Применение штамма дрожжей Komagataella pastoris в качестве реципиента для конструирования продуцентов целевого белка
Изобретение относится к биотехнологии и микробиологической промышленности, в частности к получению штамма метилотрофных дрожжей Komagataella pastoris, пригодного для конструирования на его основе штаммов-продуцентов, способных синтезировать целевой белок.
В настоящее время множество применяемых в фармацевтической отрасли субстанций получают методом микробиологического синтеза.
Для осуществления экспрессии необходимо наличие следующей системы: разрешенного к применению одноклеточного или многоклеточного организма-реципиента, пригодного для трансформации вектором, включающим последовательность ДНК, кодирующую целевой белок, и вектора для трансформации, включающего все необходимые элементы (селективные маркеры, промоторы, терминаторы, и т.д.). Применяемые в настоящее время системы экспрессии можно подразделить на прокариотические (бактериальные) и эукариотические, представляющие собой клетки дрожжей, мицелярных грибов, насекомых и млекопитающих.
Каждая из систем имеет свои технологические и экономические достоинства и недостатки. Использование бактериальных систем экспрессии, например, наиболее часто используемой в биотехнологии системы экспрессии с использованием в качестве продуцента целевого белка рекомбинантного штамма бактерий E.coli, ограничивает спектр продуцируемых белков из-за невозможности воспроизвести все посттрансляционные модификации, характерные для эукариот, такие как гликозилирование, фосфорилирование, образование дисульфидных связей между цистеиновыми остатками, цис/транс-изомеризация пролина, и т.д. [1]. Также, при использовании бактерий E.coli в качестве продуцента целевого белка, часто сталкиваются со сложностями, связанными с рефолдингом (вследствие неправильного фолдинга в цитоплазме клетки и образования нерастворимых телец включения), что делает невозможным применение E.coli для биосинтеза многих белков [1]. Синтезированный клетками E.coli негликозилированный эритропоэтин (гликопротеин, стимулятор гемапоэза), стабильность и активность которого непосредственно зависит от гликозилирования, обладает значительно меньшей стабильностью по сравнению с эритропоэтином, секретируемым О-/N-гликозилированным клетками млекопитающих [2].
Использование для секреции целевых белков клеток млекопитающих, обеспечивающих все необходимые посттрансляционные модификации, приводит к увеличению стоимости продукции в связи с необходимостью использования сложного и дорогостоящего оборудования и питательных сред, для их культивирования [3]. Использование дрожжей в качестве продуцента целевых рекомбинантных белков является компромиссным и в некоторых случаях наиболее оптимальным решением. Дрожжи являются представителями эукариот и обладают способностью ко всем вышеперечисленным посттрансляционным модификациям, характерным для эукариотических клеток, включая также секрецию белков в культуральную среду. Выделение и очистка целевого рекомбинантного продукта существенно облегчена в связи с тем, что количество эндогенных секретируемых белков дрожжей сравнительно невелико [3].
Интерес к продукции рекомбинантных белков с использованием систем экспрессии, основанных на метилотрофных дрожжах, в частности, рода Pichia, Komagataella или Hansenula [6] основан на ряде преимуществ этих систем как с научной, так и с технологический точки зрения.
Культуру метилотрофных дрожжей выращивают в ферментерах до более высоких плотностей, по сравнению с традиционно-используемыми дрожжами Saccharomyces cerevisiae, что позволяет получить более высокий уровень продукции целевого белка [5]. Метилотрофные дрожжи обладают одними из самых мощных в природе промоторов. Их отличает также альтернативный механизм гликозилирования белков, позволяющий синтезировать низкоиммуногенный целевой белок.
Наиболее часто используемыми промоторами для регуляции экспрессии гетерологичных генов являются АОХ1 (промотор алкоголь оксидазы), GAP (промотор глицеральдегид 3-фосфат дегидрогеназы), FLD (промотор формальдегид дегидрогеназы), FDH (промотор формиат дегидрогеназы), DHAS (дигидроксиацетон синтазы) [9]. Известно, что промоторы АОХ1, FLD и FDH индуцируются в ответ на присутствие в питательной среде метанола или олеата. При росте культуры на глицерин- или глюкозо-содержащей среде, перечисленные промоторы репрессированы [11]. При истощении основного источника углерода промотор АОХ1 находится в дерепрессированном состоянии, при этом его активность составляет 1-2% от активности в индуцированном состоянии [12]. GAP считается конститутивным промотором, т.е. индуцирован при росте культуры на многих субстратах, включая глицерин, глюкозу.
Существенным преимуществом метилотрофных дрожжей является уровень и качество N-гликозилирования белков. Так, в клетках P.pastoris размер углеводных цепочек составляет в среднем 8-12 остатков маннозы, и концевым остатком является низкоиммуногенный α-1,2 маннозный остаток, тогда как сахаромицеты секретируют обычно гораздо более тяжело гликозилированный белок, содержащий до 100-150 остатков на одну цепочку, концевым остатком в которой оказывается высокоиммуногенный α-1,3 маннозный остаток.
С помощью экспрессионных систем на различных метилотрофных дрожжах, получено множество субстанций для фармацевтической промышленности: поверхностный антиген гепатита В (субстанция для вакцины), антистатин, эпидермальный ростовой фактор, гирудин, α1-урокиназа, эндотелиальный ростовой фактор и др. [3]. Первый разрешенный к использованию продукт получил регистрацию FDA в декабре 2009 года.
В качестве ближайшего аналога заявляемого изобретения рассмотрим штамм метилотрофных дрожжей Komagataella pfaffii NRRL Y-48124. Штамм GS115 (фенотип his-; Invitrogen, США), являющийся его производным, широко используют в качестве экспрессионной системы, в биотехнологии [4]. Экспрессионная система на основе этого штамма позволяет получать целевые белки с высоким выходом, при использовании индуцируемого метанолом промотора алкоголь оксидазы АОХ1, привлекательность которого заключается не только в его активности, но и в его строгой регуляции [12]. В отличие от некоторых сильных промоторов S.cerevisiae, которые имеют базальный уровень экспрессии на репрессирующих субстратах, промотор АОХ1 полностью репрессирован при росте на средах, содержащих такие репрессирующие субстраты, как глюкоза или глицерин или этанол [12]. Строгая регуляция промотора является важным фактором при экспрессии целевых генов, кодирующих токсичные для клетки белки.
При использовании штамма GS115 получены такие продукты как гранулоцит-колониестимулирующий фактор в количестве до 131 мг на 1 л культуральной жидкости [6] и рекомбинантный хепсидин в количестве до 100 мг на 1 л культуральной жидкости [7]. Недостатком данного штамма является способность экспрессировать ген целевого белка, находящийся под контролем промотора АОХ1, только в присутствии индуктора-метанола, который является токсичным и легко-воспламеняющимся веществом [21], [22].
Санитарно-гигиенические требования к организации и проведению работ с метанолом накладывают серьезные ограничения на использование данного вещества в биотехнологическом производстве [16]. В связи с этим, представляется важным найти решение, которое, с одной стороны, позволило бы использовать метилотрофные дрожжи для продукции целевого белка; а с другой стороны, позволило бы не использовать метанол в качестве индуктора для экспрессии генов этого белка.
Задача заявляемого изобретения - расширить арсенал метилотрофных дрожжей, пригодных для конструирования на их основе штаммов-продуцентов, способных экспрессировать целевой белок.
Задача решена путем применения штамма дрожжей Komagataella pastoris ВКПМ Y-727 в качестве реципиента для конструирования на его основе штаммов-продуцентов, способных экспрессировать целевой белок, как в присутствии индуктора-метанола, так и в его отсутствии.
Получение и таксономическая характеристика штамма К.pastoris ВКПМ Y-727
Отвечающий вышеупомянутым требованиям штамм найден путем скрининга во Всероссийской коллекции промышленных микроорганизмов (ВКПМ). Принадлежность штамма К.pastoris ВКПМ Y-727 к метилотрофным дрожжам рода Komagataella подтверждена при помощи ростовых тестов на соответствующих субстратах - метаноле, глицерине, олеате, и др. Кроме ростовых тестов, проведено молекулярно-генетическое сравнение по следующим маркерам: нуклеотидная последовательность домена D1/D2 26S субъединицы рибосомальной РНК, нуклеотидная последовательность промотора гена алкоголь оксидазы (АОХ1). Сравнение с соответствующими нуклеотидными последовательностями штамма К.pfaffii (NRRL Y-48124) и его производного GS115 (Invitrogen, США) показало гомологию на уровне 98%.
Морфологические признаки:
При культивировании при температуре 28°С в течение 48 часов на агаризованной среде YPD следующего состава (мас.%): пептон - 2, дрожжевой экстракт - 1, глюкоза - 2, агар - 2, вода - остальное, клетки имеют овальную форму, 3-4 мкм в диаметре. Клетки почкуются. Почкование истинное, многостороннее. Истинного мицелия не образуют.
Колонии имеют следующий вид:
1) на агаризованной среде YPD колонии светло-бежевого цвета с ровным краем, матовой поверхностью, линзовидным профилем и пастообразной консистенцией;
Рост в жидкой среде YPD состава (мас.%): пептон - 2, дрожжевой экстракт - 1, глюкоза - 2, вода - остальное, при 28°С в течение 24 ч культивирования - жидкость мутная, осадок белый, коагуляции не наблюдается, пристеночных пленок не образует.
Физиолого-биохимические признаки: факультативный анаэроб. Температура роста - 20-33°С (оптимум - 28°С). рН среды культивирования - 4,8-7,4 (оптимум - 6,0).
Ассимиляция источников углерода: утилизирует глюкозу, глицерин, метанол, олеат, сорбитол, рамнозу. Не утилизирует галактозу, ксилозу, арабинозу.
Ассимиляция источников азота: утилизирует аминокислоты, сернокислый аммоний, азотнокислый аммоний.
Хранение: при температуре -70°С в 30% водном растворе глицерина. Возможно хранение на агаризованной среде YPD в течение 3 месяцев при +4°С.
Регуляторные элементы
Секвенированы следующие последовательности генома штамма К.pastoris. ВКПМ Y-727:
Промотор алкоголь оксидазы АОХ1 штамма К.pastoris. ВКПМ Y-727 [SEQ ID N1] отличается от промотора штамма - ближайшего аналога следующими нуклеотидными заменами: G->A в положении 5, С->А в положении 9, Т->А в положении 11, A->G в положении 98, G->A в положении 131, делецией G в положении 132-133, делецией С в положении 170-171, С->Т в положении 201, G->A в положении 206, делецией СС в положении 311-312, Т->А в положении 354, A->G в положении 380, G-A в положении 482, С->Т в положении 568, Т->А в положении 582, А->Т в положении 611.
Промотор глицеральдегидфосфат дегидрогеназы GAP штамма К.pastoris. ВКПМ Y-727 [SEQ ID N2] отличается от промотора штамма - ближайшего аналога следующими нуклеотидными заменами: G->A в положении 61, С->Т в положении 86, А->С в положении 180, С->Т в положении 201, Т->С в положении 202, А в положении 209, A-G в положении 211, Т->С в положении 240, делецией СТ в положении 243-244, A->G в положении 275, G->А в положении 413, Т->С в положении 450.
Изобретение проиллюстрировано следующими фигурами.
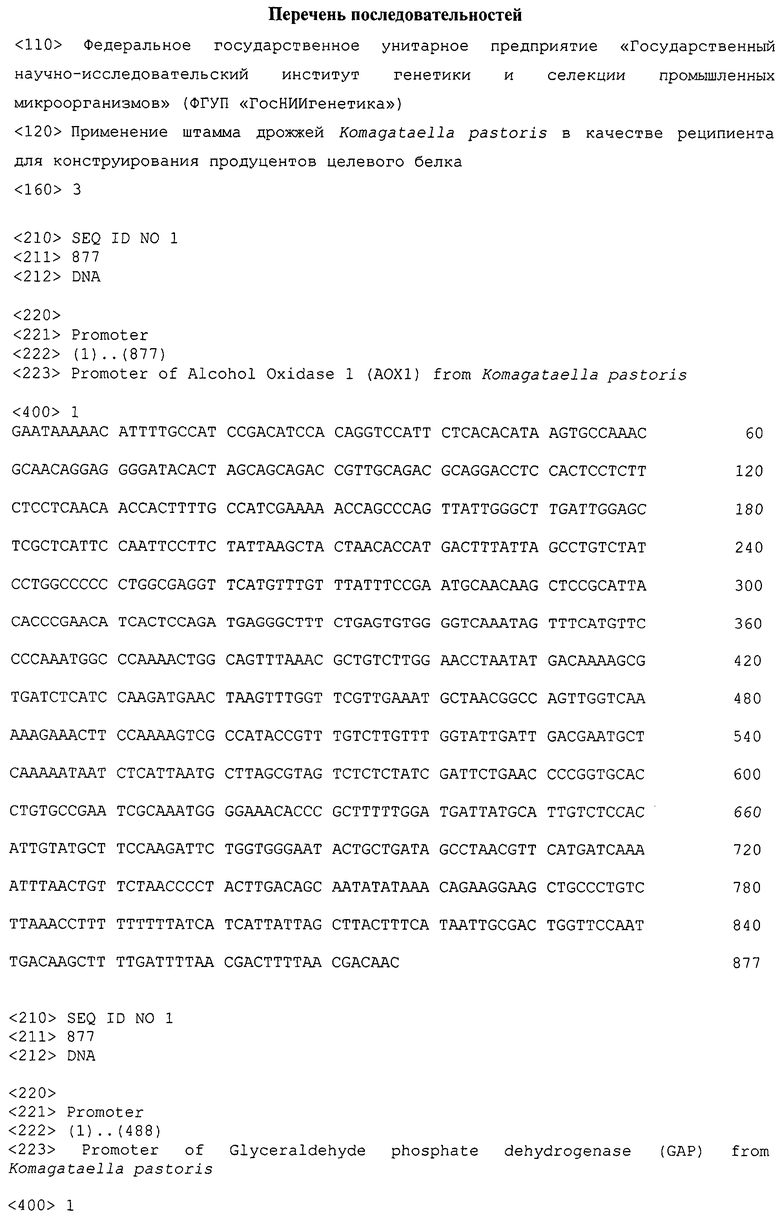
Фиг.1 Электрофореграмма клеточных лизатов штамма К.pastoris ВКПМ Y-727 и K.pfaffii GS115
Стрелками указан сигнал, которому соответствует фермент алкоголь оксидаза.
Дорожки с №№1-7 соответствуют
1. Молекулярный маркер, 70 кДа
2. Среда YPD, штамм К.pastoris. ВКПМ Y-727
3. Среда YPD, штамм K.pfaffii GS115
4. Среда YPM, штамм К.pastoris ВКПМ Y-727
5. Среда YPM, штамм K.pfaffii GS115
6. Среда YP, штамм К.pastoris. ВКПМ Y-727
7. Среда YP, штамм K.pfaffii GS115
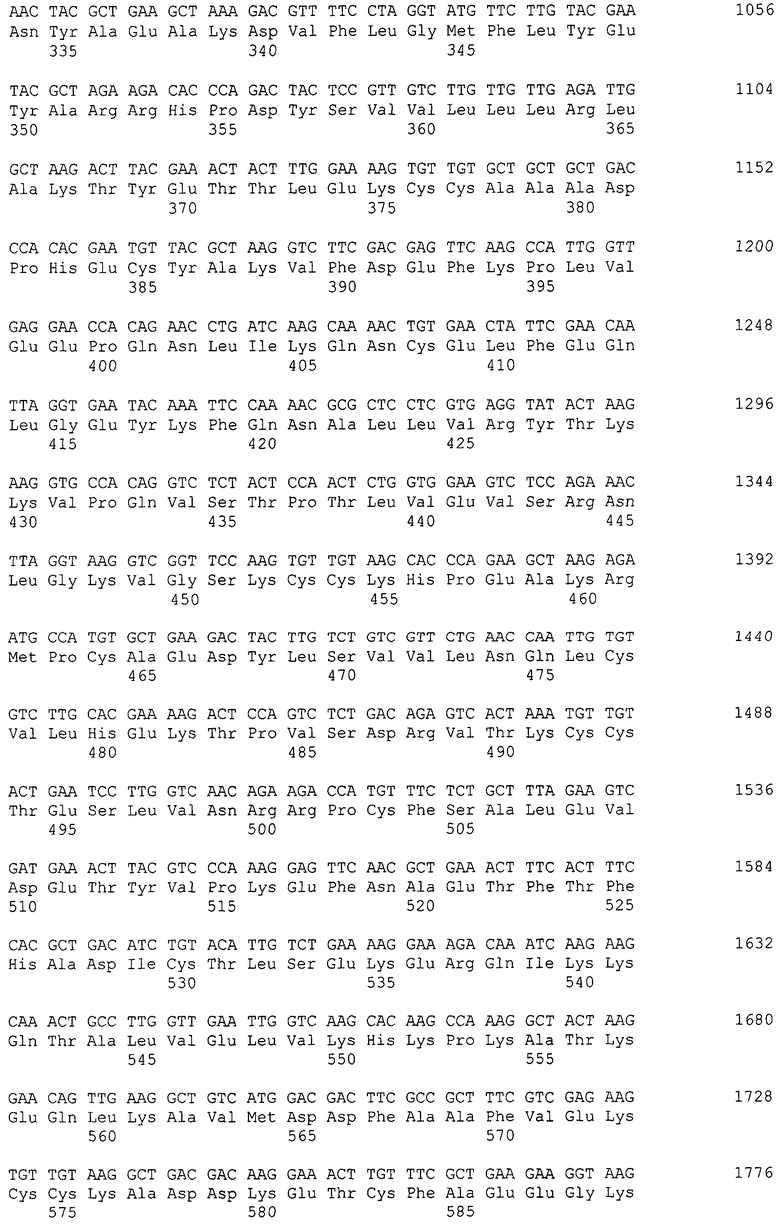
Фиг.2 Уровень активности бета-галактозидазы, продуцируемой клетками штамма К.pastoris. ВКПМ Y-727 и штамма K.pfaffii GS115
На диаграмме приведены измерения активности бета-галактозидазы при росте культур на трех различных средах: YP (имитирующая истощение источника углерода), YPM (содержащая метанол в качестве индуктора и источника углерода), YPD (содержащая глюкозу в качестве источника углерода, являющуюся репрессором промотора АОХ).
По вертикальной оси - шкала активности в Миллеровых единицах [Rose, Botstein et al.].
Темные столбцы соответствуют активности фермента в лизате клеток штамма K.pastoris ВКПМ Y-727, светлые столбцы соответствуют активности фермента в лизате клеток штамма K.pfaffii GS115.
На диаграмме планками отображено стандартное отклонение по результатам измерения активности у двух клонов.
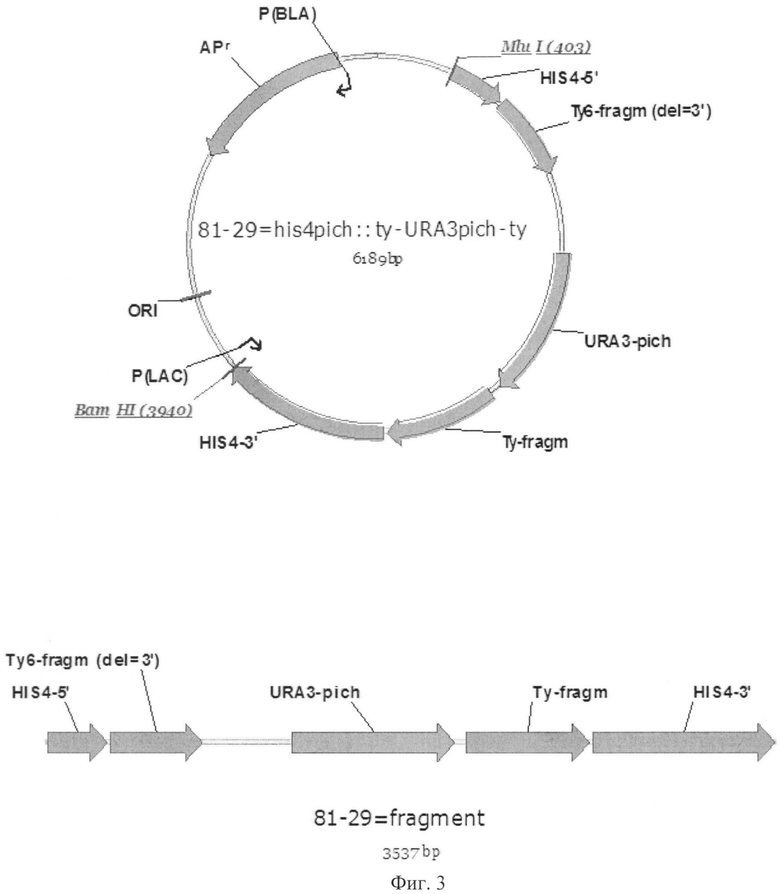
Фиг.3 Карта плазмиды 81-29. Линеаризованный фрагмент для трансформации
HIS4-5' - 5'-область гена HIS4
HIS4-3' - 3'-область гена HIS4
URA3-pich - ген URA3
ORI - бактериальный репликон
APR - ген бета-лактамазы
MluI, BamHI - сайты рестрикции соответствующих рестриктаз
Ту6-fragm - элементы, клонированные из области Ту6 ретротранспозона Ту1
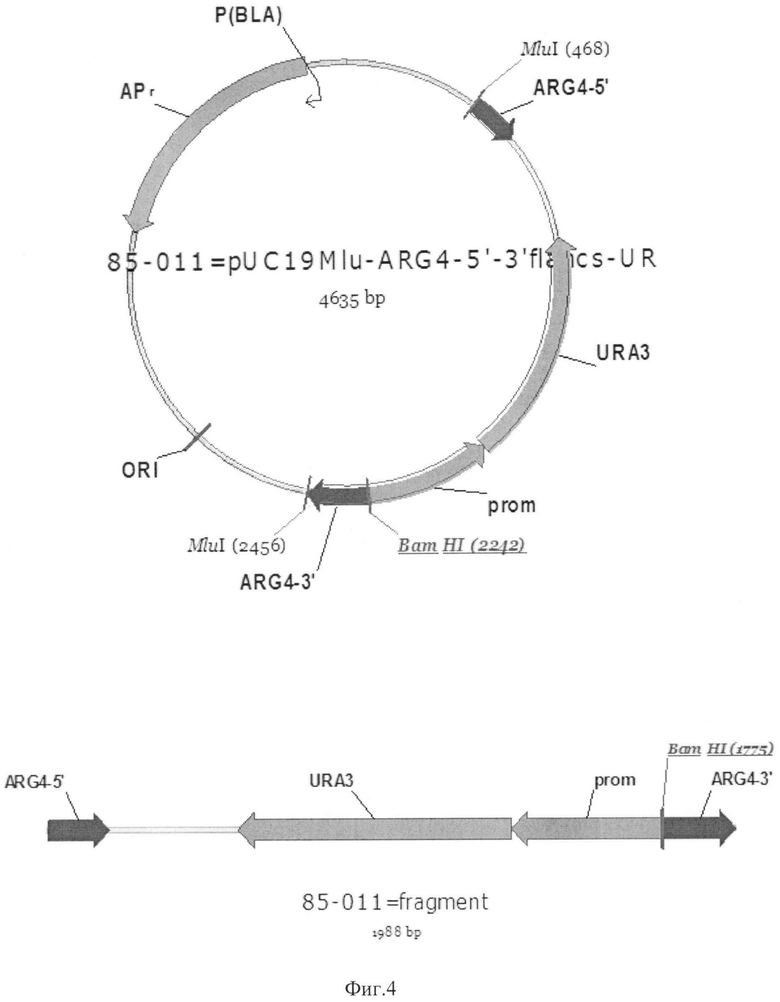
Фиг.4 Карта плазмиды 85-011. Линеаризованный фрагмент для трансформации
ARG4-5' - 5'-область гена ARG4
ARG4-3' - 3'-область гена ARG4
URA3-pich - ген URA3
Prom - регуляторный элемент гена URA3
ORI - бактериальный репликон
APR - ген бета-лактамазы
MluI, BamHI - сайты рестрикции соответствующих рестриктаз
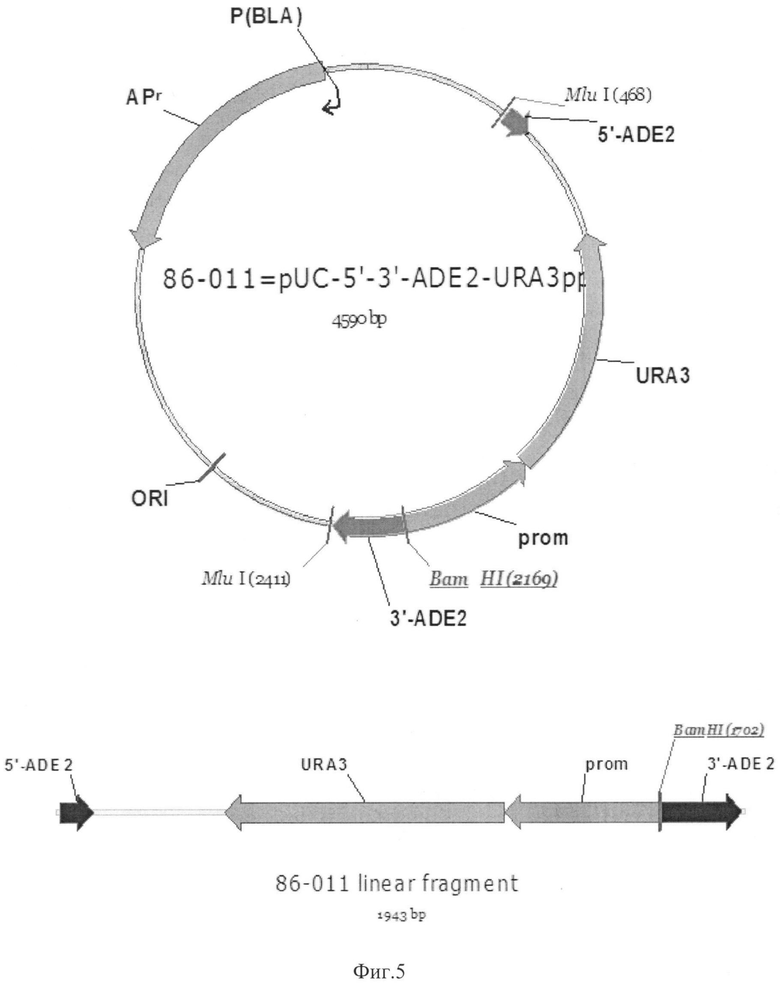
Фиг.5 Карта плазмиды 86-011. Линеаризованный фрагмент для трансформации
ADE2-5' - 5'-область гена ADE2
ADE2-3' - 3'-область гена ADE2
URA3-pich - ген URA3
Prom - регуляторный элемент гена URA3
ORI - бактериальный репликон
APR - ген бета-лактамазы
MluI, BamHI - сайты рестрикции соответствующих рестриктаз
Фиг.6а
Продукция сывороточного альбумина (HSA), нормированная на объем культуральной жидкости
По вертикальной оси - количество HSA в мг на 1 л культуральной жидкости.
Темный столбец соответствует количеству HSA в культуральной жидкости штамма К.pastoris ВКПМ Y-727. Светлый столбец соответствует количеству HSA в культуральной жидкости штамма K.pfaffii GS115. Планками отображено стандартное отклонение по измерениям у двух клонов.
ФИГ.6б
Продукция сывороточного альбумина (HSA), нормированная на единицу оптической плотности
По вертикальной оси - количество HSA в мг на единицу оптической плотности культуры (OD=1 при λ=600 нм).
Темный столбец соответствует количеству HSA в культуральной жидкости штамма К.pastoris ВКПМ Y-727. Светлый столбец соответствует количеству HSA в культуральной жидкости штамма K.pfaffii GS115. Планками отображено стандартное отклонение по измерениям у двух клонов.
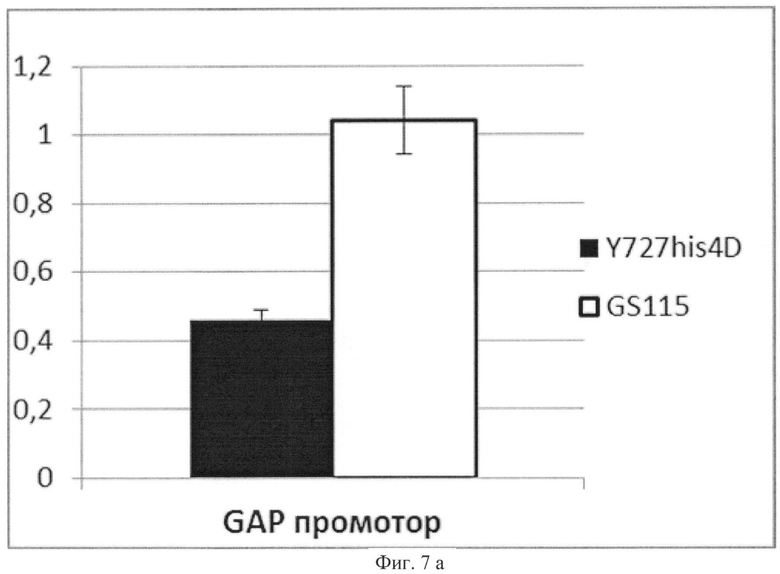
Фиг.7а
Активность инулазы, продуцируемой сравниваемыми штаммами под контролем промотора GAP
По вертикальной оси - активность инулазы в единицах активности на 1 мл культуральной жидкости на единицу оптической плотности культуры (OD=1 при λ=600 нм).
Темный столбец соответствует активности инулазы в культуральной жидкости штамма K.pastoris ВКПМ Y-727. Светлый столбец соответствует активности инулазы в культуральной жидкости штамма K.pfaffii GS115. Планками отображено стандартное отклонение по измерениям у двух клонов.
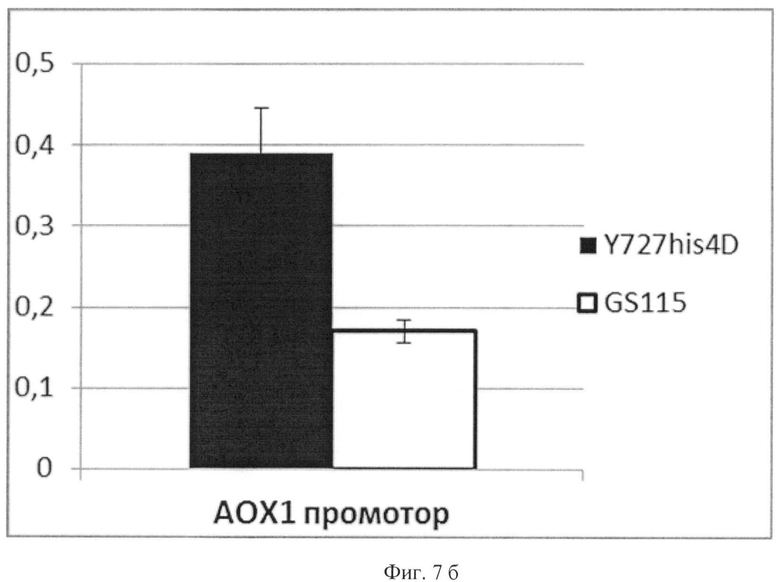
Фиг.7б
Активность инулазы, продуцируемой сравниваемыми штаммами под контролем промотора АОХ1
По вертикальной оси - активность инулазы в единицах активности на 1 мл культуральной жидкости на единицу оптической плотности культуры (OD=1 при λ=600 нм).
Темный столбец соответствует активности инулазы в культуральной жидкости штамма К.pastoris ВКПМ Y-727. Светлый столбец соответствует активности инулазы в культуральной жидкости штамма K.pfaffii GS115. Планками отображено стандартное отклонение по измерениям у двух клонов.
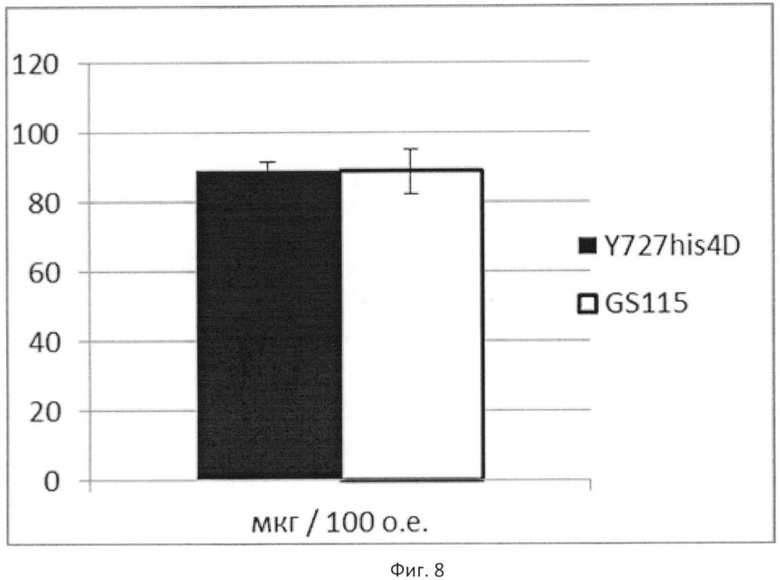
Фиг.8
Продукция поверхностного антигена гепатита В (HBsAg)
По вертикальной оси - количество HBsAg в мкг на 100 единиц оптической плотности культуры (при λ=600 нм).
Темный столбец соответствует количеству HBsAg в лизате клеток штамма К.pastoris ВКПМ Y-727. Светлый столбец соответствует количеству HBsAg в лизате клеток штамма K.pfaffii GS115. Планками отображено стандартное отклонение по измерениям у двух клонов.
Пример 1. Подтверждение автоиндукции промотора АОХ1 у штамма К.pastoris ВКПМ Y-727
Для подтверждения индукции промотора АОХ1 у штамма К.pastoris ВКПМ Y-727 без использования метанола, в условиях истощения углеродного субстрата, проведен эксперимент; в качестве контрольного штамма использован штамм К.pfaffii GS115.
Анализируемые штаммы инокулированы в пробирки со средой трех видов YPD - с репрессирующим промотор АОХ1 источником углерода - глюкозой, YPM следующего состава (мас.%) (пептон - 2, дрожжевой экстракт - 1, метанол - 0,5, вода - остальное) - с индуцирующим АОХ1 источником углерода - метанолом, YP (мас.%) (пептон - 2, дрожжевой экстракт - 1, вода - остальное) - без источника углерода. В логарифмической фазе роста образцы клеточного лизата культуры из каждой пробирки проанализированы с помощью электрофореза в денатурирующих восстанавливающих условиях в 12% полиакриламидном геле [Laemmli., 1970]. Результат представлен на электрофореграмме (фиг.1).
Из данных, представленных на электрофореграмме, по интенсивности сигнала на уровне 70 кДа, что соответствует молекулярной массе алкоголь оксидазы [13], можно судить об активности промотора алкоголь оксидазы в соответствующих условиях культивирования клеток. На Фиг.1 видно, что ген АОХ1 не экспрессируется при росте на среде с глюкозой (дорожки №2, 3), и экспрессируется при росте на среде с метанолом у обоих штаммов (дорожки №4, 5), а также экспрессируется у штамма К.pastoris ВКПМ Y-727 при росте на среде YP, то есть в условиях отсутствия углеродного субстрата. В последнем случае, уровень экспрессии составляет 60-70% от уровня экспрессии на среде с метанолом (в индуцированном состоянии), что подтверждает активацию промотора АОХ1 у штамма К.pastoris ВКПМ Y-727 в условиях роста, имитирующих истощение углеродного субстрата, без использования метанола.
Пример 2. Сравнение силы промоторов АОХ1 из штамма К.pastoris ВКПМ Y-727 и из контрольного штамма K.pfaffii GS115
Каждый из промоторов АОХ1 из штаммов К.pastoris ВКПМ Y-727 и К.pfaffii GS115 независимо друг от друга клонирован в плазмиду, содержащую ген Lac-Z (бета-галактозидазы) для оценки силы промотора АОХ1 каждого из анализируемых штаммов на основании активности продуцируемого сконструированными штаммами фермента бета-галактозидазы.
Для этого сконструированы штаммы К.pastoris Y-727/pINT-G418-AOX1Y727-LacZ и K.pfaffii GS115/pINT-G418-AOX1GS115-LacZ - продуценты бета-галактозидазы.
Конструирование плазмид
Сконструированы две плазмиды: плазмида pINT-G418-AOX1Y727-LacZ, предназначенная для интеграции в геном штамма К.pastoris ВКПМ Y-727, и плазмида pINT-G418-AOX1GS115-LacZ, предназначенная для интеграции в геном штамма K.pfaffii GS115. Обе плазмиды содержат элементы, необходимые для их амплификации в клетках E.coli: ген бета-лактамазы (в качестве селективного маркера), бактериальный репликон для обеспечения поддержания определенного уровня копийности плазмиды. Для проведения последующей процедуры трансформации дрожжевых клеток и экспрессии целевого гена Lac-Z (бета-галактозидазы), вектора содержат следующие элементы: ген neo - аминогликозид 3'-фосфотрансферазы, ген устойчивости к антибиотику G418 и канамицину [18], находящийся под контролем промотора ADH1 - в качестве селективного маркера (промотор ADH1 получили с помощью ПЦР, в качестве матрицы использовали геномную ДНК штамма S.cerevisiae YBS618, праймеры prADH1-5'-aaagatctccatccttttgttgtttcc, prADH1-3'-cttgattgtatatgagata); ген Lac-Z [17], находящийся под контролем промотора АОХ1727 (для штамма Y727) либо AOXGS115 (штамма GS115), клонированного из штамма К.pastoris ВКПМ Y-727, либо из штамма K.pfaffii GS115, соответственно, терминатор АОХ1. Промотор АОХ1727 и AOXGS115 получали при помощи ПЦР, в качестве матрицы использовали геномную ДНК штамма К.pastoris ВКПМ Y-727 либо K.pfaffii GS115, соответственно. Праймеры pr1_AOX1 (5'-ttcgtcgactaacatccaaagacgaaaggtt), pr2_AOX1 (5'-aaggtaccagatctagccatggtttggatccttcgaataattagttgttttttgatcttctcaa). Терминатор АОХ1 получали при помощи ПЦР, в качестве матрицы использовали геномную ДНК штамма К.pastoris ВКПМ Y-727 либо K.pfaffii GS115, соответственно, праймеры prAOXter-5' aatctcgaggattccagaatgccatttgcct, prAOXter3' gttgacgcgtgcacaaacgaacgtctcactt. Интегрируемая часть плазмиды фланкирована областями для встраивания в определенный локус генома - эндогенный промотор АОХ1. Для получения линеаризованных частей плазмид, предназначенных для процесса трансформации, исходные плазмиды pINT-С418-AOX1Y727-LacZ и pINT-G418-AOX1GS115-LacZ обработаны эндонуклеазой рестрикции MluI (Thermo Fisher Scientific Inc.).
Трансформация и культивирование
В результате проведения процесса трансформации по стандартному протоколу [16] и отбора трансформантов получили 2 штамма продуцента бета-галактозидазы: К.pastoris Y-727/pINT-G418-AOX1Y727-LacZ и контрольный K.pfaffii GS115/pINT-G418-AOX1GS115-LacZ, в которых ген Lac-Z (бета-галактозидазы) находится под контролем промоторов АОХ1, клонированных из соответствующих сравниваемых штаммов. Затем выбрали по 2 клона каждого штамма. Инокулят анализируемых клонов поместили в пробирки с 5 мл среды трех видов YPD с репрессирующим АОХ1 источником углерода - глюкозой, YPM - с индуцирующим АОХ1 источником углерода - метанолом, YP - без источника углерода. Культуру выращивали в ротационном шейкере-термостате (250 об/мин), при температуре 28°С. По истечении 14 часов клетки отделяли центрифугированием в настольной центрифуге, при 3000 g и разрушали с помощью стеклянных шариков для получения клеточного лизата [Miller et al.].
Оценка уровня активности бета-галактозидазы
Оценку уровня активности бета-галактозидазы, содержащейся в клеточном лизате анализируемых штаммов, осуществляли известным методом [Rose, Botstein et al.]. Из приведенных в таблице 1 и на фиг.2 данных видно, что активность бета-галактозидазы из клеточного лизата культуры штамма К.pastoris Y-727/pINT-G418-AOX1Y727-LacZ, выращенной на среде YP (без углеродного субстрата и индуктора-метанола), составляет около 70% активности бета-галактозидазы из лизата этой же культуры, выращенной на среде YPM (содержащей метанол). Это позволяет сделать вывод о том, что промотор АОХ1 в клетках К.pastoris Y-727 активирован как в присутствии индуктора-метанола, так и в его отсутствии, в отличие от экспрессионной системы контрольного штамма - ближайшего аналога K.pfaffii GS115. Уровень синтеза бета-галактозидазы у обоих штаммов в присутствии метанола отличается незначительно в пользу штамма K.pastoris ВКПМ Y-727.
Пример 3. Получение штамма К.pastoris Y727ura3mut
Конструирование плазмиды pURA3del
Вектор содержит элементы, необходимые для его амплификации в клетках E.coli: ген бета-лактамазы (в качестве селективного маркера), бактериальный репликон для обеспечения поддержания определенного уровня копийности плазмиды. Ген URA3 получили при помощи ПЦР, используя в качестве матрицы геномную ДНК данного штамма, и праймеры pr1_URA3 (5'-gataacgcgtttgacgaattgactaaagttct), pr2_URA3 (5'-gataacgcgtttgacgaattgactaaagttct). Полученный с помощью ПЦР ген клонировали в плазмиду. Далее в полученной плазмиде была искусственно нарушена рамка считывания гена URA3. После обработки плазмиды рестриктазой MluI получали фрагмент, содержащий фланги гена URA3, так, что при интеграции данного фрагмента в геном трансформируемого штамма инактивировался нативный ген URA3, что приводило к мутации штамма по синтезу урацила.
Трансформация и отбор трансформантов
Исходный штамм К.pastoris ВКПМ Y-727 трансформировали полученным фрагментом плазмиды pURA3del с целью получения мутантных клонов по гену URA3, кодирующего оротидин-5-фосфат декарбоксилазу, фермент, участвующий в синтезе de novo пиримидиновых оснований. Отбор мутантных клонов проводили на селективных чашках YPD, содержащих 5-фтороротидиновую кислоту (5'-FOA), в концентрации 500 мкг/мл агаризованной среды. В результате селекции было отобрано 5 клонов, обладавших способностью расти в присутствии данной концентрации 5-фтороротидиновой кислоты. Способность роста в присутствии данного субстрата связана с вероятной мутацией в гене URA3, продукт которого - оротидин-5-фосфат декарбоксилаза, не обладает активностью и не расщепляет субстрат (5'-FOA) с освобождением токсичного для клетки фтора. Полученные мутанты обладали фенотипом ura-. Отобранные клоны были проверены на способность роста на минимальной агаризованной среде YNB состава (мас.%) (0,17 yeast nitrogen base (Difco), 0,5 сульфат аммония, 0,4 глюкоза, 2 агароза, остальное - вода) с добавлением урацила в концентрации 50 мкг/мл среды. По всем морфологическим признакам полученные мутанты не отличались от клонов исходного штамма.
Пример 4. Получение штамма К.pastoris Y-727ura3muthis4Δ
Конструирование плазмиды 81-29
Вектор 81-29 содержит элементы, необходимые для его амплификации в клетках E.coli: ген бета-лактамазы (в качестве селективного маркера), бактериальный репликон для обеспечения поддержания определенного уровня копийности плазмиды. Ген HIS4 амплифицировали при помощи ПЦР. В качестве матрицы использовали геномную ДНК из штамма К.pastoris ВКПМ Y-727, праймеры: pr1_HIS4 (5'-ttcactcgtggatctataattgaacatgacatttcccttgctacct); pr2_HIS4 (5'-agatgccggttagatctatcgaat). Полученный с помощью ПЦР ген клонировали в исходную плазмиду. Ген URA3 получили при помощи ПЦР, используя в качестве матрицы геномную ДНК данного штамма, и праймеры pr1_URA3 (5'-gataacgcgtttgacgaattgactaaagttct), pr2_URA3 (5'-gataacgcgtttgacgaattgactaaagttct). Экспрессия клонированного в плазмиду гена URA3 находится под контролем эндогенного промотора URA3. Ген URA3 с промотором фланкирован последовательностями из области, кодирующей белок ТуА ретротранспозона Ту1, клонированными из дрожжей S.cerevisiae [19]. Полученную конструкцию вставили в середину гена HIS4 (уже клонированного в плазмиду), так, что она оказалась в окружении 5' и 3' областей гена HIS4. Данная конструкция направляет интеграцию в локус HIS4, после обработки плазмиды эндонуклеазами рестрикции MluI, BamHI (Thermo Fisher Scientific Inc.) см. (Фиг.2). Таким образом, при трансформации ауксотрофного штамма К.pastoris Y-727ura3mut линеаризованным фрагментом вектора 81-29, в результате гомологичной рекомбинации между эндогенным локусом HIS4 и данным фрагментом, фланкированным 5'- и 3'-областями гена HIS4, происходит встраивание линеаризованного фрагмента плазмиды с замещением нативного гена HIS4.
В результате получен линеаризованный фрагмент вектора 81-29, содержащий ген URA3, фланкированный последовательностями из ретротранспозона Ту1, которые в свою очередь фланкированы 5' и 3' областями гена HIS4, клонированного из штамма К.pastoris ВКПМ Y-727. Использование областей из ретротранспозона Ту1 обеспечивает выщепление нуклеотидной последовательности, заключенной между ними, что и будет использовано после трансформации.
Трансформация штамма К.pastoris Y-727ura3mut
Полученный ауксотрофный штамм К.pastoris Y727ura3mut (пример 3) трансформировали линеаризованным фрагментом вектора 81-29. В результате гомологичной рекомбинации при трансформации происходит делеция гена HIS4. Геномы полученных трансформантов содержат фрагмент вектора 81-29 с геном URA3, вместо замещенного гена HIS4. Отбор трансформантов производили на чашках с минимальной агаризованной средой YNB с гистидином, в концентрации 50 мкг/мл среды. Несколько отобранных трансформантов инокулировали в 5 мл богатой среды YPD. Культура росла в течение 24 часов, после чего данную культуру использовали в качестве инокулята для нового засева в среду YPD. После трех пересевов часть клеток отделяли центрифугированием для засева на чашки с агаризованной средой YPD, содержащей 5-фтороротовую кислоту в концентрации 500 мкг/мл среды для отбора тех клонов, у которых произошла гомологичная рекомбинация между встроившимися в локус HIS4 фрагментами Ту6, приводившая к делегированию гена URA3, заключенного между фрагментами Ту6 (Фиг.3). В результате рекомбинации отобран клон с фенотипом his-ura-, не способный к росту на среде без гистидина и урацила. Подтверждена способность этого клона к росту на минимальной агаризованной среде YNB, содержащей урацил в концентрации 50 мкг/мл среды и гистидин в концентрации 50 мкг/мл.
Один из этих клонов обозначен как штамм К.pastoris Y727ura3muthis4Δ. Полученный ауксотрофный по гистидину и урацилу штамм использован для трансформации векторами, содержащими селективные маркеры URA3 и HIS4, с последующим отбором трансформантов на соответствующих селективных средах.
Пример 5. Получение штамма К.pastoris Y-727his4Δ
Полученный ауксотрофный штамм К.pastoris s Y-727ura3muthis4Δ (пример 4) трансформировали продуктом ПЦР, содержащим ген URA3. Для ПЦР использовали праймеры Pr1_URA3 (5'-gataacgcgtttgacgaattgactaaagttct) и Pr2_URA3 (5'-atttacgcgtactccttgagtctggtcaaa). В качестве матрицы для ПЦР использовали геномную ДНК из штамма К.pastoris ВКПМ Y-727. Отбор трансформантов производили на агаризованной среде YNB с гистидином, в концентрации 50 мкг/мл среды. В результате трансформации получили клоны с фенотипом his-. Отобранные трансформанты проверили на способность роста на минимальной среде YNB с добавлением гистидина в концентрации 50 мкг/мл среды.
В результате получен штамм К.pastoris Y-727his4Δ. Полученный ауксотрофный по гистидину штамм использовался для трансформации векторами, содержащими селективный маркер HIS4, с последующим отбором трансформантов на соответствующей селективной среде.
Пример 6. Получение штамма К.pastoris Y-727arg4Δhis4Δ
Конструирование плазмиды 85-011
Вектор 85-011 содержит элементы, необходимые для его амплификации в клетках E.coli: ген бета-лактамазы (в качестве селективного маркера), бактериальный репликон для обеспечения поддержания определенного уровня копийности плазмиды. Данный вектор содержит ген URA3, находящийся под контролем нативного промотора, и фланкированный 5' и 3' областями гена ARG4, использованными для направления интеграции данного фрагмента непосредственно в локус ARG4. 5' и 3' области гена ARG4 получали при помощи ПЦР. В качестве матрицы использовали геномную ДНК штамма K.pastoris ВКПМ Y-727, праймеры:
pr1_ARG4 (5'-tctaacgcgtgtcgactaggtgttttaccctgattaga),
pr2_ARG4 (5'-ggatcctatctcgagtccggagatgctagcactaagatagctggtaataagtttaga),
pr3_ARG4 (5'-tagtgctagcatctccggactcgagataggatccatccattgactcccgttttga),
pr4_ARG4 (5'-gataacgcgtagatctcaattgtatgtactataacagttt).
Трансформация и отбор трансформантов
Штамм К.pastoris Y727ura3muthis4Δ (пример 4) трансформировали линеаризованным фрагментом вектора 85-011 (Фиг.4). Фрагмент получили в результате обработки вектора 85-011 эндонуклеазой рестрикции MluI (Фиг.4). В результате гомологичной рекомбинации при трансформации происходит встраивание фрагмента вектора 85-011 в локус ARG4 с замещением гена ARG4 (аргининсукцинатлиазы), трансформанты обладают фенотипом ura+his-arg-. Отбор трансформантов проводили на минимальной агаризованной среде YNB, содержащей гистидин в концентрации 50 мкг/мл и аргинин в концентрации 50 мкг/мл среды. Отобранные трансформанты обладали фенотипом arg-his-.
Полученный ауксотрофный по гистидину и аргинину штамм К.pastoris Y-727arg4Δhis4Δ пригоден для получения продуцентов путем трансформации векторами, содержащими селективный маркер HIS4 или ARG4.
Пример 7. Получение штамма Д. pastoris Y-727ade2Δhis4Δ
Конструирование плазмиды 86-011
Вектор 86-011 содержит элементы, необходимые для его амплификации в клетках E.coli: ген бета-лактамазы (в качестве селективного маркера), бактериальный репликон для обеспечения поддержания определенного уровня копийности плазмиды. Данный вектор содержит ген URA3, находящийся под контролем нативного промотора - фланкирован 5' и 3' областями гена ADE2 (фермента фосфорибозиламиноимидазол карбоксилазы), использованными для направления интеграции данного фрагмента непосредственно в локус ADE2. 5' и 3' области гена ADE2 получали при помощи ПЦР. В качестве матрицы использовали геномную ДНК штамма Komagataella pastoris Y727, праймеры: pr1_ADE2 (5'-tactaacgcgtgtcgacgaaagtaggcaaattagtttgt), pr2_ADE2 (5'-ctcgagtatggatcctccggagatcccgggacatgtgagctttgaaaatatctaatcgt), pr3_ADE2 (5'-acatgtcccgggatctccggaggatccatactcgagtcatcggtgttcctgtcaag), pr4_ADE2 (5'-agtaacgcgtagatcttgaaatagaaatatgattagaaaaaa).
Штамм К.pastoris Y-727ura3muthis4Δ (пример 4) трансформировали линеаризованным фрагментом вектора 86-011 (Фиг.5), который получили в результате обработки вектора 86-011 эндонуклеазой рестрикции MluI (Фиг.5). В результате гомологичной рекомбинации после трансформации происходит встраивание фрагмента вектора 86-011 в локус ADE2 с замещением гена ADE2 (фосфорибозиламиноимидазол карбоксилаза), а полученные трансформанты имеют фенотип ura+his-ade-. Отбор трансформантов проводили на минимальной агаризованной среде YNN, содержащей гистидин в концентрации 50 мкг/мл и аденин в концентрации 50 мкг/мл среды. Отобранные трансформанты обладали фенотипом ade-his-. Полученный ауксотрофный по гистидину и аденину штамм использовался для трансформацией векторами, содержащими селективный маркер HIS4 или ADE2, с последующим отбором трансформантов на соответствующей селективной среде.
Полученный ауксотрофный по гистидину и аденину штамм К.pastoris Y-727arg4Δhis4Δ пригоден для получения продуцентов путем трансформации векторами, содержащими селективный маркер HIS4 или ADE2.
Пример 8. Получение штамма К.pastoris Y727his4Δ/pPH93-AOX1Y727-HSA - продуцента человеческого сывороточного альбумина
Конструирование плазмиды
Экспрессионный интеграционный вектор рРН93-AOX1Y727-HSA содержит элементы, необходимые для его амплификации в клетках Е.coli: ген бета-лактамазы (в качестве селективного маркера), бактериальный репликон для обеспечения поддержания определенного уровня копийности плазмиды. Для трансформации дрожжевых клеток и экспрессии целевого гена, вектор содержит следующие элементы: ген HIS4 - гистидинол дегидрогеназы, клонированного из К.pastoris ВКПМ Y-727, в качестве селективного маркера, ген целевого белка - альбумина (HSA) [SEQ ID N3], находящийся под контролем промотора AOX1Y727 [SEQ ID N1], клонированного из К.pastoris ВКПМ Y-727, терминатор АОХ1, клонированный из К.pastoris ВКПМ Y-727. Интегрируемая часть плазмиды фланкирована областями для встраивания в определенный локус генома - эндогенный промотор АОХ1. Для получения линеаризованной части плазмиды, предназначенной для трансформации дрожжей К.pastoris ВКПМ Y-727, исходную плазмиду рРН93-AOX1Y727-HSA обрабатывают эндонуклеазой рестрикции MluI.
Экспрессионный интегративный вектор рРН93-AOX1GS115-HSA содержит те же элементы, что и pPH93-AOX1Y727-HSA, за исключением гена HIS4 - гистидинол дегидрогеназы, клонированного из К.pfaffii GS115 (в качестве матрицы для ПЦР использовали геномную ДНК штамма К.pfaffii GS115, праймеры pr1_HIS4, pr2_HIS4), и промотора AOX1GS115, клонированного также из К.pfaffii GS115. Для получения линеаризованной части плазмиды, предназначенной для трансформации дрожжей К.pfaffii GS115, исходную плазмиду pPH93-AOX1GS115-HSA обрабатывают эндонуклеазой рестрикции MluI.
Трансформация
Получение продуцентов секретируемого в культуральную среда белка (человеческого сывороточного альбумина, HSA) с использованием штамма К.pastoris ВКПМ Y-727 и в качестве контрольного штамма К.pfaffii GS115. В качестве реципиентных штаммов используют штамм К.pastoris Y-727his4Δ и штамм К.pfaffii GS115. В результате трансформации данных штаммов линеаризованными фрагментами плазмид рРН93-AOX1Y727-HSA и pPH93-AOX1GS115-HSA, соответственно, с последующим отбором трансформантов на селективной агаризованной среде YNB без аминокислот и сульфата аммония, получают штаммы, продуцирующие рекомбинантный сывороточный альбумин.
В результате получили следующие штаммы продуценты сывороточного альбумина:
К.pastoris Y727his4Δ/pPH93-AOX1Y727-HSA
К.pfaffii GS115/pPH93-AOX1GS115-HSA
Культивирование
Для проведения опыта выбрали по 2 клона каждого штамма. Инокулят анализируемых клонов был помещен в пробирки с 5 мл среды YPgM, следующего состава (мас.%): пептон - 2, дрожжевой экстракт - 1, глицерин 0,5, метанол - 0,5, вода - остальное. Культуру выращивали в течение 62 часов в ротационном шейкере-термостате (250 об/мин), при температуре 28°С. Каждые 24 часа проводили индукцию метанолом, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%. По истечении 62 часов биомассу отделяли центрифугированием в настольной центрифуге, при 3000 g.
Измерения концентрации альбумина в культуральной жидкости проводили методом иммуноферментного анализа на твердой подложке. В качестве подложки использовали стандартный планшет для иммуноферментного анализа с предварительно сорбированными человеческими поликлональными антителами к HSA. Данные антитела, конъюгат - человеческие поликлональные антитела к HSA, меченные пероксидазой хрена, стандарт для калибровки, а также хромоген ТМБ (3,3',5,5'-тетраметилбензидин) входили в стандартный набор Albumin, Human, BioAssay ELISA Kit (производства United States Biological, США). Анализ проводили по протоколу, описанному в инструкции к набору.
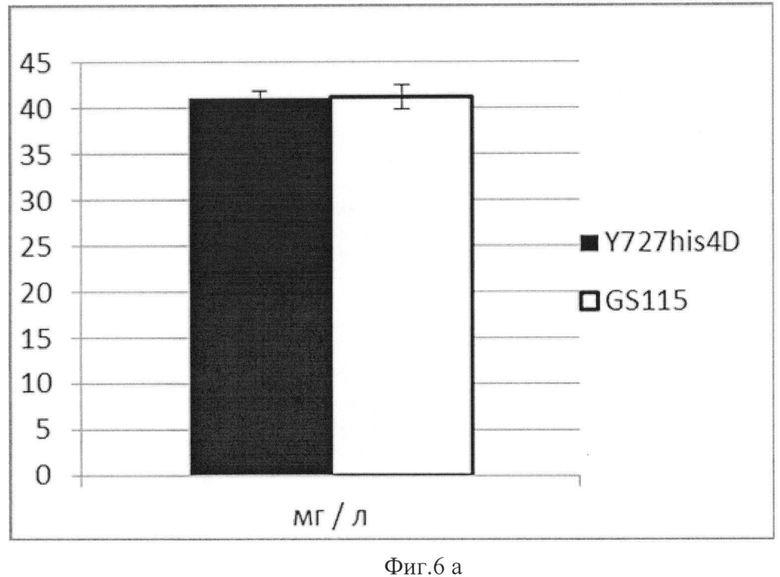
Выход целевого белка составил 41,12 (+/-0,8) мг/л культуральной жидкости или 1,93 (+/-0,2) мг на 1 оптическую единицу) для штамма К.pastoris Y-727his4Δ и 41,19 (+/-1,4) мг/л культуральной жидкости или 1,38 (+/-0,1) мг на 1 оптическую единицу для контрольного штамма K.pfaffii GS115.
Данные, представленные в таблице 2 и на Фиг.6 (а, б), свидетельствуют о том, что при нормировании на объем культуральной жидкости штамм продуцент К.pastoris Y727his4Δ/pPH93-AOX1Y727-HSA не уступает продуценту К.pfaffii GS115/рРН93-AOX1GS115-HSA, а при нормировании на оптические единицы даже превосходит его по продуктивности.
Пример 9. Получение продуцента инулазы
Инулаза - фермент, используемый в пищевой промышленности для получения фруктозы, путем расщепления β-гликозидных связей инулина (полифруктозного углевода растительного происхождения).
Конструирование плазмиды
Было сконструировано 4 экспрессионных интеграционных вектора, отличающихся способом регуляции экспрессии гена целевого белка - инулазы. Экспрессионный вектор pINT-HIS4-GAPY727-INU содержит элементы, необходимые для его амплификации в клетках E.coli: ген бета-лактамазы (в качестве селективного маркера), бактериальный репликон для обеспечения поддержания определенного уровня копийности плазмиды. Для трансформации дрожжевых клеток и экспрессии целевого гена, вектор содержит следующие элементы: ген HIS4 - гистидинол дегидрогеназы, в качестве селективного маркера, клонированный из К.pastoris ВКПМ Y727, ген целевого белка - инулазы, клонированный из Kluyveromyces marxianus [14], находящийся под контролем промотора GAPY727, клонированного из К.pastoris ВКПМ Y-727, терминатор АОХ1. Промотор GAP получили с помощью ПЦР, в качестве матрицы использовали геномную ДНК штамма K.pastoris ВКПМ Y-727, праймеры pr1_GAP (5'-aggaagcttttttgtagaaatgtcttggt), pr2_GAP (5'-ataggtacctgcagccatggtagatctttgatagttgttcaattgattgaaata).
Для получения линеаризованной части плазмиды, предназначенной для трансформации, исходную плазмиду pINT-HIS4-GAPY727-INU обрабатывают эндонуклеазой рестрикции MluI. Интегрируемая часть плазмиды фланкирована областями для встраивания в определенный локус генома - эндогенный промотор АОХ1. Экспрессионный вектор pINT-HIS4-GAPGS115INU содержит те же элементы, что и предыдущий вектор, за исключением селективного маркера, HIS4, который был клонирован из К.pfaffii GS115 (получен при помощи ПЦР, с матрицы - геномной ДНК штамма К.pfaffii GS115, праймеры pr1_HIS4, pr2_HIS4), а также промотора GAPGS115, также клонированного из К.pfaffii GS115. Промотор GAP получили с помощью ПЦР, в качестве матрицы использовали геномную ДНК штамма К.pfaffii GS115, праймеры pr1_GAP (5'-aggaagcttttttgtagaaatgtcttggt), pr2_GAP (5'-ataggtacctgcagccatggtagatctttgatagttgttcaattgattgaaata). При обработке эндонуклеазой рестрикции MluI, линеаризованный фрагмент также встраивается в локус генома АОХ1.
Экспрессионный вектор pINT-HIS4-AOX1Y727-INU содержит элементы, необходимые для его амплификации в клетках E.coli: ген бета-лактамазы (в качестве селективного маркера), бактериальный репликон для обеспечения поддержания определенного уровня копийности плазмиды. Для трансформации дрожжевых клеток и экспрессии целевого гена, вектор содержит следующие элементы: ген HIS4 - гистидинол дегидрогеназы, в качестве селективного маркера, клонированный из К.pastoris ВКПМ Y-727, ген целевого белка - инулазы, клонированный из Kluyveromyces marxianus [14], находящийся под контролем промотора AOX1Y727, клонированного из К.pastoris ВКПМ Y-727, терминатор АОХ1. Для получения линеаризованной части плазмиды, предназначенной для трансформации, исходную плазмиду pINT-HIS4-AOX1Y727-INU обрабатывают эндонуклеазой рестрикции MluI. Интегрируемая часть плазмиды фланкирована областями для встраивания в определенный локус генома - эндогенный промотор АОХ1. Экспрессионный вектор pINT-HIS4-AOX1GS115INU содержит те же элементы, что и предыдущий вектор, за исключением селективного маркера, HIS4, который был клонирован из К.pfaffii GS115, а также промотора GAPGS115, также клонированного из К.pfaffii GS115. При обработке эндонуклеазой рестрикции MluI, линеаризованный фрагмент также встраивается в локус АОХ1.
Таким образом было получено 4 плазмиды, две из которых:
pINT-HIS4-GAPY727-INU и pINT-HIS4-AOX1Y727-INU - предназначены для трансформации штамма К.pastoris Y-727his4Δ и отличаются между собой только промоторами (АОХ1 и GAP), управляющими экспрессией целевого гена инулазы у штамма К.pastoris Y-727his4Δ;
две другие pINT-HIS4-GAPGS115INU; pINT-HIS4-AOX1GS115INU - предназначены для трансформации штамма К.pfaffii GS115 и отличаются между собой также только промоторами (АОХ1 и GAP), управляющими экспрессией целевого гена инулазы у штамма К.pfaffii GS115.
Трансформация
В качестве реципиентных штаммов используют штамм К.pastoris Y-727his4Δ и штамм К.pfaffii GS115, в качестве контрольного. При этом экспрессия гена инулазы регулируется промотором АОХ1 или GAP в каждом из анализируемых штаммов-продуцентов.
В результате трансформации каждого из этих штаммов линеаризованным фрагментам плазмиды pINT-HIS4-AOX1Y727-INU или pINT-HIS4-AOX1GS115INU соответственно, с последующим отбором трансформантов на селективной агаризованной среде YNB без аминокислот с сульфатом аммония, получают штаммы, продуцирующие инулазу: К.pastoris ВКПМ Y-727/pINT-HIS4-AOX1Y727-INU и К.pfaffii GS115/pINT-HIS4-AOX1GS115INU. Экспрессия гена инулазы находится под контролем индуцируемого промотора АОХ1.
В результате трансформации каждого из этих штаммов линеаризованным фрагментам плазмиды pINT-HIS4-GAPY727-INU или pINT-HIS4-GAPGS115INU соответственно, с последующим отбором трансформантов на селективной агаризованной среде YNB без аминокислот с сульфатом аммония, получают штаммы, продуцирующие инулазу: К.pastoris ВКПМ Y727/ pINT-HIS4-GAPY727-INU и К.pfaffii GS115/pINT-HIS4-GAPGS115INU. Экспрессия гена инулазы происходит под контролем конститутивного промотора GAP.
В результате, получены следующие штаммы-продуценты инулазы:
Komagataella pastoris ВКПМ Y727/pINT-HIS4-GAPY727-INU
Komagataella pastoris ВКПМ Y727/pINT-HIS4-AOX1Y727-INU
Komagataella pfaffii GS115/pINT-HIS4-GAPGS115INU
Komagataella pfaffii GS115/pINT-HIS4-AOX1GS115INU
Определение активности инулазы
Для проведения опыта выбрали по 2 клона каждого штамма. Инокулят анализируемого клона К.pastoris Y727/pINT-HIS4-GAPY727-INU или К.pfaffii GS115/pINT-HIS4-GAPGS115INU был помещен в пробирки с 5 мл среды YPD, а инокулят клона К.pastoris ВКПМ Y-727/pINT-HIS4-AOX1Y727-INU или К.pfaffii GS115/pINT-HIS4-AOX1GS115INU - в пробирки с 5 мл среды YPgM.
Культуру выращивали в течение 40 часов в ротационном шейкере-термостате (250 об/мин), при температуре 28°С. В случае роста на среде YPgM проводили индукцию метанолом спустя 24 часа роста, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%. По истечении 40 часов отделяли биомассу центрифугированием в настольной центрифуге, при 3000 g.
Активность секретируемой инулазы определяли в культуральной жидкости, с использованием в качестве субстрата сахарозы. Инулаза гидролизует β-гликозидную связь, в результате чего образуется восстанавливающий углевод - глюкоза. Количество образовавшейся глюкозы позволяет судить об активности фермента инулазы. Количество глюкозы измеряют по оптической плотности (при λ=492 нм) реакционной смеси, после добавления 3,5-динитросалициловой кислоты, которая восстанавливается до 3-амино-5-нитросалициловой кислоты, меняя окраску [15]. Активность инулазы представлена в единицах. 1 единица активности - такое количество фермента, которое освобождает 1 мкмоль глюкозы за 1 мин, нормированное на 1 мл культуральной жидкости и далее на 1 оптическую единицу (OD595) культуры.
Из полученных данных, представленных в табл.3 и на Фиг.7 а, б, видно, что активность инулазы в случае экспрессии под контролем промотора АОХ1 выше более чем в 2 раза в штамме К.pastoris ВКПМ Y-727/pINT-HIS4-AOX1Y727-INU, по сравнению с контрольным штаммом К.pfaffii GS115/pINT-HIS4-AOX1GS115INU. И наоборот, в случае регуляции гетерологичной экспрессии промотором GAP, активность продуцируемой инулазы больше у штамма К.pfaffii GS115/pINT-HIS4-GAPGS115INU, по сравнению с K.pastoris Y727/pINT-HIS4-GAPY727-INU.
Пример 10. Получение продуцента внутриклеточного белка поверхностного антигена вируса гепатита В
Конструирование векторов pPH93-AOX1GS115-HBsAg; pPH93-AOX1Y727-HBsAg
Экспрессионный вектор pPH93-AOX1GS115-HBsAg содержит элементы, необходимые для его амплификации в клетках E.coli: ген бета-лактамазы (в качестве селективного маркера), бактериальный репликон для обеспечения поддержания определенного уровня копийности плазмиды. Для трансформации дрожжевых клеток и экспрессии целевого гена, вектор содержит следующие элементы: ген HIS4 - гистидинол дегидрогеназы, клонированного из штамма GS115 с промоторной частью, в качестве селективного маркера; ген целевого белка - поверхностного антигена вируса гепатита В (HBsAg) [20], находящийся под контролем промотора АОХ1, терминатор АОХ1, клонированные из штамма GS115. Интегрируемая часть плазмиды фланкирована областями для встраивания в определенный локус генома - эндогенный промотор АОХ1. Для получения линеаризованной части плазмиды, предназначенной для трансформации, исходную плазмиду рРН93-AOX1GS115-HBsAg обрабатывают эндонуклеазой рестрикции MluI.
Экспрессионный вектор pPH93-AOX1Y727-HBsAg содержит элементы, необходимые для его амплификации в клетках E.coli: ген бета-лактамазы (в качестве селективного маркера), бактериальный репликон для обеспечения поддержания определенного уровня копийности плазмиды. Для трансформации дрожжевых клеток и экспрессии целевого гена, вектор содержит следующие элементы: ген URA3 - оротидин декарбоксилазы, клонированного из заявляемого штамма с промоторной частью, в качестве селективного маркера; ген целевого белка - HBsAg, находящийся под контролем промотора АОХ1, терминатор АОХ1, клонированные из заявляемого штамма. Интегрируемая часть плазмиды фланкирована областями для встраивания в определенный локус генома - эндогенный промотор АОХ1. В результате обработки эндонуклеазой рестрикции MluI плазмиды рРН93-AOX1GS115-HBsAg получают линеаризованную часть плазмиды, предназначенную для трансформации реципиентного штамма.
Трансформация
В качестве реципиентных штаммов используют штамм К.pastoris Y727ura3muthis4Δ и в качестве контрольного штамм К.pfaffii GS115.
В результате трансформации штамма К.pastoris Y727ura3muthis4Δ линеаризованным фрагментом плазмиды pPH93-AOX1Y727-HBsAg, с последующим отбором трансформантов на селективной агаризованной среде YNB с содержанием гистидина в концентрации 50 мкг/мл среды, получили штамм-продуцент штамм К.pastoris Y727ura3muthis4Δ/pPH93-AOX1Y727-HBsAg, синтезирующий поверхностный антиген вируса гепатита В.
В результате трансформации штамма К.pfaffii GS115 линеаризованным фрагментом плазмиды pPH93-AOX1GS115-HBsAg, с последующим отбором трансформантов на селективной агаризованной среде YNB, получили штамм-продуцент штамм К.pfaffii GS115/pPH93-AOX1GS115-HBsAg, синтезирующий поверхностный антиген вируса гепатита В.
Определение уровня продукции поверхностного антигена вируса гепатита В
Для проведения опыта выбрали по 2 клона каждого штамма. Инокулят анализируемых клонов был помещен в пробирки с 5 мл среды YPgM. Культуру выращивали в течение 72 часов в ротационном шейкере-термостате (250 об/мин), при температуре 28°С. Каждые 24 часа проводили индукцию метанолом, путем асептического добавления 50% раствора метанола в пробирки, до конечной концентрации 0,5%. По истечении 72 часов биомассу отделяли центрифугированием в настольной центрифуге, при 3000 g.
Для определения уровня продукции использовали метод иммуноферментного анализа на твердой подложке. В качестве подложки использовали стандартный планшет для иммуноферментного анализа с предварительно сорбированными человеческими поликлональными антителами к HBsAg. Данный планшет, конъюгат - мышиные моноклональные антитела к HBsAg, меченные пероксидазой хрена, а также хромоген ТМБ (3,3',5,5'-тетраметилбензидин) входили в стандартный набор «ГепаСтрип» (пр-во ООО «Ниармедик плюс») для определения HBsAg в сыворотке крови.
Осажденные клетки разрушали в буфере (10 мМ фосфатный буфер, 500 мМ NaCl, 10 мМ ЭДТА, 2 мМ PMSF, 1% Tween 20, рН 7.2) с помощью стеклянных шариков (Sigma) на дезинтеграторе FastPrep 24 (фирмы МР). Анализировали количество HBsAg в полученном лизате. В качестве стандарта для калибровки использовали отраслевой стандартный образец HBsAg вируса гепатита В (кат. № В-331, ООО «Научно-производственное объединение Диагностические системы»). Количество HBsAg в тотальном лизате клонов определяли по интенсивности поглощения реакционной смеси при длине волны λ=492 нм. Количественно данные нормированы на 100 оптических единиц и приведены на фиг.8 и в таблице 4, из которых видно, что продукция поверхностного антигена вируса гепатита В одинакова у обоих сравниваемых штаммах в пределах погрешности измерения.
Таким образом, подтверждено эффективное применение штамма метилотрофных дрожжей К.pastoris ВКПМ Y-727 в качестве штамма-реципиента для конструирования продуцентов, способных синтезировать целевой белок. Штамм обладает следующими характеристиками, соответствующими или превосходящими характеристики штамма - ближайшего аналога:
- пригоден для синтеза различных целевых белков, в частности инулазы, сывороточного альбумина, поверхностного антигена вируса гепатита В, бета-галактозидазы, что подтверждено примерами 2, 8, 9, 10; при этом могут быть использованы как индуцируемые (АОХ1), так и конститутивные (GAP) промоторы (примеры 2, 8, 9, 10);
- в отличие от ближайшего аналога, позволяет осуществлять регулируемый промотором АОХ1 синтез целевого белка как в присутствии индуктора-метанола, так и в его отсутствии с выходом до 70% от уровня синтеза этого белка в присутствии метанола (примеры 1, 2);
- позволяет достаточно просто получать традиционно используемые при генетическом конструировании ауксотрофные мутанты, что подтверждено примерами 3, 4, 5, 6, 7.
Список научно-технической информации
1. Daly R, Milton TW Hearn (2005) Expression of heterologous proteins in Pichia pastoris: a useful experimental tool in protein engineering and production. J. Mol. Recognit. 18: 119-138.
2. Narhi LO, Arakawa T, Strickland TW. (1991). The effect of carbohydrate on the structure and stability of erythropoietin. J. Biol. Chem. 266: 23022-23026.
3. Macauley-Patrick S, Fazenda ML (2005) Heterologous protein production using the Pichia pastoris expression system. Yeast 22: 249-270.
4. Balamurugan V, Reddy GR (2006) Pichia pastoris: A notable heterologous expression system for the production of foreign proteins - Vaccines. Indian Journal of Biotechnology (6): 175-186.
5. Gellissen G (2000) Heterologous protein production in methylotrophic yeasts. Appl. Microbiol. Biotechnol. 54(6): 741-750.
6. Kurtzman CP (2009) Biotechnological strains of Komagataella (Pichia) pastoris are Komagataella phaffii as determined from multigene sequence analysis. J Ind Microbiol Biotechnol 36: 1435-1438.
7. Bhatacharya P, Pandey G (2007) Production and purification of recombinant human granulocyte-macrophage colony stimulating factor (GM-CSF) from high cell density cultures of Pichia pastoris. Bioprocess and Biosystems Engineering 30(5): 305-312.
8. Zhang H, Yuan QP (2007) Cloning and secretion expression of hepcidin in Pichia pastoris. Sheng Wu Gong Cheng Xue Bao 23(3): 381-385.
9. Yurimoto H (2011) Yeast Methylotrophy: Metabolism, Gene Regulation and Peroxisome Homeostasis. Hindawi Publishing Corporation International Journal of Microbiology Article ID 101298.
10. Conde R, Cueva R (2004) A Search for Hyperglycosylation Signals in Yeast Glycoproteins. The journal of biological chemistry (42): 43789-43798.
11. Hartner A, Glieder A (2006) Regulation of methanol utilization pathway genes in yeasts. Microbial Cell Factories 2006, 5: 39.
12. Tschopp JF, Brust PF, Cregg JM (1987) Expression of the lacZ gene from two methanol-regulated promoters in Pichia pastoris. Nucleic Acids Res. 15: 3859-3876.
13. Cregg JM, Barringer KJ (1985) Pichia pastoris as a Host System for Transformations. MOLECULAR AND CELLULAR BIOLOGY, Dec. 1985, p.3376-3385.
14. Бревнова Е.Э., Козлов Д.Г. Штамм дрожжей Saccharomyces cerevisiae - продуцент фруктозы из инулина, 1998, Биотехнология, 1, 12-20.
15. Berezin IV, Rabinovich ML Study of Applicability of quantitative kinetic spectrophotometric method for glucose determination, 1997, Biokhimiya, 42: 1631-1637.
16. Shixuan Wu, Geoffrey J. Letchworth High efficiency transformation by electroporation of Pichia pastoris pretreated with lithium acetate and dithiothreitol, BioTechniques, 2004, 36: 152-154.
17. Rose M, Botstein D Construction and use of gene fusions lacZ (b-galactosidase) which are expressed in yeast. Meth. Enzymol. 101: 167-180.
18. Oka A Sugisaki H Takanami M Nucleotide sequence of the kanamycin resistance transposon Tn903 JOURNAL J. Mol. Biol. 147 (2), 217-226 (1981).
19. Марченко А.Н., Козлов Д.Г., Свирщевская Е.В., Беневоленский С.В. Использование N-конца белка ТуА для создания гибридных вирусоподобных частиц. Биотехнология 2001, №2, 3-11.
20. Чеперегин С.Э., Арман И.П., Грановский Н.Н. "Оптимизация экспрессии гена поверхностного антигена вируса гепатита В в дрожжах". Мол. генетика, микробиология, вирусология 1990, 5, с.17-20.
21. Постановление Главного государственного санитарного врача Российской Федерации от 12 июля 2011 г. N 99 г.Москва "Об утверждении СП 2.3.3.2892-11 "Санитарно-гигиенические требования к организации и проведению работ с метанолом".
22. Государственное санитарно-эпидемиологическое нормирование Российской Федерации 4.1. Методы контроля. Химические факторы, определение концентраций химических веществ в воздухе. Газохроматографическое определение метанола в воздухе. Методические указания МУК 4.1.1046а-01 Выпуск 2, Минздрав России, Москва 2002.
| название | год | авторы | номер документа |
|---|---|---|---|
| Трансформант дрожжей Komagataella kurtzmanii, продуцирующий бета-глюканазу | 2019 |
|
RU2722563C1 |
| Способ получения дрожжей K. kurtzmanii с улучшенной способностью к усвоению сорбитола | 2023 |
|
RU2814987C1 |
| Штамм дрожжей Komagataella phaffii с инактивированным геном HIS4 - реципиент для конструирования безмаркерных штаммов-продуцентов гетерологичных белков | 2022 |
|
RU2787584C1 |
| Способ получения штамма-продуцента фосфолипазы А2 Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2S | 2020 |
|
RU2746817C1 |
| ИНТЕГРАТИВНЫЙ ПЛАЗМИДНЫЙ ВЕКТОР ДЛЯ ЭКСПРЕССИИ ГЕНОВ В ДРОЖЖАХ | 2008 |
|
RU2388823C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В ДРОЖЖАХ PICHIA PASTORIS ГЕНА ФИТАЗЫ (ВАРИАНТЫ), ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS - ПРОДУЦЕНТ ФИТАЗЫ (ВАРИАНТЫ) | 2009 |
|
RU2409670C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА ДЛЯ ЭКСПРЕССИИ В ДРОЖЖАХ PICHIA PASTORIS ГЕНА ФОСФОЛИПАЗЫ, ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS - ПРОДУЦЕНТ ФОСФОЛИПАЗЫ | 2009 |
|
RU2409671C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК РНВS, ОБЕСПЕЧИВАЮЩАЯ БИОСИНТЕЗ ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА В, СПОСОБ КОНСТРУИРОВАНИЯ ПЛАЗМИДЫ, ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS PS103 (PHBS) - ПРОДУЦЕНТ УКАЗАННОГО АНТИГЕНА | 2000 |
|
RU2201452C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Pichia pastoris - ПРОДУЦЕНТ СЕКРЕТИРУЕМОЙ ТЕРМОСТАБИЛЬНОЙ КСИЛОГЛЮКАНАЗЫ, КОДИРУЕМОЙ СИНТЕТИЧЕСКИМ ГЕНОМ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА СЕКРЕТИРУЕМОЙ ТЕРМОСТАБИЛЬНОЙ КСИЛОГЛЮКАНАЗЫ НА ОСНОВЕ ЭТОГО ШТАММА | 2015 |
|
RU2605629C1 |
| Рекомбинантная плазмида, способ её конструирования и штамм дрожжей Komagataella pastoris - продуцент иммунного интерферона-гамма собаки | 2020 |
|
RU2756852C2 |




Изобретение относится к области биотехнологии и касается применения штамма дрожжей Komagataella pastoris ВКПМ Y-727. Охарактеризовано применение штамма дрожжей Komagataella pastoris ВКПМ Y-727 в качестве реципиента для конструирования продуцентов целевого белка, необязательно включающее введение в него мутаций, обеспечивающих использование ауксотрофных селективных маркеров. Представленное решение может быть применимо при получении рекомбинантных белков без использования метанола в качестве индуктора экспрессии генов. 8 ил., 4 табл., 10 пр.
Применение штамма дрожжей Komagataella pastoris ВКПМ Y-727 в качестве реципиента для конструирования продуцентов целевого белка, необязательно включающее введение в него мутаций, обеспечивающих использование ауксотрофных селективных маркеров.
| ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS PS107(pPIC9HAbIL-2), ЯВЛЯЮЩИЙСЯ ПРОДУЦЕНТОМ ГИБРИДНОГО БЕЛКА, СОСТОЯЩЕГО ИЗ АЛЬБУМИНА ПЛАЗМЫ КРОВИ ЧЕЛОВЕКА И ИНТЕРЛЕЙКИНА-2 ЧЕЛОВЕКА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pPIC9HAbIL-2 И СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ | 2006 |
|
RU2315105C1 |
| WO 2003010288 A2, 06.02.2003 | |||

