Сведения относительно финансирования исследования государством
Данное изобретение было создано при поддержке государством грантами Министерства Здравоохранения R21 AI066010, R01 AI067703, R01 AI063503, и R03 AI083251. Правительство США имеет права на данное изобретение.
Область техники
Настоящее изобретение относится к белкам клеточной оболочки гифа Candida, к антителам, полученным в результате иммунного ответа на вакцинацию с белками клеточной оболочки гифа Candida, и к способам профилактики и/или лечения кандидоза и других бактериальных инфекций с белками клеточной оболочки гифа Candida.
Уровень техники
Все документы, указанные в настоящем изобретении, или указанные части, включены в настоящем изобретении посредством ссылок, включая предварительную заявку США с порядковым номером 61/223,005, поданную 3 июля 2009. Никакой документ, однако, не признан известным уровнем техники для заявленного изобретения.
Приблизительно 60,000 случаев диссеминированного кандидоза происходит ежегодно в Соединенных Штатах Америки [1], в результате чего затрачиваются миллиарды долларов на медицинскую помощь. С учетом 40% смертности от подобных инфекций, существует необходимость выявления новых профилактических средств или терапевтических мишеней для воздействия.
Основным иммунологическим защитным механизмом против диссеминированного кандидоза является фагоцитарный киллинг организма [2.3]. Только фагоцитарные клетки способны на непосредственный киллинг Candida in vitro [4]. Кроме того, в течение тридцати минут внутривенного введения Candida мышам, кроликам, собакам или людям, дрожжевые грибки сохраняются внутри ретикулоэндотелиальной системы, особенно в печени. Печень, с высоким содержанием макрофагов Купффера, способна очистить 99,9% дрожжевых грибков в системе воротной вены в течение одного прохода [5], что подчеркивает эффективность фагоцитарного защитного механизма против грибов. Таким образом, резистентность С. albicans к фагоцитарному киллингу является важнейшей вирулентной функцией организма.
Гликозилфосфатидилинизитол (GPI)-закрепленные белки клеточной поверхности, находятся на главной границе раздела между патогеном и хозяином, что делает эти белки возможными участниками во взаимодействии хозяин-патоген [6].
Идентификация эффекторов в регуляторном пути организма, которые участвуют в вирулентности, обеспечивает возможность для терапевтического воздействия с помощью способов или композиций, которые превосходят существующие противогрибковые средства. Идентификация белков клеточной поверхности или белков, относящихся к гифам, которые влияют на регуляторный путь, связанный с вирулентностью, является особенно перспективным потому, что характеристика белка позволяет осуществлять иммунотерапевтические методы, которые с большой вероятностью превосходят или оказывают синергическое действие с существующими противогрибковыми средствами при борьбе с кандидозной инфекцией.
Вирулентность С. albicans регулируется несколькими предполагаемыми вирулентными факторами, среди которых прилипание к компонентам хозяина и способность превращаться из дрожжевых грибков в гифы являются наиболее важными при определении патогенности. Несмотря на то, что эффективные противогрибковые средства существуют, которые являются бактерицидными по отношению к Candida, смертность, вызываемая кандидемией составляет приблизительно 38%, даже при лечении сильными противогрибковыми веществами, таким как амфотерицин В. Также, существующие вещества, такие как амфотерицин В, обычно проявляют нежелательную токсичность. Хотя дополнительные противогрибковые препараты, которые могут быть разработаны, являются менее токсичными, чем амфотерицин В, маловероятно, что эти вещества будут более эффективными. Поэтому, любая пассивная или активная иммунотерапия для лечения или профилактики диссеминированного кандидоза является преспективной альтернативой стандартной противогрибковой терапии.
Таким образом, существует необходимость в эффективных иммуногенах, которые обеспечат хозяину иммунную защиту и пассивную иммунологическую защиту против Candida и других иммуногенных подобных патогенов. Настоящее изобретение отвечает этим требованиям и также имеет соответствующие преимущества.
Сущность изобретения
Настоящее изобретение относится к полипептидным антигенам HYR1 Candida и терапевтическому применению подобных антигенов. Полипептидные антигены HYR1 настоящего изобретения могут быть использованы для лечения или профилактики инфекции Candida у субъекта.
При помощи скрининга новых систем условной гиперэкспрессии/супрессии, направленного на GPI - закрепленные белки на C. albicans, авторы изобретения идентифицировали HYR1 как вирулентный фактор. HYR1 является ко-экспрессирующим геном гифа, нулевой мутантный штамм которого не отображает каких- либо морфологических нарушений in vitro [7]. Ниже авторы приводят результаты, наглядно показывающие, что HYR1 является посредником резистентности к фагоцитарному киллингу in vitro, модулирует тканевую грибковую нагрузку in vivo и является, таким образом, мишенью вакцины для облегчения тяжести диссеминированного кандидоза.
Определения
Термин «HYR1» полипептид означает полипептид, который по существу идентичен аминокислотной последовательности SEQ ID NO:1. Желательно, чтобы полипептид HYR1 имел, по меньшей мере, 70, 75%, 80%, 85%, 90%, 95%, 99% или даже 100% идентичности с аминокислотной последовательностью SEQ ID NO:1.
Термин «фрагмент полипептида HYR1» или «фрагмент HYR1» означает фрагмент полипептида HYR1, содержащий менее, чем 937, 936 или 935 аминокислот. Преимущественно, фрагменты HYR1 составляют от 300 до 350 или от 250 до 500 аминокислот в длину. Желательно, чтобы фрагмент был менее чем 937, 936, 935, 934, 933, 932, 931 или 930, 920, 910, 900, 890, 880, 870, 860, 850, 840, 830, 820, 810, 800, 790, 780, 770, 760, 750, 740, 730, 720, 710, 700, 690, 680, 670, 660, 650, 640, 630, 620, 610, 600, 590, 580, 570, 560, 550, 540, 530, 520, 510, 500, 490, 480, 470, 460, 450, 440, 430, 420, 410, 400, 390, 380, 370, 360, 350, 340, 330, 320, 310, 300, 290, 280, 270, 260, 250, 240, 230, 220, 210, 200, 190, 180, 170, 160, 150, 140, 130, 120, 110, 100, 90, 80, 70, 60, 50, 40, 30, 25, 20, 15, или 10 аминокислот и, желательно, являлся иммуногенным. Фрагмент HYR1, например, может содержать одну или более консервативных аминокислотных замен в последовательности SEQ ID NO:2. Дополнительные желательные фрагменты HYR1 содержат одну или более консервативных аминокислотных замен в последовательности SEQ ID NO:2 и/или, по меньшей мере, одну фланкирующую аминокислоту (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 фланкирующих аминокислот) на N- и/или С- конце последовательности SEQ ID No:2. Другие предпочтительные фрагменты HYR1 содержат семь и более последовательных аминокислот последовательности SEQ ID NO:2.
Неограничивающие примеры фрагмента HYR1 включают аминокислоты 1-40, 10-50, 20-60, 30-70, 40-80, 50-90, 60-100, 70-110, 80-120, 90-130, 100-140, 110-150, 120-160, 130-170, 140-180, 150-190, 160-200, 170-210, 180-220, 190-230, 200-240, 210-250, 220-260, 230-270, 240-280, 250-290 и 260-300, 270-310, 280-320, и 290-331 последовательности SEQ ID NO:2; и эти фрагменты имеют одну или более следующих особенностей: одну или более консервативных аминокислотных замен (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или 16 консервативные аминокислотные замены) в последовательности SEQ ID NO:2; одну или более аминокислот (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или 16 аминокислот) усеченных на N- и/или С-конце последовательности SEQ ID NO:2; и, по меньшей мере, одну фланкирующую аминокислоту (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 фланкирующих аминокислот) на N- и/или С-конце последовательности SEQ ID NO:2.
Термин «практически идентичный» означает полипептид, проявляющий, по меньшей мере, 50%, желательно 60%, 70%, 75% или 80%, более желательно 85%, 90% или 95%, и наиболее желательно 99% идентичности аминокислотной последовательности к указанной аминокислотной последовательности. Длина сравниваемых последовательностей, как правило, по меньшей мере, 10 аминокислот, желательно, по меньшей мере, 15 заменимых аминокислот, более желательно, по меньшей мере, 20, 25, 50, 75, 90, 100, 150, 200, 250, 275, 300, 310, 315, 320, 325, 330, 335, 340, 345 или 350 заменимых аминокислот, и наиболее желательно, полная длина аминокислотной последовательности.
Идентичность последовательности может быть измерена при использовании компьютерной программы анализа последовательности по умолчанию (например, Sequence Analysis Software Package of the Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Avenue, Madison, WI 53705). Такая компьютерная программа может противопоставлять похожие последовательности, устанавливая степень гомологии различных замен, делеций и других модификаций. Множественные последовательности могут быть также выравнены с использованием программы Clustal W(1.4) (производство Julie D. Thompson и Toby Gibson of the European Molecular Biology Laboratory, Germany и Desmond Higgins of European Bioinformatics Institute, Cambridge, UK), путем ввода парного выравнивания режимом «медленно», параметры парного выравнивания включают штраф на внесение делеции в выравнивание 10.0 и штраф на продолжение выравнивания 0.1, а также задают сходную матрицу «blosum». Дополнительно, параметры множественного выравнивания могут включать штраф на внесение делеции в выравнивание 10.0, штраф на продолжение выравнивания 0.1, а также задают сходную матрицу «blosum», замедляют расхождения до 40% и протяженность разрыва до 8.
Термин «консервативная аминокислотная замена», используемый в данной заявке, означает замену в аминокислотной последовательности аминокислоты на другую в пределах семейства аминокислот, а именно родственных по химическим свойствам их боковых цепей.
Генетически закодированные аминокислоты можно подразделить на четыре семейства: кислые (аспартат, глутамат), основные (лизин, аргинин, гистидин), неполярные (аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан) и полярные незаряженные (глицин, аспарагин, глутамин, цистеин, серии, треонин, тирозин). Фенилаланин, триптофан и тирозин иногда группируют как ароматические аминокислоты. Аналогично, аминокислоты могут также разделять на следующие группы: кислые (аспартат, глутамат), основные (лизин, аргинин, гистидин), алифатические (глицин, аланин, валин, лейцин, изолейцин, серин, треонин), с серином и треонином не обязательно группируют отдельно как алифатическую гидроксильную; ароматические (фенилаланин, тирозин, триптофан); амиды (аспарагин, глутамин); и серосодержащие (цистен, метионин).
Любое изменение в аминокислотной последовательности, приводящее к функциональным гомологам, может быть определено путем оценки способности различных пептидов функционировать некоторым образом подобно белку дикого вида, используя стандартные способы, такие как исследования, описанные здесь.
Желательное осуществление настоящего изобретения включает, по меньшей мере, одну консервативную аминокислотную замену в аминокислотной последовательности SEQ ID NO:1 или 2; и, более желательно, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 консервативных аминокислотных замен в последовательности SEQ ID NO:1 или 2.
Термин «фланкирующая аминокислота» означает аминокислоту в полипептидной последовательности, которая непосредственно прилегающую на N-или С-конце той или иной определенной последовательности. Желательно, фланкирующая аминокислота находится на N- и/или С-конце аминокислотной последовательности SEQ ID NO:1 или 2 или их фрагмента; и более предпочтительно, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 фланкирующие аминокислоты находятся на N- и/или С-конце аминокислотной последовательности SEQ ID NO:1 или 2, или их фрагмента.
Используемый в данной заявке термин «белок слияния» относится к полипептиду, состоящему из (1) полипептида HYR1, фрагмента HYR1; и (2) партнера слияния.
Используемый в данной заявке термин «партнер слияния» относится к гетерологичной последовательности, которая может сливаться с полипептидом HYR1 или фрагментом HYR1. Примеры партнеров слияния описаны в данной заявке и включают определение маркеров, стабилизацию доменов или последовательностей, которые способствуют производству или очистке белков.
Используемый в данной заявке термин «иммунный ответ» относится к активации иммунной системы организма в ответ на антиген или возбудитель инфекции. У позвоночных он может включать, но не ограничиваться ими, одну или более следующих: мутация нативных В-клеток в В-клетках памяти; производство антител клетками плазмы (эффекторные В-клетки); индукция клеточного иммунитета; активация и освобождение цитокина CD4+ Т-клетками; активация и освобождение цитокина CD4+ Т-клетками; пополнение цитокинов и активация фагоцитарных клеток (например, макрофагов, нейтрофилов, эозинофилов); и/или активация комплемента.
Термин «иммуногенный» означает любую субстанцию, способную индуцировать иммунный ответ у субъекта.
Термин «фармацевтически приемлемая соль» означает любую нетоксичную кислотно-аддитивную соль или металлокомплекс, используемые в фармацевтической индустрии. Примеры кислотно-аддитивных солей включают органические кислоты, такие как уксусная, молочная, памовая, малеиновая, лимонная, яблочная, аскорбиновая, янтарная, бензойная, пальмитиновая, пробковая, салициловая, винная, матансульфоновая, толуолсульфокислая или трифторуксусная кислоты или в этом роде; полимерные кислоты, такие как дубильная кислота, карбоксиметилцеллюлоза или в этом роде; и неорганические кислоты, такие как солярная кислота, бромистоводородная кислота, серная кислота, фосфорная кислота или в этом роде. Металлокомплексы включают цинк, железо и тому подобное.
Термин «фармацевтически приемлемый носитель» означает любой раствор, используемый для растворения и доставки вещества субъекту. Предпочтительно, фармацевтически приемлемым носителем является солевой раствор. В предпочтительном осуществлении, фармацевтически приемлемый носитель включает адъювант. Примеры адъювантов описаны в данной заявке. Другие физиологически приемлемые носители и композиции на их основе известны специалисту в данной области техники и описаны, например, в Remington's Pharmaceutical Sciences, (20th edition), ed. A. Gennaro, 2003, Lippincott Williams & Wilkins.
Термин «выделенный» означает белок (или его фрагмент), выделенный из компонентов, которые естественным образом сопровождают его. Как правило, полипептид является по сути выделенным, когда он составляет, по меньшей мере на 60% по весу, свободный от белков и естественных органических молекул, с которыми он естественным образом связывается. Определение также распростроняется и на полипептид, отделенный от его фланкирующих аминокислот (например, для аминокислотной последовательности, выделение относится к последовательности, которая свободна от фланкирующих аминокислот, с которыми последовательность естественным образом связывается в полипептиде). Предпочтительно, полипептид, по меньшей мере, на 75%, более предпочтительно, по меньшей мере, на 90% и наиболее предпочтительно, по меньшей мере, на 99% по весу выделен. Выделенный полипептид может быть получен стандартными способами, например, путем экстракции из естественных источников (например, очистка клеток, инфицированных Candida), путем экпрессии рекомбинантных нуклеиновых кислот, кодирующих фрагмент HYR1; или слиянием его белков путем химического синтеза полипептида. Степень чистоты может быть измерена любым подходящим способом, например, колоночной хроматографией, электрофорезом в полиакриламидном геле или HPLC анализом.
Термин «терапевтически эффективное количество» означает количество иммуногенного соединения (например, полипептида, фрагмента, белка слияния или вакцины), необходимого для получения у субъекта одного или более следующих эффектов: иммунного ответа; снижения уровня инфекции Candida (например, уменьшение, по меньшей мере, на 5%, 10%, 20% или 30%; более желательно, на 40%, 50%, 60% или 70%; и наиболее желательно на 80% или 90%); ослабление (например, по меньшей мере, на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или даже 100% снижение) одного или более симптомов инфекции Candida у пациента; или повышение устойчивости к новой инфекции Candida (например, увеличение, по меньшей мере, на 5%, 10%, 20%, 30%, 40% или 50%; более желательно, на 60%, 70%, 80%, or 90%; или, наиболее желательно, на 100%, 200% или 300%).
Другие признаки и преимущества настоящего изобретения будут очевидны из следующего Подробного Описания, чертежей и формулы изобретения.
Краткое описание чертежей
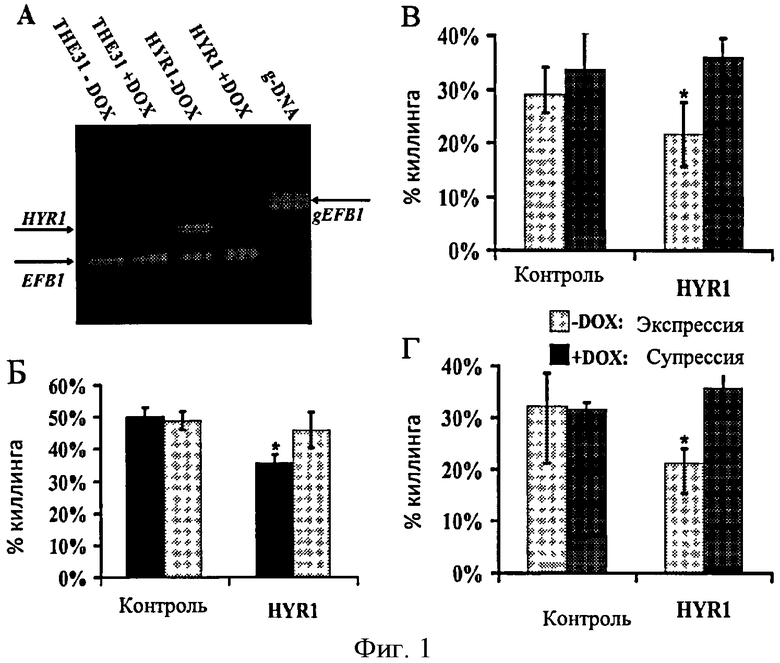
Фигура 1. Условная экспрессия HYR1 усиливает опосредованный киллинг C. albicans нейтрофилами и макрофагами. (А) Подтверждение условной сверхэкспрессии/супрессии HYR1 штамма СААН-31. Результаты RT-ПЦР демонстрируют сверхэкспрессию HYR1 гена в -DOX среде и недостаток экспрессии в+DOX среде. Фрагмент EFB1 был ко- амплифицирован и служил в качестве контроля. Отсутствие загрязнения геномной ДНК при приготовлении кДНК проверяли отсутствием 919 bp полоски, содержащейся в интроне EFB1. ТНЕ31 был контрольным штаммом дикого типа. (Б) Штаммы C. albicans выращивали в YPD с DOX (супрессия HYR1) и без DOX (сверхэкспрессия HYR1) при 30°С всю ночь и затем совместно культивировали с нейтрофилами человека; (В) C.albicans совместно культивировали с HL-60 производными нейтрофилов; (Г) с HL-60 производными макрофагов.
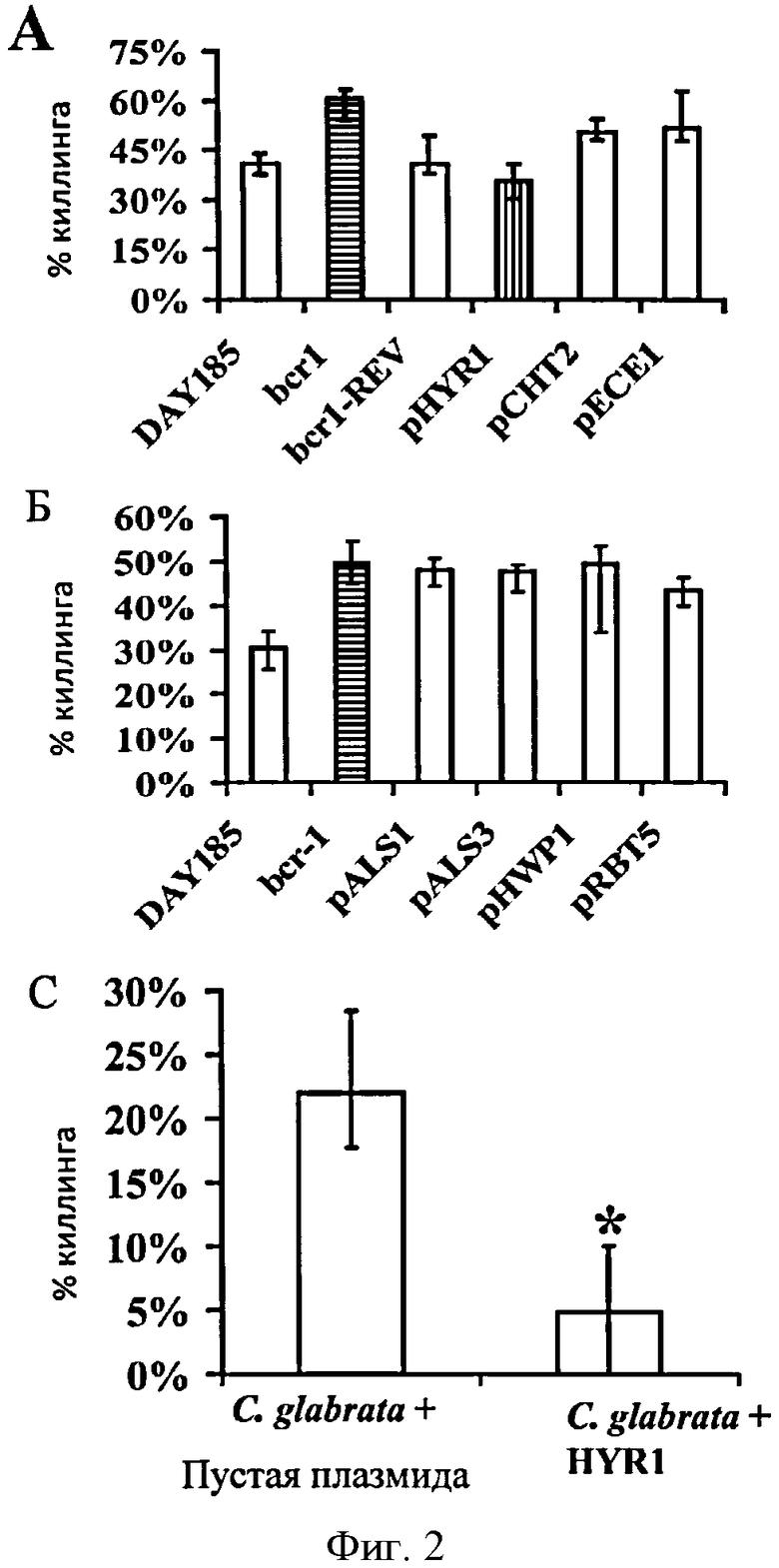
Фигура 2. Экспрессия HYR1 Candida albicans увеличивает резистентность к киллингу нейтрофилами человека в bcr1 нулевых-мутантах или штамме С. glabrata. (A и Б) Автономная экспрессия HYR1 в штамме С albicans, лишенном bcr1, полностью комплементарна повышенной восприимчивости родительского штамма к резистентности нейтрофильного киллинга. Штаммы С. albicans DAY 18 5 (дикий тип), CJN702 (bcr1 нулевой мутант) и CJN698 (BCR1 комплементарный) CJN114, CJN1153, CJN1222, CJN1259, CJN1276, CJN1281 и CJN1288 (автономно экспрессируют ALS1, ALS3, HWP1, HYR1, RBT5, СНТ2 и ЕСЕ1 в bcr1 с теневым фенотипом нулевого мутанта, соответственно) выращивают в YPD всю ночь при 30°С. Данные отображаются в виде среднего значения ± вероятность. *Р<0.04 по сравнению с диким типом и BCR1-дополненным. (В) Гетерологичная экспрессия гена HYR1 С. albicans повышает резистентность С. glabrata к HL-60 производными нейтрофилов опосредованному киллингу. *Р<0.0001.
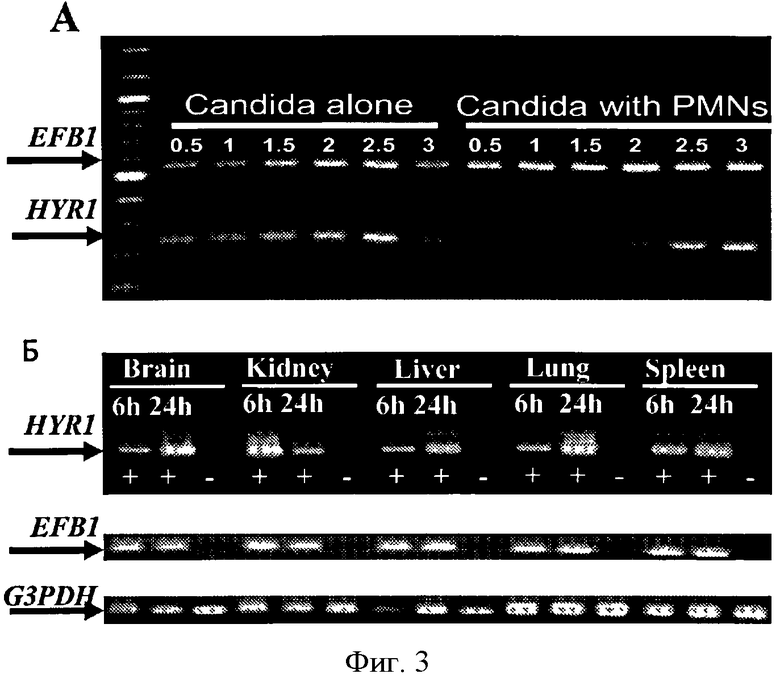
Фигура 3. Обнаружение экспрессии HYR1 при диссеминированном кандидозе, хотя эта экспрессия была изначально ингибирована нейтрофилами in vitro. (А) Почки, печень, легкие, селезенка и мозги были собраны через 6 или 24 часа после внутривенного инфицирования С. albicans. Гнездовая RT- ПЦР была использована для определения экспрессии HYR1. EFB1 С. albicans и конструктивный ген мыши G3PDH были использованы в качестве контроля. + означает инфицированные мыши, в то время как - означает неинфицированные мыши. (Б) HL-60 производные нейтрофилов ингибируют экспрессию HYR1 С. albicans. Дикий тип С. albicans культивировали в RPMI 1640 с добавлением 10% объединенной сывороткой человека. Без нейтрофилов экпрессия HYR1 была определена после получасовой индукции. С нейтрофилами, экпрессия HYR1 ингибировалась два часа.
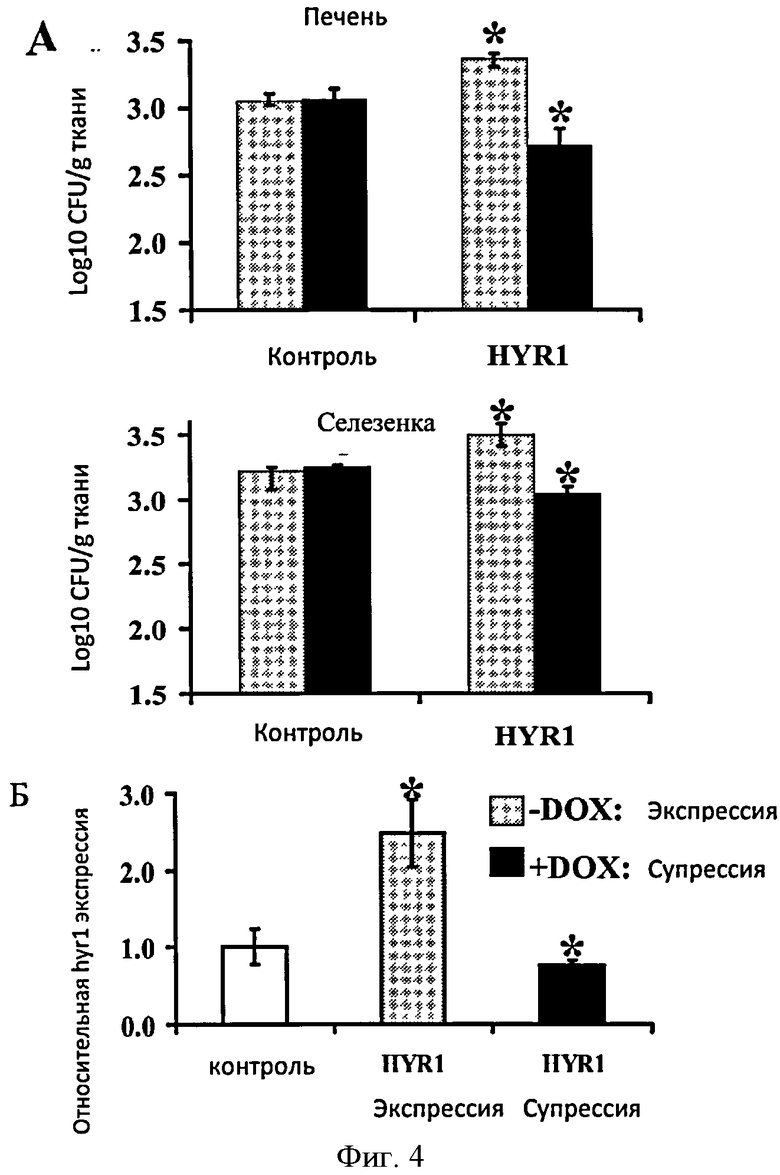
Фигура 4. Экпрессия HYR1 in vivo и его действие на грибковую нагрузку. Для определения экпрессии HYR1 в процессе диссеминированного кандидоза, почки, печень, легкие, селезенка и мозги были собраны через 6 или 24 часа после внутривенного инфицирования С. albicans. Гнездовая RT-ПЦР была использована для определения экспрессии HYR1. EFB1 С. albicans и конструктивный ген мыши G3PDH были использованы в качестве контроля. + означает инфицированные мыши, в то время как - означает неинфицированные мыши (А). Условная экспрессия HYR1 повышает грибковую нагрузку в органах с распространением тканевых фагоцитов. Нагрузка С. albicans в печени и селезенке иммунокомпетентных мышей (n=8 в группе), инфицированных HYR1 С. albicans или контрольным штаммом, выращенных в условиях условной экпресии (-DOX) или супрессии (+DOX) (А) и экпресия HYR1 in vivo (Б). Печень и селезенку собрали в тот же день после инфицирования. На оси Y отображены нижние пределы обнаружения исследования. Данные отображаются в виде среднего значения ± вероятность. *Р<0.0001 по сравнению с не экспрессирующим HYR1 штаммом (HYR1-DOX) или контрольным штаммом.
Фигура 5. Непрямая иммунофлюоресценция с анти-Hyr1p сывороткой показывает проявление экспрессии Hyr1p в гифах С, albicans. Формирование гифа было вызвано инкубацией С. albicans в RPMI 1640 в течении 90 минут. Клетки окрашивали с либо предварительно абсорбированной анти-HYR1 сывороткой hyr1 нулевого мутанта (1:1000) или анти-белком, приготовленным из сыворотки пустого плазмидного клона (отрицательный контроль) с последующей окраской Alex меченных анти-мышиных Ab.
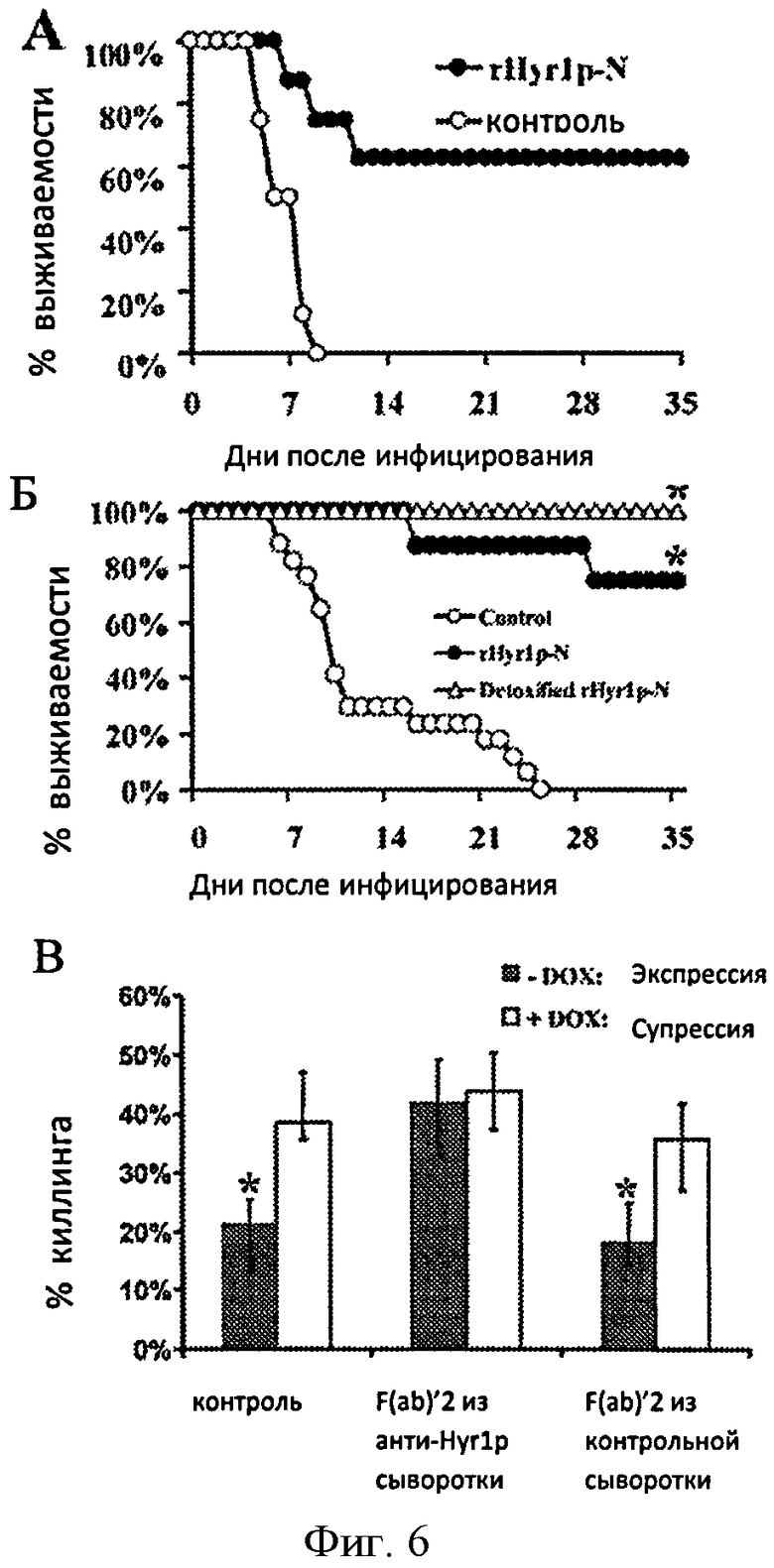
Фигура 6. Рекомбинантный N- концевой Hyr1p (rHyr1p-N) отличается защищенностью от мышинного гематогенного диссеминированного кандидоза. (А) Выживаемость мышей (8 мышей в группе), вакцинированных rHyr1p-N, смешанного с полным или неполным адъювантом Фрейда и инфицированные в хвостовую вену с 2.2×105 бластоспор SC5314 Candida albicans. (Б) Выживаемость мышей (8 мышей в группе, исключая контрольную группу, в которой 17 мышей), вакцинированных rHyr1p-N или нейтрализованным rHyr1p-N, смешанным с 0.1% гидрокисью алюминия, и инфицированных с 7×105 бластоспорами 15563 Candida albicans. *Р<0.001 логарифмический ранговый критерий. (В) Эффективность вакцинации или контроля F(ab)'2 на блокирование нейтрофильного киллинга мыши С. albicans, условно экспрессирующей или супрессирующей Hyr1. Контроль показывает исследование, выполненное при отсутствии любого F(ab)'2. Данные отображаются в виде среднего значения±вероятный диапазон. *Р=0.001 по тесту Манна-Уитни.
Подробное описание изобретения
Candida albicans является распространненным патогеном у людей. Так например, С. albicans, хотя в повседневной жизни безобидный условно-патогенный микроорганизм, в различных условиях может стать причиной целого ряда заболеваний от поверхностной кожно-слизистой инфекции, такой как вагинальный и/или орофарингиальный кандидоз, до глубокого органного поражения диссеминированным кандидозом. До вызывания болезни, грибы колонизируются в желудочно-кишечном тракте и иногда поражают кожные и слизистые оболочки. Соединение с слизистой поверхностью хозяина является ключевым требованием для этого первоначального шага. После колонизации, С. albicans попадает в кровоток путем инфицирования внутрисосудистой системы или перемещается через слизистую желудочно-кишечного тракта, поврежденную химиотерапией или стресс-язвой. Микроорганизмы затем распространяются по кровотоку, связываются и проникают внутрь васкулярного эндотелия, чтобы выйти из сосудистой сети, и попадают вглубь органов, таких как печень, селезенка и почки.
Определение и функциональная характеристика фрагмента HYR1, описанная здесь, позволяет этому полипептиду эффективно применяться в лечении кандидоза.
Природа патогенеза С. albicans, за счет присоединения к эндотельным клеткам, рассмотрена в патенте США No.5,578,309, который специально включен здесь в качестве ссылки в полном объеме. Для описания гена HYR1 и его характеристики, включая характеристику продукции гена, см. Bailey et al. (Journal of Bacteriology 178:5353-5360,1996).
Изобретение относится к вакцине, содержащей выделенный фрагмент HYR1 и, необязательно, адъювант в фармацевтически приемлемой среде. Вакцина может содержать фрагмент HYR1, полученный из вида Candida, такого как Candida albicans, Candida krusei, Candida tropicalis, Candida glabrata или Candida parapsilosis.
В изобретении применяют продукт последовательности гена HYR1 С. albicans в качестве вакцины для лечения, предупреждения или облегчения диссеминированного кандидоза. Вакцина является эффективной против различных штаммов С. albicans, a также против различных видов Candida.
Таким образом, согласно одному аспекту, изобретение относится к фрагменту HYR1 эффективному, разработанному в виде фармацевтической композиции и вводимому как вакцина с содержанием или без адъюванта. Фрагмент HYR1 может быть кандидозного происхождения и может быть получен, например, из видов, принадлежащих к классу Candida, например, Candida parapsilosis, Candida krusei, Candida glabrata и Candida tropicalis. Фрагмент HYR1 может быть получен в выделенной или очищенной форме и, таким образом, согласно одному из вариантов изобретения, фрагмент HYR1, разработанный в виде вакцины, вызывает иммунный ответ у пациента на вызванный иммунный ответ против Candida.
Изобретение также относится к способу лечения или предупреждения диссеминированного кандидоза. Способ включает введение иммуногенного количества вакцины HYR1 фрагмента. Вакцину можно вводить с содержанием или без адъюванта. Фрагмент HYR1 может быть получен из различных штаммов Candida, a также из различных видов Candida, таких как Candida albicans, Candida krusei, Candida tropicalis, Candida glabrata и Candida parapsilosis.
Эффективность вакцины заявленного изобретения против различных штаммов Candida, различных видов Candida, других бактериальных и инфекционных возбудителей и их разнообразной иммунной активности оценивают в соответствии со стандартными способами такими, как описано ниже.
Учитывая сведения и методологические руководства, приводимые в данном документе, специалисту в данной области техники очевидно, что иммуннотерапевтические способы, известные из уровня техники, могут применяться с одним или более фрагментами HYR1 в фармацевтически приемлемой композиции, вводимой в качестве вакцины с содержанием или без адъюванта. При осуществлении настоящего изобретения, термины «фармацевтическая» или «фармацевтически приемлемая» означают композиции, разработанные известными нетоксичными способами и, при необходимости, используются с носителями или добавками, которые безопасны для введения людям. Введение может производиться с использованием хорошо известных способов применения, включая, например, внутривенное, внутримышечное, внутрибрюшинное или подкожной инъекции. Указанные вакцины настоящего изобретения также могут содержать буферы, соли и другие растворители, известные специалисту в данной области техники, для сохранения активности вакцины в растворе. Аналогично, любые из большого числа адъювантов, известных из уровня техники, могут быть использованы в вакцине настоящего изобретения для выявления, активации или усиления терапевтически эффективного иммунного ответа, способного ослаблять или блокировать связывание, проникновение и/или инфицирование Candida с восприимчивой клеткой хозяина.
При этом, модификации, которые по существу не влияют на осуществление вариантов настоящего изобретения, также включены в формулу изобретения. Таким образом, следующие примеры предназначены для иллюстрации, но не ограничивающие настоящее изобретение.
Пример 1
Hyr1p Candida albicans придает резистентность к нейтрофильному киллингу и является мишенью вакцины.
Как описано выше, Candida albicans является наиболее частой причиной инвазивной грибковой инфекции у людей. Неясно, как С. albicans избегает фагоцитарной атаки и выживает в неблагоприятной кровенной среде в течение опасной для жизни системной инфекции. Применяя генетическую стратегию условной сверхэкспрессии/супрессии, авторы изобретения обнаружили, что ген HYR1 снижает фагоцитарный киллинг С. albicans in vitro и повышает тканевую грибковую нагрузку in vivo. Согласно с позитивной регуляцией транскрипционного фактора Bcr1p, HYR1 комплементарен гипервосприимчивости к фагоцитарному опосредованному киллингу bcr1 нулевого мутанта С. albicans in vitro. Кроме того, гетерологичная экспрессия HYR1 в Candida glabrata приводит организм к большей резистентности к нейтрофильному киллингу. В заключении, вакцинация рекомбинантным Hyr1p значительно защищает мышей против гематогенного диссеминированного кандидоза. Таким образом, Hyr1 является важным вирулентным фактором для С. albicans, служащим посредником резистентности к фагоцитарному киллингу. Hyr1 является, соответственно, мишенью для вакцины или других иммунологических или низкомолекулярных вмешательств для улучшения последствий диссеминированного кандидоза.
Результаты
Условная экспрессия HYR1 в бластоспорах значительно усиливает резистентность С. albicans к фагоцит- опосредованному киллингу in vitro.
Изучая функции HYR1, авторы изобретения создали условно сверхэкспрессирующий/супрессирующий штамм С. albicans, СААН-31. В СААН-31 одна аллель HYR1 контролировалась тетрациклиновым регулятором (ТК)-промотера, а другая аллель была нарушена. Полуколичественным RT-ПЦР авторы изобретения подтвердили, что HYR1 в большом количестве экспрессировался, когда бластоспоры СААН-31 росли в среде без DOX, и не определялся в присутствии DOX (Фиг.1А). Как и предполагалось, HYR1 не был обнаружен в бластоспорах дикого типа (ТНЕ31), потому что HYR1 является ко-экспрессирующим геном гиф (Фиг.1А).
Штамм СААН-31, условно сверхэкспрессирующий HYR1, и контрольный ТНЕ31 дикого типа имеют одинаковые темпы роста, независимо от присутствия или отсутствия DOX (время удвоения для контрольного штамма дикого типа без DOX=1.51±0.29 ч и с DOX=1.51±0.38 ч; время удвоения для штамма СААН-31 без DOX=1.39±0.30 ч и с DOX=1.35±0.19 ч). Авторы изобретения также определили влияние сверхэкспрессии HYR1 на нормальное накопление других GPI-закрепленных белков на клеточной поверхности. Прямой иммунофлюоресценцией авторы изобретения подтвердили, что сверхэкспрессия HYR1 не вляет на накопление GPI- закрепленных белков Alsip (данные не представлены) [8].
В продолжении многократного скрининга на вирулентно-ассоциированные фенотипы, авторы изобретения определили влияние условной сверхэкспрессии HYR1 на киллинг кандидоза фагоцитами человека. HYR1- эпрессирующий С. albicans (СААН-31-DOX) был значительно более резистентен к нейтрофил - опосредованному киллингу человека, чем дикий тип С. albicans (который не экпрессирует HYR1 в фазе бластоспор) и HYUR1-супрессорный С. albicans (СААН-31+DOX) (Фиг.1Б). Этот фенотип был не связан с DOX, в то время как киллинг не значительно различался между контрольным диким типом и HYR1-супрессорным С. albicans (+DOX).
Авторы также выполнили исследование киллинга кандидоза с использованием HL-60 клеточной линии, которая может быть дифференцирована в либо нейтрофил-подобные или макрофаг-подобные клетки [9, 10]. Как и только что собранные человеческие нейтрофилы, условная сверхэкспрессия HYR1 уменьшает киллинг бластоспор С. albicans обоими HL-60 нейтрофил-подобными (Фиг.1В) и макрофаг-подобными (Фиг.1Г) клетками in vitro.
Гипер-восприимчивость к нейтрофильному киллингу bcr1 нулевого мутанта С. albicans была дополнена экспрессией HYR1 in vitro.
Поскольку, HYR1 является геном, регулирующим последующие звенья сигнальных каскадов положительного транскрипционного регулятора Bcrip [17], авторы и изобретения предположили, что разрушение bcr1 усилит восприимчивость к нейтрофильному киллингу, при условии стимуляции дикого типа С. albicans экспрессировать HYR1 (т.е. в процессе формирования гиф). Поэтому, авторы индуцировали С. albicans в форме зародышевой трубки, инкубируя клетки в RPMI с добавлением 10% FBS при 37°С в течение 40 минут. Эти условия, как известно, индуцируют экспрессию HYR1 [18] и, в результате чего, зародышевая трубка становится достаточно короткой, что бы не формировались обширные гифы, таким образом, представляется возможность количественно описать колониеобразующие единицы (CFUs) в нашем киллинговом исследовании. Авторы изобретения сравнили нейтрофильный киллинг штамма bcr1 нулевого мутанта (CJN702), BCR1 комплементарного штамма в bcr1 теневом фенотипе нулевого мутанта (CJN698) и дикого типа штамма С. albicans (DAY185). Bcr1 нулевой мутант был гипервосприимчивым к нейтрофил-опосредованному киллингу по сравнению с BCR1-комплементарным и контрольным штаммом дикого типа (Фиг.2А). Кроме того, гипервосприимчивый к киллингу bcr1-лишенный С. albicans был полностью комплементарен автономной экспрессии HYR1 в bcr1 теневом фенотипе мутанта, но не другими клеточно-поверхностными кодирующими генами, регулирующими Bcrip [17] (Фиг.2А и 2Б).
Резистентность сверхэспресиирующего HYR1 С. albicans к фагоцитарному киллингу может быть воспроизведена гетероэкспрессией HYR1 в С. glabrata
Далее, определяя вирулентность фенотипа, опосредованного HYR1, авторы изобретения экспрессировали гетерологичный ген в BG14 С. glabrata [19], используя плазмиду pGRB2.2, несущую основной промотер PGK.1 [11]. Авторы также произвели трансформацию BG14 С. glabrata с пустой плазмидой в качестве отрицательного контроля. Экспрессия HYR1 в С. glabrata привела к 75% снижению к киллингу HL-60-производными нейтрофилов in vitro, по сравнению с С. glabrata, трансформированной с пустой плазмидой (Фиг.2В).
Нейтрофилы подавляют кандидозную экспрессию HYR1
Поскольку, условная сверхэкспрессия HYR1 присваивает Candida резистентность к нейтрофил-опосредованному киллингу, авторы изобретения применяли RT-ПЦР для исследования экспрессии HYR1 дикого типа С. albicans (SC5314) в ответ на HL-60-производные нейтрофилов in vitro. HYR1 экспрессировался в ближайшие 30 минут после воздействия среды, содержащей сыворотку, и непрерывно интенсивно экспрессировался в течение 2.5 часов в процессе культивирования (Фиг.3А). Однако, когда С. albicans подвергается воздействию HL-60-производных нейтрофилов при культивировании, даже в присутствии сыворотки, экспрессия HYR1 ингибировалась в течение 2 часов при совместном культивировании.
Дикий тип С. albicans экспрессирует HYR1 в процессе гематогенного диссеминированного кандидоза, в результате чего увеличивается тканевая грибковая нагрузка в органах с высоким содержанием фагоцитов.
Для определения экспрессии HYR1 в процессе гематогенного диссеминированного кандидоза, 5 главных органов - мозг, печень, легкое, селезенка и почки были собраны у мыши, инфицированной диким штаммом С. albicans, после 6 и 24 часов инфицирования. Улучшенное гнездовое RT-ПЦР исследование [20] применяли для оценки экспрессии HYR1 in vivo. Экспрессия HYR1 была определена во всех пяти органах (Фиг.3Б).
Сверхэкспрессия HYR1 увеличивала, а супрессия HYR1 снижала грибковую нагрузку в печени и селезенке (Фиг.4А). Кроме того, авторы изобретения подтвердили, что сверхэкспрессирующий штамм имеет значительно выше уровень HYR1, чем штамм дикого типа в печени; супрессированный штамм показывал тенденцию к снижению уровня (Фиг.4Б). В отличии от этого, экспрессия HYR1 не значительно изменяет грибковую нагрузку в почках (данные не показаны), орган без постоянных фагоцитов.
rHyr1p-N в качестве возможной вакцины
На основе анализа последовательности, Hyr1p является, предположительно, клеточно-поверхностным белком [7]. Для подтверждения этого, авторы изобретения получили рекомбинантный N- концевой Hyr1p в E.coli, трансформированных с экспрессирующим клоном, который включает 25-350 аминокислот, кодирующих последовательность (rHyr1p-N). Сыворотка от мышей, иммунизированных rHyr1p-N, была предварительно абсорбирована против hyr1 нулевого мутанта С. albicans [7], с последующим непрямым иммуно-окрашиванием гиф дикого типа. Авторы обнаружили, что клеточная оболочка гиф дикого типа С. albicans была сильно окрашена (Фиг.5), подтверждая, что их клеточная поверхность экспрессирует и, следовательно, подвергает воздействию иммунную систему.
Поскольку Hyr1p является клеточно-поверхностным белком, который придает резистентность к фагоцитарному киллингу киндидоза, авторы изобретения стремились определить его способность в качестве возможной вакцины. Мышей вакцинировали rHyr1p с добавлением адъюванта или только адъювантом. Две недели спустя после повторной иммунизации, мышей инфицировали через хвостовую вену высоко вирулентным SC5314 С. albicans. Вакцинация rHyr1p-N заметно улучшает выживаемость мышей по сравнению с вакцинированными только адъювантом (62.5 и 0% выживаемости за 35 дней, соответственно) (Фиг.6А).
Вакцинация rHyr1p-N, смешанного с полным или неполным адъювантом Фрейда или алюминием, заметно улучшает выживаемость мышей по сравнению с вакцинированными одним любым адъювантом (Фигура 6А и 6Б).
Сыворотка анти-rHyr1p усиливает нейтрофильный киллинг у мышей, непосредственно ингибируя функцию Hyr1p резистентности нейтрофилов.
Защитный эффект вакцины rHyr1p-N показывает, что сыворотка анти-rHyr1p возможно может нейтролизовать защитную функцию Hyr1p в С. albicans. Для определения могут ли антитела анти-rHyr1p непосредственно ингибировать функцию Hyr1p, авторы изобретения выделили и приготовили фрагменты F(ab)'2 из общих IgG мыши, иммунизированной rHyr1p-N или контролем (приготавливается производством из клеток E.coli, транформированных с пустой плазмидой). Авторы обнаружили, что F(ab)'2 из иммунной, но не контрольной, сыворотки способны к восстановлению нейтрофильного киллинга штамма с условной сверхэкспрессией HYR1 до уровня, приравненному к супрессированному штамму (Фигура 6В).
Заключение
В данном исследовании авторы изобретения показали, что HYR1 ко-экспрессирующий ген гиф [7, 21], кодирующий кандидозный фагоцитарный резистентный фактор. Условная экспрессия HYR1 в бластоспорах Candida является причиной грибов, более резистентных к фагоцитарному киллингу, по сравнению с бластоспорами дикого типа С. albicans. Кроме того, функция HYR1 в С. albicans повторяется в гетерологичной экспрессии гена С. glabrata in vitro. Авторы также обнаружили, что штамм, лишенный Bcr1p, транскрипционный фактор которого позитивно регулирует экспрессию HYR1 [17], проявляет усиление восприимчивости к фагоцит-опосредованному киллингу. Гипер- восприимчивость к фагоцитарному киллингу bcr1 пулевого мутанта была полностью дополнена автономной экспрессией HYR1, но не другими генами, которые кодируют GPI-белки, позитивно регулирующиеся Bcr1. Следовательно, HYR1 является геном, регулирующим последующие звенья сигнальных каскадов BCR1 в резистентном пути к фагоцитарному киллингу.
Любопытно, что HL-60 производные нейтрофилов в состоянии ингибировать экспрессию HYR1 в диком типе С. albicans в течение первоначального контакта между фагоцитами и Candida. Таким образом, динамическое взаимодействие происходит между фагоцитами хозяина и С. albicans, с помощью которого фагоциты являются посредниками задержки в экспрессии фагоцит-резистентного гена в грибах дикого типа. Это соответствует предыдущим обнаружениям того, что нейтрофилы человека задерживают формирование гифа С. albicans и экспрессию ко-экспрессирующих генов гифа[18].
Резистентность к фагоцитарному киллингу является совокупностью фенотипа, вероятно связанного с множественными факторами. Фагоциты могут убивать Candida внеклеточно или внутриклеточно. В последнее время, супероксиддисмутазу поверхности клеток С. albicans характеризуют как вирулентный фактор, который помогает грибам избежать киллинга, путем снижения высоко реактивных форм кислорода, полученных от хозяина [22]. Нейтрофилы обычно прикрепляются и распространяются по поверхности гифовых форм грибов, с учетом расширения, гифы слишком большие для полного захвата фагоцитами. Так как, С. albicans экспрессирует HYR1 в форме гифа, не исключено, что Hyr1p участвует в резистентности к фагоцитарному киллингу, предотвращая поверхностный контакт с фагоцитом. В ином случае, Hyr1p может препятствовать окислительному или неокислительному механизму фагоцитарного киллинга.
Фенотип сверхэкспрессии HYR1 in vitro был повторен in vivo. В процессе мышиной диссеминированной инфекции, сверхэкспрессия HYR1 приводила к значительному увеличению, а супрессия HYR1 приводила к значительному уменьшению тканевой грибковой нагрузки по сравнению с штаммом дикого типа в органах при наличии фагоцитов. Отсутствие фенотипа в почках вероятно отражает тот факт, что почки не имею в наличии фагоцитов и, что поступление нейтрофилов в почки в процессе летального диссеминированного кандидоза не начинается до >24 часов инфекции [15]. Однако, вакцинация rHyr1p-N приводит к значительной защите от гематогенного диссеминированного кандидоза. Эффективность, увиденная от вакцины rHyr1p-N, была больше, чем прежде увиденная у мышей, вакцинированных rAlsp-N и rAls3p-N вакцинами. Таким образом, rHyr1p-N является многообещающим кандидатом на вакцину для диссеминированного кандидоза.
Данные, полученные авторами, также демонстрируют преимущество применения условной сверхэкспрессии/супрессии, подводящей к изучению потенциальных вирулентных функций данного гена. Крупномасштабные генетические подходы к изучению вирулентности генов in vitro нуждаются в функциональном скрининговом исследовании и ограничиваются скринингом для генов, а именно экспрессирующих во время проведения исследования. Когда ген не экспрессирует значительно в штамме дикого типа при условиях использования для скринингового исследования in vitro, принудительная сверхэкспрессия гена по-прежнему предусматривает определение феномена получение - функция в исследовании, как и в случае HYR1 в процессе роста бластоспор Candida. Когда ген сильно экспрессирует, условная супрессия приводит к потери функции фенотипа в том же исследовании. Кроме того, когда сверхэкспрессия и супрессия используются одновременно, фенотип может быть усилен, как в случае с воздействия HYR1 на грибковую нагрузку печени и селезенки in vivo. Способность обнаруживать фенотип путем сравнения сверхэкспрессии и супрессии гена, позволяет условной экспрессии гена ослабить ограниченную функциональную избыточность, которая особенно досаждает передовым генетическим методикам, когда исследуются члены генетического семейства.
В заключение, авторы применяли методику условной экспрессии гена для идентификации Hyr1p как экспрессируемого поверхностью вирулентного фактора для С. albicans. Экспрессия HYR1 опосредует нейтрофильный киллинг in vitro и увеличивает тканевую грибковую нагрузку in vivo. В конечном итоге, авторы показали, что HYR1 является многообещающей мишенью вакцины, которая заслужила дальнейшей разработки, как профилактическая стратегия для диссеминированного кандидоза.
Материалы и методы
Описываемые выше результаты получают, используя следующие материалы и методы.
Штаммы и условия культивирования
Все использованные штаммы перечислены в Таблице 1, и выращивались, как описано ранее [8].
Условная сверхэкспрессия/супрессия HYR1 конструкции мутанта
Для образования условной экспрессии HYR1 штаммом, промотерную кассету HIS1-TR [8] встраивали перед одной аллелью гена HYR1 штамма ТНЕ4, получая в результате штамм СААН. URA3 локуса HIS1 в штамме СААН выпетливали с образованием СААН-1. Вторая аллель HYR1 в СААН-1 была разрушена повторным использованием кассеты URA3, образовывая штамм СААН-2, за которым следует повторение URA3, получая в результате штамм СААН-3. Фрагмент Nhe I-Pst 3.9 кб, содержащий ген URA3-IRO1, был встроен в исходный локус на геноме СААН-3, получая в результате СААН-31. Используемые праймеры описаны в Таблице 1.
Полуколичественный RT-ПЦР
Полуколичественный RT-ПЦР определяет экспрессию генов in vitro, как описано ранее. Праймеры, используемые для детекции экспрессии EFB1 EFBIa и EFBIb; праймеры, использованные для амплификации HYR1 HYR1 specific 1 и HYR1 specific 2 (Таблица 1). Для исследования воздействия нейтрофилов на экспрессию HYR1 С. albicans, 1×106 клеток SC5314 всю ночь выращивали в YPD либо совместно культивируя с 1×107 HL-60 производными нейтрофилов или, культивируя только в RPMI 1640 с добавлением 10% объединенной сывороткой человека. Образцы были взяты с 30 минутными интервалами в течение 3 часов пока извлекали РНК и производили полуколичественный RT-ПЦР.
Исследование фагоцитарного киллинга
Нейтрофилы человека были выделены, HL-60 клетки дифференцировались на нейтрофилы и макрофаги и исследование фагоцитарного киллинга производили, как описывалось ранее [8-Ю]. Вкратце, фагоциты инкубировали с грибами в течение 1 часа и затем обрабатывали ультразвуком и количественно культивировали. Процент киллинга рассчитывали путем деления количества колоний грибов после совместной инкубации с фагоцитами на количество колоний грибов, инкубированных в среде без фагоцитов. Нейтрофилы человека и HL-60 производные нейтрофилов или макрофагов исследовали в 2:1 и 20:1 соотношении фагоциты:грибы, соответственно. Для bcr1 и других мутантов, бластоспоры предварительно проращивали в течение 40 минут в RPMI 1640 с добавлением 10% FBS при 37°С перед проведением исследования.
Гетерологичная экспрессия HYR1 в BG14 С glabrata
BG14 С. glabrata была. трансформирована с любым из pGRB2.2-HYR1 экспрессионным вектором HYR1 или pGRB2.2 пустой контрольной плазмидой [11]. Кодирующая последовательность HYR1 была усилена CG-Hyr1-a и CG-Hyr1-b (Таблица 1) и клонирована в Xba I, Xho I сайты pGRB2.2 с использованием In-Fusion™ 2.0 Dry-Down PCR Cloning Kit, согласно инструкции изготовителя (Clontech Laboratories, Mountain View, CA).
Экспрессия HYR1 С. albicans в процессе гематогенной инфекции
Экспрессия HYR1 диким типом SC5314 С. albicans была исследована в процессе гематогенного диссеминированного кандидоза, как описывалось. Мозги, печень, легкие, почки и селезенка мышей BALB/C были собраны 6 и 24 часа после заражения. Использовали праймеры, перечисленные в Таблице 1. Обратная транскрипция осуществлялась с RETROscript (Ambion, Texas). Для амплификации конститутивного гена G3PDH мыши, были использованы праймеры G3PDHF и G3PDHR. Для определения HYR1 и EFB1 С. albicans, проводили два раунда ПЦР. В первом раунде использовали пару внешних праймеров (EFB1F и EFB1R для EFB1, или Р2 и Р5 для HYR1)', во втором раунде использовали определенное количество (1 мкл) продукта первого раунда ПЦР в качестве матрицы. Внутренние пары праймеров были следующие: EFB1nF и EFB1nR (для EFB1), или Р2 и Р4 (для HYR1). Все условия ПЦР были следующие: денатурация при 95°С, 2 минуты и амплификация в течение 35 циклов при 94°С, 30 с (денатурирование), 55°С, 30 с (отжиг), и 72°С, 90 с (элонгация). Для qRT-ПЦР кДНК была приготовлена, как указано выше. Оптимизация эффективности амплификации и SYBR green исследование реал-тайм RT-ПЦР проводили, как описано [12]. Конструктивно экспрессирующийся АСТ1 был использован в качестве контроля для всех реакций. Расчеты и статистические исследования проводили, как описано в ABI PRISM 7000 Sequence Detection System User Bulletin 2 (Applied Biosystems, USA).
Тканевая грибковая нагрузка
Мышам давали воду с содержанием или без DOX (2 мг/мл), разведенную с 5% раствором сахарозы, на протяжении периода исследования, исходя из: день - 3 относительно инфекции [13], и давали еду и воду без ограничения. Тканевая грибковая нагрузка производилась, как описывалось ранее [8], за исключением органов, удаленных после 1 дня заражения. Все процедуры, проводимые с мышами, были одобрены институтом использования животных и охранным комитетом, соблюдая NIH принципы.
Производство rHyr1p-N
rHyr1p-N (от 25-350 аминокислот Hyr1p) был произведен в экспрессионной системе pQE-32 Escherichia coli (Qiagen), a 6XHis меченный белок очищали, как описано в работе [14], за исключением использования HisPur Cobalt смолы (Thermo Scientific) хроматографической колонки. Эндотоксин удаляли из rHyr1p-N путем использования Detoxi-Gel Endotoxm Removing Columns (Thermo Scientific), и уровень эндотоксина определяли с Limulus Amebocyte Lysate endochrome (Charles River), согласно инструкции производителя. Используя эту процедуру, эндотоксин снижают до <0.28 EU на дозу, используемую для вакцинации.
Иммунофлюоресцентное определение клеточной локализации Hyr1p
Непрямая иммунофлюоресценция производится с применением поликлональной анти-Hyr1p антисывороки, полученной от иммунизированных rHyr1p-N мышей (от 25-350 аминокислот). Инокулят 1×107 бластоспор hyr1 нулевого штамма инкубировали в RPMI 1640 в течение 90 минут при 37°С и осаждали дважды для абсорбирования сыворотки.
Бластоспоры С. albicans (1×105) были предварительно пророщены в RPMI в течение 90 минут при 37°С и перенесены на 4-луночное предметное стекло (Nalge Nunc International Corp, IL, USA). После инкубации при 4 С в течение 30 минут, клетки блокировали 300 мкл 1,5% козьей сыворотки, окрашивали с поликлональной сывороткой в 1:100 разведении или PBS в качестве отрицательного контроля, и затем флуоресцинизотиоцианатом метили козьи анти-мышиные IgG 1:200. Клетки отображались с помощью конфокальной сканирующей лазерной микроскопии [15].
Протокол иммунизации
Все вакцинировались подкожно, в основании шеи. Восемь молодых особей (10-12 недель) C57BL/6 мышей вакцинировали с 20 мкг аффинно очищенных rHyr1p-N с полным адъювантом Фрейда и повторяли иммунизацию неполным адъювантом Фрейда (IFA) через 3 недели. Дополнительно восемь молодых особей мышей принимали только адъювант, смешанный с продуктом, полученным от трансформированных клеток Е. coli с пустой плазмидой. Четырнадцать дней после повторной иммунизации, мышей инфицировали через хвостовую вену с 5×105 клеток дикого типа SC5314 С. albicans [16].
Эффективность rHyr1p-N в защите против гематогенного диссеминированного кандидоза была также установлена с использованием алюминия (2% гидрата алюминия; Brenntag Biosector), адъюванта, утвержденного Управлением по контролю качества пищевых продуктов и лекарственных средств (FDA) для использования в организме человека. Также, для определения того, что rHyr1p-N защищает против других штаммов С. albicans, авторы применяли другой клинический штамм 15563. Для этих экспериментов, 33 мкг аффинно очищенные rHyr1p-N смешивали с 0.1% гидрата алюминия и вводили в BALB/c мышам как указано выше в день 0, после повторно иммунизировали на день 21, и затем инфицировали на день 35 с С. albicans путем инъекции в хвостовую вену. Для всех экспериментальных вакцинаций выживаемость мышей в течение 35 дней после инфицирования была использована в качестве конечной точки.
Исследование F(ab)'2 блокирования. Объединенные анти-Hyr1p или контрольная сыворотка были собраны от 5 мышей, которые вакцинировались либо rHyr1p-N или продуктом, полученным от трансформированных клеток Е. coli с пустой плазмидой. Все IgG из обоих сывороток выделяли с использованием Nab Spin Kit (Thermo Scientific). Фрагменты F(ab)'2 очищали с Pierce F(ab)'2 Preparation Kit, согласно инструкции производителя. Клетки Candida опсонизировали на льду в течение 45 минут с 5% нормальной мышиной сывороткой (Santa Cruz Biotechnology) или 5% нормальной мышиной сывороткой с добавлением 5% F(ab)'2, полученных от любой вакцинированной rHyr1p-N или контрольной IgG мыши, предварительно смешанных с нейтрофилами мыши. Исследование мышиного нейтрофильного киллинга было описано выше.
Статистический анализ
Фагоцит-опосредованный киллинг и тканевую грибковую нагрузку между различными группами сравнивали с помощью критерия U Манна-Уитни для непарного сравнения, по необходимости. Непараметрический логарифмический ранговый критерий использовался для определения различий во времени выживаемости. Значение Р<0.05 считается важным.
Олигонуклеотиды
ПРИМЕР II
rHyr1p-N является вакциной против диссеминированного кандидоза
Уровень техники: Авторы изобретения обнаружили, что сверхэкспрессия HYR1 Candida albicans опосредует резистентность к нейтрофильному киллингу in vitro. Авторы изобретения стремились определить воздействие сверхэкспрессии HYR1 на тканевую грибковую нагрузку in vivo в процессе инфекции и установить возможность вакцинации rHyr1p-N для защиты против диссеминированного кандидоза у мышей.
Методы: Мышей инфицировали через хвостовую вену сверхэкспрессирущим/супрессированным HYR1 или диким типом С. albicans. Печень и селезенку собирали 1 день после инфицирования, уровень экспрессии HYR1 и тканевую грибковую нагрузку определяли путем qRT-ПЦР и количественным культивированием, соответственно. Для вакцинации, rHyr1p-N получали в экспрессионной системе Е. coli pQE-32 и очищали, согласно инструкции производителя (Qiagen). Мышей вакцинировали с 20 мкг rHyr1p-N в полном адъюванте Фрейда (CFA), повторно иммунизировали с неполным адъювантом Фрейда (IFA) через 3 недели и инфицировали с штаммом SC5314 С.albcians через две недели после повторной иммунизации. Контрольные мыши получали адъювант с добавлением клеток, полученных из трансформированных Е. Coli с пустой плазмидой. Результаты: Сверхэкспрессия HYR1 значительно увеличивает грибковую нагрузку и в печени, и в селезенке по сравнению с контрольным штаммом. Супрессия HYR1 значительно снижает грибковую нагрузку и в печени, и в селезенке по сравнению с контрольным штаммом. Относительный уровень экспрессии HYR1 был 2.8 и 0.8 в печени, инфицированной с сверхэкспрессирующим и супрессированным штаммом по сравнению с контролем, соответственно. Вакцина rHyr1p-N приводит к 62.5% длительному выживанию инфицированных мышей, и, наоборот, 0% выживаемости у контрольных мышей.
Заключение: Экспрессия HYR1 влияет на способность С. albicans инфицировать ткани in vivo. Кроме того, вакцинация rHyr1p-N заметно защищает мышей от диссеминированного кандидоза. Вакцина rHyr1p-N полезна для предупреждения диссеминированного кандидоза.
Ссылки
1. Spellberg BJ, Filler SG, and Edwards JE, Jr. Current treatment strategies for disseminated candidiasis. Clin Infect Dis. 2006; 42:244-251.
2. Del Poeta M. Role of Phagocytosis in the Virulence of Cryptococcus neoformans. Eukaryot Cell 2004; 3:1067-1075.
3. Koh AY, Kohler JR, Coggshall KT, Van Rooijen N and Pier GB. Mucosal damage and neutropenia are required for Candida albicans dissemination. PLoS Pathog. 2008; 4:DOI:10.1371/joumal.ppat.0040035.
4. Gulay Z, Imir T. Anti-candidial activity of natural killer (NK) and lymphokine activated killer (LAK) lymphocytes in vitro. Immunobiology 1996; 195:220-230.
5. Stone HH. Studies in the pathogenesis, diagnosis, and treatment of Candida sepsis in children. J Pediatr Surg. 1974; 9:127-133.
6. Richard M, Ibata-Ombetta S, Dromer F, Bordon-Pallier F, Jouault T and Gaillardin C. Complete glycosylphosphatidylinositol anchors are required in Candida albicans for full morphogenesis, virulence and resistance to macrophages. Mol. Microbiol. 2002; 44.
7. Bailey DA, Feldmann PJ, Bovey M, Gow NA and Brown AJ. The Candida albicans HYR1 gene, which is activated in response to hyphal development, belongs to a gene family encoding yeast cell wall proteins. J Bacteriol. 1996; 178:5353-5360.
8. Fu Y, Luo G., Spellberg BJ, Edwards JE, Jr, and Ibrahim AS. Gene overexpression/suppression analysis of candidate virulence factors of Candida albicans. Eukaryot Cell. 2008;7:483-492.
9. Nusing R, Goerig M, Habenicht AJ and Ullrich V. Selective eicosanoid formation during HL-60 macrophage differentiation. Regulation of thromboxane synthase. Eur J Biochem 1993; 212:371-376.
10. Spellberg BJ, Collins M, French SW, Edwards JE, Jr., Fu Y and Ibrahim AS. A phagocytic cell line markedly improves survival of infected neutropenic mice. J Leukoc Biol 2005; 78:338-344.
11. Eiden-Plach A, Zagorc T, Heintel T, Carius Y, Breinig F. Viral preprotoxin signal sequence allows efficient secretion of green fluorescent protein by Candida glabrata, Pichia pastoris, Saccharomyces cerevsiae, and Schizosaccharomyces pombe. Appl. Environ. Microbiol 2004; 70:961-966.
12. Avrova АО, Venter E, Birch PR and Whisson SC. Profiling and quantifying differential gene transcription in Phytophthora infestans prior to and during the early stages of potato infection. Fungal Genetics & Biology 2003; 40:4-14.
13. Saville SP, Lazzell AL, Monteagudo С and Lopez-Ribot JL. Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans during infection. Eukaryot Cell 2003; 2:1053-1060.
14. Spellberg В, Ibrahim AS, Yeaman MR, et al. The antifungal vaccine derived from the recombinant N terminus of Als3p protects mice against the bacterium Staphylococcus aureus. Infect Immun. 2008; 76:4574-4580.
15. Fu Y, Ibrahim AS, Sheppard DC, Chen YC, French SW and Cutler JEea. Candida albicans Alsip: an adhesin that is a downstream effector of the EFG1 filamentation pathway. Mol Microbiol 2002; 44:61-72.
16. Ibrahim AS, Spellberg BJ, Avenissian V, Fu Y, Filler SG and Edwards JEJ. Vaccination with rAls1p-N improves survival during murine disseminated candidiasis by enhancing cell-mediated, not humoral, immunity. Infect Immun 2005; 73:999-1005.
17. Nobile CJ, Andes DR, Nett JE, et al. Critical role of Bcr1-dependent adhesins in С albicans biofilm formation in vitro and in vivo. PLoS Pathog. 2006; 2:e63.
18. Fradin C, De Groot P, MacCallum D, et al. Granulocytes govern the transcriptional response, morphology and proliferation of Candida abicans in human blood. Molecular Microbiology 2005; 56:397-415.
19. Castano I, Pan SJ, Zupancic M, Hennequin C, Dujon В and Cormack BP. Telomere length control and transcriptional regulation of subtelomeric adhesins in Candida glabrata. Mol Microbiol 2005; 55:1246-1258.
20. Schofield DA, Westwater C, Wamer T, Nicholas PJ, Paulling ЕЕ and Balish E. Hydrolytic gene expression during oroesophageal and gastric candidiasis in immunocompetent and immunodeficient gnotobiotic mice. J Infect Dis 2003; 188:591-599.
21. Kumamoto CA, Vinces MD. Contributions of hyphae and hypha-co-regulated genes to Candida albicans virulence. Cell Microbiol. 2005; 7:1546-1554.
22. Frohner IE, Bourgeois C, Yatsyk K, Majer O and Kuchler K. Candida albicans cell surface superoxide dismutases degrade host-derived reactive oxygen species to escape innate immune surveillance. Mol Microbiol 2009; 71:240-252.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ГРИБКОВЫХ И БАКТЕРИАЛЬНЫХ ПАТОГЕНОВ | 2014 |
|
RU2717306C2 |
| ПРИМЕНЕНИЕ ШТАММА ДРОЖЖЕЙ Komagataella pastoris В КАЧЕСТВЕ РЕЦИПИЕНТА ДЛЯ КОНСТРУИРОВАНИЯ ПРОДУЦЕНТОВ ЦЕЛЕВОГО БЕЛКА | 2013 |
|
RU2522479C1 |
| СПОСОБ ПОЛУЧЕНИЯ АЦЕТОИНА | 2015 |
|
RU2686614C2 |
| Способ диагностики инвазивного кандидоза и видовой идентификации его основных возбудителей методом ПЦР с гибридизационно-флуоресцентной детекцией в режиме реального времени | 2022 |
|
RU2809386C1 |
| СПОСОБ ЛЕЧЕНИЯ РЕЦИДИВИРУЮЩЕГО ВУЛЬВОВАГИНАЛЬНОГО КАНДИДОЗА | 2016 |
|
RU2642666C1 |
| Новые потенциальные противогрибковые средства на основе тиазолидин-2,4-диона и триазола | 2023 |
|
RU2814730C1 |
| ЛЕЧЕНИЕ ГРИБКОВОЙ ИНФЕКЦИИ ПРОТИВОГРИБКОВЫМИ ПРЕПАРАТАМИ ИЗ ГРУППЫ ПОЛИЕНОВ ИЛИ ИНГИБИТОРОВ β-ГЛЮКАНСИНТАЗЫ В КОМБИНАЦИИ С АНТИ-hsp90-АНТИТЕЛАМИ | 2001 |
|
RU2262952C2 |
| Lactobacillus fermentum Ess-1, DSM17851, И ЕГО ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ КАНДИДОЗА И ИНФЕКЦИЙ МОЧЕВЫХ ПУТЕЙ | 2006 |
|
RU2413761C1 |
| ШТАММ БАКТЕРИЙ Enterococcus faecium, ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ СНИЖАТЬ ОБРАЗОВАНИЕ БИОПЛЕНОК ГРИБАМИ РОДА Candida | 2015 |
|
RU2576008C2 |
| УКОРОЧЕННАЯ СЕКРЕТИРУЕМАЯ АСПАРТИЛ-ПРОТЕИНАЗА 2 | 2008 |
|
RU2493255C2 |






Изобретение относится к области биотехнологии, конкретно к рекомбинантным вакцинам против Candida albicans, и может быть использовано в медицине. Вакцина для лечения или предупреждения кандидозной инфекции содержит выделенный полипептид с SEQ ID NO: 2 в эффективном количестве, который может быть слит с партнером слияния. Вакцина также может содержать адъювант. Изобретение позволяет повысить эффективность лечения или предупреждения кандидозной инфекции. 2 н. и 9 з.п. ф-лы, 6 ил., 1 табл., 2 пр.
1. Вакцина для лечения или предупреждения кандидозной инфекции, содержащая выделенный полипептид в эффективном количестве, необязательно слитый с партнером слияния, и необязательно содержащая адъювант, отличающаяся тем, что аминокислотная последовательность указанного полипептида состоит из аминокислотной последовательности SEQ ID NO: 2.
2. Вакцина по п. 1, дополнительно содержащая адъювант.
3. Вакцина по п. 1, отличающаяся тем, что указанный полипептид слит с указанным партнером слияния.
4. Вакцина по п. 3, отличающаяся тем, что указанный партнер слияния содержит гетерологичную лидерную последовательность.
5. Вакцина по п. 3, отличающаяся тем, что указанный партнер слияния содержит метку или линкерную последовательность.
6. Вакцина по п. 5, отличающаяся тем, что указанная метка представляет собой гистидиновую метку.
7. Вакцина по п. 4 или 5, отличающаяся тем, что указанный полипептид получают из трансформированной клетки.
8. Вакцина по п. 7, отличающаяся тем, что указанная трансформированная клетка представляет собой трансформированную клетку Saccharomyces cerevisae.
9. Вакцина по п. 4 или 5, отличающаяся тем, что является вакциной для иммунизации против Candida albicans, и тем, что указанный полипептид очищен.
10. Способ лечения или предупреждения кандидозной инфекции у млекопитающих, включающий введение иммуногенного количества вакцины по п. 9.
11. Способ по п. 10, отличающийся тем, что указанная вакцина дополнительно содержит адъювант.
| US 7241613 B1, 10.07.2007 | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИМИКРОБНОГО ПЕПТИДА АРЕНИЦИНА | 2006 |
|
RU2316595C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ФУНГИЦИД, БАКТЕРИОСТАТИЧЕСКИЙ СУЛЬФОНАМИД И АНТИБАКТЕРИАЛЬНОЕ СОЕДИНЕНИЕ ДЛЯ МЕСТНОГО ПРИМЕНЕНИЯ | 2002 |
|
RU2291694C2 |
| BATES et al., Candida albicans lff11, a secreted protein required for cell wall structure and virulence | |||
| Infection & Immunity, 2007, v | |||
| Фальцовая черепица | 0 |
|
SU75A1 |
| Подаватель стеблей и кортикаторам и т.п. машинам | 1932 |
|
SU29922A1 |

