Изобретение относится к генной инженерии, биохимии, медицине и фармацевтическим технологиям и связано с созданием малотоксичных катионных дендримерных пептидов как внутриклеточных транспортеров нуклеиновых кислот (НК) различной структуры. Трансфекция клеток НК является эффективным способом введения нового гена или, наоборот, целенаправленного подавления экспрессии собственного или чужеродного гена. Последний способ, так называемая РНК-интерференция, реализуемая за счет внутриклеточного введения малой интерферирующей РНК (миРНК), может охватывать очень широкий спектр заболеваний, включая врожденные, инфекционные, онкологические и иммунные заболевания. Открытие феномена РНК-интерференции, как способа регуляции генной экспрессии, подтолкнуло исследователей и фармацевтические компании к разработке различных вариантов генно-терапевтических препаратов, содержащих в своем составе молекулы миРНК. Для проявления специфической активности НК должны проникнуть в цитоплазму клетки-мишени, но выполнение этого условия имеет ряд трудностей. Прежде всего, свободная НК в биологической среде очень быстро разрушается под действием эндогенных нуклеаз, а также и из-за высокой гидрофильности и значительного отрицательного заряда молекулы НК практически неспособны проходить мембранный барьер клетки. Поэтому основной акцент в разработке таких препаратов связан с созданием эффективных систем доставки НК.
В качестве возможных транспортеров НК исследовались вирусы, рекомбинантные вирионы и разнообразные классы веществ, включая неорганические вещества, органические амфифильные полимеры, биополимеры, липиды, липосомы, полисахариды, наночастицы и пептиды. Хотя вирусы являются эффективными проникающими агентами, однако, ввиду потенциальной биологической опасности (возможные патогенные мутации, нежелательный иммунный ответ) применения реплицирующих агентов на практике стараются избегать. Синтетические полимеры и неорганические наночастицы часто обладают высокой токсичностью и склонны накапливаться в организме. Одним из привлекательных решений с точки зрения безопасности является применение невирусных систем на основе катионных биоразлагаемых векторов петидной и/или липидной природы в виду их низкой иммуногенности и способностью к биодеструкции. Такие агенты образуют комплексы с отрицательно заряженными НК и конденсируют их в компактные наноструктуры, и это, с одной стороны, обеспечивает защиту НК от действия нуклеаз, а с другой, способствует их транслокации через клеточные мембраны за счет эндоцитоза. Обычно в составе комплекса катионный носитель находится в избытке, обеспечивая суммарный положительный заряд всего комплекса, что повышает эффективность его проникновения в клетки, т.к. поверхность клетки обычно несет отрицательный заряд.
К перспективным транспортерам НК этой группы следует отнести катионные пептиды (КП) ввиду их ряда отличительных преимуществ. К ним можно отнести: высокую трансмембранную активность, наличие четко определенного состава/структуры, стабильность при хранении, что позволяет проводить анализ чистоты и структуры современными методами (масс-спектрометрией, высокоэффективной хроматографией, ЯМР), обеспечивая необходимую стандартизацию препарата. При этом возможности современной пептидной химии обеспечивают большие возможности в дизайне пептидных структур на основе вариации аминокислотных последовательностей, что позволяет регулировать их топологию, изменять гидрофильно-липофильный баланс, распределение отрицательных и положительных зарядов, изменять энантиомерный состав. Кроме того, привлекательным является сравнительная доступность методов их синтеза и масштабируемость, производство можно осуществлять в автоматическом режиме. Таким образом, по этим параметрам пептиды, как средство доставки, имеют явное конкурентное преимущество по сравнению с липосомами. Рассматривая классификацию КП, способных проникать в клетки, следует выделить следующие классы: фрагменты белков, специфически связывающие молекулы ДНК/РНК, белковые трансдукционные домены и сигнальные структуры, природные антимикробные пептиды (чаще их фрагменты), последовательности из вирусных и бактериальных адгезионных белков и пептиды оригинального дизайна, включая пептиды из фаговых библиотек [1].
Необходимо отметить, что устойчивость линейных КП к протеолизу в биологической среде сильно ограничена, поскольку α-амидные связи аминокислотной цепи легко расщепляются вне- и внутриклеточными протеазами [2]. Более того, линейные КП с сильным положительным зарядом могут оказаться довольно токсичными и могут разрушать клеточные мембраны, вплоть до полного лизиса клеток. Эти особенности имеют решающее значение в случае применения КП в качестве компонента терапевтических агентов. Чтобы повысить протеолитическую устойчивость, предпринимались различные подходы, включая замену L-аминокислот на D-формы, введение неприродных фрагментов, химическую модификацию пептидного скелета [3,4]. В свободном виде КП из-за электростатического отталкивания заряженных атомов, особенно в случае лизин-содержащих пептидов, имеют тенденцию приобретать удлиненную конформацию, тогда как введение в них жирной кислоты способствует образованию мицелл или липосом, что сильно увеличивает их сродство к липидным мембранам [5, 6]. Однако введение сильно гидрофобных остатков значительно снижает растворимость пептидов в водной среде. Этот вопрос обычно решается путем перевода таких липопептидов в липосомы, но снова возникает проблема стабильности и хранения липосом, поскольку они существуют только в водной среде. Надо отметить, что в некоторых работах и патентах для трансфекции применяют не электростатические комплексы, а ковалентные конъюгаты НК с пептидом-носителем, причем ковалентное связывание иногда осуществляют через меркаптогруппу цистеина, которую водят в пептид, и которая способна взаимодействовать с активированной молекулой НК. Такой подход обусловлен тем, что при попадании конъюгата внутрь клетки цитоплазматические редуктазы могут расщеплять эту связь, тем самым высвобождать НК. Однако, такая система доставки нуклеиновых кислот значительно усложняет практическую реализацию метода трансфекции, поскольку необходима дополнительная непростая химическая модификация целевой НК для введения специальной SH-реактивной группы, что может нарушить функциональную специфичность НК.
Перспективной альтернативой линейным пептидам как средству доставки являются катионные дендримерные пептиды (ДКП). ДКП представляют собой разветвленные трехмерные структуры с плотным ядром и внешним слоем, состоящим из положительно-заряженных группировок. Наличие большого количества концевых заряженных групп позволяет ДКП эффективно связывать и конденсировать НК. Применение пептидных дендримеров для трансфекции в значительной степени снимает проблему протеолитической устойчивости, ввиду того, что в структуре они содержат неприродные ε-амидные связи [2, 7, 10]. Все функциональные особенности таких пептидов, в основном, сосредоточены на терминальных участках ДКП, где сосредоточен основной заряд (фиг. 1).
Надо отметить, что эти пептиды гораздо менее токсичны, чем линейные пептиды с тем же аминокислотным составом, хотя они также способны проникать через клеточные мембраны, могут взаимодействовать как с поверхностными, так и внутриклеточными клеточными мишенями [8, 9]. Самое главное, что они гораздо устойчивее к ферментативной биодеградации, что обусловлено их разветвленной структурой и присутствием неприродных амидных связей. Высокая растворимость КДП в водной среде, даже при наличии гидрофобных аминокислот имеет решающее значение при системном введении такого препарата [10]. Изменяя структуру терминальных групп, можно регулировать его свойства: заряд, гидрофобность, фармакокинетику. Введение в структуру ДКП цистеина позволяет селективно присоединять специфическую метку для исследования поведения такой молекулы в организме.
Эксперименты, проведенные нами ранее, показали, что аргинин-содержащие дендримерные пептиды, как, например, ЛТП (фиг. 1) с 8-ю концевыми остатками аргинина, обладают заметно большей трансфецирующей активностью по сравнению с линейными аналогами. Первоначально считалось, что импорт в клетку происходит путем прямого проникновения через плазматическую мембрану, однако затем было установлено, что главную роль в нем играет эндоцитоз, который включает макропиноцитоз, клатрин-опосредованный и кавеолин/рафт-опосредованный эндоцитоз. Но, вероятно, транспорт может идти параллельно по этим путям, и дальнейшее изучение показало, что импорт в клетку может идти и по эндоцитоз-независимому пути за счет взаимодействия с гепарин-сульфатом, который играет роль низко-аффинного рецептора для катионных пептидов. И чтобы обеспечивать кросс-сшивание молекул такого рецептора для интернализации пептида, очевидно, требуется некая определенная длина пептида. Экспериментально мы ранее установили, что 17-членный дендримерный пептид с концевыми аргининами (R), был намного активнее пептида, где концевой аргинин был заменен на лизин (К). Ключом к пониманию такой повышенной активности именно R-пептидов может служить особенность структуры гуанидиновой группы аргинина [17]. В водной среде гуандиновый катион, содержащий три заряженные аминогруппы, формирует планарную псевдоароматическую структуру с тремя сопряженными связями с очень высоким pK (13,6). Такой ион образует прочные водородные связи с молекулами воды, расположенными в той же плоскости, образуя, таким образом, амфифильный катион с гидрофобным анфасом. Гидрофобная плоскость/анфас катиона способна взаимодействовать с гидрофобными ароматическими аминокислотами, триптофаном, тирозином, и в тоже время такой катион также может активно связываться с анионными аминокислотами за счет координации атомов кислорода, и все это способствует дезорганизации/денатурации белковой структуры. Удивительно, что в отличие от К-пептидов, где катион имеет структуру аммония, молекулы R-пептидов в водной среде не проявляют, взаимного электростатического отталкивания. Более того, установлено, что в водном окружении наблюдается энергетически выгодное спаривание гуанидиновых групп в плоскостной ориентации (стэкинг-взаимодействие). Поэтому R-пептиды способны к образованию ассоциатов в воде, а К-пептиды, где доминируют силы отталкивания, не способны. Связывание R-пептида с мембраной на два порядка сильнее, чем для К-пептида. Так, разветвленные дендримерные конструкции, содержащие только два остатка аргинина способны связываться и проникать в клетки более эффективно, чем конструкции с двумя лизинами. Степень связывания и поглощения усиливается, если дополнительные остатки аргинина включаются в пептидную конструкцию.
Формирование комплекса между дендримером и НК предотвращает взаимодействие последних с сывороточным альбумином, которое, как известно, является основным препятствием для доставки свободных НК в ткани при генной терапии. Дендримеры могут также применяться для переноса генов с целью лечения злокачественных новообразований. Так, при генной терапии рака молочной железы дендримеры применяли для доставки канцеростатика ангиостатина и гена тканевого ингибитора металлопротеиназы (TIMP-2). В результате было показано значительное уменьшение пролиферации эндотелиальных раковых клеток. Эффективность доставки генетических конструкций в опухолевую ткань была проверена на мышах, у которых наблюдалось существенное снижение злокачественного роста. Показана эффективность дендримеров и для подавления экспрессии генов на принципе РНК-интерференции [11].
Синтез ДКП можно проводить обычными стандартными методами твердофазного синтеза. В практическом отношении такие пептиды синтезировать быстрее по сравнению с линейными, поскольку каждая стадия присоединения приводит к нарастанию числа аминокислот в геометрической прогрессии. Например, для 17-членного ДКП требуется лишь 6 стадий конденсации (фиг. 1). Единственная проблема заключается в том, что в твердофазном синтезе при высоких степенях ветвления (генерации), например, более 3-4 генераций, могут возникать стерические препятствия и замедление скорости реакций. Однако, эту проблему можно решить, используя смолы для твердофазного синтеза с низкой степенью сшивки и высокой способностью набухания, имеющие гелевую структуру (ПЭГ цепи), другой вариант - синтез целиком в растворе.
В настоящее время известны изобретения для внутриклеточной доставки НК для трансфекции, включая РНК-интерференцию, на основе дендримерных молекул. Следует отметить, что многие методы получения комплексов дендримеров с миРНК и плазмидами и протоколы их доставки в клетки опубликованы в общедоступной научной (медицинской и биохимической) литературе. Правда, большинство заявленных методов, ввиду легкости их синтеза, связано не с использованием ДКП, а дендримерных полимеров, синтетических полиамидоаминов (РАМАМ), с различными терминальными структурами. Например, в патенте PCT/GB 02/04706 от 24.04.2003 "Dendrimers for use in targeted delivery" описано использование катионных дендримеров для доставки НК, пептидов и фармацевтических агентов. В качестве дендримерной основы используются полимеры, производные диамино-этана/пропана/бутана, которые могут быть модифицированы гидрофобными, гидрофильными и амфифильными группами. Однако такие соединения имеют карбоцепную структуру без определенной четкой структуры и неспособны к биодеструкции.
Известно изобретение, описанное в патенте US 6376248 от 23.04.2002 "Peptide-enhanced transfections», в котором авторы предлагают многочисленные композиции для трансфекции клеток эукариот, включающие комплексы НК с пептидами, где нуклеиновая кислота связана ковалентно с линейным катионным пептидом с аминокислотной последовательностью ТАТ-пептида (из ВИЧ), дендримером или липопептидом. Указанные в документе дендримеры являются производными коммерческого продукта, дендримера РАМАМ (от компании Dendritech Inc.) с высокой молекулярной массой, к концевым группам которого, присоединены остатки лизина (LysDmer) или аргинина (ArgDmer). Однако известное решение обладает повышенной токсичностью, сложностью в получении, имеет ограниченный срок хранения.
Известна публикация [2], где описывается использование ДКП, вариант полилизинового дендримера, где в терминальные ветви дендримера вводят фенил-имидазольные остатки для получения амфипатической структуры. Наибольшая эффективность в плане защиты НК (на примере плазмидной ДНК) от деградации нуклеазами была получена с помощью полилизинового дендримера, имеющего степень ветвления 5 (генерации G5), этот же препарат, единственный, проявлял приемлемую трансфекционную эффективность. В другой работе этих же авторов [12] дендримеры с генерацией G5, с концевыми остатками аргинина, показывали высокую эффективность в отношении трансфекции клеток плазмидной ДНК и последующей экспрессии гена GFP (зеленый флуоресцирующий белок) или pGL3 (люциферазы). Дендример с генерацией G6 хотя и был также эффективен, но оказался намного более токсичен, вероятно из-за высокой плотности положительного заряда. Необходимо отметить, что выход при синтезе таких дендримеров довольно низкий и препараты не были использованы для подавления экспрессии генов (миРНК-интерференция).
В документе CN 102911252 В от 06.02.2013 "Cationic lipid containing peptide dendrimer, transgenic carrier and preparation method and application of transgenic carrier" описывает соединения, как носители для внутриклеточной доставки плазмид, содержащих гены GFP или pGL3. Структура описываемых носителей сдержит короткий аргинин-содержащий дендример с липидным фрагментом, содержащим холестерин. Однако, трансфекция в формате РНК-интерференции (миРНК) в патенте не приводится.
Патент RU 2127125 от 10.03.1999 («Биологически активные и/или целевые дендримерные конъюгаты») описывает плотные звездчатые полимерные конъюгаты, ассоциированные с биологическим модулятором, причем в одном из воплощений плотным звездчатым полимером является дендример, но полученный не на основе разветвленного пептида, а на основе полимеров РАМАМ, полиэтиленимина и других.
В патенте RU 2575603 от 20.02.2016 «Получение комплексов нуклеиновых кислот и поперечно-сшитых дисульфидными связями катионных компонентов, предназначенных для трансфекции и стимуляции» авторы предлагают применять композицию, содержащую полимерный носитель и молекулу-переносчик, в качестве иммуностимулирующего агента или адъюванта. В том числе заявлены олигопептиды и белки с общей формулой (Arg)1;(Lys)m;(His)n;(Orn)o;(Xaa)x, однако авторы не указывают, линейные ли это пептиды или дендримеры. В качестве НК используют одноцепочечную РНК с целью индукции врожденного или адаптивного иммунитета.
В статье [13] описываются ДКП с генерацией G3 для трансфекции клеток молекулами миРНК. Дендримерные пептиды на основе лизина содержат гидрофобный участок, представленный либо гидрофобными аминокислотами, либо жирными кислотами для того, чтобы более эффективно выходить из эндосомы в цитоплазму после интернализации комплекса НК/пептид клеткой. Среди них наиболее активные в ми-РНК-трансфекции ДКП были: (KL)8(KKL)4(KLL)2KK(C16)K(C16) и (KL)8(KKL)4(KLL)2KLLLL), Однако, нужно подчеркнуть, что синтез и очистка таких пептидов (длина 37 и 39 аминокислот + жирные кислоты) довольно сложная и крайне дорогостоящая процедура (сделанная вручную), в результате, выходы были очень низкие (менее 10 процентов).
В статье [14] описано применение полилизиновых дендримеров содержащих более 40 остатков лизина для улучшенной доставки ДНК (плазмиды) в клетки в качестве средств для противоопухолевой терапии, указывается высокая скорость и эффективность трансфекции, а также относительно малая токсичность для здоровой ткани. Их применение для РНК-интерференции не описано. Очевидно, ввиду высокой молекулярной массы выходы пептидов при твердофазном синтезе были очень низкие.
Ближайшим аналогом заявляемого изобретения является дендримерный пептид ЛТП, описанный в патенте RU 2572575 С1 от 20.01.2016, обладающий трансфекционной активностью при комплексовании с НК, низкой токсичностью в отношении клеток млекопитающих. Однако по сравнению с липосомальным средством доставки Lipofectamine®2000, используемым как золотой стандарт, ЛТП имеет трансфекционную активность на 2-3 порядка ниже.
Задачей данного изобретения является создание малотоксичных дендримерных пептидов способных обеспечивать эффективный транспорт олигонуклеиновых кислот в клетки млекопитающих и стимулировать ДНК и РНК трансфекцию с целью как селективного подавления экспрессии гена-мишени путем РНК-интерференции, так и стимулирования экспрессии целевого гена.
Поставленная задача решается за счет того, что указанное средство представляет собой средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих, содержащее эффективное количество дендримерных катионных пептидов, представляющих собой синтетические разветвленные структуры с лизинами в местах ветвлений, характеризующиеся структурной формулой:
R4nK2nX2nKnXnK(ZZ)C-Lipm, где
n=1 или 2;
R - аргинин;
K - лизин;
Х=0-5 аминокислот в любом порядке, где аминокислоты представляют собой: K (лизин), глицин (G), триптофан (W), лейцин (L), изолейцин (I), треонин (Т), пролин (Р), глутаминовую кислоту (Е), валин (V), аланин (А) или гистидин (Н);
(ZZ)C - С-концевая линейная аминокислотная последовательность, любые 1-5 аминокислот с цистеином (С) на конце;
Lipm - липофильная углеводородная цепь, где длина углеводородной цепи составляет от С6 до С18 и связана через малеимидную группу с остатком цистеина (С), где m=1 или 0 - отсутствие Lip.
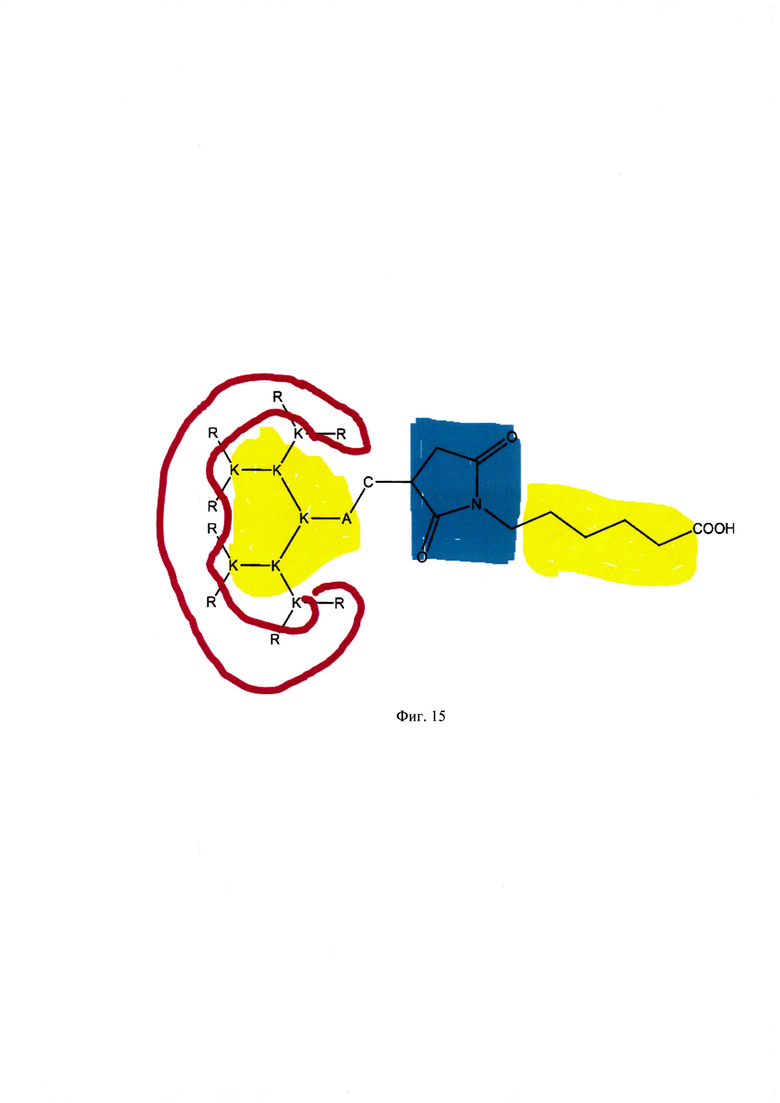
Технический результат настоящего изобретения заключается в создании модульной конструкции дендримерных пептидов, состоящей из N-концевого катионного модуля, центральной части и реакционноспособного модуля на С-конце. Катионный модуль представляет собой заряженный кластер из N-концевых аргининов и обеспечивает взаимодействие с нуклеиновыми кислотами и поверхностью клетки, заряженной отрицательно, и транслоцирование комплекса пептида с нуклеиновой кислотой в клетку. Центральная часть представляет собой гидрофобное ядро из остатков лизина, обеспечивающее дополнительное сродство с мембраной клетки и необходимую трехмерную структуру. К нему примыкает короткий гидрофильный участок, содержащий остаток цистеина, который за счет свободной тиольной группы выступает как реакционноспособный модуль, обеспечивая возможность дальнейшей модификации пептида различными молекулами. В частности, в настоящем изобретении была проведена модификация гидрофобными молекулами с длиной углеводородной цепи от С6 до С18 по тиол-малеимидной клик-реакции, что способствует повышению трансфекционной/транспортной активности дендримера. Массовое соотношение положительно-заряженного пептида к отрицательно-заряженной НК в интервале от 10 до 50 обеспечивает образование комплексов заявленных ДКП с НК, способных эффективно проникать в различные типы клеток. Таким образом, полученное средство обеспечивают доставку как плазмидной ДНК, так и малых интерферирующих РНК (миРНК), что позволяет использовать их как перспективный агент для генной терапии, включая терапию на основе РНК-интерференции.
Краткое описание графических материалов, фигур:
Фигура 1. Схема структуры дендримерного пептида ЛТП (терминальные аминокислоты представлены аргинином (R), несущим положительный заряд).
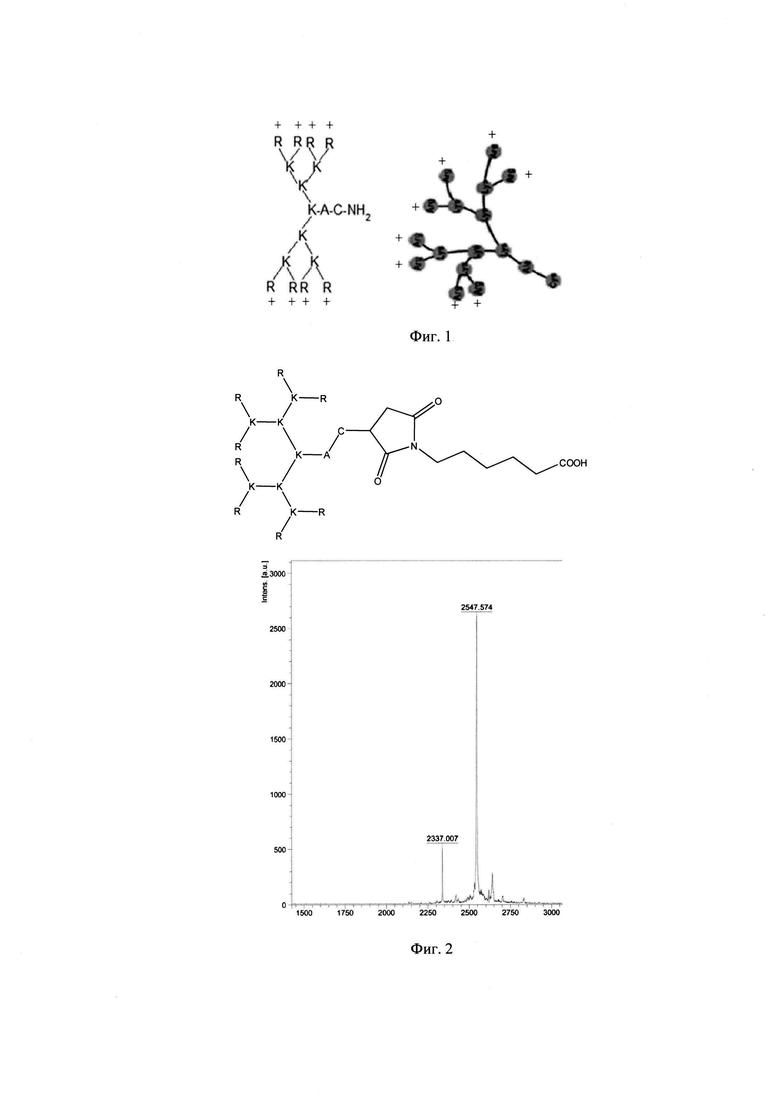
Фигура 2. Строение и масс-спектр синтезированного пептида ТА-22 (R8K4K2KAC-mal-N-5-карбоксипентил)* xTFA (трифторацетат), М=2547 г/моль, Заряд +15, Катионный амфифильный пептидный дендример (солевая форма, высокая плотность заряда).
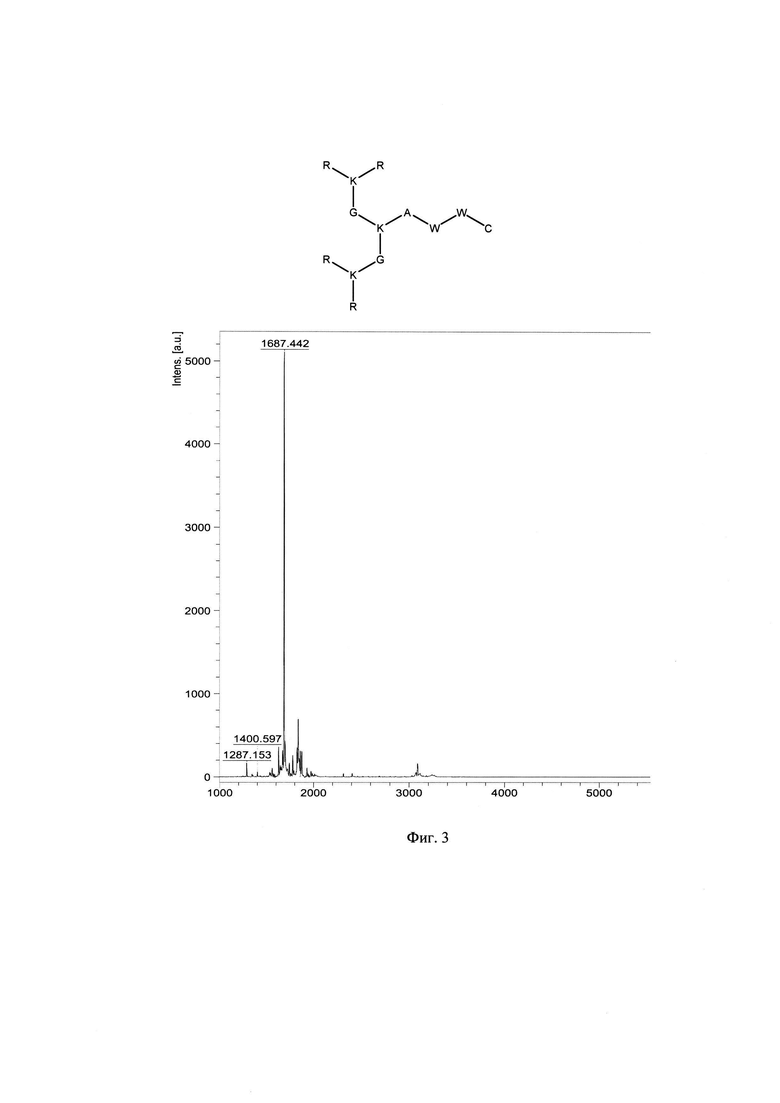
Фигура 3. Строение и масс-спектр синтезированного пептида KK-50 (R4K2G2KAWWC) xTFA, М=1687 г/моль, Заряд +8, Катионный амфифильный пептидный дендример (солевая форма, средняя плотность заряда).
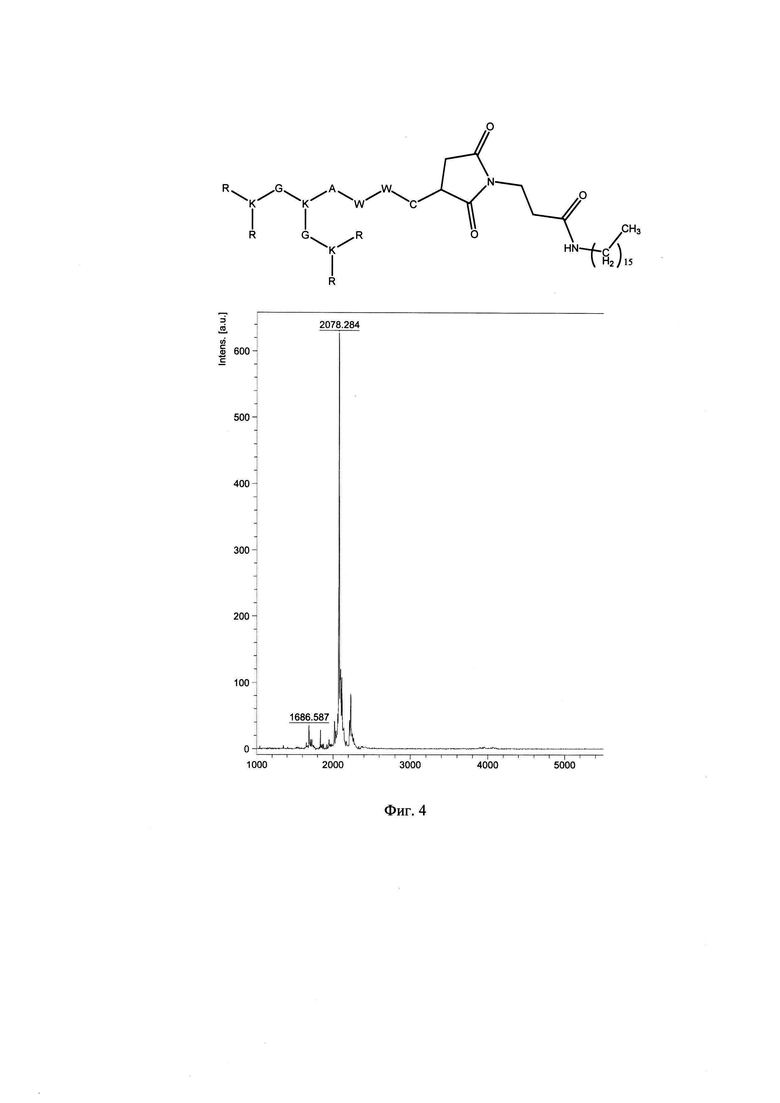
Фигура 4. Строение и масс-спектр синтезированного пептида KK-61 (R4K2G2KAWWC-mal-С2Н4-СО-NH-С16Н33)* xTFA, М=2078 г/моль, Заряд +8, Катионный амфифильный липо-пептидный дендример (солевая форма, средняя плотность заряда).
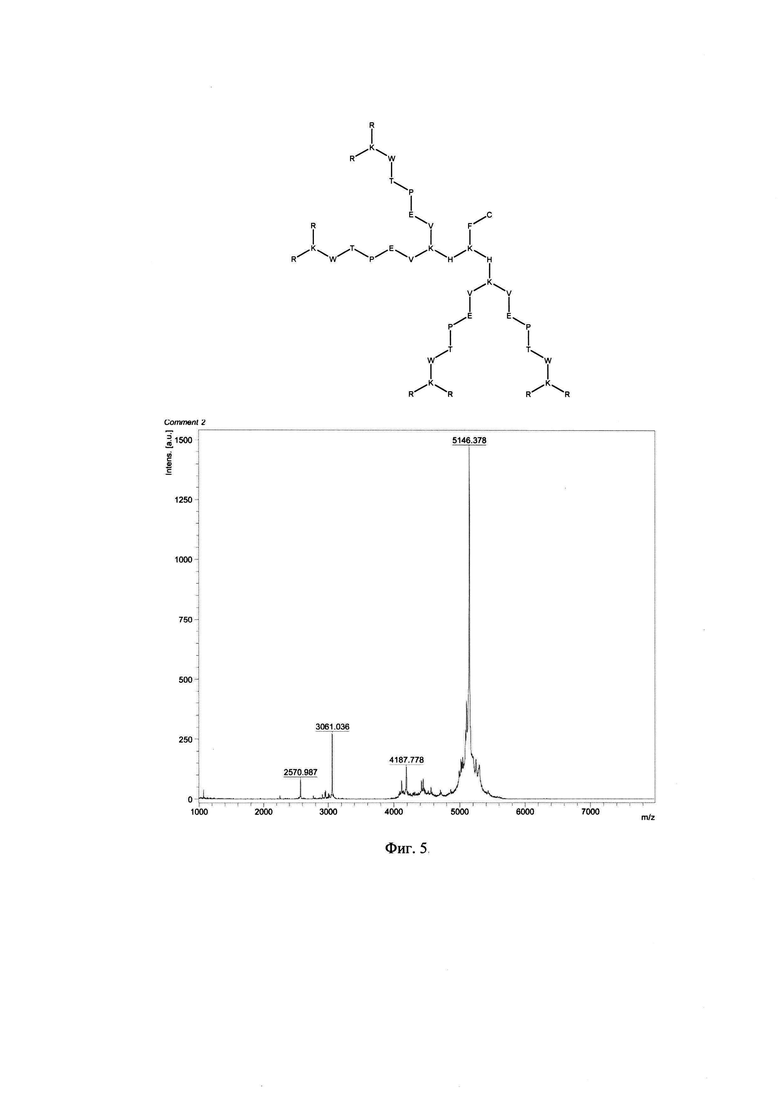
Фигура 5. Строение и масс-спектр синтезированного пептида KK-46 (R8K4W4T4P4E4V4K2H2KFC)* xTFA, М=5140 (+6Н) г/моль, Заряд +16, Катионный амфифильный пептидный дендример (солевая форма, высокая плотность заряда).
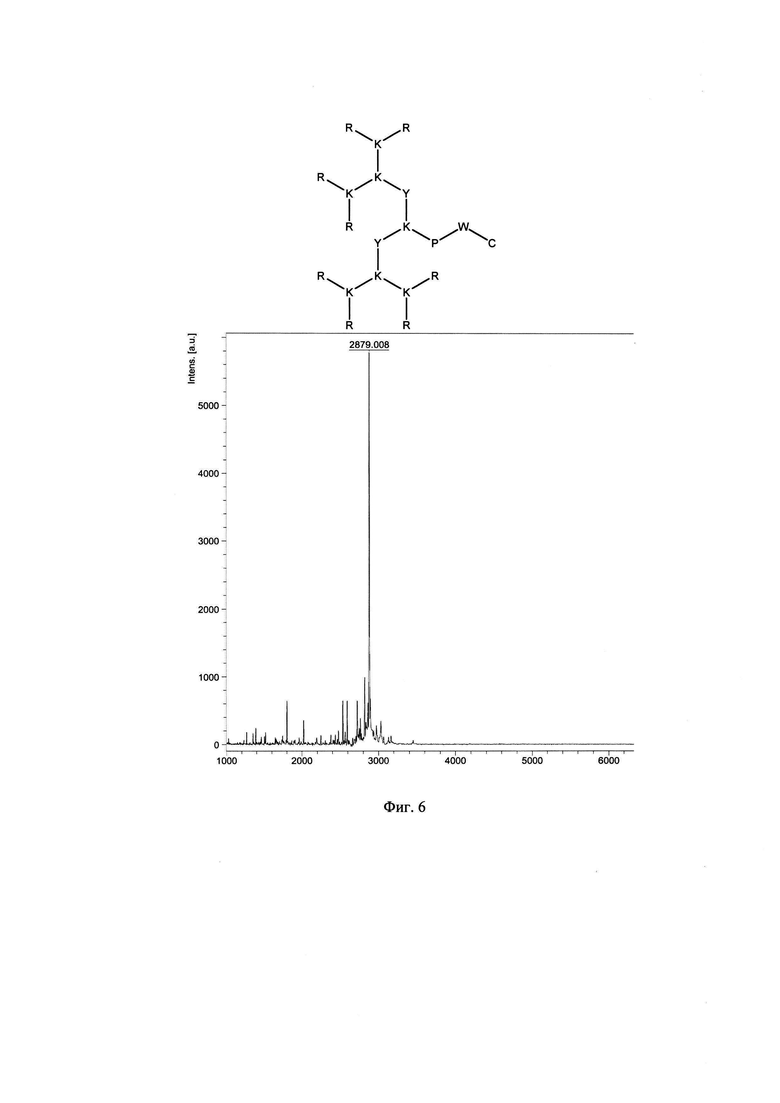
Фигура 6. Строение и масс-спектр синтезированного пептида SA-32 (R8K4K2Y2KPWC)* xTFA, М=2875 г/моль, Заряд +16, Катионный амфифильный пептидный дендример (солевая форма, высокая плотность заряда).
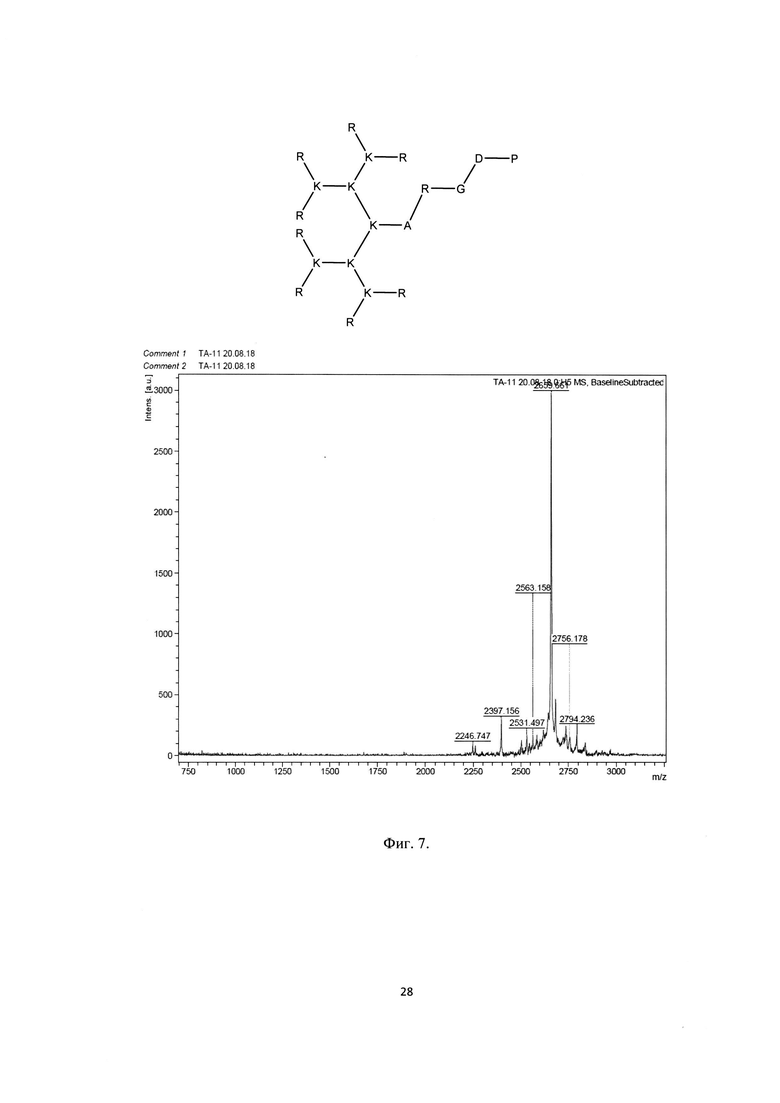
Фигура 7. Строение и масс-спектр синтезированного пептида ТА-11 (R8K4K2KARGDP)* xTFA, М=2660 г/моль, Заряд +16, Катионный амфифильный пептидный дендример (солевая форма, высокая плотность заряда).
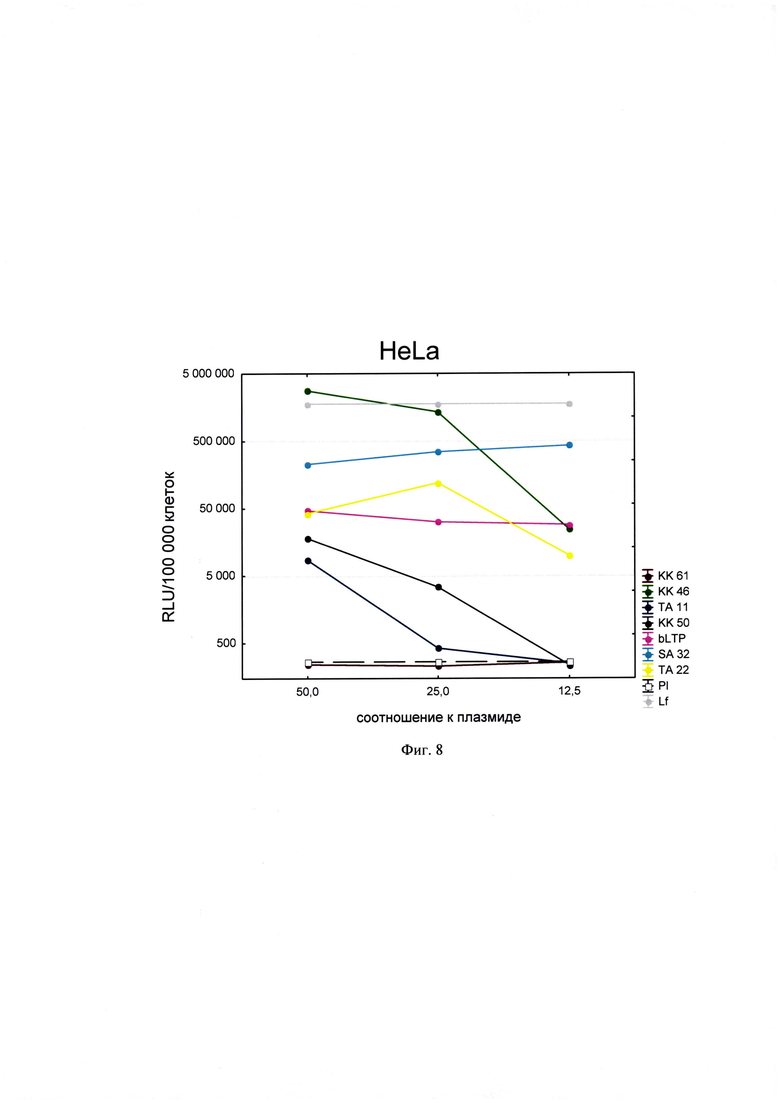
Фигура 8. Трансфекционная активность ДКП на линии клеток HeLa (карцинома шейки матки человека), где по оси (у) представлены относительные световые единицы на 100 тыс. клеток, по оси (х) - массовые соотношения пептида к НК; КК61, КК46, ТА11, KK50, SA32, ТА22 - синтезированные ДКП; bLTP - контрольный пептид, Р1 - отрицательный контроль, плазмида pGL3 без трансфекционного агента, Lf - положительный контроль, Lipofectamine®2000 в рекомендуемом производителем соотношении к плазмиде.
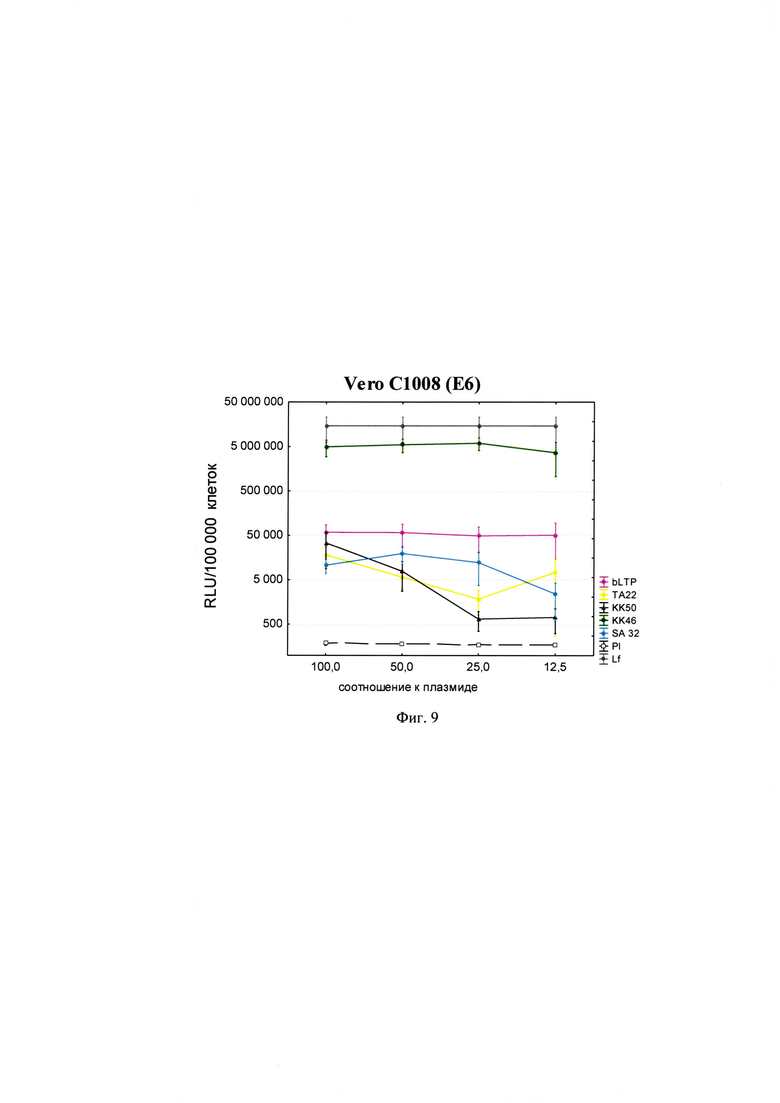
Фигура 9. Трансфекционная активность ДКП на линии клеток Vero С1008 (Е6) (почка африканской зеленой мартышки), где по оси (у) представлены относительные световые единицы на 100 тыс. клеток, по оси (х) - массовые соотношения пептида к НК; КК61, КК46, ТА11, KK50, SA32, ТА22 - синтезированные ДКП; bLTP - контрольный пептид, Р1 - отрицательный контроль, плазмида pGL3 без трансфекционного агента, Lf - положительный контроль, Lipofectamine®2000 в рекомендуемом производителем соотношении к плазмиде.
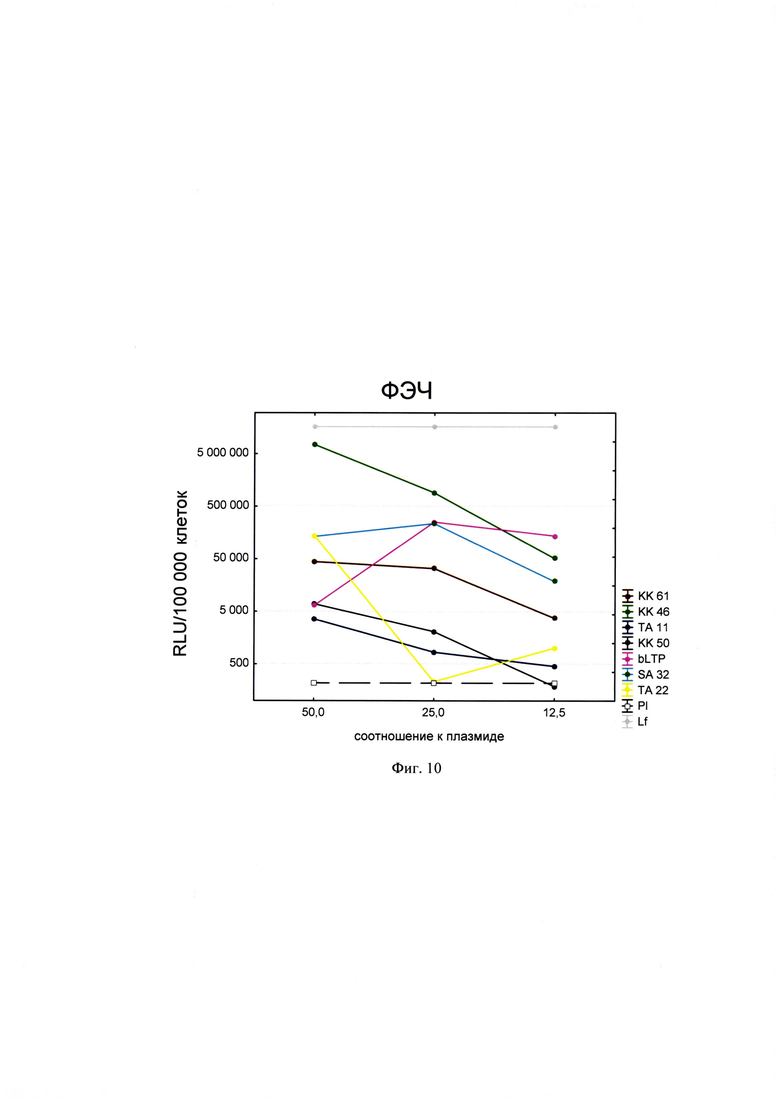
Фигура 10. Трансфекционная активность ДКП на линии клеток ФЭЧ (фибробласты эмбриона человека), где по оси (у) представлены относительные световые единицы на 100 тыс. клеток, по оси (х) - массовые соотношения пептида к НК; КК61, КК46, ТА11, KK50, SA32, ТА22 - синтезированные ДКП; bLTP - контрольный пептид, Р1 - отрицательный контроль, плазмида pGL3 без трансфекционного агента, Lf - положительный контроль, Lipofectamine®2000 в рекомендуемом производителем соотношении к плазмиде.
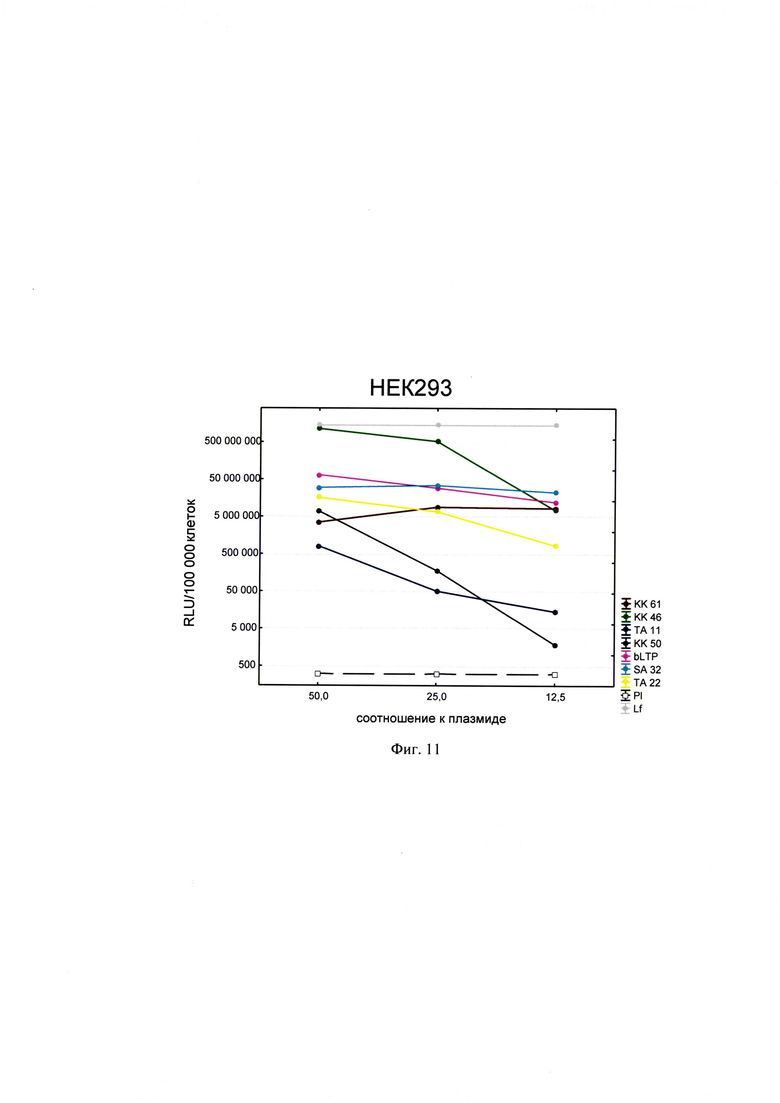
Фигура 11. Трансфекционная активность ДКП на линии клеток НЕК293 (почка эмбриона человека), где по оси (у) представлены относительные световые единицы на 100 тыс. клеток, по оси (х) - массовые соотношения пептида к НК; КК61, КК46, ТА11, KK50, SA32, ТА22 - синтезированные ДКП; bLTP - контрольный пептид, Р1 - отрицательный контроль, плазмида pGL3 без трансфекционного агента, Lf - положительный контроль, Lipofectamine®2000 в рекомендуемом производителем соотношении к плазмиде.
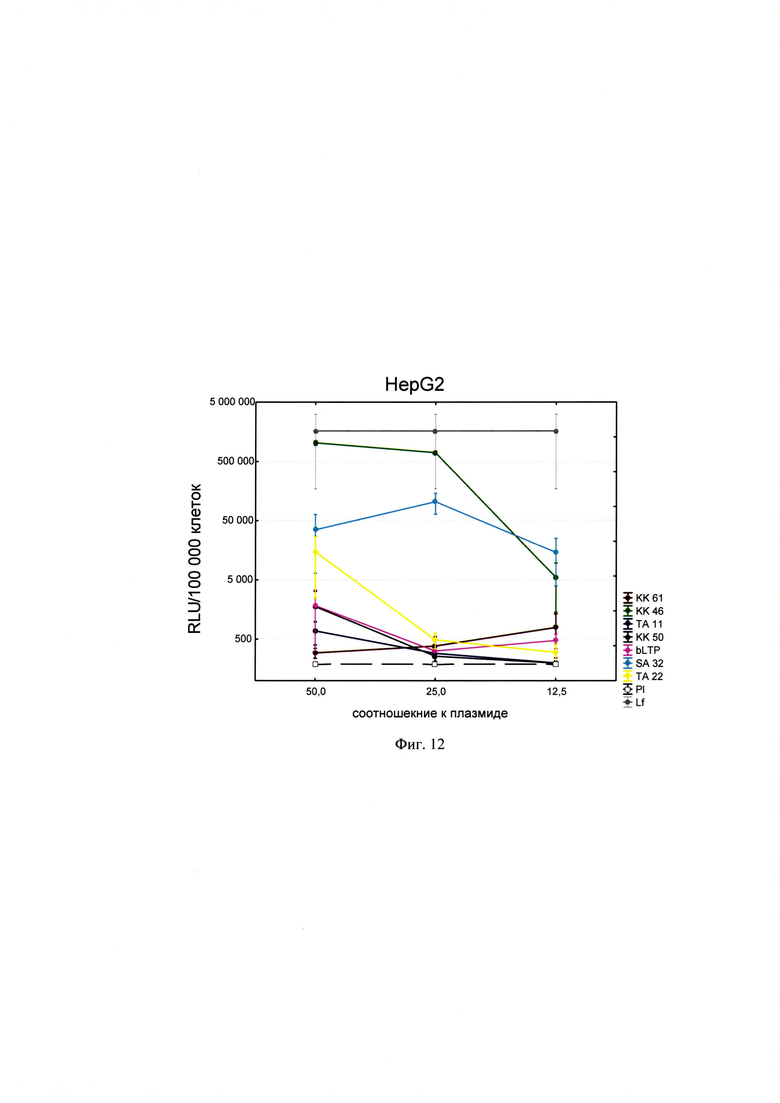
Фигура 12. Трансфекционная активность ДКП на линии клеток HepG2 (гепатоцеллюлярная карцинома человека), где по оси (у) представлены относительные световые единицы на 100 тыс. клеток, по оси (х) - массовые соотношения пептида к НК; КК61, КК46, ТА11, KK50, SA32, ТА22 - синтезированные ДКП; bLTP - контрольный пептид, Р1 - отрицательный контроль, плазмида pGL3 без трансфекционного агента, Lf - положительный контроль, Lipofectamine®2000 в рекомендуемом производителем соотношении к плазмиде.
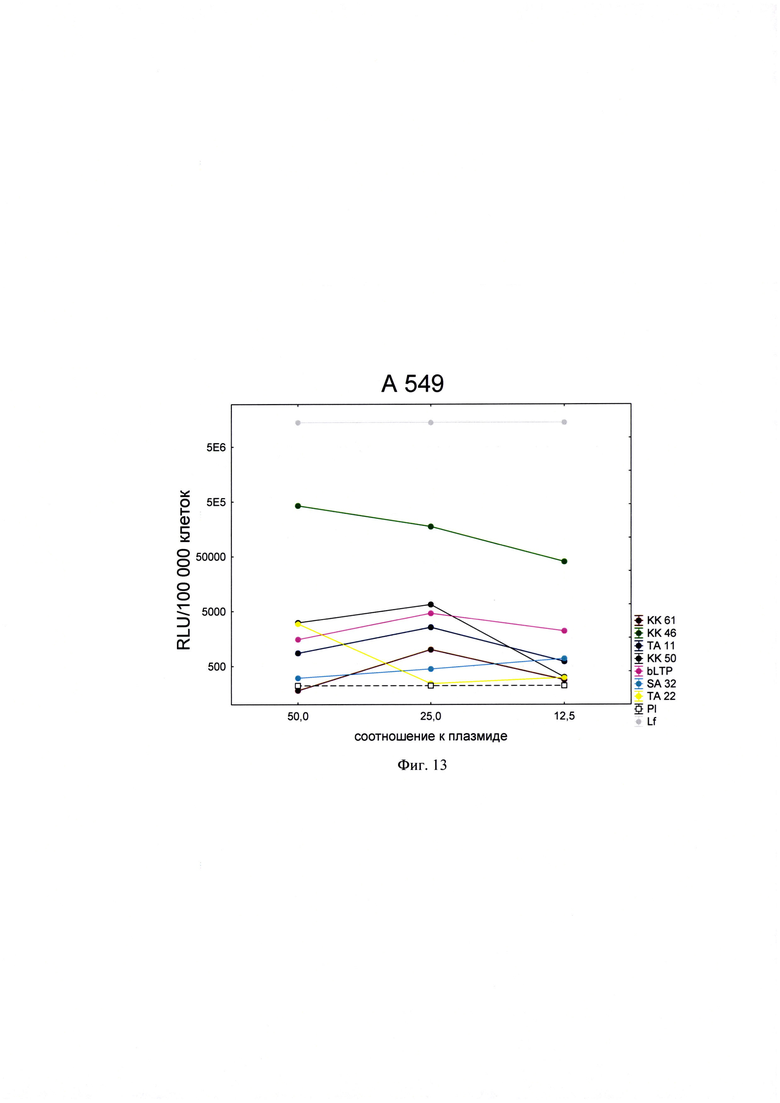
Фигура 13. Трансфекционная активность ДКП на линии клеток А549 (карцинома легкого человека), где по оси (у) представлены относительные световые единицы на 100 тыс. клеток, по оси (х) - массовые соотношения пептида к НК; КК61, КК46, ТА11, KK50, SA32, ТА22 - синтезированные ДКП; bLTP - контрольный пептид, Р1 - отрицательный контроль, плазмида pGL3 без трансфекционного агента, Lf - положительный контроль, Lipofectamine®2000 в рекомендуемом производителем соотношении к плазмиде.
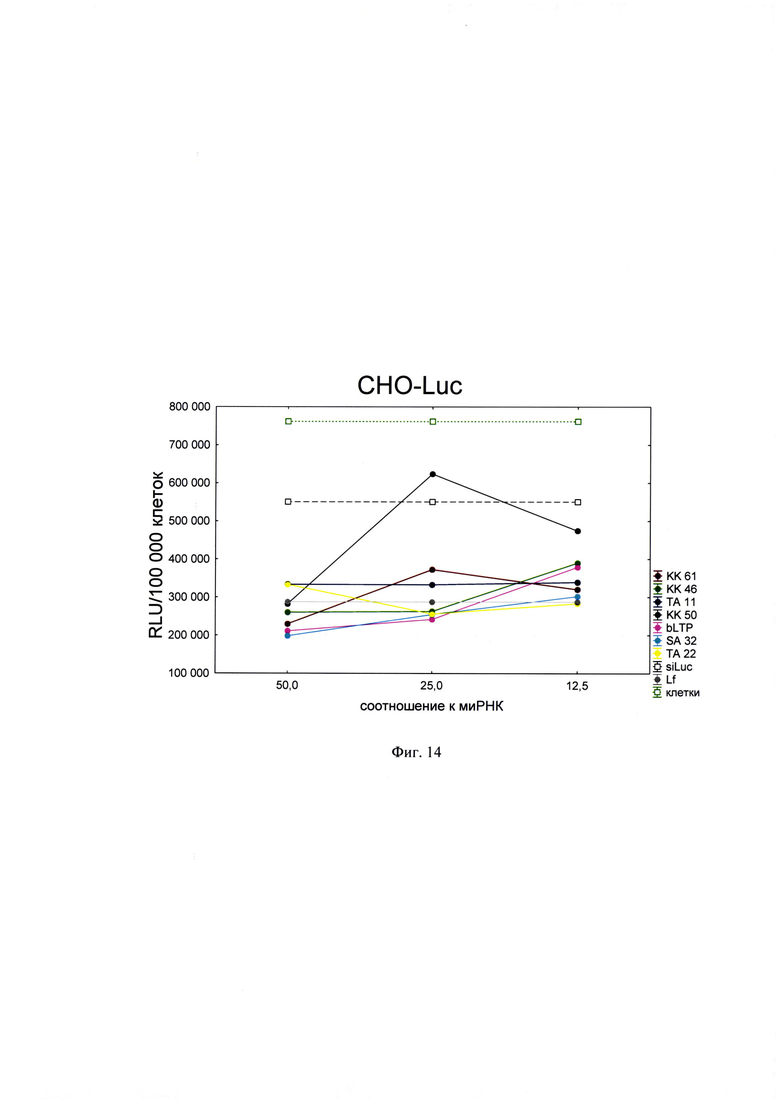
Фигура 14. Трансфекционная активность ДКП на линии клеток CHO-Luc (клеточная линия яичников китайского хомячка, трансфецированная плазмидой, кодирующей ген люциферазы), где по оси (у) представлены относительные световые единицы на 100 тыс. клеток, по оси (х) - массовые соотношения пептида к НК; КК61, КК46, ТА11, KK50, SA32, ТА22 - синтезированные ДКП; bLTP - контрольный пептид, siLuc - отрицательный контроль, миРНК siLuc без трансфекционного агента, Lf - положительный контроль, Lipofectamine®2000 в рекомендуемом производителем соотношении к миРНК.
Фигура 15. Модульная конструкция дендримерных пептидов (пример пептида ТА-22). Красным цветом выделен катионный кластер, представленный остатками L-аргинина, желтым цветом отмечены гидрофобные участки, голубым - гидрофильный фрагмент.
Изобретение иллюстрируется примерами, приведенными ниже.
Пример 1. Твердофазный синтез пептидов R4nK2nX2nKnXnK(ZZ)C-Lipm
Используемые реагенты и растворители были получены от компаний Sigma-Aldrich/Merck, Anaspec, ApexBio, Gyros Proteins, Alfa Aesar, Merck, Fluka AG, Reachem, Novabiotech, Protein Technologies Inc, Carl Roth GmbH, Scharlau, Fisher Chemical с качеством либо для пептидного синтеза, либо ВЭЖХ, либо ГЖХ/МС, либо для анализа. Для синтеза пептидов использовали автоматический синтезатор пептидов PS3 (Protein Technologies Inc., США).
Для очистки пептидов использовалась высокоэффективной жидкостная хроматография (ВЭЖХ). Одна из используемых систем состояла из бинарного насоса 321-Н (Gilson), колонки Vydac 218ТР1022 (Grace), фотометрического детектора Spectroflow 757 (Kratos Analytical), регистратора 2210 (LKB) и коллектора фракций Multirac 2111 (LKB), другая система - Серии LC-20 Prominence (Shimadzu, Япония). При элюировании использовали линейный градиент 5-70% элюента В (25 минут) при 10 мл/мин с детектированием при 226 нм. Элюент А содержал 0,1% TFA/Н2О; элюент В содержал чистый ацетонитрил (Sigma-Aldrich). Для препаративной ВЭЖХ использовали систему LC-20 Prominence Shimadzu (Япония).
Все пептиды были синтезированы с использованием методов твердофазного синтеза с Fmoc-защитой α-аминогрупп на смоле Rink Amide МВНА с помощью автоматического синтезатора или в ручном режиме, используя протоколы Fmoc-химии и N-гидроксибензотриазол/диизопропилкарбодиимидный (ГБТ/ДИК) или гексафторфосфат бензотриазол тетраметилурониевый (HBTU) методы активации аминокислот. Функциональные группы боковых цепей Fmoc-аминокислот (AnaSpec) были защищены окситрет-бутилом (Ot-Bu) для карбоксильных групп, трет-бутилом (t-Bu) для гидроксильных групп, трет-бутилоксикарбонилом (Boc) или 9-Флуоренилметоксикарбонил (Fmoc) для ε-аминогруппы лизина, тритильной группой (Trt) для SH-группы цистеина и имидазольной группы гистидина, и 2,2,4,6,7-пентаметил-дигидробензофуран-5-сульфонил (Pbf) для гуанидиновой группы аргинина. Синтез осуществляли либо вручную, либо автоматически с использованием смолы Rink-amide-МВНА (NH2-P) (NovaBiochem) и двукратных избытков активированных Fmoc-аминокислот и соответствующих реагентов.
Схема твердофазного синтеза основана на повторяющемся цикле стандартных операций (RU 2572575 С1 от 20.01.2016), который включает:
1) На старте навеску исходной смолы Rink Amide вносят в реактор, добавляют диметилформамид (ДМФА), энергично перемешивают суспензию до полного набухания смолы, операцию повторяют 3 раза по 3 мин, удаляя растворитель фильтрованием под давлением.
2) Удаление Fmoc-защиты 20% раствором 4-метилпиперидина (МПП) в ДМФА в течение 10 мин.
3) Промывку смолы ДМФА.
4) Предварительная активация 2-х кратного мольного избытка Fmoc-аминокислоты с помощью ДИК/ГБТ или HBTU в среде ДМФА или N-метилпирролидона (МП).
5) Конденсация аминокислоты путем добавления в реактор активированной Fmoc-аминокислоты в среде ДМФА или N-метилпирролидона (МП) и инкубацией в течение 1 часа.
6) Промывку смолы с пептидом ДМФА.
7) Операции по пунктам 2-6 повторяются.
Отщепление пептидов от смолы проводили в 95%-ном растворе TFA с добавлением тиоанизола, 1,2-этандитиола и деионизированной воды, в результате которого происходит полное снятие остальных защитных групп с боковых цепей (все защитные группы боковых цепей, кроме Fmoc, были лабильными для TFA). Сырые пептидные продукты осаждали диэтиловым эфиром, экстрагировали разбавленной уксусной кислотой и затем высушивали лиофильно перед очисткой. Сырой пептид очищали с помощью градиентной ВЭЖХ, используя в качестве подвижной фазы 0,1% водный раствор трифторуксусной кислоты и ацетонитрил. Заявляемые ДКП - стабильные вещества при хранении в сухом виде. Их синтез можно осуществлять, используя автоматический пептидный синтезатор или в ручном режиме.
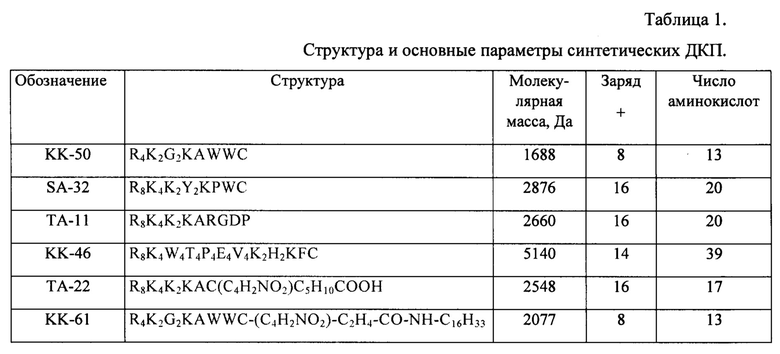
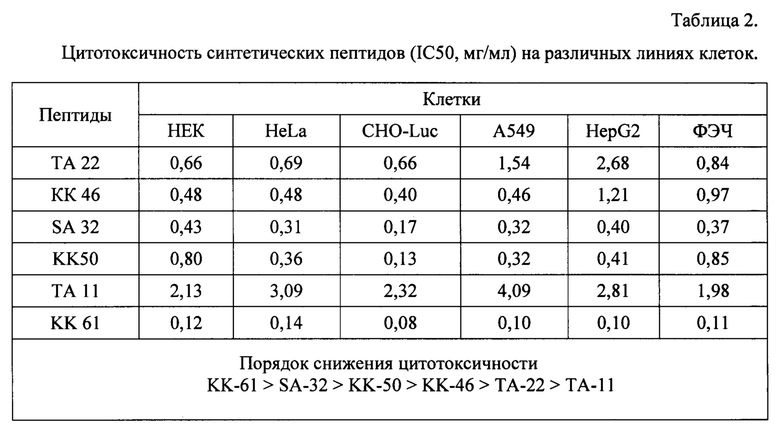
В результате были синтезированы и очищены 6 дендримерных катионных пептидов: KK-50 (R4K2G2KAWWC), SA-32 (R8K4K2Y2KPWC), ТА-11 (R8K4K2KARGDP), KK-46 (R8K4W4T4P4E4V4K2H2KFC), ТА-22 (R8K4K2KAC(C4H2NO2)C5H10COOH), KK-61 (R4K2G2KAWWC-(C4H2NO2)-C2H4-CO-NH-C16H33) (табл. 1, фиг. 2-7).
где, аргинин (R); лизин (K); глицин (G), триптофан (W), треонин (Т), пролин (Р), глутаминовую кислоту (Е), валин (V), аланин (А) или гистидин (Н);
В частности, последовательность операций при синтезе пептида ТА22 выглядит следующим образом:
Fmoc-C(Trt)-OH+NH2-P-(Rink-Amid) → Fmoc-C(Trt)-P → H-Cys(Trt)-P → Fmoc-AC(Trt)-P → H-AC(Trt)-P → Fmoc-K(Fmoc)-AC(Trt)-P → H-KAC(Trt)-P → (Fmoc-K(Fmoc))2-KAC(Trt)-P → (H-K)2KAC(Trt)-P → (Fmoc-K(Fmoc))4K2KAC(Trt)-P → K4K2KAC(Trt)-P → (FmocR(Pbf))8K4-K2KAC(Trt)-P → R8K4K2KAC-NH2 + N-малеимидгексановая кислота → R8K4K2KAC-(C4H2NO2)-C5H10COOH.
использованные аминокислоты: С - Цистеин, А - Алании, К - Лизин, R - Аргинин.
Таким образом, была создана модульная конструкция дендримерных пептидов, состоящая из N-концевого катионного модуля, центральной части и реакционноспособного модуля на С-конце. Катионный модуль представляет собой заряженный кластер из N-концевых аргининов и обеспечивает взаимодействие с нуклеиновыми кислотами и поверхностью клетки, заряженной отрицательно, и транслоцирование комплекса пептида с нуклеиновой кислотой в клетку. Центральная часть представляет собой гидрофобное ядро из остатков лизина, обеспечивающее дополнительное сродство с мембраной клетки и необходимую трехмерную структуру. К нему примыкает короткий гидрофильный участок, содержащий остаток цистеина, который за счет свободной тиольной группы выступает как реакционноспособный модуль, обеспечивая возможность дальнейшей модификации пептида различными молекулами. В частности, в настоящем изобретении была проведена модификация гидрофобными молекулами с длиной углеводородной цепи от С6 до С18 по тиол-малеимидной клик-реакции, что способствует повышению трансфекционной/транспортной активности дендримера.
Пример 2. Анализ трансфекционной активности пептида при доставке в клетки плазмидной ДНК или миРНК.
Для проведения анализа клетки Vero С1008 (Е6), HEK293, А549, HepG2, ФЭЧ, HeLa, CHO-Luc высеивали в 48-луночный планшет в количестве 4⋅104 клеток на лунку в 300 мкл соответствующей полной питательной среды (DMEM, или F12, или Игла-Мем, или RPMI, содержащей 10% фетальной бычьей сыворотки, 2% буферного раствора HEPES, 0,1% антибиотика гентамицина, 0,6% L-глутамина) накануне трансфекции и инкубировали сутки при 37°C в CO2 инкубаторе до достижения 75% конфлюентности. Дисперсию трансфекционного агента в 3 разведениях (с массовыми соотношениями пептида к НК - 50:1, 25:1 и 12,5:1) общим объемом 80 мкл, состоящего из 0,3 мкг плазмиды pGL3 или миРНК siLuc (дуплекс GGAAAGACGAUGACGGAAAtt/UUUCCGUCAUCGUCUUUCCtt) и раствора пептида в соответствующей концентрации согласно массовым соотношениям к НК, готовили в бессывороточной среде OPTIMEM. В качестве положительного контроля использовали коммерческий трансфекционный агент Lipofectamine2000 (Lf). В качестве отрицательного контроля использовали плазмиду pGL3 без носителя (Р1). В качестве агента сравнения использовали ближайший синтетический аналог - пептид ЛТП (bLTP - контрольный пептид) (RU 2572575 С1). Приготовленные смеси выдерживали 15 мин при комнатной температуре и вносили в лунки с монослоем клеток. Перед внесением трансфекционной смеси в лунках с клетками заменяли полную питательную среду на бессыворточную. Клетки инкубировали при 37°C в CO2 инкубаторе в течение 4 часов. Затем к клеткам добавляли 30 мкл 10% фетальной бычьей сыворотки. Клетки инкубировали при 37°C в CO2 инкубаторе в течение 24 часов и затем определяли люциферазную активность методом люциферазного теста. Тест проводился с использованием коммерческого набора "Luciferase Assay System" (Promega) согласно рекомендациям производителя. Для этого удаляли ростовую среду из лунок, затем добавляли 100 мкл лизирующего буфера (Glo lysis buffer, 1x, USA). Клетки выдерживали 15 минут при 37°C в СО2 инкубаторе для достижения полного лизиса. Затем соскабливали их со дна ячеек и переносили клеточную суспензию в пробирки типа eppendorf на 1,5 мл. Для осаждения клеточного дебриса полученный лизат центрифугировали 2 минуты на 10000 оборотов. Отбирали по 50 мкл супернатанта в отдельные пробирки типа eppendorf на 1,5 мл и добавляли к ним люциферазный субстрат в соотношении 1:1. Эффективность трансфекции оценивали по уровню люминисценции на Glomax 20/20 luminometr. Для всех количественных данных вычисляли среднее арифметическое (М) и стандартную ошибку среднего (m). Определяли межгрупповые различия и оценивали достоверности различий между группами с помощью критерия Стьюдента с использованием программы Statistica 8.0. Трансфекционная активность ДКП на различных линиях клеток продемонстрирована на фиг. 8-14).
Из представленных данных видно, что массовое соотношение положительно-заряженного пептида к отрицательно-заряженной НК в интервале от 10 до 50 обеспечивает образование комплексов заявленных ДКП с НК, способных эффективно проникать в различные типы клеток. Таким образом, полученное средство обеспечивают доставку как плазмидной ДНК, так и малых интерферирующих РНК (миРНК), что позволяет использовать их как перспективный агент для генной терапии, включая терапию на основе РНК-интерференции.
Пример 3. Анализ цитотоксичности пептидов.
Клетки Vero С1008 (Е6), HEK293, А549, HepG2, ФЭЧ, HeLa, CHO-Luc высеивали в 96-луночный планшет в количестве 3⋅104 клеток на лунку в 100 мкл соответствующей полной питательной среды (DMEM, или F12, или Игла-Мем, или RPMI, содержащей 10% фетальной бычьей сыворотки, 2% буферного раствора HEPES, 0,1% антибиотика гентамицина, 0,6% L-глутамина) и инкубировали сутки при 37°C в СО2 инкубаторе до 75% конфлюентности. Концентрации пептидов в лунках варьировали (5500, 2750, 1375, 688, 344, 172, 86, 43, 21, 11, 5 мкг/мл) методом последовательного разведения. Затем планшет инкубировали при 37°C в CO2-инкубаторе. Через сутки в каждую лунку добавляли по 25 мкл тетразолиевого красителя МТТ в фосфатно-солевом буфере с концентрацией 4 мг/мл и планшет помещали в CO2-инкубатор. Через 4 часа в лунки добавляли 50 мкл додецилсульфата натрия (20% ДДС в воде с 0,02N H2SO4) и инкубировали ночь. Затем определяли разницу оптической плотности раствора при 570 и 650 нм на спектрофотометре (Thermo Scientific™ Multiskan™ GO Microplate Spectrophotometer). Количество живых клеток рассчитывали как отношение оптической плотности (А) в лунке с пептидом к оптической плотности в положительном контроле (100% живых клеток) по формуле, где оба значения нормировали на отрицательный контроль (100% погибших клеток):
количество живых клеток, % = (Апептида-А-контроль)/(А+контроль-А-контроль)⋅100%
Каждое значение получали усреднением нескольких параллельных измерений в дублях. Строили график полученных значений от концентрации, определяли IC50 в мкг/мл значения представлены в таблице 2.
Анализ цитотоксичности заявляемых ДКП (ЛД50>0,2 мг/мл) указывает на их низкую токсичность.
СПИСОК ЛИТЕРАТУРЫ
1. Milletti F. Cell-penetrating peptides: classes, origin, and current landscape. Drug Discovery Today, 2012, 17:850-860.
2. Luo K., Li C, Wang G., Nie Y., He В., Wu Y., Gu Z. Peptide dendrimers as efficient and biocompatible gene delivery vectors: Synthesis and in vitro characterization. J. Control Release, 2011, 55(1):77-87. doi: 10.1016/j.jconrel.2010.10.006.
3. Takechi-Haraya Y., Saito H. Current understanding of physicochemical mechanisms for cell membrane penetration of arginine-rich cell penetrating peptides: Role of glycosaminoglycan Interactions. Current Protein and Peptide Science, 2018, 19(6), 623-630. doi: 10.2174/1389203719666180112100747.
4. Kumar V., Agrawal P., Kumar R., Bhalla S., Usmani S.S., Varshney G.C., Raghava G.P. Prediction of cell-penetrating potential of modified peptides containing natural and chemically modified residues. Frontiers in microbiology, 2018, 9, 725. doi: 10.33 89/fmicb.2018.00725.
5. Hofland H.E., Shephard L., Sullivan S.M. Formation of stable cationic lipid/DNA complexes for gene transfer. Proc. Natl. Acad. Sci. USA 1996, 93, 7305-7309. doi: 10.1073/pnas.93.14.7305.
6. Turetskiy E.A., Koloskova О.O., Nosova A.S., Shilovskiy I.P., Sebyakin Y.L., Khaitov M.R. Physicochemical properties of lipopeptide-based liposomes and their complexes with siRNA. Biomeditsinskaia khimiia, 2017, 63(5): 472-475. doi: 10.18097/PBMC20176305472.
7. Regberg J., Srimanee A., Erlandsson M., Sillard R., Dobchev D.A., Karelson M., Langel U. Rational design of a series of novel amphipathic cell-penetrating peptides. Int. J. Pharm., 2014, 464(1-2): 111-116. doi: 10.1016/j.ijpharm.2014.01.018.
8. Eggimann G.A., Blattes E., Buschor S., Biswas R., Kammer S.M., Darbre Т., Reymond J.-L. Designed cell penetrating peptide dendrimers efficiently internalize cargo into cells. Chem. Comm., 2014, 50, 7254. doi: 10.1039/c4cc02780a.
9. Shcharbin D., Shcharbina N., Bryszewska M. Recent Patents in Dendrimers for Nanomedicine: Evolution 2014. Recent Patents on Nanomedicine, 2014, 4(1):25-31.
10. Kozhikhova KV, Andreev SM, Shilovskiy IP, Timofeeva AV, Gaisina AR, et al. A novel peptide dendrimer LTP efficiently facilitates transfection of mammalian cells. Organic & Biomolecular Chemistry 16 (43), 8181-8190.
11. Wu J., Huang W., He Z. Dendrimers as Carriers for siRNA Delivery and Gene Silencing: A Review. Sci. World J., 2013, http://dx.doi.org/10.1155/2013/630654.
12. Luo K., et al. "Arginine functionalized peptide dendrimers as potential gene delivery vehicles" Biomaterials, 2012, 33(19) 4917-4927.
13. Heitz M., et al. Stereoselective pH responsive peptide dendrimers for siRNA» (Bioconjug. Chem. 2019, 30(8):2165-2182.
14. Gorzkiewicz M., Konopka M., Janaszewska A., Tarasenko I.I., … Klajnert-Maculewicz B. Application of new lysine-based peptide dendrimers D3K2 and D3G2 for gene delivery: Specific cytotoxicity to cancer cells and transfection in vitro. Bioorg. Chem., 2020, 95, 103504.
15. Conti D.S., Brewer D., Grashik J., Avasarala S., da Rocha S.R.P. Poly(amidoamine) dendrimer nanocarriers and their aerosol formulations for siRNA delivery to the lung epithelium. Mol Pharm., 2014, 11 (6): 1808-1822.
16. Ghosh S., Firdous M.S., Nath A. siRNA could be a potential therapy for COVID-19. EXCLI J., 2020, 19:528-531.
| название | год | авторы | номер документа |
|---|---|---|---|
| СРЕДСТВО ДЛЯ ВНУТРИКЛЕТОЧНОЙ ДОСТАВКИ НУКЛЕИНОВЫХ КИСЛОТ В КЛЕТКИ МЛЕКОПИТАЮЩИХ | 2014 |
|
RU2572575C1 |
| Меченые дендримерные пептиды | 2016 |
|
RU2611399C1 |
| Комбинированное лекарственное средство, обладающее противовирусным эффектом в отношении нового коронавируса SARS-CoV-2 | 2021 |
|
RU2746362C1 |
| КОМПОЗИЦИЯ ДЛЯ ПОДАВЛЕНИЯ ЭКСПРЕССИИ ГЕНА ЦИТОКИНА ИНТЕРЛЕЙКИНА-4 | 2014 |
|
RU2563989C1 |
| Композиция, подавляющая экспрессию генов интерлейкина-4 и интерлейкина-13, для терапии аллергического ринита | 2018 |
|
RU2710895C1 |
| N-галлаты синтетических пептидов с антиоксидантной активностью | 2023 |
|
RU2828796C1 |
| Катионный разветвленный пептид для ингибирования респираторных вирусов | 2022 |
|
RU2810986C2 |
| Способ получения амфифильных блок-сополимеров N,N-диметиламиноэтилметакрилата для доставки нуклеиновых кислот в живые клетки | 2014 |
|
RU2617059C2 |
| Средство для ингибирования репликации вируса SARS-CoV-2, опосредованного РНК-интерференцией | 2020 |
|
RU2733361C1 |
| МОЛЕКУЛЯРНЫЙ КОНЪЮГАТ НА ОСНОВЕ СИНТЕТИЧЕСКИХ АНАЛОГОВ ЛЮЛИБЕРИНА И ЕГО ПРИМЕНЕНИЕ В КАЧЕСТВЕ СРЕДСТВА ДОСТАВКИ ДНК В КЛЕТКИ ГОРМОН-ЧУВСТВИТЕЛЬНЫХ ОПУХОЛЕЙ (ВАРИАНТЫ) | 2007 |
|
RU2377247C2 |
Изобретение относится к области генной инженерии и биохимии, а также к медицине и фармацевтике и направлено на создание малотоксичных катионных дендримерных пептидов как внутриклеточных транспортеров нуклеиновых кислот (НК) различной структуры. Предложено средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих, представляющее собой дендримерный катионный пептид. Полученное средство обеспечивает доставку как плазмидной ДНК, так и малых интерферирующих РНК (миРНК), что позволяет использовать его как перспективный агент для генной терапии, включая терапию на основе РНК-интерференции. 15 ил., 2 табл., 3 пр.
Средство для внутриклеточной доставки нуклеиновых кислот в клетки млекопитающих, представляющее собой дендримерный катионный пептид, выбранный из:
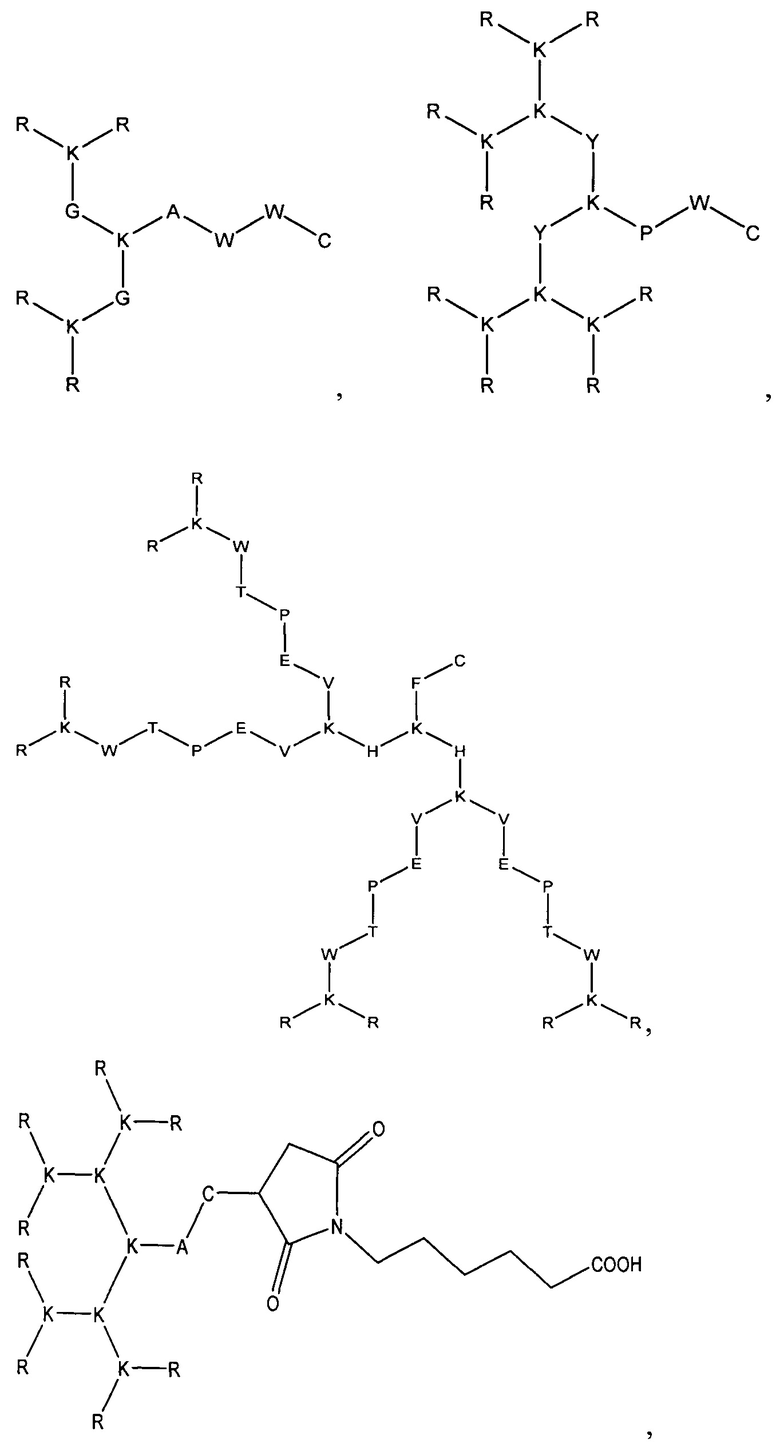
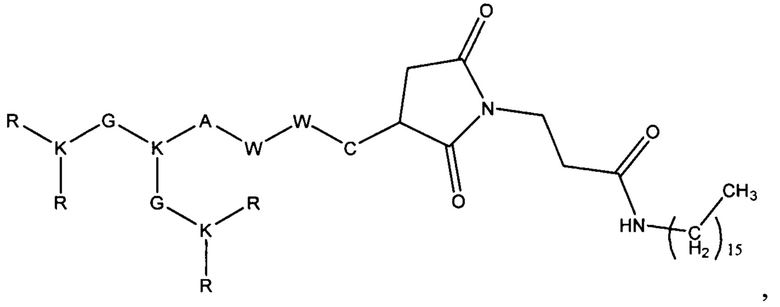
где аминокислоты представляют собой:
аргинин - R, лизин - К, глицин - G, триптофан - W, треонин - Т, пролин - Р, глутаминовая кислота - Е, валин - V, аланин - А, гистидин - Н, фенилаланин - F, тирозин - Y.
| СРЕДСТВО ДЛЯ ВНУТРИКЛЕТОЧНОЙ ДОСТАВКИ НУКЛЕИНОВЫХ КИСЛОТ В КЛЕТКИ МЛЕКОПИТАЮЩИХ | 2014 |
|
RU2572575C1 |
| EP 708637 A1, 01.05.1996 | |||
| US 20200155660 A1, 21.05.2020 | |||
| US 10029014 B2, 24.07.2018 | |||
| US 10336792 B2, 02.07.2019 | |||
| US 20030144230 A1, 31.07.2003 | |||
| Использование композиции, состоящей из катионного пептида LTP и молекул РНК против респираторных вирусов | 2015 |
|
RU2609857C1 |

