Группа изобретений относится к биотехнологии, в частности к генетической инженерии, и предназначена для биосинтеза в клетках млекопитающих полноразмерного химерного антитела, обеспечивающего экстренную профилактику мышей от вирусного клещевого энцефалита.
Вирус клещевого энцефалита, представитель семейства Flaviviridae, является высокопатогенным для человека вирусным агентом, который способен вызывать заболевание, приводящее к серьезным поражениям нервной системы [1].
В настоящее время для этиотропной терапии клещевого энцефалита применяют в основном сывороточный иммуноглобулин, получаемый из плазмы крови доноров, проживающих в природных очагах заболевания [2]. Этот препарат обладает выраженным терапевтическим эффектом особенно при среднетяжелом и тяжелом течении заболевания, причем введение препарата в 1-2 день после укуса обеспечивает значительно больший лечебный эффект, чем введение в последующие дни. Вместе с тем, препарат обладает определенными недостатками, в частности, он дефицитен, что связанно с ограниченным источником исходного материала, и содержит относительно низкий уровень специфических вируснейтрализующих антител. Кроме того, использование препаратов человеческой крови сопровождается известным биологическим риском.
В последние годы сывороточные антитела заменяют рекомбинантными антителами, получаемыми в культурах клеток. Среди рекомбинантных антител на фармацевтическом рынке преобладают химерные антитела - полноразмерные иммуноглобулины, в которых к константным доменам иммуноглобулинов человека присоединены вариабельные домены мышиных моноклональных антител (МКА) [3, 4]. При конструировании химерных антител против вируса клещевого энцефалита ключевым этапом является выбор МКА, вариабельные домены которого обладают высоким сродством к антигену и обепечивают высокиее противовирусные свойства.
Известны аналоги векторных плазмид, используемых для экспрессии химерных антител и полностью человеческих антител. К таким аналогам относятся плазмидные ДНК рСН37 и pCL37 (патент RU 2317330 С2, оп. 20.02.2008), предназначенные для экспрессии человеческого антитела против вируса осповакцины; плазмидные ДНК рСН1 и pCL1 (патент RU 2285043 С2, оп. 10.10.2006), предназначенные для экспрессии человеческого антитела против вируса Эбола; плазмидные ДНК pH1-HD и pL-HD2 (патент US 4975369 A, оп. 04.12.1990 и SU 1780541 A3, оп. 07.12.1992), предназначенные для экспрессии химерного антитела KS1/4 против антигена Ер-САМ клеток аденокарциномы человека; плазмидные ДНК pSVgptHu2VHPLAP-HuIgG1 и pSVneoHuVkPLAP-HuCk, предназначенные для экспрессии гуманизированного антитела против плацентарной щелочной фосфатазы человека (патент WO 199107500 А1, оп. 30.05.1991 и патент RU 2102479 C1, оп. 20.01.1998). Сведения о плазмидных ДНК, предназначенных для продукции химерных или гуманизированных антител против вируса клещевого энцефалита, в общедоступных источниках информации отсутствуют.
Ранее были получены и охарактеризованы МКА против гликопротеина Е - основного иммуногенного белка вируса клещевого энцефалита, причем некоторые из них, в частности антитело 14D5, способны ингибировать инфекционность вируса на культуре эукариотических клеток [5].
Наиболее близким аналогом к заявляемой группе изобретений -прототипом, является химерное антитело ch13D6, обладающее сродством к гликопротеину Е вируса клещевого энцефалита с константой аффинности 1,4·10-8 М и способное нейтрализовать инфекционность вируса клещевого энцефалита на культуре клеток [6]. Антитело ch13D6 получают в системе экспрессии, основанной на эукариотической линии НЕК293Т и двух плазмидных ДНК pD6H и pD6L, содержащих гены тяжелой и легкой цепей химерного антитела человека/мыши класса IgG1-каппа. Однако полученное химерное антитело не способно защищать модельных животных от инфекции вирусом клещевого энцефалита.
Задачей группы изобретений является получение двух полипептидов со свойствами полноразмерных легкой и тяжелой цепей иммуноглобулина человека, образующих в клетках млекопитающих химерное антитело человека/мыши класса IgG1/каппа, обладающее более высоким сродством к гликопротеину Е вируса клещевого энцефалита, а также обеспечивающее экстренную профилактику мышей от вирусного клещевого энцефалита в дозировке, не превышающей 1 мг на кг веса животного.
Технический результат: получение гликопротеина иммуноглобулинового типа, обеспечивающего экстренную профилактику мышей от вирусного клещевого энцефалита в дозировке, не превышающей 1 мг на кг веса животного.
Указанный результат достигается путем конструирования двух рекомбинантных плазмидных ДНК, одна из которых, pCLm4/hygro-14D5, кодирует синтез полипептида со свойствами легкой цепи полноразмерного химерного антитела, другая, pCHm2-14D5, - кодирует синтез полипептида со свойствами тяжелой цепи химерного антитела. Совместная трансфекция плазмидами pCLm4/hygro-14D5 и pCHm2-14D5 эукариотических клеток линии СНО-K1, обеспечивает синтез полипептида со свойствами химерного антитела, обеспечивающее экстренную профилактику мышей от вирусного клещевого энцефалита. Транзиентный биосинтез целевых полипептидов обеспечивается наличием в плазмидных ДНК pCLm4/hygro-14D5 и pCHm2-14D5 цитомегаловирусного промотора и сайта полиаденилирования гена бычьего гормона роста.
Сущность заявляемой группы изобретений заключается в следующем.
Генно-инженерными методами получают плазмиды pCLm4/hygro-14D5 и pCHm2-14D5, которые содержат искусственные гены легкой и тяжелой цепей полноразмерного химерного антитела, созданные на основе вариабельных фрагментов легких и тяжелых цепей мышиного моноклонального антитела 14D5 против вируса клещевого энцефалита и генов, кодирующих константные домены иммуноглобулинов человека класса IgG1/каппа, а также содержат цитомегаловирусный промотор и сайт полиаденилирования гена бычьего гормона роста. Сконструированные плазмидные ДНК при одновременном введении в эукариотические клетки обусловливают биосинтез полипептидов со свойствами легкой и тяжелой цепей антитела человека, которые объединяются в химерное антитело, направленное против вируса клещевого энцефалита и обеспечивающее экстренную профилактику мышей от вирусного клещевого энцефалита.
Исходным генетическим материалом для конструирования целевых плазмид служат следующие генно-инженерные исходные конструкции:
а) суммарная РНК, выделенная из гибридомных клеток, продуцирующих мышиное моноклональное антитело 14D5 [5];
б) суммарная РНК, выделенная из суммарной клеточной фракции крови здорового донора;
в) плазмидная ДНК pCL37, содержащая ген легкой цепи полноразмерного моноклонального антитела человека 1F4 против вируса осповакцины, включающая лидерную последовательность гена легкой цепи антител мыши, участок гена, кодирующего константный домен легких цепей человеческих антител класса каппа [7];
г) плазмидная ДНК рСН37, содержащая ген тяжелой цепи полноразмерного моноклонального антитела человека 1F4 против вируса осповакцины, включающая лидерную последовательность гена тяжелой цепи антитела мыши, участки генов, кодирующие константные CH1-CH2-CH3 - домены тяжелых цепей человеческих антител класса IgG1 [7];
д) плазмидная ДНК pcDNA™3.1/Hygro(+) (Invitrogen);
е) олигонуклеотидные праймеры для конструирования плазмидных
ДНК, содержащих гены легкой и тяжелой цепи антитела человека против
вируса клещевого энцефалита (в направлении 5′-3′).
1. 5' СТТ CCG GAA TTC SAR GTN MAG CTG SAG SAG TC
2. 5' GGA AGA TCT ATA GAC AGA TGG GGG TGT CGT TTT GGC
3. 5' CC GAA TTC GAY ATT GTG MTS ACM CAR WCT MCA
4. 5' GG GAA GCT TGA TAC AGT TGG TGC AGC АТС AGC
5. 5' GGC TTA TCG AAA TTA ATA CG
6. 5' TAG AAG GCA CAG TCG AGG
7. 5' GAC AAA ACT CAC АСА TGC
8. 5' CAT GAG GGT GTC СТТ GGG
9. 5' GAA АТС AAA CGT ACT GTG GCT GCA
10. 5' TGC AGC CAC AGT ACG TTT GAT TTC
11. 5' GG GAT АТС GTG ATG ACC CAG TCC CC
12. 5' CCG TTT GAT CTC AAG СТТ GGT CCC
13. 5' GAG GTG CAG CTG CTC GAG TCT GG
14. 5' GAC GGT GAC CAG GGT ACC CTG GCC
15. 5' CGC TCT AGA TCA TTT ACC CGG GGA CAG GGA GAG
16. 5' A TGC AAG CTT GAG АТС AAA CGG ACT GTG GCT GCA CCA TCT GTC
17. 5' TCG AGG АТС CCT AAC ACT CTC CCC TGT TGA AGC
18. 5' С CAG GGT ACC CTG GTC ACC GTC TCC TCA GCC TCC ACC AAG GGC CCA Т
19. 5' A TGC TCT AGA TCA TTT ACC CGG GGA CAG GGA
Полученная в результате плазмида pCLm4/hygro-14D5, кодирующая синтез в клетках млекопитающих полипептида со свойствами легкой цепи полноразмерного химерного антитела, характеризуется следующими признаками:
- имеет молекулярную массу 4,11 МДа и размер 6232 п.н.;
- кодирует гибридный белок, в котором вариабельный домен легкой цепи мышиного моноклонального антитела 14D5 соединен с константным доменом каппа-цепи иммуноглобулинов человека;
- состоит из следующих элементов:
а) NheI / ApaI - векторного фрагмента плазмиды pcDNA™3.1/Hygro(+) (Invitrogen) размером 5491 п.н., содержащего промотор-энхансер цитомегаловируса, сайт полиаденилирования и участок терминации транскрипции гена бычьего гормона роста, ген β-лактамазы (bla), ген гигромицин фосфотрансферазы (hpt);
б) NheI / ApaI - фрагмента промежуточной плазмиды pCLm4-14D5 размером 741 п.н., включающего искусственный ген, кодирующий полипептид со свойствами легкой цепи полноразмерного химерного антитела, осуществляющего протекцию против вируса клещевого энцефалита.
- содержит:
а) цитомегаловирусный (CMV) промотор и энхансер транскрипции;
б) искусственный ген, кодирующий гибридный белок, в котором вариабельный домен легкой цепи мышиного моноклонального антитела 14D5 соединен с константным доменом каппа-цепи иммуноглобулинов человека;
в) в качестве генетических маркеров ген β-лактамазы (bla), определяющий устойчивость трансформированных плазмидой pCLm4/hygro-14D5 клеток бактерий к ампициллину, и ген гигромицин фосфотрансферазы (hpt), обеспечивающий устойчивость к гигромицину В для селекции трансфицированных плазмидой pCLm4/hygro-14D5 клеток млекопитающих;
г) уникальные сайты узнавания эндонуклеазами рестрикции, имеющие следующие координаты: BglII - 13, NheI - 896, EcoRV - 980, HindIII - 1284, ApaI - 1637.
Полученная в результате плазмида pCHm2-14D5, кодирующая синтез в клетках млекопитающих полипептида со свойствами тяжелой цепи полноразмерного химерного антитела, характеризуется следующими признаками:
- имеет молекулярную массу 4,47 МДа и размер 6765 п.н.;
- кодирует гибридный белок, в котором вариабельный домен тяжелой цепи мышиного моноклонального антитела 14D5 соединен с константной частью тяжелой цепи иммуноглобулинов человека класса IgG1;
- состоит из следующих элементов:
а) NheI / XbaI - векторного фрагмента плазмиды pcDNA™3.1 (+) (Invitrogen) размером 5332 п.н., содержащего промотор-энхансер цитомегаловируса (CMV), сайт полиаденилирования и участок терминации транскрипции гена бычьего гормона роста (BGH), ген β-лактамазы (bla), ген устойчивости к неомицину (neo);
б) NheI / XhoI - фрагмента промежуточной плазмиды pCHm2 размером 104 п.н., содержащего лидерную последовательность одного из VH-генов мыши;
в) XhoI / Acc65I - фрагмента размером 312 п.н., кодирующего вариабельный домен мышиного моноклонального антитела 14D5;
г) Acc65I / XbaI - фрагмента промежуточной плазмиды pCHm2, включающего участок, кодирующий CH1-, CH2-, CH3 - домены иммуноглобулинов человека класса IgG1.
- содержит:
а) цитомегаловирусный (CMV) промотор и энхансер транскрипции;
б) искусственный ген, кодирующий гибридный белок, в котором вариабельный домен тяжелой цепи мышиного моноклонального антитела 14D5 соединен с константной частью тяжелой цепи иммуноглобулинов IgGI человека;
в) в качестве генетических маркеров ген β-лактамазы (bla), определяющий устойчивость трансформированных плазмидой pCHm2-14D5 клеток бактерий к ампициллину и ген устойчивости к неомицину (neo) для селекции трансфецированных плазмидой pCHm2-14D5 клеток млекопитающих;
г) уникальные сайты узнавания эндонуклеазами рестрикции, имеющие следующие координаты: BglII - 14, NheI - 897, XhoI - 1001, Acc65I - 1313, XbaI - 2330.
Транзиентную экспрессию химерного антитела ch14D5 осуществляют в клетках млекопитающих линии СНО-K1, трансфицированных одновременно плазмидами pCLm4/hygro-14D5 и pCHm2-14D5, обеспечивающими синтез полипептидов со свойствами полноразмерных легкой и тяжелой цепей химерного антитела, обеспечивающее экстренную профилактику мышей от вирусного клещевого энцефалита.
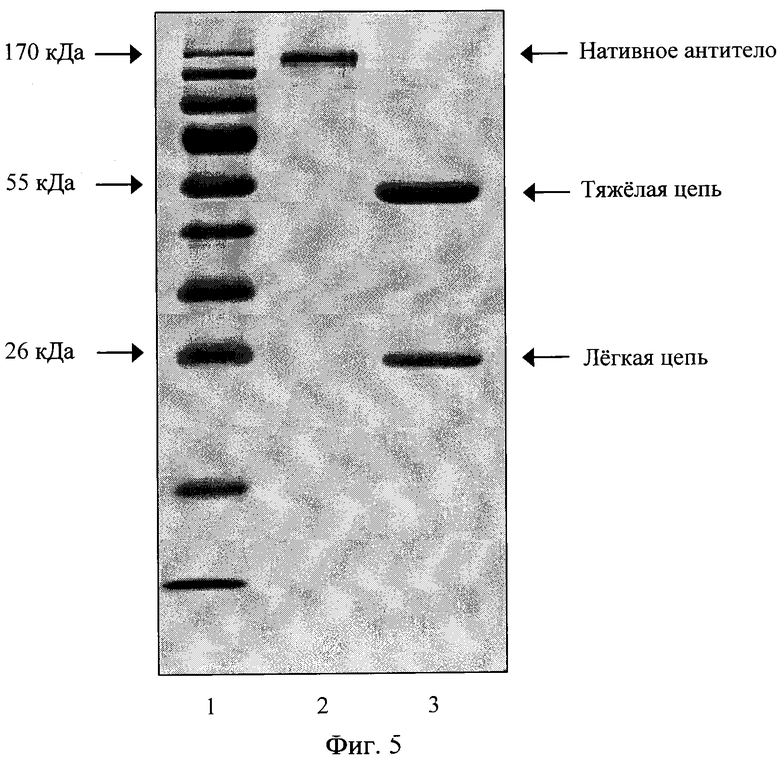
Целевое химерное антитело ch14D5 выделяют из культуральной жидкости аффинной хроматографией на сорбенте с ковалентно иммобилизованным белком A Staphylococcus aureus. Выделенное химерное антитело анализируют гель-электрофорезом в денатурирующих условиях в присутствии и в отсутствии 2-меркаптоэтанола.
Полученное химерное антитело ch14D5 обладает способностью связывать поверхностный белок Е вируса клещевого энцефалита [8] с константой аффинности 6,0·10-11 М, которую определяют методом поверхностного плазменного резонанса с использованием прибора ProteOn XPR36 (Bio-Rad).
Полученное химерное антитело ch14D5, введенное в дозировке, не превышающей 1 мг на кг веса животного, обеспечивает экстренную профилактику мышей от вирусного клещевого энцефалита, что подтверждается в экспериментах in vivo.
Таким образом, впервые получены плазмидные ДНК pCLm4/hygro-14D5 и pCHm2-14D5, обеспечивающие синтез полипептидов со свойствами полноразмерных легкой и тяжелой цепей химерного антитела, способного связывать поверхностный белок Е вируса клещевого энцефалита с константой аффинности 6,0·10-11 М и обеспечивать экстренную профилактику мышей от вирусного клещевого энцефалита при введении химерного антитела ch14D5 в дозировке, не превышающей 1 мг на кг веса животного.
Изобретение иллюстрируется следующими фигурами:
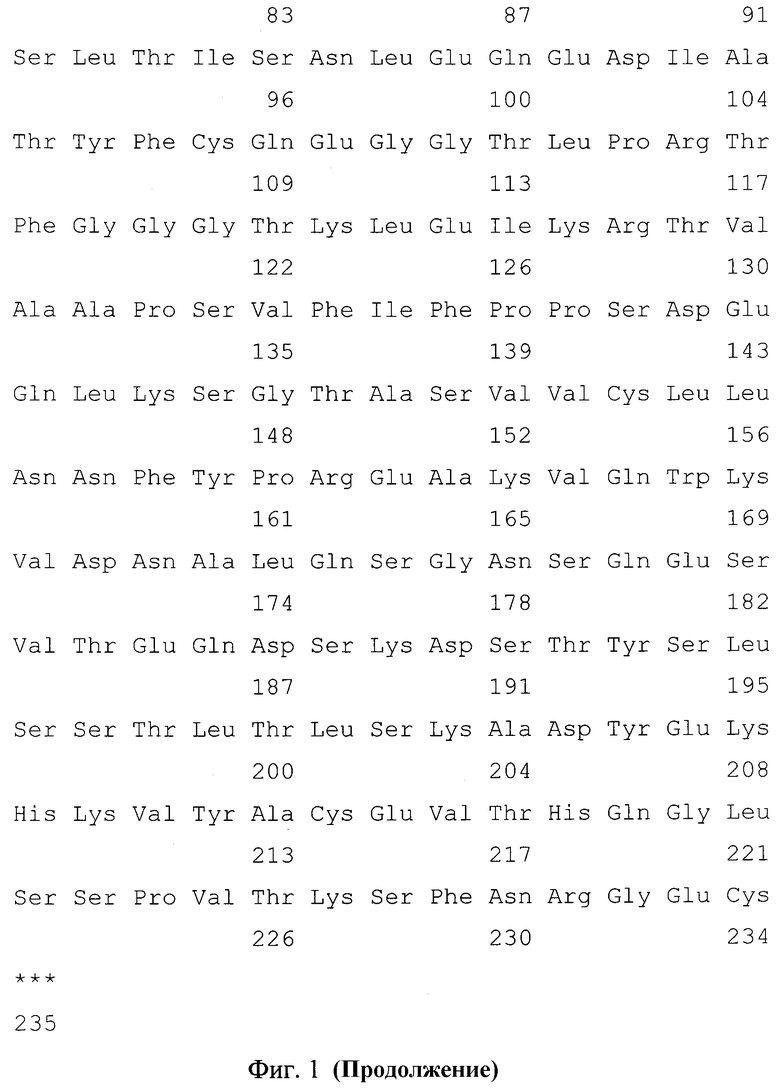
Фиг. 1. Нуклеотидная и кодируемая ею аминокислотная последовательность NheI/ApaI-фрагмента плазмиды pCLm4/hygro-14D5, кодирующая полипептид со свойствами легкой цепи химерного антитела против вируса клещевого энцефалита. Подчеркнуты сайты узнавания эндонуклеаз рестрикции, жирным шрифтом выделены инициирующий и терминирующий кодоны.
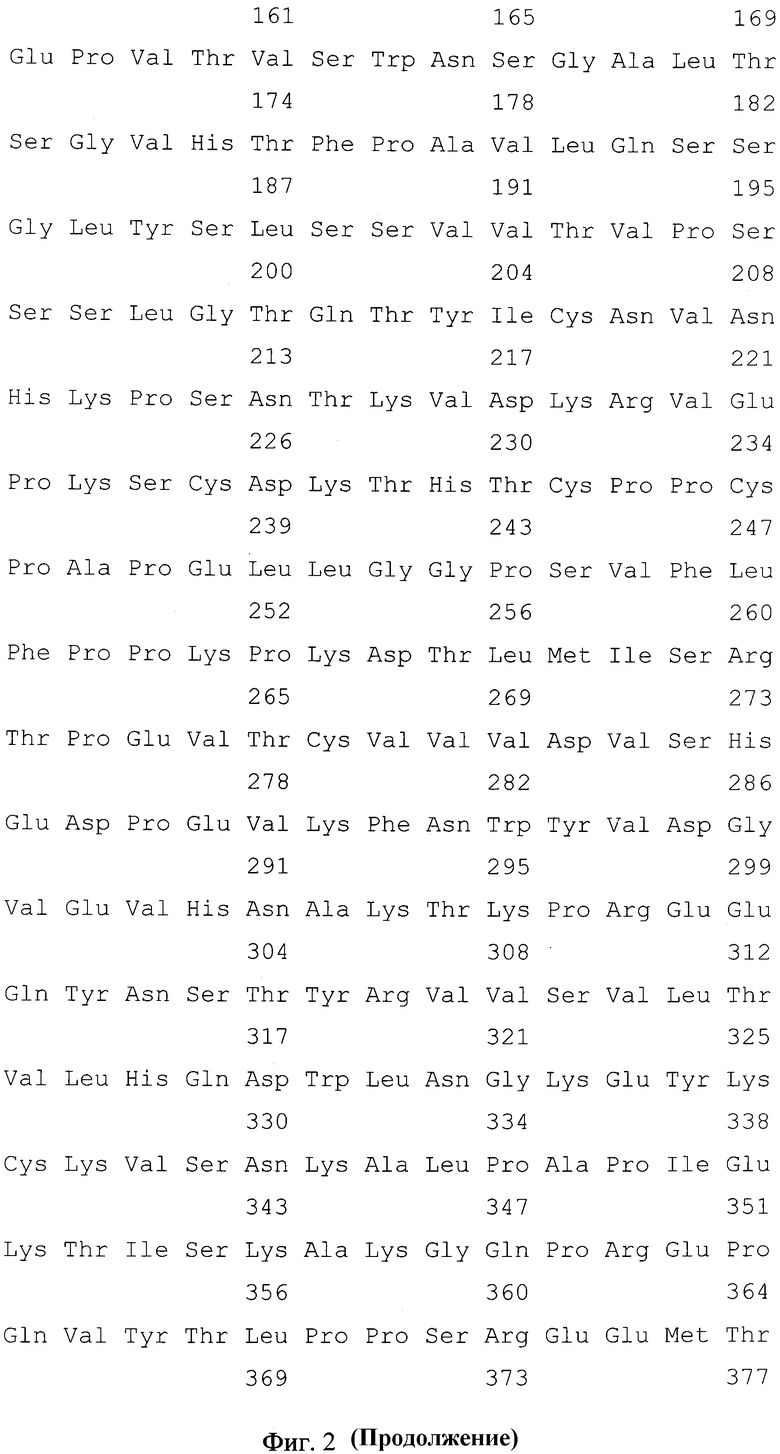
Фиг. 2. Нуклеотидная и кодируемая ею аминокислотная последовательность NheI/XbaI-фрагмента плазмиды pCHm2-14D5, кодирующая полипептид со свойствами тяжелой цепи химерного антитела против вируса клещевого энцефалита. Подчеркнуты сайты узнавания эндонуклеаз рестрикции, жирным шрифтом выделены инициирующий и терминирующий кодоны.
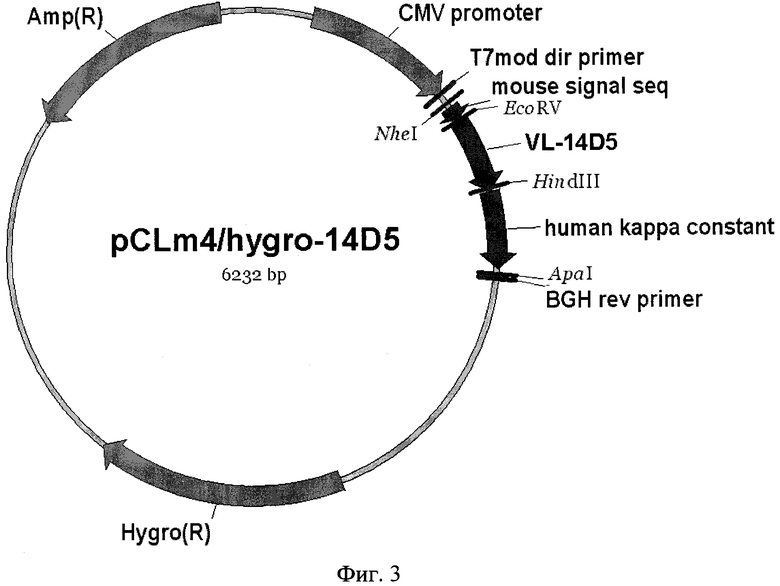
Фиг. 3. Карта плазмиды pCLm4/hygro-14D5. Указаны сайты эндонуклеаз рестрикции. Pcmv - цитомегаловирусный промотор; S - сигнальная последовательность; VL, kappa_const - вариабельный и константный участки гена легкой цепи рекомбинантного антитела; BGHpA - сайт полиаденилирования гена бычьего гормона роста.
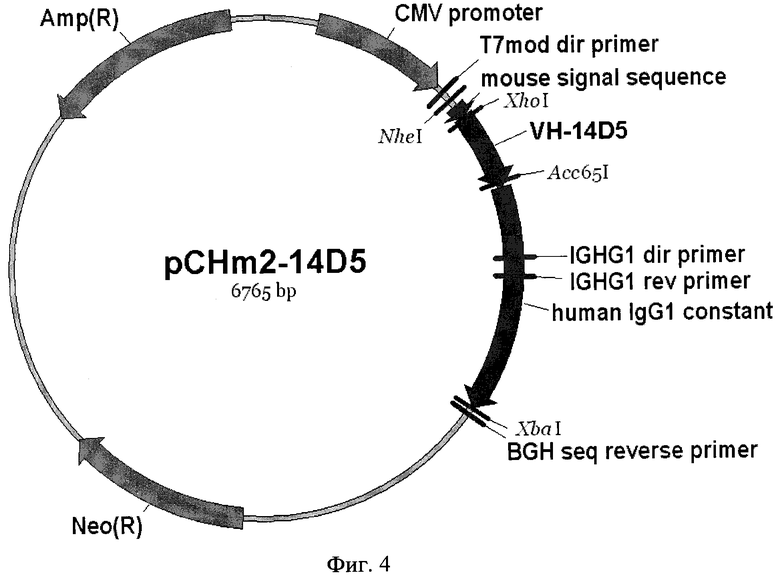
Фиг. 4. Карта плазмиды pCHm2-14D5. Указаны сайты эндонуклеаз рестрикции. Pcmv - цитомегаловирусный промотор; S - сигнальная последовательность; VH, IgG1_const. - вариабельный и константный участки гена тяжелой цепи рекомбинантного антитела; BGHpA - сайт полиаденилирования бычьего гормона роста.
Фиг. 5. Электрофореграмма очищенного химерного антитела в денатурирующем 15% полиакриламидном геле. Дорожки: 1 - маркер молекулярных масс, 2 - целевой белок, не обработанный 2-меркаптоэтанолом, 3 - целевой белок, обработанный 2-меркаптоэтанолом.
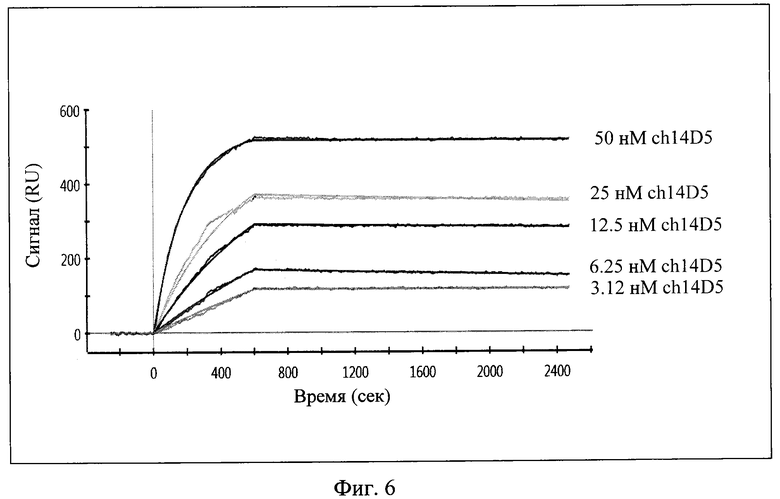
Фиг. 6. График, иллюстрирующий кинетику связывания химерного антитела ch14D5 в различных концентрациях с рекомбинантным белком Е [8]. Фрагмент 1-600 сек - фаза связывания антитела, 600-2500 сек - фаза диссоциации антитела.
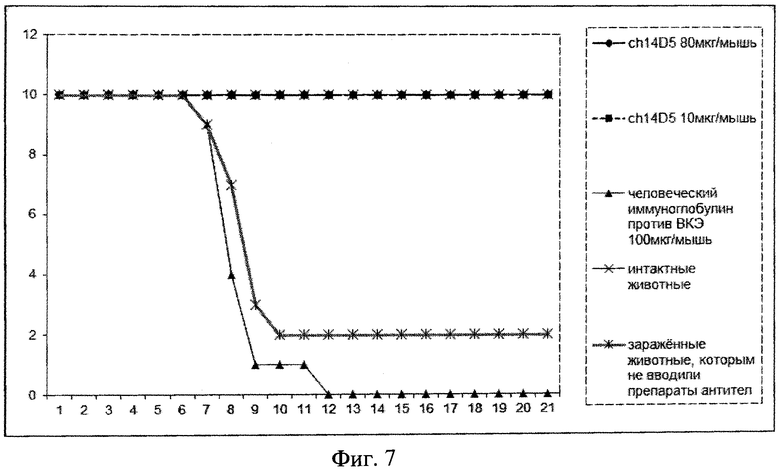
Фиг. 7. График, иллюстрирующий выживаемость мышей, инфицированных вирусом клещевого энцефалита, штамм «Абсеттаров», в дозировке 240 ЛД5о, в зависимости от введения химерного антитела ch14D5. По горизонтальной оси указаны дни после инфекции, по вертикальной - число живых мышей.
Для лучшего понимания сущности предлагаемых изобретений, они иллюстрируются следующими примерами конкретного осуществления.
Пример 1. Получение фрагментов ДНК, кодирующих вариабельные домены мышиного моноклонального антитела 14D5, и определение их нуклеотидных последовательностей.
Из гибридомных клеток, продуцирующих моноклональное антитело 14D5, выделяют суммарную РНК любым доступным способом, например, с использованием набора "RNeasy Mini Kit" (Qiagen).
На основе выделенной РНК в реакции обратной транскрипции (ОТ) -полимеразной цепной реакции (ГЩР) с использованием олигонуклеотидных праймеров 1 и 2 и набора "One step RT-PCR Kit" (Qiagen) получают фрагмент ДНК VH-14D5, кодирующий вариабельный домен тяжелой цепи мышиного моноклонального антитела 14D5, и аналогично с использованием олигонуклеотидных праймеров 3 и 4 получают фрагмент ДНК VL-14D5, кодирующий вариабельный домен легкой цепи мышиного моноклонального антитела 14D5. Продукты ОТ-ПЦР разделяют электрофоретически с использованием агарозного геля, и элюируют из геля любым доступным образом, например, с использованием набора "GeneJET™ Gel Extraction Kit" (Fermentas).
Нуклеотидные последовательности полученных фрагментов ДНК VH-14D5 и VL-14D5 определяют любым доступным способом, например, с использованием набора "BigDye® Terminator v3.1" (Applied Biosystems) и автоматического капиллярного секвенатора ABI 3730XL Genetic Analyser (Applied Biosystems). Нуклеотидные последовательности фрагментов ДНК VH-14D5 и VL-14D5, кодирующих вариабельные домены тяжелой и легкой цепей мышиного моноклонального антитела 14D5 представлены на фиг. 1 и фиг. 2.
Пример 2. Получение фрагментов ДНК, кодирующих константный домен тяжелой цепи иммуноглобулинов человека класса IgG1 и константный домен легкой цепи иммуноглобулинов человека класса каппа.
Из 1 мл крови здорового донора выделяют суммарную РНК любым доступным способом, например, с использованием набора "RNeasy Mini Kit" (Qiagen).
На основе выделенной РНК в реакции ОТ-ПЦР с использованием олигонуклеотидных праймеров 16 и 17 и набора "One step RT-PCR Kit" (Qiagen) получают фрагмент ДНК IGKC, кодирующий константный домен легкой цепи класса каппа иммуноглобулинов человека. Процедуру проводят согласно инструкции производителя, отжиг праймеров осуществляют при 55°C. Аналогично с использованием олигонуклеотидных праймеров 18 и 19 получают фрагмент ДНК IGHG1, кодирующий константный домен тяжелой цепи иммуноглобулинов человека класса IgG1. Продукты ОТ-ПЦР разделяют электрофоретически с использованием 1% агарозного геля и вырезают фрагмент геля с ДНК, соответствующей по подвижности 1000 п.н. в случае IGHG1 и 340 п.н. в случае IGKC. Элюируют ДНК из геля любым доступным образом, например, с использованием набора "GeneJET™ Gel Extraction Kit" (Fermentas).
Нуклеотидные последовательности полученных фрагментов ДНК IGHG1 и IGKC определяют любым доступным способом, например, с использованием набора "BigDye® Terminator v3.1" (Applied Biosystems) и автоматического капиллярного секвенатора ABI 3730XL Genetic Analyser (Applied Biosystems). Нуклеотидные последовательности фрагментов ДНК IGHG1 и IGKC, кодирующих константные домены тяжелой цепи иммуноглобулинов человека класса IgG1 и легкой цепи иммуноглобулинов человека класса каппа представлены на фиг. 1 и фиг. 2.
Пример 3. Конструирование плазмидной ДНК pCLm4/hygro-14D5, содержащей ген, который кодирует легкую цепь химерного антитела ch14D5.
Конструирование плазмидной ДНК pCLm4/hygro-14D5 состоит из нескольких этапов:
1. Модификация исходной плазмиды pCL37 с целью введения уникальных сайтов узнавания эндонуклеаз рестрикции. Для этого плазмидную ДНК гидролизуют эндонуклеазами BamHI и XbaI, достраивают 5′-выступающие концы векторного фрагмента до тупых при помощи Pfu-полимеразы и объединяют полученные концы с использованием ДНК-лигазы бактериофага Т4. Полученную промежуточную плазмиду pCL37deltal используют в качестве матрицы в ПЦР, которую проводят в 50 мкл реакционной смеси, содержащей 0,5 ед.акт. Taq ДНК-полимеразы (Fermentas), 0,5 ед.акт. Pfu ДНК-полимеразы (Fermentas), 12,5 пмоль каждого олигонуклеотида, 0,2 мМ каждого дезоксинуклеозидтрифосфата, 75 мМ Трис-HCl (рН 8,8 при 25°C), 20 мМ (NH4)2SO4, 0,01% детергента tween-20, 2,5 мМ MgSO4. Проводят 30 циклов по схеме: 95°C/30 сек, 52°C/30 сек, 72°C/50 сек. При этом с использованием пары праймеров 5 и 10, а также пары праймеров 6 и 9 получают два перекрывающихся фрагмента ДНК, которые после выделения используют в реакции совместной объединяющей ПЦР с образованием единого фрагмента ДНК light_delta2, представляющего собой фрагмент плазмиды pCL37, из которого удалены сайты HindIII, XhoI и XmaJI. Далее методом мегапраймера [9, 10] в ДНК-фрагмент light_delta2 вводят сайты узнавания эндонуклеаз EcoRV и HindIII в области, соответствующие концам VL-участка, кодирующего вариабельный домен легкой цепи антитела. Для этого методом ПЦР с праймеров 5 и 12 по матрице light_delta2 получают фрагмент ДНК, который после очистки достраивают по матрице light_delta2 и амплифицируют в присутствии олигонуклеотидов 5 и 6 с образованием фрагмента light_mod3. Методом ПЦР с праймеров 6 и 11 по матрице light_mod3 получают фрагмент ДНК, который после очистки достраивают по матрице light_mod3 и амплифицируют в присутствии олигонуклеотидов 5 и 6 с образованием фрагмента light_mod4. Этот фрагмент объединяют с исходной плазмидой pCL37, гидролизованной по сайтам NheI и ApaI, в реакции лигирования с образованием плазмиды pCLm4.
2. ДНК-фрагмент IGKC, полученный ранее, обрабатывают эндонуклеазами рестрикции HindIII и BamHI и лигируют с векторным фрагментом, полученным в результате гидролиза плазмиды pCLm4 этими же эндонуклеазами. В результате получают плазмидную ДНК pCLm4-IGKC.
3. Встраивание в плазмиду pCLm4 фрагмента ДНК VL-14D5, кодирующего вариабельный домен мышиного моноклонального антитела 14D5. Фрагмент VL-14D5, полученный ранее, амплифицируют методом ПНР в присутствии праймеров 11 и 12. Полученный фрагмент обрабатывают эндонуклеазами рестрикции EcoRV и HindIII и лигируют с векторным фрагментом, полученным в результате гидролиза плазмиды pCLm4-IGKC этими же эндонуклеазами. В результате получают плазмидную ДНК pCLm4-14D5.
4. Клонирование гена, кодирующего легкую цепь химерного антитела ch14D5, в плазмидную ДНК pcDNA™3.1/Hygro(+). Плазмидную ДНК pCLm4-14D5 обрабатывают эндонуклеазами NheI и ApaI, в результате чего образуется фрагмент размером 741 п.н. Этот фрагмент лигируют с векторным фрагментом ДНК, образующимся при гидролизе плазмиды pcDNA™3.1/Hygro(+) этими же эндонуклеазами. Правильность встраивания подтверждают секвенированием с использованием любого доступного метода, например, с помощью набора "BigDye® Terminator v3.1" (Applied Biosystems) и автоматического капиллярного секвенатора ABI 3730XL Genetic Analyser (Applied Biosystems). В результате получают плазмидную ДНК pCLm4/hygro-14D5, карта которой представлена на фиг. 3.
Пример 4. Конструирование плазмиды pCHm2-14D5, содержащей ген, кодирующий тяжелую цепь химерного антитела ch14D5.
Конструирование плазмиды pCHm2-14D5 проводят в несколько этапов:
1. В векторную плазмиду рСН37 вводят сайты эндонуклеаз рестрикции XhoI и Асс651. Для этого в ПЦР по матрице рСН37 с праймеров 5 и 14 получают фрагмент ДНК sigVH. Далее методом ПЦР с использованием праймеров 13 и 14 на основе матрицы sigVH получают фрагмент ДНК, который после очистки достраивают и амплифицируют в присутствии олигонуклеотидов 5 и 14 с образованием фрагмента sigVHmod. Методом ПЦР с использованием в качестве матрицы ДНК плазмиды рСН37 в присутствии праймеров 5 и 15 получают фрагмент h-pA. Затем на основе матрицы h-pA методом мегапраймера в присутствии праймеров 5 и 15 достраивают фрагмент sigVHmod до фрагмента hm2a, который содержит ген тяжелой цепи антитела с введенными по концам VH-участка сайтами эндонуклеаз XhoI и Асс651. Фрагмент hm2a объединяют с исходной плазмидой рСН37, гидролизованной по сайтам эндонуклеаз NheI и XbaI, в реакции лигирования с образованием плазмиды pCHm2.
2. ДНК-фрагмент IGHG1, полученный ранее, обрабатывают эндонуклеазами рестрикции Acc65I и XbaI и лигируют с векторным фрагментом, полученным в результате гидролиза плазмиды pCHm2 этими же эндонуклеазами. В результате получают плазмидную ДНК pCHm2-IGHG1.
3. Фрагмент VH-14D5, полученный ранее, амплифицируют методом ПЦР в присутствии праймеров 13 и 14. Полученный ПНР-фрагмент обрабатывают эндонуклеазами рестрикции XhoI и Acc65I и лигируют с векторным фрагментом, полученным в результате гидролиза плазмиды pCHm2-IGHG1 этими же эндонуклеазами. Правильность встраивания подтверждают секвенированием с использованием любого доступного метода, например, с помощью набора "BigDye® Terminator v3.1" (Applied Biosystems) и автоматического капиллярного секвенатора ABI 3730XL Genetic Analyser (Applied Biosystems). В результате получают плазмидную ДНК pCHm2-14D5, карта которой представлена на фиг. 4.
Пример 5. Получение полноразмерного химерного антитела ch14D5 в эукариотических клетках млекопитающих.
Транзиентную экспрессию полноразмерного химерного антитела против вируса клещевого энцефалита осуществляют в результате ко-трансфекции клеток яичников китайского хомячка линии СНО-К1 полученными экспрессионными плазмидами pCLm4/hygro-14D5 и pCHm2-14D5, содержащими гены легкой и тяжелой цепей антитела. Перед трансфекцией клетки СНО-К1 культивируют при 37°C в атмосфере 5% CO2 в среде DMEM (Gibco), содержащей 10% бычьей фетальной сыворотки. Клетки высевают на 6-луночные планшеты и выращивают до плотности 90-95%. Трансфекцию осуществляют с использованием реагента ″Lipofectamine 2000″ (Invitrogen) согласно инструкции производителя. Для этого в 4,5 мл среды DMEM добавляют 36 мкг ДНК pCLm4/hygro-14D5 и 36 мкг ДНК pCHm2-14D5. Одновременно смешивают 180 мкл реагента ″Lipofectamine 2000″ с 4,5 мл среды DMEM. После пятиминутной инкубации оба полученных раствора объединяют и выдерживают 20 мин при комнатной температуре. После инкубации по 0,5 мл суспензии вносят в каждую лунку планшета, содержащего клетки СНО-К1. Через 4 ч после трансфекции культуральную среду в лунках заменяют на 0,5 мл среды "DMEM F12 advanced" (Gibco) с добавлением 2% сыворотки с пониженным содержанием иммуноглобулинов (Invitrogen). Через 72 ч культуральную среду собирают и центрифугируют 15 мин при 15000 g. Супернатант отделяют, добавляют к нему азид натрия до концентрации 0,05% и используют для выделения антител.
Пример 6. Выделение рекомбинантных антител с помощью аффинной хроматографии.
Культуральную жидкость в объеме около 500 мл наносят при 4°C со скоростью 0,5 мл/мин на колонку, содержащую 5 мл сорбента "Protein A Sepharose CL-4B″ (GE Healthcare) и уравновешенную фосфатно-солевым буферным раствором (ФСБР), представляющем собой раствор 100 мМ Nad, 50 мМ Na2HPO4 (pH 7,4 при 25°C). Элюцию осуществляют при 25°C и скорости 1 мл/мин с использованием хроматографа "BioLogic LP System" (Bio-Rad). Колонку промывают 25 мл ФСБР, после чего элюируют иммуноглобулины быка, исходно присутствующие в сыворотке, 25 мл 0,1 М цитратного буфера (pH 5,0 при 25°C). Далее 15 мл 0,1 М цитратного буфера (pH 3,0 при 25°C) элюируют химерное антитело в пробирку, содержащую 1,5 мл 1 М буфера Трис-HCl (pH 8,0 при 25°C). Фракцию, содержащую химерное антитело, концентрируют с помощью концентраторов ″Amicon ultra-4″ 30 кДа (Millipore) согласно инструкции производителя, одновременно заменяя буферный раствор на ФСБР с добавлением 0,05% азида натрия. Концентрацию антитела в образце определяют спектрофотометрически при длине волны 280 нм, учитывая, что раствор иммуноглобулинов IgG в концентрации 1 мг/мл при длине оптического пути 1 см характеризуется оптической плотностью около 1,41 [11]. Очищенные рекомбинантные антитела анализируют в восстанавливающих и невосстанавливающих условиях фракционированием в 12% полиакриламидном геле с добавлением додецилсульфата натрия (фиг. 5).
Пример 7. Определение сродства химерного антитела ch14D5 к рекомбинантому аналогу гликопротеина Е вируса клещевого энцефалита.
Кинетику связывания химерного антитела ch14D5 с рекомбинантным белком Е [8] и кинетику диссоциации комплексов, а также термодинамическую константу аффинности между этими соединениями определяли методом поверхностного плазмонного резонанса с использованием прибора ProteOn XPR36 (Bio-Rad). Для этого активируют поверхность одного из вертикальных каналов биосенсорного чипа GLC (Bio-Rad) с помощью набора "ProteOn Amine Coupling Kit" (Bio-Rad) согласно методике производителя. На активированную поверхность иммобилизуют рекомбинантный белок Е из расчета 250 мкл раствора с концентрацией 30 мкг/мл белка в 10 мМ ацетатном буфере (рН 4,5 при 25°C). Оставшиеся активированные группы блокируют 1М раствором этаноламин-HCl. В качестве аналита используют последовательные двухкратные разведения химерного антитела ch14D5 в ФСБР с добавлением 0,005% детергента tween-20, максимальная концентрация антитела составляет 7,5 мкг/мл. Ассоциацию проводят в течение 600 сек при скорости потока 25 мкл/мин, затем следует диссоциация в течение 1800 сек с той же скоростью. На основе полученной сенсограммы вычисляют кинетические константы связывания компонентов и диссоциации комплексов, которые используют для вычисления равновесных константы диссоциации и константы аффинности. Полученное в эксперименте значение константы аффинности химерного антитела ch14D5 к белку Е составило 6,0·10-11 М. (фиг. 6).
Пример 8. Определение способности химерного антитела ch14D5 обеспечивать экстренную профилактику мышей от вирусного клещевого энцефалита.
Для подтверждения способности химерного антитела ch14D5 обеспечивать экстренную профилактику модельных животных от вирусного клещевого энцефалита используют мышей линии Balb/C весом 10 г. Каждая группа содержит 10 животных. Для изучения дозозависимого эффекта препарат химерного антитела ch14D5 вводят в двух дозировках, 8 мг/кг веса животного и 1 мг/кг веса животного. В качестве препаратов сравнения используют иммуноглобулин человека против клещевого энцефалита для внутримышечного введения 10% с титром специфических антител в РТГА 1:160 (НПО «Микроген», г.Томск). В качестве контроля летальности вируса используют группу инфицированных мышей, в которых после инфицирования вирусом клещевого энцефалита не вводят ни химерное антитело, ни препараты сравнения. В качестве контроля используют группу интактных мышей.
Для приготовления вируса используют суспензию головного мозга мышей-сосунков, зараженных вирусом клещевого энцефалита, штамм «Абсеттаров», хранящуюся при -30°C. После размораживания суспензию путем последовательных десятикратных разведении разводят в 107 раз в 0,9% растворе хлорида натрия (рН 7,2-7,4 при 25°C) с добавлением 2% сыворотки крупного рогатого скота. Мышам внутрибрюшинно водят по 0,2 мл суспензии в рабочем разведении. Реальную летальную дозу препарата рассчитывают по методу Рида-Менча [12]. Для этого готовят последовательные десятикратные разведения мозговой суспензии вируса от 10-7 до 10-10 и вводят мышам внутрибрюшинно по 0,2 мл (по 6 голов на каждое разведение). Реальная доза введенного вируса в эксперименте составляет 240 ЛД5о.
Определение способности химерного антитела обеспечивать экстренную профилактику мышей от вирусного клещевого энцефалита осуществляют следующим образом. Через сутки после заражения вирусом мышам внутривенно в хвостовую вену однократно вводят заранее приготовленные рабочие растворы исследуемых антител в 0,9% растворе хлорида натрия (рН 7,2-7,4 при 25°C) в дозах 8 мг/кг веса (80 мкг/мышь) и 1 мг/кг веса (10 мкг/мышь). В качестве препарата сравнения однократно внутримышечно вводят препарат иммуноглобулина человека против клещевого энцефалита для внутримышечного введения в дозировке 10 мг/кг веса (100 мкг/мышь). Мышей наблюдают в течение 21 суток после заражения вирусом клещевого энцефалита, подсчитывая смертность животных. Результаты эксперимента показывают, что однократное введение химерного антитела в дозировке 8 мг/кг веса и 1 мг/кг веса мышам, зараженным 240 ЛД50 вируса клещевого энцефалита, обеспечило 100% выживаемость животных (летальность 0%). Результаты эксперимента представлены на фиг. 7.
Таким образом, впервые сконструированы плазмиды pCLm4/hygro-14D5 и pCHm2-14D5, содержащие искусственные гены легкой и тяжелой цепей полноразмерного химерного антитела, созданные на основе вариабельных фрагментов легких и тяжелых цепей мышиного моноклонального антитела 14D5, константных генов иммуноглобулинов IgG1/каппа человека, цитомегаловирусного промотора и сайта полиаденилирования гена бычьего гормона роста. Транзиентная экспрессия в эукариотических клетках линии СНО-К1 обусловливает биосинтез полипептидов со свойствами легкой и тяжелой цепей химерного антитела, которые объединяются в антитело класса IgG1/каппа, специфически взаимодействующее поверхностным гликопротеином Е вируса клещевого энцефалита. Константа аффинности химерного антитела к рекомбинантному гликопротеину Е составляет около 6,0·10-11 М. Полученное химерное антитело ch14D5, введенное в дозировке 1 мг на кг веса животного, обеспечивает экстренную профилактику мышей от вирусного клещевого энцефалита, что делает возможным использование химерного антитела ch14D5 в качестве основы для создания лекарственных препаратов для лечения вирусного клещевого энцефалита.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. Аммосов А.Д. Клещевой энцефалит: Информационно-методическое пособие / А.Д. Аммосов. - Кольцове: Ин-т средств мед. диагностики ЗАО «Вектор-Бест», 2006. - 115 с.
2. Пеньевская Н.А., Рудаков Н.В. Эффективность применения препаратов иммуноглобулина для постэкспозиционной профилактики клещевого энцефалита в России (Обзор полувекового опыта) // Медицинская паразитология и паразитарные болезни. -2010. -N1. - С. 53-59
3. Morrison, S.L., Johnson, M.J., Herzenberg, L.A., Oi V.T. Chimeric human antibody molecules: mouse antigen-binding domains with human constant region domains. // Proc Natl Acad Sci USA. - 1984. - V. 81(21). - P. 6851-6855.
4. Boulianne G.L., Hozumi N, Shulman MJ. Production of functional chimaeric mouse/human antibody. // Nature. - 1984. - V. 312(5995). - P. 643-646.
5. Tsekhanovskaya N.A., Matveev L.E., Rubin S.G., Karavanov A.S., Pressman E.K. Epitope analysis of tick-bome encephalitis (TBE) complex viruses using monoclonal antibodies to envelope glycoprotein of TBE virus (persulcatus subtype). // Virus Res. - 1993. - V. 30(1). - P. 1-16.
6. Levanov L.N., Matveev L.E., Goncharova E.P., Lebedev L.R., Ryzhikov A.B., Yun Т.Е., Batanova T.A., Shvalov A.N., Baykov I.K., Shingarova L.N., Kirpichnikov M.P., Tikunova N.V. Chimeric antibodies against tick-borne encephalitis virus // Vaccine. - 2010. - V. 28. - P. 5265-5271.
7. Тикунова Н.В., Шингарова Л.Н., Юн Т.Э. и др. Рекомбинантная плазмидная ДНК pcL37, кодирующая полипептид со свойствами легкой цепи антитела человека против вируса осповакцины, рекомбинантная плазмидная ДНК рсН37, кодирующая полипептид со свойствами тяжелой цепи указанного антитела, и их применение. Патент РФ №2317330 С2, оп.20.02.2008
8. Нетесова Н.А., Белавин П.А., Малыгин Э.Г., Рукавишников М.Ю. Рекомбинантная плазмидная ДНК PGSDEI, кодирующая белок Е вируса клещевого энцефалита, и штамм Escherichia coli - продуцент рекомбинантного белка Е вируса клещевого энцефалита. Патент РФ 2136754,1999
9. Giebel L.B., Spritz R.A. Site-directed mutagenesis using a double-stranded DNA fragment as a PCR primer. // Nucleic Acids Res. - 1990. - V. 18(16). -P. 4947.
10. Ke S.H., Madison E.L. Rapid and efficient site-directed mutagenesis by single-tube ′megaprimer′ PCR method. // Nucleic Acids Res. - 1997. -V. 25(16). -P. 3371-3372.
11. Hale G. Isolation and purification of monoclonal antibodies from tissue culture supernatant // Monoclonal antibodies. A practical approach. Shepherd P and Dean C. (ed). Oxford University Press. 2000. P. 149-180.
12. Reed L.J., Muench, H.A simple method of estimating fifty percent endpoints. // The American Journal of Hygiene. - 1938. - V. 27. - P. 493-497.









Группа изобретений относится к биотехнологии, генной и белковой инженерии. Сконструированы плазмиды pCLm4/hygro-14D5 и pCHm2-14D5. Плазмиды обеспечивают синтез в эукариотических клетках полипептидов со свойствами легкой и тяжелой цепей химерного антитела, которые объединяются в химерное антитело ch14D5 класса IgG1/каппа. Антитело направлено против вируса клещевого энцефалита и обеспечивает экстренную профилактику мышей от вирусного клещевого энцефалита. Полученое химерное антитело ch14D5, выделенное из культуральной жидкости клеток линии СНО-К1, трансфицированных плазмидами pCLm4/hygro-14D5 и pCHm2-14D5, имеющее молекулярную массу около 150 кДа. Антитело состоит из двух одинаковых легких цепей, константная часть которых соответствует каппа-классу человеческих антител, и двух одинаковых тяжелых цепей, константная часть которых соответствует IgGI-классу человеческих антител; имеющее аминокислотную последовательность, кодируемую нуклеотидной последовательностью SEQ ID NO: 1 и SEQ ID NO: 2. 3 н.п. ф-лы, 7 ил., 8 пр.
1. Рекомбинантная плазмидная ДНК pCLm4/hygro-14D5, кодирующая полипептид со свойствами легкой цепи химерного антитела против вируса клещевого энцефалита, размером 6232 п.н. и молекулярной массой 4,11 МДа, содержащая в соответствии с физической картой, представленной на фиг.3:
- NheI/ApaI - векторный фрагмент плазмиды pcDNA™3.1/Hygro(+) (Invitrogen) размером 5491 п.н., содержащий промотор-энхансер цитомегаловируса, сайт полиаденилирования и участок терминации транскрипции гена бычьего гормона роста, ген β-лактамазы (bla), ген гигромицин фосфотрансферазы (hpt);
- NheI/ApaI - фрагмент размером 741 п.н., содержащий искусственный ген, кодирующий гибридный белок, в котором вариабельный домен легкой цепи мышиного моноклонального антитела 14D5, осуществляющего протекцию против вируса клещевого энцефалита, соединен с константным доменом каппа-цепи антител человека, имеющий нуклеотидную последовательность SEQ ID NO: 1, приведенную на фиг. 1;
- генетические маркеры: ген β-лактамазы (bla), определяющий устойчивость трансформированных плазмидой pCLm4/hygro-14D5 клеток бактерий к ампициллину, ген гигромицин фосфотрансферазы (hpt), определяющий устойчивость к гигромицину В для селекции трансфицированных плазмидой pCLm4/hygro-14D5 клеток млекопитающих;
- уникальные сайты узнавания эндонуклеазами рестрикции, имеющие следующие координаты: BglII-13, NheI-896, EcoRV-980, HindIII-1284, ApaI-1637.
2. Рекомбинантная плазмидная ДНК pCHm2-14D5, кодирующая полипептид со свойствами тяжелой цепи химерного антитела против вируса клещевого энцефалита, размером 6765 п.н. и молекулярной массой 4,47 МДа, включающая:
- NheI/XbaI - векторный фрагмент плазмиды pcDNA™3.1(+) (Invitrogen) размером 5332 п.н., содержащий промотор-энхансер цитомегаловируса CMV, сайт полиаденилирования и участок терминации транскрипции гена бычьего гормона роста BGH, ген β-лактамазы (bla), ген устойчивости к неомицину (neo);
- NheI/XbaI - фрагмент размером 1433 п.н., содержащий искусственный ген, кодирующий гибридный белок, в котором вариабельный домен тяжелой цепи мышиного моноклонального антитела 14D5, осуществляющего протекцию против вируса клещевого энцефалита, соединен с константными СН1-СН2-СН3-доменами IgG1 человека, имеющий нуклеотидную последовательность SEQ ID NO: 1, приведенную на фиг. 2;
- генетические маркеры: ген β-лактамазы (bla), определяющий устойчивость трансформированных плазмидой pCHm2-14D5 клеток бактерий к ампициллину, ген устойчивости к неомицину (neo) для селекции трансфицированных плазмидой pCHm2-14D5 клеток млекопитающих;
- уникальные сайты узнавания эндонуклеазами рестрикции, имеющие следующие координаты: BglII-14, NheI-897, XhoI-1001, Acc65I-1313, XbaI-2330.
3. Химерное антитело, способное связывать белок Е вируса клещевого энцефалита и обеспечивающее экстренную профилактику клещевого энцефалита у мышей, включающее полипептиды со свойствами легкой и тяжелой цепей химерного антитела класса IgG1/каппа, полученное совместной трансфекцией клеток млекопитающих рекомбинантными плазмидными ДНК pCLm4/hygro-14D5 и pCHm2-14D5, охарактеризованными в п. 1 и 2 формулы.
| LEVANOV L.N | |||
| et al., "Chimeric antibodies against tick-borne encephalitis virus", Vaccine | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| TSEKHANOVSKAYA N.A | |||
| et al., "Epitope analysis of tick-borne encephalitis (TBE) complex viruses using monoclonal antibodies to envelope glycoprotein of TBE virus (persulcatus | |||

