Перекрестная ссылка на родственную заявку
Эта заявка заявляет преимущество Австралийской предварительной заявки на патент № 2009900736, поданной 20 февраля 2009 года, которая включена здесь посредством ссылки в ее полном виде.
Область техники, к которой относится изобретение
Данное изобретение относится к получению живых аттенуированных бактерий, которые продолжают сохраняться в субъекте, для применения в вакцинных композициях. В частности, данное изобретение относится к удалению функции по меньшей мере одного АВС-транспортерного белка бактерии для получения аттенуированной бактерии, которая продолжает существовать в субъекте и может быть использована в вакцинной композиции.
Уровень техники
Бактериальные инфекции вызывают существенные экономические потери в животноводстве вследствие заболеваемости и смертности и оказывают существенное влияние на здоровье человека. В частности, респираторные инфекции и септицемии являются обычными инфекционными заболеваниями, которые могут распространяться в группе животных или людей (субъектов) и отрицательно влиять на здоровье этих субъектов. Это может приводить к существенным потерям продуктивности и, в конечном счете, к смерти этих субъектов. Например, патогенные бактерии, в частности, рода Mycoplasma, являются существенной причиной респираторного заболевания. Септицемия обычно вызывается E. coli.
Известно, что вакцинные композиции, содержащие аттенуированные патогенные микроорганизмы, такие как бактерии и вирусы, являются эффективными в продуцировании защитной иммунной реакции в вакцинированных животных и людях. Такие живые аттенуированные вакцины являются предпочтительными, так как после иммунизации последующее заражение патогенным организмом, на котором основана эта вакцина, приводит к быстрой повторной стимуляции иммунной реакции, первоначально индуцированной этой вакцинацией. Это вызывает ингибирование пролиферации этого патогена и предотвращает развитие клинически релевантного заболевания.
Как правило, аттенуирование патогенных организмов для использования в вакцине достигается полным или частичным удалением одного или нескольких факторов вирулентности, так что этот организм уже не является патогенным. Факторы вирулентности обычно известны как свойства, которые непосредственно вызывают заболевание и/или позволяют этому организму персистировать в хозяине. Свойства, которые усиливают вредные реакции хозяина, также обычно известны как факторы вирулентности. Типичные факторы вирулентности включают, например, токсины, прикрепляющие органеллы и механизмы иммунной инвазии. Это создает проблему, заключающуюся в том, что многие факторы вирулентности участвуют в индуцировании иммунитета, и, следовательно, удаление факторов вирулентности ухудшает иммуногенность организма, аттенуированного таким образом. Живая аттенуированная вакцина выгодным образом остается антигенной и, следовательно, способна индуцировать достаточный уровень иммунитета хозяина, будучи непатогенной. Обычно, живые аттенуированные вакцины не сохраняются в субъекте, и это может способствовать уменьшению иммуногенности живого аттенуированного организма, на котором основана эта вакцина.
Хотя некоторые факторы вирулентности, такие как токсины, являются очевидными мишенями для аттенуации, факторы, неизвестные в качестве факторов вирулентности, не являются очевидными мишенями для аттенуации. Действия аттенуации этих факторов на вирулентность организма не являются легко очевидными или предсказуемыми. Члены суперсемейства АВС-транспортеров (АТФ-связывающей кассеты) обычно не считаются факторами вирулентности. АВС-транспортеры содержат мембранные белки, которые выполняют функцию транслокации субстратов через внеклеточные и внутриклеточные мембраны. Вещества, транспортируемые АВС-транспортерами, включают пептиды, олигопептиды, белки, метаболические продукты, липиды, стерины и лекарственные средства. Сравнение последовательностей АВС-транспортеров указывает на то, что гены и белки этого суперсемейства являются консервативными среди отдаленно родственных типов (филюмов).
Данное изобретение предсказано на основе неожиданного открытия, что потеря мутации функции гена АВС-транспортера в патогенных организмах, аттенуировала патогенные организмы, хотя аттенуированные организмы оставались иммуногенными и продолжали существовать в субъекте, как было оценено в модельной системе хозяин-заболевание.
Сущность изобретения
В первом аспекте обеспечена бактерия, аттенуированная мутацией по меньшей мере в одном гене АВС-транспортера, где мутация делает соответствующий АВС-транспортерный белок нефункциональным, причем аттенуированная бактерия продолжает существовать в субъекте.
Во втором аспекте обеспечен способ аттенуации бактерий, предусматривающий мутирование по меньшей мере одного гена АВС-транспортера, причем аттенуированная бактерия продолжает существовать в субъекте.
В третьем аспекте обеспечена иммуногенная композиция, содержащая по меньшей мере одну аттенуированную бактерию первого аспекта.
В четвертом аспекте обеспечена вакцинная композиция, содержащая эффективное количество по меньшей мере одной аттенуированной бактерии первого аспекта и фармацевтически приемлемый носитель.
В пятом аспекте обеспечено применение вакцинной композиции для лечения или профилактики заболевания, причем вакцинная композиция содержит по меньшей мере одну аттенуированную бактерию первого аспекта.
В шестом аспекте обеспечен способ предотвращения или устранения заболевания в субъекте, предусматривающий введение терапевтически эффективной дозы вакцинной композиции или иммуногенной композиции субъекту, причем вакцинная композиция или иммуногенная композиция содержит по меньшей мере одну аттенуированную бактерию первого аспекта.
В седьмом аспекте обеспечен способ профилактики заболевания, предусматривающий введение терапевтически эффективной дозы вакцинной композиции или иммуногенной композиции субъекту, нуждающемуся в профилактике, причем указанная вакцинная композиция или иммуногенная композиция содержит по меньшей мере одну аттенуированную бактерию первого аспекта.
В одном варианте осуществления мутация может быть генерирована инсерцией, делецией, заменой или любой их комбинацией. Инсерционная мутация может быть получена, например, гомологичной рекомбинацией, транспозонным мутагенезом и ST-мутагенезом с последовательностью-меткой.
В одном варианте осуществления ген АВС-транспортера может быть геном, кодирующим белок АВС-пептидного транспортера. Например, CvaB, CylB, SpaB, NisT, EpiT, ComA, PedD, LcnC, McbEF, OppD и DppD и их гомологи и, в частности, ген АВС-транспортера, могут кодировать OppD.
В одном варианте осуществления бактерия может быть выбрана из группы, состоящей из Avibacterium, Bacillus, Brucella, Bartonella, Bordetella, Burkholderia, Vibrio, Escherichia, Salmonella, Clostridium, Campylobacter, Chlamydia, Coxiella, Erysipelothrix, Francisella, Listeria, Actinobacillus, Haemophilus, Helicobacter, Aeromonas, Pseudomonas, Streptococcus, Shigella, Yersinia, Mycoplasma, Mycobacterium, Mannheimia, Ornithobacterium, Rickettsia, Ureaplasma и Pasteurella. В частности, аттенуированными бактериями могут быть Avibacterium paragallinarum, Bordetella avium, Ornithobacterium rhinotracheale, Salmonella enteritidis, Pasteurella multocida, Mannheimia haemolytica, E. coli, Clostridium perfringens, Mycoplasma agalactiae, Mycoplasma anatis, Mycoplasma anseris, Mycoplasma imitans, Mycoplasma alkalescens, Mycoplasma arginini, Ureaplasma parvum, Mycoplasma arthritidis, Mycoplasma bovigenitalium, Mycoplasma bovirhinis, Mycoplasma bovis, Mycoplasma bovoculi, Mycoplasma californicum, Mycoplasma capricolum, Mycoplasma dispar, Mycoplasma felis, Mycoplasma fermentans, Mycoplasma genitalium, Mycoplasma hominis, Mycoplasma hyopneumoniae, Mycoplasma hyorhinis, Mycoplasma hyosynoviae, Mycoplasma iowae, большая колония и малая колония Mycoplasma mycoides subsp mycoides, Mycoplasma gallisepticum, Mycoplasma synoviae, Mycoplasma orale, Mycoplasma penetrans, Mycoplasma ovipneumoniae, Mycoplasma pullorum, Mycoplasma alligatorus, Mycoplasma pneumoniae, Mycoplasma maleagridis, Mycoplasma haemofelis, Mycoplasma haemominutum, Mycoplasma haematoparvum.
В одном варианте осуществления бактерией может быть птичий патогенный штамм E. coli Е956 или штамм Mycoplasma gallisepticum Ap3AS.
В одном варианте осуществления живая аттенуированная бактерия может экспрессировать гетерологичный антиген. Гетерологичный антиген может кодироваться нуклеиновой кислотой из другого патогенного организма. Нуклеиновая кислота, кодирующая гетерологичный антиген, может быть выделена из родов, выбранных из группы, включающей Avibacterium, Bacillus, Brucella, Bartonella, Bordetella, Burkholderia, Vibrio, Escherichia, Salmonella, Clostridium, Campylobacter, Chlamydia, Coxiella, Erysipelothrix, Francisella, Listeria, Actinobacillus, Haemophilus, Helicobacter, Aeromonas, Pseudomonas, Streptococcus, Shigella, Yersinia, Mycoplasma, Mycobacterium, Mannheimia, Ornithobacterium, Rickettsia, Staphylococci, Ureaplasma и Pasteurella. В частности, нуклеиновая кислота, кодирующая гетерологичный антиген, может быть получена из Avibacterium paragallinarum, Bordetella avium, Ornithobacterium rhinotracheale, Salmonella enteritidis, Pasteurella multocida, Mannheimia haemolytica, E. coli, Clostridium perfringens, Mycoplasma hyopneumoniae, Mycoplasma gallisepticum или Mycoplasma synoviae, Avibacterium paragallinarum, Bordetella avium, Ornithobacterium rhinotracheale, Mycoplasma agalactiae, Mycoplasma alkalescens, Mycoplasma anatis, Mycoplasma anseris, Mycoplasma imitans, Mycoplasma arginini, Ureaplasma parvum, Mycoplasma arthritidis, Mycoplasma bovigenitalium, Mycoplasma bovirhinis, Mycoplasma bovis, Mycoplasma bovoculi, Mycoplasma californicum, Mycoplasma capricolum, Mycoplasma dispar, Mycoplasma felis, Mycoplasma fermentans, Mycoplasma genitalium, Mycoplasma hominis, Mycoplasma hyopneumoniae, Mycoplasma hyorhinis, Mycoplasma hyosynoviae, Mycoplasma iowae, большую колонию и малую колонию Mycoplasma mycoides subsp mycoides, Mycoplasma gallisepticum, Mycoplasma synoviae, Mycoplasma orale, Mycoplasma penetrans, Mycoplasma ovipneumoniae, Mycoplasma pullorum, Mycoplasma alligatorus, Mycoplasma pneumoniae, Mycoplasma, Mycoplasma maleagridis, Mycoplasma haemofelis, Mycoplasma haemominutum, Mycoplasma haematoparvum. Нуклеиновая кислота, кодирующая гетерологичный антиген, может быть выделена из вирусов, выбранных из группы, включающей вирус болезни Ньюкасл (псевдочумы птиц), вирус инфекционного бронхита, птичий пневмовирус, вирус оспы птиц, инфекционного бурсального заболевания, вирус инфекционного ларинготрахеита, птичьего гриппа, вирус гепатита уток, вирусной чумы уток, инфекционной куриной анемии, вирус болезни Марека.
Субъект представляет собой позвоночное животное, включающее людей, коровьих, собачьих, кошачьих, козлиных, овечьих, свиных, верблюдовых, лошадиных и птичьих. Субъектом коровьих может быть корова, бык, бизон или буйвол. Субъектом собачьих может быть собака. Субъектом кошачьих может быть кошка. Субъектом козлиных может быть коза. Субъектом овечьих может быть овца. Субъектом свиных может быть свинья. Субъектом верблюдовых может быть верблюд, дромадер, лама, альпака, викунья или гуанако. Субъектом лошадиных может быть лошадь, осел, зебра или мул. Субъектом птичьих может быть любой из выращенных коммерчески или домашних представителей семейства птичьих. В частности, субъектом птичьих может быть курица (в том числе карликовые куры бентамки), индейка, утка, гусь, фазан, перепел, куропатка, голубь, цесарка, африканский страус, эму или павлин.
Вакцинные композиции и иммуногенные композиции могут дополнительно содержать по меньшей мере один фармацевтически приемлемый носитель или разбавитель, такой как вода, солевой раствор, культуральная жидкость, стабилизаторы, углеводы, белки, белоксодержащие агенты, такие как бычья сыворотка или обезжиренное молоко, и буферы или любую их комбинацию.
Стабилизатором может быть SPGA. Углеводы включают, например, сорбит, маннит, крахмал, сахарозу, глюкозу, декстран или их комбинации. Дополнительно, в качестве фармацевтически приемлемых носителей или разбавителей могут быть использованы белки, такие как альбумин или казеин, или белоксодержащие агенты, такие как бычья сыворотка или обезжиренное молоко.
Буферы для применения в качестве фармацевтически приемлемых носителей или разбавителей включают малеат, фосфат, CABS, пиперидин, глицин, цитрат, малат, формиат, сукцинат, ацетат, пропионат, пиперазин, пиридин, какодилат, сукцинат, MES, гистидин, бис-трис, фосфат, этаноламин, ADA, карбонат, ACES, PIPES, имидазол, BIS-TRIS пропан, BES, MOPS, HEPES, TES, MOPSO, MOBS, DIPSO, TAPSO, TEA, пирофосфат, HEPPSO, POPSO, трицин, гидразин, глицилглицин, TRIS, EPPS, бицин, HEPBS, TAPS, AMPD, TABS, AMPSO, таурин, борат, CHES, глицин, гидроксид аммония, CAPSO, карбонат, метиламин, пиперазин, CAPS или любую их комбинацию.
Вакцинные композиции и иммуногенные композиции могут быть лиофилизированы или высушены замораживанием.
В некоторых вариантах осуществления вакцинные композиции и иммуногенные композиции могут дополнительно содержать по меньшей мере один адъювант. Примеры адъювантов включают полный адъювант Фрейнда или неполный адъювант Фрейнда, витамин Е, неионные блоксополимеры, мурамилдипептиды, сапонины, минеральное масло, растительное масло, гидроксид алюминия карбопол, фосфат алюминия, оксид алюминия, масляные эмульсии (например, Bayol F® или Marcol 52®), сапонины или солюбилизат витамина Е или любую их комбинацию. В некоторых вариантах осуществления вакцинная композиция может содержать адъюванты, применимые, в частности, для мукозного применения, например, термолабильный токсин E. coli или холерный токсин.
Вакцинная композиция или иммуногенная композиция может вводиться интраназально, офтальмически, внутрикожно, внутрибрюшинно, внутривенно, подкожно, перорально, при помощи аэрозоля (спрей-вакцинация), через клоаку или внутримышечно. Введение в виде глазных капель или аэрозольное введение являются предпочтительными, когда субъектом является птица. Аэрозольное введение является особенно предпочтительным для введения вакцинной композиции или иммуногенной композиции большим количествам субъектов.
Иммуногенная композиция или вакцинная композиция может содержать по меньшей мере от приблизительно 103 до приблизительно 105 аттенуированных бактерий, или от приблизительно 105 до приблизительно 107 аттенуированных бактерий, или от приблизительно 107 до приблизительно 109 аттенуированных бактерий, или от приблизительно 109 до приблизительно 1011 аттенуированных бактерий, или от приблизительно 1011 до приблизительно 1013 аттенуированных бактерий, или от приблизительно 1013 до приблизительно 1015 аттенуированных бактерий, или от приблизительно 1015 до приблизительно 1017 аттенуированных бактерий, или от приблизительно 1017 до приблизительно 1019, или по меньшей мере приблизительно 1019 аттенуированных бактерий на дозу.
Определения
В контексте этой заявки, единственная форма "a", "an" и "the" включает и множественные ссылки, если контекст не указывает явно другое. Например, термин "ABC-транспортер" включает также множество АВС-транспортеров.
В контексте этого описания, термин "содержащие" обозначает "включающие в основном, но необязательно только". Кроме того, вариации слова "содержащие", такие как "содержат" и "содержит", имеют соответственно варьирующиеся значения.
В данном контексте термин "иммуногенно эффективные" обозначает, что количество живых аттенуированных бактерий, вводимых при вакцинации или при введении иммуногенной композиции, является достаточным для индукции в этом субъекте эффективной иммунной реакции против вирулентных форм этой бактерии.
В данном контексте термины "лечение" и "обработка" относятся к любому или всем применениям, которые лечат состояние или симптомы, предотвращают возникновение состояния или заболевания или другим путем задерживают или замедляют прогрессирование состояния или заболевания или другие нежелательные симптомы любым каким бы то ни было путем.
В данном контексте термины "персистируют" и "персистирующий" относятся к установлению и/или сохранению инфекции или колонизации по меньшей мере части субъекта. Персистенция включает состояние, в котором организм выживает в тканях, независимо от того, воспроизводится он или не воспроизводится. Персистирующий организм может быть или может не быть ассоциирован с клинически релевантным заболеванием.
Краткое описание фигур
Фиг.1 является схематическим планом компонентов и экспериментального протокола. А. Основная структура меченного сигнатурой (определенной последовательностью) транспозона. B. Схема для экспериментов с меченным сигнатурой (определенной последовательностью) мутагенеза (STM).
Фиг.2. Детектирование конкретных меток. Бульонные культуры, обнаруживающие изменение окраски, подвергали скринингу при помощи ПЦР, использующей пару праймеров P2/P4, для детектирования индивидуальных сигнатурных меток. Каждую пробу подвергали электрофорезу в 2% агарозном геле вместе с маркером размеров ДНК (pUC18, расщепленной HaeIII). Дорожка 1, ST-мутант, несущий метку Tag 02; Дорожка 2, ST-мутант, несущий метку Tag 03; Дорожка 3, ST-мутант, несущий метку Tag 04; Дорожка 4, ST-мутант, несущий метку Tag 06; Дорожка 5, ST-мутант, несущий метку Tag 07; Дорожка 6, отрицательный контроль. Дорожка 7, положительный контроль, плазмида, содержащая транспозон, несущий Tag 02 в качестве матрицы.
Фиг.3. Подтверждение трансформантов STM. Трансформанты, которые обнаруживали изменение окраски, подвергали скринингу при помощи ПЦР, использующей набор праймеров P2/P4, для подтверждения присутствия индивидуальной сигнатурной метки. Каждую пробу подвергали электрофорезу в 2% агарозном геле вместе со стандартом размеров ДНК pUC18, расщепленной HaeIII. Дорожка 1, рекомбинантный Mg-содержащий Tag02; Дорожка 2, трансформант Tag03; Дорожка 3, клон Tag04; Дорожка 4, трансформант Tag06; Дорожка 5, трансформант, несущий Tag07; +, положительный контроль с плазмидой Tag02 в качестве матрицы, -, отрицательный контроль без добавленной матрицы.
Фиг.4. Экспериментальная схема подтверждающего скрининга.
Фиг.5. Последовательность и Саузерн-блот-анализ ST-мутантов.
A. Электроферограммы, полученные прямым секвенированием. (I). ST-мутант 26-2, который нес единственный транспозон, со считываемой последовательностью в геном. Можно видеть сигналы смешанных последовательностей, начинающихся в точке соединения транспозона и генома штамма-хозяина в ST-мутантах 15-1 (II) и 25-1 (III).
B. Множественные инсерции, детектированные Саузерн-блоттингом. Дорожка 1, ST-мутант 15-1; Дорожка 2, ST-мутант 15-2; Дорожка 3, ST-мутант 15-3; Дорожка 4, ST-мутант 25-1; Дорожка 5, ST-мутант 25-2; Дорожка 6, ST-мутант 25-3. Стандартами размеров ДНК была ДНК фага λ, расщепленная HindIII.
Фиг.6. Распределение балльных оценок RSA и повреждений альвеолярных мешочков в птицах в исследовании вирулентности и инфективности. A. Балльная оценка RSA при 2 неделях после инфицирования. B. Балльная оценка повреждения альвеолярных мешочков. Группа 1, отрицательный контроль; Группа 2, инфицированная ST-мутантом 04-1; Группа 3, инфицированная ST-мутантом 33-1; Группа 4, инфицированная ST-мутантом 03-1; Группа 5, инфицированная ST-мутантом 26-1; Группа 6, инфицированная ST-мутантом 18-1; Группа 7, инфицированная ST-мутантом 20-1; Группа 8, инфицированная ST-мутантом 22-1; Группа 9, положительный контроль (инфицированная Ap3AS дикого типа).
Фиг.7. Распределение балльных оценок RSA и повреждений альвеолярных мешочков птиц. A. Балльная оценка RSA в день 28. B. Балльная оценка повреждения альвеолярных мешочков. Суммарная балльная оценка 6 различных альвеолярных мешочков. Группа 1, отрицательный контроль; Группа 2, положительный контроль (инфицированная Ap3AS дикого типа); Группа 3, вакцинированный контроль (вакцинированная ST-мутантом 26-1); Группа 4, вакцинированная (ST-мутантом 26-1) и зараженная (зараженная Ap3AS дикого типа).
Фиг.8. Схематическая диаграмма ПЦР-конструкций, используемых для гомологичной рекомбинации. Эти ПЦР-продукты генерировали с олигонуклеотидными праймерами, как описано. Ген канамицина амплифицировали для конструкций генов как oppA, так и dppD с использованием пары праймеров TX/TY. Генерировали плечи конструкций и дополняли дополнительными ПЦР-продуктами с использованием WS/WU и WY/WT для oppD или с использованием 60-мерных праймеров WE/WF в ПЦР для dppD.
Фиг.9. Детектирование гомологичной рекомбинации при помощи ПЦР. Олигонуклеотидные праймеры для генов dppD (Панель А) и oppD (Панель B) использовали в ПЦР для амплификации соответствующих районов в штаммах E. coli с использованием пары праймеров WS/WT для oppD и TZ/UA для dppD: E956 (дорожка 1 панелей A и B), ΔdppD (дорожка 1, панель A) и ΔoppD (дорожка 1, панель B).
Фиг.10. Саузерн-блоттинг нокаутов dppD и oppD E. coli Е956. Канамицин, зонды dppD и oppD метили радиоактивно и использовали для зондирования расщепленной рестриктазой PstI геномной ДНК E. coli Е956, ΔdppD и ΔoppD, дорожки 1, 2 и 3, соответственно.
Подробное описание
Данное изобретение относится к живым аттенуированным бактериям, которые персистируют в субъекте, и их применению в вакцинных композициях. Эти аттенуированные бактерии лишены по меньшей мере одного функционального АВС-транспортерного белка в результате мутации в нуклеиновой кислоте, кодирующей АВС-транспортерный белок. АВС-транспортерный белок может быть сделан нефункциональным любым способом, но обычно это достигается мутацией нуклеиновой кислоты, кодирующей АВС-транспортерный белок. Обычно мутация является инсерцией, делецией или мутацией со сдвигом рамки или любой их комбинацией. В некоторых вариантах осуществления живые аттенуированные бактерии могут также экспрессировать гетерологичный антиген.
В соответствии с настоящим изобретением может быть аттенуирован любой тип бактерий, хотя предпочтительной является аттенуация патогенных бактерий позвоночных животных и, в частности, патогенных бактерий птиц.
Бактерии
В одном варианте осуществления, изобретение относится к живым аттенуированным бактериям, но не ограничивается ими, родов Actinobacillus, Aeromonas, Avibacterium, Bacillus, Brucella, Bartonella, Bordetella, Burkholderia, Escherichia, Salmonella, Clostridium, Campylobacter, Chlamydia, Coxiella, Erysipelothrix, Francisella, Listeria, Actinobacillus, Haemophilus, Helicobacter, Listeria, Aeromonas, Pasteurella, Streptococcus, Shigella, Yersinia, Mycoplasma, Mycobacterium, Ornithobacterium, Mannheimia, Vibrio, Rickettsia, Pseudomonas, Staphylococci, Ureaplasma и Pasteurella для применения в вакцинной композиции. Эти бактериальные роды содержат большое количество видов, которые являются патогенными как для представителей птичьих, так и для различных других животных, в том числе человека.
В частности, аттенуированными бактериями могут быть, но не ограничиваются ими, Avibacterium paragallinarum, Bordetella avium, Ornithobacterium rhinotracheale, Salmonella enteritidis, Pasteurella multocida, Mannheimia haemolytica, E. coli, Clostridium perfringens, Mycoplasma hyopneumoniae, Mycoplasma gallisepticum, Mycoplasma synoviae, Mycoplasma agalactiae, Mycoplasma alkalescens, Mycoplasma anatis, Mycoplasma anseris, Mycoplasma imitans, Mycoplasma arginini, Ureaplasma parvum, Mycoplasma arthritidis, Mycoplasma bovigenitalium, Mycoplasma bovirhinis, Mycoplasma bovis, Mycoplasma bovoculi, Mycoplasma californicum, Mycoplasma capricolum, Mycoplasma dispar, Mycoplasma felis, Mycoplasma fermentans, Mycoplasma genitalium, Mycoplasma hominis, Mycoplasma hyopneumoniae, Mycoplasma hyorhinis, Mycoplasma hyosynoviae, Mycoplasma iowae, большая колония и малая колония Mycoplasma mycoides subsp mycoides, Mycoplasma gallisepticum, Mycoplasma synoviae, Mycoplasma orale, Mycoplasma penetrans, Mycoplasma ovipneumoniae, Mycoplasma pullorum, Mycoplasma alligatorus, Mycoplasma pneumoniae, Mycoplasma, Mycoplasma maleagridis, Mycoplasma haemofelis, Mycoplasma haemominutum, Mycoplasma haematoparvum.
В одном варианте живая аттенуированная бактерия может быть выбрана из группы, состоящей из E. coli или Mycoplasma gallisepticum.
E. coli может быть птичьей патогенной E. coli E956, и Mycoplasma gallisepticum может быть Mycoplasma gallisepticum Ap3AS.
Персистенция в субъекте
Живые аттенуированные бактерии стойко сохраняются в субъекте. Персистенция аттенуированных бактерий предпочтительно оценивается в модельной системе заболевания субъекта, но может также оцениваться в субъектах, которым были введены описанные здесь вакцинная композиция или иммуногенная композиция. Персистенция аттенуированных бактерий может оцениваться наблюдением клинических признаков и/или симптомов. Альтернативно или в добавление к этому, персистенция может оцениваться определением присутствия аттенуированной бактерии в пробе, взятой из этого субъекта. Этой пробой может быть проба ткани (например, крови, кожи, волос, внутриротового соскоба) или проба биологической секреции или экскреции, например, слизи, мочи, фекалий, слезной жидкости. Пробы могут браться при жизни субъекта или при аутопсии или вскрытии трупа. Присутствие аттенуированных бактерий в пробе может оцениваться при помощи культивирования, молекулярных способов, таких как ELISA или PCR, или любым способом, известным в данной области. Кроме того, обследование пораженных зон субъекта при аутопсии или вскрытии трупа может также позволить определение того, что аттенуированная бактерия продолжает существовать в субъекте.
ABC-транспортеры
Транспортеры АТФ-связывающей кассеты (ABC-транспортеры) образуют одно из самых больших суперсемейств белков и генов с характерными представителями во всех типах (филюмах) от простейших до человека. ABC-транспортеры являются трансмембранными белками, которые используют АТФ для транспорта веществ с определенными функциями через клеточные мембраны. Эти вещества включают метаболиты, пептиды, олигопептиды, липиды и стерины и лекарственные средства.
В бактериях ABC-транспортеры подразделяются на три основные группы на основе типа субстрата, который транслоцирует каждый из них. Этими группами являются транспортеры белков, транспортеры пептидов и системы, которые транспортируют небелковые субстраты.
В соответствии с настоящим изобретением живые аттенуированные бактерии могут быть получены мутацией по меньшей мере одного гена АВС-транспортера любого из этих классов, так что бактерия лишена по меньшей мере одного функционального АВС-транспортерного белка.
Особый интерес представляют ABC-пептидные транспортеры, которые включают, например, CvaB (E. coli), CylB (Enterococcus faecalis), SpaB (Bacillus subtilis), NisT (Lactococcus lactis 6F3), EpiT (Staphylococcus epidermidis), ComA (Streptococcus pneumoniae), PedD (Pediococcus acidilactici), LcnC (Lactococcus lactis subsp. lactis), McbEF (E coli) OppD и DppD (E. coli и M. gallisepticum) и их гомологи в других видах.
Субъекты
Предполагается, что субъектом, которому должны вводить живые аттенуированные бактерии, может быть любое позвоночное животное, включающее людей, коровьих, собачьих, кошачьих, козлиных, овечьих, свиных, верблюдовых, лошадиных и птичьих. Субъектом коровьих может быть корова, бык, бизон или буйвол. Субъектом собачьих может быть собака. Субъектом кошачьих может быть кошка. Субъектом козлиных может быть коза. Субъектом овечьих может быть овца. Субъектом свиных может быть свинья. Субъектом верблюдовых может быть верблюд, дромадер, лама, альпака, викунья или гуанако. Субъектом лошадиных может быть лошадь, осел, зебра или мул. Субъектом птичьих может быть любой из выращенных коммерчески или домашних представителей семейства птичьих. В частности, субъектом птичьих может быть курица (в том числе карликовые куры бентамки), индейка, утка, гусь, фазан, перепел, куропатка, голубь, цесарка, африканский страус, эму или павлин.
Мутации
Мутациями, используемыми для создания нефункционального АВС-транспортерного белка, могут быть инсерционные, делеционные мутации или мутации с заменами или их комбинация, при условии, что мутация приводит к нарушению экспрессии функционального АВС-транспортерного белка. АВС-транспортерные белки могут иметь множественные функции, например, связывание АТФ или связывание олигопептидов. Нефункциональный АВС-транспортерный белок включает АВС-транспортерный белок, который является дефектным по меньшей мере в одной из его функций.
Такой мутацией может быть инсерционная мутация, делеционная мутация, мутация с заменой или их комбинация, при условии, что эта мутация приводит к нарушению экспрессии функционального АВС-транспортерного белка. В предпочтительном варианте эта мутация является мутацией с нокаутом, где по меньшей мере существенная часть нуклеиновой кислоты, кодирующей АВС-транспортерный белок, делетирована. Инсерционная мутация может быть получена, например, гомологичной рекомбинацией, транспозонным мутагенезом или ST-мутагенезом (использующим в качестве меток определенные последовательности).
Живые аттенуированные бактерии для применения согласно изобретению могут быть получены различными путями. Например, живые аттенуированные бактерии могут быть получены обработкой бактерий дикого типа мутагенными агентами, такими как аналоги пуринов или пиримидинов, ультрафиолетовый свет, ионизирующая радиация, ДНК-интеркалирующие агенты, температурная обработка, транспозонный мутагенез. Эти способы не нацеливают мутации на специфические гены, и, следовательно, требуют скрининга обработанных бактерий на аттенуацию способами, известными в данной области, такими как анализ последовательности ДНК генов-мишеней в комбинации с исследованиями патогенности.
Технология рекомбинантных ДНК, использующая известные способы, такие как сайт-направленный мутагенез, может быть использована для введения мутации в заданном сайте конкретного гена, например, oppD. Сайт-направленный мутагенез может быть использован для введения мутации, такой как инцерция, делеция, замена одного или нескольких нуклеотидов, так что мутированный ген уже не экспрессирует функциональный АВС-транспортерный белок. Такие мутации могут быть, например, получены делецией ряда нуклеиновых кислот.
Делеции всего лишь единственного основания, создающие, следовательно, сдвиг рамки, могут делать АВС-белок нефункциональным. В некоторых вариантах большие количества оснований делетированы. В других вариантах могут быть делетированы большинство генов или все из генов, кодирующих АВС-транспортерный белок. Мутации, которые вводят стоп-кодон или поизводят сдвиг рамки, пригодны для получения нефункционального АВС-транспортерного белка.
Гены, кодирующие АВС-транспортерные белки, содержат не только кодирующую последовательность, но включают также регуляторные последовательности, например, промоторы. Гены включают также районы, существенные для коррекции трансляции мРНК, например, сайты связывания рибосом. Таким образом, живые аттенуированные бактерии могут содержать мутации не только в кодирующих районах, но также или альтернативно в последовательностях, существенных для транскрипции и трансляции, таких как промоторы и сайты связывания рибосом.
В отличие от аттенуированных бактерий, созданных спонтанными мутациями, аттенуированные бактерии, созданные такими мутациями, как делетирование фрагментов генов АВС-транспортеров или делетирование полных АВС-транспортерных генов или инсертирование фрагментов гетерологичной ДНК или их комбинаций, имеют преимущество, заключающееся в том, что они не ревертируются в патогенные бактерии дикого типа. Таким образом, в предпочтительном варианте осуществления изобретение обеспечивает живые аттенуированные бактерии, в которых по меньшей мере один АВС-транспортерный ген содержит инсерцию и/или делецию.
Аттенуированные бактерии, экспрессирующие гетерологичные антигены
В одном варианте осуществления живые аттенуированные бактерии могут быть использованы в качестве носителей для гетерологичных генов, которые кодируют антигены других патогенных бактерий или вирусов. Это позволяют использовать живые аттенуированные бактерии для вызывания иммунной реакции в отношении множества заболеваний.
Ген, кодирующий АВС-транспортерный белок, может быть использован в качестве сайта инсерции для гетерологичных генов. Применение гена АВС-транспортера в качестве инсерции является выгодным, так как инсерция последовательности, кодирующей гетерологичный антиген, как инактивирует АВС-транспортерный белок, так и вводит последовательность, кодирующую один или несколько гетерологичных антигенов. Конструирование таких рекомбинантных бактерий может выполняться рутинно, с использованием стандартных способов, известных в данной области.
Другой вариант осуществления изобретения относится к живым рекомбинантным бактериями, выбранным из родов Avibacterium, Staphylococcus, Escherichia, Brucella, Salmonella, Bordetella, Burkholderia, Vibrio, Haemophilus, Ornithobacterium, Mannheimia, Pasteurella, Clostridium, Campylobacter, Chlamydia, Coxiella, Erysipelothrix, Francisella, Listeria, Actinobacillus, Haemophilus, Helicobacter, Aeromonas, Pseudomonas, Shigella, Yersinia, Mycoplasma, Mycobacterium, Rickettsia, Ureaplasma и Streptococcus, которые не продуцируют функциональный АВС-транспортер и в которые инсертирована гетерологичная нуклеиновая кислота. Гетерологичная нуклеиновая кислота может кодировать антиген, выбранный из другого патогенного микроорганизма или вируса. Нуклеиновая кислота может кодировать антиген или антигены из патогенных организмов, выбранных из группы, включающей Avibacterium paragallinarum, Bordetella avium, Ornithobacterium rhinotracheale, Salmonella enteritidis, Pasteurella multocida, Mannheimia haemolytica, E. coli, Clostridium perfringens, Mycoplasma hyopneumoniae, Mycoplasma gallisepticum или Mycoplasma synoviae, Mycoplasma agalactiae, Mycoplasma alkalescens, Mycoplasma anatis, Mycoplasma anseris, Mycoplasma imitans, Mycoplasma arginini, Ureaplasma parvum, Mycoplasma arthritidis, Mycoplasma bovigenitalium, Mycoplasma bovirhinis, Mycoplasma bovis, Mycoplasma bovoculi, Mycoplasma californicum, Mycoplasma capricolum, Mycoplasma dispar, Mycoplasma felis, Mycoplasma fermentans, Mycoplasma genitalium, Mycoplasma hominis, Mycoplasma hyorhinis, Mycoplasma hyosynoviae, Mycoplasma iowae, большую колонию и малую колонию Mycoplasma mycoides subsp mycoides, Mycoplasma orale, Mycoplasma penetrans, Mycoplasma ovipneumoniae, Mycoplasma pullorum, Mycoplasma alligatorus, Mycoplasma pneumoniae, Mycoplasma, Mycoplasma maleagridis, Mycoplasma haemofelis, Mycoplasma haemominutum, Mycoplasma haematoparvum или из вирусов (не ограничивающихся ими) болезни Ньюкасл, (псевдочумы птиц), вируса инфекционного бронхита, птичьего пневмовируса, оспы птиц, инфекционного бурсального заболевания, вируса инфекционного ларинготрахеита, птичьего гриппа, вируса гепатита уток, вирусной чумы уток, инфекционной куриной анемии и вируса болезни Марека.
В другом варианте осуществления ген АВС-транспортера может быть полностью или частично заменен нуклеиновой кислотой, кодирующей белок, который выполняет функцию запуска или усиления иммунной реакции, например, интерлейкин или интерферон.
Вакцинные композиции
Живые аттенуированные бактерии пригодны для применения в вакцинных композициях. Вакцинная композиция может содержать иммуногенно эффективное количество живой аттенуированной бактерии, способной стойко сохраняться в субъекте, и фармацевтически приемлемый носитель или разбавитель. Описанные здесь вакцинная композиция или иммуногенные композиции могут вводиться интраназально, офтальмически, внутрикожно, внутрибрюшинно, внутривенно, подкожно, перорально, при помощи аэрозоля (спрей-вакцинация), через клоаку или внутримышечно. В частности, введение в виде глазных капель или аэрозоля является предпочтительным, когда субъектом являются птицы. Аэрозольное введение является особенно предпочтительным для введения вакцинной композиции или иммуногенной композиции большим количествам субъектов.
В одной форме настоящее изобретение обеспечивает живые аттенуированные вакцинные композиции для предотвращения или лечения животных и человека против инфекции бактерией, неаттенуированная форма которой содержит ген АВС-транспортера.
Фармацевтически приемлемый носитель или разбавитель может быть выбран из группы, состоящей из воды, солевого раствора, культуральной жидкости, стабилизаторов, углеводов, белков, белоксодержащих агентов, таких как бычья сыворотка или обезжиренное молоко, и буферов или их комбинации.
Стабилизатором может быть SPGA (на литр, SPGA содержит 74,62 г сахарозы, 0,52 г KH2PO4, 1,25 г K2HPO4, 0,912 г глутамата калия и 10 г сывороточного альбумина). Углеводами, применимыми в качестве фармацевтически приемлемого носителя или разбавителей, являются, например, сорбит, маннит, крахмал, сахароза, глюкоза, декстран или их комбинации.
Дополнительно, белки, такие как альбумин или казеин, или белоксодержащие агенты, такие как бычья сыворотка или обезжиренное молоко, могут быть применимы в качестве фармацевтически приемлемого носителя или разбавителей.
Буферы в качестве фармацевтически приемлемого носителя или разбавителей могут быть выбраны из группы, включающей малеат, фосфат, CABS (4-(циклогексинамино)-1-бутансульфоновую кислоту), пиперидин, глицин, цитрат, глицилглицин, малат, формиат, сукцинат, ацетат, пропионат, пиперазин, пиридин, какодилат, сукцинат, MES (гидрат 2-(N-морфолино)этансульфоновой кислоты и 4-морфолинэтансульфоновую кислоту), гистидин, бис-трис (2,2-бис(гидроксиметил)-2,2',2"-нитрилотриэтанол), фосфат, этаноламин, ADA (N-(2-ацетамидо)иминодиуксусную кислоту, N-(карбамоилметил)иминодиуксусную кислоту), карбонат, ACES (N-(2-ацетамидо)-2-аминоэтансульфоновую кислоту), PIPES (1,4-пиперазиндиэтансульфоновую кислоту), имидазол, BIS-TRIS-пропан (1,3-бис[трис(гидроксиметил)метиламино]пропан), BES (N,N-бис(2-гидроксиэтил)-2-аминоэтансульфоновую кислоту), MOPS (3-(N-морфолино)пропансульфоновую кислоту), HEPES (4-(2-гидроксиэтил)пиперазин-1-этансульфоновую кислоту), TES (2-[(2-гидрокси-1,1-бис(гидроксиметил)этил)амино]этансульфоновую кислоту), MOPSO (3-морфолино-2-гидроксипропансульфоновую кислоту), MOBS (4-(N-морфолино)бутансульфоновую кислоту), DIPSO (3-(N,N-бис[2-гидроксиэтил]амино)-2-гидроксипропансульфоновую кислоту), TAPSO (2-гидрокси-3-[трис(гидроксиметил)метиламино]-1-пропансульфоновую кислоту), TEA (триэтаноламин), пирофосфат, HEPPSO (гидрат (4-(2-гидроксиэтил)пиперазин-1-(2-гидроксипропансульфоновой кислоты)), POPSO (дегидрат пиперазин-1,4-бис(2-гидроксипропансульфоновой кислоты), трицин, гидразин, глицилглицин, TRIS (трис(гидроксиметил)аминометан), EPPS (4-(2-гидроксиэтил)-1-пиперазинпропансульфоновую кислоту), бицин, HEPBS (N-(2-гидроксиэтил)пиперазин-N'-(4-бутансульфоновую кислоту)), TAPS ([(2-гидрокси-1,1-бис(гидроксиметил)этил)амино]-1-пропансульфоновую кислоту), AMPD (2-амино-2-метил-1,3-пропандиол), TABS (N-трис(гидроксиметил)метил-4-аминобутансульфоновую кислоту), AMPSO (N-(1,1-диметил-2-гидроксиэтил)-3-амино-2-гидроксипропансульфоновую кислоту), таурин, борат, CHES (2-(циклогексиламино)этансульфоновую кислоту), AMP (2-амино-2-метил-1-пропанол), глицин, гидроксид аммония, CAPSO (3-(циклогексиламино)-2-гидрокси-1-пропансульфоновую кислоту), карбонат, метиламин, пиперазин, CAPS (3-(циклогексиламино)-1-пропансульфоновую кислоту) или любую их комбинацию.
С добавлением стабилизаторов вакцинная композиция пригодна для лиофилизации или высушивания замораживанием в соответствии со способами, известными в данной области. Таким образом, в одном варианте осуществления вакцинная композиция является высушенной вымораживанием или лиофилизированной.
В некоторых вариантах вакцинная композиция может содержать по меньшей мере одно соединение, имеющее адъювантную активность. Примеры адъювантов, подходящих для применения в вакцинных композициях, могут быть выбраны из группы, включающей полный адъювант Фрейнда или неполный адъювант Фрейнда, витамин Е, неионные блоксополимеры, мурамилдипептиды, сапонины, минеральное масло, растительное масло, гидроксид алюминия карбопол, фосфат алюминия, оксид алюминия, масляные эмульсии (например, Bayol F® или Marcol 52®), сапонины или солюбилизат витамина Е или любую их комбинацию. В некоторых вариантах осуществления эта вакцинная композиция может содержать адъюванты, применимые, в частности, для мукозного применения, например, термолабильный токсин E. coli или холерный токсин.
Вакцинные композиции данного изобретения могут быть приготовлены в виде аэрозолей (спрей-вакцин).
Вакцинные композиции данного изобретения могут быть приготовлены для офтальмического применения, например, в форме глазных капель. Например, эти глазные капли могут быть водными глазными каплями, неводными глазными каплями, суспензионными глазными каплями или эмульгированными глазными каплями. Приготовление этих глазных капель проводят суспендированием живых аттенуированных бактерий в водном растворителе, таком как стерилизованная дистиллированная вода, физиологический солевой раствор или т.п., или в неводном растворителе, таком как растительное масло, включая хлопковое масло, соевое масло, кунжутное масло, арахисовое масло, минеральное масло или т.п. Необязательно могут быть добавлены изотонирующие агенты, рН-корректирующие агенты, загустители, суспендирующие агенты, эмульгаторы и консерванты.
Изотонизирующие агенты, например, хлорид натрия, борную кислоту, нитрат натрия, нитрат калия, D-маннит, глюкозу. рН-Корректирующие агенты включают борную кислоту, безводный сульфит натрия, хлористоводородную кислоту, лимонную кислоту, цитрат натрия, уксусную кислоту, ацетат калия, карбонат натрия, буру и любые из перечисленных здесь буферов. Загустители могут быть выбраны из группы, включающей метилцеллюлозу, гидроксипропилметилцеллюлозу, поливиниловый спирт, натрий-хондроитинсульфат, поливинилпирролидон или любую их комбинацию.
Суспендирующий агент может быть выбран из группы, включающей полисорбат 80, гидрогенизированное полиоксиэтилированное касторовое масло 60, полиоксикасторовое масло или их комбинации. Дополнительно, примеры эмульгаторов включают лецитин желтка яйца и полисорбат 80. Кроме того, могут быть использованы консерванты, такие как хлорид бензалкония, хлорид бензетония, хлорбутанол, фенилэтиловый спирт, п-гидроксибензоаты или их комбинации.
Доза
Подлежащая введению доза будет, как правило, варьироваться в зависимости от возраста, массы и животного, подлежащего вакцинации, наряду со способом введения и типом патогена, против которого требуется вакцинация.
Эта вакцинная композиция может содержать любую дозу бактерий, достаточную для вызывания иммунной реакции. Например, дозы могут содержать от по меньшей мере 103 до приблизительно 105 аттенуированных бактерий, или от приблизительно 105 до приблизительно 107 аттенуированных бактерий, или от приблизительно 107 до приблизительно 109 аттенуированных бактерий, или от приблизительно 109 до приблизительно 1011 аттенуированных бактерий, или от приблизительно 1011 до приблизительно 1013 аттенуированных бактерий, или от приблизительно 1013 до приблизительно 1015 аттенуированных бактерий, или от приблизительно 1015 до приблизительно 1017 аттенуированных бактерий, или от приблизительно 1017 до приблизительно 1019, или по меньшей мере приблизительно 1019 аттенуированных бактерий.
В одном варианте осуществления, вакцинная композиция может доставляться в виде аэрозоля. Доза для обработки 1 кубического метра может содержать от по меньшей мере 106 до приблизительно 108 аттенуированных бактерий или от приблизительно 108 до приблизительно 1010 аттенуированных бактерий, или от приблизительно 1010 до приблизительно 1012, или от приблизительно 1012 до приблизительно 1014 аттенуированных бактерий, или от приблизительно 1016 до приблизительно 1018 аттенуированных бактерий, или от приблизительно 108 до приблизительно 1020 аттенуированных бактерий, или по меньшей мере приблизительно 1020 аттенуированных бактерий.
Способы введения
Вакцинные композиции данного изобретения могут вводиться интраназально, внутрикожно, через клоаку, внутривенно, внутрибрюшинно, подкожно, перорально, при помощи аэрозоля (спрей-вакцинация), в питьевой воде, в корме или внутримышечно. Для применения к домашней птице, предпочтительным является аэрозольное введение или введение в виде глазных капель.
Спрей-вакцинирование является особенно предпочтительным, так как оно делает возможной вакцинацию больших количеств субъектов простым распылением или созданием аэрозоля вакцинной композиции в присутствии подлежащих вакцинации субъектов. Распыление вакцинной композиции создает аэрозоль этой вакцинной композиции (живых аттенуированных бактерий), и при ингаляции этого аэрозоля животными живой аттенуированный вирус доставляется в животное и вторгается в дыхательные пути, имитируя распираторное заболевание. Этот способ вакцинации является особенно эффективным, так как трудоемкая, отнимающая время обработка отдельных животных не является необходимой.
Данное изобретение будет теперь описано более подробно со ссылкой на следующие конкретные примеры, которые не должны пониматься как ограничивающие каким-либо образом объем изобретения.
Примеры
Пример 1: Создание и идентификация библиотеки мутантов M. gallisepticum
Меченый сигнатурой (последовательностью-меткой) мутагенез (STM) использовали для генерирования и идентификации библиотеки мутантов M. gallisepticum. STM-стратегия показана на фиг.1. Уникальные ДНК-метки встраивают в транспозон, который используют для трансформации патогена, что приводит к случайной инсерции сигнатурной метки в геном. Время от времени, сигнатурная метка (Фиг.1A) может интегрироваться в ген и инактивировать ген с образованием инсерционной мутации. Затем используют способ отрицательного отбора для скрининга пула ST-мутантов в подходящей модели животного. Вводимый пул и ST-мутанты, извлеченные из животного, сравнивают при помощи ПЦР-амплификации индивидуальных меток с последующей гибридизацией. Этот способ идентифицирует отдельные мутанты, которые присутствуют во вводимом (перед отбором) пуле, но исчезают из пула, извлеченного после перехода в субъекта (в животное) (фиг.1В). Наиболее вероятно, что отсутствующие мутанты содержат мутации в генах, ответственных за колонизацию и рост в субъекте, и, следовательно, эти гены, по-видимому, кодируют связанные с вирулентностью детерминанты.
Материалы и способы
Бактериальный штамм и условия культивирования
M. gallisepticum (Mg) Ap3AS был первоначально выделен из альвеолярных мешочков бройлерной курицы в Австралии и является высоко патогенным. Его выращивали в модифицированном бульоне Фрея (МВ), содержащем 10% свиную сыворотку, при 37°С до поздней логарифмической фазы (рН приблизительно 6,8).
Меченные сигнатурой транспозоны
Библиотеку ST-мутантов готовили с использованием плазмиды pISM 2062.2, несущей транспозон Tn4001mod, который содержал ген устойчивости к гентамицину. Каждая сигнатурная метка состояла из уникальной олигонуклеотидной ДНК-последовательности 40 п.н. ([NK]20; N = A, C, G или T; K = G или T), фланкированной двумя неизменяемыми плечами 20 п.н., которые делают возможными амплификацию и мечение уникальных районов при помощи ПЦР с использованием пары праймеров P2/P4 или P3/P5. Сигнатурную метку клонировали в сайт рестрикции KpnI pISM 2062.2 (фиг.1А). Последовательности индивидуальных сигнатурных меток приведены в списке в таблице 1.
ДНК-последовательности сигнатурных меток, используемых в этом исследовании

# набор праймеров P2/P4 генерировали из неизменяемых плечей сигнатурных меток. Указан ожидаемый размер (п.н.) продуктов после ПЦР-амплификации.
Меченный сигнатурой мутагенез
Трансформация сигнатурными метками
Разведения Mg Ap3AS выращивали в 1,5 мл MB в течение ночи при 37°С. Разведения, обнаруживающие изменение окраски, объединяли, и клетки осаждали центрифугированием в течение 5 минут при 16000 g при комнатной температуре. Эти клетки ресуспендировали в 200 мкл охлажденного HEPES-сахарозного буфера (8 мМ HEPES, 272 мМ сахароза (pH 7,4)). Клетки осаждали центрифугированием в течение 5 минут при 16000 g при комнатной температуре и промывали еще два раза охлажденным HEPES-сахарозным буфером, затем окончательно ресуспендировали в 400 мкл HEPES-сахарозного буфера, охлажденного до 4°С.
Аликвоту клеток 100 мкл помещали на лед и добавляли 10 мкг плазмидной ДНК. Затем смесь клеток-плазмидной ДНК переносили в охлажденную gap-кювету 0,2 см (Bio-Rad) на льду и сразу же электропорировали с использованием Gene Pulser™ (Bio-Rad) с установками 2,5 кВ, устойчивостью 100 Ω и емкостью 25 мкФ. Затем электропорированные клетки осторожно ресуспендировали в 1 мл охлажденного (4°С) МВ и инкубировали при комнатной температуре в течение 10 минут с последующей инкубацией при 37°С в течение 2-3 часов, в зависимости от того, было ли изменение окраски. Пробу 500 мкл электропорированной культуры распределяли на чашке МА, содержащей гентамицин при 160 мкг/мл, осторожным поворачиванием, и избыток удаляли при помощи пастеровской пипетки. Затем чашку сушили и инкубировали при 37°С в течение 7-10 дней в герметичном сосуде.
Подтверждение ST-мутантов
Mg-колонии, растущие на чашках, отбирали с использованием пастеровской пипетки, помещали в 1 мл МВ, содержащего гентамицин (160 мкг/мл), и эту культуру инкубировали, пока среда не обнаруживала изменение окраски. Клетки из объема 200 мкл культуры осаждали центрифугированием в микрофужной пробирке при 16000 g в течение 5 минут при комнатной температуре, и осадок ресуспендировали в одной десятой исходного объема дистиллированной воды. Затем клетки нагревали при 100°С в течение 5 минут, и 2 мкл из них использовали в качестве матрицы для ПЦР. Для подтверждения присутствия ST-транспозона использовали праймеры Р2 и Р4 (таблица 2) для амплификации района в неизменяемых плечах, которые содержали уникальную последовательность (фиг.3). ПЦР проводили в реакционном объеме 20 мкл, и она содержала 2 мкл 10 × реакционного буфера, 1 мкМ каждого праймера, 200 мкМ каждого dNTP, 1,5 мМ MgCl2, 1,5 Е ДНК-полимеразы Taq (Promega) и 2 мкл матричной ДНК. Реакции выполняли в термоциклере (Omnigene, Hybaid) с одним циклом при 98°C в течение 2 минут, с последующими 35 циклами 94°C в течение 30 секунд, 50°C в течение 30 секунд и 72°С в течение 40 сек, и конечным инкубированием при 72°C в течение 7 минут. ПЦР-продукты подвергали электрофорезу в 2% агарозном геле вместе со стандартами размеров (pUC18, расщепленной HaeIII).
Олигонуклеотиды, используемые в исследованиях STM

Определение точек инсерции транспозона
Выделение ДНК из ST-мутантов
Каждый ST-трансформант выращивали в 40 мл MB, дополненного 160 мкг гентамицина/мл, при 37°С до поздней логарифмической фазы (pH приблизительно 6,8). Клетки собирали центрифугированием при 20000 g в течение 30 минут и промывали дважды в охлажденном PBS, добавляли SDS для лизиса этих клеток, и затем раствор пропускали через иглу калибра 26 для гидродинамического фрагментирования геномной ДНК. Экстракцию геномной ДНК выполняли с набором HighPure PCR (Roche) в соответствии с протоколом изготовителя, с минорными модификациями. Исходную обработку лизоцимом опускали и ДНК элюировали из колонки в объеме 50 мкл, а не 200 мкл 10 мМ Трис-буфера (рН 8,0). Количество очищенной ДНК определяли посредством электрофореза пробы 1 мкл в 0,7% агарозном геле вместе со стандартами молекулярной массы известной концентрации (ДНК фага λ, расщепленной HindIII).
Секвенирование геномной ДНК
Процедура для секвенирования геномной ДНК была адаптирована и модифицирована в соответствии с Wada (2000). Праймер IGstmGenmeF3 (таблица 2), который связывается с Tn4001, использовали для секвенирования через точку соединения транспозон-геномная ДНК и в геномную ДНК Ap3AS. Эту последовательность определяли с использованием химического способа ABI PRISM Big Dye 3.1 Terminator (Applied Biosystems Incorporated). Каждая реакционная смесь состояла из 2-3 мкг очищенной геномной ДНК, 10 мкМ праймера и 4 мкл смеси ферментов Big Dye 3.1, и циклическое секвенирование выполняли в iCycler™ (Bio-Rad) с использованием одного цикла при 95°C в течение 5 минут, с последующими 60 циклами 95°C в течение 30 секунд, 55°C в течение 30 секунд и 60°C в течение 4 минут. Затем продукт очищали в соответствии с рекомендациями изготовителя и затем анализировали с использованием капиллярного секвенатора ABI 3100 и релевантного программного обеспечения (Applied Biosystems Incorporated). Сайт инсерции для каждого ST-транспозона определяли с использованием программы BLAST (National Center for Biotechnology Information, NCBI) или версии 3.3t07 FASTA (Pearson and Lipman, 1988) для сравнения ДНК-последовательности с последовательностью генома штамма M. gallisepticum Rlow.
Детектирование Mg ST-мутантов
Синтезировали олигонуклеотиды, комплементарные каждой из 37 сигнатурных меток. Олигонуклеотиды ресуспендировали в буфере ТЕ до концентрации 100 мкМ. Объем 10 мкл раствора каждого олигонуклеотида (содержащий 3 пикомоля) наносили в виде пятен на нейлоновую мембрану Hybond-N+ (Amersham Pharmacia Biotech) с использованием прибора для микрофильтрации Bio-Dot SF® (Bio-Rad) в соответствии с инструкциями изготовителя, и затем олигонуклеотиды фиксировали на мембране УФ-сшиванием в течение 3 минут. Синтезировали ДНК-зонды, соответствующие каждой сигнатурной метке, с использованием описанной выше ПЦР-реакции, за исключением того, что олигонуклеотидные праймеры P2 и P4 были коммерчески помечены дигоксигенином (DIG) (Roche). DIG-меченные зонды гибридизовали с мембранами в буфере DIG Easy Hyb (Roche) в течение 16-18 часов при 40°C во встряхиваемой водяной бане, и затем мембраны промывали в соответствии с инструкциями изготовителя, за исключением того, что вторую промывку выполняли при 45°C. Связанные зонды детектировали с использованием набора для люминесцентного детектирования DIG (Roche), и детектирование регистрировали с использованием пленки Biomax (Kodak). Проводили также ПЦР для подтверждения присутствия гена устойчивости к гентамицину и вида трансплантата микоплазмы. Каждую ПЦР-реакцию выполняли с использованием 1,5 Е ДНК-полимеразы Taq в 25 мкл реакционной смеси, содержащей 1,5 мМ MgCl2, 200 мкМ каждого dNTP и 1 мкМ каждого праймера. Район Mg-гена 16S рРНК амплифицировали инкубированием при 95°C в течение 5 минут, затем посредством 35 циклов 95°C в течение 10 секунд, 58°C в течение 10 секунд и 72°C в течение 25 секунд. Ожидаемый продукт имел размер 219 п.н. Фрагмент гена устойчивости к гентамицину амплифицировали инкубированием при 95°C в течение 2 минут, затем посредством 28 циклов 95° в течение 30 секунд, 60°C в течение 30 секунд и 72°C в течение 15 секунд, с конечным инкубированием при 72°C в течение 5 минут. Ожидаемый размер продукта был 278 п.н.
Перекрестная гибридизация между сигнатурными метками с использованием системы детектирования DIG
Пул DIG-меченных сигнатурных меток гибридизовали с одной мембраной, содержащей 37 олигонуклеотидов с сигнатурными метками, как описано выше. Можно было ожидать, что DIG-меченные метки будут связываться только с соответствующими уникальными олигонуклеотидами, если бы не было перекрестной гибридизации между этими метками и зондами. Например, ожидалось, что пул, содержащий меченые Tag 02, 03 и 04, связывается с олигонуклеотидными метками Tag 02, 03, 04 на дот-блоте, и любые другие положительные реакции должны были рассматриваться как перекрестные реакции. Реакции перекрестной гибридизации оценивали с использованием 13 различных пулов меченых сигнатурных меток.
Предварительный эксперимент с животными
Расчет количеств жизнеспособных клеток ST-мутантов
Количество жизнеспособных клеток каждой культуры ST-мутантов, измеряемое в виде единиц изменения окраски на мл (CCU/мл), определяли с использованием следующей процедуры. В каждую лунку стерильного 96-луночного микротитрационного планшета (Nunculon, Nunc) добавляли 225 мкл стерильного MB, содержащего гентамицин (160 мкг/мл). К первому столбцу из 8 лунок добавляли 25 мкл культуры мутанта в каждую лунку, перемешивали несколько раз пипетированием, и 25 мкл из каждой лунки переносили в следующий столбец и перемешивали с использованием свежего набора кончиков пипеток. Эту последовательность серийных 10-кратных разведений повторяли до столбца 10, из которого выбрасывали 25 мкл. Столбцы лунок 11 и 12 использовали в качестве отрицательных контролей. Затем этот планшет герметизировали с использованием Linbro® plate seal (ICN Biochemicals) и инкубировали при 37°C в течение периода до 2 недель. Снижение рН среды отражало рост этой культуры и указывалось изменением окраски с красной до желтой фенолового красного индикатора в среде. Столбец 1 рассматривали как разведение 1/10, и после коррекции на первоначальные разведения культуры рассчитывали количества CCU/мл с использованием таблиц наиболее вероятных количеств. Доза каждого трансформанта, необходимая для инокуляции кур, была 1×107 CCU/мл.
План эксперимента
Шестнадцать 4-недельных не содержащих конкретного патогена (SPF) кур породы белый леггорн содержали в изоляторе из стеклопластика с положительным давлением (давлением выше атмосферного). Серию разведений десяти ST-мутантов выращивали в течение ночи в микротитрационном планшете в МВ, содержащем гентамицин при 160 мкг/мл (таблица 3). Рост каждого клона оценивали на следующий день, и 100 мкл брали из разведения, оцениваемого как разведение, находящееся в логарифмической фазе, и добавляли к пулу. Затем объединенные ST-мутанты разводили еще в десять раз в МВ, и этот объединенный пул использовали в качестве инокулята. Двенадцать из шестнадцати кур инфицировали объединенными ST-мутантами введением в виде аэрозоля. Остальные 4 птицы служили в качестве контроля в контакте и были помещены в изолятор 3 дня спустя после аэрозольного инфицирования. Шесть из инфицированных аэрозолем птиц эвтанизировали, и брали пробы спустя 14 дней после инфицирования, и то же самое выполняли в отношении остальных птиц, в том числе всех находящихся в контакте птиц, спустя 28 дней после инфицирования.
Тестированные транспозоны
Сбор и анализ проб
Пробы крови собирали у всех птиц перед аэрозольным инфицированием и перед эвтаназией и оставляли для свертывания при комнатной температуре на 3 часа. Затем собирали сыворотку центрифугированием пробы при 10000 g в течение 5 минут при комнатной температуре и затем тестировали при помощи RSA-теста для определения реакций в виде антител против M. gallisepticum. RSA-тест выполняли смешиванием 25 мкл неразведенной сыворотки и 25 мкл окрашенного коммерческого антигена Mg RSA (Intervet International) на стеклянной чашке и качанием в течение 1 минуты. Тест оценивали в баллах от 0 (отсутствие агрегации) до 4 (большие скопления антигена). Каждый тест включал как отрицательную сыворотку, так и положительную сыворотку в качестве контролей. Кур эвтанизировали ингаляцией диоксида углерода. Исследовали макроскопические повреждения альвеолярных мешочков и оценивали в баллах на тяжесть по шкале 0-3 следующим образом:
0 = отсутствие повреждений, тонкая прозрачная пленка альвеолярных мешочков;
1 = малые пятна творожистого (кавеозного) фибрина, присоединенные к этим альвеолярным мешочкам;
2 = альвеолярные мешочки кажутся мутными и утолщенными, с умеренными количествами творожистого фибрина, присоединенными к альвеолярным мешочкам;
3 = утолщенные альвеолярные мешочки и поверхность, покрытая толстым творожистым фибрином.
Оценка полбалла давалась в отношении повреждений, которые находятся между этими определениями. Регистрировали балльные оценки для альвеолярных мешочков левой и правой передней торакальной части, альвеолярных мешочков левой и правой задней торакальной части и левых и правых брюшных альвеолярных мешочков. Сумму этих шести балльных оценок использовали а качестве конечной балльной оценки для каждой отдельной птицы. Брали мазки из альвеолярных мешочков, трахеи, легкого, селезенки, печени, почки и головного мозга каждой птицы. Мазки использовали для инокуляции планшетов МА, содержащих 160 мкг гентамицина/мл, а также планшета МА без гентамицина, и затем помещали в 3 мл МВ, дополненного гентамицином при 160 мкг/мл. Планшеты MA инкубировали при 37°С и исследовали с использованием бинокулярной препаровальной лупы спустя 7-10 дней. Предварительное исследование детектировало потерю транспозона Tn4001 из некоторых мутантов во время экспериментов in vivo спустя 14 дней после инокуляции. Эти мутанты восстанавливали фенотип дикого типа, но не могли выживать при давлении отбора гентамицина. Когда количества колоний на чашках, не содержащих гентамицина, были в десть или более раз большими, чем количества колоний на чашках, содержащих гентамицин, подозревали потерю транспозона. МВ-культуры инкубировали при 37°С, и ДНК экстрагировали из бульонов, обнаруживающих изменение окраски, и использовали в качестве матрицы в ПЦР, как описано выше, для амплификации уникальных районов метки с использованием пары праймеров P2/P4.
Результаты
Подтверждение ST-мутантов
Ожидаемый размер ПЦР-продукта, генерируемого из сигнатурных меток с использованием пары праймеров P2/P4, находился между 79 и 81 п.н., за исключением размера продукта ПЦР из Tag 25, который был равен 67 п.н. Каждую плазмиду, содержащую сигнатурную метку, вводили в штамм Mg Ap3AS, и отбирали трансформанты случайным образом и выращивали в среде, дополненной гентамицином. Все МВ-культуры, обнаруживающие рост, подвергали ПЦР для детектирования присутствия ST-мутанта, содержащего уникальную сигнатурную метку (фиг.2). Устанавливали библиотеку ST-мутантов с по меньшей мере 9 трансформантами, генерированными из каждой сигнатурной метки, и использовали ее для дальнейших экспериментов.
Определение сайта инсерции транспозона
Сайт инсерции транспозона для каждого из десяти Mg ST-клонов, использованных в исследованиях, описанных в этой части, определяли прямым секвенированием геномной ДНК (таблица 3). Транспозоны инсертировались в районы, кодирующие либо уникальные, либо консервативные гипотетические белки в пяти ST-мутантах.
Оптимизированное детектирование ST-мутантов
Перекрестная гибридизация между сигнатурными метками
Большинство меток были специфическими, только с небольшой перекрестной гибридизацией с олигонуклеотидами, комплементарными другим меткам. Имелась сильная перекрестная реакция между Tag 02 и 37 (в пулах A, E, G и H). Некоторую перекрестную гибридизацию детектировали также между Tag 15 и 30 (в пулах D, I и J), но не детектировали перекрестной гибридизации с пулом В. Имелась односторонняя гибридизация между Tag 13, когда она присутствовала в пуле, и Tag 28. Это можно также видеть с Tag 29 и 23 (пулы D и H) и Tag 29 и 24 (пулы D и H).
Предварительное оценивание инфекции кур с ST-мутантами
Серологическое тестирование и макроскопическое исследование альвеолярных мешочков
Реакции анти-M. gallisepticum-антитела определяли при помощи RSA (таблица 4) до и после инфицирования. До инфицирования не детектировали M. gallisepticum-специфического антитела в птицах. Из кур, инфицированных аэрозолем, две птицы имели балльные оценки RSA, более высокие, чем балльная оценка при 2 неделях после инфицирования, и все птицы были положительными спустя 4 недели после инфицирования, с балльными оценками RSA между 1 и 4. Только один из контролей в контакте имел балльную оценку RSA 1. Слабые повреждения альвеолярных мешочков (балльная оценка 0,5) наблюдали в 2 птицах из 6 птиц спустя 2 недели после инфицирования и в 3 птицах из 6 птиц при 4 неделях после инфицирования, в то время как одна птица имела тяжелые повреждения в брюшных альвеолярных мешочках (балльная оценка 2,5). Только один из контролей в контакте имел слабые повреждения (балльная оценка 0,5). Подробные результаты показаны в таблице 4.
ST-мутанты, полученные из различных сайтов в птицах в предварительном эксперименте
* Детектированный tag ID согласно гибридизации и положительный согласно ПЦР на присутствие гена гентамицина и гена Mg 16s рРНК.
+: показывает, что Mg-колонии росли на МА или что изменение окраски наблюдали в МВ.
-: не наблюдали роста на чашке МА или в МВ.
Реверсия и идентификация выделенных ST-мутантов
MA-чашки испытывали на рост M. gallisepticum в присутствии и в отсутствие гентамицина. Не обнаруживали потери транспозона из ST-мутантов после пассажа in vivo. M. gallisepticum выделяли только на МА-чашках, инокулированных мазками альвеолярных мешочков одной птицы и трахей двух птиц при 2 неделях после инокуляции, и трахей четырех кур при 4 неделях после инфицирования. M. gallisepticum выделяли также из мазка, взятого из сердца одной птицы спустя 14 дней после инфицирования. M. gallisepticum не выделяли из мазков любого органа в контролях в контакте (таблица 4). Один культуральный бульон мазка альвеолярных мешочков, взятого при 2 неделях после инфицирования, и четыре из трахей обнаруживали изменение окраски с 3 из 10 уникальных меток, детектируемых DIG-гибридизацией из этих проб. Бульонные культуры, обнаруживающие изменение окраски, получали из мазков альвеолярных мещочков двух птиц и мазков трахей трех птиц в день 28 после инфицирования, а также из мазков других органов, в том числе легких пяти птиц, сердец четырех птиц, печеней двух птиц, почек трех птиц, селезенок четырех птиц и головного мозга одной птицы. Бульонные культуры, обнаруживающие изменение окраски, получали также из мазков трахей одной птицы, почек двух птиц и головного мозга двух птиц контроля в контакте. Однако, ПЦР, проводимые для детектирования присутствия гена устойчивости к гентамицину и гена Mg 16S рРНК, в культурах, обнаруживающих изменение окраски после 21 дня инкубирования, все были отрицательными, что позволяет предположить, что изменение окраски, наблюдаемое в этих культурах, не приводило к росту M. gallisepticum. Мутанты, несущие Tag 02, 32, 35 и 37, были действительно получены из альвеолярных мешочков, трахей этих птиц и из головного мозга одной птицы с наиболее обычными детектируемыми метками, являющимися Tag 32 и 37 (таблица 4). Несколько бульонных культур обнаруживали изменение окраски в пределах 7 дней после инкубирования, но не были детектированы метки DIG-гибридизацией (данные не показаны).
Пример 2: In vivo анализ ST-мутантов M. gallisepticum
Встраивание
STM-способ состоит из двух основных стадий: создание библиотек меченых мутантов in vitro и затем отрицательный отбор in vivo в животном. С использованием модифицированной системы гибридизационного детектирования, описанной в примере 1 выше, вводимый пул содержал только мутанты, полученные из меток, которые не гибридизуются перекрестно друг с другом, а также дают ясные сигналы для детектирования. Библиотеку ST-мутантов примера 1 генерировали с использованием 37 различных меченых транспозонов. Этот пример описывает скрининг 102 ST-мутантов, содержащих 34 различные метки, для идентификации генов, участвующих в вирулентности или необходимых для выживания in vivo, с использованием отрицательного отбора в инфицированных курах.
Материалы и способы
Определение сайтов инсерции в ST-мутантах
Геномную ДНК из каждого ST-мутанта подвергали прямому секвенированию с использованием специфического праймера (таблица 2). Для тех ST-мутантов, из которых было невозможным получение данных последовательности или которые генерировали смешанные сигналы секвенирования после конца транспозона, использовали блоттинг по Саузерну для определения количества инсерций транспозонов. Затем ST-мутанты с более чем одной инсерцией транспозона исключали из дальнейших попыток идентификации точек инсерции транспозонов. Зонды для блоттинга по Саузерну готовили с использованием DIG-меченных праймеров (P2 и P4) для амплификации района сигнатурной метки (таблица 2). Геномную ДНК ST-мутанта расщепляли в течение ночи 20 Е рестрикционной эндонуклеазы BglII (New England Biolabs) при 37°С, и полученные фрагменты разделяли в 0,7% агарозном геле в течение ночи при 2,5 В/см. Затем разделенные фрагменты переносили на нейлоновую мембрану Hybond N+, гибридизованную с DIG-меченным зондом и гибридизацию детектировали с использованием набора для люминесцентного детектирования DIG (Roche) в соответствии с инструкциями изготовителя, и люминесценцию регистрировали на пленке Biomax (Kodak).
Скрининг ST-мутантов в инфицированных птицах
Приготовление ST-мутантов
Серии десятикратных разведений каждого из 34 ST-мутантов, каждый из которых содержал отличающуюся метку, для каждого из трех инокулятов (Групп А, В и С) (102 мутанта в целом для трех экспериментальных групп) готовили в микротитрационных планшетах в МВ, дополненном гентамицином (160 мкг/мл), и инкубировали при 37°С в течение ночи (таблица 5). Разведение каждого мутанта, которое продуцировало подходящее изменение окраски, использовали для инокуляции. Брали пробу 0,1 мл подходящей культуры каждого мутанта и смешивали с культурами других мутантов для создания объединенного инокулята. Затем объединенные ST-мутанты использовали сразу же для аэрозольной инокуляции кур.
Сайты инсерции транспозонов в ST-мутантах, генерированных в этом исследовании (Инокулят А)
(% гена до точки инсерции)
Первоначальный скрининг
План эксперимента
В целом 90 четырехнедельных SPF-кур случайным образом распределяли на 3 группы, каждую из которых содержали отдельно (30 птиц на группу) в изоляторах из стеклопластика с положительным давлением (давлением выше атмосферного). Двадцать птиц в каждой группе инокулировали пулом ST-мутантов посредством аэрозоля, причем остальные 10 кур использовали в качестве контролей в контакте, которые помещали с инокулированными птицами спустя три дня после подвергания действию аэрозоля. Контроли в контакте использовали для исследования способности контактного распространения ST-мутантов. В день 14 после инфицирования брали пробы крови из десяти инокулированных кур, и их эвтанизировали и исследовали в отношении повреждений. Пробы крови тестировали на антитела против M. gallisepticum с использованием RSA-теста, как описано ниже. Всех птиц испытывали на макроскопические повреждения альвеолярных мешочков, и их оценивали на тяжесть по шкале 0-3, как описано ниже. Мазки альвеолярных мешочков и трахеи каждой птицы культивировали для получения микоплазм, как описано ниже. ДНК экстрагировали из каждой бульонной культуры, которая изменяла окраску, и использовали в качестве матрицы в ПЦР для амплификации уникального района метки. Затем ПЦР-продукты использовали в блотах по Саузерну для идентификации ST-мутантов, которые присутствовали, как описано ниже. Чашки МА проверяли после инкубации в течение 10 дней, как описано ниже. Остальных птиц эвтанизировали, обследовали и брали пробы с использованием тех же самых процедур при 28 днях после инокуляции.
Подтверждающий скрининг
Приготовление ST-мутантов M. gallisepticum
Разведения ST-мутантов, которые не были повторно выделены в первоначальном эксперименте скрининга, выращивали в МВ, содержащем 160 мкг гентамицина/мл, при 37°С в течение ночи. Разведение каждого ST-мутанта, которое вызывало подходящую степень изменения окраски, использовали для инокуляции. Пробу 0,1 мл культуры каждого мутанта смешивали с пробами других мутантов, и этот пул затем разводили в десять раз в МВ для применения в качестве нового объединенного инокулята, который использовали для инокуляции кур посредством аэрозоля.
План эксперимента
Подтверждающий эксперимент проводили для сужения количества кандидатных ST-мутантов для дефинитивного тестирования. План и способы этого эксперимента были сходными с первоначальным экспериментом скрининга, который показан на фигуре 4. В целом сорок 4-недельных кур SPF распределяли на две группы. ST-мутанты, которые не могли быть детектированы в инокулированных птицах в первоначальном эксперименте скрининга, относили к одному из двух инокулятов, каждый из которых использовали для инокуляции одной из двух групп этих птиц посредством аэрозоля. Контроли в контакте не включали в этом эксперименте. Однако всех 40 птиц помещали в тот же самый изолятор из стеклопластика с повышенным давлением, и, следовательно, каждая группа могла использоваться в качестве контроля в контакте для другой группы. Сбор проб выполняли спустя 14 дней после инокуляции с использованием тех же самых процедур, описанных ранее. Вкратце, пробы крови собирали перед инокуляцией и непосредственно перед эвтаназией. Мазки из альвеолярных мешочков и трахеи собирали post mortem для выделения микоплазм. Всех птиц исседовали на макроскопические повреждения альвеолярных мешочков. Сыворотки тестировали с использованием RSA-анализа. ДНК экстрагировали из каждой бульонной культуры, которая изменяла окраску, и использовали в качестве матрицы в ПЦР для амплификации районов уникальных меток, которые затем использовали в качестве зондов в дот-блот-гибридизационных анализах. Чашки МА проверяли после инкубации в течение 10 дней при 37°С, как описано ранее. Несколько колоний собирали из чашек МА и культивировали в МВ, дополненном гентамицином, при 37°С, пока не наблюдали изменения окраски. ДНК экстрагировали и использовали в качестве матрицы в ПЦР-анализах для подтверждения вида микоплазмы амплификацией фрагмента 219 п.н. гена Mg 16S рРНК, как описано ранее в примере 1 (фиг.4).
ПЦР-подтверждение специфических ST-мутантов, выделенных из инфицированных птиц
На основании результатов этого секвенирования конструировали ПЦР-праймеры, которые были специфическими для каждого ST-мутанта (таблица 2). Их использовали в ПЦР-анализах с праймером IGstmGenmeF3 для детектирования каждого из ST-мутантов в культурах из инокулированных птиц. ПЦР проводили в объемах 20 мкл, содержащих 2 мкл экстрагированной ДНК в качестве матрицы, 2 мкл 10 × реакционного буфера, 1 мкМ кажого праймера, 200 мкМ каждого dNTP и 1,5 Е ДНК-полимеразы Taq (Promega). ПЦР выполняли в термоциклере (Omnigene, Hybaid), с одним циклом при 95°C в течение 2 минут с последующими 35 циклами 94°C в течение 45 секунд, 52°C в течение 45 секунд и 72°C в течение 1 минуты и конечным инкубированием при 2°С в течение 7 минут.
Результаты
Картирование точек инсерции в ST-мутантах
Сайт инсерции транспозона определяли в 91 ST-мутанте с использованием способа прямого геномного секвенирования. Достоверные данные последовательности не могли быть получены для 11 мутантов (таблица 5). В большинстве случаев сайт инсерции был одним и тем же в Группах А, В и С, и единственными исключениями являются ST-мутанты 01-1, 02-2, 02-3, 04-3, 09-3 и 17-3.
ST-мутанты, которые не могли быть секвенированы непосредственно, подвергали блоттингу по Саузерну для определения количества инсерций транспозона. Секвенирующие хроматографы показали, что первые 100 п.н. данных, которые соответствуют транспозону, были данными хорошего качества, но что получали смешанные сигналы после достижения точки соединения транспозона и геномной последовательности (фиг.5А). Поскольку не было предсказано, что рестрикционная эндонуклеаза BglII расщепляет в последовательности транспозона, количество полос, связанных этим зондом, должно отражать, сколько раз этот транспозон присутствует в геномах ST-мутантов. Множественные инсерции транспозона были подтверждены в ST-мутантах 15-1, 15-2, 15-3, 25-1, 25-2, 25-3, 34-1, 34-2, 34-3, 09-3 и 17-3 посредством блоттинга по Саузерну, и мутанты, несущие одну и ту же сигнатурную метку, имеют, по-видимому, идентичные инсерции (таблицы 5 и 6, фиг.5В).
Первоначальный эксперимент скрининга
Патологические и серологические открытия
RSA-результаты из каждой экспериментальной группы показаны в таблице 7. В сыворотке из любой птицы в момент инокуляции не детектированы анти-микоплазма-антитела. Обычно большее число сывороток были положительными при 4 неделях, чем при 2 неделях после инфицирования, и антитела не были детектированы в контроле в контакте ни в одной группе. Большее число птиц имели повреждения альвеолярных мешочков при 2 неделях (2, 2 и 4 в Группах A, B и C, соответственно), чем при 4 неделях (0, 1 и 2 в Группах А, В и С, соответственно) после инокуляции. Более тяжелые повреждения альвеолярных мешочков наблюдали при 2 неделях после инокуляции (балльные оценки 0,5 в Группе А, 1,0-2,0 в Группе В и 0,5-2,5 в Группе С), за исключением того, что в Группе С одна курица имела балльную оценку повреждения 3,0 при 4 неделях после инокуляции. Больше повреждений альвеолярных мешочков было обнаружено в Группе С. Тяжесть повреждений не коррелировала хорошо с результатами RSA, т.е. некоторые птицы имели высокие балльные оценки RSA, но не имели детектируемых повреждений альвеолярных мешочков.
Классификация сайтов инсерции транспозонов в ST-мутантах в этом исследовании
Результаты RSA, балльные оценки повреждения альвеолярных мешочков и меток, детектированных в каждой экспериментальной группе в первоначальном скрининге
# Одна инокулированная птица и две птицы в контакте умерли в группе А и две птицы в контакте умерли в группе С перед post mortem.
^ Суммирование недетектированных сигнатурных меток трех групп.
Идентификация выделяемых ST-мутантов
Результаты для бульонных культур, взятых из птиц в дни 14 и 28 после инокуляции, суммированы в таблице 7. Картина выделения M. gallisepticum была сходной с картиной, наблюдаемой в предварительном эксперименте, с большими выделениями, полученными из трахей, чем из альвеолярных мешочков. Как правило, больше выделений было сделано при 2 неделях, чем при 4 неделях после инфицирования. ST-мутанты не были повторно выделены из птиц в контакте в группе В, хотя пять различных ST-мутантов были повторно выделены из птиц в контакте в группе А и две из птиц в группе С. ST-мутант, несущий Tag 20, был наиболее часто извлекаемым из альвеолярных мешочков, а также из трахей, и имел высокую степень детектирования во всех трех группах. Следующим наиболее часто выделяемым мутантом был мутант, несущий Tag 28. Шестнадцать ST-мутантов, представленных 12 различными метками, не могли быть повторно выделенными, включая ST-мутанты 02-1 (-2), 03-1 (-2/-3), 04-1 (-2), 04-3, 06-1 (-2/-3), 09-1 (-2), 09-3, 10-1 (-2/-3), 15-1 (-2/-3), 17 -1 (-2), 17-3, 22-1 (-2/-3), 23-1 (-2/-3), 25-1 (-2/-3), 33-1 (-2/-3) и 34-1 (-2/-3).
M. gallisepticum не был выделен на чашках МА, инокулированных мазками из альвеолярных мешочков или трахей любых кур в контакте. M. gallisepticum выделяли на чашках МА, инокулированных мазками, собранными спустя 2 недели после инокуляции из альвеолярных мешочков трех, одной и пяти птиц в группах А, В и С, соответственно. Их выделяли также на чашках МА, инокулированных мазками из трахей пяти кур в группе С, но не из каких-либо птиц в группах А или В. M. gallisepticum выделяли на чашках МА, инокулированных мазками, собранными из альвеолярных мешочков четырех, двух и шести птиц, и из трахей одной, двух и семи кур в группах А, В и С, соответственно, при 4 неделях после инокуляции. Почти в каждом случае, когда микоплазмы выделяли посредством агаровой культуры, их также выделяли посредством бульонной культуры. Ни в одном ST-мутанте не детектировали потерю транспозона.
Подтверждающий эксперимент скрининга
Шестнадцать ST-мутантов, которые не удалось детектировать в первоначальном эксперименте скрининга, делили на две группы. Данные последовательности в отношении сайта инсерции транспозона и блоттинг по Саузерну показали, что транспозон в ST-мутантах 04-1 и 04-2 имел один и тот же сайт инсерции, но другой сайт инсерции относительно сайта инсерции 04-3. Сайты инсерции транспозона в ST-мутантах 09-1 и 09-2 были идентичными, тогда как мутант 09-3 имел множественные инсерции. Та же самая ситуация была обнаружена также в ST-мутантах 17-1, 17-2 и 17-3. Ни один из этих мутантов не мог быть детектирован в первоначальном эксперименте скрининга, поэтому они были помещены в два разных инокулята для получения возможности их различения. Когда имелась потенциальная передача между этими группами, и эти три сигнатурные метки детектировали дот-блот-гибридизацией, могла быть использована ПЦР для идентификации мутантов, повторно выделенных из птиц.
Патологические и серологические оценивания
Одна птица умирала в группе А и одна в группе В во время этого эксперимента. Тяжелые повреждения альвеолярных мешочков (балльная оценка 2,5) наблюдали у одной птицы и слабые повреждения (0,5 и 1,0) у других двух кур в группе А. Одна птица имела слабые повреждения альвеолярных мешочков (1,0) в группе В. Перед инокуляцией не было детектировано анти-M gallisepticum-антитело. При 2 неделях после инокуляции 15 из 19 кур в группе А были RSA-положительными, тогда как в группе В 14 из 19 кур были положительными. Подробные результаты показаны в таблице 8.
Результаты RSA, балльные оценки повреждений альвеолярных мешочков и меток, детектированных в каждой экспериментальной группе в подтверждающем скрининге
# Птиц в группе А инокулировали пулом, содержащим ST-мутанты 02-2, 03-1, 04-1, 06-1, 09-1, 10-1, 15-1 и 17-2; птиц в группе В инокулировали пулом, содержащим ST-мутанты 04-3, 09-3, 17-3, 22-1, 23-1, 25-1, 33-1 и 34-1. Одна птица умерла в группах А и В перед post mortem.
^ Суммирование недетектированного ST-мутанта двух групп.
Повторное выделение ST-мутантов
M. gallisepticum более часто выделяли на чашках МА, инокулированных мазками, взятыми из трахей, чем из альвеолярных мешочков, в обеих группах. M. gallisepticum выделяли на чашках МА, инокулированных мазками альвеолярных мешочков двух птиц в обеих группах, и из мазков 18 из 19 кур в группе В.
Пять ST-мутантов извлекали из альвеолярных мешочков трех кур в группе А и три ST-мутанта извлекали из альвеолярных мешочков одной птицы в группе В. Больше ST-мутантов извлекали в МВ, инокулированном мазками, взятыми из трахей, чем из мазков, взятых из альвеолярных мешочков. В целом были повторно выделены одиннадцать ST-мутантов из кур группы А и десять были повторно выделены из восемнадцати птиц в группе В.
Наиболее частым повторно выделяемым мутантом был мутант, несущий Tag 23. Этот мутант выделяли из трахей 17 птиц и из альвеолярных мешочков двух кур. Вторым наиболее часто выделяемым мутантом был 17-2, который выделяли из трахей 12 птиц (таблица 8). ST-мутанты 02-2, 03-1 10-1 и 15-1, которые использовали для инокуляции птиц в группе А, повторно выделяли из группы B, тогда как ST-мутанты 04-3, 23-1, 25-1 и 34-1, которые использовали для инокуляции птиц в группе В, повторно выделяли из группы А (таблица 8). ST-мутанты 04-1 и 33-1 не могли быть повторно выделены ни из одной группы (таблица 8). Ни в одном ST-мутанте не была детектирована потеря транспозона.
Пример 3: Инфективность и вирулентность отобранных ST-мутантов M. gallisepticum
Получение ST-мутантов
Два ST-мутанта, 04-1 и 33-1, которые не были детектированы после первоначального или подтверждающего экспериментов скрининга, и пять, которые детектировали нечасто (ST-мутанты 03, 18, 20, 22 и 26), культивировали при 37°С в МВ, дополненном гентамицином при 160 мкг/мл, до поздней логарифмической фазы. Ap3AS дикого типа культивировали при 37°С в МВ, который не содержал гентамицина. Концентрацию каждого штамма доводили до приблизительно 1×107 CCU/мл с использованием вышеописанного способа.
Анализ вирулентности и инфективности
План эксперимента
Восемь групп четырехнедельных кур SPF содержали раздельно (20 птиц на группу) в изоляторах из стекловолокна с положительным давлением (давлением выше атмосферного). Каждую из этих шести групп инокулировали отличающимся ST-мутантом посредством аэрозольного воздействия. Птиц отрицательного контроля подвергали воздействию МВ, а птиц положительного контроля подвергали воздействию Ap3AS дикого типа. Птиц эвтанизировали при 14 днях после инфицирования, и исследования post mortem проводили, как описано выше. Сыворотки и мазки собирали из каждой птицы, и детектирование на анти-микоплазма-антитело и выделение микоплазм проводили, как описано выше, за исключением того, что мазки брали из инфицированной Ap3AS группы, которые инокулировали на чашках МА и затем помещали в МВ без гентамицина. ДНК выделяли из каждой бульонной культуры, обнаруживающей изменение окраски, и использовали в качестве матрицы в ПЦР для амплификации района уникальной метки с использованием пары праймеров P2/P4 (таблица 2). ПЦР-продукты использовали в качестве зондов в дот-блот-гибридизациях для детектирования присутствия специфических меток, как ранее описано выше. ПЦР выполняли также с использованием праймера IGstmGenmeF3 и праймера, специфического для каждого ST-мутанта, в качестве дополнительного инструмента для идентификации ST-мутантов в инокулированных птицах (таблица 2). Верхние, средние и нижние срезы трахеи брали из каждой птицы, исследовали гистопатологически и измеряли толщину слизистой оболочки.
Детектирование повторно выделенных ST-мутантов
Для детектирования каждого ST-мутанта в бульонных культурах использовали дот-блот-гибридизацию и ПЦР-амплификацию. Дот-блот-гибридизация не могла быть использована для культур из группы положительного контроля, так как Ap3AS дикого типа не содержал сигнатурную метку. Дот-блот-гибридизация описана подробно выше. Вкратце, районы этой метки в ДНК, экстрагированной из культур, обнаруживающих изменение окраски, амплифицировали с использованием набора DIG-меченных праймеров. Олигонуклеотиды, соответствующие семи сигнатурным меткам, которые идентифицировали мутанты, используемые в эксперименте, наносили в виде пятен на нейлоновую мембрану и затем использовали в гибридизациях. Набор люминесцентного детектирования DIG (Roche) использовали для детектирования гибридизации согласно инструкциям изготовителя. ПЦР-праймеры, специфические для каждого ST-мутанта, были сконструированы с использованием данных секвенирования для каждого мутанта. Пару праймеров (STM13-KE-C'-1-Rev и STM13-KF-C') использовали для подтверждения повторного выделения штамма Ap3AS дикого типа, нацеленного на район, идентифицированный при секвенировании ST-мутантов 13-1, 13-2 и 13-3 (таблица 2).
ПЦР-реакции проводили с использованием 2 мкл экстрагированной ДНК в качестве матрицы в 20 мкл реакционной смеси, содержащей 2 мкл 10 × реакционного буфера, 1 мкМ каждого праймера, 200 мкМ каждого dNTP и 1,5 Е ДНК-полимеразы Taq (Promega). ПЦР инкубировали при 95°C в течение 2 минут, с последующими 35 циклами 94°C в течение 45 секунд, 52°C в течение 45 секунд и 72°С в течение 1 минуты, с конечным инкубированием при 72°C в течение 7 минут.
Гистологическое исследование
Приготовление трахеальных срезов
Пробы верхней, средней и нижней трахеи собирали и погружали в 10% нейтрально забуференный формалин (10% формалин, 4 г NaH2PO4 и 6,5 г Na2HPO4 на литр) и выдерживали в течение по меньшей мере 24 часов для фиксации. Затем ткани обрабатывали помещением в парафиновый воск с последующей заливкой в вакууме. Срезы с толщиной 2 мкм нарезали и собирали на стеклянных предметных стеклах. После удаления воска и регидратации, срезы окрашивали гематоксилином и эозином и исследовали под световым микроскопом в отношении повреждений и для измерения толщины слизистой оболочки.
Исследование трахеальных повреждений и измерение толщины слизистой оболочки
Гистологические повреждения в верхних, средних и нижних срезах трахеи оценивали в баллах на тяжесть по шкале 0-3 следующим образом:
0 = нет значимых изменений;
0,5 = очень малые агрегаты лимфоцитов (менее 2 очагов) или очень слабая, диффузная лимфоцитарная инфильтрация;
1 = малые агрегации лимфоцитов (более 2 очагов) или минорное утолщение слизистой оболочки, вызванное диффузной инфильтрацией лимфоцитов;
2 = умеренное утолщение слизистой оболочки вследствие инфильтрации гетерофилов и лимфоцитов, а также отек, сопровождающийся дегенерацией эпителия с экссудацией или без экссудации просветов;
3 = существенное утолщение, вызванное инфильтрацией гетерофилов и лимфоцитов, и отек с плоскоклеточной метаплазией или эпителиальной дегенерацией и экссудацией просветов.
Толщину слизистой оболочки трахеи каждой птицы определяли измерением толщины в 6 точках на каждом срезе из верхней, средней и нижней трахеи из каждой птицы. Затем рассчитывали среднюю толщину для каждого из трех районов.
Статистические анализы
Медианные балльные оценки гистологических повреждений трахей для каждой экспериментальной группы сравнивали с использованием U-критериев Манна-Уитни (Minitab version 14.2 для Windows). T-критерий Стьюдента и однофакторный дисперсионный анализ (ANOVA) использовали для сравнения средней толщины слизистых оболочек трахеи. Вероятность (P-величина) ≤0,05 считалась значимой.
Электрофорез в ДСН-ПААГ
Клетки в пробе 1 мл культуры средней логарифмической фазы каждого из ST-мутантов, а также Ap3AS и ts-11-штаммов собирали центрифугированием при 16000 g в течение 5 минут, затем ресуспендировали в 1 × ДСН-ПААГ лизисном буфере и инкубировали при 100°C в течение 5 минут перед быстрым охлаждением на льду. Все белки клеток разделяли в 12,5% полиакриламидном геле вместе со стандартами молекулярных масс (Marker 12™ Wide Range Protein Standard, Novex) и затем окрашивали Кумасси бриллиантовым синим.
Результаты
Вирулентность и инфективность ST-мутантов
Клинические признаки и исследование post mortem
Повреждения альвеолярных мешочков не наблюдали в птицах, подвергнутых воздействию аэрозолей ST-мутантов 04-1 (группе 2), 33-1 (группе 3) или 22-1 (группе 8), или в птицах отрицательного контроля (группе 1). Слабые повреждения (балльная оценка 0,25) наблюдали в одной птице, инокулированной ST-мутантом 03-1 (группа 4). Из 20 птиц, инфицированных ST-мутантом 26-1 (группа 5), четыре имели слабые повреждения (0,50-1,00), в то время как повреждения наблюдали только в брюшных альвеолярных мешочках четырех птиц, подвергнутых действию ST-мутанта 18-1 (группа 6). Шесть из 20 птиц имели повреждения от слабых до тяжелых (0,50-2,50) в группе, инфицированной ST-мутантом 20-1 (группа 7), в то время как повреждения от слабых до тяжелых (0,50-3,00) наблюдали в 11 из 18 птиц с вирулентным штаммом Ap3AS (группа 9). Эти результаты суммированы в таблице 9 и показаны графически на фиг.6.
RSA-результаты, балльные оценки повреждений альвеолярных мешочков и степень повторного выделения M. gallisepticum из птиц в исследовании вирулентности и инфективности
* Результаты показаны в виде отношения количество положительных проб/собранное количество.
^ Результаты показаны в виде отношения количество проб с изменением окраски/собранное количество. Сигнатурные метки детектировали только в виде меток, несомых ST-мутантами, которые инфицировали птиц в группах 2-8.
Реакции в виде антител и повторное выделение ST-мутантов
В сыворотке любых экспериментальных птиц не детектировали анти-микоплазма-антител в момент инфицирования при помощи RSA-теста. Спустя две недели после инфицирования, реакции в виде антител не детектировали ни в одной из птиц в группах 2, 3, 5 и 8, тогда как реакция была детектируемой только в одной птице в группе 4. В противоположность этому, сильные RSA-реакции против M. gallisepticum были детектируемыми во всех птицах в группах 6 и 7. Птицы отрицательного контроля (группа 1) не обнаруживали реактивности против Mg в RSA-тесте, тогда как все птицы положительного контроля (группа 9) имели сильные RSA-реакции против Mg (таблица 9, фиг.6). M. gallisepticum не вылелялись на чашках МА, инокулированных мазками альвеолярных мешочков или трахеи любой птицы из группы 2 (инфицированной ST-мутантом 04-1) или группы 3 (инфицированной ST-мутантом 33-1) (таблица 9). M. gallisepticum не выделялись на чашках МА, инокулированных мазками альвеолярных мешочков любых птиц в группах 4 (инфицированной ST-мутантом 03-1) или 8 (инфицированной ST-мутантом 22-1), но они выделялись из трахеи двух птиц в группе 4 и одной птицы в группе 8. В группе 5 (инфицированной ST-мутантом 26-1), M. gallisepticum выделялись из альвеолярных мешочков одной птицы и трахей 6 птиц. M. gallisepticum выделялись также из трахей 17 из 20 птиц в группе 6 (инфицированной ST-мутантом 18-1) и всех птиц в группе 7 (инфицированных ST-мутантом 20-1), а также из мазков альвеолярных мешочков 5 из 20 птиц в группе 6 и 7 из 20 птиц в группе 7. В группе положительного контроля (группа 9), M. gallisepticum выделялись из альвеолярных мешочков 9 из 18 птиц и трахей 17 из 18 птиц. M. gallisepticum более часто выделялись из мазков, инкубированных в МВ, чем на чашках МА. M. gallisepticum не выделялся в МВ, инокулированном мазками альвеолярных мешочков или трахей любой птицы, подвергнутой воздействию ST-мутанта 04-1 (группа 2) (таблица 9). В других, инфицированных мутантом группах, Mg извлекали из МВ, инокулированного мазками альвеолярных мешочков одной птицы в группе 5 (ST-мутант 26-1), 7 птиц в группе 6 (ST-мутант 18-1) и 8 птиц в группе 7 (ST-мутант 20-1). MG извлекали из трахей птиц в 6 из групп, подвергнутых действию ST-мутантов, причем количество инфицированных птиц находилось в диапазоне от 2 в группе 3 (ST-мутант 33-1) до 20 в группе 7 (инфицированной ST-мутантом 20-1). Идентичность выделенных ST-мутантов M. gallisepticum подтверждали с использованием уникальных сигнатурных меток, которые они несли (таблица 9).
Трахейные повреждения и величины толщины слизистой оболочки
Медианные балльные оценки трахеальных повреждений показаны в таблице 10. Балльные оценки птиц во всех инфицированных мутантами группах, значимо отличались от балльных оценок птиц в группе положительного контроля (группе 9) (P<0,0001). Не было значимого различия между группой отрицательного контроля (группой 1) и группами 5 (инфицированной ST-мутантом 26-1) или 7 (инфицированной ST-мутантом 20-1). Более низкие балльные оценки трахеальных повреждений птиц, инфицированных ST-мутантом 18-1 (группа 6), не отличались от балльных оценок птиц в группах 2, 3, 4, 5 и 7, но значимо отличались от балльных оценок птиц в группе 8 (инфицированной ST-мутантом 22-1) и в группе отрицательного контроля (группе 1), тогда как только балльные оценки повреждений в средней трахее птиц в группе положительного контроля отличались от балльных оценок птиц в любой из других групп. Средняя толщина трахеальной слизистой оболочки показана в таблице 10. Эта средняя толщина слизистой оболочки в средней трахее не различалась значимо между любыми из инфицированных мутантами групп (группы 2-8). Однако, все они были значимо более высокими, чем средние величины толщины группы отрицательного контроля (группы 1), за исключением этих величин в группах 2 и 7, и они были значимо меньшими, чем эти величины группы положительного контроля (группы 9). Не было значимого различия между отрицательными контролями и группой 7 (инфицированной ST-мутантом 20-1) в толщине слизистой оболочки верхних и средних трахей, но было различие в нижней трахее (P=0,004).
В целом, эти положительные контроли имели значимо более тяжелые трахеальные повреждения, как оценено гистологической балльной оценкой повреждений или толщины слизистой оболочки, чем любая из групп, инфицированных ST-мутантами. Группа 4 (инфицированная ST-мутантом 03-1) была наиболее тяжело поражена среди групп, инфицированных ST-мутантами.
Балльные оценки трахеальных повреждений и величин толщины слизистой оболочки в птицах в исследовании вирулентности и инфективности
Пример 4: Оценивание защиты, обеспечиваемой вакцинацией выбранным ST-мутантом M. gallisepticum
Приготовление ST-мутантной вакцины
ST-мутант 26-1 выращивали в МВ, дополненном 160 мкг гентамицина/мл, при 37°С. Способ, используемый для определения концентрации организмов в культуре, был описан выше. Концентрацию ST-мутанта 26-1 корректировали до приблизительно 1×107 CCU/капля (22,5 мкл) для вакцинации, и затем каждую птицу вакцинировали внутриглазно каплей культуры.
План эксперимента
В целом, восемьдесят 5-недельных кур SPF случайным образом распределяли на 4 группы и содержали в изоляторах из стеклопластика при положительном давлении. ST-мутант 26-1 вводили с использованием глазных капель в день 0. Ap3AS дикого типа вводили посредством аэрозоля для заражения птиц спустя 2 недели после иммунизации. Птицы в группе 1 служили в качестве отрицательных контролей. Этих птиц вакцинировали МВ посредством глазных капель в день 0 и заражали МВ посредством аэрозоля в день 14. Птицы в группе 2 служили в качестве положительных контролей. Их вакцинировали МВ с использованием глазных капель в день 0 и инфицировали Ap3AS дикого типа посредством аэрозоля в день 14. Птицы в группе 3 служили в качестве контролей вакцинации. Этим птицам вводили ST-мутант 26-1 с использованием глазных капель в день 0. Затем МВ вводили посредством аэрозоля спустя 14 дней после вакцинации. Птицы в группе 4 служили в качестве вакцинированной и заражаемой группы. Кур иммунизировали ST-мутантом 26-1 с использованием глазных капель в день 0 и заражали Ap3AS дикого типа посредством аэрозоля в день 14.
Сбор проб из экспериментальных кур
Пробы крови собирали из всех кур перед вакцинацией, заражением и эвтаназией. Затем сыворотки собирали и тестировали при помощи RSA для определения концентраций уровней анти-микоплазма-антител. Массу тела каждой птицы также измеряли перед иммунизацией, при заражении и при post mortem. В день 28 (спустя 14 дней после заражения), кур эвтанизировали и подвергали исследованию post mortem, и брали пробы для культивирования M. gallisepticum. Шесть альвеолярных мешочков визуально оценивали на повреждения и давали балльную оценку от 0 до 3, как описано выше. Брали мазки из альвеолярных мешочков и трахей птиц и высевали штриховой разводкой на МА без добавленного гентамицина и затем переносили в МВ, не содержащий гентамицина, для выделения M. gallisepticum. МА-планшеты инкубировали при 37°С в течение 10 дней, и МВ-культуры инкубировали при 37°С, пока не наблюдали изменение окраски. Организмы из бульонов, обнаруживающих изменение окраски, осаждали центрифугированием и подвергали ПЦР-амплификации для идентификации ST-мутанта 26-1 и Ap3AS дикого типа (выше). Пробы верхнего, среднего и нижнего районов трахеи каждой птицы собирали и фиксировали в 10% нейтрально забуференном формалине в течение по меньшей мере 24 часов. После обработки и окрашивания срезы исследовали при помощи световой микроскопии. Повреждения оценивали в отношении тяжести по шкале 0-3, как описано выше. Толщину слизистой оболочки измеряли для каждого трахеального района в 6 разных точках и затем рассчитывали среднюю толщину для каждого из этих трех районов.
Повторное выделение M. gallisepticum
Пару ПЦР-праймеров, которая была специфической в отношении ST-мутанта 26-1 (IGstmGenmeF3 и STM26-Rev), и набор праймеров P2/P4, которые амплифицировали район уникальной метки, использовали для повторного выделения ST-мутанта 26-1. Пару праймеров STM26-генома и STM26-Rev связывали на каждой стороне сайта инсерции транспозона в ST-мутанте 26-1 и использовали для подтверждения повторного выделения Ap3AS дикого типа. Эта ПЦР могла легко дифференцировать ST-мутант 26-1 от исходного штамма Ap3AS (таблица 2). ПЦР проводили с 2 мкл экстрагированной ДНК в качестве матрицы в 20 мкл реакционной смеси, содержащей 2 мкл 10 × реакционного буфера, 1 мкМ каждого праймера, 200 мкМ каждого dNTP и 1,5 Е ДНК-полимеразы Taq (Promega). ПЦР инкубировали при при 95°C в течение 2 минут с последующими 35 циклами 94°C в течение 45 секунд, 52°C в течение 45 секунд и 72°C в течение 1 минуты с конечным инкубированием при 72°С в течение 7 минут. Полученные продукты разделяли в 2% агарозном геле вместе с маркерами молекулярной массы ДНК.
Статистические анализы
Средние процентные увеличения массы тела для каждой группы анализировали на различия с использованием однофакторного дисперсионного анализа ANOVA и t-критерия Стьюдента (Minitabs v14.2 для Windows). Медианные балльные оценки трахеальных повреждений сравнивали с использованием критериев U Манна-Уитни. Средние величины толщины слизистой оболочки в верхней, средней и нижней трахее сравнивали с использованием t-критерия Стьюдента и однофакторного дисперсионного анализа ANOVA. Величину Р≤0,05 считали значимой.
Результаты
Макроскопические повреждения
Макроскопические повреждения альвеолярных мешочков наблюдали только в птицах в группе положительного контроля (группе 2). В этой группе, 5 птиц имели повреждения альвеолярных мешочков, с общими балльными оценками повреждений альвеолярных мешочков в диапазоне от 0,5 до 2,5. Эти результаты суммированы в таблице 10 и показаны графически на фиг.7.
Результаты RSA-теста, балльные оценки повреждений альвеолярных мешочков и повторное выделение M. gallisepticum всех групп в исследовании иммунопротекции
# Полные данные RSA и оценки повреждений альвеолярных мешочков показаны в Приложении G.
^ Количество проб, обнаруживающих колонии M. gallisepticum, растущих на МА-чашке/общее количество собранных проб.
& ПЦР проводили для определения присутствия ST-мутанта 26-1 и штамма Ap3AS.
Серологическое тестирование и повторное выделение M. gallisepticum
Сыворотки, собранные у птиц в момент вакцинации и заражения, не содержали детектируемого анти-микоплазма-антитела. Ни одна курица в группе отрицательного контроля (группа 1) не имела никакого детектируемого сывороточного антитела против M. gallisepticum в момент post mortem. В отличие от этого, большая доля птиц в других 3 группах имела детектируемые антитела против M. gallisepticum при post mortem. Каждая птица в группе положительного контроля (группе 2) производила реакцию RSA антитела против M. gallisepticum от умеренной до сильной. Анти-микоплазма-антитела детектировали в 16 из 20 проб сыворотки из группы 3, с RSA-оценками в диапазоне от низких до умеренных. Умеренные титры анти-микоплазма-антител детектировали во всех птицах в группе 4 (вакцинированной ST-мутантом 26-1 и инфицированной штаммом дикого типа). Эти результаты суммированы в таблице 10. M. gallisepticum выделялись более часто из трахеальных мазков, чем из мазков альвеолярных мешочков, в группах 2, 3 и 4, и не выделялись из альвеолярных мешочков или трахей ни одной из птиц группы 1 (отрицательные контроли). Mg-колонии выращивали из мазков альвеолярных мешочков в одной птице как в группе 2 (только зараженные), так и группе 3 (только вакцинированные), в то время как повторное выделение достигалось из трахеи 19 и 2 птиц, соответственно. В группе 4, которой вводили ST-мутант 26-1 в качестве вакцины и затем инфицировали штаммом Ap3AS, Mg выделяли на МА-чашках из альвеолярных мешочков одной птицы и из трахей 7 птиц (таблица 10). Как и в предыдущих экспериментах, скорости повторного выделения были более высокими в МВ, чем на МА-чашках. ПЦР-амплификация подтверждала, что только микоплазмы, повторно выделенные из групп 2 и 3, были штаммом, который вводили этой группе. После ПЦР-амплификации с использованием штамм-специфической пары праймеров только микоплазмы, повторно выделенные из группы 4, были Ap3AS дикого типа. Эти результаты показаны в таблице 10.
Приросты массы кур
Не было значимого различия в приросте массы между любыми из этих групп спустя 14 дней после вакцинации. При неделе 4 (14 дней после заражения) птицы положительного контроля (группы 2) имели более низкие средние приросты массы, чем птицы в группах 1 (негативные контроли) или 3 (вакцинированные контроли) (P=0,006 и 0,027, соответственно), но не в сравнении с птицами в группе 4 (вакцинированными и зараженными) (P=0,332) (таблица 11). В периоде между заражением и post mortem (день 14 - день 28) средний прирост массы кур в группе 2 (только зараженных) был значимо более низким, чем прирост массы птиц в каждой из других групп. Средний прирост массы птиц в группе 3 (вакцинированных, незараженных) был не большим, чем прирост массы отрицательных контролей, но был значимо большим, чем прирост массы птиц в группе 4 (вакцинированных и зараженных) (P=0,016). Однако, средний прирост массы птиц в группе 4 не отличался значимо от прироста массы птиц в группе 1 (P=0,580). Эти результаты показаны в таблице 11.
Трахеальные повреждения и величины толщины слизистой оболочки
Медианные балльные оценки трахеальных повреждений показаны в таблице 12. Эти повреждения были тяжелыми в птицах в группе 2 (только зараженной). Медианные балльные оценки трахеальных повреждений в группе 2 (положительных контролях) значимо отличались от балльных оценок в других трех группах (P<0,0001). Оценки повреждений птиц в других группах не отличались значимо друг от друга. Средние величины толщины слизистой оболочки верхней, средней и нижней трахеи птиц в группе 2 (только зараженной) были значимо более высокими, чем оценки птиц в других трех группах (P<0,0001), но не было значимого различия между оценками этих птиц в этих других трех группах (таблица 12). Однако, в средней и нижней трахеях средняя толщина слизистой оболочки была меньшей в птицах в группе 4, чем в птицах в группе 1 (P=0,028) или 3 (P=0,036).
Прирост массы кур во всех экспериментальных группах в исследовании иммунопротекции
Балльные оценки трахеальных повреждений и величины толщины слизистой оболочки птиц в исследовании иммунопротекции
Пример 5: Нокаут-мутанты генов олигопептидных транспортеров dppD и oppD в птичьем патогенном штамме Escherichia coli E956 (APEC E956).
Исследования безопасности in vivo показывали статистически значимую аттенуацию oppD-нокаута как в виде балльных оценок повреждений, так и в степенях повторного выделения из птиц в сравнении с dppD-нокаутом или исходным штаммом E956. Исследования эффективности in vivo показывали статистически значимую защиту вакцинированных oppD-нокаутом птиц после заражения APEC E956.
Примеры 1 и 2 иллюстрируют количество аттенуирующих ген мутаций, включающих гены dppD и oppD, которые гомологичны другим бактериальным генам, участвующим в транспорте олигопептидов в клетку.
Поскольку все бактериальные виды имеют почти идентичные транспортные системы, могли бы быть развиты новые вакцины против других птичьих патогенов, а также патогенов других видов животных посредством продуцирования dppD-нокаутов.
Этот пример иллюстрирует нокауты генов dppD и oppD в птичьем патогенном штамме Escherichia coli E956 (APEC E956). Каждый мутант испытывали на безопасность и эффективность с использованием системы заражения.
Методология
Штаммы, среды и плазмиды E. coli
Птичий патогенный штамм Escherichia coli E956 (APEC E956) первоначально был выделен из однодневного цыпленка в бройлерном хозяйстве, и было обнаружено, что он является чувствительным к антибиотикам ампициллину, сульфафуразолу, триметоприму, хлорамфениколу, тетрациклину и канамицину. Организм размножали в бульоне Луриа-Бертани (LB) или на LB-агаре при 30°С или 37°С в течение ночи с подходящим отбором с использованием антибиотиков (ампициллина, 50 мкг/мл; канамицина, 100 мкг/мл), если не указано другое. Для стандартного клонирования и получения плазмид использовали штамм E. coli DH5α.
Использовали красную систему рекомбинации для усиления гомологичной рекомбинации между нокаут-конструкциями и геном-мишенью нокаута с использованием чувствительной к температуре плазмиды pKD46, несущей ген устойчивости к ампициллину, три гена "Красной системы" γ, λ, и exo, кодирующих Gam, Bet и Exo, соответственно. Нокаут-конструкции собирали при помощи ПЦР и лигировали в вектор pGEM-T (Promega). Плазмиду, содержащую каждую конструкцию, использовали в качестве матрицы в ПЦР для получения линейной ДНК для трансформации.
Получение нокаут-конструкций генов
Ген канамицина амплифицировали из транспозона TnphoA с использованием олигонуклеотидных праймеров TXkanFor и TYkanRev (таблица 13). Полученный ПЦР-продукт 1,064 т.п.н. очищали с использованием набора для очистки UltraClean PCR (MoBio, California USA) в соответствии с инструкциями изготовителя и лигировали с вектором pGEM-T (Promega) в соответствии с инструкциями изготовителя. Смесь лигирования использовали для трансформации E. coli DH5α электропорацией при помощи генератора импульсов Gene Pulser (BioRad) с установками 2500 В, 200 Ω, 250 мкФ, и рекомбинанты отбирали на LB-агаре, содержащем 25 мкг/мл канамицина. Трансформанты выращивали в течение ночи в LB-бульоне, содержащем 25 мкг/мл канамицина, и плазмиду экстрагировали с использованием набора (Promega) в соответствии с инструкциями изготовителя. Клонирование гена устойчивости к канамицину подтверждали анализом с использованием рестрикционных эндонуклеаз. Линейную ДНК использовали для трансформации: для dppD-нокаутов ДНК получали при помощи ПЦР с использованием пары праймеров WE и WF, каждый из которых содержит 5'-районы 40 п.н., комплементарные районам 4384-4423 п.н. и 4829-4868 п.н., соответственно, в гене dppD GenBank Accession L08399. Эти олигонуклеотидные праймеры WE и WF содержали 3'-районы 20 п.н., комплементарные гену канамицина, и при использовании в ПЦР-амплифицировали ген канамицина с 40 п.н.-концами, комплементарными ДНК dppD.
Олигонуклеотиды, используемые в этом исследовании
Получение нокаутов генов
Штаммы APEC E956 трансформировали pKD46 электропорацией с использованием генератора импульсов Bio-Rad Gene Pulser (Bio-Rad), установленного при 2500 В, 200 Ω, 250 мкФ. Рекомбинанты отбирали на агаре Луриа-Бертани, содержащем 100 мкг/мл ампициллина, с последующим инкубированием при 30°С в течение ночи. Авторы изобретения получали нокауты генов посредством гомологичной рекомбинации при помощи известных способов. Отдельную свежую колонию APEC E956/pKD46 помещали в 5 мл LB, содержащего 100 мкг/мл ампициллина, и инкубировали при встряхивании (200 об/мин) при 30°С в течение 16 часов. На следующий день гототвили разведение 1:50 ночной культуры в объеме 20 мл LB, содержащего 100 мкг/мл ампициллина, и культуры встряхивали (200 об/мин) при 30°С в конической колбе на 125 мл. При концентрации 107 клеток/мл добавляли L-арабинозу (Sigma) до конечной концентрации 1 мМ. Культуры индуцировали приблизительно 1 час при 30°С, и затем клетки подвергали тепловому шоку в течение 15 минут при 42°C и немедленно переносили на баню со смесью воды со льдом на 10 минут, затем собирали центрифугированием при 10000 × g в течение 10 минут при 4°С. Клетки ресуспендировали в 1 мл охлажденного на льду 1 мМ MOPS, содержащего 20% глицерин, переносили в центрифужную пробирку на 1,5 мл и центрифугировали при 16000 × g в течение 30 секунд при 4°С в настольной микрофуге. Супернатант удаляли и клетки ресуспендировали в 1 мл 1 мМ MOPS, содержащего 20% глицерин, и центрифугировали, как описано выше. Эту стадию повторяли еще один раз, и клетки, наконец, ресуспендировали в 100 мкл 1 мМ, содержащего 20% глицерин. Затем клетки добавляли в предварительно охлажденные кюветы для электропорации (щель 2 мм Bio-Rad), и 50 мкл клеток с 300 нг DpnI-расщепленной ДНК трансформировали с использованием вышеописанных установок. Сразу после электропорации клетки извлекали суспендированием в 0,3 мл LB, разводили в 2,7 мл LB и инкубировали при 37°C при встряхивании в течение 1,5 часов. Культуру 5× концентрировали в LB и 0,2 мл инокулировали на LB-чашки, содержащие 20 мкг/мл канамицина, и инкубировали в течение ночи при 37°С. Трансформанты отбирали и выращивали в LB, содержащем 40 мкг/мл канамицина, и нокауты генов подтверждали при помощи ПЦР и блоттинга по Саузерну.
Подтверждение штаммов с нокаутами генов dppD и oppD
Для подтверждения инсертирования гена устойчивости к канамицину в dppD или oppD, ПЦР проводили с использованием пары праймеров для dppD (TZIUA) и oppD (WSIWT) для амплификации соответствующих районов генов. Предсказанный размер ПЦР-продукта для исходного штамма APEC E956 для dppD и oppD был 0,89 т.п.н. и 0,951 т.п.н., соответственно. Предсказанным увеличением размера с инсерцией гена канамицина (1,064 т.п.н.) был размер 1,774 т.п.н. и 1,924 т.п.н. для dppD и oppD, соответственно.
Экстракция и блоттинг по Саузерну геномной ДНК
Геномную ДНК APEC E956, штаммы ΔdppD и ΔoppD получали с использованием фенола/хлороформа, как описано ранее (Sambrook et al, 2001). Геномную ДНК из штаммов E. coli расщепляли с использованием PstI, и фрагменты вместе с маркерами молекулярных масс разделяли электрофорезом в агарозном геле. После электрофореза в агарозном геле фрагменты геномной ДНК переносили на нейлоновую мембрану (Hybond-N+, GE Healthcare) капиллярным переносом. ДНК-зонды метили [γ32P]dATP с использованием набора для случайно праймированного мечения ДНК (Roche). Предварительную гибридизацию и гибридизацию проводили в буфере Черча (0,5 M Na2HPO4 [pH 7,4], 7% додецилсульфат натрия, 1 мМ ЭДТА, 1% бычий сывороточный альбумин БСА) (Church & Gilbert, 1984) в течение ночи при 54°C. Мембраны промывали в 2 × SSC (1 × SSC представляет собой 0,15 M NaCl плюс 0,015 М цитрат натрия)-0,1% додецилсульфате натрия дважды при 54°C в течение 5 минут в каждом случае и затем один раз в 0,5 × SSC, 0,1% SDS в течение 15 минут при 65°C, и авторадиографировали с использованием пленки Kodak BioMax MS при -70°C.
Безопасность нокаутов dppD и oppD APEC E956
Однодневных цыплят инфицировали штаммами APEC E956, ΔdppD и ΔoppD для оценивания патогенности каждого мутанта в сравнении с неинфицированными и APEC E956-инфицированными. В целом, покупали 95 однодневных цыплят (SPAFAS, Woodend Vic) и распределяли в три группы по 20 цыплят в каждой и одну группу с 35 цыплятами. Птиц содержали в изоляторах положительного давления и кормили ad libitum облученным коммерческим стартером (начальным рационом). Мазки клоак брали у двух цыплят из каждой группы и высевали штрихом на агар MacConkey для оценивания симбионтной E. coli. Готовили ночные культуры E956 ΔdppD, ΔoppD в питательном бульоне с 40 мкг/мл канамицина вместе с E956 без канамицина для получения концентрации 1E+10 колониеобразующих единиц/мл (кое/мл). Все группы инокулировали посредством глазных капель с вводимой 10 раз нормальной иммунизирующей дозой коммерческой вирусной вакцины Vic S вируса инфекционного бронхита (Websters, Australia). Каждая группа 2 и 3 из 20 цыплят получала 20 мл аэрозоля, содержащего 1E+10 кое/мл ΔdppD и ΔoppD), соответственно, тогда как группа 4 из 35 птиц получала 20 мл 1E+10 кое/мл E956. Цыплята в группе 1 оставались необработанными и служили в качестве отрицательного контроля. Цыплят из групп 2, 3 и 4 подвергали еще два раза действию того же самого штамма APEC в возрасте 3 и 5 дней. Все птицы были подвергнуты исследованию post mortem в возрасте 10 дней. Заболевание оценивали по макроскопической патологии, повреждения альвеолярных мешочков оценивали в соответствии с предыдущими критериями. Мазки брали из левой и правой задней и передней части альвеолярных мешочков, трахеи и асептически из печени. Мазки инокулировали на агар MacConkey с канамицином и без канамицина (40 мкг/мл) и инкубировали в течение при 37°С. На следующий день чашки обследовали на присутствие "кирпично-красных" колоний (типичный фенотип E. coli), их количества подсчитывали и регистрировали. Если количество колоний было меньше 30, то проводили ПЦР для идентификации штамма E. coli. Штаммы APEC идентифицировали с использованием пары праймеров: для dppD WA/UA, для oppD WA/WT, для E956 Sidfwdseq/bwd siderophore (таблица 13), который амплифицирует ген iucA, присутствующий на pVM01, и для симбионтной E. coli рДНК 16s амплифицировали с 16Sfwd/16Srev.
Эффективность нокаутов dppD и oppD APEC
Для оценивания эффективности вакцинного кандидата E. coli шестьдесят однодневных птиц делили на три группы из 20 птиц. Группы 2 и 3 вакцинировали при помощи аэрозоля E956 ΔdppD, ΔoppD в день один, как описано выше, тогда как группа 1 оставалась невакцинированной в качестве контроля и содержалась отдельно в изоляторах, как описано выше. В день 12 после вакцинации все группы по отдельности заражали 20 мл аэрозоля, содержащего 1E+10 кое/мл APEC E956, как описано выше, и повторно помещали в их изоляторы. После 4 дней всех птиц эвтанизировали и подвергали post mortem. Повреждения альвеолярных мешочков оценивали, как описано выше, и мазки, взятые для повторного выделения инфицирующих организмов, культивировали на агаре MacConkey с канамицином и без канамицина (40 мкг/мл).
Анализ результатов
Статистический анализ выполняли на балльных оценках скорости и степени повреждений альвеолярных мешочков, скорости выделения организмов и прироста массы птиц. Используемыми критериями были Точный критерий Фишера для скоростей повторного выделения и критерий U Манна-Уитни для оценок повреждений и изменения массы.
Результаты
Развитие и подтверждение ΔdppD и ΔoppD APEC E956
Для трансформации APEC E956/pKD46 использовали линейные ДНК-конструкции для нокаутов специфических генов, продуцируемые ПЦР. Трансформанты отбирали на LB-агаре, содержащем 40 мкг/мл канамицина, и затем проводили ПЦР для подтверждения клонов, несущих ген канамицина. Было предсказано, что размер ампликона из E956 ΔdppD, использующего пару олигонуклеотидных праймеров TZ/UA, равен 1,774 т.п.н. вследствие инсерции ДНК канамицина (1,064 т.п.н.) в сравнении с исходным штаммом E956 (0,89 т.п.н.), Фиг.8А и фиг.9, Панель А, дорожки 1 и 2, соответственно. Сходным образом, размер ампликона для E956 ΔoppD, использующего пару олигонуклеотидых праймеров WS/VT, был близким к предсказанному размеру 1,924 т.п.н. с исходным штаммом E956, продуцирующим ампликон 0,951 т.п.н., Фиг.8B и Фиг.9, Панель B, дорожки 1 и 2, соответственно.
Геномную ДНК штаммов APEC E956, ΔdppD и ΔoppD, расщепляли рестриктазой PstI, этот фермент был выбран, так как ген устойчивости к канамицину имеет единственный сайт PstI в середине гена. Радиоактивно меченные зонды dppD и oppD детектировали полосы в штаммах APEC E956, ΔdppD и ΔoppD (фиг.10). Зонд dppD связывался с фрагментом сходного размера в штаммах E956 и ΔoppD (фиг.10, дорожки 1 и 3), но с полосой слегка меньшей молекулярной массы и другой полосой 2 т.п.н. Зонд oppD связывался с фрагментом сходного размера в штаммах E956 и ΔdppD (фиг.10, дорожки 1 и 2) и с полосами 9,8 и 7 т.п.н. ΔdppD (фиг.10, дорожка 3). Зонд гена канамицина не связывался с E956, но одни и те же полосы связывались зондами dppD и oppD в штаммах ΔdppD и ΔoppD (фиг.10, дорожки 1 и 2).
Исследования вирулентности с использованием штаммов E956 ΔdppD и ΔoppD
Кур исследовали при post mortem в отношении признаков колибациллеза, исследовали шесть альвеолярных мешочков в отношении повреждений и брали мазок пробы из левой и правой передней части альвеолярного мешочка, трахеи и печени каждой птицы для повторного выделения вакцины, исходных или симбионтных организмов. Эти мазки высевали на агар MacConkey с добавленным канамицином или без добавленного канамицина.
Результаты из этих исследований обсуждаются ниже и суммированы в таблицах 14, 16 и 19 со статистическими сравнениями, выполненными между каждой из этих групп, суммированными в таблицах 15, 17, 18, 20 и 21.
Прирост массы кур в экспериментах по патогенности и эффективности
Величины вероятностей (Р) для статистического анализа приростов массы из таблицы 14
Безопасность нокаутов oppD и dppD в сравнении со штаммом E.coli E956
без антибиотиков
без антибиотиков (диапазон)
Величины вероятности (P) для статистического анализа степеней повторного выделения альвеолярных мешочков и трахей из таблицы 16
Величины вероятности (P) для статистического анализа оценок и степеней повреждений альвеолярных мешочков из таблицы 16
Исследования эффективности с использованием штамма E. coli E956, нокаутов oppD и dppD
без антибиотиков
без антибиотиков (диапазон)
Величины вероятности (P) для статистического анализа степеней повторного выделения альвеолярных мешочков и трахей из таблицы 19
Величины вероятности (P) для статистического анализа оценок и степеней повреждений альвеолярных мешочков из таблицы 19
Прирост массы птиц
Различие между процентным приростом массы в эксперименте по патогенности между E956 (143%), ΔdppD (143%) и невакцинированными курами (174%) было несущественно значимым, в то время как значимого различия не наблюдали с ΔoppD (155%) относительно невакцинированных птиц (таблица 14 и 15). В эксперименте по эффективности не было статистически значимого различия в процентном приросте массы любой группы птиц перед заражением или при post mortem (таблица 14 и 15).
Степени повторного выделения организмов на агаре MacConkey
Не было значимого различия между медианными количествами организмов, повторно выделенных из альвеолярного мешочка или трахеи контрольных и вакцинированных ΔoppD птиц в эксперименте по патогенности (таблица 16). Наблюдали более высокие количества организмов, выделенных из птиц, вакцинированных E956 или ΔdppD, чем из невакцинированных птиц (таблица 16). Результаты для эксперимента по эффективности (таблица 19) не обнаруживают различия между E956-зараженными и ΔoppD-вакцинированными птицами. Не было значимого различия в степени выделений, выполненных из трахеи, в сравнении с альвеолярными мешочками в птицах, вакцинированных либо ΔdppD, либо ΔoppD, хотя значимое увеличение в выделении организмов из трахеи в сравнении с альвеолярными мешочками наблюдали с птицами, вакцинированными E956 (P<0,05).
Степени повреждений альвеолярных мешочков
Исследование альвеолярных мешочков и органов при post mortem обнаруживало признаки колибациллеза e некоторых птиц с перигепатитом и перикардитом. В эксперименте по патогенности не было статистически значимого различия между птицами, вакцинированными либо ΔdppD, либо ΔoppD, в степени и балльной оценке повреждения альвеолярных мешочков, было меньше повреждений, наблюдаемых между ΔoppD и E956, но не было статистически значимого различия между вакцинированными ΔdppD и вакцинированными E956 птицами (таблица 16 и 18). В эксперименте по защите, сравнение степени и оценок повреждения вакцинированных ΔoppD и зараженных E956 птиц обнаруживало статистически значимое протективное действие от предыдущей вакцинации ΔoppD с медианной степенью повреждения 0,0 для вакцинатов и 1,25 для невакцинированной и зараженной группы (таблицы 19 и 21).



















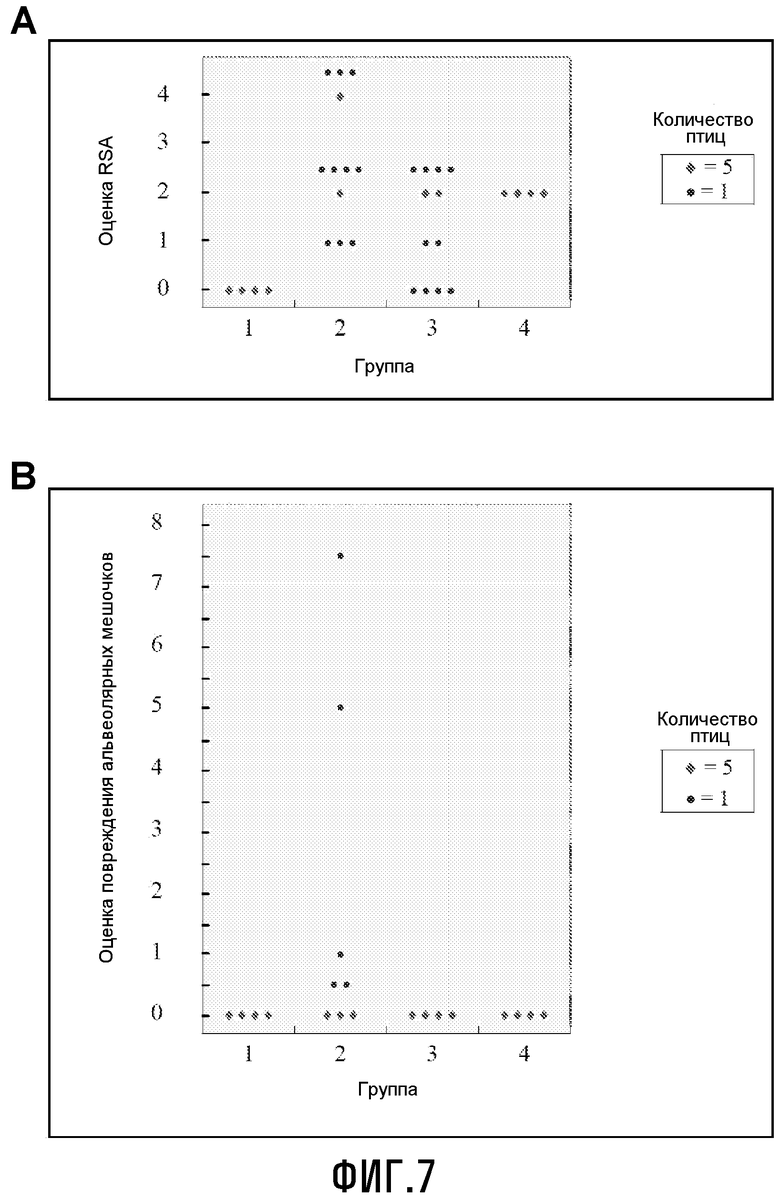
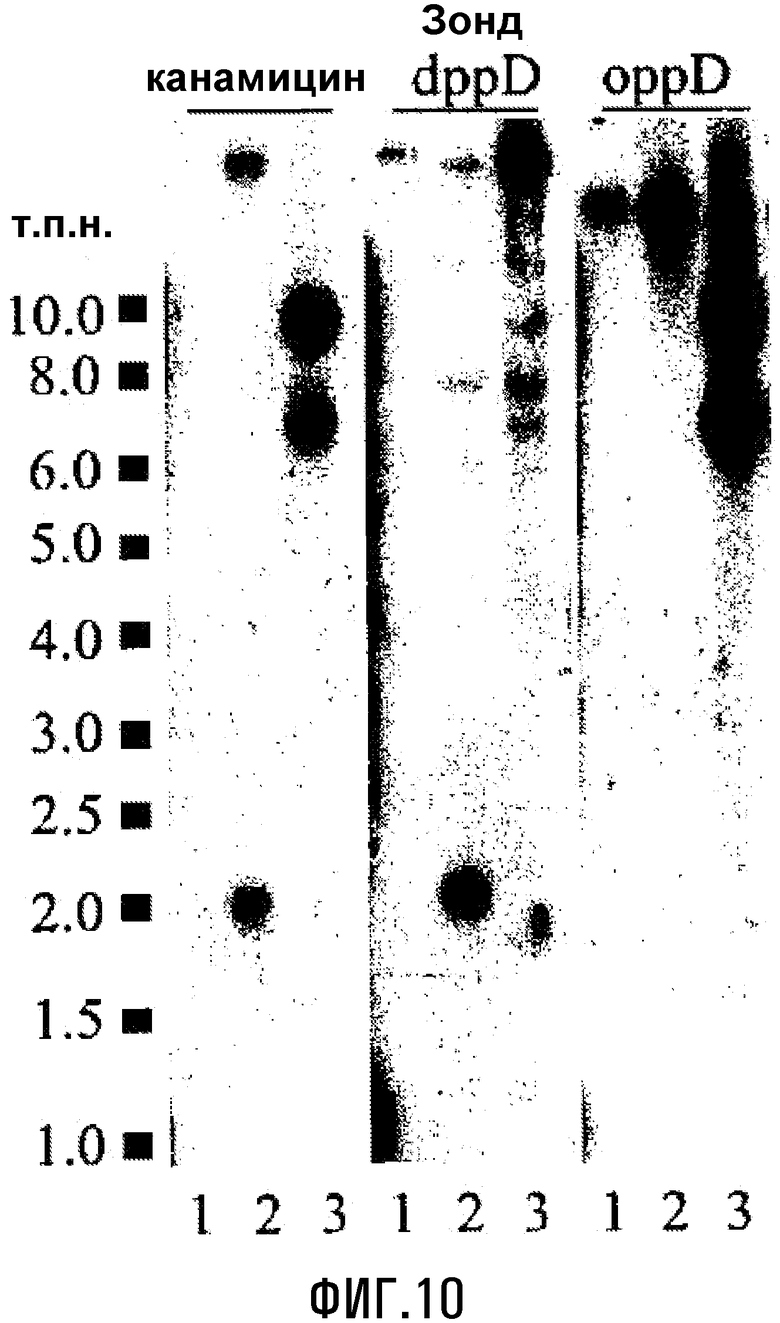
Изобретения относятся к области биотехнологии и касаются cпособа предотвращения или лечения заболевания у субъекта, вызванного патогенным организмом, путем введения вакцинной композиции, вакцинной композиции и ее применения. Охарактеризованная вакцинная композиция содержит бактерии, аттенуированные мутацией в гене, кодирующем АВС-пептидный транспортерный белок ОppD, которые могут персистировать в субъекте. Указанная мутация делает кодируемый АВС-пептидный транспортерный белок нефункциональным. Представленные изобретения могут быть применимы в иммунологии для получения и применения вакцин. 3 н. и 16 з.п. ф-лы, 10 ил., 21 табл., 5 пр.
1. Способ предотвращения или лечения заболевания у субъекта, вызванного патогенным организмом, предусматривающий введение терапевтически эффективной дозы вакцинной композиции или иммуногенной композиции субъекту, где вакцинная композиция или иммуногенная композиция содержат бактерии, аттенуированные мутацией в гене, кодирующем АВС-пептидный транспортерный белок, где:
указанная мутация делает кодируемый ABC-пептидный транспортерный белок нефункциональным,
указанные аттенуированные бактерии могут персистировать в субъекте, и
указанный ABC-пептидный транспортерный белок представляет собой OppD.
2. Способ по п. 1, где мутация генерируется инсерцией, делецией или заменой или любой их комбинацией.
3. Способ по п. 2, где инсерционная мутация выполняется любым способом или любой комбинацией способов из гомологичной рекомбинации, транспозонного мутагенеза и мутагенеза последовательности-метки.
4. Способ по п. 1, где эта бактерия выбрана из группы, включающей Avibacterium, Bacillus, Brucella, Bartonella, Bordetella, Burkholderia, Vibrio, Escherichia, Salmonella, Clostridium, Campylobacter, Chlamydia, Coxiella, Erysipelothrix, Francisella, Listeria, Actinobacillus, Haemophilus, Helicobacter, Aeromonas, Pseudomonas, Streptococcus, Shigella, Yersinia, Mycoplasma, Mycobacterium, Mannheimia, Ornithobacterium, Rickettsia, Ureaplasma и Pasteurella.
5. Способ по п. 4, где бактерией является Avibacterium paragallinarum, Bordetella avium, Ornithobacterium rhinotracheale, Salmonella enteritidis, Pasteurella multocida, Mannheimia haemolytica, E.coli, Clostridium perfringens, Mycoplasma agalactiae, Mycoplasma alkalescens, Mycoplasma anatis, Mycoplasma anseris, Mycoplasma imitans, Mycoplasma arginini, Ureaplasma parvum, Mycoplasma arthritidis, Mycoplasma bovigenitalium, Mycoplasma bovirhinis, Mycoplasma bovis, Mycoplasma bovoculi, Mycoplasma californicum, Mycoplasma capricolum, Mycoplasma dispar, Mycoplasma felis, Mycoplasma fermentans, Mycoplasma genitalium, Mycoplasma hominis, Mycoplasma hyopneumoniae, Mycoplasma hyorhinis, Mycoplasma hyosynoviae, Mycoplasma iowae, большая колония и малая колония Mycoplasma mycoides subsp mycoides, Mycoplasma gallisepticum, Mycoplasma synoviae, Mycoplasma orale, Mycoplasma penetrans, Mycoplasma ovipneumoniae, Mycoplasma pullorum, Mycoplasma alligatorus, Mycoplasma pneumoniae, Mycoplasma, Mycoplasma maleagridis, Mycoplasma haemofelis, Mycoplasma haemominutum или Mycoplasma haematoparvum.
6. Способ по п. 5, где бактерия является птичьим патогенным штаммом Е. coli Е956 или штаммом Mycoplasma gallisepticum Ap3AS.
7. Способ по п. 1, где аттенуированная бактерия экспрессирует гетерологичный антиген.
8. Способ по п. 7, где гетерологичный антиген кодируется нуклеиновой кислотой из того же самого или другого патогенного организма.
9. Способ по п. 8, где нуклеиновая кислота, кодирующая гетерологичный антиген, выделена из родов, выбранных из группы, включающей Avibacterium, Bacillus, Brucella, Bartonella, Bordetella, Burkholderia, Vibrio, Escherichia, Salmonella, Clostridium, Campylobacter, Chlamydia, Coxiella, Erysipelothrix, Francisella, Listeria, Actinobacillus, Haemophilus, Helicobacter, Aeromonas, Pseudomonas, Streptococcus, Shigella, Yersinia, Mycoplasma, Mycobacterium, Mannheimia, Ornithobacterium, Rickettsia, Ureaplasma и Pasteurella и любую их комбинацию.
10. Способ по п. 9, где нуклеиновая кислота, кодирующая гетерологичный антиген, получена из Avibacterium paragallinarum, Bordetella avium, Ornithobacterium rhinotracheale, Salmonella enteritidis, Pasteurella multocida, Mannheimia haemolytica, E.coli, Clostridium perfringens, Mycoplasma agalactiae, Mycoplasma alkalescens, Mycoplasma anatis, Mycoplasma anseris, Mycoplasma imitans, Mycoplasma arginini, Ureaplasma parvum, Mycoplasma arthritidis, Mycoplasma bovigenitalium, Mycoplasma bovirhinis, Mycoplasma bovis, Mycoplasma bovoculi, Mycoplasma californicum, Mycoplasma capricolum, Mycoplasma dispar, Mycoplasma felis, Mycoplasma fermentans, Mycoplasma genitalium, Mycoplasma hominis, Mycoplasma hyopneumoniae, Mycoplasma hyorhinis, Mycoplasma hyosynoviae, Mycoplasma iowae, большой колонии и малой колонии Mycoplasma mycoides subsp mycoides, Mycoplasma gallisepticum, Mycoplasma synoviae, Mycoplasma orale, Mycoplasma penetrans, Mycoplasma ovipneumoniae, Mycoplasma pullorum, Mycoplasma alligatorus, Mycoplasma pneumoniae, Mycoplasma, Mycoplasma maleagridis, Mycoplasma haemofelis, Mycoplasma haemominutum, Mycoplasma haematoparvum или любой их комбинации.
11. Способ по п. 8, где нуклеиновая кислота, кодирующая гетерологичный антиген, выделена из вирусов, выбранных из группы, включающей вирус болезни Ньюкасл (псевдочумы птиц), вирус инфекционного бронхита, птичий пневмовирус, вирус оспы птиц, инфекционного бурсального заболевания, вирус инфекционного ларинготрахеита, птичьего гриппа, вирус гепатита уток, чумы уток, инфекционной куриной анемии, вирус болезни Марека и любую их комбинацию.
12. Способ по любому из пп. 1-11, где субъект представляет собой позвоночное животное, выбранное из группы, включающей людей, коровьих, собачьих, кошачьих, козлиных, овечьих, свиных, верблюдовых, лошадиных и птичьих.
13. Способ по п. 12, где субъект коровьих представляет собой корову, быка, бизона или буйвола, субъект собачьих представляет собой собаку, субъект кошачьих представляет собой кошку, субъект козлиных представляет собой козу, субъект овечьих представляет собой овцу, субъект свиных представляет собой свинью, субъект верблюдовых представляет собой верблюда, дромадера, ламу, альпака, викунью или гуанако, субъект лошадиных представляет собой лошадь, осла, зебру или мула.
14. Способ по п. 12, где субъект представляет собой субъекта птичьих, выбранного из кур (в том числе карликовых кур, бентамок), индеек, уток, гусей, фазанов, перепелов, куропаток, голубей, цесарок, страусов, эму и павлинов.
15. Вакцинная композиция, пригодная для предотвращения или лечения заболевания, вызванного патогенным микроорганизмом, содержащая имуногенно эффективное количество бактерий, аттенуированных мутацией в гене, кодирующем АВС-пептидный транспортерный белок, и фармакологически приемлемый носитель, где:
указанная мутация делает кодируемый ABC-пептидный транспортерный белок нефункциональным,
указанные аттенуированные бактерии могут персистировать в субъекте, и
указанный ABC-пептидный транспортерный белок представляет собой OppD.
16. Вакцинная композиция по п. 15, где вакцинная композиция содержит, по меньшей мере, один фармацевтически приемлемый носитель или разбавитель, выбранные из группы, состоящей из воды, солевого раствора, культуральной жидкости, стабилизаторов, углеводов, белков, белоксодержащих агентов, таких как бычья сыворотка или обезжиренное молоко, и буферы, или любую их комбинацию.
17. Вакцинная композиция по п. 16, где стабилизаторы включают SPGA.
18. Вакцинная композиция по любому из пп. 15-17, где вакцинная композиция содержит, по меньшей мере, от приблизительно 103 до приблизительно 105 аттенуированных бактерий, или от приблизительно 105 до приблизительно 107 аттенуированных бактерий, или от приблизительно 107 до приблизительно 109 аттенуированных бактерий, или от приблизительно 109 до приблизительно 1011 аттенуированных бактерий, или от приблизительно 1011 до приблизительно 1013 аттенуированных бактерий, или от приблизительно 1013 до приблизительно 1015 аттенуированных бактерий, или от приблизительно 1015 до приблизительно 1017 аттенуированных бактерий, или от приблизительно 1017 до приблизительно 1019, или, по меньшей мере, приблизительно 1019 аттенуированных бактерий на дозу.
19. Применение вакцинной композиции по п. 15 для предотвращения или лечения заболевания, вызванного патогенным организмом.
| WO 2002034773 A2, 02.05.2002 | |||
| WO 2004089408 A2, 21.10.2004 | |||
| G | |||
| M | |||
| S | |||
| Rosinha et al., Identification and Characterization of a Brucella abortus ATP-Binding Cassette Transporter Homolog to Rhizobium meliloti ExsA and Its Role in Virulence and Protection in Mice, Infect Immun., 2002 September, Vol.70, No.9, p.p.5036–5044 | |||
| Helen S | |||
| Garmory et |

