ПРИТЯЗАНИЕ НА ПРИОРИТЕТ
Настоящая заявка заявляет приоритет даты регистрации предварительной патентной заявки США, серийный номер 61/319764, зарегистрированной 31 марта 2010 года для заявки "РАСТИТЕЛЬНЫЙ ПЕПТИД ГАММА-ЗЕИН ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ".
УРОВЕНЬ ТЕХНИКИ
Традиционные стратегии растениеводства для разработки новых линий растений, проявляющих конкретные признаки, являются трудоемкими и иногда непрогнозируемыми. Существующие стратегии, такие как трансформация с помощью Agrobacterium и бомбардировка частицами, в значительной степени зависят от ткани и генотипа. Пептиды, проникающие в клетки (CPP), представляют собой новый и быстро развивающийся класс коротких пептидов, которые играют важную роль в перемещении широкого диапазона транспортируемых комплексов, включающих в себя ДНК, РНК и белки, через клеточные мембраны в линиях клеток млекопитающих и человека (Schwartz and Zhang, 2000; Langel, 2002; Vives, 2002).
Несмотря на то, что CPP облегчают доставку груза в клетки млекопитающих, использование CPP в растительных клетках для изучения трансфекции ограничено многими факторами. Основное препятствие для адаптации этой технологии для растений состоит в том, что, в отличие от животных клеток, в растительных клетках существует система двойного барьера (клеточная стенка и плазматическая мембрана) для интернализации пептидов CPP и их груза. Поэтому для эффективного перемещения транспортируемых молекул в интактные растительные клетки CPP должны преодолеть два упомянутых барьера. В растительных клетках использовались CPP, но их применение основывалось на использовании агентов проницаемости для осуществления доставки транспортируемых молекул в интактные растительные клетки. CPP-опосредованная доставка маленьких молекул, нуклеиновых кислот и белков в интактные растительные клетки остается в значительной степени неисследованной и имеет преимущества для генетических и биохимических манипуляций в растительных системах in vivo и in vitro.
Наночастицы обладают уникальными свойствами, которые задействуют для использования при доставке ДНК в клетки. Наночастицы металлов, такие как наночастицы золота (Au), применяют для доставки ДНК в силу их низкой цитотоксичности и легкой функционализации с различными биологически важными лигандами. В дополнение к наночастицам металлов, для доставки молекул в клетки в качестве носителей также используют полупроводниковые наночастицы (например, квантовые точки - "QD") с размером в диапазоне 3-5 нм. ДНК и белки могут быть связаны с лигандом, присоединенным к QD посредством различных поверхностных функциональных групп (см., например, Patolsky F. Et al., J. Chem. Soc. 125, 13918 (2003)).
Наночастицы используют для доставки плазмидной ДНК в различные клетки животных. Было выявлено, что при инкубировании покрытых ДНК наночастиц с клетками, не имеющими клеточной стенки, клетки захватывают наночастицы и начинают экспрессировать любые гены, кодируемые на ДНК. Вместе с тем, на современном этапе существуют проблемы доставки растительных генов, обусловленные наличием стенок в растительных клетках, что приводит к традиционной зависимости от инвазивных способов доставки в целях генетической трансформации растений. Если желательна доставка наночастиц к клеткам, обычно имеющим клеточную стенку, то перед добавлением частиц к растительным прототипам клеточную стенку удаляют (см. Torney, F. Et al, Nature Nanotechnol. 2, (2007)). Клеточная стенка растительных клеток представляет собой труднопреодолимый барьер для доставки экзогенно находящихся молекул. Для осуществления доставки генов и маленьких молекул в растительные клетки со стенкой применялось множество инвазивных способов, таких как генная пушка (биолистика), микроинъекция, электропорация и Agrobacterium, но доставка белков достигалась только с помощью микроинъекции.
На фоне постоянно растущего объема информации о разработках в области последовательности генома растений существует острая потребность в быстром, универсальном (независимым от ткани/генотипа) способе функциональных исследований генома растений для широкого разнообразия генов и для разработки трансгенных растений, экспрессирующих важные агрономические признаки.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Описание следующих вариантов осуществления приведено совместно с системами, инструментами и способами, которые предназначены служить примерами, выполнять иллюстративные задачи и не ограничивать объем изобретения.
Один вариант осуществления настоящего изобретения относится к способу введения молекулы, представляющей интерес, в растительную клетку, имеющую клеточную стенку. Способ включает в себя предоставление растительной клетки, имеющей клеточную стенку, и взаимодействие пептида гамма-зеина с рассматриваемой молекулой для образования связанной структуры гамма-зеина. Затем клетку, имеющую клеточную стенку, и связанную структуру гамма-зеина располагают так, чтобы они контактировали друг с другом, и была возможность захвата рассматриваемой гамма-зеин связанной молекулы в клетку, имеющую клеточную стенку.
Другой вариант осуществления изобретения относится к способу экспрессии гена, и указанный способ включает в себя предоставление растительной клетки, имеющей клеточную стенку, и взаимодействие пептида гамма-зеина с рассматриваемым геном для образования гамма-зеин связанной генной структуры. Растительную клетку, имеющую клеточную стенку, и гамма-зеин связанную структуру располагают так, чтобы они контактировали друг с другом, и была возможность захвата пептида гамма-зеина и гена в растительную клетку, содержащую клеточную стенку. Затем в потомстве растения, несущего эту растительную клетку, происходит экспрессия гена.
Еще в одном варианте осуществления изобретения в растительную клетку переносится молекулярная субстанция. Способ включает в себя взаимодействие пептида гамма-зеина с плазмидной ДНК для образования гамма-зеин связанной структуры. Гамма-зеин связанная структура располагается так, чтобы она контактировала с интактной растительной клеткой, имеющей клеточную стенку, при условиях, позволяющих захватывать растительную клетку пептид гамма-зеин и ген из плазмидной ДНК.
В дополнение к примерам аспектов и вариантов осуществления, описанным выше, дополнительные аспекты и варианты осуществления станут очевидными в свете последующего описания.
КРАТКОЕ ОПИСАНИЕ ФИГУР
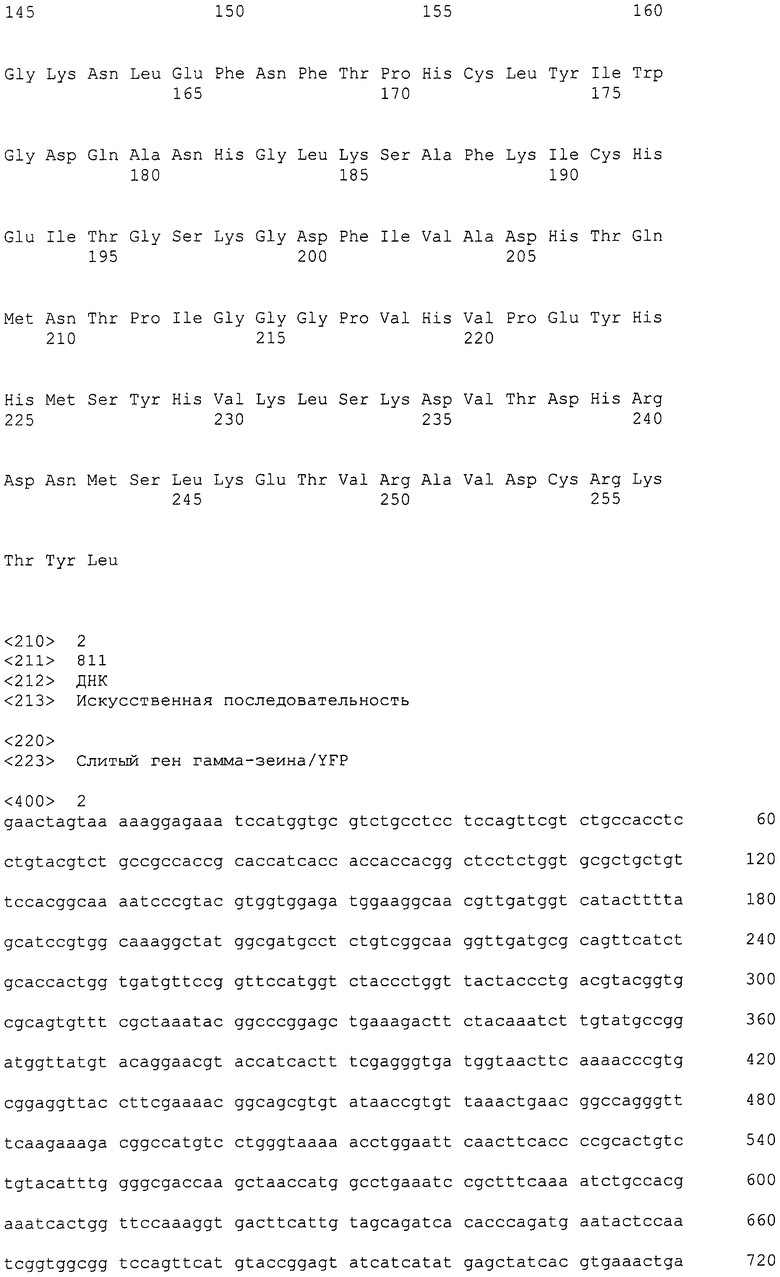
Фиг. 1 представляет карту плазмиды слитого гена гамма-зеин/YFP.
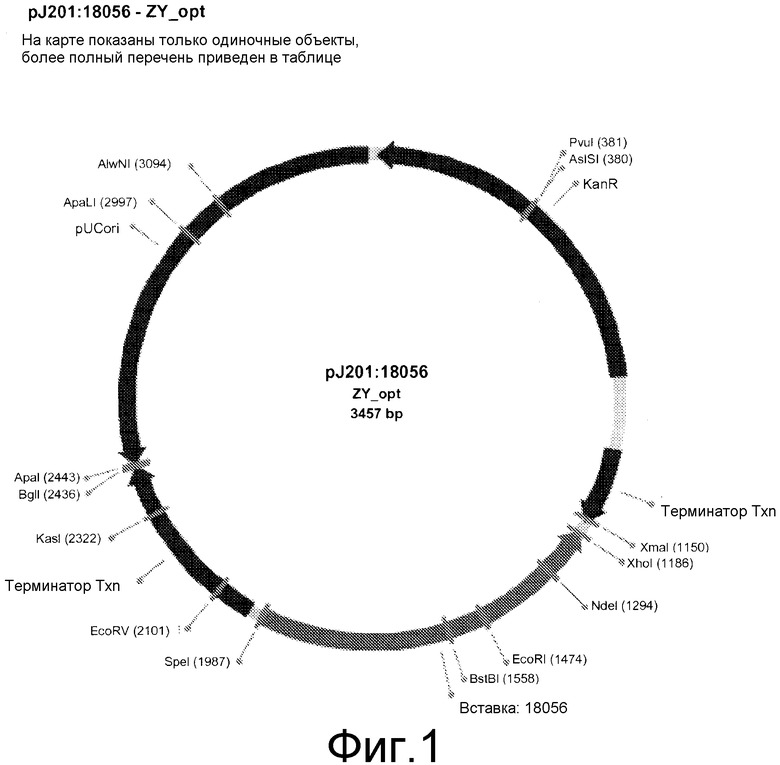
Фиг. 2 представляет Ni хелирующую хроматографию очистки слитого пептида гамма-зеин/YFP.
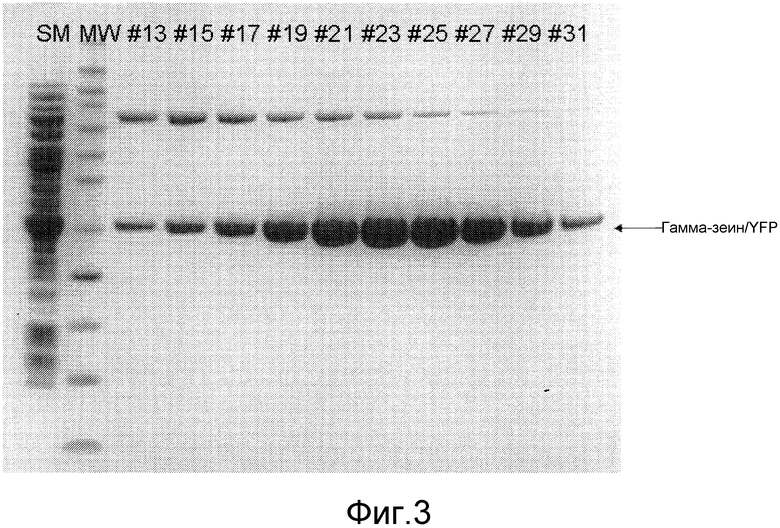
Фиг. 3 показывает профиль элюирования гамма-зеина/YFP в анализе SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия).
Фиг. 4 показывает масс-спектры пептида гамма-зеина/YFP при время-пролетной ионизации лазерной десорбцией с использованием матрицы Maldi-TOF.
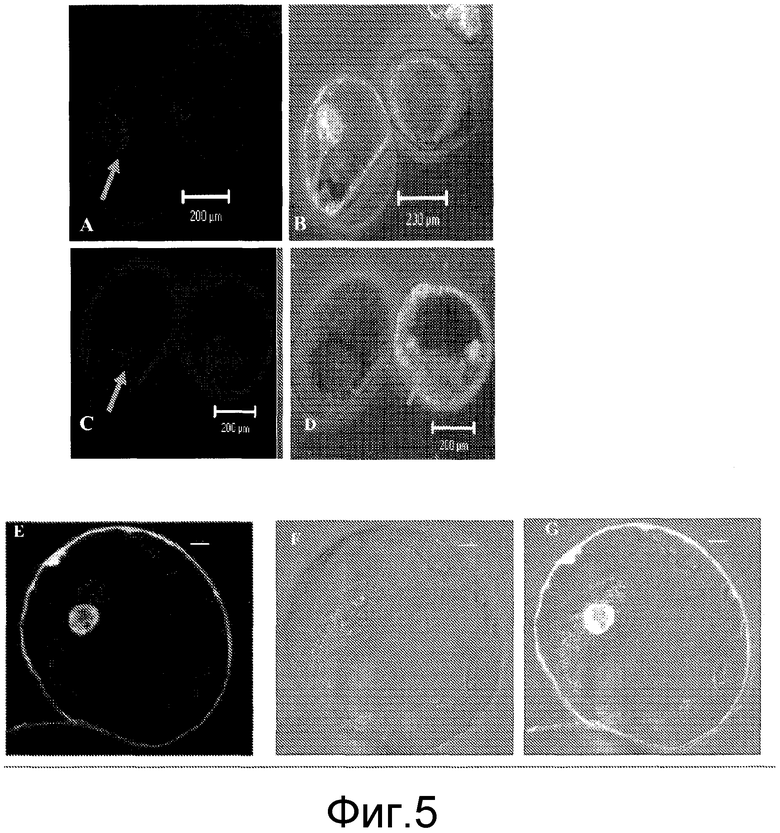
Фиг. 5 показывает конфокальные микроскопные изображения, иллюстрирующие клеточный захват и интернализацию слитого пептида: A. Одиночная клетка моркови, окрашенная Хехст 33342 с показом ядра; B. Одиночная клетка моркови, показывающая локализацию флюоресценции гамма-зеин/YFP в ядре и цитоплазме; C. Клетки табака JTNTI, окрашенные Хехст 33342 с показом ядра; D. Клетки табака JTNTl, показывающие флюоресценцию гамма-зеин/YFP в ядре и цитоплазме вместе с окрашиванием ядер синим цветом красителем Хехст 33342; E. Конфокальное изображение, показывающее слитый пептид гамма-зеин/YFP, нацеленный и на плазматическую мембрану и на ядро клеток JTNTl; и F и G, показывающие соответствующие изображения в светлом поле и наложения конфокальных/светлопольных изображений, соответственно.
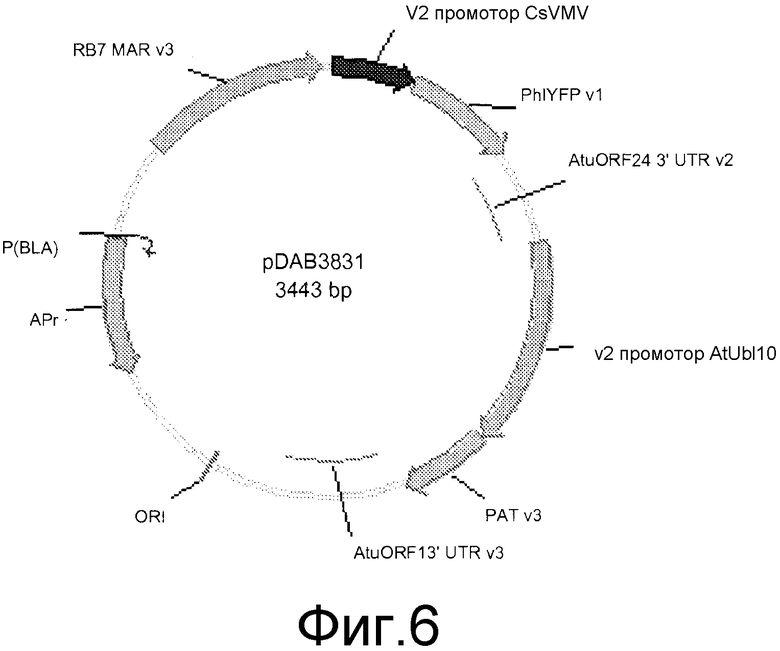
Фиг. 6 показывает карту плазмиды pDAB3831.
СПОСОБ (СПОСОБЫ) ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
В нижеследующем описании и таблицах применяется ряд терминов. Для достижения ясного и последовательного понимания описания и формулы изобретения, включающих в себя область применения таких терминов, представлены следующие определения:
обратное скрещивание. Обратное скрещивание представляет собой способ, при котором селекционер неоднократно скрещивает гибридное потомство обратно с одним из родителей, например, гибрид F1 первого поколения скрещивает с одним из родительских генотипов гибрида F1.
Зародыш. Зародыш может представлять собой маленькое растение, содержащееся в зрелом семени.
Зеин. Зеин представляет собой кукурузный белок, получаемый из кукурузной глютеновой муки. В нем не содержится аминокислот лизина и триптофана, таким образом, он не подходит в качестве единственного источника пищевого белка. Зеин нерастворим в воде и спиртах, но является растворимым в водных спиртах, гликолях и гликолевых эфирах. Зеин присутствует в виде пленки и покрытия для обеспечения влагоустойчивого барьера в орехах и зерновых продуктах. Он также используется как покрытие для кондитерских изделий и для глазури на формовых изделиях.
Наночастица. Микроскопическая частица, имеющая по меньшей мере один размер в наномасштабе, обычно меньше чем 100 нм. Наночастица, подходящая для использования в настоящем изобретении, может иметь размер в диапазоне от 1 нм до 0,4 мкм. Квантовая точка может иметь средний диаметр в пределах от 1 нм до 10 нм, предпочтительно от 2 до 4 нм. Наночастицу можно выбирать из: наночастицы золота, покрытой золотом наночастицы, пористой наночастицы, мезопористой наночастицы, кварцевой наночастицы, полимерной наночастицы, вольфрамовой наночастицы, желатиновой наночастицы, нанооболочек, наноядер, наносфер, наностержней, магнитных наночастиц и их комбинаций.
Квантовая точка. Квантовая точка представляет собой полупроводниковую наноструктуру, которая ограничивает движение электронов зоны проводимости, дырки зоны валентности или экситоны (связанные пары электронов зоны проводимости и дырки зоны валентности) во всех трех пространственных измерениях. Удержание может осуществляться благодаря электростатическим потенциалам (генерируемым внешними электродами, допированием, посторонними напряжениями), наличием пространства между различными полупроводниковыми материалами (например, в нанокристаллической системе ядро-оболочка), наличием полупроводниковой поверхности (например, полупроводникого нанокристалла) или их комбинацией. Квантовая точка может иметь дискретный квантованный спектр энергии. Соответствующие волновые функции пространственно локализованы в квантовой точке, но распространяются через многочисленные периоды кристаллической решетки. Квантовая точка содержит малое конечное число электронов зоны проводимости (порядка 1-100), дырок валентной зоны, или экситонов (то есть, конечное число элементарных электрических зарядов).
Стабильные или устойчивые трансформанты. Термины «стабильные или устойчивые трансформанты» относятся к растению, геном которого репродуктируемо передается от одного поколения к следующему поколению.
Захват. Захват относится к перемещению частицы, такой как гамма-зеин, через клеточную стенку или клеточную мембрану, при этом перемещение не происходит исключительно в результате момента, который передается частице чем-либо отличным от клетки, в которую захватывается частица. Для сравнения, примерами устройств или способов, которые вызывают перемещение частицы через клеточную стенку или клеточную мембрану исключительно в результате момента, передаваемого частице, является генная пушка, технологии биолистики, микроинъекции и/или импалефекции.
В конкретном варианте осуществления изобретение относится к использованию гамма-зеина в качестве CPP для эффективной доставки полезных грузов в интактные растительные клетки, для применения при доставке маленьких молекул, доставке биомолекул, доставке генов, визуализации и различных задач биотехнологической диагностики и функций индикации в растительных системах.
В других вариантах осуществления изобретения к молекуле гамма-зеина можно присоединять "добавочную" или "гостевую" молекулу. Это свойство можно задействовать, например, для специфичного нацеливания и редактирования молекулярных участков в клетках, в таких областях данной техники, как биомиметики, направленная доставка, для вариантов негенетически модифицированных организмов и вариантов неустойчивой трансформации у многих древесных, овощных и зерновых культур в целях достижения признака и устойчивости к болезням. Варианты осуществления изобретения можно применять также для разработки подходящих биосенсоров в растениях.
Согласно вариантам осуществления изобретения может быть представлен способ введения молекулы, представляющей интерес, в растительную клетку, содержащую клеточную стенку, и указанный способ содержит приведение гамма-зеина, содержащего молекулу, представляющую интерес, в контакт с растительный клеткой, и предоставление возможности захвата гамма-зеина через клеточную стенку растительной клетки. В конкретных аспектах изобретения гамма-зеин может обратимо или необратимо содержать молекулу, представляющую интерес, может взаимодействовать или иным образом быть связанным с молекулой, представляющей интерес, и/или нести ее.
Согласно вариантам осуществления настоящего изобретения, растительная клетка, имеющая клеточную стенку, может представлять собой любую растительную клетку, содержащую интактную и целую клеточную стенку. Примеры клеток, имеющих клеточную стенку, включают в себя без ограничения водоросли, табак, морковь, кукурузу, канолу, рапс, хлопок, пальму, арахис, сою, сахарный тростник, Oryza sp., Arabidopsis sp., и Ricinus sp., предпочтительно, табак, кукурузу, морковь, хлопок, канолу, сою и сахарный тростник; более предпочтительно табак и морковь. Варианты осуществления изобретения могут включать в себя содержащие клеточную стенку клетки из любой ткани или из любого места их выявления, включая в себя без ограничения зародыши, меристематические клетки, каллюс, пыльцу, листья, пыльники, корни, корневые кончики, цветы, семена, стручки, стебли, культуру тканей и суспензии интактных одиночных растительных клеток.
В вариантах осуществления изобретения рассматриваемая молекула может представлять собой любую молекулу, которую можно доставлять в растительную клетку согласно настоящему изобретению. Молекулы, представляющие интерес, или компоненты молекул, представляющие интерес, могут включать в себя без ограничения любые маленькие молекулы, нуклеиновые кислоты, молекулы ДНК, РНК, iРНК, miРНК, гены, плазмиды, космиды, YAC, BAC, полипептиды, ферменты, гормоны, гликопептиды, сахара, жиры, сигнальные пептиды, антитела, витамины, мессенджеры, вторичные мессенджеры, аминокислоты, цАМФ, лекарства, гербициды, фунгициды, антибиотики и/или их комбинации.
Варианты осуществления изобретения включают в себя способы профилактики или лечения болезни. Неограничивающие примеры вариантов осуществления включают в себя доставку фунгицидов, антибиотиков, и/или других лекарственных веществ в клетки, нуждающиеся в такой доставке, с помощью способов настоящего изобретения.
В аспектах изобретения связанная структура гамма-зеина может захватываться во множество органел растительной клетки. Примеры локализаций, в которые могут захватываться связанные структуры гамма-зеина, включают в себя без ограничения цитозоль, ядро, тонопласты, пластиды, этиопласты, хромопласты, лейкопласты, элайопласты, протеинопласты, амилопласты, хлоропласты и просвет двойных мембран. В других вариантах осуществления изобретения захват связанных структур гамма-зеина в клетку, содержащую клеточную стенку, может происходить посредством симпластического или апопластического пути.
Дополнительные варианты осуществления изобретения включают в себя генетически модифицированные растительные клетки и способы их получения, при этом растительные клетки имеют одну или более нуклеиновых кислот, внедренных в эти клетки посредством способов настоящего изобретения. В одном примере вариантов осуществления с помощью гамма-зеина согласно настоящему изобретению в растительную клетку, имеющую клеточную стенку, можно вводить плазмиду, содержащую ген, представляющий интерес, и селектируемый маркер. В дополнительных вариантах осуществления можно выбирать устойчивые трансформанты, в которые устойчиво внедрили ген, представляющий интерес, и/или селектируемый маркер. В альтернативных вариантах осуществления можно размножать растительную клетку, уже содержащую ген, представляющий интерес, чтобы получить другие клетки, содержащие молекулу, представляющую интерес. В других вариантах осуществления растительные клетки, уже содержащие молекулу, представляющую интерес, могут представлять собой регенерируемую клетку, которую можно использовать для регенерации целого растения, включающего в себя молекулу, представляющую интерес.
В другом аспекте настоящее изобретение относится к способам создания регенерируемых растительных клеток, содержащих молекулу, представляющую интерес, для использования в тканевой культуре. Тканевая культура предпочтительно будет способна к регенерации растений, имеющих по существу тот же генотип, что и регенерируемые клетки. Регенерируемые клетки в таких тканевых культурах могут представлять собой зародыши, прототипы, меристематические клетки, каллюс, пыльцу, листья, пыльники, корни, корневые кончики, цветы, семена, стручки или стебли. Еще один дополнительный вариант осуществления изобретения относится к растениям, регенерированным из тканевых культур по изобретению.
Альтернативно, настоящее изобретение относится к способу введения желательного признака в растительную клетку, имеющую клеточную стенку, при этом способ содержит: введение способного обеспечить желательный признак гена гамма-зеина, представляющего интерес, в растительную клетку, имеющую клеточную стенку и предоставление возможности захвата гамма-зеин связанного гена, представляющего интерес, через клеточную стенку. Примеры желательных признаков включают в себя без ограничения признаки, выбираемые из следующего: мужская стерильность, устойчивость к гербицидам, устойчивость к насекомым, устойчивость к бактериальному, грибковому и/или вирусному заболеванию, и признаки, дающие преимущества для конечного пользователя, такие как модифицированные масляные профили, измененное содержание крахмала и волокон, увеличенное содержание витаминов и аминокислот и тому подобное.
Дополнительные аспекты изобретения представляют способы создания устойчивых линий растений, содержащих целевую молекулу или ген, представляющих интерес, при этом целевая молекула или ген, представляющие интерес, сначала могут вводиться через стенку растительной клетки путем гамма-зеин опосредованной транслокации. Способы стабилизации растительных линий, измененных генетически или другим способом, известны рядовым специалистам в данной области и включают в себя без ограничения такие методики, как самовоспроизводство, обратные скрещивания, гибридное производство, скрещивание с устойчивыми популяциями и тому подобное. Все растения и растительные клетки, содержащие целевую молекулу или ген, представляющие интерес, вначале вводятся в растительную клетку (или ее предшественники) путем опосредуемого гамма-зеином переноса через клеточную стенку согласно настоящему изобретению. Преимущество состоит в том, что растительные клетки, содержащие молекулу или ген, представляющие интерес, которые сначала вводят в растение или клетку (или ее предшественники) путем опосредуемого гамма-зеином переноса через клеточную стенку, можно использовать в кросс-бридинге с другими различными растениями для получения гибридных клеток первого поколения (F1), семян и/или растений с более хорошими характеристиками и фенотипами.
В вариантах осуществления, в которых рассматриваемая молекула содержит один или более генов, ген (гены) может быть доминантной или рецессивной аллелью. Как пример, ген (гены) будет придавать такие признаки, как устойчивость к гербицидам, устойчивость к насекомым, устойчивость к бактериальной устойчивости, устойчивость к грибковым заболеваниям, устойчивость к вирусным заболеваниям, мужская фертильность, мужская стерильность, улучшенные пищевые качества и промышленное применение.
С появлением технологий молекулярной биологии, которые позволили выделять и оценивать гены, кодирующие конкретный белок или РНК-продукты (например, iРНК), у ученых в области биологии растений возник сильный интерес к созданию генома клеток, в которых могут содержаться и экспрессироваться чужие гены, или дополнительные или модифицированные версии нативных или эндогенных генов (возможно, с помощью различных промоторов), в целях изменения признаков клетки конкретным образом. Такие чужие дополнительные и/или модифицированные гены в настоящем изобретении обобщенно называются "трансгенами". За последние пятнадцать - двадцать лет были разработаны несколько способов получения трансгенных растительных клеток, и в конкретных вариантах осуществления настоящее изобретение относится к трансформированным версиям клеток и способам их получения посредством введения трансгена в растительную клетку, имеющую клеточную стенку, путем захвата связанной структуры гамма-зеина через стенку растительной клетки и мембрану. В вариантах осуществления изобретения трансген может содержаться в векторе экспрессии.
Клеточная трансформация может охватывать конструирование вектора экспрессии, кодирующего конструкцию, которая будет функционировать в конкретной клетке. Такой вектор может содержать ДНК, которая включает в себя ген под контролем регуляторного элемента, или функционально связана с регуляторным элементом (например, промотором). Вектор экспрессии может содержать один или более таких комбинаций из функционально связанного гена/регуляторного элемента. Вектор (векторы) может быть в форме плазмиды и может использоваться единственным или в комбинации с другими плазмидами для получения трансформированных клеток с помощью описанных в изобретении способов трансформации, с целью введения трансгена (трангенов) в генетический материал растительной клетки, содержащей клеточную стенку.
Векторы экспрессии для захвата посредством гамма-зеина: маркерные или репортерные гены
Векторы экспрессии могут включать в себя по меньшей мере один генетический маркер, функционально связанный с регуляторным элементом (например, промотором), который позволяет восстанавливать несущие маркер трансформированные клетки или путем отрицательной селекции (то есть, ингибированием роста клеток, которые не содержат селектируемый маркерный ген), или путем положительной селекции (то есть скринингом продукта, кодируемого генетическим маркером). В методиках трансформации известно множество селектируемых маркерных генов для трансформации, и они включают в себя, например, гены, кодирующие ферменты, которые метаболически детоксифицируют селективный химический агент, который может представлять собой антибиотик или гербицид, или гены, кодирующие измененную мишень, которая может быть нечувствительной к ингибитору. В данной области также известен ряд способов положительной селекции.
Один широко используемый селектируемый маркерный ген, подходящий для трансформации растений, может включать в себя ген неомицин-фосфотрансферазы II (nptll) под контролем растительных регуляторных сигналов, которые придают устойчивость к канамицину. См., например, Fraley et al,, Proc. Natl. Acad. Sci. USA, 80:4803 (1983). Другим обычно используемым селектируемым маркерным геном может быть ген гигромицин-фосфотрансферазы, который придает устойчивость к антибиотику гигромицину. См., например, Vanden Elzen et al., Plant Mol. Biol, 5:299 (1985). Дополнительные селектируемые маркерные гены бактериального происхождения, которые придают устойчивость к антибиотикам, включают в себя гентамицин-ацетилтрансферазу, стрептомицин-фосфотрансферазу, аминогликозид-3'-аденил-трансферазу и детерминанту устойчивости к блеомицину. См. публикации: Hayford et al., Plant Physiol. 86: 1216 (1988), Jones et al., Mol. Gen. Genet., 210:86 (1987), Svab et al., Plant Mol. Biol. 14: 197 (1990), Hille et al., Plant Mol. Biol. 7: 171 (1986). Другие селектируемые маркерные гены придают устойчивость к гербицидам, таким как глифосфат, глюфосинат или бромоксинил. См. публикации Comai et al., Nature 317:741-744 (1985), Gordon-Kamm et al., Plant Cell 2:603-618 (1990) and Stalker et al., Science 242:419-423 (1988). Селектируемые маркеры, которые придают толерантность к гербицидам, включают в себя фосфинотрицин-ацетилтрансферазу (PAT).
Другие селектируемые маркерные гены, подходящие для трансформации растений, не имеют бактериального происхождения. Эти гены включают в себя, например, мышиную дигидрофолат-редуктазу, растительную 5-энолпирувилшикимат-3-фосфат-синтазу и растительную ацетолактат-синтазу. См. публикации Eichholtz et al., Somatic Cell Mol. Genet. 13:67 (1987), Shah et al., Science 233:478 (1986), Charest et al., Plant Cell Rep. 8:643 (1990).
Для другого класса маркерных генов, подходящих для трансформации растений, необходим скрининг предположительно трансформированных растительных клеток, а не прямая генетическая селекция трансформированных клеток на устойчивость к токсичному веществу, например, к антибиотику. Эти гены особенно полезны для количественного определения или визуализации пространственной характеристики экспрессии гена в конкретных тканях, и часто их называют репортерными генами, поскольку для изучения генной экспрессии их можно сливать с геном или регуляторной последовательностью гена. Обычно используемые гены для скрининга трансформированных клеток включают в себя β-глюкуронидазу (GUS), β-галактозидазу, люциферазу и хлорамфеникол-ацетилтрансферазу. См. публикации Jefferson, R. A., Plant Mol. Biol. Rep. 5:387 (1987), Teeri et al., EMBO J. 8:343 (1989), Koncz et al Proc. Natl. Acad. Sci U.S.A. 84: 131 (1987), DeBlock et al EMBO J. 3: 1681 (1984).
В последнее время стали доступными способы визуализации активности GUS in vivo, которые не требуют разрушения растительной ткани. См. публикация Molecular Probes publication 2908, Imagene Green.TM., p. 1-4(1993) и Aleway et al., J. Cell Biol. 115: 151a (1991). Вместе с тем, эти способы in vivo для визуализации активности GUS не доказали свою полезность для восстановления трансформированных клеток из-за низкой чувствительности, высоких исходных уровней флуоресцентности и ограничений, связанных с использованием генов люциферазы в качестве селектируемых маркеров.
Позднее в качестве маркеров генной экспрессии в прокариотических и эукариотических клетках были использованы гены, кодирующие флуоресцентные белки (например, GFP, EGFP, EBFP, ECFP и YFP). См. Chalfie et al., Science 263:802 (1994). В качестве возможных для скрининга маркеров можно использовать флуоресцентные белки и мутации флуоресцентных белков.
Векторы экспрессии для захвата посредством гамма-зеина: промоторы
Гены, включенные в векторы экспрессии, должны запускаться нуклеотидными последовательностями, содержащими регуляторный элемент, например, промоторы. В настоящее время в области трансформации известны несколько типов промоторов, а также другие регуляторные элементы, которые можно использовать единственными или в комбинации с промоторами.
Используемый в изобретении термин "промотор" относится к области ДНК, которая может располагаться в 3'-5' направлении от начала транскрипции, и может участвовать в распознавании и связывании РНК-полимеразы и других белков для инициации транскрипции. "Растительным промотором" может быть промотор, способный к инициации транскрипции в растительных клетках. Примеры промоторов под контролем индивидуального развития включают в себя промоторы, которые предпочтительно инициируют транскрипцию в конкретных тканях, таких как листья, корни, семена, волокна, сосуды ксилемы, трахеиды или склеренхима. Такие промоторы называют "тканепредпочтительными". Промоторы, которые инициируют транскрипцию только в определенных тканях, называются "тканеспецифичными". Промотор, специфичный по "типу клетки", осуществляет экспрессию преимущественно в определенных типах клеток в одном или более органах, например, в сосудистых клетках корней или листьев. "Индуцибельным" промотором может быть промотор, который может регулироваться окружающей средой. Примеры условий окружающей среды, при которых может проходить транскрипция индуцибельными промоторами, включают в себя анаэробные условия или присутствие света. Тканеспецифичные, тканепредпочтительные, специфичные по типу клетки и индуцибельные промоторы составляют класс "неконститутивных" промоторов. "Конститутивный" промотор представляет собой промотор, который может быть активным в большинстве условий окружающей среды.
A. Индуцибельные промоторы
Индуцибельный промотор может быть функционально связан с геном для экспрессии в клетке. Необязательно, индуцибельный промотор может быть функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая может быть функционально связана с геном для экспрессии в клетке. При наличии индуцибельного промотора скорость транскрипции увеличивается в ответ на индуцирующий агент.
В настоящем изобретении можно использовать любой индуцибельный промотор. См. Ward et al, Plant Mol. Biol. 22:361-366 (1993). Примеры индуцибельных промоторов включают в себя без ограничения: промоторы из системы ACEI, которые реагируют на медь (Mett et al., PNAS 90:4567-4571 (1993)); ген In2 из кукурузы, который реагирует на антидоты бензолсульфонамидного гербицида (Hershey et al, Mol. Gen Genetics 227:229-237 (1991) и Gatz et al, Mol. Gen. Genetics 243:32-38 (1994)); и ген - репрессор Tet из Tnl0 (Gatz et al., Mol. Gen. Genetics 227:229-237 (1991)). Особенно полезным индуцибельным промотором может быть промотор, который реагирует на индуцирующий агент, на который растение обычно не реагирует. Примером индуцибельного промотора может быть индуцибельный промотор из гена стероидного гормона, транскрипционную активность которого можно индуцировать глюкокортикостероидным гормоном. Schena et al., Proc. Natl. Acad. Sci. USA 88:0421 (1991).
B. Конститутивные промоторы
Конститутивный промотор может быть функционально связан с геном для экспрессии в клетке, или конститутивный промотор может быть функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая может быть функционально связана с геном для экспрессии в клетке.
В настоящем изобретении можно использовать различные конститутивные промоторы. Примеры конститутивных промоторов включают в себя без ограничения: промоторы из вирусов растений, например, промотор 35S из CaMV (Odell et al., Nature 313:810-812 (1985)) и промотор из вируса прожилковой мозаики маниоки (CsVMV) (см., например, патенты США 7053205 и 6664384); промоторы из гена риса actin (McElroy et al, Plant Cell 2: 163-171 (1990)); убиквитин (Christensen et al., Plant Mol. Biol. 12:619-632 (1989) и Christensen et al., Plant Mol. Biol. 18:675-689 (1992)); pEMU (Last et al., Theor. Appl. Genet. 81:581-588 (1991)); MAS (Velten et al., EMBO J. 3:2723-2730 (1984)); и гистон H3 кукурузы (Lepetit et al, Mol. Gen. Genetics 231:276-285 (1992) и Atanassova et al., Plant Journal 2 (3): 291-300 (1992)). Особенно полезным конститутивным промотором является промотор ALS, фрагмент 5' Xbal/Ncol к структурному гену Brassica napus ALS3 (или нуклеотидная последовательность, подобная указанному фрагменту Xbal/Ncol). См. PCT заявку WO 96/30530.
C. Тканеспецифичные или тканепредпочтительные промоторы
Тканеспецифичный промотор может быть функционально связан с геном для экспрессии в клетке. Необязательно, тканеспецифичный промотор может быть функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая может быть функционально связана с геном для экспрессии в клетке. Растения, трансформированные рассматриваемым геном, функционально связанным с тканеспецифичным промотором, могут продуцировать белковый продукт трансгена исключительно или предпочтительно в конкретной ткани.
В настоящем изобретении можно использовать любой тканеспецифичный или тканепредпочтительный промотор. Примеры тканеспецифичных или тканепредпочтительных промоторов включают в себя без ограничения промотор, предпочтительный для корней, например, промотор из гена фазеолина (Murai et al., Science 23:476-482 (1983) и Sengupta-Gopalan et al., Proc. Natl. Acad. Sci. USA 82:3320-3324 (1985)); специфичный для листьев и индуцируемый светом промотор, такой как промотор из cab или rubisco (Simpson et al., EMBO J. 4(11):2723-2729 (1985) и Timko et al., Nature 318:579-582 (1985)); промотор, специфичный для пыльника, например, промотор из LAT52 (Twell et al., Mol. Gen. Genetics 217:240-245 (1989)); специфичный для пыльцы промотор, такой как Zm13 (Guerrero et al, Mol. Gen. Genetics 244: 161-168 (1993)) или предпочтительный для микроспор промотор, такой как промотор из apg (Twell et al., Sex. Plant Reprod. 6:217-224 (1993)).
Транспорт белка, продуцируемого трансгенами, в субклеточное пространство, например, в хлоропласт, вакуолю, пероксисому, глиоксисому, клеточную стенку или митохондрию, или для секреции в апопласт, может осуществляться посредством функционального присоединения нуклеотидной последовательности, кодирующей сигнальную последовательность, к 5' и/или 3'-областям гена, кодирующего рассматриваемый белок. Нацеливание последовательностей на 5' и/или 3' концах структурного гена во время синтеза и процессинга белка может определить, где может произойти окончательная компартментализация кодируемого белка. Альтернативно, такие белки, нацеленные на субклеточное пространство, могут непосредственно связываться с гамма-зеином для нацеливания молекулы, на желательное субклеточное пространство.
Присутствие сигнальной последовательности направляет полипептид или к внутриклеточному органоиду или в субклеточное пространство, или для секреции в апопласте. В данной области известно множество сигнальных последовательностей. См., например, Becker et al., Plant Mol. Biol. 20:49 (1992), Close, P. S., Master's Thesis, Iowa State University (1993), Knox, С et al., "Structure and Organization of Two Divergent Alpha-Amylase Genes from Barley", Plant Mol. Biol. 9:3-17 (1987), Lerncr ct al., Plant Physiol. 91:124-129 (1989), Fontes ct al., Plant Cell 3:483-496 (1991), Matsuoka et al., Proc. Natl. Acad. Sci. 88:834 (1991), Gould et al., J. Cell. Biol. 108:1657 (1989), Creissen et al., Plant J. 2; 129 (1991), Kalderon et al., A short amino acid sequence able to specify nuclear location, Cell 39:499-509 (1984), Steifel et al., Expression of a maize cell wall hydroxyproline-rich glycoprotein gene in early leaf and root vascular differentiation, Plant Cell 2:785-793 (1990).
Гены чужих белков и гены, представляющие агрономический интерес
С помощью трансгенных растений согласно настоящему изобретению чужой белок может быть произведен в коммерческих количествах. Таким образом, технологии селекции и размножения трансформированных растений, которые широко распространены в данной области, в результате дают множество трансгенных растений, сбор которых проводят общепринятыми способами, и затем из рассматриваемой ткани или из общей биомассы можно извлекать чужой белок. Извлечение белка из растительной биомассы можно осуществлять известными способами, которые рассмотрены, например, авторами Heney and Orr, Anal. Biochem. 114:92-6 (1981).
В аспектах изобретения трансгенное растение, предназначенное для коммерческого получения чужого белка, может представлять собой клетку или растение. В других аспектах рассматриваемая биомасса может быть семенами. Для относительно небольшого количества трансгенных растений, которые проявляют более высокие уровни экспрессии, можно создавать генетическую карту прежде всего посредством общепринятых анализов ПДРФ (полиморфизма длин рестрикционных фрагментов), ПЦР (полимеразно-цепной реакцией) и SSR (простых повторяющихся последовательностей), которые определяют приблизительную хромосомную локализацию интегрированной молекулы ДНК. В этом плане примеры методик см. в публикациях Glick and Thompson, Methods in Plant Molecular Biology and Biotechnology CRC Press, Boca Raton 269:284 (1993). Информация по картированию в отношении хромосомной локализации может быть полезной для собственной защиты рассматриваемого трансгенного растения. Если может произойти непредусмотренное размножение и скрещивание с другой зародышевой плазмой, карту области интеграции можно сравнивать с подобными картами подозрительных растений, чтобы определить наличие у последних общего происхождения с рассматриваемым растением. Сравнения карт будут охватывать гибридизации, ПДРФ, ПЦР, SSR и секвенирование, все из которых являются общепринятыми методиками.
Аналогично, в трансформированных клетках или в их потомстве можно экспрессировать гены, представляющие агрономический интерес. Более подробно, растения можно генетически конструировать с помощью способов по изобретению для экспрессии разнообразных фенотипов, представляющих агрономический интерес. Примеры генов, которые можно использовать в этом отношении, включают в себя без ограничения гены согласно нижеприведенной классификации:
1. Гены, которые придают устойчивость к вредителям или болезням, и которые кодируют:
A) Гены устойчивости к болезням растений. Защита растений часто активируется специфичным взаимодействием между продуктом гена устойчивости к болезни (R) у растения и продуктом соответствующего гена авирулентности (Avr) у патогена. Различные растения можно трансформировать клонированными генами устойчивости, чтобы создать растения, которые являются устойчивыми к определенным патогенным штаммам. См., например, Jones et al., Science 266:789 (1994) (клонирование гена томата Cf-9 для устойчивости к Cladosporium fulvum); Martin et al., Science 262: 1432 (1993) (ген Pto томата для устойчивости к Pseudomonas syringae pv. tomato, кодирующий протеинкиназу); Mindrinos et al., Cell 78: 1089 (1994) (ген Arabidops RSP2 для устойчивости к Pseudomonas syringae).
B) Ген, придающий устойчивость к вредителю, такому как цистообразующая нематода сои. См. например, заявку PCT WO 96/30517; заявку PCT WO 93/19181.
C) Белок Bacullus thuringiensis, его производное или его синтетическая полипептидная модель. См., например, публикацию Geiser et al, Gene 48: 109 (1986), в которой раскрыто клонирование и нуклеотидная последовательность гена Bt δ-эндотоксина. Кроме того, молекулы ДНК, кодирующие гены δ-эндотоксина, можно приобретать в Американской коллекции типовых культур, Manassas, Va., например, под номерами доступа ATCC №№ 40098, 67136, 31995 и 31998.
D) Лектин. См., например, публикацию Van Damme et al., Plant Molec. Biol. 24:25 (1994), в которой раскрыта нуклеотидная последовательности нескольких генов лектина, связывающего маннозу из Clivia miniata.
E) Витамин-связывающий белок, такой как авидин. См. заявку PCT US93/06487. Заявка рассматривает применение авидина и гомологов авидина в качестве ларвицидов против насекомых-вредителей.
F) Ингибитор фермента, например, протеаза или ингибитор протеиназы или ингибитор амилазы. См., например, Abe et al., J. Biol. Chem. 262: 16793 (1987) (нуклеотидная последовательность ингибитора протеиназы цистеина риса), Huub et al., Plant Molec. Biol. 21:985 (1993) (нуклеотидная последовательность кДНК, кодирующей ингибитор I протеиназы табака), Sumitani et al., Biosci. Biotech. Biochem. 57: 1243 (1993) (нуклеотидная последовательность ингибитора альфа-амилазы Streptomyces nitrosporeus) и патент США № 5494813 (авторы Hepher и Atkinson, опубликованный 27 февраля 1996 года).
G) Специфичный для насекомого гормон или феромон, такой как экдистероид или ювенильный гормон, их вариант, миметик на их основе, или их антагонист или агонист. См., например, публикацию Hammock et al., Nature 344:458 (1990), в которой раскрыта бакуловирусная экспрессия клонированной эстеразы ювенильного гормона, инактиватора ювенильного гормона.
H) Специфичный для насекомого пептид или нейропептид, который при экспрессии нарушает физиологию пораженного вредителя. Например, см. публикации Regan, J. Biol. Chem. 269:9 (1994) (в которой раскрыты результаты экспрессионного клонирования ДНК, кодирующей рецептор диуретического гормона насекомого), и Pratt et al., Biochem. Biophys. Res. Comm. 163: 1243 (1989) (возможность идентификации аллостатина в Diploptera puntata). См. также патент США № 5266317 авторов Tomalski и др., который раскрывает гены, кодирующие специфичные для насекомого паралитические нейротоксины.
I) Специфичный для насекомого яд, продуцируемый в природе змеями, осами или любым другим организмом. Например, см. публикацию Pang et al., Gene 116: 165 (1992), в которой раскрыта гетерологичная экспрессия в растениях гена, кодирующего пептид яда скорпиона.
J) Фермент, ответственный за гипераккумуляцию монотерпена, сесквитерпена, стероида, гидраксамовой кислоты, фенилпропаноидного производного или другая небелковой молекулы с инсектицидным действием.
K) Фермент, вовлеченный в модификацию, включающую в себя посттрансляционную модификацию биологически активной молекулы; например, гликолитический фермент, протеолитический фермент, липолитический фермент, нуклеаза, циклаза, трансаминаза, эстераза, гидролаза, фосфатаза, киназа, фосфорилаза, полимераза, эластаза, хитиназа и глюканаза, как природного происхождения, так и синтетические. См. заявку PCT 93/02197 авторов Scott et al., в которой раскрыта нуклеотидная последовательность гена каллазы. Молекулы ДНК, которые содержат последовательности, кодирующие хитиназу, можно получать, например, из ATCC под номерами доступа 39637 и 67152. См. также публикацию Kramer et al., Insect Biochem. Molec. Biol. 23:691 (1993), в которой рассмотрена нуклеотидная последовательность кДНК, кодирующая хитиназу табачного бражника, и Kawalleck et al., Plant Molec. Biol. 21:673 (1993), которая относится к нуклеотидной последовательность гена полиубиквитина ubi4-2 петрушки.
L) Молекула, которая активирует сигнальную трансдукцию. Например, см. публикацию Botella et al., Plant Molec. Biol. 24:757 (1994), в которой раскрыты нуклеотидные последовательности клонов кДНК кальмодулина золотистой фасоли, и Griess et al., Plant Physiol. 104: 1467 (1994), в которой раскрыта нуклеотидная последовательность клона кДНК кальмодулина кукурузы.
M) Пептид гидрофобного момента. См. заявку PCT WO 95/16776 (раскрытие производных пептида тахиплесина, которые ингибируют грибковые патогены растений), и заявку PCT WO 95/18855 (рассматривает синтетические антибактериальные пептиды, которые придают устойчивость к болезням).
N) Мембранная пермеаза, образующая канал или блокирующая канал. Например, см. раскрытие Jaynes et al., Plant Sci 89:43 (1993) гетерологичной экспрессии секропин-β-литического аналога пептида для придания трансгенным растениям табака устойчивости к Pseudomonas solanacearum.
O) Вирусно-инвазивный белок или полученный из него комплексный токсин. Например, накопление вирусных оболочечных белков в трансформированных клетках растений придает устойчивость к вирусной инфекции и/или к развитию болезни, вызванной вирусом, из которого можно получить ген оболочечного белка, а также вызванной родственными вирусами. См. Beachy et al., Ann. rev. Phytopathol. 28:451 (1990). Обусловленная оболочечным белком устойчивость передавалась при трансформации растения против вируса мозаики люцерны, вируса огуречной мозаики, вируса полосатости табака, картофельного вируса X, картофельного вируса Y, вируса гравировки табака, вируса погремковости табака и вируса табачной мозаики. Id.
P) Специфичное для насекомого антитело или иммунотоксин, полученный из антитела. Таким образом, антитело, нацеленное на важную метаболическую функцию в кишечнике насекомого, будет инактивировать фермент, на который оказано воздействие, и уничтожать насекомое. Для сравнения: Taylor et al., Тезис № 497, Седьмой Международный симпозиум по молекулярным взаимодействиям растение - микроорганизм (Эдинбург, Шотландия) (1994) (ферментативная инактивация в трансгенном табаке путем продукции фрагментов одноцепочечных антител).
Q) Вирус-специфичное антитело. См., например, Tavladoraki et al., Nature 366:469 (1993), где показано, что трансгенные растения, экспрессирующие гены рекомбинантного антитела, защищены от вирусной атаки.
R) Белок, ингибирующий индивидуальное развитие, продуцируемый в природе патогеном или паразитом. Например, грибковые эндо-α-1,4-D-полигалактуроназы способствуют грибковой колонизации и высвобождению растительного питательного вещества путем солюбилизации гомо-α-1,4-D-галактуроназы стенки растительной клетки. См. Lamb et al., Bio/Technology 10: 1436 (1992). Описание клонирования и характеристики гена, который кодирует эндополигалактуроназа - ингибирующий белок бобов, можно найти у Toubart et al., Plant J. 2:367 (1992).
S) Белок, ингибирующий индивидуальное развитие, продуцируемый в природе растением. Например, авторами Logemann et al., Bio/Technology 10:305 (1992) показано, что трансгенные растения, которые экспрессируют ген, инактивирующий рибосому ячменя, имеют повышенную устойчивость к грибковым болезням.
2. Гены, которые придают устойчивость к гербицидам:
A) Гербицид, который ингибирует растущую точку или меристему, например, имидазолинон или сульфонилмочевина. Примеры генов этого типа кодируют мутантные ферменты ALS и AHAS, как описано, например, авторами Lee et al.., EMBO J. 7: 1241 (1988), и Miki et al., Theor. Appl, Genet. 80:449 (1990), соответственно.
B) Глифосат (устойчивость передается, например, мутантными генами 5-энолпирувилшикимат-3-фосфатсиназы (EPSP) (посредством внедрения рекомбинантных нуклеиновых кислот и/или различных форм нативных генов EPSP после мутагенеза in vivo), генами aroA и ацетилглифосат-трансферазы (GAT), соответственно), другими фосфоно-соединениями, такими как гены глюфосината (фосфинотрицин-ацетилтрансфераза (PAT) из видов Streptomyces, включающих в себя Streptomyces hygroscopicus и Streptomyces viridichromogenes), и пиридинокси- или фенокси-пропионовой кислоты и циклогексоны (гены, кодирующие ингибитор ACC-азы), См., например, патент США № 4940835 Shah et al., и патент США 6248876 авторов Barry et al., раскрывающие нуклеотидные последовательности форм EPSP, которые могут придавать растениям устойчивость к глифосату. Молекулу ДНК, кодирующую мутантный ген aroA, можно получать в ATCC под инвентарным номером 39256, и нуклеотидная последовательность мутантного гена раскрыта в патенте США № 4769061 под авторством Comai. Европейская патентная заявка № 0333033 авторов Kumada et al., и патент США № 4975374 Goodman et al., раскрывают нуклеотидные последовательности генов глутаминсинтетазы, которые придают устойчивость к гербицидам, таким как L-фосфинотрицин. Нуклеотидная последовательность гена PAT представлена в европейской заявке № 0242246 авторов Leemans et al., DeGreef et al., Bio/Tecnology 7:61 (1989), описывающей производство трансгенных растений, которые экспрессируют химерные гены bar, кодирующие активность PAT. Примеры генов, придающих устойчивость к феноксипропионовым кислотам и циклогексонам, таким как сетоксидим и галоксифоп, включают в себя гены Acc 1-S1, Acc 1-S2 и Acc 1-S3, описанные в публикации Marshall et al., Theor. Appl. Genet. 83:435 (1992). Гены GAT, способные придавать устойчивость к глифосату, описаны в патенте WO 2005012515 авторов Castle et al. Гены, придающие устойчивость к 2,4-D-феноксипропионовым кислотам и гербицидам пиридилокси-ауксину описаны в патенте WO 2005107437, принадлежащем Dow AgroSciences LLC.
C) Гербицид, который ингибирует фотосинтез, такой как триазин (гены psbA и gs+) или бензонитрил (ген нитрилазы). Авторы Przibila et al., Plant Cell 3:169 (1991) описывают трансформацию Chlamydomonas плазмидами, кодирующими мутантные гены psbA. Нуклеотидные последовательности генов нитрилазы раскрыты в патенте США № 4810648, Stalker и молекулы ДНК, содержащие эти гены, доступны в ATCC, номера доступа 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион-S-трансферазу, описано авторами Hayes et al., Biochem. J. 285: 173 (1992).
3. Гены, которые придают дополнительные положительные признаки или способствуют их появлению, например:
A) Модифицированный метаболизм жирных кислот, например, путем трансформации растения с антисмысловым геном стеарил-ACP-десатуразы для повышения содержания стеариновой кислоты в растении. См. Knultzon et al., Proc. Natl. Acad. Sci. USA 89:2624 (1992).
B) Уменьшенное содержание фитата - 1) Введение гена, кодирующего фитазу, будет увеличивать расщепление фитата, добавляя больше свободного фосфата в трансформированное растение. Например, см. раскрытие нуклеотидной последовательности гена фитазы Aspergillus niger в публикации Van Hartingsveldt et al., Gene 127:87 (1993). 2) Можно вводить ген, который уменьшает содержание фитата. В кукурузе, например, это можно осуществлять путем клонирования и затем повторного введения ДНК, связанной с единственной аллелью, которая может отвечать за мутанты кукурузы, отличающиеся низким уровнем содержания фитиновой кислоты. См. Raboy et al., Maydica 35:383 (1990).
C) Модифицированный углеводный состав, получаемый, например, путем трансформации растений геном, кодирующим фермент, который изменяет структуру ветвления крахмала. См. Shiroza et al., J. Bacteol. 170:810 (1988) (нуклеотидная последовательность мутантов гена фруктозилтрансферазы из Streptococcus), Steinmetz et al, Mol. Gen. Genet. 20:220 (1985) (нуклеотидная последовательность из Bacuillus subtilis может представлять собой ген левансахаразу), Pen et al., Bio/Technology 10:292 (1992) (производство трансгенных растений, экспрессирующих Bacuillus lichenifonnmay, которые могут быть α-амилазой), Elliot et al., Plant Molec. Biol. 21:515 (1993) (нуклеотидные последовательности генов инвертазы томата), Sogaard et al., J. Biol. Chem. 268:22480 (1993) (сайт-направленный мутагенез может происходить из гена α-амилазы ячменя), и Fisher et al., Plant Physiol. 102: 1045 (1993) (крахмал-ветвящий фермент II эндосперма кукурузы).
В конкретных вариантах осуществления изобретения гамма-зеин можно сливать с наночастицами. Поверхность наночастицы можно функционализировать, и это может, например, давать возможность направленного захвата или позволять обратимое или необратимое связывание других веществ на поверхность наночастицы. В качестве неограничивающего примера поверхность наночастицы (например, наночастицы золота или квантовых точек) можно функционализировать самособранным монослоем, например, из алкантиолатов, который можно дополнительно функционализировать или дериватизировать. В дополнительных неограничивающих примерах поверхность наночастицы можно дериватизировать линкерами, которые сами по себе можно дополнительно функционализировать или дериватизировать. В одном варианте осуществления наночастица может быть пэгилирована. В других вариантах осуществления наночастица может содержать одно или более ядер (активных или неактивных), стерическую оболочку (активную или инертную), расщепляемую связь и/или нацеливающую молекулу или лиганд, или может быть мультифункционализирована вышеперечисленным. Наночастицу можно выбирать из наночастицы золота, покрытой золотом наночастицы, пористой наночастицы, мезопористой наночастицы, кварцевой наночастицы, полимерной наночастицы, вольфрамовой наночастицы, желатиновой наночастицы, нанооболочек, наноядер, наносфер, наностержней, магнитных наночастиц и их комбинаций. Аналогично, в конкретных вариантах осуществления изобретения гамма-зеин может быть слитым с квантовой точкой.
В аспектах изобретения гамма-зеин и наночастица могут захватываться в различные клеточные элементы. Примеры локализаций, в которые может захватываться наночастица, включают в себя без ограничения цитозоль, ядро, тонопласты, пластиды, этиопласты, хромопласты, лейкопласты, элайопласты, протеинопласты, амилопласты, хлоропласты и просвет двойной мембраны. В других вариантах осуществления изобретения захват наночастицы в клетку, содержащую клеточную стенку, может происходить посредством симпластического или апопластического пути. В других аспектах изобретения гамма-зеин и квантовая точка могут захватываться в вышеупомянутые различные клеточные элементы.
ПРИМЕРЫ
Настоящее изобретение дополнительно описано в следующих примерах, которые приведены в качестве иллюстрации и не предназначены каким-либо образом ограничивать изобретение.
Пример 1
Конструкция слитого белка гамма-зеин/YFP
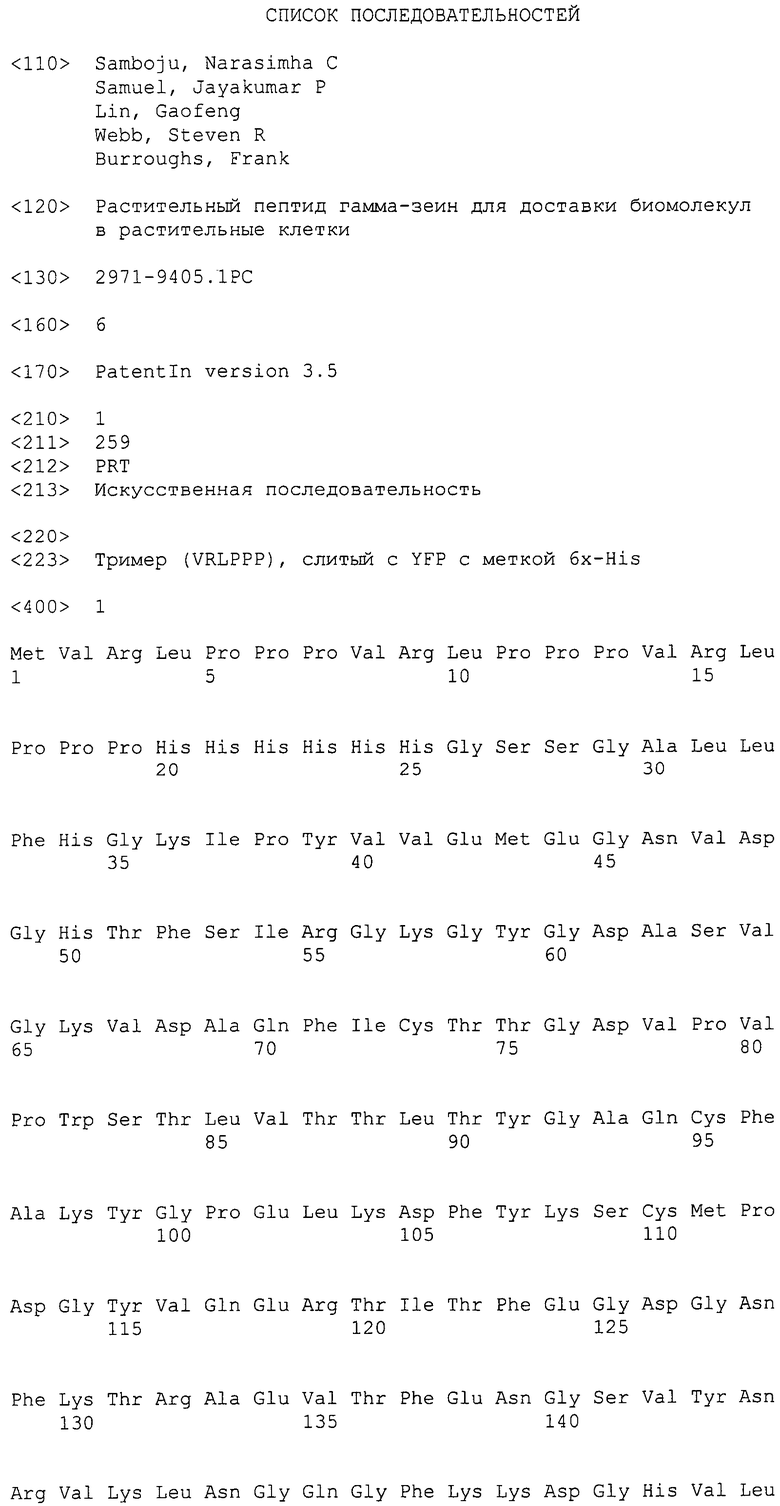
Последовательность ДНК, кодирующую N-концевую область модифицированного Zea mays белка гамма-зеина (номер доступа GenBank № AAL16977.1, GI: 16305109), сливали к N-концом желтого флюоресцентного белка (YFP) Philadium (Evrogen, Москва, Россия). Мотив гамма-зеина из шести аминокислот был модифицирован, при этом меняли второй остаток гистидин на аргинин. Эта модификация была сделана для направления слитой последовательности к ядру. Показано, что аргинин улучшил эффективность транслокации белка в цитозоль и ядро клетки. (Mitchell, et al., (2000), The Journal of Peptide Research, 56 (5):318-325). Тример из модифицированных шести аминокислот (VRLPPP) N-концевой области гамма-зеина сливали с YFP. Дополнительно, помещали метку 6x-His между кодирующей последовательностью тримера гамма-зеина и YFP. Для облегчения очистки белка добавляли мотив 6x-His. Аминокислотная последовательность этого слитого белка представлена как SEQ ID NO: 1.
Слитая генная последовательность (SEQ ID NO: 2) была химически синтезирована методикой фосфорамидитной химии с помощью автоматизированного синтезатора ДНК. Для химического синтеза указанной последовательности привлекали к DNA2.0 (Menlo Park, Калифорния). Последовательность представляла собой кодон, оптимизированный для экспрессии в Escherichia coli с помощью собственного алгоритма DNA2.0, чтобы получить "оптимизированную E. coli" нуклеотидную последовательность. Алгоритм идентифицирует кодоны, которые редко используются в желательном организме-хозяине, и заменяет эти кодоны на более часто используемые кодоны. Дополнительно, алгоритм удаляет цис-регуляторные последовательности (например, участки РНКазы, вторичную структуру РНК, участки терминации транскрипции) и лишние ферменты рестрикции. Для облегчения клонирования и экспрессии к концам слитого гена гамма-зеин/YFP добавляли дополнительные последовательности. К 5' концу последовательности гена добавляли последовательность Шайна-Дальгарно (Shine J, Dalgarno L (1975). Nature 254 (5495): 34-8). и уникальный сайт фермента рестрикции Spel. Уникальный сайт фермента рестрикции Xhol добавляли к 3' концу последовательности гена. Получаемый вектор отмечен как pJ201: 18056 (фиг. 1).
1.1 Конструирование вектора экспрессии pET
Слитую последовательность гамма-зеин/YFP клонировали в вектор экспрессии E. coli pET280 путем стандартных способов клонирования. pET280 представляет собой модифицированную версию плазмиды pET28 (Novagen, Gibbstown, NJ). Из pET28 удаляли сайт множественного клонирования и сайт связывания рибосомы, таким образом создавался pET280. Из векторов pJ201: 18056 вырезали фрагмент Spel - Xhol, содержащий кодирующие последовательности гамма-зеин/YFP, и лигировали в соответствующие сайты рестрикции вектора экспрессии pET280. Получаемую плазмиду подтверждали с помощью расщепления фермента рестрикции и секвенирования. Плазмиду трансформировали в компетентные клетки BL21 (DE3) Escherichia coli (Invitrogen, Карлсбад, Калифорния). Одиночные колонии выделяли и сохраняли в виде стокового раствора глицерина до дальнейшего использования.
1.2 Экспрессия и выделение слитого белка гамма-зеин/YFP
Слитый белок гамма-зеин/YFP индуцировали, используя следующие условия: культуры, содержащие конструкцию экспрессии pET280/гамма-зеин/YFP, выращивали до оптической плотности OD600=0,6 в 2 литрах (л) бульона Лурия-Бертани и 50 мкг/мл канамицина при 25°C. После достижения желательной OD600 культуры индуцировали 0,1 мМ IPTG (изопропил-B-D-тиогалактопиранозид) в течение 16 часов при 25°C.
Экспрессируемый слитый белок гамма-зеин/YFP выделяли и очищали. Клеточную культуру объемом 2 л центрифугировали при 24000×g в течение 10 минут, и удаляли супернатант. Ресуспендировали 5 г клеточной пасты, содержащей гамма-зеин/YFP в 100 мл холодного раствора для экстракции (0,5 М NaCl, 5% глицерин и 0,5 мл коктейля ингибитора протеазы Sigma (комплект # P8849) в фосфатно-буферном растворе ФБР). Клетки разрушали на льду с помощью ультразвука (модель Branson Sonifier 450) в течение 15 мин. Образец центрифугировали при 24000×g в течение 20 минут при 4°C, и супернатант фильтровали через фильтрационное устройство Millipore® с фильтром 0,45 мкм. К образцу добавляли имидазол в конечной концентрации 10 мМ. Лизат, содержащий гамма-зеин/YFP, со скоростью 5 мл/мин загружали в тандемную колонку 2×5 мл HisTrap™ (GE/Pharmacia, комплект # 17-5248-02) с использованием системы Akt™ Explorer 100 (GE'Pharmacia). Колонку промывали буфером (0,5 М NaCl, 10 мМ имидазол в ФБР), пока спектральная поглощательная способность A280 не достигала исходного уровня. Затем колонку промывали последовательно 10% и 20% буфером B (0,2 М имидазола в буфере A) в объемах приблизительно 4 × объем колонки (ОК).
Образец элюировали 20-100% буфером B в объеме более 10-15 ОК, и собирали 4 мл фракций (фиг. 2).
1.3 Анализ SDS-PAGE
Белковый гель прогоняли для визуализации фракционируемого белка. 10-20 мкл каждого образца загружали на сборный 10% бис-трис SDS-PAGE гель для электрофореза (Invitrogen, комплект # NP0302BOX) с использованием XCell SureLock™ Mini-Cell (Invitrogen, комплект # EI0001). Образцы прогоняли в течение 35 минут при 200V в подвижном буфере MES-SDS (Invitrogen, комплект # NP0002). Гель был окрашен Кумасси синим R-250 (Bio-Rad, комплект # 161-0436), как показано в фиг. 3. Фракции, содержащие образцы гамма-зеина/YFP, объединяли и переносили в спин-колонку Millipore с номинальным отсечением по молекулярной массе (MWCO) 10 кДа, и центрифугировали при 4000 оборотах в минуту в течение 20 минут при 4°C, используя настольную центрифугу Eppendorf (модель 5810R). После центрифугирования для буферного обмена добавляли стерильный раствор ФБР до 15 мл. Такой процесс вращения и диафильтрации повторяли три раза. Объединенные образцы гамма-зеина/YFP переносили в гофрированную диализную трубку SnakeSkin™ (Thermo Scientific, комплект # 68100) с MWCO 10 кДа, и проводили диализ против 2 л ФБР при 4°C в течение ночи, чтобы удалить остатки имидазола. Концентрации белка определяли путем анализа Брэдфорда (Bio-Rad, комплект # 500-0006) с бычьим сывороточным альбумином (БСА) в качестве стандарта (фиг. 3).
1.4 Способ идентификации белков по «отпечаткам пептидных масс» Maldi-TPF
1 мкг белковой полоски из геля SDS-PAGE (приблизительно 1 мкг) вырезали и высушивали в 25% ацетонитриле с 12,5 мМ бикарбоната аммония в Speed-Vac. Белок расщепляли трипсином (12,5 нг/мкл) при 37°C в ходе инкубации в течение ночи. Пептиды очищали с помощью C18 Zip Tip (Millipore, комплект # 2TC18S096) согласно инструкциям изготовителя. Анализы масс-спектров проводили с использованием системы биоспектрометрии Voyager (PerSeptive Biosystems, модель DE STR), и масс-спектры собирали в режиме детекции положительных ионов (фиг. 4). Данные вводили в аналитическую программу RAWS (Proteometrics Inc) для поиска идентичности пептидов.
Пример 2
Подготовка материала одиночной растительной клетки
2.1 Клетки JTNT1
Клетки JTNT1 представляют собой фотоаутотрофные клетки, выделенные из табака. За три - четыре дня до трансформации культуральную суспензию пересевали в свежую среду во флаконе объемом 250 мл путем переноса 2 мл клеток JTNl в 40 мл среды NTIB или LSBY2, содержащей 50 нМ DAS-PMTI-1 (микроканальцевый ингибитор) и 0,5-0,1% (об./об.) диметилсульфоксида. Одиночные клетки собирали или на четвертый день или на седьмой день после обработки микроканальцевым ингибитором (как описано, например, в патенте WO 2008/083233). Клетки сохранялись в минимальной среде и 5% углекислого газа. Клетки пересевались один раз каждые 14 дней путем переноса 1 мл суспензии при OD600=3,0. Эти типы клеток использовались как целевые клетки для доставки и локализации слитого белка гамма-зеин/YFP.
2.2 Клетки моркови
Регенерируемую криосохраненную линию моркови (D2-40-018) оттаивали и культивировали в среде Линсмейера-Скуга (Linsmeier-Skoog) (LS) (описание см. Nagata T., Nemoto Y., and Hasezawa S. (1992) Int. Rev. Cyto 132, 1-30). Соли для среды приобретали в PhytoTechnology Laboratories, № в каталоге L689. Суспензию активно растущей линии ставили на одну неделю, и пересевали поддерживающую линию путем переноса 2 мл PCV (упакованный объем клетки) к 58 мл суспензии 20 LSBY2 - среды при 28°C на орбитальном шейкере (Innova-3300) при 125 оборотах в минуту под рассеянным светом при семидневном культуральном цикле. Для получения одиночных клеток 1 мл PCV морковной суспензии в постоянной фазе добавляли в 30 мл суспензии LS среды с 1 мМ колхицина (Sigma, № в каталоге C3915) и культивировали в течение 7 дней. Одиночные клетки получали из 3-7 дневных культур, и они были готовы к экспериментам с трансформацией. Одиночные клетки моркови могли сохраняться в постоянной фазе до 28 дней с помощью разведения культуры через 14 дней путем добавления 60 мл свежей жидкой среды LS BY2.
Пример 3
Обработка клеток слитым белком гамма-зеин/YFP
Клетки предварительно окрашивали 15 мкг красителя Хехст в течение 10 минут при 37°C. После инкубационного периода лишний краситель удаляли. Клетки центрифугировали со скоростью 4000 оборотов в минуту в течение 5 минут и удаляли супернатант. Клетки трижды промывали 1 мл раствора ФБР, содержащим 3% сахарозу.
Слитый белок гамма-зеин/YFP добавляли в концентрации 50 мкМ к 1 мл одиночных клеток моркови или клеток JTNT1 в пробирке Эппендорф. Пробирки помещали в шейкер при 100 оборотах в минуту при 25°C в течение 60 минут.
Пример 4
Накопление слитого белка гамма-зеин/YFP в растительных клетках
Аликвотные количества 100 мкл слитого белка/клеточной смеси забирали через 10 минут, 30 минут и 60 минут для визуализации. Клетки центрифугировали при 5000 оборотов в минуту в течение пяти минут и промывали три раза ФБР. В конце клетки ресуспендировали в 100 мкл ФБР.
Контролировали эффективность слитых пептидов в отношении интернализации и локализации в субклеточные пространства растительной клетки и тканей. Клетки визуализировали на оборудовании LSM-Leica™ TCS SP2/UV или Zeiss™ LSM. Для возбуждения YFP использовали высокую числовую апертуру (1,2-1,3) с водно-иммерсионным объективом (63x) и линию лазера 488 нм или 514 нм от ионов аргона.
Конфокальные и дифференциальные интерференционно-контрастные (DIC) изображения получали с помощью лазера при 488 нм и 405 нм и фильтров LP560 от 430 до 480 нм, соответственно. Было обнаружено, что желтый сигнал флюоресценции гамма-зеина локализуется в ядрышке и цитоплазме в одиночных клетках и моркови и клетках JTNT1 через 10 минут (показано в фиг. 5).
Пример 5
Трансформация растения Arabidopsis с пептидной и плазмидной ДНК согласно протоколу “Floral dip”
Очищенный пептид гамма-зеин смешивали с плазмидной ДНК pDAB3831, показанной в фиг. 6. Добавляли 0,25 мг пептида гамма-зеин к.5 мг плазмидной ДНК и инкубировали в воде в течение 30 минут для образования комплекса пептид/ДНК. pDAB3831 несет селектируемый маркерный ген PAT, который активируется промотором убиквитин 10 (AtUbilO) Arabidopsis и ген желтого флюоресцентного белка Philadium (PhiYFP), который активируется промотором из вируса прожилковой мозаики маниоки (CsVMV). Комплекс пептид гамма-зеин/pDAB3831 и отдельную контрольную экспериментальную плазмидную ДНК pDAB3831 единственную вводили в 4 цветочных зародыша недельного возраста Arabidopsis thatiana cv Columbia. 20 мл комплекса пептид/ДНК и 20 мл контрольной ДНК смешивали с инфильтрационной средой (5% сахароза и 0,04% Silwet-77) в стеклянном лотке в течение 30 секунд. Для трансформации растения Arabidopsis использовали инфильтрационную среду, содержащую раствор пептида/ДНК, применяя модифицированный протокол Clough и Bent (Clough SJ and Bent AF, 1998. Plant J 16:735-43). После погружения инфильтрационный комплекс сохраняли при 4°C и впоследствии использовали для погружения тех же растений через два дня, применяя тот же протокол. Повторный этап трансформации проводили для увеличения частоты трансформации.
Растения покрывали пластмассовой куполообразной крышкой, чтобы поддерживать влажность в течение 24 часов. Через 24 часа крышку снимали и растения выращивали по обычной методике в Conviron® (модели CMP4030 и CMP3244, Controlled Environments Limited, Winnipeg, Manitoba, Canada) при условиях длинного дня (16 часов освещения/8 часов темноты) при интенсивности света 120-150 мкмоль/м2/с при постоянной температуре (22°C) и влажности (40-50%), и оставляли до созревания и получения семян. Семена собирали и стерилизовали. Дополнительно, проводили яровизацию семян в течение двух дней при 4°C. Для селекции положительных трансформантов семена высевали на среде MS (0,43% смесь соли Мурашиге и Скуга (Murashige и Skoog), 2,5 мМ 2-[N-морфолино]этансульфоновая кислота, 1х витаминный раствор Гамборга (Gamborg), 0,9% бакто-агар, уровень pH 5,7-5,9), содержащей 10 мкг/мл BASTA (глуфосинат аммония) и выращивали в инкубаторе (22°C, 100 мкмоль кванта m-2 s-1) в течение 7-10 дней. Предполагаемые трансформанты идентифицировали и переносили в почву до созревания.
Пример 6
Молекулярный анализ на трансгены PAT и YFP
Экстракцию gДНК от трансгенных растений Arabidopsis и gDNA от контрольных Arabidopsis экотипа Columbia дикого типа осуществляли из материала листьев 6-недельных растений, используя комплект Plant DNAZOL (Invitrogen Inc). ПЦР амплификацию генных фрагментов PAT и YFP проводили только от трансгенных растений. 50 мкл смеси для реакции ПЦР содержали 100 нг матричной ДНК, 1х реакционный буфер ExTaq (TaKaRa Bio), 0,2 мМ дНТФ, 10 пмоль каждого праймера и 0,025 ед/мкл ExTaq. Праймеры YFP показаны как SEQ ID NO: 3 и SEQ ID NO: 4. Праймеры PAT показаны как SEQ ID NO: 5 и SEQ ID NO: 6. Применяли следующие условия циклов ПЦР: 1 цикл при 96°C в течение 5 минут, 31 цикл по следующей программе ПЦР: 94°C, 15 с; 65°C, 30 с; 72°C, 1 мин., и конечное наращивание проводили при 72°C в течение 7 минут для завершения синтеза продукта. Амплифицированные фрагменты очищали из геля, используя QIAquick™ комплект для экстракции гелей (Qiagen Inc). Фрагменты ПЦР секвенировали с помощью прямого праймера PAT (SEQ ID NO: 5) и прямого праймера YFP (SEQ ID NO: 3) с использованием передовой технологии секвенирования Sanger (MWG Biotechnologies, Inc), и анализировали последовательности с помощью программного обеспечения Sequencher™.
Пример 7
Идентификация гомологичных мотивов и мутагенез последовательности гамма-зеина для повышения эффективности транслокации белка
Были идентифицированы дополнительные мотивы, которые являются гомологичными для домена гамма-зеина, и их тестировали на внутриклеточную транслокацию белка. Эти последовательности идентифицировали из современных открытых баз данных, таких как NCBI (Национальный Центр биотехнологической информации). Последовательность гамма-зеина использовали как вход для алгоритма BLAST (способ поиска основного локального выравнивания) (Altschul et al., 1997). Используя параметры настройки по умолчанию, последовательность сравнивали с депонированными в NCBI последовательностями белков. Поиск отразил несколько гомологичных последовательностей белка. Идентифицированный мотив гамма-зеина модифицировали для повышения эффективности внутриклеточной транслокации белка. Различные остатки последовательностей VRLPPP изменяли посредством сайт-направленного мутагенеза согласно протоколам. Модифицированные последовательности гамма-зеина используют для доставки белков и ДНК в растительные клетки, применяя описанные выше протоколы.
Пример 8
Нацеливание клеточных органелл с помощью химерной последовательности гамма-зеина
Проводили дополнительные эксперименты для тестирования пептидного нацеливания клеточных органелл посредством химерных нацеливающих мотивов гамма-зеин/клеточная органелла. В конкретном примере гамма-зеин и последовательность, нацеленную на хлоропласт, сливали с N-концом флуоресцентного белка, такого как YFP. Нацеливание белка YFP на хлоропласт визуализировали путем флюоресцентной микроскопии. Дополнительные нацеливающие последовательности для разных других органелл клетки (например, митохондрий, эндоплазматической сети, ядра и т.д.), которые известны в данной области, сливали в соединении с последовательностью гамма-зеина и использовали для нацеливания на конкретную органеллу клетки. На определенную органеллу клетки нацеливали как белки, так и ДНК.
Описание настоящего изобретения дано в конкретных вариантах осуществления, но вместе с тем, настоящее изобретение можно дополнительно модифицировать в пределах объема и сущности настоящего раскрытия. Таким образом, предполагается, что используя общие принципы этого изобретения, настоящая заявка охватывает его любые варианты, применения или адаптации. Дополнительно считается, что настоящая заявка охватывает такие отступления от настоящего раскрытия, которые подпадают под известную или общепринятую практику в области, к которой относится настоящее изобретение и которые входят в границы прилагаемой формулы изобретения и ее эквивалентов.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ПЕРЕНОСА МОЛЕКУЛЯРНЫХ ВЕЩЕСТВ В КЛЕТКИ РАСТЕНИЙ | 2008 |
|
RU2495935C2 |
| ПРИМЕНЕНИЕ ДЕНДРИМЕРНОЙ НАНОТЕХНОЛОГИИ ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ | 2010 |
|
RU2571928C2 |
| КОНЪЮГАТЫ КВАНТОВОЙ ТОЧКИ С ПЕПТИДОМ-НОСИТЕЛЕМ, ПРИГОДНЫЕ ДЛЯ ВИЗУАЛИЗАЦИИ И ДОСТАВКИ МОЛЕКУЛ В РАСТЕНИЯ | 2012 |
|
RU2609637C2 |
| ПОЛУЧЕНИЕ ФУНКЦИОНАЛИЗИРОВАННОЙ ЛИНЕЙНОЙ ДНК-КАССЕТЫ И ОПОСРЕДОВАННАЯ КВАНТОВЫМИ ТОЧКАМИ/НАНОЧАСТИЦАМИ ДОСТАВКА В РАСТЕНИЯ | 2011 |
|
RU2574785C2 |
| ДОСТАВКА ЛИНЕЙНОЙ МОЛЕКУЛЫ ДНК В РАСТЕНИЯ ДЛЯ СТАБИЛЬНОЙ ТРАНСФОРМАЦИИ С ПОМОЩЬЮ ПЕГИЛИРОВАННЫХ КВАНТОВЫХ ТОЧЕК | 2011 |
|
RU2575097C2 |
| СПОСОБ И КОНСТРУКТ ДЛЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА SCBV | 2012 |
|
RU2627595C2 |
| ОПОСРЕДОВАННАЯ НАНОЧАСТИЦАМИ ДОСТАВКА СИКВЕНС-СПЕЦИФИЧНЫХ НУКЛЕАЗ | 2017 |
|
RU2664865C2 |
| ОПОСРЕДОВАННАЯ НАНОЧАСТИЦАМИ ДОСТАВКА СИКВЕНС-СПЕЦИФИЧНЫХ НУКЛЕАЗ | 2010 |
|
RU2556376C2 |
| ОПОСРЕДОВАННАЯ НАНОЧАСТИЦАМИ ДОСТАВКА СИКВЕНС-СПЕЦИФИЧНЫХ НУКЛЕАЗ | 2010 |
|
RU2612156C2 |
| КОНСТРУКЦИЯ И СПОСОБ КОНСТРУИРОВАНИЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА UBI1 | 2012 |
|
RU2639538C2 |


Изобретение относится к области биохимии, в частности к способу введения нуклеиновой кислоты в растительную клетку, имеющую интактную клеточную стенку. Также раскрыты способ экспрессии гена и способ переноса плазмидной ДНК в растительную клетку. При этом способы включают: взаимодействие пептида гамма-зеина с плазмидной ДНК для образования гамма-зеин связанной структуры; и контактирование гамма-зеин связанной структуры с интактной имеющей стенку растительной клеткой для предоставления возможности захвата гамма-зеин связанной структуры клеткой, имеющей интактную клеточную стенку. Изобретение позволяет эффективно вводить нуклеиновую кислоту, представляющую интерес, в растительную клетку, имеющую интактную клеточную стенку. 3 н. и 16 з.п. ф-лы, 6 ил., 8 пр.
1. Способ введения нуклеиновой кислоты, представляющей интерес, в растительную клетку, имеющую интактную клеточную стенку, причем указанный способ содержит:
предоставление растительной клетки, имеющей интактную клеточную стенку;
взаимодействие пептида гамма-зеина с нуклеиновой кислотой, представляющей интерес, для образования гамма-зеин связанной структуры;
приведение клетки, имеющей интактную клеточную стенку, и гамма-зеин связанной структуры в контакт друг с другом; и
предоставление возможности захвата гамма-зеин связанной структуры в клетку, имеющую интактную клеточную стенку.
2. Способ согласно п.1, в котором взаимодействие пептида гамма-зеина с нуклеиновой кислотой, представляющей интерес, содержит слияние нуклеиновой кислоты, представляющей интерес, с пептидом гамма-зеином.
3. Способ согласно п.1, дополнительно содержащий предоставление возможности захвата гамма-зеин связанной структуры компартментом растительной клетки, содержащей интактную клеточную стенку.
4. Способ согласно п.3, в котором компартмент выбирают из группы, состоящей из цитозоли, ядра, тонопласта, пластиды, этиопласта, хромопласта, лейкопласта, элайопласта, протеинопласта, амилопласта, хлоропласта и просвета двойной мембраны.
5. Способ согласно п.1, в котором растительную клетку, содержащую интактную клеточную стенку, выбирают из группы, состоящей из клеток табака, моркови, кукурузы, канолы, рапса, хлопка, пальмы, арахиса, сои, Oryza sp., Arabidopsis sp., Ricinus sp. и сахарного тростника.
6. Способ согласно п.1, в котором растительная клетка происходит из ткани, выбираемой из группы, состоящей из эмбриона, меристемы, каллюса, пыльцы, листьев, пыльников, корней, корневых кончиков, цветов, семян, стручков и стеблей.
7. Способ согласно п.1, в котором пептид гамма-зеин содержит SEQ ID NO 1.
8. Способ согласно п.1, в котором нуклеиновая кислота, представляющая интерес, содержит компонент, который выбирают из группы, состоящей из нуклеиновых кислот, молекул ДНК, РНК, iPHK, генов, плазмид, космид, YAC, ВАС и их комбинаций.
9. Способ согласно п.8, в котором нуклеиновая кислота, представляющая интерес, содержит ген.
10. Способ согласно п.9, в котором ген представляет собой ген чужого белка, ген, представляющий агрономический интерес, или маркерный ген.
11. Способ согласно п.9, дополнительно содержащий селектируемые клетки, в которые устойчиво внедрили ген.
12. Способ согласно п.11, в котором селектируемые клетки являются регенерируемыми клетками.
13. Способ согласно п.12, дополнительно содержащий регенерацию фертильного растения из регенерируемых клеток.
14. Способ экспрессии гена, причем указанный способ содержит:
предоставление растительной клетки, имеющей интактную клеточную стенку;
взаимодействие пептида гамма-зеина с геном для образования гамма-зеин связанной структуры;
приведение клетки, имеющей интактную клеточную стенку, и гамма-зеин связанной структуры в контакт друг с другом;
предоставление возможности захвата гамма-зеин связанной структуры в клетку, имеющую интактную клеточную стенку; и
экспрессию гена в растительной клетке.
15. Способ согласно п.14, в котором ген экспрессируется в хлоропласте.
16. Способ согласно п.14, дополнительно содержащий селекцию клеток, устойчиво экспрессирующих ген.
17. Способ согласно п.14, в котором пептид гамма-зеин содержит SEQ ID NO 1.
18. Способ переноса плазмидной ДНК в растительную клетку, который содержит:
взаимодействие пептида гамма-зеина с плазмидной ДНК для образования гамма-зеин связанной структуры; и
контактирование гамма-зеин связанной структуры с интактной имеющей стенку растительной клеткой при условиях, позволяющих захватывать растительную клетку пептид гамма-зеин и гена из плазмидной ДНК.
19. Способ согласно п.18, в котором пептид гамма-зеин содержит SEQ ID NO 1.
| WO2009046384 A1, 09.04.2009 | |||
| CHUGH A | |||
| et al., Cell-penetrating peptides: Nanocarrier for macromolecule delivery in living cells, IUBMB LIFE, 2010, vol | |||
| Способ крашения тканей | 1922 |
|
SU62A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Переносная мусоросжигательная печь-снеготаялка | 1920 |
|
SU183A1 |
| FERNANDEZ-CORNEADO J ET AL, POTENTIAL PEPTIDE CARRIERS: AMPHIPATHIC PROLINE-RICH PEPTIDES DERIVED FROM THE N-TERMINAL DOMAINOF -ZEIN, ANGEWANDTE CHEMIE | |||
| INTERNATIONAL | |||

