Область техники, к которой относится изобретение
Настоящее изобретение относится в целом к растениям, обладающим повышенной толерантностью к имидазолиноновым гербицидам. Более конкретно настоящее изобретение относится к растениям пшеницы, полученным путем мутагенеза, и кроссбридинга, и трансформации, которые обладают повышенной толерантностью к имидазолиноновым гербицидам.
Предпосылки создания изобретения
Синтаза ацетогидроксикислот (AHAS; КФ 4.1.3.18, ацетолактатасинтаза (ALS)), кодируемая нуклеиновой кислотой Als, представляет собой первый фермент, который катализирует биохимический синтез аминокислот с разветвленными цепями, таких как валин, лейцин и изолейцин (Singh В.К. “Biosynthesis of valine, leucine и isoleucine” в: Plant amino acids, под ред. Singh B.K., изд-во Marcel Dekker Inc. New York, New York, 1999, cc.227-247). AHAS является мишенью действия четырех структурно различных семейств гербицидов, таких как сульфонилмочевины (LaRossa R.А. и Faico S.С., Trends Biotechnol 2, 1984, cc.158-161), имидазолиноны (Shaner и др., Plant Physiol 76, 1984, cc.545-546), триазолопиримидины (Subramanian и Gerwick, Inhibition of acetolactate synthase by triazolopyrimidines в: Biocatalysis in agricultural biotechnology. ACS Symposium Series, под ред. Whitaker J.R., Sonnet P.E., изд-во American Chemical Sodety. Washington, D.C., 1989, cc.277-288) и пиримидилоксибензоаты (Subramanian и др., Plant Physiol 94, 1990, cc.239-244). Гербициды из семейства имидазолинонов и сульфонилмочевин широко используют в современном сельском хозяйстве благодаря их эффективности в очень небольших нормах расхода и относительно низкой токсичности для животных. Путем ингибирования активности AHAS представители этих семейств гербицидов препятствуют дальнейшему росту и развитию чувствительных к ним растений, включая многие виды сорняков. В качестве некоторых примеров поступающих в продажу имидазолиноновых гербицидов можно привести PURSUIT® (имизетапир), SCEPTER® (имазахин) и ARSENAL® (имазапир). Примерами гербицидов из семейства сульфонилмочевин являются хлорсульфурон, метсульфурон-метил, сульфурон-метил, хлоримурон-этил, трифенсульфурон-метил, трибенурон-метил, бенсульфурон-метил, никосульфурон, этаметсульфурон-метил, римсульфурон, трифлусульфурон-метил, триасульфурон, примисульфурон-метил, циносульфурон, амидосульфурон, флузасульфурон, имазосульфурон, пиразосульфурон-этил и галосульфурон.
Благодаря их высокой эффективности и низкой токсичности, имидазолиноновые гербициды являются предпочтительными для опрыскивания верхних частей растений на большой площади их произрастания. Возможность осуществлять гербицидом опрыскивание верхних частей растений на большой площади их произрастания снижает стоимость, связанную с созданием и поддержанием плантаций, и снижает необходимость в подготовке мест произрастания перед обработкой такими химическими средствами защиты растений. Опрыскивание верхних частей требуемых толерантных видов приводит также к возможности достижения максимального потенциального урожая требуемых видов из-за отсутствия видов-конкурентов. Однако возможность применения таких методов опрыскивания верхних частей растения зависит от присутствия толерантных к имидазолинону видов требуемой растительности в области проведения обработок.
Из основных сельскохозяйственных культур некоторые виды бобовых, такие как соя, обладают природной толерантностью к имидазолиноновыми гербицидам благодаря их способности быстро метаболизировать гербициды (Shaner и Robson, Weed Sci. 33, 1985, cc.469-471). Другие культурные растения, такие как кукуруза (Newhouse и др. Plant Physiol. 100, 1992, cc.882-886) и рис (Barrett и др., Crop Safeners for Herbicides, изд-во Academic Press, New York, 1989, cc.195-220), чувствительны к действию имидазолиноновых гербицидов. Различный уровень чувствительности к имидазолиноновым гербицидам зависит от химической природы конкретного гербицида и различного метаболизма конкретного соединения в каждом растении, приводящего к превращению токсичной формы в нетоксичную (Shaner и др., Plant Physiol. 76, 1984, cc.545-546; Brown и др., Pestic. Biochm. Physiol, 27, 1987, cc.24-29). Чувствительность зависит также в большей степени от других физиологических различий растений, таких как абсорбция и транслокация (Shaner и Robson, Weed Sci. 33, 1985, cc.469-471).
Сорта культурных растений, обладающие толерантностью к имидазолинонам, сульфонилмочевинам и триазолопиримидинам, были успешно получены с использованием мутагенеза семени, микроспоры, пыльцы и каллюса таких растений, как Zea mays, Brassica napus, Glycine max и Nicotiana tabacum (Sebastian и др., Crop Sci 29, 1989, cc.1403-1408; Swanson и др., Theor. Appl. Genet 78, 1989, cc.525-530; Newhouse и др., Theor. Appl. Genet 83, 1991, cc.65-70; Sathasivan и др., Plant Physiol. 97, 1991, cc.1044-1050; Mourand и др., J. Heredity 84, 1993, cc.91-96). Во всех случаях толерантность была обусловлена индивидуальным частично доминантным ядерным геном. Ранее с помощью мутагенеза семян были получены также толерантные к имидазолинонам четыре линии растений пшеницы Triticum aestivum L. cv Rdel (Newhouse и др., Plant Physiol. 100, 1992, cc.882-886). Опыты по оценке особенностей наследования подтвердили, что толерантность обусловлена индивидуальным частично доминантным геном. Основываясь на изучении аллелей, авторы сделали заключение о том, что мутации в четырех идентифицированных линиях локализованы в одном и том же локусе. Один из генов, обусловливающих толерантность культивара Fidel, был обозначен как FS-4 (Newhouse и др., Plant Physiol. 100, 1992, cc.882-886).
Компьютерное моделирование трехмерной конформации комплекса AHAS-ингибитор позволило предсказать несколько аминокислот в предполагаемом связывающимся с ингибитором “кармане” в качестве сайтов, в которых индуцированные мутации, по-видимому, обусловливали избирательную толерантность к имидазолинонам (Ott и др., J. Mol. Biol. 263, 1996, cc.359-368). Действительно растения табака, полученные с использованием некоторых таких преднамеренно созданных мутаций в предполагаемых сайтах связывания фермента AHAS, обладали специфической толерантностью к одному классу гербицидов (Ott и др., J. Mol. Biol. 263, 1996, cc.359-368).
Толерантность растений к имидазолиноновым гербицидам описана также во многих патентах. В US 4761373, 5331107, 5304732, 6211438, 6211439 и 6222100 описано в целом применение измененных нуклеиновых кислот Als для создания толерантности к гербицидам у растений и, в частности, описаны толерантные к некоторым имидазолинонам линии кукурузы.
В US 5013659 описаны растения, имеющие обусловливающие толерантность к гербицидам мутации, которые затрагивают по меньшей мере одну аминокислоту в одной или нескольких консервативных областях. Описанные мутации кодируют либо перекрестную толерантность к имидазолинонами и сульфонилмочевинам, либо специфическую толерантность к сульфонилмочевинам, однако специфическая толерантность к имидазолинонам к настоящему времени не описана. Кроме того, в US 5731180 и US 5767361 описан выделенный ген, кодирующий одну аминокислотную замену в аминокислотной последовательности AHAS дикого типа однодольных растений, которая обусловливает специфическую толерантность к имидазолинонам.
В известном к настоящему времени уровне техники не описаны толерантные к имидазолинонам растения пшеницы Triticum turgidum или толерантные к имидазолинонам растения тритикале. К настоящему времени не описаны также толерантные к имидазолинонам растения, содержащие по меньшей мере одну измененную нуклеиновую кислоту Als Triticum turgidum. He описаны также толерантные к имидазолинонам растения пшеницы, несущие мутации в геномах, отличных от генома, из которого выведен ген FS-4. Таким образом, в данной области техники сохраняется необходимость в идентификации генов, обусловливающих толерантность к имидазолинонам, из других геномов и видов. В данной области техники существует также необходимость в создании растений пшеницы и растений тритикале, обладающих повышенной толерантностью к гербицидам, таким как имидазолинон, и содержащим по меньшей мере одну измененную нуклеиновую кислоту Als. Требуются также методы борьбы с сорняками, произрастающими вблизи таких растений пшеницы или растений тритикале. Эти композиции и способы должны позволять осуществлять опрыскивание верхних частей растений в качестве метода применения гербицидов в местах произрастания растений пшеницы или растений тритикале.
Краткое изложение сущности изобретения
В настоящем изобретении предложены растения пшеницы, несущие нуклеиновые кислоты IMI, где растение пшеницы обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа. Растения пшеницы могут содержать один, два, три или большее количество IMI-аллелей. Согласно одному варианту осуществления изобретения растение пшеницы содержит по меньшей мере одну нуклеиновую кислоту IMI. Согласно другому варианту осуществления изобретения нуклеиновую кислоту IMI выбирают из группы, включающей нуклеиновую кислоту Imi 1, нуклеиновую кислоту Imi 2 и нуклеиновую кислоту Imi 3. Согласно следующему варианту осуществления изобретения по меньшей мере одна нуклеиновая кислота IMI представляет собой нуклеиновую кислоту IMI Triticum turgidum. Согласно еще одному варианту осуществления изобретения по меньшей мере одна нуклеиновая кислота IMI представляет собой нуклеиновую кислоту IMI подвидов дурум (пшеница твердая, пшеница класса II по стандартам США). Согласно другому варианту осуществления изобретения растение пшеницы содержит несколько нуклеиновых кислот IMI, локализованных в различных геномах. Согласно следующему варианту осуществления изобретения несколько нуклеиновых кислот IMI содержат нуклеиновую кислоту Imi 2 Triticum turgidum и нуклеиновую кислоту Imi 3 Triticum turgidum. Согласно другому варианту осуществления изобретения несколько нуклеиновых кислот IMI содержат нуклеиновую кислоту Imi 2 подвидов дурум и нуклеиновую кислоту Imi 3 подвидов дурум. Предпочтительно нуклеиновые кислоты IMI кодируют белки, содержащие мутацию в консервативной аминокислотной последовательности, выбранной из группы, включающей домен А, домен В, домен С, домен D и домен Е. Более предпочтительно мутация находится в консервативном домене Е. Изобретение относится также к частям растений и семенам растений, полученным из представленных в описании растений пшеницы.
Настоящее изобретение относится также к растениям тритикале, содержащим нуклеиновые кислоты IMI, где растения тритикале обладают повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растений тритикале дикого типа. Согласно одному из вариантов осуществления изобретения растение тритикале содержит по меньшей мере одну нуклеиновую кислоту IMI. Согласно другому варианту осуществления изобретения по меньшей мере одну нуклеиновую кислоту IMI выбирают из группы, включающей нуклеиновую кислоту Imi 1, нуклеиновую кислоту Imi 2 и нуклеиновую кислоту Imi 3. Согласно следующему варианту осуществления изобретения по меньшей мере одна нуклеиновая кислота IMI представляет собой нуклеиновую кислоту IMI Triticum turgidum. Согласно еще одному варианту осуществления изобретения по меньшей мере одна нуклеиновая кислота IMI представляет собой нуклеиновую кислоту IMI подвидов дурум. Согласно другому варианту осуществления изобретения растение тритикале содержит несколько нуклеиновых кислот IMI, локализованных в различных геномах. Согласно следующему варианту осуществления изобретения несколько нуклеиновых кислот IMI содержат нуклеиновую кислоту Imi 2 Triticum turgidum и нуклеиновую кислоту Imi 3 Triticum turgidum. Согласно другому варианту осуществления изобретения несколько нуклеиновых кислот IMI содержат нуклеиновую кислоту Imi 2 подвидов дурум и нуклеиновую кислоту Imi 3 подвидов дурум. Согласно следующему варианту осуществления изобретения нуклеиновые кислоты IMI кодируют белки, содержащие мутацию в консервативной аминокислотной последовательности, выбранной из группы, включающей домен А, домен В, домен С, домен D и домен Е. Изобретение относится также к частям растений и семенам растений, полученным из представленных в описании растений тритикале.
Нуклеиновые кислоты IMI, предлагаемые в настоящем изобретении, могут содержать нуклеотидную последовательность, выбранную из группы, включающей: полинуклеотид, который имеет последовательность SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5. или SEQ ID NO:23; полинуклеотид, который кодирует полипептид, имеющий последовательность SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:24; полинуклеотид, содержащий по меньшей мере 60 последовательных нуклеотидов любого из указанных выше полинуклеотидов; и полинуклеотид, комплементарный любому из указанных выше полинуклеотидов.
Растения, предлагаемые в настоящем изобретении, могут быть трансгенными или нетрансгенными. Примеры нетрансгенных растений пшеницы, обладающих повышенной толерантностью к имидазолиноновым гербицидам, включают растения пшеницы, которые депонированы в АТСС (Американская коллекция типовых культур) под регистрационным номером для цели патентования РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4916, РТА-4917, РТА-4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922, РТА-4923 или РТА-4960; или мутант, рекомбинант или созданное с помощью генной инженерии производное растения, депонированного в АТСС под регистрационным номером для цели патентования РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4916, РТА-4917, РТА-4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922, РТА-4923 или РТА-4960; или любое потомство растения, депонированного в АТСС под регистрационным номером для цели патентования РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4916, РТА-4917, РТА-4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922, РТА-4923 или РТА-4960; или растение, являющееся потомком любого из указанных растений.
Помимо композиций, предлагаемых в настоящем изобретении, в изобретении предложено также несколько способов. Предлагаемые в изобретении способы относятся к способам модификации толерантности растений к имидазолиноновым гербицидам, которые заключаются в том, что модифицируют экспрессию нуклеиновой кислоты IMI в растении. Описаны также способы получения трансгенного растения, обладающего повышенной толерантностью к имидазолиноновому гербициду, которые заключаются в том, что трансформируют растительную клетку экспрессионным вектором, содержащим одну или несколько нуклеиновых кислот IMI, и получают растение из растительной клетки. Изобретение относится также к способу борьбы с сорняками вблизи растения, заключающемуся в том, что имидазолиноновый гербицид наносят на сорняки и на растение, где растение обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с популяцией растений дикого типа и где растение содержит одну или несколько нуклеиновых кислот IMI. В некоторых предпочтительных вариантах этих способов растения содержат несколько нуклеиновых кислот IMI, локализованных в различных геномах пшеницы.
Краткое описание чертежей
На чертежах показано:
на фиг.1 - сравнительный анализ последовательности ДНК гена Als 2, амплифицированного из геномной ДНК пшеницы подвида дурум сорта Cicco (SEQ ID NO:11), гена Als 2, амплифицированного из геномной ДНК пшеницы подвида дурум сорта Colosseo (SEQ ID NO:14), гена Als 2, амплифицированного из геномной ДНК пшеницы подвида дурум сорта Utopia (SEQ ID NO:16) и консенсусной последовательности гена Als 2 пшеницы подвида дурум (SEQ ID NO:19). Среди сортов не обнаружено полиморфизма;
на фиг.2 - сравнительный анализ последовательности ДНК гена Als 3, амплифицированного из геномной ДНК пшеницы подвида дурум сорта Ciccio (SEQ ID NO:13), гена Als 3, амплифицированного из геномной ДНК пшеницы подвида дурум сорта Colosseo (SEQ ID NO:15), гена Als 3, амплифицированного из геномной ДНК пшеницы подвида дурум сорта Utopia (SEQ ID NO:17) и консенсусной последовательности гена Als 3 пшеницы подвида дурум (SEQ ID NO:21). Среди сортов не обнаружено полиморфизма;
на фиг.3 - сравнительный анализ последовательности ДНК гена Als 2, амплифицированного из геномной ДНК пшеницы подвида дурум сорта Cicco (SEQ ID NO:11), гена Als 2, амплифицированного из геномной ДНК толерантной к имидазолинону линии СI19 (SEQ ID NO:1), гена Als 2, амплифицированного из геномной ДНК толерантной к имидазолинону линии UT15 (SEQ ID NO:7), гена Als 2, амплифицированного из геномной ДНК толерантной к имидазолинону линии UT19 (SEQ ID NO:9) и консенсусной последовательности гена Als 2 пшеницы подвида дурум (SEQ ID NO:19). Выявленный нуклеотидный полиморфизм, обусловливающий толерантность к имидазолинону линии СI19, обозначен жирным шрифтом;
на фиг.4 - сравнительный анализ выведенной аминокислотной последовательности белка, кодируемого геном Als 2 из сорта Cicco (SEQ ID NO:12), выведенной аминокислотной последовательности белка, кодируемого геном Als 2 из толерантной к имидазолинону линии СI19 (SEQ ID NO:2), выведенной аминокислотной последовательности белка, кодируемого геном Als 2 из толерантной к имидазолинону линии UT15 (SEQ ID NO:8), выведенной аминокислотной последовательности белка, кодируемого геном Als 2 из толерантной к имидазолинону линии UT19 (SEQ ID NO:10) и консенсусной последовательности Als 2 пшеницы подвида дурум (SEQ ID NO:20). Полиморфизм, обусловливающий толерантность к имидазолинону линии СI19, обозначен жирным шрифтом;
на фиг.5 - сравнительный анализ последовательности ДНК гена Als 3, амплифицированного из геномной ДНК сорта Utopia (SEQ ID NO:17), частичной полинуклеотидной последовательности Als 3, амплифицированной из геномной ДНК толерантной к имидазолинону линии UT12 (SEQ ID NO:3), гена Als 3, амплифицированного из геномной ДНК толерантной к имидазолинону линии UT15 (SEQ ID NO:5), гена Als 3, амплифицированного из геномной ДНК толерантной к имидазолинону линии UT19 (SEQ ID NO:23), и консенсусной последовательности гена Als 3 пшеницы подвида дурум (SEQ ID NO:21). Нуклеотидные полиморфизмы, обусловливающие толерантность линий к имидазолинону, обозначены жирным шрифтом;
на фиг.6 - сравнительный анализ выведенной аминокислотной последовательности белка, кодируемого геном Als 3 из сорта Utopia (SEQ ID NO:18), выведенной аминокислотной последовательности полипептида, кодируемого частичной полинуклеотидной последовательностью Als 3 из толерантной к имидазолинону линии UT12 (SEQ ID NO:4), выведенной аминокислотной последовательности белка, кодируемого геном Als 3 из толерантной к имидазолинону линии UT15 (SEQ ID NO:6), выведенной аминокислотной последовательности белка, кодируемого геном Als 3 из толерантной к имидазолинону линии UT19 (SEQ ID NO:24) и консенсусной последовательности Als 3 пшеницы подвида дурум (SEQ ID NO:22). Нуклеотидный полиморфизм, обусловливающий толерантность линии UT12 к имидазолинону, обозначен жирным шрифтом;
на фиг.7 - схематическое изображение консервативных аминокислотных последовательностей, кодируемых генами AHAS, которые участвуют в придании толерантности к различным ингибиторам AHAS. Специфические аминокислотные сайты, ответственные за толерантность, подчеркнуты (использованы с изменениями данные, полученные Devine M.D. и Eberiein С.V. “Physiological, biochemical и molecular aspects of herbicide tolerance based on altered target sites” в: Herbicid Activity: Toxicity, Biochemistry, and Molecular Biology, изд-во IOS Press, Амстердам, 1997, cc.159-185).
Подробное описание изобретения
Настоящее изобретение относится к растениям пшеницы, частям растений пшеницы и клеткам растений пшеницы, обладающим повышенной толерантностью к имидазолиноновым гербицидам. Под объем настоящего изобретения подпадают также семена, полученные на указанных растениях пшеницы, и способы борьбы с сорняками вблизи растений пшеницы, указанных в описании. Следует понимать, что в описании и формуле изобретения применение существительного в единственном числе может подразумевать также его применение во множественном числе, в зависимости от контекста, в котором оно используется. Например, при ссылке на “клетку” следует понимать, что можно использовать по меньшей мере одну клетку.
В контексте настоящего описания понятие “растение пшеницы” относится к растению, которое является представителем рода Triticum. Растения пшеницы, предлагаемые в настоящем изобретении, могут быть представителями рода Triticum, который включает (но не ограничиваясь ими), Т. aestivum, Т. turgidum, Т. timopheevii, Т. monococcum, Т. zhukovskyi и Т. urartu, а также их гибриды. Примерами подвидов Т.aestivum, подпадающими под объем настоящего изобретения, являются aestivum (пшеница обыкновенная), compactum (пшеница карликовая), macha (пшеница Маха), vavilovi (пшеница Вавилова), spelta (пшеница спельта) и sphaecrococcum (пшеница короткая). Примерами подвидов Т. turgidum, подпадающими под объем настоящего изобретения, являются turgidum, carthlicum, dicoccom, durum, paleocolchicum, polonicum, turanicum и dicoccoides. Примерами подвидов Т. monococcum subspedes, подпадающими под объем настоящего изобретения, являются monococcum (полба, пшеница однозернянка) и aegilopoides. Согласно одному из вариантов осуществления настоящего изобретения растение пшеницы является представителем подвидов Triticum turgidum и, прежде всего, представителем подвидов дурум, например представителей культиваров Ciccio, Colosseo или Utopia.
Под понятие “растение пшеницы” подпадают растения пшеницы на любой фазе созревания или развития, а также любые ткани или органы (части растения), взятые или выведенные из такого растения, если иное не следует из контекста. Части растения включают (но не ограничиваясь ими) стебли, корни, цветки, семяпочки, тычинки, листья, зародыши, области меристемы, ткань каллюса, культуры пыльника, гаметофиты, спорофиты, пыльцу, микроспоры, протопласты и т.п. Под объем настоящего изобретения подпадают также семена, образовавшиеся на растениях пшеницы, предлагаемых в настоящем изобретении. В одном из объектов изобретения семена используют для размножения в чистоте (разведение гомозигот) для получения повышенной толерантности к имидазолиноновому гербициду по сравнению с семенами сорта растения пшеницы дикого типа.
Настоящее изобретение относится также к растениями тритикале, частям растений тритикале и клеткам растений тритикале, обладающим повышенной толерантностью к имидазолиноновым гербицидам. В контексте настоящего описания понятие “растение тритикале” относится к растению, созданному путем скрещивания растения ржи (Secale cereale) либо с тетраплоидным растением пшеницы (например, Triticum turgidum), либо с гексаплоидным растением пшеницы (например, Triticum aestivum). Под объем настоящего изобретения подпадают также семена, образовавшиеся на указанных в настоящем описании растениях тритикале, и способы борьбы с сорняками вблизи указанных растений тритикале.
В настоящем изобретении описано растение пшеницы, содержащее по меньшей мере одну нуклеиновую кислоту IMI, где растение пшеницы обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом этого растения дикого типа. Растения пшеницы, предлагаемые в настоящем изобретении, могут иметь несколько нуклеиновых кислот IMI из различных геномов, поскольку эти растения могут нести более одного генома. Например, растение пшеницы Triticum turgidum несет два генома, которые обычно обозначают как геномы А и Б. Поскольку AHAS представляет собой необходимый для метаболизма фермент, можно предположить, что каждый геном имеет по меньшей мере один ген, кодирующий фермент AHAS (т.е. по меньшей мере один ген Als), как правило, в сочетании с другими метаболическими ферментами, которые приведены на известных генетических картах тетраплоидной пшеницы. В контексте настоящего описания понятие “локус гена Als” относится к положению гена Als в геноме, а понятия “ген Als” и “нуклеиновая кислота Als” относятся к нуклеиновой кислоте, кодирующей фермент AHAS. Нуклеиновая кислота Als в каждом геноме отличается по нуклеотидной последовательности от нуклеиновой кислоты Als в другом геноме. Специалист в данной области может определить исходный геном каждой нуклеиновой кислоты Als с использованием методов генетического скрещивания и/или секвенирования или путем расщепления экзонуклеазой, эти методы хорошо известны специалисту в данной области. В контексте настоящего описания понятия “нуклеиновая кислота Als 1”, “нуклеиновая кислота Als 2” и “нуклеиновая кислота Als 3” относятся к нуклеиновым кислотам Als, локализованным в трех различных геномах. В контексте настоящего описания подразумевается, что локус гена Als 3 локализован в геноме А, а локус гена Als 2 локализован в геноме Б. В контексте настоящего описания подразумевается также, что нуклеиновые кислоты IMI, полученные из генома А или Б, являются различными и их обозначают как нуклеиновые кислоты Imi 3 или Imi 2 соответственно.
В контексте настоящего описания понятие “нуклеиновая кислота IMI” относится к нуклеиновой кислоте Als, которая имеет последовательность, несущую мутацию по сравнению с нуклеиновой кислотой Als дикого типа, которая придает повышенную толерантность к имидазолинону растению, в котором происходит ее экспрессия. В контексте настоящего описания понятие “нуклеиновая кислота Imi 1”, “нуклеиновая кислота Imi 2” и “нуклеиновая кислота Imi 3” относится к нуклеиновым кислотам IMI, которые обозначают обусловливающие толерантность к имидазолинону аллели генов Als 1, Als 2 и Als 3 соответственно. Поскольку растения пшеницы имеют по 2 копии каждого генома, то растения пшеницы несут по две копии каждой конкретной нуклеиновой кислоты Als. Например, растение пшеницы Triticum turgidum несет по две копии геномов А и Б и следовательно по две копии каждого из генов Als 3 и Als 2. В контексте настоящего описания понятие “аллель IMI” относится к одной копии конкретной нуклеиновой кислоты IMI. Таким образом, в контексте настоящего описания подразумевается, что растение пшеницы может иметь две копии аллелей Imi 2, по одной из каждой двух копий генома Б.
Согласно другому варианту осуществления изобретения растение пшеницы содержит несколько нуклеиновых кислот IMI. В контексте настоящего описания при ссылке на растение, которое содержит “несколько нуклеиновых кислот IMI”, фраза “несколько нуклеиновых кислот IMI” обозначает присутствие различных нуклеиновых кислот IMI в растении и не зависит от того, является ли растение гомозиготным или гетерозиготным, в частности по локусу Als. Например, растение, содержащее несколько нуклеиновых кислот IMI, может содержать нуклеиновую кислоту Imi 2 и Imi 3 в отличие от растения, которое несет две копии нуклеиновой кислоты Imi 2.
Класс нуклеиновых кислот Imi 2 включает нуклеиновую кислоту Imi 2 из описанных ниже линий CI19, UT01, UT03, UT05, UT07, UT08, UT10, UT13, UT14, UT16, UT17 и UT20. Класс нуклеиновых кислот Imi 3 включает нуклеиновые кислоты Imi 3 из описанных ниже линий UT12, UT15 и UT19. Каждый класс Imi может включать представителей, полученных из различных видов пшеницы. Таким образом, каждый класс Imi включает нуклеиновые кислоты IMI, которые отличаются по нуклеотидной последовательности, но для которых тем не менее установлено с использованием анализов наследования, известных обычным специалистам в данной области, что они получены из одного и того же генома пшеницы или локализованы в одном и том же геноме.
Таким образом, под объем настоящего изобретения подпадает растение пшеницы, которое содержит по меньшей мере одну нуклеиновую кислоту IMI, где растение пшеницы обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа и где по меньшей мере одну нуклеиновую кислоту IMI выбирают из группы, включающей нуклеиновую кислоту Imi 1, нуклеиновую кислоту Imi 2 и нуклеиновую кислоту Imi 3. Согласно одному из вариантов осуществления изобретения растение содержит как нуклеиновую кислоту Imi 2, так и нуклеиновую кислоту Imi 3. В предпочтительном варианте осуществления изобретения нуклеиновая кислота Imi 2 имеет полинуклеотидную последовательность SEQ ID NO:1. В другом предпочтительном варианте осуществления изобретения нуклеиновая кислота Imi 3 имеет полинуклеотидную последовательность SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23.
Под объем настоящего изобретения подпадает толерантное к имидазолинону растение тритикале. В контексте настоящего описания понятие “растение тритикале” относится к растению, созданному путем скрещивания растения ржи (Secale cereale) либо с тетраплоидным растением пшеницы (например, Triticum turgidum), либо с гексаплоидным растением пшеницы (например, Triticum aestivum). В контексте настоящего описания подразумевается, что толерантное к имидазолинону растение тритикале содержит по меньшей мере одну нуклеиновую кислоту IMI, где растение тритикале обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа и где по меньшей мере одну нуклеиновую кислоту IMI выбирают из группы, включающей нуклеиновую кислоту Imi 1, нуклеиновую кислоту Imi 2 и нуклеиновую кислоту Imi 3. Согласно одному из вариантов осуществления изобретения растение содержит как нуклеиновую кислоту Imi 2, так и нуклеиновую кислоту Imi 3. В предпочтительном варианте осуществления изобретения нуклеиновая кислота Imi 2 имеет полинуклеотидную последовательность SEQ ID NO:1. В другом предпочтительном варианте осуществления изобретения нуклеиновая кислота Imi 3 имеет полинуклеотидную последовательность SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23.
В контексте настоящего описания касательно нуклеиновых кислот понятие “из” относится к нуклеиновой кислоте “локализованной” в конкретном геноме или “выведенной” из конкретного генома. Понятие “локализована в” относится к нуклеиновой кислоте, входящей в конкретный геном. В контексте настоящего описания понятие “выведена из” относится к нуклеиновой кислоте, которая была удалена или выделена из генома. Понятие “выделена” более подробно будет описано ниже.
Под объем настоящего изобретения подпадают растения пшеницы, несущие 1, 2 или 3 аллеля IMI, где растение пшеницы обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа. Аллели IMI могут содержать нуклеотидную последовательность, выбранную из группы, включающей полинуклеотид, указанный в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23; полинуклеотид, кодирующий полипептид, который имеет последовательность SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:24; полинуклеотид, содержащий по меньшей мере 60 последовательных нуклеотидов любого из вышеуказанных полинуклеотидов; и полинуклеотид, комплементарный любому из вышеуказанных полинуклеотидов. Под объем настоящего изобретения подпадают растения тритикале, несущие 1, 2 или 3 аллеля IMI, где растение тритикале обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа. Аллели IMI могут содержать нуклеотидную последовательность, выбранную из группы, включающей полинуклеотид, указанный в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23; полинуклеотид, кодирующий полипептид, который имеет последовательность SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:24; полинуклеотид, содержащий по меньшей мере 60 последовательных нуклеотидов любого из вышеуказанных полинуклеотидов; и полинуклеотид, комплементарный любому из вышеуказанных полинуклеотидов.
Согласно одному из вариантов осуществления изобретения растение пшеницы или растение тритикале содержит две различные нуклеиновые кислоты IMI, где нуклеиновые кислоты выведены или локализованы в различных геномах пшеницы. Предпочтительно две нуклеиновые кислоты представляют собой нуклеиновую кислоту Imi 2 и нуклеиновую кислоту Imi 3. Более предпочтительно нуклеиновая кислота Imi 2 содержит полинуклеотидную последовательность SEQ ID NO:1, а нуклеиновая кислота Imi 3 содержит полинуклеотидную последовательность SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23. В другом варианте осуществления растение пшеницы или растение тритикале содержит нуклеиновую кислоту IMI, где нуклеиновая кислота имеет полинуклеотидную последовательность SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23. Согласно еще одному варианту осуществления изобретения растение пшеницы содержит более двух нуклеиновых кислот IMI, где каждая нуклеиновая кислота IMI выведена из различных геномов. Предпочтительно по меньшей мере одна из нуклеиновых кислот IMI содержит полинуклеотидную последовательность, выбранную из группы, включающей SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23.
В предпочтительном варианте осуществления настоящего изобретения выделенная нуклеиновая кислота IMI кодирует аминокислотную последовательность, несущую мутацию в домене, который является консервативным для нескольких белков AHAS. Эти консервативные домены обозначены в контексте настоящего описания как домен А, домен В, домен С, домен D и домен Е. На фиг.7 показана общая локализация каждого домена в белке AHAS. Домен А содержит аминокислотную последовательность AITGQVPRRMIGT (SEQ ID NO:25). Домен В содержит аминокислотную последовательность QWED (SEQ ID NO:26). Домен С содержит аминокислотную последовательность VFAYPGGASMEIHQALTRS (SEQ ID NO:27). Домен D содержит аминокислотную последовательность AFQETP (SEQ ID NO:28). Домен Е содержит аминокислотную последовательность IPSGG (SEQ ID NO:29). В настоящем изобретении подразумевается также, что в консервативных доменах могут быть сделаны небольшие вариации, например, в растениях дурнушника остаток серина в домене заменен остатком аланина.
Таким образом, настоящее изобретение относится к растению пшеницы, несущему нуклеиновую кислоту IMI, которая кодирует аминокислотную последовательность, имеющую мутацию в консервативном домене, выбранном из группы, включающей домен А, домен В, домен С, домен D и домен Е. Согласно одному из вариантов осуществления изобретения растение пшеницы содержит нуклеиновую кислоту IMI, которая кодирует аминокислотную последовательность, имеющую мутацию в домене Е. Согласно предпочтительным вариантам осуществления изобретения мутации в консервативных доменах затрагивают положения, которые обозначены путем подчеркивания: AITGQVPRRMIGT (SEQ ID NO:25); QVVED (SEQ ID NO:26); VFAYPGGASMEIHQALTRS (SEQ ID NO:27); AFQETP (SEQ ID NO:28) и IPSGG (SEQ ID NO:29). Одной из предпочтительных замен является замена аспарагина на серин в домене Е.
Имидазолиноновые гербициды можно выбирать из группы, включающей (но не ограничиваясь ими) PURSUIT® (имизетапир), CADRE® (имазапик), RAPTOR® (имазамокс), SCEPTER® (имазахин), ASSERT® (имазетабенз), ARSENAL® (имазапир), производное любого из вышеуказанных гербицидов или смесь двух или большего количества вышеуказанных гербицидов, например, имазапир/имазамокс (ODYSSEY®). Более предпочтительно имидазолиноновый гербицид может быть выбран из группы, включающей (но не ограничиваясь ими) 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил)-4-метил-5-оксо-2-имидазолин-2-ил)-3-хинолинкарбоновую кислоту, 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-метилникотиновую кислоту и смесь метил-6-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-м-толуата и метил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-n-толуата. Предпочтительным является применение 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновой кислоты и 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновой кислоты. Особенно предпочтительным является применение 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновой кислоты.
Растения пшеницы, предлагаемые в настоящем изобретении, могут представлять собой либо трансгенные растения пшеницы, либо нетрансгенные растения пшеницы. Аналогично этому растения тритикале, предлагаемые в настоящем изобретении, могут представлять собой либо трансгенные растения тритикале, либо нетрансгенные растения тритикале. В контексте настоящего описания понятие “трансгенный” относится к любому растению, растительной клетке, каллюсу, растительной ткани или части растения, которые содержат весь рекомбинантный полинуклеотид или по меньшей мере его часть. Во многих случаях весь рекомбинантный полинуклеотид или его часть стабильно интегрированы в хромосому или представляют собой стабильный внехромосомный элемент, вследствие чего он переносится в следующие поколения. В контексте настоящего описания понятие “рекомбинантный полинуклеотид” относится к полинуклеотиду, который был изменен, перегруппирован или модифицирован с помощью генной инженерии. Примерами являются любые клонированные полинуклеотиды или полинуклеотиды, которые связаны или сочленены с гетерологичными последовательностями. Понятие “рекомбинантный” не относится к изменениям полинуклеотидов, которые являются результатом встречающихся в естественных условиях событий, таких как спонтанные мутации, или результатом неспонтанного мутагенеза с последующей избирательной селекцией. Растения, содержащие мутации, возникшие в результате неспонтанного мутагенеза и избирательной селекции, обозначены в контексте настоящего описания как нетрансгенные растения, и они подпадают под объем настоящего изобретения. В тех вариантах осуществления изобретения, в которых растение пшеницы является трансгенным и содержит несколько нуклеиновых кислот IMI, нуклеиновые кислоты могут быть получены из различных геномов или из одного и того же генома. В альтернативных вариантах осуществления изобретения, в которых растение пшеницы является нетрансгенным и содержит несколько нуклеиновых кислот IMI, нуклеиновые кислоты локализованы в различных геномах или в одном и том же геноме.
Примером нетрансгенной линии растений пшеницы, содержащих одну нуклеиновую кислоту IMI, является линия растений, депонированная в АТСС под регистрационным номером для цели патентования РТА-4960 и обозначенная в настоящем описании как линия пшеницы CI19. Линия пшеницы СI19 содержит нуклеиновую кислоту Imi 2. Нуклеотидная последовательность, соответствующая локусу гена Als 2 линии СI19, представлена в SEQ ID NO:1. Другими примерами нетрансгенных линий растений пшеницы, содержащих одну нуклеиновую кислоту IMI, являются линии растений, депонированных в АТСС под регистрационными номерами для цели патентования РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4917, РТА-4918, РТА-4920, РТА-4921, РТА-4923 и РТА-4960; и они обозначены в настоящем описании как линии UT01, UT03, UT05, UT07, UT08, UT10. UT13, UT14, UT16, UT17 и UT20 соответственно. Нуклеотидная последовательность, соответствующая локусу гена Als 2 в линиях UT01, UT03, UT05, UT07, UT08, UT10, UT13, UT14, UT16, UT17 и UT20, идентична полинуклеотидной последовательности, приведенной в SEQ ID NO:1.
Еще одним примером нетрансгенной линии растений пшеницы, содержащих одну нуклеиновую кислоту IMI, является линия растений, депонированная в АТСС под регистрационным номером для цели патентования РТА-4916 и обозначенная в настоящем описании как линия пшеницы UT12. Линия пшеницы UT12 содержит нуклеиновую кислоту Imi 3. Нуклеотидная последовательность, соответствующая локусу гена Als 3 в линии UT12, представлена в SEQ ID NO:3.
Следующим примером нетрансгенной линии растений пшеницы, содержащих одну нуклеиновую кислоту IMI, является линия растений, депонированная в АТСС под регистрационным номером для цели патентования РТА-4919 и обозначенная в настоящем описании как линия пшеницы UT15. Линия пшеницы UT15 содержит нуклеиновую кислоту Imi 3. Нуклеотидная последовательность, соответствующая локусу гена Als 3 в линии UT15, представлена в SEQ ID NO:5. Еще одним примером нетрансгенной линии растений пшеницы, содержащих одну нуклеиновую кислоту IMI, является линия растений, депонированная в АТСС под регистрационным номером для цели патентования РТА-4922 и обозначенная в настоящем описании как линия пшеницы UT12. Линия пшеницы UT12 содержит нуклеиновую кислоту Imi 3. Нуклеотидная последовательность, соответствующая локусу гена Als 3 в линии UT12, представлена в SEQ ID NO:23.
Несколько депозитов примерно по 2500 в каждой из толерантных к имидазолинону линий пшеницы были помещены в Американскую коллекцию типовых культур, Манассас, шт.Виргиния, 7 января 2003 г. и 28 января 2003 г. Эти депозиты сделаны в соответствии со сроками и условиями Будапештского договора, касающегося депонирования микроорганизмов. Депозиты должны сохраняться в течение по меньшей мере 30 лет и по меньшей в течение 5 лет после самого последнего требования о его предоставлении, полученного АТСС. Депонированным семенам присвоены регистрационные номера для цели патентования РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4916, РТА-4917, РТА-4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922, РТА-4923 и РТА-4960.
Настоящее изобретение относится к растению пшеницы, которое имеет регистрационный номер для цели патентования РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4916, РТА-4917, РТА-4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922, РТА-4923 или РТА-4960; мутанту, рекомбинанту или созданному с помощью генной инженерии производному растения, имеющего регистрационный номер для цели патентования РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4916, РТА-4917, РТА4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922, РТА-4923 или РТА-4960; любому потомку растения, имеющему регистрационный номер для цели патентования РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4916, РТА-4917, РТА-4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922. РТА-4923 или РТА-4960; и растению, которое является потомком любых этих растений. Согласно предпочтительному варианту осуществления изобретения растение пшеницы, предлагаемое в настоящем изобретении, дополнительно обладает признаками толерантности к гербицидам, характерными для растения, которое имеет регистрационный номер для цели патентования РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4916, РТА-4917, РТА-4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922, РТА-4923 и РТА-4960.
Настоящее изобретение относится также к гибридам растений пшеницы, представленных в настоящем описании, с другим растением пшеницы. Другое растение пшеницы представляет собой (но не ограничиваясь ими) Т. aestivum cv Fidel и любое другое растение пшеницы, несущее мутантный ген FS-1, FS-2, FS-3 или FS-4 (см. US 6339184 и заявку на US No. 08/474832). Предпочтительные гибриды содержат комбинацию нуклеиновых кислот Imi 1, Imi 2 и/или Imi 3.
Понятия “культивар” и “сорт” относятся к группе растений внутри одного вида, несущим общий набор характеристик или признаков, который рассматривается специалистами в данной области как достаточный для того, чтобы отличать один культивар или сорт от другого культивара или сорта. Ни для одного из указанных понятий не подразумевается, что все растения любого конкретного культивара или сорта должны быть генетически идентичными либо на уровне всего гена, либо молекулы, или что указанное конкретное растение должно быть гомозиготным во всех локусах. Культивар или сорт считаются полученными в результате “разведения гомозигот” по конкретному признаку, если при самоопылении полученного “разведением гомозигот” культивара или сорта все потомство несет указанный признак. Понятия “линейное разведение (разведение гомозигот)” или “линия” относятся к группе растений культивара, несущих общий набор характеристик или признаков, который рассматривается специалистами в данной области как достаточный для того, чтобы отличать одно линейное разведение или линию от другого линейного разведения или линии. Ни для одного из указанных понятий не подразумевается, что все растения любой конкретной линии разведения или линии должны быть генетически идентичными либо на уровне всего гена, либо на молекулярном уровне, или что указанное конкретное растение должно быть гомозиготным во всех локусах. Линия разведения или линия считаются полученными в результате “разведения гомозигот” по конкретному признаку, если при самоопылении полученной “разведением гомозигот” линии разведения или линии все потомство несет указанный признак. В контексте настоящего описания подразумевается, что признак возникает в результате мутации в гене Als растения или семени пшеницы или тритикале.
Следует понимать также, что растение пшеницы или тритикале, предлагаемое в настоящем изобретении, может содержать нуклеиновую кислоту Als дикого типа помимо нуклеиновой кислоты IMI. Подразумевается, что толерантные к имидазолинону линии могут иметь мутацию только в одном из множества изоферментов AHAS. Таким образом, настоящее изобретение относится к растению пшеницы или тритикале, которое содержит одну или несколько нуклеиновых кислот IMI в дополнение к одной или нескольким нуклеиновым кислотам Als дикого типа.
Помимо растений пшеницы и тритикале настоящее изобретение относится также к выделенным белкам и нуклеиновым кислотам IMI. Нуклеиновые кислоты содержат полинуклеотид, выбранный из группы, включающей полинуклеотид, имеющий последовательность SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23; полинуклеотид, который кодирует полипептид, имеющий последовательность SEQ ID NO:2, SEQ ID NO:4. SEQ ID NO:6 или SEQ ID NO:24; полинуклеотид, содержащий по меньшей мере 60 последовательных нуклеотидов любого из вышеуказанных полинуклеотидов; и полинуклеотид, комплементарный любому из вышеуказанных полинуклеотидов. Согласно предпочтительному варианту осуществления изобретения нуклеиновая кислота IMI содержит полинуклеотидную последовательность SEQ ID NO:1. Согласно другому предпочтительному варианту осуществления изобретения нуклеиновая кислота IMI содержит полинуклеотидную последовательность SEQ ID NO:3. Согласно еще одному предпочтительному варианту осуществления изобретения нуклеиновая кислота IMI содержит полинуклеотидную последовательность SEQ ID NO:5.
Понятие “белок AHAS” или “полипептид AHAS” относится к белку синтазы ацетогидроксикислот, а понятие “белок IMI” относится к любому белку AHAS, который является мутантом белка AHAS дикого типа и который придает повышенную толерантность к имидазолинону растению, растительной клетке, части растения, семени растения или растительной ткани при экспрессии в них.
В предпочтительном варианте осуществления изобретения белок IMI содержит полипептид, кодируемый полинуклеотидной последовательностью, содержащей SEQ ID NO:1. В другом предпочтительном варианте осуществления изобретения белок IMI содержит полипептид, кодируемый полинуклеотидной последовательностью, содержащей SEQ ID NO:3. Еще в одном предпочтительном варианте осуществления изобретения белок IMI содержит полипептид, кодируемый полинуклеотидной последовательностью, содержащей SEQ ID NO:5 или SEQ ID NO:23. В контексте настоящего описания также понятия “нуклеиновая кислота” и “полинуклеотид” относятся к РНК или ДНК, которая может быть линейной или разветвленной, одноцепочечной или двухцепочечной, или к ее гибриду. Под понятия подпадают также гибриды РНК/ДНК. Под эти понятия подпадает также нетранслируемая последовательность, локализованная как на 3'-, так и на 5'-концах кодирующей области гена: на расстоянии по меньшей мере примерно 1000 нуклеотидов указанной последовательности против хода транскрипции от 5'-конца кодирующей области и по меньшей мере примерно 200 нуклеотидов указанной последовательности по ходу транскрипции от 3'-конца кодирующей области гена. Более редкие основания, такие как инозин, 5-метилцитозин, 6-метиладенин, гипоксантин и другие, можно применять также в случае антисмысловой двухцепочечной (ds)PHK и для спаривания рибозимов. Например, установлено, что полинуклеотиды, которые содержат С-5 пропионовые аналоги уридина и цитидина, связываются с РНК с высокой аффинностью и являются эффективными антисмысловыми ингибиторами экспрессии гена. Можно осуществлять также другие модификации, такие как модификация фосфодиэфирного каркаса или 2'-гидроксигруппы в остатке сахара рибозы в РНК. Антисмысловые полинуклеотиды и рибозимы могут полностью состоять из рибонуклеотидов или могут содержать смесь рибонуклеотидов и дезоксирибонуклеотидов. Полинуклеотиды, предлагаемые в изобретении, можно получать любыми методами, в том числе с помощью геномных препаратов, препаратов кДНК, синтеза in vitro, ОТ-ПЦР и транскрипции in vitro или in vivo.
Понятие “выделенная молекула нуклеиновой кислоты” относится к молекуле, которая практически отделена от других молекул нуклеиновых кислот, которые присутствуют во встречающемся в естественных условиях источнике нуклеиновой кислоты (т.е. последовательностей, которые кодируют другие полипептиды). Предпочтительно “выделенная” нуклеиновая кислота не содержит некоторых последовательностей, которые в естественных условиях фланкируют нуклеиновую кислоту (т.е. последовательностей, локализованных на 5'- и 3'-концах нуклеиновой кислоты) в ее встречающемся в естественных условиях репликоне. Например, клонированная нуклеиновая кислота считается выделенной. Согласно различным вариантам осуществления изобретения выделенная молекула нуклеиновой кислоты IMI может содержать менее примерно 5, 4, 3, 2, 1, 0,5 или 0,1 т.п.н. нуклеотидных последовательностей, которые в естественных условиях фланкируют молекулу нуклеиновой кислоты в геномной ДНК клетки, из которой нуклеиновая кислота получена (например, клетки Triticum turgidum). Нуклеиновая кислота также считается выделенной, если она уже подвергалась обработке человеком или помещена в локус или локализована в сайте, встречающемся в естественных условиях, или ее интродуцировали в клетку путем заражения Agrobacterium, биобаллистическим методом или с помощью любого другого метода трансформации растений. Кроме того, “выделенная” молекула нуклеиновой кислоты, такая как молекула кДНК, может быть свободна от некоторого другого клеточного материала, с которым она связана в естественных условиях, или от клеточной среды, при ее получении с помощью методов рекомбинации, или от химических предшественников, или других химических веществ, когда ее получают химическим синтезом.
Конкретными исключениями, которые не подпадают под понятие “выделенные нуклеиновые кислоты”, являются: встречающиеся в естественных условиях хромосомы (такие как хромосомные препараты), библиотеки искусственных хромосом, геномные библиотеки и библиотеки кДНК, которые существуют либо в виде препарата нуклеиновой кислоты in vitro, либо в виде препарата трансфектированных/трансформированных клеток-хозяев, где клетки-хозяева либо представляют собой гетерогенный препарат in vitro, либо культивируются в виде гетерологичной популяции индивидуальных колоний. Также конкретными примерами исключений являются указанные выше библиотеки, в которых на долю конкретной нуклеиновой кислоты приходится менее 5% от общего количества нуклеиновых кислот, встроенных в векторную молекулу. Кроме того, конкретными исключениями являются препараты полной клеточной геномной ДНК или полной клеточной РНК (включая полные клеточные препараты, отделенные механическим путем или расщепленные ферментативно). Также конкретными примерами исключений являются полные клеточные препараты, которые входят в состав либо препарата in vitro, либо в виде гетерогенной смеси, разделенной с помощью электрофореза, в которой нуклеиновую кислоту, предлагаемую в изобретении, уже нельзя дополнительно отделить от гетерологичных нуклеиновых кислот, присутствующих в электрофоретической среде (например, дополнительно отделить путем вырезания индивидуальной полосы из гетерогенной популяции полос в агарозном геле или нейлоновом блоте).
Молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, например, молекулу нуклеиновой кислоты, которая содержит нуклеотидную последовательность SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23, или ее фрагмент, можно выделять с помощью стандартных методов молекулярной биологии и информации о последовательности, представленной в настоящем описании. Например, кДНК IMI Т. turgidum можно выделять из библиотеки Т. turgidum с использованием всей последовательности или фрагмента последовательности SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23. Кроме того, молекулу нуклеиновой кислоты, включающую всю последовательность или фрагмент SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23, можно выделять с помощью полимеразной цепной реакции с использованием олигонуклеотидных праймеров, созданных на основе этой последовательности. Например, мРНК можно выделять из растительных клеток (например, с помощью процедуры экстракции тиоцианатом гуанидиния, которая описана у Chirgwin и др., Biochemistry 18, 1979, cc.5294-5299), а кДНК можно получать с помощью обратной транскриптазы (например, обратной транскриптазы вируса мышиного лейкоза (MLV) Молони, поступающей в продажу от фирмы Gib-Gibco/BRL, Бетесда, шт.Мэриленд; или обратной транскриптазы вируса миелобластоза птиц (AMV), поступающей в продажу от фирмы Seikagaku America, Inc., Сент-Питерсберг, шт.Флорида). На основе нуклеотидной последовательности, представленной в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23, можно создавать синтетические олигонуклеотидные праймеры для амплификации с помощью полимеразной цепной реакции. Молекулу нуклеиновой кислоты, предлагаемую в изобретении, можно амплифицировать с использованием кДНК или в альтернативном варианте геномной ДНК в качестве матрицы и соответствующих олигонуклеотидных праймеров с помощью основанных на применении ПЦР стандартных методов амплификации. Амплифицированную таким образом молекулу нуклеиновой кислоты можно клонировать в приемлемом векторе и характеризовать с помощью анализа последовательности ДНК. Кроме того, олигонуклеотиды, соответствующие нуклеотидной последовательности IMI, можно получать с помощью стандартного синтеза, например, с использованием автоматического синтезатора ДНК.
Нуклеиновые кислоты IMI, предлагаемые в настоящем изобретении, могут содержать последовательности, кодирующие белок IMI (т.е. “кодирующие области”), а также 5'-нетранслируемые последовательности и 3'-нетранслируемые последовательности. В альтернативном варианте молекулы нуклеиновых кислот, предлагаемые в настоящем изобретении, могут содержать только кодирующие области гена IMI или могут содержать полные геномные фрагменты, выделенные из геномной ДНК. Кодирующая область этих последовательностей обозначена как “положение открытой рамки считывания (ОРС)”. Кроме того, молекула нуклеиновой кислоты, предлагаемая в изобретении, может содержать часть кодирующей области гена IMI, например фрагмент, который можно применять в качестве зонда или праймера. Нуклеотидные последовательности, полученные путем клонирования генов IMI Т. turgidum, позволяют создавать зонды и праймеры, которые можно применять для идентификации и/или клонирования гомологов IMI в других типах клеток и организмах, а также гомологов IMI из других растений пшеницы и родственных видов. Часть кодирующей области может кодировать также биологически активный фрагмент белка IMI.
В контексте настоящего описания под понятием “биологически активная часть (фрагмент)” белка IMI понимают фрагмент, например, домен/мотив белка IMI, который при производстве в растении повышает толерантность растений к имидазолиноновому гербициду по сравнению с сортом этого растения дикого типа. Методы количественной оценки повышенной толерантности к имидазолиноновым гербицидам представлены ниже в примерах. Биологически активные фрагменты белка IMI включают пептиды, выведенные из SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:24, которые состоят из меньшего количества аминокислот по сравнению с полноразмерным белком IMI и придают повышенную толерантность к имидазолиноновому гербициду при экспрессии в растении. Как правило, биологически активные фрагменты (например, пептиды, состоящие, например из 5, 10, 15, 20, 30, 35, 36, 37, 38, 39, 40, 50, 100 или большего количества аминокислот) несут домен или мотив, обладающий по меньшей мере одним видом активности белка IMI. Кроме того, другие биологически активные фрагменты, в которых другие области полипептида удалены путем делеции, можно получать с помощью методов рекомбинации и оценивать в отношении одного или нескольких видов активности, указанных в настоящем описании. Предпочтительно биологически активные фрагменты белка IMI включают один или несколько консервативных доменов, выбранных из группы, включающей домен А, домен В, домен С, домен D и домен Е, где консервативный домен несет мутацию.
Изобретение относится также к химерным или слитым полипептидам IMI. В контексте настоящего описания понятие “химерный полипептид IMI” или “слитый полипептид” относится к полипептиду IMI, функционально связанному с полипептидом, который не относится к IMI. Понятие “полипептид, который не относится к IMI”, означает полипептид, имеющий аминокислотную последовательность, которая не является практически идентичной полипептиду IMI, например, полипептид, который не представляет собой изофермент IMI, этот пептид имеет функцию, отличную от функции полипептида IMI. В контексте настоящего описания понятие “функционально связанный” касательно слитого пептида предназначено для обозначения того факта, что полипептид IMI и полипептид, который не относится к полипептиду IMI, слиты друг с другом таким образом, что обе последовательности выполняют предполагаемую функцию, присущую применяемой последовательности. Полипептид, который не относится к IMI, можно сливать с N-концом или С-концом полипептида IMI. Например, в одном варианте осуществления изобретения слитый полипептид представляет собой слитый полипептид GST-IMI, в котором последовательность IMI слита с С-концом последовательности GST. Такие слитые полипептиды могут облегчать очистку рекомбинантных полипептидов IMI. В другом варианте осуществления изобретения слитый полипептид представляет собой полипептид IMI, содержащий гетерологичную сигнальную последовательность на N-конце. В определенных клетках-хозяевах (например, в клетках-хозяевах млекопитающих) экспрессия и/или секреция полипептида IMI может возрастать благодаря применению гетерологичной сигнальной последовательности.
Выделенная молекула нуклеиновой кислоты, кодирующая полипептид IMI, который имеет последовательность, идентичную последовательности полипептида, кодируемого полинуклеотидной последовательностью SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23, можно создавать, осуществляя одну или несколько нуклеотидных замен, добавлений или делеций в нуклеотидной последовательности SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23 так, чтобы интродуцировать в кодируемый полипептид одну или несколько аминокислотных замен, добавлений или делеций. В последовательность SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23 можно интродуцировать мутации с помощью стандартных методов, таких как сайтнаправленный мутагенез и опосредуемый ПЦР мутагенез. Предпочтительно осуществляют консервативные аминокислотные замены одного или нескольких выбранных остатков заменимых аминокислот.
“Консервативная аминокислотная замена” представляет собой замену, при которой аминокислотный остаток заменяют аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, известны в данной области. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, выбранный остаток заменимой аминокислоты в полипептиде IMI предпочтительно заменяют другим аминокислотным остатком из семейства со сходными боковыми цепями. В другом варианте осуществления изобретения мутации можно интродуцировать случайно во всю или в часть кодирующей последовательности IMI, например, с помощью насыщающего мутагенеза, а полученные мутанты можно подвергать скринингу в отношении активности IMI согласно представленному в настоящем описании методу для идентификации мутантов, которые сохраняют активность IMI. После мутагенеза последовательностей SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23 кодируемый полипептид можно экспрессировать рекомбинантно и активность полипептида можно определять, анализируя толерантность к имидазолинону растения, экспрессирующего полипептид, согласно описанному ниже в примерах методу.
Для определения процента идентичности двух аминокислотных последовательностей последовательности линеаризируют для оптимизации сравнения (например, можно вводить бреши в последовательность одного полипептида для оптимального сравнительного анализа первичной структуры с другим полипептидом). Затем сравнивают аминокислотные остатки в соответствующих положениях аминокислотной последовательности. Когда в определенном положении в одной последовательности находится та же аминокислота, которая находится в соответствующем положении в другой последовательности, то молекулы являются идентичными в этом положении. Такой же тип сравнительного анализа можно применять для двух нуклеотидных последовательностей. Процент идентичности последовательностей двух последовательностей является функцией от количества идентичных положений, характерных для последовательностей (т.е. процент идентичности последовательностей = количеству идентичных положений/общее количество положений ×100). В контексте настоящего описания процент идентичности последовательностей двух нуклеотидных или полипептидных последовательностей определяют с помощью программного обеспечения Vector NTI 6.0 (PC) (InforMax, 7600 Wisconsin Ave., Бетезда, шт.Мэриленд 20814). Для определения процента идентичности двух нуклеиновых кислот используют штраф за открытие бреши 15 и штраф за удлинение бреши 6,66. Для определения процента идентичности двух полипептидов используют штраф за открытие бреши 10 и штраф за удлинение бреши 0,1. Все другие параметры задаются по умолчанию.
Следует понимать, что для определения идентичности последовательностей при сравнении последовательности ДНК с последовательностью РНК тимидиновый нуклеотид эквивалентен урациловому нуклеотиду. Выделенные полипептиды IMI, предлагаемые в настоящем изобретении, предпочтительно по меньшей мере примерно на 50-60%, предпочтительно по меньшей мере примерно на 60-70% и более предпочтительно по меньшей мере примерно на 70-75%, 75-80%, 80-85%, 85-90% или 90-95% и наиболее предпочтительно по меньшей мере примерно на 96%, 97%, 98%, 99% или более идентичны полной аминокислотной последовательности, представленной в SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:24. В другом варианте осуществления изобретения выделенные полипептиды IMI, предлагаемые в настоящем изобретении, по меньшей мере примерно на 50-60%, предпочтительно по меньшей мере примерно на 60-70% и наиболее предпочтительно по меньшей мере примерно на 70-75%; 75-80%, 80-85%, 85-90% или 90-95% и наиболее предпочтительно по меньшей мере примерно на 96%, 97%, 98%, 99% или более идентичны полной аминокислотной последовательности, представленной в SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:24. Кроме того, можно создавать оптимизированные нуклеиновые кислоты IMI. Предпочтительно оптимизированная нуклеиновая кислота IMI кодирует полипептид IMI, который модулирует толерантность растений к имидазолиноновым гербицидам, и более предпочтительно повышает толерантность растений к имидазолиноновому гербициду при ее сверхэкспрессии в растении. В контексте настоящего описания понятие “оптимизированная” относится к нуклеиновой кислоте, которую с помощью методов генной инженерии создают так, чтобы повышать ее уровень экспрессии в данном растении или организме животного. Для создания растительных оптимизированных нуклеиновых кислот IMI последовательность ДНК гена можно модифицировать так, чтобы она: 1) содержала кодоны, предпочтительные для генов, характеризующихся высоким уровнем экспрессии в растениях; 2) имела практически такое же содержание А+Т в составе нуклеотидных оснований, которое характерно для растений; 3) имела форму растительной инициирующей последовательности; 4) из нее были удалены последовательности, вызывающие дестабилизацию, несоответствующее полиаденилирование, расщепление и терминацию РНК или образующие вторичную структуру “шпилек” или сайты сплайсинга РНК. Повышенный уровень экспрессии нуклеиновых кислот IMI в растениях можно получать, используя частоту распределения наиболее часто встречающихся кодонов (наиболее предпочтительных), которая характерна для растений в целом или для конкретного растения. Методы оптимизации экспрессии нуклеиновых кислот в растениях описаны в ЕРА 0359472; ЕРА 0385962; заявке РСТ WO 91/16432; US 5380831; US 5436391; у Perlack и др., Proc. Natl. Acad. Sci. USA 88, 1991, cc.3324-3328; и Murray и др., Nucleic Acids Res. 17, 1989, cc.477-498.
В контексте настоящего описания понятие “частота предпочтительного наиболее часто встречающегося кодона” означает предпочтение, проявляемое конкретной клеткой-хозяином в отношении встречаемости нуклеотидных кодонов, специфичных для конкретной аминокислоты. Для определения частоты встречаемости конкретного кодона в гене, количество данного кодона в гене делят на общее количество всех кодонов, специфичных для одной и той же аминокислоты, в гене. Аналогично этому частоту предпочтительного наиболее часто встречающегося кодона, характерную для клетки-хозяина, можно рассчитывать по средней частоте предпочтительного наиболее часто встречающегося кодона в большом количестве генов, экспрессируемых клеткой-хозяином. Предпочтительно, чтобы такой анализ был ограничен генами с высоким уровнем экспрессии в клетке-хозяине. Процент отклонения частот предпочтительного наиболее часто встречающегося кодона в синтетическом гене по сравнению с геном, который встречается в клетке-хозяине в естественных условиях, рассчитывают, определяя сначала процент отклонения частот встречаемости индивидуального кодона от характерного для клетки-хозяина в естественных условиях, после чего определяют среднее отклонение для всех кодонов. Как указано выше, в этом расчете учитываются уникальные кодоны (т.е. ATG и TGG). В целом, общее среднее отклонение наиболее часто встречающихся кодонов в оптимизированном гене по сравнению с геном, характерным для клетки-хозяина, рассчитывают из уравнения 1А: n=1 Z Xn-Yn Хn×100 Z, где Xn обозначает частоту встречаемости кодона n в клетке-хозяине; Yn обозначает частоту встречаемости кодона n в синтетическом гене; n обозначает индивидуальный кодон, специфичный для аминокислоты; a Z обозначает общее количество кодонов. Общее отклонение частоты встречаемости кодона, А, для всех аминокислот предпочтительно должно не превышать примерно 25% и более предпочтительно составлять менее примерно 10%.
Следовательно, нуклеиновую кислоту IMI можно оптимизировать таким образом, чтобы распределение частоты встречаемости кодонов отклонялось предпочтительно не более чем на 25% от частоты встречаемости в генах, для которых характерен высокий уровень экспрессии в растениях, более предпочтительно не более чем примерно на 10%. Кроме того, принимается во внимание процентное содержание G+C вырожденного третьего основания (для однодольных растений предпочтительно наличие в этом положении G+C, а для двудольных это не является предпочтительным). Установлено также, что нуклеотид XCG (где Х обозначает А, Т, С или G) представляет собой наименее предпочтительный кодон у двудольных, а кодон ХТА отсутствует как у однодольных, так и двудольных растений. Оптимизированные нуклеиновые кислоты IMI, предлагаемые в настоящем изобретении, предпочтительно также имеют индексы исключения дуплета CG и ТА, очень близкие к индексам в выбранном растении-хозяине (т.е. Triticum turgidum). Более предпочтительно эти индексы отклоняются от индексов хозяина не более чем примерно на 10-15%.
Помимо молекул нуклеиновых кислот, кодирующих описанные выше полипептиды IMI, другим объектом изобретения являются молекулы нуклеиновых кислот, антисмысловые по отношению к ним. Считается, что антисмысловые нуклеотиды ингибируют экспрессию гена полинуклеотида-мишени в результате специфического связывания с полинуклеотидом-мишенью и воздействия на транскрипцию, сплайсинг, транспорт, трансляцию и/или стабильность полинуклеотида-мишени. Из прототипов известны методы направленного переноса антисмыслового полинуклеотида в хромосомную ДНК, в первичный транскрипт РНК или в процессированную мРНК. Предпочтительно области-мишени представляют собой сайты сплайсинга, кодоны инициации трансляции, кодоны терминации трансляции и другие последовательности внутри открытой рамки считывания.
Понятие “антисмысловой” в контексте настоящего описания относится к нуклеиновой кислоте, содержащей полинуклеотид, который комплементарен всему гену или его части, первичному транскрипту или процессированной мРНК в той степени, чтобы оказывать воздействие на экспрессию эндогенного гена. Понятие “комплементарные полинуклеотиды” относится к полинуклеотидам, которые способны к спариванию оснований в соответствии со стандартными правилами комплементарности Ватсона-Крика. В частности, пурины образуют пару оснований с пиримидинами с формированием комбинации гуанина, спаренного с цитозином (G:C), и аденина, спаренного либо с тимином (А:Т), в случае ДНК, либо аденина, спаренного с урацилом (A:U), в случае РНК. Следует понимать, что два полинуклеотида могут гибридизоваться друг с другом, даже если они не полностью комплементарны друг другу, при условии, что каждый имеет по меньшей мере одну область, практически комплементарную другой. Понятие “антисмысловая нуклеиновая кислота” относится к кассетам экспрессии одноцепочечной РНК, а также двухцепочечной ДНК, которые могут транскрибироваться с образованием антисмысловой РНК. “Активные” антисмысловые нуклеиновые кислоты представляют собой антисмысловые молекулы РНК, которые могут избирательно гибридизоваться с первичным транскриптом или мРНК, кодирующей полипептид, последовательность которого по меньшей мере на 80% идентична полипептидной последовательности SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:24.
Помимо описанных выше нуклеиновых кислот и полипептидов IMI настоящее изобретение относится также к этим нуклеиновым кислотам и полипептидам, связанным с фрагментом. Эти фрагменты включают (но не ограничиваясь ими) фрагменты для обнаружения, фрагменты для гибридизации, фрагменты для очистки, фрагменты для введения, фрагменты для осуществления реакции, фрагменты для связывания и т.п. Типичной группой нуклеиновых кислот с присоединенными фрагментами являются зонды и праймеры. Зонды и праймеры, как правило, содержат практически выделенный олигонуклеотид. Олигонуклеотид, как правило, содержит область нуклеотидной последовательности, которая гидридизуется в строгих условиях по меньшей мере примерно с 12, предпочтительно примерно с 25, более предпочтительно примерно с 40, 50 или 75 последовательными нуклеотидами смысловой цепи последовательности, представленной в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23, антисмысловой последовательности последовательности, представленной в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23, или с их встречающимися в естественных условиях мутантами. Праймеры, основой которых является нуклеотидная последовательность SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23, можно использовать в реакции ПЦР для клонирования гомологов IMI. Зонды, основой которых являются нуклеотидные последовательности IMI, можно применять для обнаружения транскриптов или геномных последовательностей, кодирующих тот же самый полипептид или гомологичные полипептиды. В предпочтительных вариантах осуществления изобретения зонд дополнительно содержит присоединенную к нему группу, служащую в качестве метки, например, метка может представлять собой радиоактивный изотоп, флуоресцентное соединение, фермент или кофактор фермента. Такие зонды можно применять в качестве компонента набора для анализа, включающего геномные маркеры, для идентификации клеток, экспрессирующих полипептид IMI, в частности, путем измерения уровня кодирующей IMI нуклеиновой кислоты в образце клеток, например, путем определения уровней мРНК IMI или определения того, является ли геномный ген IMI мутантным или удален путем делеции.
Изобретение относится также к выделенному рекомбинантному экспрессионному вектору, содержащему описанную выше нуклеиновую кислоту, где экспрессия вектора в клетке-хозяине приводит к повышенной толерантности к имидазолиноновому гербициду по сравнению с клеткой-хозяином сорта дикого типа. В контексте настоящего описания понятие “вектор” относится к молекуле нуклеиновой кислоты, обладающей способностью транспортировать другую нуклеиновую кислоту, с которой она связана. Одним из типов вектора является “плазмида”, это понятие относится к кольцевой двухцепочечной цепи ДНК, в которую встроены путем лигирования дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, в вирусный геном которого могут быть встроены путем лигирования дополнительные сегменты ДНК. Определенные векторы обладают способностью к автономной репликации в клетке-хозяине, в которую они интродуцированы (например, бактериальные векторы, имеющие бактериальный сайт инициации репликации, и эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) интегрируют в геном клетки-хозяина путем интродукции в клетку-хозяина, где они реплицируются вместе с геномом хозяина. Кроме того, определенные векторы обладают способностью обеспечивать экспрессию генов, с которыми они функционально связаны. Такие векторы обозначены в контексте настоящего описания как “экспрессионные векторы”. В целом, экспрессионные векторы, применяемые в методах рекомбинантной ДНК, часто имеют форму плазмид. В контексте настоящего описания понятия “плазмида” и “вектор” можно применять взаимозаменяемо, поскольку плазмида представляет собой наиболее широко применяемую форму вектора. Однако в настоящем изобретении подразумевается, что можно применять такие другие формы экспрессионных векторов, как вирусные векторы (например, ретровирусы с дефицитом репликации, аденовирусы и аденоассоциированные вирусы), которые обладают эквивалентными функциями.
Рекомбинантные экспрессионные векторы, предлагаемые в изобретении, содержат нуклеиновую кислоту, предлагаемую в изобретении, в форме, пригодной для экспрессии нуклеиновой кислоты в клетке-хозяине, это означает, что рекомбинантные экспрессионные векторы несут одну или несколько регуляторных последовательностей, выбранных на основе последовательностей, которые используются в клетке-хозяине для экспрессии, которые функционально связаны с нуклеотидной последовательностью, подлежащей экспрессии. Понятие “функционально связанный” касательно рекомбинантного экспрессионного вектора означает, что представляющая интерес нуклеотидная последовательность связана с регуляторной(ыми) последовательностью(ями) таким образом, чтобы происходила экспрессия нуклеотидной последовательности (например, in vitro в системе транскрипции/трансляции или в клетке-хозяине, когда вектор интродуцируют в клетку-хозяина). Понятие “регуляторная последовательность” включает промоторы, энхансеры и другие контролирующие экспрессию элементы (например, сигналы полиаденилирования). Такие регуляторные последовательности описаны, например, у Goeddel, Gene Expression Technology: Methods in Enzymology 185, изд-во Academic Press, San Diego, CA, 1990 и у Gruber и Crosby в: Methods in Plant Molecular Biology и Biotechnology, под ред. Glick и Thompson, часть 7, cc.89-108, изд-во CRC Press: Boca Raton. Florida, которые включены в настоящее описание в качестве ссылки. К регуляторным последовательностям относятся последовательности, обеспечивающие конститутивную экспрессию нуклеотидной последовательности во многих типах клеток-хозяев, и последовательности, которые обеспечивают экспрессию нуклеотидной последовательность только в определенных клетках-хозяевах или только в определенных условиях. Как должно быть очевидно специалистам в данной области, конструкция экспрессионного вектора может зависеть от таких факторов, как выбор клетки-хозяина, подлежащей трансформации, требуемый уровень экспрессии полипептида и т.д. Экспрессионные векторы, предлагаемые в изобретении, можно интродуцировать в клетки-хозяева для того, чтобы они продуцировали полипептиды или пептиды, включая слитые полипептиды или пептиды, кодируемые нуклеиновыми кислотами, указанными в контексте настоящего описания (например, полипептиды IMI, слитые полипептиды и т.д.).
В предпочтительном варианте осуществления настоящего изобретения полипептиды IMI экспрессируются в растениях и клетках растений, таких как одноклеточные растительные клетки (например, водоросли) (см. Falciatore и др., Marine Biotechnology, 1(3), 1999, cc.239-251 и приведенные в этой публикации ссылки) и растительные клетки высших растений (например, сперматофиты, такие как хлебные злаки). Полинуклеотид IMI можно интродуцировать в растительную клетку любыми методами, такими как трансфекция, трансформация или трансдукция, электропорация, бомбардировка частицами, заражение Agrobacterium, биобаллистика и т.п.
Приемлемые методы трансформации или трансфекции клеток-хозяев, включая растительные клетки, описаны у Sambrook и др. (Molecular Cloning: A Laboratory Manual, 2-е издание. Cold Spring Harbor Laboratory, изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989) и в других руководствах для лабораторных исследований, таких как Methods in Molecular Biology, том 44, 1995, Agrobacterium protocols, под ред. Gartland и Davey, изд-во Humana Press, Totowa, New Jersey. Поскольку повышенная толерантность к имидазолиноновым гербицидам является общим признаком, наследование которого является желательным для широкого разнообразия растений, таких как кукуруза, пшеница, рожь, овес, тритикале, рис, ячмень, соя, арахис, хлопчатник, семенной рапс и канола, маниок, перец, подсолнечник и бархатцы, пасленовые культуры, такие как картофель, табак, баклажан и томат, виды рода Vicia (вика), горох, люцерна, кустистые растения (кофе, какао, чай), виды семейства ивовых (Salix), деревья (масличная пальма, кокос), многолетние травы и кормовые культуры, эти полезные растения также являются предпочтительными растениями-мишенями для генетической инженерии согласно еще одному варианту осуществления настоящего изобретения. Согласно предпочтительному варианту растение представляет собой растение пшеницы. Кормовые культуры включают (но не ограничиваясь ими) пырей, канареечник канарский, костер, дикую рожь, мятлик, ежу сборную, люцерну, салфойн (Salfoin), лядвинец рогатый, клевер красно-белый, клевер красный и сладкий клевер.
Согласно одному из вариантов осуществления настоящего изобретения растение трансфектируют полинуклеотидом IMI с помощью опосредуемого Agrobacterium переноса гена. Одним из известных специалистам в данной области методов трансформации является погружение цветущего растения в раствор представителей р. Agrobacteria, где Agrobacteria содержит нуклеиновую кислоту IMI, с последующим размножением трансформированных гамет. Опосредуемую Agrobacterium трансформацию растений можно осуществлять, используя, например, штамм GV3101(pMP90) (Koncz и Schell, Mol. Gen. Genet. 204, 1986, cc.383-396) или штамм LBA4404 (фирма Clontech) Agrobacterium tumefaciens. Трансформацию можно осуществлять стандартными методами трансформации и регенерации (Deblaere и др., Nucl. Acids. Res. 13, 1994, cc.4777-4788; Gelvin Stanton В. и Schilperoort Robert A., Plant Molecular Biology Manual, 2-е изд. под ред. Dordrecht, изд-во Kluwer Academic PubL, 1995, в: Sect, Ringbuc Zentrale Signatur BT11-P ISBN 0-7923-2731-4; Glick Bernard R. и Thompson John E., Methods in Plant Molecular Biotogy and Biotechnology, Boca Raton: изд-во CRC Press, 1993 360 S., ISBN 0-8493-5164-2). Например, семенной paпс можно трансформировать, используя трансформацию семядоли или гипокотиля (Moloney и др., Plant Cell Report 8, 1989, cc.238-242; De Block и др., Plant Physiol, 91, 1989, cc.694-701). Применение антибиотиков для селекции Agrobacterium и растений зависит от бинарного вектора и штамма Agrobacterium, которые применяли для трансформации. Селекцию масличного рапса, как правило, осуществляют с помощью канамицина в качестве селектируемого маркера для растения. Опосредуемый Agrobacterium перенос гена в растение льна можно осуществлять, например, с помощью метода, описанного у Mlynarova и др., Plant Cell Report 13, 1994, cc.282-285. Кроме того, трансформацию сои можно осуществлять, например, с помощью метода, описанного в ЕР 0424047, US 5322783, ЕР 0397687, US 5376543 или US 5169770. Трансформацию кукурузы можно осуществлять с помощью бомбардировки частицами, опосредуемого полиэтиленгликолем поглощения ДНК или методом, основанным на применении волокна из карбида кремния (см., например, Freeling и Walbot “The maize handbook”, изд-во Springer, New York (1993) ISBN 3-540-97826-7). Конкретный пример трансформации кукурузы описан в US 5990387, а конкретный пример трансформации пшеницы описан в заявке РСТ WO 93/07256.
Согласно настоящему изобретению интродуцированный полинуклеотид IMI можно стабильно поддерживать в растительной клетке, если он включен в нехромосомный автономный репликон или интегрирован в хромосомы растения. В альтернативном варианте интродуцированный полинуклеотид IMI может присутствовать во внехромосомном нереплицирующемся векторе и кратковременно экспрессироваться или проявлять кратковременную активность. Согласно одному из вариантов осуществления изобретения можно создавать гомологичный рекомбинантный микроорганизм, в котором полинуклеотид IMI интегрирован в хромосому, при этом получают вектор, который содержит по меньшей мере часть гена AHAS, несущий делецию, добавление или замену, для того чтобы изменять, например нарушать функцию эндогенного гена AHAS и создавать ген IMI. Для создания точковой мутации посредством гомологичной рекомбинации можно применять гибриды ДНК-РНК для метода, известного как химера пластика (chimeraplasty) (Cole-Strauss и др., Nucleic Acids Research 27(5), 1997, cc.1323-1330 и Kmiec, Gene therapy American Scientist 87(3), 1999, cc.240-247). В данной области хорошо известны также другие методы гомологичной рекомбинации видов Triticum и их можно применять согласно настоящему изобретению.
В векторе для гомологичной рекомбинации ген IMI может быть фланкирован на 5'- и 3'-концах дополнительной молекулой нуклеиновой кислоты гена AHAS для того, чтобы позволять осуществлять гомологичную рекомбинацию между экзогенным геном IMI, который несет вектор, и эндогенным геном AHAS, в микроорганизме или растении. Дополнительная фланкирующая молекула нуклеиновой кислоты AHAS имеет длину, достаточную для успешной гомологичной рекомбинации с эндогенным геном. Как правило, в вектор интродуцируют фланкирующую ДНК (как на 5'-, так и на 3'-конец), которая имеет длину от нескольких сотен пар оснований вплоть до тысячи пар оснований (см., например, у Thomas К.R. и Capecchi M.R., Cell 51, 1987, с.503 описание векторов для гомологичной рекомбинации или у Strepp и др., PNAS, 95(8), 1998, cc.4368-4373 описание метода рекомбинации, основанного на применении кДНК, в патентах на имя Physcomitrella). Однако из-за того, что ген IMI в норме отличается от гена AHAS очень небольшим количеством аминокислот, фланкирующая последовательность не всегда является необходимой. Вектор для гомологичной рекомбинации интродуцируют в микроорганизм или растительную клетку (например, с помощью опосредуемого полиэтиленгликолем переноса ДНК) и клетки, в которых произошла гомологичная рекомбинация интродуцированного гена IMI с эндогенным геном AHAS, отбирают с помощью известных в данной области методов.
Согласно другому варианту осуществления изобретения можно получать рекомбинантные микроорганизмы, которые содержат выбранные системы, позволяющие регулировать экспрессию интродуцированного гена. Например, включение гена IMI в вектор, где он находится под контролем оперона lac, обеспечивает экспрессию гена IMI только в присутствии ИПТГ (изопропилтиогалактозид). Такие регуляторные системы хорошо известны в данной области.
Вне зависимости от того, находится ли он во внехромосомном нереплицирующемся векторе или в векторе, интегрированном в хромосому, полинуклеотид IMI предпочтительно находится в растительной кассете экспрессии. Растительная кассета экспрессии предпочтительно содержит регуляторные последовательности, обеспечивающие экспрессию гена в клетках растений, которые функционально связаны так, чтобы каждая последовательность могла проявлять свою функцию, например, терминацию транскрипции с помощью сигналов полиаденилирования. Предпочтительными сигналами полиаденилирования являются сигналы, полученные из тДНК Agrobacterium tumefaciens, например, ген 3, который известен как ген октопинсинтазы Ti-плазмиды pTiACH5 (Gielen и др., EMBO J. 3, 1984, с.835), или их функциональные эквиваленты, но также можно использовать все другие терминаторы, обладающие функциональной активностью в растениях. Поскольку экспрессия гена в растении очень часто не ограничена только уровнями транскрипции, растительная кассета экспрессии предпочтительно содержит другие функционально связанные последовательности типа энхансеров трансляции, такие как перекрывающая последовательность, содержащая 5'-нетранслируемую лидерную последовательность из вируса мозаики табака, которая повышает соотношение полипептида и РНК (Gallic и др., Nucl. Acids Research 15, 1987, cc.8693-8711). Примеры растительных кассет экспрессии включают кассеты, описанные подробно у: Becker D. и др., New plant binary vectors with selectable markers located proximal to the left border, Plant Mol. Biol. 20, 1992, cc.1195-1197; Bevan M.W., Binary Agrobacterium vectors for plant transformation, Nucl. Acid. Res. 12, 1984, cc.8711-8721; и Vectors for Gene Transfer in Higher Plants; в: Transgenic Plants, т.1, Engineering and Utilization, под ред. Kung и R. Wu, изд-во Academic Press, 1993, cc.15-38.
Экспрессируемый в растении ген должен быть функционально связан с соответствующим промотором, который обеспечивает своевременную экспрессию гена предпочтительным для определенного типа клеток или типа тканей образом. Промоторы, которые можно использовать в кассетах экспрессии, предлагаемых в изобретении, представляют собой любой промотор, который обладает способностью инициировать транскрипцию в растительной клетке. Такие промоторы включают (но не ограничиваясь ими) промоторы, которые можно получать из растений, вирусов растений и бактерий, которые несут гены, экспрессируемые в растениях, таких как Agrobacterium и Rhizobium.
Промотор может быть конститутивным, индуцибельным, обладающим предпочтением к стадии развития, типу клеток, ткани или органу. Конститутивные промоторы обладают активностью в большинстве ситуаций. Примерами конститутивных промоторов являются промоторы CaMV 198 и 35S (Odell и др., Nature 313, 1985, cc.810-812), промотор 35S штамма sX CaMV (Kay и др., Science 236, 1987, cc.1299-1302), промотор Sep 1, промотор актина риса (McElroy и др., Plant Cell 2, 1990, cc.163-171), промотор актина Arabidopsis, промотор убикитина (Christensen и др. Plant Molec. Biol. 18, 1989, cc.675-689); pEmu (Last и др., Theor. Appl. Genet 81, 1991, cc.581-588), промотор 35S вируса мозаики норичника шишковатого, промотор Smas (Velten и др., EMBO J. 3, 1984, cc.2723-2730), промотор GRP1-8, промотор дегидрогеназы циннамилового спирта (US 5683439), промоторы из Т-ДНК Agrobacterium, такие как промотор маннопинсинтазы, нопалинсинтезы и октопинсинтазы, малой субъединицы рибулозобифосфаткарбоксилазы (ssu-RUBISCO) и т.п.
Индуцибельные промоторы проявляют активность при определенных условиях окружающей среды, таких как присутствие или отсутствие питательного вещества или метаболита, тепло или холод, свет, поражение патогеном, анаэробные условия и т.п. Например, промотор hsp80 из Brassica индуцируется тепловым шоком; промотор PPDK индуцируется светом; промотор PR-1 из табака, Arabidopsis и кукурузы индуцируется заражением патогеном; а промотор Adhl индуцируется гипоксией и холодовым стрессом. Экспрессию гена в растениях можно облегчать также с помощью индуцибельного промотор (см. обзор у Gatz, Annu. Rev. Plant Physiol. Plant Mol. Biol. 48, 1997, cc.89-108). Индуцируемые химическими стимулами промоторы являются особенно предпочтительными, если требуется экспрессия гена в определенные моменты времени. Примерами таких промоторов является индуцируемый салициловой кислотой промотор (заявка РСТ WO 95/19443), индуцируемый тетрациклином промотор (Gatz и др., Plant J. 2, 1992, cc.397-404) и индуцируемый этанолом промотор (заявка РСТ WO 93/21334).
Предпочтительные для определенной стадии развития промоторы экспрессируются, прежде всего, на определенной стадии развития. Предпочтительные для определенной ткани и органа промоторы включают промоторы, которые, прежде всего, экспрессируются в определенных тканях или органах, таких как листья, корни, семена или ксилема. Примерами предпочтительных для ткани и предпочтительных для органов промоторов являются (но не ограничиваясь ими) предпочтительные для плода, предпочтительные для семяпочки, предпочтительные для мужской ткани, предпочтительные для семени, предпочтительные для интегумента, предпочтительные для клубня, предпочтительные для черешка, предпочтительные для околоплодника и предпочтительные для листа, предпочтительные для рыльца, предпочтительные для пыльцы, предпочтительные для пыльника, предпочтительные для лепестка, предпочтительные для чашелистика, предпочтительные для плодоножки, предпочтительные для стручка, предпочтительные для стебля, предпочтительные для корня промоторы и т.п. Предпочтительные для семени промоторы экспрессируются, прежде всего, в процессе развития и/или созревания семян. Например, предпочтительные для семени промоторы могут быть предпочтительными для зародыша, для эндосперма и семенного покрытия (см. Thompson и др., BioEssays 10, 1989, с.108). Примеры предпочтительных для семени промоторов включают (но не ограничиваясь ими) промотор целлюлозосинтазы (celA), Cim1, гамма-зеина, глобулина-1, зеина 19 кДа кукурузы (CZ19B1) и т.п.
Другие пригодные предпочтительные для ткани или предпочтительные для органа промоторы включают промотор гена napin из семенного рапса (US 5608152), USP-промотор из Vicia faba (Baeumlein и др., Mol Gen Genet 225(3), 1991, cc.459-467), промотор олеозина из Arabidopsis (заявка РСТ WO 98/45461), промотор фазеолина из Phaseolus vulgaris (US 5504200), Все4-промотор из Brassica (заявка РСТ А. WO 91/13980) или промотор В4 легумина (LeB4; Baeumlein и др., Plant Journal, 2(2), 1992, cc.23323-9), а также промоторы, обеспечивающие специфическую для семени экспрессию, в однодольных растениях типа кукурузы, ячменя, пшеницы, ржи, риса и т.д. Приемлемыми промоторами, которые следует упомянуть, являются промотор гена Ipt2 или Ipt1 из ячменя (заявка РСТ А. WO 95/15389 и заявка РСТ WO 95/23230) или промоторы, описанные в заявке РСТ WO 99/16890 (промоторы гена хордеина (hordein) из ячменя, гена глутелина (glutelin) из риса, гена оризина (oryzin) из риса, гена проламина (prolamin) из риса, гена глиадина (gliadin) из пшеницы, гена глутелина (glutelin) из пшеницы, гена глутелина (glutelin) из овса, гена касирина (kasirin) из сорго и гена секалина (secalin) из ржи).
Другие промоторы, которые можно применять в кассетах экспрессии, предлагаемых в изобретении, представляют собой (но не ограничиваясь ими) промотор основного белка, связывающего хлорофилл а/b, промоторы гистона, промотор Ар3, промотор β-конглицина, промотор напина, промотор тектина из сои, промотор зеина 15 кДа, промотор зеина 22 кДа, промотор зеина 27 кДа, промотор g-зеина из кукурузы, промоторы waxy, shrunken 1, shrunken 2 и bronze, промотор Zm13 (US 5086169), промоторы полигалактуроназы (PG) кукурузы (US 5412085 и 5545546) и промотор SGB6 (US 5470359), а также синтетические или другие встречающиеся в естественных условиях промоторы.
Дополнительную гибкость контроля экспрессии в растениях гетерологичного гена можно обеспечивать с помощью связывающих ДНК доменов и реагирующих элементов из гетерологичных источников (т.е. ДНК-связывающие домены из источника, отличного от растения). Примером такого ДНК-связывающего домена является ДНК-связывающий домен LexA (Brent и Ptashne, Cell 43:729-736).
Следующим объектом изобретения являются клетки-хозяева, в которые интродуцируют рекомбинантный экспрессионный вектор, предлагаемый в изобретении. Понятие “клетка-хозяин” и “рекомбинантная клетка-хозяин” в контексте настоящего описания используются взаимозаменяемо. Следует понимать, что эти понятия относятся не только к конкретной индивидуальной клетке, но и к потомству или потенциальному потомству такой клетки. Поскольку последующие поколения могут нести определенные модификации, связанные с мутацией или воздействиями окружающей среды, фактически такое потомство может быть не идентично родительской клетке, но оно тем не менее подпадает под объем настоящего изобретения. Клетка-хозяин может представлять собой прокариотическую или эукариотическую клетку. Например, полинуклеотид IMI можно экспрессировать в бактериальных клетках, таких как С. glutamicum, клетках насекомых, клетках грибов или клетках млекопитающих (таких как клетки яичника китайского хомячка (СНО) или COS-клетки), водорослях, реснитчатых, растительных клетках, грибах или других микроорганизмах типа С. glutamicum. Другие пригодные клетки-хозяева известны специалистам в данной области.
Клетку-хозяина, предлагаемую в изобретении, такую как прокариотическую или эукариотическую клетку-хозяина в культуре, можно применять для получения (т.е. экспрессии) полинуклеотида IMI. Таким образом, изобретение относится также к способам получения полипептидов IMI с помощью клеток-хозяев, предлагаемых в изобретении. Согласно одному из вариантов осуществления изобретения способ заключается в том, что культивируют клетку-хозяина, предлагаемую в изобретении (в которую интродуцирован рекомбинантный экспрессионный вектор, кодирующий полипептид IMI, или в геном которой интродуцирован ген, кодирующий полипептид дикого типа или полипептид IMI) в приемлемой среде до получения полипептида IMI. Согласно другому варианту осуществления изобретения способ заключается также в том, что выделяют полипептиды IMI из среды или из клетки-хозяина. Следующим объектом изобретения являются выделенные полипептиды IMI и их биологически активные фрагменты. Понятие “выделенный” или очищенный полипептид или его биологически активный фрагмент означает, что он не содержит какого-то клеточного материала при его получении с помощью методов рекомбинантной ДНК или химических предшественников или других химических веществ при его получении химическим синтезом. Выражение “практически не содержит клеточного материала” применимо к препаратам полипептида IMI, в которых полипептид отделен от каких-либо клеточных компонентов клеток, в которых его получают естественным путем или рекомбинантно. В этом варианте осуществления изобретения выражение “практически не содержит клеточного материала” относится к препаратам полипептида IMI, содержащим менее чем примерно 30% (в пересчете на сухую массу) не-IMI материала (обозначенного также в контексте настоящего описания как “загрязняющий (примесный) полипептид”), более предпочтительно менее чем примерно 20% не-IMI материала, еще более предпочтительно менее чем примерно 10% не-IMI материала и наиболее более предпочтительно менее чем примерно 5% не-IMI материала.
Когда полипептид IMI или его биологически активный фрагмент получают рекомбинантно, то он также предпочтительно практически не содержит культуральной среды, т.е. на долю культуральной среды приходится менее примерно 20%, более предпочтительно менее примерно 10% и наиболее предпочтительно менее примерно 5% от объема полипептидного препарата. Выражение “практически не содержит химических предшественников или других химических веществ” относится к препаратам полипептида IMI, в которых полипептид отделен от химических предшественников или других химических веществ, которые используют для синтеза полипептида. В этом варианте осуществления изобретения выражение “практически не содержит химических предшественников или других химических веществ” относится к препаратам полипептида IMI, в которых присутствует менее чем примерно 30% (в пересчете на сухую массу) химических предшественников или не-IMI химических веществ, более предпочтительно менее чем примерно 20% химических предшественников или не-IMI химических веществ, более предпочтительно менее чем примерно 10% химических предшественников или не-IMI химических веществ и наиболее предпочтительно менее чем примерно 5% химических предшественников или не-IMI химических веществ. В предпочтительных вариантах осуществления изобретения в выделенных полипептидах или их биологически активных фрагментах отсутствуют загрязняющие полипептиды из того организма, из которого получают полипептид IMI. Как правило, такие полипептиды получают путем рекомбинантной экспрессии, например полипептида IMI Triticum turgidum в растениях, отличных от Triticum turgidum, или в микроорганизмах, таких как С. glutamicum, реснитчатые, водоросли или грибы.
Полинуклеотидные и полипептидные последовательности IMI, предлагаемые в изобретении, можно применять для различных целей. Нуклеотидные и аминокислотные последовательности, предлагаемые в настоящем изобретении, можно использовать для трансформации растений, модулируя тем самым толерантность растений к имидазолиноновым гербицидам. Таким образом, изобретение относится к способу получения трансгенного растения, обладающего повышенной толерантностью к имидазолиноновому гербициду, который заключается в том, что (а) трансформируют растительную клетку одним или несколькими экспрессионными векторами, содержащими одну или несколько нуклеиновых кислот IMI, и (б) получают из растительной клетки трансгенное растение, которое обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа. Согласно одному из вариантов осуществления изобретения множество нуклеиновых кислот IMI получают из различных геномов. Настоящее изобретение относится также к способам получения трансгенного растения, обладающего повышенной толерантностью к имидазолиноновому гербициду, который заключается в том, что (а) трансформируют растительную клетку экспрессионным вектором, содержащим нуклеиновую кислоту IMI, где нуклеиновая кислота не представляет собой нуклеиновую кислоту ImI1, и (б) получают из растительной клетки трансгенное растение, которое обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа.
Настоящее изобретение относится также к способам модификации толерантности растений к имидазолиноновому гербициду, заключающемуся в том, что модифицируют экспрессию одной или нескольких нуклеиновых кислот IMI. Предпочтительно нуклеиновые кислоты локализованы в различных геномах или получены из различных геномов. Толерантность растений к имидазолиноновому гербициду можно повышать или понижать путем повышения или понижения соответственно экспрессии полинуклеотида IMI. Предпочтительно толерантность растений к имидазолиноновому гербициду повышают путем повышения экспрессии полинуклеотида IMI. Экспрессию полинуклеотида IMI можно модифицировать с помощью любого метода, известного специалистам в данной области. Можно применять способы повышения экспрессии полинуклеотида IMI, в которых используют либо трансгенное, либо нетрансгенное растение. В тех случаях, когда растение является трансгенным, растение можно трансформировать, например, вектором, содержащим любую из описанных выше кодирующих IMI нуклеиновых кислот, или растение можно трансформировать с помощью промотора, который обеспечивает экспрессию эндогенных полинуклеотидов IMI в растении. Согласно изобретению такой промотор может представлять собой тканеспецифичный или регулируемый стадией развития промотор. В альтернативном варианте в нетрансгенных растениях экспрессию эндогенного полинуклеотида IMI модифицируют с помощью индукции нативным промотором. Экспрессия полинуклеотидов, содержащих полинуклеотидную последовательность, которая представлена в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23, в растениях-мишенях может обеспечиваться одним из следующих факторов (но не ограничиваясь ими): (а) конститутивным промотором, (б) химически индуцируемым промотором и (в) созданной с помощью генной инженерии сверхэкспрессии промотора с использованием, например, полученных из “цинковых пальцев” факторов транскрипции (Greisman и Pabo, Science 275, 1997, с.657).
Согласно предпочтительному варианту осуществления изобретения транскрипцию полинуклеотида IMI модулируют с помощью факторов транскрипции “цинковых пальцев (ZFP) согласно методу, описанному у Greisman и Pabo, Science 275, 1997, с.657, и с помощью набора фирмы Sangamo Biosciences, Inc. Эти ZFP содержат как распознающий ДНК домен, так и функциональный домен, который вызывает активацию или подавление нуклеиновой кислоты-мишени, такой как нуклеиновая кислота IMI. Для этой цели можно создавать активирующие или подавляющие ZFP, которые специфично распознают описанные выше промоторы полинуклеотида IMI и используют для повышения или снижения экспрессии полинуклеотида IMI в растении, модулируя тем самым толерантность растения к гербицидам.
Как более подробно было описано выше, растения, полученные способами, предлагаемыми в настоящем изобретении, могут быть однодольными или двудольными. Растения можно выбирать из таких растений, как, например, кукуруза, пшеница, рожь, овес, тритикале, рис, ячмень, соя, арахис, хлопчатник, семенной рапс и канола, маниок, перец, подсолнечник, бархатцы, пасленовые культуры, такие как картофель, табак, баклажан и томат, виды рода Vicia (вика), горох, люцерна, кофе, какао, чай, виды семейства ивовых (Salix), масличная пальма, кокос, многолетние травы и кормовые культуры. Кормовые культуры включают (но не ограничиваясь ими) пырей, канареечник канарский, костер, дикую рожь, мятлик, ежу сборную, люцерну, салфойн (Salfoin), лядвинец рогатый, клевер красно-белый, клевер красный и сладкий клевер. В предпочтительном варианте осуществления изобретения растение представляет собой растение пшеницы. В каждом из описанных выше способов растительная клетка представляет (но не ограничиваясь ими) протопласт, гаметопродуцирующую клетку и клетку, из которой регенерируется целое растение. В контексте настоящего описания понятие “трансгенный” относится к любому растению, растительной клетке, каллюсу, растительной ткани или части растения, которые содержат всю или часть по меньшей мере одного рекомбинантного полинуклеотида. Во многих случаях весь рекомбинантной полинуклеотид или его часть стабильно интегрированы в хромосому или представляют собой стабильный внехромосомный элемент, такой, который переходит в следующие поколения.
Как указано выше, настоящее изобретение относится к композициям и способам повышения толерантности к имидазолинонам растения пшеницы или его семени по сравнению с сортом растения или семени дикого типа. В предпочтительном варианте осуществления изобретения толерантность к имидазолинонам растения пшеницы или его семени повышают так, что растение или семя могут выдерживать обработку имидазолиноновым гербицидом предпочтительно с использованием его нормы расхода примерно 10-400 г д.в. на га, более предпочтительно 20-160 г д.в. на га и наиболее предпочтительно 40-80 г д.в. на га. В контексте настоящего описания понятие “выдерживать” обработку имидазолиноновым гербицидом означает, что растение либо не погибает, либо не повреждается при такой обработке.
Кроме того, в настоящем изобретении предложен способ борьбы с сорняками вблизи растений пшеницы или растений тритикале, заключающийся в том, что обрабатывают имидазолиноновым гербицидом сорняки и растения пшеницы или тритикале, где растение пшеницы или тритикале обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом дикого типа растения пшеницы или тритикале и где растение пшеницы или тритикале содержит одну или несколько нуклеиновых кислот IMI. Согласно одному из вариантов осуществления изобретения растение пшеницы или тритикале содержит множество нуклеиновых кислот IMI, локализованных в различных геномах или полученных из различных геномов, где нуклеиновые кислоты IMI выбирают из группы, включающей нуклеиновую кислоту Imi 1, нуклеиновую кислоту Imi 2 и нуклеиновую кислоту Imi 3. В другом варианте осуществления изобретения растение содержит нуклеиновую кислоту Imi 2 и нуклеиновую кислоту Imi 3. В результате придания растениям пшеницы и тритикале повышенной толерантности к имидазолинону можно применять широкое разнообразие композиций для защиты растений пшеницы и тритикале от сорняков, что способствует росту растений и снижает конкуренцию за питательные вещества. Имидазолиноновый гербицид можно применять индивидуально для предвсходовой, послевсходовой, предпосевной обработки и обработки в процессе посева с целью борьбы с сорняками в областях, окружающих растения пшеницы, указанные в изобретении, или можно применять препаративную форму имидазолинонового гербицида, которая содержит другие добавки. Имидазолиноновый гербицид можно использовать также для обработки семян. Добавки, входящие в состав препаративной формы имидазолинонового гербицида, включают другие гербициды, поверхностно-активные вещества, адъюванты, вещества, повышающие смачивающую способность, прилипатели, стабилизаторы или т.п. Препаративная форма имидазолинонового гербицида может представлять собой мокрый или сухой препарат и может представлять собой (но не ограничиваясь ими) текучие порошки, эмульгирующиеся концентраты и жидкие концентраты. Имидазолиноновый гербицид и препаративные формы гербицида можно применять с помощью общепринятых методов, например, путем опрыскивания, орошения, опыливания или т.п.
В настоящем описании приведены ссылки на различные публикации. Описание всех этих публикаций и ссылок, процитированных в этих публикациях, во всей их полноте включено в настоящее описание в качестве ссылки для того, чтобы более подробно описать уровень техники, к которой относится изобретение.
Следует понимать также, что предыдущее описание относится к предпочтительным вариантам осуществления настоящего изобретения и что можно осуществлять многочисленные изменения без отклонения от объема изобретения. Ниже изобретение проиллюстрировано на примерах, которые никоим образом не ограничивают его объем. Напротив, следует ясно понимать, что можно реализовывать различные другие варианты осуществления изобретения, его модификации и эквиваленты, которые после изучения настоящего описания специалисты в данной области могут предложить сами без отклонения от сущности настоящего изобретения и/или объема формулы изобретения.
Примеры
Пример 1
Мутагенез и селекция толерантных линий пшеницы дурум
Толерантные к имидазолинону линии пшеницы получали путем мутации и последующих стандартных методов селекции. Сначала мутагенез семян вызывали путем обработки семян пшеницы сорта дурум с использованием 3 или 3,5 мл ЭМС (этилметансульфонат) на литр в течение 2 ч и предварительного замачивания в проточной воде в течение 5,5 ч, после чего их промывали дистиллированной водой. В процессе ЭМС-обработки семена встряхивали через каждые 10-15 мин. После 2-часовой ЭМС-обработки мутаген сливали и заменяли фосфатным буфером (0,001М, рН 3,5). Затем семена обрабатывали азидом натрия (2 мл/л 1М маточного раствора), в процессе обработки семена встряхивали периодически через 1 ч. Жидкость сливали и семена дважды промывали дистиллированной водой, спускали воду и выкладывали для сушки на поддоны в теплице в течение 24-36 ч, после чего высаживали в полевых условиях во влажную почву.
Растения поколения M1, полученные из обработанных семян, собирали полностью и высаживали полученные семена растений поколения М2. Растения поколения М2 обрабатывали гербицидом Raptor с использованием нормы расхода 10 унций/акр (88,6 г имазамокса/га) на стадии трех настоящих листьев. Растения, выжившие после обработки гербицидом, переносили в теплицу для получения семян поколения М3.
Линии М2:3 подвергали скринингу в теплице с использованием либо 10 унций/акр (88,6 г имазамокса/га), либо 12 унций/акр (106,3 г имазамокса/га) гербицида Raptor. Гербицид применяли на стадии трех настоящих листьев. Семена М3:4 получали из наиболее толерантных растений поколения М3.
Пример 2
Толерантность линий пшеницы дурум к имидазолиноновым гербицидам
Полевые опыты (1):
Девять линий М4:6, полученных из четырех линий растений М2 (СI19, CI32, CI37, СО12) оценивали в опыте, проведенном с дублированием в одной области выращивания пшеницы дурум в Италии. 75 г/га имазамокса применяли для обработки 21-25 растений на стадии роста ВВСН. Как правило, норма расхода 35 г/га приводила к практически 100%-ной гибели чувствительных растений пшеницы. Воздействие на выращиваемую культуру (общее повреждение) варьировалось от 0 до 13% через 21 день после обработки (ДПО) и от 0 до 17% через 43 ДПО. Урожай (в %) от урожая необработанной линии варьировался в пределах от 85 до 102%.
Полевые опыты (2):
110 линий М3:4, выведенных из 60 растений поколения М2, подвергали скринингу с использованием норм расхода имазомокса 71 г/га и 160 г/га. Количество линий М4 на растение М2 варьировалось от 1 до 20. Толерантность растений линий М3:4 была сравнима с толерантностью растений той же самой линии на необработанных делянках, а также сорта растений дикого типа на обработанных делянках, из которого были выведены некоторые из указанных растений. Результаты обобщены в таблице 1.
Все тестированные линии выживали при обеих нормах расхода гербицида, в то время как все растения дикого типа погибали при обеих нормах расхода. На основе сравнения с линией дикого типа все тестированные линии характеризовались выраженной толерантностью к применяемым нормам расхода, прежде всего, в отношении уменьшения высоты растения по отношению к растениям на необработанных делянках. Норма расхода 35 г/га имазамокса приводила к гибели чувствительных растений пшеницы дурум. Таким образом, тестированные линии оказались толерантными к нормам, превышающим указанную норму от почти 3 до более чем 4 раз.
Оценка толерантности в баллах линий М3:4, выведенных из 14 различных растений поколения М2, обработанных двумя различными нормами расхода имазамокса
2,3Цифры представляют собой нормы расхода гербицида в г/га
Цифры в таблице представляют собой диапазон реакций линий М3:4, выведенных из каждого указанного растения поколения М2
Опыт в теплице
В опыте в теплице проводили оценку 15 линий пшеницы дурум, каждая из которых была получена из различных растений поколения М2, и двух линий пшеницы дикого типа в отношении толерантности к имидазолиноновому гербициду имазамоксу, применяемому в нормах расхода 100 и 160 г/га. Оценки толерантности проводили на 14 и 21 день после обработки. Повреждение оценивали по 0-9-балльной шкале, где 0 соответствовал отсутствию повреждений и 9 соответствовал гибели растения. Результаты обобщены в таблице 2.
У всех линий была выявлена более высокая толерантность по сравнению с линиями дикого типа, проявившаяся в том, что все растения дикого типа погибли через 21 день после обработки, т.е. к тому времени, когда даже линии, у которых через 14 дней были обнаружены серьезные повреждения, начинали восстанавливаться. Норма расхода имазамокса 35 г/га приводила к гибели чувствительных растений пшеницы дурум. Таким образом, все 15 линий, выведенные путем мутагенеза, обладали очень высокой толерантностью по отношению к имазамоксу.
Осредненные баллы повреждения растений потомства, выведенного из 15 растений пшеницы дурум поколения М2 и одной линии пшеницы дурум дикого типа, обработанных двумя различными нормами расхода
Числа в таблице представляют собой средние значения, полученные для 24 растений для конкретного вида обработки. Растения оценивали по 0-9-балльной шкале, где 0 соответствует отсутствию повреждений и 9 соответствует гибели растения.
Пример 3
Биохимическая основа толерантности
Фермент, который является мишенью для имидазолиноновых гербицидов, представляет собой синтазу ацетогидроксикислот (AHAS), первый каталитический фермент в пути биохимического синтеза аминокислот с разветвленной цепью валина, лейцина и изолейцина. Предполагается, что гербицид связывается с сайтами в “кармане” фермента, но не связывается с активным сайтом.
Активность in vitro фермента AHAS, экстрагированного из растения, можно оценивать биохимическими методами. В таблице 3 проиллюстрировано влияние добавления различных концентраций имидазолинонового гербицида, такого как имазамокс, на активность белка AHAS, экстрагированного из растений пшеницы дурум дикого типа (линия UT). Даже при добавлении относительно небольших концентраций активность AHAS быстро снижалась.
В таблице 3 приведены также данные о активности AHAS в нескольких толерантных к имидазолиноновому гербициду линий, полученных из М2. Ингибирование активности оказалось существенно более низким при использовании более низких концентраций имазамокса и даже при использовании наиболее высоких концентраций активность, как правило, составляла от одной трети до одной второй от активности в контроле. Эти данные в совокупности с данными о толерантности, полученными в теплице и в полевых опытах, по-видимому, подтверждают гипотезу о том, что изменение в результате мутагенеза по меньшей мере одного гена AHAS в геноме пшеницы дурум приводит к образованию белка AHAS с пониженной способностью ингибироваться имазамоксом.
Результаты анализа активности AHAS in vitro, выраженные в процентах по отношению к контролю, для 13 линий пшеницы дурум и контрольной линии дикого типа (UT) при воздействии различных концентраций имазамокса
Пример 4
Молекулярная основа толерантности
Определение характеристик на молекулярном уровне толерантных к имидазолинону линий подтвердило наличие специфических мутаций в генах, кодирующих фермент AHAS (Als 2 и Als 3). Толерантная к имидазолинону линия СI19 имела замену пары оснований (гуанина на аденин) в гене Als 2, что приводило к замене серина на аспарагин в домене Е фермента AHAS. Линия СI19 не содержала никаких мутаций в гене Als 3. Аналогично этому толерантные к имидазолинону линии UT01, UT03, UT05, UT07, UT08, UT10, UT13, UT14, UT16, UT17, UT20 все содержали последовательность гена Als 3 дикого типа и замену пары оснований (гуанина на аденин) в гене Als 2.
Толерантная к имидазолинону линия UT12 содержала в гене Als 3 замену пары оснований (гуанина на аденин), что приводило к замене серина на аспарагин в домене Е фермента AHAS. Линия UT12 не содержала никаких мутаций в гене Als 2.
Толерантные к имидазолинону линии Utopia UT15 и UT19 содержали в гене Als 3 новые мутации, а именно замену пары оснований (гуанина на аденин), что приводило к замене аминокислоты аланина на треонин в N-концевой области фермента AHAS. Толерантная к имидазолинону линия Utopia UT15 содержала также в гене Als 2 замену пары оснований (тимина на цитозин), что не приводило к аминокислотной замене.
Пример 5
Создание толерантных к имидазолинону растений пшеницы
Толерантные к имидазолинону растения пшеницы получали методом, описанным у Ishida и др., Natura Biotech., 14, 1996, cc.745-750. Незрелые зародыши размером 1-2 мм выделяли через 10-15 дней после опыления и стерилизовали этанолом и 30%-ным раствором хлорокса. Незрелые зародыши заражали клетками Agrobacterium, несущими представляющую интерес конструкцию в векторе фирмы Japan Tobacco на LS-среде для заражения и затем на LS-среде для совместного культивирования в течение 3-7 дней (все среды получали от Japan Tobacco) согласно методу, описанному у Ishida и др., Natura Biotech., 14, 1996, cc.745-750. Затем эксплантаты переносили на LS-среду, содержащую от 0,05 до 0,1 мкМ PURSUIT®, и культивировали при слабом освещении в течение 1-2 недель. Активно растущие каллюсы переносили для 2-й и 3-й селекции на LS-среду, дополненную 0,5-1,0 мкМ имазетапиром (PURSUIT®), и культивировали в течение 2-3 недель. После 3-й селекции каллюсы переносили на среду для регенерации, дополненную 0,25-0,75 мкМ имазетапиром (PURSUIT®), на три недели. Затем проростки переносили на LS-среду для выращивания половинной крепости и культивировали в течение трех недель перед пересадкой в почву и выращиванием в теплице. Предполагаемые трансгенные растения опрыскивали имазамоксом (25-50 г/га) (RAPTOR®) для уничтожения одичавших культурных растений.















































Растения пшеницы или тритикале, содержащие одну или несколько нуклеиновых кислот IMI Triticum turgidum, обладают повышенной толерантностью к имидазолиноновому гербициду. Раскрываются также семена, образующиеся на указанных растениях пшеницы и тритикале, и способы борьбы с сорняками вблизи этих растений пшеницы и растений тритикале. 20 н. и 49 з.п. ф-лы, 23 ил., 3 табл.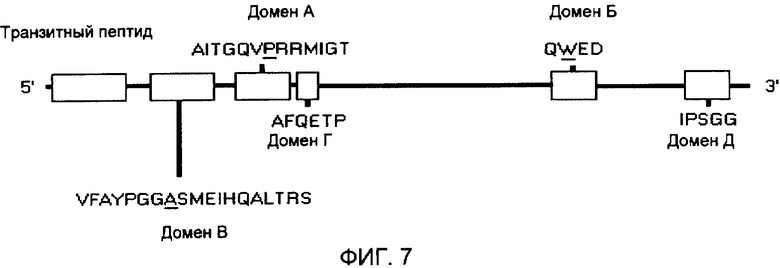
1. Растение пшеницы, содержащее по меньшей мере одну нуклеиновую кислоту IMI Triticum turgidum, выбранную из группы, включающей:
(а) нуклеиновую кислоту Imi 2 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене Е, приводящую к замене серина на аспарагин в белке IMI по сравнению с белком AHAS дикого типа;
(б) нуклеиновую кислоту Imi 3 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене Е, приводящую к замене серина на аспарагин в белке IMI по сравнению с белком AHAS дикого типа;
(в) нуклеиновую кислоту Imi 3 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене С, приводящую к замене аланина на треонин в белке IMI по сравнению с белком AHAS дикого типа;
где нуклеиновая кислота IMI Triticum turgidum обусловливает повышенную толерантность растений к имидазолиноновому гербициду по сравнению с сортом растения дикого типа, причем это растение пшеницы является трансгенным растением или растением, полученным с помощью способа, включающего мутагенез.
2. Растение пшеницы по п.1, где растение содержит нуклеиновую кислоту Imi 2 Triticum turgidum пo (a).
3. Растение пшеницы по п.1, где растение содержит нуклеиновую кислоту Imi 3 Triticum turgidum по (б).
4. Растение пшеницы по п.1, где растение содержит нуклеиновую кислоту Imi 3 Triticum turgidum пo (в).
5. Растение пшеницы по п.1, где нуклеиновая кислота IMI Triticum turgidum представляет собой нуклеиновую кислоту IMI подвидов Durum.
6. Растение пшеницы по п.1, где растение содержит первую нуклеиновую кислоту IMI Triticum turgidum и вторую нуклеиновую кислоту IMI Triticum turgidum, причем первая нуклеиновая кислота IMI Triticum turgidum представляет собой нуклеиновую кислоту IMI Triticum turgidum (а) и вторая нуклеиновая кислота IMI Triticum turgidum представляет собой нуклеиновую кислоту IMI Triticum turgidum (б) или (в).
7. Растение пшеницы по п.1, где нуклеиновая кислота IMI Triticum turgidum содержит полинуклеотидную последовательность, выбранную из группы, включающей:
(I) полинуклеотиды, содержащие нуклеотидную последовательность, представленную в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23;
(II) полинуклеотиды, кодирующие полипептид, который содержит аминокислотную последовательность, указанную в SEQ ID NO:2, SEQ ID NO:6 или SEQ ID NO:24;
(III) полинуклеотиды, кодирующие белок IMI, аминокислотная последовательность которого по меньшей мере на 95% идентична полной аминокислотной последовательности SEQ ID NO:2, где белок IMI содержит мутацию в домене Е, которая приводит к замене серина аспарагином в белке IMI по сравнению с AHAS белком дикого типа;
(IV) полинуклеотиды, кодирующие белок IMI, аминокислотная последовательность которого по меньшей мере на 95% идентична полной аминокислотной последовательности SEQ ID NO:6 или SEQ ID NO:24, где белок IMI содержит мутацию в домене С, которая приводит к замене аланина треонином в белке IMI по сравнению с AHAS белком дикого типа.
8. Растение пшеницы по п.1, где нуклеиновая кислота Imi 2 Triticum turgidum содержит полинуклеотидную последовательность, представленную в SEQ ID NO:1.
9. Растение пшеницы по п.1, где нуклеиновая кислота Imi 3 Triticum turgidum содержит полинуклеотидную последовательность, представленную в SEQ ID NO:3.
10. Растение пшеницы по п.1, где нуклеиновая кислота Imi 3 Triticum turgidum содержит полинуклеотидную последовательность, представленную в SEQ ID NO:5 или SEQ ID NO:23.
11. Растение пшеницы по п.1, содержащее нуклеиновые кислоты IMI Triticum turgidum.
12. Растение пшеницы по п.1, содержащее нуклеиновую кислоту Imi 2 Triticum turgidum и нуклеиновую кислоту Imi 3 Triticum turgidum.
13. Растение пшеницы по п.1, где имидазолиноновый гербицид выбирают из группы, включающей 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-3-хинолинкарбоновую кислоту, 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-метилникотиновую кислоту и смесь метил-6-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-мета-толуата и метил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-пара-толуата и их смеси.
14. Растение пшеницы по п.1, где имидазолиноновый гербицид представляет собой 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту.
15. Растение пшеницы по п.1, где имидазолиноновый гербицид представляет собой 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновую кислоту.
16. Часть растения пшеницы по п.1, где часть растения включает по меньшей мере одну нуклеиновую кислоту IMI.
17. Растительная клетка растения пшеницы по п.1, где растительная клетка включает по меньшей мере одну нуклеиновую кислоту IMI.
18. Семя, образовавшееся на растении пшеницы по п.1, где это семя содержит по меньшей мере одну нуклеиновую кислоту IMI.
19. Семя по п.18 для получения гомозигот с повышенной толерантностью к имидазолиноновому гербициду по сравнению с семенем растения пшеницы сорта дикого типа.
20. Растение пшеницы, включающее по меньшей мере одну нуклеиновую кислоту, выбранную из группы, которая включает
(I) полинуклеотиды, содержащие нуклеотидную последовательность, представленную в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23;
(II) полинуклеотиды, кодирующие полипептид, который содержит аминокислотную последовательность, указанную в SEQ ID NO:2, SEQ ID NO:6 или SEQ ID NO:24,
и где растение пшеницы обладает толерантностью к гербицидам, которая характерна для растения любой из линий UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19, UT20 или CI19, репрезентативные образцы семян которых депонированы в АТСС (Американская коллекция типовых культур) для целей патентования под регистрационным номером РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914. РТА-4915. РТА-4916. РТА-4917, РТА-4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922, РТА-4923 или РТА-4960, где
(а) растение пшеницы является рекомбинантным или созданным с помощью генной инженерии производным растения любой линии UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19;
(б) растение пшеницы, являющееся любым потомком растения любой линии UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19; или
(в) растение пшеницы, являющееся потомком любого из растений, указанных в (а)-(в).
21. Растение пшеницы по п.20, где растение пшеницы представляет собой растение пшеницы вида Triticum turgidum.
22. Растение пшеницы по п.20, где растение обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа.
23. Растение пшеницы по п.22, где имидазолиноновый гербицид выбирают из группы, включающей 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-3-хинолинкарбоновую кислоту, 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-метилникотиновую кислоту и смесь метил-6-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-мета-толуата и метил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-пара-толуата и их смеси.
24. Растение пшеницы по п.22, где имидазолиноновый гербицид представляет собой 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту.
25. Растение пшеницы по п.22, где имидазолиноновый гербицид представляет собой 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновую кислоту.
26. Часть растения пшеницы по п.20, где часть растения обладает признаками толерантности к гербицидам.
27. Растительная клетка растения пшеницы по п.20, где растительная клетка обладает признаками толерантности к гербицидам.
28. Семя, образовавшееся на растении пшеницы по п.20, где семя обладает признаками толерантности к гербицидам.
29. Семя по п.28 для получения гомозигот с повышенной толерантностью к имидазолиноновому гербициду по сравнению с семенем растения пшеницы сорта дикого типа.
30. Растение тритикале, содержащее по меньшей мере одну нуклеиновую кислоту IMI Triticum turgidum, выбранную из группы, включающей:
(а) нуклеиновую кислоту Imi 2 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене Е, приводящую к замене серина на аспарагин в белке IMI по сравнению с белком AHAS дикого типа;
(б) нуклеиновую кислоту Imi 3 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене Е, приводящую к замене серина на аспарагин в белке IMI по сравнению с белком AHAS дикого типа;
(в) нуклеиновую кислоту Imi 3 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене С, приводящую к замене аланина на треонин в белке IMI по сравнению с белком AHAS дикого типа;
где нуклеиновая кислота IMI Triticum turgidum обусловливает повышенную толерантность растений к имидазолиноновому гербициду по сравнению с сортом растения дикого типа, причем это растение тритикале является трансгенным растением или растением, полученным с помощью способа, включающего мутагенез.
31. Растение тритикале по п.30, где растение содержит нуклеиновую кислоту Imi 2 Triticum turgidum пo (a).
32. Растение тритикале по п.30, где растение содержит нуклеиновую кислоту Imi 3 Triticum turgidum по (б).
33. Растение тритикале по п.30, где растение содержит нуклеиновую кислоту Imi 3 Triticum turgidum по (в).
34. Растение тритикале по п.30, где нуклеиновая кислота IMI Triticum turgidum представляет собой нуклеиновую кислоту IMI подвидов Durum.
35. Растение тритикале по п.30, где растение содержит первую нуклеиновую кислоту IMI Triticum turgidum и вторую нуклеиновую кислоту IMI Triticum turgidum, первая нуклеиновая кислота IMI Triticum turgidum представляет собой нуклеиновую кислоту IMI Triticum turgidum (а), и вторая нуклеиновая кислота IMI Triticum turgidum представляет собой нуклеиновую кислоту IMI Triticum turgidum (б) или (в).
36. Растение тритикале по п.30, где нуклеиновая кислота IMI Triticum turgidum содержит полинуклеотидную последовательность, выбранную из группы, включающей:
(I) полинуклеотиды, содержащие последовательность, представленную в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23;
(II) полинуклеотиды, кодирующие полипептид, содержащий аминокислотную последовательность, указанную в SEQ ID NO:2, SEQ ID NO:6 или SEQ ID NO:24;
(III) полинуклеотид, кодирующий белок IMI, аминокислотная последовательность которого по меньшей мере на 95% идентична полной аминокислотной последовательности SEQ ID NO:2, где белок IMI содержит мутацию в домене Е, которая приводит к замене серина аспарагином в белке IMI, по сравнению с AHAS белком дикого типа;
(IV) полинуклеотид, кодирующий белок IMI, аминокислотная последовательность которого по меньшей мере на 95% идентична полной аминокислотной последовательности SEQ ID NO:6 или SEQ ID NO:24, где белок IMI содержит мутацию в домене С, которая приводит к замене аланина треонином в белке IMI по сравнению с AHAS белком дикого типа.
37. Растение тритикале по п.36, где нуклеиновая кислота Imi 2 Triticum turgidum содержит полинуклеотидную последовательность, представленную в SEQ ID NO:1.
38. Растение тритикале по п.36, где нуклеиновая кислота Imi 3 Triticum turgidum содержит полинуклеотидную последовательность, представленную в SEQ ID NO:3.
39. Растение тритикале по п.36, где нуклеиновая кислота Imi 3 Triticum turgidum содержит полинуклеотидную последовательность, представленную в SEQ ID NO: 5 или SEQ ID NO:23.
40. Растение тритикале по п.36, содержащее две нуклеиновые кислоты IMI Triticum turgidum.
41. Часть растения тритикале по п.30, где часть растения включает по меньшей мере одну нуклеиновую кислоту IMI.
42. Растительная клетка растения тритикале по п.30, где растительная клетка включает по меньшей мере одну нуклеиновую кислоту IMI.
43. Семя, образовавшееся на растении тритикале по п.30, где семя содержит по меньшей мере одну нуклеиновую кислоту IMI Triticum turgidum.
44. Семя по п.42 для получения гомозигот с повышенной толерантностью к имидазолиноновому гербициду по сравнению с семенем растения тритикале сорта дикого типа.
45. Растение тритикале, включающее по меньшей мере одну нуклеиновую кислоту, выбранную из группы, которая содержит
(I) полинуклеотиды, включающие нуклеотидную последовательность, представленную в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23;
(II) полинуклеотиды, кодирующие полипептид, который содержит аминокислотную последовательность, указанную в SEQ ID NO:2, SEQ ID NO:6 или SEQ ID NO:24,
и где растение тритикале обладает толерантностью к гербицидам, которая характерна для растения, обладающего толерантностью к гербицидам, характерным для растения любой из линий UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19, UT20 или CI19, репрезентативные образцы семян которых депонированы в АТСС (Американская коллекция типовых культур) для целей патентования под регистрационным номером РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4916, РТА-4917, РТА-4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922, РТА-4923 или РТА-4960, где
(а) растение тритикале является рекомбинантным или созданным с помощью генной инженерии производным растения любой из линий UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19, и Т20 или С119;
(б) растение тритикале, являющееся любым потомком растения любой из линий UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19, и Т20 или С119; или
(в) растение тритикале, являющееся потомком любого из растений, указанных в (а)-(б).
46. Растение тритикале по п.45, где растение обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа.
47. Часть растения тритикале по п.45, где часть растения обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа.
48. Растительная клетка растения тритикале по п.45, где растение обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа.
49. Семя, образовавшееся на растении тритикале по п.45, где семя обладает признаками толерантности к гербицидам.
50. Семя по п.49 для получения гомозигот, обладающих повышенной толерантностью к имидазолиноновому гербициду по сравнению с семенем растения тритикале сорта дикого типа.
51. Выделенная нуклеиновая кислота IMI, где нуклеиновая кислота содержит полинуклеотид, выбранный из группы, включающей:
(а) полинуклеотиды, содержащие нуклеотидную последовательность, представленную в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23;
(б) полинуклеотиды, кодирующие полипептид, который содержит аминокислотную последовательность, указанную в SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:24; и
(в) полинуклеотиды, кодирующие белок IMI, аминокислотная последовательность которого по меньшей мере на 95% идентична полной аминокислотной последовательности SEQ ID NO:2, где белок IMI содержит мутацию в домене Е, которая приводит к замене серина аспарагином в белке IMI, по сравнению с AHAS белком дикого типа;
(г) полинуклеотиды, кодирующие белок IMI, аминокислотная последовательность которого по меньшей мере на 95% идентична полной аминокислотной последовательности SEQ ID NO:6 или SEQ ID NO:24, где белок IMI содержит мутацию в домене С, которая приводит к замене аланина треонином в белке IMI по сравнению с AHAS белком дикого типа;
(д) полинуклеотиды, комплементарные любому полинуклеотиду, указанному выше в (а)-(г),
где нуклеиновая кислота IMI или ее комплемент кодирует белок IMI.
52. Выделенная нуклеиновая кислота IMI по п.51, где нуклеиновая кислота содержит полинуклеотид, представленный в SEQ ID NO:1.
53. Выделенная нуклеиновая кислота IMI по п.51, где нуклеиновая кислота содержит полинуклеотид, представленный в SEQ ID NO:3.
54. Выделенная нуклеиновая кислота IMI по п.51, где нуклеиновая кислота содержит полинуклеотид, представленный в SEQ ID NO:5 или SEQ ID NO:23.
55. Выделенная нуклеиновая кислота IMI по п.51, где нуклеиновая кислота содержит полинуклеотид, кодирующий полипептид, который представлен в SEQ ID NO:2.
56. Выделенная нуклеиновая кислота IMI по п.51, где нуклеиновая кислота содержит полинуклеотид, кодирующий полипептид, который представлен в SEQ ID NO:6 или 24.
57. Способ борьбы с сорняками вблизи растения, заключающийся в том, что имидазолиноновый гербицид наносят на сорняки и на растение, где растение обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа, и где растение содержит по меньшей мере одну нуклеиновую кислоту IMI Triticum turgidum, выбранную из группы, включающей:
(а) нуклеиновую кислоту Imi 2 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене Е, приводящую к замене серина на аспарагин в белке IMI по сравнению с белком AHAS дикого типа;
(б) нуклеиновую кислоту Imi 3 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене Е, приводящую к замене серина на аспарагин в белке IMI по сравнению с белком AHAS дикого типа;
(в) нуклеиновую кислоту Imi 3 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене С, приводящую к замене аланина на треонин в белке IMI по сравнению с белком AHAS дикого типа, причем это растение является трансгенным растением или растением, полученным с помощью способа, включающего мутагенез.
58. Способ по п.57, где растение содержит две нуклеиновые кислоты IMI Triticum turgidum.
59. Способ по п.57, где растение содержит нуклеиновую кислоту Imi 2 Triticum turgidum и нуклеиновую кислоту Imi 3 Triticum turgidum.
60. Способ по п.57, где имидазолиноновый гербицид выбирают из группы, включающей 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-3-хинолинкарбоновую кислоту, 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-метилникотиновую кислоту и смесь метил-6-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-мета-толуата и метил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-пара-толуата или их смеси.
61. Способ по п.57, где имидазолиноновый гербицид представляет собой 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту.
62. Способ по п.57, где имидазолиноновый гербицид представляет собой 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновую кислоту.
63. Способ по п.57, где нуклеиновая кислота IMI Triticum turgidum выбрана из группы, включающей:
(I) полинуклеотиды, содержащие нуклеотидную последовательность, представленную в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23;
(II) полинуклеотиды, кодирующие полипептид, включающий аминокислотную последовательность, указанную в SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:24;
(III) полинуклеотиды, кодирующие белок IMI, аминокислотная последовательность которого по меньшей мере на 95% идентична полной аминокислотной последовательности SEQ ID NO:2, где белок IMI содержит мутацию в домене Е, которая приводит к замене серина аспарагином в белке IMI по сравнению с AHAS белком дикого типа;
(IV) полинуклеотиды, кодирующие белок IMI, аминокислотная последовательность которого по меньшей мере на 95% идентична полной аминокислотной последовательности SEQ ID NO:6, где белок IMI содержит мутацию в домене С, которая приводит к замене аланина треонином в белке IMI по сравнению с AHAS белком дикого типа.
64. Способ получения трансгенного растения, обладающего повышенной толерантностью к имидазолиноновому гербициду, заключающийся в том, что:
(а) трансформируют растительную клетку одним или несколькими экспрессионными векторами, содержащими по меньшей мере одну нуклеиновую кислоту IMI Triticum turgidum; и
(б) получают из растительной клетки трансгенное растение, обладающее повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа;
где нуклеиновая кислота IMI Triticum turgidum выбрана из группы, включающей:
(а) нуклеиновую кислоту Imi 2 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене Е, приводящую к замене серина на аспарагин в белке IMI по сравнению с белком AHAS дикого типа;
(б) нуклеиновую кислоту Imi 3 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене Е, приводящую к замене серина на аспарагин в белке IMI по сравнению с белком AHAS дикого типа;
(в) нуклеиновую кислоту Imi 3 Triticum turgidum, кодирующую белок IMI, который содержит мутацию в домене С, приводящую к замене аланина на треонин в белке IMI по сравнению с белком AHAS дикого типа.
65. Способ по п.64, в котором нуклеиновая кислота IMI Triticum turgidum содержит полинуклеотидную последовательность, выбранную из группы, включающей:
(i) полинуклеотиды, включающие нуклеотидную последовательность, представленную в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23;
(ii) полинуклеотиды, кодирующие полипептид, представленный в SEQ ID NO:2, SEQ ID NO:6 или SEQ ID NO:24;
(iii) полинуклеотиды, кодирующие белок IMI, аминокислотная последовательность которого по меньшей мере на 95% идентична полной аминокислотной последовательности SEQ ID NO:2, где белок IMI содержит мутацию в домене Е, которая приводит к замене серина аспарагином в белке IMI по сравнению с AHAS белком дикого типа;
(iv) полинуклеотиды, кодирующие белок IMI, аминокислотная последовательность которого по меньшей мере на 95% идентична полной аминокислотной последовательности SEQ ID NO:6, где белок IMI содержит мутацию в домене С, которая приводит к замене аланина треонином в белке IMI по сравнению с AHAS белком дикого типа.
66. Способ борьбы с сорняками вблизи растения, заключающийся в том, что имидазолиноновый гербицид наносят на сорняки и на растение, где растение обладает повышенной толерантностью к имидазолиноновому гербициду по сравнению с сортом растения дикого типа, и где растение обладает признаками устойчивости к гербицидам растения, характерного для растения, любой из линий UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19, UT20 или CI19, репрезентативные образцы семян которых депонированы в АТСС (Американская коллекция типовых культур) для целей патентования под регистрационным номером РТА-4910, РТА-4911, РТА-4912, РТА-4913, РТА-4914, РТА-4915, РТА-4916, РТА-4917, РТА-4918, РТА-4919, РТА-4920, РТА-4921, РТА-4922, РТА-4923 или РТА-4960, где
(а) растение является растением пшеницы любой из линий UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19, UT20 или CI19;
(б) растение является растением пшеницы, рекомбинантным или созданным с помощью генной инженерии производным растения, любой из линий UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19, UT20 или CI19;
(в) растение является любым потомком растения пшеницы любой из линий UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19, UT20 или CI19;
(г) растение является растением тритикале, рекомбинантным или созданным с помощью генной инженерии производным растения любой из линий UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19, UT20 или CI19;
(д) растение является любым потомком растения тритикале любой из линий UT01, UT03, UT05, UT07, UT08, UT10, UT12, UT13, UT14, UT15, UT16, UT17, UT19, UT20 или СI19; или
(е) растение является потомком любого из растений, указанных в (а)-(д),
где указанное растение включает по меньшей мере одну нуклеиновую кислоту
IMI, выбранную из группы, включающей
(I) полинуклеотиды, содержащие нуклеотидную последовательность, представленную в SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:23; и
(II) полинуклеотиды, кодирующие полипептид, включающий аминокислотную последовательность, представленную в SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6 или SEQ ID NO:24;
67. Способ по п.66, где имидазолиноновый гербицид выбирают из группы, включающей 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-3 хинолинкарбоновую кислоту, 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновую кислоту, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-метилникотиновую кислоту и смесь метил-6-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-мета-толуата и метил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-пара-толуата или их смеси.
68. Способ по п.66, где имидазолиноновый гербицид представляет собой 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)никотиновую кислоту.
69. Способ по п.66, где имидазолиноновый гербицид представляет собой 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)никотиновую кислоту.
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |

