течение 5 минут инкубации при температуре точно или приблизительно 50°С, например от 48 до 52°С. Например, в течение 1, 2 или 3 минут инкубации при 50°С. ДНКаза по изобретению может быть по существу необратимо инактивирована при меньших температурах или в течение более коротких периодов, но в соответствии с изобретением нагревание в течение 5 минут при приблизительно 50°С должно быть достаточным для по существу необратимой инактивации фермента. Специалисту в данной области техники должно быть понятно, что изменение одного из этих двух параметров может быть компенсировано путем корректировки другого. Например, увеличение температуры инактивации может дать возможность для уменьшения времени инкубации. Наоборот, увеличение длительности инкубации может дать возможность для использования меньшей температуре инактивации. Например, ДНКаза в соответствии с изобретением может быть инактивирована в течение 1 или 2 минут инкубации при температуре 55°С, в течение 3 минут инкубации при температуре 52°С, в течение 4 минут инкубации при температуре 51°С, в течение 10 минут инкубации при температуре 49°С или в течение 15 минут инкубации при температуре 48°С. Безусловно, как понятно специалисту в данной области техники и представлено в примерах, когда ДНКазу по изобретению используют в способах по изобретению, можно использовать длительность инкубации больше пяти минут и температуру инактивации больше чем приблизительно 50°С при практическом применении (например, инактивация может осуществляться при каждой из температур 50°С, 55°С, 65°С или 94°С в течение каждого периода из 15, 30 или 60 минут; 60°С в течение 15 минут; или 95°С в течение 10 минут). Тем не менее, в соответствии с изобретением ДНКаза должна демонстрировать существенную инактивацию при инкубации при температуре точно или приблизительно 50°С в течение 5 минут.
Температуры и времена инактивации ДНКазы должны быть оценены путем инкубации ДНКазы в растворе, который имитирует типичную ПЦР, или буфере для обратной транскриптазы (например 25 мМ Tris/HCl, pH 8,5, 5 мМ MgCl2). EDTA предпочтительно должна отсутствовать. ДНКаза должна быть представлена в концентрации от приблизительно 0,01 Ед./мкл до 10 Ед./мкл, предпочтительно от 0,05 до 5 Ед./мкл, например от 0,5 до 1,5 Ед./мкл.
Инактивация при любой данной температуре может быть усилена с точки зрения длительности и/или скорости путем присутствия агента, восстанавливающего дисульфидные связи (т.е. агента, который ингибирует и/или разрушает дисульфидные связи между двумя или более чем двумя цистеиновыми остатками в белке) в инактивирующем буфере. Примеры таких агентов включают DTT, 2-меркаптоэтанол, 2-меркаптоэтиламин·НСl, TCEP·HCl (Трис(2-карбоксиэтил)фосфин гидрохлорид), N-этилмалеимид. Предпочтительным является DTT. Альтернативно, агент, восстанавливающий дисульфидные связи (например, DTT), можно использовать для уменьшения температуры инактивации, которая требуется для конкретной длительности стадии инактивации. Специалист в данной области техники сможет определить подходящие для его целей концентрации агента, восстанавливающего дисульфидные связи, которые могут улучшить инактивацию, но не будут оказывать вредного воздействия на последующие реакции. Например, DTT для удобства может быть включен на стадии в концентрации от 0,05 до 50 мМ. DTT стандартно используется в реакциях обратной транскрипции в концентрации от приблизительно 1 до 10 мМ и часто используется в реакциях ПЦР.
Предпочтительно инактивация ДНКазы в способах по изобретению происходит при концентрации DTT от 0,1 до 10 мМ, предпочтительно от 0,5 до 5 мМ и наиболее предпочтительно от 1 до 2 мМ. Для стандартной оценки температуры инактивация предпочтительно используется буфер 25 мМ Tris/HCl, рН 8,5, 5 мМ MgCl2 и 1 мМ DTT.
Линейная двухцепочечная ДНК и суперспирализованная кольцевая ДНК представляют собой субстраты фермента по изобретению. Фермент обладает небольшой, незначительной или по существу не обнаруживаемой активностью в отношении одноцепочечной ДНК, такой как праймеры для амплификации/обратной транскрипции. Другими словами, ДНКаза по существу специфична в отношении двухцепочечной ДНК.
Под "по существу специфичной в отношении двухцепочечной ДНК" понимают, что ДНКаза расщепляет двухцепочечную ДНК, но обладает небольшой, незначительной или по существу не обнаруживаемой активностью в отношении одноцепочечной ДНК в концентрациях от 0,01 до 0,05 Ед./мкл. Предпочтительно отсутствует обнаруживаемая активность в отношении одноцепочечной ДНК в таких концентрациях. Специалист в данной области техники может легко поставить эксперимент для сравнения относительной активности ДНКазы в отношении одно- и двухцепочечной ДНК. Anisimova et al. (BMC Biochemistry, 2008, 9:14) описали такой эксперимент. Кратко, 2 единицы Кунитца ДНКазы в тесте инкубировали с ДНК фагом М13 (одноцепочечным) или ДНК фагом лямбда (двухцепочечным) в 50 мМ Tris-HCl, рН 7,5, 5 мМ MgCl2 (конечный реакционный объем 30 мкл) в течение одного часа, и продукты разделяли на агарозном геле с бромистым этидием. Активность против одноцепочечной и/или двухцепочечной ДНК обнаруживали по положению полос относительно необработанных контролей. Еще один подход более подробно описан в примере 6. В этом подходе специфичность ДНКазы в отношении двух- и одноцепочечной ДНК может быть тестирована путем измерения увеличения флуоресценции с олигонуклеотидов, меченных флуорофором FAM (флуоресцеин) по 5' концу, и TAMRA по 3' концу. Испущенный свет с FAM абсорбируется (гасится) TAMRA, когда две метки находятся рядом друг с другом. Расщепление олигонуклеотида ДНКазой приводит в результате к разделению FAM и TAMRA и увеличению флуоресценции с FAM, которое может быть измерено во флуориметре с длиной волны возбуждения 485 нм и длиной волны излучения 520 нм. Субстрат-двухцепочечная ДНК может быть приготовлена путем смешивания меченого олигонуклеотида со вторым олигонуклеотидом, комплементарным меченому олигонуклеотиду. Безусловно, что аналогично могут использоваться другие подходящие пары флуорофоров.
В случае реакции обратной транскрипции эти характеристики дают возможность для включения ДНКазы в реакционную смесь для обратной транскрипции, содержащей образец РНК, праймеры, нуклеотиды, обратную транскриптазу и буферы (т.е. полную реакционную смесь) и быструю деградацию перенесенного материала и геномной ДНК, например при комнатной температуре. Эти характеристики также дают возможность для включения ДНКазы в полную смесь для реакции амплификации, основанной на одностадийной обратной транскрипции.
Эти характеристики также дают возможность для включения ДНКазы в смесь для реакции амплификации, содержащей праймеры, нуклеотиды, ДНК-полимеразу и буферы, и быструю деградацию перенесенного материала, например, при комнатной температуре.
Благоприятным образом, термолабильная ДНКаза по изобретению является полностью функциональной в полной смеси для реакции амплификации и совместима со стандартными реагентами для амплификации in vitro и условиями. Фермент также должен обладать способностью удаления подходящих количеств загрязняющей геномной ДНК и/или перенесенной ДНК из реакционной смеси, обычно фг- или пг-уровни, но предпочтительно до 1 нг. Предпочтительно ДНКаза способна подвергнуть дергадации всю перенесенную ДНК в течение 5 минут при комнатной температуре, более предпочтительно в течение 3 минут, наиболее предпочтительно в течение 2 минут.
Увеличение температуры реакционной смеси до температуры инактивации ДНКазы по изобретению (приблизительно 50°С) в течение короткого времени (например, 5 минут) необратимо инактивирует ДНКазу по изобретению.
В случае реакций обратной транскрипции это для удобства можно совместить со стадией обратной транскрипции. В случае реакций амплификации ДНК (включая ПЦР с горячим стартом) амплифицируемые и анализируемые образцы нуклеиновой кислоты (т.е. нуклеиновой кислоты-мишени) затем могут быть добавлены, и амплификация начнется. Даже когда температура реакционной смеси падает во время термоциклирования и после амплификации или обратной транскрипции, копии последовательности-мишени не деградируют, поскольку ДНКаза необратимо инактивирована. В соответствии с настоящим изобретением особенно благоприятно, чтобы обратная транскриптаза и/или ДНК-полимераза могли быть включены в реакционные смеси для обратной транскрипции и/или амплификации во время прохождения стадии удаления загрязнения и последующей инактивации. Это является результатом мягких условий, приводящих в результате к инактивации ДНКазы (приблизительно 50°С в течение 5 минут), таким образом, что дополнительный потенциальный источник загрязнения устраняется.
Предпочтительно ДНКаза обладает минимальной нуклеазной активностью в отношении цепи ДНК цепь дуплекса ДНК:РНК. Под "минимальной" понимают, что ДНКаза обладает нуклеазной активностью в отношении цепи ДНК дуплекса ДНК:РНК, которая составляет менее чем 40% относительно ее активности в отношении двухцепочечной ДНК. Предпочтительно ДНКаза обладает активностью в отношении дуплексов ДНК:РНК, которая составляет менее чем 30% или менее чем 20% относительно ее активности в отношении двухцепочечной ДНК.
Именно эти конкретные характеристики предпочтительных ДНКаз по изобретению (т.е. быстрая по существу необратимая инактивация, специфичность в отношении двойной цепи и предпочтительно минимальная нуклеазная активность в отношении дуплекса ДНК:РНК) делают эти ДНКазы исключительно подходящими для удаления загрязнения в одностадийных протоколах амплификации с обратной транскрипцией. Последнее происходит потому, что полная реакционная смесь может быть очищена от загрязнений без вероятности нежелательной деградации продуктов амплификации или обратной транскрипции в одну стадию и без необходимости добавления дополнительных материалов. Последнее минимизирует риск загрязнения (включая риск заражения геномной ДНК) без потери чувствительности путем нежелательного расщепления исходного продукта обратной транскрипции и/или продукта амплификации.
Фермент ДНКаза, используемая в вышеприведенных способах, сама составляет еще один аспект изобретения. Таким образом, в этом аспекте изобретению предложена ДНКаза, которая по существу необратимо инактивируется путем нагревания при температуре приблизительно 50°С в течение 5 минут и которая по существу специфична в отношении двухцепочечной ДНК. Предпочтительно этих характеристик инактивации достигают в отсутствие EDTA).
Хотя понятно, что любая термолабильная ДНКаза, обладающая описанными выше характеристиками, может быть подходящей для применения в способах по изобретению, модифицированные ДНКазы, полученные из ДНКазы Pandalus borealis или похожей ДНКазы другого, предпочтительно морского организма, в которых конкретный остаток пролина модифицирован, удален или замещен, образуют еще один аспект настоящего изобретения. Организм может представлять собой прокариотический или эукариотический организм. Под "прокариотическим" понимают любой организм, лишенный клеточного ядра, т.е. любой организм доменов бактерий и архей. Предпочтительно организм представляет собой бактерию. Более предпочтительно организм представляет собой эукариота, например организм, классифицированный в таксономических царствах животных, растений, грибов или простейших, например организм типа/деления Acanthocephala, Acoelomorpha, Annelida, Arthropoda, Brachiopoda, Bryozoa, Chaetognatha, Chordata, Cnidaria, Ctenophora, Cycliophora, Echinodermata, Echiura, Entoprocta, Gastrotricha, Gnathostomulida, Hemichordata, Kinorhyncha, Loricifera, Micrognathozoa, Mollusca, Nematoda, Nematomorpha, Nemertea, Onychophora, Orthonectida, Phoronida, Placozoa, Platyhelminthes, Porifera, Priapulida, Rhombozoa, Rotifera, Sipuncula, Xenoturbellida, Anthocerotophyta, Bryophyta, Marchantiophyta, Lycopodiophyta, Pteridophyta, Pteridospermatophyta, Coniferophyta, Cycadophyta, Ginkgophyta, Gnetophyta, Anthophyta (или Magnoliophyta), Chytridiomycota, Deuteromycota, Zygomycota, GIomeromycota, Ascomycota или Basidiomycota. Также достойны внимания организмы царства животных, например беспозвоночные и позвоночные. Более предпочтительно организм выбран из организмов типа Arthropoda, например организм в подтипах Crustacea, Hexpoda, Chelicerata или Myriapoda, например организм в классах Crustacea в Branchiopoda, Remipedia, Cephalocarida, Maxillopoda, Ostracoda или Malacostraca, предпочтительно Malacostraca и более предпочтительно организм в порядке Decapoda. Организм может быть классифицирован в семейство Pandalidae, например в родах Anachlorocurtis, Atlantopandalus, Austropandalus, Calipandalus, Chelonika, Chlorocurtis, Chlorotocella, Chlorotocus, Dichelopandalus, Dorodotes, Heterocarpus, Miropandalus, Notopandalus, Pandalina, Pandalopsis, Pandalus, Pantomus, Peripandalus, Plesionika, Procletes, Pseudopandalus или Stylopandalus; семейство Lithodidae, например в родах Cryptolithodes, Glyptolithodes, Lithodes, Lopholithodes, Neolithodes, Paralithodes, Paralomis, Phyllolithodes или Rhinolithode; или семейство Penaeidae, например в родах: Farfantepenaeus, Fenneropenaeus, Litopenaeus или Marsupenaeus. Организм предпочтительно представляет собой организм, который эволюционировал таким образом, чтобы жить в холодной окружающей среде, например холодной среде морей или водоемов. Организм предпочтительно выбран из, например, Paralithodes camtschaticus (королевский краб), Marsupenaeus japonicus (креветка курума) или Penaeusjaponicus. В других воплощениях ДНКаза получена из видов организмов, которые не представляют собой прокариоты, например не представляет собой бактерию, например не представляют собой психротрофную бактерию.
Также в объем настоящего изобретения включены ферментативно активные фрагменты этих модифицированных ДНКаз.
Таким образом, в еще одном аспекте изобретения предложена ДНКаза или ее ферментативно активный фрагмент, где указанная ДНКаза имеет последовательность SEQ ID NO: 1 или последовательность, которая по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, 80%, 90% или 95%, например, по меньшей мере на 98% идентична ей, но где остаток пролина в положении 237 в SEQ ID NO: 1, или эквивалентный пролин в другой последовательности модифицирован, удален или замещен, где указанная ДНКаза или ее ферментативно активный фрагмент по существу необратимо инактивируются путем нагревания при температуре приблизительно 50°С в течение 5 мин и которая по существу специфична в отношении двухцепочечной ДНК.
SEQ ID NO: 1 представляет собой аминокислотную последовательность транслированного фрагмента кДНК для ДНКазы Pandalus borealis. Последовательность кДНК представлена в SEQ ID NO: 2. SEQ ID NO: 1 содержит сигнальную пептидную последовательность MIGRTTFIALFVKVLTIWSFTKG (SEQ ID NO: 9). Зрелая форма ДНКазы Panda/us borealis представлена на SEQ ID NO: 5 (т.е. последовательность SEQ ID NO: 1 без сигнального пептида (SEQ ID NO: 9). Таким образом, пролин в положении 237 в SEQ ID NO: 1 представляет собой то же самое положение, как пролин в положении 214 в SEQ ID NO: 5.
Таким образом, в изобретении также предложена ДНКаза или ее ферментативно активный фрагмент, где указанная ДНКаза имеет последовательность SEQ ID NO: 5 или последовательность, которая по меньшей мере на 60%, предпочтительно по меньшей мере на 70%, 80%, 90% или 95%, например, по меньшей мере на 98% идентична ей, но где остаток пролина в положении 214 в SEQ ID NO: 5 или эквивалентный пролин в другой последовательности модифицирован, удален или замещен, где указанная ДНКаза или ее ферментативно активный фрагмент по существу необратимо инактивирован путем нагревания при температуре приблизительно 50°С в течение 5 мин и которая по существу специфична в отношении двухцепочечной ДНК.
Ферментативно активные фрагменты и варианты SEQ ID NO: 1 демонстрируют по меньшей мере 70%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 95% и наиболее предпочтительно по меньшей мере 99% ферментативной функции зрелого фермента SEQ ID NO: 5 (т.е. способность гидролизовать фосфодиэфирную связь в скелете ДНК без специфичности в отношении нуклеотидной последовательность). Как обсуждается, активность ДНКазы может быть легко оценена с использованием стандартных способов.
Процентная доля идентичности последовательности в соответствии с изобретением может быть рассчитана с использованием любого из широко доступных алгоритмов (например, с использованием программы совмещения множества последовательностей Clustal W2 (http://www.ebi.ac.uk/Tools/clustalW2) с использованием параметров по умолчанию (штраф за пробел в ДНК = 15,0; штраф за удлинение пробела в ДНК = 6,66; матрица ДНК = идентичность; штраф за пробел в белке = 10,0; штраф за удлинение пробела в белке = 0,2; матрица белка = Gonnet; Белок/ДНК ENDGAP = -1; Белок/ДНК GAPDIST = 4).
"Эквивалентные пролиновые остатки в других последовательностях", отличающихся от SEQ ID NO: 1 или 5, легко могут быть идентифицированы с использованием стандартных способов совмещения последовательности, таких как Clustal W2, с получением совмещений, таких как представленные на Фиг.11.
Предпочтительно ДНКаза по изобретению или ее фрагмент имеют последовательность ДНКазы, полученной из видов, классифицированных в любую из упомянутых выше таксономических групп, например типа Arthropoda или подтипов Crustacea, Hexpoda, Chelicerata и Myriapoda, например Pandalus borealis, Paralithodes camtschaticus (королевский краб), Marsupenaeusjaponicus (креветка курума) или Penaeus japonicas, но где эквивалент пролинового остатка в положении 237 в SEQ ID NO: 1 модифицирован, удален или замещен. Предпочтительной является ДНКаза из Pandalus borealis, где эквивалент пролинового остатка в положении 237 в SEQ ID NO: 1 модифицирован, удален или замещен.
В наиболее предпочтительном воплощении ДНКаза по изобретению имеет аминокислотную последовательность SEQ ID NO: 3 или 7.
Под "замещением" пролина (например, по остатку 237 в SEQ ID NO: 1 / остатку 214 в SEQ ID NO: 5) понимают, что пролиновый остаток заменен на другую встречающуюся в природе аминокислоту, типично кодируемую генетически, или аминокислотный аналог. Предпочтительно пролин заменен на аланин, глицин, серин или цистеин.
Под "модификацией" пролина понимают, что стереохимические свойства остатка пролина изменены, например, путем замещения его боковой цепи отличающейся группой, модифицирующей саму композицию боковой цепи или замещающей водород, противоположный боковой цепи отличающейся боковой группой.
В изобретении также предложены молекулы нуклеиновой кислоты, кодирующие ДНКазы по изобретению. Нуклеотидные последовательности, соответствующие аминокислотной последовательности SEQ ID NO: 3 и 7, раскрыты в SEQ ID NO: 4 и 8. Вырожденность генетического кода означает, что SEQ ID NO: 4 и 8 представляют собой лишь две из множества возможных нуклеотидных последовательностей.
В изобретении также предложено применение описанных выше конкретных ДНКаз в качестве агента для удаления загрязнения в способах амплификации нуклеиновой кислоты. Применение конкретных описанных выше ДНКаз в описанных здесь способах удаления загрязнения представляет собой особенно предпочтительное воплощение изобретения.
Способ выделения и очистки описанной выше ДНКазы или ее ферментативно активного фрагмента представляет собой еще один аспект настоящего изобретения. Таким образом, в этом аспекте изобретения предложен такой способ, включающий экспрессию указанной ДНКазы или ее фрагмента в подходящей клетке-хозяине (например, Pichia pastoris; E.coli; S. cereviciae, клетках насекомых, инфицированных бакуловирусом), и последующее выделение ДНКазы из указанных клеток-хозяев и/или сред, в которых выращивают указанные клетки. Экспрессия указанной ДНКазы или ее фрагмента может быть достигнута путем включения в подходящую клетку-хозяина экспрессирующегося вектора, кодирующего указанную ДНКазу или ее фрагмент, например экспрессирующийся вектор, содержащий молекулу нуклеиновой кислоты, кодирующую аминокислотную последовательности SEQ ID NO: 3 и 7, например молекулу нуклеиновой кислоты, содержащую нуклеотидные последовательности SEQ ID NO: 4 или 8. Клетки-хозяева, содержащие эти экспрессирующиеся кассеты и молекулы нуклеиновой кислоты, также входят в объем изобретения.
Фермент ДНКаза может быть отделен или выделен из клеток-хозяев/культуральных сред с использованием любого из способов очистки белка, известных в области техники и широко описанных в литературе, или любой их комбинации. Такие способы могут включать, например, осаждение, ультрафильтрацию, диализ, различные хроматографические способы, например гель-фильтрацию, ионообменную хроматографию, аффинную хроматографию, электрофорез, центрифугирование и т.д.
Аналогично, экстракт из клеток-хозяев также может быть приготовлен с использованием способов, хорошо известных в области техники, например гомогенизации, замораживания-оттаивания и т.д, и из этого экстракта может быть очищена ДНКаза по изобретению.
Обнаружено, что протокол очистки, основанный на комбинации ионообменной хроматографии и аффинной хроматографии, например, на колонке сефарозы, например красной сефарозы (Pharmacia Biotech, Sweden) или голубой сефарозы (GE Healthcare), может легко использоваться для выделения фермента.
Более конкретно, экстракт можно подвергать ионообменной хроматографии, и белок элюируют градиентом NaCl. Фракции, содержащие активность ДНКазы, могут быть подвергнуты диализу и затем нанесены на аффинную колонку с последующей окончательной элюцией NaCl.
В настоящем изобретении также предложены наборы, которые содержат по меньшей мере ДНКазу по изобретению. Наборы также могут содержать некоторые или все необходимые реагенты, буферы, ферменты и т.д. для осуществления реакции амплификации нуклеиновой кислоты и/или обратной транскрипции. Более конкретно, набор может содержать нуклеотидтрифосфаты (включающие dNTPαS для SDA), олигонуклеотидные праймеры, обратные транскриптазы, предпочтительно способные функционировать при 50°С, ДНК-полимеразы, предпочтительно термостабильную полимеразу, такую как Taq полимераза или Bst полимераза (и их варианты с горячим стартом) или, в случае ЛРА, ДНК лигазу (предпочтительно термостабильную ДНК лигазу, такую как Ampligase®, или раскрытую в US 6280998, выделенную из Pyrococcus furiosus) или рестрикционный фермент (предпочтительно термостабильный рестрикционный фермент, такой как BsoB1). ДНКаза может находиться в одном компартменте с обратной транскриптазой, ДНК-полимеразой, цепьзамещающей полимеразой или ЛЦР лигазой.
В настоящем изобретении также предложены композиции, содержащие ДНКазу по изобретению и один или более чем один необходимый реагент для проведения реакции и способов амплификации нуклеиновой кислоты и/или реакции обратной транскрипции, например, компоненты, описанные выше. Типично, такие композиции являются водными и забуферены стандартным буфером, таким как Tris, HEPES и т.д.
Способы обратной транскрипции, безусловно, в настоящее время представляют собой стандарт в области техники и могут быть осуществлены с использованием любого известного или стандартного реагента и способов.
В типичном протоколе обратной транскрипции стадия удаления загрязнения может просто включать инкубацию реакционной смеси для обратной транскрипции, содержащей ДНКазу, в течение короткого периода времени, например от 1 до 30 минут, при комнатной температуре, для удобства от 2 до 15 минут. Время этой инкубации не является критичным и может варьировать в зависимости от конкретной используемой ДНКазы и концентрации и других компонентов реакционной смеси. Температура может представлять собой любую температуру, при которой фермент является активным, т.е. ниже температуры инактивации (например, 37°С), для удобства температура является комнатной.
Такая реакционная смесь может, как упомянуто выше, содержать все необходимые реагенты для реакции обратной транскрипции.
Типичная смесь для обратной транскрипции может, например, включать:
В вышеприведенном типичном примере можно использовать любую комбинацию объемов стерильной дистиллированной воды и экспериментальной матрицы, а также общий объем реакционной смеси (включающей буфер, dNTP, праймеры, ферменты и растворы MgCl2), равный 50-100 мкл. Тем не менее, альтернативные конечные объемы можно использовать в зависимости от выбора для достижения, например, близких или других желаемых конечных концентраций реагентов. Можно использовать любой удобный или имеющийся в продаже буфер для обратной транскрипции. Подходящий 5Х буфер для обратной транскрипции может быть 250 мМ Tris-HCl (pH 8,5 при 25°С), 40 мМ MgCl2, 150 мМ KCl, 5 мМ DTT. Буфер для обратной транскрипции может быть приобретен в Fermentas.
В зависимости от уровня потенциального загрязнения количество необходимой ДНКазы может варьировать. Для короткой стадии инкубации (0-15 минут при комнатной температуре), как правило, более чем достаточно 2,0 Ед./50 мкл реакционной смеси. Подходящим является от 0,1 до 2,0 Ед./50 мкл реакционной смеси, и предпочтительна активность приблизительно 0,5 Ед./50 мкл реакционной смеси, например от 0,2 до 1,0 Ед./50 мкл реакционной смеси). В концентрации 2,0 Ед./50 мкл реакционной смеси обнаруживается некоторая активность ДНКазы в отношении двухцепочечной молекулы, и, таким образом, предпочтительны приведенные выше активности. Одна единица фермента определяется как количество, которое в анализе Кунитца или модифицированном Ямамото анализе Кунитца (выше) увеличивает поглощение при 260 нм на 0,001 в минуту. После инкубации ДНКазу инактивируют путем нагревания реакционной смеси. Для удобства это может быть осуществлено путем нагревания на стадии обратной транскрипции, например, приблизительно 50°С в течение 30 минут.
Для удобства способ амплификации, включающий стадию удаления загрязнения с использованием ДНКазы по изобретению, включает ПЦР или основан на ней. Способы ПЦР являются стандартными в области техники и могут быть осуществлены с использованием любого известного или стандартного реагента и способов.
В типичном протоколе реакции ПЦР стадия удаления загрязнения может просто включать инкубацию смеси для реакции амплификации, содержащей ДНКазу, в течение короткого периода времени, например от 1 до 10 минут, при комнатной температуре, для удобства от 2 до 5 минут. Время этой инкубации не является критичным и может варьировать в зависимости от конкретной используемой ДНКазы и концентрации, и других компонентов реакционной смеси. Температура может представлять собой любую температуру, при которой фермент является активным, т.е. ниже температуры инактивации (например, 37°С), для удобства температура является комнатной.
Такая реакционная смесь может, как упомянуто выше, содержать все необходимые реагенты для реакции амплификации помимо матрицы, т.е. амплифицируемой нуклеиновой кислоты-мишени.
Типичная смесь для реакции амплификации путем ПЦР может, например, включать:
В вышеприведенном типичном примере может использоваться любая комбинация объемов стерильной дистиллированной воды и экспериментальной матрицы, а также общий объем реакционной смеси (включающей буфер, dNTP, праймеры, ферменты и растворы MgCl2), равный 25-50 мкл. Тем не менее, альтернативные конечные объемы можно использовать в зависимости от выбора для достижения, например, близких или других желаемых конечных концентраций реагентов. Можно использовать любой удобный или имеющийся в продаже буфер для ПЦР.
После удаления загрязнения ДНКазу инактивируют путем нагревания реакционной смеси. Для удобства это может быть осуществлено путем нагревания в первом цикле ПЦР.
Оптимальное осуществление способа ПЦР зависит от выбора температуры, времени при данной температуре и длительности времени между температурами для каждой стадии цикла. Типичный профиль цитирования для использования ДНКазы для деградации загрязняющей дцДНК перед амплификацией путем ПЦР новой добавленной нуклеиновой кислоты-мишени является следующим: (а) от 0 до 10 минут инкубации с ДНКазой при комнатной температуре; (б) 2 минуты инактивации ДНКазы при 94°С; (в) добавление матрицы; 1 минута плавления ДНК при 94°С; (г) 15 секунд отжига праймера при 50-65°С; (д) 30 секунд удлинения праймера при 72°С; (е) 10 секунд плавления ДНК при 94°С; и стадии (г)-(е) повторяют столько раз, сколько необходимо для достижения желаемого уровня амплификации.
Как упомянуто ранее, ДНКаза по изобретению особенно подходит для одностадийной амплификации реакции с обратной транскрипцией, например ПЦР с обратной транскрипцией. Такие протоколы хорошо известны в области техники, но для полноты типичная смесь для ПЦР с обратной транскрипцией может например включать:
Здесь применимы вышеприведенное обсуждение относительно объемов и буферов для реакций обратной транскрипции и ПЦР. После удаления загрязнения реакцию обратной транскрипции осуществляют при температуре, при которой ДНКаза инактивируется (например, 50°С в течение одного часа). В течение этой стадии ДНКаза инактивируется. Последнее означает, что продукт кДНК не будет деградировать по мере его получения, и когда начнется реакция ПЦР, также не будет происходить деградация этого продукта. После реакции обратной транскрипции осуществляют реакцию ПЦР без дополнительного добавления в реакционную смесь путем воздействия на реакционные сосуды профилем цитирования, таким как описанный выше профиль.
Краткое описание графических материалов
Изобретение далее будет описано при помощи не ограничивающих его объем примеров со ссылкой на следующие графические материалы, на которых:
На Фиг.1 продемонстрированы фотографии множества агарозных гелей, отражающих активность ДНКазы в соответствии с SEQ ID NO: 7 и ДНКазы Pandalus borealis дикого типа (SEQ ID NO: 6), которые инактивировали в присутствие или отсуствие DTT при 50, 55, 65 или 94°С в течение 15, 30 или 60 минут, в отношении плазмидной ДНК при 37°С в течение 3 часов.
На Фиг.2 продемонстрирована динамика по времени инактивации ДНКазы в соответствии с SEQ ID NO: 7 при 55°С в присутствии DTT.
На Фиг.3 продемонстрирована динамика по времени инактивации ДНКазы в соответствии с SEQ ID NO: 7 при 50°С в присутствии DTT.
На Фиг.4 продемонстрирована последовательность аминокислот зрелой формы Р214А мутантной ДНКазы Pandalus borealis по изобретению (SEQ ID NO: 7).
На Фиг.5 продемонстрирована кодирующая нуклеотидная последовательность мутанта Р214А зрелой формы ДНКазы Pandalus borealis (SEQ ID NO: 8).
На Фиг.6 продемонстрирована нуклеотидная последовательность и аминокислотная последовательность Р237А мутантной ДНКазы Pandalus borealis по изобретению (SEQ ID NO: 3 и 4). Эта аминокислотная последовательность включает сигнальный пептид MIGRTTFIALFVKVLTIWSFTKG (SEQ ID NO: 9).
На Фиг.7 продемонстрирована аминокислотная последовательность зрелой формы ДНКазы Pandalus borealis (SEQ ID NO: 5).
На Фиг.8 продемонстрирована кодирующая нуклеотидная последовательность зрелой формы ДНКазы Pandalus borealis (SEQ ID NO: 6).
На Фиг.9 продемонстрирована нуклеотидная последовательность кДНК и транслированная аминокислотная последовательность ДНКазы Pandalus borealis (SEQ ID NO: 1 и 2). Эта аминокислотная последовательность включает сигнальный пептид MIGRTTFIALFVKVLTIWSFTKG (SEQ ID NO: 9).
На Фиг.10 продемонстрировано действие ДНКазы Pandalus borealis дикого типа и мутанта Р214А на эффективность одностадийной ОТ-ПЦР.
На Фиг.11 продемонстрировано совмещение аминокислотной последовательности ДНКазы королевского краба (Paralithodes camtschaticus) (SEQ ID NO: 15) и ДНКазы Pandalus borealis (SEQ ID NO: 5). 65,7% идентичность с совпадением по 379 остаткам; Оценка: 1384,0; Частота пропусков: 0,0%.
На Фиг.12 продемонстрировано действие увеличивающихся концентраций мутанта Р214А на протокол количественной ПЦР.
На Фиг.13 продемонстрировано сравнение термолабильности ДНКазы I и мутанта Р214А путем измерения ингибирующих действий обработанных температурой ферментов на протокол количественной ПЦР.
На Фиг.14 продемонстрирована степень удаления внесенной ДНК из реакционной смеси для количественной ПЦР при увеличении количества мутанта Р214А.
На Фиг.15 продемонстрировано действие увеличивающейся концентрации мутанта Р214А на одностадийную ОТ-ПЦР.
На Фиг.16 продемонстрировано действие мутанта Р214А на нематричные контроли для кПЦР;
и на которых
SEQ ID NO: 1 представляет собой аминокислотную последовательность транслированного фрагмента нуклеотидной последовательности кДНК ДНКазы Panda/us borealis.
SEQ ID NO: 2 представляет собой нуклеотидную последовательность кДНК ДНКазы Pandalus borealis.
SEQ ID NO: 3 представляет собой аминокислотную последовательность мутанта Р237А ДНКазы Pandalus borealis.
SEQ ID NO: 4 представляет собой кодирующую нуклеотидную последовательность мутанта Р237А ДНКазы Pandalus borealis.
SEQ ID NO: 5 представляет собой аминокислотную последовательность зрелой формы ДНКазы Pandalus borealis.
SEQ ID NO: 6 представляет собой кодирующую нуклеотидную последовательность зрелой формы ДНКазы Pandalus borealis.
SEQ ID NO: 7 представляет собой аминокислотную последовательность мутанта Р214А зрелой формы ДНКазы Pandalus borealis.
SEQ ID NO: 8 представляет собой кодирующую нуклеотидную последовательность мутанта Р214А зрелой формы ДНКазы Pandalus borealis.
SEQ ID NO: 9 представляет собой аминокислотную последовательность сигнального пептида ДНКазы Pandalus borealis.
SEQ ID NO: 10 представляет собой меченный 5' FAM и 3' TAMRA олигонуклеотид для измерения ДНКазной активности.
SEQ ID NO: 11 представляет собой комплементарную последовательность SEQ ID NO: 10.
SEQ ID NO: 12 представляет собой прямой праймер для амплификации фрагмента гена Е.coli 23SrРНК.
SEQ ID NO: 13 представляет собой обратный праймер для амплификации фрагмента гена Е.coli 23SrРНК.
SEQ ID NO: 14 представляет собой 5' FAM и 3' BHQ меченный олигонуклеотидный праймер, комплементарный фрагменту гена Е.coli 23SrРНК между областями, комплементарными SEQ ID NO: 13 и SEQ ID NO: 14.
SEQ ID NO: 15 представляет собой аминокислотную последовательность ДНКазы королевского краба (Paralithodes camtschaticus).
Свободный текст перечня последовательностей
SEQ ID NO: 10
<223> 5' FAM и 3' TAMRA меченный олигонуклеотидный праймер для измерения ДНКазной активности.
SEQ ID NO: 11
<223> комплементарная последовательность SEQ ID NO: 10.
SEQ ID NO: 12
<223> прямой праймер для амплификации фрагмента гена Е.coli 23SrРНК.
SEQ ID NO: 13
<223> обратный праймер для амплификации фрагмента гена Е.соli 23SrРНК.
SEQ ID NO: 14
<223> 5' FAM и 3' BHQ меченный олигонуклеотидный праймер, комплементарный к фрагменту гена Е.соli 23SrРНК между областями, комплементарными SEQ ID NO: 13 и SEQ ID NO: 14.
ПРИМЕР 1 - Измерение активности ДНКазы
Анализ Кунитца
Активность ДНКазы может быть исследована в соответствии со способом Кунитца (Kunitz, М., 1950, Crystalline Deoxyribonuclease, II, Digestion of Thymus Nucleic Acid. The Kinetics of Reaction. J. Gen. Physiol., 33, 363-377). Десять мкл препарата фермента добавляют к 50 мкг ДНК тимуса ягнят в 100 мМ ацетате натрия, рН 5,0,5 мМ MgCl2, в конечном объеме 1 мл. Смесь инкубируют при 25°С и увеличение поглощения измеряют при 260 нм. 1 Ед. = 0,001 OD260 увеличение × мин-1.
Модифицированный Ямамото анализ Кунитца
Модифицированный анализ Кунитца, представляющий собой анализ по конечной точке, описанный Yamamoto (Yamamoto, M. 1971. Purification and some properties of an acid deoxyribonuclease from testes of Chinook salmon Oncorhynchus tshawytscha. Biochim Biophys Acta, 228, 95-104) представляет собой более чувствительный вариант анализа Кунитца и рассматривается как более подходящий для измерения остаточной активности ДНКазы после инактивации. Десять мкл фермента добавляют к 200 мкгДНКтимуса ягнят в 20 мМ Tris/HCl, pH 8,0, 5 мМ MgCl2, в конечном объеме 1 мл. Смесь инкубируют при 37°С в течение 20 минут. Затем добавляют 0,5 мл охлажденной на льду 12% HClO4, тщательно смешивают и оставляют на льду в течение 20 минут. Пробирки центрифугируют на полной скорости в центрифуге Eppendorf в течение 10 минут. Определяют поглощение при 260 нм, из которого рассчитывают Единицы. 1 Ед. = 0,001 OD260 увеличение × мин-1.
ПРИМЕР 2 - Мутация ДНКазы Panda/us borealis
Осуществляли мутацию ДНКазы Pandalus borealis (SEQ ID NO: 5) по остатку 214 (соответствующему остатку 237 в SEQ ID NO: 1) с использованием набора для мутагенеза Quick-change™ от Invitrogen и указаний производителя. Пролин представляет собой остаток дикого типа, и аланин представлял собой замещающий остаток. На Фиг.4-9 продемонстрированы аминокислотные и нуклеотидные последовательности варианта дикого типа и мутантного варианта ДНКазы Pandalus borealis. Мутантов секвенировали и выяснили, что они являются правильными, и трансформировали в Pichia paston's. Получали трансформант, который демонстрировал хорошую экспрессию, близкую к дикому типу. Тесты исходной инактивации мутанта Р214А демонстрировали его более легкую инактивацию при 55°С по сравнению с ДНКазой дикого типа.
Рекомбинантный клон Pichia paston's, содержащий экспрессирующуюся кассету мутанта Р214А ДНКазы, затем экспрессировали в ферментере объемом один литр. Ферментацию осуществляли, как описано в Pichia fermentation process guidelines, Invitrogen. Ферментат (приблизительно 1 л) центрифугировали при 4500 g в течение 15 минут для удаления клеток и супернатант выливали в новую колбу. pH затем доводили до 8 путем добавления 0,5 M NaOH и затем центрифугировали при 4500 g в течение 15 минут для удаления осаждаемых солей. Новый супернатант наконец фильтровали через фильтр Whatman GF/F.
Белок Р214А ДНКазу исходно очищали с использованием анионообменной хроматографии. рН доводили и отфильтрованный супернатант (1150 мл) наносили на колонку Q-Sepharose FF (2,6/10), уравновешенную 25 мМ Tris/HCl pH 8,5 мМ MgCl2, 0,25 М NaCl (Буфер А). Колонку затем промывали 19 объемами колонки буера А и затем белок Р214А элюировали буфером 25 мМ Tris/HCl рН 8,5 мМ MgCl2, 0,5 М NaCl. Собирали фракции по 10 мл. Используемая скорость потока составила 10 мл/мин. Фракции, содержащие белок Р214А, отбирали путем измерения активности в соответствии со способом Кунитца, описанным в примере 1.
Отобранные фракции объединяли, подвергали диализу в 10 мМ Tris/HCl, рН 7,5,5 мМ MgCl2 (Буфер Б) при 4°С. Объем диализированного образца доводили до 200 мл с использованием того же самого буфера и затем наносили на колонку Blue Sepharose FF (5,0/10), уравновешенную буфером Б. Колонку промывали двумя объемами колонки буфера Б и белок Р214А ДНКазу элюировали с использованием буфер В + 0,25 М NaCl и фракции по 10 мл собирали. Используемая скорость потока составила 10 мл/мин. Наконец, фракции, содержащие Р214А, отбирали путем измерения активности, как описано выше, объединяли и концентрировали.
ПРИМЕР 3 - Определение остаточной активности после инактивации при различных температурах
Для определения того, полностью ли инактивируется мутант Р214А путем температурного воздействия, оценивали целостность ПЦР-продукта в присутствии инактивированного мутанта Р214А или фермента дикого типа.
Фермент (0,8 Е Р214А, или 1,5 U wt (дикого типа)) добавляли в пробирки для ПЦР, содержащие общий объем 20 мкл в 25 мМ Tris/HCl, рН 8,5 мМ MgCl2, ±1 мМ буфер DTT. Ферменты инактивировали теплом в течение 15, 30 и 60 минут при различных температурах и пробирки затем помещали в лед. Добавляли 0,5 мкг очищенного приблизительно 500 п.о. продукта ПЦР и реакционные смеси инкубировали в течение 3 часов при 37°С. Наконец реакции анализировали с использованием электрофореза в агарозном геле. Отрицательный контроль (без фермента) и положительный контроль (100 × разведенного фермента, добавляемого после стадии тепловой инактивации) обрабатывали тем же самым путем, как вышеприведенные реакционные смеси.
На Фиг.1 обобщены эксперименты по тепловой инактивации мутанта Р214А по сравнению с ферментом дикого типа при 50°С, 55°С, 65°С и 94°С. Контроль без фермента (-) демонстрирует интактный продукт ПЦР, тогда как положительный контроль (+), в 100 раз разведенный фермент, не инактивированный теплом, иллюстрирует действие 1% остаточной активности.
В результате экспериментов с контролем отсутствие видимой деградации продукта ПЦР указывает на меньше чем 0,01% остаточную активность (результаты не представлены), которая представляет собой предел обнаружения при использовании приблизительно 1 Ед. фермента в этом анализе. При 50°С и 55°С лишь мутант Р214А полностью инактивируется теплом, демонстрируя действие замены Р214А. Добавление DTT (1 мМ в этом эксперименте) необходимо для полной инактивации обоих ферментов. Лишь при инкубации в течение 60 мин при 94°С полная инактивация теплом обнаруживается в отсутствие DTT.
ПРИМЕР 4 - Динамика по времени инактивации Р214А при 50°С и 55°С
Если ДНКазу используют для удаления загрязнения полной реакционной смеси для обратной транскриптазы, то важно, чтобы она инактивировалась ранее на стадии обратной транскриптазы. Если нуклеаза не инактивирована немедленно, то она может начать расщеплять кДНК и оказывать негативное воздействие на продукты обратной транскрипции. Особенно важно, если обратная транскрипция представляет собой часть количественного анализа для измерения количества РНК в образце. Поэтому анализировали инактивацию мутанта Р214А ДНКазы при 50°С и 55°С в течение более коротких периодов времени.
Р214А, 12,5-125 Е, разбавляли в типичном буфере для RT (обратной транскрипции) (50 мМ Tris/HCl, рН 8,3, 50 мМ KCl, 5 мМ MgCl2, 5 мМ DTT) в общем объеме 25 мкл. Образцы инкубировали при 50 и 55°С в аппарате для ПЦР в течение 0-5 минут. Затем остаточную активность измеряли с использованием модифицированного анализа Кунитца, как описано в примере 1.
Как можно видеть на Фиг.2 и 3, мутант Р214А может быть полностью инактивирован в течение 1 минуты при 55°С и почти полностью инактивирован в течение одной минуты при 50°С.
ПРИМЕР 5 - Влияние ДНКазы дикого типа и ДНКазы Panda/us borealis и мутанта Р214А на эффективность одностадийной ОТ-ПЦР
Одностадийные реакции амплификации qOT-ПЦР осуществляли с использованием Brilliant QOT-PCR Master Mix Kit 1-Step (Stratagene) и термоциклирования и обнаружения в Smart Cycler II (Cepheid).
Использовали реакционную смесь (25 мкл), содержащую 12,5 мкл мастер-смесь 2Х QOT-PCR, 1,25 мкл 20х смеси праймер/зонд (GAPDH HS99999905_m1, Applied Biosystems), 0,1 мкл обратной транскриптазы Stratascript, 1 мкл фермента ДНКазы. В качестве матрицы использовали 1 мкл (1 нг/мкл) Stratagene QPCR Human Reference Total RNA (Stratagene). Каждую реакционную смесь предварительно инкубировали при 30°С в течение 15 минут. Затем осуществляли одностадийную ПЦР с обратной транскрипцией при 50°С в течение 30 мин, 95°С в течение 10 мин, а затем 45 циклов при 94°С в течение 15 с, 60°С в течение 1 мин.
Как представлено на фиг.10, небольшое действие или отсутствие действия на эффективность ОТ-ПЦР обнаружено в образцах, содержащих мутант Р214А. С другой стороны, нуклеаза дикого типа серьезно влияет на эффективность ОТ-ПЦР.
ПРИМЕР 6 -Анализ дц/оцДНК специфичностей в отношении Р214А:
Специфичность двух- и одноцепочечной ДНК в отношении Р214А тестировали путем измерения флуоресценции из олигонуклеотидов, меченных флуорофором FAM (флуоресцеин) по 5' концу и TAMRA по 3' концу. Расщепление олигонуклеотида нуклеазой может привести в результате к увеличению флуоресценции FAM, которое измеряют во флуориметре с длиной волны возбуждения 485 нм и длиной волны эмиссии 520 нм. Субстрат-двухцепочечную ДНК готовили путем смешивания меченого олигонуклеотида со вторым олигонуклеотидом, который комплементарен меченому олигонуклеотиду.
37 Единиц мутанта Р214А добавляли в реакционную смесь, содержащую 50 мМ Tris/HCl, pH 8,0, 5 мМ MgCl2 и 0,2 мкМ меченого олигонуклеотида (DNAsub) (общий объем 100 мкл). Смесь инкубировали при 25°С в микропланшете с белыми лунками для флуороскопии.
Аналогично, 0,2 мкМ комплементарного олигонуклеотида, compDNAsub, добавляли в реакционную смесь, как указано выше, для образования субстрата-двухцепочечной ДНК. 0,01 Е мутанта Р214А затем добавляли к реакционной смеси.
Флуоресценцию со временем измеряли в приборе Victor3 и активность расчитывали как исходное увеличение флуоресценции в минуту, скорректированное на увеличение флуоресценции без нуклеазы (реакция с холостой пробой), и выражали в виде (единиц флуоресценции/минуту)/Единиц Кунитца.
В отношении субстрата двухцепочечной ДНК результат составил 211922 (единиц флуоресценции/мин)/Единиц Кунитца и в отношении субстрата одноцепочечной ДНК результат составил 10,4 (единиц флуоресценции/мин)/Единиц Кунитца. Соответственно, субстрат двухцепочечная ДНК деградирует со скоростью, в 20366 быстрее одноцепочечной ДНК.
Олигонуклеотиды:
DNAsub: 5'-FAM-CGCCATCGGAGGTTC-TAMRA-3' [SEQ ID NO: 10]
compDNAsub: 5'-GAACCTCCGATGGCG-3' [SEQ ID NO: 11]
ПРИМЕР 7 - удаление загрязнения из дПЦР с использованием мутанта Р214А
Материалы и методы
Геномную ДНК Escherichia coli ТОР10 выделяли с использованием набора DNeasy Blood and Tissue Kit (Qiagen) и концентрацию ДНК измеряли с использованием набора Quant-iT dsDNA BR assay kit и флуориметра Qubit (Life Technologies). Для детекции и количественной оценки геномной ДНК Е.coli с использованием количественной ПЦР (кПЦР) небольшую область высококонсервативного гена 23S rРНК использовали для конструирования праймера/зонда, как описано в (Smith GJ III et al.; 1999; Biomethods 26(3): 518-22, 524, 526). Этот ген представлен в семи копиях в геноме Е.coli.
В общем, кПЦР осуществляли в Smart Cycler II (Cepheid) в 25 мкл реакционной смеси, содержащей 12,5 мкл 2х мастер-смеси Brilliant qПЦР (Stratagene) или мастер-смесь TaqMan Gene Expression (Applied Biosystems), 3 мкМ каждого праймера и 1 мкМ зонда, 1 мМ DTT, 1 или 10 пг геномной ДНК Е.coli, и различные количества мутантного фермента Р214А или ДНКазы I (Sigma). Реакцию кПЦР осуществляли следующим образом: 95°С в течение 10 минут, затем 40 циклов при 95°С в течение 15 секунд и 60°С в течение 1 минуты.
Праймеры/зонд:
Ecoli_23S_fwd: 5'-GAAAGGCGCGCGATACAG-3' [SEQ ID NO: 12]
Ecoli_23S_rev: 5'-GTCCCGCCCTACTCATCG A-3' [SEQ ID NO: 13]
Ecoli_23S_probe: 5'-FAM-CCCCGTACACAAAAATGCACATGCTG-BHQ-3' [SEQ ID NO: 14]
Действие мутанта Р214А на целостность праймера/зонда (оцДНК)
В этом эксперименте тестировали, оказывает ли мутант Р214А ингибирующее действие на протокол кПЦР путем деградации праймеров/зонда в смеси кПЦР (т.е. деградации одноцепочечной ДНК).
Описанные выше реакционные смеси (мастер-смесь Brilliant qPCR) готовили без матрицы (Е.соli. геномная) ДНК и инкубировали при 37°С в течение 10 минут. Стадию инактивации при 95°С в течение 10 минут осуществляли перед добавлением 10 пг геномной ДНК Е.coli в качестве матрицы. Амплификацию путем кПЦР затем осуществляли, как описано выше.
Стадия инкубации при 37°С давала возможность мутанту Р214А катализировать эту деградацию ДНК. Инкубация при 95°С в течение 10 минут полностью инактивирует мутант перед добавлением матрицы ДНК. Таким образом, результат любого ингибирования кПЦР может быть лишь следствием нуклеазной активности против оцДНК (праймеры/зонд деградировали). Как представлено на фиг.12, на результат кПЦР не влияло добавление до 1 Ед. мутанта Р214А, свидетельствуя о том, что отсутствовала измеряемая активность против праймеров/зонда в реакционной смеси для кПЦР.
Сравнение термолабильности ДНКазы I и мутанта Р214А в протоколе для кПЦР
Описанные реакционные смеси готовили без матрицы ДНК в присутствии или отсутствии ДНКазы I (1 Ед.) или Р214А (1 Ед.). За стадией инкубации при 37°С в течение 10 минут затем следовала стадия инактивации при 50°С или 55°С в течение 15 минут. 1 пг геномной ДНК E.соli затем добавляли в смеси и кПЦР осуществляли, как описано выше.
Для оценки различных реакционных условий-смесей в экспериментах кПЦР реакционные смеси инкубировали при комнатной температуре в течение 15 мин перед амплификацией. Какая-либо остаточная активность ДНКазы в реакционной смеси деградирует матрицу ДНК и ингибирует результаты кПЦР.
Как проиллюстрировано на фиг.13, мутант Р214А не ингибирует кПЦР по сравнению с контрольной реакцией (Контроль (-Enz); фермент не добавлен) и, таким образом, может рассматриваться как полностью инактивированный на стадии инкубации в течение 15 мин при 50°С в этом эксперименте. Фермент ДНКаза I не инактивирована, и Ct смещается более чем на 8, указывая на то, что высокая остаточная активность после стадии инактивации или/и активность против праймеров/зонда в реакционной смеси.
Удаление загрязняющей ДНК из реакционной смеси для кПЦР
Для тестирования способности мутанта Р214А удалять "загрязняющую" ДНК, в различные количества мутанта Р214А в описанных выше реакционных смесях для кПЦР (мастер-смесь TaqMan Gene Expression) добавляли по 1 пг геномной ДНК E.coli. Реакционные смеси затем инкубировали в течение 10 минут при 37°С и затем инкубировали при 60°С в течение 15 минут. Результаты представлены на фиг.14. Как можно видеть, 0,25 Ед. или больше мутанта Р214А на 25 мкл реакционной смеси приводили к увеличению Ct более чем на 8. Последнее свидетельствует о более чем 250-кратном уменьшении концентрации загрязняющей ДНК.
Дополнительно к обсуждавшимся выше индивидуальным результатам следует отметить, что никакие из матричных контролей (NТС) не приводили к положительным результатам для мастер-смеси Brilliant qPCR (Stratagene), и мастер-смеси TaqMan Gene Expression (Applied Biosystems). Последнее иллюстрирует проблему загрязняющей ДНК в смесях кПЦР при использовании универсальных праймеров, нацеленных на бактериальную ДНК или ДНК E.соli для обнаружения или диагностики бактерий.
ПРИМЕР 8 - Действие мутанта Р214А на эффективность одностадийной ОТ-ПЦР.
Эксперимент, описанный в Примере 5, повторяли с несколькими концентрациями мутанта Р214А для исследования того, как увеличенные количества фермента влияли на чувствительность реакции ОТ-ПЦР. Тестировали пять различных концентраций в диапазоне от 0 до 1 Ед. ДНКазы, и результаты отображены на фиг.15. Использование 0,1-0,5 Ед. ДНКазы не влияло на чувствительность ОТ-ПЦР. Использование 1 Ед. фермента уменьшало чувствительность с Ct 1,5.
ПРИМЕР 9 - Удаление загрязняющей бактериальной ДНК из коммерческих продуктов ПЦР
Показано (Пример 7), что следы бактериальной ДНК часто присутствуют в коммерческих реакционных смесях для амплификации нуклеиновой кислоты (так называемые "мастер-смеси"). В экспериментах кПЦР по обнаружению патогенов это часто представляет собой проблему, поскольку амплификация этих примесей ведет к ложноположительным результатам, включая контроли без матрицы (NTC). В этом примере 1 Ед. ДНКазы мутанта Р214А добавляли к мастер-смесям для кПЦР различных производителей и предварительно инкубировали в течение 10 минут при 37°С. После этого мастер-смеси инкубировали при 60°С в течение минут и сравнивали с необработанными мастер-смесями в реакции кПЦР, как описано в примере 7. Матрицу не добавляли к реакционным смесям. Результаты представлены на фиг.16, и можно видеть, что лишь мастер-смеси, проинкубированные с ферментом, приводят к отрицательным NTC.















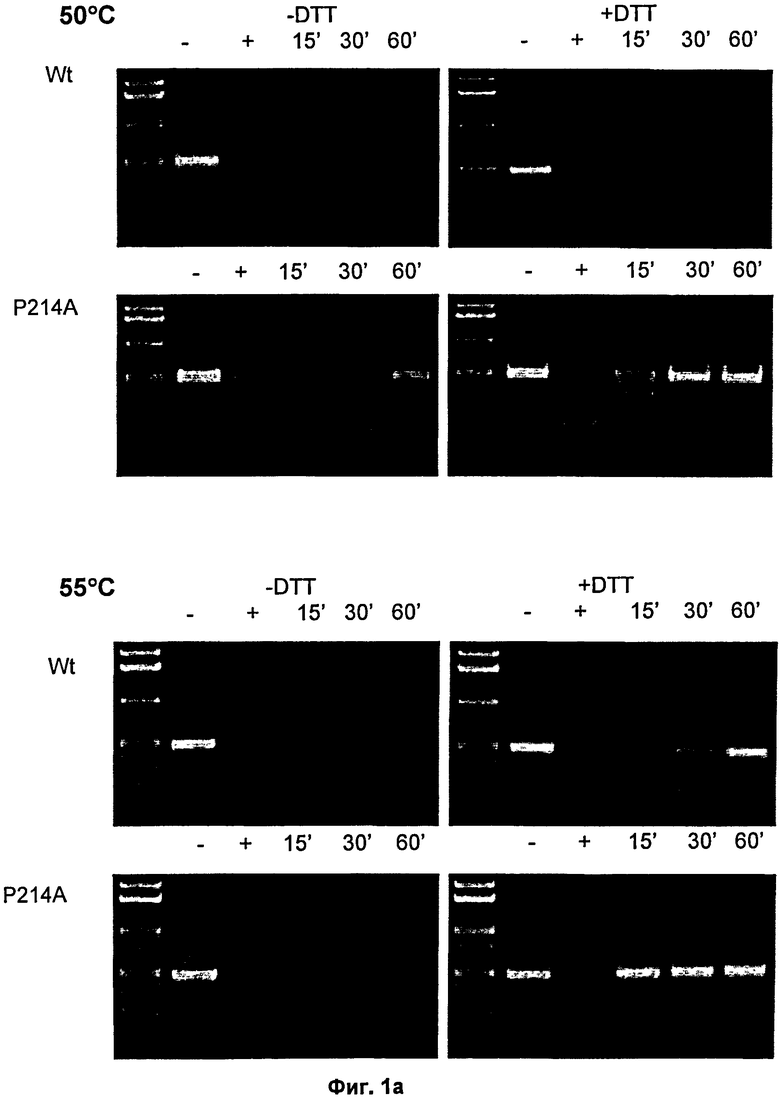
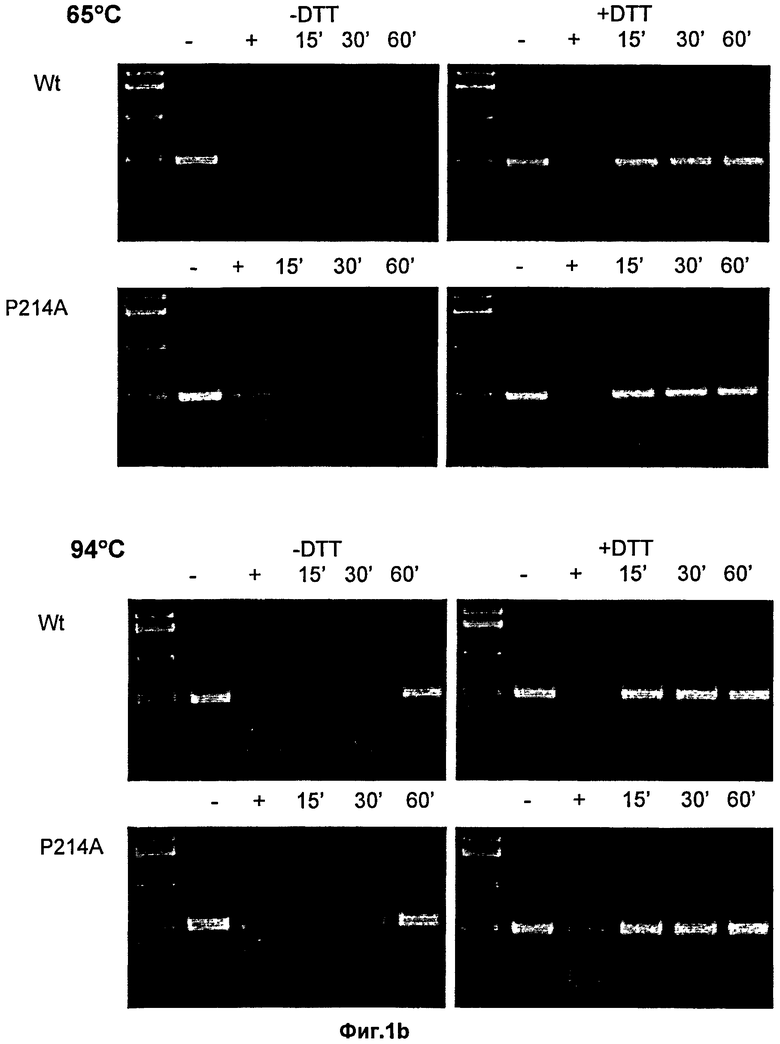
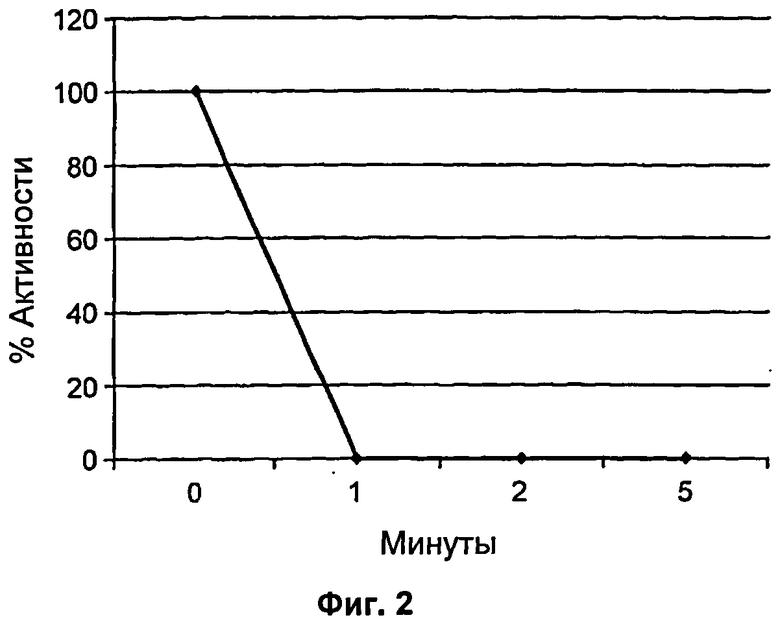
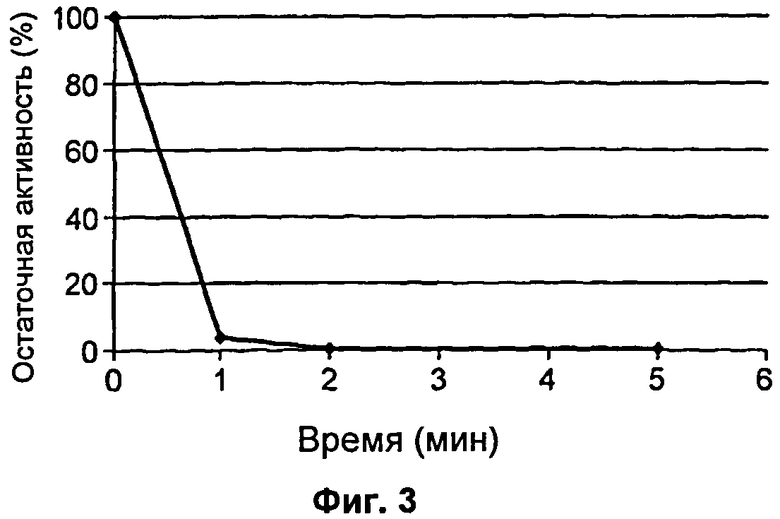



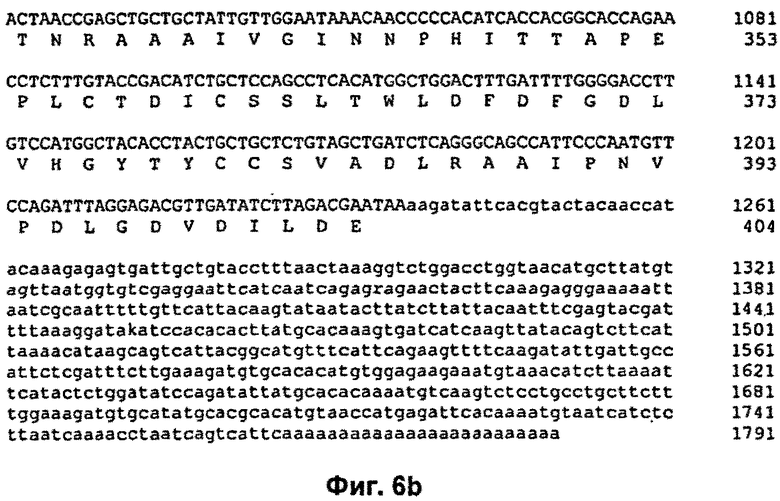



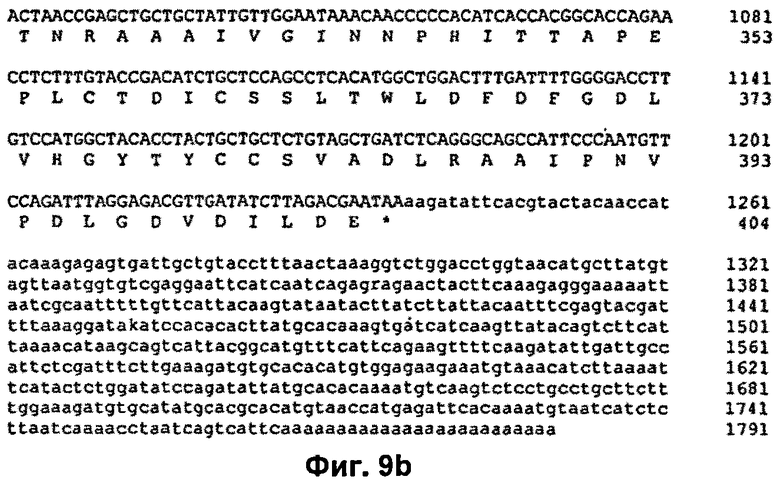
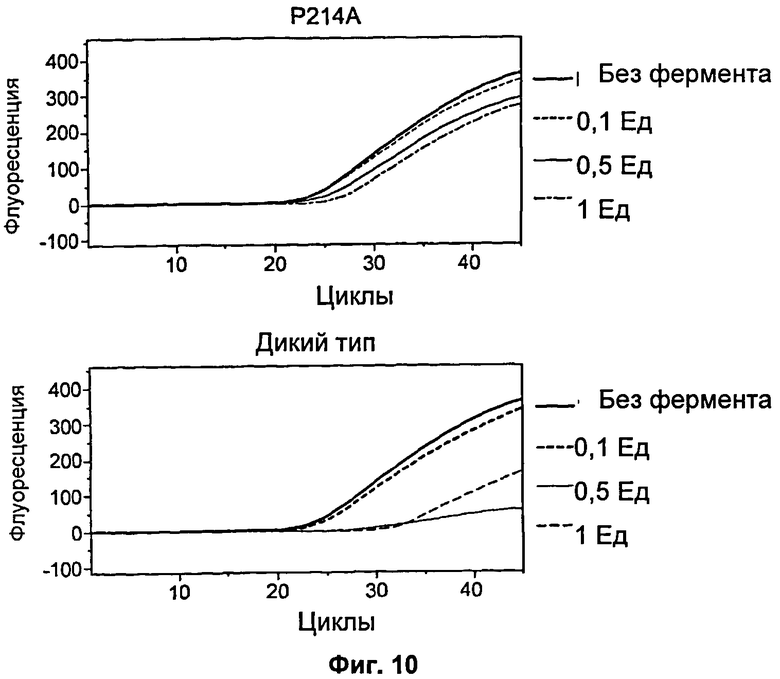

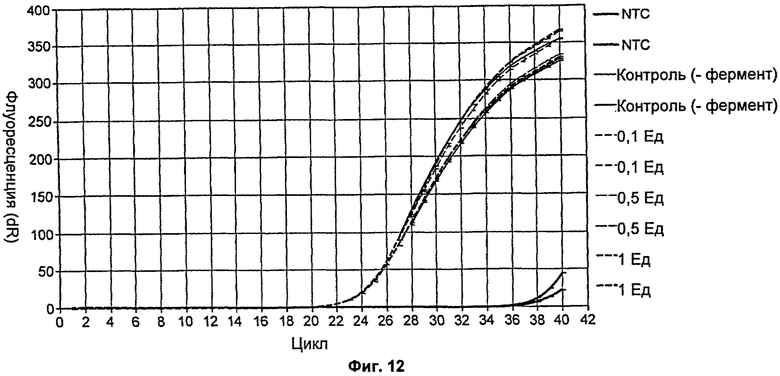
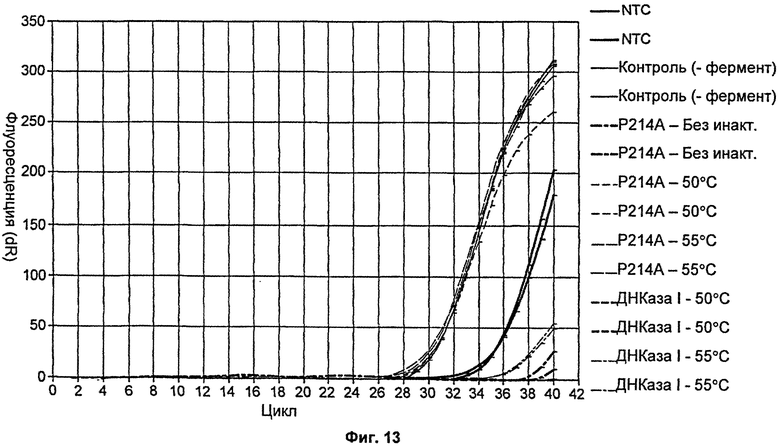
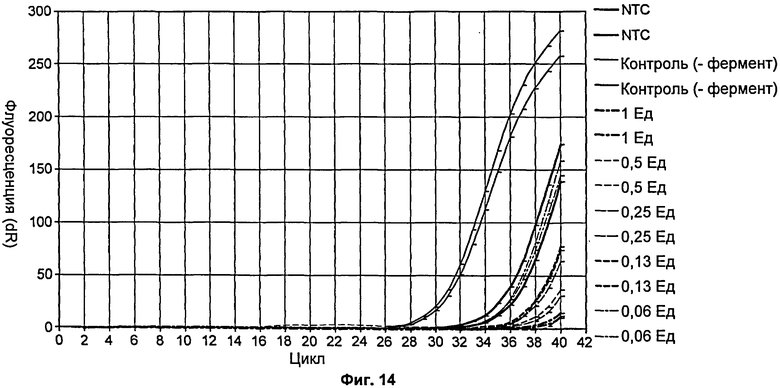
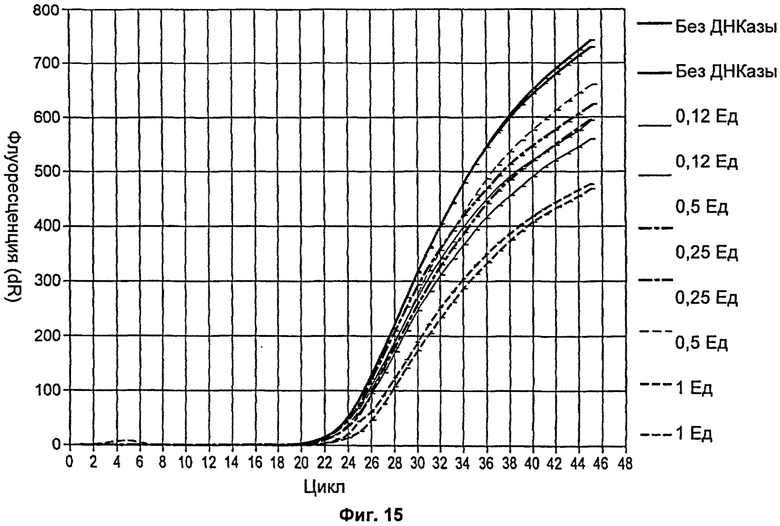
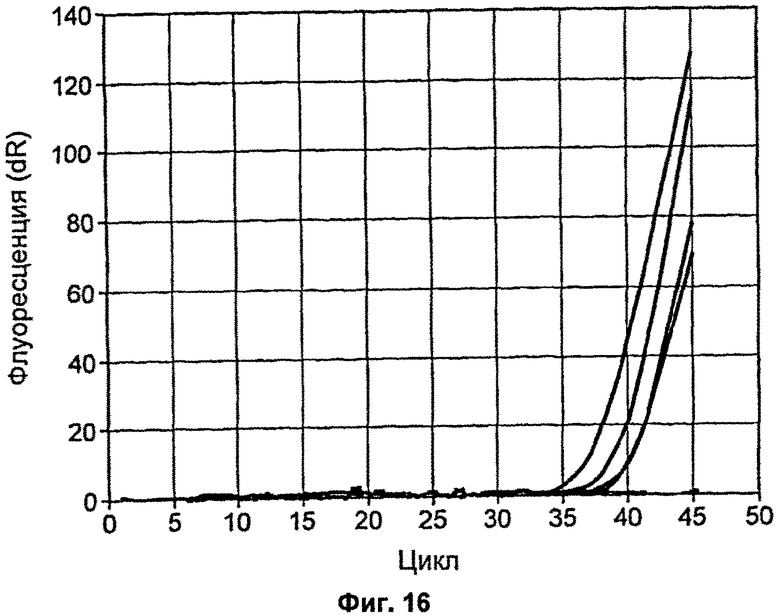
Изобретение относится к биотехнологии и представляет собой ДНКазу или ее ферментативно активный фрагмент, способы удаления загрязняющей нуклеиновой кислоты из реакции обратной транскрипции, в которых используется данная ДНКаза, нуклеиновую кислоту, кодирующую указанную ДНКазу, и наборы или композиции, содержащие указанную ДНКазу. Указанная ДНКаза представляет собой ДНКазу, которая имеет последовательность SEQ ID NO: 1, или ферментативно активный вариант SEQ ID NO: 1, имеющий последовательность, которая по меньшей мере на 95% идентична ей, но где пролиновый остаток в положении 237 в SEQ ID NO: 1 или эквивалентный пролин в других последовательностях замещен, и где указанная ДНКаза или ее ферментативно активный фрагмент по существу необратимо инактивируются путем нагревания при температуре примерно 50°С в течение 5 мин в буфере, состоящем из 25 мМ Tris/HCI, pH 8,5, 5 мМ MgCl2 и 1 мМ DTT, и которые по существу специфичны в отношении двухцепочечной ДНК. Предложенное изобретение позволяет с высокой эффективностью удалять загрязняющую ДНК из реакционных смесей для обратной транскрипции, препаратов ДНК-полимеразы с горячим стартом и реакционных смесей для ПЦР с горячим стартом. 13 н. и 22 з.п. ф-лы, 19 ил., 9 пр.
1. ДНКаза или ее ферментативно активный фрагмент, где указанная ДНКаза представляет собой:
1) ДНКазу, которая имеет последовательность в соответствии с SEQ ID NO: 1, или
2) ферментативно активный вариант SEQ ID NO: 1, имеющий последовательность, которая по меньшей мере на 95% идентична ей, но где пролиновый остаток в положении 237 в SEQ ID NO: 1 или эквивалентный пролин в других последовательностях замещен, и где указанная ДНКаза или ее ферментативно активный фрагмент по существу необратимо инактивируются путем нагревания при температуре примерно 50°C в течение 5 мин в буфере, состоящем из 25 мМ Tris/HCl, pH 8,5, 5 мМ MgCl2 и 1 мМ DTT, и которые по существу специфичны в отношении двухцепочечной ДНК.
2. ДНКаза или ее ферментативно активный фрагмент по п. 1, где указанная ДНКаза представляет собой:
1) ДНКазу, которая имеет последовательность в соответствии с SEQ ID NO: 5, или
2) ферментативно активный вариант SEQ ID NO: 5, имеющий последовательность, которая по меньшей мере на 95% идентична ей, но где пролиновый остаток в положении 214 в SEQ ID NO: 5 или эквивалентный пролин в других последовательностях замещен, и где указанная ДНКаза или ее ферментативно активный фрагмент по существу необратимо инактивируются путем нагревания при температуре примерно 50°C в течение 5 мин в буфере, состоящем из 25 мМ Tris/HCl, pH 8,5, 5 мМ MgCl2 и 1 мМ DTT, и которые по существу специфичны в отношении двухцепочечной ДНК.
3. ДНКаза или ее фрагмент по п. 1, где указанная ДНКаза имеет последовательность, которая по меньшей мере на 98% идентична SEQ ID NO: 1, но где эквивалент пролинового остатка в положении 237 в SEQ ID NO: 1 замещен.
4. ДНКаза или ее фрагмент по п. 3, где указанная ДНКаза имеет последовательность SEQ ID NO: 1, но где пролиновый остаток в положении 237 замещен.
5. ДНКаза или ее фрагмент по п. 2, где указанная ДНКаза имеет последовательность, которая по меньшей мере на 98% идентична SEQ ID NO: 5, но где эквивалент пролинового остатка в положении 214 в SEQ ID NO: 5 замещен.
6. ДНКаза или ее фрагмент по п. 5, где указанная ДНКаза имеет последовательность SEQ ID NO: 5, но где пролиновый остаток в положении 214 замещен.
7. ДНКаза или ее фрагмент по любому из пп. 1-3 или 5, где указанная ДНКаза имеет последовательность ДНКазы, полученной из видов типа Arthropodoa, но где пролиновый остаток, эквивалентный пролину в положении 237 в SEQ ID NO: 1, замещен.
8. ДНКаза или ее фрагмент по п. 7, имеющие последовательность ДНКазы, полученной из видов подтипа, выбранного из Crustacea, Hexpoda, Chelicerata или Myriapoda, но где пролиновый остаток, эквивалентный пролину в положении 237 в SEQ ID NO: 1, замещен.
9. ДНКаза или ее фрагмент по п. 7, имеющие последовательность ДНКазы, полученной из видов, выбранных из Pandalus borealis, Paralithodes camtschaticus (королевский краб), Marspenus japonicus (креветка kuruma) или Penaeus japonicus, но где пролиновый остаток, эквивалентный пролину в положении 237 в SEQ ID NO: 1, замещен.
10. ДНКаза или ее фрагмент по п. 9, имеющие последовательность ДНКазы, полученной из Pandalus borealis, но где пролиновый остаток, эквивалентный пролину в положении 237 в SEQ ID NO: 1, замещен.
11. ДНКаза или ее фрагмент по любому из пп. 1-6, где указанный пролиновый остаток замещен аланином, глицином, серином или цистеином.
12. ДНКаза или ее фрагмент по п. 11, где указанный пролиновый остаток замещен аланином.
13. ДНКаза по п. 1 или 2, имеющая последовательность в соответствии с SEQ ID NO: 3, SEQ ID NO: 7, ферментативно активный фрагмент SEQ ID NO: 3 или ферментативно активный фрагмент SEQ ID NO: 7.
14. Молекула нуклеиновой кислоты, кодирующая ДНКазу или ее фрагмент по любому из пп. 1-13.
15. Молекула нуклеиновой кислоты по п. 14, содержащая нуклеотидную последовательность в соответствии с SEQ ID NO: 4 или SEQ ID NO: 8.
16. Способ удаления загрязняющей нуклеиновой кислоты из реакции обратной транскрипции, включающий приведение смеси для реакции обратной транскрипции или ее компонентов в контакт с ДНКазой, которая по существу специфична в отношении двухцепочечной ДНК, в условиях, которые дают возможность для расщепления любой загрязняющей двухцепочечной ДНК, отличающийся тем, что указанная ДНКаза представляет собой ДНКазу или ее фрагмент по любому из пп. 1-13.
17. Способ удаления загрязняющей нуклеиновой кислоты из полимеразной цепной реакции (ПЦР) с горячим стартом, где указанная реакция представляет собой барьерную комбинацию для ПЦР с горячим стартом и/или включает ДНК-полимеразу с горячим стартом, включающий приведение комбинации/смеси для ПЦР с горячим стартом или ее компонентов в контакт с ДНКазой, которая по существу специфична в отношении двухцепочечной ДНК, в условиях, которые дают возможность для расщепления любой загрязняющей двухцепочечной ДНК, отличающийся тем, что указанная ДНКаза представляет собой ДНКазу или ее фрагмент по любому из пп. 1-13.
18. Способ по п. 16, где после расщепления любой загрязняющей двухцепочечной ДНК смесь, содержащую ДНКазу или ее фрагмент, нагревают для инактивации ДНКазы или ее фрагмента.
19. Способ по п. 17, где после расщепления любой загрязняющей двухцепочечной ДНК смесь, содержащую ДНКазу или ее фрагмент, нагревают для инактивации ДНКазы или ее фрагмента.
20. Способ обратной транскрипции in vitro нуклеиновой кислоты-мишени, включающий стадию обработки реакционной смеси для обратной транскрипции ДНКазой, которая по существу специфична в отношении двухцепочечной ДНК, перед началом реакции обратной транскрипции, отличающийся тем, что указанная ДНКаза представляет собой ДНКазу или ее фрагмент по любому из пп. 1-13.
21. Способ по п. 18, где реакционная смесь для обратной транскрипции представляет собой полную смесь и ее нагревают при рабочей температуре фермента для обратной транскрипции для инактивации ДНКазы или ее фрагмента.
22. Способ по п. 20, где реакционная смесь для обратной транскрипции представляет собой полную смесь и ее нагревают при рабочей температуре фермента для обратной транскрипции для инактивации ДНКазы или ее фрагмента.
23. Способ по любому из пп. 20-22, при котором после стадии обработки осуществляют один или более чем один цикл, включающий одну или более чем одну из следующих стадий:
(1) отжиг праймера,
(2) удлинение цепи относительно полинуклеотида с праймером,
(3) плавление полинуклеотидного дуплекса.
24. Способ по любому из пп. 16 и 20-22, где за указанной реакцией обратной транскрипции следует реакция амплификации нуклеиновой кислоты.
25. Способ по п. 24, где указанная реакция амплификации нуклеиновой кислоты представляет собой ПЦР, лигазную цепную реакцию (ЛЦР), амплификацию с вытеснением цепи (SDA), самоподдерживающуюся репликацию последовательности (3SR) или изотермальную петлевую амплификацию (ИПА), предпочтительно ПЦР.
26. Способ по п. 24, где реакцию обратной транскрипции и реакцию амплификации осуществляют в одном реакционном сосуде.
27. Способ ПЦР с горячим стартом, где указанная реакция представляет собой барьерную комбинацию для ПЦР с горячим стартом и/или включает ДНК-полимеразу с горячим стартом, включающий стадию обработки комбинации/смеси для ПЦР с горячим стартом или ДНК-полимеразы с горячим стартом ДНКазой, которая по существу специфична в отношении двухцепочечной ДНК, перед началом ПЦР с горячим стартом, отличающийся тем, что указанная ДНКаза представляет собой ДНКазу или ее фрагмент по любому из пп. 1-13.
28. Способ по любому из пп. 25-27, при котором дополнительно осуществляют множество циклов, включающих одну или более чем одну из следующих стадий:
(1) плавление полинуклеотидного дуплекса,
(2) отжиг праймера,
(3) удлинение цепи относительно полинуклеотида с праймером.
29. Применение ДНКазы или ее ферментативно активного фрагмента по любому из пп. 1-13 в качестве агента для удаления загрязнения в способе амплификации нуклеиновой кислоты.
30. Применение ДНКазы или ее ферментативно активного фрагмента по любому из пп. 1-13 в способе по любому из пп. 16-28.
31. Способ выделения и очистки ДНКазы или ее ферментативно активного фрагмента по любому из пп. 1-13, включающий экспрессию указанной ДНКазы или ее фрагмента в подходящей клетке-хозяине и затем выделение ДНКазы или ее фрагмента из указанных клеток-хозяев и/или сред, в которых выращивали указанные клетки.
32. Набор для осуществления способа по любому из пп. 16-19 и 27, содержащий ДНКазу или ее фрагмент по любому из пп. 1-13; и возможно один или более чем один из следующих:
(1) нуклеотидтрифосфат;
(2) олигонуклеотидный праймер;
(3) фермент обратной транскрипции;
(4) ДНК-полимераза;
(5) ДНК-лигаза; и
(6) рестрикционный фермент.
33. Композиция для осуществления способа по любому из пп. 16-19 и 27, содержащая ДНКазу или ее фрагмент по любому из пп. 1-13; и возможно один или более чем один из следующих:
(1) нуклеотидтрифосфат;
(2) олигонуклеотидный праймер;
(3) фермент обратной транскрипции;
(4) ДНК-полимераза;
(5) ДНК-лигаза; и
(6) рестрикционный фермент.
34. Набор для осуществления способа по любому из пп. 20-23, содержащий ДНКазу или ее фрагмент по любому из пп. 1-13, фермент обратной транскрипции; и возможно один или более чем один из следующих:
(1) нуклеотидтрифосфат;
(2) олигонуклеотидный праймер;
(3) ДНК-полимераза;
(4) ДНК-лигаза; и
(5) рестрикционный фермент.
35. Композиция для осуществления способа по любому из пп. 20-23, содержащая ДНКазу или ее фрагмент по любому из пп. 1-13, фермент обратной транскрипции; и возможно один или более чем один из следующих:
(1) нуклеотидтрифосфат;
(2) олигонуклеотидный праймер;
(3) ДНК-полимераза;
(4) ДНК-лигаза; и
(5) рестрикционный фермент.
| ANISIMOVA V.E | |||
| ET AL., Thermolabile duplex-specific nuclease, BIOTECHNOLOGY LETTERS, 2009, v | |||
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Способ модулирования для радиотелефона | 1921 |
|
SU251A1 |
| Двусторонний центробежный распылитель жидкости | 1981 |
|
SU1003913A1 |
| US 20020042052 A1, 11.04.2002 | |||
| WO 1999007887 A3, 18.02.1999 | |||
| RU 94046424 A1, 10.03.1997 | |||

