Настоящее изобретение относится к области молекулярной биологии, в частности к синтезу полинуклеотидов. Настоящее изобретение относится также фактически к чистой термостабильной экзонуклеазе, клонированию и экспрессии термостабильной экзонуклеазы III в Е.coli и ее применению в реакциях амплификации. Изобретение благоприятствует высокой точности амплификации ДНК в условиях, которые допускают очистку от переброса и синтеза длиномерных продуктов. Изобретение может найти применение для различных промышленных, медицинских и судебно-медицинских целей.
Синтез нуклеиновых кислот in vitro обычно осуществляют в присутствии ДНК-полимераз с дополнительными полипептидами или без дополнительных полипептидов. ДНК-полимеразы представляют собой семейство ферментов, участвующих в репликации и репарации ДНК. Широкие исследования проведены по выделению ДНК-полимераз из мезофильных микроорганизмов, таких, как Е.coli, например, Bessman et al. (1957), J. Biol. Chem. 223:171-177 и Buttin and Kornberg (1966), J. Biol. Chem. 241: 5419-5427.
Проводились также исследования по выделению и очистке ДНК-полимераз из термофилов, таких, как Thermus aquaticus. В публикации Chien А. et al. (1976), J. Bacteriol. 127:1550-1557, описаны выделение и очистка ДНК-полимеразы с температурным оптимумом 80°С из штамма YT1 Thermus aquaticus. В патенте США №4889818 описана очищенная термостабильная ДНК-полимераза из Т.aquaticus, полимераза Taq, имеющая молекулярную массу примерно 86000-90000 Да. Кроме того, заявка на Европейский патент №0258017 раскрывает полимеразу Taq в качестве предпочтительного фермента для применения в процессе PCR (полимеразной цепной реакции).
Исследования указывают на то, что в то время как ДНК-полимераза Taq обладает 5'-3'-полимераза-зависимой функцией экзонуклеазы, она не обладает функцией 3'-5'-экзонуклеазы III (Lawyer F.C. et al. (1989), J. Biol. Chem. 264:6427-6437; Bernard A. et al. (1989), Cell, 59:219). 3'-5'-Экзонуклеазную активность ДНК-полимераз обычно называют «корректирующей активностью». 3'-5'-Экзонуклеазная активность удаляет основания, которые ошибочно спарены на 3'-конце праймер-матричного дуплекса. Наличие 3'-5'-экзонуклеазной активности может считаться преимуществом, поскольку она приводит к повышению точности репликации цепей нуклеиновых кислот и к элонгации преждевременно терминированных продуктов. Поскольку полимераза Taq не способна удалять ошибочно спаренные концы праймеров, она предрасположена к ошибкам при включении оснований, что делает ее применение в некоторых случаях нежелательным. Например, попытка клонировать амплифицированный ген является проблематичной, поскольку любая копия гена может содержать ошибку из-за случайного ошибочного включения. В зависимости от цикла, при котором происходит ошибка (например, в цикле ранней репликации), полностью амплифицированная ДНК могла бы содержать ошибочно включенное основание, приводя таким образом к мутированному генному продукту.
Существует несколько термостабильных ДНК-полимераз, известных в этой области, которые обладают 3'-5'-экзонуклеазной активностью подобно полимеразам типа В из термофильных Archaebacteria, которые используются для высокоточной амплификации ДНК. Термостабильные полимеразы, обладающие 3'-5'-экзонуклеазной активностью, могут быть выделены или клонированы от Pyrococcus (очищенная термостабильная ДНК-полимераза Pyrococcus furiosus, Mathur E., фирма Stratagene, Международная заявка 92/09689, патент США №5545552; очищенная термостабильная ДНК-полимераза из Pyrococcus species. Comb D.G. et al., фирма New England Biolabs, Inc., Европейский патент №0547359; организация и нуклеотидная последовательность гена ДНК-полимеразы из археонных Pyrococcus furiosus, Uemori Т. et al. (1993), Nucl. Acids Res., 21:259-265), из Pyrodictium spec. (термостабильная полимераза нуклеиновых кислот, Gelfand D.H., фирма Hoffmann-La Roche AG, Европейский патент №0624641; очищенная термостабильная полимераза нуклеиновых кислот и кодирующие ДНК последовательности из разновидностей Pyrodictium species, Gelfand D.H., фирма Hoffmann-La Roche Inc., патент США №5491086), из Thermococcus (например, термостабильная ДНК-полимераза из разновидностей Thermococcus TY, Niehaus F. et al., Международная заявка 97/35988; очищенная ДНК-полимераза из Thermococcus barossii, Luhm R.A., фирма Pharmacia Biotech, Inc., Международная заявка 96/22389; ДНК-полимераза из Thermococcus barossii с промежуточной экзонуклеазной активностью и повышенной долговременной стабильностью при высокой температуре, применяемая для секвенирования ДНК, полимеразной цепной реакции и т.д., Dhennezel О.В., фирма Pharmacia Biotech Inc., Международная заявка №96/22389; очищенная термостабильная ДНК-полимераза из Thermococcus litoralis для применения в манипуляциях с ДНК, Comb D.G., фирма New England Biolabs, Inc., патент США №5322785, Европейский патент №0455430; рекомбинантная термостабильная ДНК-полимераза из Archaebacteria, Comb D.G., фирма New England Biolabs, Inc., патент США №5352778, Европейский патент №0547920, Европейский патент №0701000; новая выделенная термостабильная ДНК-полимераза, полученная из Thermococcus gorgonarius, Angerer B. et al., фирма Boehringer Mannheim GmbH, Международная заявка №98/14590).
Другая возможность проведения PCR в присутствии корректирующей функции состоит в применении смеси полимеразных ферментов, причем одна полимераза обладает такой корректирующей активностью (например, термостабильная ДНК-полимераза с повышенной термостабильностью и увеличенной протяженностью и эффективностью праймера, Barnes W.M., патент США №5436149, Европейский патент №0693078; новые полимеразные композиции и их применение, Sorge J.A., фирма Stratagene, Международная заявка №95/16028). Обычная практика состоит в использовании композиции термостабильной ДНК-полимеразы, содержащей основной компонент, по меньшей мере одну термостабильную ДНК-полимеразу, которая лишена 3'-5'-экзонуклеазной активности, и минорный компонент, обладающий 3'-5'-экзонуклеазной активностью, например, полимеразу Taq и ДНК-полимеразу Pfu. В этих смесях последовательное ферментативное действие осуществляется с помощью фермента типа pol I, подобного полимеразе Taq, корректирующая функция - с помощью термостабильной полимеразы типа В, подобной полимеразе Pfu. Высокая точность синтеза ДНК является одним из желаемых параметров при амплификации нуклеиновой кислоты, другой важной характеристикой является возможность освобождения от примесей.
Полимеразная цепная реакция может амплифицировать одну молекулу свыше миллиарда раз. Таким образом, даже минимальные количества примесей могут амплифицироваться и привести к ложно положительному результату. Такие примеси часто являются продуктами предшествующих амплификаций с помощью PCR (внесенные примеси). Поэтому исследователи создали метод, позволяющий избежать такого загрязнения.
Методика состоит в замене ТТР (тимидинтрифосфата) на dUTP (дезоксиуридинтрифосфат) при амплификации методом PCR с целью получения урацилсодержащей ДНК (U-ДНК). Последующая обработка реакционных смесей PCR урацил-ДНК-гликозилазой (UNG) перед амплификацией методом PCR приводит к деградации примесной нуклеиновой кислоты и делает ее непригодной для амплификации. Включение dUTP может быть легко осуществлено в присутствии термостабильной полимеразы типа pol I, но не полимеразы типа В (G.Slupphaug et al. (1993), Anal. Biochem. 211:164-169). Низкий уровень включения dUTP полимеразой типа В ограничивает ее применение в лабораториях, в которых такой же тип матрицы повторно анализируют амплификацией методом PCR.
Термостабильные ДНК-полимеразы, обладающие 3'-5'-экзонуклеазной активностью, были выделены также из эубактериальных штаммов, подобных Thermotoga (термофильные ДНК-полимеразы из Thermotoga neapolitana, Slater M.R. et al., фирма Promega Corporation, Международная заявка №96/41014; клонированные ДНК-полимеразы из Thermotoga neapolitana и его мутантов, Hughes A.J. et al., фирма Life Technologies, Inc., Международная заявка №96/10640; очищенная термостабильная полимераза нуклеиновых кислот из Thermotoga maritima, Gelfand D.H. et al., фирма CETUS Corporation, Международная заявка №92/03556). Эти ферменты обладают высокой 3'-5'-экзонуклеазной активностью, которая способна элиминировать ошибочно включенные или ошибочно спаренные основания. Генно-инженерный вариант этого фермента коммерчески доступен как ULTma, ДНК-полимераза, которая может быть использована в PCR без дополнительных полипептидов. Данный фермент способен удалять ошибочно включенные основания, включать dUTP, но качество этих его функций по неизвестным причинам не выше, чем у полимеразы Taq (точность репликации в полимеразной цепной реакции, Diaz R.S. et al. Braz. J. Med. Biol. Res. (1988), 31:1239-1242; точность ДНК-полимеразы Pfu и других термостабильных ДНК-полимераз в ПЦР, Cline et al., Nucleic Acids Res. (1996), 24:3546-3551).
Для высокоточного синтеза ДНК альтернативой применению полимераз типа В или содержащих их смесей является применение холофермента, термофильной ДНК-полимеразы III, комплекса из 18 полипептидных цепей. Такие комплексы идентичны бактериальным хромосомным репликазам, содержащим все факторы, необходимые для синтеза цепи ДНК в несколько сот тысяч оснований или хромосом полностью. Десять различных субъединиц этого фермента, некоторые из которых присутствуют в виде множественных копий, могут быть получены рекомбинантными методами, реконструированы и использованы для синтеза ДНК in vitro. В качестве возможного использования этих комплексов предложена амплификация PCR нуклеиновых кислот из нескольких тысяч до сотен тысяч пар оснований (Фермент, полученный из термофильных организмов, который функционирует как хромосомная репликаза, его получение и применение, Yurieva О. et al., The Rockefeller University, Международная заявка №98/45452; холофермент, новая термофильная полимераза III, McHenry С., фирма ENZYCO Inc., Международная заявка №99/13060).
Цель данного изобретения состояла в создании высокоточной системы PCR, которая предпочтительно сопутствующим образом была способна включать dUTP. Настоящее изобретение обеспечивает термостабильный фермент, обладающий 3'-экзонуклеазной активностью, но фактически лишенный активности ДНК-полимеразы, тогда как этот фермент повышает качество процесса амплификации при добавлении ко второму ферменту, обладающему полимеразной активностью. Обеспечиваемый фермент может вырезать ошибочно спаренные концы праймеров, давая возможность второму ферменту, обладающему полимеразной активностью, как, например, полимеразе Taq, разъединять и восстанавливать элонгацию в ходе процесса синтеза ДНК. Фермент согласно изобретению способен к взаимодействию в качестве корректирующего фермента со вторым ферментом, обладающим полимеразной активностью. Ферментом, который, как было показано, применим для этой цели, является, например термостабильная экзонуклеаза III. Предпочтительной является экзонуклеаза III, действующая в направлении от 3' к 5', отщепляя 5'-фосфат, оставляя 3'-гидроксильные группы, и идеально действующая только на двухцепочечной ДНК. ДНК-полимер азы с 3'-5'-экзонуклеазными функциями активны на двух- и одноцепочечной ДНК. Последняя активность может привести к деградации праймера, что является нежелательным в пробах с PCR. Предпочтительно, чтобы фермент был активен при 70°С до 80°С, достаточно устойчив для того, чтобы выдержать циклы денатурации, и неактивен при более низких температурах, не вызывая деградацию продуктов PCR после завершения процесса PCR. Ферменты, обладающие этими свойствами, могут быть получены из термофильных эубактерий или родственных ферментов из термофильных архебактерий. Геномы трех термостабильных архебактерий секвенированы, Methanococcus jannaschii (Complete Genome Sequence of the Methanogenic Archaeon, Methanococcus jannaschii, Bult C.J. et al. (1996) Science 273:1058-1072), Methanobacterium Thermoautotrophicum (Complete genomic sequence of Methanobacterium thermoautotrophicum ΔH: Functional Analysis and Comparative Genomics, Smith D.R. et al., J. of Bacteriol. (1997), 179: 7135-7155) и Archaeoglobus fulgidus (The complete genome sequence of the hyperthermophilie, sulfate-reducing archaeon Archaeoglobus fulgidus, Klenk H.-P. et al. (1997) Nature 390:364-370).
В частности, обеспечивается термостабильный фермент, получаемый из Archaeoglobus fulgidus, который катализирует деградацию ошибочно спаренных концов праймеров полинуклеотидов в направлении 3'-5' в двухцепочечной ДНК. Ген, кодирующий термостабильную экзонуклеазу III, получаемую из Archaeoglobus fulgidus (Afu), клонировали, экспрессировали в Е.coli и выделяли. Фермент активен в условиях инкубации и температуры, применяемых в PCR. Фермент поддерживает ДНК-полимеразу, подобную Taq, в осуществлении синтеза ДНК при низкой частоте ошибок и синтеза продуктов более чем 3 т.н. на геномной ДНК - верхний предел продуктов, синтезируемых полимеразой Taq, - с хорошими выходами в присутствии или в отсутствие dUTP в реакционной смеси. Предпочтительно используют 50-500 нг экзонуклеазы III, получаемой из Afu, на 2.5 единицы полимеразы Taq для того, чтобы получить оптимальный режим PCR. Более предпочтительно использовать в PCR от 67 нг до 380 нг экзонуклеазы III, получаемой из Afu, на 2.5 единицы полимеразы Taq.
Таким образом, фермент согласно изобретению способен взаимодействовать с полимеразой Taq в качестве корректирующего фермента. Преимущество использования фермента согласно изобретению по сравнению с другими ферментами состоит в том, что фермент согласно изобретению предпочтительно активен на двухцепочечной ДНК. Термостабильный фермент данного изобретения может быть применен для любой цели, при которой такая ферментативная активность является необходимой или желательной. В особо предпочтительном варианте осуществления изобретения фермент применяют в комбинации с термостабильной ДНК-полимеразой в реакции амплификации нуклеиновой кислоты, известной как PCR, чтобы удалить ошибочно спаренные концы праймеров, которые приводят к преждевременным остановкам, обеспечить концы праймеров, которые более эффективно элонгируются полимеразой, исправить ошибки включения оснований и дать возможность полимеразе образовать протяженные продукты PCR.
Кроме того, объектом настоящего изобретения является композиция, включающая в себя первый термостабильный фермент, обладающий 3'-экзонуклеазной активностью, но фактически лишенный активности ДНК-полимеразы, и второй фермент, обладающий полимеразной активостью, поскольку точность процесса амплификации повышается при применении этой композиции по сравнению с применением одного только второго фермента. Термостабильный фермент согласно изобретению, обладающий 3'-экзонуклеазной активностью, но фактически лишенный активности ДНК-полимеразы, включает в себя также соответствующие ферменты, обладающие пониженной активностью ДНК-полимеразы или совсем не имеющие такой активности. Пониженная ДНК-полимеразная активность согласно изобретению означает менее чем 50% упомянутой активности фермента, обладающего активностью ДНК-полимеразы. В предпочтительном варианте осуществления изобретения второй фермент композиции согласно изобретению лишен корректирующей активности. В особо предпочтительном варианте вторым ферментом является полимераза Taq.
Другим объектом настоящего изобретения является способ синтеза ДНК с использованием смеси, включающей в себя первый термостабильный фермент, обладающий 3'-экзонуклеазной активностью, но фактически лишенный активности ДНК-полимеразы, и второй фермент, обладающий полимеразной активностью. В соответствии с этим способом преждевременно терминированные цепи вырезаются путем деградации от 3' к 5'. По этому способу удаляются ошибочно спаренные концы или праймера, или растущей цепи.
Изобретение далее включает в себя способ, соответствующий приведенному выше описанию, в котором в реакционной смеси присутствует dUTP, заменяя частично или полностью ТТР. Предпочтительным по данному способу является использование урацил-ДНК-гликозилазы (UDG или UNG) для деградации примесных нуклеиновых кислот.
Предпочтительно по данному способу, когда смесь, состоящая из первого термостабильного фермента, обладающего 3'-экзонуклеазной активностью, но фактически лишенного активности ДНК-полимеразы, и второго фермента, обладающего полимеразной активностью, образует продукты PCR с более низкой частотой ошибок по сравнению с продуктами PCR, полученными в присутствии второго фермента, обладающего полимеразной активностью, в отсутствие первого термостабильного фермента, обладающего 3'-экзонуклеазной активностью, но фактически лишенного активности ДНК-полимеразы. Способ, при котором смесь первого термостабильного фермента, обладающего 3'-экзонуклеазной активностью, но фактически лишенного активности ДНК-полимеразы, и второго фермента, обладающего полимеразной активностью, образует продукты PCR большей протяженности по сравнению с продуктами PCR, полученными в присутствии второго фермента, обладающего полимеразной активностью, в отсутствие первого термостабильного фермента, обладающего 3'-экзонуклеазной активностью, но фактически лишенного активности ДНК-полимеразы. Кроме того, первый термостабильный фермент, обладающий 3'-экзонуклеазной активностью, но фактически лишенный активности ДНК-полимеразы, является родственным экзонуклеазе III из Е.coli, но термостабильным по данному способу. Дополнительным вариантом воплощения описанного выше способа является способ, при котором получают продукты PCR с тупыми концами.
Объектом настоящего изобретения являются также способы получения термостабильного фермента согласно изобретению, обладающего 3'-экзонуклеазной активностью, но фактически лишенного активности ДНК-полимеразы, и средства и материалы для получения данного фермента, как, например, векторы и клетки-хозяева (например, DSM (Немецкая коллекция микроорганизмов) №-13021).
Следующие примеры предлагаются с целью иллюстрации, но не ограничения объекта изобретения.
Подробное описание предпочтительных вариантов осуществления изобретения
Краткое описание чертежей.
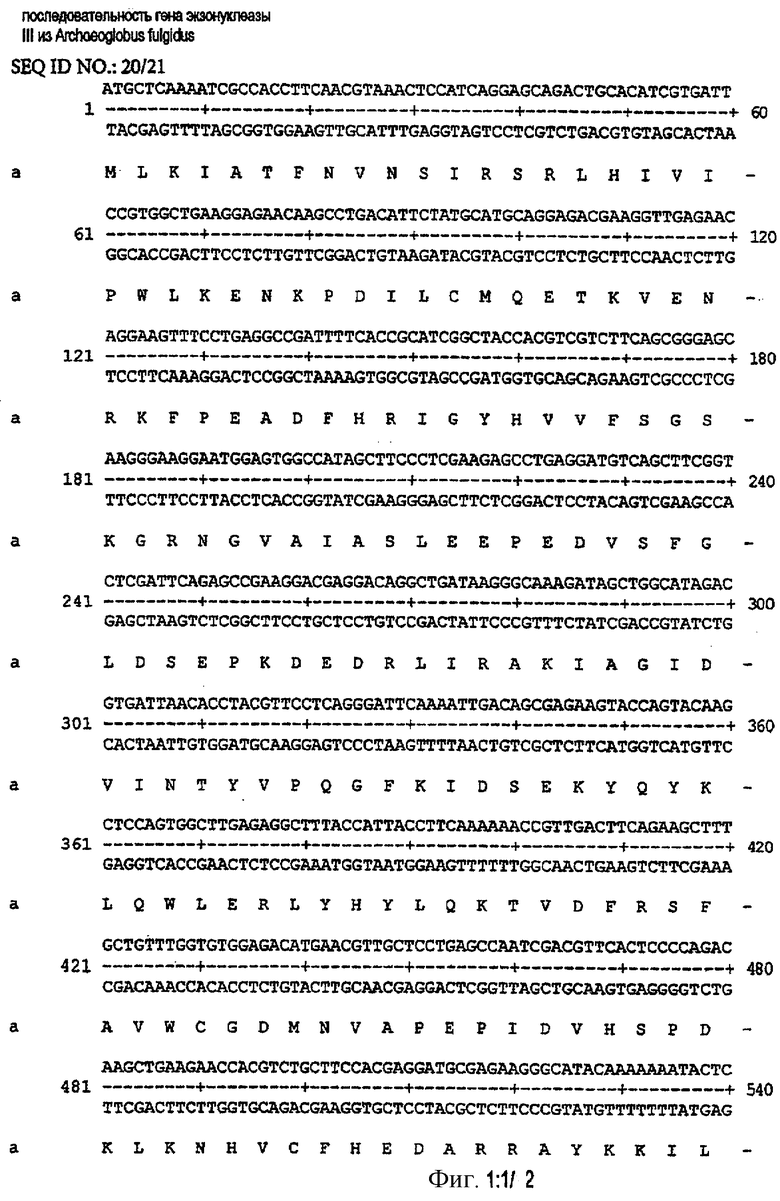
Фиг.1.
Последовательность ДНК и установленная аминокислотная последовательность гена, кодирующего ДНК-полимеразу из экзонуклеазы III Archaeoglobus fulgidus.
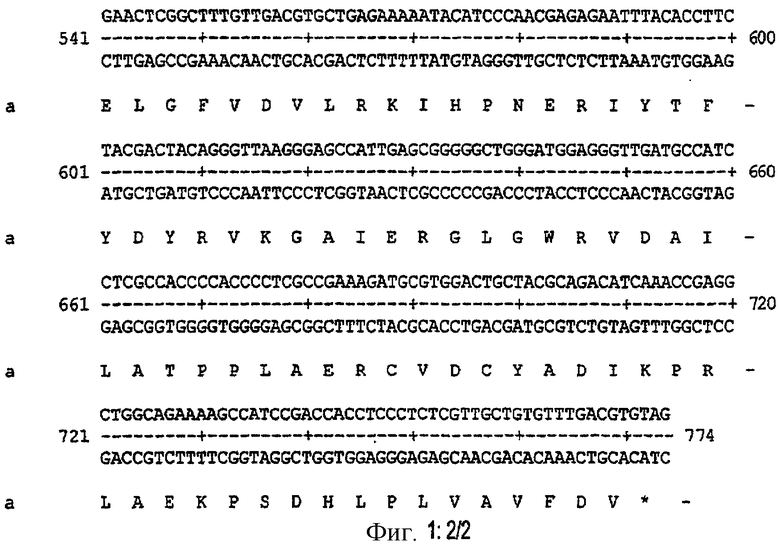
Фиг.2.
Устойчивость к тепловой денатурации рекомбинантной экзонуклеазы III из Archaeoglobus fulgidus, экспрессированной в Е.coli, как описано в примере V.
Дорожка 1: инкубация при 50°С.
Дорожка 2: инкубация при 60°С.
Дорожка 3: инкубация при 70°С.
Дорожка 4: инкубация при 80°С.
Дорожка 5: инкубация при 90°С.
Дорожка 6: экстракт клетки-хозяина Е.coil, не трансформированный геном, кодирующим экзонуклеазу III Afu.
Дорожка 7: экзонуклеаза III из Е.coli.
Дорожка 8: маркер молекулярной массы.
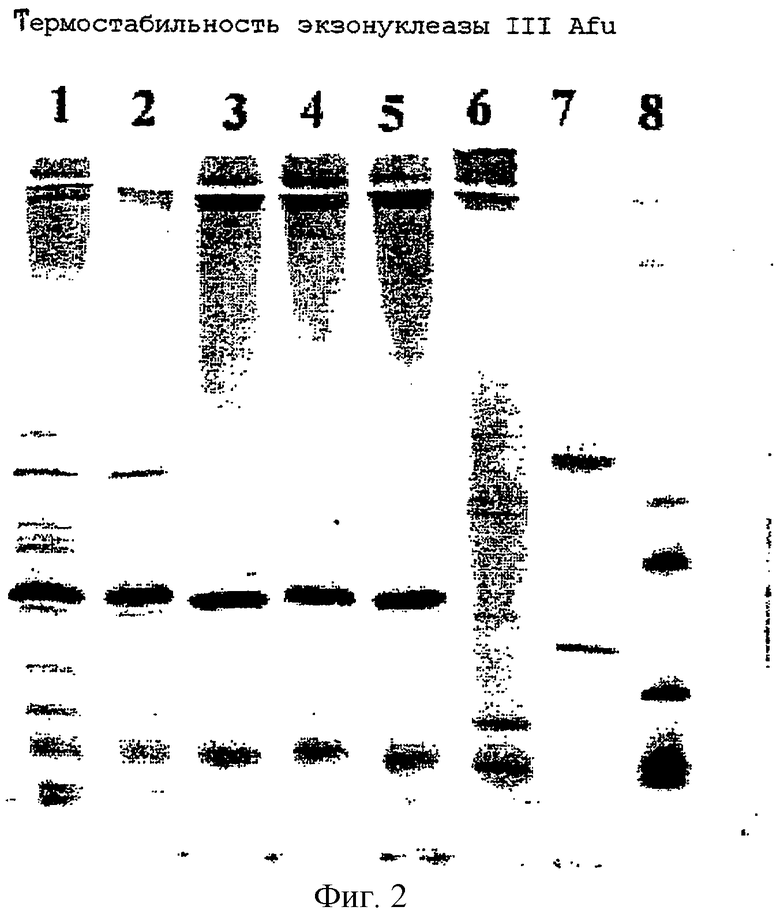
Фиг.3.
Экзонуклеазная активность экзонуклеазы III Afu на фрагментах ДНК, как описано в примере VI.
Дорожка 1: 10 единиц экзонуклеазы III из Е.coli, инкубация при 37°С.
Дорожка 2: 50 нг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 3: 100 нг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 4: 150 нг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 5: 100 нг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 6: 200 нг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 7: 300 нг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 8: 250 нг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 9: 750 нг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 10: 1 мкг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 11: 500 нг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 12: 1 мкг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 13: 1,5 мкг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 14: 1,5 мкг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 15: 3 мкг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 16: 4,5 мкг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 17: 7,6 мкг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 18: 15,2 мкг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 19: 22,8 мкг экзонуклеазы III Afu, инкубация при 72°С.
Дорожка 2: без добавления экзонуклеазы.
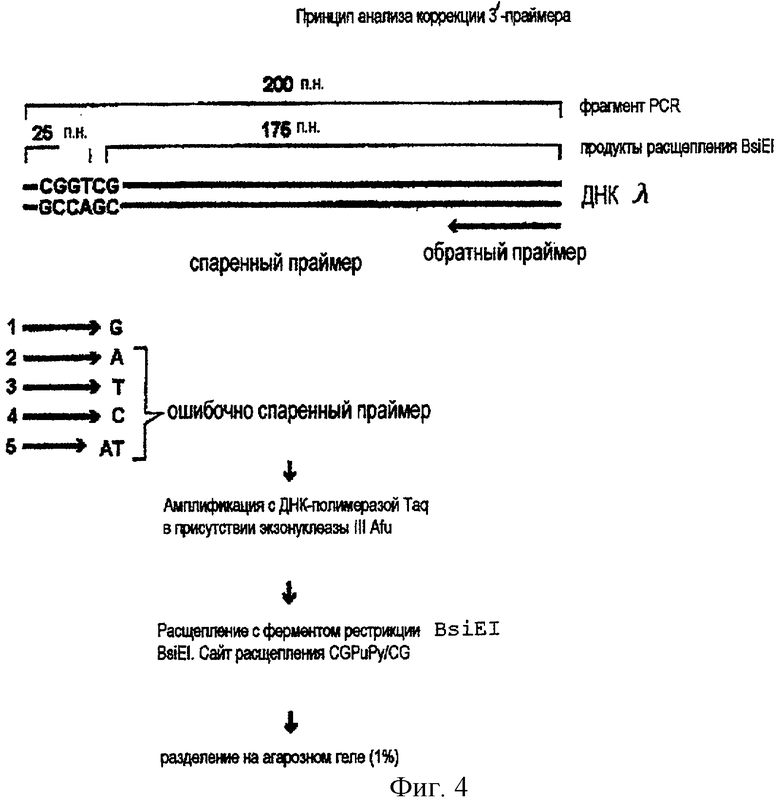
Фиг.4.
Принцип коррекционного анализа ошибочного спаривания.
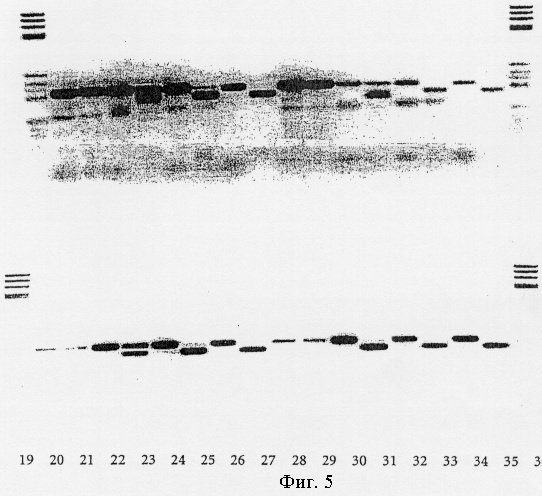
Фиг.5.
Коррекция ошибочного спаривания праймера в PCR, как описано в примере VII.
Дорожка 1: маркер V молекулярной массы ДНК (ROCHE Molecular Biochemicals №821705).
Дорожка 2: G:А-ошибочно спаренный праймер, амплификация с ДНК-полимеразой Taq.
Дорожка 3: как в дорожке 2, но с последующим расщеплением рестриктазой BsiEI.
Дорожка 4: G:А-ошибочно спаренный праймер, амплификация с системой PCR Expand HiFi.
Дорожка 5: как в дорожке 4, но с последующим расщеплением с BsiEI.
Дорожка 6: G:А-ошибочно спаренный праймер, амплификация с полимеразой Taq/экзонуклеазой III Afu.
Дорожка 7: как в дорожке 6, но с последующим расщеплением с BsiEI.
Дорожка 8: G:А-ошибочно спаренный праймер, амплификация с ДНК-полимеразой Tgo.
Дорожка 9: как в дорожке 8, но с последующим расщеплением с BsiEI.
Дорожка 10: G:Т-ошибочно спаренный праймер, амплификация с ДНК-полимеразой Taq.
Дорожка 11: как в дорожке 10, но с последующим расщеплением с BsiEI.
Дорожка 12: G:Т-ошибочно спаренный праймер, амплификация с системой PCR Expand HiFi.
Дорожка 13: как в дорожке 12, но с последующим расщеплением с BsiEI.
Дорожка 14: G:Т-ошибочно спаренный праймер, амплификация с полимеразой Taq/экзонуклеазой III Afu.
Дорожка 15: как в дорожке 14, но с последующим расщеплением с BsiEI.
Дорожка 16: G:Т-ошибочно спаренный праймер, амплификация с ДНК-полимеразой Tgo.
Дорожка 17: как в дорожке 16, но с последующим расщеплением с BsiEI.
Дорожка 18: маркер V молекулярной массы ДНК.
Дорожка 19: маркер V молекулярной массы ДНК.
Дорожка 20: G:С-ошибочно спаренный праймер, амплификация с ДНК-полимеразой Taq.
Дорожка 21: как в дорожке 20, но с последующим расщеплением с BsiEI.
Дорожка 22: G:С-ошибочно спаренный праймер, амплификация с системой PCR Expand HiFi.
Дорожка 23: как в дорожке 22, но с последующим расщеплением с BsiEI.
Дорожка 24: G:С-ошибочно спаренный праймер, амплификация с полимеразой Taq/экзонуклеазой III Afu.
Дорожка 25: как в дорожке 24, но с последующим расщеплением с BsiEI.
Дорожка 26: G:С-ошибочно спаренный праймер, амплификация с ДНК-полимеразой Tgo.
Дорожка 27: как в дорожке 26, но с последующим расщеплением с BsiEI.
Дорожка 28: CG:АТ-ошибочно спаренный праймер, ДНК-полимераза Taq.
Дорожка 29: как в дорожке 28, но с последующим расщеплением с BsiEI.
Дорожка 30: CG:АТ-ошибочно спаренный праймер, система PCR Expand HiFi.
Дорожка 31: как в дорожке 2, но с последующим расщеплением с BsiEI.
Дорожка 32: CG:AT-ошибочно спаренный праймер, полимераза Taq/экзонуклеаза III Afu.
Дорожка 33: как в дорожке 2, но с последующим расщеплением с BsiEI.
Дорожка 34: CG:АТ-ошибочно спаренный праймер, амплификация с ДНК-полимеразой Tgo.
Дорожка 35: как в дорожке 2, но с последующим расщеплением с BsiEI.
Дорожка 36: маркер V молекулярной массы ДНК.
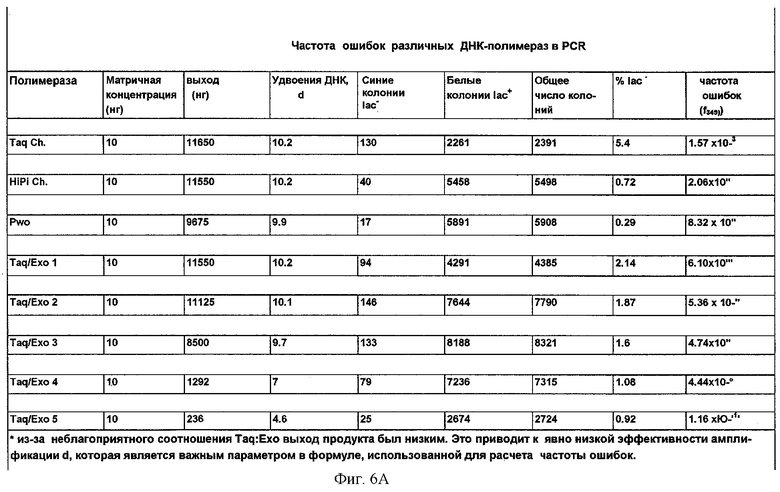
Фиг.6А.
Частота ошибок различных полимераз в PCR.
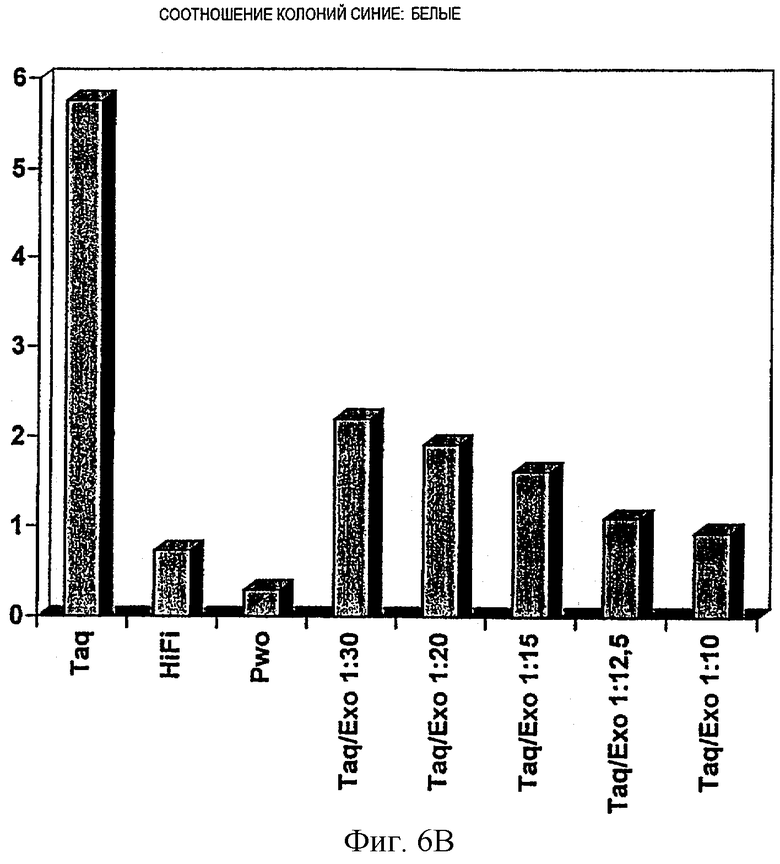
Фиг.6В.
Повышение точности экзонуклеазой III Afu, присутствующей в смеси PCR, как описано в примере VIII.
Колонии с соотношением синие:белые блотировали, и различные смеси ДНК-полимеразы Taq и экзонуклеазы III Afu (Taq/экзо 1:30, Taq/экзо 1:20, Taq/экзо 1:15, Taq/экзо 1:12.5, Taq/экзо 1:10, соответствующие 2.5 ед. ДНК-полимеразы Taq, смешанной со 125 нг, 175 нг, 250 нг, 375 нг и 500 нг экзонуклеазы III Afu, соответственно) исследовали в сравнении с ДНК-полимеразой Taq (Taq), системой PCR Expand HiFi (HiFi) и ДНК-полимеразой Pwo (Pwo).
Фиг.7.
Включение dUTP смесью ДНК-полимеразы Taq/экзонуклеазы III Afu, как описано в примере IX.
Дорожка 1: маркер XIV молекулярной массы ДНК (Roche Molecular Biochemicals №1721933).
Дорожка 2: амплификация с 2.5 ед. ДНК-полимеразы Taq.
Дорожка 3: амплификация с 2.5 ед. ДНК-полимеразы Taq и 125 нг экзонуклеазы III Afu.
Дорожка 4: амплификация с 2.5 ед. ДНК-полимеразы Taq и 250 нг экзонуклеазы III Afu.
Дорожка 5: амплификация с 2.5 ед. ДНК-полимеразы Taq и 375 нг экзонуклеазы III Afu.
Дорожка 6: амплификация с 2.5 ед. ДНК-полимеразы Taq и 500 нг экзонуклеазы III Afu.
Фиг.8.
Деградация продуктов PCR, содержащих dUTP, урацил-ДНК-гликозилазой, как описано в примере IX.
Дорожка 1: маркер XIV молекулярной массы ДНК (Roche Molecular Biochemicals №1721933).
Дорожка 2: 1 мкл продукта амплификации, полученного с ДНК-полимеразой Taq, 125 нг экзонуклеазы III Afu и последующей обработкой UNG и теплом.
Дорожка 3: 2 мкл продукта амплификации, полученного с ДНК-полимеразой Taq, 125 нг экзонуклеазы III Afu и последующей обработкой UNG и теплом.
Дорожка 4: 3 мкл продукта амплификации, полученного с ДНК-полимеразой Taq, 125 нг экзонуклеазы III Afu и последующей обработкой UNG и теплом.
Дорожка 5: 4 мкл продукта амплификации, полученного с ДНК-полимеразой Taq, 125 нг экзонуклеазы III Afu и последующей обработкой UNG и теплом.
Дорожка 6: 5 мкл продукта амплификации, полученного с ДНК-полимеразой Taq, 125 нг экзонуклеазы III Afu и последующей обработкой UNG или теплом.
Дорожка 7: 5 мкл продукта амплификации, полученного с ДНК-полимеразой Taq, 125 нг экзонуклеазы III Afu без последующей обработки UNG и теплом.
Дорожка 8: 5 мкл продукта амплификации, полученного с ДНК-полимеразой Taq, 125 нг экзонуклеазы III Afu без последующей обработки UNG, но с обработкой теплом.
Дорожка 9: маркер XIV молекулярной массы ДНК (Roche Molecular Biochemicals №1721933).
Фиг.9.
Влияние экзонуклеазы III Afu на протяженность продукта PCR.
Смесь ДНК-полимеразы Taq/экзонуклеазы III Afu анализировали на человеческой геномной ДНК, как описано в примере X.
Дорожка 1: фрагмент tPA 9.3 т.н. со смесью Taq/экзо III.
Дорожка 2: фрагмент tPA 9.3 т.н. с Taq-Pol.
Дорожка 3: фрагмент tPA 12 т.н. со смесью Taq/экзо III.
Дорожка 4: фрагмент tPA 12 т.н. с Taq/Pol.
Дорожка 5: фрагмент tPA 15 т.н. со смесью Taq/экзо III.
Дорожка 6: фрагмент tPA 15 т.н. с Taq-Pol.
Фиг.10.
Термостабильная экзонуклеаза III может быть заменена мутантной полимеразой с пониженной полимеразной активностью, но повышенной 3'-экзонуклеазной активностью, как описано в примере XI.
Дорожка 1: маркер молекулярной массы.
Дорожка 2: реакция 1, полимераза Taq, фрагмент 4.8 т.н.
Дорожка 3: реакция 2, полимераза Taq плюс мутантная полимераза Taq, фрагмент 4.8 т.н.
Дорожка 4: реакция 3, без полимеразы Taq, мутантная полимераза Taq, фрагмент 4.8 т.н.
Дорожка 5: реакция 4, полимераза Taq плюс экзонуклеаза III Afu, фрагмент 4.8 т.н.
Дорожка 6: реакция 5, полимераза Taq, фрагмент 9.3 т.н.
Дорожка 7: реакция 6, полимераза Taq плюс мутантная полимераза Taq, фрагмент 9.3 т.н.
Дорожка 8: реакция 7, без полимеразы Taq, мутантная полимераза Taq, фрагмент 9.3 т.н.
Дорожка 9: реакция 8, полимераза Taq плюс экзонуклеаза III Afu, фрагмент 9.3 т.н.
Дорожка 10: маркер молекулярной массы.
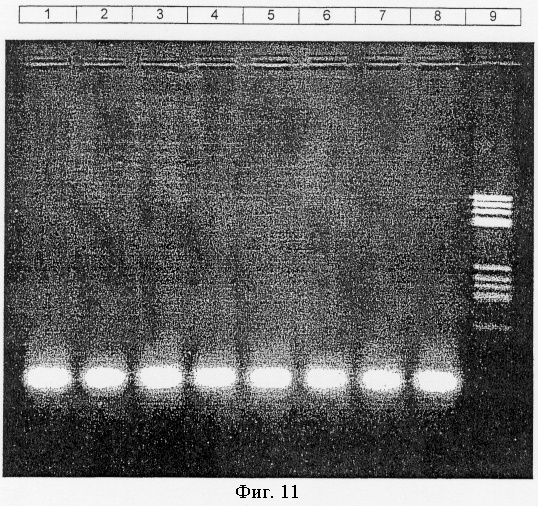
Фиг.11.
Экзонуклеаза III Afu неактивна на линейной одноцепочечной ДНК, как описано в примере XII.
Дорожка 1: экзонуклеаза III Afu, без инкубации.
Дорожка 2: экзонуклеаза III Afu, 1 ч при 65°С.
Дорожка 3: экзонуклеаза III Afu, 2 ч при 65°С.
Дорожка 4: экзонуклеаза III Afu, 3 ч при 65°С.
Дорожка 5: экзонуклеаза III Afu, 4 ч при 65°С.
Дорожка 6: экзонуклеаза III Afu, 5 ч при 65°С.
Дорожка 7: реакционный буфер без фермента, без инкубации.
Дорожка 8: реакционный буфер без фермента, 5 ч при 65°С.
Дорожка 9: маркер молекулярной массы.
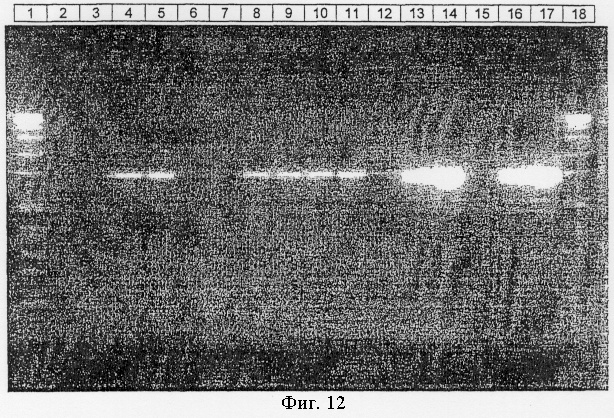
Фиг.12.
Сравнение экзонуклеазы III Afu с термостабильной полимеразой типа В по деградирующей праймер активности, как описано в примере XIII.
Дорожка 1: маркер молекулярной массы.
Дорожка 2: 1 ед. Tgo, проинкубирована (реакция 1).
Дорожка 3: 1.5 ед. Tgo, преинкубирована (реакция 2).
Дорожка 4: 1 ед. Tgo, без преинкубации (реакция 3).
Дорожка 5: 1.5 ед. Tgo, без преинкубации (реакция 4).
Дорожка 6: 1 ед. Tgo, преинкубирована в отсутствие dNTPs (дезоксинуклеозидтрифосфатов) (реакция 5).
Дорожка 7: 1.5 ед. Tgo, принкубирована в отсутствие dNTPs (реакция 6).
Дорожка 8: 1 ед. Tgo, без преинкубации в отсутствие dNTPs (реакция 7).
Дорожка 9: 1.5 ед. Tgo, без преинкубации в отсутствие dNTPs (реакция 8).
Дорожка 10: 1 ед. Tgo, преинкубирована в отсутствие dNTPs с добавлением дополнительного праймера (реакция 9).
Дорожка 11: 1.5 ед. Tgo, преинкубирована в отсутствие dNTPs с добавлением дополнительного праймера (реакция 10).
Дорожка 12: полимераза Taq, преинкубирована (реакция 11).
Дорожка 13: Taq плюс 37.5 нг экзонуклеазы III Afu, проинкубированы (реакция 12).
Дорожка 14: Taq плюс 75 нг экзонуклеазы III Afu, проинкубированы (реакция 13).
Дорожка 15: полимераза Taq, без преинкубации (реакция 14).
Дорожка 16: Taq плюс 37,5 нг экзонуклеазы III Afu, без преинкубации (реакция 15).
Дорожка 17: Taq плюс 75 нг экзонуклеазы III Afu, без преинкубации (реакция 16).
Дорожка 18: маркер молекулярной массы.
Пример I
Выделение кодирующих последовательностей
Предпочтительным термостабильным ферментом согласно изобретению является чрезвычайно термостабильная экзодезоксирибонуклеаза, получаемая из Archaeoglobus fulgidus штамм VC-16 (DSM №4304). Штамм выделен из морских гидротермальных организмов на Острове вулканов и Stufe di Nerone, Неаполь, Италия (Stetter К.О. et al., Science (1987) 236:822-824). Данный организм является чрезвычайно термофильной, метаболизирующей серу архебактерией с ростом в интервале между 60°С и 95°С и оптимумом при 83°С (Klenk H.P. et al., Nature (1997) 390:364-370). Геномная последовательность внесена в банк данных TIGR. Ген, предположительно кодирующий экзонуклеазу III (xthA), имеет номер доступа AF0580. Кажущаяся молекулярная масса экзодезоксирибонуклеазы, получаемой из Archaeoglobus fulgidus, составляет примерно 32000 Да при сопоставлении с белковыми стандартами известной молекулярной массы (SDS-PAGE) (электрофорез в полиакриламидном геле с додецилсульфатом натрия). Точная молекулярная масса термостабильного фермента согласно изобретению может быть определена по кодирующей последовательности гена экзодезоксирибонуклеазы III из Archaeoglobus fulgidus.
Пример II
Клонирование гена, кодирующего экзонуклеазу III из Archaeoglobus fulgidus
Для выделения хромосомной ДНК из Archaeoglobus fulgidus использовали около 6 мл клеточной культуры DSM №4304.
Создавали следующие праймеры с рестриционными сайтами, совместимыми с множественным клонирующим сайтом желаемого экспрессирующего вектора и комплементарные N- и С-концам гена экзонуклеазы III из Archaeoglobus fulgidus:
SEQ ID NO.: 1 (последовательность с идентификационным №1):
N-конец (сайт BamHI): 5'-GAAACGAGGATCCATGCTCAAAATCGCCACC-3'
SEQ ID NO.: 2
5'-TTG TTC ACT GCA GCT АСА CGT CAA АСА CAG С-3'
С-конец (сайт PstI):
Сначала клетки собирали повторным центрифугированием при 5000 об/мин в одной пробирке Эппендорфа на 2 мл. Выделение ДНК можно проводить любым методом, описанным для выделения из бактериальных клеток. В данном случае геномную ДНК из Archaeoglobus fulgidus получали с набором High PureTM для матричной PCR (ROCHE Diagnostics GmbH, №1796828). Таким способом получали примерно 6 мкг хромосомной ДНК с концентрацией 72 нг/мкл.
Проводили PCR с праймерами, описанными выше, в высокоточной системе PCR ExpandTM (ROCHE Diagnostics GmbH, №1732641) и 100 нг геномной ДНК из Archaeoglobus fulgidus на пробирку в четырех идентичных препаратах. Проводили PCR при следующих условиях:
1×94°С, 2 мин;
10×94°С, 10 сек; 54°С, 30 сек; 68°С, 3 мин;
20×94°С, 10 сек; 54°С, 30 сек; 68°С, 3 мин с циклом элонгации 20 сек для каждого цикла;
1×68°С, 7 мин.
После прибавления MgCl2 до конечной концентрации 10 мМ продукт PCR расщепляли с рестриктазами BamHI и PstI, 10 единиц каждой при 37°С в течение 2 часов. Продукты реакции разделяли на низкоплавком агарозном геле. После электрофореза соответствующие полосы вырезали, слои геля объединяли, расплавляли, фрагменты ДНК выделяли расщеплением геля с помощью агаразы и осаждением с этанолом. Высушенный осадок растворяли в 30 мкл воды.
Соответствующий экспрессирующий вектор, в данном случае pDS56_T, расщепляли с помощью тех же самых ферментов рестрикции, которые использовали для вставки, и очищали тем же методом.
После лигирования вставки и вектора с помощью набора для быстрого лигирования ДНК (ROCHE Diagnostics GmbH, №1635...) плазмиду трансформировали в экспрессионную хозяйскую плазмиду Е.coli 392 pUBS520 (Brinkmann U. et al. (1989) Gene, 85:109-114).
Плазмидную ДНК транформантов выделяли, используя набор для выделения плазмид High PureTM (ROCHE Diagnostics GmbH, №1754777) и характеризовали расщеплением с рестриктазами BamHI и PstI и электрофорезом в агарозном геле.
Положительные трансформанты E.coli pUBS520 ExoIII хранили в глицерине при -70°С. Последовательность гена, кодирующего экзонуклеазу III, подтверждали секвенированием ДНК. Результаты показаны на фиг.1.
Клонирование и экспрессия экзонуклеазы III из Archaeoglobus fulgidus или других термофильных организмов, можно также осуществить другими методами, известными специалистам (см., например, Sambrook et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbour Lab., 1989).
Пример III
Экспрессия рекомбинантной экзонуклеазы III Afu
Трансформант из примера I культивировали в ферментере в обогащенной среде, содержащей соответствующий антибиотик. Клетки собирали при оптической плотности [A540] 5,5 с помощью центрифугирования и замораживали до использования или лизировали обработкой лизоцимом для получения сырого клеточного экстракта с активностью экзонуклеазы III из Archaeoglobus fulgidus.
Сырой экстракт с активностью экзонуклеазы III из Archaeoglobus fulgidus очищали способом, описанным в примере IV, или другими методами очистки, такими как аффинная хроматография, ионообменная хроматография или хроматография гидрофобного взаимодействия.
Пример IV
Очистка рекомбинантной экзонуклеазы III Afu
Выращивали Е.coli pUBS520 Exo III (DSM №13021) из примера I в ферментере на 10 л в среде, содержащей триптон (20 г/л), дрожжевой экстракт (10 г/л), хлорид натрия (5 г/л) и ампициллин (100 мг/л) при 37°С, индуцированной IPTG (изопропил-β-D-тиогалактопиранозидом) (0.3 мМ) при среднеэкспоненциальной фазе роста, и инкубировали дополнительные 4 часа. Примерно 45 г клеток собирали центрифугированием и хранили при -70°С. Оттаивали 2 г клеток и суспендировали в 4 мл буфера А (40 мМ Трис/HCl, рН 7.5; 0.1 мМ EDTA (этилендиаминтетрауксусная кислота); 7 мМ 2-меркаптоэтанол; 1 мМ Pefabloc SC). Клетки лизировали при перемешивании добавлением 1.2 мг лизоцима в течение 30 минут при 4°С, и добавление 4.56 мг дезоксихолата натрия в течение 10 минут при комнатной температуре сопровождалось выдержкой в течение 20 минут при 0°С. В сырой экстракт добавляли хлористый калий до концентрации 750 мМ, нагревали в течение 15 минут при 72°С и центрифугировали для удаления денатурированного белка.
Нагревание до температуры 90°С также возможно без разрушения (денатурации) экзонуклеазы III из Archaeoglobus fulgidus. Супернатант диализовали против буфера В (буфер А, содержащий 10% глицерина), доводили концентрацию хлорида магния до 10 мМ и наносили на колонку с синим трисакрилом М (SERVA, №67031) размером 1×7 см и объемом слоя 5.5 мл, уравновешенную буфером В. Колонку промывали 16.5 мл буфера В, и элюировали белок экзонуклеазы 82 мл линейного градиента от 0 до 3 М хлорида натрия в буфере В. Фракции с колонки анализировали на содержание белка экзодезоксирибонуклеазы из Archaeoglobus fulgidus с использованием PAGE (электрофореза в полиакриламидном геле) с градиентом 10-15% SDS (додецилсульфат натрия). Активные фракции, 16.5 мл, собирали, концентрировали с помощью Aquacide II (Calbiochem №17851) и диализовали против буфера С для хранения (10 мМ Трис/HCl, рН 7.9; 10 мМ 2-меркаптоэтанол; 0.1 М EDTA; 50 мМ KCl; 50% глицерин). После диализа прибавляли Thesit (полиоксиэтилен-9-лауриловый эфир) и Nonidet P40 [(октилфенокси)полиэтоксиэтанол] до конечной концентрации каждого 0.5%. Данный препарат хранили при -20°С.
Полученная экзонуклеаза III из Archaeoglobus fulgidus имела чистоту 95%, как было установлено SDS-гель-электрофорезом. Выход белка составил 50 мг на 2.3 г клеточной массы (сырой массы).
Пример V
Термостабильность рекомбинантной экзонуклеазы III из Archaeoglobus fulgidus
Термостабильность экзонуклеазы III из Archaeoglobus fulgidus, клонированной, как описано в примере II, определяли, анализируя устойчивость к тепловой денатурации. После лизиса, как описано в примере IV, 100 мкл сырого экстракта центрифугировали при 15000 об/мин в течение 10 минут в центрифуге Эппендорфа. Аликвоты супернатанта вносили в пять новых пробирок Эппендорфа. Пробирки инкубировали в течение 10 минут при различных температурах: 50°С, 60°С, 70°С, 80°С и 90°С. После центрифугирования, как описано выше, аликвоты супернатанта анализировали электрофорезом на градиенте геля 10-15% SDS-PAGE. Как показано на фиг.2, количество белка экзонуклеазы III из Archaeoglobus fulgidus после инкубации при 90°С было таким, как и в образцах, обработанных при более низких температурах. Тепловая денатурация не вызвала значительных потерь. Из этого результата можно сделать вывод о том, что время полураспада при 90°С составляет более десяти минут.
Пример VI
Активность экзонуклеазы III из Archaeoglobus fulgidus
Экзонуклеаза III катализирует поэтапное удаление мононуклеотидов с 3'-гидроксильного конца дуплексной ДНК (Rogers G.S. and Weiss В (1980) Methods Enzymol. 65:201-211). При каждом событии связывания удаляется ограниченное число нуклеотидов. Предпочтительным субстратом являются тупые или удаленные 3'-концы. Фермент неактивен на одноцепочечной ДНК, и 3'-выступающие концы более устойчивы к расщеплению. Маркер VI молекулярной массы ДНК (ROCHE Molecular Biochemicals, №1062590) состоит из расщепленной рестриктазой BglI плазмиды pBR328, смешанной с расщепленной рестриктазой HinfI плазмидой pBR328. Продукты расщепления с Hinfl имеют 3'-рецессивные концы и, как ожидается, являются предпочтительными субстратами для деградации экзонуклеазой III, продукты расщепления с BglI имеют 3'-выступающие концы с 3 выступающими основаниями и должны быть более устойчивы к расщеплению экзонуклеазой III.
Последовательные разбавления экзонуклеазы III из Archaeoglobus fulgidus из примера IV инкубировали в течение 2 часов при 72°С с 0.5 мкг маркера VI молекулярной массы ДНК (ROCHE Molecular Biochemicals, №1062590) в 25 мкл следующего инкубационного буфера: 10 мМ Трис/HCl, рН 8.0; 5 мМ хлорид магния; 1 мМ 2-меркаптоэтанол; 100 мМ хлорид натрия с парафиновым покрытием. В качестве контроля включали 10 единиц экзонуклеазы III из Е.coli (ROCHE Molecular Biochemicals, №779709). Контрольную реакцию проводили при 37°С. После прибавления 5 мкл останавливающего раствора (0.2% агароза, 60 мМ EDTA, 10 мМ Трис-HCl, рН 7.8, 10% глицерин, 0.01% бромфеноловый синий) смеси разделяли на 1% агарозном геле. Результат показан на фиг.3. Экзонуклеаза III Afu различает два различных типа субстрата. Предпочтительным субстратом является фрагмент с 3'-рецессивными концами (например, фрагмент с 1766 п.н.), а 3'-выступающие концы (например, фрагменты с 2176 п.н., 1230 п.н., 1033 п.н.) более устойчивы к деградации. При более высоком содержании белка субстрат деградируется до такой степени, как в дорожке 1, где проанализированы продукты экзонуклеазы III из Е.coil. С увеличением количества белка экзонуклеазы III Afu остается лишь малое количество ДНК-субстрата (дорожки 15-19), удерживание остающихся фрагментов может объясняться наличием в препарате ДНК-связывающих белков в качестве примесей.
Пример VII
Коррекция ошибочно спаренного праймера в PCR с экзонуклеазой III Afu
Репарационную эффективность смеси экзонуклеаза III Afu/полимераза Taq в ходе PCR исследовали с 3'-концевыми ошибочно спаренными праймерами, принцип анализа показан на фиг.4. Для амплификации методом PCR используют набор праймеров, в которых прямой праймер имеет один или два нуклеотида на 3'-конце, которые не могут образовать пару с матричной ДНК. Вырезание ошибочно спаренного праймерного конца и амплификация репарированного праймера дает продукт, который впоследствии может быть расщеплен эндонуклеазой рестрикции BsiEI, тогда как продукт, возникающий от ошибочно спаренного праймера, устойчив к расщеплению.
Использовали праймерные последовательности:
1. обратная 5'-GGT TAT CGA AAT CAG ССА CAG CG-3'
(SEQ ID NO.: 3)
Проводили PCR, используя 2.5 ед. ДНК-полимеразы Taq (ROCHE Diagnostics GmbH, №1435094), 0.25 мкг экзонуклеазы III из Archaeoglobus fulgidus из примера IV, 10 нг ДНК бактериофага λ, 0.4 мкМ каждого праймера, 200 мкМ dNTPs, 1.5 мМ хлорид магния, 50 мМ Трис-HCl, рН 9.2, 16 мМ сульфат аммония. Осуществляли PCR в объеме 50 мкл в следующих условиях:
1×94°С, 2 мин;
40×94°С, 10 сек; 60°С, 30 сек; 72°С, 1 мин;
1×72°С, 7 мин.
Функцию смеси экзонуклеаза /полимераза Taq сопоставляли с контролями, как, например, 2.5 ед. ДНК-полимеразы Taq, 0.3 ед. ДНК-полимеразы Tgo (ROCHE Diagnostics GmbH) и с 0.75 мкл высокоточной системы PCR ExpandTM (ROCHE Diagnostics GmbH, №1732641). Как следует из успешного расщепления продуктов PCR с BsiEI, экзонуклеаза III Afu проявила корректирующую активность в отношении всех описанных ошибочных спариваний с эффективностью 90-100% (фиг.5). ДНК-полимераза Taq, как ожидалось, не проявила корректирующей активности, тогда как ДНК-полимераза Tgo с ее 3'-5'-экзонуклеазной активностью также корректировала полностью. Высокоточная система PCR ExpandTM проявила 100% корректирующую активность в отношении только двух ошибочно спаренных оснований. Другие ошибочные спаривания репарировались с эффективностью приблизительно 50%.
Пример VIII
Точность смесей экзонуклеазы III Afu/ДНК-полимеразы Tag в процессе PCR
Точность смесей экзонуклеазы III Afu/ДНК-полимеразы Taq в процессе PCR определяли в анализе, основанном на амплификации, циркуляризации и трансформации pUC19-производного плазмиды pUCIQ17, содержащей функциональный аллель lac Iq (Frey В. and Suppmann В. (1995) Biochemica, 2:34-35). Получаемые при PCR мутации в lac I являются результатом дерепрессии экспрессии lac Zα и последующего образования функционального фермента β-галактозидазы, который может быть легко обнаружен на индикаторных пластинах с X-Gal (5-бром-4-хлор-3-индолил-β-D-галактопиранозидом). Частоту ошибок смесей полимераза Taq/экзонуклеаза Afu, определяемую в основанном на lac I анализе точности PCR, устанавливали в сравнении с ДНК-полимеразой Taq и высокоточной системой PCR ExpandTM (Roche Molecular Biochemicals) и ДНК-полимеразой Pwo (Roche Molecular Biochemicals) в качестве контролей.
Плазмида pUCIQ17 была линеаризована расщеплением с DraII для того, чтобы служить субстратом для амплификации с исследуемыми ферментами методом PCR.
Оба использованных праймера имели сайты ClaI на своих первичных концах.
SEQ ID NO.:8
Праймер 1: 5'-AGCTTATCGATGGCACTTTTCGGGGAAATGTGCG-3'
Seq ID NO.:9
Праймер 2: 5'-AGCTTATCGATAAGCGGATGCCGGGAGCAGACAAGC-3'
Протяженность образующегося продукта PCR составляет 3493 п.н.
Проводили PCR в конечном объеме 50 мкл в присутствии 1.5 мМ хлорида магния, 50 мМ Трис-HCl, рН 8.5 (25°С), 12.5 мМ сульфата аммония, 35 мМ хлорида калия, 200 мкМ dNTPs и 2.5 ед. полимеразы Taq и 125 нг, 175 нг, 250 нг, 375 нг и 500 нг, соответственно, экзонуклеазы III Afu.
Условия цикла были следующие:
1×денатурация матрицы в течение 2 мин при 95°С


После окончания PCR продукты осаждали PEG (полиэтиленгликолем) (Barnes W.M. (1992) Gene, 112:229), ДНК расщепляли рестриктазой Clal и очищали электрофорезом в агарозном геле. Выделенную ДНК лигировали, используя набор для быстрого лигирования ДНК (Roche Molecular Biochemicals), и продукты лигирования, трансформированные в Е.coli DH5α, помещали в планшеты TN Amp X-Gal. α-Комплементарный штамм DH5α E.coli, трансформированный образующейся плазмидой pUCIQ17 (3632 п.н.), образовывал белые (lacI+) колонии в планшетах TN (1.5% бактотриптон, 1% хлорид натрия, 1.5% агар), содержащих ампициллин (100 мкг/мл) и X-Gal (0.004% об./мас.). Мутации приводили к образованию синих колоний.
После инкубации в течение ночи при 37°С подсчитывали белые колонии. Частоту ошибок (f) на п.н. рассчитывали с использованием преобразованного уравнения, как опубликовано Keohavong and Thilly (Keohavong P. and Thilly W. (1989) PNAS USA, 86:9253):
F=-lnF/d×b п.н.,
где F означает фракцию белых колоний:
F=белые (lac+) колонии/общее число колоний;
d - означает число удвоений ДНК:
2d=ДНК на выходе/ДНК на входе;
и b означает эффективный целевой размер (1080 п.н.) гена lac I, который составляет 349 п.н. в соответствии с Provost et al. (Provost et al. (1993) Mut. Res. 288:133).
Результаты, показанные на фиг.6А и 6В, демонстрируют, что присутствие в реакционной смеси термостабильной экзонуклеазы III приводит к меньшей частоте ошибок. В зависимости от соотношения полимеразы и экзонуклеазы, частота ошибок снижается. Точность, достигнутая с наиболее оптимальной смесью полимеразы Taq/экзонуклеазы III Afu (4.44×10-6), находится в том же интервале, что и для смеси Taq/Pwo (Expand HiFi; 2.06×10-6). Оценка оптимальных буферных условий будет дополнительно улучшать качество. Соотношение между полимеразой и экзонуклеазой должно быть оптимизировано. Большие количества экзонуклеазы уменьшают выход продукта, очевидно понижая эффективность амплификации (соотношение Taq/Exo 1:10 соответствует 2.5 ед. полимеразы Taq и 500 нг экзонуклеазы III Afu).
Точность данной системы в дальнейшем может быть оптимизирована с использованием общеизвестных знаний в этой области, например изменением компонентов буфера, оптимизацией концентрации отдельных компонентов или изменением условий цикла.
Пример IX
Включение dUTP в присутствии экзонуклеазы III Afu в ходе PCR
Исследовали смесь экзонуклеазы Afu/полимеразы Taq для синтеза ДНК при полном замещении ТТР на dUTP. Сравнение включения или ТТР, или dUTP проводили в PCR, используя 2.5 ед. ДНК-полимеразы Taq в присутствии 0.125 мкг, 0.25 мкг, 0.375 мкг и 0.5 мкг экзонуклеазы III из Archaeoglobus fulgidus из примера IV, на нативной человеческой геномной ДНК в качестве матрицы и с использованием гена β-глобина в качестве мишени. Использовали следующие праймеры:
прямой: 5'-TGG TTG AAT ТСА ТАТ АТС TTA GAG GGA GGG С-3'
(SEQ ID NO.:10)
обратный: 5'-TGT GTC TGC AGA AAA CAT CAA GGG TCC CAT А-3'
(SEQ ID NO.: 11)
Проводили PCR в объеме 50 мкл при следующих условиях цикла:
1×94°С, 2 мин;
40×94°С, 10 сек; 60°С, 30 сек; 72°С, 1 мин;
1×72°С, 7 мин.
Аликвоты продуктов PCR разделяли на агарозном геле. Как показано на фиг.7, с данной системой матрица/праймер синтез ДНК в присутствии dUTP возможен вплоть до 375 нг экзонуклеазы III Afu. Включение dUTP может быть дополнительно доказано обработкой урацил-ДНК-гликозилазой (ROCHE Diagnostics GmbH, №1775367) аликвот продуктов PCR в течение 30 минут при температуре окружающей среды и последующей инкубацией в течение 5 минут при 95°С с целью расщепления полинуклеотидов в апуриновых сайтах, которые приводят к полной деградации фрагментов. Анализ продуктов реакции электрофорезом в агарозном геле показан на фиг.8.
Пример X
Влияние экзонуклеазы III Afu на длину продукта PCR
Полимераза Taq способна синтезировать продукты PCR длиной до 3 т.н. на геномных матрицах. Для того чтобы оценить способность смеси полимераза Taq/экзонуклеаза Afu синтезировать более длинные продукты, ферментную смесь анализировали на человеческой геномной ДНК в качестве матрицы с тремя парами праймеров, созданных для амплификации продуктов длиной 9.3 т.н., 12 т.н. и 15 т.н. Буферная система была выбрана из расширенной длинноматричной системы PCR (Roche Molecular Biochemicals. Cat. №1681834). Реакции проводили в объеме 50 мкл с 250 нг человеческой геномной ДНК, 220 нг каждого праймера, 350 мкМ dTNPs и 2.5 ед. полимеразы Taq и 62.5 нг экзонуклеазы Afu в условиях, приведенных в таблице 1.
отжиг при 65°С, 30 сек элонгация при 68°С, 8 мин
отжиг при 65°С, 30 сек элонгация при 68°С, 8 мин плюс элонгация цикла в 20 сек на цикл
отжиг при 62°С, 30 сек элонгация при 68°С, 12 мин
отжиг при 62°С, 30 сек элонгация при 68°С, 12 мин плюс элонгация цикла в 20 сек на цикл
Праймеры, специфичные для амплификации используемых генов. tPA:
Праймер 7а прямой: 5'-GGA AGT АСА GCT CAG AGT TCT GCA GCA CCC CTG С-3' (SEQ ID NO.:12)
Праймер 14а обратный: 5'-САА AGT CAT GCG GCC АТС GTT CAG АСА САС С-3' (SEQ ID NO.:13)
Праймер 1 прямой: 5'-ССТ ТСА CTG TCT GCC ТАА СТС СТТ CGT GTG ТСС С-3' (SEQ ID NO.:14)
Праймер 2 обратный: 5'-ACTGTGCTTCCTGACCCATGGCAGAAGCGCCTTC-3' (SEQ ID NO.:15)
Праймер 3 обратный: 5'-CCTTCTAGAGTCAACTCTAGATGTGGACTTAGAG-3' (SEQ ID NO.:16)
Как показано на фиг.9, со смесью полимераза Taq/экзонуклеаза Afu можно синтезировать продукты длиной по меньшей мере 15 т.н.
Пример XI
Термостабильная экзонуклеаза III может быть заменена мутантом полимеразы со сниженной полимеразной активностью, но с повышенной 3'-экзонуклеазной активностью
ДНК-полимераза из Thermococcuss aggregans (Taq), описанная Niehaus F., Frey В. and Antranikian G в международной заявке №97/35988 или Gene (1997) 204 (1-2), 153-8, с заменой аминокислоты в положении 385, где тирозин заменен аспарагином (Boehlke et al., подписано для публикации и Международная заявка на патент №00105155.6), проявляет всего 6.4% полимеразной активности, но 205% экзонуклеазной активности ДНК-полимеразы дикого типа. Данный фермент был использован для демонстрации того, что изобретение не ограничено ферментами типа экзонуклеазы III, но включает в себя также и другие типы ферментов, привносящие 3'-экзонуклеазную активность.
Реакции проводили в объеме 50 мкл с 200 нг человеческой геномной ДНК, 200 мкМ dNTP, 220 нг каждого праймера и расширенного HiFi буфера, включающего в себя Mg++ для реакций 1-4, или расширенного длинноматричного буфера 1 для реакций 5-8 (фиг.10). Для того чтобы амплифицировать фрагмент гена tPA в 4.8 т.н., в реакциях 1-4 использовали праймер tPA 7a прямой (5'-GGAAGTACAGCTCAGAGTTCTGCAGCACCCCTGC-3', SEQ ID NO.:12) и tPA 10а обратный (5'-GAT GCG AAA CTG AGG CTG GCT GTA CTG ТСТ С-3', SEQ ID NO.:17). Для того чтобы амплифицировать фрагмент гена tPA в 9.3 т.н., в реакциях 5-8 использовали праймер tPA 7а прямой и tPA 14а обратный (5'-САА AGT CAT GCG GCC ATC GTT CAG АСА САС С-3', SEQ ID NO.:13). Прибавляли 2.5 ед. полимеразы Taq в реакции 1, 2, 4, 5, 6 и 8, но не в реакции 3 и 7, которые использовали как отрицательные контроли. Прибавляли 11 нг мутанта полимеразы Taq в реакции 2, 3, 6 и 7 и 150 нг экзонуклеазы III Afu в реакции 4 и 8.
Программы циклов, использованные для реакций 1-4:
1×94°С, 2 мин
10×94°С, 10 сек
62°С, 30 сек
68°С, 4 мин
20×94°С, 10 сек
62°С, 30 сек
68°С, 4 мин плюс элонгация цикла в 20 сек на цикл
1×68°С, 7 мин
для реакций 5-8:
1×94°С, 2 мин
10×94°С, 10 сек
65°С, 30 сек
68°С, 8 мин
20×94°С, 10 сек
65°С, 30 сек
68°С, 8 мин плюс элонгация цикла в 20 сек на цикл
1×68°С, 7 мин
Продукты PCR анализировали на 1% агарозном геле, содержащем бромистый этидий (фиг.10). Из данных следует, что полимераза Taq способна амплифицировать фрагмент в 4.8 т.н., но с низким выходом. Сочетание полимеразы Taq с мутантом полимеразы Taq или с экзонуклеазой III Afu приводит к заметному увеличению выхода продукта. Сам по себе мутантный фермент полимераза Taq не способен синтезировать этот продукт.
Подобные результаты получены с системой 9.3 т.н. При использовании одной полимеразы Taq никакой продукт не определяется. В комбинации с мутантом полимеразы Taq или экзонуклеазой III Afu ожидаемый продукт PCR получают с высоким выходом.
Данные результаты показывают, что полимераза Taq не способна амплифицировать фрагменты ДНК в несколько т.н. от геномной ДНК, и поддерживают гипотезу Barnes (Barnes W.M. (1994) Proc. Natl. Acad. Sci. USA, 91:2216-2220) о том, что ограничение длины при амплификации методом PCR вызвано низкой эффективностью удлинения в сайтах включения ошибочно спаренных пар оснований. После удаления ошибочно спаренного нуклеотида на праймерном конце полимераза Taq способна снова участвовать в синтезе ДНК. Законченная нуклеотидная цепь как полноразмерный продукт может затем служить матрицей для связывания с праймером в последующих циклах.
Пример XII
Экзонуклеаза III Afu неактивна на линейной одноцепочечной ДНК
Реакции проводили в объеме 50 мкл с 270 нг экзонуклеазы III Afu, 5 мкг 49-мерного олигонуклеотида в расширенном HiFi PCR буфере с хлоридом магния и инкубировали в течение 0, 1, 2, 3, 4 и 5 ч при 65°С. После добавления 10 мкл раствора протеиназы К (20 мг/мл) образцы инкубировали в течение 20 минут при 37°С. Продукты реакции анализировали на 3.5% агарозном геле, содержащем бромистый этидий.
Результаты представлены на фиг.11. Из них следует, что нуклеиновая кислота имеет один и тот же размер во всех дорожках. Продукт, полученный после инкубации вплоть до 5 часов (дорожка 6) с экзонуклеазой III Afu, имеет тот же размер, что и контроли (дорожки 1, 7 и 8). Не наблюдалось ни значительного снижения интенсивности полноразмерного олигонуклеотида, ни размытости, происходящей от продуктов деградации.
Пример XIII
Сравнение экзонуклеазы III Afu с термостабильной полимеразой типа В по активности деградации праймера
Сообщается, что термостабильные полимеразы типа В обладают одно- и двухцепочечной нуклеазной активностью (Kong Н. et al. (1993) J. Biol. Chem. 268:1965-1975). Подобная активность способна разрушать молекулы праймеров, независимо от того, гибридизованы ли они с матрицей или являются одноцепочечными. Замена в реакционной смеси термостабильной полимеразы типа В на термостабильную экзонуклеазу могла бы иметь преимущество в отношении стабильности одноцепочечного праймера или других нуклеиновых кислот, присутствующих в реакционной смеси.
Для того чтобы исследовать активность деградации праймера, реакционные смеси без матричной ДНК инкубировали в течение 1 часа при 72°С, затем прибавляли ДНК и проводили PCR. Результаты сопоставляли с реакциями, содержащими полимеразу TgO в качестве примера термостабильной полимеразы типа В (Angerer В. et al. Международная заявка №98/14590). В качестве контроля использовали те же смеси без предварительной инкубации. Результаты суммированы в таблице 2.
В качестве мишени для амплификации был выбран фрагмент гена р53, праймерами служили: p53I GTC ССА AGC ААТ GGA TGA Т-3' (SEQ ID NO.:18) и p53II 5'-TGGAAACTTTCCACTTGAT-3'(SEQ ID NO.:19). Проводили PCR в объеме 50 мкл. Реакции №1-10 содержали 200 нг человеческой геномной ДНК, 40 пмоль каждого праймера, 10 мМ Трис-HCl, рН 8.5, 17.5 мМ сульфат аммония, 1.25 мМ хлорид магния, 0.5% Твин, 2.5% ДМСО, 250 мкг/мл BSA (бычьего сывороточного альбумина) и 1 ед. (реакции №1, 3, 5, 7 и 9) или 1.5 ед. (реакции №2, 4, 6, 8 и 10) полимеразы Tgo и 200 мкМ dNTPs.
Реакции №11-16 содержали 2.5 ед. полимеразы Taq, расширенного HiFi буфера с Mg++, 40 пмоль праймера, 200 мкМ dNTPs, 100 нг человеческой геномной ДНК. Реакции №12 и 15 содержали 37.5 нг экзонуклеазы III Afu, реакции №13 и 16 содержали 75 нг экзонуклеазы III Afu.
Как указано в таблице 2, реакции 1, 2, 5, 6 и 11-13 инкубировали в течение 1 часа при 72°С в отсутствие матричной ДНК. Матричную ДНК прибавляли перед началом PCR. Реакции 5, 6, 9 и 10 предварительно инкубировали в отсутствие нуклеотидов, реакции 9 и 10 дополняли добавочными 40 пмоль праймера после этапа преинкубации. Из-за 5'-экзонуклеазной активности полимеразы Taq фермент добавляли в реакции 11-13 после преинкубации.
Условия PCR:
1×94°С, 2 мин
35×94°С, 10 сек
55°С, 30 сек
72°С, 4 мин
1×72°С, 10 мин
Продукты реакции анализировали на агарозном геле и окрашивали бромистым этидием (фиг.12).
Когда полимеразу Taq инкубировали с праймером в отсутствие матричной ДНК (реакции 1, 2, 5 и 6) и сравнивали с соответствующими реакциями без преинкубации (3, 4, 7 и 8), наблюдалось четкое отличие. Преинкубация приводит к сильно заниженному продукту PCR, очевидно воздействуя по меньшей мере на один существенный компонент, наиболее вероятно, праймер PCR. Дополнительное добавление 40 пмоль праймера PCR (реакции 9 и 10) после преинкубации приводит к сильным сигналам с интенсивностями, сопоставимыми с контрольной реакцией без преинкубации. Отсюда следует, что полимераза Tgo, термостабильная полимераза типа В, разрушает праймер PCR в отсутствие матрицы независимо от того, присутствуют ли dNTPs или нет.
Продукты PCR, полученные в реакциях 12 и 13, в которых праймеры были проинкубированы с экзонуклеазой III Afu до добавления матричной ДНК и полимеразы Taq, дают такие же полосы, как и полученные в реакциях 15 и 16, в которых этап преинкубации не проводили. На основании подобных сильных полос интенсивностей можно заключить, что происходит слабая или не происходит совсем деградация праймера и что одноцепочечные олигонуклеотиды являются плохими субстратами экзонуклеазы III Afu. На основании сильных полос интенсивностей или повышенных выходов продуктов PCR можно заключить, что фермент повышает точность процесса амплификации.











Изобретение относится к области биохимии и молекулярной биологии и может быть использовано в аналитических системах любого назначения, основанных на ПЦР-амплификации ДНК. Предложена композиция для осуществления реакции амплификации, состоящая из двух термостабильных ферментов, один из которых (основной компонент) представляет собой обычно используемую в ПЦР ДНК-полимеразу, а второй (минорный компонент) - 3'-экзонуклеазу из Archeoglobus fulgidus (экзонуклеазу III Afu). Способ ПЦР-амплификации, осуществляемый с использованием данной композиции, отличается большей точностью копирования, чем амплификация, проводимая только в присутствии ДНК-полимеразы, и, соответственно, повышает достоверность результатов тест-систем с его использованием. 2 н. и 8 з.п. ф-лы, 13 ил., 2 табл.
Конвенционный приоритет установлен от 28.09.1999 в соответствии с датой подачи заявки 99119268.3 в Европейское Патентное ведомство.
| EMBL SEQUENCE DATABASE, Ac | |||
| СПОСОБ СВЕРТЫВАНИЯ ЗОЛЕЙ И ТОНКИХ СУСПЕНЗИЙ СУЛЬФИДОВ ТЯЖЕЛЫХ МЕТАЛЛОВ | 1924 |
|
SU1064A1 |
| WO 9913060 A, 18.03.1999 | |||
| Коллектор текучей среды высокого давления | 1980 |
|
SU870832A1 |

