Изобретение относится к области медицины и генной терапии, и может быть использовано для лечения аллергической бронхиальной астмы и аллергического ринита путем регуляции генной экспрессии про-воспалительного цитокина интерлейкина-4 (IL-4).
Предпосылками изобретения явилось следующее. Бронхиальная астма (БА) - это хроническое воспалительное заболевание дыхательных путей, характеризующееся приступами удушья, экспираторной одышки, кашля, свистящих хрипов и эмфиземой легких (Global Inititative for Asthma). В основе этих симптомов лежат специфические (иммунные) и неспецифические механизмы, к которым относятся нарушения равновесия между отделами вегетативной нервной системы, повышенная способность клеток отвечать высвобождением медиаторов на неспецифические стимулы создавая при этом состояние гиперреактивности различных систем организма.
Многие пациенты с астмой имеют атопию, т.е. они генетически предрасположены к синтезу антител IgE после контакта с аллергенами, такими как аллерген клещей домашней пыли, пыльца растений и перхоть животных и др. Известно, что около 60-70% случаев заболевания БА связано с атопической предрасположенностью (Humbert М., Menz G., Ying S. et al. The immunopathology of extrinsic (atopic) and intrinsic (non-atopic) asthma: more similarities than differences. Immunol. Today, 1999; 20: 528-533). Повторяющиеся контакты с такими аллергенами могут индуцировать IgE-опосредованную реакцию гиперчувствительности, которая характеризуется проницаемостью кровеносных сосудов, расширением сосудов, сокращением гладкой мускулатуры, бронхоконстрикцией и воспалением дыхательных путей. При этом, дыхательные пути астматиков подвергаются структурным изменениям (ремоделированию), с изменением в эпителии, увеличением гладкой мускулатуры, отложением белков внеклеточного матрикса и гиперплазией дыхательных путей.
В настоящее время астма представляет глобальную проблему здравоохранения. По данным Всемирной организации здравоохранения (ВОЗ) на 2008 год, в мире около 300 миллионов человек, страдающих этим заболеванием, из которых примерно 5-6 млн. - в субрегионе ВОЗ Euro-C (к нему отнесена и Россия) (www.ginasthma.org). По оценкам ВОЗ, ежегодно астма обусловливает потерю 15 миллионов так называемых DALY (Disability Adjusted Life Year - дословно «год жизни, измененный или потерянный в связи с нетрудоспособностью»), что составляет 1% от общего всемирного ущерба от болезней.
По данным отчета GINA (Global Initiative for Asthma) (www.ginasthma.org), страны с наибольшей распространенностью бронхиальной астмы - это Великобритания (частота бронхиальной астмы в популяции - от 18,4% до 15,3% в зависимости от региона), Новая Зеландия - 15,1%, Австралия - 14,7%, Ирландия - 14,6%, Канада - 14,1%, Перу - 13%, Тринидад и Тобаго - 12,6%, Коста-Рика - 11,9%, Бразилия - 11,4%, США - 10,9%, Фиджи - 10,5%, Парагвай - 9,7%, Уругвай - 9,5%, Израиль - 9%, Барбадос - 8,9%, Панама - 8,8%, Кувейт - 8,5%, Украина - 8,3%, Эквадор - 8,2%, ЮАР - 8,1%, Чехия - 8%, Финляндия - 8%, Мальта - 8%, Республика Берег Слоновой Кости - 7,8%, Колумбия - 7,4%, Турция - 7,4%, Кения - 7%, Германия - 6,9%, Франция - 6,8%, Норвегия - 6,8%, Япония - 6,7%, Швеция - 6,5%, Таиланд - 6,5%, Филиппины - 6,2%, ОАЭ - 6,2%, Бельгия - 6%, Австрия - 5,8%, Испания - 5,7%, Саудовская Аравия - 5,6%, Аргентина - 5,5%, Иран - 5,5%, Эстония - 5,4%, Нигерия - 5,4%, Чили - 5,1%. Распространенность астмы в России - 5-7% (Revised Global Burden of Disease (GBD) 2008).
Смертность от БА значительно варьирует в разных странах, по-видимому, не коррелируя с распространенностью этой нозологии. Так, например, в США в 2002 г. от бронхиальной астмы умерло более 4000 чел. (14 чел. на 1 млн. населения) (National Surveillance for Asthma), а в субрегионе ВОЗ SEARO-D за тот же период смертность составила почти 77000 чел. (более 59 чел. на 1 млн). По данным специалистов, в мире от астмы умирают 250 тысяч человек в год (Revised Global Burden of Disease (GBD)).
Таким образом, БА оказывает выраженные отрицательные эффекты на общественное здоровье и экономику всего мира. В настоящее время ведутся многочисленные клинические и фундаментальные исследования, конечной целью которых является уменьшение бремени этого заболевания.
На сегодняшний день поддерживающая терапия астмы включает применение противовоспалительных лекарств, включая ингаляцию кортикостероидов, применение ингибиторов лейкотриенов (5-липоксигеназные ингибиторы и антагонисты лейкотриеновых рецепторов), и бронходилататоров. Эти препараты снижают проявления симптомов астмы и являются достаточными для лечения значительной доли пациентов, но они не воздействуют на первопричины патогенетических звеньев заболевания. Для лечения атопической БА успешно применяют аллерген-специфическую иммунотерапию (АСИТ), которая является единственным патогенетическим видом лечения атопических аллергических болезней, влияющим на саму природу заболевания. Однако АСИТ имеет ряд недостатков, основным из которых является опасность возникновения местных и системных реакций на проводимое лечение за счет сохранения B-клеточных эпитопов в препаратах нативного аллергена. B-клеточные эпитопы аллергена обусловливают IgE-опосредованное высвобождение медиаторов (гистамина, лейкотриенов и др.) из клеток-мишеней аллергии (в основном тучных клеток, базофилов крови) в основном на начальных этапах лечения, что приводит к серьезным осложнениям лечения (в ряде случаев с угрозой для жизни) примерно у 3,7% пациентов (156. Williams А.Р., Krishna М.Т., Frew A.J. The safety of immunotherapy // Clin Exp Allergy. - 2004. -34. - P. 513-514).
Распространенность аллергического ринита в популяции человека в 3-5 раз выше, чем аллергической бронхиальной астмы, однако выявляемость данного заболевания низкая по причине редких обращений пациентов с соответствующими симптомами к врачам-специалистам. Стоит отметить, что развитие аллергической бронхиальной астмы в подавляющем большинстве случаев начинается с развития аллергического ринита, поскольку данные патологии имеют сходный молекулярный механизм. Это подтверждается и тем фактом, что 75-90% пациентов с аллергической бронхиальной астмой, страдают также и от аллергического ринита.
Таким образом, разработка новых подходов к лечению и профилактике аллергической бронхиальной астмы и ринита остается актуальной задачей.
Один из базовых подходов к лечению аллергической бронхиальной астмы и ринита это анти-IL-4 терапия, где применяются различные ингибиторы как самого IL-4, так и компонентов его сигнального пути. Еще два десятилетия назад было предположено, что IL-4 играет ключевую роль в патогенезе астмы (Steinke J.W. and Borish L. Th2 cytokines and asthma. Interleukin-4: its role in the pathogenesis of asthma, and targeting it for asthma treatment with interleukin-4 receptor antagonists // Respir. Res. 2001. V. 2. P. 66-70). Было показано, что астматики имеют повышенный уровень белка IL-4 как в сыворотке крови, так и в бронхоальвеолярном лаваже (БАЛ) (Daher S., Santos L.М., Sole D., De Lima M.G., Naspitz С.K., Musatti C.C. Interleukin-4 and soluble CD23 serum levels in asthmatic atopic children // J. Investig. Allergol. Clin. Immunol. 1995. V. 5. P. 251-254), а также увеличенный уровень mRNA IL-4 и самого белка в бронхиальной биопсии пациентов (Humbert М., Durham S.R., Ying S., Kimmitt P., Barkans J., Assoufi В., Pfister R., Menz G., Robinson D.S., Kay A.B., Corrigan C. J. IL-4 and IL-5 mRNA and protein in bronchial biopsies from patients with atopic and nonatopic asthma: evidence against "intrinsic" asthma being a distinct immunopathologic entity // Am. J. Respir. Crit Care Med. 1996. V. 154. P. 1497-1504). Также важным доказательством участия IL-4 в патогенезе бронхиальной астмы являлся тот факто, что аэрозольное введение IL-4 индивидуумам с легкой астмой индуцировало у них гиперреактивность бронхов (ГРБ) и эозинофилию (Shi H.Z., Deng J.M., Xu Н., Nong Z.X., Xiao C.Q., Liu Z.M., Qin S.M., Jiang H.X., Liu G.N., Chen Y.Q. Effect of inhaled interleukin-4 on airway hyperreactivity in asthmatics // Am. J. Respir. Crit Care Med. V. 157. 1998. P. 1818-1821).
Для выяснения точной роли IL-4 в патогенезе астмы были использованы экспериментальные модели на животных, таких как мыши, крысы, морские свинки и приматы. Особый вклад в понимание биологической роли IL-4 внесли эксперименты с трансгенными мышами, у которых был полностью инактивирован ген il-4. Стандартное моделирование признаков бронхиальной астмы на таких IL-4-дефектных мышах показало, что происходит редукция аллергического воспаления, что выражалось в снижении количества эозинофилов в ткани легких и перибронхиального воспаления в сравнении с мышами, имеющих функциональный ген il-4. Кроме того, IL-4-дефектные мыши не продуцировали аллерген-специфические IgE антитела (Ат) и не развивали ГРБ (Brusselle G.G., Kips J.C., Tavernier J.H., van der Heyden J.G., Cuvelier C.A., Pauwels R.A., Bluethmann H. Attenuation of allergic airway inflammation in IL-4 deficient mice // Clin. Exp. Allergy. 1994. V. 24. P. 73-80).
Экспериментально установлено, что в патогенезе бронхиальной астмы IL-4 обеспечивает дифференциацию про-аллергические Th2-клеток из нативных Th0-клеток, индуцирует переключение синтеза B-клетками антител с IgM на про-аллергические IgE. В последствии синтезированные IgE антитела способствуют высвобождению тучными клетками и базофилами крови про-воспалительных медиаторов гистамина, лейкотриенов, простагландинов. В нескольких независимых исследованиях было установлено, что IL-4, вместе с IL-13, увеличивают экспрессию в эндотелии адгезионной молекулы VCAM-1, которая способствует миграции про-воспалительных клеток эозинофилов, T-лимфоцитов, моноцитов и базофилов, привлекаемых хемокинами тучных клеток, тем самым IL-4 вносит вклад в локальное воспаление в дыхательных путях (Schleimer R.P., Sterbinsky S.A., Kaiser J., Bickel C.A., Klunk D.A., Tomioka K., Newman W., Luscinskas F.W., Gimbrone M.A., Jr., McIntyre B.W.. IL-4 induces adherence of human eosinophils and basophils but not neutrophils to endothelium. Association with expression of VCAM-1 // J. Immunol. 1992. V. 148. P. 1086-1092).
Что касается источников IL-4 в организме, то этот цитокин продуцируется Th2 клетками, но может высвобождаться также и тучными, базофилами, эозинофилами и альвеолярными макрофагами (Gessner А., Mohrs K., Mohrs M. Mast cells, basophils, and eosinophils acquire constitutive IL-4 and IL-13 transcripts during lineage differentiation that are sufficient for rapid cytokine production // J. Immunol. 2005. V. 174. P. 1063-1072). IL-4 специфически связывается с IL-4Rα цепью, которая экспрессируется на T-лимфоцитах, B-лимфоцитах, эозинофилах, фагоцитах, эндотелиальных клетках, легочных фибробластах, эпителиальных клетках бронхов, и клетках гладкой мускулатуры (Kotsimbos Т.С., Ghaffar О., Minshall Е.М., Humbert М., Durham S.R., Pfister R., Menz G., Kay A.B., Hamid Q.A. Expression of the IL-4 receptor alpha-subunit is increased in bronchial biopsy specimens from atopic and nonatopic asthmatic subjects // J. Allergy Clin. Immunol. 1998. V. 102. P. 859-866).
IL-4Rα вовлечен в образование трех рецепторных комплексов: IL-4Rα/(γc)/IL-4 (Рецептор I типа), IL-4Rα/IL-13Rα1/IL-4 и IL-4Rα/IL-13Rα1/IL-13 комплекс (рецепторы II типа). Рецептор I типа связывается исключительно с IL-4, комплекс II типа связывается как с IL-4, так и с IL-13. Это объясняет перекрывание некоторых биологических функций IL-4 и IL-13. Рецептор I типа типично встречается у гематопоэтических клетках и вовлечен в развитие Th2-ответа, тогда как рецептор II типа обнаруживается как у гематопоэтических так и у негематопоэтических клеток. Передача сигнала IL-4, и IL-13, посредством рецептора II типа на негематопоэтических клетках дыхательных путей, таких как клетки гладкой мускулатуры и эпителиальные клетки, может напрямую индуцировать ГРБ и секрецию слизи (Kuperman D.A., Schleimer R.P. Interleukin-4, interleukin-13, signal transducer and activator of transcription factor 6, and allergic asthma // Curr. Mol. Med. 2008. V. 8. P. 384-392).
Связывание лиганда (IL-4 или IL-13) с I или II типом рецептора инициирует активацию протеинкиназ семейства Janus (Jak). В частности, IL-4Rα, ус и IL-13Rα1 активируют Jak1, Jak3 и Tyk2 соответственно. Это, в свою очередь, приводит к активации нескольких внутриклеточных сигналов посредством фосфорилирования специфических тирозиновых остатков в цитоплазматическом домене IL-4Rα, и к фосфорилированию фактора транскрипции - белка Stat6. Фосфорилированный Stat6 димеризуется и транслоцируется в ядро, где связывается со специфическими участками ДНК и индуцирует так называемые IL-4/IL-13-стимулируемые гены, вовлеченные в Th2 ответ, переключение синтеза с IgM на IgE, ГРБ и продукцию слизи. Более того субъединица IL-13Rα1 в комплексе II типа может также инициировать другие сигнальные пути, привлекая другие белки Stat (например, Stat3, Stat1). При этом происходит развитие Stat6-независимой, но IL-13-зависимой ГРБ, что было показано на мышиных моделях хронической бронхиальной астмы.
Таким образом, экспериментально доказана роль гена IL-4 в формировании бронхиальной астмы, что делает данный ген привлекательной мишенью при разработке новых лекарственный препаратов.
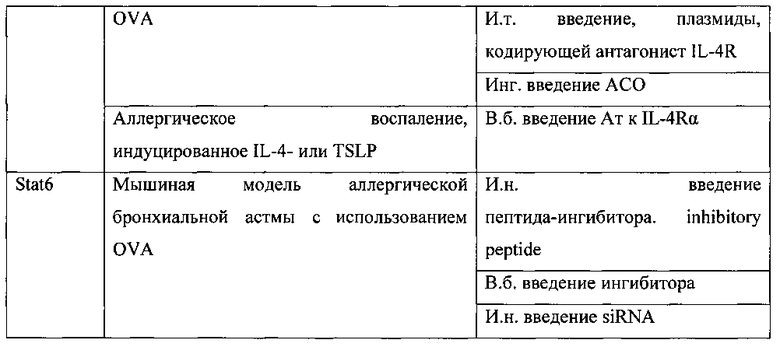
К настоящему времени создан ряд лекарственных средств и терапевтических подходов для подавления данного гена, которые апробированы в доклинических и клинических исследованиях. Так, например, растворимый белок-рецептор - sIL-4R является естественным белком - секретируемой формой рецептора IL-4R, который содержит внеклеточную часть цепи IL-4Rα, но утративший трансмембранный и цитоплазматический домены, благодаря этому sIL-4R способен связываться с IL-4, но при этом не активирует сигнальный путь, т.е. функционирует как «ловушка», связывающаяся с IL-4 и нейтрализующая его (табл. 1, табл. 2). В экспериментах на мышиной модели астмы, sIL-4R значительно ингибировал ГРБ и продукцию IgE при введении в период сенсибилизации. Более того, когда sIL-4R вводился во время стадии провокации он предотвращал позднюю фазу воспаления в легких, что выражалось в репрессии VCAM-1, инфильтрации эозинофилов в дыхательные пути и снижении гиперсекреции слизи, но при этом не происходило улучшения ГРБ, которая оставалась высокой). Это исследование показало, что блокирование IL-4 может иметь клинический эффект, даже после сенсибилизации организма к аллергену (Renz Н., Bradley K., Enssle K., Loader J.Е., Larsen G.L., Gelfand E.W. Prevention of the development of immediate hypersensitivity and airway hyperresponsiveness following in vivo treatment with soluble IL-4 receptor. Int. Arch. Allergy Immunol. 1996. V. 109. P. 167-176).
Терапевтический потенциал ингаляции рекомбинантного человеческого sIL-4R (Nuvance) был многообещающим на начальной фазе клинических испытаний (Borish L.C., Nelson Н.S., Corren J., Bensch G., Busse W.W., Whitmore J.В., Agosti J.M. Efficacy of soluble IL-4 receptor for the treatment of adults with asthma // J. Allergy Clin. Immunol // 2001. V. 107. P. 963-970). С пролонгированным периодом полу-жизни sIL-4R (примерно 5 дней), применялся для ингаляционной терапии курсом один раз в неделю. Фазы I/II испытаний с прерыванием приема кортикостероидов пациентами, показало, что одиночное ингаляционное введение sIL-4R было безопасным и эффективным при астме средней тяжести, не наблюдалось токсического эффекта, происходило значительное улучшение показателя объема форсированного выдоха в 1 секунду (forced expiratory volume in 1 second (FEV1)), пациентам требовалось принимать меньше β2-агонистов, и снижался уровень воспаления в легких, измеренный по количеству выдыхаемого оксида азота (NO).
Последующие исследования с многократным введением sIL-4R в течение 12 недель по оценке длительной безопасности и эффективности также показали многообещающие результаты при лечении астмы средней тяжести (Immunex Phase II Efficacy Study, 2005). Однако дальнейшие клинические испытания, на астматиках, которые принимали только β2-агонисты, не показали клинического эффекта на FEV1 или симптомы астмы. Отсутствие клинических эффектов наблюдалось и в другом исследовании, когда пациенты принимали sIL-4R при уменьшении приема кортикостероидов. В последующем было показано, что sIL-4Rα стабилизирует взаимодействие IL-13 со своим рецептором, тем самым активируя сигнальный путь IL-13. Это может быть объяснением почему клинические испытания sIL-4R потерпели неудачу (Andrews A.L., Holloway J.W, Holgate S.Т., Davies D.E. IL-4 receptor alpha is an important modulator of IL-4 and IL-13 receptor binding: implications for the development of therapeutic targets // J. Immunol. 2006. V. 176. P. 7456-7461).
Другой подход для блокирования IL-4 это использование гуманизированных моноклональных антител (мАт), например, препарат Pascolizumab, (табл. 1, табл. 2). Препарат Pascolizumab представляет собой мышиные мАт (3B9) со специфичностью к человеческому IL-4, которые были гуманизированы с целью снижения иммуногенности. На мышиных моделях астмы было показано снижение уровня IgE и ГРБ после введения анти-IL-4-мАт. (Zhou C.Y., Crocker I.С, Koenig G., Romero F.A., Townley R.G. Anti-interleukin-4 inhibits immunoglobulin E production in a murine model of atopic asthma // J. Asthma. 1997. V. 34. P. 195-201). Исследования in vitro показали, что Pascolizumab нейтрализовал активность IL-4 в клеточных линиях человека и ингибировал Th2-процесс, например, синтез IL-5, и продукцию IgE. Изучение фармакокинетики и хронической безопасности Pascolizumab in vivo на обезьянах показало хорошую переносимость без побочных эффектов (Hart Т.K., Blackburn М.N., Brigham-Burke М., Dede K., Al-Mahdi N., Zia-Amirhosseini P., Cook R.M. Preclinical efficacy and safety of pascolizumab (SB 240683): a humanized antiinterleukin-4 antibody with therapeutic potential in asthma // Clin. Exp. Immunol. 2002. V. 130. P. 93-100).
Фаза I клинических испытаний при однократном внутривенном введении пациентам с астмой средней тяжести также показала хорошую переносимость препарата, который имел период полу-жизни более чем 2 недели (Shames R.S., Vexler V., Lane N., McClellan M., Shi J., Keller S., The safety and pharmacokinetics of SB240683 (anti-IL-4 humanized monoclonal antibody) in patients with mild to moderate asthma. J. Allergy Clin. Immunol. 2001. V. 163.). Однако последующая II фаза клинических испытаний была прекращена, т.к. Pascolizumab не обеспечивал клинического эффекта даже при многократном введении астматикам, не принимающим кортикостероиды (PDL BioPharma, Inc. Pilot Study, 2008).
Альтернативой использованию моноклональных антител (мАт) служит стратегия применения антисмысловых олигонуклеотидов (АСО). АСО осуществляют деградацию молекул мРНК гена с комплементарной последовательностью. Поэтому в отличие от мАт, которые нейтрализуют IL-4 на белковом уровне, АСО осуществляют сайленсинг на уровне мРНК. Этот подход может быть интересной альтернативой, т.к. несколько копий белка может считываться с одной молекулы мРНК. На крысиной модели астмы с помощью рекомбинантного адено-ассоциированного вируса осуществлялось системное введение АСО, подавляющих IL-4. В результате наблюдали значительное снижение уровня IL-4 в БАЛ, аллергического воспаления и ремоделирования (Cao Y., Zeng D., Song Q., Сао С, Xie M., Liu X., Xiong S., Xu Y., Xiong W. The effects of antisense interleukin-4 gene transferred by recombinant adeno-associated virus vector on the airway remodeling in allergic rats // J. Asthma. 2010. V. 47. P. 951-958). К настоящему моменту данный подход еще не опробован в клинических испытаниях.
Перекрывающиеся биологические функции IL-13, объясняющееся способностью данного цитокина связываться с рецепторными цепями IL-4, свидетельствуют о перспективности стратегии по его блокированию с помощью мАт. Такие мАт против IL-13 использовались как при профилактическом, так при терапевтическом режиме введения на мышиной модели бронхиальной астмы. Анти-IL-13-мАт ингибировали аллергическое воспаление, гиперплазию бокаловидных клеток и ремоделирование у мышей. Однако, тогда как при профилактическом протоколе введения мАт снижалась ГРБ, при терапевтическом введении препарат не ингибировал этот важный клинический признак бронхиальной астмы (Tomlinson K.L., Davies G.С, Sutton D.J., Palframan R.Т. Neutralisation of interleukin-13 in mice prevents airway pathology caused by chronic exposure to house dust mite // PLoS. One. 2010. V. 5).
Некоторые успешные исследования на обезьянах и овцах позволили начать клинические испытания анти-IL-13 Ат субкласса IgG1 (IMA-638 и IMA-026) (Bree A., Schlerman F.J., Wadanoli М., Tchistiakova L., Marquette K., Tan X.Y., Jacobson B.A., Widom A., Cook T.A., Wood N., Vunnum S., Krykbaev R., Xu X., Donaldson D.D., Goldman S.J., Sypek J., Kasaian M.T. IL-13 blockade reduces lung inflammation after Ascaris suum challenge in cynomolgus monkeys // J. Allergy Clin. Immunol. 2007. V. 119. P. 1251-1257). IMA-638 и IMA-026, связываются с разными эпитопами IL-13 и ингибируют аллерген-индуцированный ответ. Антитело IMA-638 блокирует связывание IL-13 с цепью IL-4Rα, а IMA-026 с цепями IL-13Rα1 и IL-13Rα2. Они были тестированы на пациентах с мягкой астмой (табл. 2). Показано, что подкожное введение IMA-638 и IMA-026 на день 1 и 8 не влияло на ГРБ и эозинофилию в слюне, но IMA-638 кратковременно снижал раннюю и позднюю фазы аллергического ответа (Gauvreau G.М., Boulet L.Р., Cockcroft D.W., Fitzgerald J.М., Carlsten С., Davis В.Е., Deschesnes F., Duong M., Durn В.L., Howie K.J., Hui L., Kasaian M.Т., Killian K.J., Strinich Т.X., Watson R.M., Zhou Y N. S., Raible D., O′Byrne P.M. Effects of interleukin-13 blockade on allergen-induced airway responses in mild atopic asthma // Am. J. Respir. Crit Care Med. 2011. V. 183. P. 1007-1014). Однако, при более длительном введении мАт пациентам с неконтролируемой астмой, (12 недель) которые получали кортикостеройды, не было эффекта IMA-638 на симптомы БА в сравнении с плацебо (Study Evaluating the Effect of IMA-638, 2009).
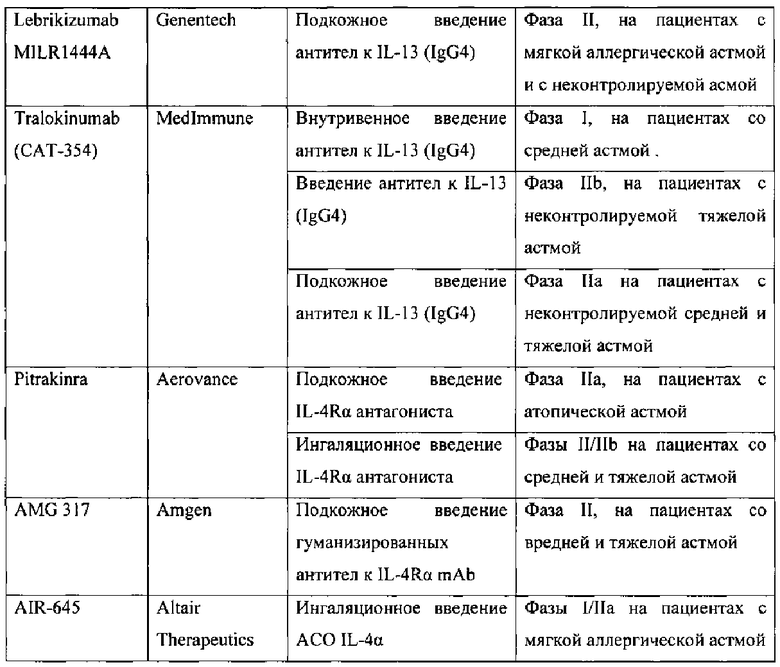
Другой препарат - Lebrikizumab - представляет собой гуманизированные мАт субкласса IgG4, также направленные против IL-13. Lebrikizumab тестировался на пациентах как с мягкой астмой, так и с неконтролируемой астмой посредством подкожного введения в течение нескольких месяцев (вплоть до 6 месяцев) (Genentech, A Study to Evaluate MILR1444A, 2010; Genentech, A Study of Lebrikizumab, 2011; Genentech, A Study to Evaluate Lebrikizumab (MILR1444A) in Adult Patient, 2011). Согласно рабочей гипотезе анти-IL-13 терапия могла бы быть полезной для пациентов с повышенной активностью IL-13. Подгруппа пациентов у которых был доказан Th2 статус (повышенный уровень IgE и эозинофилов в крови, увеличенный уровень периостина - белка, секретируемого в ответ на IL-13) (Corren et al., 2011; Genentech, A Study of Lebrikizumab (MILR1444A) in Adult Patients 2011) хорошо отвечала на препарат. У таких пациентов Lebrikizumab значительно улучшал форсированный объем выдоха (FEV1) на 5.5% к 12-ой неделе. В связи с этим измерение уровня периостина у астматиков могло бы использоваться как биомаркер для поиска пациентов, отвечающих на Lebrikizumab. Несмотря на успех препарата на ограниченной группе пациентов Lebrikizumab не приводил к значительному улучшению большинства симптомов БА (Corren J., Lemanske R.F., Hanania N.A., Korenblat P.E., Parsey M.V., Arron J.R., Harris J.M., Scheerens H., Wu L.C, Su Z., Mosesova S., Eisner M.D., Bohen S.P., Matthews J.G. Lebrikizumab treatment in adults with asthma // N. Engl. J. Med. 2011. V. 365. P. 1088-1098).
Еще один препарат, основанный на гуманизированных мАт субкласса IgG4 - Tralikinumab (САТ-354) - показал эффективность в доклинических моделях, а в клинических испытаниях был безопасен и хорошо переносим как для здоровых индивидуумов, так и для астматиков. Препарат Tralikinumab в настоящий момент находиться на II фазе клинических испытаний (Medimmune, A Phase 2b, Randomized, Double-blind Study 2012; Medimmune, Study to Evaluate the Safety and Efficacy 2011).
Одна из причин неудач анти-IL-4 терапии - это взаимозаменяемость IL-4 и интерлейкина-13 (IL-13), когда при подавлении только IL-4 его функцию может выполнять IL-13. Потому некоторые исследования были сфокусированы на блокировании цепи IL-4Rα, которая входит в состав рецепторов как для IL-4, так и IL-13 и инициирует Stat6-опосредованный сигнальный путь. Для инактивации IL-4Rα разработано несколько подходов: использование IL-4-мутеина (Pitrakinra), использование мАт против IL-4Rα (AMG317) и не так давно стали применять АСО (AIR-645).
Pitrakinra - это рекомбинантная форма человеческого IL-4 с двумя функциональными мутациями в позиции 121 (замена аргинина на аспарагиновую кислоту) и 124 (замена тирозина на аспарагиновую кислоту). Была продемонстрирована высокая афинность связывания Pitrakinra с IL-4Rα цепью, в результате чего препарат действует как конкурентный антагонист и блокирует связывание обоих цитокинов (IL-4 IL-13) с IL-4Rα. Эффективность такого подхода изначально оценена на мышиной модели астмы, где показано ингибирование продукции Th2 цитокинов, эозинофильного воспаления, гиперплазии бокаловидных клеток, ГРБ и специфического IgE. У обезьян, двойной антагонизм IL-4/IL-13 посредством Pitrakinra снижал ГРБ и имел тенденцию к снижению эозинофильного воспаления (Tomlinson K.L., Davies G.С., Sutton D.J., Palframan R.Т. Neutralisation of interleukin-13 in mice prevents airway pathology caused by chronic exposure to house dust mite // PLoS. One. 2010. V. 5).
Многообещающие результаты в доклинических испытаниях были основаниями для проведения клинических исследований Pitrakinra. Два независимых клинических исследования фазы IIa изучили эффект препарата (при подкожном и аэрозольном введении) на позднюю фазу аллергического ответа у атопических астматиков. Независимо от пути введения, препарат Pitrakinra улучшал FEV1, в то же время ингаляционное введение имело большую эффективность в сравнении с подкожным (Wenzel S., Wilbraham D., Fuller R., Getz E.В., Longphre M. Effect of an interleukin-4 variant on late phase asthmatic response to allergen challenge in asthmatic patients: results of two phase 2a studies // Lancet. 2007. V. 370. P. 1422-1431). На текущий момент описана фармакодинамика, фармакокинетика, клиническая эффективность и безопасность препарата (Antoniu S.A., Cojocaru I. Pitrakinra for asthma // Expert. Opin. Biol. Ther. 2010. V. 10. P. 1609-1615). В фазе IIb клинических исследований на астматиках со средней и тяжелой формой астмы изучалась способность Pitrakinra снижать осложнения при постепенном уменьшении приема β-агонистов и кортикостеройдов. Как оказалось, Pitrakinra не имел значительного эффекта в общей популяции астматиков со средней и тяжелой формой, но у подгруппы пациентов с эозинофильной астмой снижались осложнения на 37% (Aerovance Inc. Press release. Phase 2b Clinical Trial, 2010; Aerovance, Inc. A Study of the Treatment-Sparing Effects, 2011). Подобные результаты были получены и при испытании препарата Mepolizumab (мАт против IL-5), который был неэффективен в смешанной (гетерогенной) популяции астматиков, но показал свою эффективность в подгруппе пациентов с тяжелой эозинофильной астмой.
AMG317 - это гуманизированные мАт против IL-4Rα человека, предотвращают связывание как IL-4 так и IL-13 с IL-4Rα (фиг. 1). Доклинические исследования на мышах показали, что AMG317 снижает воспаление дыхательных путей, ГРБ и гиперплазию бокаловидных клеток. В фазах I и II клинических испытаний изучена фармакокинетика и фармакодинамика AMG317 при однократном и многократном введениях посредством внутривенного и подкожного путей. Длительное подкожное введение AMG317 в течение нескольких месяцев показало хорошую переносимость препарата. Фаза II исследований на пациентах со средней и тяжелой астмой выявила отсутствие клинической эффективности во всех группах пациентов (Amgen Inc. Phase II Study, 2009).
Снижение экспрессии IL-4Rα посредством антисмысловых олигонуклеотидов (АСО) приводило к подавлению признаков БА на мышиных моделях. В фазе I клинических исследований была показана хорошая переносимость препарата здоровыми индивидуумами и пациентами с мягкой астмой, что дает возможность его использования для ежедневной терапии (Altair Therapeutics, Inc. A Phase 1 Study 2011; Seguin and Ferrari 2009). Однако в фазе II клинических испытаний препарат не проявил достаточного клинического эффекта, в частности на показатель FEV1, чтобы оправдать дальнейшее развитие (Altair Therapeutics, Inc. Study Evaluating the Effects of AIR645, 2010).
Другим подходом блокирования биологической активности обоих цитокинов IL-4 и IL-13, является подавление фактора Stat6, который необходим для транскрипции генов, отвечающих на IL-4 и IL-13. В доклинических исследованиях использовались несколько разных подходов для подавления Stat6, а именно: использование ингибирующего пептида (AS1517499), и молекул малых интерферирующих РНК (миРНК). Применение обоих подходов в доклинических исследованиях было успешным, т.к. на мышиных моделях продемонстрировано снижение аллергического воспаления дыхательных путей, снижение гиперплазии бокаловидных клеток, продукции слизи и ГРБ (Tian X.R., Tian X.L., Во J.P., Li S.G., Liu Z.L., Niu B. Inhibition of allergic airway inflammation by antisense-induced blockade of STAT6 expression // Chin Med. J. (Engl.) 2011. V. 124. P. 26-31) (табл. 1). Хотя данные исследования подтверждают перспективность мишени Stat6 для лечения астмы, к настоящему моменту нет клинических данных.


Несмотря на существование разных способов антиастматической терапии, все еще существует большое количество пациентов с низким качеством жизни по причине неконтролируемой астмы. Поэтому разработка новых дополнительных способов лечения заболевания остается актуальной. За последние 15 лет получены новые данные о молекулярных механизмах БА, усовершенствована технология получения мАт и открыто явление РНК интерференции. Все это привело к созданию концепции анти-IL-4 терапии для лечения БА. Хотя блокирование IL-4 в доклинических исследованиях показало многообещающие результаты, однако не были оправданы ожидания от клинических испытаний. Этим неудачам есть несколько объяснений. Во-первых, информация о роли IL-4 в астме и в Th2-дифференциации постоянно дополняется. Изначально сигнальный путь IL-4/IL-4Rα/Stat6 считался ключевым в развитии CD4+ T-клеток, a IL-4 единственным инициатором Th2-дифференциации. Сейчас установлено, что Th2-дифференциация представляет собой более сложный процесс, а сигнальный путь IL-4/IL-4Rα/Stat6 может быть необязательным, т.к. открыты иные пути Th2-дифференциации (например, пути опосредуемые IL-25, IL-33). Поэтому стратегия подавление исключительно IL-4 оказалась не оправданной. Во-вторых, блокирование IL-4 для предотвращения Th2-дифференциации может быть успешным в краткосрочных мышиных моделях при терапевтическом введении анти-IL4 препаратов, но в клинической практике пациенты с астмой уже имеют сформированный Th2-ответ, поэтому не удается экстраполировать результаты доклинических исследований на человека.
Обе причины неудачных клинических испытаний в той или иной степени преодолимы. Во-первых, в настоящее время астма у человека больше не рассматривается как исключительно аллергическое Th2-опосредованное, эозинофильное заболевание, но подразделяется на отдельные клинические и молекулярные фенотипы. Такое фенотипическое деление астмы позволит оптимально подбирать группу пациентов, для которых анти-IL-4 терапия будет эффективной. Во-вторых, для корректного проведения доклинических испытаний необходимы более адекватные модели на животных. Учитывая многообразие фенотипов астмы, сложилась необходимость в создании разнообразных мышиных моделей, например, не только с эозинофильным, но и с нейтрофильным воспалением, а также необходимость использования причинно-значимых аллергенов при моделировании. Более того исследуемые препараты в доклинических испытаниях необходимо вводить не в профилактическом, а в терапевтическом режиме, т.е. после того как у животных разовьются признаки БА. Для этих целей разрабатываются хронические модели астмы.
Таким образом, будущее развитие анти-IL-4 терапии будет связано с использованием адекватных моделей для доклинических исследованиях, а также с тщательным отбором респондентов из гетерогенной популяции астматиков для клинических исследований.
Недавнее открытие явления интерференции РНК (РНКи) - одного из основных механизмов посттранскрипционной регуляции генов, дало возможность использовать данный феномен для создания новых лекарственных средств, способных эффективно и безопасно подавлять экспрессию генов-мишеней. РНКи позволяет специфически «выключать» экспрессию генов-мишеней с помощью молекул siRNA (small interfering RNA - малых интерферирующих РНК). Для эффективной доставки siRNA в клетки-мишени применяют специальные молекулы-носители, формирующие комплексы с siRNA. Комплексование приводит к компактизации молекул РНК, их защите от деградации клеточными РНКазами, что в сочетании с позитивным суммарным зарядом, позволяет им эффективно проникать внутрь клетки и достигать нужной мишени.
Главными преимуществами использования препаратов на основе РНКи являются высокая специфичность подавления экспрессии генов, вовлеченных в патогенез, а также высокая эффективность их подавления (до 90%), т.к. вводимые siRNA способны действовать в крайне низких концентрациях. Кроме того, привлекательной является сравнительная дешевизна методики. Синтез олигонуклеотидов в настоящее время вполне доступен и прост в масштабировании. Этот факт дает препаратам, созданным на базе, siRNA, важное конкурентное преимущество, например, по сравнению с моноклональными антителами.
Известны изобретения для лечения аллергических заболеваний, включая аллергическую бронхиальную астму и ринит путем блокирования экспрессии гена про-воспалительного цитокина IL-4.
Известно применение моноклональных и биспецифических антител, которые специфически связываются с IL-4 и/или IL-13 (патент ЕР 2574630). Однако эффективность такого применения не высока, поскольку моноклональные антитела действуют на белковом (т.е. посттрансляционном уровне), в то же время препараты на основе интерференции РНК, к таковым относится и заявляемое изобретение, действуют на уровне мРНК (т.е. на посттранскрипционном уровне), что позволяет более эффективно блокировать ген-мишень за счет того, что с одной копии мРНК может синтезироваться более 1000 копий белка.
Помимо ингибирования путем непосредственного связывания цитокина, можно снижать его активность посредством блокирования соответствующего рецептора. Известно, что сигнальный путь IL-4 опосредован гетеродимерным комплексом двух белков рецептора: IL-4Rα и γ-цепи IL-2-рецептора. Создание моноклонального антитела (imAb) Х2/45, направленного против внеклеточного домена IL-4Rα позволило предположить его для ингибирования IL-4-опосредованных реакций, в том числе и для создания нового способа терапии бронхиальной астмы. Этот подход реализован в одном из изобретений (патент WO 2009121847), который относится к области применения антител или антигенсвязывающих фрагментов, специфичных для человеческого рецептора к IL-4. В данном изобретении также используются моноклональные антитела со всеми присущими им недостатками и ограничениями, описанными выше.
Известно, что связывание IgE с FcεRI-рецепторами эффекторных клеток (тучные клетки, базофилы) индуцирует высвобождение медиаторов воспаления (гистамин, лейкотриены) и вызывает острые симптомы аллергического воспаления. Таким образом, взаимодействие IgE-FcεRI является одним из ключевых в патогенезе бронхиальной астмы. Этот факт был использован при создании изобретения для терапии аллергических воспалений в том числе бронхиальной астмы. Изобретение заключается в использовании Fab фрагмента антитела, который ингибирует IgE-FcεRI взаимодействия (патент WO 2002079257).
Открытие РНК интерференции дало новые возможности для регуляции экспрессии патогенетически-значимых генов, в том числе и генов, вовлеченных в инициацию и развитие БА. РНКи активируется молекулами siRNA, которые имеют нуклеотидную последовательность комплементарную гену-мишени. Несмотря на сравнительно недавнее открытие РНКи уже имеются изобретения по применению данного подхода к терапии бронхиальной астмы. Известно, что CD23 участвует в активации T-лимфоцитов, которые в свою очередь вовлечены в патогенез бронхиальной астмы. Этот факт был использован при создании изобретения (патент US 8461125) для лечения бронхиальной астмы, заключающееся в использовании молекул siRNA для блокирования гена CD23.
Еще одно изобретение заключается в использовании молекул siRNA для снижения синтеза антител класса IgE, как основных про-аллергических медиаторов (патент US 20110112169).
Однако эти изобретения (US 8461125, US 20110112169) направленны на подавление синтеза антител IgE, или препятствие связывания IgE антител с клетками мишенами, в то время как заявляемое изобретение направлено на подавление IL-4, с активации которого запускаются данные процесс, а такде патогенез бронхиальной астмы. В связи с этим ожидается, что заявляемое изобретение будет более эффективно подавлять признаки бронхиальной астмы.
Таким образом, прямых аналогов заявляемого изобретения, представляющего собой композицию, состоящую из катионного дендримерного пептида - LTP и двух молекул siRNA против гена IL-4 для подавления аллергического воспаления путем блокировки экспрессии гена про-воспалительного цитокина IL-4 не выявлено.
Задачей предлагаемого изобретения является создание малотоксичного средства для лечения бронхиальной астмы.
Техническим результатом предлагаемого изобретения является создание малотоксичной композиции, подавляющая экспрессию про-воспалительного гена IL-4.
Указанная задача решается, а технический результат достигается, за счет того, что композиция для подавления экспрессия гена цитокина интерлейкина-4 (IL-4) посредством механизма интерференции РНК состоит из катионного дендримерного пептида LTP с формулой (Arg)8(Lys)4(Lys)2Lys-Ala-Cys-NH2, выступающего в качестве носителя, и двух молекул siRNA с формулами 5-UUGAUGAUCUCCUGUAAGGtt-3 (SEQ ID NO 57) и 5-AAAGAUGUCUGUUACGGUCtt-3 (SEQ ID NO 59).
При отношении количества положительных зарядов пептида к количеству отрицательных зарядов нуклеиновых кислот от 24 до 96 композиция для подавления экспрессия гена цитокина интерлейкина-4 (IL-4) способна эффективно трансфецировать различные типы клеток.
При LD50 равным 325 мг/кг композиция для подавления экспрессия гена цитокина интерлейкина-4 (IL-4) является малотоксичной.
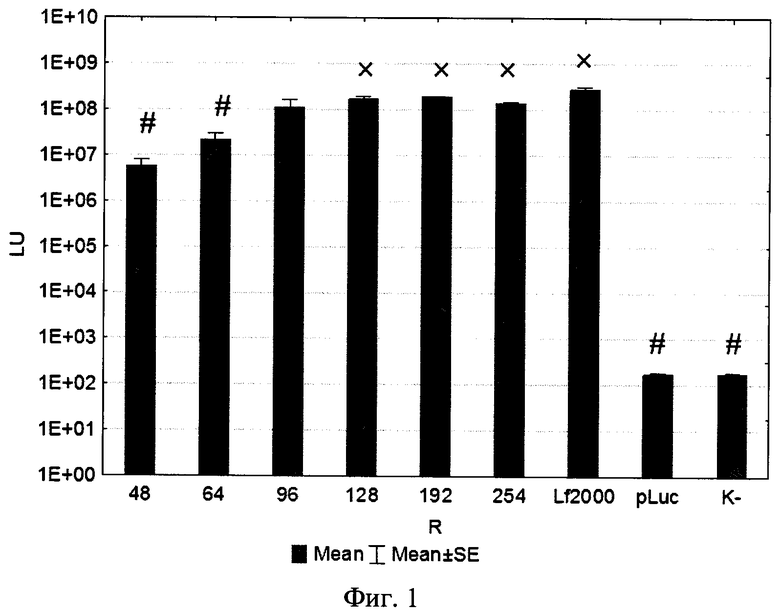
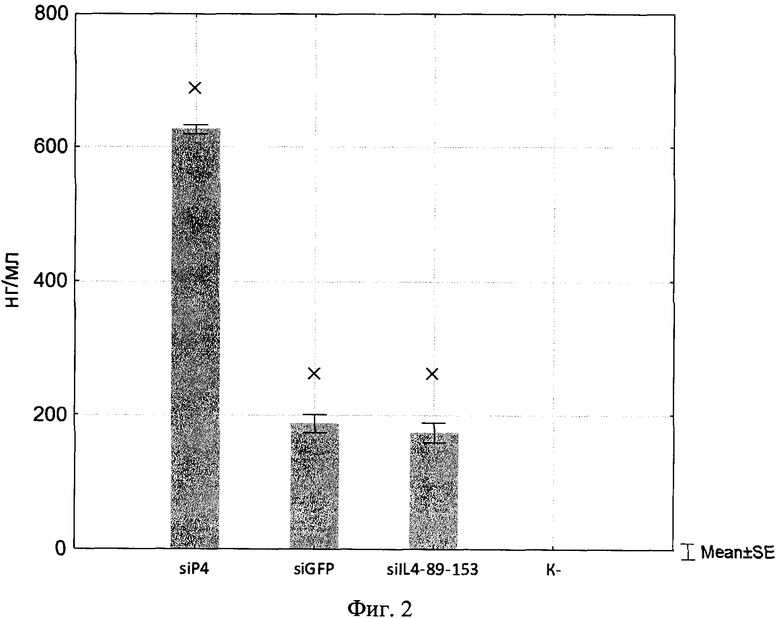
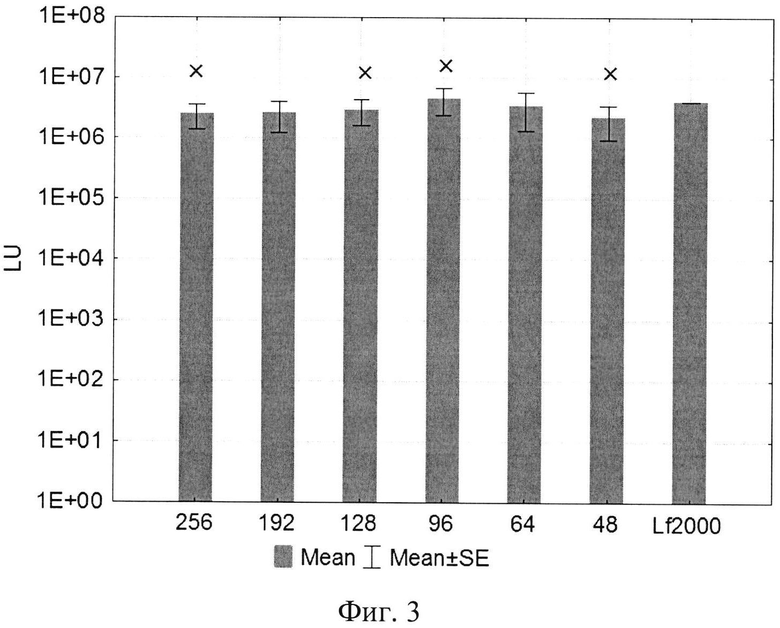
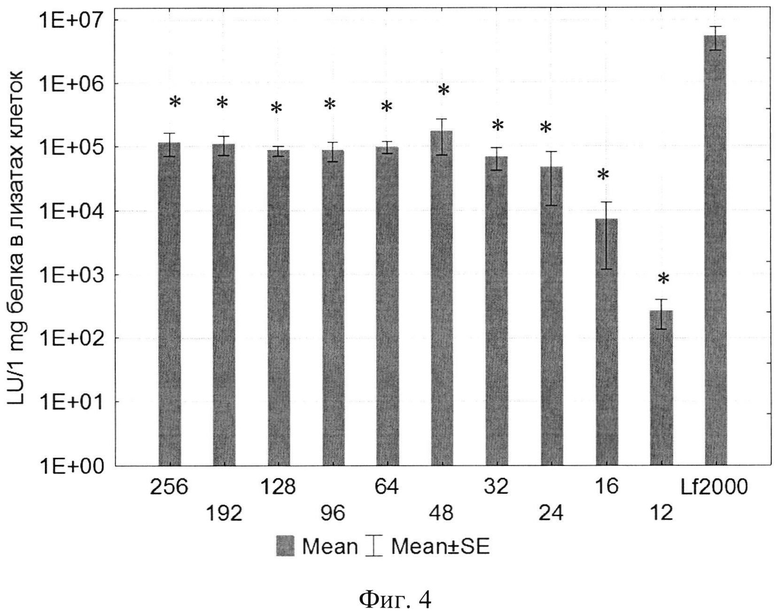
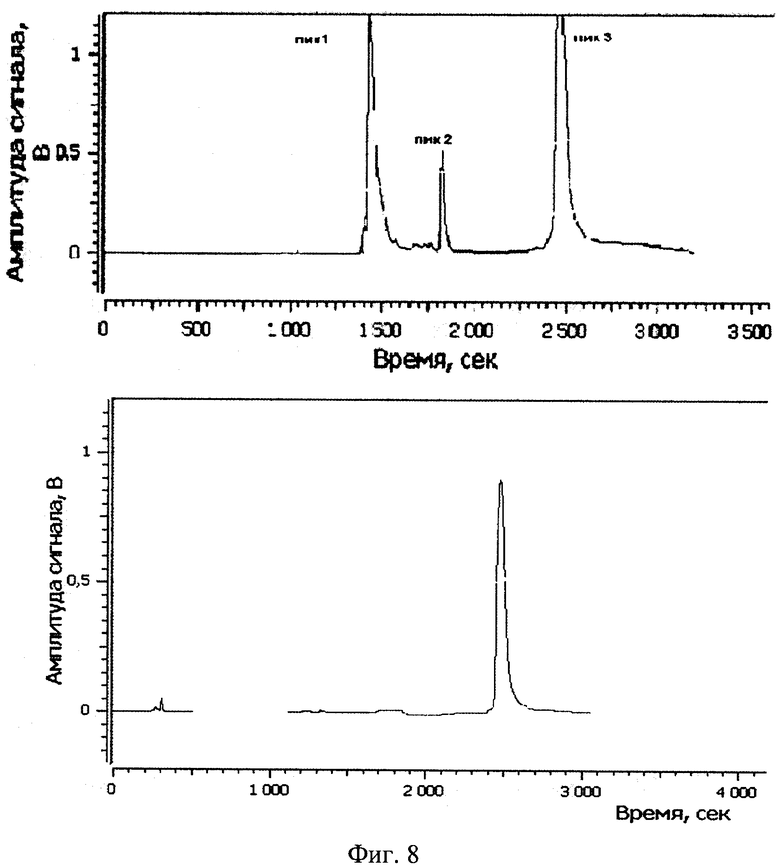
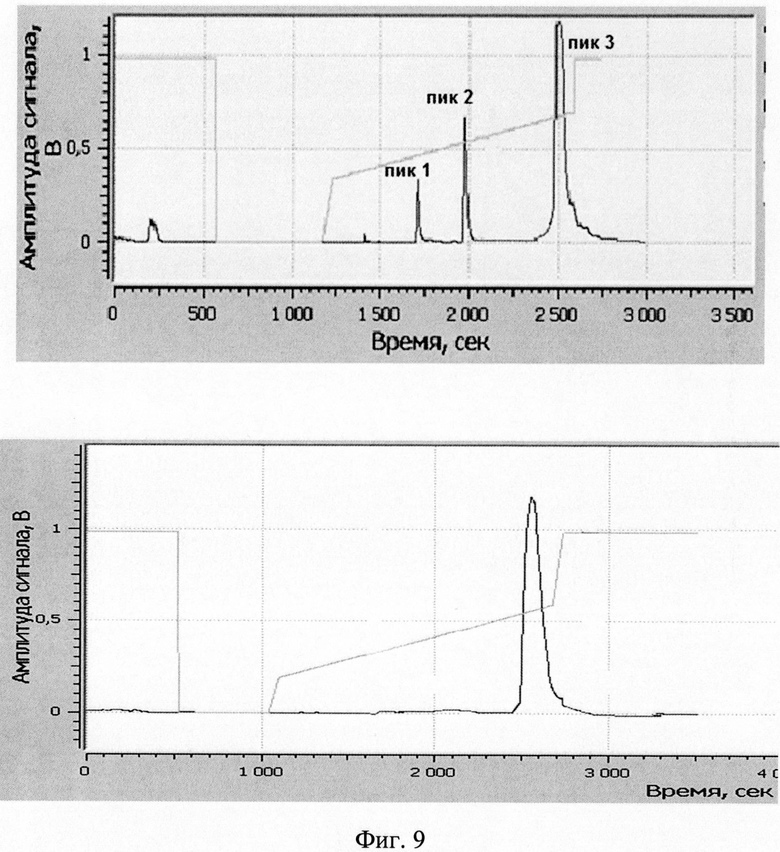
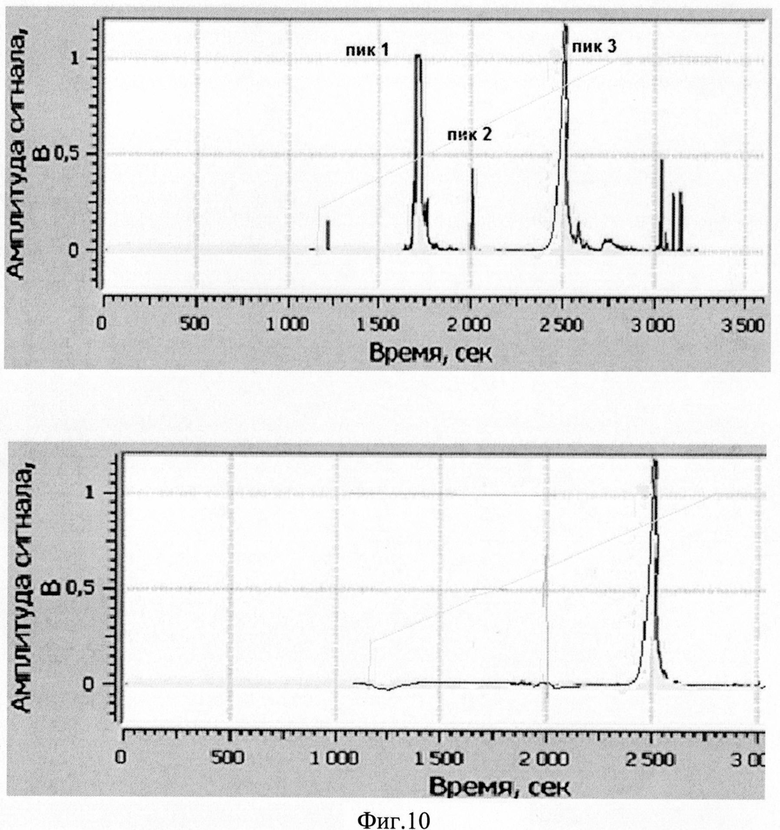
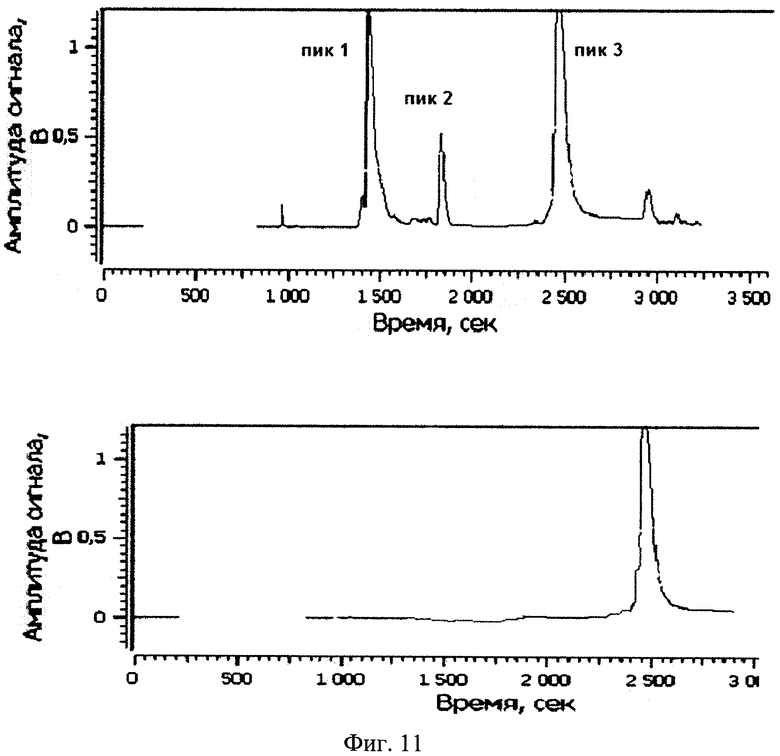
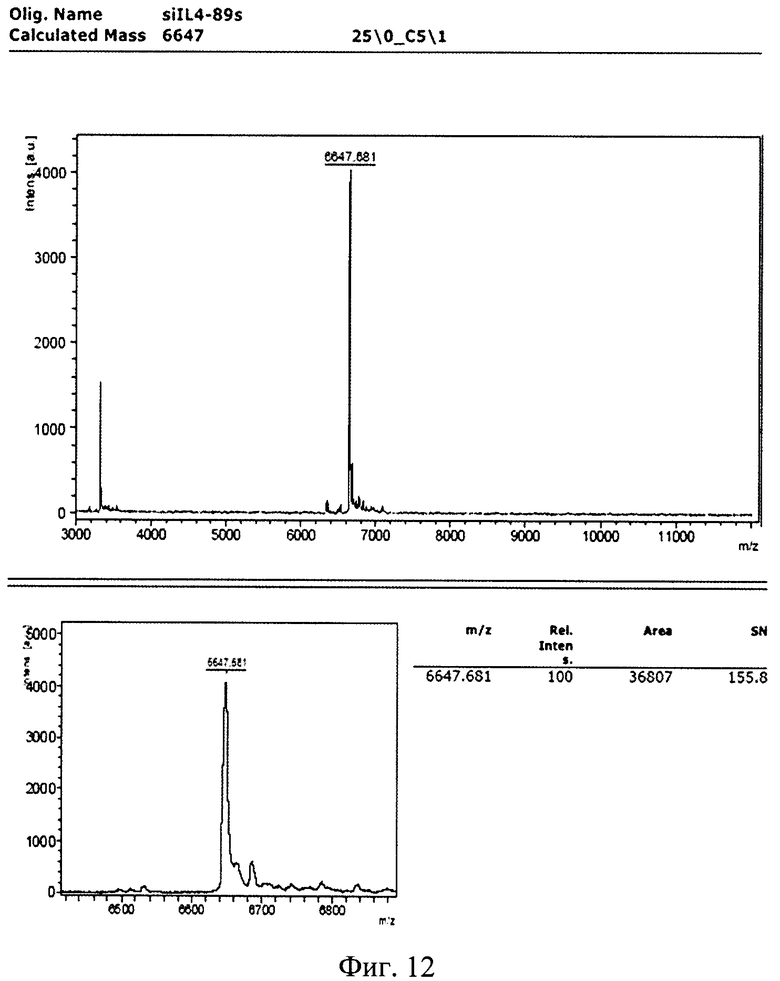
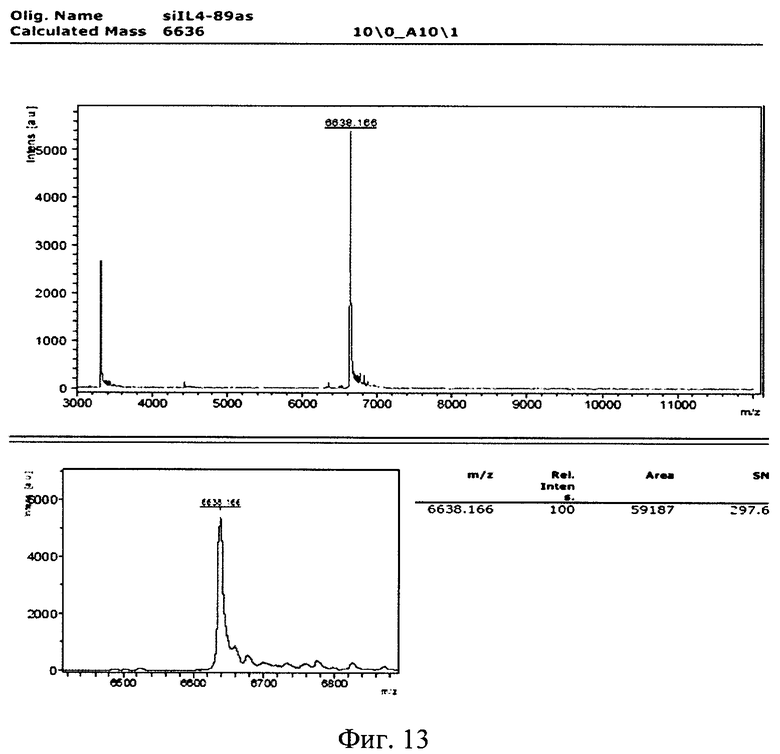
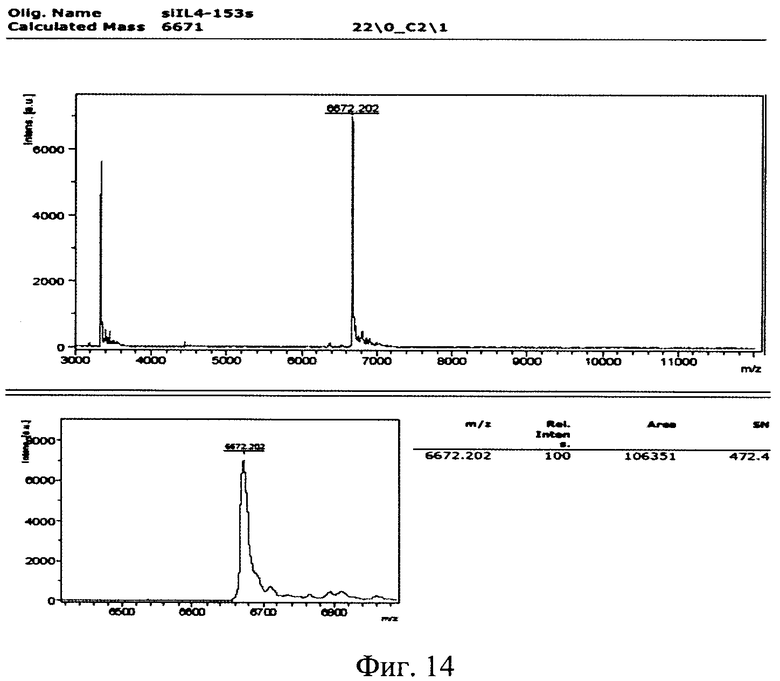
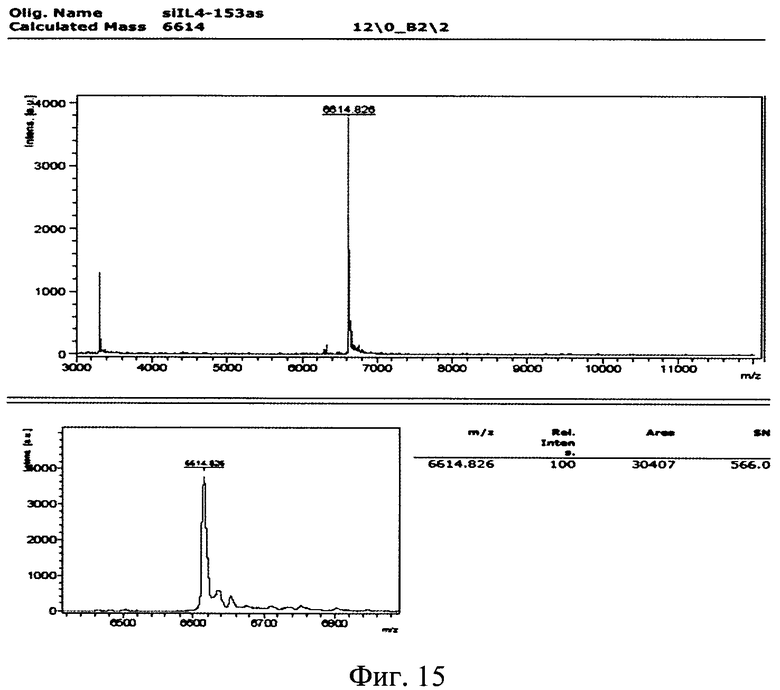
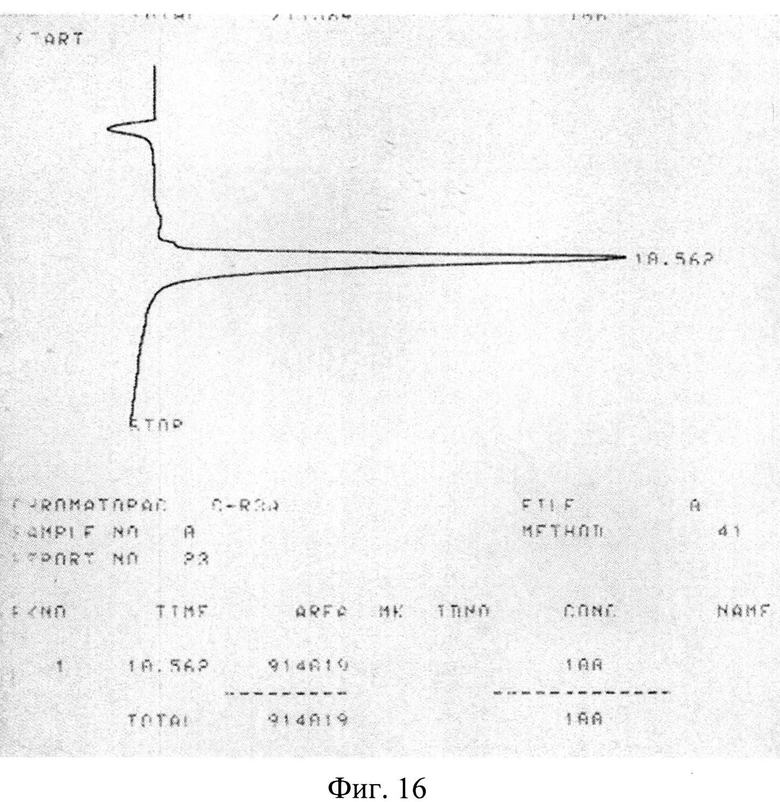
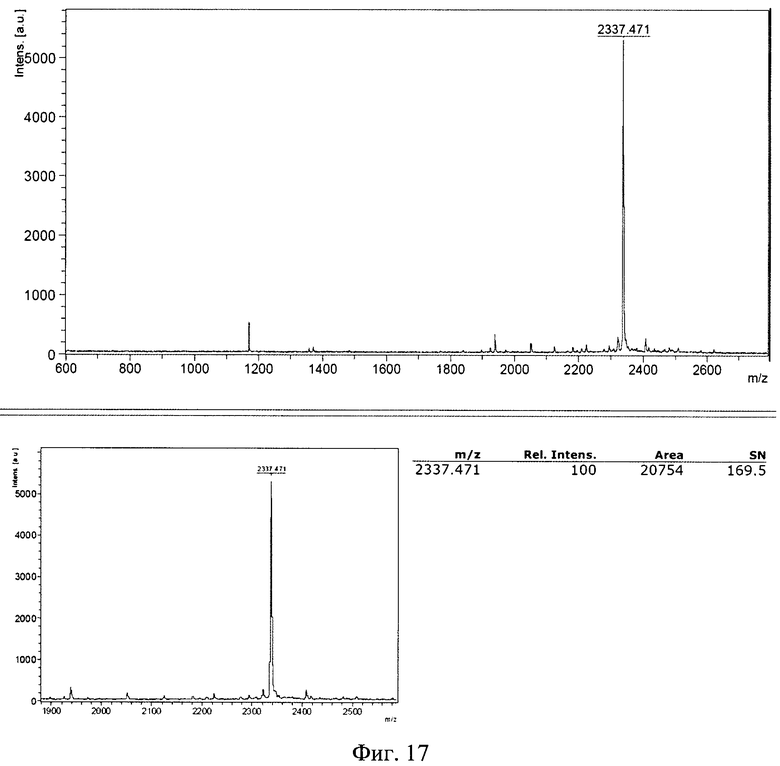
Изобретение поясняется рисунками, где на фиг. 1 представлена диаграмма эффективности трансфекции клеток HEK293T с использованием LTP, при этом условные обозначения приняты следующие: R - соотношение количества положительных зарядов LTP к количеству отрицательных зарядов плазмиды pGL3; pLuc - плазмида pGL3, кодирующая ген люциферазы luc; LU - световые единицы (light units); # - достоверно отличает от Lf2000 (Lipofectamine2000 (Invitrogen, США), использованный в качестве положительного контроля); × - достоверно отличается от pLuc (плазмида pGL3 без каких-либо трансфекционных реагентов); на фиг. 2 представлена диаграмма изменения концентрации IL-4 в супернатантах клеток HEK293T при котрансфекции молекул siRNA и рекомбинантной плазмиды pUCHR IL4 IRES GFP, где приняты следующие условные обозначения: × - достоверно отличается от siP4; на фиг. 3 представлена эффективность трасфекции клеток HelaHI с использованием LTP; на фиг. 4 представлена эффективность трасфекции клеток A549 с использованием LTP; на фиг. 5 изображены защитные группировки 2′-гидроксильной группы мономера РНК; на фиг. 6 изображены мономеры с защитными группировками ТОМ фирмы Glen Research Corporation; на фиг. 7 изображены мономеры с защитными группировками TBDMS фирмы Glen Research; на фиг. 8 приведена хроматограмма образцов РНК-олигонуклеотидов - миРНК siIL4-89s; на фиг. 9 - хроматограмма образцов РНК-олигонуклеотидов - миРНК siIL4-89as; на фиг. 10 - хроматограмма образцов РНК-олигонуклеотидов - миРНК siIL4-153s; на фиг. 11 - хроматограмма образцов РНК-олигонуклеотидов - миРНК siIL4-153as; на фиг. 12 приведены масс-спектры, подтверждающие чистоту и структуру РНК-олигонуклеотидов siIL4-89s; на фиг. 13 приведены масс-спектры, подтверждающие чистоту и структуру РНК-олигонуклеотидов siIL4-89as; на фиг. 14 приведены масс-спектры, подтверждающие чистоту и структуру РНК-олигонуклеотидов siIL4-153s; на фиг. 15 приведены масс-спектры, подтверждающие чистоту и структуру РНК-олигонуклеотидов siIL4-153as; на фиг. 16 приведена аналитическая высокоэффективная жидкостная хроматография (ВЭЖХ) очищенного пептида LTP; на фиг. 17 приведен масс-спектр очищенного пептида LTP (MALDI-TOF, на приборе BRUKER Microflex LT); на фиг. 18 показана диаграмма подавления экспрессии гена про-воспалительного цитокина IL-4 посредством композиции siIL4-89-153/LTP.
Выбор оптимальных молекул siRNA для композиции siIL4-89-153/LTP осуществлялся следующим образом. Для выбора оптимальных последовательностей молекул siRNA было использовано специализированное программное обеспечение OligoWalk, которое рассчитывает термодинамические параметры гибридизации РНК-олигонуклеотидов, предсказывает их свободную энергию связывания с мРНК-мишенью
(http://rna.urmc.rochester.edu/cgi-bin/server_exe/oligowalk/oligowalk_form.cgi). Метод селекции эффективных вариантов миРНК описан в статье (Lu Z.J. and Mathews D.H., 2008).
Для предсказания оптимальных вариантов siRNA, способных подавлять ген il-4 человека, была использована кодирующая часть нуклеотидной последовательности данного гена, опубликованная в базе данных GeneBank (Номер: M13982.1), ее последовательность представлена далее (SEQ ID NO 1):

С использованием программы OligoWalk было спрогнозировано 55 различных вариантов молекул siRNA, последовательности смысловых цепей и их возможная эффективность представлены в таблице (таб. 3). Серым цветом в таблице выделены выбранные варианты.
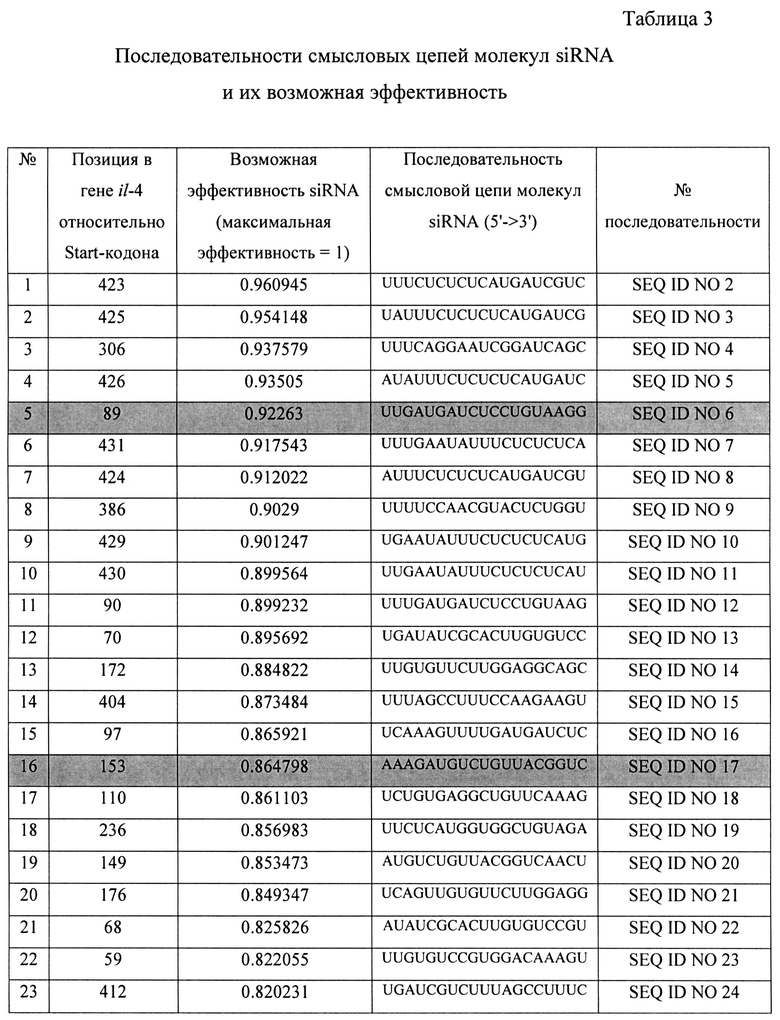
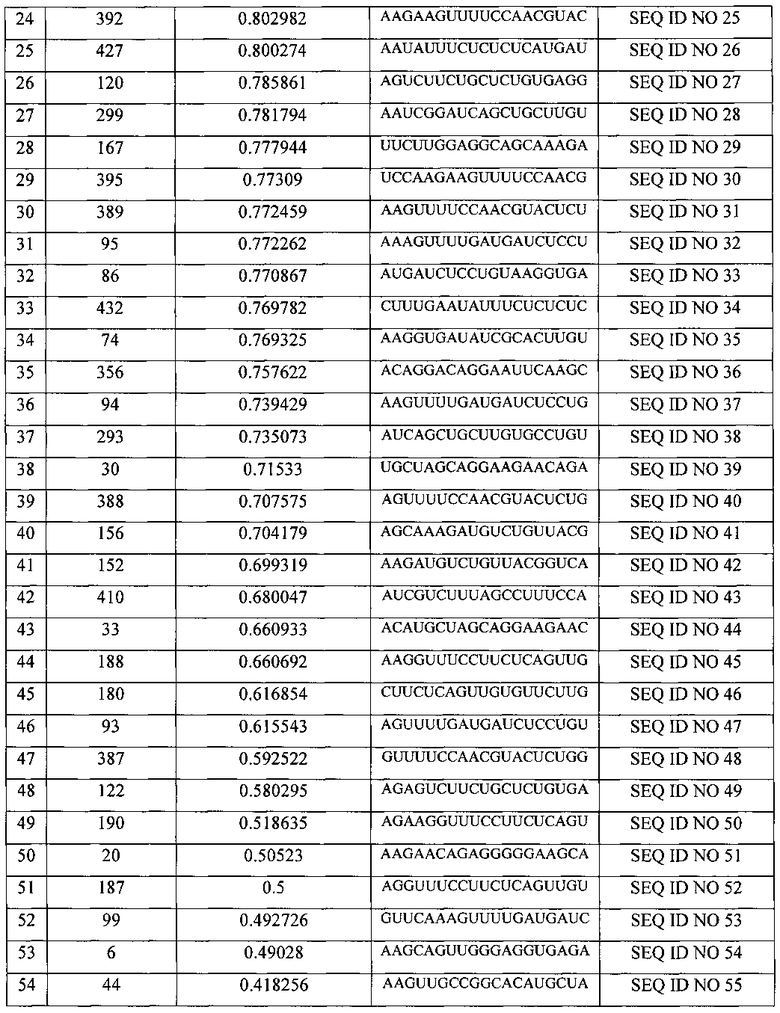

Из представленных в таблице 3 вариантов siRNA были выбраны варианты №5 и №16, они получили названия siIL4-89 и siIL4-153, соответственно. Выбор именно этих вариантов миРНК основан на рекомендациях, описанных в публикациях (Elbashir SM, Harborth J, Lendeckel W, Yalcin A, Weber K, Tuschl T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature. 2001 V. 411(6836). P. 494-8.). Согласно этим рекомендациям положение участка отжига на мРНК гена-мишени (il-4 человека) не должен находиться ближе 50 пн относительно Start-кодона гена, т.к. в данном участке мРНК-мишени инициируется трансляция, которая сопряжена с привлечением различных инициирующих белковых факторов, что снижает пространственную доступность данного участка для молекул миРНК, и как следствие понижает их активность. Рекомендуется избегать повторяющихся более 4 раз нуклеотидов таких как «АААА» и «GGGG». Рекомендуется изучить нуклеотидную последовательность миРНК на предмет сходства с другими генами в геноме человека, чтобы избежать подавления экспрессии иных генов, что может вызвать побочные эффекты лекарственного средства, в составе которого будет данная композиция. Также рекомендовано для 19-нуклеотидной последовательности спрогнозированной программой OligoWalk на 3′-конец РНК-олгонуклеотидов добавить динуклеотид «TT». С учетом этих рекомендаций были спроектированы последовательности четырех РНК-олигонуклеотидов, составляющих две молекулы siRNA для дальнейшей наработки. Их последовательности представлены в таблице 4.


Таким образом, выбрано 2 молекулы siRNA, каждая из которых состоит из двух РНК-олигонуклеотидов смысловой и антисмысловой полярности. Каждая из двух молекул siRNA способна независимо связываться с различными участками молекулы мРНК гена il-4, что повысит эффективность подавления гена-мишени.
Более того, известно, что для гена il-4 человека характерен полиморфизм нуклеотидной последовательности, т.е. в человеческой популяции присутствуют индивидуумы, у которых нуклеотидная последовательность в различных участках гена незначительно отличается. Все это может значительно снизить эффективность лекарственных средств, созданных на основе данной композиции. Иными словами, индивидуумы-носители полифонных мутаций в гене il-4 могут оказаться нечувствительными к такому препарату (хотя и с незначительной вероятностью), которое будет содержать в своем составе лишь одну молекулу siRNA. Поэтому в заявляемом изобретении применяется два варианта молекул siRNA, способных независимо связываться с разными участками мРНК гена il-4, что повысит универсальность лекарственных средств на основе данной композиции.
Пример 1. Для экспериментального подтверждения способности спроектированных молекул siRNA подавлять экспрессию про-воспалительного гена IL-4 человека был проведен соответствующий эксперимент in vitro с использованием культуры клеток HEK293T (эмбриональная почка человека) и рекомбинантной плазмиды pUCHR IL4 IRES GFP, способной экспрессировать в этих клетках IL-4 человека.
Для оценки эффективности подавления IL-4 молекулами siRNA (siIL4-89 и siIL4-153) 0,75×105 клеток HEK293T трансфецировали смесью, состоящей из 0,25 мкг плазмиды pUCHR IL4 IRES GFP, 0,5 мкг молекул siRNA и 1,5 мкл Lipofectamine2000 (Invitrogen, США) в 80 мкл optiMEM (Gibco, США). В качестве положительного контроля использовали siRNA, направленную против мРНК-gfp, которая (siGFP) подавляет экспрессию гена GFP, находящегося в составе трансфецируемой плазмиды. Так как клонированный ген IL-4 мыши и ген GFP находятся под общим промотором и транскрибируются в виде единого мРНК-транскрипта, то siGFP, подавляет и экспрессию IL-4. В качестве отрицательного контроля в эксперименте использовали siRNA против мРНК гена p респираторно-синцитиального вируса (RSV) (siP), которая не имеет сходства с последовательностями генов IL-4 и GFP. Последовательности siRNA использованные в экспериментах представлены в таблице 5.
Сутки после котрансфекции образцы клеточных супернатантов были собраны, и концентрация IL-4 была определена в них методом ИФА. Согласно полученным данным siGFP снижала экспрессию IL-4 на 70% в сравнении с неспецифической siP4, в то же время смесь siIL4-89 и siIL4-153 также снижала экспрессию IL-4, но 73% (фиг. 2).


Таким образом, экспериментально была подтверждена способность спроектированных молекул siRNA (siIL4-89 и siIL4-153) эффективно подавлять экспрессию про-воспалительного гена IL-4.
Затем проводили выбор оптимальной структуры катионного дендримерного пептида - LTP.
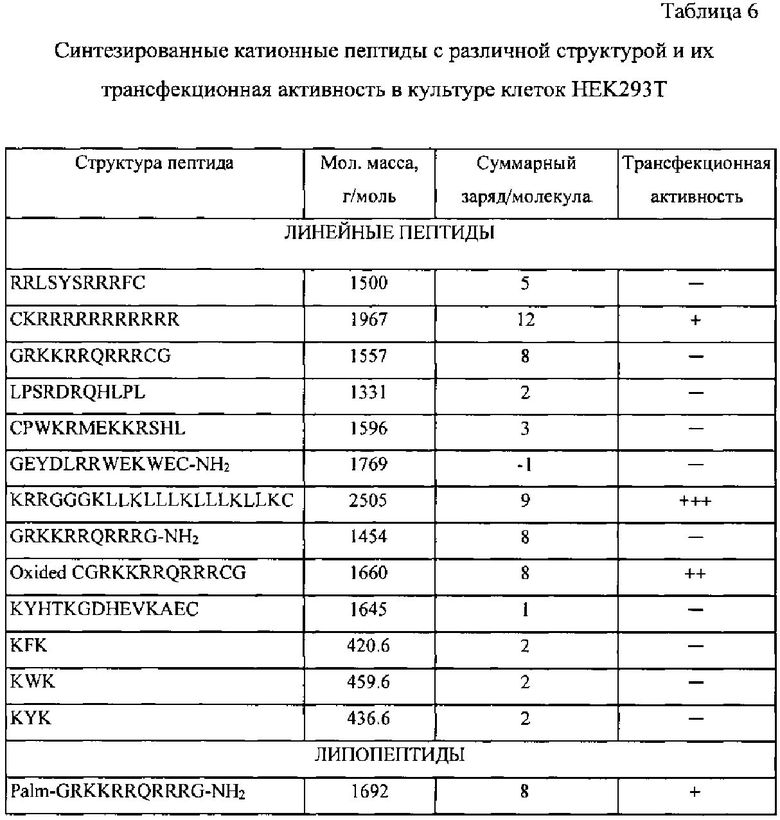
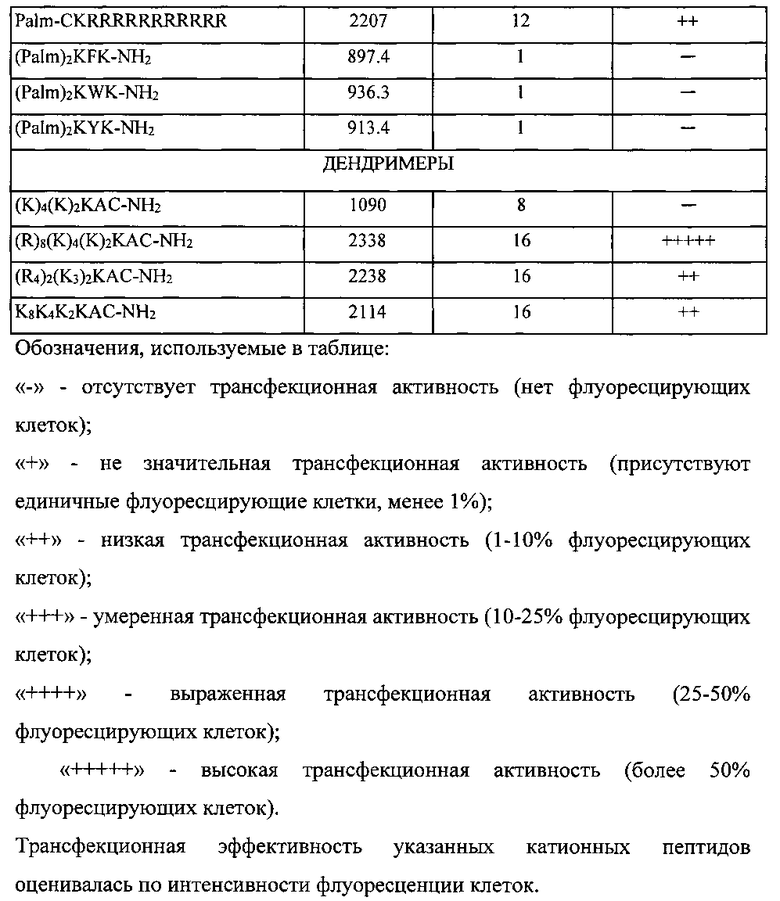
Помимо двух типов молекул siRNA (siIL4-89 и siIL4-153) в состав заявляемого изобретения входит катионный дендримерный пептид LTP, который выполняет роль носителя для молекул siRNA. Оптимальная структура пептида была выбрана в результате теоретического анализа известных литературных данных, анализе функционирования природных транспортных белков и экспериментальной проверке различных структурных особенностей пептидов. Проведенный нами предварительный анализ обусловил выбор 22 катионных пептидных последовательностей в качестве предполагаемых эффективных носителей молекул siRNA (таблица 6). Выбранные структуры можно подразделить на три основные группы: линейные пептиды, липопептиды, содержащие привязанный к N-концу остаток пальмитиновой кислоты, и - дендримерные пептиды, имеющие ветвления по аминогруппам лизина. Дальнейший выбор оптимальной структуры носителя базировался на результатах экспериментов по трансфекции клеточных культур.
Для этого все соединения с различной структурой, представленные в таблице 6 были синтезированы твердофазным методом, очищены и использованы в экспериментах по изучению эффективности трансфекции. Трансфекционная активность синтезированных структур оценивалась в эксперименте in vitro с использованием клеточной линии HEK293T (эмбриональная почка человека). Различные концентрации ситезированных пептидов индивидуально смешивались с одинаковым количеством (0,25 мкг) плазмидной ДНК, несущей репортерный ген GFP (зеленый флуоресцирующий белок); плазмида pUCHR IRES GFP. Конечный объем смеси = 80 мкл питательной среды optiMEM (Gibco, США), не содержащей антибиотики и эмбриональную телячью сыворотку (ЭТС). Смесь пептида и ДНК инкубировалась при комнатной температуре в течение 30 минут, при этом происходило образование электростатического комплекса пепетид/ДНК. После этого весь объем смеси (80 мкл) был внесен в предварительно подготовленную культуру клеток HEK293T. Культура клеток готовилась заранее в 48-луночном плейте (Costar, США): количество клеток 75000 в объеме 300 мкл полной питательно среды ДМЕМ (Gibco, США), содержащая 10% ЭТС (Hyclone, США) и 80 мг/л антибиотика Гентамицина (Gibco, США). Далее после внесения смеси в лунки с клетками их инкубировали 48 часов при 37 C в 5% атмосфере CO2. После инкубации осуществляли оценку эффективности проникновения плазмиды pUCHR IRES GFP методом флуоресцентной микроскопии. Эффективность проникновение ДНК в клетки была прямо пропорциональная интенсивности флуоресценции клеток, т.е. чем эффективнее пептид способствует внутриклеточному проникновению ДНК, тем выше интенсивность флуоресценции.
По результатам данного исследования (таблица 6) можно заключить, что наибольшую трансфекционную активность из изученных пептидов проявлял дендримерный катионный пептид с формулой (R)8(K)4(K)2KAC-NH2, обозначенный нами как LTP, входящий в состав композиции siIL4-89-153/LTP, которая является предметом данного изобретения.
Выбор оптимального массового соотношения компонентов в композиции проводили следующим образом.
Поскольку катионный носитель LTP и молекулы IL-4 (siIL4-89-153/LTP) являются уникальным соединениями, синтезированным впервые, то для подбора оптимального соотношения LTP/siRNA целесообразно использовать плазмидную ДНК, несущую репортерный ген luc, кодирующий фермент - люциферазу светлячков. Такая система удобна тем, что эффективность трансфекции будет напрямую связана с уровнем люминесценции клетки, и он легко измеряется специальным люминометром. Так как химические структуры ДНК и РНК очень близки, а количество отрицательных зарядов на 1 моль в ДНК и РНК идентичны, то эффективность комплексов носитель/ДНК коррелирует с эффективностью комплексов носитель/siRNA (Faizuloev et al., 2012). Поэтому оптимизация соотношения LTP/siRNA была проведена с использованием плазмидной ДНК (pGL3), кодирующей репортерный ген люциферазы светлячка (luc).
Для этого культуру клеток HEK293T засевают в 48-луночный плейт в полной среде ДМЕМ, которая содержит 10% эмбриональной телячий сыворотки (ЭТС), 300 мг/л Глутамина-L и 60 мг/мл Гентамицина в количестве 75 тыс. клеток на лунку в объеме 300 мкл полной среды ДМЕМ и культивируют при 37 C в 5% атмосфере CO2 до образования монослоя 75% конфлуентности (1 сутки).
Комплекс pGL3 и LTP готовился среде optiMEM (Gibco) не содержащей ЭТС и антибиотиков. Для этого необходимое количество LTP смешивали с 40 мкл optiMEM в отдельной пробирке, а 0,25 мкг pGL3 также смешивали с 40 мкл optiMEM в отдельной пробирке. Затем содержимое пробирки с pGL3 смешивали с содержимым пробирки с LTP, таким образом суммарный объем комплекса составил 80 мкл. Для достижения равновесного состояния комплекса LTP/ДНК смесь инкубировали при комнатной температуре в течение 25-30 минут избегая попадания прямых солнечных лучей или других источников ультрафиолетового излучения.
Готовили несколько вариантов комплекса с различным соотношением количества положительных зарядов LTP к количеству отрицательных зарядов pGL3 (R).
R=N(+)/N(-),
где R - соотношение количества положительных зарядов LTP к количеству отрицательных зарядов pGL3, N(+) - количество положительных зарядов LTP, N(-) - количество отрицательных зарядов pGL3.
Известно, что количество отрицательных зарядов на 1 мкг pGL3 (N(-))=17,3×1014, а что количество положительных зарядов на 1 мкг LTP (N(-))=39,3×1014.
Подготовленный и сформированный комплекс LTP/pGL3 в объеме 80 мкл optiMEM вносили к клеткам в лунки 48-луночного плейта по каплям и инкубировали при 37 C в 5% атмосфере CO2 в течение 2 суток.
После инкубации клеток с комплексом надосадочную среду удаляли из лунок плейта, а монослой клеток лизировали в 60 мкл специального буфера Luciferase Cell Culture Lysis Reagent (Promega). Лизат клеток переносили в отдельные пробирки на 1,5 мл и центрифугировали и осуществляли цикл замораживания оттаивания для лучшего лизиса клеток; замораживание проводили в течение ночи при -70 C, а оттаивание в течение 10-15 мин при 37 C в твердотельном термостате Далее лизаты клеток вортексировали и центрифугировали при 10000 об/мин при 4 C в течение 2 минут для удаления клеточного дебриза. Надосадок переносили в отдельные пробирки на 1,5 мл и проводили измерение люциферазной активности каждого образца путем смешивания 50 мкл лизата с 50 мкл люциферина - субстрата для люфиферазы. При наличие в образце фермента люциферазы происходит излучение света, интенсивность которого детектировалось люминометром (Promega).
Эффективность трансфекции культуры клеток HEK293T с использованием LTP была сравнима с коммерческим реагентом Lipofectamine2000. Оптимальное соотношение R при трансфекции клеток HEK293T находилось в пределах от 96 до 254 (фиг. 1). При трансфекции клеток HelaHI (эпителиоидная карцинома шейки матки) оптимальные соотношения (R) находились в пределах от 48 до 254 (рис. 3). При изучении эффективности трансфекции клеток A549 (карцинома легкого) показано, что оптимальная соотношение (R) находится в пределах от 32 до 254 (фиг. 4).
На фиг. 4 использованы следующие условные обозначения:
R - соотношение количества положительных зарядов LTP к количеству отрицательных зарядов плазмиды pGL3;
pLuc - плазмида pGL3, кодирующая ген люциферазы luc;
LU - световые единицы (light units);
# - достоверно отличает от Lf2000 (Lipofectamine2000 (Invitrogen, США), использованный в качестве положительного контроля);
× - достоверно отличается от pLuc (плазмида pGL3 без каких-либо трансфекционных реагентов).
Также определена эффективность трасфекции клеток HelaHI с использованием LTP, результаты которой преведены на фиг. 3, где
R - соотношение количества положительных зарядов LTP к количеству отрицательных зарядов плазмиды pGL3;
pLuc - плазмида pGL3, кодирующая ген люциферазы luc;
LU - световые единицы (light units);
Lf2000 - Lipofectamine2000 (Invitrogen, США), используется в качестве положительного контроля;
× - достоверно отличается от pLuc (плазмида pGL3 без каких-либо трансфекционных реагентов).
Также была определена эффективность трасфекции клеток A549 с использованием LTP, результаты которой приведены на фиг. 4, где
R - соотношение количества положительных зарядов LTP к количеству отрицательных зарядов плазмиды pGL3;
pLuc - плазмида pGL3, кодирующая ген люциферазы luc;
LU - световые единицы (light units);
Lf2000 - Lipofectamine2000 (Invitrogen, США), используется в качестве положительного контроля;
* - достоверно отличается от pLuc (плазмида pGL3 без каких-либо трансфекционных реагентов).
Таким образом, суммировав результаты трасфекции трех различных клеточных линий можно заключить, что наиболее оптимальное соотношение LTP/siRNA будет находиться в пределах от 48 до 128. С точки зрения экономической целесообразности соотношение зарядов R=48, что соответствует соотношению LTP/siRNA 25/1 по массе является наиболее оптимальным, т.к. выбор такого состава композиции позволит эффективно трасфецировать различные типы клеток с минимальными затратами LTP.
Получение композиции siIL4-89-153/LTP осуществлялось следующим способом.
Синтез молекул siRNA (siIL4-89 и siIL4-153), входящих в состав композиции siIL4-89-153/LTP, осуществляют амидофосфатным методом твердофазного синтеза. Суть метода заключается в добавлении одного нуклеотидного звена к иммобилизованному защищенному нуклеозиду или олигонуклеотиду. Нуклеозидный компонент связан ковалентно с нерастворимым полимером, а нуклеотидный компонент и необходимые реагенты находятся в растворе. В качестве полимерной подложки использовали стекло Glen 20-2030-10 dT-CPG-500 °А. 3′-Концевой нуклеозидный остаток связывается с такими матрицами через сукцинильное спейсерное звено. Никаких стадий очистки не проводится до тех пор, пока полностью синтезированная олигомерная последовательность не будет удалена с твердой фазы.
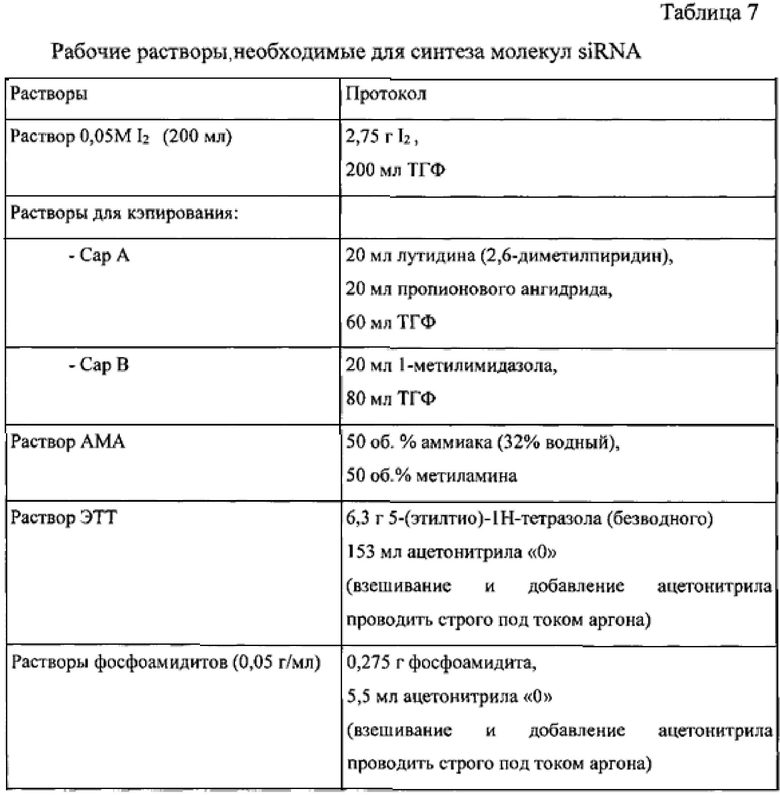
Синтез проводится на ДНК-синтезаторе Poligen 2000 с 12 колонками. Управление и программирование осуществляют с помощью вынесенного компьютера. Приготовление рабочих растворов для синтеза проводится по протоколу, указанному в таблице 7.

Для синтеза РНК-олигонуклеотидов, составляющих молекул siRNA проводился подбор мономеров. В синтезе РНК используются строительные блоки с «дополнительной» 2′-гидроксильной группой, не участвующей в синтезе. Ее защищают трет-бутилдиметилсилильной (TBDMS) или триизопропилсилилоксиметильной (ТОМ) группой (фиг. 5, 6).
ТОМ-защита обладает более низким стерическим фактором, чем TBMDS, что обеспечивает высокую эффективность сцепления мономеров. Еще одной особенностью ТОМ является то, что во время основных стадий синтеза практически исключена ее миграция из 2′ в 3′ положение, что сводит к минимуму получение небиологических активных связей.
Были проведены параллельно синтезы, с использованием мономеров с ТОМ- и TBMDS-защитами.
В обоих случаях синтез прошел с высокой эффективностью, но, исходя из экономической целесообразности, для дальнейшей работы были выбраны мономеры с TBMDS-защитой. Спектры полученных РНК-олигонуклеотидов для обоих случаев идентичны, что свидетельствует об идентичности продуктов.
После синтеза РНК-олигонуклеотидов у них были удалены все защитные группы:
1. 5′-концевая DMT-группа (удаляется после очистки олигонуклеотида методом ВЭЖХ);
2. Ацильные и бензильные защитные группы;
3. 3′-цианэтильные группы, защищающие межнуклеозидные фосфатные группы;
4. 2′-O-Si-связь
Отщепление олигонуклеотида от носителя, удаление защитных групп оснований и цианэтильных защитных групп проводили одновременно под действием водного раствора аммиак/метиламин.
Для удаления защиты с 2′-конца (с сохранением 5′-DMT-группы) использовали следующую методику (количества реактивов указаны на 250 мг олигонуклеотида).
Сначала полностью растворяли РНК-олигонуклеотид в 115 мкл диметилсульфоксида DMSO (для лучшей растворимости нагревать в течение 5 мин при температуре 65°C).
Затем добавляли 60 мкл триэтиламина TEA к раствору DMSO/олигонуклеотид и аккуратно перемешивали. После этого добавляли 75 мкл TEA*3HF и нагревали раствор при 65°C в течение 2,5 часов. После чего раствор немедленно охлаждали.
При подготовке олигонуклеотидов к очистке их предварительно обессоливали осаждением растворов по следующей методике:
1. Добавить 5 мкл перхлората лития LiClO4 на 1 мкл раствора олигонуклеотида.
2. Центрифугировать 10 мин при 12,5000 rpm
3. Отобрать надосадочную жидкость (перхлорат лития), используя стерильный наконечник.
4. Промыть осадок ацетоном 2-3 раза, пока раствор не станет прозрачным.
5. Отобрать надосадочную жидкость (ацетон), используя стерильный наконечник.
6. Высушить под высоким вакуумом для удаления следов ацетона.
Очистка РНК-олигонуклеотидов проводилась методом обращено-фазовой ВЭЖХ (высокоэффективной жидкостной хроматографии). Такой метод очистки был выбран исходя из того, что только целевой компонент (синтетический РНК-олигонуклеотид) содержит на 5-конце гидрофобную диметокситритильную группу. Все побочные продукты с более низкой молекулярной массой, которые могли появиться в результате твердофазного синтеза, не несут на 5′-конце диметокситритильной группы. Оптимальным методом выделения для такого компонента является градиентная хроматография на обращено-фазовой колонке C18 с полярным растворителем (50% ацетонитрил) в качестве элюента, где целевой компонент будет обладать наибольшим временем удерживания и соответственно может легко быть отделен от нецелевых продуктов. Подбор градиента состава подвижной фазы в зависимости от времени подбирался экспериментально. Критерием отбора было оптимальное разделение целевого компонента от остальных при наименьшем времени очистки.
Очистка синтетических РНК-олигонуклеотидов проводилась с использованием препаративной хроматографической системы, оснащенной препаративной колонкой (250×10 мм), содержащей 5 мкм-сорбент с обращенной C18 фазой, градиентным насосом, детектором с переменной длиной волны (установлена длина волны 260 нм), коллектором фракций, самописцем, компьютером для управления насосом, расчета времени удерживания и площади пиков.
Хроматограммы образцов РНК-олигонуклеотидов (фиг. 8-11) содержат 3 основных пика, первые два из которых обладают низким временем удерживания, характерным для компонентов, не содержащих диметокситритильной группы, и не являются целевыми, что дополнительно было подтверждено методом масс-спектрометрии. Пик №3, содержащий требуемый РНК-олигонуклеотид (что подтверждено данными масс-спектрометрии) и обладающий наибольшим временем удерживания и площадью, собирали в автоклавированные пробирки на 1,5 мл так, чтобы побочные пики и «плечи» не попали в пробирку. Пробирки с собранными целевыми фракциями высушивали на роторной сушке в течение 12-15 часов (как изложено ниже). Аликвоту целевого компонента объемом 20 мкл повторно хроматографировали в тех же условиях на аналитической колонке (Jupiter 5u C18 300 4,6×250 мм). Аналитические хроматограммы содержали один пик целевого компонента. Результаты хроматографической очистки РНК-олигонуклеотидов и аналитические хроматограммы целевого пика представлены на фиг. 17.
Для высушивания растворов РНК-олигонуклеотидов использовали систему роторной сушки, время высушивания 12-15 часов.
Для анализа структуры синтезированных РНК-олгонуклеотидов использовался МАЛДИ-масс-спектрометр BRTJKER microflex LT. Детекция велась в режиме положительных ионов. Оптимальная энергия импульсов азотного лазера составляла 70 мкрДж. Детекция ионов велась в диапазоне 3-12 кДа (г/моль). Для сополимеризации образца использовался раствор матрицы 3-НРА (3-гидроксипиколиновая кислота) в деионизованной воде в концентрации 30 мг/мл с двузамещенным цитратом аммония в концентрации 0,5 мг/мл.
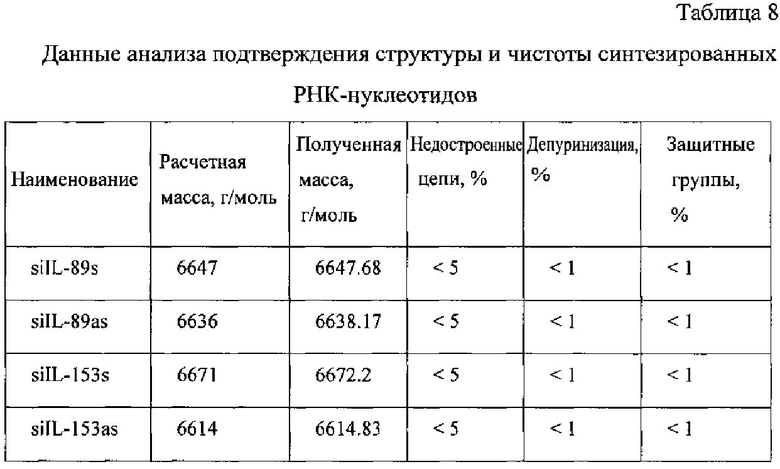
Полученные масс-спектры приведены на фиг. 12-15. Наблюдаемая масса однозарядного иона для каждого образца соответствует расчетной с погрешностью, не превышающей 0.1%. Наблюдаемые пики с массой в два раза меньшей массе пика с максимальной интенсивностью соответствуют двузарядным ионам целевого пика, являются особенностью метода и в анализе не учитываются.
Пики с массой ниже массы основного пика на 329,21 Да, 305,18 Да, 345,21 Да, 306,20 Да, 304,20 Да, 109,09 Да, 124,1 Да, 133,11 Да, 149,11 Да, как и пики с массой выше искомой на 114,2 Да, 42,04 Да не обнаружены. Соответственно был сделан вывод о низком содержании или отсутствии недостроенных цепей, последовательностей, несущих депуринизованные основания, либо несущие защитные группы.
Сводные данные по анализу полученных масс-спектров представлены в таблице 8, представленной далее. Чистота анализируемых РНК-олигонуклеотидов не ниже 90%.
Синтез LTP проводили твердофазным методом, используя стандартный протокол Fmoc-химии и N-гидроксибензотриазол/диизопропилкарбодиимидный (ГБТ/ДИП) метод активации Fmoc-аминокислот. В качестве полимера-носителя использовали коммерческие реактивы: смолу Rink Amide МВНА, или Wang. В качестве исходных производных аминокислот применяли: Fmoc-Cys(Trt)-OH, Fmoc-Ala-OH, Fmoc-Lys(Boc)-OH, Fmoc-Arg(Pbf)-OH. Стандартный цикл включет промывку (ДМФА), удаление Fmoc-защиты (20% пиперидин в ДМФА), предварительное активирование Fmoc-аминокислоты (ДИП/ГБТ) и кондесацию в среде ДМФА/N-метилпирролидон при 2-кратном избытке карбоксильного компонента (~0,5 часа). Контроль над полнотой реакции осуществляли нингидриновым методом, и при необходимости реакцию конденсации повторяли. Конечные пептиды отщепляют от полимера трифторуксусной кислотой в присутствии скавенджеров (тиоанизол, этандитиол, фенол, диметилсульфид).
Сырой продукт высаждали сухим серным эфиром, экстрагировали водной уксусной кислотой и экстракт лиофилизируют. Очистку пептида проводили методом высокоэффективной жидкостной хроматографии (ВЭЖХ) (фиг. 16), например, на колонке Grace Vydac 218 ТР54, а структуру очищенного пептида подтверждали масс-спектрометрией (фиг. 17).
Пример 2. Полученные параметры стандартного образца LTP: время удерживания на аналитической колонке: 10,1±0,2 мин., масс-спектр (MALDI-TOF): m/z=2337,471±1 при расчетной массе: 2337,98 (для амида). Аналитическая высокоэффективная жидкостная хроматография (ВЭЖХ) очищенного пептида LTP была проведена следующим образом. Для аналитических целей была использована аналитическая колонка Grace Vydac 218ТР54 (150×4.6 мм), содержащая 5 мкм сорбент C18. Подвижная фаза: 0,1% водный раствор трифторуксусной кислоты (А) и ацетонитрил (Б) - смесь, 1 мл/мин. Градиент, % состав: 0-2 мин (А - 100%); 2-25 мин (Б -линейно от 0 до 70%); 25-27 мин (Б - 70%); 27-29 мин (Б - линейно от 70 до 0%); 29-35 мин (А - 100%).
После синтеза четырех РНК-олигонуклеотидов проводили их попарный отжиг, чтобы получить два типа молекул siRNA (siIL4-89-153/LTP). Под отжигом олигонуклеотида понимается смешивание водных растворов эквимолярных количеств сенс- и антисенс-олигонуклеотидов, при этом между ними образуются водородные связи и два олигонуклеотида образуют прочный дуплекс. Для этого водные растворы с известной концентрацией сенс- и антисенс-РНК-олигонуклеотидов смешивают в эквимолярных количествах, и прогревают при 80 C в течение 10 мин после чего раствору дают остывать до комнатной температуры в течении 1 часа. Затем водный р-ор образовавшихся после отжига молекул siRNA лиофильно высушивали для получения порошкообразной формы.
Получение композиции siIL4-89-153/LTP из компонентов LTP и двух видов молекул siRNA (siIL4-89 и siIL4-153) заключается в следующем:
1. готовят раствор LTP в фосфатно-солевом буфере (ФСБ), pH=7,4+0,4 М NaCl концентрацией 25 мг/мл и раствор смеси молекул siIL4-89 и siIL4-153 концентрацией 1 мг/мл в ФСБ pH=7,4+0,4 М NaCl.
2. водные растворы LTP (25 мг/мл) и siRNA (1 мг/мл) смешиваются в объемных соотношениях 1:1 соответственно. Смешивание осуществляют в стеклянной колбе.
3. Смесь растворов инкубируется в течении 30 минут при комнатной температуре (+21 C). При этом происходит образование комплекса LTP/siRNA в массовых соотношениях 25/1 концентрацией 13 мг/мл.
Таким образом, в результате синтеза компоненты композиции siIL4-89-153/LTP, пептид siRNA и молекулы siRNA (siIL4-89 и siIL4-153) изначально получают независимо, после чего проводят процедуру комплексообразования LTP/siRNA.
Было показано, что композиция siIL4-89-153/LTP эффективно подавляет экспрессию про-воспалительного цитокина IL-4.
Пример 3. В предварительных исследованиях было показано, что созданные молекулы siIL4-89 и siIL4-153 эффективно подавляли экспрессию гена IL-4. Чтобы подтвердить, что непосредственно композиция также siIL4-89-153/LTP подавляет экспрессию IL-4, были проведены дополнительные эксперименты. Для этого культуру клеток HEK293T засевали в 4 8-луночный плейт в полной среде DMEM, которая содержала 10% эмбриональной телячий сыворотки (ЭТС), 300 мг/л Глутамина-L и 60 мг/мл Гентамицина в количестве 75 тыс.клеток на лунку в объеме 300 мкл полной среды DMEM и культивируют при 37°C в 5% атмосфере CO2 до образования монослоя 75% конфлуентности (примерно 1 сутки). Готовили смесь, состоящую из 25 мкг LTP с 1,0 мкг плазмиды pUCHR IL4 IRES GFP (соотношение 25:1 по массе) в 40 мкл среды optiMEM, не содержащей ЭТС и антибиотиков в отдельной пробирке. Отдельно готовили несколько образцов композиции siIL4-89-153/LTP (130 мкг, 65 мкг, 33 мкг) растворяя их в 40 мкл optiMEM в отдельных пробирках. Затем содержимое пробирки со смесью пептида с плазмидой (LTP/pUCHR IL4 IRES GFP) смешивали с содержимым пробир с композицией siIL4-89-153/LTP, таким образом, суммарный объем составлял 80 мкл. Для достижения равновесного состояния смеси ее инкубировали при комнатной температуре в течение 25-30 минут, избегая попадания прямых солнечных лучей или других источников ультрафиолетового излучения. В качестве контроля сравнения использовали образец без добавления композиции. В качестве отрицательного контроля использовали среду optiMEM без добавления плазмиды или трансфекционных агентов.
Далее, подготовленные трансфекционные смеси и контроли в объеме 80 мкл optiMEM вносили к клеткам в лунки 48-луночного плейта по каплям и инкубировали при 37°C в 5% атмосфере CO2 в течение 2 суток.
После инкубации клеток с среду удаляли из лунок плейта и хранили при -70 C до анализа из методом иммуноферментного анализа (ИФА) на содержание IL-4. Уровень экспрессии IL-4 определяли с использованием коммерческого набора Human IL-4 ELISA Set (BD). Для этого на подложку (96-луночный плейт) сорбировали связывающие антитела (Anti-human IL-4) в разведении 1/250 от начальной концентрации по 100 мкл раствора на лунку. Инкубировали в течение ночи при +4°C. Плейт отмывали отмывочным буфером (1 л PBS/ 0,5 мл Tween 20) 3 раза и вносили обезжиренное молоко по 200 мкл на лунку, инкубировали 2 часа при комнатной температуре. Плейт отмывали 2 раза и вносили супернатанты (без разведения, 1/10, 1/100, 1/1000). Инкубировали в течение часа при комнатной температуре. Далее плейт отмывали 3 раза, после чего вносили рабочий раствор (детектирующие антитела против IL-4 в разведении 1/1000 + стрептавидиновый коньюгат в разведении 1/250) по 100 мкл на лунку и инкубировали 1 час при комнатной температуре. Затем плейт отмывали 6 раз отмывочным буфером и вносили субстрат (тетра-метил-бензидин) по 100 мкл на лунку, после чего инкубировали 25-30 минут в темноте. Реакцию останавливают добавлением 50 мкл 2 н H2SO4 и фиксировали результат с помощью спектрофотометра Reader MPR1 при длинах волн 450 и 630 нм.
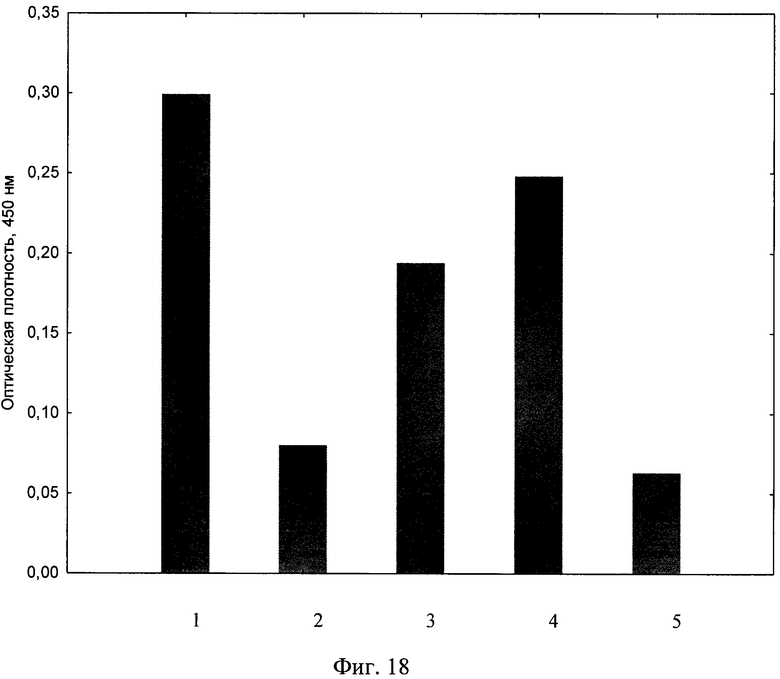
Интенсивность экспрессии IL-4 определяли по показателю оптической плотности раствора. Отрицательный контроль, определяющий фоновый уровень реакции, не превышает 0,07. В образце положительного контроля (клетки инкубировались без добавления композиции siIL4-89-153/LTP) наблюдался высокий уровень экспрессии IL-4 в сравнении с контролем (фиг. 18). Однако, добавление в трансфекционную смесь композиции siIL4-89-153/LTP, значительно ослабило экспрессию IL-4 клетками. При этом эффект подавления IL-4 был дозозависимым, т.е. при уменьшении концентрации композиции siIL4-89-153/LTP происходило восстановление экспрессии IL-4. Таким образом, установлено, что композиция дозозависимо подавляет экспрессию гена про-воспалительного цитокина IL-4.
На фиг. 18 показано подавление экспрессии гена про-воспалительного цитокина IL-4 посредством композиции siIL4-89-153/LTP,
где
1 - экспрессия гена IL-4 клетками HEK293T,
2 - экспрессия гена IL-4 клетками HEK293T, после добавление 130 мкг композиции siIL4-89-153/LTP,
3 - экспрессия гена IL-4 клетками HEK293T, после добавление 65 мкг композиции siIL4-89-153/LTP,
4 - экспрессия гена IL-4 клетками HEK293T, после добавление 33 мкг композиции siIL4-89-153/LTP.
5 - экспрессия гена IL-4 клетками HEK293T без добавления каких-либо трансфекционных агентов и нуклеиновых кислот.
Было показано и доказано, что композиция siIL4-89-153/LTP мало токсична.
Пример 4. Была изучена острая токсичность композиции siIL4-89-153/LTP при внутрибрюшинном введении мышам в высоких дозах. Исследование выполнено на половозрелых мышах (самки) при однократном внутрибрюшинном введении в максимально возможной для введения дозе. Предполагаемая терапевтическая доза композиции siIL4-89-153/LTP находится в пределах 3,25 мг для взрослого, т.е. максимум 0,05 мг/кг. Чтобы оценить летальные дозы композиция siIL4-89-153/LTP была введен мышам в значительной передозировке в 13000 раз (650 мкг/кг). Препарат вводился внутрибрюшинно, поскольку данный способ обеспечивает высокий уровень всасывания. Для данного исследования выбраны мыши (CBA×C57BL/6)F1 как вид, общепринятый для токсикологических исследований.
Дизайн исследования острой токсичности на половозрелых мышах (CBA×C57BL/6)F1 представлен в таблице 9.

 .
.
Токсичность регистрировали по появлению признаков нарушения здоровья животных при клиническом осмотре в клетке, в руках и на открытой площадке (поза, поведение, тремор, аномальные движения, внешний вид, состояние шерсти, диарея, пилоэрекция, исследовательская активность, общая подвижность), а также по приросту массы тела. Исследуемые показатели у мышей:
1. Масса тела каждые на 0, 1, 7 и 14 сутки.
2. Клинический осмотр - ежедневно. Параметры оценки интоксикации (двигательная активность, координация движений, ориентировочные реакции, тонус скелетных мышц, реакция на тактильные и болевые раздражители, частота дыхания, окраска слизистых оболочек, состояние волосяного покрова, консистенция фекальных масс).
Клинический осмотр заключался в наблюдении за животными для выявления отклонений в состоянии здоровья. Его проводили дважды в первые сутки после введения тестируемого препарата, затем регулярно один раз в день вплоть до некропсии. Клинический осмотр каждого животного проводили в течение первых трех часов после введения и далее один раз в сутки. Осмотр животного проводили в клетке содержания, в руках и на стандартной открытой площадке. Ниже указаны признаки, которые описываются при оценке токсичности тестируемых веществ.
При изучении острой токсичности массу тела определяли перед введением, далее на 1,3, 7, 10 и 14 сутки.
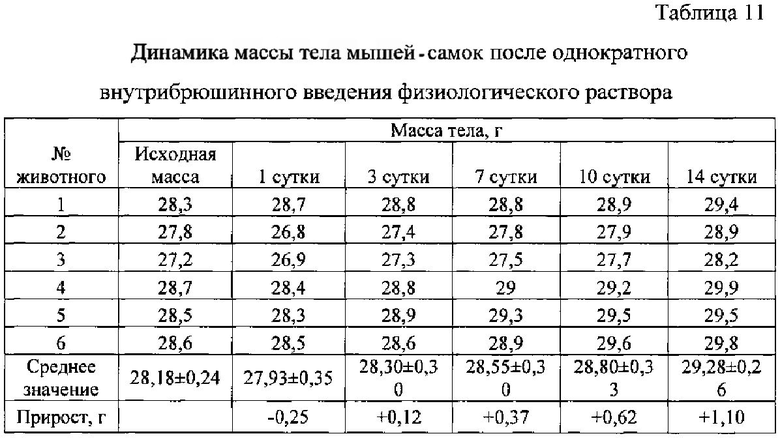
Результаты наблюдений показали, что на протяжении всего исследования гибели животных в контрольной группе, получавшей физиологический раствор, не наблюдалось.
Выживаемость и масса тела подопытных животных представлены в таблицах ниже (табл. 10-14).
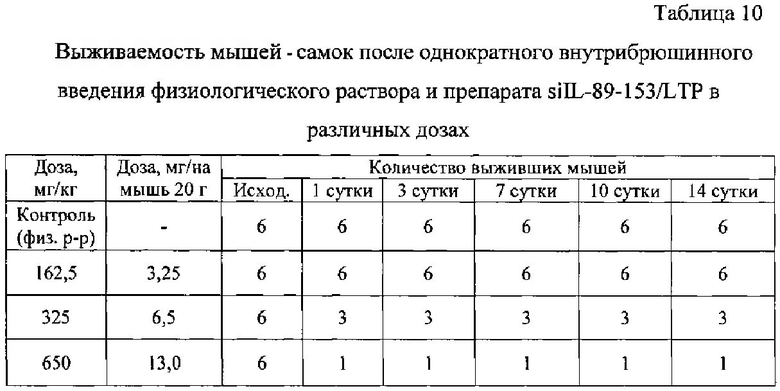
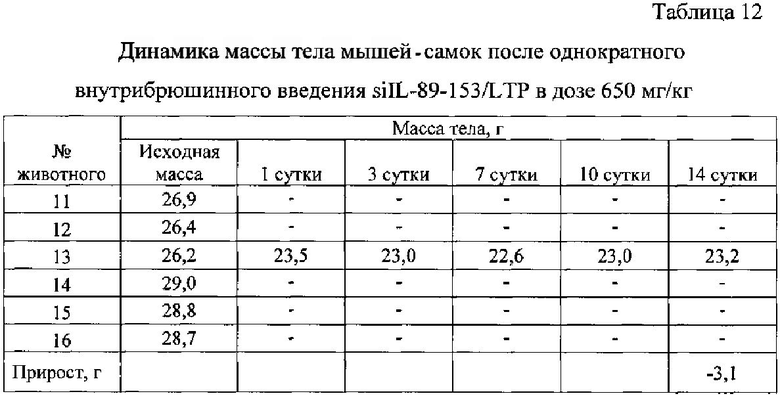

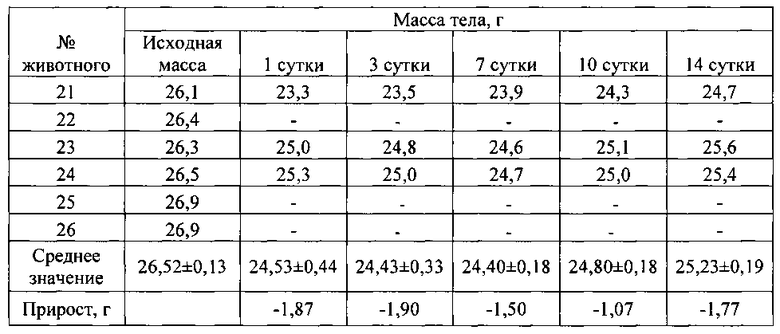
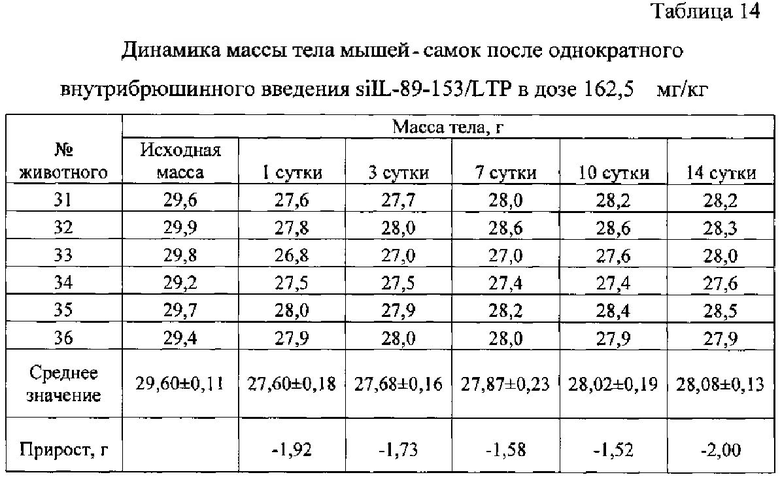
Из таблиц 10-14 видно, что однократное внутрибрюшинное введение мышам-самкам композиции siIL-89-153/LTP в дозе 162,5 мг/кг, не вызвало гибели животных, то есть препарат переносим. Интоксикация проявлялась в снижении массы тела, сильной диуретическом эффекте, взъерошенности шерстного покрова и судорогах задних конечностей. Признаки выраженной интоксикации наблюдались в течение первых суток после введения и сохранялись в течение трех дней. На 14 сутки масса тела подопытных животных была ниже, чем контрольных.
При однократном внутрибрюшинном введении мышам самкам препарата siIL-89-153/LTP LD84 составляет 650 мг/кг, a LD50=325 мг/кг, что позволяет отнести данную композицию к малотоксичным соединениям по классификации токсичности (IV класс токсичности, К.К. Сидоров, 1973).
Таким образом была установлена доза близкая к летальной для композиции siIL-89-153/LTP при внутрибрюшинном введении: LD84=650 мг/кг, гибель животных наступает в первые часы. Выявлено, что LD50 при внутрибрюшинном введении мышам составляет 325 мг/кг, что в 6 500 раз превышает разовую предполагаемую дозу для человека (0,05 мг/кг). По классификации токсичности веществ (по К.К. Сидорову) композиция siIL-89-153/LTP относится к IV классу малотоксичных соединений. Также установлено, что максимальная переносимая мышами доза композиции siIL-89-153/LTP при внутрибрюшинном введении - 162,5 мг/кг.
Таким образом, было доказано, что заявляемое изобретение - малотоксичная композиция siIL4-89-153/LTP, подавляющая экспрессию про-воспалительного гена IL-4, может применяться в составе лекарственных препаратов для лечения IL-4-опосредованных патологий таких как аллергическая бронхиальная астма и аллергический ринит.
Состав композиции siIL4-89-153/LTP был установлен в результате теоретического анализа структурных особенностей гена IL-4 человека и известных пептидных конструкций, а также в результате экспериментальных исследований. Проведенные эксперименты показали, что выбранные и синтезированные молекулы siRNA (siIL4-89 и siIL4-153) значительно снижали экспрессию гена-мишени (IL-4), а выбранный и синтезированный катионный пептид LTP эффективно доставлял нуклеиновые кислоты в различные типы клеток млекопитающих при соотношении LTP/нуклеиновая кислота 25/1 по массе. Также экспериментально подтверждено, что композиция siIL4-89-153/LTP эффективно подавляла экспрессию IL-4 человека. В исследования токсичности созданной композиции siIL4-89-153/LTP установлена полу-летальная доза LD50=325 мг/кг, что позволяет отнести ее к малотоксичным соединениям по классификации токсичности (IV класс токсичности, К.К. Сидоров, 1973).
| название | год | авторы | номер документа |
|---|---|---|---|
| Композиция, подавляющая экспрессию генов интерлейкина-4 и интерлейкина-13, для терапии аллергического ринита | 2018 |
|
RU2710895C1 |
| Способ лечения аллергической бронхиальной астмы, основанный на подавлении экспрессии генов цитокинов IL-4 и IL-13 с использованием молекул миРНК | 2016 |
|
RU2615463C1 |
| СПОСОБ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ БРОНХИАЛЬНОЙ АСТМЫ, ОСЛОЖНЯЮЩИХ ЕЕ РЕСПИРАТОРНЫХ ВИРУСНЫХ ИНФЕКЦИЙ И ДРУГИХ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ ДЫХАТЕЛЬНЫХ ПУТЕЙ | 2012 |
|
RU2526146C2 |
| Использование композиции, состоящей из катионного пептида LTP и молекул РНК против респираторных вирусов | 2015 |
|
RU2609857C1 |
| СРЕДСТВО ДЛЯ ВНУТРИКЛЕТОЧНОЙ ДОСТАВКИ НУКЛЕИНОВЫХ КИСЛОТ В КЛЕТКИ МЛЕКОПИТАЮЩИХ | 2014 |
|
RU2572575C1 |
| Пептиды для внутриклеточной доставки нуклеиновых кислот | 2020 |
|
RU2771605C2 |
| Комбинированное лекарственное средство, обладающее противовирусным эффектом в отношении нового коронавируса SARS-CoV-2 | 2021 |
|
RU2746362C1 |
| Меченые дендримерные пептиды | 2016 |
|
RU2611399C1 |
| Способ оценки эффективности проведения аллерген-специфической иммунотерапии при аллергическом рините | 2019 |
|
RU2700788C1 |
| Средство для подавления экспрессии генов, связанных с накоплением холестерина макрофагами человека | 2018 |
|
RU2698199C1 |















Изобретение относится к медицине и касается композиции для подавления экспрессии гена цитокина интерлейкина-4 посредством механизма интерференции РНК, состоящей из катионного дендримерного пептида LTP с формулой (Arg)8(Lys)4(Lys)2Lys-Ala-Cys-NH2, выступающего в качестве носителя, и двух молекул siRNA с формулами 5-UUGAUGAUCUCCUGUAAGGtt-3 и 5-AAAGAUGUCUGUUACGGUCtt-3. Изобретение обеспечивает создание малотоксичной композиции, подавляющей экспрессию гена интерлейкина-4. 2 з.п. ф-лы, 18 ил., 14 табл., 4 пр.
1. Композиция для подавления экспрессии гена цитокина интерлейкина-4 посредством механизма интерференции РНК, состоящая из катионного дендримерного пептида LTP с формулой (Arg)8(Lys)4(Lys)2Lys-Ala-Cys-NH2, выступающего в качестве носителя, и двух молекул siRNA с формулами 5-UUGAUGAUCUCCUGUAAGGtt-3 (SEQ ID NO 57) и 5-AAAGAUGUCUGUUACGGUCtt-3 (SEQ ID NO 59).
2. Композиция для подавления экспрессии гена цитокина интерлейкина-4 по п. 1, отличающаяся тем, что при отношении количества положительных зарядов пептида к количеству отрицательных зарядов нуклеиновых кислот от 24 до 96 она способна эффективно трансфецировать различные типы клеток.
3. Композиция для подавления экспрессии гена цитокина интерлейкина-4 по п. 1, отличающаяся тем, что при LD50, равном 325 мг/кг, она является малотоксичной.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| TRIPATHI P., et al., A novel mechanism for ERK-dependent regulation of IL4 transcription during human Th2-cell differentiation | |||
| Immunol Cell Biol | |||
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |

