Изобретение относится к пероральным вакцинам на основе генетически аттенуированных (т.е. с ослабленной вирулентностью) микроорганизмов и к самим микроорганизмам, в частности к аттенуированным штаммам Salmonella.
В 1950 г. Bacon el al. (Br. 1. Exp. Path. 31: 714 - 724) продемонстрировали, что некоторые ауксотрофные мутанты S.typhi в мышах аттенуируются в сравнении с родительским штаммом. Некоторые из этих штаммов включали мутации в ароматическом и биосинтетическом пути пурина. Известно также, что другие мутации, такие, как Ihy A, аттенуируют (ослабляют вирулентность) бактерии.
В 1981 г. авторы Hosielh и Stocker (Nature 241:238 - 39) сообщили о конструировании мутанта S.typhimurium aro A. Мутация aro A создавалась с использованием мутагенеза транспозона Тп 10 для конструирования штаммов S. hyphimurium, несущих неревертирующие (необратимые) патологические изменения в гене aro A. Данный ген кодирует фермент 5-энолпирувилшикимат-3-фосфат синтазу ключевой фермент в ароматическом био-синтетическом метаболическом пути организма, который отсутствует у млекопитающих. Мутанты aro A, следовательно, зависимы от экзогенных ароматических соединений, включающих ароматические аминокислоты, n-амино-бензойную кислоту и 2,4-дигидроксибензоат, для роста.
На инбредных мышах было показано, что мутанты S.typhimurium ara A аттенуируются в них, и обнаружено, что они являются эффективными живыми вакцинами в мышах против сальмонеллеза, когда они доставляются орально или парэнтерально.
Если при приготовлении вакцины микроорганизм предполагается использовать в живом виде, факторы или причины безопасности диктуют, чтобы микроорганизм аттенуировался по крайней мере двумя мутациями, предпочтительно в отдельных частях генома. При этом важно, чтобы такой микроорганизм не ревертировал обратно к вирулентной родительской форме, и считается, что вероятность этого, случающегося с одной мутацией, является незначительной. Однако риск возврата к прежнему состоянию, происходящего в штамме, таящем мутации в двух дискретных генах, расположенных в различных местах генома, является незначительным.
Таким образом, считается, что двойной мутант данного сорта является гораздо более безопасным кандидатом для использования в вакцине.
В европейской патентной заявке N 184086 (Дзе Борд оф Трастиз оф дзе Леланд Станфорд Юниор и Юниверсити; изобретатель - Брюс Стокер) описывает конструирование неревертирующего штамма S.typhi, который таит неревертирующие мутации aro A и pur A. Неревертирующие (необратимые) мутации представляют собой мутации, которые не могут быть восстановлены в одну стадию. Генные мутации данного вида включают инверсии и делеции последовательно ДНК, которая составляет часть гена.
В своих экспериментах авторы показали, что внутривенное назначение неревертирующих aro A pur A мутантов S.typhimurium плохо действует в части защиты BAIhB/c мышей от внутривенного введения. Кроме того, показано, что эти мутанты являются неэффективными в смысле защиты BALB/c мышей, когда они назначаются оральным путем (O'Callaghan et al., 1988 Inf ect. Immun. 56, 419 - 423).
Мутации aro A и pur A могут быть приготовлены с использованием транспозонов. Они являются последовательностями ДНК от 750 пар оснований до многих тысяч нуклеотидных пар, которые могут интегрировать их генетический материал в различные положения в бактериальной хромосоме. Некоторые транспозоны кодируют ген, который сообщает антибиотическую устойчивость организму, содержащему транспозон. Когда инсерция происходит в каком-то локусе (местоположения гена), непрерывность гена часто прерывается и это приводит в результате к потере функции гена. При частоте примерно 10-8 (клеток) генерацию транспозоны определенно исключаются (делетируются) из гена. Это восстанавливает функцию гена, однако более часто происходит нечеткая эксцизия. Это не восстанавливает генную функцию и часто ведет к неревертирующей мутации.
Некоторая часть работы, осуществленной в поддержку данной заявки, сосредоточена на S. typhi, являющемся причиной брюшного тифа людей. S. typhi в основном является патогенным организмом людей и, таким образом, не подходит для экспериментальной работы с большинством животных. Исследования на животных проводятся с использованием мышей в качестве модели и организма S. typhimurium, близко родственного организма, который вызывает "тифо-подобное" заболевание мышей и крупного рогатого скота. Описание данной модели мышей можно найти в работе Collins, 1974, Bacteriol Rev. 38, 371.
Для того, чтобы микроорганизм рассматривался на предмет использования в вакцине, он должен обладать следующими свойствами:
достаточно аттенуироваться, так чтобы он по существу был не способен вызвать заражение, связанное с неаттенуированным микроорганизмом;
быть по существу не способным к возвращению к вирулентности;
быть способным вызывать иммунность у животных, инокулированных данным организмом и таким образом обеспечивать защиту от последующего введения вещества вирулентным штаммом.
При этом считают, что живые микроорганизмы, описанные в известных источниках и предлагаемые к использованию в качестве вакцин, не отвечают всем необходимым требованиям, отмеченным выше. Поэтому остается все еще необходимость разработки живой вакцины, которая не имела бы недостатков известного уровня техники, поскольку показано (Collins, Bacteriol. Rev, 1974), что обычно живые бактерии показывают более высокую эффективность иммунизации, чем убитые бактерии. Настоящие изобретатели показали, что введение неревертирующей мутации в каждый из двух дискретных генов в ароматическом пути бактерий дает подходящий организм для использования при изготовлении живой вакцины.
Таким образом, согласно первому аспекту изобретения обеспечивается способ получения аттенуированного штамма Salmonella для использования в качестве вакцины, предусматривающий введение второй неревертируемой мутации в хромосому исходного штамма бактерий Salmonella, содержащих первую неревертируемую мутацию в гене aro A, отличающийся тем, что вторую неревертируемую мутацию вводят в ген aro, отличный от гена, содержащего первую мутацию.
При этом имеется по крайней мере десять генов, вовлеченных в синтез хоризмата, узлового соединения в биосинтетическом пути ароматических аминокислот. Некоторые из них располагаются на хромосомной карте в широко различающихся местоположениях на бактериальном геноме, т.е. aro A (5-энолпирувилшикимат-3-фосфат синтаза) aro C (хоризмат синтаза) aro D (3-дигидрохинат дегидратаза) и aro E (шикимат дегидрогеназа). Предпочтительно вторая мутация согласно изобретению вводится в arо C или aro E гены. Согласно трем воплощениям изобретение предоставляет aro A aro E мутантные бактерии, aro A aro C мутантные бактерии и aro A aro D мутантные бактерии, хотя в сфере настоящего изобретения находятся и другие двойные aro мутанты.
Изобретение применимо к различным штаммам Salmonella, например S. typhi, вызывающим брюшной тиф у людей, S. typhimurium вызывающим сальмонеллез у некоторых видов животных; S. enteritidis, вызывающим пищевые отравления у людей, S. cholerasuis, являющихся причиной сальмонеллеза у свиней и S. dublin.
Согласно предпочтительному воплощению изобретения предоставляется штамм S. typhi Ty 2, таящий или aro A aro C, или aro A aro E, или aro A aro D-неревертирующие мутации.
Конструирование S.typhi Ty 2 aro A описывается в MGG 207, 402 (Dougan et al. ). Неревертирующие мутации генерируются трансдукцией (или переносом) LT2 aro A: Tn 10 маркера в штамм S. typhi Ty 2. Tn 10 транспозон несет ген, кодирующий устойчивость к тетрациклину. При этом выбираются трансдуктанты, которые являются устойчивыми к тетрациклину при росте колоний на соответствующей среде. Дальнейший отбор производится путем скрининга тех организмов, которые имеют ген устойчивости к тетрациклину и которые являются также ароматически зависимыми.
Альтернативный метод введения делеции в S. typhi aro A ген (или другие S. typhi aro гены) включает транспозоновый мутагенез клонированного S. typhi aro A гена, рекомбинирование видоизмененного (мутированного) гена в S. typhi хромосому, заменяющую ген дикого типа мутантом и отбор или селекцию неопределенной эксцизии транспозона. Данный метод устраняет введение на S. typhi ДНК в штамм вакцины.
В основном имеется несколько способов введения второй мутации во второй ген. Один способ является транспозонным мутагенезом. Он включает инсерцию (включение) транспозируемого элемента во второй ген, а затем достижение его неопределенной эксцизии бактерий таким же самым образом, как описано выше для конструирования первой мутации. Введение мутации в еще один aro ген дает двойной aro мутант. Это фенотипически не отличимо от одинарного aro мутанта. Таким образом, для комплементаци первого мутированного aro гена клонированная копия одного из генов интродуцируется на плазмиду в организме, и организм проверяется на зависимость от ароматического соединения с использованием соответствующей селекционной среды.
Трансформацию исходного штамма бактерий, используя плазмидную ДНК, содержащую инактивированный ген aro (отличный от инактивированного гена aro A) и селективный маркер, с последующей селекцией штамма, у которого инактивированный ген встроен в хромосому путем рекомбинации. Более специфично этот другой способ включает клонирование второго гена в вектор, например плазмиду или космиду, а затем введение способного к селекции маркерного гена в клонированный второй ген, в то же самое время инактивируя этот ген. Плазмида, несущая инактивированный ген и иной селекцируемый маркер, может вводиться в организм с помощью известных приемов. Затем с помощью подходящей селекции можно идентифицировать мутант, в котором неактивированный ген рекомбинировался в собственную хромосому организмов, а собственная копия гена организмов утеряна. В особенности используемым вектором является вектор, который не стабилен в организме и спонтанно теряется. Мутированный ген в плазмиде и собственная копия гена организмов в его хромосоме могут быть обменены с помощью генетического перекрестного метода. Дополнительные методы устраняют (элиминируют) введение чужеродного ДНК в вакцинные штаммы в сайте мутации.
Мутанты aro A aro C и aro B aro E и aro A aro D изобретения являются достаточно аттенуированными, чтобы быть по существу безопасными для использования в вакцинах, и являются достаточно иммуногенными, чтобы вызывать иммунную реакцию в пациенте, которая будет обеспечивать защиту пациента от последующего введения веществ вирулентным штаммом.
Штаммы изобретения могут производиться методами генной инженерии, так, чтобы экспрессировать антигены из одного или более различных патогенов. Такие патогены могут быть вирусными, бактериальными, протозойными или более высокоорганизованными паразитическими организмами. Патогены могут заражать, как людей, так и других млекопитающих, и могут быть селективными видами или даже специфичными видами. Такие штаммы могут затем составлять основу би- или поливалентной вакцины. Примеры полезных антигенов включают антигены E. coli тепло-лабильный токсин B подгруппа (LT - B) E. coli K 88, FM DV (Fool and Mouth) пептиды, протеины вирусного гриппа, 69 Kd протеин из B. perfussis. Другие антигены, которые могли бы полезным образом экспрессироваться, представляют антигены из Chlamydia, трематод, микоплазмы, круглых червей, ленточных червей, вируса бешенства и ротавируса.
Эти антигены могут получаться путем введения гена или генов, кодирующих их, в экспрессионные касcеты. Касcеты экспрессии включают ДНК последовательности в дополнение к последовательности, кодирующей структурный ген, который кодируют транскрипционную и трансляционную области инициирования и терминации. Кассета экспрессии может также включать регуляторные области. Такие экспрессионные кассеты хорошо известны в данной области техники, и квалифицированный специалист вполне может сконструировать их. Экспрессионной кассетой может быть сконструированная или встречающаяся в природе плазмида. Например, аттенуированной методом генной инженерии Salmonella, которая экспрессирует чужеродный антиген, можно найти в патентной заявке EP (на европейский патент) N 0127153 A/SSVI/Веллкам/. Экспрессионная кассета также может быть получена инженерными методами для обеспечения возможности введения гетерологического гена в бактериальную хромосому.
Еще одна бивалентная вакцина, включающая аттенуированный микроорганизм Salmonella typhi, способный экспрессировать подгруппу B тепло-лабильного энтеротоксина E. coli, была описана авторами Clements et al. (Infection and Immunity, 46, N 2, ноябрь 1984, с.564 - 569). Ty21a использовался для экспрессии других антигенов, таких, как антиген формы I Shigella sonnei (Formal et al. Infection and Immunity 34, 746 - 750).
Аттенуированная бактерия, описанная здесь, следовательно, может быть трансформирована экспрессионной кассетой, кодирующей антиген патогена, в котором при использовании названный антиген экспрессируется указанной аттенуированной бактерией.
Фармацевтическая композиция, которая включает аттенуированные бактерии, описанные здесь, может быть получена в сочетании с фармацевтически приемлемым носителем. Предпочтительно фармацевтической композицией является композиция вакцины.
Вакцина благоприятным образом представлена в лиофилизованном виде, например в форме капсулы для орального назначения пациенту. Такие капсулы могут быть снабжены энтерическим покрытием, включающим, например, Eudragate "S" Eudragate "L" ацетат целлюлозы, фталат целлюлозы или гидроксипропилметилцеллюлозу. Эти капсулы могут использоваться как таковые или альтернативно перед назначением, лиофилизованный материал может преобразовываться, например, в виде суспензи. Преобразование (реконституирование) успешно проводится в буфере для защиты аттенуированных бактерий и вакцины от желудочной кислотности, перед каждым назначением вакцины назначается препарат бикарбоната натрия. Альтернативно вакцина может приготавливаться для парэнтерального назначения, интраназального или интрамаммарного (внутрь молочной железы) назначения.
Способ профилактики бактериальной инфекции включает назначение пациенту эффективной дозы описанной выше вакцины. Применяемая в таком способе дозировка зависит от различных клинических факторов, включающих размер и вес пациента, тип сформированной вакцины. Однако в случае аттенуированной S. typhi доза, включающая назначение 109 - 1011 организмов S. typhi на дозу, обычно удобна для взрослого человека весом 70 кг.
Ниже представлены примеры, включающие экспериментальные подробности в соответствии с настоящим изобретением. При этом очевидно понятно, что эти примеры никоим образом не предназначены для ограничения изобретения.
Конструирование вакцинных штаммов S. typhi aro A, aro E и aro A, aro C и aro A aro D.
Все штаммы конструировались с использованием исходного штамма S. typhi Ty2 aro A, описанного подробно ранее и являющегося объектом работы Douqan et al., Mol. Gen. Genel (1987) 207:402 - 405.
Пример 1. S. typhi Ty2 aro A aro E
Штамм S. typhimurium LT2 aro E: : Tn 10 получался из Центра запасов Salmonella в Калгари. Он был первоначально выделен I. Roth. Для приготовления лизата на LT2 aro E::Tn10 выращивался фаг P 22. Лизат фага P 22 использовался для трансдуцирования S. typhi Ty2 aro A селективного по устойчивости к тетрациклину. В этот момент плазмида, кодирующая клонированный aro A ген, из S. typhimurium C5 для комплементации aro A мутации вводилась в S. typhi aro A штамм. S. typhi aro A, aro E::Tn 10, несущий клонированный aro A ген был фенотипически зависимым от ароматических соединений (обычно требуемый aro мутантами). Элемент Tn 10 удалялся из aro E гена с помощью отбора разновидностей чувствительных к тетрациклину на среде Bochner, приема, используемого ранее (Bochner et al, I. Bacteriol, 143, 929 - 933). Мутанты S. typhi aro A aro E, таящие клонированный aro A ген проверялись на зависимость от ароматических соединений с использованием минимальной среды. Ароматически зависимые колонии отбирались и обширно проверялись на aro A реверсию путем помещения 1011 организма на минимальную среду с недостатком ароматических соединений и инкубирования среды при 37oC и проверки в течение 5 дней на ревертантные колонии. Колонии, которые стабильно были aro E несмотря на тщательный отсев, подвергались разведению /размножению/ для отбора вариантов, которые самопроизвольно теряли клонированный aro A ген.
Отбирался один мутант S. typhi aro A aro E, который депонировался в Национальной коллекции Культур, 61 авеню Колиндейл, Лондон NW 9 5HT под номером 12164, датированным 25 ноября 1987 г, в соответствии с условиями Будапештского договора.
Пример 2. S. typhi Ty 2 aro A aro C.
В качестве исходного штамма использовался штамм S. typhi Ty 2 aro A. Клонировался aro C ген S. typhi Ty 2 с использованием козмид для комплементации E. coli aro C с использованием методов Honn и Collins (Gene 11:291 - 298/1978). Козмида aro C подвергалась мутагенезу транспозоном Tn 5 и субклонировалась для размещения небольшого фрагмента ДНК, кодирующего клонированный aro C ген. Клонированный aro C ген инактивировался с помощью клонирования гена устойчивости к металлу ртути в кодирующую область aro C. Плазмида, несущая инактивированный aro C, вводилась в S. typhi aro A. Данная плазмида также содержит ген устойчивости к ампициллину. С помощью отбора устойчивости к ртути и чувствительности к ампициллину можно идентифицировать мутант, в котором инактивированный aro C рекомбинирован в хромосому S. typhi для генерирования S. typhi aro A aro C мутанта.
При этом осуществлялось альтернативное конструирование, которое включало использование гена устойчивости к канамицину (Km-P) вместо гена устойчивости к ртути.
S. typhi Ty 2 aro A aro C Km-P депонировался в Национальной Коллекции культур, 61 авеню Колиндейл, Лондон NW9 5HT под номером 12165, датированным 25 ноября 1987 в соответствии с условиями Будапештского договора.
Пример 3. Конструирование двойного мутанта Salmonella typhi aro A aro D.
В качестве исходного штамма использовался S. typhi aro A. Конструирование S. typhi aro A aro D достигалось трансдуцированием штамма лизатом фага Р22, полученного с использованием донорного штамма LT2 aro D 553::Tn 10, и отбора на устойчивость к тетрациклину. Один изолят очищался и использовался для приготовления чувствительных к тетрациклину производных путем отбора на среде Bochner. Некоторые из них очищались и трансформировались плазмидой pAB51 (aro A+) для комплементации делеции aro A. Один из чувствительных к тетрациклину изолятов, который был стабильно зависимым от ароматических соединений, когда таил в себе данную плазмиду, обозначался S. typhi Ty 2 aro A aro D.
Все сконструированные штаммы S.typhi, таящие мутации в различных aro генах, все еще продуцировали Vi антиген, были "O" неагглютинируемыми, 09 - агглютинируемыми после кипячения и были жгутикового типа Ho. Один такой штамм aro A aro D депонировался в Национальной коллекции культур, 61 авеню Колиндейл, Лондон, под номером, датированным в соответствии с условиями Будапештского договора.
Пример 4. Конструирование двойных aro мутантов в S.typhimurium, S.dublin и S. cholerasuis.
Делеция aro A вводилась в S. typhimurium SL 1344, S. dublin, S. cholerasuis с использованием метода McFarland и Slocker. Лизат фага, полученный из штамма TT 472, использовался для трансдукции всех штаммов Salmonella с отбором колоний, устойчивых к тетрациклину. Штамм TT 472 несет Tn 10, вставленный в Ser C, который находится в верховье и в том же опероне, что и aro A. Устойчивые к тетрациклину трансдуктанты были зависимыми от ароматических соединений, серина и пиридоксина. Кроме того, приготавливался еще один лизат P 22, выращенный на SL 5254, который имеет известную делецию в aro A. Он использовался для трансдукции устойчивых к тетрациклину штаммов, которые представляли Ser C::Tn 10, и трансдуктанты отбирались на минимальной среде, испытывающей недостаток серина и пиридоксина, но содержащей ароматические соединения. Колонии, растущие на минимальной среде с добавлением ароматических соединений, но в отсутствии серина и пиридоксина, были чувствительными к тетрациклину и зависимыми от ароматических соединений.
Пример 4а. Конструирование S.typhimurium aro A aro C.
S. typhimurium aro A aro C конструировался сначала путем передвижения стабильной aro C мутации из авирулентного штамма S. typhimurium SA 2018 в вирулентный для мышей штамм S. typhimurium SL 1344 с использованием ряда трансдукций. P 22-трансдуцирующий лизат, полученный с использованием штамма S. typhimurium SGSC 592 Ze 608::Tn 10, использовался для трансдукции Tn 10 в штамм SA2018. Tn 10 в штамме SGSC 592 на 40% связан с aro C геном дикого типа, тогда как штамм SA 2018 таит в себе стабильную aro C мутацию. Устойчивые к тетрациклину трансдуктанты собирались на минимальной среде и было обнаружено, что некоторые колонии были зависимыми от ароматических соединений. Одни из этих изолятов очищался и использовался для приготовления еще одного лизата фага P22.
Данный лизат использовался для трансдукции S. typhimurium SL 1344, и снова идентифицировались несколько устойчивых к тетрациклину, зависимых от ароматических соединений трансдуктантов. Один изолят использовался для приготовления чувствительных к тетрациклину производных с помощью отбора на среде Bochner. Один чувствительный к тетрациклину зависимый от ароматических соединений изолят очищался, и в него вводилась делеция aro A с использованием метода McFarland и Slocker, как описано ранее. Для подтверждения того, что данный изолят представлял aro A aro C, он трансформировался или плазмидой pAB51 (aro A+) или PTMC12H (aro C+), и было найдено, что он является зависимым от ароматических соединений, когда он таит в себе любую из этих плазмид.
Пример 4b. Конструирование S. typhimurium aro A aro D.
S.typhimurium aro A aro D конструировался с помощью введения aro D делеции в штамм SL 1344 aro A. Это достигалось с помощью трансдукции штамма лизатом фага P22, приготовленным с использованием донорного штамма LT2 aro D 553: : Tn 10 и путем отбора на устойчивость к тетрациклину. Один изолят очищался и использовался для приготовления чувствительных к тетрациклину производных с помощью отбора на среде Bochner. Несколько из них очищались и трансформировались плазмидой pAB51 (Aro A+) для комплементации делеции aro A. Один из чувствительных к тетрациклину изолятов, который был стабильно зависимым от ароматических соединений, когда он таил в себе данную плазмиду, обозначался SL1344 aro A aro D.
Пример 4c. Конструирование S. typhimurium aro A aro E.
S. typhimurium aro A aro E конструировался с помощью введения aro E делеции в штамм SL 1344 aro A. Это достигалось с помощью трансдукции штамма лизатом фага P22, приготовленным с использованием донорного штамма LT2 aro E: : Tn 10, и отбора на устойчивость к тетрациклину. Один изолят очищался и использовался для приготовления чувствительных к тетрациклину производных с помощью отбора на среде Bochner. Несколько из них очищались и трансформировались pAB51 (aro A+) для комплементации aro A делеции. Один из чувствительных к тетрациклину изолятов, который был стабильно зависимым от ароматических соединений, когда он таил в себе данную плазмиду, обозначался SL1344 aro A aro E.
Пример 5. Производные S. dublin и S. cholerasuis aro A aro D конструировались таким же образом, как производные SL1344 aro A aro D, как показано в примере 4b.
Свойства ин виво аттенуированных штаммов S. typhimurium
Подробности о штаммах S. typhimurium, использованных в данной работе, показаны в табл. 1.
Заражение мышей и определение бактерий в органах мышей
Для определения числа организмов в различных органах после tnt внутривенной инокуляции S. typhimurium, печень и селезенка гомогенизировались, как описано в работе Hormaeche, CE, Immunology, 37, 311 - 318. Подсчет жизнеспособных особей производился на этих гомогенатах с использованием в качестве ростовой среды L-агара и выражался в числах как геометрические средние значения ± два стандартных отклонения от среднего для четырех мышей. В примерах 6, 7 и 8 использовались восприимчивые от природы к Salmonella BALB/c мыши в возрасте 8 - 10 нед. Величина ЛД50 при внутривенном введении (в.в) для вирулентных штаммов получалась с помощью инъецирования групп из 5 мышей серийными десятикратными разбавлениями, приготовленными в буферированном фосфатом физиологическом растворе с pH 7,2 (PBS), ночных культур L-бульона, собранных с помощью центрифугирования и повторно суспендированных в PBS с получением концентрации 109 - 1011 бактерий на мл. Это были серийно десятикратно разбавленные в PBS препараты, причем верхняя доза составляла 0,2 мл чистой суспензии, даваемой в.в. или орально с использованием 8 - 10 мышей в группе. Смертные случаи регистрировались на протяжении последующих четырех недель, и вычислялась величина ЛД50 с использованием метода Reed и Muench Am. I. Hyg. 27: 493 - 497, 1983. Для в.в. - инокуляции мышей инъецировали 0,2 мл бактериальной суспензии в хвостовую вену.
Для оральной инокуляции бактерии назначались в объеме 0,2 мл слегка анестезированным эфиром мышам с помощью кормления через желудочный зонд, как описано ранее /Microbial Pathogenesis 2: 211 - 221.
Пример 6. Аттенуирование вирулентных штаммов S.typhimurium с помощью введения стабильных aro мутаций и других ауксотрофных мутаций.
S. typhimurium HWSH и SL1344 оба представляли вирулентные для мышей штаммы с величиной ЛД50 менее чем 10 организмов после в.в. введения мышам BALB/c. Величины ЛД50 при внутривенном введении определялись для отобранных ауксотрофных производных этих штаммов на мышах BALB/c (табл. 2). Все aro A и pur A мутанты были вполне аттенуированными по сравнению с родительскими штаммами. HWSH aro A и SL3261 (SL1344 aro A) имели величины ЛД50 log 7.4 и log 7.1 соответственно, что вполне согласовывалось с опубликованными данными. Производные pur A и aro A pur A были даже более аттенуированными. Штамм HWSH pur E был значительно менее аттенуированным, хотя было найдено, что измеренная величина ЛД50 значительно варьировала от эксперимента к эксперименту. Например, иногда мыши, которым давалось такое небольшое количество, как 100 организмов, умирали, тогда как другие, которым давалось 104 - 105 организмов, выживали. Мутант aro C aro A был менее аттенуированным, чем мутант aro A pur A, имеющий ЛД50 6.5. Данная цифра была той же самой для одинарного aro C мутанта. При оральном введении ароматические или пуриновые мутанты не убивали мышей, даже когда им давались такие высокие дозы, как 1010 организмов.
Пример 7. Устойчивость одинарных и двойных aro мутантов S. typhimurium после в.в. инокуляции мышей BALB/c.
Все штаммы S. typhimurium обнаруживали очень сходный характер устойчивости, при этом никаких сальмонелл не обнаружено в селезенке на 56 день (чертеж).
Аттенуированные штаммы S. typhimurium в виде перорально назначаемых вакцин.
Пример 8. Определялась способность орально ауксотрофных штаммов S.typhimurium защищать мышей BALB/c от орального введения вещества вирулентным S. typhimurium (SL1344). Мышей сначала заражали орально аукстрофными мутантами в количестве между 109 - 1010, а затем иммунизированным мышам орально вводили через четыре недели родительский вирулентный штамм.
Результаты, приведенные в табл. 3, ясно показывают, что ни pur A, ни aro A pur A мутанты не обеспечивали какой-либо значительной защиты от орального введения, тогда как мутанты aro A, aro C и aro A aro C обеспечивали существенную защиту против орального введения провокационной пробы как через 28, так и через 70 дней после иммунизации.
Пример 9. Препаративная форма. Организм S. typhi изобретения предпочтительно представлен в форме таблеток для введения через рот.
Ингредиент - мг/таблетку
Таблетки (основная масса)
1) Подвергнутый сушке вымораживанием эксципиент носитель, содержащий 109 - 1010 Salmonella typhi - 70,0
2) Двуокись кремния /Аэросил 200/ - 0,5
3) Дипак /97% сахароза/ - 235,0
4) Поперечно сшитый Повидон /Коллиндон С/ - 7,0
5) Микрокристаллическая целлюлоза /Авицел pH 102/ - 35,0
6) Стеарат магния - 2,5
Покрытие - 350,0
7) Opadry Enteric, OY-P-7156
8) Фталат поливинилацетата + Диэтилфталат - 35,0 - 385,0
Приготавливается носитель, содержащий 5% сахарозы, 1% глютанат натрия и 1% бакто-каситон в водном растворителе. Организм S.typhi суспендируется в данном носителе, а затем подвергается сушке вымораживанием.
Высушенный вымораживанием материал смешивается с Аэросилом 200, и смешанная смесь просеивается через сито. Просеянный порошок смешивается с веществами Дипак, Коллиндон CL, Авицел pH 102 и стеаратом магния в смесителе. Данная смесь прессуется в таблетки для последующего нанесения энтерического (кишечного покрытия).
Специалист в данной области техники очевидно сознает, что многие из ингредиентов в данной препаративной форме могут заменяться функциональными эквивалентными фармацевтически приемлемыми эксципиентами /носителями/.

Способ обеспечивает получение штаммов бактерий рода Salmonella, содержащих неревертируемые мутации в генах, ответственных за образование метаболитов ароматических соединений. Мутации затрагивают гены aro A , aro C или aro E; их вводят путем транспозонного мутагенеза или трансформацией с помощью плазмидной ДНК, включающей инактивированный ген aro. Такие штаммы могут эффективно использоваться для приготовления вакцин. 4 з.п. ф-лы, 1 ил., 3 табл.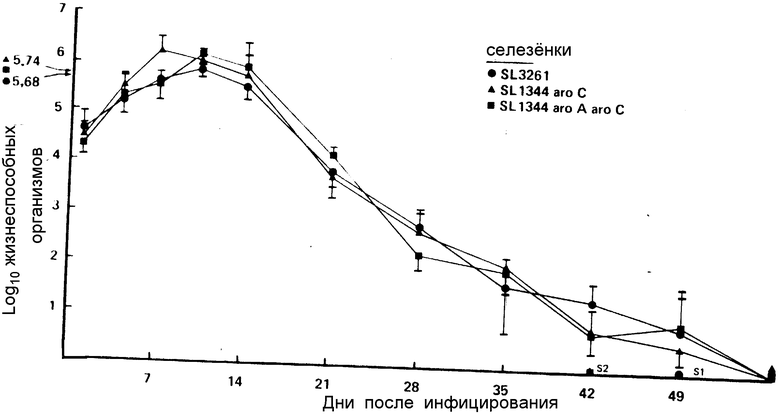
| EP, заявка, 0184086, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
