Область техники
[1] Настоящее изобретение относится к способу детекции множества нуклеиновых кислот-мишеней, с использованием множества температур детекции.
Уровень техники
[2] Для детекции нуклеиновых кислот-мишеней, широко используют способы детекции в реальном времени, способные к детекции нуклеиновых кислот-мишеней, с мониторированием амплификации мишени способом с детекцией в реальном времени. В способах детекции в реальном времени, как правило, используют меченые зонды или праймеры, специфически гибридизующиеся с нуклеиновыми кислотами-мишенями.
[3] Примеры способов с использованием гибридизации между мечеными зондами и нуклеиновыми кислотами-мишенями включают способ молекулярного маяка с использованием зондов с двойной меткой со структурой шпильки (Tyagi et al., Nature Biotechnology v.14 MARCH 1996), способ HyBeacon (French D J et al., Mol. Cell Probes, 15(6):363-374 (2001)), способ гибридизации зондов с использованием двух зондов, где каждый является меченным как донор и акцептор (Bernard et al., 147-148 Clin. Chem. 2000; 46), и способ Lux с использованием олигонуклеотидов с одиночной меткой (Патент США No. 7537886). Способ TaqMan (Патенты США No. 5210015 и 5538848) с использованием расщепления зонда с двойной меткой посредством 5′-нуклеазной активности ДНК полимеразы также широко используют в данной области техники.
[4] Примеры способов с использованием меченых праймеров включают способ с использованием праймера Sunrise (Nazarenko et al., 2516-2521 Nucleic Acids Research, 1997, v.25 no.12, и Патент США No. 6117635), способ с использованием праймера скорпион (Whitcombe et al., 804-807, Nature Biotechnology v.17 AUGUST 1999 и Патент США No. 6326145) и способ с использованием праймера TSG (WO 2011/078441).
[5] В качестве альтернативных способов, предложены способы детекции в реальном времени с использованием дуплексов, сформированных в зависимости от присутствия нуклеиновых кислот-мишеней: анализ Invader (Патенты США No. 5691142, 6358691 и 6194149), способ PTOCE (расщепления и удлинения PTO) (WO 2012/096523), способ PCE-SH (гибридизации сигнального олигонуклеотида, зависящей от расщепления и удлинения PTO) (WO 2013/115442) и способ PCE-NH (отсутствия гибридизации, зависящего от расщепления и удлинения PTO) (PCT/KR2013/012312).
[6] Поскольку вышеописанные общепринятые способы детекции в реальном времени могут детектировать только одиночную нуклеиновую кислоту-мишень с одиночной меткой, количество нуклеиновых кислот-мишеней, которые можно одновременно детектировать в одной реакции, ограничено количеством меток, которые можно использовать (например, 5 или менее).
[7] Хотя анализ плавления можно использовать для детекции множества нуклеиновых кислот-мишеней с использованием одной метки, он имеет недостаток более длительного времени выполнения, чем способы с детекцией в реальном времени.
[8] В то же время, в последние годы, предложены способы, которые могут детектировать множество нуклеиновых кислот-мишеней в реальном времени с использованием одного типа метки, с использованием детекции сигнала при различных температурах детекции (WO 2015/147370, WO 2015/147377, WO 2015/147382, WO 2015/147412). Однако, среди таких различных температур детекции, при относительно более низкой температуре детекции, эти способы также позволяют нуклеиновым кислотам-мишеням, имеющим относительно более высокую температуру детекции, поддаваться детекции вместе с нуклеиновыми кислотами-мишенями, имеющими относительно более низкую температуру детекции, таким образом, делая необходимым способ получения различия между сигналами, детектированными при различных температурах детекции (например, различия между сигналом, детектированным при относительно более низкой температуре детекции, и сигналом, детектированным при относительно более высокой температуре детекции).
[9] Соответственно, существует потребность в разработке нового метода или способа для детекции множества нуклеиновых кислот-мишеней, имеющего сильно улучшенное удобство и высокую эффективность.
[10]
[11] На всем протяжении настоящей заявки, приведены ссылки на различные патенты и публикации, и цитаты представлены в скобках. Полное содержание этих патентов и публикаций, таким образом, приведено в настоящем описании в качестве ссылки для более полного описания настоящего изобретения и существующего уровня техники в области, к которой относится настоящее изобретение.
Описание изобретения
Техническая проблема
[12] Авторы настоящего изобретения прикладывали усилия для разработки способа детекции множества нуклеиновых кислот-мишеней посредством использования одного типа метки и одного типа детектора в одном реакционном сосуде. В результате, авторы настоящего изобретения подтвердили, что, с использованием множества температур детекции, посредством коррекции температур детекции и механизмов образования сигнала таким образом, что только один сигнал, показательный для присутствия одной последовательности-мишени нуклеиновой кислоты, обеспечивают при одной температуре детекции, является возможной детекция множества нуклеиновых кислот-мишеней способом с детекцией в реальном времени посредством использования одного типа метки и одного типа детектора в одном реакционном сосуде, с улучшенным удобством и с высокой экономической эффективностью и эффективностью.
[13] Таким образом, целью настоящего изобретения является предоставление способа детекции n нуклеиновых кислот-мишеней в образце.
[14] Другой целью настоящего изобретения является предоставление набора для детекции n нуклеиновых кислот-мишеней в образце.
Решение проблемы
[15] В соответствии с одним аспектом, настоящее изобретение относится к способу детекции n нуклеиновых кислот-мишеней в образце, включающему: (a) детекцию сигналов при n температурах детекции, с инкубацией в то же время с n композициями для детекции нуклеиновых кислот-мишеней, образца, как подозревают, содержащего по меньшей мере одну из n нуклеиновых кислот-мишеней, в реакционном сосуде; где n представляет собой целое число 2 или более, где инкубация включает множество циклов, и детекцию сигналов проводят по меньшей мере в одном из множества циклов, где каждая из n композиций для детекции нуклеиновых кислот-мишеней обеспечивает изменение сигнала при соответствующей температуре детекции среди n температур детекции в присутствии соответствующей нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие соответствующей нуклеиновой кислоты-мишени, где композиция для детекции iй нуклеиновой кислоты-мишени среди n композиций для детекции нуклеиновых кислот-мишеней обеспечивает изменение сигнала при iй температуре детекции среди n температур детекции и обеспечивает постоянный сигнал при других температурах детекции в присутствии iй нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие iй нуклеиновой кислоты-мишени, и где i представляет собой целое число от 1 до n, и iя температура детекции является более низкой, чем (i+1)я температура детекции; и (b) определение присутствия n нуклеиновых кислот-мишеней из сигналов, детектированных на стадии (a), где присутствие iй нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при iй температуре детекции.
[16] В соответствии с вариантом осуществления настоящего изобретения, в температурном диапазоне, покрывающем все из n температур детекции, композиция для детекции iй нуклеиновой кислоты-мишени имеет температурный диапазон изменения сигнала (SChTR), в котором сигнал изменяется в зависимости от присутствия iй нуклеиновой кислоты-мишени, и температурный диапазон постоянного сигнала (SCoTR), в котором сигнал является постоянным, даже в присутствии iй нуклеиновой кислоты-мишени.
[17] В соответствии с вариантом осуществления настоящего изобретения, композиция для детекции iй нуклеиновой кислоты-мишени имеет один или два температурных диапазона постоянного сигнала.
[18] В соответствии с вариантом осуществления настоящего изобретения, композиция для детекции iй нуклеиновой кислоты-мишени представляет собой любую из: (i) композиции с нижним изменением сигнала (UnderSC), имеющей характеристику, что температурный диапазон изменения сигнала является более низким, чем температурный диапазон постоянного сигнала; (ii) композиции с верхним изменением сигнала (OverSC), имеющей характеристику, что температурный диапазон изменения сигнала является более высоким, чем температурный диапазон постоянного сигнала; и (iii) композиции с промежуточным изменением сигнала (InterSC), имеющей характеристику, что температурный диапазон изменения сигнала является более высоким, чем один из двух температурных диапазонов постоянного сигнала, и более низким, чем другой из двух температурных диапазонов постоянного сигнала.
[19] В соответствии с вариантом осуществления настоящего изобретения, iя температура детекции выбрана в рамках температурного диапазона изменения сигнала композиции для детекции iй нуклеиновой кислоты-мишени, где iя температура детекции не включена в температурные диапазоны изменения сигнала композиций для детекции других нуклеиновых кислот-мишеней.
[20] В соответствии с вариантом осуществления настоящего изобретения, температурный диапазон изменения сигнала композиции для детекции iй нуклеиновой кислоты-мишени частично перекрывается с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей соседнюю температуру детекции, и не перекрывается с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей температуру детекции, не являющуюся соседней с ней.
[21] В соответствии с вариантом осуществления настоящего изобретения, когда n представляет собой 2, композиция для детекции первой нуклеиновой кислоты-мишени представляет собой композицию UnderSC или композицию InterSC, и композиция для детекции второй нуклеиновой кислоты-мишени представляет собой композицию InterSC или композицию OverSC.
[22] В соответствии с вариантом осуществления настоящего изобретения, когда n представляет собой 3 или более, композиция для детекции первой нуклеиновой кислоты-мишени представляет собой композицию UnderSC или композицию InterSC, композиция для детекции nй нуклеиновой кислоты-мишени представляет собой композицию InterSC или композицию OverSC, и каждая из композиций для детекции нуклеиновых кислот-мишеней, отличных от первой нуклеиновой кислоты-мишени и nй нуклеиновой кислоты-мишени, представляет собой композицию InterSC.
[23] В соответствии с вариантом осуществления настоящего изобретения, композиция для детекции iй нуклеиновой кислоты-мишени содержит метку, которая обеспечивает сигнал, зависимый от присутствия iй нуклеиновой кислоты-мишени.
[24] В соответствии с вариантом осуществления настоящего изобретения, метка является связанной с олигонуклеотидом или встраивается в олигонуклеотид в ходе инкубации.
[25] В соответствии с вариантом осуществления настоящего изобретения, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала.
[26] В соответствии с вариантом осуществления настоящего изобретения, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала, и композиция для детекции iй нуклеиновой кислоты-мишени обеспечивает сигнал от метки, когда дуплекс, обеспечивающий изменение сигнала, находится в связанной форме.
[27] В соответствии с вариантом осуществления настоящего изобретения, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала, и композиция для детекции iй нуклеиновой кислоты-мишени обеспечивает сигнал от метки, когда дуплекс, обеспечивающий изменение сигнала, находится в диссоциированной форме.
[28] В соответствии с вариантом осуществления настоящего изобретения, дуплекс, обеспечивающий изменение сигнала, был изначально включен в композицию для детекции iй нуклеиновой кислоты-мишени.
[29] В соответствии с вариантом осуществления настоящего изобретения, дуплекс, обеспечивающий изменение сигнала, образуется посредством гибридизации между связанным с меткой олигонуклеотидом и олигонуклеотидом, способным к гибридизации со связанным с меткой олигонуклеотидом.
[30] В соответствии с вариантом осуществления настоящего изобретения, дуплекс, обеспечивающий изменение сигнала, образуется посредством инкубации.
[31] В соответствии с вариантом осуществления настоящего изобретения, дуплекс, обеспечивающий изменение сигнала, образуется посредством гибридизации между связанным с меткой олигонуклеотидом и нуклеиновой кислотой-мишенью.
[32] В соответствии с вариантом осуществления настоящего изобретения, дуплекс, обеспечивающий изменение сигнала, образуется посредством реакции расщепления, зависимой от присутствия нуклеиновой кислоты-мишени.
[33] В соответствии с вариантом осуществления настоящего изобретения, композиция для детекции нуклеиновой кислоты-мишени содержит метящий олигонуклеотид, который гибридизуется с нуклеиновой кислотой-мишенью, и реакция расщепления, зависимая от присутствия нуклеиновой кислоты-мишени, включает расщепление метящего олигонуклеотида.
[34] В соответствии с вариантом осуществления настоящего изобретения, дуплекс, обеспечивающий изменение сигнала, представляет собой дуплекс одиночного типирования или дуплексы множественного типирования.
[35] В соответствии с вариантом осуществления настоящего изобретения, когда дуплекс, обеспечивающий изменение сигнала, представляет собой дуплекс одиночного типирования, количество дуплекса одиночного типирования изменяется в зависимости от присутствия нуклеиновой кислоты-мишени, таким образом, изменяя сигнал.
[36] В соответствии с вариантом осуществления настоящего изобретения, когда дуплекс, обеспечивающий изменение сигнала, представляет собой дуплексы множественного типирования, соотношение количеств между дуплексами множественного типирования изменяется в зависимости от присутствия нуклеиновой кислоты-мишени, таким образом, изменяя сигнал.
[37] В соответствии с вариантом осуществления настоящего изобретения, когда дуплекс представляет собой дуплексы множественного типирования, значения Tm дуплексов отличаются друг от друга.
[38] В соответствии с вариантом осуществления настоящего изобретения, по меньшей мере два из дуплексов множественного типирования содержат одну и ту же одиночную цепь.
[39] В соответствии с вариантом осуществления настоящего изобретения, дуплекс, обеспечивающий изменение сигнала, содержит метку.
[40] В соответствии с вариантом осуществления настоящего изобретения, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала, и температурный диапазон изменения сигнала композиции для детекции iй нуклеиновой кислоты-мишени определяют, в зависимости от длины и/или последовательности дуплекса.
[41] В соответствии с вариантом осуществления настоящего изобретения, детекцию сигналов проводят по меньшей мере в двух из множества циклов.
[42] В соответствии с вариантом осуществления настоящего изобретения, изменение сигнала измеряют с использованием сигналов, детектированных по меньшей мере в двух из множества циклов.
[43] В соответствии с вариантом осуществления настоящего изобретения, изменение сигнала при iй температуре детекции измеряют с использованием сигнала, детектированного по меньшей мере в одном из множества циклов, и контрольного значения сигнала.
[44] В соответствии с вариантом осуществления настоящего изобретения, контрольное значение сигнала получают в результате реакции в отсутствие iй нуклеиновой кислоты-мишени.
[45] В соответствии с вариантом осуществления настоящего изобретения, детекцию сигнала при каждой из n температур детекции проводят с использованием одного типа детектора.
[46] В соответствии с вариантом осуществления настоящего изобретения, сигналы, детектированные при n температурах детекции, не являются отличимыми друг от друга посредством одного типа детектора.
[47] В соответствии с вариантом осуществления настоящего изобретения, инкубация включает реакцию амплификации нуклеиновой кислоты.
[48] В соответствии с вариантом осуществления настоящего изобретения, реакция амплификации нуклеиновой кислоты представляет собой полимеразную цепную реакцию (ПЦР).
[49] В соответствии с другим аспектом, настоящее изобретение относится к способу детекции двух нуклеиновых кислот-мишеней в образце, включающему:
[50] (a) детекцию сигналов при первой температуре детекции и второй температуре детекции, с инкубацией в то же время образца, как подозревают, содержащего по меньшей мере одну из двух нуклеиновых кислот-мишеней, с композицией для детекции первой нуклеиновой кислоты-мишени и композицией для детекции второй нуклеиновой кислоты-мишени в реакционном сосуде; где инкубация включает множество циклов, и детекцию сигналов проводят по меньшей мере в одном из множества циклов, где композиция для детекции первой нуклеиновой кислоты-мишени обеспечивает изменение сигнала при первой температуре детекции и обеспечивает постоянный сигнал при второй температуре детекции в присутствии первой нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие первой нуклеиновой кислоты-мишени; и композиция для детекции второй нуклеиновой кислоты- мишени обеспечивает изменение сигнала при второй температуре детекции и обеспечивает постоянный сигнал при первой температуре детекции в присутствии второй нуклеиновой кислоты-мишени, изменение сигнала указывает на присутствие второй нуклеиновой кислоты-мишени, и где первая температура детекции является более низкой, чем вторая температура детекции, и (b) определение присутствия двух нуклеиновых кислот-мишеней из сигналов, детектированных на стадии (a); где присутствие первой нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при первой температуре детекции, и присутствие второй нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при второй температуре детекции.
[51] В соответствии с другим аспектом, настоящее изобретение относится к способу детекции трех нуклеиновых кислот-мишеней в образце, включающему:
[52] (a) детекцию сигналов при первой температуре детекции, второй температуре детекции и третьей температуре детекции, с инкубацией в то же время образца, как подозревают, содержащего по меньшей мере одну из трех нуклеиновых кислот-мишеней, с композицией для детекции первой нуклеиновой кислоты-мишени, композицией для детекции второй нуклеиновой кислоты-мишени и композицией для детекции третьей нуклеиновой кислоты-мишени в реакционном сосуде; где инкубация включает множество циклов, и детекцию сигналов проводят по меньшей мере в одном из множества циклов, где композиция для детекции первой нуклеиновой кислоты-мишени обеспечивает изменение сигнала при первой температуре детекции и обеспечивает постоянный сигнал при второй температуре детекции и третьей температуре детекции в присутствии первой нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие первой нуклеиновой кислоты-мишени; композиция для детекции второй нуклеиновой кислоты-мишени обеспечивает изменение сигнала при второй температуре детекции и обеспечивает постоянный сигнал при первой температуре детекции и третьей температуре детекции в присутствии второй нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие второй нуклеиновой кислоты-мишени; и композиция для детекции третьей нуклеиновой кислоты-мишени обеспечивает изменение сигнала при третьей температуре детекции и обеспечивает постоянный сигнал при первой температуре детекции и второй температуре детекции в присутствии третьей нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие третьей нуклеиновой кислоты-мишени, и где первая температура детекции является более низкой, чем вторая температура детекции, и вторая температура детекции является более низкой, чем третья температура детекции, и (b) определение присутствия трех нуклеиновых кислот-мишеней из сигналов, детектированных на стадии (a), где присутствие первой нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при первой температуре детекции, присутствие второй нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при второй температуре детекции, и присутствие третьей нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при третьей температуре детекции.
[53] В соответствии с другим аспектом, настоящее изобретение относится к набору, содержащему n композиций для детекции n нуклеиновых кислот-мишеней в образце, где n представляет собой целое число 2 или более, где каждая из n композиций для детекции n нуклеиновых кислот-мишеней обеспечивает изменение сигнала при соответствующей температуре детекции среди n температур детекции, где изменение сигнала указывает на присутствие соответствующей нуклеиновой кислоты-мишени, где композиция для детекции iй нуклеиновой кислоты-мишени среди n нуклеиновых кислот-мишеней обеспечивает изменение сигнала при iй температуре детекции среди n температур детекции и обеспечивает постоянный сигнал при других температурах детекции в присутствии iй нуклеиновой кислоты-мишени, и где i представляет собой целое число от 1 до n, и iя температура детекции является более низкой, чем (i+1)я температура детекции.
Обеспечивающие преимущество эффекты изобретения
[54] Признаки и преимущества настоящего изобретения обобщены следующим образом:
[55] (a) Настоящее изобретение относится к способу детекции множества нуклеиновых кислот-мишеней посредством только одного типа метки в одном реакционном сосуде посредством использования множества температур детекции, отличающемуся обеспечением изменения сигнала, зависимого от присутствия соответствующей нуклеиновой кислоты-мишени, при соответствующей температуре детекции каждой из нуклеиновых кислот-мишеней.
[56] (b) Интересно, что авторы настоящего изобретения обнаружили, что различные механизмы образования сигналов для детекции нуклеиновых кислот-мишеней, хорошо известные в данной области, включают температурный диапазон, в котором сигнал изменяется в зависимости от присутствия нуклеиновой кислоты-мишени (то есть, температурный диапазон изменения сигнала), и температурный диапазон, в котором сигнал не изменяется, даже в присутствии нуклеиновой кислоты-мишени (то есть, температурный диапазон постоянного сигнала), и могут быть разделены по категориям на три типа, в соответствии с количеством и/или порядком этих температурных диапазонов изменения сигнала и температурных диапазонов постоянного сигнала. Авторы настоящего изобретения также обнаружили, что соответствующая комбинация этих трех типов механизмов образования сигнала является полезной в детекции множества нуклеиновых кислот-мишеней.
[57] (c) В частности, когда механизм образования сигнала, включающий один температурный диапазон изменения сигнала и два температурных диапазона постоянного сигнала (то есть, механизм образования сигнала, приемлемый для композиции InterSC для детекции нуклеиновой кислоты-мишени), применяют по настоящему изобретению, количество нуклеиновых кислот-мишеней, поддающихся детекции посредством способа в соответствии с настоящим изобретением, далее увеличивается.
[58] (d) Посредством принятия одного или нескольких из вышеописанных трех типов механизмов образования сигнала, и посредством коррекции температурных диапазонов изменения сигнала из одного или нескольких механизмов образования сигнала таким образом, что только сигнал, указывающий на присутствие одной нуклеиновой кислоты-мишени, обеспечивают при каждой температуре детекции, настоящее изобретение имеет те преимущества, что присутствие конкретной нуклеиновой кислоты-мишени можно определять только посредством изменения сигнала, измеренного при конкретной температуре детекции, без необходимости учитывать изменения сигнала при других температурах детекции (например, температурах детекции, отличных от конкретной температуры детекции, то есть, температурах детекции, показывающих изменения сигнала, указывающие на присутствие других нуклеиновых кислот-мишеней). В соответствии с настоящим изобретением, в способе в соответствии с настоящим изобретением используют n композиций для детекции n различных нуклеиновых кислот-мишеней, где каждая композиция соответствует каждой из нуклеиновых кислот-мишеней. В одном варианте осуществления, для каждой из n композиций для детекции нуклеиновых кислот-мишеней принимают один из трех типов механизмов образования сигнала, так что изменение сигнала, указывающее на присутствие соответствующей нуклеиновой кислоты-мишени, обеспечивают только при соответствующей температуре детекции среди n температур детекции.
[59] (e) Способ в соответствии с настоящим изобретением позволяет детекцию множества нуклеиновых кислот-мишеней в реальном времени, посредством использования только одного типа метки в одном реакционном сосуде. Общепринятые способы с использованием одного типа метки подвергают анализу плавления после амплификации мишени, чтобы таким образом детектировать множество нуклеиновых кислот-мишеней. В отличие от этого, способ по настоящему изобретению не требует анализа кривой плавления после амплификации мишени, даже с использованием одного типа метки, и таким образом, может заметно уменьшать время анализа.
Краткое описание чертежей
[60] На ФИГ. 1a - 1h показаны примеры механизмов образования сигнала, приемлемых для композиции UnderSC. На каждом чертеже, «(a)» представляет собой до инкубации в присутствии или в отсутствие нуклеиновой кислоты-мишени, или после инкубации в отсутствие нуклеиновой кислоты-мишени, и «(b)» представляет собой после инкубации в присутствии нуклеиновой кислоты-мишени.
[61] На ФИГ. 2a - 2c показаны примеры механизмов образования сигнала, приемлемых для композиции OverSC. На каждом чертеже, «(a)» представляет собой до инкубации в присутствии или в отсутствие нуклеиновой кислоты-мишени, или после инкубации в отсутствие нуклеиновой кислоты-мишени, и «(b)» представляет собой после инкубации в присутствии нуклеиновой кислоты-мишени.
[62] На ФИГ. 3a - 3b показаны примеры механизмов образования сигнала, приемлемых для композиции InterSC. На каждом чертеже, «(a)» представляет собой до инкубации в присутствии или в отсутствие нуклеиновой кислоты-мишени, или после инкубации в отсутствие нуклеиновой кислоты-мишени, и «(b)» представляет собой после инкубации в присутствии нуклеиновой кислоты-мишени.
[63] На ФИГ. 4 показаны температурные диапазоны, каждый из которых является селективным в качестве температуры детекции, в случае, когда SChTR не перекрываются друг с другом, или когда SChTR частично перекрываются друг с другом.
[64] На ФИГ. 5 схематически показан механизм образования сигнала для способа 1 на основе PTOCE, который является приемлемым для композиции UnderSC, используемой в примерах.
[65] Как изображено на чертеже, CTO имеет репортерную молекулу и молекулу гасителя в своей служащей матрицей части.
[66] На ФИГ. 6 схематически показан механизм образования сигнала для способа 2 на основе PTOCE , который является приемлемым для композиции InterSC, используемой в примерах.
[67] Как изображено на чертеже, PTO имеет молекулу гасителя в своей 5′-метящей части, и CTO имеет репортерную молекулу в своей связывающей части.
[68] На ФИГ. 7 схематически показан механизм образования сигнала для способа двойного гашения, который является приемлемым для композиции OverSC, используемой в примерах.
[69] Как изображено на чертеже, PTO имеет первую молекулу гасителя и репортерную молекулу, и CQO имеет вторую молекулу гасителя.
[70] На ФИГ. 8 схематически показаны механизмы образования сигнала для двух композиций, композиции UnderSC и композиции InterSC, используемых в комбинации 1 в примерах.
[71] На ФИГ. 9 схематически показаны механизмы образования сигнала для двух композиций, композиции UnderSC и композиции OverSC, используемых в комбинации 2 в примерах.
[72] На ФИГ. 10 схематически показаны механизмы образования сигнала для двух композиций, композиции InterSC и композиции InterSC, используемых в комбинации 3 в примерах.
[73] На ФИГ. 11 схематически показаны механизмы образования сигнала для двух композиций, композиции InterSC и композиции OverSC, используемых в комбинации 4 в примерах.
[74] На ФИГ. 12 схематически показаны механизмы образования сигнала для трех композиций, композиции UnderSC, композиции InterSC и композиции OverSC, используемых в комбинации 5 в примерах.
[75] На ФИГ. 13 показаны результаты ПЦР с детекцией в реальном времени для комбинации 1 в примерах.
[76] На чертеже, «мишень 1» представляет геномную ДНК Chlamydia trachomatis (CT), «мишень 2» представляет Neisseria gonorrhoeae (NG), «NTC» представляет контроль без матрицы, и «RFU» представляет относительные единицы флуоресценции.
[77] На ФИГ. 14 показаны результаты ПЦР с детекцией в реальном времени для комбинации 2 в примерах.
[78] На чертеже, «мишень 1» представляет геномную ДНК Chlamydia trachomatis (CT), «мишень 2» представляет Ureaplasma parvum (UP), «NTC» представляет контроль без матрицы, и «RFU» представляет относительные единицы флуоресценции.
[79] На ФИГ. 15 показаны результаты ПЦР с детекцией в реальном времени для комбинации 3 в примерах.
[80] На чертеже, «мишень 1» представляет геномную ДНК Chlamydia trachomatis (CT), «мишень 2» представляет Neisseria gonorrhoeae (NG), «NTC» представляет контроль без матрицы, и «RFU» представляет относительные единицы флуоресценции. На ФИГ. 16 показаны результаты ПЦР с детекцией в реальном времени для комбинации 4 в примерах.
[81] На чертеже, «мишень 1» представляет геномную ДНК Neisseria gonorrhoeae (NG), «мишень 2» представляет Ureaplasma parvum (UP), «NTC» представляет контроль без матрицы, и «RFU» представляет относительные единицы флуоресценции.
[82] На ФИГ. 17a и 17b показаны результаты ПЦР с детекцией в реальном времени для комбинации 5 в примерах.
[83] На каждом чертеже, «мишень 1» представляет геномную ДНК Chlamydia trachomatis (CT), «мишень 2» представляет Neisseria gonorrhoeae (NG), «мишень 3» представляет Ureaplasma parvum (UP), «NTC» представляет контроль без матрицы, и «RFU» представляет относительные единицы флуоресценции.
Наилучший способ осуществления изобретения
[84] Авторы настоящего изобретения обнаружили, что различные механизмы образования сигнала для детекции нуклеиновой кислоты-мишени, хорошо известные в данной области, включают температурный диапазон, в котором сигнал изменяется, в зависимости от присутствия нуклеиновой кислоты-мишени (то есть, температурный диапазон изменения сигнала), и температурный диапазон, в котором нет изменения сигнала, даже в присутствии нуклеиновой кислоты-мишени (то есть, температурный диапазон постоянного сигнала), и могут быть разделены по категориям на три типа, в соответствии с количеством и/или порядком этих температурных диапазонов изменения сигнала и температурных диапазонов постоянного сигнала. Посредством применения различных комбинаций этих трех типов механизмов образования сигнала к детекции нуклеиновых кислот-мишеней, авторы настоящего изобретения разработали новый способ для детекции множества нуклеиновых кислот-мишеней посредством использования одного типа метки и одного типа детектора в одном реакционном сосуде.
[85]
[86] I. Способ детекции нуклеиновых кислот-мишеней
[87]
[88] В соответствии с одним аспектом, настоящее изобретение относится к способу детекции n нуклеиновых кислот-мишеней в образце, включающему: (a) детекцию сигналов при n температурах детекции, с инкубацией в то же время с n композициями для детекции нуклеиновых кислот-мишеней, образца, как подозревают, содержащего по меньшей мере одну из n нуклеиновых кислот-мишеней, в реакционном сосуде; где n представляет собой целое число 2 или более, где инкубация включает множество циклов, и детекцию сигналов проводят по меньшей мере в одном из множества циклов, где каждая из n композиций для детекции нуклеиновых кислот-мишеней обеспечивает изменение сигнала при соответствующей температуре детекции среди n температур детекции в присутствии соответствующей нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие соответствующей нуклеиновой кислоты-мишени, где композиция для детекции iй нуклеиновой кислоты-мишени среди n композиций для детекции нуклеиновых кислот-мишеней обеспечивает изменение сигнала при iй температуре детекции среди n температур детекции и обеспечивает постоянный сигнал при других температурах детекции в присутствии iй нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие iй нуклеиновой кислоты-мишени, и где i представляет собой целое число от 1 до n, и iя температура детекции является более низкой, чем (i+1)я температура детекции; и (b) определение присутствия n нуклеиновых кислот-мишеней из сигналов, детектированных на стадии (a), где присутствие iй нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при iй температуре детекции.
[89]
[90] Настоящее изобретение можно описать для каждой стадии следующим образом:
[91]
[92] Стадия (a): Инкубация и детекция сигналов
[93] Сначала, в одном реакционном сосуде, образец, как подозревают, содержащий по меньшей мере одну из n нуклеиновых кислот-мишеней, смешивают и инкубируют с n композициями для детекции нуклеиновых кислот-мишеней.
[94] В одном варианте осуществления, n представляет собой целое число 2 или более. Например, n может представлять собой 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40, 45 или 50, но не является ограниченным этим.
[95] Термин «нуклеиновая кислота-мишень», «последовательность-мишень нуклеиновой кислоты» или «последовательность-мишень», в рамках изобретения, относится к последовательности нуклеиновой кислоты, подлежащей детекции или количественной оценке. Последовательность-мишень нуклеиновой кислоты включает одноцепочечную, так же как двухцепочечную. Последовательность-мишень нуклеиновой кислоты включает не только последовательность, вновь образованную в реакции, но также последовательность, изначально присутствующую в образце нуклеиновой кислоты.
[96] Нуклеиновая кислота-мишень включает любые молекулы ДНК (гДНК и кДНК) и РНК, и их гибриды (химерные нуклеиновые кислоты). Последовательность может находиться в двухцепочечной или одноцепочечной форме.
[97] Нуклеиновые кислоты-мишени включают любую природную нуклеиновую кислоту прокариот, эукариот (например, простейших и паразитов, грибов, дрожжей, высших растений, низших животных и высших животных, включая млекопитающих и человека) или вирусов (например, вируса герпеса, HIV, вируса гриппа, вируса Эпштейна-Барр, вируса гепатита, вируса полиомиелита и т.д.), или вироида. Кроме того, молекула нуклеиновой кислоты может представлять собой любую молекулу нуклеиновой кислоты, которая получена или может быть получена посредством рекомбинации, или любую молекулу нуклеиновой кислоты, которая химически синтезирована или может быть химически синтезирована. Таким образом, последовательность нуклеиновой кислоты может быть или может не быть обнаружена в природе. Последовательность-мишень нуклеиновой кислоты может иметь известную или неизвестную последовательность.
[98] В одном варианте осуществления, n нуклеиновых кислот-мишеней могут включать вариант нуклеотида. Например, одна из n нуклеиновых кислот-мишеней может включать один тип варианта нуклеотида, и другая из n нуклеиновых кислот-мишеней может включать отличный тип варианта нуклеотида.
[99] В рамках изобретения, термин «вариант нуклеотида» относится к любым заменам, делециям или вставкам одного или множества нуклеотидов в последовательности ДНК в конкретной локализации среди непрерывных фрагментов ДНК. Такие непрерывные фрагменты ДНК включают ген или любую другую часть хромосомы. Эти варианты нуклеотидов могут представлять собой мутантные или полиморфные аллельные варианты. Например, вариант нуклеотида, детектированный по настоящему изобретению, включает однонуклеотидный полиморфизм (SNP), мутацию, делецию, вставку, замену и транслокацию. Примеры варианта нуклеотида включают многочисленные варианты в геноме человека (например, варианты в гене MTHFR (метилентетрагидрофолатредуктазы)), варианты, вовлеченные в устойчивость патогенов к лекарственному средству, и вызывающие онкогенез варианты. В рамках изобретения, термин «вариант нуклеотида» включает любой вариант в конкретной локализации в последовательности нуклеиновой кислоты. То есть, термин «вариант нуклеотида» включает дикий тип и любую его мутацию, в конкретной локализации в последовательности нуклеиновой кислоты.
[100] В рамках изобретения, термин «образец» относится к клеткам, тканям или жидкости из биологического источника, или к любой другой среде, для которой можно доказать, что ее можно использовать по настоящему изобретению, и включает вирус, бактерии, ткани, клетки, кровь, сыворотку, плазму, лимфу, молоко, мочу, фекалии, внутриглазную жидкость, слюну, сперму, экстракт головного мозга, спинномозговую жидкость, аппендикс, экстракты ткани селезенки и миндалин, амниотическую жидкость, асциты и небиологические образцы (например, пищи и воды). Кроме того, образец включает природную молекулу нуклеиновой кислоты, выделенную из биологического источника, и синтезированные молекулы нуклеиновых кислот.
[101] Настоящее изобретение используют для определения того, присутствует ли по меньшей мере одна из n нуклеиновых кислот-мишеней в образце. Например, когда n представляет собой 2, настоящее изобретение можно использовать для определения того, присутствует ли по меньшей мере одна из первой нуклеиновой кислоты-мишени и второй нуклеиновой кислоты-мишени в образце. В качестве другого примера, когда n представляет собой 3, настоящее изобретение можно использовать для определения того, присутствует ли по меньшей мере одна среди первой нуклеиновой кислоты-мишени, второй нуклеиновой кислоты-мишени и третьей нуклеиновой кислоты-мишени в образце.
[102] В одном варианте осуществления, реакция при инкубации относится к любой реакции, которая индуцирует изменение сигнала, в зависимости от присутствия соответствующей нуклеиновой кислоты-мишени при соответствующей температуре детекции, по мере того, как каждая из нуклеиновых кислот-мишеней вступает в реакцию с соответствующей композицией для детекции нуклеиновой кислоты-мишени.
[103] В одном варианте осуществления, инкубация включает множество циклов.
[104] В одном варианте осуществления, инкубация на стадии (a) может включать реакцию амплификации, и может включать, например, реакцию амплификации сигнала и/или реакцию амплификации нуклеиновой кислоты.
[105] В одном варианте осуществления, реакция амплификации включает множество циклов.
[106] В одном варианте осуществления, инкубацию на стадии (a) проводят в условиях, позволяющих амплификацию мишени и изменение сигнала посредством композиции для детекции нуклеиновой кислоты-мишени. Такие условия включают температуру, концентрацию соли и pH раствора.
[107] В одном варианте осуществления, инкубацию на стадии (a) проводят в процессе амплификации сигнала без амплификации нуклеиновой кислоты.
[108] В одном варианте осуществления, сигнал можно амплифицировать одновременно с амплификацией мишени. Альтернативно, сигнал можно амплифицировать без амплификации мишени.
[109] В одном варианте осуществления, изменение сигнала происходит в ходе процесса, включающего амплификацию сигнала и амплификацию мишени.
[110] В одном варианте осуществления, амплификацию нуклеиновой кислоты-мишени можно проводить посредством полимеразной цепной реакции (ПЦР). ПЦР широко используют в данной области для амплификации нуклеиновых кислот-мишеней, и она включает циклы денатурации нуклеиновых кислот-мишеней, отжига (гибридизации) между нуклеиновыми кислотами-мишенями и праймерами, и удлинения праймеров (Mullis et al., Патенты США No. 4683195, 4683202 и 4800159; Saiki et al., (1985) Science 230, 1350-1354).
[111] В одном варианте осуществления, амплификацию нуклеиновых кислот-мишеней можно проводить посредством лигазной цепной реакции (LCR) (Патенты США No. 4683195 и 4683202; PCR Protocols: A Guide to Methods and Applications (Innis et al., eds, 1990)), амплификации с вытеснением цепи (SDA) (Walker, et al., Nucleic Acids Res. 20(7):1691-6 (1992); Walker PCR Methods Appl 3(1):1-6 (1993)), опосредованной транскрипцией амплификации (Phyffer, et al., J. Clin. Microbiol. 34:834-841 (1996); Vuorinen, et al., J. Clin. Microbiol. 33:1856-1859 (1995)), зависимой от хеликазы амплификации (HAD) (M. Vincent, Y. Xu and H. Kong, EMBO Rep., 2004, 5, 795-800), основанной на последовательности нуклеиновой кислоты амплификации (NASBA) (Compton, Nature 350(6313):91-2 (1991)), амплификации по типу катящегося кольца (RCA) (Lisby, Mol. Biotechnol. 12(1):75-99 (1999); Hatch et al., Genet. Anal. 15(2):35-40 (1999)), Q-бета-репликазы (Lizardi et al., BiolTechnology 6:1197 (1988)), петлевой изотермической амплификации (LAMP) (Y. Mori, H. Kanda and T. Notomi, J. Infect. Chemother., 2013, 19, 404-411) или рекомбиназной полимеразной амплификации (RPA) (J. Li, J. Macdonald and F. von Stetten, Analyst, 2018, 144, 31-67).
[112] Различные ДНК-полимеразы могут быть использованы в реакции амплификации и включают фрагмент «Кленова» ДНК-полимеразы I E. coli, термостабильную ДНК-полимеразу и ДНК-полимеразу бактериофага T7. В частности, полимераза представляет собой термостабильную ДНК-полимеразу, которая может быть получена из различных бактерий, включающих Thermus aquaticus (Taq), Thermus thermophilus (Tth), Thermus filiformis, Thermis flavus, Thermococcus literalis и Pyrococcus furiosus (Pfu). Большинство из вышеуказанных полимераз можно выделять как есть из бактерий или можно закупать из коммерческих источников.
[113] Вышеописанным способом амплификации можно амплифицировать нуклеиновую кислоту-мишень и/или сигнал посредством повторения серий реакций в присутствии или в отсутствие изменения температуры. Звено амплификации, включающее повторение таких серий реакций, обозначено как «цикл». Цикл можно выражать как количество повторений или длительность, в зависимости от используемого способа амплификации.
[114] В одном варианте осуществления, серии реакций можно проводить последовательно. Например, для ПЦР, после денатурации нуклеиновых кислот-мишеней (то есть, матриц), отжиг праймеров и впоследствии, удлинение праймеров можно проводить последовательно. В этом случае, цикл можно выражать как количество повторений.
[115] В одном варианте осуществления, серии реакций можно проводить одновременно. Например, в LAMP, представляющей собой анализ изотермической амплификации, гибридизация праймеров может происходить на некоторых матрицах среди множества матриц, в то время как на некоторых других матрицах, праймеры могли быть уже гибридизованы, так что может происходить удлинение праймеров. В этом случае, цикл можно выражать как длительность. В частности, 1 цикл может составлять 5 секунд, 10 секунд, 1 минуту, 2 минуты, 3 минуты, 4 минуты, 5 минут, 10 минут, 20 минут, 30 минут, 1 час или 2 часа.
[116] В одном варианте осуществления, инкубацию можно проводить в течение множества циклов, что позволяет измерение изменения сигнала, зависимого от присутствия нуклеиновой кислоты-мишени. Например, множество циклов может включать 2-100 циклов, 2-90 циклов, 2-80 циклов, 2-70 циклов, 2-60 циклов, 2-50 циклов, 2-40 циклов, 2-30 циклов, 2-20 циклов, 2-10 циклов, 5-100 циклов, 5-90 циклов, 5-80 циклов, 5-70 циклов, 5-60 циклов, 5-50 циклов, 5-40 циклов, 5-30 циклов, 5-20 циклов, 5-10 циклов, 10-100 циклов, 10-90 циклов, 10-80 циклов, 10-70 циклов, 10-60 циклов, 10-50 циклов, 10-40 циклов, 10-30 циклов, 10-20 циклов, 20-100 циклов, 20-90 циклов, 20-80 циклов, 20-70 циклов, 20-60 циклов, 20-50 циклов, 20-40 циклов или 20-30 циклов, и в частности, может включать 10 циклов, 15 циклов, 20 циклов, 25 циклов, 30 циклов, 35 циклов, 40 циклов, 45 циклов или 50 циклов.
[117] В одном варианте осуществления, детекцию сигналов можно проводить в каждом цикле, в избранных нескольких циклах или в конечной точке инкубации реакции, включающей множество циклов.
[118] В одном варианте осуществления, реакция амплификации нуклеиновой кислоты-мишени может представлять собой реакцию амплификации множества нуклеиновых кислот-мишеней.
[119] Термин «реакция амплификации множества нуклеиновых кислот-мишеней», в рамках изобретения, относится к реакции, в которой амплифицируют две или более нуклеиновых кислот, в качестве мишеней, в одном реакционном сосуде. Реакция амплификации множества нуклеиновых кислот-мишеней относится к реакции, в которой амплифицируют две или более нуклеиновых кислот совместно. Например, в реакции амплификации множества нуклеиновых кислот-мишеней можно амплифицировать, в одной реакции, 2 или более, 3 или более, 4 или более, 5 или более, 6 или более, 7 или более, 8 или более, 9 или более, 10 или более, 20 или более, 30 или более, 40 или более, или 50 или более нуклеиновых кислот-мишеней совместно.
[120] В одном варианте осуществления, способом по настоящему изобретению можно детектировать 2-50, 2-40, 2-30, 2-20, 2-15, 2-12, 2-10, 2-9, 2-8, 2-7, 2-6, 2-5, 3-50, 3-40, 3-30, 3-20, 3-15, 3-12, 3-10, 3-9, 3-8, 3-7, 3-6, 3-5, 4-50, 4-40, 4-30, 4-20, 4-15, 4-12, 4-10, 4-9, 4-8, 4-7, 4-6 или 4-5 нуклеиновых кислот-мишеней посредством использования одного типа метки в одном реакционном сосуде.
[121] В одном варианте осуществления, способ по настоящему изобретению, при использовании множества типов меток, может детектировать большее количество нуклеиновых кислот-мишеней, чем количество нуклеиновых кислот-мишеней, поддающихся детекции с использованием одного типа метки, в соответствии со способом по настоящему изобретению, и например, может являться способным к детекции большего количества нуклеиновых кислот-мишеней, в количество раз увеличения количества используемых типов меток.
[122] В соответствии с настоящим изобретением, в способе в соответствии с настоящим изобретением используют n различных композиций для детекции n нуклеиновых кислот-мишеней, каждая из которых соответствует своей соответствующей нуклеиновой кислоте-мишени.
[123] Например, когда n представляет собой 2, используют композицию для детекции первой нуклеиновой кислоты-мишени и композицию для детекции второй нуклеиновой кислоты-мишени; когда n представляет собой 3, используют композицию для детекции первой нуклеиновой кислоты-мишени, композицию для детекции второй нуклеиновой кислоты-мишени и композицию для детекции третьей нуклеиновой кислоты-мишени; и когда n представляет собой 4, используют композицию для детекции первой нуклеиновой кислоты-мишени - композицию для детекции четвертой нуклеиновой кислоты-мишени.
[124] В одном варианте осуществления, каждая из n композиций для детекции нуклеиновых кислот-мишеней обеспечивает изменение сигнала при соответствующей температуре детекции среди n температур детекции, где изменение сигнала указывает на присутствие соответствующей нуклеиновой кислоты-мишени.
[125] Например, композиция для детекции iй нуклеиновой кислоты-мишени среди n нуклеиновых кислот-мишеней обеспечивает изменение сигнала при iй температуре детекции среди n температур детекции и обеспечивает постоянный сигнал при других температурах детекции в присутствии iй нуклеиновой кислоты-мишени.
[126] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени, в присутствии iй нуклеиновой кислоты-мишени, обеспечивает изменение сигнала при амплификации нуклеиновой кислоты-мишени при iй температуре детекции (то есть, изменение iго сигнала) с обеспечением в то же время отсутствия изменения сигнала при других температурах детекции, даже по мере амплификации нуклеиновой кислоты-мишени (то есть, сигнал является постоянным).
[127] Термин «iй сигнал», в рамках изобретения, относится к сигналу, обеспечиваемому при iй температуре детекции посредством композиции для детекции iй нуклеиновой кислоты-мишени, который использован взаимозаменяемо с «сигнал при iй температуре детекции».
[128] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени, когда находится в отсутствие iй нуклеиновой кислоты-мишени, обеспечивает отсутствие изменения сигнала, то есть, обеспечивает постоянный сигнал при iй температуре детекции в ходе инкубации реакции (например, реакции амплификации нуклеиновой кислоты-мишени). В одном варианте осуществления, при детекции n нуклеиновых кислот-мишеней, iй сигнал может означать сигнал, обеспечиваемый посредством n композиций для детекции нуклеиновых кислот-мишеней, включая композицию для детекции iй нуклеиновой кислоты-мишени, при iй температуре детекции.
[129] В одном варианте осуществления, i представляет собой целое число от 1 до n, и iя температура детекции является более низкой, чем (i+1)я температура детекции. В одном варианте осуществления, когда i представляет собой n, существует i+1 температур детекции (то есть, n+1 температур детекции). Например, когда n представляет собой 3, i представляет собой целое число от 1 до 3, и существует первая температура детекции, вторая температура детекции и третья температура детекции, где первая температура детекции является более низкой, чем вторая температура детекции, и вторая температура детекции является более низкой, чем третья температура детекции.
[130] В конкретном варианте осуществления, когда n представляет собой 2, композиция для детекции первой нуклеиновой кислоты-мишени обеспечивает изменение сигнала при первой температуре детекции и обеспечивает постоянный сигнал при других температурах детекции, то есть, при второй температуре детекции, в присутствии первой нуклеиновой кислоты-мишени; и композиция для детекции второй нуклеиновой кислоты-мишени обеспечивает изменение сигнала при второй температуре детекции и обеспечивает постоянный сигнал при других температурах детекции, то есть, при первой температуре детекции, в присутствии второй нуклеиновой кислоты-мишени,.
[131] В другом конкретном варианте осуществления, когда n представляет собой 3, композиция для детекции первой нуклеиновой кислоты-мишени обеспечивает изменение сигнала при первой температуре детекции и обеспечивает постоянный сигнал при других температурах детекции, то есть, при второй температуре детекции и третьей температуре детекции, в присутствии первой нуклеиновой кислоты-мишени; композиция для детекции второй нуклеиновой кислоты-мишени обеспечивает изменение сигнала при второй температуре детекции и обеспечивает постоянный сигнал при других температурах детекции, то есть, при первой температуре детекции и третьей температуре детекции, в присутствии второй нуклеиновой кислоты-мишени; и композиция для детекции третьей нуклеиновой кислоты-мишени обеспечивает изменение сигнала при третьей температуре детекции и обеспечивает постоянный сигнал при других температурах детекции, то есть, при первой температуре детекции и второй температуре детекции, в присутствии третьей нуклеиновой кислоты-мишени.
[132] В одном варианте осуществления, изменение сигнала включает «образование или угасание сигнала» и «увеличение или уменьшение сигнала» от метки.
[133] В настоящем описании, изменение сигнала означает значимое «изменение сигнала» то есть, значимое изменение сигнала, где изменение сигнала указывает на присутствие нуклеиновой кислоты-мишени. Например, значимое изменение сигнала, то есть, изменение сигнала, которое указывает на присутствие нуклеиновой кислоты-мишени, может относиться к образованию или угасанию сигнала, имеющему отличную интенсивность, по сравнению с фоновой интенсивностью сигнала или интенсивностью сигнала в отсутствие нуклеиновой кислоты-мишени, или может относиться к существенному увеличению или уменьшению интенсивности сигнала, указывающему на присутствие нуклеиновой кислоты-мишени, по мере амплификации нуклеиновой кислоты-мишени и/или сигнала в ходе инкубации реакции на стадии (a).
[134] В одном варианте осуществления, изменение сигнала может возникать в зависимости от присутствия нуклеиновой кислоты-мишени. Например, изменение сигнала может возникать по мере образования или угасания сигнала, указывающего на присутствие нуклеиновой кислоты-мишени.
[135] В одном варианте осуществления, изменение сигнала может возникать при амплификации нуклеиновой кислоты-мишени. То есть, изменение сигнала может возникать по мере увеличения количества нуклеиновой кислоты-мишени при амплификации нуклеиновой кислоты-мишени. Например, при амплификации нуклеиновой кислоты-мишени, сигнал, указывающий на присутствие нуклеиновой кислоты-мишени, можно увеличивать или уменьшать, таким образом, индуцируя изменение сигнала.
[136] В одном варианте осуществления, изменение сигнала может возникать при амплификации сигнала, зависимой от нуклеиновой кислоты-мишени. То есть, при амплификации сигнала, зависимой от нуклеиновой кислоты-мишени, интенсивность сигнала изменяется, таким образом, изменяя сигнал. Например, по мере амплификации сигнала, зависимой от присутствия нуклеиновой кислоты-мишени, сигнал, указывающий на присутствие нуклеиновой кислоты-мишени, можно увеличивать или уменьшать, таким образом, индуцируя изменение сигнала.
[137] Термин «постоянный сигнал», в рамках изобретения, относится к отсутствию существенного изменения сигнала в ходе инкубации реакции (например, реакции амплификации нуклеиновой кислоты-мишени). То есть, термин «постоянный сигнал» относится ко всем или к любому паттернам сигнала, отличным от значимых изменений сигнала, вызванных амплификацией присутствующей нуклеиновой кислоты-мишени. Конкретно, постоянный сигнал означает отсутствие изменения сигнала. Например, если сигнал в ходе реакции амплификации не превышает фоновую интенсивность сигнала или интенсивность сигнала в отсутствие нуклеиновой кислоты-мишени, это можно выражать как «сигнал является постоянным». В настоящем описании, постоянный сигнал может быть взаимозаменяемо использован как сигнал, который не изменяется, или сигнал, для которого не показано изменение.
[138] В настоящем описании, обозначение «изменение сигнала» и/или «постоянный сигнал» основано на сигналах, детектированных при одной и той же температуре в ходе реакции амплификации нуклеиновой кислоты с использованием одной и той же композиции для детекции нуклеиновой кислоты-мишени. Например, обозначение «изменение сигнала» и/или «постоянный сигнал» используют на основании различия между значениями сигнала, детектированными при одной и той же температуре с использованием n композиций для детекции нуклеиновых кислот-мишеней, и более конкретно, «изменение сигнала» и/или «постоянный сигнал» обозначают, на этом основании, как «изменение сигнала» и/или «постоянный сигнал» на основании (i) различия между значениями сигнала, детектированными при одной и той же температуре за множество циклов, или (ii) различия между «контрольным значением сигнала», описанным ниже, и значением сигнала, детектированным при той же температуре, что и температура, для которой установлено контрольное значение сигнала. То есть, обозначение «изменение сигнала» и/или «постоянный сигнал» не основано на различии между значениями сигнала, детектированными при различных температурах.
[139]
[140] В настоящем описании, для обеспечения изменений сигналов для нуклеиновых кислот-мишеней, используют композиции для детекции нуклеиновых кислот-мишеней. Каждую из нуклеиновых кислот-мишеней детектируют посредством соответствующей композиции для детекции нуклеиновой кислоты-мишени.
[141] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени включает метку, которая обеспечивает сигнал в зависимости от присутствия iй нуклеиновой кислоты-мишени.
[142] В одном варианте осуществления, метка является связанной с олигонуклеотидом, или встраивается в олигонуклеотид в ходе инкубации (например, реакции амплификации нуклеиновой кислоты). То есть, композиция для детекции нуклеиновой кислоты-мишени может изначально включать связанный с меткой олигонуклеотид, или может предоставлять связанный с меткой олигонуклеотид, по мере встраивания метки во вновь образованный олигонуклеотид (например, удлиняемую цепь) в ходе инкубации реакции.
[143] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени включает метку для встраивания, которая встраивается в олигонуклеотид в ходе инкубации и обеспечивает сигнал в зависимости от присутствия iй нуклеиновой кислоты-мишени.
[144] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет связанный с меткой олигонуклеотид, который служит для обеспечения сигнала в зависимости от присутствия iй нуклеиновой кислоты-мишени.
[145] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени изначально включает связанный с меткой олигонуклеотид, который служит для обеспечения сигнала в зависимости от присутствия iй нуклеиновой кислоты-мишени. Альтернативно, композиция для детекции iй нуклеиновой кислоты-мишени может включать олигонуклеотид и метку, которая обеспечивает сигнал в зависимости от присутствия нуклеиновой кислоты-мишени, и по мере встраивания метки в олигонуклеотид в ходе инкубации реакции (например, реакции амплификации нуклеиновой кислоты), предоставляет связанный с меткой олигонуклеотид, который служит для обеспечения сигнала в зависимости от присутствия нуклеиновой кислоты-мишени.
[146] В рамках изобретения, термин «связанный с меткой олигонуклеотид» относится к олигонуклеотиду, вовлеченному в образование сигнала, подвергаемого детекции.
[147] В одном варианте осуществления, связанный с меткой олигонуклеотид может содержать олигонуклеотид, который специфически гибридизуется с нуклеиновой кислотой-мишенью (например, зонд или праймер); когда зонд или праймер, гибридизованный с нуклеиновой кислотой-мишенью, расщепляется для высвобождения фрагмента, связанный с меткой олигонуклеотид может содержать связывающий олигонуклеотид, который специфически гибридизуется с фрагментом; когда фрагмент, гибридизованный со связывающим олигонуклеотидом, удлиняется для формирования удлиненной цепи, связанный с меткой олигонуклеотид может содержать олигонуклеотид, который специфически гибридизуется с удлиненной цепью, олигонуклеотид, который получен посредством встраивания метки в ходе удлинения фрагмента, олигонуклеотид, который специфически гибридизуется с связывающим олигонуклеотидом, и их комбинацию.
[148] В соответствии с одним вариантом осуществления, связанный с меткой олигонуклеотид включает олигонуклеотид, вовлеченный в фактическое образование сигнала. Например, гибридизация или отсутствие гибридизации между связанным с меткой олигонуклеотидом и другим олигонуклеотидом (например, олигонуклеотидом, содержащим нуклеотидную последовательность, комплементарную связанному с меткой олигонуклеотиду или нуклеиновой кислоте-мишени) определяет образование сигнала.
[149] В одном варианте осуществления, связанный с меткой олигонуклеотид может представлять собой «зонд», известный в данной области. Термин «зонд», в рамках изобретения, относится к одноцепочечной молекуле нуклеиновой кислоты, содержащей одну или несколько частей, по существу комплементарных последовательности-мишени нуклеиновой кислоты. В соответствии с вариантом осуществления настоящего изобретения, 3'-конец зонда является «блокированным», чтобы запретить его удлинение. Блокирование можно осуществлять в соответствии с общепринятыми способами. Например, блокирование можно проводить посредством добавления к 3′-гидроксильной группе последнего нуклеотида химической группы, такой как биотин, метки, фосфатные группы, алкильные группы, ненуклеотидные линкеры, фосфоротиоатные или алкандиольные остатки. Альтернативно, блокирование можно проводить посредством удаления 3′-гидроксильной группы последнего нуклеотида или с использованием нуклеотида без 3′-гидроксильной группы, такого как дидезоксинуклеотид.
[150] В соответствии с вариантом осуществления, связанный с меткой олигонуклеотид может состоять из по меньшей мере одного олигонуклеотида. В соответствии с вариантом осуществления настоящего изобретения, когда связанный с меткой олигонуклеотид состоит из множества олигонуклеотидов, связанный с меткой олигонуклеотид может являться меченным различными способами. Например, все или часть множества олигонуклеотидов могут иметь по меньшей мере одну метку.
[151] В одном варианте осуществления, интерактивные метки могут представлять собой интерактивные двойные метки, включающие одну репортерную молекулу и одну молекулу гасителя.
[152] В одном варианте осуществления, интерактивные метки могут представлять собой интерактивные метки, включающие по меньшей мере одну репортерную молекулу и по меньшей мере одну молекулу гасителя. В частности, интерактивные метки могут представлять собой интерактивные двойные метки, включающие одну репортерную молекулу и одну молекулу гасителя. Или, интерактивные метки могут представлять собой интерактивные метки, включающие одну репортерную молекулу и две молекулы гасителя.
[153] В одном варианте осуществления, когда метка представляет собой одиночную метку, одиночная метка может являться связанной с одним олигонуклеотидом.
[154] В одном варианте осуществления, когда метка представляет собой интерактивные метки, интерактивные метки могут представлять собой интерактивные метки, включающие по меньшей мере одну репортерную молекулу и по меньшей мере одну молекулу гасителя, где все интерактивные метки могут являться связанными с одним олигонуклеотидом или могут являться связанными с каждым из множества олигонуклеотидов.
[155] Например, одиночная метка включает флуоресцентную метку, люминесцентную метку, хемилюминесцентную метку, электрохимическую метку и металлическую метку. В одном варианте осуществления, одиночная метка обеспечивает различные сигналы (например, различные силы сигналов), в зависимости от ее присутствия на двойной цепи или одиночной цепи. В одном варианте осуществления, одиночная метка представляет собой флуоресцентную метку. Предпочтительные типы и участки связывания одиночных флуоресцентных меток, используемых в настоящем описании, описаны в Патентах США No. 7537886 и 7348141, полное содержание объяснения которых приведено в настоящем описании в качестве ссылки. Например, одиночная флуоресцентная метка включает метку на основе JOE, FAM, TAMRA, ROX и флуоресцеина. Одиночная метка может являться связанной с олигонуклеотидом посредством различных способов. Например, метка может являться связанной с зондом посредством спейсера, содержащего атомы углерода (например, 3-углеродного спейсера, 6-углеродного спейсера или 12-углеродного спейсера).
[156] В качестве репрезентативной системы интерактивной метки, система метки FRET (резонансного переноса энергии флуоресценции) включает флуоресцентную репортерную молекулу (донорную молекулу) и молекулу гасителя (акцепторную молекулу). В FRET, донор энергии является флуоресцентным, но акцептор энергии может являться флуоресцентным или нефлуоресцентным. В другой форме систем интерактивной метки, донор энергии является нефлуоресцентным, например, хромофором, и акцептор энергии является флуоресцентным. В другой форме систем интерактивной метки, донор энергии является люминесцентным, например, биолюминесцентным, хемилюминесцентным, электрохемилюминесцентным, и акцептор является флуоресцентным. Система интерактивной метки включает пару меток на основе «опосредованного контактом гашения» (Salvatore et al., Nucleic Acids Research, 2002 (30) no.21 e122 и Johansson et al., J. AM. CHEM. SOC 2002 (124) pp 6950-6956). Система интерактивной метки включает любую или все системы метки, которые индуцируют изменения сигнала посредством взаимодействий между по меньшей мере двумя молекулами (например, красителями).
[157] Репортерная молекула и молекула гасителя, которые можно использовать по настоящему изобретению, могут включать любые молекулы, известные в данной области. Примеры таких молекул являются следующими: Cy2™ (506), YO-PRO™-1 (509), YOYO™-1 (509), кальцеин (517), FITC (518), FluorX™ (519), Alexa™ (520), родамин 110 (520), орегонский зеленый™ 500 (522), орегонский зеленый™ 488 (524), RiboGreen™ (525), родаминовый зеленый™ (527), родамин 123 (529), магниевый зеленый™ (531), кальциевый зеленый™ (533), TO-PRO™-1 (533), TOTO1 (533), JOE (548), BODIPY530/550 (550), Dil (565), BODIPY TMR (568), BODIPY558/568 (568), BODIPY564/570 (570), Cy3™ (570), Alexa™ 546 (570), TRITC (572), магниевый оранжевый™ (575), фикоэритрин R&B (575), родамин-фаллоидин (575), кальциевый оранжевый™ (576), пиронин Y (580), родамин B (580), TAMRA (582), родаминовый красный™ (590), Cy3.5™ (596), ROX (608), кальциевый карминный™ (615), Alexa™ 594 (615), техасский красный (615), нильский красный (628), YO-PRO™-3 (631), YOYO™-3 (631), R-фикоцианин (642), C-Фикоцианин (648), TO-PRO™-3 (660), TOTO3 (660), DiD DilC(5) (665), Cy5™ (670), тиадикарбоцианин (671), Cy5.5 (694), HEX (556), TET (536), Biosearch синий (447), CAL Fluor золотой 540 (544), CAL Fluor оранжевый 560 (559), CAL Fluor красный 590 (591), CAL Fluor красный 610 (610), CAL Fluor красный 635 (637), FAM (520), флуоресцеин (520), флуоресцеин-C3 (520), Pulsar 650 (566), Quasar 570 (667), Quasar 670 (705) и Quasar 705 (610). Числа в скобках представляют собой максимальную длину волны излучения в нанометрах. Предпочтительно, репортерная молекула и молекула гасителя включают метку на основе JOE, FAM, TAMRA, ROX и флуоресцеина.
[158] Подходящие пары репортер-гаситель описаны во множестве следующих публикаций: Pesce et al., editors, Fluorescence Spectroscopy (Marcel Dekker, New York, 1971); White et al., Fluorescence Analysis: A Practical Approach (Marcel Dekker, New York, 1970); Berlman, Handbook of Fluorescence Spectra of Aromatic Molecules, 2nd Edition (Academic Press, New York, 1971); Griffiths, Color AND Constitution of Organic Molecules (Academic Press, New York, 1976); Bishop, editor, Indicators (Pergamon Press, Oxford, 1972); Haugland, Handbook of Fluorescent Probes and Research Chemicals (Molecular Probes, Eugene, 1992); Pringsheim, Fluorescence and Phosphorescence (Interscience Publishers, New York, 1949); Haugland, R. P., Handbook of Fluorescent Probes and Research Chemicals, 6th Edition (Molecular Probes, Eugene, Oreg., 1996); Патенты США No. 3996345 и 4351760.
[159] В одном варианте осуществления, метку для встраивания можно использовать в процессе встраивания метки в ходе удлинения праймера, для образования сигнала (например, Plexor technology, Sherrill C B et al., Journal of the American Chemical Society, 126:4550-45569 (2004)). Кроме того, метку для встраивания можно использовать в образовании сигнала посредством дуплекса, сформированного способом, зависимым от расщепления олигонуклеотида-посредника, гибридизованного с последовательностью-мишенью нуклеиновой кислоты.
[160] В одном варианте осуществления, метка для встраивания может являться, как правило, связанной с нуклеотидом. Кроме того, можно использовать нуклеотид, имеющий неприродное основание.
[161] В рамках изобретения, термин «неприродное основание» относится к производным природных оснований, таких как аденин (A), гуанин (G), тимин (T), цитозин (C) и урацил (U), которые являются способными к формированию пар оснований с водородными связями. Термин «неприродное основание», в рамках изобретения, включает основания, имеющие паттерны спаривания оснований, отличные от природных оснований в качестве родительских соединений, как описано, например, в Патентах США No. 5432272, 5965364, 6001983 и 6037120. Спаривание оснований между неприродными основаниями включает две или три водородные связи, как для природных оснований. Спаривание оснований между неприродными основаниями также формируется специфическим образом. Конкретные примеры неприродных оснований включают следующие основания в комбинациях пар оснований: изо-C/изо-G, изо-dC/ изо-dG, Z/P, V/J, K/X, H/J, Pa/Ds, Pa/Q, Pn/Ds, Pn/Dss, Px/Ds, NaM/5SICS, 5FM/5SICS и M/N (см. Патенты США No. 5432272; 5965364; 6001983; 6037120; 6140496; 6627456; 6617106; и 7422850; и Filip Wojciechowski et al., Chem. Soc. Rev., 2011, 40, 5669-5679).
[162] Общепринятый способ детекции множества нуклеиновых кислот-мишеней требует использования различных типов флуоресцентных меток для различных нуклеиновых кислот-мишеней, или даже при использовании одного типа флуоресцентной метки, эти способы имеют тот недостаток, что они требуют дополнительного анализа, такого как анализ кривой плавления. В отличие от таких способов, посредством использования композиции для детекции нуклеиновой кислоты-мишени, которая предоставляет дуплекс, способом в соответствии с настоящим изобретением можно детектировать множество нуклеиновых кислот-мишеней в способе с детекцией в реальном времени с использованием одного типа метки (например, одиночной флуоресцентной метки) без дополнительного анализа, такого как анализ плавления.
[163] В одном варианте осуществления, каждая из n композиций для детекции нуклеиновых кислот-мишеней предоставляет один или несколько дуплексов.
[164] Термин «дуплекс», в рамках изобретения, предназначен для включения как дуплекса в связанной форме, так и дуплекса в диссоциированной форме. То есть, указанный термин может относиться к двум одноцепочечным молекулам нуклеиновых кислот, имеющим последовательности, частично или полностью комплементарные друг другу, так что, в условиях гибридизации, они могут гибридизоваться друг с другом, чтобы иметь структуру дуплекса. Например, в зависимости от температуры детекции, все или часть дуплексов могут находится в связанной форме или в диссоциированной форме.
[165] Термин «связывание или диссоциация» имеет такое же значение, как термин «гибридизация или денатурация».
[166] Выражение «композиция для детекции нуклеиновой кислоты-мишени предоставляет дуплекс», в рамках изобретения, может означать предоставление дуплекса в связанной форме и/или дуплекса в диссоциированной форме. Подобным образом, выражение «композиция для детекции нуклеиновой кислоты-мишени образует дуплекс в ходе инкубации», в рамках изобретения, может означать образование дуплекса в связанной форме и/или дуплекса в диссоциированной форме в ходе инкубации реакции.
[167] В одном варианте осуществления, по меньшей мере один из дуплексов, предоставленных посредством композиции для детекции нуклеиновой кислоты-мишени, представляет собой дуплекс, обеспечивающий сигнал. В частности, дуплекс представляет собой дуплекс, обеспечивающий изменение сигнала. То есть, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий сигнал, и в частности, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала в зависимости от присутствия iй нуклеиновой кислоты-мишени.
[168] Термин «дуплекс обеспечивающий сигнал», в рамках изобретения, относится к дуплексу, способному к предоставлению сигнала, который можно различать, в зависимости от того, находится ли дуплекс в связанной форме или диссоциированной форме. Например, это означает, что дуплекс в связанной форме образует (или гасит) сигнал, и дуплекс в диссоциированной форме гасит (или образует) сигнал.
[169] В одном варианте осуществления, дуплекс, обеспечивающий сигнал, может включать по меньшей мере одну метку. В частности, по меньшей мере одна метка является связанной с по меньшей мере одной цепью из двух одиночных цепей, составляющих дуплекс. Например, дуплекс, обеспечивающий сигнал, включает одиночную метку, и в этом случае, одиночная метка является связанной с любой одной цепью из двух одиночных цепей, составляющих дуплекс. В качестве другого примера, дуплекс, обеспечивающий сигнал, включает интерактивные метки, и в этом случае, все интерактивные метки являются связанными с одной из двух цепей, составляющих дуплекс, обеспечивающий сигнал, или одна из интерактивных меток является связанной с любой из двух одиночных цепей, и другая из интерактивных меток является связанной с другой из двух одиночных цепей.
[170] В рамках изобретения, термин «дуплекс, обеспечивающий изменение сигнала», относится к дуплексу, обеспечивающему изменение сигнала, показательное для присутствия нуклеиновой кислоты-мишени, по мере изменения количества дуплекса, обеспечивающего изменение сигнала в зависимости от присутствия нуклеиновой кислоты-мишени. В частности, в способе на основе PTOCE, описанном ниже, удлиненный дуплекс, включающий метку, образованный в зависимости от присутствия нуклеиновой кислоты-мишени, является примером дуплекса, обеспечивающего изменение сигнала, описанного в настоящем описании.
[171] В одном варианте осуществления, дуплекс, обеспечивающий изменение сигнала, включает метку. В частности, по меньшей мере одна метка является связанной с по меньшей мере одной цепью из двух одиночных цепей, составляющих дуплекс. Например, дуплекс, обеспечивающий изменение сигнала, включает одиночную метку, и в этом случае, одиночная метка является связанной с любой одной цепью из двух одиночных цепей, составляющих дуплекс. В качестве другого примера, дуплекс, обеспечивающий изменение сигнала, включает интерактивные метки, и в этом случае, все интерактивные метки являются связанными с одной из двух цепей, составляющих дуплекс, обеспечивающий изменение сигнала, или одна из интерактивных меток является связанной с любой из двух одиночных цепей, и другая из интерактивных меток является связанной с другой из двух одиночных цепей.
[172] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала.
[173] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени обеспечивает сигнал от метки, когда дуплекс, обеспечивающий изменение сигнала, присутствует в связанной форме. То есть, композиция для детекции iй нуклеиновой кислоты-мишени обеспечивает сигнал, зависящий от связывания двух одноцепочечных молекул нуклеиновых кислот, составляющих дуплекс.
[174] В альтернативном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени обеспечивает сигнал от метки, когда дуплекс, обеспечивающий изменение сигнала, присутствует в диссоциированной форме. То есть, композиция для детекции iй нуклеиновой кислоты-мишени обеспечивает сигнал, зависящий от диссоциации двух одноцепочечных молекул нуклеиновых кислот, составляющих дуплекс.
[175] В одном варианте осуществления, связывание или диссоциация дуплекса могут быть вызваны температурой.
[176] В одном варианте осуществления, дуплекс, обеспечивающий изменение сигнала, может представлять собой дуплекс, который был изначально (исходно) включен в композицию для детекции нуклеиновой кислоты-мишени.
[177] В одном варианте осуществления, когда дуплекс, обеспечивающий изменение сигнала, был включен в композицию для детекции нуклеиновой кислоты-мишени, дуплекс может быть образован посредством гибридизации между связанным с меткой олигонуклеотидом и олигонуклеотидом, способным к гибридизации со связанным с меткой олигонуклеотидом. Например, зонд Инь-Янь, описанный ниже, представляет собой пример дуплекса, обеспечивающего изменение сигнала в зависимости от присутствия нуклеиновой кислоты-мишени, и был изначально включен в композицию для детекции нуклеиновой кислоты-мишени.
[178] В одном варианте осуществления, когда дуплекс, обеспечивающий изменение сигнала, был изначально включен в композицию для детекции нуклеиновой кислоты-мишени, количество дуплекса, обеспечивающее изменение сигнала, изменяется, в частности, уменьшается, способом, зависимым от присутствия нуклеиновой кислоты-мишени, таким образом, обеспечивая изменение сигнала. Например, для зонда Инь-Янь, изначально включенного в композицию для детекции нуклеиновой кислоты-мишени, по мере амплификации нуклеиновой кислоты-мишени, одна из двух одиночных цепей, составляющих зонд Инь-Янь, спаривается с амплифицированной нуклеиновой кислотой-мишенью для образования нового дуплекса, и количество зонда Инь-Янь уменьшается, таким образом обеспечивая изменение сигнала в зависимости от присутствия нуклеиновой кислоты-мишени.
[179] В одном варианте осуществления, дуплекс, обеспечивающий изменение сигнала, может представлять собой дуплекс, вновь предоставленный посредством композиции для детекции нуклеиновой кислоты-мишени в ходе инкубации реакции.
[180] В одном варианте осуществления, дуплекс, обеспечивающий изменение сигнала, образованный в ходе инкубации реакции, может быть предоставлен посредством гибридизации между связанным с меткой олигонуклеотидом и нуклеиновой кислотой-мишенью.
[181] Сигналы в результате формирования дуплекса между связанным с меткой олигонуклеотидом и нуклеиновой кислотой-мишенью могут быть образованы посредством различных способов, включая способ скорпиона (Whitcombe et al., Nature Biotechnology 17:804-807 (1999)), способ Sunrise (или Amplifluor) (Nazarenko et al., Nucleic Acids Research, 25(12):2516-2521 (1997), и Патент США No. 6117635), способ LUX (Патент США No. 7537886), способ Plexor (Sherrill CB, et al., Journal of the American Chemical Society, 126:4550-4556 (2004)), способ молекулярного маяка (Tyagi et al., Nature Biotechnology v.14 MARCH 1996), способ Hybeacon (French DJ et al., Mol. Cell Probes, 15(6):363-374 (2001)), способ гибридизации соседних зондов (Bernard P.S. et al., Anal. Biochem., 273:221 (1999)) и способ LNA (Патент США No. 6977295).
[182] В одном варианте осуществления, дуплекс, обеспечивающий изменение сигнала, образованный в ходе инкубации реакции, может представлять собой дуплекс, образованный посредством реакции расщепления, зависимой от присутствия нуклеиновой кислоты-мишени.
[183] Для вышеуказанной реакции, можно использовать 5'-нуклеазу и 3'-нуклеазу, в частности, ДНК-полимеразу, имеющую 5'-нуклеазную активность, ДНК-полимеразу, имеющую 3'-нуклеазную активность, или нуклеазу FEN.
[184] В одном варианте осуществления, изменение сигнала, образуется посредством дуплекса, образованного способом, зависящим от специфического расщепления олигонуклеотида-посредника, гибридизованного с нуклеиновой кислотой-мишенью.
[185] В рамках изобретения, термин «олигонуклеотид-посредник» относится к олигонуклеотиду, опосредующему образование дуплекса, не включающего нуклеиновую кислоту-мишень.
[186] В одном варианте осуществления, только расщепление олигонуклеотида-посредника не образует сигнал, но после гибридизации и расщепления олигонуклеотида-посредника, фрагмент (продукт расщепления), полученный посредством расщепления, вовлекается в непрерывную реакцию для образования сигнала.
[187] В одном варианте осуществления, только гибридизация или расщепление олигонуклеотида-посредника не образует сигнал.
[188] В одном варианте осуществления, олигонуклеотид-посредник включает олигонуклеотид, который гибридизуется с последовательность-мишенью нуклеиновой кислоты и расщепляется для высвобождения фрагмента, таким образом, опосредуя образование дуплекса.
[189] В одном варианте осуществления, фрагмент опосредует образование дуплекса посредством удлинения фрагмента на связывающем олигонуклеотиде.
[190] В соответствии с вариантом осуществления, олигонуклеотид-посредник содержит (i) нацеливающую часть, содержащую гибридизующуюся нуклеотидную последовательность, комплементарную последовательности-мишени нуклеиновой кислоты, и (ii) метящую часть, содержащую нуклеотидную последовательность, не комплементарную последовательности-мишени нуклеиновой кислоты.
[191] В одном варианте осуществления, композиция для детекции нуклеиновой кислоты-мишени может включать метящий олигонуклеотид, который гибридизуется с нуклеиновой кислотой-мишенью, и реакция расщепления, зависимая от присутствия нуклеиновой кислоты-мишени, может включать расщепление метящего олигонуклеотида. Метящий олигонуклеотид соответствует примеру олигонуклеотида-посредника, описанного выше.
[192] В соответствии с вариантом осуществления, расщепление олигонуклеотида-посредника высвобождает фрагмент, и фрагмент специфически гибридизуется со связывающим олигонуклеотидом и удлиняется на связывающем олигонуклеотиде. Когда связывающий олигонуклеотид содержит метку, связывающий олигонуклеотид соответствует примеру связанного с меткой олигонуклеотида, описанного в настоящем описании.
[193] В соответствии с вариантом осуществления, олигонуклеотид-посредник, гибридизованный с последовательностью-мишенью нуклеиновой кислоты, расщепляется и высвобождает фрагмент, фрагмент специфически гибридизуется со связывающим олигонуклеотидом, и фрагмент удлиняется для формирования удлиненной цепи, и это индуцирует формирование удлиненного дуплекса между удлиненной цепью и связывающим олигонуклеотидом, таким образом, обеспечение сигнала, показательного для присутствия последовательности-мишени нуклеиновой кислоты.
[194] В одном варианте осуществления, сигнал, показательный для присутствия нуклеиновой кислоты-мишени, можно обеспечивать посредством (i) по меньшей мере одной метки, связанной с фрагментом и/или связывающим олигонуклеотидом, (ii) метки, встроенной в удлиненный дуплекс в ходе реакции удлинения, и (iii) метки, встроенной в удлиненный дуплекс в ходе реакции удлинения, и метки, связанной с фрагментом и/или связывающим олигонуклеотидом.
[195] В соответствии с вариантом осуществления, когда используют третий олигонуклеотид, включающий гибридизующуюся нуклеотидную последовательность, комплементарную удлиненной цепи, гибридизация удлиненной цепи и третьего олигонуклеотида формирует другой тип дуплекса, таким образом, предоставляя сигнал, указывающий на присутствие нуклеиновой кислоты-мишени (например, PCE-SH). В этом случае, другой тип дуплекса представляет собой дуплекс, предоставляющий изменение сигнала.
[196] В одном варианте осуществления, дуплекс, сформированный посредством реакции расщепления, зависимой от присутствия нуклеиновой кислоты-мишени, не представляет собой дуплекс, обеспечивающий изменение сигнала, но может представлять собой дуплекс, изменяющий содержание дуплекса, обеспечивающего изменение сигнала. В этом случае, дуплекс, обеспечивающий изменение сигнала, был изначально включен в композицию для детекции нуклеиновой кислоты-мишени. В соответствии с вариантом осуществления, когда используют дополнительный олигонуклеотид, включающий гибридизующуюся нуклеотидную последовательность, комплементарную связывающему олигонуклеотиду, дуплекс между дополнительным олигонуклеотидом и связывающим олигонуклеотидом представляет собой дуплекс, обеспечивающий изменение сигнала, и количество дуплекса, обеспечивающего изменение сигнала, изменяется (например, уменьшается) посредством образования дуплекса между удлиненной цепью и связывающим олигонуклеотидом, таким образом обеспечивая изменение сигнала, показательное для присутствия нуклеиновой кислоты-мишени. Например, в способе PCE-NH, дуплекс между дополнительным олигонуклеотидом и связывающим олигонуклеотидом представляет собой дуплекс, обеспечивающий изменение сигнала, который был изначально включен в композицию для детекции нуклеиновой кислоты-мишени, и связывающий олигонуклеотид, составляющий дуплекс, обеспечивающий изменение сигнала, используют для образования нового дуплекса (например, удлиненного дуплекса) посредством реакции расщепления, зависимой от присутствия нуклеиновой кислоты-мишени, таким образом, уменьшая количество дуплекса, обеспечивающего изменение сигнала, для обеспечения изменения сигнала.
[197] В соответствии с вариантом осуществления, фрагмент, удлиненная цепь, связывающий олигонуклеотид, дополнительный олигонуклеотид или их комбинация могут действовать в качестве связанного с меткой олигонуклеотида.
[198] Сигналы посредством дуплекса, сформированного способом, зависимым от расщепления олигонуклеотида-посредника, могут быть образованы посредством различных способов, включая способ PTOCE (расщепления и удлинения PTO) (WO 2012/096523), способ PCE-SH (гибридизации сигнального олигонуклеотида, зависящей от расщепления и удлинения PTO) (WO 2013/115442) и способ PCE-NH (отсутствия гибридизации, зависящего от расщепления и удлинения PTO) (WO 2014/104818).
[199] Применительно к терминам, описанным в вышеуказанных ссылках, соответствующие примеры олигонуклеотидов являются следующими: олигонуклеотид-посредник соответствует PTO (служащему зондом и метящему олигонуклеотиду), связывающий олигонуклеотид соответствует CTO (связывающему и служащему матрицей олигонуклеотиду), и дополнительный олигонуклеотид соответствует SO (сигнальному олигонуклеотиду) или HO (гибридизующемуся олигонуклеотиду). SO, HO, CTO, удлиненная цепь или их комбинация могут играть роль связанного с меткой олигонуклеотида.
[200] В одном варианте осуществления, сигналы посредством дуплекса, сформированного способом, зависимым от расщепления олигонуклеотида-посредника, включают сигнал, обеспечиваемый по мере уменьшения количества других дуплексов посредством дуплекса, сформированного способом, зависимым от расщепления олигонуклеотида-посредника (например, PCE-NH).
[201]
[202] В одном варианте осуществления, когда сигналы посредством дуплекса, образованного способом, зависимым от расщепления олигонуклеотида-посредника, получают посредством способа PTOCE, вышележащий олигонуклеотид, служащий зондом и метящий олигонуклеотид (PTO), содержащий гибридизующуюся нуклеотидную последовательность, комплементарную нуклеиновой кислоте-мишени, связывающий и служащий матрицей олигонуклеотид (CTO), соответствующую метку и зависимую от матрицы ДНК-полимеразу, имеющую 5'-нуклеазную активность, можно включать в реакцию, и композиция для детекции нуклеиновой кислоты-мишени может включать эти компоненты.
[203] PTO содержит (i) 3′-нацеливающую часть, содержащую гибридизующуюся нуклеотидную последовательность, комплементарную последовательности-мишени нуклеиновой кислоты, и (ii) 5′-метящую часть, содержащую нуклеотидную последовательность, не комплементарную последовательности-мишени нуклеиновой кислоты. CTO содержит, в направлении от 3′ до 5′, (i) связывающую часть, содержащую нуклеотидную последовательность, комплементарную 5′-метящей части или части 5′-метящей части PTO, и (ii) служащую матрицей часть, содержащую нуклеотидную последовательность, не комплементарную 5′-метящей части и 3′-нецеливающей части PTO.
[204] Конкретный пример образования сигнала посредством способа PTOCE включает следующие стадии:
[205] (a) гибридизации нуклеиновой кислоты-мишени с вышележащим олигонуклеотидом и PTO; (b) приведение продукта со стадии (a) в контакт с ферментом, имеющим 5’-нуклеазную активность, в условиях для расщепления PTO; где вышележащий олигонуклеотид или его удлиненная цепь индуцирует расщепление PTO посредством фермента, имеющего 5’-нуклеазную активность, таким образом, что расщепление высвобождает фрагмент, содержащий 5’-метящую часть или часть 5’-метящей части PTO; (c) гибридизации фрагмента, высвобожденного из PTO, с CTO; где фрагмент, высвобожденный из PTO, гибридизуется со связывающей частью CTO; (d) проведения реакции удлинения с использованием продукта, полученного на стадии (c), и зависимой от матрицы полимеразы нуклеиновой кислоты; где фрагмент, гибридизованный со связывающей частью CTO, удлиняется с формированием удлиненного дуплекса; где удлиненный дуплекс имеет значение Tm, поддающееся коррекции посредством (i) последовательности и/или длины фрагмента, (ii) последовательности и/или длины CTO, или (iii) последовательности и/или длины фрагмента и последовательности и/или длины CTO; и удлиненный дуплекс обеспечивает сигнал мишени посредством по меньшей мере одной метки, связанной с фрагментом и/или CTO; и (e) детекции удлиненного дуплекса посредством измерения сигнала мишени при предопределенной температуре, при которой удлиненный дуплекс сохраняет свою двухцепочечную форму, где присутствие удлиненного дуплекса указывает на присутствие нуклеиновой кислоты-мишени. В этом случае, способ дополнительно включает повтор всех или части стадий (a)-(e) с включением денатурации между повторяющимися циклами.
[206] В фразе «денатурация между повторяющимися циклами», термин «денатурация» относится к разделению двухцепочечной молекулы нуклеиновой кислоты на одноцепочечные молекулы нуклеиновой кислоты.
[207] На стадии (a) в способе PTOCE, набор праймеров для амплификации нуклеиновой кислоты-мишени можно использовать вместо вышележащего олигонуклеотида. В этом случае, способ дополнительно включает повтор всех или части стадий (a)-(e) с денатурацией между повторяющимися циклами.
[208] Способ PTOCE можно классифицировать как способ, в котором фрагмент PTO, гибридизованный с CTO, удлиняют для формирования удлиненной цепи, и удлиненную цепь детектируют. Способ PTOCE отличается тем, что формирование удлиненной цепи детектируют с использованием дуплекса между удлиненной цепью и CTO.
[209] Существуют другие способы детекции формирования удлиненной цепи. Например, формирование удлиненной цепи можно детектировать с использованием олигонуклеотида, специфически гибридизованного с удлиненной цепью (например, анализ PCE-SH). В таком способе, можно обеспечивать сигнал от (i) метки, связанной с олигонуклеотидом, специфически гибридизованным с удлиненной цепью, или (ii) метки, связанной с олигонуклеотидом, специфически гибридизованным с удлиненной цепью, и метки, связанной с фрагментом PTO.
[210] Альтернативно, детекцию формирования удлиненной цепи проводят посредством других способов (например, анализа PCE-NH) для детекции изменения количества дуплекса между CTO и олигонуклеотидом, способным к специфической гибридизации с CTO. Такое изменение считают показательным для присутствия нуклеиновой кислоты-мишени. Можно обеспечивать сигнал от (i) метки, связанной с олигонуклеотидом, способным к гибридизации с CTO, (ii) метки, связанной с CTO, или (iii) метки, связанной с олигонуклеотидом, способным к гибридизации с CTO, и метки, связанной с CTO.
[211] В соответствии с вариантом осуществления, олигонуклеотид, способный к специфической гибридизации с CTO, имеет перекрывающуюся последовательность с фрагментом PTO.
[212] В соответствии с вариантом осуществления, связанный с меткой олигонуклеотид включает олигонуклеотид, способный к специфической гибридизации с удлиненной цепью (например, анализ PCE-SH), и олигонуклеотид, способный к специфической гибридизации с CTO (например, анализ PCE-NH).
[213] Способ на основе PTOCE, как правило, сопровождается формированием удлиненной цепи, которое является зависимым от присутствия нуклеиновой кислоты-мишени. Термин «способ на основе PTOCE» предназначен, в рамках изобретения, для включения различных способов обеспечения сигналов, включающих формирование удлиненной цепи посредством расщепления и удлинения PTO.
[214] Один пример образования сигнала посредством способа на основе PTOCE включает следующие стадии: (a) гибридизации нуклеиновой кислоты-мишени с вышележащим олигонуклеотидом и PTO; (b) приведение продукта со стадии (a) в контакт с ферментом, имеющим 5’-нуклеазную активность, в условиях для расщепления PTO, где вышележащий олигонуклеотид или его удлиненная цепь индуцирует расщепление PTO посредством фермента, имеющего 5’-нуклеазную активность, и расщепление высвобождает фрагмент, содержащий 5’-метящую часть или часть 5’-метящей части PTO; (c) гибридизации фрагмента, высвобожденного из PTO, с CTO, где фрагмент, высвобожденный из PTO, гибридизуется со связывающей частью CTO; (d) проведения реакции удлинения с использованием продукта со стадии (c) и зависимой от матрицы полимеразы нуклеиновой кислоты, где фрагмент, гибридизованный со связывающей частью CTO, удлиняется, формируя удлиненный дуплекс; и (e) детекции формирования удлиненного дуплекса посредством измерения сигнала, образованного в зависимости от присутствия удлиненной цепи. На стадии (a), набор праймеров для амплификации нуклеиновой кислоты-мишени можно использовать вместо вышележащего олигонуклеотида. В этом случае, способ дополнительно включает повтор всех или части стадий (a)-(e) с денатурацией между повторяющимися циклами.
[215] Другой пример механизмов образования сигнала, зависимых от формирования дуплекса посредством расщепления олигонуклеотида-посредника, представляет собой способ C-метки (Корейский пат. No. 1961642). В этом способе используют праймер (далее в настоящем описании обозначенный как праймер с C-меткой), имеющий структуру, которая последовательно включает случайную последовательность нуклеиновой кислоты, не комплементарную нуклеиновой кислоте-мишени, последовательность узнавания рестрикционного фермента и последовательность нуклеиновой кислоты, комплементарную последовательности-мишени нуклеиновой кислоты, в качестве олигонуклеотида-посредника. Продукт амплификации, сформированный как праймер с C-меткой, гибридизованный с нуклеиновой кислотой-мишенью и удлиненный, включает последовательность узнавания рестрикционного фермента и последовательность, комплементарную случайной последовательности нуклеиновой кислоты. Далее, по мере того, как рестрикционный фермент расщепляет продукт амплификации, таким образом, высвобождая фрагмент метки, комплементарный случайной последовательности нуклеиновой кислоты, и фрагмент метки специфически гибридизуется со связывающим олигонуклеотидом с формированием дуплекса, таким образом предоставляя сигнал, показательный для присутствия последовательности-мишени нуклеиновой кислоты.
[216] В одном варианте осуществления, изменение сигнала можно получать посредством диссоциации дуплекса, сформированного после расщепления меченого олигонуклеотида, гибридизованного с нуклеиновой кислотой-мишенью, которую проводят способом, зависимым от присутствия нуклеиновой кислоты-мишени.
[217] Другой пример механизмов образования сигнала, зависимых от формирования дуплекса посредством расщепления олигонуклеотида-посредника, включает анализ двойного гашения в комбинации с анализом плавления (WO 2016/101959).
[218] Применительно к терминам, описанным в вышеуказанной публикации, соответствующие примеры олигонуклеотидов являются следующими: олигонуклеотид-посредник соответствует служащему зондом и метящему олигонуклеотиду (PTO), и связывающий олигонуклеотид соответствует связывающему и гасящему олигонуклеотиду (CQO).
[219] В одном варианте осуществления, конкретный вариант осуществления образования сигнала посредством анализа двойного гашения, включает следующие стадии:
[220] (a) гибридизации последовательности-мишени нуклеиновой кислоты с PTO; где PTO содержит (i) нацеливающую часть, содержащую нуклеотидную последовательность, по существу комплементарную последовательности-мишени нуклеиновой кислоты, (ii) определяющую температуру плавления область (MTDR), содержащую нуклеотидную последовательность, не комплементарную последовательности-мишени нуклеиновой кислоты, и (iii) по меньшей мере один набор интерактивных меток, содержащий по меньшей мере один флуорофор и по меньшей мере один гаситель;
[221] (b) гибридизации PTO с CQO;
[222] где CQO содержит (i) связывающую часть, содержащую нуклеотидную последовательность, обратно комплементарную MTDR из PTO, и (ii) по меньшей мере одну гасящую молекулу, где MTDR имеет конфигурацию для гибридизации со связывающей частью CQO для формирования дуплекса с меткой;
[223] (c) приведения дуплекса с меткой в контакт с ферментом, имеющим нуклеазную активность; где фермент, имеющий нуклеазную активность, индуцирует расщепление дуплекса с меткой, когда дуплекс с меткой гибридизуется с последовательностью-мишенью нуклеиновой кислоты, таким образом, высвобождая фрагмент активированного дуплекса с меткой, содержащий фрагмент PTO, содержащий MTDR, гибридизованный со связывающей частью CQO, и по меньшей мере один флуорофор;
[224] (d) плавления и/или гибридизации указанного фрагмента активированного дуплекса с меткой для получения сигнала от по меньшей мере одного флуорофора; и
[225] (e) детекции фрагмента активированного дуплекса с меткой посредством измерения сигнала от по меньшей мере одного флуорофора, где сигнал является показателем присутствия последовательности-мишени нуклеиновой кислоты. В одном варианте осуществления, порядок стадий (a) - (c) можно изменять. Например, стадии можно проводить в следующем порядке: гибридизация PTO с CQO (стадия (b)) - гибридизация последовательности-мишени нуклеиновой кислоты с дуплексом с меткой (стадия (a)) - расщепление дуплекса с меткой, гибридизованного с последовательностью-мишенью нуклеиновой кислоты, посредством фермента, имеющего нуклеазную активность (стадия (c)); или гибридизация PTO с последовательностью-мишенью нуклеиновой кислоты (стадия (a)) - высвобождение активированного фрагмента PTO, включающего по меньшей мере один флуорофор и MTDR, посредством фермента, имеющего нуклеазную активность (стадия (c)) - гибридизация активированного PTO с CQO (стадия (b)).
[226] В одном варианте осуществления, дуплекс, обеспечивающий изменение сигнала, может представлять собой дуплекс одиночного типирования или дуплексы множественного типирования. Конкретно, когда дуплекс, обеспечивающий изменение сигнала, представляет собой дуплекс одиночного типирования, количество дуплексов может представлять собой 1, и когда дуплекс, обеспечивающий изменение сигнала, представляет собой дуплексы множественного типирования, количество дуплексов может представлять собой 2, 3, 4, 5, 6, 7, 8, 9, 10, 15 или 20, более конкретно, может представлять собой 2, 3 или 4, и более конкретно, может представлять собой 2 или 3.
[227] В одном варианте осуществления, дуплекс одиночного типирования или любой дуплекс среди дуплексов множественного типирования включает метку.
[228] В одном варианте осуществления, может присутствовать дуплекс, в котором две различные одиночные цепи гибридизуются с различными участками на одной и той же одиночной цепи. Например, как показано на ФИГ. 1c, способ гибридизации соседних зондов, использующий два зонда, представляет собой способ, который обеспечивает сигнал, зависящий от того, гибридизованы ли два зонда в различных участках, соседних друг с другом, на одной цепи (например, нуклеиновой кислоты-мишени). В этом случае, поскольку способ, в соответствии с настоящим изобретением, обеспечивает одинаковый сигнал, когда по меньшей мере один из двух зондов диссоциирует, дуплекс, предоставляющий сигнал, является одиночным, и таким образом, способ, в соответствии с настоящим изобретением, можно классифицировать как способ, предоставляющий дуплекс одиночного типирования.
[229] В одном варианте осуществления, когда дуплекс, обеспечивающий изменение сигнала, представляет собой дуплекс одиночного типирования, количество дуплекса одиночного типирования изменяется, в зависимости от присутствия нуклеиновой кислоты-мишени, таким образом, изменяя сигнал. Например, со ссылкой на чертежи, способ молекулярного маяка на ФИГ. 1a, способ зонд LUX на ФИГ. 1b, способ гибридного зонда на ФИГ. 1c, и способ зонда Инь-Янь на ФИГ. 1d, способ на основе PTOCE на ФИГ. 1e, ФИГ. 1f, ФИГ. 1g, ФИГ. 1h, ФИГ. 2a и ФИГ. 2c, и способ двойного гашения на ФИГ. 2b, представляют собой примеры предоставления дуплекса одиночного типирования.
[230] В частности, в случае ФИГ. 1a, ФИГ. 1b, ФИГ. 1c, ФИГ. 1f, ФИГ. 1g, ФИГ. 1h, ФИГ. 2a, ФИГ. 2b и ФИГ. 2c, дуплекс одиночного типирования образуется в ходе инкубации, и количество дуплекса одиночного типирования увеличивается в ходе инкубации (например, в ходе реакции амплификации), обеспечивая изменение сигнала. В случае ФИГ. 1d и ФИГ. 1e, дуплекс одиночного типирования был изначально включен в композицию для детекции нуклеиновой кислоты-мишени, и количество дуплекса одиночного типирования уменьшается в ходе инкубации реакции, таким образом, обеспечивая изменение сигнала.
[231] В одном варианте осуществления, когда дуплекс представляет собой дуплексы множественного типирования, соотношение количеств между дуплексами множественного типирования изменяется в зависимости от присутствия нуклеиновой кислоты-мишени, таким образом, изменяя сигнал. Например, со ссылкой на чертежи, способ на основе PTOCE на ФИГ. 3a и ФИГ. 3b представляет собой пример предоставления дуплексов множественного типирования (в частности, двух типов дуплексов). На ФИГ. 3a и ФИГ. 3b, один из двух типов дуплексов был изначально включен в композицию для детекции нуклеиновой кислоты-мишени, и другой образуется в ходе инкубации реакции, и в ходе инкубации реакции (например, в ходе реакции амплификации) соотношение количеств между двумя типами дуплексов изменяется, таким образом, обеспечивая изменение сигнала. В частности, по мере того, как количество дуплекса, изначально включенного в композицию для детекции нуклеиновой кислоты-мишени, уменьшается, и количество дуплекса, вновь образованного в ходе инкубации реакции, увеличивается, соотношение количеств между этими дуплексами изменяется.
[232] В одном варианте осуществления, по меньшей мере два из дуплексов множественного типирования включают одну и ту же одиночную цепь. Например, со ссылкой на ФИГ. 3a и ФИГ. 3b, в двух различных типах дуплексов, одна из двух одиночных цепей, составляющих каждый дуплекс, представляет собой одну и ту же цепь. В частности, CTO и PTO (то есть, нерасщепленный PTO), где PTO (или CTO) имеет связанные с ним интерактивные двойные метки, или PTO имеет одну из интерактивных двойных меток, и CTO имеет другую из интерактивных двойных меток, составляют первый дуплекс, который обеспечивает изменение сигнала и был изначально включен в композицию для детекции нуклеиновой кислоты-мишени, и в ходе инкубации реакции, PTO расщепляется в зависимости от присутствия нуклеиновой кислоты-мишени, для высвобождения фрагмента, и высвобожденный фрагмент гибридизуется с CTO для образования удлиненной цепи, и вновь образованная удлиненная цепь и CTO составляют второй дуплекс, обеспечивающий изменение сигнала. В данном случае, первый дуплекс и второй дуплекс включают одну и ту же одиночную цепь (то есть, CTO).
[233] В одном варианте осуществления, значения Tm дуплексов множественного типирования отличаются друг от друга. Например, значения Tm дуплексов отличаются друг от друга по меньшей мере на 2°C, 3°C, 4°C, 5°C, 7°C, 8°C, 9°C, 10°C, 11°C, 12°C, 13°C, 14°C, 15°C или 20°C.
[234] В одном варианте осуществления, количество дуплекса относится к сумме количества дуплекса в состоянии, в котором две цепи нуклеиновой кислоты, составляющие дуплекс, являются диссоциированными (то есть, дуплекс в диссоциированной форме), и количества дуплекса в состоянии, в котором две цепи нуклеиновой кислоты являются гибридизованными (то есть, дуплекс в связанной форме).
[235] В одном варианте осуществления, когда дуплексы множественного типирования включают одну и ту же одиночную цепь, одна и та же одиночная цепь включена в первый дуплекс, изначально включенный в композицию для детекции нуклеиновой кислоты-мишени, и в ходе инкубации реакции, может образовываться новый второй дуплекс, включающий ту же самую одиночную цепь. В этом случае, можно считать, что одна и та же одиночная цепь, включенная в первый дуплекс, расходуется, в то же время образуя второй дуплекс в ходе инкубации реакции, и в результате, количество первого дуплекса, включающего ту же самую одиночную цепь, уменьшается, и количество второго дуплекса, включающего ту же самую одиночную цепь, увеличивается.
[236] В одном варианте осуществления, любая одна из n композиций для детекции нуклеиновых кислот-мишеней может амплифицировать соответствующую нуклеиновую кислоту-мишень.
[237] В одном варианте осуществления, любая из n композиций для детекции нуклеиновых кислот-мишеней может включать амплифицирующий олигонуклеотид, который служит для амплификации соответствующей нуклеиновой кислоты-мишени. В одном варианте осуществления, амплифицирующий олигонуклеотид может являться таким же, как связанный с меткой олигонуклеотид.
[238] В рамках изобретения, термин «амплифицирующий олигонуклеотид» относится к любым олигонуклеотидам, служащим для амплификации нуклеиновых кислот-мишеней.
[239] В одном варианте осуществления, амплифицирующий олигонуклеотид может представлять собой «праймер», известный в данной области. В рамках изобретения, термин «праймер» относится к олигонуклеотиду, который является способным действовать в качестве точки инициации синтеза при помещении в условия, в которых индуцируется синтез продукта удлинения праймера, который является комплементарным цепи нуклеиновой кислоты-мишени (матрице), т.е., в присутствии нуклеотидов и средства для полимеризации, такого как ДНК-полимераза, и при подходящих температуре и pH. Праймер должен являться достаточно длинным, чтобы служить затравкой для синтеза продуктов удлинения в присутствии средства для полимеризации. Подходящая длина праймера определяется множеством факторов, включая температуру, область применения и источник праймера.
[240] Праймер может включать прямой праймер (также обозначенный как вышележащий праймер или вышележащий олигонуклеотид), обратный праймер (также обозначенный как нижележащий праймер или нижележащий олигонуклеотид), или оба. Амплифицирующий олигонуклеотид может представлять собой олигонуклеотид, который имеет структуру, известную в данной области, или может быть синтезирован посредством способа, известного в данной области.
[241] То, что амплифицирующий олигонуклеотид и связанный с меткой олигонуклеотид являются одинаковыми, означает, что один олигонуклеотид одновременно действует в качестве амплифицирующего олигонуклеотида, который осуществляет амплификацию нуклеиновой кислоты-мишени, и в качестве связанного с меткой олигонуклеотида, который образует сигнал в присутствии нуклеиновой кислоты-мишени. В качестве одного примера, связанный с меткой олигонуклеотид может подвергаться гибридизации с нуклеиновой кислотой-мишенью и удлинению, таким образом, образуя сигнал.
[242] Авторы настоящего изобретения обнаружили, что различные механизмы образования сигналов с использованием дуплекса включают температурный диапазон, в котором сигнал изменяется в зависимости от присутствия нуклеиновой кислоты-мишени (то есть, температурный диапазон изменения сигнала), и температурный диапазон, в котором сигнал не изменяется, даже в присутствии нуклеиновой кислоты-мишени (то есть, температурный диапазон постоянного сигнала). Кроме того, авторы настоящего изобретения обнаружили, что способы, не предоставляющие дуплекс, обеспечивающий сигнал, такие как анализ с зондом TaqMan, не имеют температурного диапазона постоянного сигнала.
[243] Кроме того, авторы настоящего изобретения обнаружили, что хорошо известные механизмы образования сигнала можно классифицировать на три типа, в зависимости от количества и/или порядка температурных диапазонов изменения сигнала и температурных диапазонов постоянного сигнала, и посредством использования различных комбинаций этих трех типов механизмов образования сигнала, разработали новый способ, который является способным к детекции множества нуклеиновых кислот-мишеней способом с детекцией в реальном времени, с использованием метки одиночного типирования, без анализа плавления после амплификации.
[244] В одном варианте осуществления, в температурном диапазоне, покрывающем все из n температур детекции, композиция для детекции iй нуклеиновой кислоты-мишени имеет «температурный диапазон изменения сигнала», в котором сигнал изменяется в зависимости от присутствия iй нуклеиновой кислоты-мишени, и «температурный диапазон постоянного сигнала», в котором сигнал является постоянным, даже в присутствии iй нуклеиновой кислоты-мишени.
[245] В одном варианте осуществления, среди n композиций для детекции нуклеиновых кислот-мишеней, композиция для детекции iй нуклеиновой кислоты-мишени может иметь «температурный диапазон изменения сигнала», в котором сигнал изменяется по мере амплификации нуклеиновой кислоты-мишени, и «температурный диапазон постоянного сигнала», в котором сигнал является постоянным, даже по мере амплификации нуклеиновой кислоты-мишени.
[246] В одном варианте осуществления, температурный диапазон изменения сигнала представляет собой температурный диапазон, в котором различие значения сигнала (например, интенсивность сигнала) образуется в зависимости от присутствия нуклеиновой кислоты-мишени.
[247] В одном варианте осуществления, температурный диапазон изменения сигнала представляет собой температурный диапазон, в котором значение сигнала изменяется, в зависимости от уровня амплификации нуклеиновой кислоты-мишени (например, количества амплифицированной нуклеиновой кислоты-мишени).
[248] В одном варианте осуществления, температурный диапазон постоянного сигнала представляет собой температурный диапазон, в котором значение сигнала не изменяется, независимо от присутствия нуклеиновой кислоты-мишени. То есть, температурный диапазон постоянного сигнала представляет собой температурный диапазон, в котором не присутствует различий между значением сигнала в присутствии нуклеиновой кислоты-мишени и значением сигнала в отсутствие нуклеиновой кислоты-мишени.
[249] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени может иметь температурный диапазон изменения сигнала и температурный диапазон постоянного сигнала в непрерывном или прерывистом порядке.
[250] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени может иметь один или два температурных диапазона постоянного сигнала.
[251] В одном варианте осуществления, iя температура детекции может быть выбрана в рамках температурного диапазона изменения сигнала композиции для детекции iй нуклеиновой кислоты-мишени. В этом случае, в настоящем описании, композиция для детекции iй нуклеиновой кислоты-мишени обозначена как имеющая iю температуру детекции. Кроме того, iя нуклеиновая кислота-мишень, соответствующая композиции для детекции iй нуклеиновой кислоты-мишени, может быть обозначена как нуклеиновая кислота-мишень, имеющая iю температуру детекции.
[252] В соответствии с вариантом осуществления, одну температуру детекции, которую определяют посредством композиции для детекции соответствующей нуклеиновой кислоты-мишени, приписывают одной нуклеиновой кислоты-мишени.
[253] В одном варианте осуществления, температурные диапазоны изменения сигнала n композиций для детекции нуклеиновых кислот-мишеней не перекрываются друг с другом. В этом случае, для каждой из n температур детекции, любая температура может быть выбрана в качестве температуры детекции, при условии, что она находится в рамках соответствующего температурного диапазона изменения сигнала.
[254] В одном варианте осуществления, iя температура детекции выбрана в рамках температурного диапазона изменения сигнала композиции для детекции iй нуклеиновой кислоты-мишени, и iя температура детекции не включена в температурные диапазоны изменения сигнала композиций для детекции других нуклеиновых кислот-мишеней.
[255] В одном варианте осуществления, температурный диапазон изменения сигнала любой композиции среди композиций для детекции нуклеиновых кислот-мишеней может перекрываться с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей соседнюю температуру детекции, в то же время, на перекрываясь с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей температуру детекции, не являющуюся соседней с ней. В этом случае, температуру детекции композиции для детекции нуклеиновой кислоты-мишени, которая имеет температурный диапазон изменения сигнала, перекрывающийся с температурным диапазоном изменения сигнала композиции для детекции другой нуклеиновой кислоты-мишени, выбирают в рамках температурного диапазона из температурного диапазона изменения сигнала, который не перекрывается с температурным диапазоном изменения сигнала композиции для детекции другой нуклеиновой кислоты-мишени. Таким образом, посредством выбора температуры детекции в рамках температурного диапазона, который не перекрывается между двумя температурными диапазонами изменения сигнала, только сигнал, показательный для присутствия одной конкретной нуклеиновой кислоты-мишени, можно обеспечивать при одной температуре детекции (см. ФИГ. 4).
[256] В одном варианте осуществления, температурный диапазон изменения сигнала любой из композиции для детекции нуклеиновых кислот-мишеней может перекрываться с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей соседнюю температуру детекции, но любой один из двух температурных диапазонов изменения сигнала не включен полностью в другой температурный диапазон изменения сигнала.
[257] Термин «соседняя температура детекции» использован в настоящем описании для обозначения последовательных температур детекции среди n температур детекции, и например, соседняя температура детекции с iй температурой детекции представляет собой (i-1)ю температуру детекции или (i+1)ю температуру детекции.
[258] В одном варианте осуществления, температурный диапазон изменения сигнала композиции для детекции iй нуклеиновой кислоты-мишени может частично перекрываться с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей соседнюю температуру детекции, в то же время, на перекрываясь с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей температуру детекции, не являющуюся соседней с ней.
[259] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени может иметь один температурный диапазон изменения сигнала и один температурный диапазон постоянного сигнала.
[260] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени может иметь один температурный диапазон изменения сигнала и два температурных диапазона постоянного сигнала.
[261] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени может представлять собой любую из: (i) композиции с нижним изменением сигнала (UnderSC), имеющей характеристику, что температурный диапазон изменения сигнала является более низким, чем температурный диапазон постоянного сигнала; (ii) композиции с верхним изменением сигнала (OverSC), имеющей характеристику, что температурный диапазон изменения сигнала является более высоким, чем температурный диапазон постоянного сигнала; и (iii) композиции с промежуточным изменением сигнала (InterSC), имеющей характеристику, что температурный диапазон изменения сигнала является более высоким, чем один из двух температурных диапазонов постоянного сигнала, и более низкой, чем другой из двух температурных диапазонов постоянного сигнала.
[262] В одном варианте осуществления, выражение «один температурный диапазон является более низким, чем другой температурный диапазон», использованное применительно к температурному диапазону изменения сигнала и температурному диапазону постоянного сигнала композиции для детекции нуклеиновой кислоты-мишени, означает, что самая высокая температура одного температурного диапазона является более низкой, чем самая низкая температура другого температурного диапазона. Кроме того, выражение «один температурный диапазон является более высоким, чем другой температурный диапазон», означает, что самая низкая температура одного температурного диапазона является более высокой, чем самая высокая температура другого температурного диапазона. Например, то, что температурный диапазон постоянного сигнала является более высоким, чем температурный диапазон изменения сигнала, означает, что самая низкая температура в рамках температурного диапазона постоянного сигнала является более высокой, чем самая высокая температура в рамках температурного диапазона изменения сигнала.
[263] В одном варианте осуществления, температурный диапазон изменения сигнала композиции для детекции iй нуклеиновой кислоты-мишени можно определять в зависимости от длины и/или последовательности дуплекса, обеспечивающего изменение сигнала.
[264] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс одиночного типирования, композиция для детекции нуклеиновой кислоты-мишени может иметь один температурный диапазон изменения сигнала и один температурный диапазон постоянного сигнала. Температурный диапазон изменения сигнала и температурный диапазон постоянного сигнала можно определять в зависимости от длины и/или последовательности дуплекса одиночного типирования.
[265] В одном варианте осуществления, когда композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплексы множественного типирования, в частности, два типа дуплексов, композиция для детекции нуклеиновой кислоты-мишени может иметь один температурный диапазон изменения сигнала и два температурных диапазона постоянного сигнала. Температурный диапазон изменения сигнала и температурные диапазоны постоянного сигнала можно определять в зависимости от длин и/или последовательностей двух типов дуплексов.
[266] В любой из n композиций для детекции нуклеиновых кислот-мишеней, в рамках изобретения, можно использовать различные механизмы образования сигналов, описанные выше.
[267] В одном варианте осуществления, вышеописанные различные механизмы образования сигналов можно использовать для любой из композиции UnderSC, композиции InterSC и композиции OverSC.
[268] В одном варианте осуществления, даже когда механизм образования сигнала композиции для детекции нуклеиновой кислоты-мишени является одинаковым, композиции для детекции нуклеиновых кислот-мишеней, включающие олигонуклеотиды различных последовательностей, можно считать отличными друг от друга. Различные композиции для детекции нуклеиновых кислот имеют температуры детекции, отличные друг от друга.
[269] Более конкретно, со ссылкой на чертежи, описаны примеры механизмов образования сигнала, приемлемые для композиции UnderSC, композиции InterSC и композиции OverSC, но без ограничения этим.
[270] (i) Механизм образования сигнала, приемлемый для композиции UnderSC для детекции нуклеиновой кислоты-мишени
[271] На ФИГ. 1a - ФИГ. 1h показаны различные механизмы образования сигналов, приемлемые для композиции UnderSC.
[272] Механизмы образования сигнала на ФИГ. 1a и ФИГ. 1h имеют один температурный диапазон изменения сигнала, в котором сигнал изменяется при амплификации нуклеиновой кислоты-мишени, и один температурный диапазон постоянного сигнала, в котором сигнал является постоянным, даже по мере амплификации нуклеиновой кислоты-мишени, где температурный диапазон изменения сигнала является более низким, чем температурный диапазон постоянного сигнала.
[273] В одном варианте осуществления, как в способе молекулярного маяка на ФИГ. 1a, способе LUX на ФИГ. 1b и способе гибридизации зонда на ФИГ. 1c, различные механизмы образования сигналов, обеспечивающие изменение сигнала от дуплекса (то есть, дуплекса, обеспечивающего изменение сигнала) посредством гибридизации между олигонуклеотидом, связанным с меткой (например, одиночной меткой или интерактивными метками), и нуклеиновой кислотой-мишенью, можно использовать для композиции UnderSC, для детекции нуклеиновой кислоты-мишени. В настоящем описании, дуплексы, обеспечивающие изменения сигнала, представляют собой дуплексы, образованные в ходе инкубации реакции.
[274] В одном варианте осуществления, в качестве механизма образования сигнала для композиции UnderSC, для детекции нуклеиновой кислоты-мишени, можно использовать различные механизмы образования сигналов, такие как показанные в способе зонда Инь-Янь на ФИГ. 1d и способе PCE-NH на ФИГ. 1e, в которых композиция для детекции нуклеиновой кислоты-мишени изначально включает дуплекс обеспечивающий изменение сигнала, и одна из двух одиночных цепей, составляющих дуплекс, обеспечивающий изменение сигнала, спаривается с нуклеиновой кислотой-мишенью, таким образом, формируя новый дуплекс, или новый дуплекс формируется по мере того, как одна из двух одиночных цепей, составляющих дуплекс, обеспечивающий изменение сигнала, спаривается с одной из двух цепей, составляющих дуплекс, сформированный посредством реакции расщепления, зависимой от гибридизации нуклеиновой кислоты-мишени и олигонуклеотида-посредника, так что количество дуплекса, обеспечивающего изменение сигнала, изменяется (в частности, уменьшается), таким образом, обеспечивая изменение сигнала.
[275] В одном варианте осуществления, различные механизмы образования сигналов, такие как показанные в способе на основе PTOCE на ФИГ. 1f, ФИГ. 1g и ФИГ. 1h, в котором обеспечивают изменение сигнала от дуплекса, обеспечивающего изменение сигнала, сформированного посредством реакции расщепления, зависимой от гибридизации олигонуклеотида-посредника и нуклеиновой кислоты-мишени, можно использовать для композиции UnderSC. В данном случае, дуплексы, обеспечивающие изменения сигнала, представляют собой дуплексы, образованные в ходе инкубации реакции, и в частности, метка дуплекса, обеспечивающего изменение сигнала на ФИГ. 1f (то есть, удлиненного дуплекса) встраивается в ходе инкубации реакции.
[276] (ii) Механизм образования сигнала, приемлемый для композиции OverSC, для детекции нуклеиновой кислоты-мишени
[277] На ФИГ. 2a-ФИГ. 2c показаны различные механизмы образования сигналов, приемлемые для композиции OverSC.
[278] Механизмы образования сигнала на ФИГ. 2a и ФИГ. 2c имеют один температурный диапазон изменения сигнала, в котором сигнал изменяется при амплификации нуклеиновой кислоты-мишени, и один температурный диапазон постоянного сигнала, в котором сигнал является постоянным, даже по мере амплификации нуклеиновой кислоты-мишени, где температурный диапазон изменения сигнала является более высоким, чем температурный диапазон постоянного сигнала.
[279] В одном варианте осуществления, различные механизмы образования сигналов, такие как показанные в способах на основе PTOCE на ФИГ. 2a и ФИГ. 2c, и способ двойного гашения на ФИГ. 2b, в котором обеспечивают изменение сигнала от дуплекса, обеспечивающего изменение сигнала, сформированного посредством реакции расщепления, зависимой от гибридизации олигонуклеотида-посредника и нуклеиновой кислоты-мишени , можно использовать для композиции UnderSC. В данном случае, дуплексы, обеспечивающие изменения сигнала, представляют собой дуплексы, образованные в ходе инкубации реакции, и в частности, метка дуплекса, обеспечивающего изменение сигнала на ФИГ. 2c (то есть, удлиненного дуплекса), встраивается в ходе инкубации реакции.
[280] (iii) Механизм образования сигнала, приемлемый для композиции InterSC, для детекции нуклеиновой кислоты-мишени
[281] На ФИГ. 3a - ФИГ. 3b показаны различные механизмы образования сигналов, приемлемые для композиции InterSC.
[282] Механизмы образования сигнала на ФИГ. 3a и ФИГ. 3b имеют один температурный диапазон изменения сигнала, в котором сигнал изменяется по мере амплификации нуклеиновой кислоты-мишени, и два температурных диапазона постоянного сигнала, в котором сигнал является постоянным, даже по мере амплификации нуклеиновой кислоты-мишени, где температурный диапазон изменения сигнала является более высоким, чем один из двух температурных диапазонов постоянного сигнала, и более низким, чем другой из двух температурных диапазонов постоянного сигнала.
[283] В одном варианте осуществления, различные механизмы образования сигналов, обеспечивающие изменение сигнала от дуплекса, предоставляющего множество типов изменения сигнала, можно использовать для композиции InterSC.
[284] Например, как в способах на основе PTOCE, показанных на ФИГ. 3a и ФИГ. 3b, можно использовать механизм, в котором обеспечивают изменение сигнала от двух типов дуплексов, предоставляющих изменение сигнала, и в частности, в способах на ФИГ. 3a и ФИГ. 3b, один из двух типов дуплексов был изначально включен в композицию для детекции нуклеиновой кислоты-мишени, и другой образуется посредством реакции расщепления, зависимой от гибридизации олигонуклеотида-посредника и нуклеиновой кислоты-мишени, и по мере прогрессирования инкубации реакции, соотношение количеств между этими двумя типами дуплексов изменяется, таким образом, обеспечивая изменение сигнала. В настоящем описании, два типа дуплексов имеют отличные друг от друга значения Tm.
[285] В одном варианте осуществления, множество дуплексов могут иметь отличные друг от друга значения Tm.
[286] В одном варианте осуществления, композиция InterSC имеет один температурный диапазон изменения сигнала, в котором сигнал изменяется в зависимости от присутствия нуклеиновой кислоты-мишени, и два температурных диапазона постоянного сигнала, в которых сигнал является постоянным, даже в присутствии нуклеиновой кислоты-мишени, где температурный диапазон изменения сигнала является более высоким, чем один из двух температурных диапазонов постоянного сигнала, и более низким, чем другой из двух температурных диапазонов постоянного сигнала.
[287] В одном варианте осуществления, композиция InterSC предоставляет дуплексы множественного типирования, обеспечивающие изменение сигнала.
[288] В одном варианте осуществления, дуплексы множественного типирования имеют отличные друг от друга значения Tm.
[289] В одном варианте осуществления, температурный диапазон изменения сигнала и температурные диапазоны постоянного сигнала композиции InterSC можно корректировать посредством коррекции значений Tm дуплексов множественного типирования, и нуклеиновую кислоту-мишень можно детектировать с использованием температуры детекции, выбранной из температурного диапазона изменения сигнала.
[290] В одном варианте осуществления, соотношение количеств между дуплексами множественного типирования изменяется в зависимости от присутствия нуклеиновой кислоты-мишени, таким образом, изменяя сигнал.
[291] В одном варианте осуществления, композиция InterSC предоставляет два дуплекса, обеспечивающих изменение сигнала. В зависимости от присутствия нуклеиновой кислоты-мишени, соотношение количеств между двумя дуплексами изменяется, приводя к изменению сигнала. Два дуплекса имеют различные значения Tm.
[292] В одном варианте осуществления, один (например, дуплекс с относительно низкой Tm) из двух дуплексов, обеспечивающих изменение сигнала, изначально включен в композицию InterSC, и другой (например, дуплекс с относительно высокой Tm) образуется в ходе инкубации реакции.
[293] В одном варианте осуществления, одна из двух одиночных цепей, составляющих каждый дуплекс, в двух дуплексах представляет собой одну и ту же цепь. В частности, по мере того, как количество дуплекса, изначально включенного в композицию для детекции нуклеиновой кислоты-мишени, уменьшается, и количество дуплекса, вновь образованного в ходе инкубации реакции увеличивается, соотношение количеств между этими дуплексами изменяется. То есть, количество первого дуплекса, изначально включенного в композицию InterSC, из двух дуплексов, содержащих одну и ту же цепь, уменьшается посредством второго дуплекса, вновь образованного в ходе инкубации реакции, и в ходе инкубации, количество второго дуплекса увеличивается, и соотношение количеств между ними изменяется.
[294] В одном варианте осуществления, любой из дуплексов, предоставленных посредством композиции InterSC, представляет собой дуплекс, предоставленный посредством реакции расщепления, зависимой от присутствия нуклеиновой кислоты-мишени. Например, дуплекс, который является вновь образованным в ходе инкубации реакции, может являться образованным посредством реакции расщепления (например, способом на основе PTOCE), зависимой от присутствия нуклеиновой кислоты-мишени.
[295] В одном варианте осуществления, каждый из двух дуплексов содержит интерактивные двойные метки (например, репортерную молекулу и молекулу гасителя). В частности, (i) когда два дуплекса находятся в связанной форме, молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы, и когда два дуплекса находятся в диссоциированной форме, молекула гасителя отделена от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы (см. ФИГ. 3a), или (ii) когда два дуплекса находятся в связанной форме, молекула гасителя отделена от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы, и когда два дуплекса находятся в диссоциированной форме, молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы.(см. ФИГ. 3b)
[296] Механизмы образования сигнала, приемлемые для композиции InterSC, можно использовать для детекции одной нуклеиновой кислоты-мишени, так же как можно использовать для детекции множества нуклеиновых кислот-мишеней посредством использования множества композиций InterSC и коррекции каждого температурного диапазона изменения сигнала множества композиций InterSC.
[297] Как описано выше, композиции для детекции нуклеиновых кислот-мишеней, даже когда их механизм образования сигнала является одинаковым, могут иметь различный порядок температурного диапазона изменения сигнала и температурного диапазона постоянного сигнала, в зависимости от (i) типа метки (например, одиночная метка, интерактивные метки и т.д.), (ii) типа связанного с меткой олигонуклеотида (например, PTO, CTO и т.д.), и (iii) локализации, в которой связана метка, и/или способа, посредством которого связана метка (была ли метка изначально связана с олигонуклеотидом до инкубации реакции, или связана с олигонуклеотидом посредством встраивания в него в ходе инкубации реакции и т.д.), и т.п. Соответственно, одинаковый механизм образования сигнала можно использовать для любой из композиции UnderSC, композиции InterSC и композиции OverSC. Например, в способе на основе PTOCE, механизм, посредством которого присутствие нуклеиновой кислоты-мишени определяют по сигналу от удлинения и расщепления PTO, может являться одинаковым, но случай, в котором интерактивные двойные метки все являются связанными с CTO (например, ФИГ. 1g, ФИГ. 1h) и случай, в котором одна из интерактивных меток является связанной с PTO, и другая из интерактивных меток является связанной с CTO (например, ФИГ. 3a), можно использовать в качестве механизма образования сигнала для композиции UnderSC и композиции InterSC, соответственно. Кроме того, случай, в котором интерактивные двойные метки встроены (например, ФИГ. 2c) и случай, в котором она изначально связана с олигонуклеотидом (например, ФИГ. 3b), можно использовать в качестве механизмов образования сигнала для композиции OverSC и композиции InterSC для детекции нуклеиновой кислоты-мишени, соответственно.
[298] В конкретном варианте осуществления, как в вышеописанном способе на основе PTOCE, конкретно, в способе на основе PTOCE, в котором CTO, со связанными с ним интерактивные двойными метками, формирует дуплекс с удлиненной цепью фрагмента PTO, расщепленного в зависимости от присутствия нуклеиновой кислоты-мишени, таким образом, обеспечивая сигнал, можно принимать в качестве механизма образования сигнала для композиции UnderSC (см. ФИГ. 5). В данном случае, температурный диапазон изменения сигнала и температурный диапазон постоянного сигнала композиции UnderSC можно контролировать посредством коррекции значения Tm удлиненного дуплекса (то есть, дуплекса между связанным с интерактивной двойной меткой CTO и удлиненной цепью).
[299] В конкретном варианте осуществления, вышеописанный способ двойного гашения можно принимать в качестве механизма образования сигнала для композиции OverSC (см. ФИГ. 7). В данном случае, температурный диапазон изменения сигнала и температурный диапазон постоянного сигнала композиции OverSC можно контролировать посредством коррекции значения Tm дуплекса с меткой (то есть, фрагмента активированного дуплекса с меткой).
[300] В конкретном варианте осуществления, вышеописанный способ на основе PTOCE, конкретно, способ на основе PTOCE с использованием PTO со связанной с ним одной из интерактивных двойных меток и CTO со связанной с ним другой из интерактивных двойных меток, можно принимать в качестве механизма образования сигнала для композиции InterSC (см. ФИГ. 6). В данном случае, температурный диапазон изменения сигнала и температурный диапазон постоянного сигнала композиции InterSC можно контролировать посредством коррекции значения Tm дуплекса между CTO и метящей частью нерасщепленного PTO, и значения Tm удлиненного дуплекса между CTO и удлиненной цепью фрагмента PTO, расщепленного, в зависимости от присутствия нуклеиновой кислоты-мишени.
[301] В одном варианте осуществления, когда n представляет собой 2, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию UnderSC или композицию InterSC, и композиция для детекции второй нуклеиновой кислоты-мишени может представлять собой композицию InterSC или композицию OverSC.
[302] В конкретном варианте осуществления, когда n представляет собой 2, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию UnderSC, и композиция для детекции второй нуклеиновой кислоты-мишени может представлять собой композицию InterSC. Например, как показано на ФИГ. 8, для механизма образования сигнала композиции для детекции первой нуклеиновой кислоты-мишени, можно использовать способ на основе PTOCE, в котором используют CTO, связанный с интерактивными двойными метками, и для механизма образования сигнала композиции для детекции второй нуклеиновой кислоты-мишени, можно использовать способ на основе PTOCE, в котором одна из интерактивных двойных меток является связанной с PTO, и другая является связанной с CTO. В одном варианте осуществления, первую температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой дуплекс, предоставленный посредством композиции для детекции первой нуклеиновой кислоты-мишени (то есть, удлиненный дуплекс), и два типа дуплексов, предоставленных посредством композиции для детекции второй нуклеиновой кислоты-мишени (то есть, дуплекс между нерасщепленным PTO и CTO, и удлиненный дуплекс), все находятся в связанной форме, и вторую температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой удлиненный дуплекс, предоставленный посредством композиции для детекции первой нуклеиновой кислоты-мишени, и дуплекс между нерасщепленным PTO и CTO, предоставленный посредством композиции для детекции второй нуклеиновой кислоты-мишени, находятся в диссоциированной форме, и при которой удлиненный дуплекс, предоставленный посредством композиции для детекции второй нуклеиновой кислоты-мишени, находится в связанной форме. Композиция для детекции первой нуклеиновой кислоты-мишени, в присутствии первой нуклеиновой кислоты-мишени, обеспечивает изменение сигнала при первой температуре детекции (то есть, по мере того, как удлиненный дуплекс в связанной форме образуется при амплификации нуклеиновой кислоты-мишени, так что молекула гасителя отделяется от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы), и обеспечивает постоянный сигнал при второй температуре детекции (то есть, удлиненный дуплекс, даже когда образовался, полностью находится в диссоциированной форме, так что молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы), и композиция для детекции второй нуклеиновой кислоты-мишени, в присутствии второй нуклеиновой кислоты-мишени, обеспечивает постоянный сигнал при первой температуре детекции (то есть, два типа дуплексов оба находятся в связанной форме, так что молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы) и обеспечивает изменение сигнала при второй температуре детекции (дуплекс между нерасщепленным PTO и CTO расходуется, и удлиненный дуплекс в связанной форме образуется, так что молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы).
[303] В конкретном варианте осуществления, когда n представляет собой 2, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию UnderSC, и композиция для детекции второй нуклеиновой кислоты-мишени может представлять собой композицию OverSC. Например, как показано на ФИГ. 9, для механизма образования сигнала композиции для детекции первой нуклеиновой кислоты-мишени, можно использовать способ на основе PTOCE с использованием CTO, связанного с интерактивными двойными метками, и для механизма образования сигнала композиции для детекции второй нуклеиновой кислоты-мишени, можно использовать способ двойного гашения. В одном варианте осуществления, первую температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой дуплекс, предоставленный посредством композиции для детекции первой нуклеиновой кислоты-мишени (то есть, удлиненный дуплекс), и дуплекс, предоставленный посредством композиции для детекции второй нуклеиновой кислоты-мишени (то есть, дуплекс с меткой), находятся в связанной форме, и вторую температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой удлиненный дуплекс и дуплекс с меткой оба находятся в диссоциированной форме. В этом случае, композиция для детекции первой нуклеиновой кислоты-мишени, в присутствии первой нуклеиновой кислоты-мишени, обеспечивает изменение сигнала при первой температуре детекции (то есть, удлиненный дуплекс в связанной форме образуется при амплификации нуклеиновой кислоты-мишени, так что молекула гасителя отделяется от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы) и обеспечивает постоянный сигнал при второй температуре детекции (то есть, удлиненный дуплекс, даже когда образовался, полностью находится в диссоциированной форме, так что молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы); композиция для детекции второй нуклеиновой кислоты-мишени, в присутствии второй нуклеиновой кислоты-мишени, обеспечивает постоянный сигнал при первой температуре детекции (то есть, в дуплексе с меткой в связанной форме (нерасщепленном дуплексе с меткой и фрагменте активированного дуплекса с меткой), по меньшей мере один из двух гасителей находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы) и обеспечивает изменение сигнала при второй температуре детекции (то есть, фрагмент активированного дуплекса с меткой, образованный при амплификации нуклеиновой кислоты-мишени, находится в диссоциированной форме, так что обе из двух молекул гасителя отделены от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы).
[304] В конкретном варианте осуществления, когда n представляет собой 2, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию InterSC, и композиция для детекции второй нуклеиновой кислоты-мишени может представлять собой композицию InterSC. Например, как показано на ФИГ. 10, в качестве механизма образования сигнала композиции для детекции первой нуклеиновой кислоты-мишени и композиции для детекции второй нуклеиновой кислоты-мишени, можно использовать способ на основе PTOCE, в котором одна из интерактивных двойных меток является связанной с PTO, и другая - с CTO. В одном варианте осуществления, первую температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой, среди двух типов дуплексов, предоставленных посредством композиции для детекции первой нуклеиновой кислоты-мишени (то есть, удлиненного дуплекса и дуплекса между нерасщепленным PTO и CTO), и двух типов дуплексов, предоставленных посредством композиции для детекции второй нуклеиновой кислоты-мишени (то есть, удлиненного дуплекса и дуплекса между нерасщепленным PTO и CTO), все другие дуплексы, за исключением дуплекса между нерасщепленным PTO и CTO, предоставленного посредством композиции для детекции первой нуклеиновой кислоты-мишени, находятся в связанной форме, и дуплекс между нерасщепленным PTO и CTO, предоставленный посредством композиции для детекции первой нуклеиновой кислоты-мишени, находится в диссоциированной форме; и вторую температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой, среди двух типов дуплексов, предоставленных посредством композиции для детекции первой нуклеиновой кислоты-мишени, и двух типов дуплексов, предоставленных посредством композиции для детекции второй нуклеиновой кислоты-мишени, только удлиненный дуплекс, предоставленный посредством композиции для детекции второй нуклеиновой кислоты-мишени, находится в связанной форме, и остальные из других дуплексов все находятся в диссоциированной форме. Композиция для детекции первой нуклеиновой кислоты-мишени, в присутствии первой нуклеиновой кислоты-мишени, обеспечивает изменение сигнала при первой температуре детекции (то есть, по мере того, как удлиненный дуплекс в связанной форме образуется при амплификации нуклеиновой кислоты-мишени, молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы), и обеспечивает постоянный сигнал при второй температуре детекции (то есть, удлиненный дуплекс, даже когда образовался, полностью находится в диссоциированной форме, так что молекула гасителя отделена от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы); и композиция для детекции второй нуклеиновой кислоты-мишени, в присутствии второй нуклеиновой кислоты-мишени, обеспечивает постоянный сигнал при первой температуре детекции (то есть, два типа дуплексов оба находятся в связанной форме, так что молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы) и обеспечивает изменение сигнала при второй температуре детекции (по мере того, как дуплекс между нерасщепленным PTO и CTO расходуется, и удлиненный дуплекс в связанной форме образуется, молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы).
[305] В конкретном варианте осуществления, когда n представляет собой 2, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию InterSC, и композиция для детекции второй нуклеиновой кислоты-мишени может представлять собой композицию OverSC. Например, как показано на ФИГ. 11, для механизма образования сигнала композиции для детекции первой нуклеиновой кислоты-мишени, можно использовать способ на основе PTOCE, в котором одна из двойных меток является связанной с PTO и другая - с CTO, и для механизма образования сигнала композиции для детекции второй нуклеиновой кислоты-мишени, можно использовать способ двойного гашения. В одном варианте осуществления, первую температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой, среди двух типов дуплексов, предоставленных посредством композиции для детекции первой нуклеиновой кислоты-мишени (то есть, удлиненного дуплекса и дуплекса между нерасщепленным PTO и CTO), и фрагмента активированного дуплекса с меткой, предоставленного посредством композиции для детекции второй нуклеиновой кислоты-мишени, все другие дуплексы, за исключением дуплекса между нерасщепленным PTO и CTO, предоставленного посредством композиции для детекции первой нуклеиновой кислоты-мишени, все находятся в связанной форме, и дуплекс между нерасщепленным PTO и CTO, предоставленный посредством композиции для детекции первой нуклеиновой кислоты-мишени, находится в диссоциированной форме; и вторую температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой два типа дуплексов, предоставленных посредством композиции для детекции первой нуклеиновой кислоты-мишени (удлиненный дуплекс и дуплекс между нерасщепленным PTO и CTO), и фрагмент активированного дуплекса с меткой, предоставленного посредством композиции для детекции второй нуклеиновой кислоты-мишени, все находятся в диссоциированной форме. В этом случае, композиция для детекции первой нуклеиновой кислоты-мишени, в присутствии первой нуклеиновой кислоты-мишени, обеспечивает изменение сигнала при первой температуре детекции (то есть, по мере того, как удлиненный дуплекс в связанной форме образуется при амплификации нуклеиновой кислоты-мишени, молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы) и обеспечивает постоянный сигнал при второй температуре детекции (то есть, удлиненный дуплекс, даже когда образовался, полностью находится в диссоциированной форме, так что молекула гасителя отделена от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы); композиция для детекции второй нуклеиновой кислоты-мишени, в присутствии второй нуклеиновой кислоты-мишени, обеспечивает постоянный сигнал при первой температуре детекции (то есть, в дуплексе с меткой в связанной форме (нерасщепленном дуплексе с меткой и фрагменте активированного дуплекса с меткой), по меньшей мере один из двух гасителей находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы), и обеспечивает изменение сигнала при второй температуре детекции (то есть, фрагмент активированного дуплекса с меткой, образованный при амплификации нуклеиновой кислоты-мишени, находится в диссоциированной форме, так что обе из двух молекул гасителя отделены от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы).
[306] В одном варианте осуществления, когда n представляет собой 3 или более, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию UnderSC или композицию InterSC, композиция для детекции nй нуклеиновой кислоты-мишени может представлять собой композицию InterSC или композицию OverSC, и композиция(и) для детекции нуклеиновых кислот-мишеней, отличных от первой нуклеиновой кислоты-мишени и nй нуклеиновой кислоты-мишени, может представлять собой композицию InterSC.
[307] В конкретном варианте осуществления, когда n представляет собой 3, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию UnderSC, композиция для детекции второй нуклеиновой кислоты-мишени может представлять собой композицию InterSC, и композиция для детекции третьей нуклеиновой кислоты-мишени может представлять собой композицию OverSC. Например, как показано на ФИГ. 12, для механизма образования сигнала композиции для детекции первой нуклеиновой кислоты-мишени, можно использовать способ на основе PTOCE, в котором используют CTO, связанный с интерактивными двойными метками, и для механизма образования сигнала композиции для детекции второй нуклеиновой кислоты-мишени, можно использовать способ на основе PTOCE, в котором одна из интерактивных двойных меток является связанной с PTO, и другая является связанной с CTO, и для механизма образования сигнала композиции для детекции третьей нуклеиновой кислоты-мишени, можно использовать способ двойного гашения. В одном варианте осуществления, первую температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой дуплексы (в частности, дуплексы, обеспечивающие изменения сигнала) предоставленные посредством композиций для детекции первой - третьей нуклеиновых кислот-мишеней, все находятся в связанной форме, вторую температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой удлиненный дуплекс, предоставленный посредством композиции для детекции первой нуклеиновой кислоты-мишени, и дуплекс между нерасщепленным PTO и CTO, предоставленный посредством композиции для детекции второй нуклеиновой кислоты-мишени, находятся в диссоциированной форме, и другие дуплексы (то есть, удлиненные дуплексы, предоставленные посредством композиции для детекции второй нуклеиновой кислоты-мишени, и дуплексы с меткой, предоставленные посредством композиции для детекции третьей нуклеиновой кислоты-мишени) находятся в связанной форме, и третью температуру детекции можно выбирать, чтобы она представляла собой температуру, при которой все дуплексы находятся в диссоциированной форме. В таком случае, композиция для детекции первой нуклеиновой кислоты-мишени, в присутствии первой нуклеиновой кислоты-мишени, обеспечивает изменение сигнала при первой температуре детекции (то есть, удлиненный дуплекс в связанной форме образуется при амплификации нуклеиновой кислоты-мишени, так что молекула гасителя отделяется от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы), и обеспечивает постоянный сигнал при второй и третьей температурах детекции (то есть, удлиненный дуплекс, даже когда образовался, полностью находится в диссоциированной форме, так что молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы); композиция для детекции второй нуклеиновой кислоты-мишени, в присутствии второй нуклеиновой кислоты-мишени, обеспечивает постоянный сигнал при первой и третьей температурах детекции (то есть, два типа дуплексов оба находятся в связанной форме, так что молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы, или два типа дуплексов оба находятся в диссоциированной форме, так что молекула гасителя отделяется от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы), и обеспечивает изменение сигнала при второй температуре детекции (по мере того, как дуплекс между нерасщепленным PTO и CTO расходуется, и удлиненный дуплекс в связанной форме образуется, молекула гасителя находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы); и композиция для детекции третьей нуклеиновой кислоты-мишени, в присутствии третьей нуклеиновой кислоты-мишени, обеспечивает постоянный сигнал при первой и второй температурах детекции (то есть, в дуплексе с меткой в связанной форме (нерасщепленном дуплексе с меткой и фрагменте активированного дуплекса с меткой) по меньшей мере один из двух гасителей находится в тесной близости с репортерной молекулой, таким образом, гася сигнал от репортерной молекулы), и обеспечивает изменение сигнала при третьей температуре детекции (то есть, активированный дуплекс с меткой, образованный в результате амплификация нуклеиновой кислоты-мишени, находится в диссоциированной форме, так что две молекулы гасителя обе отделены от репортерной молекулы, таким образом, не гася сигнал от репортерной молекулы).
[308] В конкретном варианте осуществления, когда n представляет собой 3, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию UnderSC, композиция для детекции второй нуклеиновой кислоты-мишени может представлять собой композицию InterSC, и композиция для детекции третьей нуклеиновой кислоты-мишени может представлять собой композицию InterSC.
[309] В конкретном варианте осуществления, когда n представляет собой 3, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию InterSC, композиция для детекции второй нуклеиновой кислоты-мишени может представлять собой композицию InterSC, и композиция для детекции третьей нуклеиновой кислоты-мишени может представлять собой композицию OverSC.
[310] В конкретном варианте осуществления, когда n представляет собой 3, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию InterSC, композиция для детекции второй нуклеиновой кислоты-мишени может представлять собой композицию InterSC, и композиция для детекции третьей нуклеиновой кислоты-мишени может представлять собой композицию InterSC.
[311] В конкретном варианте осуществления, когда n представляет собой 4, композиция для детекции первой нуклеиновой кислоты-мишени может представлять собой композицию UnderSC или InterSC, композиция для детекции второй нуклеиновой кислоты-мишени и композиция для детекции третьей нуклеиновой кислоты-мишени обе могут представлять собой композицию InterSC, и композиция для детекции четвертой нуклеиновой кислоты-мишени может представляет собой композицию InterSC или OverSC. В способе, в соответствии с настоящим изобретением, используют тот факт, что температурный диапазон изменения сигнала существует в зависимости от механизма образования сигнала композиции для детекции нуклеиновой кислоты-мишени.
[312] В одном варианте осуществления, температуры детекции в соответствии с настоящим изобретением можно предварительно определять в свете температурного диапазона изменения сигнала каждой из n композиций для детекции нуклеиновых кислот-мишеней.
[313] В одном варианте осуществления, температурный диапазон изменения сигнала любой из n композиций для детекции нуклеиновых кислот-мишеней можно определять в зависимости от длины и/или последовательности дуплекса. То есть, посредством коррекции значения Tm дуплекса, температурный диапазон изменения сигнала можно предварительно определять.
[314] В одном варианте осуществления, когда изменение сигнала образуется посредством связанного с меткой олигонуклеотида, который специфически гибридизуется с нуклеиновой кислотой-мишенью (например, зонда LUX, зонда на основе молекулярного маяка, зонда HyBeacon, гибридизующихся соседних зондов и т.д.), детекции сигналов можно достигать при предопределенной температуре детекции посредством коррекции значения Tm связанного с меткой олигонуклеотида.
[315] В одном варианте осуществления, когда праймер скорпион используют в качестве связанного с меткой олигонуклеотида, детекцию сигналов успешно проводят при предопределенной температуре, посредством коррекции значения Tm части, которая гибридизуется с удлиненной цепью.
[316] В одном варианте осуществления, когда сигнал образуется посредством дуплекса, сформированного на основании присутствия последовательности-мишени нуклеиновой кислоты, детекцию сигналов успешно проводят при предопределенной температуре посредством коррекции значения Tm дуплекса. Например, когда сигнал, образуется посредством способа PTOCE, детекции сигналов успешно достигают при предопределенной температуре посредством коррекции значения Tm удлиненного дуплекса, сформированного посредством удлинения фрагмента PTO на CTO.
[317] Способ на основе PTOCE имеет то преимущество, что является простым контроль значения Tm дуплекса или значения Tm третьего продукта гибридизации, на гибридизацию которого влияет дуплекс.
[318] Как описано выше, температуру детекции определяют с учетом температурного диапазона изменения сигнала, который изменяется, в зависимости от дуплекса, предоставленного посредством композиции для детекции нуклеиновой кислоты-мишени.
[319] В одном варианте осуществления, температуру детекции любой из n композиций для детекции n нуклеиновых кислот-мишеней можно предварительно определять в рамках температурного диапазона изменения сигнала, который не перекрывается с температурным диапазоном изменения сигнала других композиций (см. ФИГ. 4).
[320] В одном варианте осуществления, температуры детекции, приписанные композициям для детекции нуклеиновых кислот-мишеней, отличаются друг от друга на по меньшей мере 2°C, 3°C, 4°C, 5°C, 7°C, 8°C, 9°C, 10°C, 11°C, 12°C, 15°C или 20°C, или более.
[321] В одном варианте осуществления, n температур детекции можно выбирать из температурного диапазона 45°C-97°C, 45°C-96°C, 45°C-95°C, 45°C-94°C, 45°C-93°C, 45°C-92°C, 45°C-91°C, 45°C-90°C, 46°C-97°C, 46°C-96°C, 46°C-95°C, 46°C-94°C, 46°C-93°C, 46°C-92°C, 46°C-91°C, 46°C-90°C, 47°C-97°C, 47°C-96°C, 47°C-95°C, 47°C-94°C, 47°C-93°C, 47°C-92°C, 47°C-91°C, 47°C-90°C, 48°C-97°C, 48°C-96°C, 48°C-95°C, 48°C-94°C, 48°C-93°C, 48°C-92°C, 48°C-91°C, 48°C-90°C, 49°C-97°C, 49°C-96°C, 49°C-95°C, 49°C-94°C, 49°C-93°C, 49°C-92°C, 49°C-91°C, 49°C-90°C, 50°C-97°C, 50°C-96°C, 50°C-95°C, 50°C-94°C, 50°C-93°C, 50°C-92°C, 50°C-91°C или 50°C-90°C.
[322] Например, самую высокую температуру детекции (то есть, nю температуру детекции) среди вышеуказанных температур детекции можно выбирать из температурного диапазона 70°C-97°C, 70°C-95°C, 70°C-93°C, 70°C-90°C, 73°C-97°C, 73°C-95°C, 73°C-93°C, 73°C-90°C, 75°C-97°C, 75°C-95°C, 75°C-93°C, 75°C-90°C, 78°C-97°C, 78°C-95°C, 78°C-93°C, 78°C-90°C, 80°C-97°C, 80°C-95°C, 80°C-93°C, 80°C-90°C, 83°C-97°C, 83°C-95°C, 83°C-93°C, 83°C-90°C, 85°C-97°C, 85°C-95°C, 85°C-93°C или 85°C-90°C.
[323] Например, самую низкую температуру детекции (то есть, первую температуру детекции) среди n температур детекции можно выбирать из температурного диапазона 45°C-70°C, 45°C-68°C, 45°C-65°C, 45°C-63°C, 45°C-60°C, 45°C-58°C, 45°C-55°C, 48°C-70°C, 48°C-68°C, 48°C-65°C, 48°C-63°C, 48°C-60°C, 48°C-58°C, 48°C-55°C, 50°C-70°C, 50°C-68°C, 50°C-65°C, 50°C-63°C, 50°C-60°C, 50°C-58°C или 50°C-55°C.
[324] Например, промежуточную температуру детекции (например, от второй температуры детекции до (n-1)й температуры детекции) среди n температур детекции можно выбирать из температурного диапазона 55°C-85°C, 55°C-83°C, 55°C-80°C, 55°C-78°C, 55°C-7,5°C, 55°C-73°C, 55°C-70°C, 55°C-68°C, 55°C-65°C, 55°C-63°C, 55°C-60°C, 58°C-85°C, 58°C-83°C, 58°C-80°C, 58°C-78°C, 58°C-75°C, 58°C-73°C, 58°C-70°C, 58°C-68°C, 58°C-65°C, 58°C-63°C, 58°C-60°C, 60°C-85°C, 60°C-83°C, 60°C-80°C, 60°C-78°C, 60°C-75°C, 60°C-73°C, 60°C-70°C, 60°C-68°C, 60°C-65°C, 60°C-63°C, 63°C-85°C, 63°C-83°C, 63°C-80°C, 63°C-78°C, 63°C-75°C, 63°C-73°C, 63°C-70°C, 63°C-68°C, 63°C-65°C, 65°C-85°C, 65°C-83°C, 65°C-80°C, 65°C-78°C, 65°C-75°C, 65°C-73°C, 65°C-70°C, 65°C-68°C, 68°C-85°C, 68°C-83°C, 68°C-80°C, 68°C-78°C, 68°C-75°C, 68°C-73°C, 68°C-70°C, 70°C-85°C, 70°C-83°C, 70°C-80°C, 70°C-78°C, 70°C-75°C или 70°C-73°C.
[325] В соответствии с вариантом осуществления, n нуклеиновых кислот-мишеней, соответственно, приписывают n температурам детекции, n композиций для детекции n нуклеиновых кислот-мишеней, подходящих для температур детекции, получают, и затем можно проводить стадию (a).
[326] В одном варианте осуществления, когда n представляет собой 3, первую температуру детекции можно выбирать из температурного диапазона 50°C-60°C, вторую температуру детекции можно выбирать из температурного диапазона 65°C-75°C, и третью температуру детекции можно выбирать из температурного диапазона 80°C-95°C.
[327] На стадии (a), сигналы детектируют при n температурах детекции в ходе инкубации.
[328] В одном варианте осуществления, детекцию сигналов можно проводить в каждом цикле или избранных циклах, или в конечной точке реакции.
[329] В одном варианте осуществления, детекцию сигналов можно проводить по меньшей мере в одном цикле. Например, сигналы можно детектировать при n температурах детекции в одном избранном цикле, или можно детектировать при n температурах детекции в каждом из двух избранных циклов. Например, когда n представляет собой 3, и сигналы детектируют в цикле 1 и цикле 30, сигналы (то есть, первый сигнал, второй сигнал и третий сигнал) детектируют при первой температуре детекции, второй температуре детекции и третьей температуре детекции в цикле 1, и сигналы детектируют при первой температуре детекции, второй температуре детекции и третьей температуре детекции в цикле 30.
[330] В одном варианте осуществления, детекцию сигналов можно проводить по меньшей мере в двух циклах.
[331] В одном варианте осуществления, изменение сигнала можно измерять с использованием сигналов, детектированных по меньшей мере в двух циклах. Например, амплификацию нуклеиновых кислот можно проводить на протяжении 30 циклов, 40 циклов, 45 циклов или 50 циклов ПЦР, и в каждом цикле, сигналы можно измерять при n температурах детекции. Затем, значения сигналов, детектированные при каждой температуре детекции в множестве циклов, можно изображать в форме кривой амплификации (набора точек данных циклов и RFU в циклах) при каждой температуре детекции. В качестве конкретного примера, когда n представляет собой 3, когда каждая нуклеиновая кислота-мишень присутствует, можно получать кривую амплификации при первой температуре детекции, кривую амплификации при второй температуре детекции и кривую амплификации при третьей температуре детекции, и изменение сигнала можно идентифицировать из кривых амплификации.
[332] Термин «кривая амплификации», в рамках изобретения, относится к кривой, полученной в результате реакции образования сигнала, в частности, реакции амплификации аналита-мишени (в частности, нуклеиновой кислоты-мишени). Кривая амплификации включает кривую, полученную в результате реакции в присутствии нуклеиновой кислоты-мишени в образце, и кривую или линию, полученную в результате реакции в отсутствие нуклеиновой кислоты-мишени в образце.
[333] В одном варианте осуществления, изменение сигнала и/или постоянный сигнал можно измерять по индикатору, показательному для амплификации нуклеиновой кислоты-мишени.
[334] В рамках изобретения, термин «индикатор, показательный для амплификации», относится к любому индикатору, тесно связанному с возникновением амплификации нуклеиновой кислоты-мишени, который можно получать из сигнала, обеспеченного на стадии (a). Индикатор может относиться к значению, полученному в зависимости от амплификации нуклеиновой кислоты-мишени. Индикатор может представлять собой индикатор, обеспечивающий большее значение, по мере того, как амплификация нуклеиновой кислоты-мишени увеличивается (то есть, по мере того, как количество нуклеиновой кислоты-мишени увеличивается), или может представлять собой индикатор, обеспечивающий меньшее значение, по мере того, как амплификация увеличивается. Индикатор может представлять собой любой индикатор, при условии, что он показывает амплификацию.
[335] В одном варианте осуществления, индикатор, показательный для амплификации, может включать индикатор, полученный из кривой амплификации или кривой плавления. В частности, индикатор может включать значение сигнала (например, RFU) в конкретном цикле, значение сигнала в каждом цикле, различие значений сигналов между конкретными циклами, или различие между контрольным значением сигнала и значением сигнала в конкретном цикле на кривой амплификации, или высоту, ширину или площадь максимального пика плавления на кривой плавления. В одном варианте осуществления, индикатор может представлять собой значение Ct (порогового цикла), ΔRFU (например, различие RFU в двух циклах, различие между контрольными RFU и RFU в конкретном цикле и т.д.), соотношение RFU (например, соотношение RFU в двух циклах или соотношение RFU между контрольными RFU и RFU в конкретном цикле и т.д.), и высоту/площадь/ширину пика плавления (например, высоту/площадь/ширину максимального пика на кривой плавления), но без ограничения этим.
[336] В соответствии с вариантом осуществления, индикатор, показательный для амплификации, представляет собой значение Ct. Значение Ct может представлять собой точку пересечения между кривой амплификации и пороговой линией. Концепция значения Ct хорошо известна в данной области.
[337] В соответствии с вариантом осуществления, индикатор, показательный для амплификации, представляет собой ΔRFU или соотношение RFU между значениями RFU, полученными в реакции амплификации. Например, индикатор представляет собой различие (вычитание) или соотношение между RFU в двух циклах, или различие (вычитание) или соотношение между RFU в конкретном цикле и контрольными RFU.
[338] В соответствии с одним вариантом осуществления, индикатор, показательный для амплификации, представляет собой площадь или ширину пика плавления. Площадь или ширина пика плавления относится к площади или ширине максимального пика в производной функции кривой плавления, полученной для анализа плавления. Площадь или ширина пика плавления широко известны в данной области.
[339] В одном варианте осуществления, детекцию сигналов можно проводить в одном цикле. В этом случае, является сложным измерение изменения сигнала посредством значения сигнала, детектированного только в одном цикле. Таким образом, изменение сигнала можно детектировать с использованием отдельного контрольного значения сигнала.
[340] В способе в соответствии с настоящим изобретением используют тот факт, что композиция для детекции нуклеиновой кислоты-мишени обеспечивает изменение сигнала в зависимости от присутствия нуклеиновой кислоты-мишени только при соответствующей температуре детекции. В соответствии с одним вариантом осуществления, способом в соответствии с настоящим изобретением можно измерять изменение сигнала с использованием значений сигнала, детектированных при температурах детекции по меньшей мере в двух циклах. В другом варианте осуществления, способом в соответствии с настоящим изобретением можно измерять изменение сигнала посредством использования значения сигнала при температуре детекции, детектированного в одном цикле (то есть, значения сигнала, детектированного на стадии (a)), и контрольного значения сигнала.
[341] В одном варианте осуществления, когда сигналы детектируют в множестве циклов на стадии (a), первый цикл и конечный цикл, при которых детектируют сигналы, можно выбирать, чтобы они были отделены друг от друга посредством по меньшей мере 1 цикла - 20 циклов между ними. В частности, первый цикл и конечный цикл, при которых детектируют сигналы, можно выбирать, чтобы они были отделены друг от друга посредством 1 цикла, 2 циклов, 3 циклов, 4 циклов, 5 циклов, 6 циклов, 7 циклов, 8 циклов, 9 циклов, 10 циклов, 11 циклов, 12 циклов, 13 циклов, 14 циклов, 15 циклов, 16 циклов, 17 циклов, 18 циклов, 19 циклов, или 20 циклов или более, и более конкретно, посредством 5 циклов, 10 циклов, 15 циклов, 20 циклов или 30 циклов, или более.
[342] В одном варианте осуществления, детекцию сигналов можно проводить в любом из промежуточных циклов включая область экспоненциальной фазы, или в любом из поздних циклов, включая область плато. Например, сигналы можно детектировать в двух циклах, где один цикл представляет собой любой из начальных циклов, включая начальную область, и другой цикл представляет собой любой из промежуточных или поздних циклов, или сигналы можно детектировать в двух циклах, где один цикл представляет собой любой из промежуточных циклов, и другой цикл представляет собой любой из промежуточных или поздних циклов.
[343] В одном варианте осуществления, начальные циклы включают циклы вплоть до цикла, соседнего с значением, полученным посредством деления конечного цикла на 3. Например, когда конечный цикл представляет собой 45, результат деления 45 на 3 представляет собой 15, и таким образом, начальные циклы можно определять, как представляющие собой от цикла 1 до цикла 20, цикл 1 - цикл 15, цикл 1 - цикл 10, или цикл 1 - цикл 5. промежуточные циклы можно определять, как представляющие собой циклы, соседние с значением, полученным посредством деления конечного цикла на 2. Например, когда конечный цикл представляет собой цикл 45, результат деления 45 на 2 представляет собой 22,5, и таким образом, промежуточные циклы можно определять, как представляющие собой от цикла 16 до цикла 30, цикл 18 - цикл 30, цикл 20 - цикл 30, цикл 16 - цикл 27, цикл 18 - цикл 27, цикл 20 - цикл 27, цикл 16 - цикл 25, цикл 18 - цикл 25 или цикл 20 - цикл 25. Поздние циклы можно определять, как представляющие собой конечный цикл реакции амплификации, или циклы, соседние с конечным циклом. Например, когда конечный цикл представляет собой цикл 45, поздние циклы можно определять, как представляющие собой от цикла 31 до цикла 45, цикл 35 - цикл 45, цикл 38 - цикл 45, цикл 40 - цикл 45 или цикл 43 - цикл 45. Начальные циклы, промежуточные циклы и поздние циклы могут изменяться в зависимости от конечного количества циклов реакции амплификации. В частности, на стадии (a), цикл, в котором детектируют сигнал, может представлять собой любой из поздних циклов (например, конечный цикл), и цикл, в котором детектируют контрольный сигнал, может представлять собой любой из начальных циклов (например, цикл 1) являющейся положительным контролем реакции.
[344] В одном варианте осуществления, изменение сигнала можно измерять с использованием сигнала, детектированного в по меньшей мере одном цикле, и «контрольного значения сигнала». Контрольное значение сигнала может относиться к значению, которое может подтверждать изменение сигнала, зависимое от присутствия нуклеиновой кислоты-мишени, посредством отдельной реакции.
[345] В одном варианте осуществления, контрольное значение сигнала можно получать из реакции в отсутствие соответствующей нуклеиновой кислоты-мишени при соответствующей температуре детекции. Например, контрольное значение сигнала может представлять собой «значение сигнала при температуре детекции» в отсутствие нуклеиновой кислоты-мишени.
[346] В одном варианте осуществления, существует n контрольных значений сигнала для n температур детекции.
[347] В одном варианте осуществления, «значение сигнала при температуре детекции (например, iй температуре детекции)», детектированное в отсутствие нуклеиновой кислоты-мишени (например, iй нуклеиновой кислоты-мишени), можно получать посредством отдельной являющейся отрицательным контролем реакции.
[348] В одном варианте осуществления, контрольное значение сигнала можно получать посредством проведения являющейся отрицательным контролем реакции в то же самое время или отдельно от способа в соответствии с настоящим изобретением.
[349] В одном варианте осуществления, контрольное значение сигнала можно получать посредством являющейся отрицательным контролем реакции. В соответствии с одним конкретным вариантом осуществления, контрольное значение сигнала при iй температуре детекции можно получать посредством смешивания образца, свободного от iй нуклеиновой кислоты-мишени (например, дистиллированной воды), с n композициями для детекции нуклеиновых кислот-мишеней и детекции сигналов при iй температуре детекции, с амплификацией в то же время нуклеиновых кислот. В данном случае, детекцию сигналов можно проводить в любом цикле. В частности, значение сигнала, детектированное в любом из начальных циклов являющейся отрицательным контролем реакции, можно использовать в качестве контрольного значения сигнала, или значение сигнала, детектированное в любом из поздних циклов являющейся отрицательным контролем реакции, можно использовать в качестве контрольного значения сигнала. Более конкретно, значение сигнала, детектированное в том же цикле, что и цикл, в котором детектируют сигнал на стадии (a), можно использовать в качестве контрольного значения сигнала.
[350] В одном варианте осуществления, контрольное значение сигнала можно получать посредством являющейся положительным контролем реакции. В соответствии с конкретным вариантом осуществления, контрольное значение сигнала можно получать посредством смешивания образца, содержащего iю нуклеиновую кислоту-мишень, с композицией для детекции iй нуклеиновой кислоты-мишени, и детекции сигнала при iй температуре детекции, с амплификацией в то же время нуклеиновых кислот.
[351] Когда контрольное значение сигнала получают посредством являющейся положительным контролем реакции, цикл, в котором детектируют значение сигнала, может представлять собой цикл в начальной области являющейся положительным контролем реакции. Начальная область относится к области, в которой сигнал (например, флуоресцентный сигнал) остается по существу постоянным в ходе начальных циклов реакции амплификации (например, ПЦР). В этой области, уровень продуктов амплификации является недостаточным, чтобы поддаваться детекции, большинство флуоресцентных сигналов в этой области приписывают флуоресцентному сигналу, присущему реакционному образцу, и фоновому сигналу, включающему флуоресцентные сигналы самой системы измерения. То есть, значение сигнала, детектированное в цикле в начальной области являющейся положительным контролем реакции, является по существу идентичным контрольному значению сигнала, полученному из реакции в отсутствие нуклеиновой кислоты-мишени (например, являющейся отрицательным контролем реакции).
[352] В одном варианте осуществления, изменение сигнала можно измерять посредством различия между контрольным значением сигнала и значением сигнала, детектированным на стадии (a).
[353] В одном варианте осуществления, контрольное значение сигнала может представлять собой пороговое значение, предварительно определенное из являющейся отрицательным контролем реакции, принимая во внимание фоновый сигнал и чувствительность детектора или характеристики используемых меток. С использованием порогового значения, можно определять значимость изменения сигнала. Пороговое значение можно определять посредством известного способа установки порога. Например, пороговое значение можно определять с учетом фонового сигнала, чувствительности, характеристик метки, изменчивости сигнала для детектора или пределы ошибки, и т.п.
[354] В одном варианте осуществления, когда пороговое значение используют в качестве контрольного значения сигнала, если значение сигнала, детектированное на стадии (a), является равным или большим, чем пороговое значение, можно определять, что сигнал изменился.
[355] В одном варианте осуществления, детекцию сигналов при каждой из n температур детекции можно проводить с использованием одного типа детектора.
[356] В одном варианте осуществления, один тип детектора представляет собой один детектор. В одном варианте осуществления, сигналы от меток в каждом из связанных с меткой олигонуклеотидов, включенных в n композиций для детекции нуклеиновых кислот-мишеней, не являются отличимыми друг от друга посредством одного типа детектора, применительно к каждой нуклеиновой кислоте-мишени.
[357] В рамках изобретения, одиночная метка или один тип флуоресцентной метки относится к флуоресцентной метке, имеющей идентичные или по существу идентичные характеристики сигнала (например, оптические характеристики, длину волны излучения и электрические сигналы).
[358] Например, FAM и CAL Fluor 610 обеспечивают различные типы сигналов. В настоящем описании, одиночная метка или один тип флуоресцентной метки означает, что сигналы от флуоресцентной метки не являются отличимыми друг от друга, с использованием канала детекции. Такая одиночная метка или один тип флуоресцентной метки не зависит от химической структуры флуоресцентной метки, и таким образом, даже когда две флуоресцентные метки имеют отличные друг от друга химические структуры, если они не являются отличимыми с использованием канала детекции, их рассматривают как один тип.
[359] В соответствии с настоящим изобретением, сигналы, образованные из n композиций для детекции нуклеиновых кислот-мишеней, включающих общий один тип флуоресцентной метки, не являются отличимыми посредством одного канала детекции.
[360] Термин «канал детекции», в рамках изобретения, относится к средствам для детекции сигнала от одного типа флуоресцентной метки. Термоциклеры, применимые в данной области, например, ABI 7500 (Applied Biosystems), QuantStudio (Applied Biosystems), CFX96 (Bio-Rad Laboratories), Cobas z 480 (Roche), LightCycler (Roche) и т.д., включают несколько каналов (например, светодиодов) для детекции сигналов от нескольких различных типов флуоресцентных меток, и эти каналы соответствуют каналу детекции в рамках изобретения.
[361] Канал детекции, используемый по настоящему изобретению, включает средства для детекции сигналов. Например, канал детекции может представлять собой светодиод, способный к детекции флуоресцентного сигнала при конкретной длине волны.
[362] В соответствии с вариантом осуществления, сигналы, детектированные при n температурах детекции, не являются отличимыми друг от друга посредством одного типа детектора.
[363]
[364] Стадия (b): Определение присутствие нуклеиновой кислоты-мишени
[365] После детекции сигналов, присутствие n нуклеиновых кислот-мишеней определяют из сигналов, детектированных на стадии (a).
[366] В одном варианте осуществления, присутствие iй нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при iй температуре детекции. Например, изменение сигнала измеряют из сигналов, детектированных при iй температуре детекции, для определения присутствия iй нуклеиновой кислоты-мишени.
[367] В одном варианте осуществления, когда измеряют изменение сигнала при iй температуре детекции, можно определять, что iя нуклеиновая кислота-мишень присутствует.
[368] В одном варианте осуществления, если сигнал является постоянным при iй температуре детекции, можно определять, что iя нуклеиновая кислота-мишень отсутствует.
[369] В одном варианте осуществления, изменение сигнала можно измерять с использованием сигналов, детектированных по меньшей мере в двух циклах, или сигнала, детектированного по меньшей мере в одном цикле, и «контрольного значения сигнала».
[370] Определение присутствия нуклеиновой кислоты-мишени из сигналов, детектированных при каждой температуре детекции, можно проводить посредством способа, описанного на стадии (a), для измерения изменения сигнала, например, способа с использованием метки, показательной для амплификации, и других различных способов, известных в данной области.
[371] В конкретном варианте осуществления, когда n представляет собой 3, и детекцию сигналов проводят в цикле 10, цикле 20 и цикле 30, присутствие первой нуклеиновой кислоты-мишени можно определять из сигналов, детектированных при первой температуре детекции (первого сигнала в цикле 10, первого сигнала в цикле 20 и первого сигнала в цикле 30), присутствие второй нуклеиновой кислоты-мишени можно определять из сигналов, детектированных при второй температуре детекции (второго сигнала в цикле 10, второго сигнала в цикле 20 и второго сигнала в цикле 30), и присутствие третьей нуклеиновой кислоты-мишени можно определять из сигналов, детектированных при третьей температуре детекции (третьего сигнала в цикле 10, третьего сигнала в цикле 20 и третьего сигнала в цикле 30).
[372] В конкретном варианте осуществления, когда n представляет собой 4, и детекцию сигналов проводят в цикле 30, присутствие первой нуклеиновой кислоты-мишени определяют из сигнала, детектированного при первой температуре детекции (то есть, первого сигнала в цикле 30), и контрольного значения сигнала, присутствие второй нуклеиновой кислоты-мишени определяют из сигнала, детектированного при второй температуре детекции (то есть, второго сигнала в цикле 30), и контрольного значения сигнала, и присутствие третьей нуклеиновой кислоты-мишени определяют из сигнала, детектированного при третьей температуре детекции (то есть, третьего сигнала в цикле 30), и контрольного значения сигнала.
[373] В одном варианте осуществления, контрольное значение сигнала можно получать посредством отдельной являющейся отрицательным контролем реакции или являющейся положительным контролем реакции.
[374] В одном варианте осуществления, способ в соответствии с настоящим изобретением можно осуществлять вместе с являющейся отрицательным контролем реакцией. Значение сигнала, детектированное в являющейся отрицательным контролем реакции, можно использовать в качестве контрольного значения сигнала. Например, сигнал, детектированный в одном цикле (например, конечном цикле) при iй температуре детекции в реакции, содержащей композицию для детекции iй нуклеиновой кислоты-мишени, можно сравнивать с сигналом, детектированным в том же самом цикле (например, конечном цикле) при одной и той же температуре детекции (то есть, iй температуре детекции) в являющейся отрицательным контролем реакции, для определения того, изменился или нет сигнал.
[375] В конкретном варианте осуществления, когда n представляет собой 3, и сигнал детектируют в цикле 30, присутствие первой нуклеиновой кислоты-мишени можно определять из сигнала, детектированного при первой температуре детекции (то есть, первого сигнала в цикле 30), и первого контрольного значения сигнала (например, сигнала при первой температуре детекции, детектированного в цикле 30 являющейся отрицательным контролем реакции), присутствие второй нуклеиновой кислоты-мишени можно определять из сигнала, детектированного при второй температуре детекции (то есть, второго сигнала в цикле 30), и второго контрольного значения сигнала (например, сигнала при второй температуре детекции, детектированного в цикле 30 являющейся отрицательным контролем реакции), и присутствие третьей нуклеиновой кислоты-мишени можно определять из сигнала, детектированного при третьей температуре детекции (то есть, третьего сигнала в цикле 30), и третьего контрольного значения сигнала (например, сигнала при третьей температуре детекции, детектированного в цикле 30 являющейся отрицательным контролем реакции).
[376] В одном варианте осуществления, способ в соответствии с настоящим изобретением можно осуществлять вместе с являющейся положительным контролем реакцией. Значение сигнала, детектированное в являющейся положительным контролем реакции, можно использовать в качестве контрольного значения сигнала. Например, сигнал, детектированный в одном цикле при iй температуре детекции, например, в цикле 30, первый сигнал, детектированный в цикле 30, можно сравнивать с сигналом, детектированным при iй температуре детекции в являющейся положительным контролем реакции в цикле до цикла 30, например, цикле 1, для определения того, изменился ли сигнал.
[377] В одном варианте осуществления, когда детекцию сигнала проводят в одном цикле на стадии (a), и изменение сигнала измеряют с использованием контрольного значения сигнала, полученного посредством являющейся положительным контролем реакции, детекцию сигнала в являющейся положительным контролем реакции для получения контрольного значения сигнала можно проводить в цикле, составляющем по меньшей мере 30 циклов, 20 циклов, 10 циклов или 5 циклов до цикла, в котором детекцию сигнала проводят на стадии (a).
[378] В конкретном варианте осуществления, когда n представляет собой 3, и детекцию сигнала проводят в цикле 30, присутствие первой нуклеиновой кислоты-мишени можно определять из сигнала, детектированного при первой температуре детекции (то есть, первого сигнала в цикле 30), и первого контрольного значения сигнала (например, сигнала при первой температуре детекции, детектированного в цикле 1 являющейся положительным контролем реакции первой нуклеиновой кислоты-мишени), присутствие второй нуклеиновой кислоты-мишени можно определять из сигнала, детектированного при второй температуре детекции (то есть, второго сигнала в цикле 30) и второго контрольного значения сигнала (например, сигнала при второй температуре детекции, детектированного в цикле 1 являющейся положительным контролем реакции второй нуклеиновой кислоты-мишени), и присутствие третьей нуклеиновой кислоты-мишени можно определять из сигнала, детектированного при третьей температуре детекции (то есть, третьего сигнала в цикле 30), и третьего контрольного значения сигнала (например, сигнала при третьей температуре детекции, детектированного в цикле 1 являющейся положительным контролем реакции третьей нуклеиновой кислоты-мишени).
[379]
[380] В соответствии с другим аспектом, настоящее изобретение относится к способу детекции двух нуклеиновых кислот-мишеней в образце, включающему:
[381] (a) детекцию сигналов при первой температуре детекции и второй температуре детекции, с инкубацией в то же время образца, как подозревают, содержащего по меньшей мере одну из двух нуклеиновых кислот-мишеней, с композицией для детекции первой нуклеиновой кислоты-мишени и композицией для детекции второй нуклеиновой кислоты-мишени в реакционном сосуде; где инкубация включает множество циклов, и детекцию сигналов проводят по меньшей мере в одном из множества циклов, где композиция для детекции первой нуклеиновой кислоты-мишени обеспечивает изменение сигнала при первой температуре детекции и обеспечивает постоянный сигнал при второй температуре детекции в присутствии первой нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие первой нуклеиновой кислоты-мишени; и композиция для детекции второй нуклеиновой кислоты-мишени обеспечивает изменение сигнала при второй температуре детекции и обеспечивает постоянный сигнал при первой температуре детекции в присутствии второй нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие второй нуклеиновой кислоты-мишени, и где первая температура детекции является более низкой, чем вторая температура детекции, и (b) определение присутствия двух нуклеиновых кислот-мишеней из сигналов, детектированных на стадии (a); где присутствие первой нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при первой температуре детекции, и присутствие второй нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при второй температуре детекции.
[382]
[383] В соответствии с другим аспектом, настоящее изобретение относится к способу детекции трех нуклеиновых кислот-мишеней в образце, включающему:
[384] (a) детекцию сигналов при первой температуре детекции, второй температуре детекции и третьей температуре детекции, с инкубацией в то же время образца, как подозревают, содержащего по меньшей мере одну из трех нуклеиновых кислот-мишеней, с композицией для детекции первой нуклеиновой кислоты-мишени, композицией для детекции второй нуклеиновой кислоты-мишени и композицией для детекции третьей нуклеиновой кислоты-мишени в реакционном сосуде; где инкубация включает множество циклов, и детекцию сигналов проводят по меньшей мере в одном из множества циклов, где композиция для детекции первой нуклеиновой кислоты-мишени обеспечивает изменение сигнала при первой температуре детекции и обеспечивает постоянный сигнал при второй температуре детекции и третьей температуре детекции в присутствии первой нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие первой нуклеиновой кислоты-мишени; композиция для детекции второй нуклеиновой кислоты-мишени обеспечивает изменение сигнала при второй температуре детекции и обеспечивает постоянный сигнал при первой температуре детекции и третьей температуре детекции в присутствии второй нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие второй нуклеиновой кислоты-мишени; и композиция для детекции третьей нуклеиновой кислоты-мишени обеспечивает изменение сигнала при третьей температуре детекции и обеспечивает постоянный сигнал при первой температуре детекции и второй температуре детекции в присутствии третьей нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие третьей нуклеиновой кислоты-мишени, и где первая температура детекции является более низкой, чем вторая температура детекции, и вторая температура детекции является более низкой, чем третья температура детекции, и (b) определение присутствия трех нуклеиновых кислот-мишеней из сигналов, детектированных на стадии (a), где присутствие первой нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при первой температуре детекции, присутствие второй нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при второй температуре детекции, и присутствие третьей нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при третьей температуре детекции.
[385]
[386] Поскольку второй вариант осуществления и третий вариант осуществления настоящего изобретения следуют таким же принципам, что и первый вариант осуществления настоящего изобретения, описанный выше, признаки, общие между этими вариантами осуществления, не описаны избыточным образом, в интересах ясности настоящей заявки.
[387]
[388] II. Набор для детекции нуклеиновых кислот-мишеней
[389] В соответствии с другим аспектом, настоящее изобретение относится к набору, содержащему n композиций для детекции n нуклеиновых кислот-мишеней в образце, где n представляет собой целое число 2 или более, где каждая из n композиций для детекции n нуклеиновых кислот-мишеней обеспечивает изменение сигнала при соответствующей температуре детекции среди n температур детекции, где изменение сигнала указывает на присутствие соответствующей нуклеиновой кислоты-мишени, где композиция для детекции iй нуклеиновой кислоты-мишени среди n нуклеиновых кислот-мишеней обеспечивает изменение сигнала при iй температуре детекции среди n температур детекции и обеспечивает постоянный сигнал при других температурах детекции в присутствии iй нуклеиновой кислоты-мишени, и где i представляет собой целое число от 1 до n, и iя температура детекции является более низкой, чем (i+1)я температура детекции.
[390]
[391] Поскольку набор по настоящему изобретению получают для обеспечения возможности осуществления способов по настоящему изобретению, признаки, общие между ними, не описаны избыточным образом, в интересах ясности настоящей заявки.
[392]
[393] В соответствии с вариантом осуществления, в температурном диапазоне, покрывающем все из n температур детекции, композиция для детекции iй нуклеиновой кислоты-мишени имеет «температурный диапазон изменения сигнала», в котором сигнал изменяется, в зависимости от присутствия iй нуклеиновой кислоты-мишени, и «температурный диапазон постоянного сигнала», в котором сигнал является постоянным, даже в присутствии iй нуклеиновой кислоты-мишени.
[394] В соответствии с вариантом осуществления, композиция для детекции iй нуклеиновой кислоты-мишени имеет один или два температурных диапазона постоянного сигнала.
[395] В соответствии с вариантом осуществления, композиция для детекции iй нуклеиновой кислоты-мишени представляет собой любую из: (i) композиции с нижним изменением сигнала (UnderSC), имеющей характеристику, что температурный диапазон изменения сигнала является более низким, чем температурный диапазон постоянного сигнала; (ii) композиции с верхним изменением сигнала (OverSC), имеющей характеристику, что температурный диапазон изменения сигнала является более высоким, чем температурный диапазон постоянного сигнала; и (iii) композиции с промежуточным изменением сигнала (InterSC), имеющей характеристику, что температурный диапазон изменения сигнала является более высоким, чем один из двух температурных диапазонов постоянного сигнала, и более низкой, чем другой из двух температурных диапазонов постоянного сигнала.
[396] В соответствии с вариантом осуществления, iя температура детекции выбрана в рамках температурного диапазона изменения сигнала композиции для детекции iй нуклеиновой кислоты-мишени, и iя температура детекции не включена в температурные диапазоны изменения сигнала композиций для детекции других нуклеиновых кислот-мишеней.
[397] В соответствии с вариантом осуществления, температурный диапазон изменения сигнала композиции для детекции iй нуклеиновой кислоты-мишени может частично перекрываться с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей соседнюю температуру детекции, и не перекрывается с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей температуру детекции, не являющуюся соседней с ней.
[398] В соответствии с вариантом осуществления, когда n представляет собой 2, композиция для детекции первой нуклеиновой кислоты-мишени представляет собой композицию UnderSC или композицию InterSC, и композиция для детекции второй нуклеиновой кислоты-мишени представляет собой композицию InterSC или композицию OverSC.
[399] В соответствии с вариантом осуществления, когда n представляет собой 3 или более, композиция для детекции первой нуклеиновой кислоты-мишени представляет собой композицию UnderSC или композицию InterSC, композиция для детекции nй нуклеиновой кислоты-мишени представляет собой композицию InterSC или композицию OverSC, и композиции для детекции нуклеиновых кислот-мишеней, отличных от первой нуклеиновой кислоты-мишени и nй нуклеиновой кислоты-мишени, представляют собой композицию InterSC.
[400] В соответствии с вариантом осуществления, композиция для детекции iй нуклеиновой кислоты-мишени включает метку, предоставляющую сигнал в зависимости от присутствия iй нуклеиновой кислоты-мишени.
[401] В соответствии с вариантом осуществления, метка является связанной с олигонуклеотидом или встраивается в олигонуклеотид в ходе инкубации набора с образцом.
[402] В соответствии с вариантом осуществления, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала.
[403] В соответствии с вариантом осуществления, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала, и когда дуплекс существует в форме дуплекса в связанной форме или в диссоциированной форме, композиция для детекции iй нуклеиновой кислоты-мишени обеспечивает сигнал от метки.
[404] В соответствии с вариантом осуществления, сигналы, обеспечиваемые посредством n композиций для детекции n нуклеиновых кислот-мишеней, не являются отличимыми друг от друга посредством одного типа детектора.
[405] В одном варианте осуществления, сигналы, указывающие на присутствие каждой нуклеиновой кислоты-мишени, обеспечиваемые посредством n композиций для детекции нуклеиновых кислот-мишеней, не являются отличимыми друг от друга посредством одного типа детектора. Например, все из меток, включенных в n композиций для детекции нуклеиновых кислот-мишеней, представляют собой идентичный тип метки (например, одну флуоресцентную метку).
[406] В одном варианте осуществления, композиция для детекции iй нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала, и температурный диапазон изменения сигнала композиции для детекции iй нуклеиновой кислоты-мишени определяют в зависимости от длины и/или последовательности дуплекса.
[407] В соответствии с вариантом осуществления, набор, кроме того, содержит инструкцию, описывающую способ по настоящему изобретению.
[408] В соответствии с вариантом осуществления, инструкции для описания или практического осуществления способов по настоящему изобретению могут быть записаны на подходящей среде для записи. Например, инструкции могут быть напечатаны на субстрате, таком как бумага и пластик. В других вариантах осуществления, инструкции могут быть представлены в форме файла электронного хранения данных, присутствующего на подходящей машиночитаемой среде для хранения информации, такой как CD-ROM и дискета. В других вариантах осуществления, действующие инструкции могут не присутствовать в наборе, однако, предоставлены средства для получения инструкций из удаленного источника, например, через интернет. Примером этого варианта осуществления является набор, включающий веб-адрес, по которому инструкции можно просматривать и/или с которого инструкции можно загружать.
[409] Все наборы, описанные в настоящем описании выше, могут, необязательно, включать реагенты, необходимые для проведения реакций амплификации нуклеиновой кислоты-мишени (например, реакций ПЦР), такие как буферы, кофакторы ДНК-полимеразы и дезоксирибонуклеотид-5-трифосфаты. Необязательно, наборы могут также включать различные молекулы полинуклеотидов, обратную транскриптазу, различные буферы и реагенты, и антитела, ингибирующие активность ДНК-полимеразы. Наборы могут также включать реагенты, необходимые для проведения являющихся положительным и отрицательным контролем реакций. Оптимальные количества реагентов, подлежащих использованию в данной реакции, может легко определять специалист в данной области, пользуясь преимуществом настоящего описания. Вышеописанные составляющие наборов могут находиться в отдельных контейнерах, или множество составляющих могут находиться в одном контейнере.
[410]
[411] Далее в настоящем описании, настоящее изобретение описано более подробно посредством вариантов осуществления. Следующие варианты осуществления представлены для более подробного описания настоящего изобретения, и станет очевидным специалисту в области, к которой относится настоящее изобретение, что объем настоящего изобретения, как предложено в прилагаемой формуле изобретения, не является ограниченным следующими вариантами осуществления.
[412]
[413] ПРИМЕРЫ
[414]
[415] Авторы настоящего изобретения подтвердили, можно ли множество нуклеиновых кислот-мишеней детектировать в реальном времени с использованием одинакового типа, то есть, одного типа метки, в соответствии со способом по настоящему изобретению, посредством использования различных комбинаций композиции UnderSC для детекции нуклеиновой кислоты-мишени, композиции InterSC для детекции нуклеиновой кислоты-мишени и композиции OverSC для детекции нуклеиновой кислоты-мишени.
[416] В частности, среди различных механизмов образования сигналов, приемлемых для трех типов композиций для детекции нуклеиновых кислот-мишеней, способ на основе PTOCE, как на ФИГ. 5, с использованием CTO, с которым связаны все из интерактивных двойных меток (в настоящем описании ниже, обозначенный как способ-1 на основе PTOCE), использовали для композиции UnderSC; способ на основе PTOCE, как на ФИГ. 6, в котором одна из интерактивных двойных меток является связанной с 5’-метящей частью PTO, и другая является связанной со связывающей частью CTO (далее в настоящем описании обозначенный как способ-2 на основе PTOCE) использовали для композиции InterSC; и способ двойного гашения, как показано на ФИГ. 7, использовали для композиции OverSC.
[417] Для матриц множества нуклеиновых кислот-мишеней, использовали геномную ДНК Chlamydia trachomatis (CT) (Номер доступа: ATCC VR-1500, Koram Deo Lab), Neisseria gonorrhoeae (NG) (Номер доступа: ATCC 700825, Koram Deo Lab) и Ureaplasma parvum (UP) (Номер доступа: ATCC 27815, Koram Deo Lab). Как показано в таблице 1 ниже, вышеуказанные три нуклеиновые кислоты-мишени детектировали посредством организации различных комбинаций вышеуказанных 3 типов композиций и температур детекции. Комбинации 1-4 представляют собой случаи, когда количество нуклеиновых кислот-мишеней (n) представляет собой 2, и комбинация 5 представляет собой случай, когда количество нуклеиновых кислот-мишеней (n) представляет собой 3.
[418]
[419] [Таблица 1]
(°C)
[420]
[421] Пример 1: Получение олигонуклеотидов для композиции для детекции нуклеиновых кислот-мишеней
[422]
[423] <Комбинация 1>
[424] Первую температуру детекции для детекции первого изменения сигнала, показательного для присутствия первой нуклеиновой кислоты-мишени, CT, устанавливали на 50°C, и вторую температуру детекции для детекции второго изменения сигнала, показательного для присутствия второй нуклеиновой кислоты-мишени, NG, устанавливали на 72°C. Затем, олигонуклеотиды для композиции для детекции нуклеиновой кислоты-мишени CT и композиции для детекции нуклеиновой кислоты-мишени NG, получали, как показано в таблице 2 ниже.
[425] Композиция для детекции нуклеиновой кислоты-мишени CT включает пару праймеров, PTO и CTO, связанных с репортерной молекулой (CAL Fluor красным 610) и молекулой гасителя (BHQ-2), где последовательности PTO и CTO были разработаны таким образом, что дуплекс между PTO (то есть, нерасщепленным PTO) и CTO находится в диссоциированной форме как при первой, так и при второй температурах детекции, и удлиненный дуплекс, сформированный в зависимости от присутствия CT, находится в связанной форме при первой температуре детекции и в диссоциированной форме при второй температуре детекции (см. ФИГ. 8).
[426] Композиция для детекции нуклеиновой кислоты-мишени NG включает пару праймеров, PTO, имеющий молекулу гасителя (BHQ-2), связанную с его метящей частью, и CTO, имеющий репортерную молекулу (CAL Fluor красный 610), связанную с его связывающей частью, где последовательности PTO и CTO были разработаны таким образом, что дуплекс между PTO (то есть, нерасщепленным PTO) и CTO находится в связанной форме при первой температуре детекции, и в диссоциированной форме при второй температуре детекции, и удлиненный дуплекс, сформированный в зависимости от присутствия NG, находится в связанной форме как при первой, так и при второй температурах детекции (см. ФИГ. 8).
[427]
[428] <Комбинация 2>
[429] Первую температуру детекции для детекции первого изменения сигнала, показательного для присутствия первой нуклеиновой кислоты-мишени, CT, устанавливали на 57°C, и вторую температуру детекции для детекции второго изменения сигнала, показательного для присутствия второй нуклеиновой кислоты-мишени, UP, устанавливали на 85°C. Затем, олигонуклеотиды для композиции для детекции нуклеиновой кислоты-мишени CT и композиции для детекции нуклеиновой кислоты-мишени UP, получали, как показано в таблице 2 ниже.
[430] Композиция для детекции нуклеиновой кислоты-мишени CT включает пару праймеров, PTO и CTO, связанных с репортерной молекулой (CAL Fluor красным 610) и молекулой гасителя (BHQ-2), где последовательности PTO и CTO были разработаны таким образом, что дуплекс между PTO (то есть, нерасщепленным PTO) и CTO находится в диссоциированной форме как при первой, так и при второй температурах детекции, и удлиненный дуплекс, сформированный в зависимости от присутствия CT, находится в связанной форме при первой температуре детекции, и в диссоциированной форме при второй температуре детекции (см. ФИГ. 9).
[431] Композиция для детекции нуклеиновой кислоты-мишени UP включает пару праймеров, PTO, связанный с репортерной молекулой (CAL Fluor красным 610) и первой молекулой гасителя (BHQ-2), и CQO, связанный с второй молекулой гасителя (BHQ-2), где последовательности PTO и CQO были разработаны таким образом, что дуплекс с меткой между PTO (то есть, нерасщепленным PTO) и CQO находится в связанной форме при первой температуре детекции, и в диссоциированной форме при второй температуре детекции. То есть, последовательности PTO и CQO были разработаны таким образом, что дуплекс с меткой между PTO и CQO расщепляется в зависимости от присутствия нуклеиновой кислоты-мишени UP для получения фрагмента активированного дуплекса с меткой, и фрагмент активированного дуплекса с меткой находится в связанной форме при первой температуре детекции, и в диссоциированной форме при второй температуре детекции (см. ФИГ. 9).
[432]
[433] <Комбинация 3>
[434] Первая температура детекции для детекции первого изменения сигнала, показательного для присутствия первой нуклеиновой кислоты-мишени, CT, составляла 57°C, и вторую температуру детекции для детекции второго изменения сигнала, показательного для присутствия второй нуклеиновой кислоты-мишени, NG, устанавливали на 72°C. Затем, олигонуклеотиды для композиции для детекции нуклеиновой кислоты-мишени CT и композиции для детекции нуклеиновой кислоты-мишени NG, получали, как показано в таблице 2 ниже.
[435] Композиция для детекции нуклеиновой кислоты-мишени CT включает пару праймеров, PTO, имеющий молекулу гасителя (BHQ-2), связанную с его метящей частью, и CTO, имеющий репортерную молекулу (CAL Fluor красный 610), связанную с его связывающей частью, где последовательности PTO и CTO были разработаны таким образом, что дуплекс между PTO (то есть, нерасщепленным PTO) и CTO находится в диссоциированной форме при первой и второй температурах детекции, и удлиненный дуплекс, сформированный в зависимости от присутствия CT, находится в связанной форме при первой температуре детекции и в диссоциированной форме при второй температуре детекции (см. ФИГ. 10).
[436] Композиция для детекции нуклеиновой кислоты-мишени NG включает пару праймеров, PTO, имеющий молекулу гасителя (BHQ-2), связанную с его метящей частью, и CTO, имеющий репортерную молекулу (CAL Fluor красный 610), связанную с его связывающей частью, где последовательности PTO и CTO были разработаны таким образом, что дуплекс между PTO (то есть, нерасщепленным PTO) и CTO находится в связанной форме при первой температуре детекции, и в диссоциированной форме при второй температуре детекции, и удлиненный дуплекс, сформированный в зависимости от присутствия NG, находится в связанной форме как при первой, так и при второй температурах детекции (см. ФИГ. 10).
[437]
[438] <Комбинация 4>
[439] Первую температуру детекции для детекции первого изменения сигнала, показательного для присутствия первой нуклеиновой кислоты-мишени, NG, устанавливали на 72°C, и вторую температуру детекции для детекции второго изменения сигнала, показательного для присутствия второй нуклеиновой кислоты-мишени, UP, устанавливали на 95°C. Затем, олигонуклеотиды для композиции для детекции нуклеиновой кислоты-мишени NG и композиции для детекции нуклеиновой кислоты-мишени UP, получали, как показано в таблице 2 ниже.
[440] Композиция для детекции нуклеиновой кислоты-мишени NG включает пару праймеров, PTO, имеющий молекулу гасителя (BHQ-2), связанную с его метящей частью, и CTO, имеющий репортерную молекулу (CAL Fluor красный 610), связанную с его связывающей частью, где последовательности PTO и CTO были разработаны таким образом, что дуплекс между PTO (то есть, нерасщепленным PTO) и CTO находится в диссоциированной форме при первой и второй температурах детекции, и удлиненный дуплекс, сформированный в зависимости от присутствия NG, находится в связанной форме при первой температуре детекции, и в диссоциированной форме при второй температуре детекции (см. ФИГ. 11).
[441] Композиция для детекции нуклеиновой кислоты-мишени UP включает пару праймеров, PTO, связанный с репортерной молекулой (CAL Fluor красным 610) и первой молекулой гасителя (BHQ-2), и CQO, связанный с второй молекулой гасителя (BHQ-2), где последовательности PTO и CQO были разработаны таким образом, что дуплекс с меткой между PTO (то есть, нерасщепленным PTO) и CQO находится в связанной форме при первой температуре детекции, и в диссоциированной форме при второй температуре детекции. То есть, последовательности PTO и CQO были разработаны таким образом, что дуплекс с меткой между PTO и CQO расщепляется в зависимости от присутствия нуклеиновой кислоты-мишени UP, для получения фрагмента активированного дуплекса с меткой, и фрагмент активированного дуплекса с меткой находится в связанной форме при первой температуре детекции, и в диссоциированной форме при второй температуре детекции (см. ФИГ. 11).
[442]
[443] <Комбинация 5>
[444] Первую температуру детекции для детекции первого изменения сигнала, показательного для присутствия первой нуклеиновой кислоты-мишени, CT, устанавливали на 50°C, вторую температуру детекции для детекции второго изменения сигнала, показательного для присутствия второй нуклеиновой кислоты-мишени, NG, устанавливали на 72°C, и третью температуру детекции для детекции третьего изменения сигнала, показательного для присутствия третьей нуклеиновой кислоты-мишени, UP, устанавливали на 95°C. Затем, олигонуклеотиды для композиции для детекции нуклеиновой кислоты-мишени CT, композиции для детекции NG и композиции для детекции нуклеиновой кислоты-мишени UP, получали, как показано в таблице 2 ниже.
[445] Композиция для детекции нуклеиновой кислоты-мишени CT включает пару праймеров, PTO и CTO, связанных с репортерной молекулой (CAL Fluor красным 610) и молекулой гасителя (BHQ-2), где последовательности PTO и CTO были разработаны таким образом, что дуплекс между PTO (то есть, нерасщепленным PTO) и CTO находится в диссоциированной форме при всех из первой - третьей температур детекции, и удлиненный дуплекс, сформированный в зависимости от присутствия CT, находится в связанной форме при первой температуре детекции, и в диссоциированной форме при второй и третьей температурах детекции (см. ФИГ. 12).
[446] Композиция для детекции нуклеиновой кислоты-мишени NG включает пару праймеров, PTO, имеющий молекулу гасителя (BHQ-2) связанную с его метящей частью, и CTO, имеющий репортерную молекулу (CAL Fluor красный 610), связанную с его связывающей частью, где последовательности PTO и CTO были разработаны таким образом, что дуплекс между PTO (то есть, нерасщепленным PTO) и CTO находится в связанной форме при первой температуре детекции, и в диссоциированной форме при второй и третьей температурах детекции, и удлиненный дуплекс, сформированный в зависимости от присутствия NG, находится в связанной форме при первой и второй температурах детекции, и в диссоциированной форме при третьей температуре детекции (см. ФИГ. 12).
[447] Композиция для детекции нуклеиновой кислоты-мишени UP включает пару праймеров, PTO, связанный с репортерной молекулой (CAL Fluor красным 610) и первой молекулой гасителя (BHQ-2), и CQO, связанный с второй молекулой гасителя (BHQ-2), где последовательности PTO и CQO были разработаны таким образом, что дуплекс с меткой между PTO (то есть, нерасщепленным PTO) и CQO находится в связанной форме при первой и второй температурах детекции, и в диссоциированной форме при третьей температуре детекции. То есть, последовательности PTO и CQO были разработаны таким образом, что дуплекс с меткой между PTO и CQO расщепляется в зависимости от присутствия нуклеиновой кислоты-мишени UP для получения фрагмента активированного дуплекса с меткой, и фрагмент активированного дуплекса с меткой находится в связанной форме при первой и второй температурах детекции, и в диссоциированной форме при третьей температуре детекции (см. ФИГ. 12).
[448]
[449] 3′-конец PTO и 3'-конец CTO в способе-1 на основе PTOCE являлись блокированными посредством спейсера C3 для запрещения реакций удлинения посредством ДНК полимеразы.
[450]
[451] [Таблица 2]
[452] Подчеркнуто: Метящая часть PTO
[453]
[454] Пример 2: Получение реакционных смесей
[455] С использованием олигонуклеотидов из комбинаций 1-5, полученных в примере 1 выше, реакционные смеси получали следующим образом. ДНК-полимеразу Taq, имеющую 5'-нуклеазную активность, включали в реакционные смеси для удлинения прямых праймеров и обратных праймеров, и расщепления олигонуклеотида.
[456]
[457] <Реакционная смесь из комбинации 1>
[458] Нуклеиновые кислоты-мишени (пробирка 1: 500 пг геномной ДНК CT; пробирка 2: 500 пг геномной ДНК NG; пробирка 3: смесь 500 пг геномной ДНК CT и 500 пг геномной ДНК NG, и пробирка 4: дистиллированная вода (отрицательный контроль)) смешивали с: 5 пмоль прямого праймера (SEQ ID NO: 1), 5 пмоль обратного праймера (SEQ ID NO: 2), 3 пмоль PTO (SEQ ID NO: 3) и 1 пмоль CTO (SEQ ID NO: 4), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени CT; и 5 пмоль прямого праймера (SEQ ID NO: 5), 5 пмоль обратного праймера (SEQ ID NO: 6), 1 пмоль PTO (SEQ ID NO: 7) и 1 пмоль CTO (SEQ ID NO: 8), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени NG, и затем объединяли с 5 мкл 4X готовой реакционной смеси (конечной, 200 мкМ dNTP, 2 мМ MgCl2, 2 ед. Taq) (Enzynomics, Korea), для получения реакционной смеси в конечном объеме 20 мкл.
[459]
[460] <Реакционная смесь из комбинации 2>
[461] Нуклеиновые кислоты-мишени (пробирка 1: 50 пг геномной ДНК CT; пробирка 2: 50 пг геномной ДНК UP; пробирка 3: смесь 50 пг геномной ДНК CT и 50 пг геномной ДНК UP, и пробирка 4: дистиллированная вода (отрицательный контроль)) смешивали с: 5 пмоль прямого праймера (SEQ ID NO: 1), 5 пмоль обратного праймера (SEQ ID NO: 2), 3 пмоль PTO (SEQ ID NO: 3) и 1 пмоль CTO (SEQ ID NO: 4), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени CT; и 5 пмоль прямого праймера (SEQ ID NO: 9), 5 пмоль обратного праймера (SEQ ID NO: 10), 1 пмоль PTO (SEQ ID NO: 11) и 5 пмоль CQO (SEQ ID NO: 12), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени UP, и затем объединяли с 5 мкл 4X готовой реакционной смеси (конечной, 200 мкМ dNTP, 2 мМ MgCl2, 2 ед. ДНК-полимеразы Taq) (Enzynomics, Korea), для получения реакционной смеси в конечном объеме 20 мкл.
[462]
[463] <Реакционная смесь из комбинации 3>
[464] Нуклеиновые кислоты-мишени (пробирка 1: 500 пг геномной ДНК CT; пробирка 2: 500 пг геномной ДНК NG; пробирка 3: смесь 500 пг геномной ДНК CT и 500 пг геномной ДНК NG, и пробирка 4: дистиллированная вода (отрицательный контроль)) смешивали с: 5 пмоль прямого праймера (SEQ ID NO: 1), 5 пмоль обратного праймера (SEQ ID NO: 2), 5 пмоль PTO (SEQ ID NO: 13) и 2 пмоль CTO (SEQ ID NO: 14), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени CT; и 5 пмоль прямого праймера (SEQ ID NO: 5), 5 пмоль обратного праймера (SEQ ID NO: 6), 3 пмоль PTO (SEQ ID NO: 7) и 1 пмоль CTO (SEQ ID NO: 8), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени NG, и затем объединяли с 5 мкл 4X готовой реакционной смеси (конечной, 200 мкМ dNTP, 2 мМ MgCl2, 2 ед. ДНК-полимеразы Taq) (Enzynomics, Korea), для получения реакционной смеси в конечном объеме 20 мкл.
[465]
[466] <Реакционная смесь из комбинации 4>
[467] Нуклеиновые кислоты-мишени (пробирка 1: 500 пг геномной ДНК NG; пробирка 2: 500 пг геномной ДНК UP; пробирка 3: смесь 500 пг геномной ДНК NG и 500 пг геномной ДНК UP, и пробирка 4: дистиллированная вода (отрицательный контроль)) смешивали с: 5 пмоль прямого праймера (SEQ ID NO: 5), 5 пмоль обратного праймера (SEQ ID NO: 6), 1 пмоль PTO (SEQ ID NO: 7) и 1 пмоль CTO (SEQ ID NO: 8), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени NG; и 5 пмоль прямого праймера (SEQ ID NO: 9), 5 пмоль обратного праймера (SEQ ID NO: 10), 0,5 пмоль PTO (SEQ ID NO: 11), и 3 пмоль CQO (SEQ ID NO: 12), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени UP, и затем объединяли с 5 мкл 4X готовой реакционной смеси (конечной, 200 мкМ dNTP, 2 мМ MgCl2, 2 ед. ДНК-полимеразы Taq) (Enzynomics, Korea), для получения реакционной смеси в конечном объеме 20 мкл.
[468]
[469] <Реакционная смесь из комбинации 5>
[470] Нуклеиновые кислоты-мишени (пробирка 1: 50 пг геномной ДНК CT; пробирка 2: 50 пг геномной ДНК NG; пробирка 3: 50 пг геномной ДНК UP; пробирка 4: смесь 50 пг геномной ДНК CT и 50 пг геномной ДНК NG; пробирка 5: смесь 50 пг геномной ДНК CT и 50 пг геномной ДНК UP; пробирка 6: смесь 50 пг геномной ДНК NG и 50 пг геномной ДНК UP; пробирка 7: смесь 50 пг геномной ДНК CT, 50 пг геномной ДНК NG и 50 пг геномной ДНК UP; пробирка 8: дистиллированная вода (отрицательный контроль)) смешивали с: 5 пмоль прямого праймера (SEQ ID NO: 1), 5 пмоль обратного праймера (SEQ ID NO: 2), 3 пмоль PTO (SEQ ID NO: 3), и 1 пмоль CTO (SEQ ID NO: 4), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени CT; 5 пмоль прямого праймера (SEQ ID NO: 5), 5 пмоль обратного праймера (SEQ ID NO: 6), 1 пмоль PTO (SEQ ID NO: 7) и 1 пмоль CTO (SEQ ID NO: 8), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени NG; и 5 пмоль прямого праймера (SEQ ID NO: 9), 5 пмоль обратного праймера (SEQ ID NO: 10), 0,5 пмоль PTO (SEQ ID NO: 11) и 3 пмоль CQO (SEQ ID NO: 12), в качестве олигонуклеотидов для нуклеиновой кислоты-мишени UP, и затем объединяли с 5 мкл 4X готовой реакционной смеси (конечной, 200 мкМ dNTP, 2 мМ MgCl2, 2 ед. ДНК-полимеразы Taq) (Enzynomics, Korea), для получения реакционной смеси в конечном объеме 20 мкл.
[471]
[472] Пример 3: ПЦР с детекцией в реальном времени
[473] Затем, ПЦР с детекцией в реальном времени проводили с использованием реакционных смесей из комбинации 1 - комбинации 5, полученных в примере 2.
[474]
[475] <ПЦР с детекцией в реальном времени с использованием реакционной смеси из комбинации 1>
[476] Пробирку, содержащую реакционную смесь из комбинации 1, помещали в термоциклер с детекцией в реальном времени (термоциклер CFX96 с детекцией в реальном времени, Bio-Rad) и инкубировали при 50°C в течение 4 минут, и денатурировали при 95°C в течение 15 минут, и затем подвергали 50 повторениям цикла из 15 секунд при 57°C, 1 секунды при 50°C, 10 секунд при 72°C, 1 секунды при 85°C и 10 секунд при 95°C. Детекцию сигналов проводили при 50°C (первая температура детекции) и 72°C (вторая температура детекции) в каждом цикле.
[477] Изменение сигнала измеряли посредством сравнения между случаями, когда нуклеиновая кислота-мишень присутствует, и когда она не присутствует. В этой связи, на основании значения сигнала отрицательного контроля (то есть, RFU представляет собой 0), если значение сигнала при первой температуре детекции больше или равно установленному пороговому значению 300, сигнал считали изменившимся, и если значение сигнала при второй температуре детекции было меньше или равно установленному пороговому значению -300, сигнал считали изменившимся.
[478] В результате, как показано на ФИГ. 13, в пробирке 1, содержащей нуклеиновую кислоту-мишень CT, изменение сигнала идентифицировано при первой температуре детекции, и в пробирке 2, содержащей нуклеиновую кислоту-мишень NG, изменение сигнала идентифицировано при второй температуре детекции, и в пробирке 3, содержащей нуклеиновые кислоты-мишени CT и NG, изменение сигнала идентифицировано при первой температуре детекции и второй температуре детекции, соответственно.
[479] Между тем, отрицательный контроль, в пробирке 4, обеспечивал постоянный сигнал в ходе реакций амплификации при обеих первой температуре детекции и второй температуре детекции. То есть, идентифицировано отсутствие изменения сигнала.
[480]
[481] [Таблица 3]
[482] Пробирка 1: 500 пг геномной ДНК CT;
[483] Пробирка 2: 500 пг геномной ДНК NG;
[484] Пробирка 3: смесь 500 пг геномной ДНК CT и 500 пг геномной ДНК NG;
[485] Пробирка 4: Отрицательный контроль;
[486] N/A: Неприменимо
[487]
[488] Также, как показано в таблице 3, не присутствовало различия между значением Ct (порогового цикла) (28,53), показательным для присутствия CT, в пробирке 1, содержащей только нуклеиновую кислоту-мишень CT, и значением Ct (28,27), показательным для присутствия CT, в пробирке 3, содержащей обе нуклеиновые кислоты-мишени CT и NG, и не присутствовало различия между значением Ct (29,59), показательным для присутствия NG, в пробирке 2, содержащей только нуклеиновую кислоту-мишень NG, и значением Ct (29,28), показательным для присутствия NG, в пробирке 3, содержащей обе нуклеиновые кислоты-мишени CT и NG.
[489] Эти результаты показывают, что, при каждой температуре детекции, обеспечивается только изменение сигнала, которое является зависимым от присутствия соответствующей нуклеиновой кислоты-мишени, без обеспечения изменений сигналов, зависимых от присутствия других нуклеиновых кислот-мишеней. То есть, эти результаты показывают, что каждую из множества различных нуклеиновых кислот-мишеней можно независимо детектировать при соответствующей ей температуре детекции. Соответственно, способ в соответствии с настоящим изобретением имеет те преимущества, что им можно определять присутствие конкретной нуклеиновой кислоты-мишени посредством только изменения сигнала, детектированного при конкретной температуре детекции, без учета изменений сигналов, детектированных при температурах детекции, отличных от конкретной температуры детекции (например, температурах детекции, отличных от конкретной температуры детекции, то есть, температурах детекции, обеспечивающих изменения сигналов, показательные для присутствия других нуклеиновых кислот-мишеней).
[490]
[491] <ПЦР с детекцией в реальном времени с использованием реакционной смеси из комбинации 2>
[492] Пробирку, содержащую реакционную смесь из комбинации 2, помещали в термоциклер с детекцией в реальном времени (термоциклер CFX96 с детекцией в реальном времени) и инкубировали при 50°C в течение 4 минут, и денатурировали при 95°C в течение 15 минут, и затем подвергали 50 повторениям цикла из 15 секунд при 57°C, 1 секунды при 50°C, 10 секунд при 72°C, 1 секунды при 85°C и 10 секунд при 95°C. Детекцию сигналов проводили при 57°C (первая температура детекции) и 85°C (вторая температура детекции) в каждом цикле.
[493] Изменение сигнала измеряли посредством сравнения между случаями, когда нуклеиновая кислота-мишень присутствует, и когда она не присутствует. В этой связи, на основании значения сигнала отрицательного контроля (то есть, RFU представляет собой 0), если значение сигнала при первой температуре детекции и второй температуре детекции больше или равно установленному пороговому значению 300, сигнал считали изменившимся.
[494] В результате, как показано на ФИГ. 14, в пробирке 1, содержащей нуклеиновую кислоту-мишень CT, изменение сигнала идентифицировано при первой температуре детекции, и в пробирке 2, содержащей нуклеиновую кислоту-мишень UP, изменение сигнала идентифицировано при второй температуре детекции, и в пробирке 3, содержащей нуклеиновые кислоты-мишени CT и UP, изменение сигнала идентифицировано при первой температуре детекции и второй температуре детекции, соответственно.
[495] Между тем, для отрицательного контроля, пробирки 4, показан постоянный сигнал в ходе реакций амплификации при обеих первой температуре детекции и второй температуре детекции. То есть, детектировано отсутствие изменения сигнала.
[496]
[497] [Таблица 4]
[498] Пробирка 1: 50 пг геномной ДНК CT;
[499] Пробирка 2: 50 пг геномной ДНК UP;
[500] Пробирка 3: смесь 50 пг геномной ДНК CT и 50 пг геномной ДНК UP;
[501] Пробирка 4: Отрицательный контроль;
[502] N/A: Неприменимо
[503]
[504] Также, как показано в таблице 4, не присутствовало различия между значением Ct (порогового цикла) (34,29), показательным для присутствия CT, в пробирке 1, содержащей только нуклеиновую кислоту-мишень CT, и значением Ct (33,83), показательным для присутствия CT, в пробирке 3, содержащей обе нуклеиновые кислоты-мишени CT и UP, и не присутствовало различия между значением Ct (33,93), показательным для присутствия UP, в пробирке 2, содержащей только нуклеиновую кислоту-мишень UP, и значением Ct (33,47), показательным для присутствия UP, в пробирке 3, содержащей обе нуклеиновые кислоты-мишени CT и UP.
[505] Эти результаты показывают, что, как описано выше, каждую из множества различных нуклеиновых кислот-мишеней можно независимо детектировать при соответствующей ей температуре детекции.
[506]
[507] <ПЦР с детекцией в реальном времени с использованием реакционной смеси из комбинации 3>
[508] Пробирку, содержащую реакционную смесь из комбинации 3, помещали в термоциклер с детекцией в реальном времени (термоциклер CFX96 с детекцией в реальном времени) и инкубировали при 50°C в течение 4 минут, и денатурировали при 95°C в течение 15 минут, и затем подвергали 50 повторениям процесса реакции из 15 секунд при 57°C, 1 секунды при 50°C, 10 секунд при 72°C, 1 секунды при 85°C и 10 секунд при 95°C. Детекцию сигналов проводили при 57°C (первая температура детекции) и 72°C (вторая температура детекции) в каждом цикле.
[509] Изменение сигнала измеряли посредством сравнения между случаями, когда нуклеиновая кислота-мишень присутствует, и когда она не присутствует. В этой связи, на основании значения сигнала отрицательного контроля (то есть, RFU представляет собой 0), если значение сигнала при первой температуре детекции и второй температуре детекции было меньше или равно установленному пороговому значению -300, сигнал считали изменившимся.
[510] В результате, как показано на ФИГ. 15, в пробирке 1, содержащей нуклеиновую кислоту-мишень CT, изменение сигнала идентифицировано при первой температуре детекции, и в пробирке 2, содержащей нуклеиновую кислоту-мишень NG, изменение сигнала идентифицировано при второй температуре детекции, и в пробирке 3, содержащей нуклеиновые кислоты-мишени CT и NG, изменение сигнала идентифицировано при первой температуре детекции и второй температуре детекции, соответственно.
[511] Между тем, для отрицательного контроля, пробирки 4, показан постоянный сигнал в ходе реакций амплификации при обеих первой температуре детекции и второй температуре детекции. То есть, детектировано отсутствие изменения сигнала.
[512]
[513] [Таблица 5]
[514] Пробирка 1: 500 пг геномной ДНК CT;
[515] Пробирка 2: 500 пг геномной ДНК NG;
[516] Пробирка 3: смесь 500 пг геномной ДНК CT и 500 пг геномной ДНК NG;
[517] Пробирка 4: Отрицательный контроль;
[518] N/A: Неприменимо
[519]
[520] Также, как показано в таблице 5 выше, не присутствовало различия между значением Ct (порогового цикла) (30,62), показательным для присутствия CT, в пробирке 1, содержащей только нуклеиновую кислоту-мишень CT, и значением Ct (30,33), показательным для присутствия CT, в пробирке 3, содержащей обе нуклеиновые кислоты-мишени CT и NG, и не присутствовало различия между значением Ct (28,33), показательным для присутствия NG, в пробирке 2, содержащей только нуклеиновую кислоту-мишень NG, и значением Ct (28,17), показательным для присутствия NG, в пробирке 3, содержащей обе нуклеиновые кислоты-мишени CT и NG.
[521] Эти результаты показывают, что, как описано выше, каждую из множества различных нуклеиновых кислот-мишеней можно независимо детектировать при соответствующей ей температуре детекции.
[522]
[523] <ПЦР с детекцией в реальном времени с использованием реакционной смеси из комбинации 4>
[524] Пробирку, содержащую реакционную смесь из комбинации 4, помещали в термоциклер с детекцией в реальном времени (термоциклер CFX96 с детекцией в реальном времени) и инкубировали при 50°C в течение 4 минут, и денатурировали при 95°C в течение 15 минут, и затем подвергали 50 повторениям процесса реакции из 15 секунд при 57°C, 1 секунды при 50°C, 10 секунд при 72°C, 1 секунды при 85°C и 10 секунд при 95°C. Детекцию сигналов проводили при 72°C (первая температура детекции) и 95°C (вторая температура детекции) в каждом цикле.
[525] Изменение сигнала измеряли посредством сравнения между случаями, когда нуклеиновая кислота-мишень присутствует, и когда она не присутствует. В этой связи, на основании значения сигнала отрицательного контроля (то есть, RFU представляет собой 0), если значение сигнала при первой температуре детекции меньше или равно установленному пороговому значению -300, сигнал считали изменившимся, и если значение сигнала при второй температуре детекции больше или равно установленному пороговому значению 300, сигнал считали изменившимся.
[526] В результате, как показано на ФИГ. 16, в пробирке 1, содержащей нуклеиновую кислоту-мишень NG, изменение сигнала идентифицировано при первой температуре детекции, и в пробирке 2, содержащей нуклеиновую кислоту-мишень UP, изменение сигнала идентифицировано при второй температуре детекции, и в пробирке 3, содержащей нуклеиновые кислоты-мишени NG и UP, изменение сигнала идентифицировано при первой температуре детекции и второй температуре детекции, соответственно.
[527] Между тем, для отрицательного контроля, пробирки 4, показан постоянный сигнал в ходе реакций амплификации при обеих первой температуре детекции и второй температуре детекции. То есть, детектировано отсутствие изменения сигнала.
[528]
[529] [Таблица 6]
[530] Пробирка 1: 500 пг геномной ДНК NG;
[531] Пробирка 2: 500 пг геномной ДНК UP;
[532] Пробирка 3: смесь 500 пг геномной ДНК NG и 500 пг геномной ДНК UP;
[533] Пробирка 4: Отрицательный контроль;
[534] N/A: Неприменимо
[535]
[536] Также, как показано в таблице 6, не присутствовало различия между значением Ct (порогового цикла) (29,62), показательным для присутствия NG в пробирке 1, содержащей только нуклеиновую кислоту-мишень NG, и значением Ct (30,72), показательным для присутствия NG, в пробирке 3, содержащей обе нуклеиновые кислоты-мишени NG и UP, и не присутствовало различия между значением Ct (33,42), показательным для присутствия UP, в пробирке 2, содержащей только нуклеиновую кислоту-мишень UP, и значением Ct (32,68), показательным для присутствия UP, в пробирке 3, содержащей обе нуклеиновые кислоты-мишени NG и UP.
[537] Эти результаты показывают, что, как описано выше, каждую из множества различных последовательностей нуклеиновых кислот-мишеней можно независимо детектировать при соответствующей ей температуре детекции.
[538]
[539] <ПЦР с детекцией в реальном времени с использованием реакционной смеси из комбинации 5>
[540] Пробирку, содержащую реакционную смесь из комбинации 5, помещали в термоциклер с детекцией в реальном времени (термоциклер CFX96 с детекцией в реальном времени) и инкубировали при 50°C в течение 4 минут, и денатурировали при 95°C в течение 15 минут, и затем подвергали 50 повторениям процесса реакции из 15 секунд при 57°C, 1 секунды при 50°C, 10 секунд при 72°C, 1 секунды при 85°C и 10 секунд при 95°C. Детекцию сигналов проводили при 50°C (первая температура детекции), 72°C (вторая температура детекции) и 95°C (третья температура детекции) в каждом цикле.
[541] Изменение сигнала измеряли посредством сравнения между случаями, когда нуклеиновая кислота-мишень присутствует, и когда она не присутствует. В этой связи, на основании значения сигнала отрицательного контроля (то есть, RFU представляет собой 0), если значение сигнала при первой температуре детекции и третьей температуре детекции было больше или равно установленному пороговому значению 300, сигнал считали изменившимся, и если значение сигнала при второй температуре детекции было меньше или равно установленному пороговому значению -300, сигнал считали изменившимся.
[542] В результате, как показано на ФИГ. 17a и ФИГ. 17b, в пробирке 1, содержащей нуклеиновую кислоту-мишень CT, изменение сигнала идентифицировано при первой температуре детекции, и в пробирке 2, содержащей нуклеиновую кислоту-мишень NG, изменение сигнала идентифицировано при второй температуре детекции, и в пробирке 3, содержащей нуклеиновую кислоту-мишень UP, изменение сигнала идентифицировано при третьей температуре детекции.
[543] Также, в пробирке 4 - пробирке 7, каждая из которых содержала по меньшей мере две из CT, NG и UP, изменение сигнала идентифицировано при соответствующей температуре детекции, в зависимости от нуклеиновых кислот-мишеней, присутствующих в пробирке.
[544] Между тем, для отрицательного контроля, пробирки 8, показан постоянный сигнал в ходе реакций амплификации при всех из первой, второй и третьей температур детекции. То есть, детектировано отсутствие изменения сигнала.
[545]
[546] [Таблица 7]
[547] Пробирка 1: 50 пг геномной ДНК CT;
[548] Пробирка 2: 50 пг геномной ДНК NG;
[549] Пробирка 3: 50 пг геномной ДНК UO;
[550] Пробирка 4: смесь 50 пг геномной ДНК CT и 50 пг геномной ДНК NG;
[551] Пробирка 5: смесь 50 пг геномной ДНК CT и 50 пг геномной ДНК UP;
[552] Пробирка 6: смесь 50 пг геномной ДНК NG и 50 пг геномной ДНК UP;
[553] Пробирка 7: смесь 50 пг геномной ДНК CT, 50 пг геномной ДНК NG и 50 пг геномной ДНК UP;
[554] Пробирка 8: Отрицательный контроль;
[555] N/A: Неприменимо
[556]
[557] Также, как показано в таблице 7 выше, не присутствовало различия значения Ct, когда конкретная нуклеиновая кислота-мишень присутствовала отдельно, и когда она присутствовала вместе с другими нуклеиновыми кислотами-мишенями (CT: 32,92 против 33,03 против 32,47 против 33,34; NG: 33,00 против 33,23 против 33,99 против 34,12; UP: 35,46 против 34,99 против 35,91 против 35,32).
[558] Эти результаты показывают, что, как описано выше, каждую из множества различных нуклеиновых кислот-мишеней можно независимо детектировать при соответствующей ей температуре детекции.
[559]
[560] Между тем, как описано выше в настоящем описании, температуру детекции можно выбирать в рамках температурного диапазона изменения сигнала композиции для детекции нуклеиновой кислоты-мишени. Кроме того, температуру детекции можно выбирать с учетом температурных диапазонов изменения сигнала других композиций для детекции нуклеиновых кислот-мишеней, используемых совместно.
[561] Для подтверждения того, что температуру детекции можно выбирать в различных точках в рамках температурного диапазона изменения сигнала, авторы настоящего изобретения устанавливали различную температуру детекции для CT в комбинации 2 и комбинации 5, с использованием в то же время одинаковой композиции в качестве композиции для детекции CT. То есть, температуру детекции для CT из комбинации 2 устанавливали на 57°C, и температуру детекции для CT из комбинации 5 устанавливали на 50°C. В результате, обнаружено, что комбинация 2 и комбинация 5 обе обеспечивают изменение сигнала, показательное для присутствия нуклеиновой кислоты-мишени CT, только при соответствующих ей температурах детекции (то есть, 57°C и 50°C). Также, в комбинации 2 и комбинации 5, устанавливали различную температуру детекции для UP, с использованием в то же время одинаковой композиции в качестве композиции для детекции UP. То есть, температуру детекции для UP из комбинации 2 устанавливали на 85°C, и температуру детекции для UP из комбинации 5 устанавливали на 95°C. В результате, обнаружено также, что изменение сигнала, показательное для присутствия нуклеиновой кислоты-мишени UP, было обеспечено только при связанных с ней соответствующих температурах детекции (85°C и 95°C).
[562] Кроме того, как описано выше в настоящем описании, температурный диапазон изменения сигнала и температурный диапазон постоянного сигнала можно контролировать посредством коррекции длины и/или последовательности дуплекса (например, коррекции Tm дуплекса), предоставленного посредством композиции для детекции нуклеиновой кислоты-мишени.
[563] Для подтверждения того, что температурный диапазон изменения сигнала и температурный диапазон постоянного сигнала композиции для детекции нуклеиновой кислоты-мишени можно контролировать посредством коррекции Tm дуплекса, авторы настоящего изобретения использовали композиции InterSC с одинаковым механизмом образования сигнала для различных нуклеиновых кислот-мишеней (CT и NG) в комбинации 3, и разработали последовательности олигонуклеотидов таким образом, что CT предоставляет дуплекс с относительно низкой Tm, и NG предоставляет дуплекс с относительно высокой Tm, так что даже с использованием композиций с одинаковым механизмом образования сигнала, детекцию можно осуществлять при различных температурах детекции (то есть, первой температуре детекции и второй температуре детекции).
[564] В результате, как показано в результате ПЦР с детекцией в реальном времени для комбинации 3, обнаружено, что каждая температура детекции обеспечивает только изменение сигнала, показательное для соответствующей ей нуклеиновой кислоты-мишени.
[565] Таким образом, в соответствии с настоящим изобретением, множество нуклеиновых кислот-мишеней можно детектировать в способе с детекцией в реальном времени с использованием одного типа метки, посредством использования по меньшей мере одного из вышеописанных трех типов механизмов образования сигнала и коррекции температурного диапазона изменения сигнала таким образом, что только сигнал, показательный для присутствия одной нуклеиновой кислоты-мишени, обеспечивают при каждой температуре детекции.
[566]
[567] При наличии описания предпочтительных вариантов осуществления настоящего изобретения, следует понимать, что их варианты и модификации, попадающие в содержание изобретения, могут стать очевидными специалисту в данной области. Таким образом, объем настоящего изобретения следует определять по пунктам прилагаемой формулы изобретения и их эквивалентам.
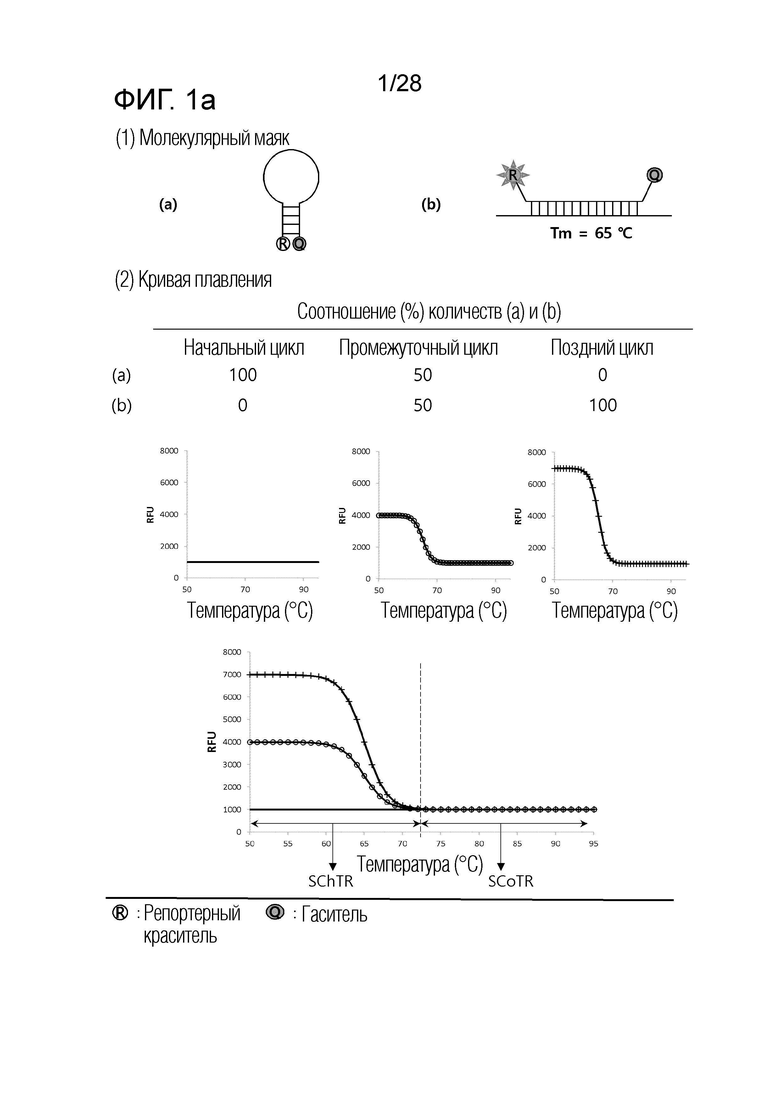
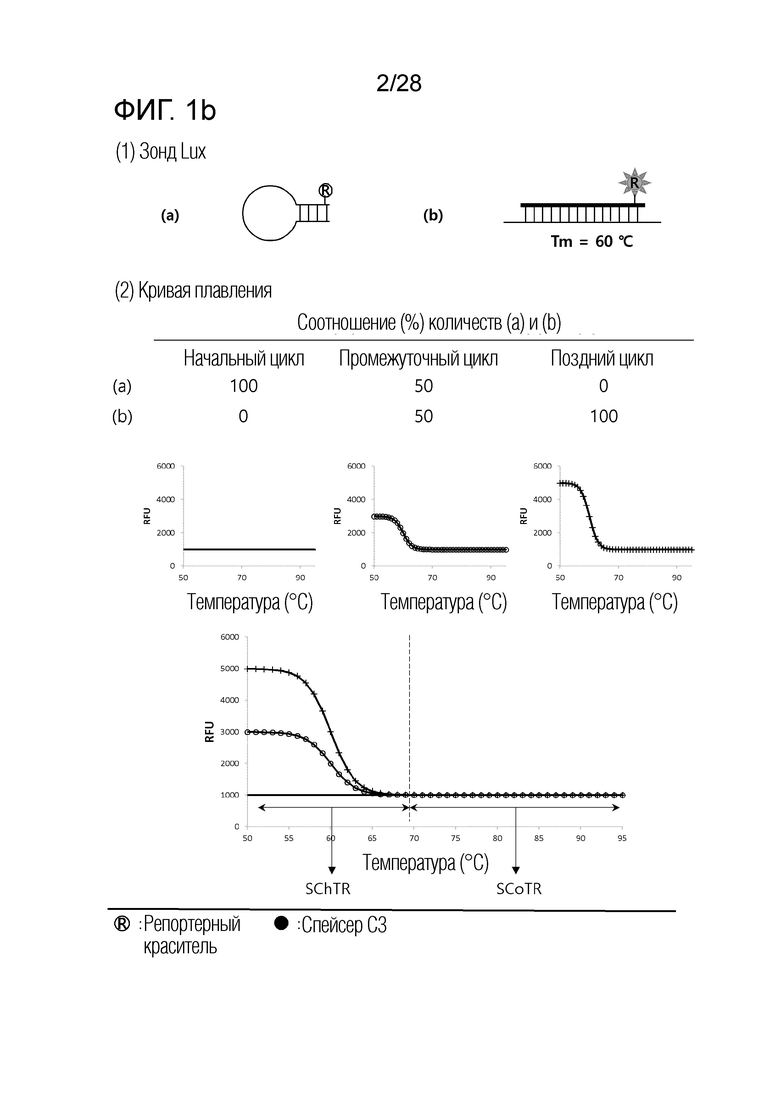
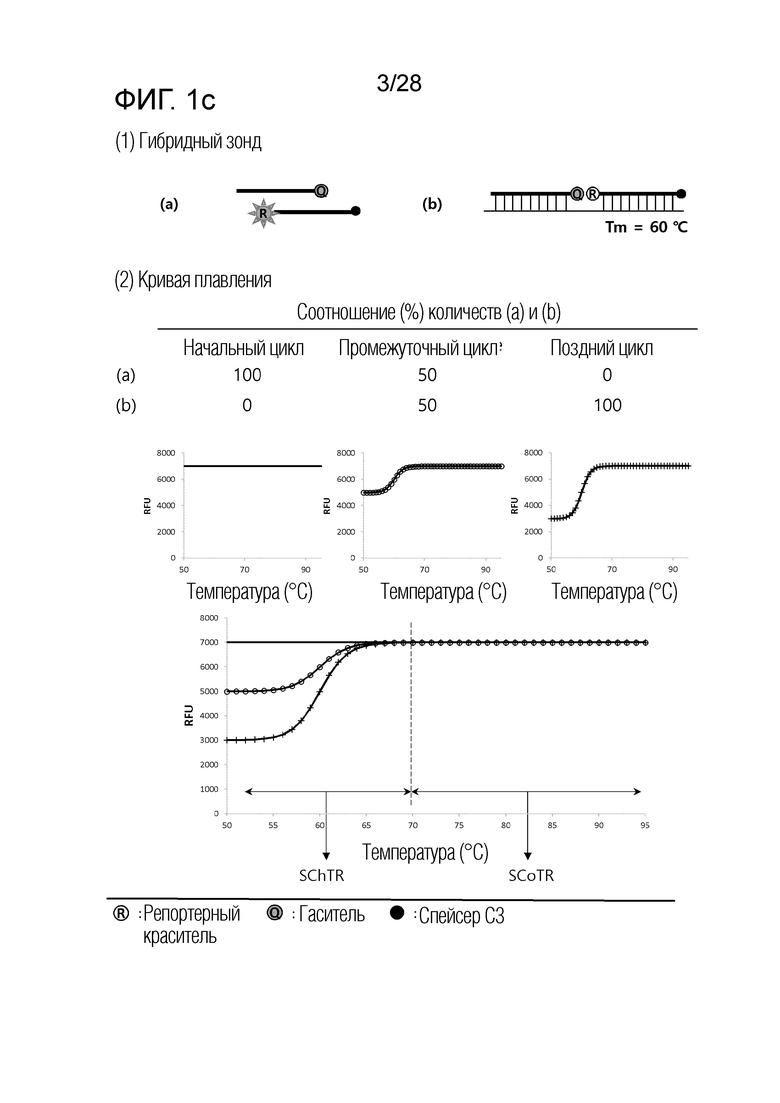
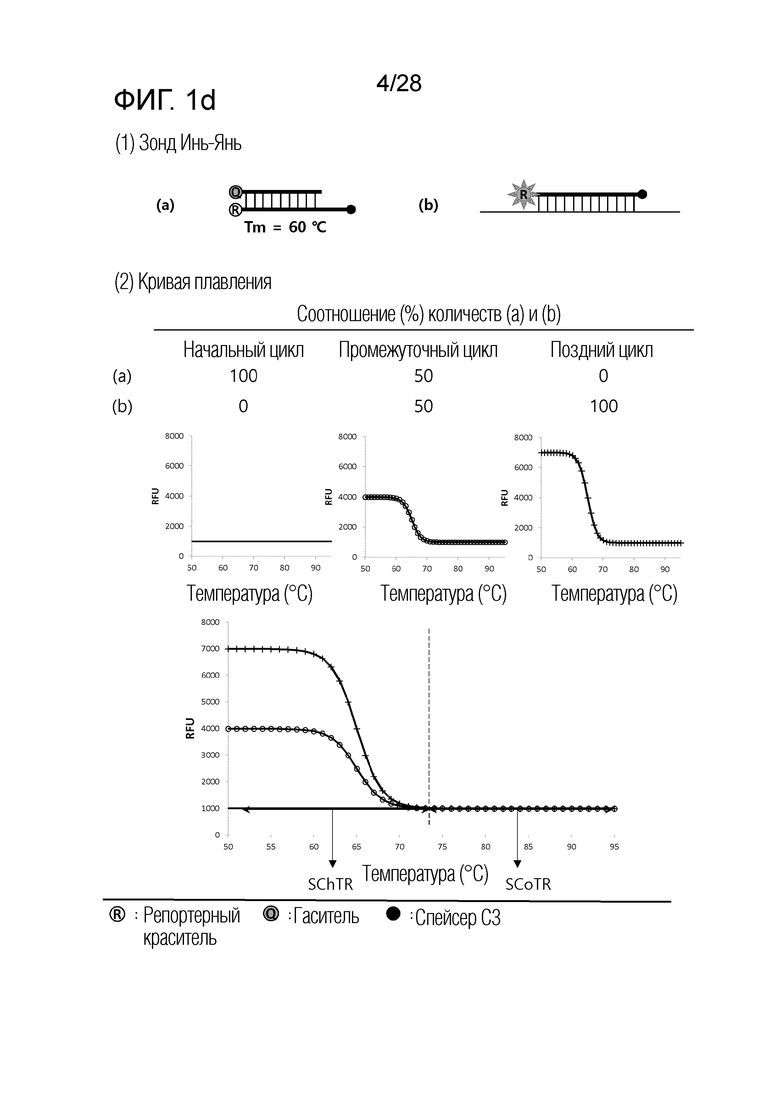
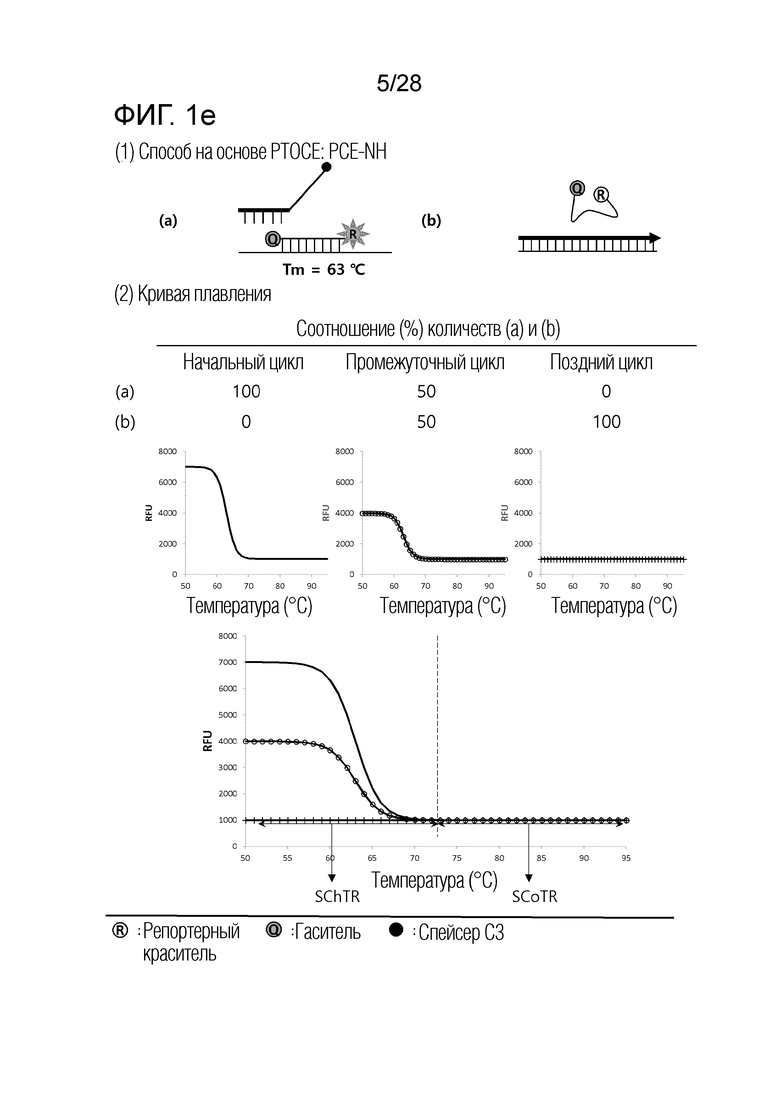
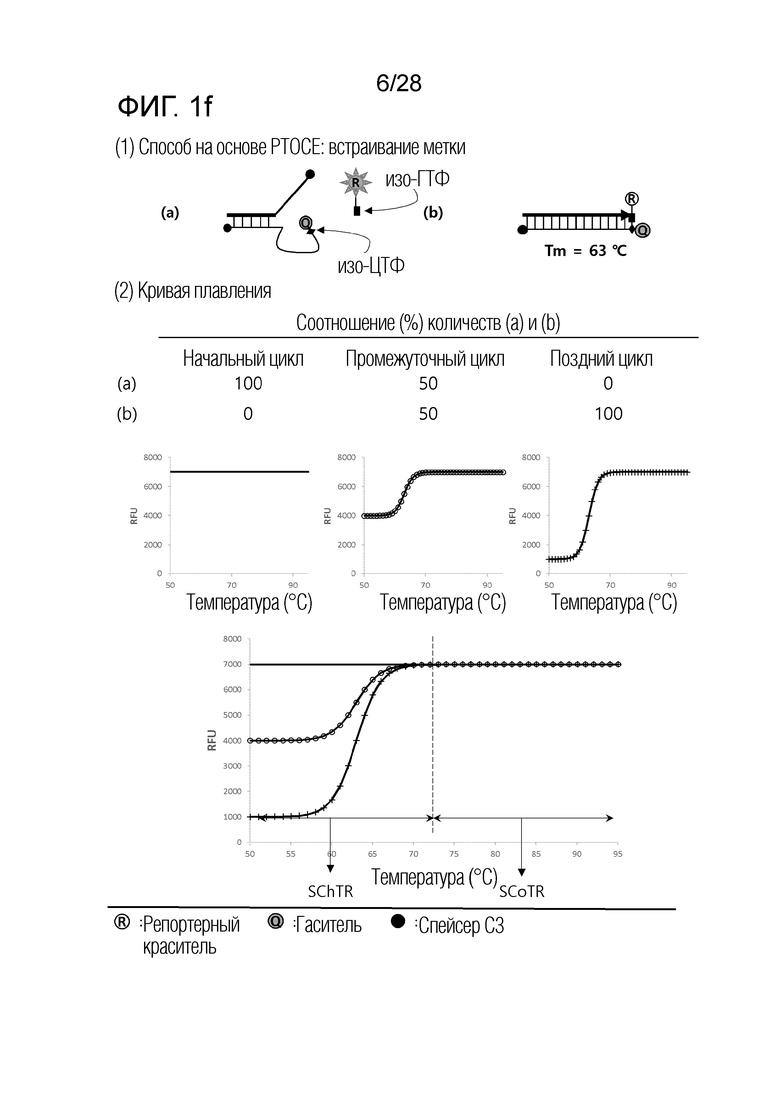
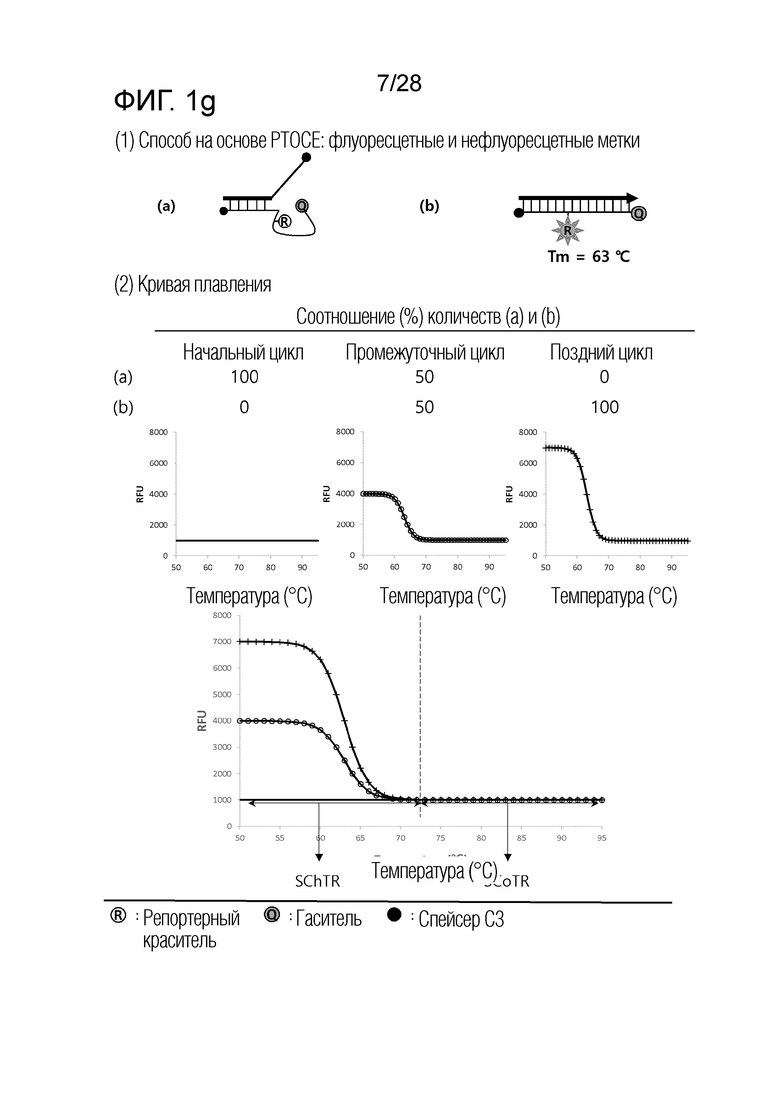
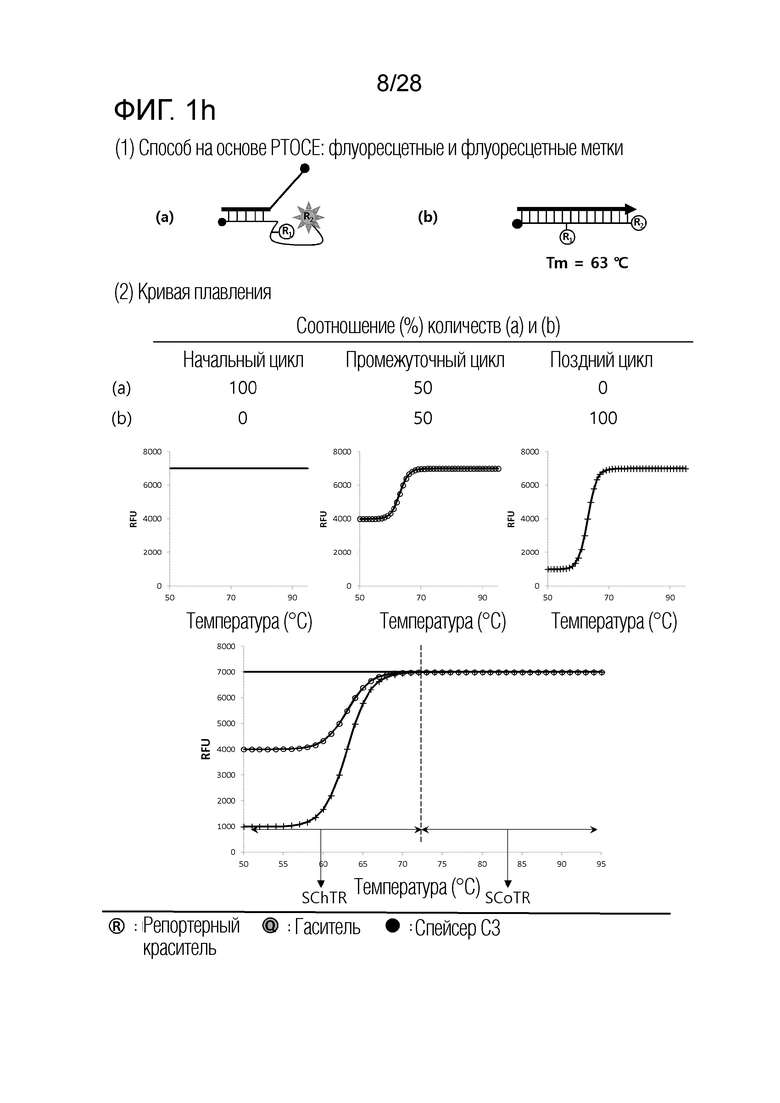

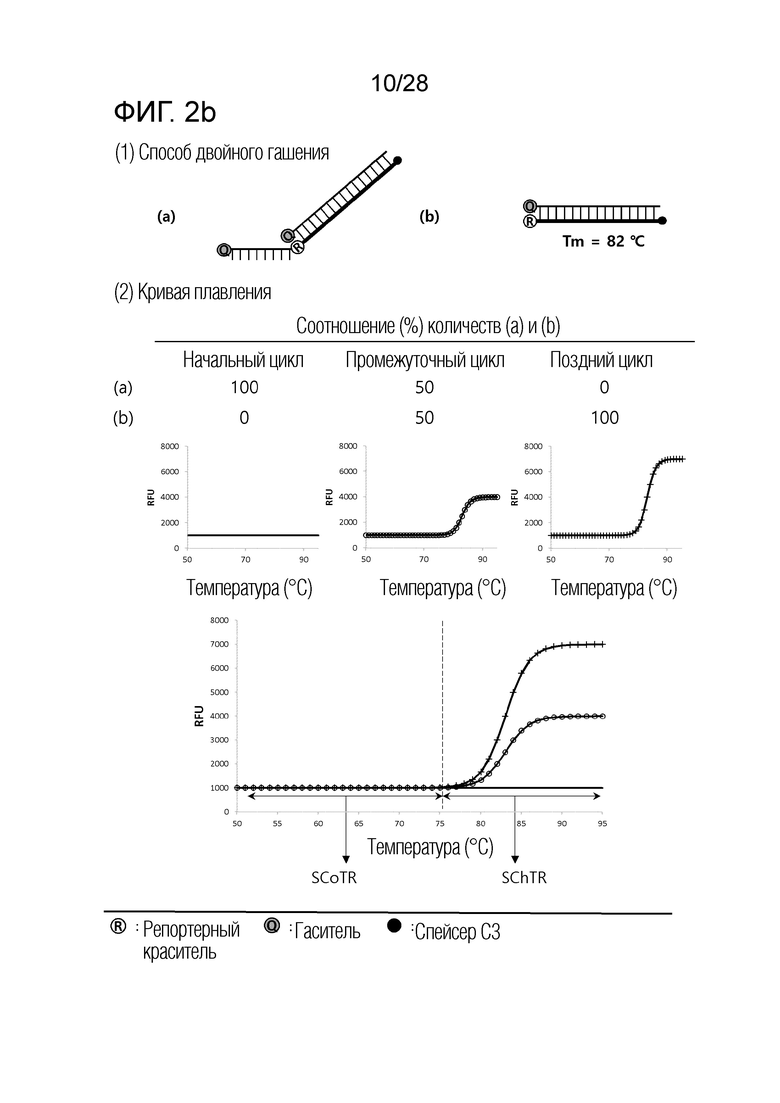
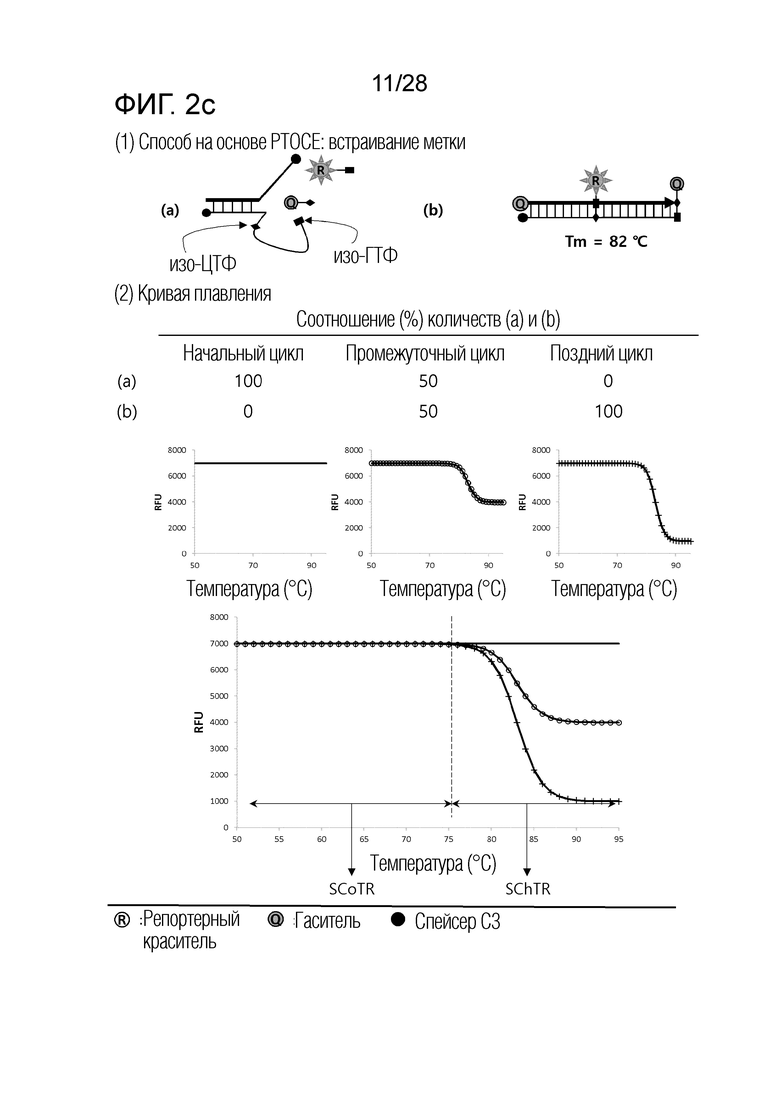
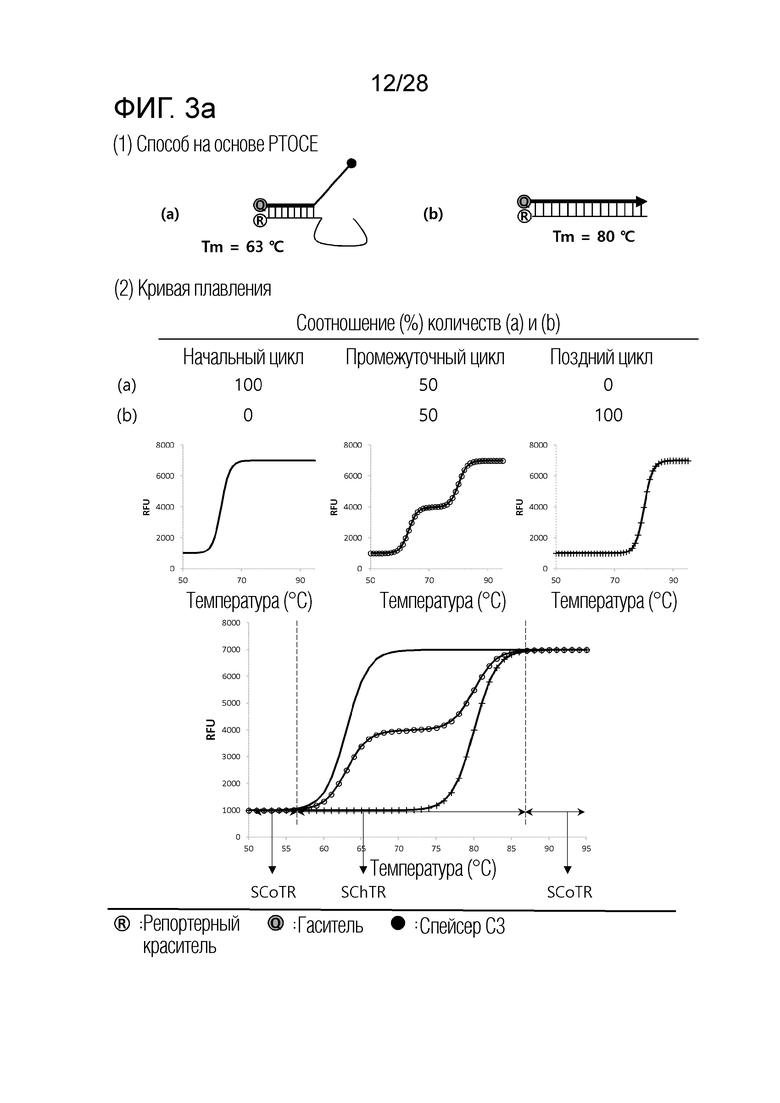
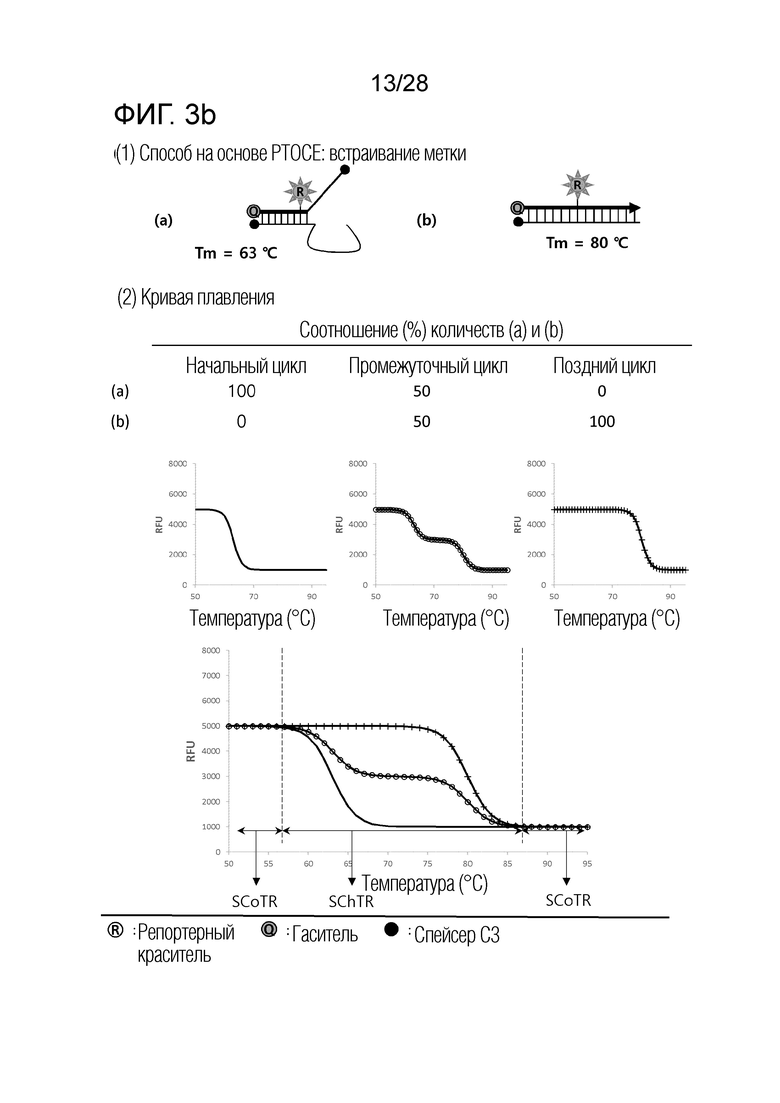
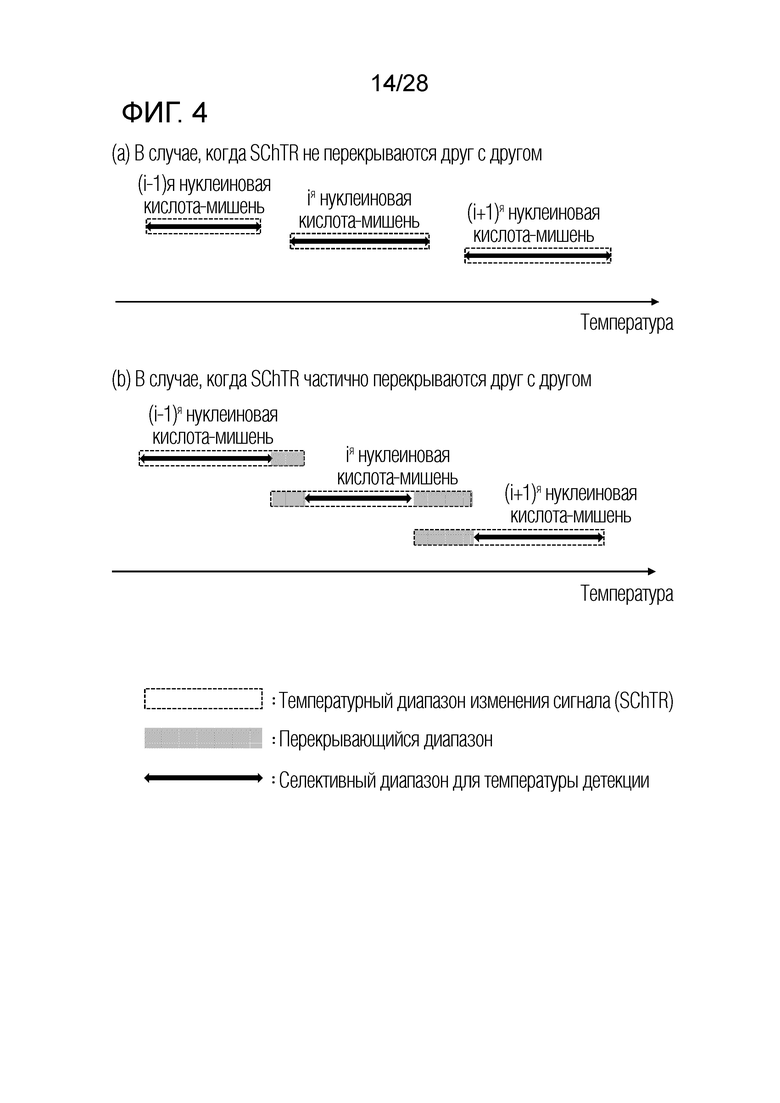
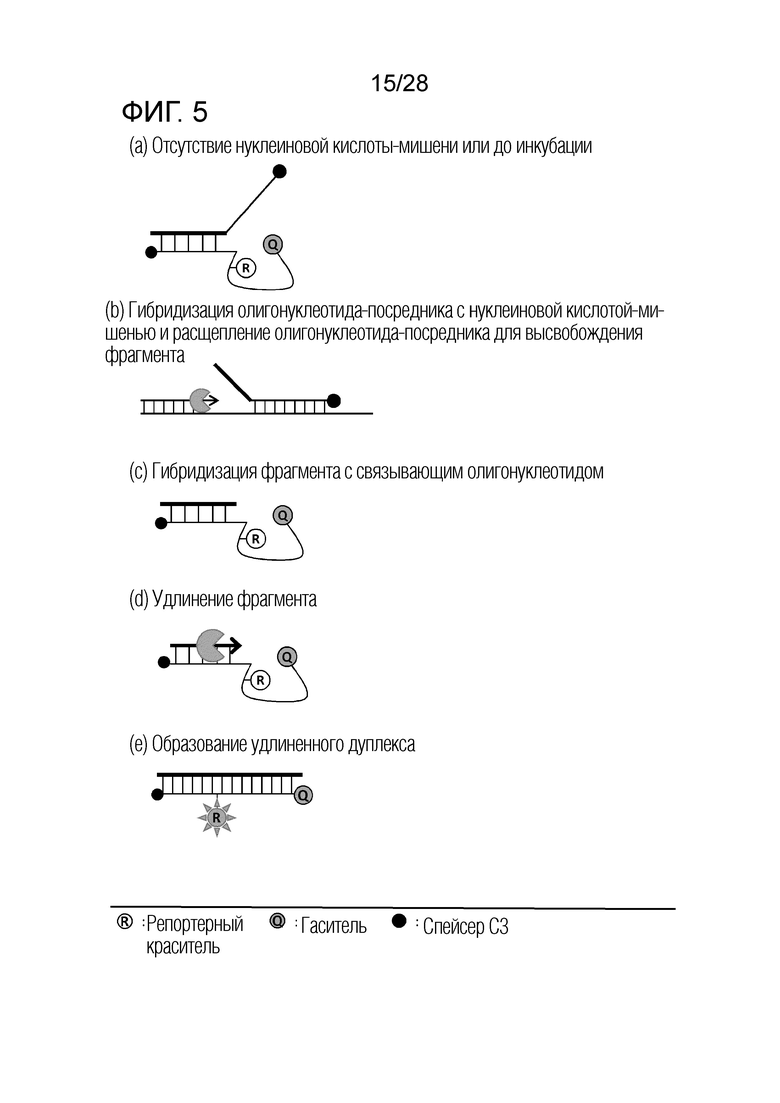
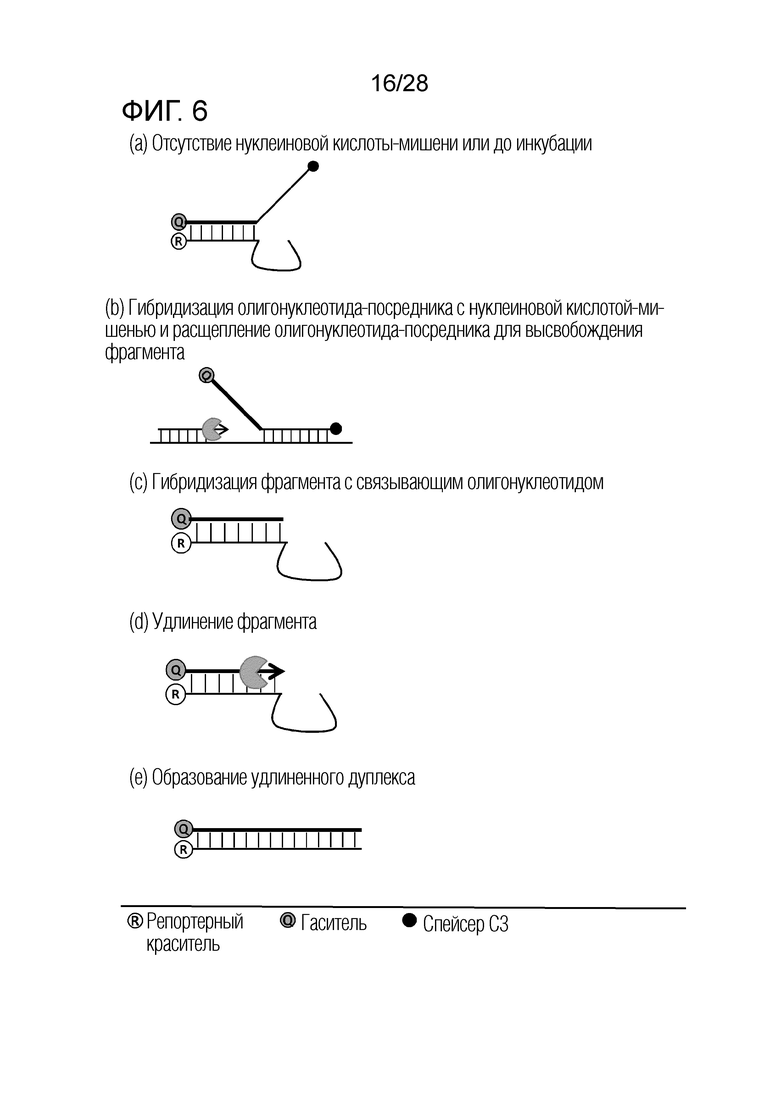
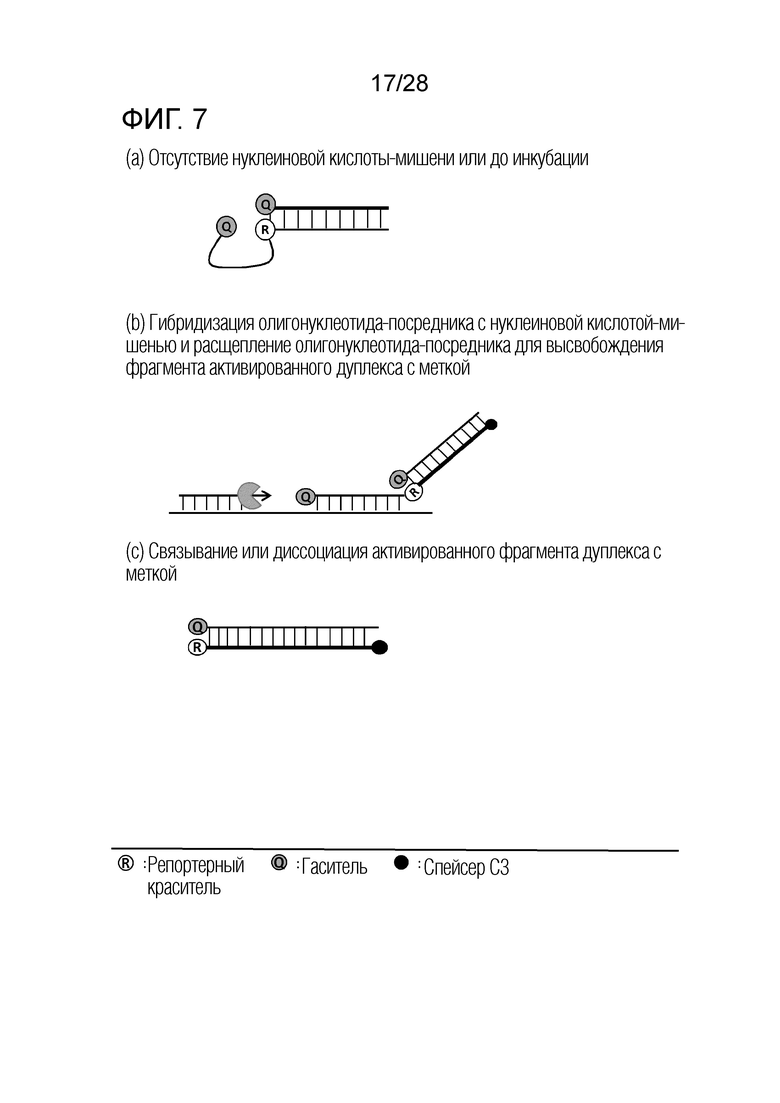
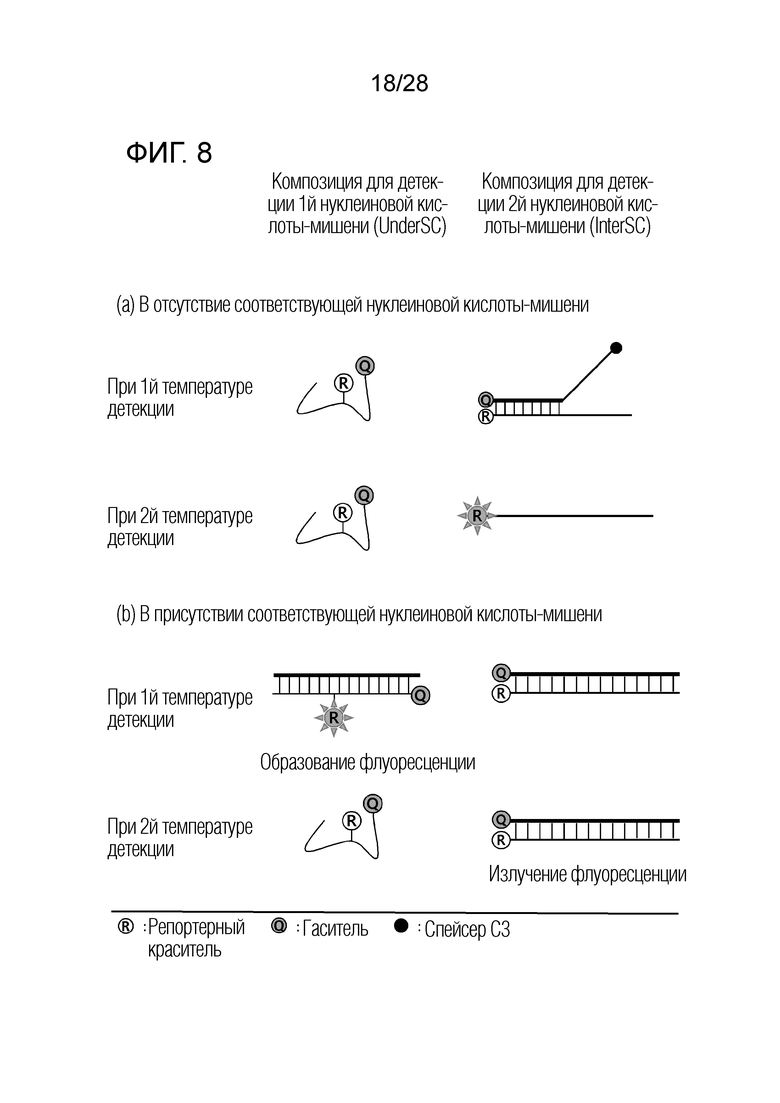
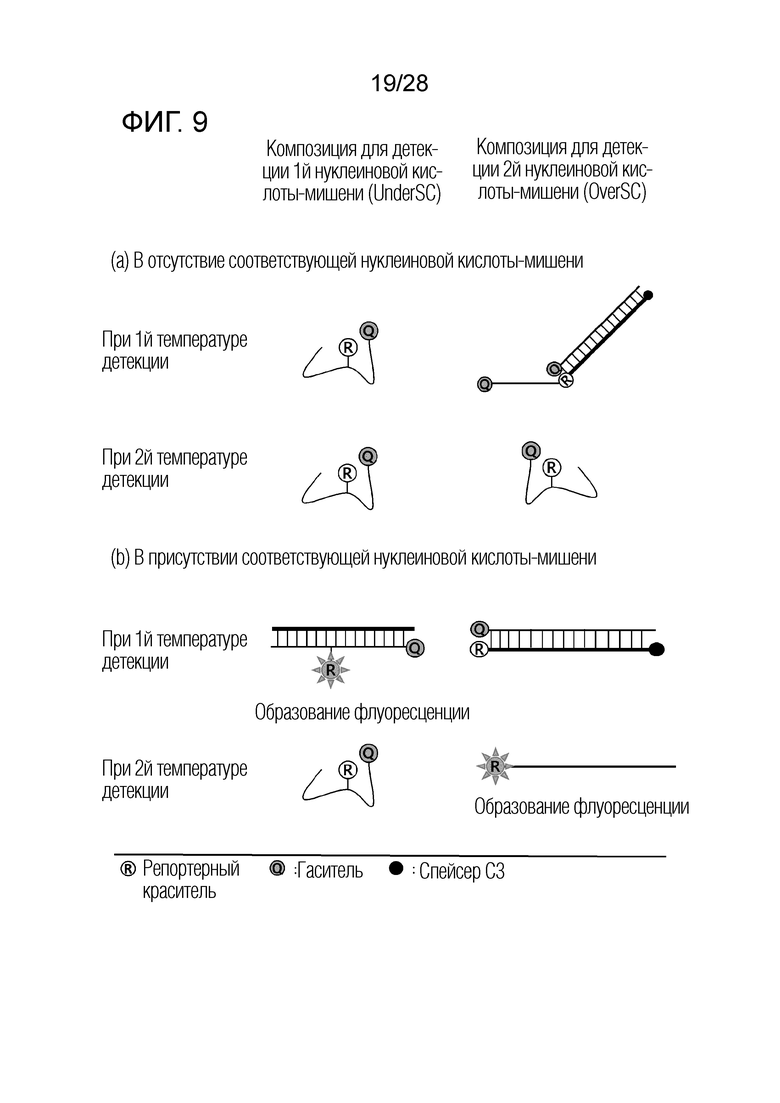
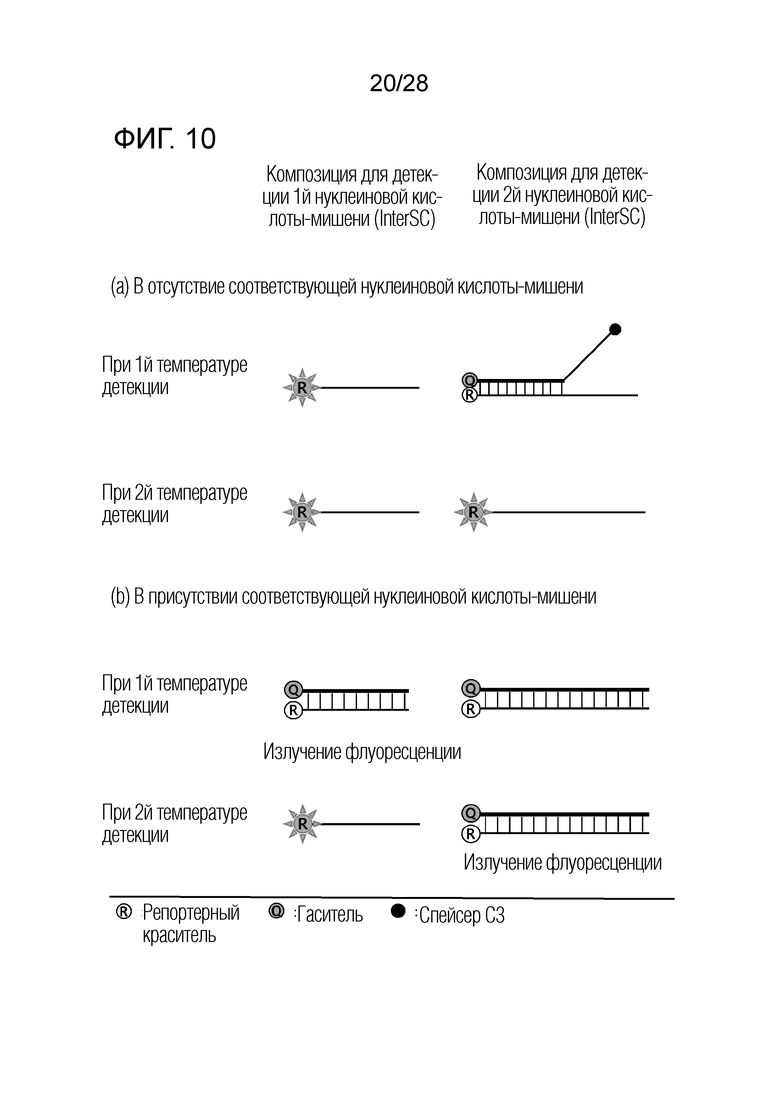
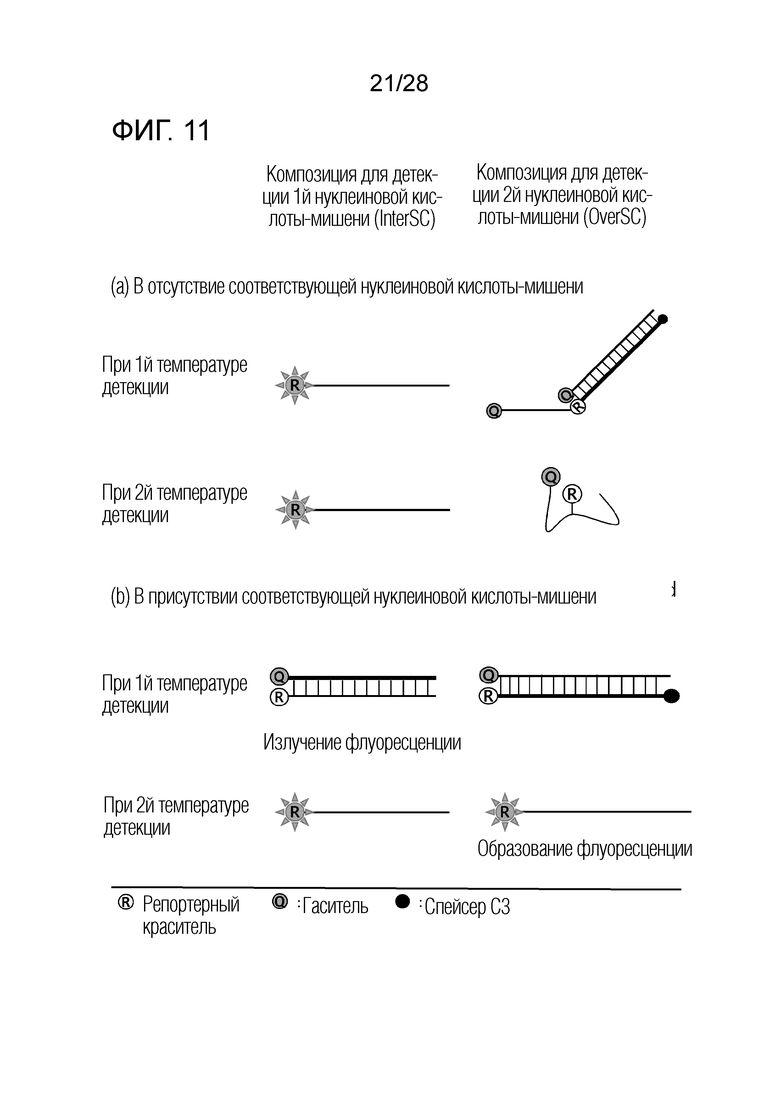
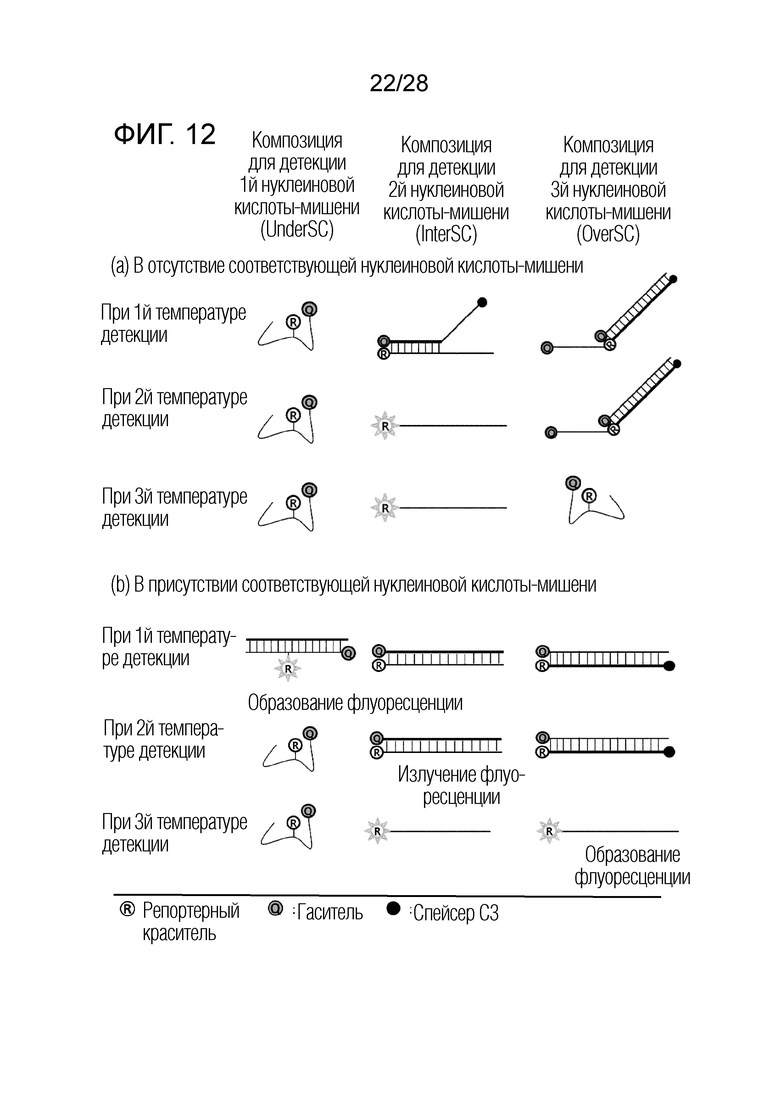
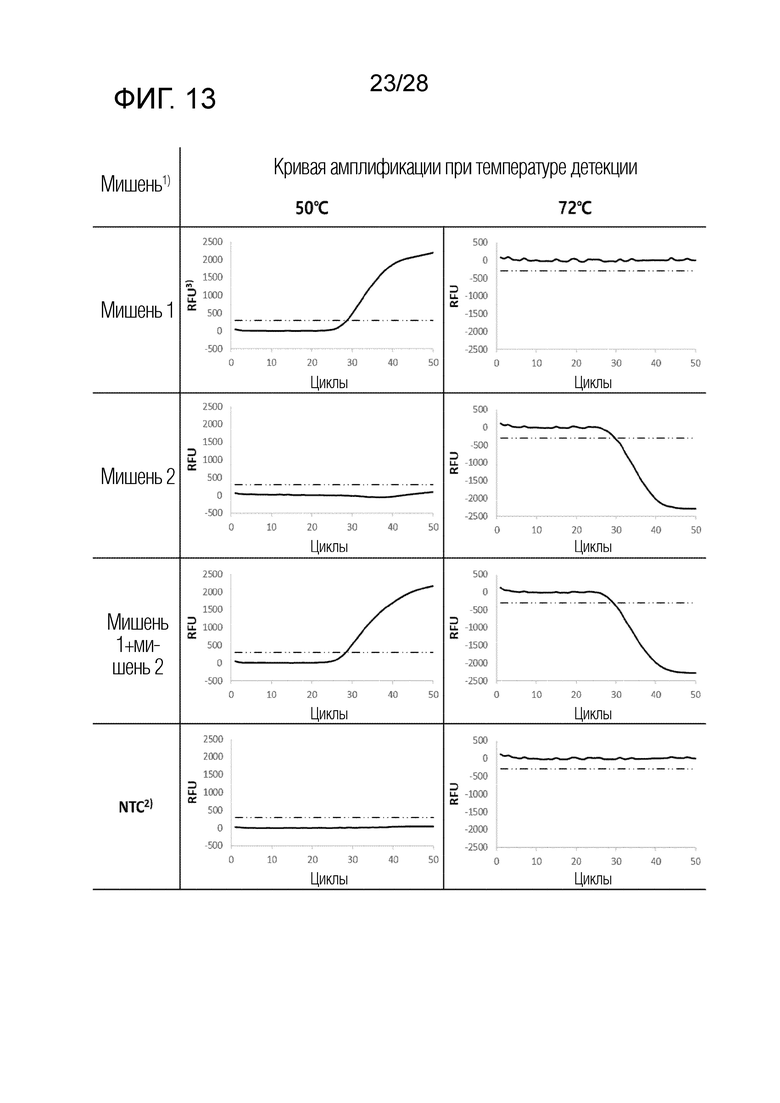
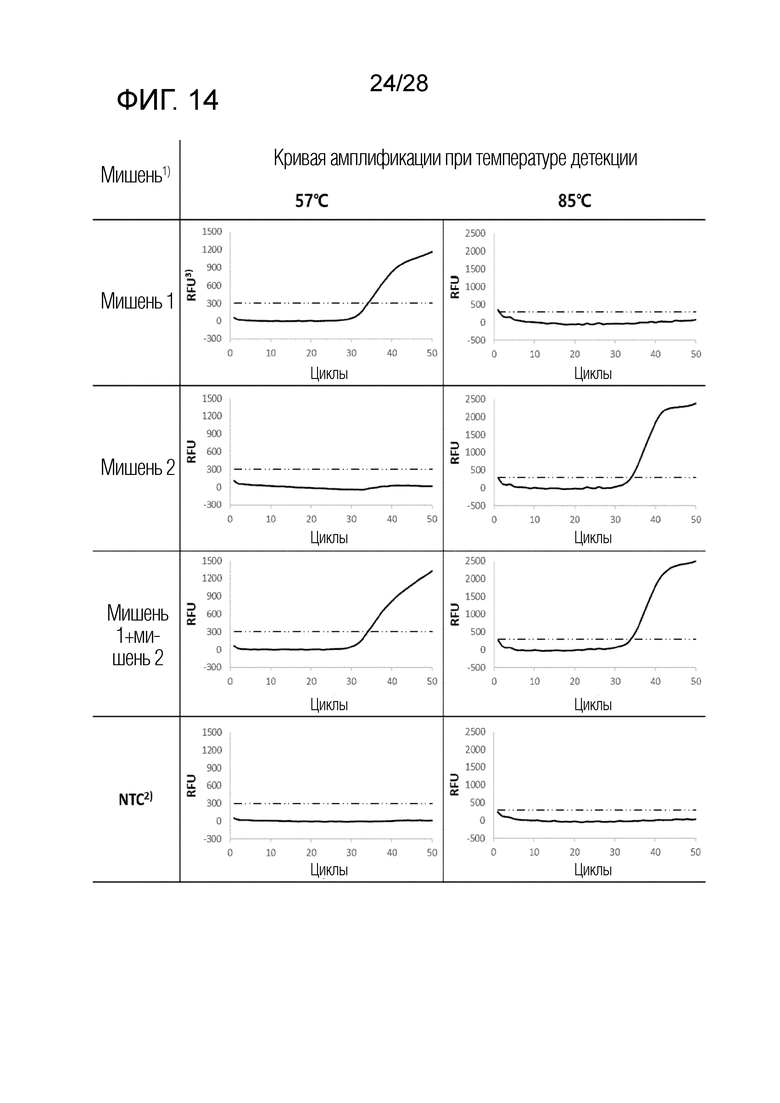
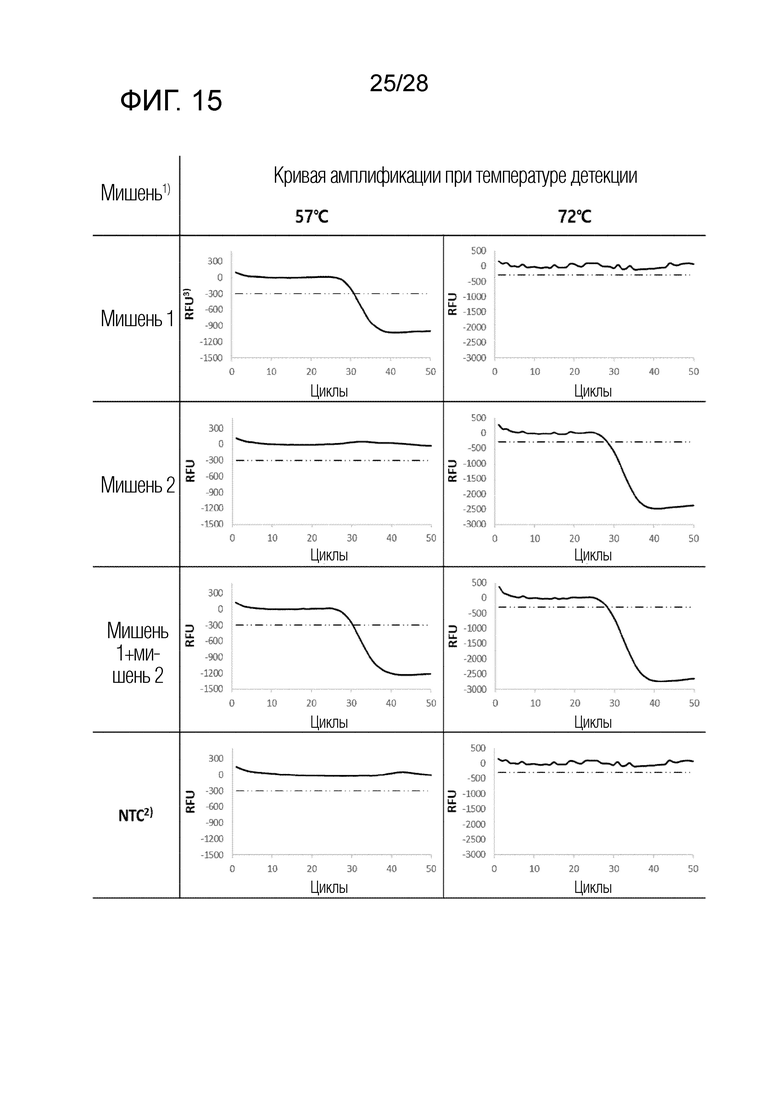
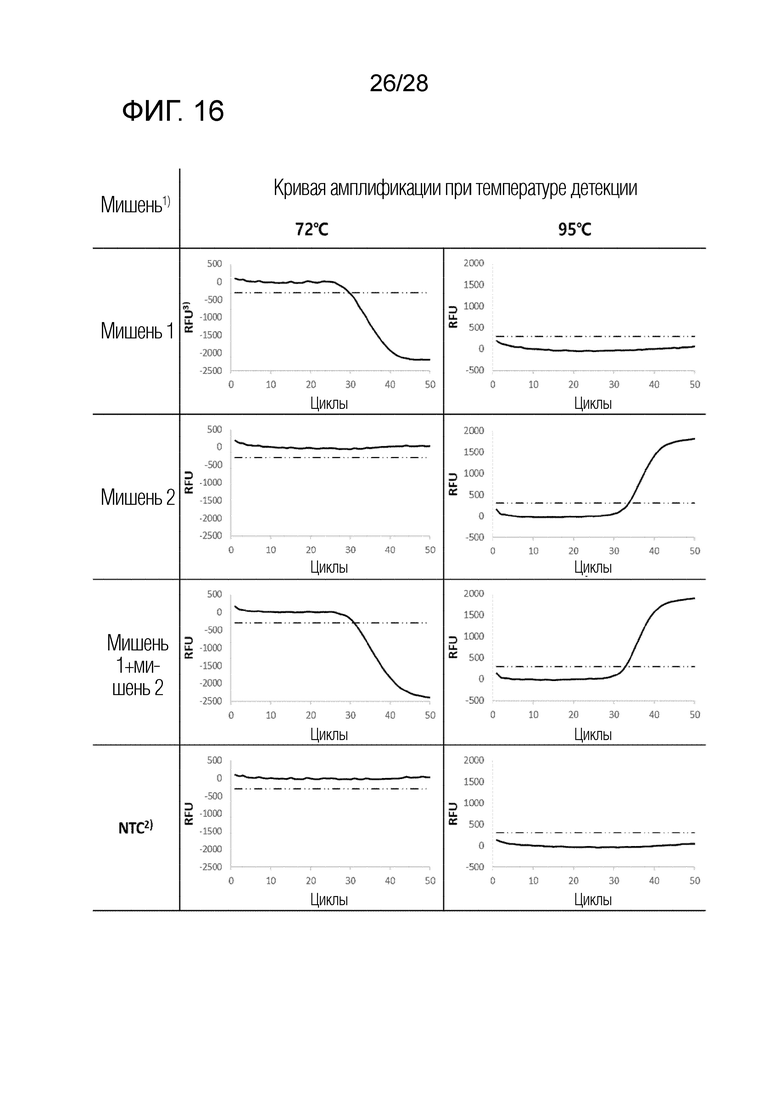
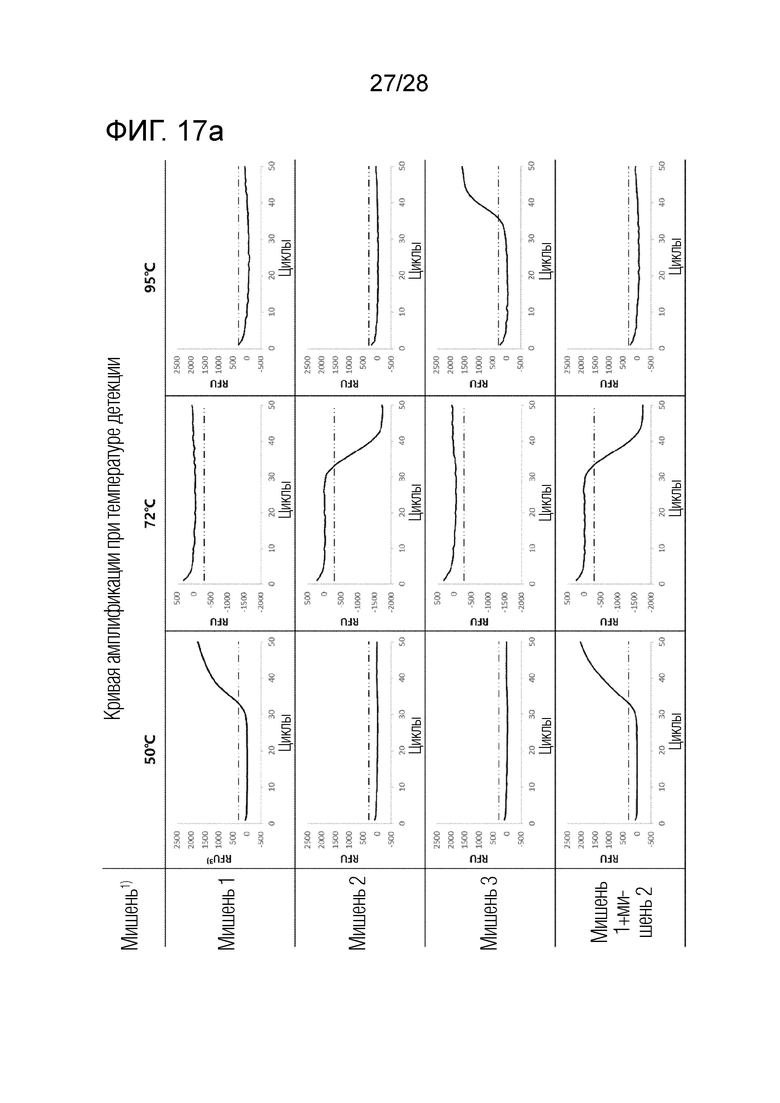
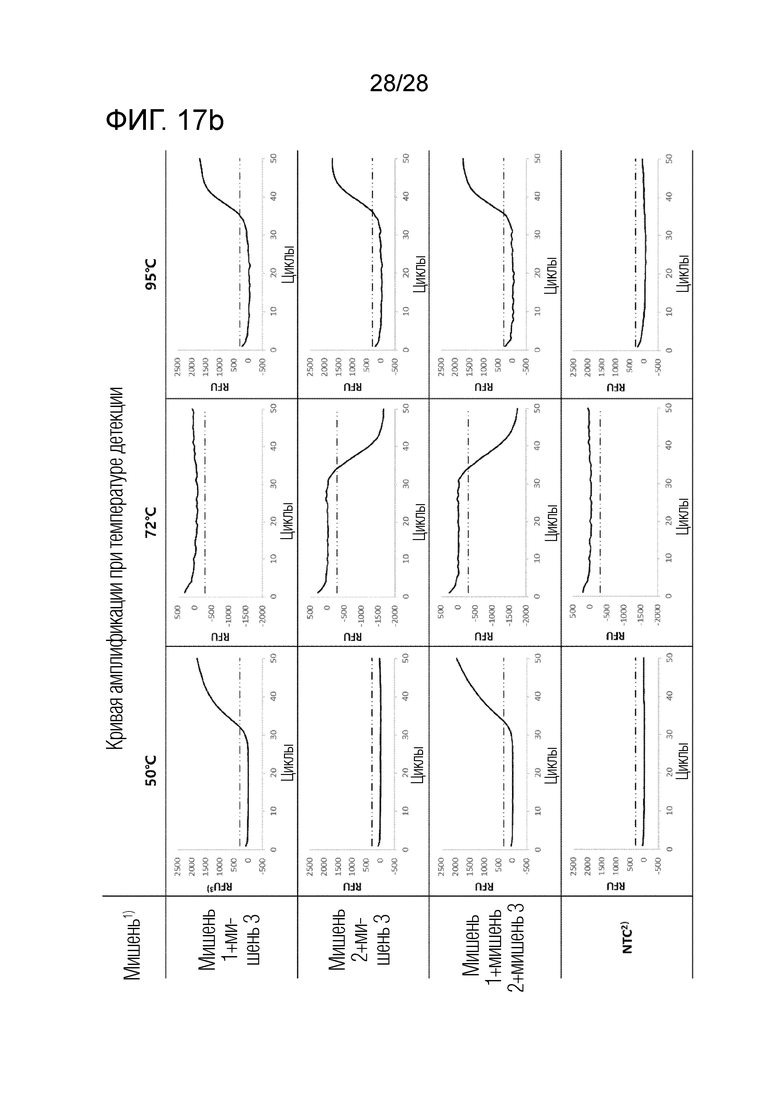
Изобретение относится к биотехнологии. Описан способ детекции множества нуклеиновых кислот-мишеней посредством только одного типа метки в одном реакционном сосуде посредством использования множества температур детекции. Обеспечивают изменение сигнала, зависимого от присутствия соответствующей нуклеиновой кислоты-мишени, при соответствующей температуре детекции каждой из нуклеиновых кислот-мишеней. Способ по настоящему изобретению позволяет не анализировать кривую плавления после амплификации мишени, даже с использованием одного типа метки, и таким образом может заметно уменьшать время анализа. 4 н. и 36 з.п. ф-лы, 17 ил., 7 табл.
1. Способ детекции n нуклеиновых кислот-мишеней в образце,
включающий:
(a) детекцию сигналов при n температурах детекции, с инкубацией в то же время с n композициями для детекции нуклеиновых кислот-мишеней, образца, как подозревают, содержащего по меньшей мере одну из n нуклеиновых кислот-мишеней, в реакционном сосуде;
где n представляет собой целое число 2 или более, где инкубация включает множество циклов, и детекцию сигналов проводят по меньшей мере в одном из множества циклов, где каждая из n композиций для детекции нуклеиновых кислот-мишеней обеспечивает изменение сигнала при соответствующей температуре детекции среди n температур детекции в присутствии соответствующей нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие соответствующей нуклеиновой кислоты-мишени, где композиция для детекции i-й нуклеиновой кислоты-мишени среди n композиций для детекции нуклеиновых кислот-мишеней обеспечивает изменение сигнала при i-й температуре детекции среди n температур детекции и обеспечивает постоянный сигнал при других температурах детекции в присутствии i-й нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие i-й нуклеиновой кислоты-мишени, и
где i представляет собой целое число от 1 до n, и i-я температура детекции является более низкой, чем (i+1)-я температура детекции; и
(b) определение присутствия n нуклеиновых кислот-мишеней из сигналов, детектированных на стадии (a), где присутствие i-й нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при i-й температуре детекции.
2. Способ по п.1, где в температурном диапазоне, покрывающем все из n температур детекции, композиция для детекции i-й нуклеиновой кислоты-мишени имеет температурный диапазон изменения сигнала (SChTR), в котором сигнал изменяется в зависимости от присутствия i-й нуклеиновой кислоты-мишени, и один или два температурных диапазона постоянного сигнала (SCoTR), в которых сигнал является постоянным, даже в присутствии i-й нуклеиновой кислоты-мишени.
3. Способ по п.2, где композиция для детекции i-й нуклеиновой кислоты-мишени представляет собой любую из:
(i) композиции с нижним изменением сигнала (UnderSC), имеющей характеристику, что температурный диапазон изменения сигнала является более низким, чем температурный диапазон постоянного сигнала;
(ii) композиции с верхним изменением сигнала (OverSC), имеющей характеристику, что температурный диапазон изменения сигнала является более высоким, чем температурный диапазон постоянного сигнала; и
(iii) композиции с промежуточным изменением сигнала (InterSC), имеющей характеристику, что температурный диапазон изменения сигнала является более высоким, чем один из двух температурных диапазонов постоянного сигнала, и более низким, чем другой из двух температурных диапазонов постоянного сигнала.
4. Способ по п.2, где i-я температура детекции выбрана в рамках температурного диапазона изменения сигнала композиции для детекции i-й нуклеиновой кислоты-мишени, где i-я температура детекции не включена в температурные диапазоны изменения сигнала композиций для детекции других нуклеиновых кислот-мишеней.
5. Способ по п.2, где температурный диапазон изменения сигнала композиции для детекции i-й нуклеиновой кислоты-мишени частично перекрывается с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей соседнюю температуру детекции, и не перекрывается с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей температуру детекции, не являющуюся соседней с ней.
6. Способ по п.3, где, (i) когда n представляет собой 2, композиция для детекции первой нуклеиновой кислоты-мишени представляет собой композицию UnderSC или композицию InterSC, и композиция для детекции второй нуклеиновой кислоты-мишени представляет собой композицию InterSC или композицию OverSC, или
(ii) когда n представляет собой 3 или более, композиция для детекции первой нуклеиновой кислоты-мишени представляет собой композицию UnderSC или композицию InterSC, композиция для детекции n-й нуклеиновой кислоты-мишени представляет собой композицию InterSC или композицию OverSC, и каждая из композиций для детекции нуклеиновых кислот-мишеней, отличных от первой нуклеиновой кислоты-мишени и n-й нуклеиновой кислоты-мишени, представляет собой композицию InterSC.
7. Способ по п.1, где композиция для детекции i-й нуклеиновой кислоты-мишени содержит метку, которая обеспечивает сигнал, зависимый от присутствия i-й нуклеиновой кислоты-мишени.
8. Способ по п.7, где метка является связанной с олигонуклеотидом или встраивается в олигонуклеотид в ходе инкубации.
9. Способ по п.1, где композиция для детекции i-й нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала; где дуплекс, обеспечивающий изменение сигнала, содержит метку.
10. Способ по п.7, где композиция для детекции i-й нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала, и композиция для детекции i-й нуклеиновой кислоты-мишени обеспечивает сигнал от метки, когда дуплекс, обеспечивающий изменение сигнала, находится в связанной форме или в диссоциированной форме.
11. Способ по п.9, где дуплекс, обеспечивающий изменение сигнала, был изначально включен в композицию для детекции i-й нуклеиновой кислоты-мишени.
12. Способ по п.11, где дуплекс, обеспечивающий изменение сигнала, образуется посредством гибридизации между связанным с меткой олигонуклеотидом и олигонуклеотидом, способным к гибридизации со связанным с меткой олигонуклеотидом.
13. Способ по п.9, где дуплекс, обеспечивающий изменение сигнала, образуется посредством инкубации.
14. Способ по п.13, где дуплекс, обеспечивающий изменение сигнала, образуется посредством (i) гибридизации между связанным с меткой олигонуклеотидом и нуклеиновой кислотой-мишенью или (ii) реакции расщепления, зависимой от присутствия нуклеиновой кислоты-мишени.
15. Способ по п.14, где композиция для детекции нуклеиновой кислоты-мишени содержит метящий олигонуклеотид, который гибридизуется с нуклеиновой кислотой-мишенью, и реакция расщепления, зависимая от присутствия нуклеиновой кислоты-мишени, включает расщепление метящего олигонуклеотида.
16. Способ по п.9, где дуплекс, обеспечивающий изменение сигнала, представляет собой дуплекс одиночного типирования или дуплексы множественного типирования.
17. Способ по п.16, где, (i) когда дуплекс, обеспечивающий изменение сигнала, представляет собой дуплекс одиночного типирования, количество дуплекса одиночного типирования изменяется в зависимости от присутствия нуклеиновой кислоты-мишени, таким образом изменяя сигнал, или (ii) когда дуплекс, обеспечивающий изменение сигнала, представляет собой дуплексы множественного типирования, соотношение количеств между дуплексами множественного типирования изменяется в зависимости от присутствия нуклеиновой кислоты-мишени, таким образом изменяя сигнал.
18. Способ по п.16, где, когда дуплекс представляет собой дуплексы множественного типирования, значения Tm дуплексов отличаются друг от друга.
19. Способ по п.4, где композиция для детекции i-й нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала, и температурный диапазон изменения сигнала композиции для детекции i-й нуклеиновой кислоты-мишени определяют в зависимости от длины и/или последовательности дуплекса.
20. Способ по п.1, где детекцию сигналов проводят по меньшей мере в двух из множества циклов.
21. Способ по п.20, где изменение сигнала измеряют с использованием сигналов, детектированных по меньшей мере в двух из множества циклов.
22. Способ по п.1, где изменение сигнала при i-й температуре детекции измеряют с использованием сигнала, детектированного по меньшей мере в одном из множества циклов, и контрольного значения сигнала.
23. Способ по п.22, где контрольное значение сигнала получают в результате реакции в отсутствие i-й нуклеиновой кислоты-мишени.
24. Способ по п.1, где детекцию сигнала при каждой из n температур детекции проводят с использованием одного типа детектора.
25. Способ по п.24, где сигналы, детектированные при n температурах детекции, не являются отличимыми друг от друга посредством одного типа детектора.
26. Способ по п.1, где инкубация включает реакцию амплификации нуклеиновой кислоты.
27. Способ детекции двух нуклеиновых кислот-мишеней в образце, включающий:
(a) детекцию сигналов при первой температуре детекции и второй температуре детекции, с инкубацией в то же время образца, как подозревают, содержащего по меньшей мере одну из двух нуклеиновых кислот-мишеней, с композицией для детекции первой нуклеиновой кислоты-мишени и композицией для детекции второй нуклеиновой кислоты-мишени в реакционном сосуде;
где инкубация включает множество циклов, и детекцию сигналов проводят по меньшей мере в одном из множества циклов, где композиция для детекции первой нуклеиновой кислоты-мишени обеспечивает изменение сигнала при первой температуре детекции и обеспечивает постоянный сигнал при второй температуре детекции в присутствии первой нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие первой нуклеиновой кислоты-мишени; и композиция для детекции второй нуклеиновой кислоты-мишени обеспечивает изменение сигнала при второй температуре детекции и обеспечивает постоянный сигнал при первой температуре детекции в присутствии второй нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие второй нуклеиновой кислоты-мишени, и где первая температура детекции является более низкой, чем вторая температура детекции, и
(b) определение присутствия двух нуклеиновых кислот-мишеней из сигналов, детектированных на стадии (a); где присутствие первой нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при первой температуре детекции, и присутствие второй нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при второй температуре детекции.
28. Способ детекции трех нуклеиновых кислот-мишеней в образце, включающий:
(a) детекцию сигналов при первой температуре детекции, второй температуре детекции и третьей температуре детекции, с инкубацией в то же время образца, как подозревают, содержащего по меньшей мере одну из трех нуклеиновых кислот-мишеней, с композицией для детекции первой нуклеиновой кислоты-мишени, композицией для детекции второй нуклеиновой кислоты-мишени и композицией для детекции третьей нуклеиновой кислоты-мишени в реакционном сосуде;
где инкубация включает множество циклов, и детекцию сигналов проводят по меньшей мере в одном из множества циклов, где композиция для детекции первой нуклеиновой кислоты-мишени обеспечивает изменение сигнала при первой температуре детекции и обеспечивает постоянный сигнал при второй температуре детекции и третьей температуре детекции в присутствии первой нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие первой нуклеиновой кислоты-мишени; композиция для детекции второй нуклеиновой кислоты-мишени обеспечивает изменение сигнала при второй температуре детекции и обеспечивает постоянный сигнал при первой температуре детекции и третьей температуре детекции в присутствии второй нуклеиновой кислоты- мишени, где изменение сигнала указывает на присутствие второй нуклеиновой кислоты-мишени; и композиция для детекции третьей нуклеиновой кислоты-мишени обеспечивает изменение сигнала при третьей температуре детекции и обеспечивает постоянный сигнал
при первой температуре детекции и второй температуре детекции в присутствии третьей нуклеиновой кислоты-мишени, где изменение сигнала указывает на присутствие третьей нуклеиновой кислоты-мишени, и где первая температура детекции является более низкой, чем вторая температура детекции, и вторая температура детекции является более низкой, чем третья температура детекции, и
(b) определение присутствия трех нуклеиновых кислот-мишеней из сигналов, детектированных на стадии (a), где присутствие первой нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при первой температуре детекции, присутствие второй нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при второй температуре детекции, и присутствие третьей нуклеиновой кислоты-мишени определяют посредством изменения сигнала, детектированного при третьей температуре детекции.
29. Набор для применения в осуществлении способа по любому из пп.1-26 формулы изобретения, содержащий n композиций для детекции n нуклеиновых кислот-мишеней в образце, где n представляет собой целое число 2 или более, где каждая из n композиций для детекции n нуклеиновых кислот-мишеней обеспечивает изменение сигнала при соответствующей температуре детекции среди n температур детекции, где изменение сигнала указывает на присутствие соответствующей нуклеиновой кислоты-мишени, где композиция для детекции i-й нуклеиновой кислоты-мишени среди n нуклеиновых кислот-мишеней обеспечивает изменение сигнала при i-й температуре детекции среди n температур детекции и обеспечивает постоянный сигнал при других температурах детекции в присутствии i-й нуклеиновой кислоты-мишени, и где i представляет собой целое число от 1 до n, и i-я температура детекции является более низкой, чем (i+1)-я температура детекции.
30. Набор по п.29, где в температурном диапазоне, покрывающем все из n температур детекции, композиция для детекции i-й нуклеиновой кислоты-мишени имеет температурный диапазон изменения сигнала (SChTR), в котором сигнал изменяется
в зависимости от присутствия i-й нуклеиновой кислоты-мишени, и один или два температурных диапазона постоянного сигнала (SCoTR), в которых сигнал является постоянным, даже в присутствии i-й нуклеиновой кислоты-мишени.
31. Набор по п.30, где композиция для детекции i-й нуклеиновой кислоты-мишени представляет собой любую из:
(i) композиции с нижним изменением сигнала (UnderSC), имеющей характеристику, что температурный диапазон изменения сигнала является более низким, чем температурный диапазон постоянного сигнала;
(ii) композиции с верхним изменением сигнала (OverSC), имеющей характеристику, что температурный диапазон изменения сигнала является более высоким, чем температурный диапазон постоянного сигнала; и
(iii) композиции с промежуточным изменением сигнала (InterSC), имеющей характеристику, что температурный диапазон изменения сигнала является более высоким, чем один из двух температурных диапазонов постоянного сигнала, и более низким,
чем другой из двух температурных диапазонов постоянного сигнала.
32. Набор по п.30, где i-я температура детекции выбрана в рамках температурного диапазона изменения сигнала композиции для детекции i-й нуклеиновой кислоты-мишени, где i-я температура детекции не включена в температурные диапазоны изменения сигнала композиций для детекции других нуклеиновых кислот-мишеней.
33. Набор по п.30, где температурный диапазон изменения сигнала композиции для детекции i-й нуклеиновой кислоты-мишени частично перекрывается с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей соседнюю температуру детекции, и не перекрывается с температурным диапазоном изменения сигнала композиции для детекции нуклеиновой кислоты-мишени, имеющей температуру детекции, не являющуюся соседней с ней.
34. Набор по п.31, где, (i) когда n представляет собой 2, композиция для детекции первой нуклеиновой кислоты-мишени представляет собой композицию UnderSC или композицию InterSC, и композиция для детекции второй нуклеиновой кислоты-мишени представляет собой композицию InterSC или композицию OverSC, или
(ii) когда n представляет собой 3 или более, композиция для детекции первой нуклеиновой кислоты-мишени представляет собой композицию UnderSC или композицию InterSC, композиция для детекции n-й нуклеиновой кислоты-мишени представляет собой композицию InterSC или композицию OverSC, и каждая из композиций для детекции нуклеиновых кислот-мишеней, отличных от первой нуклеиновой кислоты-мишени и n-й нуклеиновой кислоты-мишени, представляет собой композицию InterSC.
35. Набор по п.29, где композиция для детекции i-й нуклеиновой кислоты-мишени содержит метку, которая обеспечивает сигнал, зависимый от присутствия i-й нуклеиновой кислоты-мишени.
36. Набор по п.35, где метка является связанной с олигонуклеотидом или встраивается в олигонуклеотид в ходе инкубации набора и образца.
37. Набор по п.29, где композиция для детекции i-й нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала.
38. Набор по п.35, где композиция для детекции i-й нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала, и композиция для детекции i-й нуклеиновой кислоты-мишени обеспечивает сигнал от метки, когда дуплекс,
обеспечивающий изменение сигнала, находится в связанной форме или в диссоциированной форме.
39. Набор по п.29, где композиция для детекции i-й нуклеиновой кислоты-мишени предоставляет дуплекс, обеспечивающий изменение сигнала, и температурный диапазон изменения сигнала композиции для детекции i-й нуклеиновой кислоты-мишени определяют в зависимости от длины и/или последовательности дуплекса.
40. Набор по п.35, где сигналы, обеспечиваемые посредством n композиций для детекции n нуклеиновых кислот-мишеней, не являются отличимыми друг от друга посредством одного типа детектора.
| US 20170247750 A1, 31.08.2017 | |||
| US 20210040542 A1, 11.02.2021 | |||
| ВИРТУАЛЬНЫЕ НАБОРЫ ФРАГМЕНТОВ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ | 2004 |
|
RU2390561C2 |
| ДЕТЕКЦИЯ НУКЛЕИНОВЫХ КИСЛОТ СПОСОБОМ, ОСНОВАННЫМ НА СВЯЗЫВАНИИ МИШЕНЕСПЕЦИФИЧНОГО ГИБРИДА | 2006 |
|
RU2437939C2 |
