Изобретение относится к области генной инженерии и биохимии, а также к медицине и фармацевтике.
В настоящее время разрабатывается и испытывается несколько десятков лекарственных средств, предназначенных для генной терапии онкологических, воспалительных, инфекционных заболеваний, а также заболеваний, связанных с нарушением обмена веществ. В составе таких генно-терапевтических препаратов присутствуют нуклеиновые кислоты (НК) в виде молекул дезоксирибонуклеиновой кислоты (ДНК) и рибонуклеиновой кислоты (РНК). Также в последнее время большой интерес представляет разработка РНК и ДНК вакцин, например, в отношении вируса гепатита С. Данные вакцинные препараты также в своем составе содержат НК. К тому же открытие феномена - РНК-интерференции, как способа регуляции генной экспрессии, существенно подтолкнуло исследователей и фармацевтические компании к разработке генно-терапевтических препаратов, содержащих в своем составе молекулы миРНК (малые интерферирующие РНК).
Однако все подобные препараты способны проявлять свою биологическую активность только после проникновения содержащихся в них молекул НК в цитоплазму или ядро клеток. В то же время, известно, что самопроизвольно ни ДНК, ни РНК не способны преодолевать цитоплазматическую или ядерную мембрану. В связи с этим, главным препятствием для внедрения генно-терапевтических препаратов в медицинскую практику является отсутствие эффективных, малотоксичных средств доставки НК в клетки-мишени.
Также известно, что «голая» нуклеиновая кислота может быть локально доставлена в отдельные органы и ткани, например, мышцы или печень, с помощью физических методов, таких как гидродинамические инъекции или «бомбардировка» молекулами ДНК. Помимо низкой эффективности эти методы невозможно использовать для системной доставки генетического материала, кроме того, они являются экономически затратными. Поэтому большее внимание уделяется исследованиям по созданию систем доставки как вирусной, так и невирусной природы.
Из имеющихся способов, вирусные системы пока являются наиболее эффективными транспортными средствами доставки нуклеиновых кислот. Однако, несмотря на значительные усилия в этой сфере за последние 15 лет, реализация их в клинической практике осложняется, что связано с их недостатками. В частности, вирусные векторы вызывают нежелательный иммунный ответ, а также приводят к инсерционному мутагенезу. Уже имеется значительный негативный опыт использования вирусных векторов у человека. Так, например, 18-летний волонтер Гелсингер умер во время клинических исследований генно-терапевтического препарата, где в качестве вектора использовался аденовирус. Также известен печальный эпизод, когда у 30% детей развилась лейкемия после использования препарата, сконструированного на основе ретровируса и предназначенного для лечения врожденного иммунодефицита.
Поэтому наиболее привлекательным, с точки зрения безопасности, решением является разработка невирусных систем доставки молекул ДНК и РНК на основе катионных липосом и пептидов. Этим соединениям присущи такие важные свойства, как неинфекционность, низкая иммуногенность и биодеградируемость. Как липосомы, так и пептиды способны образовывать комплексы с отрицательно заряженными НК и конденсировать их в компактные наноструктуры. Комплексование, с одной стороны, обеспечивает защиту НК от действия нуклеаз, а с другой, способствует их транслокации через клеточные мембраны, обычно, путем эндоцитоза. В качестве НК-связывающих агентов могут выступать катионные синтетические полимеры (например, полиэтиленимины), основные полиаминокислоты (полилизины, полиаргинины), хитозаны, катионные пептиды различного состава (например, пептид из белка ТАТ-ВИЧ) и липосомы. Степень компактизации НК, т.е. физический размер формируемого комплекса НК/носитель, сильно влияет на эффективность его транслокации через клеточную мембрану. Эффективность также зависит от суммарного заряда комплекса, который, в свою очередь, определяется отношением количества положительных зарядов носителя к количеству отрицательных фосфатных групп НК (величина R). Обычно в составе комплекса катионный носитель находится в избытке, обеспечивая суммарный позитивный заряд всего комплекса, что повышает эффективность его проникновения в клетки, т.к. поверхность клетки обычно несет отрицательный заряд. Кроме того, при взаимодействии носителя с НК происходит увеличение размера комплекса носитель/НК по сравнению с исходными размерами отдельных компонентов до их взаимодействия, при этом оптимальным для эффективного проникновения в клетки размером формируемых наночастиц является не более 200-300 нм.
В настоящее время липосомы, по сравнению с пептидами, наиболее широко используются в качестве стандартных носителей для трансфекции клеток в научных исследованиях (in vitro), но высокий уровень токсичности и низкая стабильность делает их малоприменимыми в медицине. Тем не менее на данный момент имеется ряд липосомных соединений, разработанных зарубежными фармацевтическими компаниями Alnylam, Silence Therapeutics, Tekmira и Calando Pharmaceuticals, которые успешно проходят клинические исследования. Анализируя результаты доклинических и клинических исследований липосом, можно заключить, что высокоэффективные варианты, разработанные на сегодняшний день, как правило, оказываются достаточно токсичными. Поэтому их использование у человека разрешено лишь для пациентов с онкологическими заболеваниями и гепатитом C, поскольку для лечения данных патологий допускается применение лекарственных средств с низким коэффициентом безопасности (например, химиотерапия). Таким образом, создание эффективных и при этом малотоксичных средств доставки молекул ДНК и РНК в клетки продолжает оставаться актуальной задачей для медицины.
В связи с высокой токсичностью липосомных конструкций наиболее перспективными выглядят менее токсичные и более стабильные катионные пептидные носители. К тому же в живых организмах трансмембранный транспорт обычно обеспечивают похожие специализированные белки, содержащие позитивно-заряженные аминокислотные последовательности. Сами по себе такие последовательности представляют собой позитивно заряженные пептиды и проявляют высокую транспортную активность. Их обозначают термином cell-penetrating peptides (СРР), т.е. проникающие пептиды (ПП). Среди различных изученных ПП одни представляют собой фрагменты природных белков, другие построены из комбинаций различных участков белка, дизайн некоторых ПП основан на чисто умозрительных гипотезах с учетом структурных соображений. Общим для всех ПП является наличие положительного заряда при физиологическом значении рН, их длина составляет 8-30 аминокислот. Установлено, что транспорт ПП через клеточную мембрану осуществляется рецептор-независимым путем, преимущественно путем эндоцитоза.
Главными преимуществами использования синтетических пептидов над липосомами являются их низкая токсичность ввиду их быстрой деградации в клетке-мишени, и практически неограниченные возможности в дизайне структур, которые могут представлять собой линейные, циклические и дендримерные пептиды с желаемым суммарным зарядом, трехмерной структурой и энантиомерным составом. Кроме того, привлекательной является их устойчивость при хранении, сравнительная дешевизна методов их синтеза и масштабируемость производства, причем в полностью в автоматическом режиме. По этим параметрам пептиды имеют явное конкурентное преимущество по сравнению с липосомами как средством доставки.
Одним из перспективных вариантов доставки НК являются дендримерные соединения. Дендримеры представляют собой разветвленные трехмерные структуры с плотным ядром и внешним слоем, состоящим из одних и тех же заряженных группировок. По аналогии с липоплексами и полиплексами их называют еще дендриплексами. Наличие большого количества концевых заряженных групп они способны эффективно связывать НК. Например, молекула дендримера на основе полипропиленимина содержит 64 (100%) концевых аминогрупп (5-я генерация, DAB 64), способных акцептировать протоны от НК при нейтральном рН. Их использование в качестве агентов доставки молекул НК в значительной степени сосредоточено на использовании полиамидоаминов (РАМАМ). В 1993 г. впервые были опубликованы результаты исследований о том, что молекулы ДНК, содержащие гены люциферазы и β-галактозидазы, могут быть доставлены в клетки с помощью РАМАМ-дендримеров. В 1996 г. появились данные исследований об эффективности трансфекции генетического материала при помощи дендримеров на различных клеточных линиях. Было показано, что различные поколения протонированных дендримеров взаимодействуют с отрицательно заряженными плазмидными ДНК, и этот комплекс стабилен при физиологических условиях даже в присутствии додецилсульфата натрия. Результативность трансфекции зависела от типа дендримеров, в некоторых случаях эффективность трансфекции в 10-100 раз превышала эффективность известных коммерческих катионных липидов.
Дендримеры могут также применяться для переноса генов с целью лечения злокачественных новообразований. При генетической терапии рост опухоли может быть заторможен с помощью контроля ангиогенеза. Так, при исследовании эффективности дендримеров как носителей генов при раке молочной железы РАМАМ-дендримеры ассоциировали с 36-мерными анионными олигомерами для доставки ангиостатина и генов тканевого ингибитора металлопротеиназы (TIMP-2). В результате трансфекции in vitro при разработке заявляемого изобретения было показано значительное уменьшение пролиферации эндотелиальных клеток и торможение развития эндотелиальных и раковых клеток. На конечном этапе работы трансфер генов в опухолевую область был проверен на мышах in vivo, у которых наблюдалось существенное снижение роста опухолей.
Также было показано, что формирование дендриплекса между дендримером и НК предотвращает их взаимодействие с сывороточным альбумином, которое, как известно, является основным препятствием для доставки свободных НК в пораженные органы и ткани при генной терапии.
Таким образом, катионные дендримерные пептидные носители представляют собой перспективный класс соединений для внутриклеточной доставки молекул ДНК и РНК при использовании в составе генно-терапевтических лекарственных средств.
Известны изобретения для внутриклеточной доставки НК.
Так, первые патенты по доставке молекул НК были связаны с использованием дендримеров РАМАМ-типа, полученные German and Szoka (США) в 1998, и в дальнейшем они служили основой для многочисленных модификаций при получении препаратов для доставки генов (см. “Recent patents in dendrimers for nanomedicine: evolution 2014 - Recent Patents on Nanomedicine, 2014, 4 (1), 000-0001877-9123/14 © 2014 Bentham Science Publishers).
Также из уровня техники известно, что аргинин-содержащие пептиды и ковалентно связанные с молекулами НК хорошо проникают в клетки и обеспечивают их трансфекцию с эффективностью, даже лучшей, чем известные проникающие пептиды из природных белков, таких как ТАТ и Пенетратин (Luo К, Li С, Wang G, Nie Υ, Не В, Wu Y, Gu Ζ. Peptide dendrimers as efficient and biocompatible gene delivery vectors: Synthesis and in vitro characterization. J. Control Release, 2011, 55(1):77-87). Причем дополнительное введение в терминальные ветви дендримера остатков глицина или ε-аминогексановой кислоты и гистидина может повышать трансфецирующую активность данных конъюгатов, по-видимому, за счет увеличения гибкости пептидных цепей и менее жесткому связыванию НК с дендримером. Тем не менее в данном изобретении, как и в ряду других, осуществляется не электростатическое формирование комплекса НК/носитель, а их жесткое ковалентное взаимодействие. Такой принцип формирования комплекса значительно усложняет практическую реализацию изобретения ввиду необходимости дополнительной химической модификации молекул носителя и НК, с последующей очисткой комплекса, что сопряжено с потерями и снижением выхода конечного продукта.
Известно изобретение, описанное в патенте US 6376248 “Peptide-enhanced transfections», в котором авторы предлагают многочисленные композиции для трансфекции клеток эукариот, включающие комплексы НК с пептидами, где нуклеиновая кислота связана ковалентно с линейным катионным пептидом с аминокислотной последовательностью ТАТ-пептида (из ВИЧ), дендримером или липопептидом. Описанные в патенте дендримеры являются производными коммерческого дендримера РАМАМ (от компании Dendritech Inc.) с высокой молекулярной массой, к концевым группам которого присоединены остатки лизина (LysDmer) или аргинина (ArgDmer). Однако известное решение обладает повышенной токсичностью, сложно в получении, имеет ограниченный срок хранения.
Анализ заявляемого изобретения в сравнении с известным и широко используемым соединением, таким как Lipofectamine®2000 и дендримерами, описанными в патенте US 6376248, показал, что заявляемое средство обладает следующими преимуществами:
1) Благодаря наличию в его составе только природных аминокислот, заявляемое средство, так называемый лизин-транспортный пептид (LTP), обладает низкой токсичностью в отношении клеток млекопитающих, по сравнению с липосомами Lipofectamine®2000.
2) Заявляемое средство (LTP), в отличие от Lipofectamine®2000, обладает способностью доставлять в различные типы клеток млекопитающих нуклеиновые кислоты, что позволяет рассматривать его как перспективный агент для трансфекции клеток молекулами РНК и ДНК.
3) Для доставки НК в клетки требуется лишь простое смешивание водных растворов НК и заявляемого средства LTP, после чего происходит электростатическое формирование комплекса HK/LTP, т.е. в отличие от дендримеров, описанных в патенте US 6376248, не требуется сложной процедуры приготовления ковалентного комплекса (конъюгата).
4) Заявляемое средство, в отличие от липосом Lipofectamine®2000, стабильно при хранении в сухом виде.
5) Синтез LTP в сравнении с липосомами Lipofectamine®2000 и дендримеров, описанных в патенте US 6376248, легко осуществлять с использованием автоматического пептидного синтезатора.
6) В отличие от липосом Lipofectamine®2000 и дендримеров, описанных в патенте US 6376248 В1, пептид LTP содержит свободную меркаптогруппу в С-концевом цистеине, что позволяет присоединять к нему репортерные метки, необходимые для изучения фармакокинетики, которая является необходимым этапом при доклинических исследованиях.
Известен носитель для направленной доставки нуклеиновых кислот в клетки, экспрессирующие рецептор CXCR4, состоящий из последовательности-лиганда к рецептору CXCR4 с последовательностью аминокислот KPVSLSYRSPSRFFESH, линкерного участка из двух молекул G-аминогексановой кислоты, соединяющей последовательность-лиганд с последовательностью для компактизации нуклеиновых кислот, последовательности, обеспечивающей компактизацию нуклеиновых кислот и выход комплекса из эндосом CHRRRRRRHC (патент РФ №2522810), выбранный в качестве ближайшего аналога. Однако известный носитель по указанному изобретению имеет большие размеры, что не позволяет эффективно осуществить доставку нуклеиновых кислот в клетки млекопитающих, не имеющих рецептора CXCR4. Также известный носитель имеет иную структуру и иной механизм проникновения (рецептор-опосредованный), нежели заявляемое средство LTP.
Поиск по источникам научно-технической и патентной литературы показал, что заявляемое средство LTP в доступных источниках не описано.
Задачей изобретения является создание малотоксичного эффективного средства для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих.
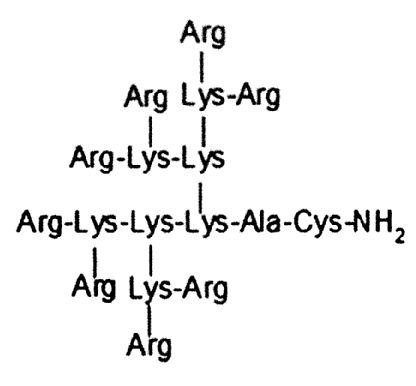
Поставленная задача решается за счет того, что средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих представляет собой катионный дендримерный пептид, содержащий 8 остатков аргинина, 7 остатков лизина, аланин и цистеин со структурной формулой (Arg)8(Lys)4(Lys)2LysAlaCys-NH2.
При отношении количества положительных зарядов к количеству отрицательных фосфатных групп нуклеиновых кислот, равном или выше 8, средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих оно полностью взаимодействует с плазмидной ДНК.
При отношении количества положительных зарядов к количеству отрицательных фосфатных групп нуклеиновых кислот от 24 до 96 средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих способно эффективно трансфецировать различные типы клеток.
При ЛД50 для 0,8-1,5 мг/мл средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих является малотоксичным.
Изобретение поясняется чертежами.
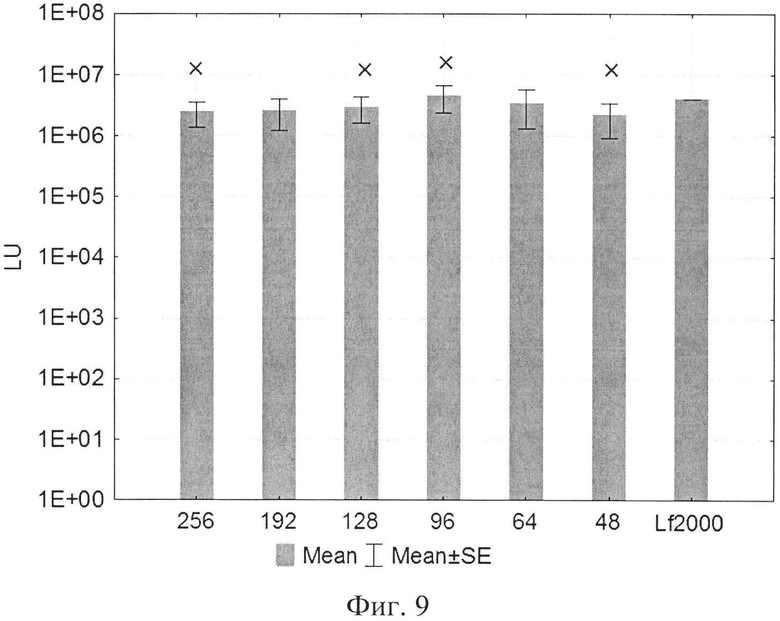
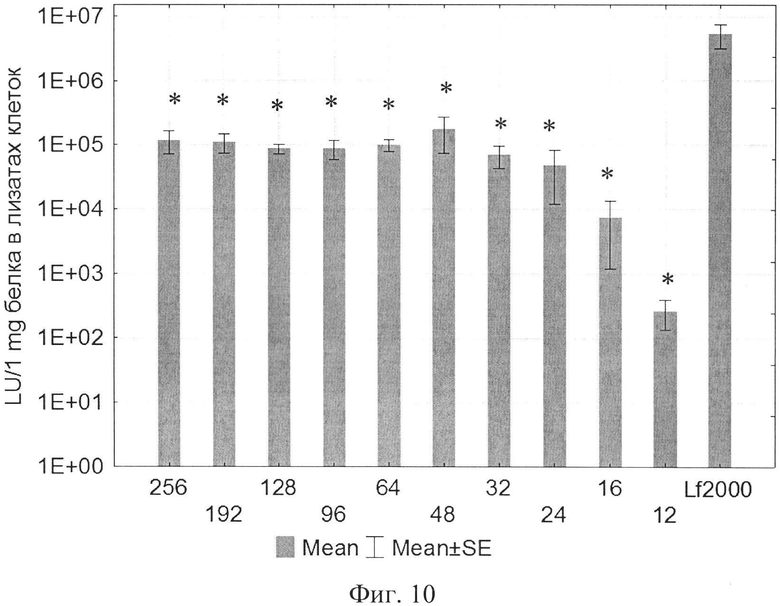
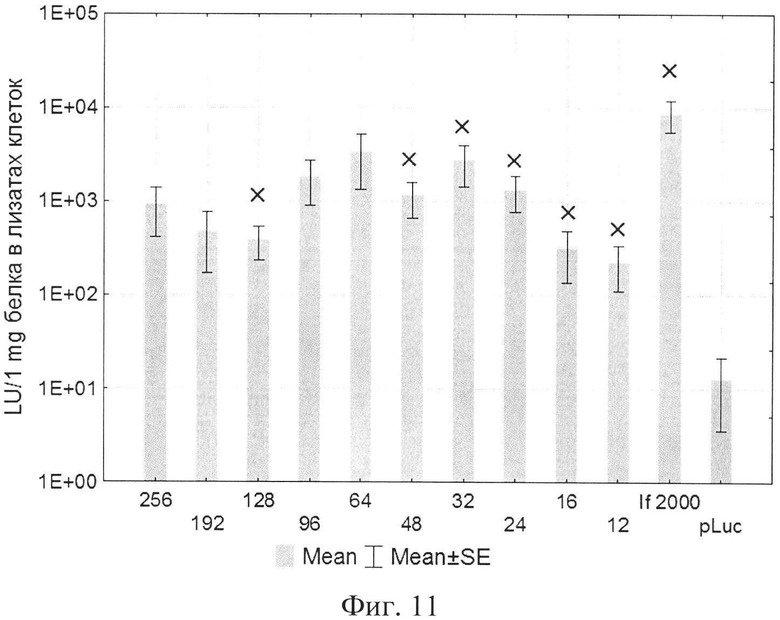
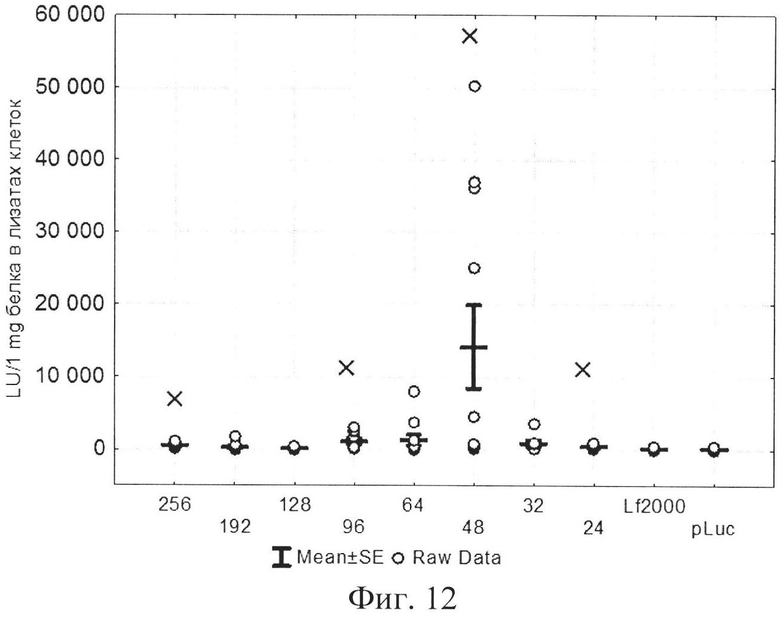
На фиг. 1 представлена структура катионного дендримерного пептида LTP в двух видах: А - аминокислоты представлены в трехбуквенном обозначении, Б - аминокислоты представлены в однобуквенном обозначении, на фиг. 2 представлены копии полученных результатов при аналитической высокоэффективной жидкостной хроматографии (ВЭЖХ) очищенного пептида LTP, на фиг. 3 представлены копии масс-спектра очищенного пептида LTP (MALDI-TOF, прибор BRUKER Microflex LT), на фиг. 4 представлены химические формулы основных реагентов, применяемых в синтезе LTP, на фиг. 5 представлена схема цикла основных операций твердофазного синтеза пептида, на фиг. 6 представлены микрофотографии LTP и смеси LTP с плазмидой pGL3 (LTP/pGL3), на фиг. 7 представлены фотографии интенсивности окраски плазмидной ДНК (pGL3) и комплекса LTP/pGL3 после электрофоретического разделения в 1% агарозном геле, на фиг. 8 представлена диаграмма эффективности трансфекции клеток 293Т с использованием LTP, на фиг. 9 представлена диаграмма эффективности трансфекции клеток HelaHI с использованием LTP, на фиг. 10 представлена диаграмма эффективности трансфекции клеток А549 с использованием LTP, на фиг. 11 представлена диаграмма эффективности трансфекции клеток МТ4 с использованием LTP, на фиг. 12 представлена диаграмма эффективности трансфекции не стимулированных клеток селезенки мыши с использованием LTP.
Средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих представляет собой катионный дендримерный пептид, при этом он содержит 8 остатков аргинина, 7 остатков лизина, аланин и цистеин со структурной формулой (Arg)8(Lys)4(Lys)2LysAlaCys-NH2. На фиг. 1 представлена структура заявленного катионного дендримерного пептида.
Было установлено, что средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих полностью взаимодействует с плазмидной ДНК, если отношение количества положительных зарядов к количеству отрицательных фосфатных групп нуклеиновых кислот равно или выше 8.
Также было установлено, что средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих при отношении количества положительных зарядов к количеству отрицательных фосфатных групп нуклеиновых кислот от 24 до 96 способно эффективно трансфецировать различные типы клеток.
В результате проведенных экспериментов было доказано, что заявленное средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих при ЛД50 для 0,8-1,5 мг/мл является малотоксичным.
В предлагаемом изобретении был проведен выбор оптимальной структуры катионного дендримерного пептида - LTP.
Пока не существует определенного алгоритма выбора оптимальной конструкции носителя НК, поскольку трансфекция - сложный, многофакторный процесс, многие ступени которого еще плохо изучены, особенно механизмы транслокации клеточной мембраны и внутриклеточных барьеров. Поэтому дизайн таких структур, в том числе катионных дендримерных пептидов, базируется на обзоре известных литературных данных, анализе функционирования природных транспортных белков и экспериментальной проверке изменений различных структурных особенностей. Проведенный предварительный анализ обусловил выбор 22 катионных пептидных последовательностей в качестве предполагаемых эффективных носителей молекул НК (таблица 1).
Выбранные структуры можно подразделить на три основные группы: линейные пептиды, липопептиды, содержащие привязанный к N-концу остаток пальмитиновой кислоты, и - дендримерные пептиды, имеющие ветвления по аминогруппам лизина. Дальнейший выбор оптимальной структуры носителя базировался на результатах экспериментов по трансфекции клеточных культур.
Для этого все соединения с различной структурой, представленные в таблице 1, были синтезированы твердофазным методом, очищены и использованы в экспериментах по изучению эффективности трансфекции. Трансфекционная активность синтезированных структур оценивалась в эксперименте in vitro с использованием клеточной линии HEK293Т (эмбриональная почка человека). Различные концентрации синтезированных пептидов индивидуально смешивались с одинаковым количеством (0,25 мкг) плазмидной ДНК, несущей репортерный ген GFP (зеленый флуоресцирующий белок); плазмида pUCHR IRES GFP. Конечный объем смеси = 80 мкл питательной среды optiMEM (Gibco, США), не содержащей антибиотики и эмбриональную телячью сыворотку (ЭТС). Смесь пептида и ДНК инкубировалась при комнатной температуре в течение 30 минут, при этом происходило образование электростатического комплекса пепетид/ДНК. После этого весь объем смеси (80 мкл) был внесен в предварительно подготовленную культуру клеток НЕК293Т. Культура клеток готовилась заранее в 48-луночном плейте (Costar, США): количество клеток 75000 в объеме 300 мкл полной питательно среды ДМЕМ (Gibco, США), содержащая 10% ЭТС (Hyclone, США) и 80 мг/л антибиотика Гентамицина (Gibco, США). Далее после внесения смеси в лунки с клетками их инкубировали 48 часов при 37 C в 5% атмосфере СО2. После инкубации осуществляли оценку эффективности проникновения плазмиды pUCHR IRES GFP методом флуоресцентной микроскопии. Эффективность проникновение ДНК в клетки была прямо пропорциональная интенсивности флуоресценции клеток, т.е. чем эффективнее пептид способствует внутриклеточному проникновению ДНК, тем выше интенсивность флуоресценции.
По результатам данного исследования (таб. 1) можно заключить, что наибольшую трансфекционную активность из изученных пептидов проявлял дендримерный катионный пептид с формулой (R)8(K)4(K)2KAC-NH2, обозначенный нами как LTP, являющийся объектом данного изобретения.

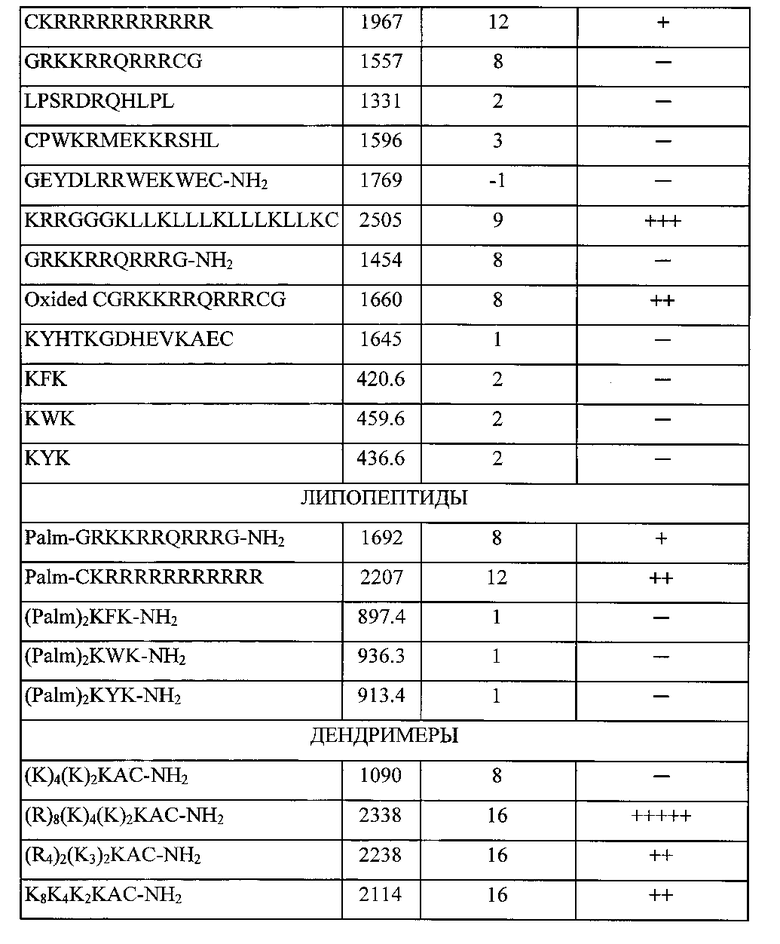
Обозначения, используемые в таблице 1:
«-» - отсутствует трансфекционная активность (нет флуоресцирующих клеток);
«+» - незначительная трансфекционная активность (присутствуют единичные флуоресцирующие клетки, менее 1%);
«++» - низкая трансфекционная активность (1-10% флуоресцирующих клеток);
«+++» - умеренная трансфекционная активность (10-25% флуоресцирующих клеток);
«++++» - выраженная трансфекционная активность (25-50% флуоресцирующих клеток);
«+++++» - высокая трансфекционная активность (более 50% флуоресцирующих клеток);
Трансфекционная эффективность указанных катионных пептидов оценивалась по интенсивности флуоресценции клеток.
Таким образом, структурная особенность заявленного изобретения такова (фиг. 1), что катионный дендримерный пептид содержит 16 гуанидиновых групп на внешнем слое. Благодаря этому, в водном растворе и в широком диапазоне pH пептид имеет сильный позитивный заряд (+16), способствующий его комплексованию с отрицательно заряженной нуклеиновой кислотой и взаимодействию с клеткой.
Синтез и очистку катионного дендримерного пептида LTP проводят твердофазным методом, используя стандартный протокол Fmoc-химии и N-гидроксибензотриазол/диизопропилкарбодиимидный (ГБТ/ДИП) метод активации Fmoc-аминокислот. В качестве полимера-носителя используют коммерческие реактивы: смолу Rink Amide МВНА, или Wang. В качестве исходных производных аминокислот применяют: Fmoc-Cys(Trt)-OH, Fmoc-Ala-OH, Fmoc-Lys(Boc)-OH, Fmoc-Arg(Pbf)-OH (фиг. 4). Стандартный цикл включет промывку (ДМФА), удаление Fmoc-защиты (20% пиперидин в ДМФА), предварительное активирование Fmoc-аминокислоты (ДИП/ГБТ) и конденсацию в среде ДМФА/N-метилпирролидон при 2-кратном избытке карбоксильного компонента (~0,5 часа). Контроль над полнотой реакции осуществляют нингидриновым методом и при необходимости реакцию конденсации повторяют. Конечные пептиды отщепляют от полимера трифторуксусной кислотой в присутствии скавенджеров (тиоанизол, этандитиол, фенол, диметилсульфид).
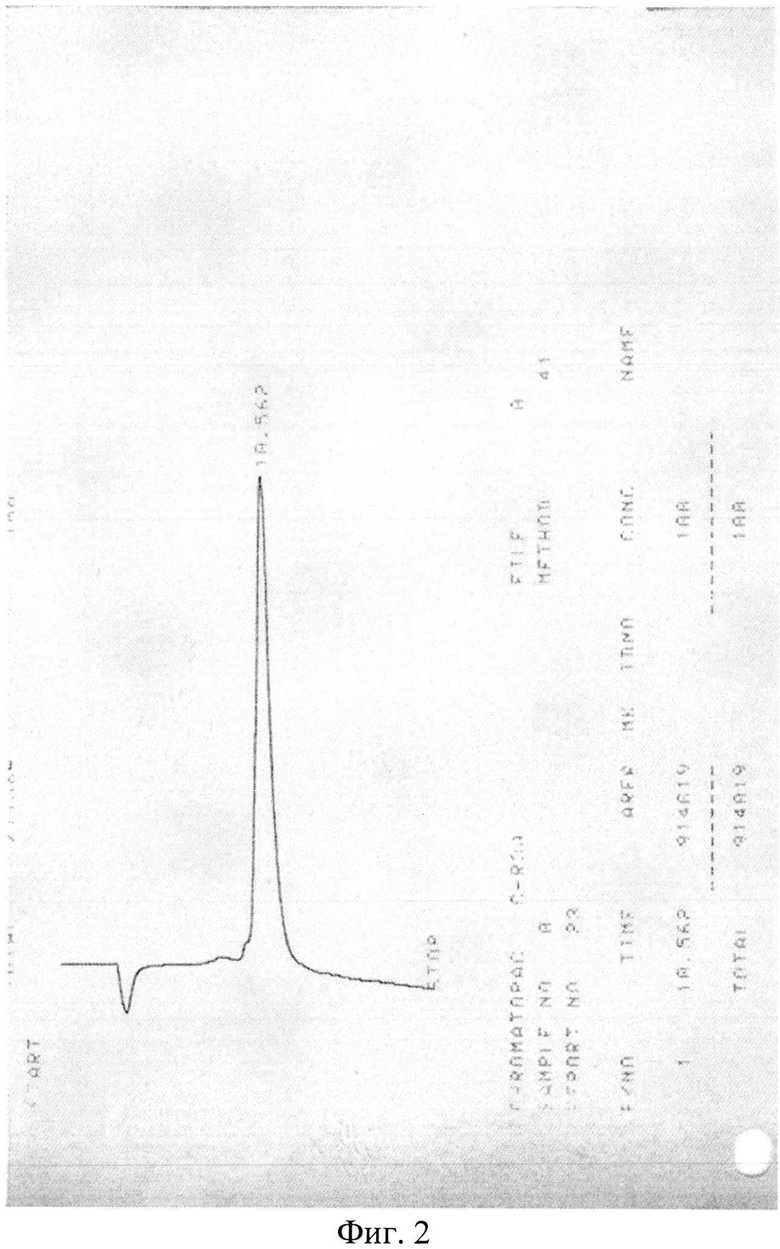
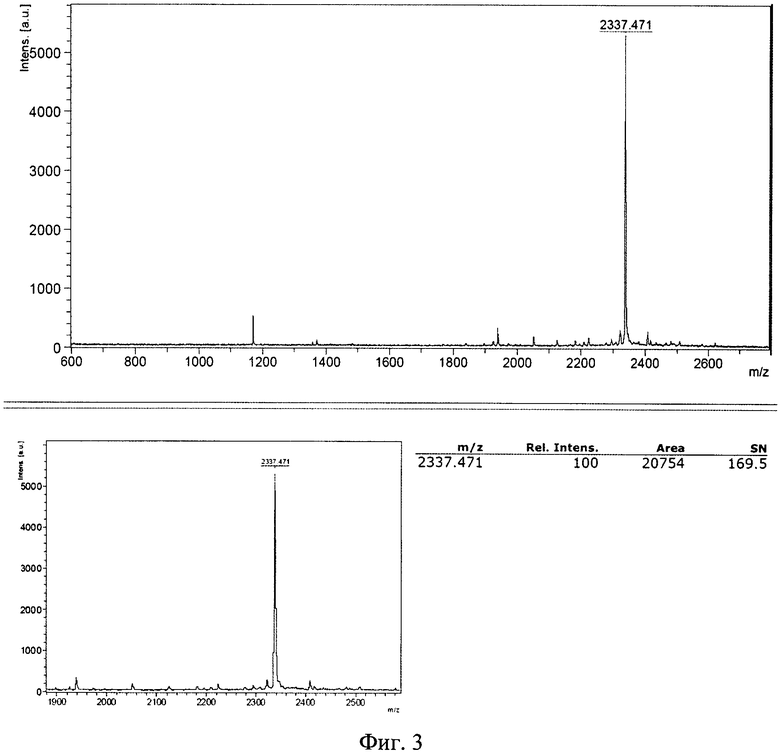
Сырой продукт высаждают сухим серным эфиром, экстрагируют водной уксусной кислотой и экстракт лиофилизируют. Очистку пептида проводят методом высокоэффективной жидкостной хроматографии (ВЭЖХ) (фиг. 2), например, на колонке Grace Vydac 218 ТР54, а структуру очищенного пептида подтверждают масс-спектрометрией (фиг. 3).
Полученные параметры стандартного образца LTP: время удерживания на аналитической колонке: 10,1±0,2 мин, масс-спектр (MALDI-TOF): m/z=2337,471±1 при расчетной массе: 2337,98.
Для аналитической высокоэффективной жидкостной хроматографии (ВЭЖХ) очищенного пептида LTP была использована аналитическая колонка Grace Vydac 218ТР54 (150×4.6 мм), содержащая 5 мкм сорбент С18. Подвижная фаза: 0,1% водный раствор трифторуксусной кислоты (А) и ацетонитрил (Б) - смесь, 1 мл/мин. Градиент, % состав: 0-2 мин (А - 100%); 2-25 мин (Б - линейно от 0 до 70%); 25-27 мин (Б - 70%); 27-29 мин (Б - линейно от 70 до 0%); 29-35 мин (А - 100%).
Пример 1.
Синтез осуществлялся следующим образом.
На фиг. 4 представлены химические формулы основных реагентов, применяемых в синтезе LTP.
Схема твердофазного синтеза основана на повторяющемся цикле стандартных операций (фиг. 5), который включает:
1) На старте навеску исходной смолы Rink Amide вносят в реактор, добавляют ДМФА, энергично перемешивают суспензию до полного набухания смолы, операцию повторяют 3 раза по 3 мин, удаляя растворитель фильтрованием через пористое дно под вакуумом или под давлением.
2) Удаление Fmoc-защиты 20% раствором 4-метилпиперидина (МПП) в течение 10 мин.
3) Промывка смолы.
4) Добавление в реактор активированной Fmoc-аминокислоты (см. 5.2.6) в среде ДМФА/г4-метилпирролидон (МП) для присоединения очередной Fmoc-аминокислоты и инкубацией в течение 30 мин.
5) Промывка смолы.
6) Анализ на полноту реакции присоединения нингидриновым методом (см. пункт 5.2.9),
Затем операции по пунктам 2-6 повторяются. Если нингидриновый тест показывает наличие свободной аминогруппы (окрашивание зерен смолы), то операции по пунктам 4 и 5 повторяются, пока нингидриновый тест не покажет отрицательной реакции.
На фиг. 5 представлена схема цикла основных операций твердофазного синтеза пептида. Использованы следующие обозначения: Fmoc-AA -Fmoc-аминокислота, n=1, 2, 3 и т.д..
Нингидриновый тест (анализ полноты реакции конденсации) проводят путем отбора пробы смолы (1-3 мг гранул смолы) в стеклянную пробирку и добавления к ней 0.5 мл раствора нингидрина (2% раствор в н-бутаноле) выдерживанием пробирки в термостате при 110°С 5 мин. Отсутствие окрашивания гранул смолы (малиновый цвет) свидетельствует о полноте (~100%) прохождения реакции. При наличии окрашивания гранул реакцию конденсации (п. 3) повторяют.
Количества реагентов рассчитаны на синтез LTP, исходя из загрузки в реактор 3,0 г смолы Fmoc-RinkAmide (1.74 ммоль Fmoc - из расчета 0,58 ммоля/г смолы, фирма AnaSpec).
Arg8Lys4Lys2LysAla-Cys-NH2
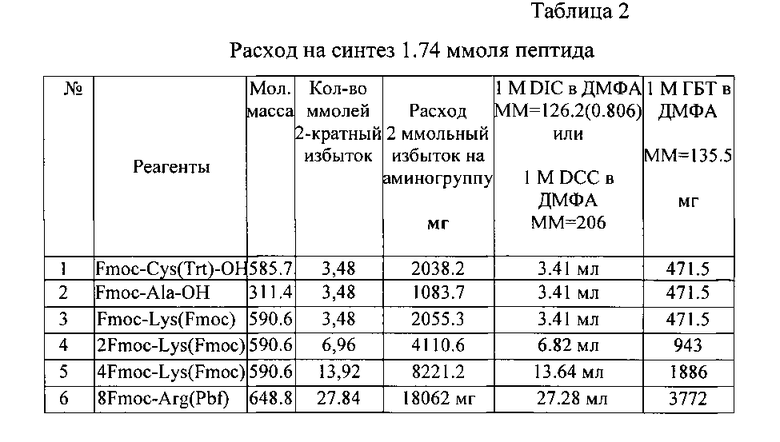
После последней конденсации (присоединения Fmoc-Arg(Pbf)-OH) стандартную промывку дополняют промывкой этанолом и диэтиловым эфиром и затем высушиванием конечной Fmoc-пептидил-смолы в вакууме до постоянного веса.
Отщепление пептида от смолы проводят в отдельном сосуде путем перемешивания суспензии высушенной Fmoc-пептидил-смолы в смеси 2,25 г кристаллического фенола, 0,75 мл 1,2-этандитиола, 1,5 мл тиоанизола, 1,5 мл диет, воды и 30 мл трифторуксусной кислоты в течение 3 часов при комнатной температуре. Затем суспензию фильтруют через стеклянную фильтровальную воронку с плотным пористым диском (№3) в круглодонную колбу. Осадок на фильтре дополнительно промывают небольшими порциями этанола и объединенный фильтрат, содержащий пептид, смешивают с 200 мл сухого диэтилового эфира. В результате последней операции образуется осадок, суспензию отстаивают, надосадочную жидкость декантируют и сливают в отходы. Остаток опять суспендируют в 100 мл диэтилового эфира, суспензию отстаивают, надосадочную жидкость декантируют и сливают в отходы. Остаток высушивают в вакууме и растворяют в 50 мл уксусной кислоты для экстракции свободного пептида и сырой экстракт фильтруют через стеклянный фильтр №3. Полученный фильтрат высушивают лиофильно (сублимационная сушка), получая в итоге сырой пептид.
Первичную очистку сырого пептида проводят путем гель-проникающей хроматографии, используя стеклянную колонку ПСК24/500 (диаметр 24 мм, длина 500 мм) с наполнителем Toyopearl HW-40 (сшитый гидроксилированный метакрилатный полимер), используя в качестве подвижной фазы 0,1% водный раствор аммиака, скорость потока 60 мл/час. Фракции вещества, выходящие с колонки, детектируют с помощью спектрофотометрического детектора при длине волны 254 нм. В колонку вносят фильтрованный (0,45 мкм фильтр) раствор 120 мг сырого пептида в 4 мл 0,1% водном растворе аммиака с концентрацией (30 мг/мл). Фракцию, соответствующую первому пику (зона свободного объема колонки), отделяют, замораживают и высушивают лиофильно, получая 84 мг полусырого пептида.
Полусырой пептид очищают путем градиентной высокоэффективной хроматографии (ВЭЖХ), используя препаративную хроматографическую систему, оснащенную препаративной колонкой (250×22 мм), содержащей 10 мкм-сорбент с обращенной С18 фазой (Vydac 218ТР1022), бинарным градиентным насосом, детектором с переменной длиной волны (длина волны установлена на 226 нм), коллектором фракций, самописцем, компьютером для управления насосом, расчета времени удерживания и площади пиков. Температура колонки поддерживается на уровне комнатной, скорость потока 10 мл/мин.
Состав подвижной фаза изменяется во времени согласно ниже приведенной диаграмме и таблице (таблица 3).
Хроматограмма полусырого пептида показывает наличие трех мажорных фракций (пиков). Данные масс-спектрометрии показывали, что фракция, содержащая требуемый пептид LTP с мол. массой 2338, соответствует пику с временем удерживания около 10 мин. Фракцию с временем удерживания 9,975 мин выделяли, высушивали лиофильно и сухой остаток повторно хроматографировали в тех же условиях. Аналитическая хроматограмма показывала один пик с временем удерживания около 10 мин (фиг. 2). Содержание пептида определяют по относительной площади пика к суммарной площади всех пиков.
Анализ гомогенности пептида проводят на той же хроматографической системе, используя аналитическую ВЭЖХ-колонку (5 мкм-сорбент Vydac 218ТР54, размеры колонки 4,6×250 мм). Содержание основного пика (площадь), время удерживания около 10 мин должно быть не менее 93%.
Подтверждение подлинности структуры пептида проводят следующим образом. Анализ проводят путем масс-спектрометрии, используя метод МАЛДИ (MALDI), матрично-активированную лазерную десорбцию/ионизацию. Масс-спектр очищенного пептида должен показывать преимущественное наличие молекулярного иона с m/z=2338±1 Да (фиг. 3).
Пример 2
Взаимодействие катионного дендримерного пептида LTP с нуклеиновыми кислотами.
О способности дендримерного пептида LTP взаимодействовать с НК можно судить по данным электронной микроскопии, путем сравнения микрофотографий пептида LTP и электростатического комплекса LTP/HK. Для этого были получены микрофотографии раствора LTP в концентрации 1,12 мкг/мл в стерильном физиологическом растворе (ПанЭко, Россия) и микрофотокрафии смеси LTP с НК, концентрацией 1,12 мкг/мл (для LTP) + 0,02 мкг/мг (для НК), где в качестве НК выступала плазмида pGL3 (Invitrogen, США). Анализ микрофотографий позволят судить об увеличении размера частиц, запечатленных в смеси LTP с НК в сравнении с LTP. По всей видимости, такое увеличение в размерах частиц происходит за счет взаимодействия LTP с плазмидной ДНК (pGL3) (фиг. 6). Таким образом, можно предположить, что катионный пептид LTP способен взаимодействовать с НК и формировать электростатический комплекс LTP/HK.
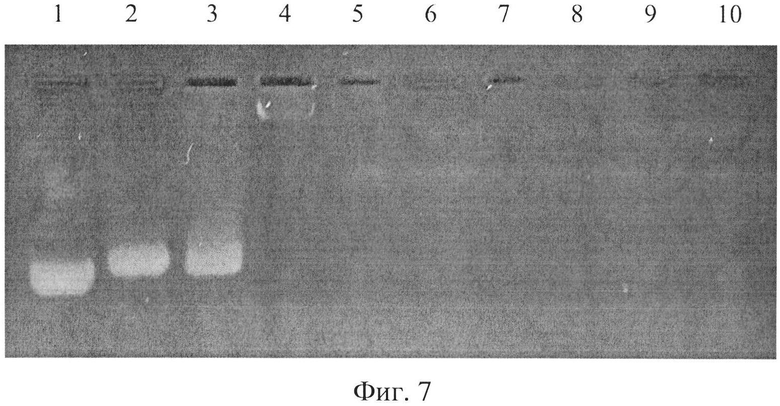
Кроме того, о взаимодействии дендримерного пептида LTP с НК можно судить по снижению интенсивности окрашивания плазмидной ДНК (pGL3) красителем этидиум бромидом. Известно, что краситель этидиум бромид окрашивает только свободную ДНК, не связанную ДНК. Для оценки влияния LTP на интенсивность окрашивания ДНК (плазмиды pGL3) был проведен следующий эксперимент. Смешивались различные концентрации пептида LTP с одинаковым количеством 0,5 мкг плазмидной ДНК (pGL3). В качестве контроля использовали 0,5 мкг pGL3 без добавления LTP. После смешивания LTP с pGL3 смесь инкубировали 30 минут при комнатной температуре для образования электростатического комплекса LTP/pGL3. После чего смесь подвергали разделению в 1% агарозном геле, содержащем 10 мкг/мл красителя этидиум бромида. Разделение осуществлялось при напряжении 80V в течение 1 часа. При этом свободная ДНК (заряженная отрицательно) двигалась к катоду (сверху вниз на фиг. 7), и при движении ДНК через гель происходило ее окрашивание этидиум бромидом, находящимся в составе геля. В то же время если происходит взаимодействие LTP с ДНК и образование электростатического комплекса LTP/ДНК, то ДНК в составе данного комплекса окрашиваться не будет. В результате оказалось, что по мере увеличения концентрации LTP в комплексе LTP/pGL3 происходит снижение интенсивности окраски ДНК (дорожки 3-10 на фиг. 7), что свидетельствует о уменьшении количества несвязавшейся ДНК в образцах (фиг. 5). Окраска ДНК отсутствует на дорожках 6-10 (фиг. 5), что свидетельствует об отсутствии несвязавшейся ДНК в данных образцах, т.е. вся ДНК полностью связывается с LTP при R=8 и более.
Дорожка 1 соответствует 0,5 мкг плазмидной ДНК - pGL3. Дорожки 2-10 соответствуют комплексу LTP/pGL3, состоящему из 0,5 мкг pGL3 и LTP при различной величине R (дорожка 2 - R=0,5; дорожка 3 - R=1; дорожка 4 - R=2; дорожка 5 - R=4; дорожка 6 - R=8; дорожка 7 - R=16; дорожка 8 - R=32; дорожка 8 - R=62; дорожка 9 - R=128; дорожка 10 - R=256). R - соотношение количества позитивных зарядов LTP к количеству отрицательных зарядов плазмидной ДНК pGL3 в комплекса LTP/pGL3.
Пример 3
Эффективная трансфекция различных типов клеток млекопитающих с помощью катионного дендримерного пептида LTP.
О способности катионного пептида LTP трансфецировать различные типы клеток свидетельствуют данные экспериментов in vitro. Для анализа трансфекционной активности катионный пептид в смешивали с плазмидной ДНК (pGL3), несущей репортерный ген люциферазы в различных соотношениях позитивного заряда LTP к отрицательному заряду молекул ДНК (величина R). Смесь пептида LTP и 0,2 мкг pGL3 готовили в объеме 80 мкл питательной среды optiMEM, после чего инкубировали при комнатной температуре в течение 30 минут для образования электростатического комплекса LTP/pGL3. После этого раствор комплекса вносили в различные типы клеток, подготовленные заранее. В экспериментах использовали следующие типы клеток млекопитающих различного происхождения: НЕК293Т - эмбриональная почка человека в количестве 75000 клеток, HelaHI - эпителиоидная карцинома шейки матки в количестве 75000 клеток, А549 - карцинома легкого в количестве 75000 клеток, МТ4 - Τ-клетки человека, выделенные от пациента с Т-клеточной лейкемией в количестве 300000 клеток, не стимулированные клетки селезенки мышей линии BALB/c в количестве 1000000 клеток. В качестве положительного контроля использовали коммерческий реагент липосомной природы Lipofectamine200 (Invitrogen, США). После внесения образов в клетки их инкубировали в течение 2 суток при температуре 37С в 5% атмосфере СO2. После инкубации клетки лизировали в 70-300 мкл буфера GLO lysis buffer (Promega, США) в течение 5-10 минут при комнатной температуре, после чего лизаты клеток центрифугировали при 10000 оборотах в минуту в течение 3-5 минут для осаждения клеточного дебриза. Надосадок в количестве 50 мкл отбирали в отдельные микропробирки для последующего анализа на количественное содержание в них фермента люциферазы. Для этого к 50 мкл лизата добавляли реагент люциферин (Promega, США) в количестве 50 мкл после чего полученная смесь излучала свет, интенсивность которого детектировалась с использование люминометра (Promega, США). Чем выше интенсивность свечения образца, тем выше содержание в нем фермента люциферазы, который экспрессируется после проникновения в клетки плазмидной ДНК - pGL3. Т.е. чем выше интенсивность излучения, тем выше трансфекционная активность образца.
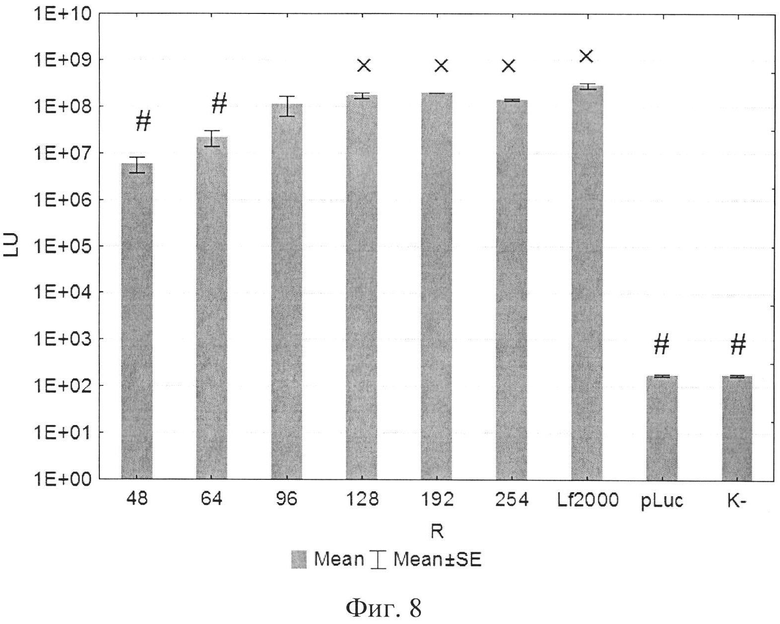
В экспериментах по трансфекции культуры клеток HEK293Т показано, что активность LTP была сравнима с коммерческим реагентом Lipofectamine2000. Оптимальное соотношение R находилось в пределах от 96 до 254 (фиг. 8). При трансфекции клеток HelaHI также наблюдалась выраженная трансфекционная активность LTP с оптимальной величиной R в пределах от 48 до 254 (фиг. 9). При изучении эффективности трансфекции клеток А549 показано, что оптимальное соотношение (R) находится в пределах от 32 до 254 (фиг. 10). При изучении эффективности трансфекции клеток МТ4 показано, что оптимальная соотношение (R) находится в пределах от 24 до 96 (фиг. 11). Также дендримерный катионный пептид LTP способен трансфецировать нестимулированные клетки селезенки мышей линии BALB/c (фиг. 12) с оптимальным R от 32 до 96.
На фиг. 8 представлена диаграмма эффективности трансфекции клеток 293Т с использованием LTP, где
R - соотношение количества положительных зарядов LTP к количеству отрицательных зарядов плазмиды pGL3;
pLuc - плазмида pGL3, кодирующая ген люциферазы luc;
LU - световые единицы (light units);
# - достоверно отличает от Lf2000 (Lipofectamine2000 (Invitrogen, США), использованный в качестве положительного контроля);
× - достоверно отличается от pLuc (плазмида pGL3 без каких-либо трансфекционных реагентов).
На фиг. 9 приняты следующие обозначения:
R - соотношение количества положительных зарядов LTP к количеству отрицательных зарядов плазмиды pGL3;
pLuc - плазмида pGL3, кодирующая ген люциферазы luc.
LU - световые единицы (light units);
Lf2000 - Lipofectamine2000 (Invitrogen, США), используется в качестве положительного контроля;
× - достоверно отличается от pLuc (плазмида pGL3 без каких-либо трансфекционных реагентов).
На фиг. 10, представляющей собой диаграмму эффективности трансфекции клеток А549 с использованием LTP, приняты следующие обозначения:
R - соотношение количества положительных зарядов LTP к количеству отрицательных зарядов плазмиды pGL3;
pLuc - плазмида pGL3, кодирующая ген люциферазы luc;
LU - световые единицы (light units);
Lf2000 - Lipofectamine2000 (Invitrogen, США), используется в качестве положительного контроля;
* - достоверно отличается от pLuc (плазмида pGL3 без каких-либо трансфекционных реагентов).
На фиг. 11 представлена диаграмма эффективности трансфекции клеток МТ4 с использованием LTP, где
R - соотношение количества положительных зарядов LTP к количеству отрицательных зарядов плазмиды pGL3;
pLuc - плазмида pGL3, кодирующая ген люциферазы luc;
LU - световые единицы (light units);
Lf2000 - Lipofectamine2000 (Invitrogen, США), используется в качестве положительного контроля;
× - достоверно отличается от pLuc (плазмида pGL3 без каких-либо трансфекционных реагентов).
На фиг. 12 представлена диаграмма эффективности трансфекции нестимулированных клеток селезенки мыши с использованием LTP, где
R - соотношение количества положительных зарядов LTP к количеству отрицательных зарядов плазмиды pGL3;
pLuc - плазмида pGL3, кодирующая ген люциферазы luc;
LU - световые единицы (light units);
Lf2000 - Lipofectamine2000 (Invitrogen, США), используется в качестве положительного контроля;
× - достоверно отличается от pLuc (плазмида pGL3 без каких-либо трансфекционных реагентов).
Таким образом, суммировав результаты трансфекции пяти различных типов клеток, можно заключить, что наиболее оптимальное соотношение LTP и НК находится в пределах от 24 до 96. Использование данных соотношений позволяет эффективно трансфецировать различные типы клеток.
Пример 4
Катионный дендримерный пептид LTP является малотоксичным соединением.
Катионный пептид LTP является малотоксичным соединением, о чем свидетельствуют данные, полученные в МТТ-тесте с использованием культуры клеток НЕК293Т (эмбриональная почка человека). Для изучения токсичности LTP в МТТ-тесте клетки НЕК293Т засевали в 96-луночный планшет (Costar, США) в 100 мкл полной питательной среды ДМЕМ (Gibco, США) в количестве 30000 клеток/лунку и инкубировали его в СO2-инкубаторе при температуре 37°C в течение 1 суток для образования монослоя клеток. После этого в клетки вносили 8 двоичных разведений пептида LTP, начиная с концентрации 3 мг/мл, и инкубировали совместно с клетками в течение 1 суток в тех же условиях. После инкубации к клеткам добавляли краситель МТТ (Sigma, США) в объеме 50 мкл и инкубировали в тех же условиях в течение 4 часов. При этом краситель окрашивал жизнеспособные клетки, таким образом интенсивность окраски лунок, в которых происходила гибель клеток, снижалась. Для высвобождения красителя из клеток в питательный раствор клетки лизировали добавлением 50 мкл раствора SDS с последующей инкубацией в течение суток в тех же условиях. Интенсивность окраски раствора в лунках измеряли с помощью планшетного фотометра (BioRad, США) при длинах волн 570 и 610 нм. В итоге по данным МТТ-теста ЛД50 для LTP колеблется в пределах 800-1500 мкг/мл. В то же время липосомные трансфекционные соединения более токсичны, их ЛД50 колеблется в пределах 7-40 мкг/мл. Таким образом заявленное изобретение - LTP является малотоксичным соединением.
Таким образом, вышеизложенное подтверждает, что заявленное изобретение обеспечивает транспорт молекул нуклеиновых кислот в цитоплазму и ядро различных типов клеток млекопитающих.
Заявленное изобретение относится к классу малотоксичных соединений.
Заявленное изобретение в дальнейшем может быть использовано в качестве нетоксичного носителя молекул ДНК и РНК в составе генно-терапевтических лекарственных средств для лечения различных заболеваний.
| название | год | авторы | номер документа |
|---|---|---|---|
| Пептиды для внутриклеточной доставки нуклеиновых кислот | 2020 |
|
RU2771605C2 |
| Меченые дендримерные пептиды | 2016 |
|
RU2611399C1 |
| КОМПОЗИЦИЯ ДЛЯ ПОДАВЛЕНИЯ ЭКСПРЕССИИ ГЕНА ЦИТОКИНА ИНТЕРЛЕЙКИНА-4 | 2014 |
|
RU2563989C1 |
| Комбинированное лекарственное средство, обладающее противовирусным эффектом в отношении нового коронавируса SARS-CoV-2 | 2021 |
|
RU2746362C1 |
| Композиция, подавляющая экспрессию генов интерлейкина-4 и интерлейкина-13, для терапии аллергического ринита | 2018 |
|
RU2710895C1 |
| КОМПОЗИЦИИ ВАКЦИН И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2600798C2 |
| ПРИМЕНЕНИЕ ДЕНДРИМЕРНОЙ НАНОТЕХНОЛОГИИ ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ | 2010 |
|
RU2571928C2 |
| N-галлаты синтетических пептидов с антиоксидантной активностью | 2023 |
|
RU2828796C1 |
| Гистоны и биодеградируемые липиды как средство для доставки нуклеиновых кислот в клетки эукариот | 2015 |
|
RU2637371C2 |
| БИОДЕГРАДИРУЕМЫЕ КАТИОННЫЕ ПОЛИМЕРЫ | 2005 |
|
RU2440380C2 |


Изобретение относится к области генной инженерии и биохимии, конкретно к созданию малотоксичного катионного дендримерного пептида со структурной формулой (Arg)8(Lys)4(Lys)2LysAlaCys-NH2 (Фиг. 1), и может быть использовано в медицине и фармацевтике. Заявленный дендримерный пептид используют в качестве средства для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих. При отношении количества положительных зарядов к количеству отрицательных фосфатных групп нуклеиновых кислот от 24 до 96 дендримерный пептид способен эффективно проникать в различные типы клеток млекопитающих и полностью взаимодействовать с плазмидной ДНК. При ЛД50 для 0,8-1,5 мг/мл средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих является малотоксичным. 1 з.п. ф-лы, 12 ил., 3 табл., 4 пр.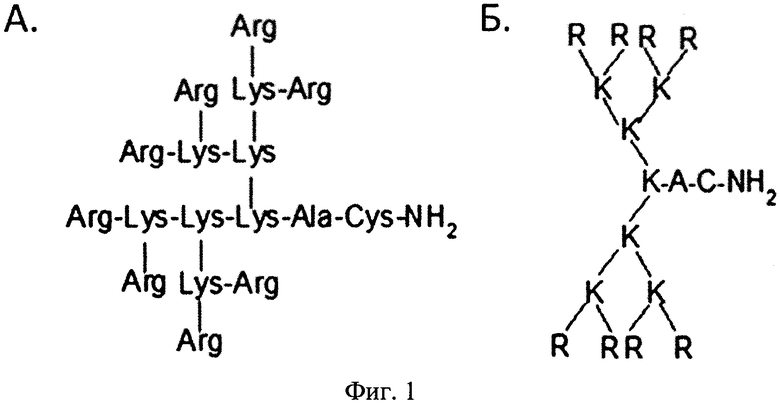
1. Средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих, представляющее собой катионный дендримерный пептид, отличающийся тем, что он содержит 8 остатков аргинина, 7 остатков лизина, аланин и цистеин со структурной формулой:
и при отношении количества положительных зарядов катионного дендримерного пептида к количеству фосфатных групп нуклеиновых кислот от 24 до 96 эффективно проникает в различные типы клеток.
2. Средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих по п. 1, отличающееся тем, что при ЛД50 для 0,8-1,5 мг/мл является малотоксичным.
| НОСИТЕЛЬ ДЛЯ НАПРАВЛЕННОЙ ДОСТАВКИ НУКЛЕИНОВЫХ КИСЛОТ В КЛЕТКИ, ЭКСПРЕССИРУЮЩИЕ РЕЦЕПТОР CXCR4 | 2012 |
|
RU2522810C1 |
| LUO К | |||
| et al., Peptide dendrimers as efficient and biocompatible gene delivery vectors: Synthesis and in vitro characterization, J | |||
| Control Release, 2011, v.155, n.1, p | |||
| Спускная труба при плотине | 0 |
|
SU77A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| US 6376248 B2, 23.04.2002 | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |

