Настоящее изобретение относится к области ингибирования ферментов, в частности к (поли)пептидным ингибиторам c-Jun-аминотерминальной киназы (JNK). Настоящее изобретение относится также к способам повышения титров антител к указанным (поли)пептидным ингибиторам, а также к соответствующим антителам и продуцирующим их клеткам.
c-Jun-аминотерминальная киназа (JNK) является представителем группы активируемых стрессом митогенактивируемых протеин-(МАР)-киназ. Эти киназы принимают участие в контроле роста и дифференцировки клеток и более конкретно, в ответе клеток на стимулы окружающей среды. Путь трансдукции сигнала JNK активируется в ответ на вызываемый окружающей средой стресс и в этом участвуют несколько классов рецепторов клеточной поверхности. Эти рецепторы могут представлять собой рецепторы цитокинов, рецепторы серпентина и рецепторы тирозинкиназ. В клетках млекопитающих JNK принимает участие в биологических процессах, таких как онкогенная трансформация и опосредование адаптивных ответов на связанный с окружающей средой стресс.JNK ассоциирована также с модулированием иммунных ответов, таких как созревание и дифференцировка иммунных клеток, а также с воздействием на запрограммированную гибель клеток, выявленных иммунной системой в качестве мишеней для деструкции. Известно, что митогенактивируемая протеинкиназа (МАРК) р38альфа обладает способностью осуществлять негативную регуляцию клеточной пролиферации путем антагонистического воздействия на путь JNK-c-Jun. Таким образом, митогенактивируемая протеинкиназа (МАРК) р38альфа, по-видимому, может обладать активностью, заключающейся в подавлении пролиферации здоровых и раковых клеток (см., например, Hui и др. Nature Genetics, т.39, №6, июнь 2007 г.). Установлено также, что c-Jun-N-терминальная киназа (JNK) участвует в возникновении нейропатической боли, вызываемой лигатурой спинального нерва (SNL), при этом SNL индуцирует медленную и постоянную активацию JNK, в частности JNK1, в то же время установлено, что активация митогенактивируемой протеинкиназы р38 в микроглии спинного мозга после SNL, снижается практически до исходного уровня через 21 день (Zhuang и др., The Journal of Neuroscience, 26(13), 29 марта 2006 г., cc.3551-3560).
Ингибиторы пути передачи сигналов JNK уже известны в данной области, в частности, к ним относятся, например, расположенные против хода транскрипции ингибиторы киназ (например, СЕР-1347), малые химические ингибиторы JNK (SP600125 и AS601245), которые оказывают непосредственное воздействие на киназную активность, например, конкурируя за сайт связывания АТФ протеинкиназы, и пептидные ингибиторы взаимодействия между JNK и ее субстратами (см., например, Kuan и др. Current Drug Targets - CNS & Neurological Disorders, т.4, №1, февраль 2005 г., cc.63-67; WO 2007/031280; все документы включены в настоящее описание в качестве ссылки). В WO 2007/031280 описаны малые обладающие способностью проникать в клетку слитые пептиды, которые содержат последовательность так называемого ТАТ-транспортера, выведенную из основной обеспечивающей транспорт последовательности белка ТАТ ВИЧ, и аминокислотную ингибирующую последовательность IB1.
В WO 2007/031280 описаны также две конкретные последовательности, L-TAT-IB1 (GRKKRRQRRRPPRPKRPTTLNLFPQVPRSQD, в настоящем описании SEQ ID NO: 196) и D-TAT-IB1 (dqsrpvqpflnlttprkprpprrrqrrkkrg; в настоящем описании SEQ ID NO: 197), последняя представляет собой ретро-инвертированную последовательность (т.е. изомер линейной полипептидной последовательности, в котором направление последовательности изменено на противоположное и хиральность каждого аминокислотного остатка является инвертированной) L-TAT-IB1. Благодаря присутствию последовательности ВИЧ ТАТ, выведенной из транспортной последовательности, эти слитые пептиды более эффективно транспортируются в клетки-мишени, где они сохраняют эффективность вплоть до протеолитического расщепления.
Поскольку АТФ-зависимые пептидные ингибиторы JNK, как правило, являются более специфическими ингибиторами, их часто отбирают, прежде всего, по наличию способности ингибировать JNK. Однако даже описанные в WO 2007/031280 пептидные ингибиторы не являются оптимальными. Например, соединение L-TAT-IB1 (в настоящем описании его последовательность представлена в SEQ ID NO: 196), которое состоит только из L-аминокислот, подвергается быстрому протеолитическому расщеплению. Для решения указанной проблемы авторы изобретения, представленного в WO 2007/031280, предложили также соединение D-TAT-IB1 (в настоящем описании его последовательность представлена в SEQ ID NO: 197), которое содержит D-аминокислоты. Более точно, D-TAT-IB1 представляет собой ретро-инвертированную последовательность L-TAT-IB1. Включение D-аминокислот является сложным, поскольку изменение стереохимии может приводить к утрате функции. Подход, основанный на применении ретро-инвертированных пептидов, можно применять для снижения указанного риска, поскольку используют I) только D-аминокислоты, II) но в инвертированной пептидной последовательности может оказаться более вероятным получать приемлемую конформацию, аналогичную исходному пептиду, чем при включении одной или нескольких D-аминокислот в исходную последовательность. Тем не менее, в случае, который описан в WO 2007/031280, этот подход привел к значительному снижению ингибирующей активности по сравнению с L-TAT-IB1 (см. фиг.4). Кроме того, ретро-инвертированный пептид является очень устойчивым к протеолитическому расщеплению, что делает контролируемое расщепление, например, в экспериментах, чувствительных к временным параметрам, маловероятным.
Таким образом, в данной области все еще сохраняется необходимость в пептидных ингибиторах JNK, которые являются более стабильными, чем, например L-TAT-IB1 (в настоящем описании его последовательность представлена в SEQ ID NO: 196). С другой стороны, существует потребность в пептидных ингибиторах JNK, которые являются более активными и при этом менее стабильными, чем, например D-TAT-IB1 (в настоящем описании его последовательность представлена в SEQ ID NO: 197).
Таким образом, задачу, положенную в основу настоящего изобретения, можно решать с помощью дополнительных (пептидных) ингибиторов JNK, которые предпочтительно являются менее чувствительными к протеолитическому расщеплению, чем соединение L-TAT-IB1, описанное в WO 2007/031280, но предпочтительно одновременно являются более чувствительными к протеолитическому расщеплению и/или более активными, чем соединение D-TAT-IB1, описанное в WO 2007/031280.
Задача, положенная в основу настоящего изобретения, решена заявителями с помощью объекта изобретения, изложенного в прилагаемой формуле изобретения.
Ниже представлено краткое описание прилагаемых чертежей. Чертежи предназначены для более подробной иллюстрации настоящего изобретения. Однако они никоим образом не направлены на ограничение объема объекта изобретения.
На чертежах показано:
на фиг.1 - иллюстрация ингибирующего действия нескольких ингибиторов JNK, предлагаемых в настоящем изобретении, для изучения которых in vitro применяли AlphaScreen-анализ (Amplified Luminescence Proximity Homogenous Assay (гомогенный анализ усиленной люминесценции в ближней области));
на фиг.1А - данные об ингибировании JNK1 с помощью соединений, последовательности которых представлены в SEQ ID NO: 193, 2, 3, 5, 6 и 7;
на фиг.1Б - данные об ингибировании JNK2 с помощью соединений, последовательности которых представлены в SEQ ID NO: 193, 2, 3, 5, 6 и 7;
на фиг.1В - данные об ингибировании JNK3 с помощью соединений, последовательности которых представлены в SEQ ID NO: 193, 2, 3, 5, 6 и 7;
на фиг.2 - таблица, в которой проиллюстрировано ингибирующее действие нескольких ингибиторов JNK (SEQ ID NO: 193, 2, 3, 5, 6 и 7), предлагаемых в настоящем изобретении. Представлены значения IC50, находящиеся в нМ-диапазоне, соответствующие стандартные отклонения от среднего значения и количество осуществленных экспериментов (n);
на фиг.3 - иллюстрация ингибирующего действия нескольких ингибиторов JNK, предлагаемых в настоящем изобретении, которые представляли собой слитые белки, включающие (поли)пептидную последовательность ингибитора JNK и последовательность-транспортер. Ингибирующую активность определяли in vitro с помощью AlphaScreen-анализа (гомогенный анализ усиленной люминесценции в ближней области);
на фиг.3А - данные об ингибировании JNK1 с помощью соединений, последовательности которых представлены в SEQ ID NO: 194, 195, 172, 200, 46, 173, 174, 175, 176, 177, 178, 179, 180, 181 и 197;
на фиг.3Б - данные об ингибировании JNK2 с помощью соединений, последовательности которых представлены в SEQ ID NO: 194, 195, 172, 200, 46, 173, 174, 175, 176, 177, 178, 179, 180, 181 и 197;
на фиг.3В - данные об ингибировании JNK3 с помощью соединений, последовательности которых представлены в SEQ ID NO: 194, 195, 172, 200, 46, 173, 174, 175, 176, 177, 178, 179, 180, 181 и 197;
на фиг.3Г - данные об ингибировании JNK1 с помощью соединений, последовательности которых представлены в SEQ ID NO: 194, 195, 172, 200, 46, 182, 183, 184, 185, 186, 187, 188, 189, 190 и 197;
на фиг.3Д - данные об ингибировании JNK2 с помощью соединений, последовательности которых представлены в SEQ ID NO: 194, 195, 172, 200, 46, 182, 183, 184, 185, 186, 187, 188, 189, 190 и 197;
на фиг.3Е - данные об ингибировании JNK3 с помощью соединений, последовательности которых представлены в SEQ ID NO: 194, 195, 172, 200, 46, 182, 183, 184, 185, 186, 187, 188, 189, 190 и 197;
на фиг.4 - таблица, в которой проиллюстрировано ингибирующее действие нескольких ингибиторов JNK, предлагаемых в настоящем изобретении, которые представляли собой слитые белки, включающие (поли)пептидную последовательность ингибитора JNK и последовательность-транспортер. Представлены значения IC50, находящиеся в нМ-диапазоне, соответствующие стандартные отклонения от среднего значения (СКО) и количество осуществленных экспериментов (n);
на фиг.5 - данные о стабильности ингибиторов JNK, последовательности которых представлены в SEQ ID NO: 172, 196 и 197, в 50% человеческой сыворотке. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 196, полностью расщеплялся на аминокислотные остатки в течение 6 ч (А). Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, полностью расщеплялся только через 14 дней (Б). Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 197, сохранял стабильность в течение периода времени, составлявшего вплоть до 30 дней (Б);
на фиг.6 - результаты экспериментов по интернализации с использованием выведенных из ТАТ конструкций транспортеров, имеющих схему D-аминокислот/L-аминокислот, которая представлена в SEQ ID NO: 30. Проанализированные последовательности транспортеров соответствуют SEQ ID NO: 52-94 плюс SEQ ID NO: 45, 47, 46, 43 и 99 (фиг.6а) и SEQ ID NO: 100-147 (фиг.6б). Продемонстрировано, что все транспортеры, несущие консенсусную последовательность rXXXrXXXr (SEQ ID NO: 31), обладали повышенной способностью к интернализации по сравнению с транспортером L-TAT (SEQ ID NO: 43). Клетки линии Hela инкубировали в течение 24 ч в 96-луночном планшете с соответствующими транспортерами в концентрации 10 мМ. Затем клетки отмывали дважды кислым буфером (0,2М глицин, 0,15М NaCl, pH 3,0) и дважды ЗФР. Клетки разрушали путем добавления буфера для лизиса RIPA. Затем относительное количество интернализированного пептида определяли, оценивая интенсивность флуоресценции (планшет-ридер Fusion Alpha; фирма PerkinElmer) каждого экстракта после вычитания фонового уровня;
на фиг.7 - данные, иллюстрирующие, что ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, блокирует индуцированное ЛПС высвобождение цитокинов и хемокинов в клетках ТНР1, дифференцированных в макрофаги с помощью ФМА (форболмиристатацетат). На фиг.7А - данные о высвобождении TNFα (ТНР1-ФМА, 6 ч, 3 нг/мл ЛПС); на фиг.7Б - данные о высвобождении TNFα (ТНР1-ФМА, 6 ч, 10 нг/мл ЛПС); на фиг.7В - данные о высвобождении IL6 (ТНР1-ФМА, 6 ч, 10 нг/мл ЛПС); на фиг.7Г - данные о высвобождении МСР1 (ТНР1-ФМА, 6 ч, 3 нг/мл ЛПС);
на фиг.8 - данные, иллюстрирующие, что ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, блокирует индуцированное ЛПС высвобождение IL6 в дифференцированных в макрофаги клетках ТНР1 с более высокой эффективностью, чем D-TAT-IB1 (SEQ ID NO: 197), dTAT (SEQ ID NO: 45) и SP 600125. ЛПС добавляли в течение 6 ч (10 нг/мл);
на фиг.9 - данные, иллюстрирующие, что ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, блокирует индуцированное ЛПС высвобождение TNFα в дифференцированных в макрофаги клетках ТНР1 с более высокой эффективностью, чем D-TAT-IB1 (SEQ ID NO: 197), dTAT (SEQ ID NO: 45) и SP 600125. ЛПС добавляли в течение 6 ч (10 нг/мл);
на фиг.10 - данные, иллюстрирующие, что ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, блокирует индуцированное ЛПС высвобождение IL-6 в макрофагах, полученных в результате дифференцировки с помощью обработки ФМА, с более высокой эффективностью, чем D-TAT-IB1 (SEQ ID NO: 197) и L-TAT-IB1 (SEQ ID NO: 196). ЛПС добавляли в течение 6 ч;
на фиг.11 - данные, иллюстрирующие, что ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, блокирует индуцированное ЛПС высвобождение TNFα в макрофагах, полученных в результате дифференцировки с помощью обработки ФМА, с более высокой эффективностью, чем D-TAT-IB1 (SEQ ID NO: 197) и L-TAT-IB1 (SEQ ID NO: 196);
на фиг.12 - данные, иллюстрирующие, что ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, блокирует индуцированное ЛПС высвобождение TNFα в первичных клетках цельной крови крыс при его использовании в концентрации 3 нг/мл. Представлены результаты для контроля, 1 мкМ SEQ ID NO: 172, 3 мкМ SEQ ID NO: 172 и 10 мкМ SEQ ID NO: 172 при применении ЛПС в различных концентрациях (нг/мл);
на фиг.13 - данные, иллюстрирующие, что ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, блокирует секрецию IL2 первичными человеческими Т-клетками в ответ на обработку ФМА/иономицином;
на фиг.14 - данные, иллюстрирующие, что ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, блокирует секрецию IL2 первичными человеческими Т-клетками в ответ на стимуляцию CD3/CD28. Применяемые ингибиторы JNK имели последовательности, представленные в SEQ ID NO: 172 и 197;
на фиг.15 - данные, иллюстрирующие зависящее от дозы ингибирование ингибитором JNK последовательность которого представлена в SEQ ID NO: 172, индуцированного CD3/CD28 высвобождения IL-2 в первичных выделенных из лимфатических узлов крыс Т-клетках. Контрольную крысу умерщвляли и изымали лимфатические узлы. Т-клетки дополнительно очищали (с помощью негативной селекции с использованием магнитного поля) и высевали в 96-луночные планшеты из расчета 200000 клеток/лунку. Клетки обрабатывали антителом к крысиному CD3 и антителом к крысиному CD28 (2 мкг/мл). Добавляли в культуры ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, за 1 ч до обработки CD3/CD28 и высвобождение IL-2 оценивали в супернатанте через 24 ч после обработки;
на фиг.16 - данные, иллюстрирующие зависящее от дозы ингибирование индуцированного CD3/CD28 высвобождения IL-2 в первичных выделенных из лимфатических узлов крыс Т-клеток: сравнение нескольких ингибиторов JNK, а именно, ингибиторов, которые имели последовательности, представленные в SEQ ID NO: 172, 197, и SP600125;
на фиг.17 - данные, иллюстрирующие зависящее от дозы ингибирование высвобождения IL-2 в цельной крови крыс, стимулированной ФМА + иономицином. Добавляли ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, в трех различных концентрациях, а именно 1, 3 и 10 мкМ за 1 ч до стимуляции ФМА + иономицином. В течение 4 ч добавляли активаторы в трех дозах (25/500 нг/мл, 50/750 нг/мл и 50/1000 нг/мл). Оценивали высвобождение IL-2 в супернатанте. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, в концентрации 10 мкМ обладал способностью эффективно снижать индуцированное ФМА-иономицином высвобождение IL-2 при применении активаторов в трех тестированных концентрациях;
на фиг.18 - данные по оценке ингибирования JNK и высвобождения IL-6 в цельной крови человека. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, добавляли в трех различных концентрациях, а именно 1, 3 и 10 мкМ, за 1 ч до стимуляции цельной крови ЛПС (0,02 нг/мл) в течение 4 ч. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, обладал способностью снижать индуцированное ЛПС высвобождение IL-6 в зависимости от дозы;
на фиг.19 - данные по оценке ингибирования JNK и высвобождения IL-2 в цельной крови человека. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, добавляли в трех различных концентрациях, а именно 1, 3 и 10 мкМ, за 1 ч до стимуляции цельной крови ФМА + иономицином (25/700 нг/мл, 50/800 нг/мл и 50/1000 нг/мл) в течение 4 ч. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, обладал способностью снижать индуцированное ФМА + иономицином высвобождение IL-2 в зависимости от дозы;
на фиг.20 - данные по оценке ингибирования JNK и высвобождения IFN-γ в цельной крови человека. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, добавляли в трех различных концентрациях, а именно 1, 3 и 10 мкМ, за 1 ч до стимуляции цельной крови ФМА + иономицином (25/700 нг/мл, 50/800 нг/мл и 50/1000 нг/мл) в течение 4 ч. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, обладал способностью снижать индуцированное ФМА + иономицином высвобождение IFN-γ в зависимости от дозы;
на фиг.21 - данные по оценке ингибирования JNK и высвобождения TNF-α в цельной крови человека. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, добавляли в трех различных концентрациях, а именно 1, 3 и 10 мкМ, за 1 ч до стимуляции цельной крови ФМА + иономицином (25/700 нг/мл, 50/800 нг/мл и 50/1000 нг/мл) в течение 4 ч. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, обладал способностью снижать индуцированное ФМА + иономицином высвобождение TNF-α в зависимости от дозы;
на фиг.22 - данные по оценке ингибирования JNK и высвобождения TNF-α в цельной крови человека. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, добавляли в трех различных концентрациях, а именно 1, 3 и 10 мкМ, за 1 ч до стимуляции цельной крови фитогемагглютинином-L (ФГА-L (5 мкг/мл) в течение 3 дней. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, обладал способностью снижать индуцированное ФГА-L высвобождение TNF-α в зависимости от дозы;
на фиг.23 - данные по оценке ингибирования JNK и высвобождения IL-2 в цельной крови человека. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, добавляли в трех различных концентрациях, а именно 1, 3 и 10 мкМ, за 1 ч до стимуляции цельной крови ФГА-L (5 мкг/мл) в течение 3 дней. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, обладал способностью снижать индуцированное ФГА-L высвобождение IL-2 в зависимости от дозы;
на фиг.24 - данные по оценке ингибирования JNK и высвобождения TNF-α в цельной крови человека. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, добавляли в трех различных концентрациях, а именно 1, 3 и 10 мкМ, за 1 ч до стимуляции цельной крови антителами к CD3+/-CD28 (2 мкг/мл) в течение 3 дней. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, обладал способностью снижать индуцированное CD3/CD28 высвобождение TNF-α в зависимости от дозы.
Ингибиторы JNK
Первым объектом настоящего изобретения является ингибитор JNK, который содержит ингибирующую (поли)пептидную последовательность, имеющую следующую общую формулу:
X1-X2-X3-R-X4-X5-X6-L-X7-L-X8 (SEQ ID NO: 1),
в которой X1 обозначает аминокислоту, выбранную из аминокислот R, Р, Q и r,
в которой Х2 обозначает аминокислоту, выбранную из аминокислот R, Р, G и r,
в которой Х3 обозначает аминокислоту, выбранную из аминокислот К, R, k и r,
в которой Х4 обозначает аминокислоту, выбранную из аминокислот Р и К,
в которой Х5 обозначает аминокислоту, выбранную из аминокислот Т, a, s, q, k, или Х5 отсутствует,
в которой Х6 обозначает аминокислоту, выбранную из аминокислот Т, D и А,
в которой Х7 обозначает аминокислоту, выбранную из аминокислот N, n, r и К; и
в которой Х8 обозначает аминокислоту, выбранную из аминокислот F, f и w,
при условии, что по меньшей мере одна, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или шесть аминокислот, выбранных из группы, включающей X1, Х2, Х3, Х5, Х7 и Х8, представляет(ют) собой D-аминокислоту(ы), предпочтительно при условии, что по меньшей мере одна, по меньшей мере две, по меньшей мере три или четыре аминокислоты, выбранные из группы, включающей Х3, Х5, Х7 и Х8, представляет(ют) собой D-аминокислоту(ы).
Ингибирующая (поли)пептидная последовательность ингибитора JNK, предлагаемого в настоящем изобретении, содержит L-аминокислоты и в большинстве вариантов осуществления изобретения содержит D-аминокислоты. Если специально не указано иное, то в контексте настоящего описания остатки L-аминокислот обозначают заглавными буквами, а остатки D-аминокислот обозначают прописными буквами. Глицин может быть обозначает заглавными или прописными буквами (поскольку не существует D- или L-глицина). Аминокислотные последовательности, представленные в настоящем описании, всегда указаны в направлении от N- конца к С-концу (слева направо), если специально не указано иное. Представленная аминокислотная последовательность может быть модифицированной или немодифицированной на С- и/или N-конце, например, ацетилирована на С-конце и/или амидирована или модифицирована с помощью цистеинамида на N-конце. Очевидные указанные возможные, но совершенно необязательные модификации на С- и/или N-конце аминокислотных последовательностей, представленных в настоящем описании, конкретно не обозначены.
Ингибиторы JNK, предлагаемые в настоящем изобретении, представляют собой (поли)пептидные ингибиторы c-Jun-N-терминальной киназы (JNK). Указанные ингибиторы ингибируют киназную активность c-Jun N-терминальной киназы (JNK), т.е. препятствуют фосфорилированию или снижают уровень фосфорилирования субстратов JNK, таких как c-Jun, ATF2 и/или Elk-1. Специалисту в данной области должно быть очевидно, что в контексте настоящего описания понятие «ингибитор» не относится к соединениям, которые необратимо разрушают молекулу c-Jun-N-терминальной киназы (JNK) и/или элиминируют киназную активность. Кроме того, в контексте настоящего описания понятие «ингибирование активности JNK» относится к ингибированию киназной активности c-Jun-N-терминальной киназы (JNK).
Кроме того, согласно настоящему описанию ингибитор JNK содержит по меньшей мере одну функциональную единицу полимера, состоящего из аминокислот, т.е. имеющего (поли)пептидную последовательность. Кроме того, указанный по меньшей мере один функциональный полимер, состоящий из аминокислот, обладает способностью ингибировать активность JNK. Аминокислотные мономеры указанной ингибирующей (поли)пептидной последовательности, как правило, сцеплены друг с другом через пептидные связи, но допустимы также (химические) модификации указанной(ых) пептидной(ых) связи(ей) или остатков боковых цепей, при условии, что ингибирующая активность (ингибирование активности JNK) не утрачивается полностью, т.е. образовавшаяся химическая субстанция все еще классифицируется в качестве функционального ингибитора JNK согласно настоящему описанию. Понятие «(поли)пептид» не рассматривается как ограничивающее длину (поли)пептидной единицы. Предпочтительно ингибирующая (поли)пептидная последовательность ингибиторов JNK, предлагаемых в настоящем изобретении, состоит менее чем из 500, 490, 480, 470, 460, 450, 440, 430, 420, 410, 400, 390, 380, 370, 360, 350, 340, 330, 320, 310, 300, 290, 280, 270, 260, 250, 240, 230, 220, 210, 200, 190, 180, 170, 160, 150, 140, 130, 120, 110, 100, 95, 90, 85, 80, 75, 70, 65, 60, 55, 50, 49, 48, 47, 46, 45, 44, 43, 42, 41, 40, 39, 38, 37, 36, 35, 34, 33, 32, 31, 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13 или менее чем из 12 аминокислот. Предпочтительно ингибирующая (поли)пептидная последовательность не содержит менее 10 аминокислотных остатков, более предпочтительно не содержит менее 11 аминокислотных остатков.
Кроме того, «ингибитор JNK», предлагаемый в настоящем изобретении, ингибирует активность JNK, например, касательно ингибирования опосредуемого человеческой JNK фосфорилирования субстрата, представляющего собой c-Jun (SEQ ID NO: 198), что характеризуется следующими значения IC50:
а) менее 3000 нМ, более предпочтительно менее 2000 нМ, еще более предпочтительно менее 1000 нМ, еще более предпочтительно менее 500 нМ, еще более предпочтительно менее 250 нМ, еще более предпочтительно менее 200 нМ, еще более предпочтительно менее 150 нМ, наиболее предпочтительно менее 100 нМ, касательно ингибирования человеческой JNK1,
б) менее 3000 нМ, более предпочтительно менее 2000 нМ, еще более предпочтительно менее 1000 нМ, еще более предпочтительно менее 500 нМ, еще более предпочтительно менее 250 нМ, еще более предпочтительно менее 200 нМ, еще более предпочтительно менее 150 нМ, наиболее предпочтительно менее 100 нМ, касательно ингибирования человеческой JNK2, и/или
в) менее 3000 нМ, более предпочтительно менее 2000 нМ, еще более предпочтительно менее 1000 нМ, еще более предпочтительно менее 500 нМ, еще более предпочтительно менее 250 нМ, еще более предпочтительно менее 200 нМ, еще более предпочтительно менее 150 нМ, наиболее предпочтительно менее 100 нМ, касательно ингибирования человеческой JNK3.
Для некоторых вариантов применения предпочтительно, чтобы ингибитор ингибировал указанную выше человеческую JNK2 и/или человеческую JNK3, но не ингибировал указанную выше JNK1.
Специалист в данной области легко может определить ингибируется ли или нет активность JNK. В данной области известно несколько методов. Одним из примеров является радиоактивный анализ киназ или нерадиоактивный анализ киназ (например, AlphaScreen-анализ; см., например, Guenat и др., J Biomol Screen, 11, 2006, cc.1015-1026).
Таким образом, ингибитор JNK, предлагаемый в настоящем изобретении, может, например, содержать ингибирующую (поли)пептидную последовательность, представленную в любой из SEQ ID NO: 2-27 (см. таблицу 1).
Ингибитор JNK, предлагаемый в настоящем изобретении, может представлять собой также ингибитор JNK (вариант), содержащий ингибирующую (поли)пептидную последовательность, которая по меньшей мере на 50%, более предпочтительно по меньшей мере на 55%, более предпочтительно по меньшей мере на 60%, более предпочтительно по меньшей мере на 65%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% идентична последовательности, выбранной из последовательностей, представленных в SEQ ID NO: 1-27, в частности, последовательности, представленной в SEQ ID NO: 8,
при условии, что относительно соответствующей последовательности, выбранной из SEQ ID NO: 1-27, в указанной идентичной ингибирующей (поли)пептидной последовательности
а) сохраняется остаток L-аргинина (R) в положении 4,
б) сохраняются два остатка L-лейцина (L) в положениях 8 и 10 (положения 7 и 9 в SEQ ID NO: 25-27),
в) присутствует(ют) одна, две, три, четыре, пять или шесть D-аминокислота(от) в указанных положениях, соответствующих аминокислотам, выбранным из группы, включающей X1, Х2, Х3, Х5, Х7 и Х8 в SEQ ID NO: 1, и в указанных положениях в SEQ ID NO: 2-27, более предпочтительно присутствует(ют) одна, две, три или четыре D-аминокислота(ты) в указанных положениях, соответствующих аминокислотам, выбранным из группы, включающей ХЗ, Х5, Х7 и Х8 в SEQ ID NO: 1, и в указанных положениях в SEQ ID NO: 2-27, и
г) она все еще обладает способностью ингибировать активность JNK (т.е. представляет собой ингибитор JNK, как он указан в настоящем описании).
Таким образом, для вариантов, представленных в настоящем описании (в частности, вариантов ингибитора JNK, которые содержат ингибирующую (поли)пептидную последовательность, удовлетворяющую указанному выше определению, характерна определенная степень идентичности последовательности с последовательностью, выбранной из SEQ ID NO: 1-27), предпочтительно характерна идентичность последовательностей с соответствующей референс-последовательностью, составляющая менее 100%.
В свете указанного определения и также для ясности изложения, в таблице 1 подчеркнуты остатки, которые нельзя изменять в вариантах ингибиторов JNK, содержащих последовательности, представленные в SEQ ID NO: 1-27 (см. подпункты а) и б) в указанном выше определении).
Неидентичные аминокислоты предпочтительно являются результатом осуществления консервативных аминокислотных замен.
Консервативные аминокислотные замены согласно настоящему описанию могут включать замены аминокислотных остатков в пределах группы остатков, обладающих в достаточной степени сходными физико-химическими свойствами, в результате чего при замене представителями группы должна сохраняться биологическая активность молекулы (см., например, Grantham R., Science 185, 1974, cc.862-864). В частности, консервативные аминокислотные замены предпочтительно представляют собой замены, при которых аминокислоты, относятся к одному и тому же классу аминокислот (например, основные аминокислоты, кислые аминокислоты, полярные аминокислоты, аминокислоты с алифатической боковой цепью, аминокислоты с положительно или отрицательно заряженными боковыми цепями, аминокислоты с ароматическими группами в боковых цепях, аминокислоты, боковые цепи которых могут участвовать в образовании водородных связей, например, боковые цепи которых несут гидроксильную функцию, и т.д.). В контексте настоящего описания консервативная замена представляет собой, например, замену остатка основной аминокислоты (Lys, Arg, His) на остаток другой основной аминокислоты (Lys, Arg, His), замену остатка алифатической аминокислоты (Gly, Ala, Val, Leu, Ile) на остаток другой алифатической аминокислоты, замену остатка ароматической аминокислоты (Phe, Tyr, Trp) на остаток другой ароматической аминокислоты, замену треонина на серии или лейцина на изолейцин. Другие консервативные аминокислотные замены должны быть хорошо известны специалисту в данной области. Предпочтительно должна сохраняться изомерная форма, К предпочтительно заменяют на R или Н, a k предпочтительно заменяют на r и h.
Другие возможные замены, подпадающие под указанное выше определение вариантов ингибитора JNK, представляют собой, например, варианты, в которых:
а) один, два или большее количество X1, Х2, Х3, Х4, Х5, Х6, Х7 и/или Х8 в SEQ ID NO: 1 или аминокислоты в соответствующий положениях в соответствующей последовательности, выбранной из SEQ ID NO: 2-27, заменяют на А или а,
б) X1 или Х8 в SEQ ID NO: 1 или аминокислоту в соответствующем положении в соответствующей последовательности, выбранной из SEQ ID NO: 2-27, удаляют путем делеции;
в) Х5 в SEQ ID NO: 1 или аминокислота в соответствующем положении в соответствующей последовательности, выбранной из SEQ ID NO: 2-27, обозначает Е, Y, L, V, F или К;
г) Х5 в SEQ ID NO: 1 или аминокислота в соответствующем положении в соответствующей последовательности, выбранной из SEQ ID NO: 2-27, обозначает Е, L, V, F или К; или
д) один, два или три X1, Х2, Х3 в SEQ ID NO: 1 или аминокислоты в соответствующих положениях в соответствующей последовательности, выбранной из SEQ ID NO: 2-27, обозначают нейтральные аминокислоты.
В контексте настоящего описания понятие «% идентичности последовательностей» означает следующее: две подлежащие сравнению последовательности выравнивают (подвергают сравнительному анализу), добиваясь максимальной корреляции между последовательностями. Эта процедура может включать встраивание «брешей» в одну или обе последовательности для повышения степени выравнивания (эффективности сравнительного анализа первичной структуры последовательностей). Затем % идентичности можно определять по всей длине каждой подлежащей сравнению последовательности (так называемый глобальный сравнительный анализ первичной структуры), что является более приемлемым для последовательностей, имеющих одинаковую или близкую длину, или для более коротких участков определенной длины (так называемый локальный сравнительный анализ первичной структуры), что является наиболее приемлемым для последовательностей, имеющих разную длину. В указанном выше контексте подразумевается, что аминокислотная последовательность, имеющая последовательность, «характеризующуюся идентичностью последовательности», составляющей, например, по меньшей мере 95%, с запрашиваемой аминокислотной последовательностью, означает, что последовательность рассматриваемой аминокислотной последовательности идентична запрашиваемой последовательности за исключением того, что рассматриваемая аминокислотная последовательность может включать вплоть до 5 аминокислотных изменений на каждые 100 аминокислот запрашиваемой аминокислотной последовательности. Другими словами, для получения аминокислотной последовательности, идентичной по меньшей мере на 95% запрашиваемой аминокислотной последовательности, вплоть до 5% (5 из 100) аминокислотных остатков в рассматриваемой последовательности могут быть встроены или заменены на другую аминокислоту или удалены в результате делеции. Для целей определения идентичности последовательностей замена L-аминокислоты на D-аминокислоту (и наоборот) рассматриваются, как приводящая к получению неидентичного остатка, даже, если он представляет собой только лишь D- (или L-) изомер точно такой же аминокислоты.
Методы сравнения идентичности и гомологии двух или большего количества последовательностей хорошо известны в данной области. Процент идентичности двух последовательностей можно определять, например, с помощью математического алгоритма. Предпочтительным примером математического алгоритма, который можно применять, является (но, не ограничиваясь только им) алгоритм, описанный у Karlin и др., PNAS USA, 90, 1993, cc.5873-5877. Указанный алгоритм входит в семейство программ BLAST, например, в программу BLAST или NBLAST (см. также у Altschul и др., J. Мо1. Biol. 215, 1990, cc.403-410 или Altschul др., Nucleic Acids Res, 25, 1997, cc.3389-3402), которые доступны на домашней странице NCBI на сайте в сети Интернет ncbi.nlm.nih.gov) и FASTA (Pearson, Methods Enzymol. 183, 1990, cc.63-98; Pearson и Lipman, Proc. Natl. Acad. Sci. U.S.A. 85, 1988, cc.2444-2448.). С помощью этих программ можно идентифицировать последовательности, идентичные в определенной степени другим последовательностям. Кроме того, для определения % идентичности двух полипептидных последовательностей можно использовать пакет программ анализа последовательностей Группы компьютерной генетики Биотехнологического центра Университета Висконсина (Wisconsin Sequence Analysis Package, версия 9.1. (Devereux и др., Nucleic Acids Res. 12, 1984, cc. 387-395), например, для определения % идентичности двух полипептидных последовательностей можно применять программы BESTFIT и GAP. В программе BESTFIT использован алгоритм «локальной гомологии» (Smith и Waterman, J. Мо1. Biol. 147, 1981, cc.195-197), и она позволяет находить одну характеризующуюся наиболее высоким уровнем сходства область в двух последовательностях.
В целом, ингибитор JNK, предлагаемый в настоящем изобретении, может содержать помимо ингибирующей (поли)пептидной последовательности, указанной выше, дополнительные последовательности, домены, метки (например, флуоресцентные или радиоактивные метки), эпитопы и т.д., если при этом не теряется активность в отношении ингибирования JNK, указанная в настоящем описании. Например, ингибитор JNK, предлагаемый в настоящем изобретении, может содержать также последовательность-транспортер. «Последовательность транспортер» в контексте настоящего описания означает (поли)пептидную последовательность, обеспечивающую транслокацию присоединенной к ней молекулы через биологические мембраны. Таким образом, ингибитор JNK, предлагаемый в настоящем изобретении, содержащий последовательность-транспортер, предпочтительно обладает способностью транслоцироваться через биологические мембраны. В результате ингибитор JNK, предлагаемый в настоящем изобретении, с большей легкостью проникает в клетку, клеточный субкомпартмент и/или в ядро клетки.
Указанная последовательность-транспортер может быть, например, присоединена (например, непосредственно) к N-концу или (например, непосредственно) к С-концу ингибирующей (поли)пептидной последовательности ингибитора JNK. Последовательность-транспортер и ингибирующая (поли)пептидная последовательность могут быть также пространственно отделены друг от друга, например, могут быть разделены промежуточными последовательностями. Подразумевается также, что последовательность-транспортер может располагаться в молекуле JNK полностью не в том месте, в котором находится ингибирующая (поли)пептидная последовательность, в частности, если ингибитор JNK представляет собой более сложную молекулу (например, содержит несколько доменов, представляет собой мультимерный конъюгат и т.д.). Предполагается также, что последовательность-транспортер и ингибирующая (поли)пептидная последовательность могут перекрываться, если при этом сохраняется ингибирующая активность в отношении JNK. Примеры указанных перекрывающихся последовательностей дополнительны представлены ниже.
Последовательности-транспортеры, которые можно применять в сочетании с ингибитором JNK, предлагаемым в настоящем изобретении, можно выбирать (но, не ограничиваясь только ими) из последовательностей-транспортеров, полученных из ТАТ ВИЧ, например, нативных белков, таких, например, как ТАТ-белок (например, описанный в US 5804604 и 5674980, каждый из указанных документов включен в настоящее описание в качестве ссылки), VP22 HSV (вирус герпеса простого. Herpes simplex) (описан, например, в WO 97/05265; у Elliott и O'Hare, Cell, 88, 1997, cc.223-233), невирусных белков (Jackson и др., Proc.Natl. Acad. Sci. USA, 89, 1992, cc.10691-10695), последовательностей-транспортеров, полученных из белков генов Antennapedia, прежде всего из Drosophila antennapedia (например, последовательность-носитель Antennapedia), FGF, лактоферрина и т.д, или выведенных из основных пептидов, например, пептидов, состоящих из 5-15 аминокислот, предпочтительно 10-12 аминокислот, и содержащих по меньшей мере 80%, более предпочтительно 85% или даже 90% основных аминокислот, таких, например, как аргинин, лизин и/или гистидин, или можно выбирать, например, из богатых аргинином пептидных последовательностей, таких как RRRRRRRRR (R9; SEQ ID NO: 152), RRRRRRRR (R8; SEQ ID NO: 153), RRRRRRR (R7; SEQ ID NO: 154), RRRRRR (R6, SEQ ID NO: 155), RRRRR (R5, SEQ ID NO: 156) и т.д., из VP22, из белков или пептидов PTD-4, из RGD-K16, из РЕРТ1/2 или белков или пептидов РЕРТ1/2, из SynB3 и/или белков или пептидов SynB3, из ингибиторов PC, из белков или пептидов, выведенных из Р21, или из белков или пептидов JNKI.
Примерами последовательностей-транспортеров, которые можно применять в сочетании с ингибитором JNK, предлагаемым в настоящем изобретении, являются, в частности (но, не ограничиваясь только ими) состоящие из основных аминокислот последовательности-транспортеры, полученные из белка ТАТ ВИЧ-1. Предпочтительно состоящая из основных аминокислот последовательность-транспортер, полученная из белка ТАТ ВИЧ-1, может включать последовательности из белка ТАТ вируса иммунодефицита человека ВИЧ-1, например, описанную, в частности, в US 5804604 и 5,674,980, каждый из которых включен в настоящее описание в качестве ссылки. В этом контексте полноразмерный белок ТАТ ВИЧ-1 состоит из 86 аминокислотных остатков, кодируемых двумя экзонами гена ТАТ ВИЧ. Аминокислоты ТАТ 1-72 кодируются экзоном 1, а аминокислоты 73-86 кодируются экзоном 2. Полноразмерный белок ТАТ отличается наличием основной области, которая содержит два остатка лизина и шесть остатков аргинина (аминокислоты 49-57), и богатой цистеином области, которая содержит семь остатков цистеина (аминокислоты 22-37). Основная область (т.е. аминокислоты 49-57), вероятно, является важной для ядерной локализации (Ruben S. и др., J. Virol. 63, 1989, cc.1-8; Hauber J. и др., J. Virol. 63, 1989, cc.1181-1187). Богатая цистеином область опосредует формирование связанных металлом димеров in vitro (Frankel A.D. и др., Science 240, 1988, cc.70-73; Frankel A.D. и др., Proc. Natl. Acad. Sci USA 85, 1988, cc.6297-6300) и имеет решающее значение для его активности в качестве трансактиватора (Garcia J.А. и др., ЕМВО J. 7, 1988, с.3143; Sadaie M.R. и др., J. Virol. 63, 1989, c.1). Также как и в других регуляторных белках, N-концевая область может принимать участие в защите от внутриклеточных протеаз (Bachmair А. и др., Cell 56, 1989, cc.1019-1032). Предпочтительно последовательности-транспортеры на основе ТАТ, предназначенные для применения в ингибиторе JNK, предлагаемом в настоящем изобретении, предпочтительно отличаются наличием аминокислотной последовательности основной области ТАТ (аминокислоты 49-57 встречающегося в естественных условиях белка ТАТ); отсутствием аминокислотной последовательности богатой цистеином области ТАТ (аминокислоты 22-36 встречающегося в естественных условиях белка ТАТ) и отсутствием кодируемого экзоном 2 карбоксиконцевого домена ТАТ (аминокислоты 73-86 встречающегося в естественных условиях белка ТАТ). Более предпочтительно последовательность-транспортер в ингибиторе JNK, предлагаемом в настоящем изобретении, можно выбирать из аминокислотной последовательности, которая содержит остатки ТАТ 48-57 или 49-57, или ее вариантов.
Предпочтительно последовательность-транспортер, входящая в конкретный ингибитор JNK, предлагаемый в настоящем изобретении, включает также D-аминокислоты, например, для повышения устойчивости к действию протеаз. Наиболее предпочтительными являются последовательности-транспортеры, которые отличаются специфическим порядком расположения, характеризующимся чередованием D- и L-аминокислот. Указанный порядок чередования D- и L-аминокислот (мотив) может иметь следующую схему (но, не ограничиваясь только ими), представленную в любой из SEQ ID NO: 28-30:
d1LLLxdmLLLydn (SEQ ID NO: 28);
dLLLd(LLLd)a (SEQ ID NO: 29); и/или
dLLLdLLLd (SEQ ID NO: 30);
где d обозначает D-аминокислоту;
L обозначает L-аминокислоту;
А обозначает 0-3, предпочтительно 0-2, более предпочтительно 0, 1,2 или 3, еще более предпочтительно 0, 1 или 2 и наиболее предпочтительно 1;
l, m и n каждый независимо друг от друга обозначает 1 или 2, предпочтительно 1;
х и у каждый независимо друг от друга обозначает 0, 1 или 2, предпочтительно 1.
Указанный порядок расположения D- и L-аминокислот (мотив) становится именно таким при синтезе последовательности-транспортера, т.е. когда аминокислотная последовательность (т.е. тип остатков боковой цепи) остается неизмененной, а чередуются соответствующие изомеры. Например, известная последовательность-транспортер, полученная из ТАТ ВИЧ, представляет собой RKKRRQRRR (SEQ ID NO: 43). Применяя порядок расположения D-/L-аминокислот, указанный для SEQ ID NO: 30, можно получать последовательность rKKRrQRRr (SEQ ID NO: 46).
В конкретном варианте осуществления изобретения последовательность-транспортер ингибитора JNK, предлагаемого в настоящем изобретении, может содержать по меньшей мере одну последовательность, представленную в rXXXrXXXr (SEQ ID NO: 31), в которой:
r обозначает D-энантиомерный аргинин;
Х обозначает любую L-аминокислоту (включая глицин);
и в которой Х каждый можно выбирать индивидуально и независимо от любого другого Х в SEQ ID NO: 31. Предпочтительно по меньшей мере 4 из 6 X, представляющих собой L-аминокислоты, в SEQ ID NO: 31 обозначают К или R. В другом варианте осуществления изобретения ингибитор JNK, предлагаемый в настоящем изобретении, содержит последовательность-транспортер rX1X2X3rX4X5X6r (SEQ ID NO: 32), в которой X1 обозначает К, Х2 обозначает К, Х3 обозначает R и Х4, Х5 и Х6 обозначают любую L-аминокислоту (включая глицин), выбранную независимо друг от друга. Аналогично этому, последовательность-транспортер ингибитора JNK, предлагаемого в настоящем изобретении, может содержать последовательность rX1X2X3rX4X5X6r (SEQ ID NO: 33), в которой Х4 обозначает Q, Х5 обозначает R, Х6 обозначает R и Х1, Х2 и Х3 обозначают любую L-аминокислоту (включая глицин), выбранную независимо друг от друга. Предлагаемый в изобретении ингибитор JNK может содержать также последовательность rX1X2X3rX4X5X6r (SEQ ID NO: 34), в которой один, два, три, четыре, пять или шесть аминокислотных остатков Х выбраны из группы, в которой: X1 обозначает К, Х2 обозначает К, Х3 обозначает R, Х4 обозначает Q, Х5 обозначает R, Х6 обозначает R, а остальные аминокислотные остатки X, которые не выбраны из указанной выше группы, могут представлять собой любую L-аминокислоту (включая глицин), и их каждый выбирают независимо друг от друга. При этом X1 предпочтительно обозначает Y и/или Х4 предпочтительно обозначает К или R.
Примеры последовательностей-транспортеров, предназначенных для применения в молекуле ингибитора JNK, предлагаемой в изобретении, можно выбирать (но, не ограничиваясь только ими) из последовательностей, представленных ниже в таблице 2 (SEQ ID NO: 31-170), или из любых их фрагментов или вариантов, или химически модифицированных производных (которые предпочтительно сохраняют функциональную способность обеспечивать транслокацию через биологическую мембрану).
Как отмечалось выше, последовательности-транспортеры можно выбирать также из фрагментов или вариантов последовательностей, указанных выше в таблице 2 (при условии, что такой фрагмент или вариант предпочтительно обладает функциональной способностью обеспечивать транслокацию через биологические мембраны). В этом конкретном контексте варианты и/или фрагменты указанных последовательностей-транспортеров предпочтительно содержат пептидную последовательность, которая идентична по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80% или по меньшей мере на 85%, предпочтительно по меньшей мере на 90%, более предпочтительно по меньшей мере на 95% и наиболее предпочтительно по меньшей мере на 99% полноразмерной последовательности указанной последовательности-транспортера, представленной в таблице 2. В этом конкретном контексте под «фрагментом» последовательности-транспортера, указанной выше в таблице 2, предпочтительно подразумевается ее укороченная последовательность, т.е. аминокислотная последовательность, которая на N-конце, С-конце и/или внутри последовательности укорочена по сравнению с аминокислотной последовательностью исходной последовательности.
Кроме того, под «вариантом» последовательности-транспортера или ее фрагмента, как они определены выше, предпочтительно подразумевается последовательность, где аминокислотная последовательность варианта отличается от исходной последовательности-транспортера или ее фрагмента, указанных в настоящем описании, одной или несколькими мутацией(ями), такой(ими) как одна или несколько замен (или при необходимости инсерций и/или делеций) аминокислот. Предпочтительно варианты последовательности-транспортера, указанной выше в настоящем описании, обладают такой же биологической функцией или специфической активностью, что и соответствующая исходная последовательность, т.е. способностью осуществлять транспорт, например, в клетки или ядра. В этом контексте вариант последовательности-транспортера, указанной выше в настоящем описании, может включать примерно 1-50, 1-20, еще более предпочтительно 1-10 и наиболее предпочтительно 1-5, 4, 3, 2 или 1 аминокислотное(их) изменение(ий). Варианты последовательности-транспортера, указанной выше в настоящем описании, могут предпочтительно содержать консервативные аминокислотные замены. Концепция консервативных аминокислотных замен известна в данной области и уже описана выше для ингибирующей JNK (поли)пептидной последовательности и, ее соответственно можно использовать и в этом случае.
Длина последовательности-транспортера, которая включена в ингибитор JNK, предлагаемый в настоящем изобретении, может варьироваться. Согласно некоторым вариантам осуществления изобретения последовательность-транспортер ингибитора JNK, предлагаемого в настоящем изобретении, состоит менее чем из 150, менее чем из 140, менее чем из 130, менее чем из 120, менее чем из 110, менее чем из 100, менее чем из 90, менее чем из 80, менее чем из 70, менее чем из 60, менее чем из 50, менее чем из 40, менее чем из 30, менее чем из 20 и/или менее чем из 10 аминокислот.
Специалист в данной области легко может определить, обладает ли функциональной активностью конкретная последовательность-транспортер в контексте ингибитора JNK, предлагаемого в настоящем изобретении. Например, ингибитор JNK, который содержит домен-транспортер, можно сливать с меткой, например, с флуоресцентным белком, таким как GFP, радиоактивной меткой, ферментом, флуорофором, эпитопом и т.д., которую легко выявлять в клетке. Затем ингибитором JNK, содержащим последовательность-транспортер и метку, трансфектируют клетку или добавляют его в клеточный супернатант, и таким путем можно оценивать проницаемость клеточных мембран с помощью стандартных биофизических и биохимических методов (например, с помощью проточной цитометрии, (иммуно)флуоресцентной микроскопии и т.д.).
Конкретные примеры ингибиторов JNK, предлагаемых в настоящем изобретении, которые содержат последовательность-транспортер, представлены в таблице 3:
Как отмечалось выше, в конкретных вариантах осуществления настоящего изобретения последовательность-транспортер и ингибирующая (поли)пептидная последовательность могут перекрываться. Другими словами, N-конец последовательности-транспортера может перекрываться с С-концом ингибирующей (поли)пептидной последовательности или С-конец последовательности-транспортера может перекрываться с N-концом ингибирующей (поли)пептидной последовательности. Последний вариант осуществления изобретения является наиболее предпочтительным. Предпочтительно последовательность-транспортер перекрывается на одном, двух или трех аминокислотных остатках с ингибирующей (поли)пептидной последовательностью. При таком сценарии конкретная последовательность-транспортер может перекрываться последовательностью, представленной в SEQ ID NO: 1, или ее соответствующими вариантами в положении 1 (X1), положениях 1 и 2 (X1, Х2), положениях 1, 2 и 3 (X1, Х2, Х3).
SEQ ID NO: 174, 175, 178, 179, 180, 181, 182, 183, 184, 188, 189 и 190 представляют собой репрезентативные примеры ингибиторов JNK, предлагаемых в настоящем изобретении, в которых последовательность-транспортер и ингибирующая (поли)пептидная последовательность перекрываются, например, rKKRrQRRrRPTTLNLf (SEQ ID NO: 174) представляет собой последовательность, образованную в результате перекрывания последовательности, представленной в SEQ ID NO: 46 (подчеркнуто), с последовательностью, представленной в SEQ ID NO: 11 (курсив).
В целом, ингибитор JNK, предлагаемый в настоящем изобретении, можно выбирать также из числа ингибиторов JNK, которые представляют собой вариант любого из ингибиторов JNK, которые имеют последовательность, представленную в SEQ ID NO: 171-190. Предпочтительно последовательность указанного варианта идентична по меньшей мере на 50%, более предпочтительно по меньшей мере на 55%, более предпочтительно по меньшей мере на 60%, более предпочтительно по меньшей мере на 65%, более предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 75%, более предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90%, наиболее предпочтительно по меньшей мере на 95% последовательности, представленной в SEQ ID NO: 171-190, прежде всего в SEQ ID NO: 172,
при условии, что касательно ингибирующей (поли)пептидной последовательности, которая входит в SEQ ID NO: 171-190 (см. в качестве эталона ингибирующую (поли)пептидную последовательность, представленную в SEQ ID NO: 1, и конкретные примеры последовательностей, представленных в SEQ ID NO: 2-27)), в такой обладающей указанной степенью идентичности последовательности
а) сохраняется остаток L-аргинина (R) в положении 4 в ингибирующей (поли)пептидной последовательности,
б) сохраняются два остатка L-лейцина (L) в положениях 8 и 10 (положения 7 и 9 в SEQ ID NO: 25-27) в ингибирующей (поли)пептидной последовательности,
в) присутствует(ют) по меньшей мере одна, две, три, четыре, пять или шесть D-аминокислота(от) в указанных положениях, соответствующих аминокислотам, выбранным из группы, включающей X1, Х2, Х3, Х5, Х7 и Х8 в SEQ ID NO: 1, и в указанных положениях в SEQ ID NO: 2-27, более предпочтительно присутствует(ют) по меньшей мере одна, две, три или четыре D-аминокислота(ы) в указанных положениях, соответствующих аминокислотам, выбранным из группы, включающей Х3, Х5, Х7 и Х8 в SEQ ID NO: 1, и в указанных положениях в SEQ ID NO: 2-27, и
г) она все еще ингибирует активность JNK (т.е. представляет собой ингибитор JNK, как он указан в настоящем описании).
В свете указанного определения и также для ясности изложения, в таблице 3 подчеркнуты остатки, которые нельзя изменять в вариантах ингибиторов JNK, содержащих SEQ ID NO: 171-190 (см. подпункты а) и б) в указанном выше определении).
Неидентичные аминокислоты в вариантах ингибиторов JNK, которые содержат последовательности, представленные в SEQ ID NO: 171-190, предпочтительно являются результатом осуществления консервативных аминокислотных замен (см. выше). При этом можно осуществлять также другие возможные замены, указанные выше, в вариантах ингибиторов JNK, которые содержат последовательности, представленные SEQ ID NO: 171-190. Аналогично этому, настоящее изобретение относится также к вариантам любого из ингибиторов JNK, содержащие последовательности, которые представлены SEQ ID NO: 171-190, имеющие отличие от исходной последовательности, которые не присутствуют в ингибирующей (поли)пептидной последовательности или присутствуют не только в ингибирующей (поли)пептидной последовательности, но отличаются присутствием вариантных остатков в последовательности-транспортере. Варианты и фрагменты последовательностей-транспортеров описаны конкретно выше.
Как отмечалось ранее, последовательность-транспортер и ингибирующая JNK (поли)пептидная последовательность ингибиторов JNK, предложенных в настоящем изобретении, не обязательно должны быть непосредственно связаны друг с другом. Они могут быть пространственно отделены, например, с помощью промежуточных (поли)пептидных последовательностей. Предпочтительные промежуточные последовательности, разделяющие ингибирующие (поли)пептидные последовательности и другие (функциональные) последовательности, такие как последовательности-транспортеры, состоят из коротких пептидных последовательностей, которые содержат менее 10 аминокислот, типа гексамера, пентамера, тетрамера, трипептида или даже только дипептида или одного аминокислотного остатка. Наиболее предпочтительная промежуточная последовательность представляет собой одну, две или большее количество копий, таких как два остатка пролина, два остатка глицина, два остатка аргинина и/или два остатка лизина, все либо только в форме L-аминокислот, либо все только в форме D-аминокислот, или в виде смеси D-и L-аминокислот. Естественно, можно применять также другие пептидные спейсерные последовательности.
Наиболее предпочтительный предлагаемый в настоящем изобретении ингибитор JNK содержит последовательность, представленную в SEQ ID NO: 8 (или последовательность, отличающуюся идентичностью последовательности с SEQ ID NO: 8 в том объеме и пределах, которые дополнительно описаны выше) и последовательность-транспортер. Последовательность-транспортер предпочтительно выбирают из любой из SEQ ID NO: 31-170 или их указанных в настоящем описании вариантов, еще более предпочтительно из любой из SEQ ID NO: 31-34 и 46-151. Наиболее предпочтительным вариантом ингибитора JNK, предлагаемого в настоящем изобретении, является ингибитор JNK, содержащий последовательности, представленные в SEQ ID NO: 8 и SEQ ID NO: 46 (или последовательности, отличающиеся идентичностью последовательности с ними в том объеме и пределах, которые дополнительно описаны выше). Предпочтительным примером является ингибитор JNK, который содержит последовательность, представленную в SEQ ID NO: 172, или ее соответствующие варианты, которые имеют вариации в последовательности-транспортере и/или в ингибирующей (поли)пептидной последовательности, которые указаны в настоящем описании.
Следующим объектом настоящего изобретения является ингибитор JNK, который содержит
а) ингибирующий (поли)пептид, содержащий последовательность, выбранную из группы последовательностей, которая включает RPTTLNLF (SEQ ID NO: 191), KRPTTLNLF (SEQ ID NO: 192), RRPTTLNLF и/или RPKRPTTLNLF (SEQ ID NO: 193), и
б) последовательность-транспортер, предпочтительно последовательность-транспортер, выбранную из группы последовательностей-транспортеров, которые указаны в таблице 2, или их вариантов/фрагментов, еще более предпочтительно последовательность-транспортер, выбранную из последовательностей, представленных в SEQ ID NO: 31-34 и 46-151, или соответствующих их вариантов или фрагментов.
Последовательность-транспортер и ингибирующая (поли)пептидная последовательность могут перекрываться. Согласно этому варианту осуществления изобретения из предпочтительных последовательностей-транспортеров следует упомянуть прежде всего последовательность-транспортер, которая представлена в SEQ ID NO: 46, предпочтительно сцепленную (например, непосредственно) с N-концом ингибирующей (поли)пептидной последовательности.
Ингибитор JNK, предлагаемый в настоящем изобретении, может представлять собой также ингибитор JNK, который содержит или состоит из последовательности GRKKRRQRRRPPKRPTTLNLFPQVPRSQD (SEQ ID NO: 194) или последовательности GRKKRRQRRRPTTLNLFPQVPRSQD (SEQ ID NO: 195).
Следующим объектом настоящего изобретения является (поли)пептид, который содержит последовательность-транспортер, выбранную из группы последовательностей, которая включает rKKRrQRr (SEQ ID NO: 148), rKKRrQRrK (SEQ ID NO: 149) и/или rKKRrQRrR (SEQ ID NO: 150).
В контексте настоящего описания фраза «содержит определенную последовательность или определенную SEQ ID NO:», как правило, означает, что присутствует (по меньшей мере) одна копия указанной последовательности, например, в молекуле ингибитора JNK. Например, одной ингибирующей (поли)пептидной последовательности, как правило, достаточно для достижения требуемого ингибирования активности JNK. Однако под объем изобретения естественно подпадает также применение двух или большего количества копий соответствующей последовательности (например, две или большее количество копий ингибирующей (поли)пептидной последовательности различных типов и/или одного и того же типа и/или две или большее количество копий последовательности-транспортера различных типов и/или одного и того же типа), которые можно использовать, если при, в целом, этом не аннулируется способность полученной молекулы ингибировать активность JNK (т.е. соответствующая молекула все еще представляет собой ингибитор JNK, указанный в настоящем описании).
Предлагаемые в изобретении ингибиторы JNK можно получать или создавать с помощью хорошо известных в данной области методов, например, с помощью химического синтеза, включающего твердофазный пептидный синтез с использованием стратегии, основанной на применении групп Fmoc (9-флуоренилметилоксикарбонил), т.е. путем осуществления последовательных циклов удаления защитной группы Fmoc и циклов сочетания Fmoc-аминокислота. Услуги по осуществлению такого пептидного синтеза предоставляются многими компаниями, такими, например, как компания PolyPeptide (Страсбург, Франция).
Антитела
Следующим объектом настоящего изобретения является получение антител к ингибиторам JNK, предлагаемым в настоящем изобретении, т.е. способ получения антител, распознающих ингибиторы JNK, предлагаемые в настоящем изобретении. Методы получения антител очень хорошо известны в данной области.
Таким образом, настоящее изобретение относится также к способу иммунизации животного кроме человека ингибитором JNK, предлагаемым в настоящем изобретении, где способ включает следующую стадию, на которой:
- обрабатывают (иммунизируют) животное кроме человека, в котором можно получать антитела,
в частности, млекопитающее кроме человека,
более предпочтительно такое животное, как коза и грызуны, например, мышь, крыса и кролик,
ингибитором JNK, предлагаемым в настоящем изобретении,
более предпочтительно ингибитором JNK, который содержит или состоит из (поли)пептида, имеющего последовательность, выбранную из любой из последовательностей, представленных в SEQ ID NO: 1-27.
В контексте настоящего описания подразумевается, что «иммунизация» не является терапевтической по своей природе, поскольку ингибиторы JNK, предлагаемые в настоящем изобретении, не являются патогенами (т.е. в данном случае отсутствует потребность в терапии).
Настоящее изобретение относится также к способу получения (поликлонального) антитела, распознающего ингибитор JNK, предлагаемый в настоящем изобретении, где способ включает следующую стадию, на которой:
- выделяют из животного кроме человека, в котором можно получать антитела,
в частности, из млекопитающего кроме человека,
более предпочтительно из такого животного, как коза и грызуны,
например, мышь, крыса и кролик,
которое ранее было обработано (иммунизировано) ингибитором JNK, предлагаемым в настоящем изобретении,
более предпочтительно ингибитором JNK, который содержит или
состоит из (поли)пептида, имеющего последовательность, выбранную
из любой из последовательностей, представленных в SEQ ID NO: 1-27, (поликлональное) антитело, распознающее указанный ингибитор JNK.
Настоящее изобретение относится также к способу выделения клетки, продуцирующей антитело, которое распознает ингибитор JNK, предлагаемый в настоящем изобретении, где способ включает следующую стадию, на которой:
- выделяют из животного кроме человека, в котором можно получать антитела,
в частности, млекопитающего кроме человека,
более предпочтительно такого животного, как коза и грызуны,
например, мышь, крыса и кролик,
которое ранее было обработано (иммунизировано) ингибитором JNK, предлагаемым в настоящем изобретении,
более предпочтительно ингибитором JNK, который содержит или
состоит из (поли)пептида, имеющего последовательность, выбранную
из любой из последовательностей, представленных в SEQ ID NO: 1-27, клетку, продуцирующую указанное антитело, которое распознает указанный ингибитор JNK, и
необязательно иммортализуют указанную клетку.
Настоящее изобретение относится также к способу получения (моноклонального) антитела, распознающего ингибитор JNK, предлагаемый в настоящем изобретении, где способ включает следующую стадию, на которой:
- выделяют антитело, которое распознает ингибитор JNK, предлагаемый в настоящем изобретении,
более предпочтительно которое распознает ингибитор JNK, состоящий
из (поли)пептида, который имеет последовательность, выбранную из
любой из последовательностей, представленных в SEQ ID NO: 1-27,
из супернатанта клеточной культуры клетки, продуцирующей указанное антитело, где клетка необязательно является иммортализованной.
Специалисту в данной области должно быть очевидно, что способ иммунизации животного кроме человека и способ получения (поликлонального) антитела, представленный в настоящем описании, следует осуществлять последовательно. Аналогично этому, способ иммунизации животного кроме человека, способ выделения клетки, продуцирующей антитело, и способ получения (моноклонального) антитела можно объединять.
Следующим объектом настоящего изобретения является антитело, которое можно получать (и/или полученное) с помощью способов, предлагаемых в настоящем изобретении, получения поликлонального или моноклонального антитела, где антитело распознает по меньшей мере один (поли)пептид, который содержит или состоит из последовательности, выбранной из любой из последовательностей, представленных в SEQ ID NO: 1-27, но предпочтительно не распознает (или по меньшей мере распознает в значительно пониженной например, по меньшей мере на один порядок, степени,) практически такой же (поли)пептид, в котором L-аминокислоты присутствуют вместо D-аминокислот в соответствующем участке последовательности, представленной в SEQ ID NO: 1-27. Предпочтительно указанное антитело может распознавать ингибитор JNK, предлагаемый в настоящем изобретении, но не может распознавать (или может распознавать по меньшей мере в значительно пониженной, например, по меньшей мере на один порядок, степени) (поли)пептид, который содержит последовательность RPKRPTTLNLF (SEQ ID NO: 193)). Наиболее предпочтительное антитело (моноклональное или поликлональное) может распознавать ингибитор JNK, который содержит последовательность SEQ ID NO: 8 (например, ингибитор JNK, содержащий последовательность SEQ ID NO: 172), но не может распознавать (или может распознавать по меньшей мере в значительно пониженной, например, по меньшей мере на один порядок, степени) (поли)пептид, который содержит очень сходную последовательность, в которой L-аминокислоты присутствуют вместо D-аминокислот.
Наиболее предпочтительными являются указанные поликлональные или моноклональные антитела, которые распознают (поли)пептид, содержащий SEQ ID NO: 172, но не распознают (или распознают по меньшей мере в значительно пониженной, например, по меньшей мере на один порядок, степени) (поли)пептид, содержащий последовательность RKKRRQRRRRPKRPATLNLF (SEQ ID NO: 199).
Настоящее изобретение относится также к клетке, выделенной с помощью описанного выше конкретного способа выделения клетки, продуцирующей антитело, которое распознает ингибитор JNK, предлагаемый в настоящем изобретении, где клетка продуцирует антитело, которое предпочтительно распознает по меньшей мере один (поли)пептид, выбранный из любой из последовательностей, представленных в SEQ ID NO: 1-27, но не распознает практически такой же (поли)пептид, в котором L-аминокислоты присутствуют вместо D-аминокислот в последовательности, соответствующей SEQ ID NO: 1, (например, может распознавать (поли)пептид, который содержит последовательность RPkRPaTLNLf (SEQ ID NO: 8), но не может распознавать (или по меньшей мере может распознавать в значительно пониженной, например, по меньшей мере на один порядок, степени) (поли)пептид, который содержит последовательность RPKRPTTLNLF (SEQ ID NO: 193).
Настоящее изобретение относится также к созданию антител к конкретным последовательностям-транспортерам, что позволяет осуществлять идентификацию, например, ингибиторов JNK, представленных в таблице 3. Таким образом, все аспекты (моноклональные или поликлональные антитела; способы их создания, продуцирующие их клетки и т.д.), обсужденные выше для антител, распознающих ингибитор JNK, предлагаемый в настоящем изобретении (в, частности, по меньшей мере один (поли)пептид, содержащий или состоящий из последовательности, выбранной из любой из последовательностей, представленных в SEQ ID NO: 1-27), можно использовать также в контексте (поли)пептида, который содержит или состоит из последовательности, выбранной из любой из последовательностей, представленных в SEQ ID NO: 31-34 и 46-151. Естественно, референс-последовательность, которая не распознается (или распознается по меньшей мере в значительно пониженной, например, по меньшей мере на один порядок, степени) в этом контексте и в этом случае представляет собой практически сходную последовательность, однако в которой присутствуют L-аминокислоты вместо D-аминокислот в соответствующем участке последовательности-транспортера.
Методы тестирования аффинности связывания (моноклональных и/или поликлональных) антител хорошо известны в данной области. Одним из путей, среди прочего, является получение характеристик аффинности связывания антитела с помощью сэндвич-ELISA с использованием пептида-мишени, а также отрицательных контролей (например, такого же пептида, состоящего только из L-аминокислот). Предел чувствительности ELISA можно рассчитывать (но, не ограничиваясь только указанным методом) с помощью контрольных повторностей следующим образом:
предел чувствительности ELISA=среднее значение (отрицательный контроль)+(3 × стандартное отклонение для отрицательного контроля).
Если в образце полученное значение ниже или равно пределу чувствительности ELISA, то тестируемое антитело может рассматриваться как не обладающее аффинностью к пептиду-мишени. Если в образце полученное значение превышает предел чувствительности ELISA, то тестируемое антитело может рассматриваться как обладающее аффинностью к пептиду-мишени. Кроме того, чем вышеуказанное значение, тем больше аффинность тестируемого антитела к мишени.
Коммерческие услуги по получению моноклональных или поликлональных антител предоставляются, например, такой компанией, как Eurogentec (Серен, Бельгия).
Все процитированные ссылки включены в настоящее описание в качестве ссылки.
Примеры
Ниже представлены конкретные примеры, иллюстрирующие различные варианты осуществления и объекты изобретения. Однако объем настоящего изобретения не ограничен представленными в настоящем описании конкретными вариантами его осуществления. Так, специалистам в данной области, исходя из представленного выше описания, прилагаемых чертежей и приведенных ниже примеров, должны быть очевидны различные модификации изобретения помимо указанных в настоящем описании. Все такие модификации подпадают под объем прилагаемой формулы изобретения.
Пример 1: Синтез ингибитора JNK, последовательность которого представлена в SEQ ID NO: 172
В качестве приведенного с целью иллюстрации примера ниже представлен синтез ингибитора JNK, последовательность которого представлена в SEQ ID NO: 172. Специалисту в данной области хорошо известно, что указанный синтез можно также применять и легко адаптировать для синтеза любого другого ингибитора JNK, предлагаемого в настоящем изобретении.
Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, получали с помощью твердофазного пептидного синтеза на основе стратегии, предусматривающей применение Fmoc (9-флуоренилметилоксикарбонил). Линкер между пептидом и смолой представлял собой амидный линкер Ринка (пара-[Fmoc-2,3-диметоксибензил]феноксиуксусная кислота). Пептид синтезировали путем осуществления последовательных циклов удаления защитной группы Fmoc и сочетания Fmoc-аминокислота. После завершения синтеза полный пептид отщепляли с помощью трифторуксусной кислоты (ТФК), непосредственно получая неочищенный С-концевой амид, который затем очищали с помощью препаративной ЖХВР с обращенной фазой. Очищенные фракции объединяли, получая гомогенную партию, которую обрабатывали с помощью ионообменной хроматографии, получая ацетат. Затем пептид сушили сушкой вымораживанием.
1.1. Твердофазный синтез пептида
Если не указано иное, то процесс получения осуществляли при комнатной температуре (22°С±7°С) в среде с фильтрацией воздуха. Масштаб синтеза был следующим: применяли 0,7 ммоля исходной аминокислоты на смоле, при этом ожидаемый выход очищенного пептида составлял примерно 1 г. Синтез осуществляли вручную в реакторе объемом 30-50 мл, снабженном фриттированным диском, при механическом перемешивании и/или барботировании азотом.
1.2 Приготовление смолы
Пара-метилбензгидриламидную смолу (МВНА-смола) сначала промывали смесью дихлорметан/диметилформамид/диизопропилэтиламин в атмосфере азота. Затем промытую смолу сшивали с амидным линкером Ринка (пара-[Fmoc-2,3-диметоксибензил]феноксиуксусная кислота) в смеси: РуВОВ (гексафторфосфат бензотриазол-1-илокситриспирролидинофосфония)/диизопропилэтиламин/1-гидроксибензотриазол, получая Fmoc-амид Ринка-МВНА-смолу.
1.3 Сочетание аминокислот
Аминокислоты сочетали со смолой, используя следующий цикл:
Осуществляли удаление защитной группы у Fmoc-амид Ринка-МВНА-смолы, осуществляя отмывку в 35 об.% пиперидина/диметилформамида, с последующей обработкой диметилформамидом. Реакция удаления защитной группы протекала в течение примерно 16 мин. Защищенные с помощью Fmoc аминокислоты (например, 2 экв. аминокислоты и HOBt (1-гидроксибензотриазол) в диметилформамиде/дихлорметане (50/50) добавляли к смоле, после чего добавляли 2 экв. агента для сочетания, такого как диизопропилкарбодиимид (DIC). Реакция сочетания протекала в течение периода времени, составлявшего от 1 ч до целой ночи, в зависимости от реакционной способности соответствующей добавляемой аминокислоты. Применяли объемы из расчета 0,5 мл/100 мг конъюгата пептид-смола и корректировали их после каждого цикла. После завершения реакции сочетания смолу отмывали трижды с помощью ДМФ. Завершение реакции сочетания оценивали с помощью нингидринового теста (или теста 1 по Кайзеру) для первичных аминов и хлоранилового теста 2 для вторичных аминов. В некоторых случаях хлораниловый тест можно объединять с нингидриновым тестом для более надежного контроля качества. В том случае, когда тест свидетельствовал о неполном завершении реакции сочетания, сочетание повторяли с использованием меньшего избытка (0,5-1 экв.) аминокислоты, PYBOP, HOBT в диметилформамиде/дихлорметане и диизопропилэтиламина. Функциональность смолы оценивали, и она составляла, как правило, 0,6-0,2 мэкв./г в зависимости от исходной загрузки смолы. После сочетания последней аминокислоты, как правило, осуществляли удаление защитной группы у пептида-смолы и затем отмывали 5 раз в ДХМ перед сушкой в печи в вакууме при 30°С. После сушки конъюгата пептид-смола рассчитывали выход твердофазного синтеза в виде соотношения увеличения массы конъюгата пептид-смола по сравнению с теоретическим увеличением массы, рассчитанным, исходя из начальной загрузки смолы. Выход может быть близким к 100%.
1.4 Расщепление и удаление защитной группы
Пептид отщепляли от смолы в смеси, содержащей трифторуксусную кислоту/1,2-этандитиол/тиоанизол/воду/фенол (88/2,2/4,4/4,4/7 об/об), которую обозначают также как ТФК/К-реагент, в течение 4 ч при комнатной температуре. Реакционный объем составлял 1 мл/100 мг пептидной смолы. В процессе добавления смолы к реагенту температуру смеси регулировали, поддерживая на уровне ниже 30°С.
1.5 Экстракция пептида из смолы
Пептид экстрагировали из смолы путем фильтрации через фриттированный диск. После концентрирования на роторном испарителе до 1/3 объема пептид осаждали с помощью холодного простого метил-трет-бутилового эфира и фильтровали. Затем неочищенный пептид сушили в вакууме при 30°С.
1.6 Очистка с помощью препаративной ЖХВР
Затем неочищенный пептид очищали с помощью ЖХВР с обращенной фазой до чистоты ≥95%. Очищенные фракции концентрировали на роторном испарителе и подвергали сушке вымораживанием.
1.7 Ионообменная хроматография
Концентрированные высушенные вымораживанием пулы очищенного пептида, который имел последовательность, представленную в SEQ ID NO: 172, растворяли в воде и очищали с помощью ионообменной хроматографии на смоле Dowex в ацетатной форме, 50-100 меш.
Для синтеза применяли следующие исходные реагенты:
Аналогичным методом можно получать другие ингибиторы JNK, предлагаемые в настоящем изобретении.
Пример 2: Ингибирующее действие отобранных ингибиторов JNK, предлагаемых в настоящем изобретении
Ниже изложена стандартная процедура оценки ингибирующего действия ингибиторов JNK, предлагаемых в настоящем изобретении. Метод позволяет осуществлять оценку in vitro с использованием стандартизованного нерадиоактивного анализа способности соединения-кандидата снижать фосфорилирование специфического для JNK субстрата c-Jun. Кроме того, проиллюстрирован метод определения ингибирующего действия (IC50) в отношении JNK и величины Ki выбранного соединения. Этот метод можно применять для решения вопроса о том, ингибирует или не ингибирует соединение-кандидат активность JNK, и специалисту в данной области должно быть очевидно, каким образом можно адаптировать приведенные ниже методы для его конкретных целей и требований. 2.1 Материалы Реагенты и планшет для AlphaScreen-анализа:
- His-JNKI (ref 14-327, фирма Upstate, 10 мкг в 100 мкл: концентрация: 2,2 мкМ), конечная концентрация 5 нМ,
- His-JNK2 (ref 14-329, фирма Upstate, 10 мкг в 100 мкл: концентрация: 2 мкМ), конечная концентрация 5 нМ,
- His-JNK3 (ref 14-501, фирма Upstate, 10 мкг в 100 мкл: концентрация: 1,88 мкМ), конечная концентрация 5 нМ,
- антитело к фосфо-cJun (ref 06-828, фирма Upstate, партия DAM1503356, концентрация: 44,5 мкМ), конечная концентрация 10 нМ,
- биотин-cJun (29-67):
последовательность: биотин-SNPKILKQSMTLNLADPVGSLKPHLRAKNSDLLTSPDVG (SEQ ID NO: 198), партия 100509 (молекулярная масса 4382,11, чистота 99,28%), растворенный в Н2О, концентрация: 10 мМ) конечная концентрация 30 нМ,
- АТФ (ref AS001A, фирма Invitrogen, партия 50860 В, концентрация 100 мМ)) конечная концентрация 5 мкМ,
- SAD-гранулы (ref 6760617М, фирма PerkinElmer, партия 540-460-А, концентрация 5 мг/мл) конечная концентрация 20 мкг/мл,
- AprotA-гранулы (ref 6760617М, фирма PerkinElmer, партия 540-460-А, концентрация 5 мг/мл), конечная концентрация 20 мкг/мл,
- 384-луночный белый планшет типа Optiplate (ref 6007299, фирма PerkinElmer, партия 654280/2008),
- 96-луночный планшет для разведения пептидов (ref 82.1581, фирма Sarstedt),
- TopSeals-A (ref 6005185, фирма Perkin Elmer, партия 65673),
- устройство для определения биолюминисцентного переноса энергии:
- биолюминисцентный перенос энергии определяли с помощью планшет-ридера Fusion Alpha (фирма Perkin Elmer).
Пипетки:
- Электронную EDP3-пипетку 20-300 (Ref 17007243; фирма Rainin) применяли для внесения в планшет смеси фермент-антитело, смеси субстрат-АТФ и гранул.
- Многоканальное устройство типа PIPETMAN® Ultra 8×20 (Ref 21040; фирма Gilson) применяли для внесения в планшет соединений-ингибиторов.
Буферы и растворы
- Буфер для киназы: 20 мМ Трис-основание, рН 7,4, 10 мМ MgCl2, 1 мМ ДТТ, 100 мкМ Na3VO4, 0,01% Твин, (1% ДМСО),
- Стоп-буфер: 20 мМ Трис-основание, рН 7,4, 200 мМ NaCl, 80 мМ ЭДТК-К (значение рН доведено до 8 с использованием КОН вместо NaOH), 0,3% БСА,
- Буфер для разведения киназы JNK: 50 мМ Трис-основание, рН 7,4, 150 мМ NaCl, 0,1 мМ ЭГТК, 0,03% Brij-35, 270 мМ сахароза, 0,1% β-меркаптоэтанола.
2.2 Метод
Для оценки ингибирующего действия пептидов осуществляли стандартный AlphaScreen-анализ (см., например, Guenat и др., J Biomol Screen, 11, 2006, cc.1015-1026). Приготавливали различные компоненты и последовательно смешивали согласно представленной рекомендации. Планшеты запечатывали и инкубировали следующим образом:
Для того, чтобы избежать загрязнения, смеси добавляли с помощью пипетки в различные углы лунки. После заполнения планшета каждой смесью планшет встряхивали (держа одну сторону и постукивая противоположной стороной по столу), давая смеси стекать по стенкам лунок.
Биолюминисцентный перенос энергии определяли с помощью планшет-ридера Fusion Alpha (фирма Perkin Elmer).
Все компоненты обязательно тестировали по меньшей мере в трех повторностях в трех независимых экспериментах для каждой изоформы JNK. Приемлемыми концентрациями соединений, подлежащих тестированию, были следующие: 0, 0,03 нМ, 0,1 нМ, 0,3 нМ, 1 нМ, 3 нМ, 10 нМ, 30 нМ, 100 нМ, 300 нМ, 1 мкМ, 3 мкМ, 10 мкМ, 30 мкМ и 100 мкМ. В качестве контролей использовали либо образцы без JNK, либо образцы без субстрата (c-Jun).
Приготовления смесей
JNK1, JNK2 и JNK3, 5 нМ,
биотин-cJun, 30 нМ,
АТФ, 5 мкМ; антитело к фосфо-cJun (S63), 10 нМ,
гранулы SAD/AprotA, 20 мкг/мл.
Конечная концентрация антитела = 10 нм (антитела к фосфо-cJun (S63)).
Применяемая для определения часть: [смесь]×5 (5 мкл в конечном объеме 25 мкл).
Концентрация маточного раствора = 44,5 мкМ (ref 06-828, фирма Upstate, партия DAM1503356)
10 нМ → 50 нМ в буфере для киназ.
Конечная концентрация JNK1, JNK2 и JNK3=5 нМ.
Применяемая для реакции часть: [смесь]×3 (5 мкл в конечном объеме 15 мкл).
Концентрация маточного раствора=2,2 мкМ для JNK1 (ref 14-327, фирма Upstate, партия D7KN022CU),
2,0 мкМ для JNK2 (ref 14-329, фирма Upstate, партия 33221 CU),
1,88 мкМ для JNK3 (ref 14-501, фирма Upstate, партия D7CN041CU).
5 нМ → 15 нМ в буфере для антитела.
Ингибитор:
Применяемая для реакции часть: [смесь]×3 (5 мкл в конечном объеме 15 мкл).
Концентрация маточного раствора = 10 мМ.
100 мкм → 300 мкМ в буфере для киназ,
30 мкМ → 90 мкМ в буфере для киназ,
10 мкМ → 30 мкМ в буфере для киназ,
0,03 нМ → 0,09 нМ в буфере для киназ,
и 0 нМ → буфер для киназ.
Приготавливали две серии 10-кратных серийных разведении в 96-луночном планшете, первую серию начинали с концентрации 300 мкМ и доводили до 0 нМ, вторую серию начинали с 90 мкМ и доводили до 0,03 нМ. Пептиды добавляли в 384-луночные планшеты с помощью 8-канальной мультипипетки (ref F 14401, фирма Gilson, 8×20).
Конечная концентрация АТФ=5 мкМ.
Применяемая для реакции часть: [смесь]×3 (5 мкл в конечном объеме 15 мкл).
Концентрация маточного раствора = 100 мМ (ref AS001A, фирма Invitrogen, партия 50860 В).
5 мкМ → 15 мкМ в буфере для киназ.
Конечная концентрация биотин-c-Jun = 30 нМ.
Применяемая для реакции часть: [смесь]×3 (5 мкл в конечном объеме 15 мкл).
Концентрация маточного раствора = 10 мМ.
30 нМ → 30 нМ в АТФ-буфере.
Гранулы: конечная концентрация SAD/A ProtA=20 мкг/мл (светочувствительные).
Применяемая для определения часть: [смесь]×2,5 (10 мкл в конечном объеме 25 мкл).
Концентрация маточного раствора = 5 мг/мл → 20 мкг/мл 50 мкг/мл в стон-буфере.
Перемешивание в темной комнате (зеленый свет) или в темноте.
Анализ кривых для расчета величины IC50:
Анализ осуществляли с использованием программы GraphPad Prism4 с помощью следующего уравнения:
Сигмовидная зависимость доза-ответ (без ограничений).
Y = минимальное значение + (максимальное значение-минимальное значение)/(1+10((LogEC50-X)))
Выпадающие значения исключали с помощью теста Граббса.
Сравнение значений IC50 :
Анализ осуществляли с помощью программы GraphPad с использованием следующих критериев: однонаправленный дисперсионный анализ с последующим использованием критерия Тьюки для множественных сравнений. Результат рассматривали как статистически значимый при Р<0,05.
Величины Km АТФ для JNK и Km пептида, специфического в отношении биотин-cJun, определяли с помощью AlphaScreen-анализа с использованием стандартизации результатов.
Для расчета величин Ki можно применять следующее математическое соотношение между величинами Ki и IC50: Ki=IC50/(1+(концентрация субстрата/Km субстрата).
Пример 3: Эксперименты по интернализации и анализ полученных результатов
3.1 Материалы и методы, применяемые при проведении экспериментов по анализу поглощения
а) Клеточная линия:
Применяемая в этом эксперименте клеточная линия представляла собой HL-60 (Ref CCL-240, АТСС, партия 116523)
б) Культуральная среда и планшеты
Среда RPMI (Ref 21875-091, фирма Invitrogen, партия 8296) или DMEM (Ref 41965, фирма Invitrogen, партия 13481), дополненная 05.05.2008 г. следующими компонентами:
10% FBS (Ref A64906-0098, фирма РАА, партия А 15-151): декомплементированная при 56°С в течение 30 мин 04.04.2008 г.;
1 мМ пируват натрия (Ref S8636, фирма Sigma, партия 56К2386);
пенициллин (100 ед./мл)/стрептомицин (100 мкг/мл) (Ref P4333, фирма Sigma, партия 106К2321).
ЗФР 10×(Ref 70011, фирма Invitrogen, партия 8277): разведенный до 1 × стерильной Н2О.
Трипсин-0,05% ЭДТК (Ref L-11660, фирма РАА, партия L66007-1194).
6-луночные культуральные планшеты (Ref 140675, фирма Nunc, партия 102613),
24-луночные культуральные планшеты (Ref 142475, фирма Nunc, партия 095849),
96-луночные культуральные планшеты (Ref 167008, фирма Nunc, партия 083310),
96-луночные планшеты для дозирования белков (Ref 82.1581, фирма Sarstedt),
96-луночные планшеты для оценки флуоресценции (Ref 6005279, фирма Perkin Elmer).
в) Растворы
Поли-D-лизиновый раствор для сенсибилизации (нанесения покрытия) (фирма Sigma P9011, партия 095К5104): концентрация 25 мкг/мл, конечное разведение в 1×ЗФР.
Кислый буфер для отмывки: 0,2М глицин, 0,15М NaCl, pH 3,0.
Буфер для лизиса Ripa: 10 мМ NaH2PO4, pH 7,2, 150 мМ NaCl, 1% Тритон X-100, 1 мМ ЭДТК, pH 8,0, 200 мкМ Na3VO2, 0,1% ДСН, 1 × коктейль ингибиторов протеаз (Ref 11873580001, фирма Roche, партия 13732700).
г) Микроскопия и анализ с помощью флуоресцентного планшет-ридера
Выявление и подсчет клеток осуществляли с помощью инвертированного микроскопа (Axiovert 40 CFL; фирма Zeiss; 20×).
Флуоресценцию определяли с помощью планшет-ридера Fusion Alpha (фирма Perkin Elmer).
д) Метод
Интернализацию меченного с помощью ФИТЦ пептида изучали с использованием суспензионной культуры клеток. Клетки высевали в покрытые поли-DL-лизином планшеты в концентрации 1×106 клеток/мл. Затем планшеты инкубировали в течение 24 ч при 37°С, 5% СО2 и относительной влажности 100% перед добавлением пептида в известной концентрации. После добавления пептида клетки инкубировали в течение 30 мин, 1, 6 или 24 ч при 37°С, 5% СО2 и относительной влажности 100%. Затем клетки отмывали дважды кислым буфером (глицин 0,2 М, NaCl 0,15M, рН 3,0) для удаления адсорбированного на клеточной поверхности пептида (см. Kameyama и др., Biopolymers, 88, 2007, cc.98-107). Применяли кислый буфер, поскольку пептиды, богатые основными аминокислотами, сильно адсорбировались на поверхности клеток, что часто приводило к завышенной оценке количества интернализованого пептида. Поэтому для удаления адсорбированных на поверхности клеток пептидов применяли отмывку клеток кислым буфером. При определении поглощения клетками проникающих в клетку пептидных конъюгатов Fab осуществляли кислотную отмывку с последующими двумя отмывками с помощью ЗФР. Клетки расщепляли, добавляя буфер для лизиса RIPA. Относительное количество интернализованного пептида затем определяли на основе флуоресценции после вычитания фонового уровня и стандартизации относительно содержания белка.
Таким образом, осуществляли следующие стадии:
1. культивирование клеток;
2. кислотную отмывку и экстрагирование клеток;
3. анализ интернализации пептидов с помощью флуоресцентного планшет-ридера.
е) Культивирование клеток и обработка пептидами
6-Луночные культуральные планшеты покрывали 3 мл поли-D-Lys (фирма Sigma, P9011; 25 мкг/мл в ЗФР), 24-луночные - 600 мкл, а 96-луночные планшеты - 125 мкл и инкубировали в течение 4 ч при 37°С, 5% СО2 и относительной влажности 100%.
Через 4 ч планшеты дважды отмывали 3,5 мл ЗФР, 700 мкл или 150 мкл ЗФР в случае 6-, 24- или 96-луночных планшетов соответственно.
Клетки высевали в планшеты в 2,4 мл среды (RPMI) с плотностью 1000000 клеток/мл для получения суспензии клеток. После инокуляции планшеты инкубировали при 37°С, 5% СО2 и относительной влажности 100% в течение 24 ч перед добавлением пептида. Прикрепленные (прилипшие) клетки, плотность которых должна составлять 90-95% в день обработки, высевали в среду DMEM:
Клетки обрабатывали взятым в требуемой концентрации меченным с помощью ФИТЦ пептидом (маточный раствор с концентрацией 10 мМ в Н2О).
После добавления пептида клетки инкубировали в течение 0-24 ч (например, 30 мин, 1, 6 или 24 ч) при 37°С, 5% СО2 и относительной влажности 100%.
Кислотная отмывка и получение клеточных экстрактов:
Экстракты охлаждали на льду.
Клетки в суспензии (или клетки, не прикрепившиеся к планшету):
Переносили клетки в пробирку «Falcon 15 мл». Для получения максимального количества клеток отмывали планшет 1 мл ЗФР.
Собирали клетки в течение 2 мин максимум при 2400 об/мин.
Суспендировали клетки в 1 мл холодного ЗФР.
Переносили клетки в сенсибилизированную «пробирку Эппендорфа» (покрытую 1 мл поли-D-Lys, на 4 ч и отмывают дважды 1 мл ЗФР).
Отмывали трижды 1 мл холодного кислого буфера для отмывки и центрифугировали в течение 2 мин при максимальной скорости 2400 об/мин. Применяли меры предосторожности для предупреждения распластывания клеток в «пробирке Эппендорфа».
Отмывали дважды 1 мл холодного ЗФР для нейтрализации.
Добавляли 50 мкл буфера для лизиса RIPA.
Инкубировали в течение 30 мин - 1 ч на льду при перемешивании.
Прикрепившиеся клетки
Отмывали трижды, используя по 3 мл, 1 мл или 200 мкл (для 6-, 24- или 96-луночных планшетов соответственно) холодного кислого буфера для отмывки. Применяли меры предосторожности для предупреждения отлипания клеток от планшета.
Отмывали дважды с помощью 1 мл холодного ЗФР (для 6-, 24- или 96-луночных планшетов соответственно) для нейтрализации.
Добавляли 50 мкл буфера для лизиса RIPA.
Инкубировали в течение 30 мин-1 ч на льду при перемешивании.
Отскабливали клетки с помощью холодного лезвия. 24- и 96-луночные планшеты непосредственно центрифугировали при 4000 об/мин при 4°С в течение 15 мин для удаления клеточного дебриса. Затем супернатанты (100 или 50 мл соответственно для 24- или 96-луночных планшетов) переносили непосредственно в темные 96-луночные планшеты. Планшеты считывали с помощью флуоресцентного планшет-ридера (Fusion Alpha, фирма Perkin Elmer).
Переносили лизат в сенсибилизированную «пробирку Эппендорфа» (которую покрывали 1 мл поли-D-Lys на 4 ч, и отмывали дважды 1 мл ЗФР).
Затем лизированные клетки центрифугировали в течение 30 мин при 10000 g при 4°С для удаления клеточного дебриса.
Удаляли супернатант и хранили при -80°С в сенсибилизированной «пробирке Эппендорфа» (покрытой 1 мл поли-D-Lys, в течение 4 ч и отмывали дважды 1 мл ЗФР).
Анализ интернализации пептидов с помощью флуоресцентного планшет-ридера:
Содержание белка в каждом белковом экстракте определяли с помощью стандартного ВСА-анализа (набор №23225, фирма Pierce) согласно инструкциям производителя.
Определяли относительную флуоресценцию каждого образца после считывания 10 мкл каждого образца с помощью флуоресцентного планшет-ридера (Fusion Alpha, фирма Perkin Elmer), вычитали фоновый уровень и стандартизовали относительно концентрации белка.
3.2 Эксперименты по поглощению
Зависящую от времени интернализацию (поглощение) меченных с помощью ФИТЦ полученных из ТАТ конструкций транспортеров в клетки клеточной линии HL-60 осуществляли с использованием указанных выше методов с использованием пептидных последовательностей-транспортеров, которые представлены в SEQ ID NO: 52-96, 43 и 45-47. Эти последовательности приведены ниже в таблице 4.
В приведенной выше таблице D-аминокислоты обозначены прописной буквой «d» перед соответствующим аминокислотным остатком (например, dR обозначает D-Arg).
Для некоторых последовательностей при использовании первого подхода, к сожалению, не удалось осуществить синтез по техническим причинам. Эти последовательности сокращенно обозначены на фиг.6 как 1, 2, 3, 4, 5, 6, 7, 8, 43, 52, 53, 54, 55, 56, 57, 85, 86, 87, 88, 89 и 90. Однако остальные последовательности применяли в экспериментах по интернализации.
Результаты представлены на фиг.6.
Как продемонстрировано на фиг.6, после 24-часовой инкубации все транспортеры с консенсусной последовательностью rXXXrXXXr (SEQ ID NO: 31) отличались более высокой способностью к интернализации, чем транспортер L-TAT (SEQ ID NO: 43). Клетки линии Hela инкубировали в течение 24 ч в 96-луночном планшете с 10 мМ полученными из r3-L-TAT транспортерами. Затем клетки отмывали дважды кислым буфером (0,2 М глицин, 0,15М NaCl, pH 3,0) и дважды ЗФР. Клетки расщепляли, добавляя буфер для лизиса RIPA. Затем определяли относительное количество интернализованного пептида, считывая интенсивность флуоресценции (ридер Fusion Alpha, фирма Perkin Elmer) для каждого экстракта, с последующим вычитанием фонового уровня.
Как продемонстрировано на фиг.6, одно из положений, вероятно, имеет решающее значение для наиболее высокой активности транспортера и улучшенных кинетических характеристик транспортной активности: а именно, Y в положении 2 (пептид №91, соответствующий SEQ ID NO: 142).
На основе результатов, полученных в этом эксперименте, можно сделать следующие выводы:
- после 24-часовой инкубации все транспортеры с консенсусной последовательностью rXXXrXXXr (SEQ ID NO: 31) (см. таблицу 2 для выбора возможных последовательностей) отличались более высокой способностью к интернализации, чем транспортер L-TAT (SEQ ID NO: 43) (фиг.6). Эти результаты полностью валидируют консенсусную последовательность rXXXrXXXr (SEQ ID NO: 31);
- одно из положений имеет решающее значение для обеспечения наиболее высокой активности транспортера (см. также фиг.6): а именно, Y в положении 2 (последовательность 91, соответствующая SEQ ID NO: 142).
Следовательно такие полученные из ТАТ последовательности, представленные в таблице 4, которые отличаются наличием Y в положении 2, являются предпочтительными, прежде всего в том случае, когда последовательность состоит из 9 аминокислот и имеет консенсусную последовательность rXXXrXXXr (SEQ ID NO: 31).
Пример 4: Оценка высвобождения цитокинов и хемокинов
Ниже описана процедура оценки количества высвободившихся человеческих цитокинов нескольких типов после индуцированной лигандом секреции из человеческих клеток (кровь, WBC, РВМС, очищенные первичные лимфоциты, клеточные линии и т.д.).
Для этой цели применяли метод сэндвич-ELISA, который позволяет измерять количество антигена между двумя слоями антител (т.е. между иммобилизованным (улавливающим) антителом и идентифицирующим антителом). Подлежащий оценке антиген должен содержать по меньшей мере два антигенных сайта, которые обладают способностью связываться с антителом, поскольку в формировании сэндвича принимают участие по меньшей мере два антитела. В системе сэндвич-ELISA в качестве иммобилизованного антитела и идентифицирующего антитела можно применять как моноклональные, так и поликлональные антитела. Моноклональные антитела распознают единичный эпитоп, что позволяет осуществлять точное определение и количественную оценку небольших различий в антигене. Поликлональное антитело часто используют в качестве улавливающего антитела для сорбции максимально возможного количества антигена. Преимуществом сэндвич-ELISA является то, что образец не требуется очищать перед анализом, и анализ может быть очень чувствительным (вплоть до 2-5 раз более чувствительным, чем прямой или косвенный анализ).
Метод можно применять для определения действия ингибиторов JNK, предлагаемых в настоящем изобретении, in vitro/на культуре клеток. При применении в нетоксичных дозах об эффективности соединения можно судить по снижению уровней цитокинов (изменение оптической плотности (абсорбция при 450 нм)) по сравнению с необработанными образцами и при мониторинге с помощью ELISA. Результаты выражают в нг/мл.
4.1 Материал
96-Луночный планшет:
для сбора супернатантов (Ref 82.1581, фирма Sarstedt),
для ELISA (F96 maxisorp, Ref 442404, фирма Nunc)
TopSeal-A: материал для запечатывания 96-луночного планшета (Ref 600585, фирма PerkinElmer).
Реагенты для ELISA
Сенсибилизирующий буфер для ELISA: 0,1М Na-карбонат, рН 9,5 (=7,13 г NaHCO3 (ref 71627, фирма Fluka)+1,59 г Na2CO3 (ref 71345, фирма Fluka) в 1 л Н2О, доведение значения рН до 9,5 с помощью концентрированного NaOH.
Отмывочный буфер для ELISA: 1×ЗФР+0,01% Твин 20. Приготавливают 1 л 1×ЗФР (10×ЗФР: ref 70011, фирма GIBCO) и медленно добавляют при перемешивании с помощью магнитной мешалки 100 мкл Твин 20 (refP1379, фирма Sigma).
Разбавитель для осуществления анализа: 1×ЗФР+10% FBS (RefA15-151, фирма РАА, декомплементированная при 56°С в течение 30 мин).
DAKO ТМВ (ref S1599, фирма DAKO): поступающий в продажу раствор субстрата.
Стоп-раствор: 1М Н3РО4 (→ для приготовления 200 мл = 177 мл Н2О+23 мл H3PO4, 85% (ref 345245, фирма Aldrich).
Набор для ELISA (реагенты для 20 планшетов)
IFN-γ: Человеческий IFN-γ, набор для ELISA, BD OptEIA™ (ref 555142, фирма DB).
IL-1β: Человеческий IL-1β, набор для ELISA II, BD OptEIA™ (ref 557953, фирма BD).
IL-10: Человеческий IL-10, набор для ELISA II, BD OptEIA™ (ref 555157, фирма DB).
IL-12: Человеческий IL-12 (p70), набор для ELISA, BD OptEIA™ (ref 555183, фирма DB).
IL-15: Человеческий IL-15, набор для ELISA, BD OptEIA™ (ref 559268, фирма DB).
IL-2: Человеческий IL-2, набор для ELISA, BD OptEIA™ (ref 555190, фирма DB).
IL-4: Человеческий IL-4, набор для ELISA, BD OptEIA™ (ref 555194, фирма DB).
IL-5: Человеческий IL-5, набор для ELISA, BD OptEIA™ (ref 555202, фирма DB).
IL-6: Человеческий IL-6, набор для ELISA I, BD OptEIA™ (ref 555220, фирма DB).
IL-8: Человеческий IL-8, набор для ELISA, BD OptEIA™ (ref 555244, фирма DB).
MCP-1: Человеческий МСР-1, набор для ELISA BD OptEIA™ (ref 555179, фирма BD).
TNF-α: Человеческий TNF, набор для ELISA, BD OptEIA™ (ref 555212, фирма DB).
Измерение абсорбции: Абсорбцию измеряли с помощью планшет-ридера Fusion Alpha (фирма Perkin Elmer).
Пипетки: автоматические пипетки-микродозаторы, цифровые пипетки или многоканальные пипетки.
4.2 Метод
Приготовление образцов
Образцы представляли собой супернатант культуральной среды культивируемых человеческих клеток (как правило, цельная кровь, WBC, РВМС, очищенный подтип WBC, линии злокачественных клеток). Удаляли любые (микро)частицы центрифугированием (400 g, 5 мин, 4°С) и применяли непосредственно для анализа или хранили образцы при ≤ -20°С. Необходимо избегать повторения циклов замораживания-оттаивания.
За 1 ч до применения образцы размораживали на льду и их центрифугировали. На стадии 11 разводили образцы в разбавителе для анализа непосредственно в планшете (добавляли сначала разбавитель для анализа, затем образцы и пипетировали вверх-вниз):
Приготовление стандарта
После нагревания лиофилизированного стандарта до комнатной температуры осторожно открывали пузырек, избегая потери продукта. Восстанавливали лиофилизированный стандарт соответствующим объемом деонизированной воды, получая маточный раствор стандарта. Давали стандарту уравновеситься в течение по меньшей мере 15 мин перед приготовлением разведении. Осторожно встряхивали для смешения. После восстановления немедленно разделяли стандарт на аликвоты, которые вносили в полипропиленовые пузырьки по 50 мкл на пузырек и замораживали при -20°С и хранили в течение периода времени вплоть до 6 месяцев. При необходимости хранили при 2-8°С вплоть до 8 ч перед разделением на аликовоты/замораживанием. Не оставляли восстановленный стандарт при комнатной температуре.
Непосредственно перед использованием для построения стандартной кривой по десяти точкам получали 2-кратные серийные разведения в реагенте, обозначенном как разбавитель. В качестве наиболее высокой концентрации стандарта рекомендована концентрация 4000 пг/мл.
Приготовление смеси для обнаружения
Одностадийная инкубация реагентов биотин/SAv. Добавляли требуемый объем идентифицирующего антитела в разбавитель, применяемый для анализа. За 15 мин до использования добавляли в требуемом количестве реагент, представляющий собой фермент, встряхивали и хорошо перемешивали. Рекомендации по разведению для каждой партии представлены в инструкции/сертификате для каждой партии. После применения отбрасывали любой оставшийся применявшийся для обнаружения реагент («рабочий детектор»).
Сенсибилизация с помощью улавливающего антитела
1. Сенсибилизировали лунки титрационного ПВХ-микропланшета, используя по 100 мкл/лунку улавливающего антитела, разведенного в буфере для сенсибилизации. Рекомендации по разведению предназначенного для сенсибилизации антитела представлены в инструкции/сертификате для каждой партии.
2. Накрывали планшет адгезивным пластиком и инкубировали в течение ночи при 4°С.
3. Удаляли сенсибилизирующий раствор и отмывали планшет, заполняя лунки 150 мкл буфера для отмывки.
4. Растворы или буфер для отмывки удаляли путем стряхивания с планшета над раковиной.
5. Повторяли процесс дважды, осуществляя в целом три отмывки.
6. После последней отмывки удаляли оставшийся буфер для отмывки, помещая планшет на бумажное полотенце. Блокада
7. Блокировали оставшиеся связывающие белок сайты в сенсибилизированных лунках, добавляя по 100 мкл реагента, представляющего собой разбавитель, на лунку.
8. Накрывали планшет адгезивным пластиком и инкубировали в течение 1 ч при комнатной температуре.
9. В процессе осуществления инкубации начинали приготовление стандарта.
Добавление образцов
10. Осуществляли одну отмывку, такую же, которая описана на стадии 3, используя 150 мкл буфера для отмывки. После этого планшеты были готовы для добавления образцов.
11. Добавляли в каждую лунку по 50 мкл соответствующим образом разведенных образцов в разбавителе для осуществления анализа. Для получения надежных количественных результатов всегда осуществляли сравнение сигнала неизвестных образцов с сигналом стандартной кривой. Для гарантии точности при исследовании каждого цитокина следует вводить стандарты (в трех повторностях) и контроль.
12. Накрывали планшета адгезивным пластиком и инкубировали в течение 2 ч при комнатной температуре.
Инкубация с идентифицирующим антителом и вторичным антителом
13. Отмывали планшет 4 раза, используя по 150 мкл буфера для отмывки, аналогично осуществлению стадии 3.
14. Добавляли в каждую лунку по 50 мкл смеси для обнаружения (идентифицирующее антитело + вторичное антитело к стрептавидину, конъюгированное с HRP, в разбавителе для анализа) в рекомендованных разведениях (см. инструкцию/сертификат для каждой партии).
15. Накрывали планшет адгезивным пластиком и инкубировали в течение 1 ч при комнатной температуре, предохраняя от действия света.
16. Отмывали планшет 6 раз, используя по 150 мкл буфера для отмывки, аналогично осуществлению стадии 3.
17. Добавляли в каждую лунку по 50 мкл ТМВ-раствора фирмы DAKO, инкубировали в течение 15-20 мин при комнатной температуре в темноте, не запечатывая.
18. Добавляли в каждую лунку по 50 мкл стоп-раствора. Осторожно постукивали по планшету для гарантии полного перемешивания.
19. Перемешивали планшет в течение 5 мин при 500 об/мин на смесителе для планшетов.
20. Определяли оптическую плотность при 450 нм (программа: Cytokine ELISA на планшет-ридере Fusion Alpha).
Анализ данных
Усредняли полученные в трех повторностях данные для каждого применяемого в качестве контроля стандарта и каждого образца. Вычитали среднюю оптическую плотность (О.П.) нулевого стандарта. Строили стандартную кривую зависимости log концентрации цитокина от log О.П. и с помощью регрессионного анализа можно было строить линию, соответствующую наилучшему подбору. Если образцы разводили, то концентрацию, полученную исходя из стандартной кривой, следует умножать на фактор разведения. Стандартную кривую следует создавать для каждого набора проанализированных образцов. Выпадающие значения исключали с помощью теста Граббса. Затем отбрасывали данные, которые не находились в интервале двух С.К.О. Результаты независимых экспериментов учитывали, если данные, полученные для положительного контроля, не противоречили ранее полученным. Результаты независимых экспериментов объединяли (N>3).
Данные представляют собой высвобождение цитокинов в пг/мл или в виде % в сравнении с индуцированным состоянием в отсутствии обработки ингибитором.
Пример 5: Дифференцировка ТНР1 - стимулирование высвобождения цитокинов
Ниже описана процедура индукции производства цитокинов в дифференцированных посредством ФМА человеческих ТНР1-клетках, подвергнутых обработке ЛПС в течение 6 ч, для оценки способности ингибиторов JNK, предлагаемых в настоящем изобретении, в частности ингибитора JNK, имеющего последовательность, представленную в SEQ ID NO: 172, снижать индуцированное стимуляцией высвобождение цитокинов. ТНР1-клетки стимулировали ex vivo различными лигандами для получения данных о высвобождении цитокинов. При применении в нетоксичных дозах эффективность ингибитора JNK определяли по снижению уровней цитокинов по сравнению с необработанными образцами, осуществляя мониторинг с помощью ELISA. Токсичность соединения оценивали по снижению способности клеток восстанавливать соль тетразолия (MTS) с образованием формазана, что приводило к появлению фиолетового цвета.
Процедура:
А. Материал
- Клеточная линия: ТНР-1 (Ref TIB-202, ATCC, партия 57731475).
- Культуральная среда, реагенты и планшеты
Среда RPMI (Ref 21875-091, фирма Invitrogen, дополненная следующими компонентами:
10% FBS (Ref A15-151, фирма РАА): декомплементированная при 56°С в течение 30 мин,
10 мМ Hepes (Ref H0887, фирма Sigma),
50 мкМ β-меркаптоэтанол (Ref 63690, фирма Fluka: концентрация маточного раствора 14,3 М): добавляли 560 мкл аликвот по 50 мМ в ЗФР, хранение при -20°С),
1 мМ пируват натрия (Ref S8636, фирма Sigma),
пенициллин (100 ед./мл)/стрептомицин (100 мкг/мл) (Ref P4333, фирма Sigma).
Затем среду RPMI фильтровали через фильтр с размером пор 0,22 мкм (Ref SCGPU05RE, фирма Millipore).
10×ЗФР (Ref 70011, фирма Invitrogen): разведенный до 1 × с помощью стерильной H2O.
ДМСО: Ref 41444, фирма Fluka.
ФМА (форбол-12-миристат-13-ацета, Ref P1585, фирма Sigma, концентрация 1 мМ = 616,8 мкг/мл в ДМСО, при -20°С). Применяют непосредственно в конечной концентрации 100 нМ в RPMI (1 мкл в 10 мл среды).
ЛПС, ультрачистый (липополисахарид, Ref tlrl-ekLPS, фирма Invivogen, концентрация 5 мг/мл): маточный раствор ЛПС: 3 мкг/мл в ЗФР при 4°С.применяют непосредственно для приготовления 4 × концентрированного раствора (40 нг/мл) в RPMI-среде (минимум 1800 мкл/планшет; для 5 планшетов: 125 мкл ЛПС (3 мкг/мл)+9250 мкл RPMI).
96-луночный планшет:
для прикрепленной культуры клеток (Ref 167008, фирма Nunc),
для сбора супернатантов (Ref 82.1581, фирма Sarstedt),
для ELISA (F96 maxisorp, Ref 442404, фирма Nunc).
Растворы для сенсибилизации: поли-D-лизин (Ref P9011, фирма Sigma): 25 мкг/мл, окончательное разведение в 1×ЗФР
Реагенты и наборы для ELISA
Сенсибилизирующий буфер для ELISA: 0,1М Na-карбонат, рН 9,5 (=7,13 г NaHCO3 (ref 71627, фирма Fluka)+1,59 г Na2CO3 (ref 71345, фирма Fluka) в 1 л Н2О, доведение значения рН до 9,5 с помощью концентрированного NaOH.
Отмывочный буфер для ELISA: 1×ЗФР+0,01% Твин 20 (ref PI 379, фирма Sigma, партия 094К0052) (приготавливают 1 л 1×ЗФР и медленно добавляют при перемешивании с помощью магнитной мешалки 100 мкл Твин 20.
Разбавитель для осуществления анализа: 1×ЗФР+10% FBS (Ref A15-151, фирма РАА, декомплементированная при 56°С в течение 30 мин).
DAKO ТМВ (ref S 1599, фирма DAKO): поступающий в продажу раствор субстрата.
Стоп-раствор: 1М H3PO4 (→ для приготовления 200 мл = 177 мл Н2О+23 мл H3PO4, 85% (ref 345245, фирма Aldrich).
TNFα: набор для ELISA для оценки человеческого TNF, BD OptEIA (ref 555212, фирма DB).
- Оценка цитотоксичности: реагент CellTiter 96 (ref G3581, фирма Promega).
- Применяемое в качестве контроля соединение: SP600125 (ref ALX-270-339-М025, фирма Alexis, концентрация: 20 мМ в ДМСО).
Измерение абсорбции: Абсорбцию измеряли с помощью планшет-ридера Fusion Alpha (фирма Perkin Elmer).
Пипетки: автоматические пипетки-микродозаторы, цифровые пипетки или многоканальные пипетки.
TopSeal-A: материал для запечатывания 96-лучночного планшета (Ref 600585, фирма PerkinElmer).
Б. Метод
Сенсибилизация лунок
Планшеты сенсибилизировали с помощью 200 мкл поли-D-лизина (1 х) и инкубировали в течение 2 ч при 37°С, 5% CO2 и относительной влажности 100%.
Посев клеток
Через 2 ч лунки отмывали дважды с помощью 200 мкл 1×ЗФР (либо немедленно, либо выдерживали с 200 мкл 1°ЗФР при 37°С до применения, но не более 3 дней).
Подсчитывали количество клеток. Отбирали требуемое количество клеток и ресуспендировали в количестве среды, необходимом для получения разведения:
1000000 клеток/мл. Добавляли 100 нМ ФМА для индукции дифференцировки ТНР1 из суспензии моноцитов в прикрепленные макрофаги. Клетки высевали в лунки в 100 мкл среды, используя плотность посева 100000 клеток/лунку. После инокуляции планшеты инкубировали при 37°С, 5% СО2 и относительной влажности 100% в течение 3 дней, после чего добавляли экспериментальные лекарственные средства.
Обработка клеток
Через 3 дня прикрепившиеся клетки выявляли с помощью микроскопа. Содержащие ФМА среды аспирировали и заменяли на 100 мкл свежей RPMI-среды без ФМА (без стадии отмывки с помощью 1×ЗФР).
Экспериментальное лекарственное средство приготавливали в концентрации 10 мМ в Н2О или ДМСО и хранили при -80°С. Каждый раз за день до использования одну аликвоту ингибиторов JNK размораживали и разводили до получения 4 × концентрированного раствора (120 мкМ) в среде RPMI, а затем до требуемой концентрации в среде RPMI. SP600125 разводили до получения 4 × концентрированного раствора (40 мкМ) в среде RPMI, а затем до требуемой концентрации в среде RPMI, содержащей 0,8% ДМСО.
Планшеты обрабатывали 50 мкл среды или раствора, содержащего 4 × конечную требуемую концентрацию лекарственного средства (конечная концентрация соединения-ингибитора JNK: 0, 100 нМ, 1, 3, 10 или 30 мкМ или конечная концентрация применяемого в качестве положительного контроля соединения SP600: 125 0, 10, 100 нМ, 1, 3 или 10 мкМ). После добавления лекарственного средства планшеты инкубировали в течение еще 1 ч при 37°С, 5% CO2 и относительной влажности 100%.
Через 1 ч секрецию TNFα индуцировали, добавляя 50 мкл 4 × концентрированного разведения ультрачистого ЛПС (конечная концентрация 3 нг/мл).
Анализ
Через 6 ч 100 мкл супернатанта переносили в новые 96-луночные планшеты. Указанные планшеты запечатывали и хранили при -20° до анализа с помощью ELISA (см., например, пример 4) секреции цитокинов.
Цитотоксическое действие соединений оценивали по абсорбции MTS (см., например, пример 4) и клетки исследовали с помощью инвертированного микроскопа (Axiovert 40 CFL; фирма Zeiss; 10×).
Анализ данных
Анализ данных осуществляли согласно методу, описанному для ELISA (см. пример 4). В целом, метод состоял в следующем:
При осуществлении ELISA: Усредняли полученные в трех повторностях данные для каждого применяемого в качестве контроля стандарта и каждого образца. Вычитали среднюю оптическую плотность (О.П.) нулевого стандарта. Строили стандартную кривую зависимости log концентрации цитокина от log О.П. и с помощью регрессионного анализа можно было строить линию, соответствующую наилучшему подбору. Если образцы разводили, то концентрацию, полученную исходя из стандартной кривой, следует умножать на фактор разведения. Стандартную кривую следует создавать для каждого набора проанализированных образцов. Выпадающие значения исключали с помощью теста Граббса. Затем отбрасывали данные, которые не находились в интервале двух С.К.О. Результаты независимых экспериментов учитывали, если данные, полученные для положительного контроля, не противоречили ранее полученным данным. Результаты независимых экспериментов объединяли (N>3).
При оценке цитотоксического действия: Для каждого планшета каждого независимого эксперимента, который принимали в расчет при анализе эксперимента по высвобождению цитокина, среднюю абсорбцию при применении только среды рассматривали в качестве фонового уровня и вычитали из каждой величины абсорбции. Среднее значение, полученное по трем повторностям для необработанных клеток, для каждого соединения принимали за показатель 100% жизнеспособности. Среднее значение, полученное в трех повторностях для каждого соединения, стандартизовали относительно 100%. Резко выделяющиеся значения исключали с помощью теста Граббса. Затем отбрасывали данные, которые не находились в интервале двух С.К.О. Результаты независимых экспериментов объединяли (N>3).
Все статистические анализы сравнения вариантов осуществляли с помощью программы GraphPad с использованием следующих критериев:
однонаправленный дисперсионный анализ с последующим использованием критерия множественных сравнений Тьюки. Результаты рассматривали как статистически значимые при Р<0,05.
Пример 6: Воздействие ингибитора JNK, имеющего последовательность SEQ ID NO: 172, на высвобождение TNFα в первичных клетках крови крысы или человека
Цельную кровь получали из анестезированной крысы или здоровых доноров с помощью иглы для венепункции, соединенной с предварительно калиброванной вакуумной пробиркой, которая содержала цитрат натрия. Содержимое пробирок осторожно перемешивали путем 7-8 переворачиваний; и затем пробирки выдерживали при КТ до стимуляции. Ингибитор JNK, имеющий последовательность SEQ ID NO: 172, приготавливали в виде концентрированной в 6 раз субстанции в ЗФР и добавляли из расчета 30 мкл смеси/лунку в 96-луночный планшет. Цельную кровь разводили в соотношении 1:2 в ЗФР и в каждую лунку добавляли по 120 мкл разведенной крови, в которой ранее был добавлен либо только ЗФР, либо ингибитор JNK, имеющий последовательность SEQ ID NO: 172. Цельную кровь инкубировали при 37°С; 85 об/мин (орбитальный инкубатор фирмы SI500, фирма Stuart) в течение 60 мин. Приготавливали активаторы (ЛПС), вносили из расчета 30 мкл ЛПС, концентрированного в 6 раз/лунку. После инкубации в течение 60 мин ЛПС добавляли к крови, кровь перемешивали пипетированием вверх-вниз и затем выдерживали в течение 4 ч при перемешивании (85 об/мин), при 37°С. После инкубации в течение 4 ч планшеты центрифугировали примерно при 770 g, при 4°С в течение 15 мин в предварительно охлажденной центрифуге. В конце процедуры собирали супернатанты и выдерживали при -20°С вплоть до осуществления количественной оценки цитокинов. Затем количественно оценивали уровни цитокинов (IL-6, IL-2, IFNγ и TNFα) с использованием стандартных наборов для ELISA (например, фирма R&D Systems: DuoSet Elisas; или фирмы BD Biosciences: BD Opteia Set Elisa). Результаты выражали в виде мг оцениваемого цитокина/мл супернатанта.
Аналогичный эксперимент осуществляли с использованием в качестве активатора/стимулятора ФМА + иономицина вместо ЛПС.
Пример 7: Время полужизни конкретных ингибиторов JNK, представленных в настоящем описании
Оценивали расщепление ингибиторов JNK, последовательности которых представлены в SEQ ID NO: 196, 197 и 172 (конечная концентрация 0,1 мМ) в человеческой сыворотке (10 и 50% в 1×ЗФР). Эксперимент осуществляли согласно методу, описанному у Tugyi и др,, Proc Nati Acad Sci USA, 2005, cc.413-418. Содержание нерасщепленного пептида определяли с помощью UPLC (сверхпроизводительная жидкостная хроматография)-МС. Стабильность SEQ ID NO: 196, 197 и 172 определяли идентично, но с использованием двух различных анализов. В то время как ингибитор JNK, последовательность которого представлена в SEQ ID NO: 196, полностью расщеплялся на аминокислотные остатки в течение 6 ч, ингибитор JNK, последовательность которого представлена в SEQ ID NO: 172, полностью расщеплялся только через 14 дней. Ингибитор JNK, последовательность которого представлена в SEQ ID NO: 197, сохранял стабильность в течение более чем 30 дней.
Пример 8: Зависящее от дозы ингибирование ингибитором JNK, имеющим последовательность SEQ ID NO: 172, индуцированного CD3/CD28 высвобождения IL-2 в крысиных первичных Т-клетках
Контрольных животных умерщвляли, изымали лимфатические узлы (LN) и выдерживали в полной RPMI-среде. LN растирали в полной RPMI-среде на фильтре с размером отверстий 70 мкм с помощью 5-миллилитрового поршня. Добавляли несколько капель среды для поддерживания сетчатого фильтра во влажном состоянии. Клетки центрифугировали в течение 7 мин при 450 g и 4°С. Дебрис ресуспендировали в течение 5 мл в свежей среде. Клетки вновь пропускали через сетчатый фильтр для клеток. Подсчитывали клетки в аликвоте, клетки центрифугировали вновь в течение 10 мин при 1400 об/мин при 4°С. Клетки ресуспендировали в MACS-буфере (80 мкл MACS-буфера на 10 клеток). Добавляли по 10 мкл микрогранул с антителом к крысиному ГКГ на 10 миллионов клеток, клетки инкубировали в течение 15 мин при 4°-8°С. Клетки отмывали с помощью 15 мл MACS-буфера и центрифугировали в течение 7 мин при 700 g и 4°С. Дебрис ресуспендировали в 500 мкл MACS-буфера на 108 клеток. Одну LS-колонку, которую помещали в магнитное поле MACS-сепаратора, применяли для оценки образцов от одного животного. Колонку сначала промывали 3 мл MACS-буфера. Одну пробирку помещали под колонку на лед для сбора клеток (т.е. Т-клеток)(использовали негативную селекцию, поэтому собирали элюат). Добавляли клеточную суспензию и собирали элюат на льду. Колонку отмывали трижды 3 мл MACS-буфера. Элюированные Т-клетки центрифугировали в течение 7 мин при 700 g и 4°С. Подсчитывали количество ресуспендированных клеток и высевали с плотностью 200000 клеток/лунку в 100 мкл полной среды. Планшеты предварительно сенсибилизировали за 1 день до начала эксперимента, используя 2 мкг/мл антитела к CD3, а в день эксперимента планшеты отмывали трижды ЗФР. Клетки обрабатывали 100 мкл (поли)пептида ингибитора JNK (SEQ ID NO: 172), концентрировали в два раза за 1 ч до активации лигандами. Через 1 ч после предварительной обработки (поли)пептидом ингибитора JNK (SEQ ID NO: 172), клетки стимулировали, используя 2 мкг/мл антитела к CD28 в течение 24 ч. Через 24 ч после стимуляции супернатант собирали и хранили при -20°С до анализа. Затем количественно оценивали уровни цитокинов с использованием стандартных наборов для ELISA. Результаты выражали в пг оцениваемого цитокина/мл супернатанта.
В дополнительном эксперименте использовали протокол, практически соответствующий вышеуказанному, но помимо (поли)пептида ингибитора JNK, имеющего последовательность SEQ ID NO: 172, оценивали также ингибиторы JNK, имеющие последовательность SEQ ID NO: 197, и молекулу лекарственного средства SP600125, что позволяло сравнивать воздействие указанных ингибиторов на ингибирование индуцированного CD3/CD28 высвобождение IL-2.
Пример 9: Воздействие ингибитора JNK на высвобождение TNFα/IL-2 в человеческой цельной крови
Цельную кровь здоровых добровольцев собирали с помощью иглы для венопункции, соединенной с предварительно калиброванной вакуумной пробиркой, которая содержала цитрат натрия. Содержимое пробирок осторожно перемешивали путем 7-8 переворачиваний; и затем пробирки выдерживали при КТ до осуществления стимуляции. Добавляли по 350 мкл RPMI+P/S в 1,2-миллилитровый 96-луночный планшет. Приготавливали концентрированный в 10 раз ингибитор, имеющий последовательность SEQ ID NO: 172, в среде RPMI+P/S (50 мкл на лунку). Добавляли по 50 мкл в 1,2-миллилитровые 96-луночные планшеты. Затем вносили по 50 мкл цельной крови в каждую лунку, в которую предварительно были добавлены либо только среда, либо ингибитор JNK. Цельную кровь инкубировали при 37°С, 5% CO2 в течение 60 мин. Получали и добавляли по 50 мкл/лунку лигандов, разведенных в RPMI+P/S, что соответствовало конечному 10-кратному разведению концентрата. После инкубации в течение 60 мин добавляли лиганд; затем содержимое лунок перемешивали пипетированием крови вверх-вниз. Цельную кровь инкубировали в течение 3 дней при 37°С (содержимое лунок перемешивали пипетированием содержимого каждой лунки вверх-вниз один раз в день). В конце периода инкубации содержимое планшетов перемешивали и затем центрифугировали при 2500 об/мин, 4°С в течение 15 мин в предварительно охлажденной центрифуге. Затем количественно оценивали уровни цитокинов с использованием стандартных наборов для ELISA. Результаты выражали в пг оцениваемого цитокина/мл супернатанта.
Осуществляли аналогичный эксперимент с небольшими модификациями. В случае стимуляции с помощью CD3/CD8 для сенсибилизации применяли антитело к CD3 в концентрации 2 мкг/мл в ЗФР в течение ночи при 4°С. В день проведения эксперимента лунки отмывали трижды ЗФР и выдерживали в ЗФР при 37°С до использования. Антитело к CD28 добавляли через 1 ч после SEQ ID NO: 172 в конечной концентрации 2 мкг/мл; супернатанты собирали через 3 дня после стимуляции.





























































































































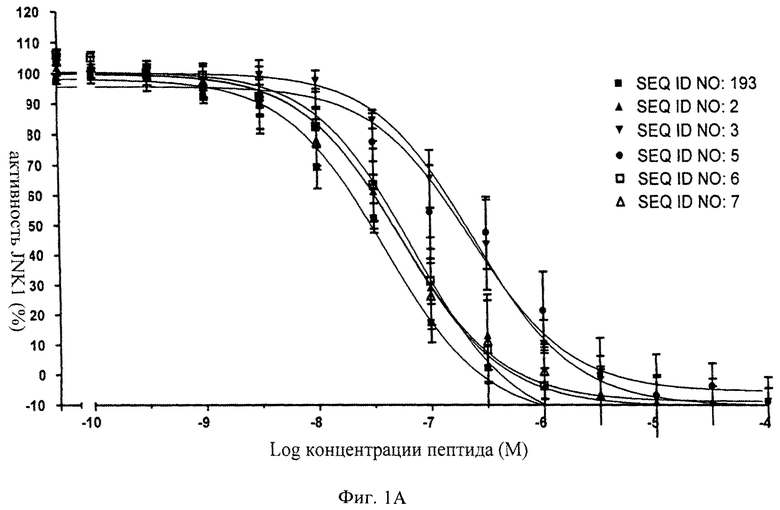
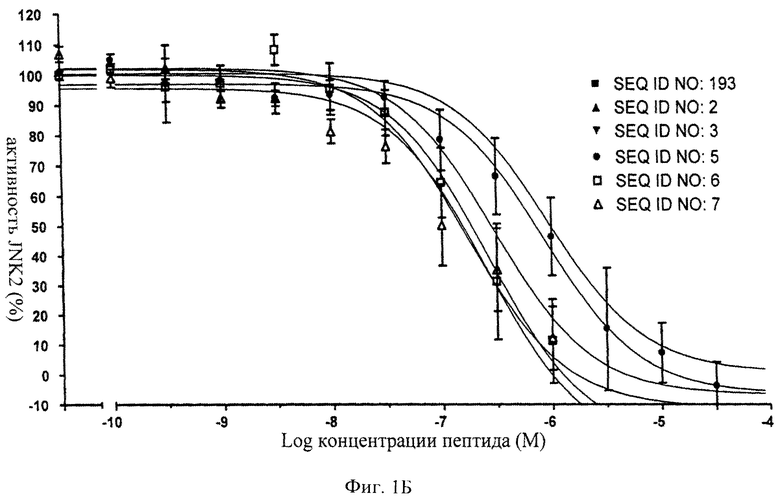
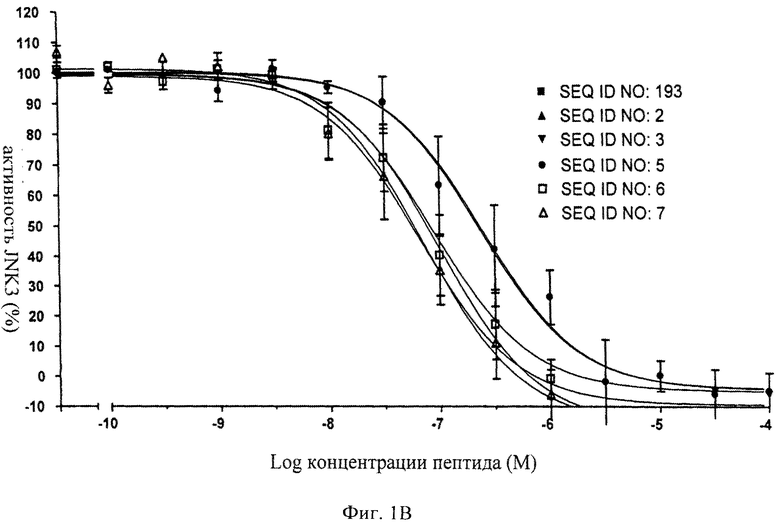
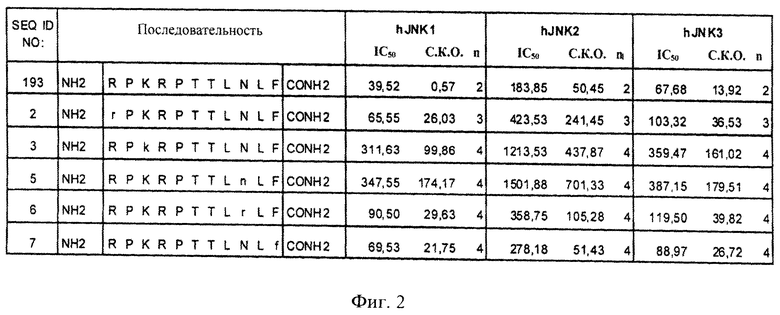

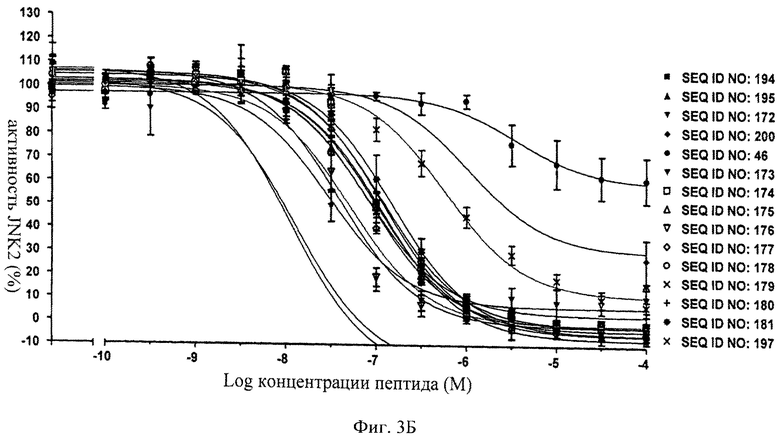



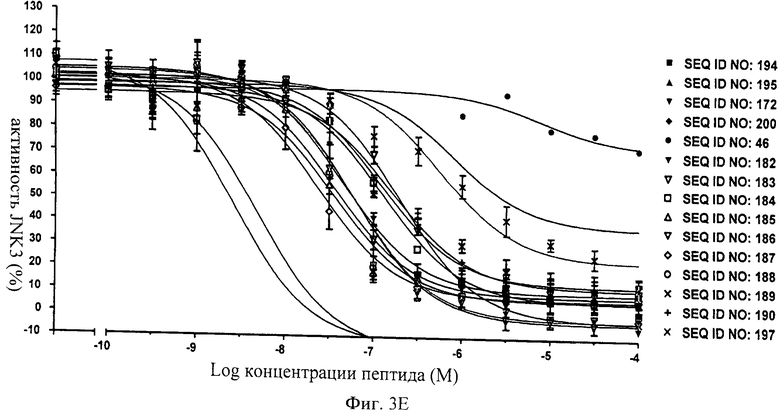
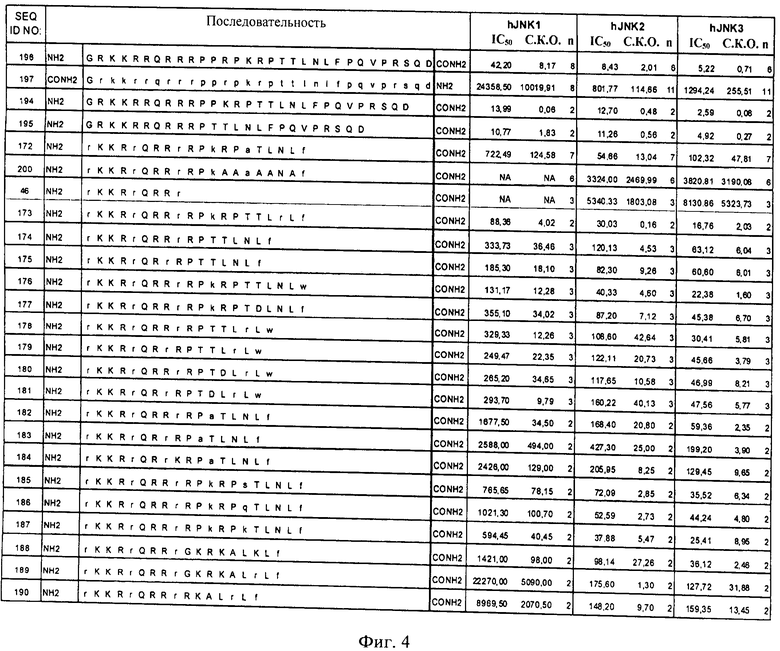


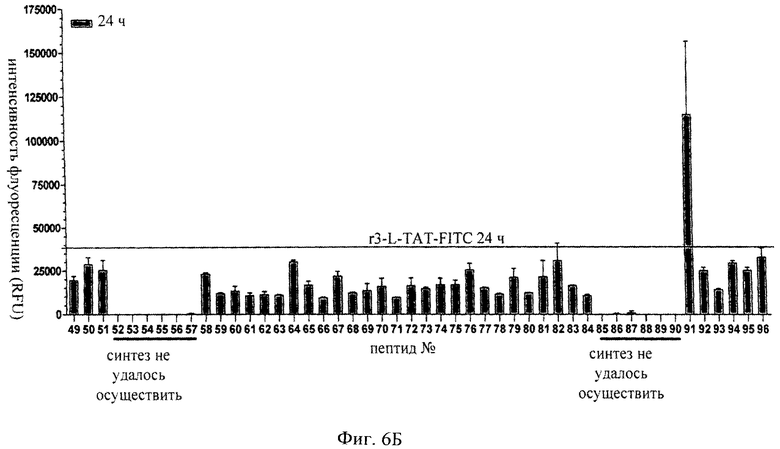

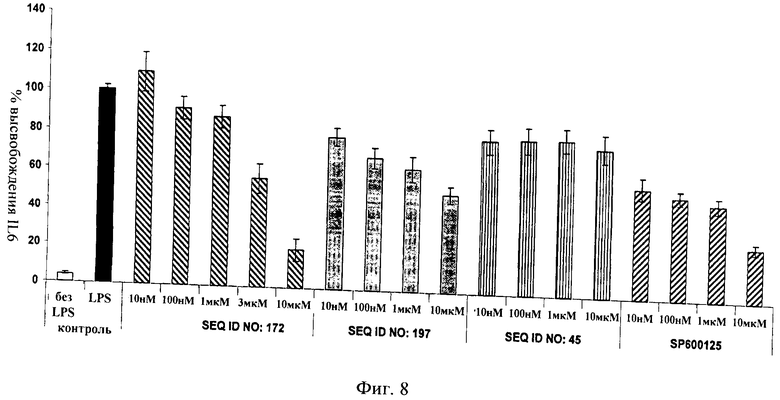
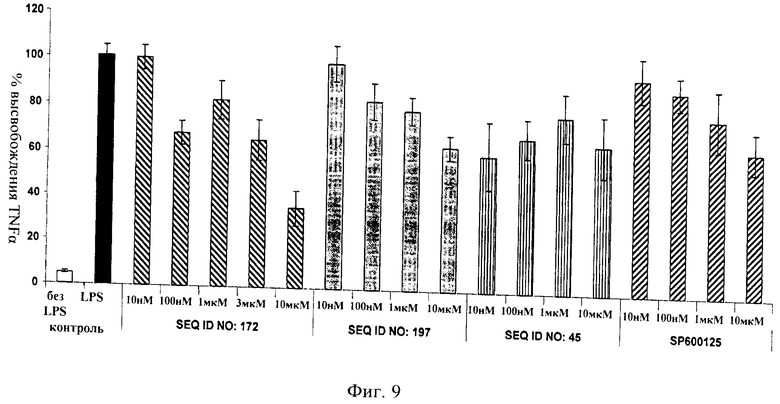


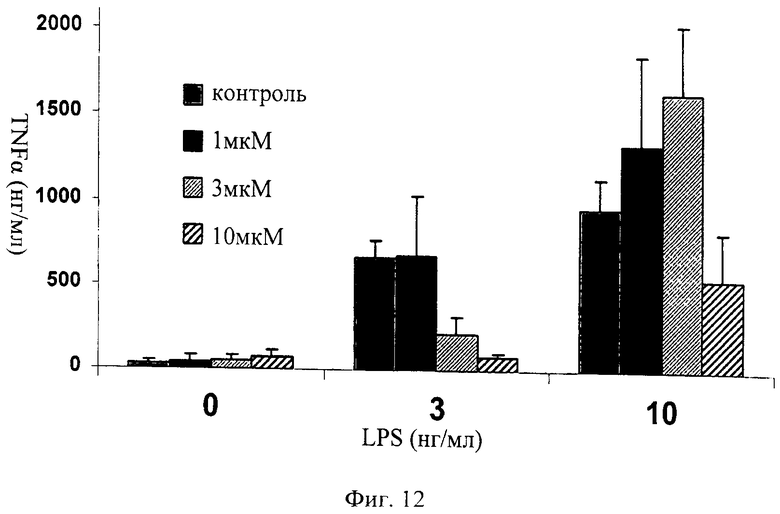
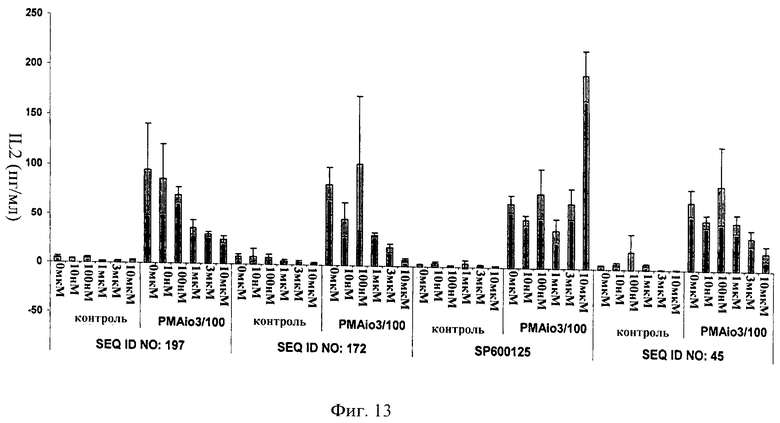
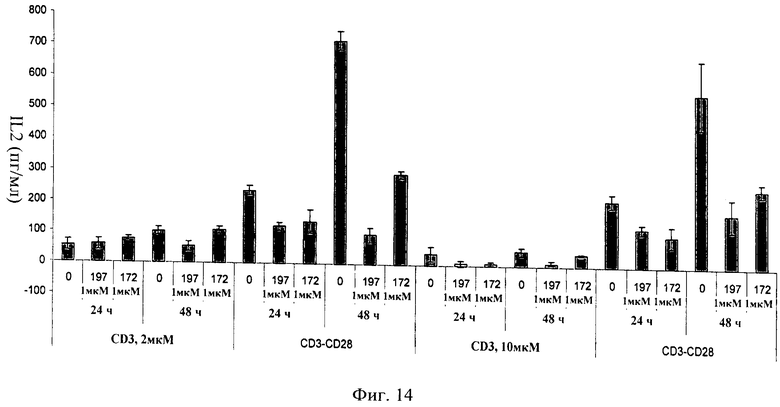
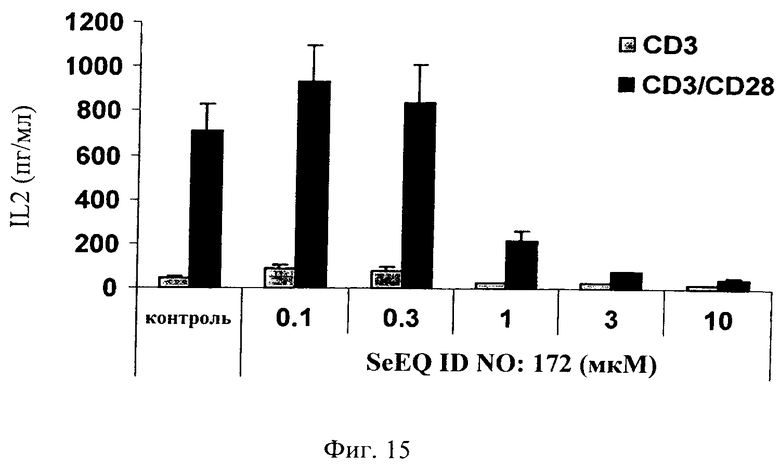
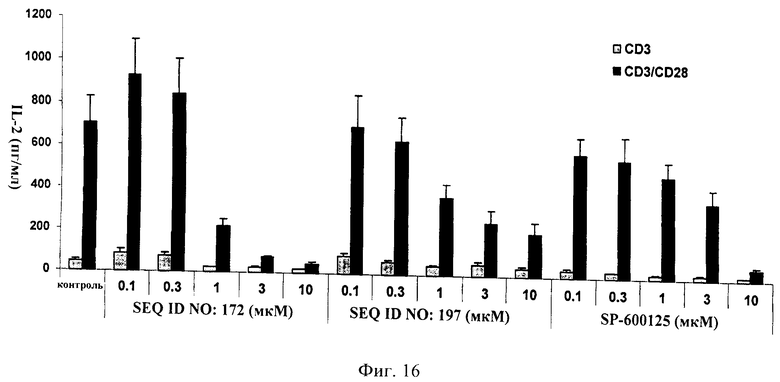


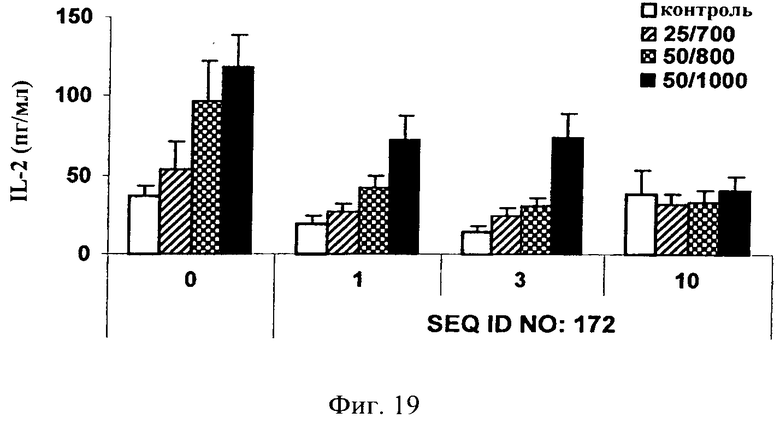
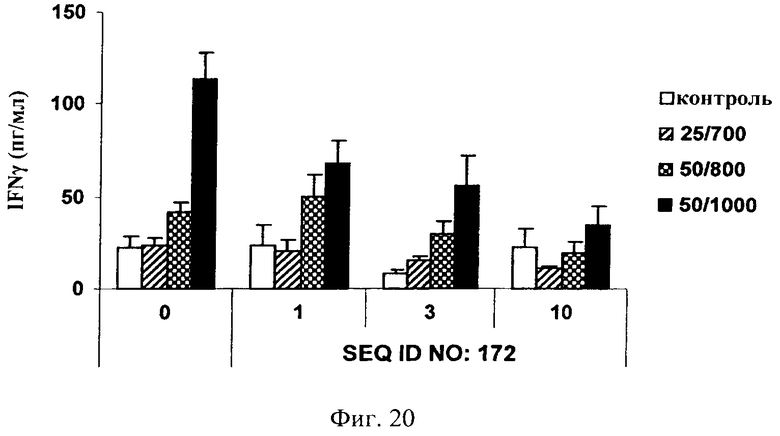
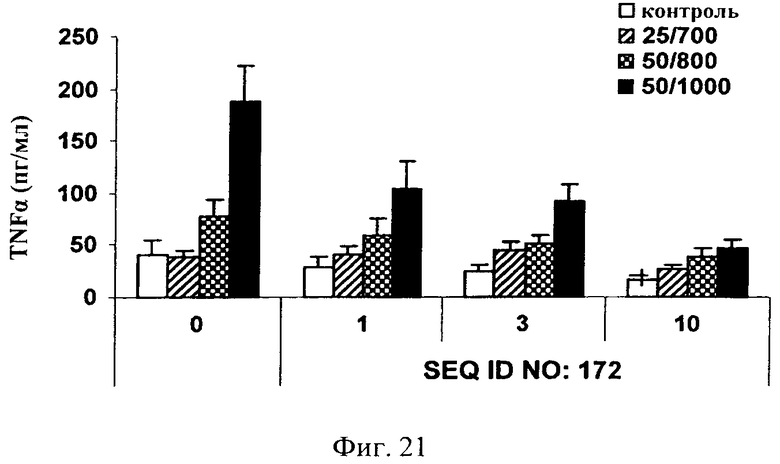
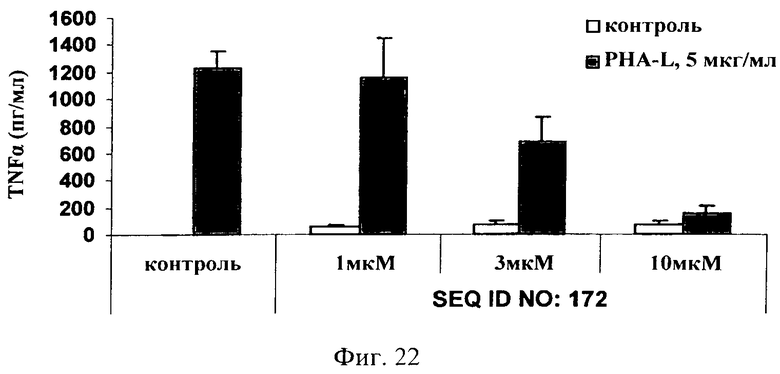
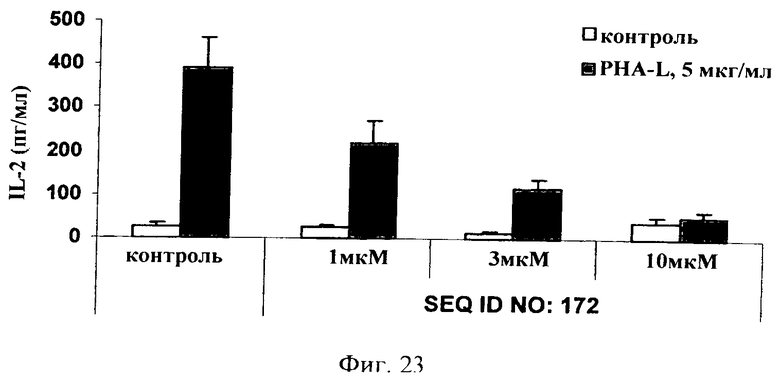
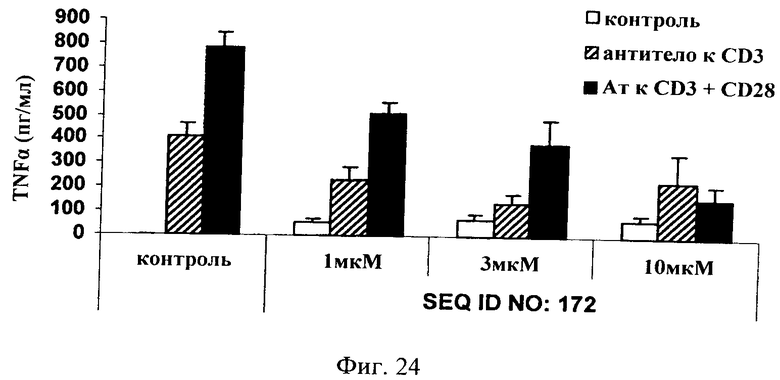
Изобретение относится к новым молекулам-ингибиторам JNK, способам получения антител к указанным молекулам-ингибиторам JNK, а также к соответствующим антителам и клеткам, продуцирующим указанные антитела. 7 н. и 13 з.п. ф-лы, 32 ил., 4 табл., 9 пр.
1. Ингибитор JNK, выбранный из группы, включающей:
ингибитор JNK, который содержит ингибирующую (поли)пептидную последовательность, имеющую следующую общую формулу:
X1-X2-X3-R-X4-X5-X6-L-X7-L-X8 (SEQ ID NO: 1),
в которой X1 обозначает аминокислоту, выбранную из аминокислот R, Р, Q и r,
в которой Х2 обозначает аминокислоту, выбранную из аминокислот R, Р, G и r,
в которой Х3 обозначает аминокислоту, выбранную из аминокислот К, R, k и r,
в которой Х4 обозначает аминокислоту, выбранную из аминокислот Р и К,
в которой Х5 обозначает аминокислоту, выбранную из аминокислот Т, a, s, q, k, или Х5 отсутствует,
в которой Х6 обозначает аминокислоту, выбранную из аминокислот Т, D и А,
в которой Х7 обозначает аминокислоту, выбранную из аминокислот N, n, r и К; и
в которой Х8 обозначает аминокислоту, выбранную из аминокислот F, f и w,
в которой аминокислотный остаток, обозначенный заглавными буквами, соответствует L-аминокислоте, а аминокислотный остаток, обозначенный прописными буквами, соответствует остатку D-аминокислоты,
при условии, что по меньшей мере одна аминокислота, выбранная из группы, включающей X1, Х2, Х3, Х5, Х7 и Х8, представляет(ют) собой D-аминокислоту(ы).
2. Ингибитор JNK по п. 1, в котором по меньшей мере одна из аминокислот, выбранных из группы, включающей Х3, Х5, Х7 и Х8, представляет(ют) собой D-аминокислоту(ы).
3. Ингибитор JNK по п. 1 или 2, в котором ингибирующая (поли)пептидная последовательность выбрана из любой из последовательностей, представленных в SEQ ID NO: 2-27.
4. Ингибитор JNK по п. 1 или 2, где ингибитор JNK содержит ингибирующую (поли)пептидную последовательность, идентичную по меньшей мере на 80% последовательности, выбранной из любой из SEQ ID NO: 2-27.
5. Ингибитор JNK по п. 1 или 2, где ингибитор JNK содержит SEQ ID NO: 8 или ингибирующую (поли)пептидную последовательность, которая идентична по меньшей мере на 80% последовательности SEQ ID NO: 8.
6. Ингибитор JNK по п. 1 или 2, где ингибитор JNK содержит последовательность-транспортер.
7. Ингибитор JNK по п. 6, в котором ингибирующая (поли)пептидная последовательность и последовательность-транспортер перекрываются.
8. Ингибитор JNK по п. 6, в котором последовательность-транспортер содержит последовательность, состоящую из чередующихся D- и L-аминокислот, представленных в любой из последовательностей SEQ ID NO: 28-30.
9. Ингибитор JNK по п. 6, в котором последовательность-транспортер выбрана из любой из последовательностей SEQ ID NO: 31-170.
10. Ингибитор JNK по п. 6, в котором последовательность-транспортер выбрана из любой из последовательностей SEQ ID NO: 31-34, 46, 47 и 52-151.
11. Ингибитор JNK по п. 6, в котором последовательность-транспортер расположена непосредственно на N-конце или непосредственно на С-конце ингибирующей (поли)пептидной последовательности.
12. Ингибитор JNK по п. 6, где ингибитор JNK содержит:
а) последовательность, представленную в любой из SEQ ID NO: 171-190, или
б) последовательность, которая идентична по меньшей мере на 50% последовательности, представленной по меньшей мере в одной из SEQ ID NO: 171-190, при условии, что в указанной последовательности, которая идентична любой из последовательностей, представленных в SEQ ID NO: 171-190:
I) сохраняется остаток L-аргинина (R) в положении 4 в участке последовательности, соответствующем SEQ ID NO: 1,
II) сохраняются два остатка L-лейцина (L) в участке последовательности, соответствующем SEQ ID NO: 1, и
III) присутствует по меньшей мере одна D-аминокислота в положениях X1, Х2, Х3, Х5, Х7 или Х8 в участке последовательности, соответствующем SEQ ID NO: 1.
13. Ингибитор JNK по п. 6, где ингибитор JNK содержит:
а) последовательность, представленную в SEQ ID NO: 172, или
б) последовательность, которая идентична на 50% последовательности, представленной в SEQ ID NO: 172, при условии, что в указанной последовательности, которая идентична на 50% последовательности, представленной в SEQ ID NO: 172,
I) сохраняется остаток L-аргинина (R) в положении 4 в участке последовательности, соответствующем SEQ ID NO: 1,
II) сохраняются два остатка L-лейцина (L) в участке последовательности, соответствующем SEQ ID NO: 1, и
III) присутствует по меньшей мере одна D-аминокислота в положениях X1, Х2, Х3, Х5, Х7 или Х8 в участке последовательности, соответствующем SEQ ID NO: 1.
14. Ингибитор JNK по п. 13, где ингибитор JNK состоит из последовательности, представленной в SEQ ID NO: 172.
15. Способ иммунизации животного, кроме человека, ингибитором JNK по одному из пп. 1-14, заключающийся в том, что осуществляют следующую стадию:
- обрабатывают (иммунизируют) животное, кроме человека, в котором можно получать антитела,
в частности, млекопитающее, кроме человека,
более предпочтительно такое животное, как коза и грызуны, например мышь, крыса и кролик,
ингибитором JNK по одному из пп. 1-14,
более предпочтительно ингибитором JNK, который содержит (поли)пептид, имеющий последовательность, выбранную из любой из последовательностей, представленных в SEQ ID NO: 1-27.
16. Способ получения (поликлонального) антитела, распознающего ингибитор JNK по одному из пп. 1-14, заключающийся в том, что осуществляют следующую стадию:
- выделяют из животного, кроме человека, в котором можно получать антитела,
в частности, млекопитающего, кроме человека,
более предпочтительно такого животного, как коза и грызуны, например мышь, крыса и кролик,
которое ранее было обработано (иммунизировано) ингибитором JNK по одному из пп. 1-14,
более предпочтительно ингибитором JNK, который содержит (поли)пептид, имеющий последовательность, выбранную из любой из последовательностей, представленных в SEQ IDNO: 1-27,
(поликлональное) антитело, распознающее указанный ингибитор JNK.
17. Способ выделения клетки, продуцирующей антитело, которое распознает ингибитор JNK по одному из пп. 1-14, заключающийся в том, что осуществляют следующую стадию:
- выделяют из животного, кроме человека, в котором можно получать антитела,
в частности, млекопитающего, кроме человека,
более предпочтительно такого животного, как коза и грызуны, например мышь, крыса и кролик,
которое ранее было обработано (иммунизировано) ингибитором JNK по одному из пп. 1-14,
более предпочтительно ингибитором JNK, который содержит (поли)пептид, имеющий последовательность, выбранную из любой из последовательностей, представленных в SEQ ID NO: 1-27,
клетку, продуцирующую указанное антитело, которое распознает указанный ингибитор JNK, и
необязательно иммортализуют указанную клетку.
18. Способ получения (моноклонального) антитела, распознающего ингибитор JNK по одному из пп. 1-14, заключающийся в том, что осуществляют следующую стадию:
- выделяют антитело, которое распознает ингибитор JNK по одному из пп. 1-14,
более предпочтительно которое распознает ингибитор JNK, состоящий из (поли)пептида, который имеет последовательность, выбранную из любой из последовательностей, представленных в SEQ ID NO: 1-27,
из супернатанта клеточной культуры клетки, продуцирующей указанное антитело, где клетка необязательно является иммортализованной.
19. Антитело, которое можно получать согласно любому способу по п. 16 или 18, где антитело распознает по меньшей мере один (поли)пептид, выбранный из любой из последовательностей, которые представлены в SEQ ID NO: 1-27, но не распознает практически такой же (поли)пептид, в котором L-аминокислоты присутствуют вместо D-аминокислот.
20. Клетка, которую можно получать с помощью способа по п. 17, где клетка продуцирует антитело по п. 19.
| WO 2009143865 A1, 2009-12-03 | |||
| Прибор для отбирания средней пробы зерна через отверстие в нижней грани сточной трубы | 1927 |
|
SU14330A1 |
| WO 2010072228 A1, 01.07.2010 | |||
| WO 2010072405 A1, 01.07.2010 | |||
| BONNY C ET AL: "Cell-permeable peptide inhibitors of JNK: novel blockers of beta-cell death", DIABETES, AMERICAN DIABETES ASSOCIATION, US, vol | |||
| Устройство для выпрямления многофазного тока | 1923 |
|
SU50A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| PAN JING ET AL: "Small peptide | |||

