Перекрестная ссылка на родственные заявки
Данная заявка испрашивает приоритет на основании предварительной заявки на патент США № 60/311282, поданной 9 августа 2001 г.
Область техники
Настоящее изобретение относится в целом к растениям с повышенной устойчивостью к имидазолиноновым гербицидам. В частности настоящее изобретение относится к растениям пшеницы, полученным при помощи мутагенеза, кроссбридинга и трансформации, которые обладают повышенной устойчивостью к имидазолиноновым гербицидам.
Предпосылки изобретения
Синтаза ацетогидроксикислоты (AHAS; EC 4.1.3.18) является первым ферментом, катализирующим биохимический синтез валина, лейцина и изолейцина в аминокислотах с разветвленной цепью (Singh B.K., 1999, Biosynthesis of valine, leucine and isoleucine in: Singh B.K. (Ed) Plant amino acids. Marcel Dekker Inc. New York, New York. Pg 227-247). На AHAS воздействуют четыре разных в структурном отношении семейства гербицидов, включающих сульфонилмочевины (LaRossa RA and Falco SC, 1984 Trends Biotechnol 2:158-161), имидазолиноны (Shaner et al., 1984 Plant Physiol 76:545-546), триазолопиримидины (Subramanian and Gerwick, 1989 Inhibition of acetolactate synthase by triazolopyrimidines in (ed) Whitaker JR, Sonnet PE Biocatalysis in agricultural biotechnology. ACS Symposium Series, American Chemical Society, Washington, D.C. Pg 277-288) и пиримидилоксибензоаты (Subramanian et al., 1990 Plant Physiol 94: 239-244). Гербициды на основе имидазолинона и сульфонилмочевины широко применяются в современном сельском хозяйстве благодаря их эффективности при очень низких нормах внесения и относительной нетоксичности для животных. Подавляя активность AHAS, указанные семейства гербицидов предотвращают дальнейший рост и развитие восприимчивых к ним растений, включая многие виды сорняков. Несколькими примерами промышленно производимых имидазолиноновых гербицидов являются PURSUIT® (имазетапир), SCEPTER® (имазахин) и ARSENAL® (имазапир). Примеры гербицидов на основе сульфонилмочевины включают хлорсульфурон, метсульфуронметил, сульфометуронметил, хлоримуронэтил, тифенсульфуронметил, трибенуронметил, бенсульфуронметил, никосульфурон, этаметсульфуронметил, римсульфурон, трифлусульфуронметил, триасульфурон, примисульфуронметил, циносульфурон, амидосульфурон, флузасульфурон, имазосульфурон, пиразосульфуронэтил и галогенсульфурон.
Благодаря высокой эффективности и низкой токсичности имидазолиноновые гербициды применяют для опрыскивания растительности на больших площадях. Возможность опрыскивания гербицидом растительности на больших площадях сокращает затраты, связанные с возделыванием и содержанием сельскохозяйственных культур, и уменьшает потребность в подготовке участка к использованию таких химикатов. Кроме того, опрыскивание желательных устойчивых видов растений позволяет максимально увеличить урожайность требуемых культур благодаря отсутствию конкурирующих видов. Однако возможность применения таких методов опрыскивания зависит от наличия устойчивых к имидазолинону видов растительности на опрыскиваемом участке.
Среди главных сельскохозяйственных культур некоторые бобовые культуры, такие как соя, обладают природной устойчивостью к имидазолиноновым гербицидам благодаря их способности быстро метаболизировать гербицидные соединения (Shaner and Robinson, 1985 Weed Sci. 33:469-471). Другие сельскохозяйственные культуры, такие как кукуруза (Newhouse et al., 1992, Plant Physiol. 100:882-886) и рис (Barrette et al., 1989 Crop Safeners for Herbicides, Academic Press Nes York, pp. 195-220), в некоторой степени восприимчивы к воздействию имидазолиноновых гербицидов. Различная восприимчивость к имидазолиноновым гербицидам зависит от химической природы конкретного гербицида и разного превращения соединения из токсичной в нетоксичную форму, происходящего в каждом растении в процессе обмена веществ (Shaner et al., 1984 Plant Physiol. 76:545-546; Brown et al., 1987 Pestic. Biochim. Physiol. 27:24-29). Другие физиологические различия растений, такие как поглощение и передвижение веществ, также имеют важное значение для восприимчивости к определенным веществам (Shaner and Robinson, 1985 Weed Sci. 33:469-471).
Сорта сельскохозяйственных культур, устойчивые к имидазолинонам, сульфонилмочевинам и триазолопиримидинам, успешно получены при помощи мутагенеза семян, микроспор, пыльцы и каллюсов в таких видах, как Zea mays, Arabidopsis thaliana, Brassica napus, Glycine max и Nicotiana tabacum (Sebastian et al., 1989 Crop Sci. 29:1403-1408; Swanson et al., 1989 Theor. Appl. Genet. 78:525-530; Newhouse et al., 1991 Theor. Appl. Genet. 83:65-70; Sathasivan et al., 1991 Plant Physiol. 97: 1044-1050; Mourand et al., 1993 J. Heredity 84:91-96). Во всех случаях устойчивость вызывает один частично доминантный ядерный ген. В результате мутагенеза семян Triticum aestivum L. сорта Fidel (Newhouse et al., 1992 Plant Physiol. 100:882-886) ранее были выделены четыре вида растений пшеницы, устойчивых к имидазолинону. Исследования наследственности подтвердили, что устойчивость придает один, частично доминантный ген. В результате исследования аллелей авторы пришли к выводу, что мутации в четырех идентифицированных линиях расположены в одном и том же локусе. Один из генов устойчивости в сорте Fidel получил название FS-4 (Newhouse et al., 1992 Plant Physiol. 100:882-886).
Компьютерное моделирование трехмерной структуры комплекса AHAS-ингибитор позволяет прогнозировать наличие нескольких аминокислот в предполагаемом "кармане" связывания ингибитора в качестве сайтов, где индуцированные мутации, по-видимому, вызывают избирательную устойчивость к имидазолинонам (Ott et al., 1996 J. Mol. Biol. 263:359-368). Растения пшеницы, полученные в результате некоторых из указанных рационально программируемых мутаций на предполагаемых сайтах связывания фермента AHAS, действительно обладают специфической устойчивостью к одному классу гербицидов (Ott et al., 1996 J. Mol. Biol. 263:359-368).
Устойчивость растений к имидазолиноновым гербицидам описана также в ряде патентов. В патентах США №№ 4761373, 5331107, 5304732, 6211438, 6211439 и 6222100 дано общее описание применения измененного гена AHAS для создания растений, устойчивых к гербицидам, и, в частности, рассмотрены некоторые сорта зерновых культур, устойчивые к имидазолинону. В патенте США № 5013659 описаны растения с устойчивостью к гербицидам, которые имеют мутации, по меньшей мере, одной аминокислоты в одной или более консервативных областях. Мутации, описанные в вышеуказанном патенте, кодируют перекрестную устойчивость к имидазолинонам и сульфонилмочевинам или специфическую устойчивость к сульфонилмочевине, но в данном патенте не рассмотрена специфическая устойчивость к имидазолинону. Кроме того, в патентах США №№ 5731180 и 5767361 описан выделенный ген с заменой одной аминокислоты в аминокислотной последовательности AHAS однодольных растений дикого типа, который определяет специфическую устойчивость к имидазолинону.
До настоящего времени не были известны устойчивые к имидазолинону растения пшеницы, имеющие более одного измененного гена AHAS. В научной литературе ранее также не были описаны устойчивые к имидазолинону растения пшеницы, имеющие мутации в геномах, не являющихся геномом, из которого выделен ген FS-4. Поэтому необходимо идентифицировать гены устойчивости к имидазолинону из дополнительных геномов. Кроме того, необходимо получить растения пшеницы, обладающие повышенной устойчивостью к гербицидам, таким как имидазолинон, и содержащие более одного измененного гена AHAS. Существует потребность также в способах борьбы с сорняками, произрастающими рядом с такими растениями пшеницы. Такие композиции и способы позволяют применять методы опрыскивания при внесении гербицидов на участки, засеянные пшеницей.
Краткое ОПИСАНИЕ изобретения
Настоящее изобретение относится к растениям пшеницы, содержащим нуклеиновые кислоты IMI, которые обладают повышенной устойчивостью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа. Растения пшеницы могут содержать одну, две, три или большее число нуклеиновых кислот IMI. В одном варианте осуществления изобретения растение пшеницы включает множество нуклеиновых кислот IMI, расположенных в разных геномах. Нуклеиновые кислоты IMI предпочтительно кодируют белки, включающие мутацию в консервативной аминокислотной последовательности, выбираемой из группы, состоящей из домена А, домена В, домена С, домена D и домена Е. Более предпочтительно мутация находится в консервативном домене Е или консервативном домене С. Кроме того, настоящее изобретение относится к частям растения и семенам растения, полученным из описанных растений пшеницы. В другом варианте осуществления изобретения растение пшеницы включает нуклеиновую кислоту IMI, которая не является нуклеиновой кислотой Imi1. Например, нуклеиновая кислота IMI может быть нуклеиновой кислотой Imi2 или Imi3.
Нуклеиновые кислоты IMI по настоящему изобретению могут включать нуклеотидную последовательность, выбираемую из группы, состоящей из полинуклеотида SEQ ID NO: 1; полинуклеотида SEQ ID NO: 3; полинуклеотидной последовательности, кодирующей полипептид SEQ ID NO: 2; полинуклеотидной последовательности, кодирующей полипептид SEQ ID NO: 4; полинуклеотида, содержащего, по меньшей мере, 60 последовательно расположенных нуклеотидов любого из вышеуказанных полинуклеотидов; и полинуклеотида, комплементарного любым вышеуказанным полинуклеотидам.
Растения по настоящему изобретению могут быть трансгенными или нетрансгенными. Примеры нетрансгенных растений пшеницы с повышенной устойчивостью к имидазолиноновым гербицидам включают растение пшеницы, представленное под идентификационным номером патентного депозитария АТСС РТА-3953 или РТА-3955; или мутантное, рекомбинантное или генетически сконструированное производное растения, представленного под идентификационным номером патентного депозитария АТСС РТА-3953 или РТА-3955; или любое потомство растения, представленного под идентификационным номером патентного депозитария АТСС РТА-3953 или РТА-3955; или растение, являющееся потомством любого из указанных растений.
Помимо композиций в объем настоящего изобретения входят несколько способов. В данной заявке на патент описаны способы модификации устойчивости растения к имидазолиноновому гербициду, включающие модификацию экспрессии нуклеиновой кислоты IMI в растении. Кроме того, в данной заявке описаны способы получения трансгенного растения с повышенной устойчивостью к имидазолиноновому гербициду, включающие трансформацию растительной клетки экспрессирующим вектором, содержащим одну или более нуклеиновых кислот IMI, и выращивание растения из растительной клетки. Данное изобретение далее относится к способу борьбы с сорняками, произрастающими рядом с растениями пшеницы, который включает нанесение имидазолинонового гербицида на сорняки и растение пшеницы, при этом растение пшеницы обладает повышенной устойчивостью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа пшеницы, при этом растение содержит одну или более нуклеиновых кислот IMI. В некоторых предпочтительных вариантах осуществления указанных способов растения содержат множество нуклеиновых кислот IMI, находящихся в разных геномах пшеницы.
Краткое описание чертежей
На фиг.1 представлена таблица, в которой приведены результаты оценки устойчивости к имазамоксу для одного вида растений в исходной популяции и популяции F1, полученной в результате реципрокного скрещивания устойчивых линий с CDC Teal. Указанные величины означают число растений, подвергнутых оценке в каждом фенотипическом классе. Родительские линии выделены жирным шрифтом. Число исследованных родительских линий включает линии, исследованные в популяциях F2.
На фиг.2 представлена таблица, в которой показана реакция на имазамокс в популяциях F2 и BCF1, полученных в результате скрещивания устойчивых линий с CDC Teal, и результаты проверок на соответствие по критерию хи-квадрат однолокусной и двухлокусной моделей (15А × Teal) для оценки устойчивости. Символы, использованные на фиг.2, имеют следующие значения: а - значение Р критерия хи-квадрат (1 степень свободы) означает вероятность того, что отклонения от соотношения испытанных растений являются случайными. Значения Р критерия хи-квадрат выше 0,05 указывают на то, что полученные значения не отличаются на значимом уровне от ожидаемых результатов; b - значение Р критерия хи-квадрат означает вероятность того, что отклонения в популяциях F2, полученных в результате реципрокного скрещивания CDC Teal с устойчивыми линиями, являются случайными. Значения критерия хи-квадрат выше 0,05 указывают на то, что реципрокные популяции F2 являются гомогенными, и данные для двух реципрокных популяций объединены; с - CDC Teal использован в качестве родительской формы, с которой гибрид скрещивается вновь; d - соотношения испытанных растений основаны на результатах поколения F2; и е - значение Р критерия хи-квадрат (1 степень свободы) для соотношения BCF1.
На фиг.3 представлена таблица, в которой приведены результаты оценки устойчивости к имазамоксу в семействах F2:3, полученных в результате скрещивания устойчивых линий с CDC Teal, и результаты проверок на соответствие по критерию хи-квадрат однолокусной и двухлокусной моделей (15А × Teal) для оценки устойчивости. Символы, использованные на фиг.3, имеют следующие значения: а - исследованные соотношения расщепления семейства основаны на результатах популяций F2 и BCF1; b - значение Р критерия хи-квадрат (2 степени свободы) представляет вероятность того, что отклонения от соотношения испытанных растений являются случайными. Значения Р критерия хи-квадрат выше 0,05 указывают на то, что полученные значения не отличаются на значимом уровне от ожидаемых результатов.
На фиг.4 представлена таблица, в которой приведены результаты оценки устойчивости к имазамоксу для одного растения в поколениях F2, полученных в результате интеркросса устойчивых линий. Соотношения испытанных растений, вычисленные по критерию хи-квадрат, основаны на результатах для семейств F2 и F2:3, полученных путем скрещивания устойчивых линий с CDC Teal. Соотношение испытанных растений, равное 15:1, относится к двухлокусной модели, и соотношение испытанных растений, равное 63:1, относится к трехлокусной модели. Символ "a", использованный на фиг.4, имеет следующее значение: значение Р критерия хи-квадрат (1 степень свободы) представляет вероятность того, что отклонения от соотношения испытанных растений являются случайными. Значения Р критерия хи-квадрат выше 0,05 указывают на то, что полученные значения не отличаются на значимом уровне от ожидаемых результатов.
На фиг.5 представлена таблица, в которой приведены результаты оценки устойчивости к имазамоксу в семействах F2:3, полученных в результате расщепления интеркроссов устойчивых линий. Символы, использованные на фиг.5, имеют следующие значения: а - соотношения испытанных растений в процессе расщепления семейства основаны на результатах исследованных популяций F2; b - значение Р критерия хи-квадрат (2 степени свободы) представляет вероятность того, что отклонения от соотношения испытанных растений являются случайными. Значения Р критерия хи-квадрат выше 0,05 указывают на то, что полученные значения не отличаются на значимом уровне от ожидаемых результатов.
На фиг.6 представлена таблица, в которой сравнивают процентные значения неингибированной активности AHAS in vitro в четырех линиях пшеницы в присутствии возрастающих концентраций имазамокса, являющегося имидазолиноновым гербицидом. Teal представляет линию дикого типа, не имеющую устойчивости к имидазолиноновым гербицидам, в то время как линия BW755 содержит мутантный ген FS4.
На фиг.7 представлена таблица, в которой сравнивают результаты повреждения, выявленные у трех генотипов пшеницы после обработки 10-кратной или 30-кратной нормой имазамокса. Однократная норма составляет 20 г/га. BW755 содержит мутантный ген FS4. 15А/11А представляет основную массу самоопыленного потомства, полученного в результате скрещивания Teal11A и Teal15A. Данная популяция не является еще гомозиготной во всех трех неаллельных локусах.
На фиг.8 показано сопоставление последовательностей ДНК частичных генов пшеницы Als1 и Imi1, амплифицированных из геномной ДНК: CDC Teal (ряд 2; SEQ ID NO: 15 и SEQ ID NO: 16), BW755 (ряд 3; SEQ ID NO: 17 и SEQ ID NO: 18), TealIMI 10A (ряд 4; SEQ ID NO: 19 и SEQ ID NO: 20), TealIMI 11A (ряд 5; SEQ ID NO: 21 и SEQ ID NO: 22) и TealIMI 15A (ряд 6; SEQ ID NO: 23 и SEQ ID NO: 24). Частичные последовательности сопоставляют с полной последовательностью гена ALS риса (ряд 1; SEQ ID NO: 13 и SEQ ID NO: 14), полученной из банка генов GenBank (№ доступа АВО49822) и транслированной в белковые последовательности (представленные над последовательностями ДНК). Пять высококонсервативных аминокислотных доменов, которые, как известно, включают мутации, придающие устойчивость к ингибиторам AHAS, указаны жирным шрифтом. Следует отметить, что замена гуанина аденином в BW755, TealIMI 10A и TealIMI 15A вызывает замену серина аспарагином (серин627 в рисе) в домене IPSGG (домен Е) гена Als1. Поэтому гены устойчивости, присутствующие в растениях BW755, TealIMI 10A и TealIMI 15A, представлены в виде части класса Imi1. Указанные гены устойчивости Teal определяются как TealIMI1 10А и TealIMI1 15A.
На фиг.9 показано сопоставление последовательностей ДНК частичных генов пшеницы Als2 и Imi2, амплифицированных из геномной ДНК: CDC Teal (ряд 2; SEQ ID NO: 25 и SEQ ID NO: 26), BW755 (ряд 3; SEQ ID NO: 27 и SEQ ID NO: 28), TealIMI 10A (ряд 4; SEQ ID NO: 29 и SEQ ID NO: 30), TealIMI 11A (ряд 5; SEQ ID NO: 31 и SEQ ID NO: 32) и TealIMI 15A (ряд 6; SEQ ID NO: 33 и SEQ ID NO: 34). Частичные последовательности AHAS сопоставляют с полной последовательностью AHAS риса (ряд 1; SEQ ID NO: 13 и SEQ ID NO: 14), полученной из банка генов GenBank (№ доступа АВ049822) и транслированной в белковые последовательности (представленные над последовательностями ДНК). Пять высококонсервативных доменов, которые, как известно, включают мутации, придающие устойчивость к ингибиторам AHAS, указаны жирным шрифтом. Следует отметить, что замена гуанина аденином в TealIMI 11A вызывает замену серина аспарагином (серин627 в рисе) в домене IPSGG гена Als2. Поэтому ген устойчивости, присутствующий в растении TealIMI 11A, представлен в виде части класса нуклеиновых кислот Imi2. Указанный ген устойчивости Teal определяется как TealIMI2 11A.
На фиг.10 показана частичная последовательность ДНК TealIMI1 15A (SEQ ID NO: 1) и выведенная аминокислотная последовательность указанной последовательности (SEQ ID NO: 2).
На фиг.11 показана частичная последовательность ДНК TealIMI2 11А (SEQ ID NO: 3) и выведенная аминокислотная последовательность указанной последовательности (SEQ ID NO: 4).
На фиг.12 показана последовательность нуклеиновых кислот дикого типа Teal ALS1 ORF (SEQ ID NO: 5), Teal ALS2 ORF (SEQ ID NO: 6), Teal ALS3 ORF (SEQ ID NO: 7).
На фиг.13 показано схематическое изображение консервативных аминокислотных последовательностей в генах AHAS, определяющих устойчивость к разным ингибиторам AHAS. Специфический аминокислотный сайт, ответственный за устойчивость, подчеркнут. (Modified from Devine, M.D. and Eberlein, C.V., 1997 Physiological, biochemical and molecular aspects of herbicide resistance based on altered target sites in Herbicide Activity: Toxicity, Biochemistry, and Molecular Biology, IOS Press Amsterdam, p. 159-185).
Подробное описание изобретения
Настоящее изобретение относится к растениям пшеницы, частям и клеткам растений пшеницы, обладающим повышенной устойчивостью к имидазолиноновым гербицидам. В объем настоящего изобретения входят также семена, продуцируемые растениями пшеницы, и способы борьбы с сорняками, произрастающими рядом с описанными растениями пшеницы. Следует понимать, что в данном описании изобретения и формуле изобретения используемый в единственном числе термин в зависимости от контекста, так, например, ссылка на клетку, означает, что может использоваться, по меньшей мере, одна клетка.
В используемом здесь значении термин "растение пшеницы" означает растение, относящееся к роду Triticum. Растения пшеницы по настоящему изобретению могут быть членами рода Triticum, включающего, не ограничиваясь ими, T. aestivum, T. turgidum, T. timopheevii, T. monococcum, T. zhukovskyi, T. urartu и их гибриды. Примерами подвидов T. aestivum, входящих в объем настоящего изобретения, являются aestivum (пшеница обыкновенная), compactum (пшеница плотная), macha (пшеница мача), vavilovi (пшеница вавилонская) spelta и sphaecrococcum (пшеница карликовая). Примерами подвидов T. turgidum, входящих в объем настоящего изобретения, являются turgidum, carthlicum, dicoccon, durum, paleocolchicum, polonicum, turanicum и dicoccoides. Примерами подвидов T. monococcum, входящих в объем настоящего изобретения, являются monococcum (пшеница однозернянка) и aegilopoides. В одном варианте осуществления настоящего изобретения растение пшеницы относится к виду Triticum aestivum и, в частности, представляет культурный сорт CDC Teal.
Термин "растение пшеницы" охватывает растения пшеницы на любой стадии зрелости или развития, а также любые ткани или органы (части растения), полученные из любого такого растения, за исключением особо оговоренных случаев. Части растения включают, не ограничиваясь ими, стебли, корни, цветки, семяпочки, тычинки, листья, завязи, меристематические области, каллюсную ткань, пыльники, гаметофиты, спорофиты, пыльцу, микроспоры, протопласты и тому подобные. В объем настоящего изобретения входят также семена, полученные из растений пшеницы по настоящему изобретению. В одном варианте осуществления изобретения семена используют для истинной селекции для получения повышенной устойчивости к имидазолиноновому гербициду по сравнению с семенами сорта дикого типа пшеницы.
В настоящем изобретении описано растение пшеницы, содержащее одну или более нуклеиновых кислот IMI, которое обладает повышенной устойчивостью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа. В используемом здесь значении термин "нуклеиновая кислота IMI" означает нуклеиновую кислоту, мутированную из нуклеиновой кислоты AHAS в растении пшеницы дикого типа, которая придает повышенную устойчивость к имидазолинону растению, в котором она транскрибирована. Нуклеиновые кислоты AHAS пшеницы Teal дикого типа показаны в SEQ ID NO: 5 (Teal ALS1 ORF), SEQ ID NO: 6 (Teal ALS2 ORF) и SEQ ID NO: 7 (Teal ALS3 ORF). В одном варианте осуществления изобретения растение пшеницы включает множество нуклеиновых кислот IMI. При использовании для описания нуклеиновых кислот IMI термин "множество" относится к нуклеиновым кислотам IMI, имеющим разные нуклеотидные последовательности, а не к простому увеличению числа одной нуклеиновой кислоты IMI. Например, нуклеиновые кислоты IMI могут отличаться вследствие того, что они получены из разных геномов или находятся в разных геномах пшеницы.
Растения пшеницы по настоящему изобретению могут иметь множество нуклеиновых кислот IMI из разных геномов, так как указанные растения могут содержать более одного генома. Например, растение пшеницы Triticum aestivum имеет три генома, которые иногда определяют как геномы А, В и D. Так как AHAS является необходимым метаболическим ферментом, можно предположить, что каждый геном имеет, по меньшей мере, один ген, кодирующий фермент AHAS, обычно обнаруживаемый вместе с другими метаболическими ферментами в картированной гексаплоидной пшенице. Нуклеиновая кислота AHAS в каждом геноме может и обычно отличается своей нуклеотидной последовательностью от нуклеиновой кислоты AHAS в другом геноме. Специалист в данной области может определить геном происхождения каждой нуклеиновой кислоты AHAS методами генетического скрещивания и/или секвенирования или расщепления экзонуклеазой, известными в данной области и описанными в нижеследующем примере 2. В соответствии с целями данного изобретения нуклеиновые кислоты IMI, полученные из геномов А, В или D, получили названия нуклеиновых кислот Imi1, Imi2 или Imi3.
В данном описании изобретения не утверждается, что какой-либо определенный класс нуклеиновых кислот Imi связан с каким-либо определенным геномом А, В или D. Например, здесь не утверждается, что нуклеиновые кислоты Imi1 связаны с нуклеиновыми кислотами генома А, что нуклеиновые кислоты Imi2 связаны с нуклеиновыми кислотами генома В и т.д. Названия Imi1, Imi2 и Imi3 просто показывают, что нуклеиновые кислоты IMI в каждом таком классе не расщепляются независимо, в то время как две нуклеиновые кислоты IMI из разных классов расщепляются независимо и поэтому могут быть получены из разных геномов пшеницы. Класс нуклеиновых кислот Imi1 включает ген FS-4, описанный Newhouse et al. (1992 Plant Physiol. 100:882-886), и ген TealIMI1 15A, который более подробно рассмотрен ниже. Класс нуклеиновых кислот Imi2 включает рассматриваемый ниже ген TealIMI2 11A. Каждый класс Imi может включать члены из разных видов пшеницы. Поэтому каждый класс Imi включает нуклеиновые кислоты IMI, которые отличаются своей нуклеотидной последовательностью, но, тем не менее, происходят или находятся в одном и том же геноме пшеницы, о чем свидетельствуют результаты исследований наследственности, приведенные в нижеследующих примерах и известные специалистам в данной области.
Таким образом, в объем настоящего изобретения входит растение пшеницы, включающее одну или более нуклеиновых кислот IMI, которое обладает повышенной устойчивостью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа и в котором одна или более нуклеиновых кислот IMI выбраны из группы, состоящей из нуклеиновой кислоты Imi1, Imi2 и Imi3. В одном варианте осуществления изобретения растение включает нуклеиновые кислоты Imi1 и Imi3. В предпочтительном варианте осуществления изобретения нуклеиновая кислота Imi1 включает полинуклеотидную последовательность, представленную SEQ ID NO: 1. В другом варианте осуществления изобретения указанное растение включает нуклеиновую кислоту Imi2. В предпочтительном варианте осуществления изобретения нуклеиновая кислота Imi2 включает полинуклеотидную последовательность, представленную SEQ ID NO: 3.
При использовании применительно к нуклеиновым кислотам термин "полученная из" относится к нуклеиновой кислоте, "расположенной в" определенном геноме или "полученной из" определенного генома. Термин "расположенный в" относится к нуклеиновой кислоте, находящейся в пределах данного определенного генома. При использовании применительно к геному термин "полученный из" относится к нуклеиновой кислоте, которая извлечена или выделена из генома. Термин "выделенный" более подробно рассматривается ниже.
В другом варианте осуществления изобретения растение пшеницы включает нуклеиновую кислоту IMI, которая не является нуклеиновой кислотой Imi1. Термин "не является Imi1" означает нуклеиновую кислоту IMI, которая не относится к вышеописанному классу нуклеиновых кислот Imi1. Примеры нуклеиновых кислот класса Imi1 приведены в рядах 3, 4 и 5 на фиг.8. Один пример нуклеиновой кислоты, не являющейся Imi1, показан в ряду 5 на фиг.8. Поэтому в предпочтительном варианте осуществления изобретения растение пшеницы включает нуклеиновую кислоту IMI, включающую полинуклеотидную последовательность, кодирующую полипептид SEQ ID NO: 4. Полинуклеотидная последовательность может включать последовательность, представленную SEQ ID NO: 3.
Настоящее изобретение включает растения пшеницы, содержащие одну, две, три или большее число нуклеиновых кислот IMI, которые обладают повышенной устойчивостью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа. Нуклеиновые кислоты IMI могут включать нуклеотидную последовательность, выбираемую из группы, состоящей из полинуклеотида SEQ ID NO: 1, полинуклеотида SEQ ID NO: 3, полинуклеотидной последовательности, кодирующей полипептид SEQ ID NO: 2, полинуклеотидной последовательности, кодирующей полипептид SEQ ID NO: 4, полинуклеотида, содержащего, по меньшей мере, 60 последовательно расположенных нуклеотидов из любых вышеуказанных полинуклеотидов, и полинуклеотида, комплементарного любым из вышеуказанных полинуклеотидов.
Имидазолиноновые гербициды включают, не ограничиваясь ими, PURSUIT® (имазетапир), CADRE® (имазапик), RAPTOR® (имазамокс), SCEPTER® (имазахин), ASSERT® (имазетабенз), ARSENAL® (имазапир), производное любого вышеуказанного гербицида или смесь двух или большего числа вышеуказанных гербицидов, например имазапир/имазамокс (ODYSSEY®). Более конкретно имидазолиноновые гербициды включают, не ограничиваясь ими, 2-(4-изопропил)-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил)-4-метил-5-оксо-2-имидазолин-2-ил)-3-хинолинкарбоновую кислоту, 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-метилникотиновую кислоту и смесь метил 6-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-м-толуата и метил 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-п-толуата. 5-Этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновая кислота и 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновая кислота являются предпочтительными для использования. 2-(4-Изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновая кислота является особенно предпочтительной для использования.
В одном варианте осуществления изобретения растение пшеницы включает две нуклеиновые кислоты IMI, полученные из или находящиеся в разных геномах пшеницы. Одна из двух нуклеиновых кислот предпочтительно является нуклеиновой кислотой Imi1 и более предпочтительно включает полинуклеотидную последовательность SEQ ID NO: 1. В другом варианте осуществления изобретения растение пшеницы включает одну нуклеиновую кислоту IMI, которая включает одну полинуклеотидную последовательность SEQ ID NO: 1 или SEQ ID NO: 3. В другом варианте осуществления изобретения растение пшеницы включает три или более нуклеиновых кислот IMI, которые получены из разных геномов. По меньшей мере, одна из трех нуклеиновых кислот IMI предпочтительно включает полинуклеотидную последовательность, выбираемую из группы, состоящей из SEQ ID NO: 1 и SEQ ID NO: 3.
В предпочтительном варианте осуществления настоящего изобретения одна или более нуклеиновых кислот IMI, содержащихся в растении, кодируют аминокислотную последовательность, включающую мутацию в домене, который является консервативным для нескольких белков AHAS. Указанные консервативные домены определяются как домен А, домен В, домен С, домен D и домен Е. На фиг.13 показано общее расположение каждого домена в белке AHAS. В используемом здесь значении домен А содержит аминокислотную последовательность AITGQVPRRMIGT (SEQ ID NO: 8); домен В содержит аминокислотную последовательность QWED (SEQ ID NO: 9); домен С содержит аминокислотную последовательность VFAYPGGASMEIHQALTRS (SEQ ID NO: 10); домен D содержит аминокислотную последовательность AFQETP (SEQ ID NO: 11); домен Е содержит аминокислотную последовательность IPSGG (SEQ ID NO: 12). Настоящее изобретение также предполагает, что консервативные домены могут иметь незначительные вариации, например в растениях куколя посевного остаток серина в домене Е заменен остатком аланина.
Таким образом, настоящее изобретение включает растения пшеницы, включающие нуклеиновую кислоту IMI, кодирующую аминокислотную последовательность с мутацией в консервативном домене, выбираемом из группы, состоящей из домена А, домена В, домена С, домена D и домена Е. В одном варианте осуществления изобретения растение пшеницы включает нуклеиновую кислоту IMI, кодирующую аминокислотную последовательность с мутацией в домене Е. В других предпочтительных вариантах осуществления изобретения мутации в консервативных доменах локализованы на подчеркнутых участках: AITGQVPRRMIGT (SEQ ID NO: 8); QWED (SEQ ID NO: 9); VFAYPGGASMEIHQALTRS (SEQ ID NO: 10); AFQETP (SEQ ID NO: 11) и IPSGG (SEQ ID NO: 12). Одной предпочтительной заменой является замена серина аспарагином в домене Е (SEQ ID NO: 12).
Рассматриваемые растения пшеницы могут быть трансгенными или нетрансгенными растениями пшеницы. В используемом здесь значении термин "трансгенный" относится к любому растению, растительной клетке, каллюсу, растительной ткани или части растения, которые содержат, по меньшей мере, один полный или частичный рекомбинантный полинуклеотид. Во многих случаях осуществляют стабильную интеграцию полного или частичного рекомбинантного полинуклеотида в хромосому или в стабильный внехромосомный элемент, так чтобы он передавался последующим поколениям. В соответствии с целями данного изобретения термин "рекомбинантный полинуклеотид" означает полинуклеотид, который изменен, перестроен или модифицирован методами генной инженерии. Примеры указанного полинуклеотида включают любой клонированный полинуклеотид или полинуклеотиды, связанные или присоединенные к гетерологичным последовательностям. Термин "рекомбинантный" не относится к изменениям полинуклеотидов, возникающим в результате природных явлений, таких как спонтанные мутации, или неспонтанного мутагенеза последующим селекционным размножением. Растения, имеющие мутации, возникающие вследствие неспонтанного мутагенеза и селекционного размножения, определяются в данном описании изобретения как нетрансгенные растения и входят в объем настоящего изобретения. В вариантах осуществления изобретения, где растение пшеницы является трансгенным и включает множество нуклеиновых кислот IMI, нуклеиновые кислоты могут быть получены из разных геномов или из одного генома. Альтернативно в вариантах осуществления изобретения, где растение пшеницы является нетрансгенным и включает множество нуклеиновых кислот IMI, нуклеиновые кислоты находятся в разных геномах.
Примером нетрансгенного растения пшеницы, включающего одну нуклеиновую кислоту IMI, является сорт, депонированный в АТСС под идентификационным номером патентного депозитария РТА-3953 и определяемый здесь как сорт пшеницы TealIMI 11A. Сорт пшеницы TealIMI 11A содержит нуклеиновую кислоту Imi2. Частичная нуклеотидная и выведенная аминокислотная последовательности, соответствующие гену TealIMI2 11A, представлены соответственно в SEQ ID NO: 3 и SEQ ID NO: 4. Единственную часть последовательностей, не входящих в SEQ ID NO: 3 и SEQ ID NO: 4, представляют последовательности, кодирующие и соответствующие сигнальной последовательности, отщепляемой от зрелого белка TealIMI2 11A.
Соответственно в SEQ ID NO: 4 представлена полноразмерная выведенная последовательность зрелого белка TealIMI2 11A.
Примером сорта растения пшеницы, включающего две нуклеиновые кислоты IMI в разных геномах, является сорт, депонированный в АТСС под идентификационным номером патентного депозитария РТА-3955 и определяемый здесь как сорт пшеницы TealIMI 15A. Сорт пшеницы TealIMI 15A содержит нуклеиновые кислоты Imi1 и Imi3. Нуклеиновая кислота Imi1 включает мутацию, которая вызывает замену серина аспарагином в белке IMI, кодируемом указанной нуклеиновой кислотой. Мутированные гены AHAS обозначены здесь как TealIMI1 15A и TealIMI3 15A. Частичные нуклеотидные и выведенные аминокислотные последовательности, соответствующие гену TealIMI1 15A, представлены соответственно в SEQ ID NO: 1 и SEQ ID NO: 2. Единственную часть последовательностей, не входящих в SEQ ID NO: 1 и SEQ ID NO: 2, представляют последовательности, кодирующие и соответствующие примерно 100-150 парам оснований у 5'-конца и примерно 5 парам оснований у 3'-конца кодирующей области.
В Американскую коллекцию типовых культур, Manassas, Virginia, 3 января 2002 г. было отдельно депонировано 2500 семян сортов пшеницы TealIMI 11A и TealIMI 15A. Указанные депозиты созданы в соответствии с нормами и правилами Будапештского договора о депонировании микроорганизмов. Депонирование было осуществлено на срок не менее тридцати лет и, по меньшей мере, на пять лет после последнего запроса к АТСС о предоставлении образца из хранилища. Депонированные семена представлены под идентификационными номерами патентных депозитариев РТА-3953 (TealIMI 11A) и РТА-3955 (TealIMI 15A).
В объем настоящего изобретения входит растение пшеницы, представленное под идентификационным номером патентного депозитария РТА-3953 или РТА-3955; мутантное, рекомбинантное или генетически сконструированное производное растения, представленного под идентификационным номером патентного депозитария РТА-3953 или РТА-3955; любое потомство растения, представленного под идентификационным номером патентного депозитария РТА-3953 или РТА-3955; и растение, являющееся потомством любого из указанных растений. В предпочтительном варианте осуществления изобретения растение пшеницы по настоящему изобретению дополнительно имеет характеристики устойчивости к гербицидам, присущие растению, представленному под идентификационным номером патентного депозитария РТА-3953 или РТА-3955.
В объем настоящего изобретения входят также гибриды вышеописанных сортов пшеницы TealIMI 11A и TealIMI 15A. В примере 5 рассмотрены гибриды TealIMI 11A/TealIMI 15A, обладающие повышенной устойчивостью к имидазолиноновому гербициду. Настоящее изобретение далее относится к гибридам сортов пшеницы TealIMI 11A или TealIMI 15A и других сортов пшеницы. Другие сорта пшеницы включают, не ограничиваясь ими, T. aestivum L. сорта Fidel и любой сорт пшеницы, содержащий мутантный ген FS-1, FS-2, FS-3 или FS-4 (См. патент США № 6339184 и заявку на патент США № 08/474832). В предпочтительном варианте осуществления изобретения растение пшеницы представляет гибрид сорта TealIMI 11A и сорта Fidel FS-4. Гибриды TealIMI 11A/FS-4 включают нуклеиновые кислоты Imi1 и Imi2. Гибрид сортов TealIMI 11A и Fidel, содержащий ген FS-4, входит в объем настоящего изобретения и депонирован в Американскую коллекцию типовых культур, Manassas, Virginia, 3 января 2002 г. Указанный депозит создан в соответствии с нормами и правилами Будапештского договора о депонировании микроорганизмов. Депонирование было осуществлено на срок не менее тридцати лет и, по меньшей мере, на пять лет после последнего запроса к АТСС о предоставлении образца из хранилища. Депонированные семена представлены под идентификационным номером патентного депозитария РТА-3954.
Термины "культурный сорт" и "сорт" означают группу растений одного вида, обладающих общим набором характеристик или признаков, которые признаны специалистами в данной области достаточными для отличия одного сорта или культурного сорта от другого сорта или культурного сорта. Ни один из терминов не предполагает, что все растения любого данного сорта или культурного сорта являются генетически идентичными в отношении всего гена или на молекулярном уровне либо что любое данное растение является гомозиготным во всех локусах. Сорт или культурный сорт считается результатом "истинной селекции" определенного признака, если у самоопыляющегося сорта или культурного сорта все потомство имеет указанный признак. В настоящем изобретении признак возникает в результате мутации гена AHAS растения или семени пшеницы.
Следует отметить, что растение пшеницы по настоящему изобретению помимо гена IMI может включать немутированный, или дикого типа, ген AHAS. Как показано в примере 4, предполагается, что сорт пшеницы TealIMI 11A содержит мутацию только в одном из множества изоферментов AHAS и сорт пшеницы TealIMI 15A содержит мутацию только в двух из множества изоферментов AHAS. Поэтому в объем настоящего изобретения входит растение пшеницы, включающее одну или более нуклеиновых кислот IMI помимо одной или более немутированных или дикого типа нуклеиновых кислот AHAS.
Помимо растений пшеницы настоящее изобретение относится к выделенным белкам и нуклеиновым кислотам IMI. Нуклеиновые кислоты включают полинуклеотид, выбираемый из группы, состоящей из полинуклеотида SEQ ID NO: 1, полинуклеотида SEQ ID NO: 3, полинуклеотидной последовательности, кодирующей полипептид SEQ ID NO: 2, полинуклеотидной последовательности, кодирующей полипептид SEQ ID NO: 4, полинуклеотида, содержащего, по меньшей мере, 60 последовательно расположенных нуклеотидов из любых вышеуказанных полинуклеотидов, и полинуклеотида, комплементарного любым вышеуказанным полинуклеотидам. В предпочтительном варианте осуществления изобретения нуклеиновая кислота IMI включает полинуклеотидную последовательность, кодирующую полипептид SEQ ID NO: 2 или SEQ ID NO: 4. В другом предпочтительном варианте осуществления изобретения нуклеиновая кислота IMI включает полинуклеотидную последовательность SEQ ID NO: 1 или SEQ ID NO: 3.
Термин "белок AHAS" означает белок-синтазу ацетогидроксикислот, и термин "белок IMI" означает любой белок AHAS, который мутировал из белка AHAS дикого типа и придает повышенную устойчивость к имидазолинону растению, растительной клетке, части растения, семени растения или растительной ткани, в которых он экспрессирован. В предпочтительном варианте осуществления изобретения белок IMI включает полипептид SEQ ID NO: 2 или SEQ ID NO: 4. В используемом здесь значении термины "нуклеиновая кислота" и "полинуклеотид" означают РНК или ДНК, которая является линейной или разветвленной, одноцепочечной или двухцепочечной, или их гибрид. Данный термин также означает гибриды РНК/ДНК. Указанные термины означают также нетранслируемую последовательность, находящуюся как у 3'-конца, так и у 5'-конца кодирующей области гена: по меньшей мере, около 1000 нуклеотидов последовательности выше 5'-конца кодирующей области и, по меньшей мере, около 200 нуклеотидов последовательности ниже 3'-конца кодирующей области гена. Реже встречающиеся основания, такие как инозин, 5-метилцитозин, 6-метиладенин, гипоксантин и другие, могут быть также использованы для антисмыслового дцРНК и рибозимного спаривания. Например, установлено, что полинуклеотиды, содержащие С-5 пропиновые аналоги уридина и цитидина, могут связываться с РНК с высокой степенью сродства и являются сильнодействующими антисмысловыми ингибиторами экспрессии генов. Можно также произвести другие модификации, такие как изменение фосфодиэфирного скелета или 2'-гидроксильной группы в рибозосахарной группе РНК. Антисмысловые полинуклеотиды и рибозимы могут полностью состоять из рибонуклеотидов или могут содержать смесь рибонуклеотидов и дезоксирибонуклеотидов. Полинуклеотиды по данному изобретению можно получить любыми способами, включая геномные препараты, препараты кДНК, синтез in vitro, ОТ-ПЦР и транскрипцию in vitro или in vivo.
"Выделенная" молекула нуклеиновой кислоты представляет такую молекулу, которая по существу отделена от других молекул нуклеиновой кислоты, присутствующих в природном источнике нуклеиновой кислоты (то есть последовательности, кодирующие другие полипептиды). "Выделенная" нуклеиновая кислота предпочтительно не имеет некоторых последовательностей, которые в естественных условиях фланкируют нуклеиновую кислоту (то есть последовательности, находящиеся у 5'-конца и 3'-конца нуклеиновой кислоты) в природном репликоне. Например, клонированная нуклеиновая кислота считается выделенной. В разных вариантах осуществления изобретения выделенная молекула нуклеиновой кислоты IMI может содержать нуклеотидные последовательности длиной менее примерно 5 т.п.о., 4 т.п.о., 3 т.п.о., 2 т.п.о., 1 т.п.о., 0,5 т.п.о. или 0,1 т.п.о, которые в естественных условиях фланкируют молекулу нуклеиновой кислоты в геномной ДНК клетки, из которой получена данная нуклеиновая кислота (например, клетка Triticum aestivum). Нуклеиновая кислота также считается выделенной, если она изменена в результате вмешательства человека или помещена в локус или участок, не являющийся ее природным сайтом, или если указанная нуклеиновая кислота внесена в клетку агроинфицированием или высокоскоростной биолистической трансфекцией. Кроме того, "выделенная" молекула нуклеиновой кислоты, такая как молекула кДНК, может не иметь некоторых других клеточных веществ, с которыми она обычно бывает связана, или культуральной среды при получении методами генной инженерии или химических предшественников, или других химических веществ в случае химического синтеза.
Из определения термина "выделенные нуклеиновые кислоты" особо исключаются природные хромосомы (такие как хромосомные препараты), библиотеки искусственных хромосом, геномные библиотеки и библиотеки кДНК, которые существуют в виде препаратов нуклеиновых кислот in vitro или в виде препаратов трансфицированных/трансформированных клеток-хозяев, где клетки-хозяева представляют собой гетерогенный препарат in vitro или культивируются в виде гетерогенной популяции отдельных колоний. Из определения выделенной нуклеиновой кислоты исключены также вышеуказанные библиотеки, в которых специфическая нуклеиновая кислота составляет менее 5% от общего числа вставок нуклеиновых кислот в векторных молекулах. Кроме того, из данного определения исключена геномная ДНК клетки или препараты РНК клетки (включая цельноклеточные препараты, подвергнутые механическому фрагментированию или ферментативному расщеплению). Далее из указанного определения особо исключены цельноклеточные препараты, представляющие препарат in vitro или гетерогенную смесь, разделенную электрофорезом, в которой нуклеиновая кислота по данному изобретению не отделена от гетерологичных нуклеиновых кислот в среде для электрофореза (например, путем вырезания одной полосы из гетерогенной популяции, полос, в агарозном геле или найлоновом блотте).
Молекулу нуклеиновой кислоты по настоящему изобретению, например молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность SEQ ID NO: 1, SEQ ID NO: 3 или ее часть, можно выделить стандартными методами, применяемыми в молекулярной биологии, и на основании информации о последовательностях, приводимой в данном описании изобретения. Например, кДНК IMI T. aestivum можно выделить из библиотеки T. aestivum, используя полную или частичную последовательность SEQ ID NO: 1 или SEQ ID NO: 3. Кроме того, молекулу нуклеиновой кислоты, содержащую полную или частичную SEQ ID NO: 1 или SEQ ID NO: 3, можно выделить при помощи полимеразной цепной реакции с использованием олигонуклеотидных затравок, созданных на основе данной последовательности. Например, мРНК можно выделить из растительных клеток (например, методом экстракции гуанидинтиоционатом, описанным Chirgwin et al., 1979 Biochemistry 18: 5294-5299), и кДНК можно получить при помощи обратной транскриптазы (например, обратной транскриптазы Moloney MLV фирмы Gibco/BRL, Bethesda, MD; или обратной транскриптазы AMV фирмы Seikagaku America, Inc., St. Petersburg, FL). Синтетические олигонуклеотидные затравки для амплификации при помощи полимеразной цепной реакции можно создать на основе нуклеотидной последовательности, представленной в SEQ ID NO: 1 или SEQ ID NO: 3. Молекулу нуклеиновой кислоты по данному изобретению можно амплифицировать, используя кДНК или альтернативно геномную ДНК в качестве матрицы и соответствующие олигонуклеотидные затравки, стандартными методами амплификации при помощи ПЦР. Амплифицированную таким образом молекулу нуклеиновой кислоты можно клонировать в соответствующий вектор и исследовать, выполняя анализ последовательности ДНК. Кроме того, олигонуклеотиды, соответствующие нуклеотидной последовательности IMI, можно получить стандартными методами синтеза, например в автоматическом синтезаторе ДНК.
Нуклеиновые кислоты IMI по настоящему изобретению могут включать последовательности, кодирующие белок IMI (то есть "кодирующие области"), а также 5'-нетранслируемые последовательности и 3'-нетранслируемые последовательности. Альтернативно молекулы нуклеиновой кислоты по настоящему изобретению могут включать только кодирующие области гена IMI или могут содержать целые геномные фрагменты, выделенные из геномной ДНК. Кодирующая область указанных последовательностей определяется как "положение ORF". Кроме того, молекула нуклеиновой кислоты по данному изобретению может включать часть кодирующей области гена IMI, например фрагмент, который можно использовать в качестве зонда или затравки. Нуклеотидные последовательности, обнаруженные в результате клонирования генов IMI из T.aestivum, позволяют получить зонды и затравки, предназначенные для идентификации и/или клонирования гомологов IMI в других типах клеток и организмах, а также гомологов IMI из других растений пшеницы и родственных видов. Часть кодирующей области может также кодировать биологически активный фрагмент белка IMI.
В используемом здесь значении термин "биологически активная часть" белка IMI означает такую часть, как домен/мотив белка IMI, которая при продуцировании в растении повышает устойчивость данного растения к имидазолиноновому гербициду по сравнению с сортом растения дикого типа. Способы количественного определения повышенной устойчивости к имидазолиноновым гербицидам представлены в нижеследующих примерах. Биологически активные части белка IMI включают пептиды, кодируемые полинуклеотидными последовательностями, включающими SEQ ID NO: 1 или SEQ ID NO: 3, которые содержат меньше аминокислот, чем полноразмерный белок IMI, и придают повышенную устойчивость к имидазолиноновому гербициду при экспрессии в растении. Биологически активные части (например, пептиды, имеющие длину, равную, например, 5, 10, 15, 20, 30, 35, 36, 37, 38, 39, 40, 50, 100 или более аминокислотам) обычно включают домен или мотив, характеризующийся, по меньшей мере, одной активностью белка IMI. Кроме того, методами генной инженерии можно получить другие биологически активные части, в которых делетированы другие области полипептида, и исследовать их на наличие одной или более описанных активностей. Биологически активные части белка IMI предпочтительно включают один или более консервативных доменов, имеющих мутацию, которые выбирают из группы, состоящей из домена А, домена В, домена С, домена D и домена Е.
Данное изобретение относится также к химерным или слитым полипептидам IMI. В используемом здесь значении термин "химерный полипептид" или "слитый полипептид" IMI включает полипептид IMI, функционально связанный с полипептидом, не являющимся IMI. Термин "полипептид, не являющийся IMI" означает полипептид, имеющий аминокислотную последовательность, которая по существу не идентична полипептиду IMI, например полипептид, который не является изоферментом IMI и который выполняет другую функцию по сравнению с полипептидом IMI. Применительно к слитому полипептиду термин "функционально связанный" означает, что полипептид IMI и полипептид, не являющийся IMI, слиты друг с другом таким образом, что обе последовательности выполняют функцию, свойственную используемой последовательности. Полипептид, не являющийся IMI, может быть слит с N-концом или С-концом полипептида IMI. Например, в одном варианте осуществления изобретения слитый полипептид представляет собой слитый полипептид GST-IMI, в котором последовательность IMI слита с С-концом последовательности GST. Такие слитые полипептиды могут облегчить очистку рекомбинантных полипептидов IMI. В другом варианте осуществления изобретения слитый полипептид является полипептидом IMI, содержащим гетерологичную сигнальную последовательность у N-конца. В некоторых клетках-хозяевах (например, клетках-хозяевах млекопитающих) экспрессию и/или секрецию полипептида IMI можно увеличить благодаря использованию гетерологичной сигнальной последовательности.
Выделенную молекулу нуклеиновой кислоты, кодирующую полипептид IMI, имеющий последовательность, идентичную последовательности полипептида, кодируемого полинуклеотидной последовательностью SEQ ID NO: 1 или SEQ ID NO: 3, можно получить путем внесения одной или более замен, добавления или делеции нуклеотидов в нуклеотидную последовательность SEQ ID NO: 1 или SEQ ID NO: 3 таким образом, что в кодируемый полипептид вносят одну или более аминокислотных замен, добавлений или делеций. Мутации можно внести в последовательности SEQ ID NO: 1 или SEQ ID NO: 3 стандартными методами, такими как сайтнаправленный мутагенез и ПЦР-опосредуемый мутагенез. В консервативных аминокислотах замены предпочтительно производят в положении одного или более предполагаемых несущественных аминокислотных остатков.
Термин "консервативная аминокислотная замена" означает замену одного аминокислотного остатка другим аминокислотным остатком, имеющим подобную боковую цепь. В данной области определены семейства аминокислотных остатков, имеющих схожие боковые цепи. Такие семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, предполагаемый несущественный аминокислотный остаток в полипептиде IMI предпочтительно заменяют другим аминокислотным остатком из семейства с такими же боковыми цепями. Альтернативно в другом варианте осуществления изобретения мутации могут быть произведены произвольно в любом месте всей или части кодирующей последовательности IMI, например, при помощи насыщающего мутагенеза, после чего полученные мутанты можно исследовать с целью выявления требуемой активности IMI для идентификации мутантов, сохраняющих активность IMI. После мутагенеза последовательности SEQ ID NO: 1 или SEQ ID NO: 3 кодируемый полипептид может быть рекомбинантно экспрессирован, при этом активность полипептида можно определить, анализируя устойчивость к имидазолинону у растения, экспрессирующего полипептид, как это описано в нижеследующих примерах.
Для определения процента идентичности двух аминокислотных последовательностей (например, SEQ ID NO: 2 или SEQ ID NO: 4 и их мутантная форма) их сопоставляют с целью оптимального сравнения (например, в последовательность одного полипептида могут быть введены гэпы (пробелы) для оптимального сопоставления с другим полипептидом). Затем сопоставляют аминокислотные остатки в соответствующих положениях аминокислот. Если в каком-либо положении одной последовательности (например, SEQ ID NO: 2 или SEQ ID NO: 4) находится такой же аминокислотный остаток, что и в соответствующем положении другой последовательности (например, мутантная форма SEQ ID NO: 2 или SEQ ID NO: 4), то молекулы являются идентичными в данном положении. Подобным образом можно сравнить две последовательности нуклеиновых кислот. Процент идентичности последовательностей соответствует числу одинаковых положений в последовательностях (то есть процент идентичности последовательностей=число одинаковых положений/общее число положений × 100). В соответствии с целями данного изобретения процент идентичности двух нуклеотидных или полипептидных последовательностей определяют при помощи программного обеспечения Vector NTI 6.0 (РС) (InforMax, 7600 Wisconsin Ave., Bethesda, MD 20814). Для определения процента идентичности двух нуклеиновых кислот начисляют 15 штрафных очков за образование гэпа и 6,66 штрафного очка за удлинение гэпа. Для определения процента идентичности двух полипептидов начисляют 10 штрафных очков за образование гэпа и 0,1 штрафного очка за удлинение гэпа. Все другие параметры являются значениями, задаваемыми по умолчанию. Следует отметить, что для определения идентичности последовательностей при сравнении последовательности ДНК с последовательностью РНК нуклеотид тимидин эквивалентен нуклеотиду урацилу. Предпочтительно выделенные полипептиды IMI, входящие в объем настоящего изобретения, по меньшей мере, примерно на 50-60%, предпочтительно, по меньшей мере, примерно на 60-70%, более предпочтительно, по меньшей мере, примерно на 70-75%, 75-80%, 80-85%, 85-90% или 90-95% и наиболее предпочтительно, по меньшей мере, примерно на 96%, 97%, 98%, 99% или более процентов идентичны полной аминокислотной последовательности, кодируемой полинуклеотидной последовательностью, представленной SEQ ID NO: 1 или SEQ ID NO: 3. В другом варианте осуществления изобретения выделенные полипептиды IMI, входящие в объем настоящего изобретения, по меньшей мере, примерно на 50-60%, предпочтительно, по меньшей мере, примерно на 60-70%, более предпочтительно, по меньшей мере, примерно на 70-75%, 75-80%, 80-85%, 85-90% или 90-95% и наиболее предпочтительно, по меньшей мере, примерно на 96%, 97%, 98%, 99% или более процентов идентичны полной аминокислотной последовательности, представленной SEQ ID NO: 2 или SEQ ID NO: 4.
Кроме того, можно создать оптимизированные нуклеиновые кислоты IMI. Оптимизированная нуклеиновая кислота IMI предпочтительно кодирует полипептид IMI, который модулирует устойчивость растения к имидазолиноновым гербицидам и более предпочтительно повышает устойчивость растения к имидазолиноновому гербициду при сверхэкспрессии в растении. В используемом здесь значении термин "оптимизированный" относится к нуклеиновой кислоте, генетически сконструированной для увеличения ее экспрессии в данном растении или животном. Для получения оптимизированных нуклеиновых кислот IMI в растении последовательность ДНК гена может быть модифицирована так, чтобы она 1) включала кодоны, предпочтительные для генов растений с высокой экспрессией; 2) включала такое содержание А+Т, которое, по существу, соответствует обнаруженному в растениях; 3) образовывала инициирующую последовательность в растениях; 4) не содержала последовательностей, вызывающих дестабилизацию, неадекватное полиаденилирование, деградацию и терминацию РНК, или последовательностей, образующих "шпильки" вторичной структуры или сайты сплайсинга РНК. Повышенная экспрессия нуклеиновых кислот IMI в растениях может быть достигнута благодаря частоте распределения используемых кодонов в растениях вообще или в определенном растении в частности. Способы оптимизации экспрессии нуклеиновых кислот в растениях описаны в ЕРА 0359472 и 0385962; заявке РСТ WO 91/16432; патентах США №№ 5380831 и 5436391; Perlack et al., 1991 Proc. Natl. Acad. Sci. USA 88:3324-3328; и Murray et al., 1989 Nucleic Acids Res. 17:477-498.
В используемом здесь значении термин "частота предпочтительного использования кодонов" означает предпочтительное использование специфической клеткой-хозяином нуклеотидных кодонов для определения соответствующей аминокислоты. Чтобы определить частоту использования конкретного кодона в гене, количество повторов данного кодона в гене делят на общее количество повторов всех кодонов, определяющих одну и ту же аминокислоту в гене. Аналогичным образом частоту предпочтительного использования кодонов, характерную для клетки-хозяина, можно вычислить путем усреднения частоты предпочтительного использования кодонов в большом количестве генов, экспрессированных клеткой-хозяином. Желательно ограничить указанный анализ генами, высоко экспрессированными в клетке-хозяине. Процентное отклонение частоты предпочтительного использования кодона для синтетического гена от используемого клеткой-хозяином высчитывают, определяя сначала процентное отклонение частоты использования одного кодона от аналогичного кодона клетки-хозяина с последующим определением среднего отклонения для всех кодонов. В соответствии с определением такое вычисление относится к уникальным кодонам (то есть ATG и TGG). В общих чертах, общее среднее отклонение использования кодона оптимизированным геном от подобного кодона клетки-хозяина высчитывают при помощи уравнения 1А=n=1 Z Xn - YnXn × 100 Z, где Xn=частота использования кодона n в клетке-хозяине; Yn=частота использования кодона n в синтетическом гене, n означает отдельный кодон, определяющий аминокислоту, и Z означает общее число кодонов. Общее отклонение частоты использования кодона А для всех аминокислот должно предпочтительно быть менее примерно 25% и более предпочтительно менее примерно 10%.
Следовательно, нуклеиновая кислота IMI может быть оптимизирована таким образом, чтобы частота распределения используемых кодонов отклонялась предпочтительно не более чем на 25% от аналогичного значения для генов растения, экспрессирующихся на высоком уровне, и более предпочтительно не более чем примерно на 10%. Кроме того, следует учитывать процентное содержание G+C в пропущенном третьем основании (в однодольных растениях в указанном положении предпочтительно находится G+C, что не характерно для двудольных растений). Также известно, что кодон XCG (где Х означает А, Т, С или G) является наименее предпочтительным кодоном в двудольных растениях, при этом кодон ХТА не используется ни в однодольных, ни в двудольных растениях. Оптимизированные нуклеиновые кислоты IMI по данному изобретению также предпочтительно имеют индексы избегания дублетов CG и ТА, предельно приближающиеся к показателям выбранного растения-хозяина (то есть Triticum aestivum). Более предпочтительно указанные индексы отклоняются от показателей хозяина не более чем примерно на 10-15%.
Помимо молекул нуклеиновой кислоты, кодирующих вышеописанные полипептиды IMI, другим объектом данного изобретения являются выделенные антисмысловые молекулы нуклеиновой кислоты. Считается, что антисмысловые полинуклеотиды ингибируют экспрессию гена полинуклеотида-мишени в результате специфического связывания с полинуклеотидом-мишенью, препятствуя транскрипции, сплайсингу, транспорту, трансляции и/или устойчивости полинуклеотида-мишени. Ранее были описаны способы направленной доставки антисмыслового полинуклеотида к хромосомной ДНК, первичному транскрипту РНК или процессированной мРНК. Области-мишени предпочтительно включают сайты сплайсинга, инициирующие трансляцию кодоны, терминирующие трансляцию кодоны и другие последовательности в открытой рамке считывания.
Термин "антисмысловая" в соответствии с целями данного изобретения относится к нуклеиновой кислоте, включающей полинуклеотид, который является достаточно комплементарным всему гену или его части, первичному транскрипту или процессированной мРНК для того, чтобы препятствовать экспрессии эндогенного гена. "Комплементарные" полинуклеотиды являются такими полинуклеотидами, которые способны обеспечивать спаривание оснований в соответствии со стандартными правилами комплементарности Уотсона-Крика. В частности, пурины образуют пару оснований с пиримидинами, в результате чего происходит спаривание гуанина с цитозином (G:C) и аденина с тимином (А:Т) в случае ДНК или аденина с урацилом (A:U) в случае РНК. Очевидно, что два полинуклеотида могут гибридизировать друг с другом, даже если они не являются полностью комплементарными друг другу, при условии, что каждый имеет, по меньшей мере, одну область, которая по существу комплементарна другой. Термин "антисмысловая нуклеиновая кислота" означает экспрессирующие кластеры одноцепочечной РНК и двухцепочечной ДНК, которые могут быть транскрибированы с образованием антисмысловой РНК. "Активные" антисмысловые нуклеиновые кислоты представляют антисмысловые молекулы РНК, способные избирательно гибридизировать с первичным транскриптом или мРНК, кодирующей полипептид, характеризующийся, по меньшей мере, 80% идентичностью последовательности с полипептидом, кодируемым полинуклеотидной последовательностью SEQ ID NO: 1 или SEQ ID NO: 3.
Помимо вышеописанных нуклеиновых кислот и полипептидов IMI настоящее изобретение относится к нуклеиновым кислотам и полипептидам, присоединенным к одной из нижеследующих частей молекулы. Такие части включают, не ограничиваясь ими, части, обеспечивающие обнаружение, гибридизацию, очистку, доставку, взаимодействие, связывание и подобные функции. Типичная группа нуклеиновых кислот, имеющих присоединенные части, представляет зонды и затравки. Зонды и затравки обычно включают по существу выделенный олигонуклеотид. Олигонуклеотид обычно включает область нуклеотидной последовательности, которая гибридизирует в строгих условиях, по меньшей мере, примерно с 12, предпочтительно примерно с 25, более предпочтительно примерно с 40, 50 или 75 последовательно расположенными нуклеотидами смысловой цепи последовательности, приведенной в SEQ ID NO: 1 или SEQ ID NO: 3, антисмысловой цепи последовательности, приведенной в SEQ ID NO: 1 или SEQ ID NO: 3, или их природными мутантами. Затравки, созданные на основе нуклеотидной последовательности SEQ ID NO: 1 или SEQ ID NO: 3, можно использовать при осуществлении реакций ПЦР для клонирования гомологов IMI. Зонды, созданные на основе нуклеотидных последовательностей IMI, можно использовать для обнаружения транскриптов или геномных последовательностей, кодирующих одинаковые или гомологичные полипептиды. В предпочтительных вариантах осуществления изобретения зонд дополнительно содержит присоединенную меченую группу, например, меченая группа может представлять радиоизотоп, флуоресцирующее соединение, фермент или ферментный кофактор. Такие зонды можно использовать в составе тест-набора, включающего геномный маркер, для идентификации клеток, экспрессирующих полипептид IMI, например, путем измерения содержания IMI-кодирующей нуклеиновой кислоты в образце клеток, например, для обнаружения уровней мРНК IMI или определения того, мутировал ген геномной IMI или был делетирован.
Данное изобретение далее относится к выделенному рекомбинантному экспрессирующему вектору, включающему вышеописанную нуклеиновую кислоту IMI, экспрессия которого в клетке-хозяине вызывает повышенную устойчивость к имидазолиноновому гербициду по сравнению с диким сортом клетки-хозяина. В используемом здесь значении термин "вектор" означает молекулу нуклеиновой кислоты, способную переносить другую нуклеиновую кислоту, с которой она связана. Одним типом вектора является "плазмида", которая представляет кольцевую двухцепочечную петлю ДНК, с которой лигированы дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы с вирусным геномом. Некоторые векторы способны автономно реплицироваться в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальный механизм репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) встраивают в геном клетки-хозяина при введении в клетку-хозяин, в результате чего они реплицируют вместе с геномом хозяина. Кроме того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы определяются как "экспрессирующие векторы". Как правило, экспрессирующие векторы, используемые при осуществлении методов рекомбинантных ДНК, часто находятся в форме плазмид. В данном описании изобретения термины "плазмида" и "вектор" имеют взаимозаменяемые значения, так как плазмида является наиболее широко используемой формой вектора. Однако в объем данного изобретения входят и другие формы экспрессирующих векторов, такие как вирусные векторы (например, ретровирусы с дефектом репликации, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции.
Рекомбинантные экспрессирующие векторы по данному изобретению включают нуклеиновую кислоту по данному изобретению в форме, пригодной для экспрессии нуклеиновой кислоты в клетке-хозяине, из чего следует, что рекомбинантные экспрессирующие векторы имеют одну или более регуляторных последовательностей, выбираемых в зависимости от клеток-хозяев, используемых для экспрессии, которые функционально связаны с экспрессируемой последовательностью нуклеиновой кислоты. Применительно к рекомбинантному экспрессирующему вектору термин "функционально связанный" означает, что представляющая интерес нуклеотидная последовательность связана с одной или более регуляторными последовательностями таким образом, что это делает возможной экспрессию нуклеотидной последовательности (например, в транскрипционной/трансляционной системе in vitro или в клетке-хозяине при введении вектора в клетку-хозяин). Термин "регуляторная последовательность" означает промоторы, энхансеры и другие элементы регуляции экспрессии (например сигналы полиаденилирования). Такие регуляторные последовательности описаны, например, в издании Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990) или Gruber and Crosby, in: Methods in Plant Molecular Biology and Biotechnology, eds. Glick and Thompson, Chapter 7, 89-108, CRC Press: Boca Raton, Florida, которые включены в данное описание изобретения в качестве ссылки. Регуляторными последовательностями являются такие последовательности, которые управляют структурной экспрессией нуклеотидной последовательности во многих типах клеток-хозяев и которые управляют экспрессией нуклеотидной последовательности только в определенных клетках-хозяевах или в определенных условиях. Специалистам в данной области должно быть известно, что конструкция экспрессирующего вектора может зависеть от таких факторов, как выбор трансформируемой клетки-хозяина, уровня экспрессии требуемого полипептида и т.д. Экспрессирующие векторы по данному изобретению можно вводить в клетки-хозяева для продуцирования полипептидов или пептидов, включая слитые полипептиды или пептиды, кодируемые описанными нуклеиновыми кислотами (например, полипептиды IMI, слитые полипептиды и т.д.).
В предпочтительном варианте осуществления настоящего изобретения полипептиды IMI экспрессированы в растениях и растительных клетках, таких как клетки одноклеточных растений (например, водоросли) (см. Falciatore et al., 1999 Marine Biotechnology 1(3):239-251 и приведенные в указанной статье ссылки) и растительные клетки высших растений (например, семенные растения, такие как сельскохозяйственные культуры). Полинуклеотид IMI может быть "введен" в растительную клетку любым способом, включая трансфекцию, трансформацию или трансдукцию, электропорацию, бомбардировку частицами, высокоскоростную биолистическую трансфекцию, агроинфицирование и тому подобные. Один метод трансформации, известный специалистам в данной области, представляет погружение цветущего растения в раствор агробактерий, содержащих нуклеиновую кислоту IMI, с последуюшей селекцией трансформированных гамет.
Другие приемлемые методы трансформации или трансфекции клеток-хозяев, включая растительные клетки, представлены в издании Sambrook, et al. (Molecular Cloning: A Laboratory Manual. 2nd, ed., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989) и других лабораторных руководствах, таких как Methods in Molecular Biology, 1995, Vol.44, Agrobacterium protocols, ed: Gartland and Davey, Humana Press, Totowa, New Jersey. Так как повышенная устойчивость к имидазолиноновым гербицидам является общим признаком, желательно наследуемым рядом растений, таких как кукуруза, пшеница, рожь, овес, тритикале, рис, ячмень, соя, арахис, хлопок, рапс и канола, маниок, перец, подсолнечник и бархатцы, пасленовые культуры, такие как картофель, табак, баклажан и томат, вид Vicia, горох, люцерна, кустарниковые растения (кофе, какао, чай), вид Salix, деревья (гвинейская масличная пальма, кокосовая пальма), многолетние травы и фуражные культуры, указанные сельскохозяйственные культуры также являются предпочтительными растениями-мишенями, подлежащими модификации методами генетической инженерии, и представляют еще один вариант осуществления настоящего изобретения. Фуражные культуры включают, не ограничиваясь ими, пырей, канареечник канарский, костер, плевел, пырей жесткий, ежу сборную, люцерну, воробейник полевой, лядвенец рогатый, клевер гибридный, клевер луговой и донник.
В одном варианте осуществления настоящего изобретения трансфекция полинуклеотида IMI в растение достигается путем переноса гена, опосредуемого агробактериями. Опосредуемая агробактериями трансформация растения может быть произведена, например, при помощи штамма Agrobacterium tumefaciens GV3101 (pMP90) (Koncz and Schell, 1986 Mol. Gen. Genet. 204:383-396) или LBA4404 (Clontech). Трансформация может быть выполнена стандартными методами трансформации и регенерации (Deblaere et al., 1994 Nucl. Acids. Res. 13:4777-4788; Gelvin, Stanton B. and Schilperoort, Robert A, Plant Molecular Biology Manual, 2nd Ed. - Dordrecht: Kluwer Academic Publ., 1995. - in Sect., Ringbuc Zentrale Signatur: BT11-P ISBN 0-7923-2731-4; Glick, Bernard R. and Thompson, Johg E., Methods in Plant Molecular Biology and Biotechnology, Boca Raton: CRC Press, 1993 360 S., ISBN 0-8493-5164-2). Например, рапс можно трансформировать путем трансформации семядоли или гипокотиля (Moloney et al., 1989 Plant cell Report 8:238-242; De Block et al., 1989 Plant Physiol. 91:694-701). Использование антибиотиков для выбора агробактерий и растений зависит от бинарного вектора и штамма агробактерий, используемого для трансформации. Селекцию рапса обычно проводят, используя канамицин в качестве селектируемого маркера растения. Опосредуемый агробактериями перенос гена в лен можно произвести, например, при помощи метода, описанного в статье Mlynarova et al., 1994 Plant Cell Report 13:282-285. Кроме того, сою можно трансформировать, например, методом, описанным в европейском патенте № 0424047, патенте США № 5322783, европейском патенте № 0397687, патентах США №№ 5376543 или 5169770. Кукурузу можно трансформировать бомбардировкой частицами, поглощением ДНК, опосредуемым полиэтиленгликолем, или методом на основе карбидкремниевых волокон. (См., например, Freeling and Walbot "The maize handbook" Springer Verlag: New York (1993) ISBN 3-540-97826-7.) Типичный пример трансформации кукурузы приведен в патенте США № 5990387, и типичный пример трансформации пшеницы приведен в заявке РСТ WO 93/07256.
В соответствии с настоящим изобретением введенный полинуклеотид IMI может устойчиво сохраняться в растительной клетке, если он встроен во внехромосомный автономный репликон или хромосомы растения. Альтернативно введенный полинуклеотид IMI может находиться во внехромосомном нереплицирующем векторе, будучи временно экспрессированным или временно активным. В одном варианте осуществления изобретения может быть создан гомологичный рекомбинантный микроорганизм, в хромосому которого встроен полинуклеотид IMI, получен вектор, содержащий, по меньшей мере, часть гена AHAS с делецией, добавлением или заменой, предназначенными для изменения, например, функционального разрушения эндогенного гена AHAS и создания гена IMI. Для получения точковой мутации путем гомологичной рекомбинации можно использовать гибриды ДНК-РНК при осуществлении метода, известного как химеропластика (Cole-Strauss et al., 1999 Nucleic Acids Research 27(5):1323-1330 and Kmiec, 1999 Gene therapy American Scientist 87(3):240-247). Другие методы гомологичной рекомбинации вида Triticum также известны в данной области и могут быть использованы в данном изобретении.
В векторе гомологичной рекомбинации ген IMI может быть фланкирован у 5'-конца и 3'-конца дополнительной молекулой нуклеиновой кислоты гена AHAS для обеспечения гомологичной рекомбинации между экзогенным геном IMI, переносимым вектором, и эндогенным геном AHAS в микроорганизме или растении. Дополнительная фланкирующая молекула нуклеиновой кислоты AHAS имеет длину, достаточную для успешной гомологичной рекомбинации с эндогенным геном. Обычно в вектор встраивают фланкирующую ДНК длиной от нескольких сотен до тысяч пар оснований (как у 5'-конца, так и у 3'-конца) (см., например, Thomas, K.R., and Capecchi, M.R., 1987 Cell 51:503 для ознакомлением с описанием векторов гомологичной рекомбинации или Strepp et al., 1998 PNAS, 95(8):4368-4373 для ознакомления с рекомбинацией на основе кДНК в Physcomitrella patens). Однако поскольку ген IMI обычно отличается от гена AHAS незначительным числом аминокислот, фланкирующая последовательность не всегда является необходимой. Вектор гомологичной рекомбинации вводят в микроорганизм или растительную клетку (например, при помощи ДНК, опосредуемой полиэтиленгликолем), при этом клетки, в которых происходит гомологичная рекомбинация введенного гена IMI с эндогенным геном AHAS, выбирают методами, известными в данной области.
В другом варианте осуществления изобретения можно получить рекомбинантные микроорганизмы, содержащие выбранные системы, обеспечивающие регулируемую экспрессию введенного гена. Например, введение гена IMI в вектор, передающий функцию контроля lac-оперону, делает возможной экспрессию гена IMI только в присутствии IPTG. Такие регуляторные системы хорошо известны в данной области.
Полинуклеотид IMI, присутствующий во внехромосомном нереплицирующем векторе или векторе, встроенном в хромосому, предпочтительно находится в экспрессирующем кластере растения. Экспрессирующий кластер растения предпочтительно содержит регуляторные последовательности, способные регулировать экспрессию генов в растительных клетках, которые функционально связаны таким образом, что каждая последовательность может выполнять свою функцию, например терминацию транскрипции сигналами полиаденилирования. Предпочтительными сигналами полиаденилирования являются сигналы, формируемые тДНК Agrobacterium tumefaciens, например, геном 3, известным как октопинсинтаза Ti-плазмиды pTiACH5 (Gielen et al., 1984 EMBO J. 3:835), или его функциональными эквивалентами, а также всеми другими терминаторами, функционально активными в растениях. Так как экспрессирующий ген растения очень часто не ограничен на транскрипционных уровнях, экспрессирующий кластер растения предпочтительно содержит другие функционально связанные последовательности, подобные энхансерам трансляции, такие как ускоряющая последовательность, содержащая 5'-нетранслируемую лидерную последовательность вируса мозаики табака, увеличивающую выход полипептида относительно РНК (Gallie et al., 1987 Nucl. Acids Research 15:8693-8711). Примеры экспрессирующих векторов растений включают векторы, подробно описанные в изданиях Becker, D. et al., 1992 New plant binary vectors with selectable markers located proximal to the left border, Plant Mol. Biol. 20:1195-1197; Bevan, M.W., 1984 Binary Agrobacterium vectors for plant transformation, Nucl. Acid. Res. 12:8711-8721; and Vectors for Gene Transfer in Higher Plants; in: Transgenic Plants, Vol.1, Engineering and Utilization, eds.; Kung and R. Wu, Academic Press, 1993, S. 15-38.
Экспрессирующий ген растения должен быть функционально связан с соответствующим промотором, вызывающим своевременную и специфическую для клетки или ткани экспрессию гена. Промоторы, используемые в экспрессирующих кластерах по данному изобретению, включают любой промотор, способный инициировать транскрипцию в растительной клетке. Указанные промоторы включают, не ограничиваясь ими, такие промоторы, которые могут быть получены из растений, растительных вирусов и бактерий, содержащих гены, экспрессируемые в растениях, в частности Agrobacterium и Rhizobium.
Промотор может быть конститутивным, индуцируемым, специфическим для стадии развития, клетко-, ткане- или органспецифическим. Конститутивные промоторы являются активными в большинстве условий. Примеры конститутивных промоторов включают промоторы CaMV 19S и 35S (Odell et al. 1985 Nature 313:810-812), промотор sX CaMV 35S (Kay et al. 1987 Science 236:1299-1302), промотор Sep1, актиновый промотор риса (MsElroy et al. 1990 Plant Cell 2:163-171), актиновый промотор Arabidopsis, убихитановый промотор (Christensen et al. 1989 Plant Molec Biol. 18:675-689); pEmu (Last et al. 1991 Theor Appl Genet 81:581-588), промотор 35S вируса мозаики норичника шишковатого, промотор Smas (Velten et al. 1984 EMBO J. 3:2723-2730), промотор GRP1-8, промотор, представляющий (циннамилалкоголь)дегидрогеназу (патент США № 5683439), промоторы, полученные из Т-ДНК Agrobacterium, такие как маннопинсинтаза, нопалинсинтаза и октопинсинтаза, промотор, представляющий небольшую субъединицу рибулозобифосфаткарбоксилазы (ssuRUBISCO), и тому подобные.
Индуцируемые промоторы являются активными в определенных окружающих условиях, например при наличии или отсутствии питательного вещества или метаболита, тепла или холода, света, воздействия патогенных микроорганизмов, анаэробных условий и тому подобных. Например, промотор hsp80, полученный из Brassica, индуцируется тепловым шоком, промотор PPDK индуцируется светом, промотор PR-1, полученный из табака, Arabidopsis, и кукурузы, индуцируется инфекцией, вызываемой патогенным микроорганизмом, и промотор Adh1 индуцируется гипоксией и стрессом, вызываемым холодом. Индуцируемый промотор может также облегчать экспрессию растительного гена (см. Gatz, 1997 Annu. Rev. Plant Physiol. Plant Mol. Biol. 48:89-108). Химически индуцируемые промоторы особенно пригодны в тех случаях, когда желательно, чтобы экспрессия гена происходила в зависимости от времени. Примерами таких промоторов являются промотор, индуцируемый салициловой кислотой (заявка РТС WO 95/19443), промотор, индуцируемый тетрациклином (Gatz et al., 1992 Plant J. 2:397-404) и промотор, индуцируемый этанолом (заявка РСТ WO 93/21334).
Промоторы, специфические для стадии развития, предпочтительно экспрессируются на определенных стадиях развития. Ткане- и органспецифические промоторы включают такие промоторы, которые предпочтительно экспрессируются в определенных тканях или органах, таких как листья, корни, семена или ксилема. Ткане- и органспецифические промоторы включают, не ограничиваясь ими, промоторы, специфические для плодов, семяпочки, мужской ткани, семени, наружного покрова, клубня, побега, околоплодника и листа, рыльца, пыльцы, пыльника, лепестка, чашелистика, цветоножки, стручка, стебля, корня и тому подобных. Семяспецифические промоторы предпочтительно экспрессируются во время развития и/или прорастания семени. Например, семяспецифические промоторы могут предпочтительно воздействовать на завязь, эндосперму и семенную оболочку. См. Thompson et al. 1989 BioEssays 10:108. Примеры семяспецифических промоторов включают, не ограничиваясь ими, целлюлозосинтазу (celA), Cim1, гамма-зеин, глобулин-1, зеин кукурузы массой 19 кДа (cZ19B1) и тому подобные.
Другие подходящие ткане- или органспецифические промоторы включают промотор напин-гена, полученный из семян рапса (патент США № 5608152), USP-промотор, полученный из Vicia faba (Baeumlein et al., 1991 Mol Gen Genet. 225(3):459-67), олеозиновый промотор, полученный из Arabidopsis (заявка РСТ WO 98/45461), фазеолиновый промотор, полученный из Phaseolus vulgaris (патент США № 5504200), Все4-промотор, полученный из Brassica (заявка РСТ WO 91/13980) или промотор бобовых В4 (LeB4; Baeumlein et al., 1992 Plant Journal, 2(2): 233-9), а также промоторы, обеспечивающие семяспецифическую экспрессию в однодольных растениях, таких как кукуруза, ячмень, пшеница, рожь, рис и т.д. Подходящими промоторами является промотор lpt2 или lpt1-гена, полученный из ячменя (заявка РСТ WO 95/15389 и заявка РСТ WO 95/23230), или промоторы, описанные в заявке РСТ WO 99/16890 (промоторы, полученные из гена гордеина ячменя, гена глютелина риса, гена оризина риса, гена проламина риса, гена глиадина пшеницы, гена глютелина пшеницы, гена глютелина овса, гена казирина сорго и гена секалина ржи).
Другие промоторы, пригодные для использования в экспрессирующих кластерах по данному изобретению, включают, не ограничиваясь ими, промотор белка, связывающего основной хлорофилл a/b, гистоновые промоторы, промотор Ар3, конглициновый промотор, напиновый промотор, лектиновый промотор сои, промотор зеина кукурузы 15 кДа, промотор зеина 22 кДа, промотор зеина 27 кДа, промотор g-зеина, воскообразные промоторы, shrunken 1, shrunken 2 и бронзовые промоторы, промотор Zm13 (патент США № 5086169), полигалактуроназные промоторы кукурузы (PG) (патенты США №№ 5412085 и 5545546) и промотор SGB6 (патент США № 5470359), а также синтетические или другие природные промоторы.
Дополнительная гибкость при регулировании экспрессии гетерологичного гена в растениях может быть достигнута при использовании ДНК-связывающих доменов и чувствительных элементов, полученных из гетерологичных источников (то есть ДНК-связывающие домены из нерастительных источников). Примером такого гетерологичного ДНК-связывающего домена является ДНК-связывающий домен LexA (Brent and Prashne, Cell 43:729-736 (1985)).
Другим объектом настоящего изобретения являются клетки-хозяева, в которые вводят рекомбинантный экспрессирующий вектор по данному изобретению. Термины "клетка-хозяин" и "рекомбинантная клетка-хозяин" являются взаимозаменяемыми. Следует отметить, что такие термины означают не только конкретную клетку, но и потомство или потенциальное потомство такой клетки. Так как в последующих поколениях могут возникать определенные модификации вследствие мутации или под воздействием окружающих условий, такое потомство фактически не может быть идентично родительской клетке, и все же такое потомство входит в определение используемого здесь термина. Клетка-хозяин может быть прокариотической или эукариотической клеткой. Например, полинуклеотид IMI может быть экспрессирован в бактериальных клетках, таких как C. glutamicum, клетках насекомых, клетках грибов или клетках млекопитающих (таких как клетки яичника китайского хомячка (СНО) или клетки COS), водорослях, ресничных, растительных клетках, грибах или других микроорганизмах, таких как C. glutamicum. В данной области известны другие приемлемые клетки-хозяева.
Клетка-хозяин по данному изобретению, в частности прокариотическая или эукариотическая клетка-хозяин в культуре, может быть использована для продуцирования (то есть экспрессии) полинуклеотида IMI. Настоящее изобретение далее относится к способам продуцирования полипептидов IMI с использованием клеток-хозяев по данному изобретению. В одном варианте осуществления изобретения способ включают культивирование клетки-хозяина по данному изобретению (в которую введен рекомбинантный экспрессирующий вектор, кодирующий полипептид IMI, или в геном которой введен ген, кодирующий полипептид дикого типа или полипептид IMI) в приемлемой среде до продуцирования полипептида IMI. В другом варианте осуществления изобретения способ дополнительно включает выделение полипептидов IMI из среды или клетки-хозяина. Другим объектом настоящего изобретения являются выделенные полипептиды IMI и их биологически активные части. "Выделенный" или "очищенный" полипептид или его биологически активная часть не содержат клеточного вещества при продуцировании методами генной инженерии, химических предшественников или других химических веществ, образующихся в процессе химического синтеза. Термин "по существу не содержащий клеточного материала" относится к препаратам полипептида IMI, в которых полипептид отделен от клеточных компонентов клеток, в которых он продуцирован естественным путем или рекомбинантными методами. В одном варианте осуществления изобретения термин "по существу не содержащий клеточного материала" относится к препаратам полипептида IMI, содержащим менее примерно 30% (по сухой массе) вещества, не являющегося IMI (определяемого также как "загрязняющий полипептид"), более предпочтительно менее примерно 20% вещества, не являющегося IMI, еще предпочтительнее менее примерно 10% вещества, не являющегося IMI, и наиболее предпочтительно менее примерно 5% вещества, не являющегося IMI.
Полипептид IMI или его биологически активная часть, продуцируемые методами генной инженерии, также предпочтительно не содержат культуральной среды, то есть культуральная среда составляет менее примерно 20%, более предпочтительно менее примерно 10% и наиболее предпочтительно менее примерно 5% от объема полипептидного препарата. Термин "по существу не содержащий химических предшественников или других химических веществ" относится к препаратам полипептида IMI, в которых полипептид отделен от химических предшественников или других химических веществ, участвующих в синтезе полипептида. В одном варианте осуществления изобретения термин "по существу не содержащий химических предшественников или других химических веществ" относится к препаратам полипептида IMI, содержащим менее примерно 30% (по сухой массе) химических предшественников или химических веществ, не являющихся IMI, более предпочтительно менее примерно 20% химических предшественников или химических веществ, не являющихся IMI, еще предпочтительнее менее примерно 10% химических предшественников или химических веществ, не являющихся IMI, и наиболее предпочтительно менее примерно 5% химических предшественников или химических веществ, не являющихся IMI. В предпочтительных вариантах осуществления изобретения выделенные полипептиды или их биологически активные части не имеют загрязняющих полипептидов из того же организма, из которого был выделен полипептид IMI. Такие полипептиды обычно продуцируются путем рекомбинантной экспрессии, например полипептид IMI Triticum aestivum в растениях, не являющихся Triticum aestivum, или в микроорганизмах, таких как C. glutamicum, ресничные, водоросли или грибы.
Полинуклеотидные и полипептидные последовательности IMI по настоящему изобретению имеют ряд применений. Нуклеотидные и аминокислотные последовательности по настоящему изобретению можно использовать для трансформации растений, в частности для модуляции устойчивости растения к имидазолиноновым гербицидам. Таким образом, данное изобретение относится к способу получения трансгенного растения с повышенной устойчивостью к имидазолиноновому гербициду, который включает (а) трансформацию растительной клетки одним или более экспрессирующими векторами, включающими одну или более нуклеиновых кислот IMI, и (b) получение из растительной клетки трансгенного растения с повышенной устойчивостью к имидазолиноновому гербициду по сравнению с сортом дикого типа растения. В одном варианте осуществления изобретения множество нуклеиновых кислот IMI выделяют из разных геномов. В объем настоящего изобретения входят способы получения трансгенного растения с повышенной устойчивостью к имидазолиноновому гербициду, включающие (а) трансформацию растительной клетки экспрессирующим вектором, содержащим нуклеиновую кислоту IMI, которая не является нуклеиновой кислотой Imi1, и (b) получение из растительной клетки трансгенного растения с повышенной устойчивостью к имидазолиноновому гербициду по сравнению с диким сортом растения.
Настоящее изобретение относится к способам модификации устойчивости растения к имидазолиноновому гербициду, которые включают модификацию экспрессии одной или более нуклеиновых кислот IMI. Нуклеиновые кислоты предпочтительно находятся в разных геномах или получены из разных геномов. Устойчивость растения к имидазолиноновому гербициду можно повысить или понизить, что достигается соответственно увеличением или уменьшением экспрессии полинуклеотида IMI. Устойчивость растения к имидазолиноновому гербициду предпочтительно повышают, увеличивая экспрессию полинуклеотида IMI. Экспрессию полинуклеотида IMI можно модифицировать любым способом, известным специалистам в данной области. Способы увеличения экспрессии полинуклеотидов IMI применяют в зависимости от того, является растение трансгенным или нетрансгенным. Трансгенное растение можно трансформировать, например, при помощи вектора, содержащего любые из вышеописанных IMI-кодирующих нуклеиновых кислот, или промотора, регулирующего экспрессию эндогенных полинуклеотидов IMI в растении. В соответствии с данным изобретением такой промотор может быть тканеспецифическим или активируемым в зависимости от стадии развития. Альтернативно в нетрансгенных растениях экспрессия эндогенного полинуклеотида IMI может быть модифицирована путем индукции нативного промотора. Экспрессию полинуклеотидов, включающих SEQ ID NO: 1 или SEQ ID NO: 3, в растения-мишени можно осуществлять с помощью одного из нижеследующих компонентов, не ограничиваясь ими: (а) конститутивного промотора, (b) химически индуцируемого промотора и (с) сконструированного промотора вместе с факторами транскрипции, например, цинксодержащей пальцеобразной области (Greisman and Pabo, 1997 Science 275:657).
В предпочтительном варианте осуществления изобретения транскрипцию полинуклеотида IMI модулируют при помощи факторов транскрипции цинксодержащей пальцеобразной области (ZFP), описанных в статье Greisman and Pabo, 1997 Science 275:657 и созданных компанией Sangamo Biosciences, Inc. Указанные факторы ZFP содержат как домен распознавания ДНК, так и функциональный домен, вызывающий активацию или подавление нуклеиновой кислоты-мишени, такой как нуклеиновая кислота IMI. Таким образом могут быть получены активирующие или подавляющие факторы ZFP, которые специфически распознают вышеописанные промоторы полинуклеотидов IMI и могут быть использованы для увеличения или уменьшения экспрессии полинуклеотида IMI в растении, в результате чего изменяется устойчивость растения к гербициду.
Как более подробно описано выше, растения, получаемые способами по настоящему изобретению, могут быть однодольными или двудольными. Растения могут быть выбраны из таких растений, как кукуруза, пшеница, рожь, овес, тритикале, рис, ячмень, соя, арахис, хлопок, рапс, канола, маниок, перец, подсолнечник, бархатцы, пасленовые культуры, картофель, табак, баклажан, томат, вид Vicia, горох, люцерна, кофе, какао, чай, вид Salix, гвинейская масличная пальма, кокосовая пальма, многолетние травы и фуражные культуры. Фуражные культуры включают, не ограничиваясь ими, пырей, канареечник канарский, костер, плевел, пырей жесткий, ежу сборную, люцерну, воробейник полевой, лядвенец рогатый, клевер гибридный, клевер луговой и донник. В предпочтительном варианте осуществления изобретения растение является растением пшеницы. Во всех вышеописанных способах растительная клетка включает, не ограничиваясь ими, протопласт, гаметаобразующую клетку и клетку, которая регенерирует с образованием целого растения. В используемом здесь значении термин "трансгенный" относится к любому растению, растительной клетке, каллюсу, растительной ткани или части растения, которые содержат, по меньшей мере, один полный или частичный рекомбинантный полинуклеотид. Во многих случаях осуществляют стабильную интеграцию полного или частичного рекомбинантного полинуклеотида в хромосому или стабильный внехромосомный элемент, так чтобы он передавался последующим поколениям.
Как описано выше, настоящее изобретение относится к композициям и способам увеличения устойчивости к имидазолинону у растения или семени пшеницы по сравнению с диким сортом растения или семени. В предпочтительном варианте осуществления изобретения устойчивость к имидазолинону у растения или семени пшеницы увеличивается настолько, что растение или семя может выдерживать воздействие имидазолинонового гербицида, применяемого в количестве предпочтительно около 10-400 г а.и. (грамм активного ингредиента)/га, более предпочтительно 20-160 г а.и./га и наиболее предпочтительно 40-80 г а.и./га. В используемом здесь значении термин "выдерживать" воздействие имидазолинонового гербицида означает, что растение не гибнет или не повреждается в результате такого воздействия.
Кроме того, настоящее изобретение относится к способу борьбы с сорняками, произрастающими рядом с растением пшеницы, который включает нанесение имидазолинонового гербицида на сорняки и растение пшеницы, при этом растение пшеницы обладает повышенной устойчивостью к имидазолиноновому гербициду по сравнению с диким сортом пшеницы и при этом растение включает одну или более нуклеиновых кислот IMI. В одном варианте осуществления изобретения растение включает множество нуклеиновых кислот IMI, находящихся в разных геномах или полученных из разных геномов. В другом варианте осуществления изобретения растение содержит нуклеиновую кислоту, не являющуюся Imi1. Благодаря созданию пшеницы с повышенной устойчивостью к имидазолинону можно использовать разнообразные препараты для защиты растений пшеницы от сорняков, в результате чего усиливается рост растений и уменьшается конкуренция за питательные вещества. Имидазолиноновый гербицид можно использовать отдельно для довсходового, послевсходового, предпосевного и посевного уничтожения сорняков на участках, окружающих растения пшеницы, либо можно применять препарат имидазолинонового гербицида, содержащий другие добавки. Имидазолиноновый гербицид можно также использовать для обработки семян. Препарат имидазолинонового гербицида может содержать такие добавки, как другие гербициды, детергенты, адъюванты, распределяющие вещества, связывающие вещества, стабилизирующие вещества или тому подобные. Имидазолиноновый гербицид может быть мокрым или сухим препаратом и может включать, не ограничиваясь ими, текучие порошки, эмульгируемые концентраты и жидкие концентраты. Препараты, содержащие имидазолиноновый гербицид и другие гербициды, можно наносить известными методами, такими как опрыскивание, орошение, поливки или тому подобные.
В данном описании изобретения сделаны ссылки на разные публикации. Все вышеуказанные публикации и ссылки, приведенные в таких публикациях, полностью включены в данную заявку в качестве ссылки для более полного описания уровня техники в области, к которой относится настоящее изобретение.
Следует отметить, что все вышеизложенное относится к предпочтительным вариантам осуществления настоящего изобретения, в которые могут быть внесены многочисленные изменения, не выходящие за пределы объема данного изобретения. Изобретение далее проиллюстрировано нижеследующими примерами, которые никоим образом не ограничивают объем изобретения. Наоборот, совершенно очевидно, что специалисты в данной области могут предложить разные другие варианты осуществления изобретения, модификации и эквиваленты, не выходящие за пределы сущности настоящего изобретения и/или объема прилагаемой формулы изобретения.
Примеры
Пример 1
Мутагенез и селекция устойчивых линий пшеницы
Примерно 40000 семян Triticum aestivum L. сорта CDC Teal (Hughes and Hucl, 1993 Can. J. Plant Sci. 73:193-197) подвергают мутагенезу при помощи модифицированных способов, описанных Вашингтоном и Сиарсом (Washington and Sears, 1970). Семена предварительно замачивают в дистиллированной воде в течение четырех часов и обрабатывают 0,3% EMS в течение шести часов. Затем семена непрерывно промывают водопроводной водой в течение семи часов и сушат в течение примерно четырех часов до посева в поле. Растения М1 оплодотворяются самоопылением, и собранные семена хранят россыпью. На следующий год в поле выращивают примерно 2×106 растений М2, которые опрыскивают в фазе двух листьев имазамоксом при норме внесения 40 г а.и./га в разбрызгиваемом объеме 100 л/га. В раствор для опрыскивания добавляют 0,05% (об./об.) смешиваемого адъюванта. Отбирают шесть линий, устойчивых к имазамоксу, которые обозначают как линии 1А, 9А, 10А, 11А, 15А и 16А. Поколения М3 и М4 выращивают в помещении для проращивания семян и отбирают растения, устойчивые к имазамоксу, обрабатывая их вышеуказанным веществом при норме внесения 20 г/га. Устойчивые растения отбирают в поколении М5 после нанесения 40 г/га на поле. Семя М5 является гомозиготным в отношении данного признака, так как результаты испытания потомства свидетельствуют об отсутствии различий в устойчивости к имазамоксу.
Пример 2
Способы, используемые для определения наследования и аллелизма генов IMI
Для определения генетического контроля устойчивости к имазамоксу в шести линиях пшеницы производят реципрокные скрещивания шести устойчивых гомозиготных линий М6 и CDC Teal (сорт, восприимчивый к имазамоксу). Растения F1, произвольно отобранные во всех скрещиваниях, подвергают обратному скрещиванию с CDC Teal, получая при этом обратноскрещенные популяции (ВС)F1. Для исследования аллелизма производят все возможные интеркроссы шести мутантов и SWP965001 (Grandin/3*Fidel-FS-4). SWP965001 представляет линию яровой пшеницы, которая является гомозиготной для аллеля FS-4. Родительские генотипы выращивают в помещении для проращивания семян с 16-часовым фотопериодом при дневной температуре 24°С и ночной температуре 16°С. Колосья, вышедшие в трубку на 3/4 высоты, стерилизуют и опыляют через 2-3 дня после эмаскуляции. Растения F2, произвольно выбранные из всех расщепляющих скрещиваний, оплодотворяют самоопылением с образованием семейств F2:3. Родительские растения, растения F1, BCF1, F2 и семейства F2:3 испытывают с целью определения их реакции на имазамокс. Все эксперименты выполняют в помещении для проращивания семян с 16-часовым фотопериодом, дневной температурой 23°С и ночной температурой 16°С. Все эксперименты выполняют по совершенно рандомизированной схеме. В экспериментах по испытанию семейств F2:3 усилия направлены на рандомизацию как внутри семейств, так и между семействами. Популяции F1 и F2 исследуют в одном эксперименте вместе с родительскими генотипами и сортом CDC Teal, используемыми в качестве контрольных образцов. Популяции BCF1 и F2:3 исследуют в двух отдельных экспериментах вместе с соответствующими родительскими генотипами, используемыми в качестве контрольных образцов.
Растения, выращиваемые в ящиках для выгонки рассады с 8×16 ячейками, обрабатывают гербицидом в фазе двух листьев при помощи передвижного кабельного опрыскивателя, распыляющего 100 л/га. Имазамокс наносят на растения при норме 20 г а.и./га, используя насадку 8001 EVS, под давлением 275 кПа. К раствору гербицида перед применением добавляют связывающее поверхностно-активное вещество (0,05% об./об.). Через пятнадцать дней после нанесения гербицида производят оценку растений, сравнивая их реакции с реакциями родительских растений, и на основании полученных результатов определяют растения как устойчивые, промежуточные или чувствительные. Устойчивые растения фенотипически не подвергаются воздействию гербицида после произведенной обработки, в то время как у промежуточных растений прекращается рост первых двух листьев, наблюдается потемнение (темно-зеленая пигментация) листьев и появляются колеоптильные побеги. У чувствительных растений не образуются новые листья, наблюдается обширный хлороз листьев, и в конце концов растение погибает. Для анализа расщепляющихся популяций по Менделю растения оценивают по категориям устойчивой и неустойчивой реакции и испытывают на соответствие разным моделям с 1 геном, 2 генами и 3 генами по методу хи-квадрат. На основании данных для растений F2 и BCF1 промежуточные реакции включены в категорию устойчивой реакции. Поправка Йейтса на непрерывность использована для проверки по критерию хи-квадрат при выполнения анализа методом хи-квадрат только с одной степенью свободы (Steele and Torrie 1980 Principles and procedures of statistics, McGraw-Hill, New York, New York, pp 633).
Пример 3
Результаты наследования генов IMI
Все устойчивые родительские формы образуют аналогичный фенотип при опрыскивании 20 г а.и./га имазамокса (фиг.1). Реципрокные скрещивания устойчивых линий и чувствительной родительской формы (CDC Teal) позволяют получить растения F1, устойчивые к воздействию имазамокса (фиг.1), из чего следует, что устойчивость к имазамоксу является ядерным, а не цитоплазматическим признаком. За исключением скрещивания 15А × Teal растения F1, полученные из устойчивых линий, скрещенных с сортом CDC Teal, характеризуются промежуточной реакцией (фиг.1). Так как растения F1 в фенотипическом отношении занимают промежуточное положение между двумя родительскими формами, можно сделать вывод о том, что устойчивость к имазамоксу у указанных линий является частично доминантным признаком (фиг.1). Результаты генетического анализа устойчивости к имидазолинонам и сульфонилмочевинам у Arabidopsis thaliana (Haughn and Somerville, 1986 Mol. Gen. Genet. 204:430-434), Zea mays (Newhouse et al., 1991 Theor. Appl. Genet. 83:65-70), Brassica napus (Swanson et al., 1989 Theor. Appl. Gen. 78:525-530) и Glycine max (Sebastian et al., 1989 Crop Sci. 29:1403-1408) также указывают на наличие одного частично доминантного ядерного гена.
Четырнадцать растений F1, полученных в результате скрещивания 15А × Teal, оценены как устойчивые (фиг.1). Оценка популяций F2, полученных в результате указанного скрещивания, показывает, что два независимо расщепляющихся локуса придают устойчивость в данном генотипе (фиг.2). Так как растения F1 имеют два гетерозиготных устойчивых локуса, следует ожидать наличия устойчивой реакции. Если каждый из указанный локусов в отдельности придает частичную доминантность, то два гетерозиготных локуса должны вызывать устойчивую реакцию. Свансон и др. (Swanson et al., 1989) объединили два полудоминантных устойчивых к имидазолинону аллеля из Brassica napus, представляющих два несцепленных гена, получив при этом гибрид F1, характеризующийся более высокой устойчивостью к имидазолинону по сравнению с отдельными гетерозиготными линиями. Вышеуказанные авторы пришли к выводу, что механизмы устойчивости являются аддитивными и более высокий уровень устойчивости наблюдается в линиях, имеющих более одного аллеля устойчивости.
Анализ цитоплазматического наследования выполняли в поколении F2, исследуя гомогенность отклонений от соотношений расщепления для двух популяций F2 реципрокного скрещивания. Проверка на соответствие по критерию хи-квадрат не выявила существенных отклонений от популяций реципрокного скрещивания, что подтверждает отсутствие цитоплазматического наследования (фиг.2). Поскольку цитоплазматическое наследование отсутствует, данные для двух популяций реципрокного скрещивания объединяют и вычисляют общее значение критерия хи-квадрат для собранных данных F2 (фиг.2).
За исключением скрещенной линии Teal × 15A все популяции F2, полученные в результате скрещивания устойчивых и чувствительных линий, характеризуются хорошим соотношением чувствительности растений, равным 3:1, что указывает на расщепление одного главного гена устойчивости к имазамоксу (фиг.2). При скрещивании растений F1 с чувствительной родительской формой образуются популяции BCF1, характеризующиеся хорошим соответствием соотношению устойчивые растения : чувствительные растения, равному 1:1, что подтверждает гипотезу об одном локусе (фиг.2). Данные для популяции F2, полученные в результате скрещивания 15А × Teal, соответствуют соотношению устойчивые растения : чувствительные растения, равному 15:1 (Р=0,08), что указывает на расщепление двух независимых комплементарных генов (фиг.2). Популяция BCF1 характеризуется хорошим соответствием соотношению устойчивые растения : чувствительные растения, равному 3:1, при значении Р критерия хи-квадрат, равном 0,35, что подтверждает результаты, полученные для F2 (фиг.2).
Так как из данных, полученных для F2, следует, что устойчивость в линиях 1А, 9А, 10А, 11А и 16А контролируется одним главным геном, семейства F2:3 должны расщепляться в соответствии с соотношением гомозиготное устойчивое семейство : расщепляющееся семейство : гомозиготное чувствительное семейство, равным 1:2:1. Оценка семейств F2:3 показывает, что скрещенные формы Teal x 1A, Teal x 9A, Teal x 10A, Teal x 11A и Teal x 16A соответствуют соотношению устойчивое семейство : расщепляющееся семейство : чувствительное семейство F2:3, равному 1:2:1, со значениями Р критерия хи-квадрат, равными соответственно 0,64, 0,66, 0,52, 0,40 и 0,94 (фиг.3). Указанные результаты подтверждают полученные для F2 и BCF1 данные о том, что устойчивость в линиях 1А, 10А, 9А, 11А и устойчивость в линии 16А контролируется одним главным геном. Указанный паттерн наследования сопоставим с другими результатами, которые свидетельствуют о генетическом контроле устойчивости к гербицидам, ингибирующим AHAS. В настоящее время почти все мутации в растениях, сообщающие устойчивость к имидазолинонам, показывают, что признак устойчивости контролирует один частично доминантный ген. В Triticum aestivum, Zea Mays, Glycine max, Arabidopsis thaliana и Nicotiana tabacum устойчивость к ингибиторам AHAS наследуется в виде одного частично доминантного ядерного гена (Newhouse et al. 1991; Newhouse et al. 1992; Chaleff and Ray, 1984 Science 223:1148-1151; Sathasivan et al., 1991 Plant Physiol. 97:1044-1050). Устойчивость растений к гербицидам, ингибирующим AHAS, возникает главным образом вследствие одной точковой мутации гена, кодирующего фермент AHAS (Harms et al. 1992, Mol. Gen. Genet. 233:427-435; Winder and Spalding, 1988 Mol. Gen. Genet. 238:394-399).
Данные для F2, полученные в результате скрещивания Teal x 15A, хорошо соответствуют соотношению устойчивые растения : чувствительные растения, равному 15:1, что указывает на расщепление двух независимо расщепляющихся локусов (фиг.2). В подобном случае семейства F2:3 должны расщепляться в соответствии с соотношением устойчивое семейство : расщепляющееся семейство : чувствительное семейство F2:3, равным 7:8:1. Семейства F2:3, полученные в результате скрещивания 15А × Teal, соответствуют ожидаемому соотношению 7:8:1 (фиг.3), подтверждая результаты для популяций F2 и BCF1, свидетельствующие о том, что устойчивость в 15А придается двумя независимыми локусами. Насколько известно авторам изобретения, это первый описанный в научной литературе случай, когда два независимо расщепляющихся аллеля, устойчивых к имидазолинону, идентифицированы в одной линии после мутагенеза семян.
Пример 4
Результаты аллелизма генов IMI
Для определения аллельного взаимодействия генов устойчивости оценивают все возможные интеркроссы между устойчивыми линиями. В популяциях F2, полученных в результате интеркроссов линий SWP965001, 1A, 9A, 10A, 15A и 16А, не обнаружены чувствительные растения (фиг.4). Так как указанные популяции не расщеплялись, гены устойчивости в указанных линиях или являются аллелями в локусе FS-4, или очень близко связаны с ними. Поскольку указанные популяции не расщеплялись в поколении F2, не производилась оценка семейств F2:3, полученных в результате указанных скрещиваний.
Все интеркроссы, включающие линию 11А, расщепляются в поколении F2, что указывает на наличие одного гена устойчивости в 11А (фиг.4). Если два независимо расщепляющихся гена устойчивости присутствуют в результате скрещивания двух линий, каждая из которых имеет один ген устойчивости, в поколении F2 можно ожидать соотношения устойчивые растения : чувствительные растения, равного 15:1. В поколении F2 скрещенные формы SWP965001 × 11A, 1A × 11A, 10A × 11A и 16А × 11А соответствуют ожидаемому соотношению устойчивые растения : чувствительные растения, равному 15:1, что предполагает независимое расщепление двух главных генов устойчивости (фиг.4). Соотношения в семействе F2:3, полученные в результате указанных скрещиваний, также находятся в хорошем соответствии с соотношением устойчивые растения : расщепляющиеся растения : чувствительные растения, что подтверждает результаты, полученные в поколении F2 (фиг.5). В результате скрещивания 11А×9А получают расщепляющуюся популяцию F2, но данное соотношение не соответствует соотношению расщепления, равному 15:1, вследствие большого числа чувствительных расщепляющихся растений. Были исследованы другие гипотезы о двух генах, и все они были признаны в высшей степени значимыми (данные не приведены). Оценки семейств F2:3, полученных в результате указанного скрещивания, однако, хорошо соответствуют соотношению расщепления 7:8:1, что указывает на расщепление двух независимых генов (фиг.5). Полученные результаты подтверждают, что ген устойчивости в 11А уникален по отношению к генам в линиях SWP96001, 1А, 9А, 10А и 16А.
Скрещивание 11А×15А вызывает образование расщепляющейся популяции F2. Так как 15А имеет два гена устойчивости, один из которых аллелен FS-4, то расщепляющаяся популяция F2 в скрещенной форме 11А×15А должна указывать на наличие трех расщепляющихся генов. Расщепляющиеся поколения, полученные в результате скрещивания 15А×11А, испытывали на расщепление трех независимых локусов. Растения F2 соответствуют ожидаемому соотношению устойчивые растения : чувствительные растения, равному 63:1, что свидетельствует о расщеплении трех независимых локусов (фиг.4). Полученные результаты позволяют предположить, что вторая мутация в 15А не является аллельной гену устойчивости в 11А. Семейства F2:3 не исследовали, так как в каждом семействе необходимо было исследовать более 330 растений для гарантии требуемой точности испытания (Hanson, 1959 Agron. J. 51:711-716).
Идентифицированы три независимых локуса устойчивости, каждый из который имеет аллель, определяющий устойчивость к имазамоксу. В научной литературе опубликованы правила, рекомендуемые для обозначения локуса и аллеля гена (McIntosh et al., 1998 Catalogue of Gene Symbols. Volume 5, Proceedings of the 9th International Wheat Genetics Symposium. Saskatoon, Saskatchewan). Локусам неаллельного гена фермента, катализирующим одинаковую реакцию, должен быть присвоен тот же символ, соответствующий обычному названию фермента. Обычным названием AHAS является ALS. При отсутствии данных, необходимых для обозначения локусов для специфических хромосом и геномов, указанные локусы должны быть обозначены в последовательном порядке. Обозначение фенотипа, получаемого при изменении гена с образованием нового аллеля, должно отражать данный фенотип. Таким образом, следует предположить, что аллель устойчивости к имидазолинону FS-4 должен быть обозначен как Imi1 и локус, в котором он находится, должен быть обозначен как Als1. Символ Imi служит для обозначения устойчивости к имидазолинону. Данное обозначение показывает, что указанный ген представляет доминантный признак и является первым идентифицированным аллелем. Данные, полученные для расщепляющихся популяций F2 и F2:3, позволяют предположить, что 15А и 11А имеют два новых независимых аллеля устойчивости в разных локусах (фигуры 2 и 3). Указанные аллели могут быть обозначены Imi2 для мутации 11А в локусе Als2 и Imi3 для второй мутации 15А в локусе Als3.
В данном изобретении идентифицированы три независимо расщепляющихся аллеля, придающих устойчивость к имазамоксу, а именно Imi1 (1А, 9А, 10А, 15А и 16А), Imi2 (11А) и Imi3 (15А). Можно предположить, что каждый из трех идентифицированных аллелей ассоциирован с разным структурным геном, кодирующим устойчивые к гербицидам формы AHAS. Так как пшеница является гексаплоидом, следует ожидать наличия нескольких локусов AHAS. Обнаружены другие полиплоидные виды, имеющие более одной копии AHAS. В Nicotiana tabacum, который является аллотетраплоидом, идентифицированы и исследованы два гена AHAS (Mazur et al. 1987). Чалефф и Рей (Chaleff and Ray, 1984) идентифицировали два независимо расщепляющихся аллеля устойчивости к сульфонилмочевине в Nicotiana tabacum, каждый из который кодирует измененную форму AHAS. Zea mays имеет два структурно экспрессированных идентичных гена AHAS (Fang et al., 1992 Plant Mol. Biol. 18:1185-1187). В аллотетраплоиде Brassica napus и Gossypium hirsutum имеется мультигенное семейство AHAS, состоящее из пяти и шести членов соответственно (Rutledge et al., 1991 Mol Gen. Genet. 229:31-40; Grula et al., 1995 Plant Mol. Biol. 28:837-846). Более высокие уровни устойчивости к гербицидам наблюдаются в полиплоидных видах при наличии множества аллелей устойчивости. Свансон и др. (Swanson et al., 1989 Theor. Appl. Gen. 78:525-530) объединили два уникальных аллеля устойчивости к имидазолинону из двух гомозиготных линий Brassica napus, получив потомство с более высоким уровнем устойчивости по сравнению с каждой гомозиготной линией в отдельности. Кризон и Чалефф (Creason and Chaleff, 1988 Theor. App.; Genet. 76: 177-182) идентифицировали растения Nicotiana tabacum, которые являются гомозиготными для двух мутаций, придающих устойчивость к сульфонилмочевинам. Растения, гомозиготные для обеих мутаций, обладают пятикратной устойчивостью к некорневой обработке хлорсульфуроном по сравнению с растениями, гомозиготными для каждой отдельной мутации. Настоящее изобретение позволяет увеличить уровни устойчивости к имидазолиноновому гербициду у пшеницы путем объединения любых двух или всех трех аллелей устойчивости.
Пример 5
Устойчивость к имидазолиноновым гербицидам у Teal11A, Teal15A и гибрида Teal11A/15A
Повышенная устойчивость Teal 11A и Teal 15A при внесении 20 г/га имазамокса была продемонстрирована в предыдущих примерах в результате выполнения исследований наследования, позволяющих отличать устойчивые растения от чувствительных родительских и расщепляющихся растений. Установлено, что Teal 11A придает уровни устойчивости к имидазолиноновым гербицидам, аналогичные уровням, придаваемым мутацией FS4 в сорте Fidel, при сравнении, производимом в теплице и в полевых условиях. Сходство устойчивости обнаружено также при сравнении in vitro активности AHAS, экстрагированной из устойчивых растений. Такая ситуация вполне возможна, так как устойчивость Teal 11A, Teal 15A и FS4 обусловлена мутацией в ферменте AHAS, делающей его устойчивым к подавлению имидазолиноновыми гербицидами. На фиг.6 показано, что активность фермента AHAS, экстрагированного из Teal 11A и BW755, линии, содержащей FS4, одинаково изменяется при увеличении нормы внесения имазамокса, при этом оба сорта характеризуются более высоким процентным содержанием активного (устойчивого) фермента при самой высокой концентрации имазамокса по сравнению с контрольным видом Teal дикого типа.
Наличие двух нуклеиновых кислот IMI в Teal 15A обеспечивает повышенную устойчивость к имидазолиноновым гербицидам по сравнению с такой линией, как BW755, имеющей только одну нуклеиновую кислоту IMI. Повышенная устойчивость находит отражение в меньшем повреждении растений при более высоких нормах внесения гербицида и наличии более высокой активности неингибированного фермента AHAS. На фиг.7 показано, что при внесении 10-кратного количества имазамокса (200 г/га) все обработанные растения с одним геном были повреждены, в то время как растения с двумя генами не были повреждены. При выполнении анализа активности AHAS in vitro при всех концентрациях имазамокса (фиг.6) и особенно при самых высоких концентрациях сорт Teal15A характеризовался более высоким процентным содержанием активного (устойчивого) фермента по сравнению с линиями Teal11A и BW755 с одним геном.
При объединении трех неаллельных генов, каждый из которых придает устойчивость к имидазолиноновым гербицидам, достигается более высокая устойчивость, чем при наличии только двух неаллельных генов (фиг.7). При 30-кратной норме внесения гербицида, соответствующей 600 г/га имазамокса, более половины растений во все еще расщепляющейся самоопыляемой популяции Teal15A, скрещенной с Teal11A, не были повреждены, в то время как все растения гомозиготной популяции Teal15A были повреждены.







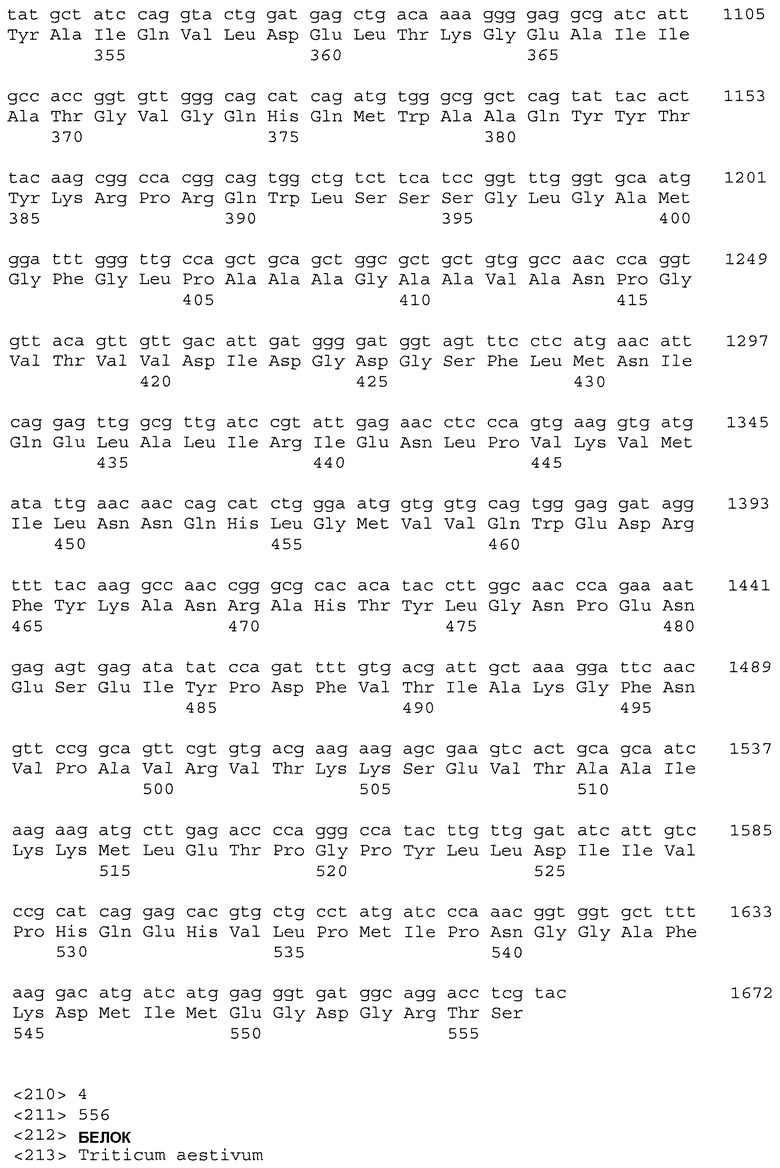
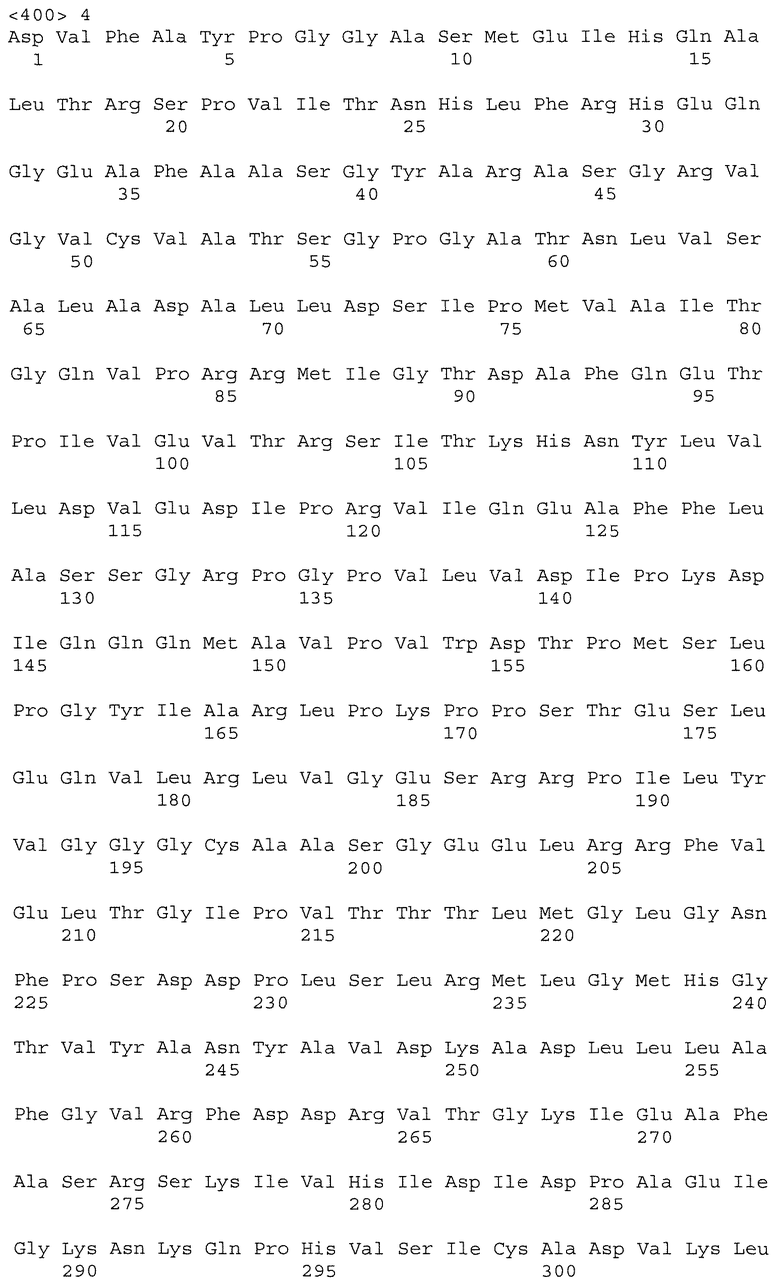






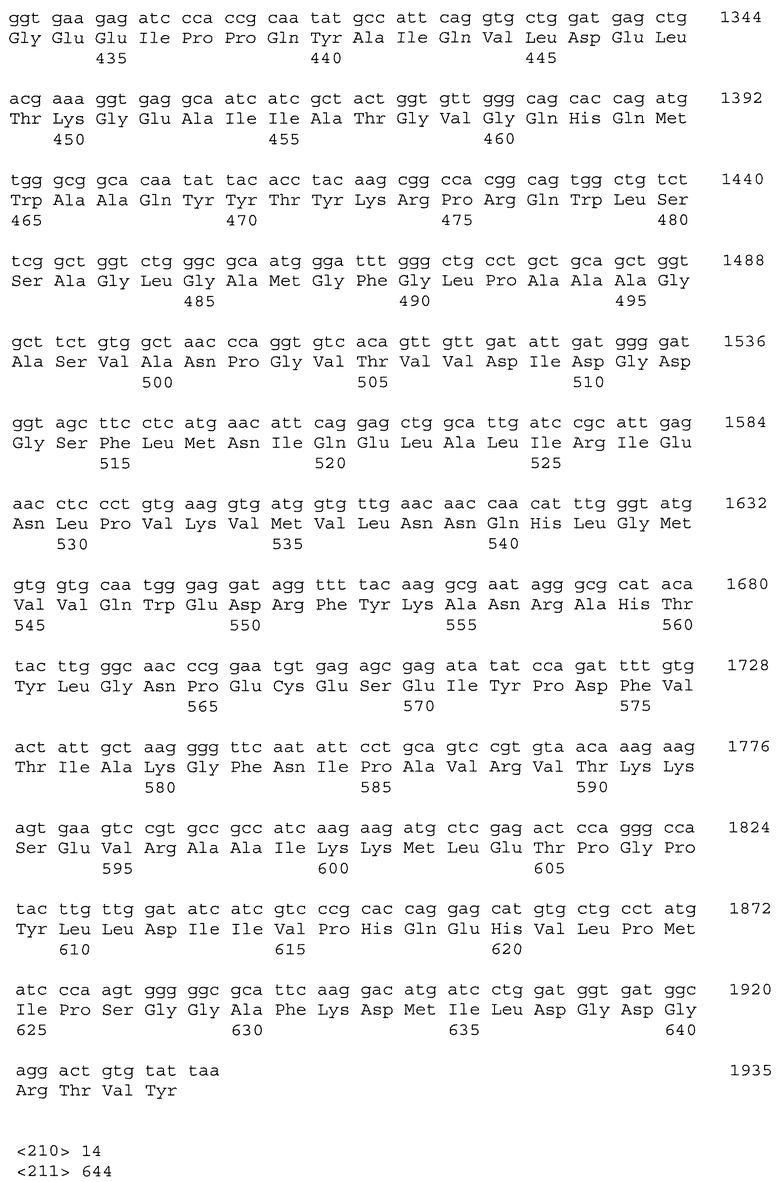




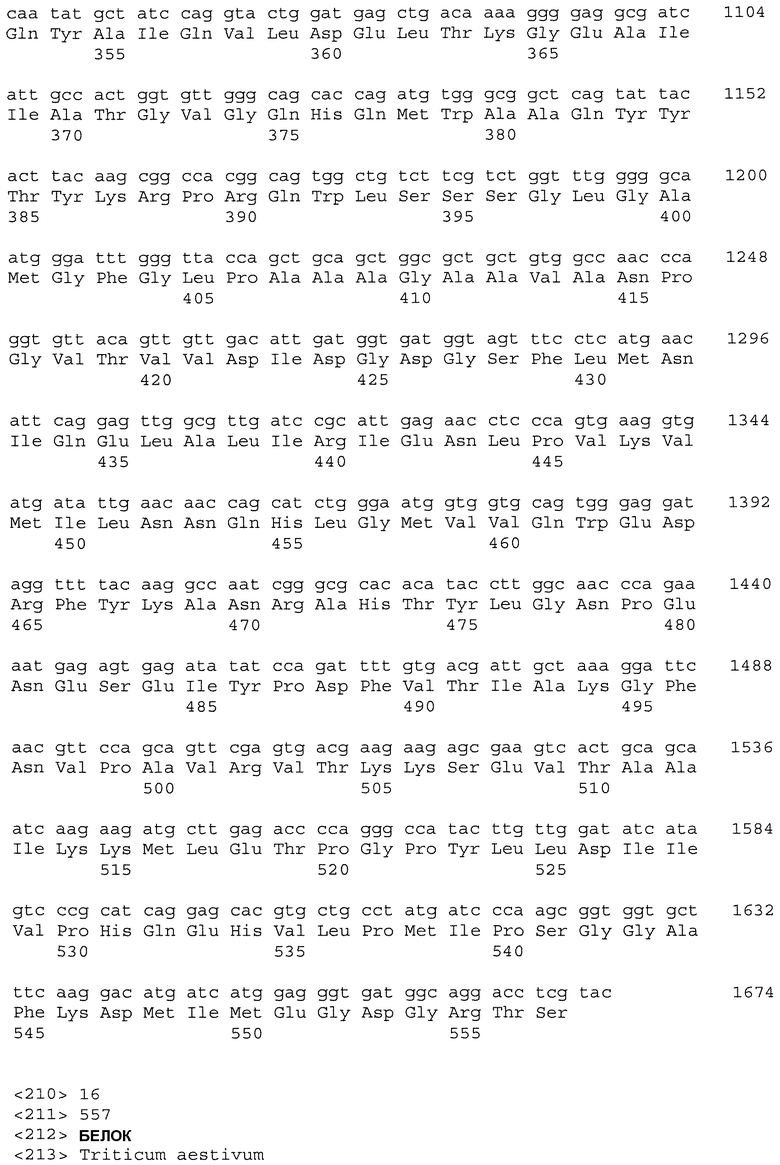







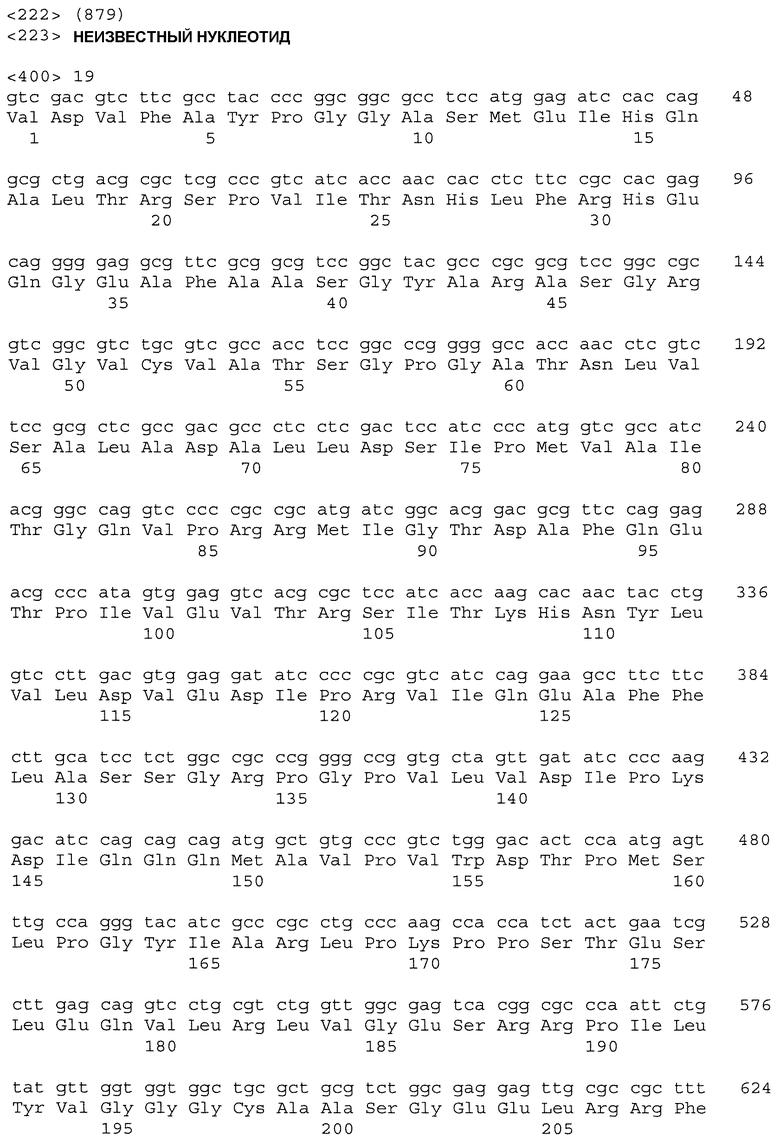





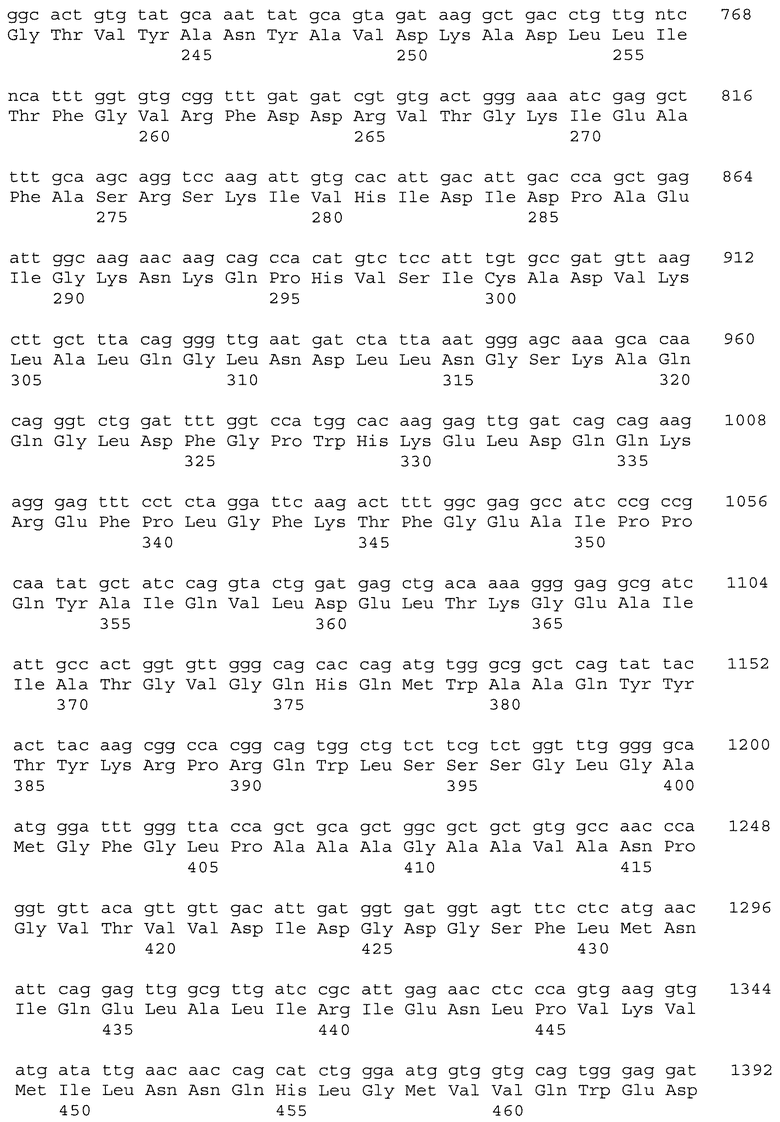



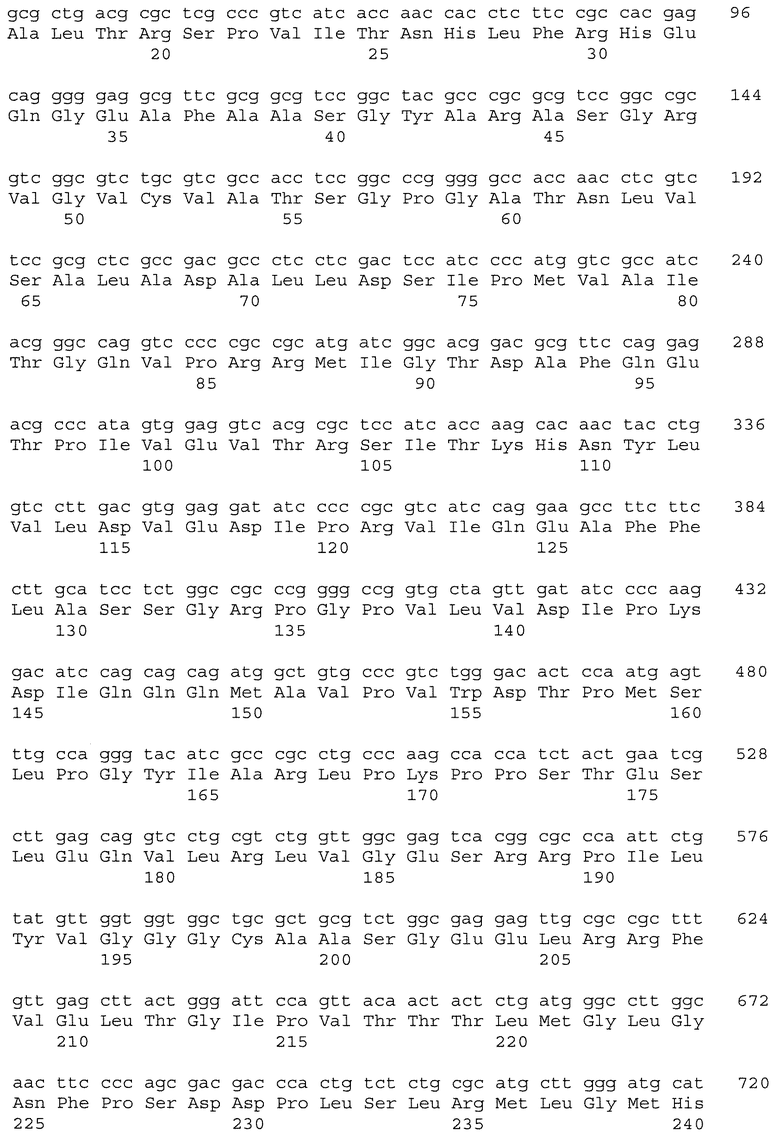


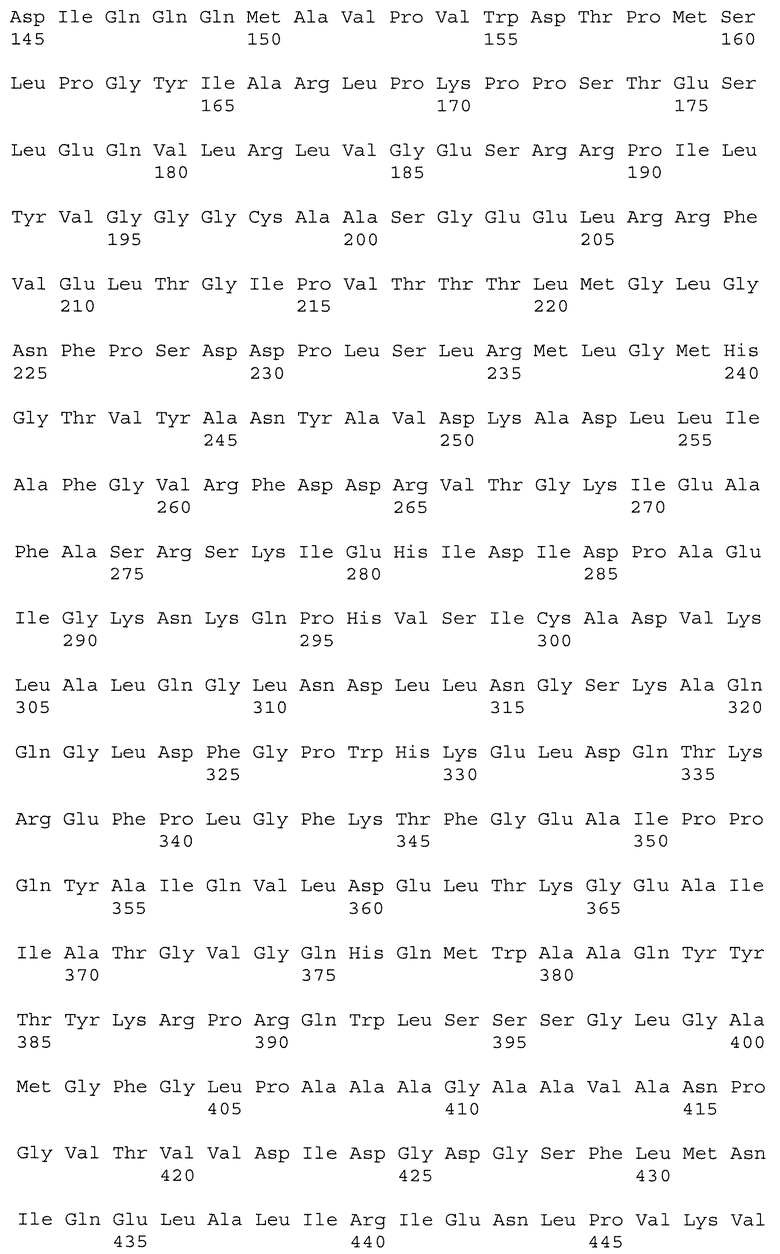
























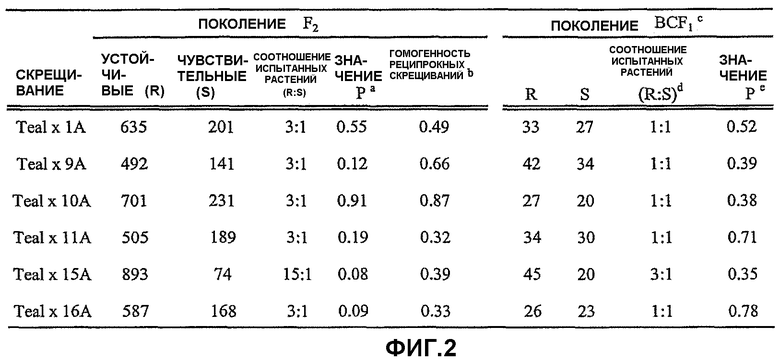
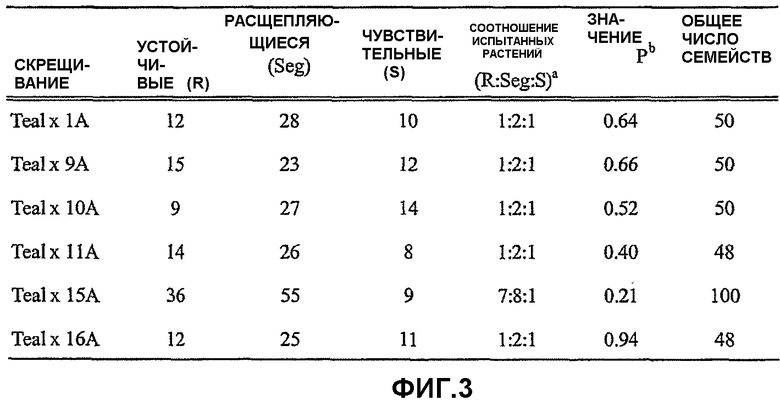
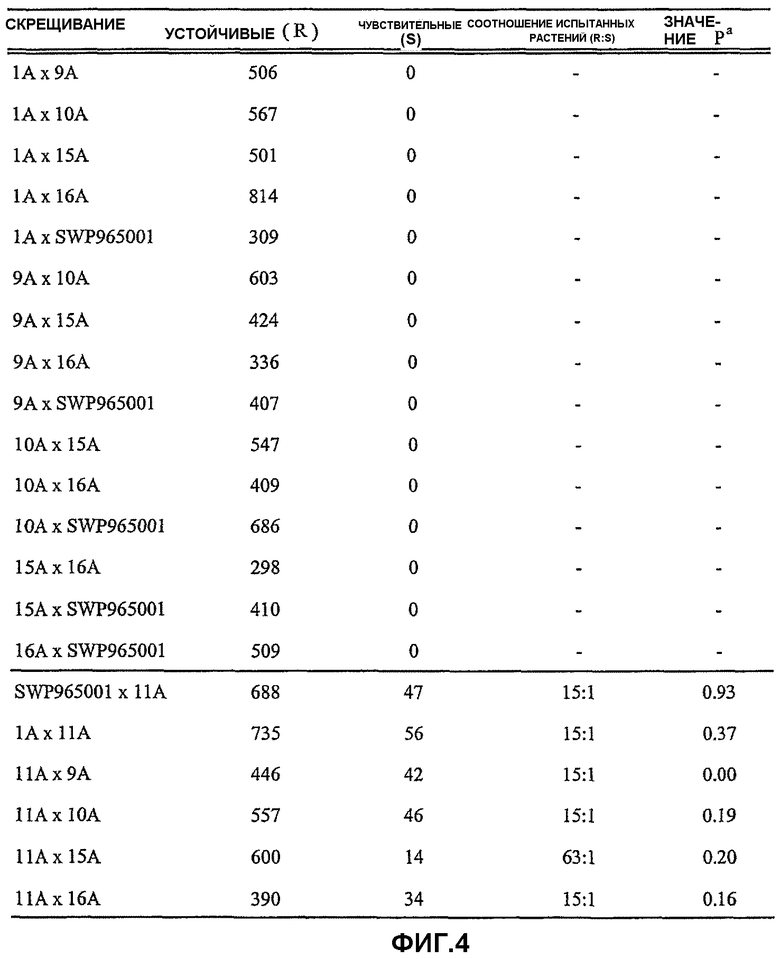

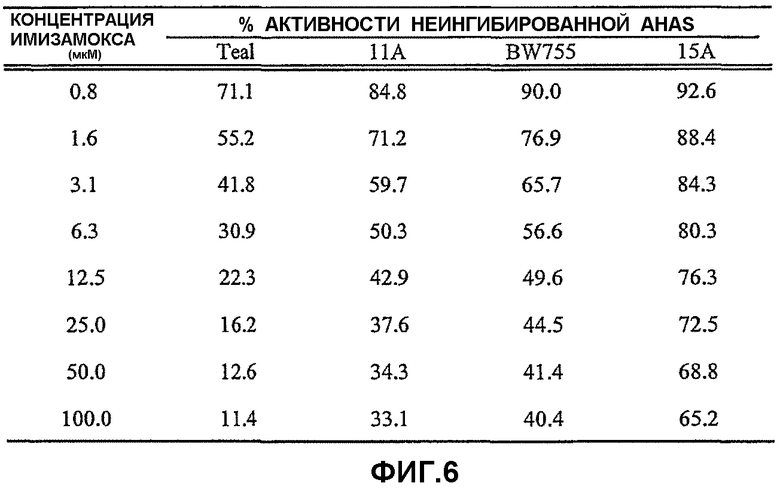





























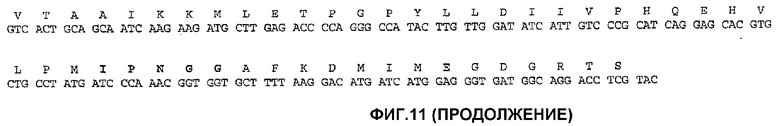






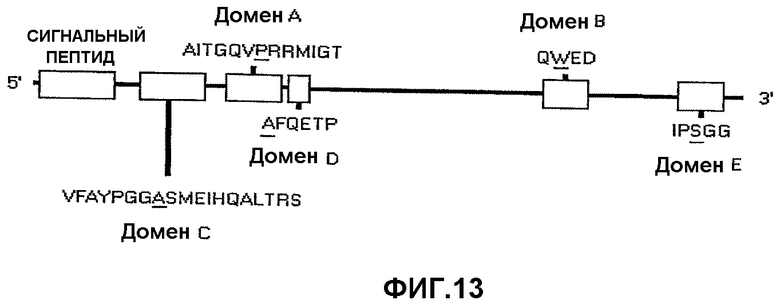
Семена растения обрабатывают этил метансульфонатом в течение 6 часов, затем просушивают или проращивают. Полученные растения M1 самоопыляют, получают семена, из которых выращивают растения пшеницы М2. Растения М2 обрабатывают имазамоксом и отбирают обладающие повышенной устойчивостью к нему по сравнению с растениями дикого типа. В другом варианте растения трансформируют экспрессирующим вектором, содержащим одну или более нуклеиновых кислот IMI. Указанные нуклеиновые кислоты предпочтительно находятся в разных геномах или получены из разных геномов. Полученные растения обладают повышенной устойчивостью к имидазолинону и могут быть использованы в условиях борьбы с сорняками посредством гербицида. 18 н. и 29 з.п. ф-лы, 13 ил.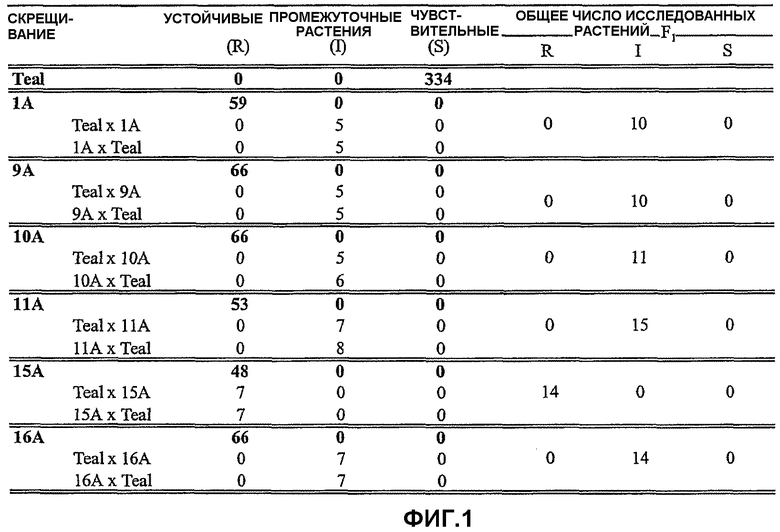
a) полинуклеотида, содержащего SEQ ID NO:1;
b) полинуклеотида, содержащего SEQ ID NO:3;
c) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:2;
d) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:4;
e) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:2, где в домене Е полипептида серин заменен на аспарагин; и
f) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:4, где в домене Е полипептида серин заменен на аспарагин.
или растение представляет собой потомка, по меньшей мере, одного растения, выбранного из группы, состоящей из растения с идентификационным номером патентного депозитария АТСС РТА-3954 и растения с идентификационным номером патентного депозитария РТА-3953, где указанный потомок содержит, по меньшей мере, одну указанную нуклеиновую кислоту IMI.
a) предварительное вымачивание семян в дистиллированной воде в течение четырех часов;
b) обработку семян пшеницы 0,3%-ным EMS (этил метансульфонат) в течение шести часов;
c) последовательную промывку пшеницы водопроводной водой в течение семи часов;
d) просушку семян пшеницы в течение приблизительно четырех часов перед посевом;
e) прорастание растений пшеницы M1 из семени пшеницы;
f) самоопыление растений пшеницы M1 с получением семян пшеницы М2;
g) посев семян М2 с прорастанием растений пшеницы М2 с получением из них двупольной стадии;
h) распыление на растения пшеницы имазамокса в количестве 40 г а.и./га; и
i) отбор растений пшеницы М2 с повышенной устойчивостью к имазамоксу по сравнению с разновидностью растения дикого типа.
а) полинуклеотида, содержащего SEQ ID NO:1;
b) полинуклеотида, содержащего SEQ ID NO:3;
c) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:2;
d) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:4;
e) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:2, где в домене Е полипептида серин заменен на аспарагин; и
f) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:4, где в домене Е полипептида серин заменен на аспарагин;
g) полинуклеотида, комплементарного полинуклеотиду по любому из а)-f).
a) полинуклеотида, содержащего SEQ ID NO:1;
b) полинуклеотида, содержащего SEQ ID NO:3;
c) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:2;
d) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:4;
e) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:2, где в домене Е полипептида серии заменен на аспарагин; и
f) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:4, где в домене Е полипептида серии заменен на аспарагин.
а) трансформацию растительной клетки одним или несколькими экспрессирующими векторами, содержащими, по крайней мере, одну нуклеиновую кислоту IMI, где нуклеиновая кислота выбрана из группы, состоящей из
i) полинуклеотида, содержащего SEQ ID NO:1;
ii) полинуклеотида, содержащего SEQ ID NO:3;
iii) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:2;
iv) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:4;
v) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:2, где в домене Е полипептида серин заменен на аспарагин; и
vi) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:4, где в домене Е полипептида серии заменен на аспарагин,
b) получение из растительной клетки трансгенного растения с повышенной устойчивостью к имидазолиноновому гербициду по сравнению с разновидностью растения дикого типа.
a) кроссбридинг первого растения пшеницы со вторым растением пшеницы, где первое растение пшеницы обладает повышенной устойчивостью к имидазолиноновому гербициду по сравнению с разновидностью растения пшеницы дикого типа;
b) получение потомства растений пшеницы от кроссбридинга; и
c) обработку потомства растений пшеницы имидазолиноновым гербицидом;
где первое растение пшеницы представляет собой растение пшеницы, содержащее нуклеиновую кислоту IMI, выбранную из группы, состоящей из
i) полинуклеотида, содержащего SEQ ID NO:1;
ii) полинуклеотида, содержащего SEQ ID NO:3;
iii) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:2;
iv) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:4;
v) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:2, где в домене Е полипептида серии заменен на аспарагин; и
vi) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:4, где в домене Е полипептида серии заменен на аспарагин.
a) кроссбридинг первого растения пшеницы со вторым растением пшеницы, где первое растение пшеницы обладает повышенной устойчивостью к имидазолиноновому гербициду по сравнению с разновидностью растения пшеницы дикого типа;
b) получение потомства растений пшеницы от кроссбридинга; и
c) обработку потомства растений пшеницы имидазолиноновым гербицидом;
где первое растение пшеницы представляет собой растение пшеницы по п.21.
i) полинуклеотида, содержащего SEQ ID NO:1;
ii) полинуклеотида, содержащего SEQ ID NO:3;
iii) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:2;
iv) полинуклеотида, кодирующего полипептид, содержащий SEQ ID NO:4;
v) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:2, где в домене Е полипептида серии заменен на аспарагин; и
vi) полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, которая, по меньшей мере, на 90% идентична SEQ ID NO:4, где в домене Е полипептида серин заменен на аспарагин.
| Устройство для определения параметров дыхания | 1972 |
|
SU508161A3 |
| PLANT PHYSIOLOGY, vol.100, no.2, 1992, p.882-886 | |||
| US 5928937 A, 27.07.1999 | |||
| ЕР 0525384 A, 03.02.1993 | |||
| СПОСОБ ПОЛУЧЕНИЯ МУТАНТНОЙ ZEA MAYS | 1990 |
|
RU2128426C1 |

