РОДСТВЕННЫЕ ЗАЯВКИ НА ПОЛУЧЕНИЕ ПАТЕНТА
[0001] Настоящая заявка испрашивает приоритет согласно предварительным заявкам на патент США US 61/372,072, поданной 9 августа 2010, US 61/307,412, поданной 23 февраля, 2010 и US 61/285,149, поданной 9 декабря, 2009, каждая из которых имеет название «Модуляция экспрессии HSP47», и которые включены в настоящее описание в полном объеме посредством ссылки и для всех целей.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка включает список последовательностей, который обозначен 220-PCT1_ST25_07-Dec-10.txt, при этом копия этого списка в стандарте ASCII, созданная 7 декабря, 2010, размером 533 kb, включена в настоящее описание в полном объеме посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0003] Предложены композиции и способы модуляции экспрессиии hsp47.
УРОВЕНЬ ТЕХНИКИ
[0004] В работе Sato, Y., et al. описано введение липосом, соединенных с витамином А, для доставки малых интерферирующих РНК (siRNA) против gp46, крысиного гомолога белка теплового шока 47 человека, крысиной животной модели цирроза печени. Sato, Y., et al., Nature Biotechnology, vol. 26(4), p.431-442 (2008).
[0005] Chen, J-J., et al. описывал трансфекцию образцов келоидных рубцов человека HSP47-shRNA (малые РНК, образующие шпильки) для исследования пролиферации фибробластов келоидных рубцов. Chen, J-J., et al., British Journal of Dermatology, vol. 156, p.1188-1195 (2007).
[0006] В публикации РСТ патента № WO 2006/068232 описан носитель лекарственного средства, специфичный в отношении астроцитов, который включает производное ретиноевой кислоты и/или аналог витамина А.
КРАТКОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
[0007] В настоящем описании обеспечиваются композиции, способы и наборы для модуляции экспрессии генов-мишеней. В различных аспектах и вариантах настоящего изобретения композиции, способы и наборы, которые обеспечиваются в настоящем описании, модулируют экспрессию белка теплового шока 47 (hsp47), который также известен как SERPINH1. Указанные композиции, способы и наборы могут включать применение молекул нуклеиновых кислот (например, малые интерферирующие нуклеиновые кислоты (siНК), короткие интерферирующие РНК (siРНК), двухцепочечные РНК (dsPHK), микро-РНК (miPHK) или короткие РНК, образующие шпильки, (shPHK)), которые связывают последовательность нуклеотидов (например, последовательность мРНК), кодирующую hsp47, например, кодирующую последовательность мРНК hsp47 человека, примером которой является SEQ ID NO:1. Согласно некоторым предпочтительным вариантам реализации композиции, способы и наборы согласно настоящему описанию подавляют экспрессию hsp47. Например, обеспечиваются молекулы, siНК (например, молекулы dsHK длины RISC или молекулы dsHK длины Dicer), которые снижают или подавляют экспрессию hsp47. Также обеспечиваются композиции, способы и наборы для лечения и/или предотвращения заболеваний, состояний или нарушений, вызываемых hsp47, таких как фиброз печени, цирроз печени, пневмофиброз, включая фиброз легких (в том числе ИЛФ), фиброз почек в результате любых заболеваний (например, ХБП, включая ТХПН), перитонеальный фиброз, хроническое повреждение печени, фибриллогенез, фибротические заболевания других органов, аномальные рубцы (келоидные рубцы) в результате любых возможных типов повреждений кожи: случайных и ятрогенных (операции); склеродермия; кардиофиброз, неудачная фильтрующая операция при глаукоме, а также кишечные спайки.
[0008] Согласно одному аспекту обеспечиваются молекулы нуклеиновых кислот (например, молекулы siНК), в которых (а) молекула нуклеиновой кислоты содержит смысловую цепь и антисмысловую цепь; (b) каждая цепь молекулы нуклеиновой кислоты имеет независимо 15-49 нуклеотидов в длину; (с) последовательность 15-49 нуклеотидов антисмысловой цепи комплементарна последовательности мРНК, кодирующей hsp47 человека (например, SEQ ID NO:1); и (d) последовательность 15-49 нуклеотидов смысловой цепи комплементарна последовательности антисмысловой цепи и содержит последовательность 15-49 нуклеотидов мРНК, кодирующей hsp47 человека (например, SEQ ID NO:1).
[0009] Согласно некоторым вариантам реализации, последовательность антисмысловой цепи, которая комплементарна последовательности мРНК, кодирующей hsp47 человека, включает последовательность, комплементарную последовательности между нуклеотидами 600-800; или 801-899; или 900-1000; или 1001-1300 SEQ ID NO:1; или между нуклеотидами 650-730; или 900-975 SEQ ID NO:1. Согласно некоторым вариантам реализации антисмысловая цепь включает последовательность, которая комплементарна последовательности мРНК, кодирующей hsp47 человека, соответвующей нуклеотидам 674-693 SEQ ID NO:1 или ее части; или нуклеотидам 698-716 SEQ ID NO:1 или ее части; или нуклеотидам 698-722 SEQ ID NO:1 или ее части; или нуклеотидам 701-720 SEQ ID NO:1 или ее части; или нуклеотидам 920-939 SEQ ID NO:1 или ее части; или нуклеотидам 963-982 SEQ ID NO:1 или ее части; или нуклеотидам 947-972 SEQ ID NO:1 или ее части; или нуклеотидам 948-966 SEQ ID NO:1 или ее части; или нуклеотидам 945-969 of SEQ ID NO:1 или ее части; или нуклеотидам 945-963 SEQ ID NO:1 или ее части.
[0010] Согласно некоторым вариантам реализации антисмысловая цепь молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию включает последовательность, соответствующую SEQ ID NO:4 или ее части; или SEQ ID NO:6 или ее части; или SEQ ID NO:8 или ее части; или SEQ ID NO:10 или ее части; или SEQ ID NO:12 или ее части; или SEQ ID NO:14 или ее части; или SEQ ID NO:16 или ее части; или SEQ ID NO:18 или ее части; или SEQ ID NO:20 или ее части; или SEQ ID NO:22 или ее части; или SEQ ID NO:24 или ее части; или SEQ ID NO:26 или ее части; или SEQ ID NO:28 или ее части; или SEQ ID NO:30 или ее части; или SEQ ID NO:32 или ее части; или SEQ ID NO:34 или ее части; или SEQ ID NO:36 или ее части; или SEQ ID NO:38 или ее части; или SEQ ID NO:40 или ее части; или SEQ ID NO:42 или ее части; или SEQ ID NO:44 или ее части; или SEQ ID NO:46 или ее части; или SEQ ID NO:48 или ее части; или SEQ ID NO:50 или ее части; или SEQ ID NO:52 или ее части; или SEQ ID NO:54 или ее части; или SEQ ID NO:56 или ее части; или SEQ ID NO:58 или ее части. Согласно некоторым вариантам реализации смысловая цепь молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию включает последовательность, соответствующую SEQ ID NO:3 или ее части; или SEQ ID NO:5 или ее части; или SEQ ID NO:7 или ее части; или SEQ ID NO:9 или ее части; или SEQ ID NO:11 или ее части; или SEQ ID NO:13 или ее части; или SEQ ID NO:15 или ее части; или SEQ ID NO:17 или ее части; или SEQ ID NO:19 или ее части; или SEQ ID NO:21 или ее части; или SEQ ID NO:23 или ее части; или SEQ ID NO:25 или ее части; или SEQ ID NO:27 или ее части; или SEQ ID NO:29 или ее части; или SEQ ID NO:31 или ее части; или SEQ ID NO:33 или ее части; или SEQ ID NO:35 или ее части; или SEQ ID NO:37 или ее части; или SEQ ID NO:39 или ее части; или SEQ ID NO:41 или ее части; или SEQ ID NO:43 или ее части; или SEQ ID NO:45 или ее части; или SEQ ID NO:47 или ее части; или SEQ ID NO:49 или ее части; или SEQ ID NO:51 или ее части; или SEQ ID NO:53 или ее части; или SEQ ID NO:55 или ее части; или SEQ ID NO:57 или ее части.
[0011] Согласно некоторым предпочтительным вариантам реализации антисмысловая цепь молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию включает последовательность, соответствующую любой из антисмысловых последовательностей, представленных в Таблице А-19. Согласно некоторым предпочтительным вариантам реализации антисмысловую цепь и смысловую цепь выбирают из пар последовательностей, показанных в Таблице А-19. Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных в SERPINH1_4, SERPINH1_12, SERPINH1_18, SERPINH1_30, SERPINH1_58 и SERPINH1_88. Согласно некоторым вариантам реализации антисмысловую цепь и смысловую цепь выбирают из пар последовательностей, представленных в SERPINH1_4 (SEQ ID NOS:195 и 220), SERPINH1_12 (SEQ ID NOS:196 и 221), SERPINH1_30 (SEQ ID NOS:199 и 224), и SERPINH1_58 (SEQ ID NOS:208 и 233).
[0012] Согласно некоторым вариантам реализации антисмысловая и смысловая цепи молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию включают пары последовательностей, представленные в SERPINH1_4 (SEQ ID NOS:195 и 220). Согласно некоторым вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию включает антисмысловую и смысловую цепи пар последовательностей, представленных в SERPINH1_12 (SEQ ID NOS:196 и 221). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию включают пары последовательностей, представленные в SERPINH1_30 (SEQ ID NOS:199 и 224). Согласно некоторым вариантам реализации молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию она включает антисмысловую и смысловую цепи пар последовательностей, представленных в SERPINH1_58 (SEQ ID NOS:208 и 233).
[0013] Согласно некоторым вариантам реализации антисмысловая цепь молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию включает последовательность, соответствующую любой из антисмысловых последовательностей, представленных в любой из Таблиц В или С.
[0014] Согласно некоторым предпочтительным вариантам реализации антисмысловая цепь молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию включает последовательность, соответствующую любой антисмысловой последовательности, представленной в Таблице А-18. Согласно некоторым предпочтительным вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, показанных в Таблице А-18. Согласно некоторым вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию включает антисмысловую и смысловую цепи, выбранные из пар последовательностей, представленных в SERPINH1_2 (SEQ ID NOS:60 и 127), SERPINH1_6 (SEQ ID NOS:63 и 130), SERPINH1_11 (SEQ ID NOS:68 и 135), SERPINH1_13 (SEQ ID NOS:69 и 136), SERPINH1_45 (SEQ ID NOS:97 и 164), SERPINH1_45a (SEQ ID NOS:98 и 165), SERPINH1_51 (SEQ ID NOS:101 и 168), SERPINH1_52 (SEQ ID NOS:102 и 169) или SERPINH1_86 (SEQ ID NOS:123 и 190). Согласно некоторым предпочтительным вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных в SERPINH1_2 (SEQ ID NOS:60 и 127), SERPINH1_6 (SEQ ID NOS:63 и 130), SERPINH1_45a (SEQ ID NOS:98 и 165) и SERPINH1_51 (SEQ ID NOS:101 и 168).
[0015] Согласно некоторым предпочтительным вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию включает антисмысловую и смысловую цепи, выбранные из пар последовательностей, представленных в SERPINH1_2 (SEQ ID NOS:60 и 127). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи включают пары последовательностей, представленные в SERPINH1_6 (SEQ ID NOS:63 и 130). Согласно некоторым вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию включает антисмысловую и смысловую цепи, выбранные из пар последовательностей, представленных SERPINH1_11 (SEQ ID NOS:68 и 135). Согласно некоторым вариантам реализации the antisense and sense strands are the sequence pairs set forth in SERPINH1_13 (SEQ ID NOS:69 и 136). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленных в SERPINH1_45 (SEQ ID NOS:97 и 164). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленных в SERPINH1_45a (SEQ ID NOS:98 и 165). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_51 (SEQ ID NOS:101 и 168).
[0016] Согласно некоторым вариантам реализации антисмысловая цепь молекулы нуклеиновой кислоты(например, молекула siНК) согласно настоящему описанию включает последовательность, соответствующую любой из антисмысловых последовательностей, показанных в любой из Таблиц D или Е.
[0017] Согласно различным вариантам реализации молекул нуклеиновых кислот (например, молекул siНК) согласно настоящему описанию антисмысловая цепь может иметь 15-49 нуклеотидов в длину (например, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48 или 49 нуклеотидов в длину); или 17-35 нуклеотидов в длину; или 17-30 нуклеотидов в длину; или 15-25 нуклеотидов в длину; или 18-25 нуклеотидов в длину; или 18-23 нуклеотидов в длину; или 19-21 нуклеотидов в длину; или 25-30 нуклеотидов в длину; или 26-28 нуклеотидов в длину. Согласно некоторым вариантам реализации молекул нуклеиновых кислот (например, молекул siНК) согласно настоящему описанию антисмысловая цепь может иметь 19 нуклеотидов в длину. Аналогично, смысловая цепь молекул нуклеиновых кислот (например, молекул siНК) согласно настоящему описанию может иметь 15-49 нуклеотидов в длину (например, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48 или 49 нуклеотидов в длину); или 17-35 нуклеотидов в длину; или 17-30 нуклеотидов в длину; или 15-25 нуклеотидов в длину; или 18-25 нуклеотидов в длину; или 18-23 нуклеотидов в длину; или 19-21 нуклеотидов в длину; или 25-30 нуклеотидов в длину; или 26-28 нуклеотидов в длину. Согласно некоторым вариантам реализации молекул нуклеиновых кислот (например, молекул siНК) согласно настоящему описанию, смысловая цепь может иметь 19 нуклеотидов в длину. Согласно некоторым вариантам реализации молекул нуклеиновых кислот (например, молекул siНК) согласно настоящему описанию антисмысловая цепь и смысловая цепь могут иметь 19 нуклеотидов в длину. Двунитевой участок молекул нуклеиновых кислот (например, молекул siНК) согласно настоящему описанию может иметь 15-49 нуклеотидов в длину (например, приблизительно 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48 или 49 нуклеотидов в длину), 15-35 нуклеотидов в длину; или 15-30 нуклеотидов в длину или приблизительно 15-25 нуклеотидов в длину; или 17-25 нуклеотидов в длину; или 17-23 нуклеотидов в длину; или 17-21 нуклеотидов в длину; или 25-30 нуклеотидов в длину; или 25-28 нуклеотидов в длину. Согласно различным вариантам реализации молекул нуклеиновых кислот (например, молекул siНК) согласно настоящему описанию двунитевой участок может иметь 19 нуклеотидов в длину.
[0018] Согласно некоторым вариантам реализации смысловая и антисмысловая цепи нуклеиновой кислоты (например, молекулы нуклеиновой кислоты siНК) согласно настоящему описанию являются отдельными полинуклеотидными цепями. Согласно некоторым вариантам реализации, отдельная антисмысловая и смысловая цепи образуют двухцепочечную структуру посредством водородных связей, например, путем связывания пар оснований по Уотсону-Крику. Согласно некоторым вариантам реализации смысловая и антисмысловая цепи являются отдельными цепями, которые ковалентно связаны друг с другом. Согласно другим вариантам реализации смысловая и антисмысловая цепи являются частью одной цепи полинуклеотидов, имеющей как смысловой, так и антисмысловой участок; согласно некоторым предпочтительным вариантам реализации указанная полинуклеотидная цепь имеет шпилечную структуру.
[0019] Согласно некоторым вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) представляет собой молекулу двухцепочечной нуклеиновой кислоты (dsHK), которая имеет симметричные выступающие («липкие») концы и имеет тупой конец с обоих концов. Согласно другим вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) представляет собой молекулу dsHK, которая имеет симметричные выступающие концы и имеет выступающий конец с обеих сторон молекулы dsHK; предпочтительно указанная молекула имеет выступающие концы длиной 1, 2, 3, 4, 5, 6, 7 или 8 нуклеотидов; предпочтительно молекула имеет выступающие концы из 2 нуклеотидов. Согласно некоторым вариантам реализации выступающие концы представляют собой 5' выступающие концы; согласно другим вариантам реализации выступающие концы представляют собой 3' выступающие концы. Согласно некоторым вариантам реализации нуклеотиды на выступающих концах модифованы с помощью модификаций согласно настоящему описанию. Согласно некоторым вариантам реализации нуклеотиды на выступающих концах представляют собой 2'-дезоксинуклеотиды.
[0020] Согласно некоторым предпочтительным вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) представляет собой молекулу dsHK, которая является ассимметричной по выступающим концам и имеет тупой конец с на одном конце молекулы и выступающий конец на другом конце молекулы. Согласно некоторым вариантам реализации выступающий конец имеет длину 1, 2, 3, 4, 5, 6, 7 или 8 нуклеотидов; предпочтительно выступающий конец составляет 2 нуклеотида. Согласно некоторым предпочтительным вариантам реализации ассимметричная молекула dsHK имеет 3'-выступающий конец (например, 3'-выступающий конец из 2 нуклеотидов) на одной стороне двунитевого участка, образованного на смысловой цепи, и тупой конец на другом конце молекулы. Согласно некоторым предпочтительным вариантам реализации ассимметричная молекула dsHK имеет 5'-выступающий конец (например, выступающий 5'-конец длиной 2 нуклеотида) на одной стороне дуплекса, образованного на смысловой цепи, и тупой конец на другой стороне молекулы. Согласно другим предпочтительным вариантам реализации ассимметричная молекула dsHK имеет выступающий 3'-конец (например, выступающий 3'-конец длиной 2 нуклеотида) на одной стороне двунитевого участка, образованного на антисмысловой цепи, и тупой конец на другой стороне молекулы. Согласно некоторым предпочтительным вариантам реализации ассимметричная молекула dsHK имеет выступающий 5'-конец (например, выступающий 5'-конец длиной 2 нуклеотида) на одной стороне двунитевого участка, образованного на антисмысловой цепи, и тупой конец на другой стороне молекулы. Согласно некоторым предпочтительным вариантам реализации выступающие концы представляют собой 2'-дезоксинуклеотида.
[0021] Согласно некоторым вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) имеет шпилечную структуру (имеющую смысловую и антисмысловую цепи на одном полинуклеотиде) со структурой петли на одном конце и тупым концом на втором конце. Согласно некоторым вариантам реализации молекула нуклеиновой кислоты имеет шпилечную структуру со структурой петли на одном конце и выступающим вторым концом (например, выступающим концом длиной 1, 2, 3, 4, 5, 6, 7 или 8 нуклеотидов); согласно некоторым вариантам реализации выступающий конец представляет собой выступающий 3'-конец; согласно некоторым вариантам реализации выступающий конец представляет собой выступающий 5'-конец; согласно некоторым вариантам реализации выступающий конец находится на смысловой цепи; согласно некоторым вариантам реализации выступающий конец находится на антисмысловой цепи.
[0022] Согласно некоторым предпочтительным вариантам реализации молекулу нуклеиновой кислоты выбирают из молекул нуклеиновых кислот, показанных в Таблице 1.
[0023] Молекулы нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию могут включать одну или несколько модификаций или модифицированных нуклеотидов, например, согласно настоящему описанию. Например, молекула нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию может включать модифицированный нуклеотид, имеющий модифицированный остаток сахара; модифицированный нуклеотид, имеющий модифицированное нуклеиновое основание, или модифицированный нуклеотид, имеющий модифицированную фосфатную группу. Аналогично, молекула нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию может включать модифицированный фосфодиэфирный остов и/или может включать модифицированную концевую фосфатную группу.
[0024] Молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию могут иметь один или несколько нуклеотидов, которые включают модифицированную сахарную группу, например, согласно настоящему описанию. Согласно некоторым предпочтительным вариантам реализации модифицированную сахарную группу выбирают из группы, состоящей из 2'-O-метила, 2'-метоксиэтокси, 2'-дезокси, 2'-фтора, 2'-аллила, 2'-O-[2-(метиламино)-2-оксиэтила], 4'-тио, 4'-(СН2)2-O-2'-мостика, 2'-закрытой нуклеиновой кислоты и 2'-O-(N-метилкарбамата).
[0025] Молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию могут иметь одно или несколько модифицированных нуклеиновых оснований, например, согласно настоящему описанию, которые предпочтительно могут представлять собой нуклеиновые основания, выбранные из группы, состоящей из ксантина, гипоксантина, 2-аминоаденина, 6-метила и других алкильных производных аденина и гуанина, 2-пропила и других алкильных производных аденина и гуанина, 5-галоурацила и цитозина, 5-пропинилурацила и цитозина, 6-азо урацила, цитозина и тимина, 5-урацила (псевдоурацила), 4-тиоурацила, 8-гало, амино, тиола, тиоалкила, гидроксила и других замещенных в положении 8 аденинов и гуанинов, 5-трифторметила и других 5-замещенных урацилов и цитозинов, 7-метилгуанина и ациклических нуклеотидов.
[0026] Молекулы нуклеиновой кислоты (например, молекулы siНК) согласно настоящему описанию могут иметь одну или более модификаций фосфодиэфирного остова, например, согласно настоящему описанию. Согласно некоторым предпочтительным вариантам реализации фосфодиэфирная связь модифицирована посредством замещения фофодиэфирной связи на фосфотиоатную, 3'-(или -5')дезокси-3'-(или-5')тио-фосфотиоатную, фосфодитиоатную, фосфоселенатную, 3'-(или -5')дезокси фосфинатную, борано фосфатную, 3'-(или -5')дезокси-3'-(или 5'-)амино фосфоамидатную, гидроген фосфонатную, боранофосфатэфирную, фосфоамидатную, алкил или арил фосфонатную и фосфотриэфирную или фосфористую связи.
[0027] Согласно различным вариантам реализации предлагаемые молекулы нуклеиновой кислоты (например, молекулы siНК) могут включать одну или несколько модификаций в смысловой цепи, но не в антисмысловой цепи. Согласно некоторым вариантам реализации обеспечиваются молекулы нуклеиновой кислоты (например, молекулы siНК), которые включают одну или несколько модификаций в антисмысловой цепи, но не в смысловой цепи. Согласно некоторым вариантам реализации предлагаемые молекулы нуклеиновой кислоты (например, молекулы siНК) включают одну или несколько модификаций как в смысловой цепи, так и в антисмысловой цепи.
[0028] Согласно некоторым вариантам реализации, в котором предлагаемые молекулы нуклеиновой кислоты (например, молекулы siНК) имеют модификации, смысловая цепь включает последовательность чередующихся модифицированных и немодифицированных нуклеотидов, и/или антисмысловая цепь включает последовательность чередующихся модифицированных и не модифицированных нуклеотидов; согласно некоторым предпочтительным типам таких вариантов реализации указанная модификация представляет собой 2'-O-метил (2' метокси или 2'ОМе) сахарную группу. Последовательность чередующихся модифицированных и немодифицированных нуклеотидов может начинаться с модифицированного нуклеотида на 5'-конце или 3'-конце одной из цепей; например. Последовательность чередующихся модифицированных и немодифицированных нуклеотидов может начинаться с модифицированного нуклеотида на 5'-конце или 3'-конце смысловой цепи, и/или Последовательность чередующихся модифицированных и немодифицированных нуклеотидов может начинаться с модифицированного нуклеотида на 5'-конце или 3'-конце антисмысловой цепи. Если и антисмысловая и смысловая цепи включают последовательность чередующихся модифицированных нуклеотидов, последовательность модифицированных нуклеотидов может иметь такую конфигурацию, что модифицированные нуклеотиды в смысловой цепи распорложены напротив модифицированных нуклеотидов в антисмысловой цепи, или может происходит сдвиг по фазе в указанной последовательности, такой что модифицированные нуклеотиды смысловой цепи находятся напротив немодифицированных нуклеотидов антисмысловой цепи и наоборот.
[0029] Молекулы нуклеиновых кислот (например, молекулы siHK) согласно настоящему описанию могут включать 1-3 (т.е. 1, 2 или 3) дезоксинуклеотида на 3′-конце смысловой и/или антисмысловой цепи.
[0030] Молекулы нуклеиновой кислоты (например, молекулы siHK) согласно настоящему описанию могут включать фосфатную группу на 5′-конце смысловой и/или антисмысловой цепи.
Согласно одному аспекту обеспечиваются двухцепочечные молекулы нуклеиновой кислоты, имеющие структуру (А1):
(А1) 5′ (N)x-Z 3′ (антисмысловая цепь)
3′ Z′-(N')y-z′′ 5′ (смысловая цепь)
где каждое из N и N′ представляет собой нуклеотид, который может быть немодифицированным или модифицированным, или представляет собой нетрадиционную функциональную группу;
где каждое из (N)x и (N′)y представляет собой олигонуклеотид, в котором каждый следующий N или N′ соединен со следующим N или N′ ковалентной связью;
где каждое из Ζ и Z′ независимости может присутствовать или отсутствовать, но при наличии независимо содержит 1-5 следующих друг за другом нуклеотидов или других функциональных групп или их комбинацию, ковалентно присоединенные к 3′-концу цепи, в которой он присутствует;
где z′′ может присутствовать или отсутствовать, но при наличии представляет собой блокирующую группу, ковалетно присоединенную к 5′-концу (N′)y;
где каждое из x и y независимо представляет собой целое число от 18 до 40; где последовательность (N′)y комплементарна последовательности (N)x; и при этом (N)x включает антисмысловую последовательность к SEQ ID ΝΟ:1. Согласно некоторым вариантам реализации (N)x включает антисмысловой олигонуклеотид, представленный в Таблице А-19. Согласно другим вариантам реализации (N)x выбирают из антисмыслового нуклеотида, представленного в Таблицах D или С.
[0031] Согласно некоторым вариантам реализации ковалентная связь, соединяющая каждые соседние N или N', представляет собой фосфодиэфирную связь.
[0032] Согласно некоторым вариантам реализации x=y, и каждое из x и y равно 19, 20, 21, 22 или 23. Согласно различным вариантам реализации х=у=19.
[0033] Согласно некоторым вариантам реализации молекул нуклеиновых кислот (например, молекул siНК) согласно настоящему описанию двухцепочечная молекула нуклеиновой кислоты представляет собой siРНК, siНК или miPHK.
[0034] Согласно некоторым вариантам реализации, антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных в SERPINH1_4 (SEQ ID NOS:195 и 220), SERPINH1_2 (SEQ ID NOS:196 и 221), SERPINH1_30 (SEQ ID NOS:199 и 224) и SERPINH1_58 (SEQ ID NOS:208 и 233).
[0035] Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_4 (SEQ ID NOS:195 и 220). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_12 (SEQ ID NOS:196 и 221). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_30 (SEQ ID NOS:199 и 224). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_58 (SEQ ID NOS:208 и 233).
[0036] Согласно некоторым вариантам реализации двухцепочечные молекулы нуклеиновой кислоты содержат ДНК группу или несоответствие мишени в положении 1 антисмысловой цепи (5'-конец). Такая структура описана ниже. Согласно одному варианту реализации обеспечиваются модифицированные молекулы нуклеиновых кислот, имеющие структуру (А2), изложенную ниже:
(А2) 5' N1-(N)x-Z 3'(антисмысловая цепь)
3' Z'-N2-(N')y-z" 5'(смысловая цепь)
где каждое из N2, N и N' представляет собой немодифицированный или модифицированный рибонуклеотид или нетрадиционную группу;
где каждое из (N)x и (N')y представляет собой олигонуклотид, в котором каждый последующий N или N' соединен с прилегающим N или N' ковалентной связью;
где каждый из x и y независимо представляет собой целое число от 17 до 39;
где последовательность (N')y имеет комплементарность последовательности (N)x, и (N)x имеет комплементарность непрерывной последовательности в РНК-мишени;
где N1 ковалентно соединена с (N)x и не соответствует РНК-мишени или представляет собой ДНК группу, комплементарную РНК-мишени;
где N1 представляет собой группу, выбранную из группы, состоящей из природного или модифицированного уридина, дезоксирибоуридина, риботимидина, дезоксириботимидина, аденозина или дезоксиаденозина;
где z" может присутствовать или отсутствовать, но при наличии представляет собой блокирующую группу, ковалентно присоединенную к 5'-концу N2-(N')y; и
где каждое из Z и Z' независимо друг от друга присутствует или отсутствует, но при наличии независимо представляет 1-5 последовательных нуклеотидов, последовательных не являющихся нуклеотидами групп или их комбинацию, ковалентно соединенные с 3'-концом цепи, в которой оно присутствует.
[0037] Согласно некоторым вариантам реализации последовательность (N')y полностью комплементарна последовательности (N)x. Согласно различным вариантам реализации последовательность N2-(N')y комплементарна последовательности N1-(N)x. Согласно некоторым вариантам реализации (N)x содержит антисмысловую последовательность, которая полностью комплементарна приблизительно 17-39 следующим друг за другом нуклеотидам в РНК-мишени. Согласно другим вариантам реализации (N)x содержит антисмысловую последовательность, которая существенно комплементарна приблизительно 17-39 следующим друг за другом нуклеотидам в РНК-мишени.
[0038] Согласно некоторым вариантам реализации N1 и N2 образуют пару оснований по Уотсону-Крику. Согласно некоторым вариантам реализации N1 и N2 образуют пару оснований не по Уотсону-Крику. Согласно некоторым вариантам реализации образуется пара оснований между рибонуклеотидом и дезоксирибонуклеотидом.
[0039] Согласно некоторым вариантам реализации x=y=18, x=y=19 или x=y=20. Согласно предпочтительным вариантам реализации x=y=18. Если х=18 в N1-(N)x, N1 обозначает положение 1, и положения 2-19 включены в (N)18. Если у=18 в N2-(N')y, N2. обозначает положение 19, и положения 1-18 включены в (N')18.
[0040] Согласно некоторым вариантам реализации N1 ковалентно связано с (N)x и не соответствует РНК-мишени. Согласно различным вариантам реализации N1 ковалентно связано с (N)x и представляет собой ДНК группу, комплементарную РНК-мишени.
[0041] Согласно некоторьм вариантам реализации уридин в положении 1 антисмысловой цепи замещен на N1, выбранное из аденозина, дезоксиаденозина, дезоксиуридина (dU), риботимидина или дезокситимидина. Согласно различным вариантам реализации N1 выбирают из аденозина, дезоксиаденозина или дезоксиуридина.
[0042] Согласно некоторым вариантам реализации гуанозин в положении 1 антисмысловой цепи замещен на N1, выбранное из аденозина, дезоксиаденозина, уридина, дезоксиуридина, риботимидина или дезокситимидина. Согласно различным вариантам реализации N1 выбирают из аденозина, дезоксиаденозина, уридина или дезоксиуридина.
[0043] Согласно некоторым вариантам реализации цитидин в положении 1 антисмысловой цепи замещен на N1, выбранный из аденозина, дезоксиаденозина, уридина, дезоксиуридина, риботимидина или дезокситимидина. Согласно различным вариантам реализации N1 выбирают из аденозина, дезоксиаденозина, уридина или дезоксиуридина.
[0044] Согласно некоторым вариантам реализации аденозин в положении 1 антисмысловой цепи замещен на N1, выбранный из дезоксиаденозина, дезоксиуридина, риботимидина или дезокситимидина. Согласно различным вариантам реализации N1 выбирают из дезоксиаденозина или дезоксиуридина.
[0045] Согласно некоторым вариантам реализации N1 и N2 образуют пару оснований между уридином или дезоксиуридином, и аденозином или дезоксиаденозином. Согласно другим вариантам реализации N1 и N2 образуют пару оснований между дезоксиуридином и аденозином.
[0046] Согласно некоторым вариантам реализации двухцепочечная молекула нуклеиновой кислоты представляет собой siРНК, siНК или miPHK. Двухцепочечные молекулы нуклеиновой кислоты согласно настоящему описанию также обозначают термином «дуплексы».
[0047] Согласно некоторым вариантам реализации (N)x включает антисмысловой олигонуклеотид, представленный в Таблице А-18. Согласно некоторым вариантам реализации x=y=18, и N1-(N)x включает антисмысловой олигонуклеотид, представленный в Таблице А-18. Согласно некоторым вариантам реализации x=y=19 или x=y=20. Согласно некоторым предпочтительным вариантам реализации х=у=18. Согласно некоторым вариантам реализации х=y=18, и последовательности N1-(N)x и N2-(N')y выбирают из пар олигонуклеотидов, представленных в Таблице А-18. Согласно некоторым вариантам реализации х=y=18, и последовательности N1-(N)x и N2-(N')y выбирают из пар олигонуклеотидов, представленных в Таблицах D и Е. Согласно некоторым вариантам реализации антисмысловую цепь и смысловую цепь выбирают из пар последовательностей, представленных в SERPINH1_2 (SEQ ID NOS:60 и 127), SERPINH1_6 (SEQ ID NOS:63 и 130), SERPINH1_1 (SEQ ID NOS:68 и 135), SERPINH1_3 (SEQ ID NOS:69 и 136), SERPINH1_45 (SEQ ID NOS:97 и 164), SERPINH1_45a (SEQ ID NOS:98 и 165), SERPINH1_51 (SEQ ID NOS:101 и 168), SERPINH1_51a (SEQ ID NOS:105 и 172), SERPINH1_52 (SEQ ID NOS:102 и 169) или SERPINH1_86 (SEQ ID NOS:123 и 190). Согласно некоторым предпочтительным вариантам реализации антисмысловую цепь и смысловую цепь выбирают из пар последовательностей, представленных в SERPINH1_2 (SEQ ID NOS:60 и 127), SERPINH1_6 (SEQ ID NOS:63 и 130), SERPINH1_45a (SEQ ID NOS:98 и 165), SERPINH1_51 (SEQ ID NOS:101 и 168) и SERPINH1_51a (SEQ ID NOS:105 и 172).
[0048] Согласно некоторым предпочтительным вариантам реализации антисмысловую цепь и смысловую цепь выбирают из пар последовательностей, представленных в SERPINH1_2 (SEQ ID NOS:60 и 127). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_6 (SEQ ID NOS:63 и 130). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_11(SEQ ID NOS:68 и 135). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_13 (SEQ ID NOS:69 и 136). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_45 (SEQ ID NOS: 97 и 164). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_45a (SEQ ID NOS: 98 и 165). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_51 (SEQ ID NOS: 101 и 168). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_51a (SEQ ID NOS: 105 and 172). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_52 (SEQ ID NOS: 102 и 169). Согласно некоторым вариантам реализации антисмысловая и смысловая цепи представляют собой пары последовательностей, представленные в SERPINH1_86 (SEQ ID NOS: 123 и 190). Согласно некоторым предпочтительным вариантам реализации антисмысловую цепь и смысловую цепь выбирают из пар последовательностей, представленных в SERPINH1_2 (SEQ ID NOS: 60 и 127), SERPINH1_6 (SEQ ID NOS: 63 и 130), SERPINH1_45a (SEQ ID NOS: 98 и 165), SERPINH1_51 (SEQ ID NOS: 101 и 168) и SERPINH1_51a (SEQ ID NOS: 105 и 172).
[0049] Согласно некоторым вариантам реализации N1 и N2 образуют пару оснований по Уотсону-Крику. Согласно другим вариантам реализации N1 и N2 образуют пару оснований не по Уотсону-Крику. Согласно некоторым вариантам реализации N1 представляет собой модифицированный рибоаденозин или модифицированный рибоуридин.
[0050] Согласно некоторым вариантам реализации N1 и N2 образуют пару оснований по Уотсону-Крику. Согласно другим вариантам реализации N1 и N2 образуют пару оснований не по Уотсону-Крику. Согласно некоторым вариантам реализации N1 выбирают из группы, состоящей из рибоаденозина, модифицированного рибоаденозина, дезоксирибоаденозина, модифицированного дезоксирибоаденозина. Согласно другим вариантам реализации N1 выбирают из группы, состоящей из рибоуридина, дезоксирибоуридина, модифицированного рибоуридина и модифицированного дезоксирибоуридина.
[0051] Согласно некоторым вариантам реализации положение 1 в антисмысловой цепи (5′-конец) содержит дезоксирибоуридин (dU) или аденозин. Согласно некоторым вариантам реализации N1 выбирают из группы, состоящей из рибоаденозина, модифицированного рибоаденозина, дезоксирибоаденозина, модифицированного дезоксирибоаденозина, и N2 выбирают из группы, состоящей из рибоуридина, дезоксирибоуридина, модифицированного рибоуридина и модифицированного дезоксирибоуридина. Согласно некоторым вариантам реализации N1 выбирают из группы, состоящей из рибоаденозина и модифицированного рибоаденозина, и N2 выбирают из группы, состоящей из рибоуридина и модифицированного рибоуридина.
[0052] Согласно некоторым вариантам реализации N1 выбирают из группы, состоящей из рибоуридина, дезоксирибоуридина, модифицированного рибоуридина и модифицированного дезоксирибоуридина, и N2 выбирают из группы, состоящей из рибоаденозина, модифицированного рибоаденозина, дезоксирибоаденозина, модифицированного дезоксирибоаденозина. Согласно некоторым вариантам реализации N1 выбирают из группы, состоящей из рибоуридина и дезоксирибоуридина, и N2 выбирают из группы, состоящей из рибоаденозина и модифицированного рибоаденозина. Согласно некоторым вариантам реализации N1 представляет собой рибоуридин, и N2 представляет собой рибоаденозин. Согласно некоторым вариантам реализации N1 представляет собой дезоксирибоуридин, и N2 представляет собой рибоаденозин.
[0053] Согласно некоторым вариантам реализации Структуры (А2), N1 включает модифицированный рибоурацил с 2'ОМе-сахаром или модифицированный рибоаденозин с 2'ОМе-сахаром. Согласно некоторым вариантам реализации структуры (А), N2 включает модифицированный рибонуклеотид или дезоксирибонуклеотид с 2'ОМе-сахаром.
[0054] Согласно некоторым вариантам реализации Структуры (А2), N1 включает модифицированный рибоурацил с 2'ОМе-сахаром или модифицированный рибоцитозин с 2'ОМе-сахаром. Согласно некоторым вариантам реализации структуры (А), N2 включает модифицированный рибонуклеотид с 2'ОМе сахаром.
[0055] Согласно некоторым вариантам реализации N и N' представляет собой немодифицированный нуклеотид. Согласно некоторым вариантам реализации по меньшей мере один из N или N' включает химически модифицированный нуклеотид или нетрадиционную группу. Согласно некоторым вариантам реализации нетрадиционную группу выбирают из зеркального нуклеотида, остатка рибозы без основания и остатка дезоксирибозы без основания. Согласно некоторым вариантам реализации нетрадиционная группа представляет собой зеркальный нуклеотид, предпочтительно группу L-ДНК. Согласно некоторым вариантам реализации по меньшей мере один из N или N′ включает модифицированный рибонуклеотид с 2′ОМе сахаром.
[0056] Согласно некоторым вариантам реализации последовательность (N′)y полностью комплементарна последовательности (N)x. Согласно другим вариантам реализации последовательность (N′)y существенно комплементарна последовательности (N)x.
[0057] Согласно некоторым вариантам реализации (N)x включает антисмысловую последовательность, которая полностью комплементарна приблизительно 17-39 следующим друг за другом нуклеотидам в мРНК-мишени. Согласно другим вариантам реализации (N)x включает антисмысловую последовательность, которая существенно комплементарна приблизительно 17-39 следующим друг за другом нуклеотидам в мРНК-мишени.
[0058] Согласно некоторым вариантам реализации Структуры А1 и Структуры А2 соединение имеет тупые концы, например, при этом как Z, так и Z′ отсутствуют. Согласно другому варианту реализации по меньшей мере один из Ζ или Z′ присутствует.Ζ и Z′ независимо друг от друга включают один или несколько ковалентно связанных модифицированных и/или немодифицированных нуклеотидов, включая дезоксирибонуклеотиды и рибонуклеотиды или нетрадиционную группу, например, инвертированную группу дезоксирибозы или рибозы без основания; не-нуклеотидную С3, С4 или С5-группу, амино-6 группу, зеркальный нуклеотид т.п. Согласно некоторым вариантам реализации Ζ и Z′ независимо друг от друга включают С3-группу или амино-С6-группу. Согласно некоторым вариантам реализации Z′ отсутствует, и Ζ присутствует и включает не-нуклеотидную С3-группу. Согласно некоторым вариантам реализации Ζ отсутствует, и Z′ присутствует и включает не-нуклеотидную С3-группу.
[0059] Согласно некоторым вариантам реализации Структуры А1 и Структуры А2, каждый N состоит из немодифицированного рибонуклеотида. Согласно некоторым вариантам реализации Структуры А1 и Структуры А2, каждая N′ состоит из немодифицированного нуклеотида. Согласно предпочтительным вариантам реализации, по меньшей мере один из N и N' является немодифицированным рибонуклеотидом или нетрадиционной группой.
[0060] Согласно другим вариантам реализации соединение Структуры А1 или Структуры А2 включает по меньшей мере один рибонуклеотид, модифицированный в остатке сахара. Согласно некоторым вариантам реализации соединение включает модификацию в положении 2′ остатка сахара. Согласно некоторым вариантам реализации модификация в положении 2′ включает наличие амино, фтор, алкокси или алкильной группы. Согласно некоторым вариантам реализации модификация 2′ включает алкоксигруппу. Согласно предпочтительным вариантам реализации алкокси-группа представляет собой метоксигруппу (также известную как 2′-O-метил; 2′ОМе; 2′-ОСН3). Согласно некоторым вариантам реализации соединение нуклеиновой кислоты включает чередующиеся рибонуклеотиды, модифицированные 2′ОМе по остатку сахара, на антисмысловой и/или смысловой цепи. Согласно другим вариантам реализации соединение включает модифицированные рибонуклеотиды с 2′ОМе-сахаром в антисмысловой цепи, только (Ν)x или Ν1-(Ν)x. Согласно некоторым вариантам реализации средний рибонуклеотид антисмысловой цепи, например, рибонуклеотид в положении 10 в 19-мерной цепи, является не модифицированным. Согласно различным вариантам реализации соединение нуклеиновой кислоты включает по меньшей мере 5 чередующихся модифицированных и немодифицированных с 2′ОМе-сахаром рибонуклеотидов. Согласно дополнительным вариантам реализации соединение Структуры А1 или Структуры А2 включает модифицированные рибонуклеотиды в чередующихся положениях, при этом каждый рибонуклеотид на 5′ и 3′-концах (Ν)x или Ν1-(Ν)x модифицированы в их остатках сахара и каждый рибонуклеотид на 5′- и 3′-концах (N′)y или N2-(N′)y не модифицированы в их остатках сахара.
[0061] Согласно некоторым вариантам реализации двухцепочечная молекула включает одну или более из следующих модификаций
a) N в по меньшей мере одном из положений 5, 6, 7, 8 или 9 от 5′-конца антисмысловой цепи выбирают из 2′5′ нуклеотида или зеркального нуклеотида;
b) N′ в по меньшей мере одном из положений 9 или 10 от 5′-конца смысловой цепи выбирают из 2′5′ нуклеотида и псевдоуридина; и
c) N′ в 4, 5 или 6 последовательных положениях на 3′-конце (N′)y содержит 2′5′ нуклеотид.
[0062] Согласно некоторым вариантам реализации двухцепочечная молекула включает комбинацию следующих модификаций:
a) антисмысловая цепь включает 2′5′ нуклеотид или зеркальный нуклеотид в по меньшей мере одном из положений 5, 6, 7, 8 или 9 от 5′-конца; и
b) смысловая цепь включает по меньшей мере один из 2′5′ нуклеотидов и псевдоуридина в положениях 9 или 10 от 5′-конца.
[0063] Согласно некоторым вариантам реализации двухцепочечная молекула включает комбинацию следующих модификаций:
a) антисмысловая цепь включает 2′5′ нуклеотид или зеркальный нуклеотид в по меньшей мере одном из положений 5, 6, 7, 8 или 9 от 5′-конца; и
b) смысловая цепь включает 4, 5 или 6 последовательных 2′5′ нуклеотидов в предпоследнем положении на 3′-конце или в 3′-концевом положении.
[0064] Согласно некоторым вариантам реализации смысловая цепь [(Ν)x или N1-(N)x] включает 1, 2, 3, 4, 5, 6, 7, 8 или 9 модифицированных рибонуклеотидов с 2′ОМе-сахаром. Согласно некоторым вариантам реализации антисмысловая цепь включает модифицированные рибонуклеотиды с 2′ОМе-сахаром в положениях 2, 4, 6, 8, 11, 13, 15, 17 и 19. Согласно другим вариантам реализации антисмысловая цепь включает модифицированные рибонуклеотиды с 2′ОМе-сахаром в положениях 1, 3, 5, 7, 9, 11, 13, 15, 17 и 19. Согласно другим вариантам реализации антисмысловая цепь включает модифицированные рибонуклеотиды с 2′ОМе-сахаром в положениях 3, 5, 7, 9, 11, 13, 15, 17 и 19. Согласно некоторым вариантам реализации антисмысловая цепь включает один или несколько модифицированных пиримидинов с 2′ОМе-сахаром. Согласно некоторым вариантам реализации все пиримидиновые нуклеотиды в антисмысловой цепи являются модифицированными по 2′ОМе-остаткам сахара. Согласно некоторым вариантам реализации смысловая цепь включает модифицированные пиримидины с 2′ОМе-сахаром.
[0065] Согласно некоторым вариантам реализации Структуры А1 и Структуры А2, ни смысловая цепь, ни антисмысловая цепь не фосфорилированы на 3′- и 5′-концах. Согласно другим вариантам реализации и смысловая цепь, и антисмысловая цепь фосфорилированы на 3′-концах.
[0066] Согласно некоторым вариантам реализации Структуры А1 и Структуры А2 (N′)y включает по меньшей мере одну нетрадиционную группу, выбранную из зеркального нуклеотида, 2′5′ нуклеотида и TNA. Согласно некоторым вариантам реализации нетрадиционная группа представляет собой зеркальный нуклеотид. Согласно различным вариантам реализации зеркальный нуклеотид выбирают среди L-рибонуклеотида (L-PHK) и L-дезоксирибонуклеотида (L-ДНК). Согласно предпочтительным вариантам реализации зеркальный нуклеотид представляет собой L-ДНК. Согласно некоторым вариантам реализации смысловая цепь содержит нетрадиционную группу в положении 9 или 10 (от 5'-конца). Согласно предпочтительным вариантам реализации смысловая цепь включает нетрадиционную группу в положении 9 (от 5'-конца). Согласно некоторым вариантам реализации смысловая цепь имеет 19 нуклеотидов в длину и содержит 4, 5 или 6 последовательных функциональных нестандартных групп в положениях 15 (от 5'-конца). Согласно некоторым вариантам реализации смысловая цепь включает 4 последовательных 2'5' рибонуклеотида в положениях 15, 16, 17 и 18. Согласно некоторым вариантам реализации смысловая цепь включает 5 последовательных 2'5' рибонуклеотидов в положениях 15, 16, 17, 18 и 19. Согласно различным вариантам реализации смысловая цепь также содержит Z'. Согласно некоторым вариантам реализации Z' включает С3ОН группу или C3Pi группу.
[0067] Согласно некоторым вариантам реализации Структуры A1 (N')y включает по меньшей мере одну L-ДНК группу. Согласно некоторым вариантам реализации х=y=19, и (N')y состоит из немодифицированных рибонуклеотидов в положениях 1-17 и 19 и одной L-ДНК в предпоследнем положении на 3'-конце (положение 18). Согласно другим вариантам реализации х=y=19, и (N')y состоит из немодифицированных рибонуклеотидов в положениях 1-16 и 19 и двух последовательных L-ДНК в предпоследнем положении на 3'-конце (положения 17 и 18). Согласно различным вариантам реализации нетрадиционная группа представляет собой нуклеотид, соединенный с прилегающим нуклеотидом посредством 2'-5' межнуклеотидной фосфатной связи. Согласно различным вариантам реализации (N')y включает 2, 3, 4, 5, или 6 последовательных рибонуклеотидов на 3'-конце, соединенных 2'-5' межнуклеотидными связями. Согласно одному варианту реализации четыре последовательных нуклеотида на 3'-конце (N')y соединены посредством трех 2'-5' фосфодиэфирных связей, при этом один или несколько 2'-5' нуклеотидов, которые образуют 2'-5' фосфодиэфирные связи, также включает модификацию 3'-O-метил сахар (3'ОМе). Предпочтительно 3'-концевой нуклеотид (N')y содержит модификацию 2'ОМе сахар. Согласно некоторым вариантам реализации х=y=19, и (N')y включает два или более последовательных нуклеотидов в положениях 15, 16, 17, 18 и 19, и включает нуклеотид, соединенный с прилегающим нуклеотидом 2'-5' межнуклеотидной связью (2'-5' нуклеотид). Согласно различным вариантам реализации нуклеотид, образующий 2'-5' межнуклеотидную связь, включает нуклеотид 3'дезоксирибозы или 3' метокси нуклеотид (3' Н или 3'ОМе вместо 3' ОН). Согласно некоторым вариантам реализации х=y=19, и (N')y включает 2'-5' нуклеотиды в положениях 15, 16 и 17, таким образом, что прилегающие нуклеотиды соединены 2'-5' межнуклеотидной связью между положениями 15-16, 16-17 и 17-18; или в положениях 15, 16, 17, 18 и 19, таким образом, что прилегающие нуклеотиды соединены 2'-5' межнуклеотидной связью между положениями 15-16, 16-17, 17-18 и 18-19, и 3'ОН доступен в 3'-концевом нуклеотиде или в положениях 16, 17 и 18 таким образом, что прилегающие нуклеотиды соединены 2'-5' межнуклеотидной связью между положениями 16-17, 17-18 и 18-19. Согласно некоторым вариантам реализации х=y=19, и (N')y включает 2'-5'нуклеотиды в положениях 16 и 17 или в положениях 17 и 18 или в положениях 15 и 17,таким образом, что прилегающие нуклеотиды соединены 2'-5' межнуклеотидной связью между положениями 16-17 и 17-18 или между положениями 17-18 и 18-19 или между положениями 15-16 и 17-18, соответственно. Согласно другим вариантам реализации пиримидиновые нуклеотиды (rU, rC) в (N')y замещены на нуклеотиды, соединенные с прилегающим нуклеотидом 2'-5' межнуклеотидной связью. Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, показанных на SERPINH1_4, SERPINH1_12, SERPINH1_18, SERPINH1_30, SERPINH1_58 или SERPINH1_88, и x=y-19, и (N')y содержит пять последовательных нуклеотидов на 3'-конце, соединенных четырьмя 2'-5' связями, в частности, связями между положениями нуклеотидов 15-16, 16-17, 17-18 и 18-19.
[0068] Согласно некоторым вариантам реализации связи включают фосфодиэфирные связи. Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_4, SERPINH1_12, SERPINH1_8, SERPINH1_30, SERPINH1_58 или SERPINH1_88, и х=y=19, и (N')у содержит пять последовательных нуклеотидов на 3'-конце, соединенных четырьмя 2'-5' связями, и дополнительно также включает Z' и z' независимо выбранные из инвертированной группы без основания, и С3 алкил [С3; 1,3-пропандиол моно(дигидрофосфат)] блокирующей группы. С3 алкил блокирующая группа ковалентно соединена с 3' или 5'-концевым нуклеотидом. Согласно некоторым вариантам реализации 3'-концевая С3 блокирующая группа также содержит 3'-фосфат. Согласно некоторым вариантам реализации 3'-концевая С3 блокирующая группа также содержит 3'-концевую гидроксигруппу.
[0069] Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_4, SERPINH1_12, SERPINH1_18, SERPINH1_30, SERPINH1_58 или SERPINH1_88, и x=y=19, и (N')y включает L-ДНК в положении 18; и (N')y дополнительно также включает Z' и z', независимо выбранные из инвертированной группы без основания, и С3 алкил [С3; 1,3-пропандиол моно(дигидрофосфат)] блокирующей группы.
[0070] Согласно некоторым вариантам реализации (N')y включает 3'-концевой фосфат. Согласно некоторым вариантам реализации (N')y включает 3'-концевой гидроксил.
[0071] Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_4, SERPINH1_12, SERPINH1_18, SERPINH1_30, SERPINH1_58 или SERPINH1_88, и x=y=19, и (N)x включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях 1, 3, 5, 7, 9, 11, 13, 15, 17, 19 или в положениях 2, 4, 6, 8, 11, 13, 15, 17, 19. Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_4, SERPINH1_12, SERPINH1_18, SERPINH1_30, SERPINH1_58 и SERPINH1_88, и x=y=19, и (N)x включает модифицированные пиримидины с 2'ОМе сахаром. Согласно некоторым вариантам реализации все пиримидины в (N)x включают модификацию сахара 2'ОМе.
[0072] Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_2, SERPINH1_6, SERPINH1_11, SERPINH1_13, SERPINH1_45, SERPINH1_45a, SERPINH1_51, SERPIN51a, SERPINH1_52 или SERPINH1_86, и х=y=18, и N2 представляет собой группу рибоаденозина.
[0073] Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_2, SERPINH1_6, SERPINH1_11, SERPINH1_13, SERPINH1_45, SERPINH1_45a, SERPINH1_51, SERPIN51a, SERPINH1_52 или SERPINH1_86, и x=y=18, и N2-(N')y включает пять последовательных нуклеотидов на 3'-конце, соединенных четырьмя 2'-5'-связями, в частности, связями между нуклеотидов в положениях 15-16, 16-17, 17-18 и 18-19. Согласно некоторым вариантам реализации связи включают фосфодиэфирные связи.
[0074] Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_2, SERPINH1_6, SERPINH1_11, SERPINH1_13, SERPINH1_45, SERPINH1_45a, SERPINH1_51, SERPINH1_51a, SERPINH1_52 или SERPINH1_86, и x=y=18, и N2-(N′)y включает пять последовательных нуклеотидов на 3′-конце, соединенных четырьмя 2′-5′-связями, и дополнительно также включает Z′ и z′, независимо выбранные из инвертированной группы без основания, и С3 алкил [С3; 1,3-пропандиол моно(дигидрофосфат)] блокирующей группы.
[0075] Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_2, SERPINH1_6, SERPINH1_11, SERPINH1_13, SERPINH1_45, SERPINH1_45a, SERPINH1_51, SERPINH1_51a, SERPINH1_52 или SERPINH1_86, и x=y=18, и N2-(N′)y включает L-ДНК в положении 18; и (N′)y дополнительно также включает Z′ и z′, независимо выбранные из инвертированной группы, лишенной основания, и С3 алкил [С3; 1,3-пропандиол моно(дигидрофосфат)] блокирующей группы.
[0076] Согласно некоторым вариантам реализации N2-(N')y содержит 3′-концевой фосфат. Согласно некоторым вариантам реализации N2-(N′)y содержит 3′-концевой гидроксил.
[0077] Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_2, SERPINH1_6, SERPINH1_11, SERPINH1_13, SERPINH1_45, SERPINH1_45a, SERPINH1_51, SERPINH1_51a, SERPINH1_52 или SERPINH1_86, и x=y=18, и N′-(N)x включает модифицированные рибонуклеотиды с 2′ОМе сахаром в положениях 1, 3, 5, 7, 9, 11, 13, 15,17, 19 или в положениях 1, 3, 5, 9, 11,13,15,17, 19, или в положениях 3, 5, 9,11,13,15, 17, или в положениях 2, 4, 6, 8, 11, 13, 15, 17, 19. Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_2, SERPINH1_6, SERPINH1_11, SERPINH1_13, SERPINH1_45, SERPINH1_45a, SERPINH1_51, SERPINH1_51a, SERPINH1_52 или SERPINH1_86, и x=y=18, и N1-(N)x включает модифицированные рибонуклеотиды с 2′ОМе сахаром в положениях 11, 13, 15, 17 и 19 (от 5′-конца). Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_2, SERPINH1_6, SERPINH1_11, SERPINH1_13, SERPINH1_45, SERPINH1_45a, SERPINH1_51, SERPINH1_51a, SERPINH1_52 или SERPINH1_86, и x=y=18, и N′-(N)x ключает модифицированные рибонуклеотиды с 2′ОМе сахаром в положениях 1, 3, 5, 7, 9, 11, 13, 15, 17, 19 или в положениях 3, 5, 7, 9, 11, 13, 15, 17, 19. Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_2, SERPINH1_6, SERPINH1_11, SERPINH1_13, SERPINH1_45, SERPINH1_45a, SERPINH1_51, SERPINH1_51a, SERPINH1_52 или SERPINH1_86, и x=y=18, и N1-(N)x включает модифицированные рибонуклеотиды с 2′ОМе сахаром в положениях 2, 4, 6, 8, 11,13, 15,17, 19.
[0078] Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар последовательностей, представленных на SERPINH1_2, SERPINH1_6, SERPINH1_11, SERPINH1_13, SERPINH1_45, SERPINH1_45a, SERPINH1_51, SERPINH1_51a, SERPINH1_52 или SERPINH1_86, и x=y=18, и N1-(N)x включает модифицированные пиримидины с 2′ОМе сахаром. Согласно некоторым вариантам реализации все пиримидины в (N)x включают модификацию сахара 2′ОМе. Согласно некоторым вариантам реализации антисмысловая цепь также включает L-ДНК или 2′-5′ нуклеотид в положении 5, 6 или 7 (5′>3′). Согласно другим вариантам реализации антисмысловая цепь также включает рибонуклеотид, который образует 2′5′ межнуклеотидную связь между рибонуклеотидами в положениях 5-6 или 6-7 (5′>3′)
[0079] Согласно дополнительным вариантам реализации Ν1-(Ν)x также включает Ζ, при этом Ζ включает ненуклеклеотидный выступающий конец. Согласно некоторым вариантам реализации не состоящий из нуклеотидов выступающий конец представляет собой С3-С3 [1,3-пропандиол моно(дигидрофосфат)]2.
[0080] Согласно некоторым вариантам реализации Структуры А2, (N′)y включает по меньшей мере одну группу L-ДНК. Согласно некоторым вариантам реализации х=у=18, и (N′)y состоит из немодифицированных нуклеотидов в положениях 1-16 и 18 и одной L-ДНК в предпоследнем положении на 3′-конце (положение 17). Согласно другим вариантам реализации х=у=18, и (N′)y состоит из немодифицированных нуклеотидов в положении 1-15 и 18 и двух последовательных L-ДНК в предпоследнем положении на 3′-конце (положения 16 и 17). Согласно различным вариантам реализации нетрадиционная группа представляет собой нуклеотид, соединенный с прилегающим нуклеотидом с помощью 2′-5′ межнуклеотидной фосфорной связи. Согласно различным вариантам реализации (N′)y включает 2, 3, 4, 5 или 6 последовательных рибонуклеотидов на 3′-конце, соединенных 2′-5′ межнуклеотидными связями. Согласно одному варианту реализации четыре последовательных нуклеотида на 3′-конце (N′)y соединены с помощью трех 2'-5' фосфодиэфирных связей, при этом один или несколько 2'-5' нуклеотидов, которые образуют 2'-5' фосфодиэфирные связи, также включают модификацию сахара 3'-O-метил (3'ОМе). Предпочтительно 3'-концевой нуклеотид (N')y включает модификацию 2'ОМе сахара. Согласно некоторым вариантам реализации x=y=18, и в (N')y два или несколько последовательных нуклеотидов в положениях 14, 15, 16, 17 и 18 включают нуклеотид, соединенный с прилегающим нуклеотидом с помощью 2'-5' межнуклеотидной связи. Согласно различным вариантам реализации, нуклеотид, образующий 2'-5' межнуклеотидную связь, включает 3' дезоксирибозу или 3' метоксинуклеотид. Согласно некоторым вариантам реализации х=y=18, и (N')y включает нуклеотид, соединенный с прилегающим нуклеотидом с помощью 2'-5' межнуклеотидной связи между положениями 15-16, 16-17 и 17-18 или между положениями 16-17 и 17-18. Согласно некоторым вариантам реализации х=y=18, и (N')y включает нуклеотид, соединенный с прилегающим нуклеотидом с помощью 2'-5' межнуклеотидной связи между положениями 14-15, 15-16, 16-17 и 17-18 или между положениями 15-16, 16-17 и 17-18 или между положениями 16-17 и 17-18 или между положениями 17-18 или между положениями 15-16 и 17-18. Согласно другим вариантам реализации пиримидиновые нуклеотиды (rU, rC) в (N')y замещены на нуклеотиды, соединенные с прилегающими нуклеотидами 2'-5' межнуклеотидной связью.
[0081] Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар олигонуклеотидов, представленных в Таблице А-18 и обозначенных в настоящем описании как SERPINH1_2 (SEQ ID NOS:60 и 127), SERPINH1_6 (SEQ ID NOS:63 и 130), SERPINH1_45a (SEQ ID NOS:98 и 165), SERPINH1_51 (SEQ ID NOS:101 и 168) и SERPINH1_51a (SEQ ID NOS:105 и 172).
[0082] Согласно некоторым вариантам реализации двухцепочечная молекула нуклеиновой кислоты включает антисмысловую цепь, представленную на SEQ ID NO:127, и смысловую цепь, представленную на SEQ ID NO:60; обозначенные в настоящем описании как SERPINH1_2. Согласно некоторым вариантам реализации двухцепочечная молекула нуклеиновой кислоты имеет структуру
в которой каждая «|» обозначает спаривание оснований между рибонуклеотидами;
в которой А, С, G, U независимо друг от друга представляют собой модифицированный или немодифицированный рибонуклеотид, или нетрадиционную группу;
в которой Z и Z' могут независимо друг от друга присутствовать или отсутствовать, но при наличии независимо друг от друга представляют собой 1-5 последовательных нуклеотидов или не являющихся нуклеотидами групп или их сочетание, ковалентно соединенные с 3'-концом цепи, в которой они находятся; и
в которой z" может присутствовать или отсутствовать, но при наличии представляет собой блокирующую группу, ковалентно сединенную с 5'-концом N2-(N')y.
[0083] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:127) включает один или несколько модифицированных пиримидинов и/или пуринов с 2'ОМе сахаром, 2'-5' рибонуклеотид в положении5, 6, 7 или 8, и нуклеотидный или не состоящий из нуклеотидов 3'-выступающий конец. Согласно некоторым вариантам реализации смысловая цепь (SEQ ID NO:60) включает 4 или 5 последовательных 2'5' нуклеотидов на 3'-концевом или в предпоследнем положениях, нуклеотидную или не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и блокирующую группу, ковалентно соединенную с 5'-концом. Согласно другим вариантам реализации смысловая цепь (SEQ ID NO:60) включает один или несколько 2'ОМе пиримидинов, нуклеотидную или не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и блокирующую группу, ковалентно соединенную с 5'-концом.
[0084] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:127) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 15, 17 и 19, 2'-5' рибонуклеотид в положении 7, и C3Pi-С3ОН группу, ковалентно соединенную с 3'-концом; и смысловую цепь (SEQ ID NO:60) выбирают из смысловой цепи, которая включает
а) 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19, С3ОН не состоящий из нуклеотидов выступающий 3'-конец; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; или
б) 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19, 3'-концевой фосфат, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; или
в) модифицированные рибонуклеотиды 2'ОМе сахаром в положениях (5'>3') 5, 7, 13, и 16; 2'5' рибонуклеотид в положении 18; С3-ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; или
г) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 7, 13, 16 и 18; 2'5' рибонуклеотид в положении 9; С3ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; или
д) рибонуклеотиды 2'-5' в положениях 15, 16, 17, 18 и 19: С3-Pi группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0085] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:127) включает модифицированные нуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 15, 17, 19, 2'-5' рибонуклеотид в положении 7 и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; и смысловая цепь (SEQ ID NO:60) включает 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19: С3 выступающий 3'-конец; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0086] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:127) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 15, 17, 19, 2'-5' рибонуклеотид в положении 7 и C3Pi-C3OH выступающий 3'-конец; и смысловая цепь (SEQ ID NO:60) включает 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19: 3'-концевой фосфат; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0087] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:127) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 15, 17, 19, 2'-5' рибонуклеотид в положении 7 и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; и смысловая цепь (SEQ ID NO:60) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях(5'>3') 5, 7, 13 и 16; 2'-5' рибонуклеотид в положении 18; С3-ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0088] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:127) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 15, 17, 19, 2'-5' рибонуклеотид в положении 7 и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; и смысловая цепь (SEQ ID NO:60) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 7, 13, 16 и 18; 2'-5' рибонуклеотид в положении 9; С3-ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0089] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:127) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 15, 17, 19, 2'-5' рибонуклеотид в положении 7 и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; и смысловая цепь (SEQ ID NO:60) включает 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19: С3-Pi группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0090] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:127) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 13, 15, 17, 19 и С3-С3 выступающий 3'-конец; и смысловая цепь (SEQ ID NO:60) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 7, 9, 13, 16 и 18; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0091] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:60) включает 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19: 3'-концевой фосфат, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом, и антисмысловая цепь (SEQ ID NO:127) включает антисмысловую цепь, выбранную из:
а) модифицированных рибонуклеотидов с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 7, 9, 11, 13, 15, 17, 19 и C3Pi-C3OH группы, ковалентно соединенной с 3'-концом; или
б) модифицированных рибонуклеотидов с 2'ОМе сахаром в положениях (5'>3') 1, 3, 6, 8, 10, 12, 14, 17, 18 и C3Pi-C3OH группы, ковалентно соединенной с 3'-концом.
[0092] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, которая включает антисмысловую цепь, изложенную в SEQ ID NO:130, и смысловую цепь, изложенную в SEQ ID NO:63; обозначаемые в настоящем описании как SERPINH1_6. Согласно некоторым вариантам реализации указанный дуплекс содержит структуру
на которой «|» обозначает спаривание оснований между рибонуклеотидами;
на которой А, С, G, U независимо друг от друга представляют собой немодифицированный или модифицированный рибонуклеотид или нетрадиционную группу;
на которой Z и Z' могут независимо друг от друга присутствовать или отсутствовать, но при наличии представляют собой независимо 1-5 последовательных нуклеотидов или не являющихся нуклеотидами групп или их комбинацию, ковалентно соединенных с 3'-концом цепи, в которой они находятся; и
на которой z" может присутствовать или отсутствовать, но при наличии представляет собой блокирующую группу, ковалентно соединенную с 5'-концом N2-(N')y.
[0093] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:63) включает один или больше модифицированных пиримидина с 2'ОМе сахаром; нуклеотидный или не состоящий из нуклеотидов выступающий 3'-конец; и блокирующую группу, ковалентно соединенную с 5'-концом. Согласно некоторым вариантам реализации антисмысловая цепь (SEQ ID NO:130) включает один или более модифицированных пиримидина с 2'ОМе сахаром, нуклеотидную или не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и блокирующую группу, ковалентно соединенную с 5'-концом.
[0094] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:63) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 14 и 18; С3ОН или C3Pi группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловую цепь (SEQ ID NO:130) выбирают из антисмысловой цепи, которая включает:
а) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 13, 15 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; или
б) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 7, 9, 12, 13 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; или
в) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 9, 11, 13, 15 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; или
г) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 9, 11, 13, 15 и 17; dU в положении 1; 2'-5' рибонуклеотид в положении7; и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0095] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:63) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 14 и 18; С3-ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:130) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 13, 15 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0096] Согласно некоторым вариантам реализации обеспечивается молекула дуплекса олигонуклеотидов, в которой смысловая цепь (SEQ ID NO:63) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 14 и 18 и, возможно, в положении 2; С3-ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:130) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 7, 9, 12, 13 и 17; 2'-5' рибонуклеотид в положении 7; и C3:Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0097] Согласно некоторым вариантам реализации обеспечивается молекула дуплекса олигонуклеотидов, в которой смысловая цепь (SEQ ID NO:63) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 14 и 18; С3-ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловую цепь (SEQ ID NO:130) выбирают из антисмысловой цепи, которая включает:
а) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 13, 15 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3Pi или C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; или
б) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 7, 9, 12, 13 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3Pi или C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0098] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:63) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 14 и 18; С3-ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:130) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 13. 15 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0099] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:63) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 14 и 18; С3-ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:130) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5,7, 9, 12,13 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH выступающий 3'-конец.
[0100] Согласно некоторым вариантам реализации дуплекс включает антисмысловую цепь, изложенную на SEQ ID NO:165, и смысловую цепь, изложенную на SEQ ID NO:98; обозначаемую в настоящем описании как SERPINH1_45a. Согласно некоторым вариантам реализации указанная дуплекс содержит структуру
на которой «|» обозначает спаривание оснований между рибонуклеотидами;
на которой А, С, G, U независимо друг от друга представляют собой немодифицированный или модифицированный рибонуклеотид или нетрадиционную группу;
на которой Z и Z' могут независимо друг от друга присутствовать или отсутствовать, но при наличии независимо друг от друга представляет собой 1-5 последовательных нуклеотидных или не являющихся нуклеотидами группы их сочетание, ковалентно соединенные с 3'-концом цепи, в которой они находятся; и
на которой z" может присутствовать или отсутствовать, но при наличии представляет собой блокирующую группу, ковалентно соединенную с 5'-концом N2-(N')y.
[0101] Согласно некоторым вариантам реализации смысловая цепь (SEQ ID NO:98) включает 2'-5' рибонуклеотиды в положениях (5'>3') 15, 16, 17 и 18 или 15, 16, 17, 18 и 19: нуклеотид или не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и блокирующую группу, ковалентно соединенную с 5'-концом. Согласно некоторым вариантам реализации антисмысловая цепь (SEQ ID NO:165) включает модифицированный пиримидин или пурины с 2'ОМе сахаром, 2'-5' нуклеотид в положении5, 6, 7 и 8 (5'>3'); и нуклеотид или не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом.
[0102] Согласно некоторым вариантам реализации смысловая цепь (SEQ ID NO:98) включает 2'-5' рибонуклеотиды в положениях (5'>3') 15, 16, 17, 18 и 19: C3Pi или С3-ОН 3'-концевую не являющуюся нуклеотидом группу и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:165) включает антисмысловую цепь, выбранную из одной из:
а) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 4, 6, 8, 11, 13, 15, 17 и 19; 2'-5' рибонуклеотид в положении 7 и C3Pi-C3Pi или C3Pi-C3OH выступающий 3'-конец; или
б) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 4, 6, 8, 11,13,15,17 и 19 и C3Pi-C3Pi или C3Pi-C3OH выступающий 3'-конец;
в) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 13, 15, 17 и 19; 2'-5' рибонуклеотид в положении 7 и C3Pi-C3Pi или C3Pi-C3OH выступающий 3'-конец; или
г) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 7, 9, 11, 13, 15, 17 и 19 и C3Pi-C3Pi или C3Pi-C3OH выступающий 3'-конец.
[0103] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:98) включает 2'-5' рибонуклеотиды в положениях (5'>3') 15, 16, 17, 18 и 19: С3-ОН 3'-концевую группу и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:165) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 4, 6, 8, 11, 13, 15, 17 и 19; 2'-5' рибонуклеотид в положении 7 и С3Р1-СОН выступающий 3'-конец.
[0104] Согласно некоторым вариантам реализации двухцепочечная молекула нуклеиновой кислоты включает антисмысловую цепь, представленную на SEQ ID NO:168, и смысловую цепь, представленную на SEQ ID NO:101; обозначенные в настоящем описании как SERPINH1_51. Согласно некоторым вариантам реализации указанная двойная спираль содержит структуру
на которой «|» обозначает спаривание оснований между рибонуклеотидами;
на которой А, С, G, U независимо друг от друга представляют собой немодифицированный или модифицированный рибонуклеотид или нетрадиционную группу;
на которой Z и Z' могут независимо друг от друга присутствовать или отсутствовать, но при наличии представляют независимо друг от друга 1-5 последовательных нуклеотидов или не являющихся нуклеотидами групп или их комбинацию, ковалентно соединенных с 3'-концом цепи, в которой они находятся; и
на которой z" может присутствовать или отсутствовать, но при наличии представляет собой блокирующую группу, ковалентно соединенную с 5'-концом N2-(N')y.
[0105] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:101) включает модифицированные пиримидины с 2'ОМе сахаром, возможно, 2'-5' рибонуклеотид в положении 9 или 10; нуклеотид или не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и, возможно, блокирующую группу, ковалентно соединенную с 5'-концом. Согласно некоторым вариантам реализации антисмысловая цепь (SEQ ID NO:168) включает модифицированный пиримидин и/или пурины с 2'ОМе сахаром, 2'-5' нуклеотид в положении5, 6, 7 или 8 (5'>3'); и нуклеотид или не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом.
[0106] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:101) включает модифицированные пиримидины с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, возможно, 2'-5' рибонуклеотид в положении 9 или 10, C3Pi или С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловую цепь (SEQ ID NO:168) выбирают из антисмысловой цепи, которая включает:
а) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 8 и 15, 2'5' рибонуклеотид в положении 6 или 7; выступающий конец C3Pi-C3OH, ковалентно соединенный с 3'-концом; или
б) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 4, 8, 13 и 15, 2'5' рибонуклеотид в положении 6 или 7; выступающий конец C3Pi-C3OH, ковалентно соединенный с 3'-концом; или
в) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1,4, 8, 11 и 15, 2'5' рибонуклеотид в положении 6; выступающий конец C3Pi-C3OH, ковалентно соединенный с 3'-концом; или
г) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 8, 12, 13 и 15; 2'5' рибонуклеотид в положении 6; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0107] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:101) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, возможно, 2'-5' рибонуклеотид в положении 9, С3-ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:168) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 8 и 15, 2'5' рибонуклеотид в положении 6; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0108] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:101) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, возможно, 2'-5' рибонуклеотид в положении 9, С3-ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:168) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 4, 8, 13 и 15, 2'5' рибонуклеотид в положении 6; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0109] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:101) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, 2'-5' рибонуклеотид в положении 9, С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:168) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 4, 8, 11 и 15, 2'5' рибонуклеотид в положении 6; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0110] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:101) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, 2'-5' рибонуклеотид в положении 9, С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3′-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5′-концом; и антисмысловая цепь (SEQ ID NO:168) включает модифицированные рибонуклеотиды с 2′OMe сахаром в положениях (5′>3′) 1, 3, 8, 12, 13 и 15; 2′5′ рибонуклеотид в положении 6; C3Pi-C3OH группу, ковалентно соединенную с 3′-концом.
[0111] Согласно некоторым вариантам реализации двухцепочечная молекула нуклеиновой кислоты включает антисмысловую цепь, представленную на SEQ ID NO:172, и смысловую цепь, представленную на SEQ ID NO:105; обозначенные в настоящем описании как SERPINH1_51a. Согласно некоторым вариантам реализации указанный дуплекс содержит структуру
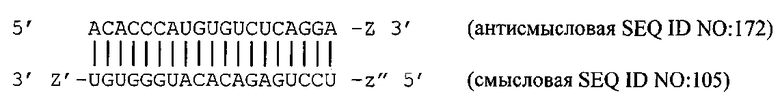
на которой «|» обозначает спаривание оснований между рибонуклеотидами;
на которой А, С, G, U независимо друг от друга представляют собой немодифицированный или модифицированный рибонуклеотид или нетрадиционную группу;
на которой Ζ и Z′ могут независимо друг от друга присутствовать или отсутствовать, но при наличии независимо друг от друга представляют собой 1-5 последовательных нуклеотидов или не являющихся нуклеотидами групп или их комбинацию, ковалентно соединенных с 3′-концом цепи, в которой они находятся; и
на которой z′′ может присутствовать или отсутствовать, но при наличии представляет собой блокирующую группу, ковалентно соединенную с 5′-концом N2-(N′)y.
[0112] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:105) включает модифицированные пиримидины с 2′ОМе сахаром, возможно, 2′-5′ рибонуклеотид в положении 9 или 10; нуклеотид или не являющуюся нуклеотидом группу, ковалентно соединенную с 3′-концом, и, возможно, блокирующую группу, ковалентно соединенную с 5′-концом. Согласно некоторым вариантам реализации антисмысловая цепь (SEQ ID NO:172) включает модифицированный пиримидин и/или пурины с 2′ОМе сахаром, 2′-5′ нуклеотид в положении 5, 6, 7 или 8 (5′>3′); и
нуклеотидную или не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом.
[0113] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:105) включает модифицированные пиримидины с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, возможно, 2'-5' рибонуклеотид в положении 9 или 10, C3Pi или С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловую цепь (SEQ ID NO:172) выбирают из антисмысловой цепи, которая включает
а) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 8 и 15, 2'5' рибонуклеотид в положении 6 или 7; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; или
б) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 8, 13 и 15, 2'5' рибонуклеотид в положении 6 или 7; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; или
в) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 8, 11 и 15, 2'5' рибонуклеотид в положении 6; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; или
г) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 8, 12, 13 и 15; 2'5' рибонуклеотид в положении 6; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0114] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:105) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, возможно, 2'-5' рибонуклеотид в положении 9, С3-ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:172) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 8 и 15, 2'5' рибонуклеотид в положении 6; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0115] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:105) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, возможно, 2'-5'рибонуклеотид в положении 9, С3-ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:172) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 8, 13 и 15, 2'5' рибонуклеотид в положении 6; а С3Р1-С3ОН группу, ковалентно соединенную с 3'-концом.
[0116] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:105) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, 2'-5' рибонуклеотид в положении 9, С3-ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:172) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 8, 11 и 15, 2'5' рибонуклеотид в положении 6; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0117] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:105) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, 2'-5' рибонуклеотид в положении 9, С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:172) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 8, 12, 13 и 15; 2'5' рибонуклеотид положении 6; C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0118] Согласно некоторым вариантам реализации антисмысловую и смысловую цепи выбирают из пар олигонуклеотидов, представленных в Таблице А-19, и обозначенных в настоящем описании как SERPINH1_4 (SEQ ID NOS:195 и 220) и SERPINH1_12 (SEQ ID NOS:196H221).
[0119] Согласно некоторым вариантам реализации двухцепочечная молекула нуклеиновой кислоты включает антисмысловую цепь, представленную на SEQ ID NO:220, и смысловую цепь, представленную на SEQ ID NO:194; обозначенные в настоящем описании как SERPINH1_4. Согласно некоторым вариантам реализации двухцепочечная молекула нуклеиновой кислоты имеет структуру
на которой «|» обозначает спаривание оснований между рибонуклеотидами;
на которой А, С, G, U независимо друг от друга представляют собой немодифицированный или модифицированный рибонуклеотид или нетрадиционную группу;
на которой Z и Z' могут независимо друг от друга присутствовать или отсутствовать, но при наличии представляют собой независимо друг от друга 1-5 последовательных нуклеотидов или не являющихся нуклеотидами групп или их комбинацию, ковалентно соединенных с 3'-концом цепи, в которой они находятся; и
на которой z" может присутствовать или отсутствовать, но при наличии представляет собой блокирующую группу, ковалентно соединенную с 5'-концом N2-(N')y.
[0120] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:220) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 9, 11, 15, 17 и 19, 2'-5' рибонуклеотид в положении 7, и С3Р1-С3ОН группу, ковалентно соединенную с 3'-концом; и смысловую цепь (SEQ ID NO:195) выбирают из смысловой цепи, которая включает:
а) 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19, С3ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; или
б) 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19, 3'-концевой фосфат; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; или
в) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 5, 7, 13 и 16; 2'5' рибонуклеотид в положении 18; С3ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; или
г) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 7, 13, 16 и 18; 2'5' рибонуклеотид в положении 9; С3ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; или
д) 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19: C3Pi группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0121] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:220) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 9, 11, 15, 17, 19, 2'-5' рибонуклеотид в положении 7 и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; и смысловая цепь (SEQ ID NO:195) включает 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19: С3 группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0122] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:220) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 9, 11, 15, 17, 19, 2'-5' рибонуклеотид в положении 7 и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; и смысловая цепь (SEQ ID NO:195) включает 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19: 3'-концевой фосфат; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0123] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:220) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 9, 11, 15, 17, 19, 2'-5' рибонуклеотид в положении 7 и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; и смысловая цепь (SEQ ID NO:195) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 5, 7, 13 и 16; 2'-5' рибонуклеотид в положении 18; С3ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0124] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:220) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 9, 11, 15, 17, 19, 2'-5' рибонуклеотид в положении 7 и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; и смысловая цепь (SEQ ID NO:195) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 7, 13, 16 и 18; 2'-5' рибонуклеотид в положении 9; С3ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0125] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:220) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 9, 11, 15, 17, 19, 2'-5' рибонуклеотид в положении 7 и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; и смысловая цепь (SEQ ID NO:195) включает 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19: C3Pi группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0126] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой антисмысловая цепь (SEQ ID NO:220) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 13, 15, 17, 19 и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; и смысловая цепь (SEQ ID NO:195) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 7, 9, 13, 16 и 18; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом.
[0127] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:195) включает 2'-5' рибонуклеотиды в положениях 15, 16, 17, 18 и 19: 3'-концевой фосфат и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом, и антисмысловая цепь (SEQ ID NO:220) включает антисмысловую цепь, выбранную из одного из следующего:
а) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 7, 9, 11, 13, 15, 17, 19 и C3Pi-C3OH группа, ковалентно соединенная с 3'-концом; или
б) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 6, 8, 10, 12, 14, 17, 18 и C3Pi-C3OH группа, ковалентно соединенная с 3'-концом.
[0128] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, которая включает антисмысловую цепь, представленную на SEQ ID NO:130, и смысловую цепь, представленную на SEQ ID NO:63; обозначенные в настоящем описании как SERPINH1_12. Согласно некоторым вариантам реализации указанная двойная спираль содержит структуру
в которой «|» обозначает спаривание оснований между рибонуклеотидами;
в которой А, С, G, U независимо друг от друга представляют собой немодифицированный или модифицированный рибонуклеотид или нетрадиционную группу;
в которой Z и Z' могут независимо друг от друга присутствовать или отсутствовать, но при наличии независимо друг от друга представляют собой 1-5 последовательных нуклеотидов или не являющихся нуклеотидами групп или их комбинацию, ковалентно соединенных с 3'-концом цепи, в которой они находятся; и
в которой z" может присутствовать или отсутствовать, но при наличии представляет собой блокирующую группу, ковалентно соединенную с 5'-концом N2-(N')y.
[0129] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:196) включает один или несколько модифицированных пиримидинов с 2'ОМе сахаром; нуклеотидный или не состоящий из нуклеотидов выступающий 3'-конец; и блокирующую группу, ковалентно соединенную с 5'-концом. Согласно некоторым вариантам реализации антисмысловая цепь (SEQ ID NO:221) включает один или несколько модифицированных пиримидинов с 2'ОМе сахаром, нуклеотид или не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и блокирующую группу, ковалентно соединенную с 5'-концом.
[0130] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:196) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 14 и 18; С3ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловую цепь (SEQ ID NO:221) выбирают из антисмысловой цепи, которая включает
а) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 9, 11, 13, 15 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом; или
б) модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 7, 9, 12, 13 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0131] Согласно некоторым вариантам реализации обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:196) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 14 и 18; С3-ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:221) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 9, 11, 13, 15 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0132] Согласно некоторым вариантам реализации обеспечивается молекула дуплекса олигонуклеотидов, в которой смысловая цепь (SEQ ID NO:196) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 14 и 18 и, возможно, в положении 2; С3-ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь (SEQ ID NO:221) включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 3, 5, 7, 9, 12, 13 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3'-концом.
[0133] Согласно некоторым вариантам реализации обеспечивается молекула дуплекса олигонуклеотидов, в которой смысловая цепь (SEQ ID NO:196) включает модифицированные рибонуклеотиды с 2′ОМе сахаром в положениях (5′>3′) 14 и 18; С3-ОН группу, ковалентно соединенную с 3′-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5′-концом; и антисмысловую цепь (SEQ ID NO:221) выбирают из антисмысловой цепи, которая включает:
а) модифицированные рибонуклеотиды с 2′ОМе сахаром в положениях (5′>3′) 3, 5, 9, 11, 13, 15 и 17; 2′-5′ рибонуклеотид в положении7; и C3Pi-C3OH группу, ковалентно соединенную с 3′-концом; или
б) модифицированные рибонуклеотиды с 2′ОМе сахаром в положениях (5′>3′) 3, 5, 7, 9, 12, 13 и 17; 2′-5′ рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3′-концом.
[0134] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:196) включает модифицированные рибонуклеотиды с 2′ОМе сахаром в положениях (5′>3′) 14 и 18; С3-ОН группу, ковалентно соединенную с 3′-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5′-концом; и антисмысловая цепь (SEQ ID NO:221) включает модифицированные рибонуклеотиды с 2′ОМе сахаром в положениях (5′>3′) 1, 3, 5, 9, 11, 13, 15 и 17; 2′-5′ рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3′-концом.
[0135] Обеспечивается молекула двухцепочечной нуклеиновой кислоты, в которой смысловая цепь (SEQ ID NO:196) включает модифицированные рибонуклеотиды с 2′ОМе сахаром в положениях (5′>3′) 14 и 18; С3-ОН группу, ковалентно соединенную с 3′-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5′-концом; и антисмысловая цепь (SEQ ID NO:221) включает модифицированные рибонуклеотиды с 2′OMe сахаром в положениях (5′>3′) 1, 3, 5,7, 9, 12, 13 и 17; 2′-5′ рибонуклеотид в положении 7; и C3Pi-C3OH группу, ковалентно соединенную с 3′-концом.
[0136] Согласно другим вариантам реализации Структур А1 и А2 (N′)y включает 1-8 модифицированных нуклеотидов, при этом модифицированный рибонуклеотид представляет собой нуклеотид ДНК. Согласно некоторым вариантам реализации (N')y включает 1, 2, 3, 4, 5, 6, 7 или до 8 групп ДНК.
[0137] Согласно некоторым вариантам реализации присутствует Z или Z' и независимо друг от друга включает две ненуклеотидные группы.
[0138] Согласно дополнительным вариантам реализации присутствуют Z и Z' и каждая из них независимо друг от друга включает две ненуклеотидные группы.
[0139] Согласно некоторым вариантам реализации каждый из Z и Z' включает группу без основания, например, группу дезоксирибозы без основания (обозначаемую как «dAb») или группу рибозы без основания (обозначаемую как «rAb»). Согласно некоторым вариантам реализации каждый из Z и/или Z' включает две ковалентно связанные группы, лишенные основания, и представляет собой, например, dAb-dAb или rAb-rAb или dAb-rAb или rAb-dAb, где каждая группа коваленто соединена с прилегающей группой, предпочтительно, с помощью фосфорной связи. Согласно некоторым вариантам реализации фосфорная связь включает фосфотиоатную, фосфоноацетатную или фосфодиэфирную связь. Согласно предпочтительным вариантам реализации фосфорная связь включает фосфодиэфирную связь.
[0140] Согласно некоторым вариантам реализации каждый из Z и/или Z' независимо друг от друга включает алкильную группу, возможно, пропановую группу [(СН2)3] (С3) или их производное, включая пропанол (С3-ОН) и фосфорные производные пропандиола ("С3-3'Pi"). Согласно некоторым вариантам реализации каждый из Z и/или Z' включает две алкильные группы, ковалентно соединенные с 3'-концом антисмысловой цепи или смысловой цепи посредством фосфодиэфирной или фосфотиоатной связи, и ковалентно соединенные друг с другом посредством фосфодиэфирной или фосфотиоатной связи, и в некоторых примерах представляет собой C3Pi-C3Pi или C3Pi-C3OH. 3'-конец антисмысловой цепи и/или 3'-конец смысловой цепи коваленто соединен с группой С3 посредством фосфорной связи, и группа С3 ковалентно конъюгирована с С3-ОН группой посредством фосфорной связи. Согласно некоторым вариантам реализации фосфорные связи включают фосфодиэфирную, фосфоноацетатную или фосфодиэфирную связь. Согласно предпочтительным вариантам реализации фосфорная связь включает фосфодиэфирную связь.
[0141] Согласно различным вариантам реализации Структуры А1 или Структуры А2, Z и Z' отсутствуют. Согласно другим вариантам реализации присутствует каждый из Z и/или Z'. Согласно некоторым вариантам реализации каждый из Z и/или Z' независимо друг от друга включает С2, С3, С4, С5 или С6 алкильную группу, возможно, С3 [пропан, -(СН2)3-] группу или ее производное, включая пропанол (С3-ОН/С3ОН), пропандиол, и фосфодиэфирное производное пропандиола («C3Pi»). Согласно предпочтительным вариантам реализации каждый из Z и/или Z' включает две углеводородные группы и в некоторых примерах представляет собой С3Р1-С3ОН или C3Pi-C3Pi. Каждый С3 ковалентно соединен с прилегающим С3 с помощью ковалентной связи, предпочтительно, фосфорной связи. Согласно некоторым вариантам реализации фосфорные связи включают фосфодиэфирную, фосфоноацетатную или фосфодиэфирную связь.
[0142] Согласно определенным вариантам реализации x=y=19, и Z содержит выступающий конец по меньшей мере с одним С3 алкилом. Согласно некоторым вариантам реализации выступающий конец С3-С3 ковалентно соединен с 3'-концом (N)x или (N')y посредством ковалентной связи, предпочтительно фосфодиэфирной связи. Согласно некоторым вариантам реализации связь между первым С3 и вторым С3 представляет собой фосфодиэфирную связь. Согласно некоторым вариантам реализации не состоящий из нуклеотидов выступающий 3'- конец представляет собой C3Pi-C3Pi. Согласно некоторым вариантам реализации не состоящий из нуклеотидов выступающий 3'-конец представляет собой C3Pi-C3Ps. Согласно некоторым вариантам реализации не состоящий из нуклеотидов выступающий 3'-конец представляет собой C3Pi-C3OH (ОН обозначает гидроксил). Согласно некоторым вариантам реализации не состоящий из нуклеотидов выступающий 3'-конец представляет собой C3Pi-C3OH.
[0143] Согласно различным вариантам реализации алкильная группа содержит производное алкила, включая С3 алкил, С4 алкил, С5 алкил или С6 алкильную группу, содержающую концевую гидрокси, амино или фосфатную группу. Согласно некоторым вариантам реализации алкильная группа представляет собой производную группу С3 алкила или С3 алкила. Согласно некоторым вариантам реализации С3 алкильная группа содержит пропанол, пропилфосфат, пропилфосфотиоат или их комбинацию. С3 алкильная группа соединена с 3'-концом (N')y и/или 3'-концом (N)x с помощью фосфодиэфирной связи. Согласно некоторым вариантам реализации алкильная группа содержит пропанол, пропилфосфат или пропил фосфотиоат. Согласно некоторым вариантам реализации каждый из Z и Z' независимо выбирают из пропанола, пропил фосфата, пропил фосфотиоата их комбинаций или их кратных количеств, в частности 2 или 3 ковалентно соединенных пропанола, пропилфосфата, пропил фосфотиоата или их комбинаций. Согласно некоторым вариантам реализации каждый из Z и Z' независимо выбирают из пропилфосфата, пропил фосфотиоата, пропил фосфопропанола; пропил фосфо-пропил фосфотиоата; пропилфосфо-пропил фосфата; (пропил фофата)3, (пропил фосфата)2-пропанола, (пропил фосфата)2-пропил фосфотиоата. Любую группу, конъюгированную с пропаном или пропанолом, можно включить в Z или Z'.
[0144] Примеры структур примеров 3'-концевых не являющихся нуклеотидами групп приведены ниже:
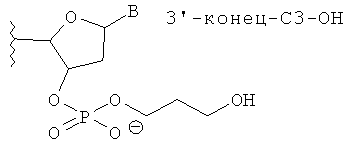
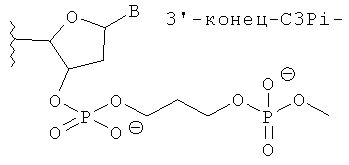
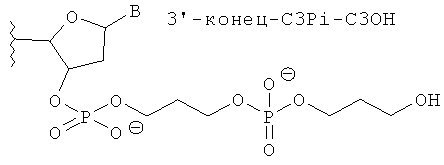
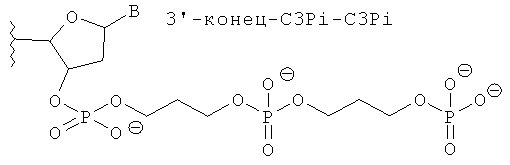

[0145] Согласно некоторым вариантам реализации каждого из Z и Z' независимо выбирают из пропанола, пропил фосфата, пропил фосфотиоата их комбинаций или их кратных количеств.
[0146] Согласно некоторым вариантам реализации каждого из Z и Z' независимо выбирают из пропил фосфата, пропил фосфотиоата, пропил фосфопропанола; пропил фосфо-пропил фосфотиоата; пропилфосфо-пропил фосфата; (пропил фосфата)3, (пропил фосфата)2-пропанола, (пропил фосфата)2-пропил фосфотиоата. Любую группу, конъюгированную с пропаном или пропанолом, можно включить в Z или Z'.
[0147] Согласно дополнительным вариантам реализации каждый из Z и/или Z' включает комбинацию группы без основания и немодифицированного дезоксирибонуклеотида или рибонуклеотида или комбинацию группы углевода и немодифицированного дезоксирибонуклеотида или рибонуклеотида или комбинацию группы без основания (дезоксирибо или рибо) и группы углевода. Согласно таким вариантам реализации каждый из Z и/или Z' включает С3-rAb или С3-dAb, при этом каждая группа ковалентно связана с прилегающей группой посредством фосфорной связи, предпочтительно, фосфодэифирной, фосфотиоатной или фосфотиоацетатной связи.
[0148] Согласно некоторым вариантам реализации молекулы нуклеиновой кислоты согласно настоящему описанию включают смысловую последовательность олигонуклеотида, выбранную из любого из олигонуклеотидов под номерами 2-67 или 68-92, представленными ниже в Таблицах А-18 и А-19, соответственно.
[0149] Согласно некоторым предпочтительным вариантам реализации соединения, обеспечиваемые в настоящем описании, включают Соединение_1, Соединение_2, Соединение_3, Соединение_4, Соединение_5, Соединение_6, Соединение_7, Соединение_8 и Соединение_9 согласно настоящему описанию.
[0150] Согласно некоторым вариантам реализации (таким как, например, Соединение_1, Соединение_5 и Соединение_6 согласно настоящему описанию) обеспечиваются 19 мерные двухцепочечные молекулы нуклеиновой кислоты, в которых антисмысловая цепь представляет собой SEQ ID NO:127, и смысловая цепь представляет собой SEQ ID NO:60. Согласно некоторым вариантам реализации, обеспечиваются 19 мерные двухцепочечные молекулы нуклеиновой кислоты, в которых антисмысловая цепь представляет собой SEQ ID NO:127 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром, 2'-5' рибонуклеотид в по меньшей мере одном из положений 1, 5, 6 или 7, и 3'-концевую не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом; и смысловая цепь представляет собой SEQ ID NO:60 и включает по меньшей мере один 2'5' рибонуклеотид или модифицированный рибонуклеотид с 2'ОМе сахаром, не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и блокирующую группу, ковалентно соединенную с 5'-концом. Согласно некоторым вариантам реализации, обеспечивается 19 мерная двухцепочечная молекула нуклеиновой кислоты, в которой антисмысловая цепь представляет собой SEQ ID NO:127 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях 3, 5, 9, 11, 13, 15, 17 и 19 (5'>3'), 2'-5' рибонуклеотид в положении 7, и 3'-концевую С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом; и смысловая цепь представляет собой SEQ ID NO:60 и включает 5 последовательных 2'5' рибонуклеотидов в 3'-концевых положениях 15, 16, 17, 18 и 19 (5'>3'), C3Pi не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу без основания, ковалентно соединенную с 5'-концом.
[0151] Согласно одному варианту реализации обеспечивается Соединение_1, которое представляет собой 19 мерную двухцепочечную молекулу нуклеиновой кислоты, в которой антисмысловая цепь представляет собой SEQ ID NO:127 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях 3, 5, 9, 11, 13, 15, 17 и 19 (5'>3'), 2'-5' рибонуклеотид в положении 7, и C3Pi-C3OH не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом; и смысловая цепь представляет собой SEQ ID NO:60 и включает 5 последовательных 2'5' рибонуклеотидов в 3'-концевых положениях 15, 16, 17, 18 и 19 (5'>3'), C3Pi не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инветированную группу без основания, ковалентно соединенную с 5'-концом; и которая также включает модифицированный рибонуклеотид с 2'ОМе сахаром в положении 1 антисмысловой цепи.
[0152] Согласно одному варианту реализации, обеспечивается Соединение_6, которое представляет собой 19 мерную двухцепочечную молекулу нуклеиновой кислоты, в которой антисмысловая цепь представляет собой SEQ ID NO:127 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях 3, 5, 9, 11, 13, 15, 17 и 19 (5'>3'), 2'-5' рибонуклеотид в положении 7, и C3Pi-C3OH не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом; и смысловая цепь представляет собой SEQ ID NO:60 и включает 5 последовательных 2'5' рибонуклеотидов в 3'-концевых положениях 15, 16, 17, 18 и 19 (5'>3'), C3Pi не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу без основания, ковалентно соединенную с 5'-концом; и которая также включает 2'5' рибонуклеотид в положении 1 антисмысловой цепи.
[0153] Согласно одному варианту реализации обеспечивается Соединение_5, которое представляет собой 19 мерную двухцепочечную молекулу нуклеиновой кислоты, в которой антисмысловая цепь представляет собой SEQ ID NO:127 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях 1, 3, 5, 9, 11, 13, 15, 17 и 19 (5'>3'), 2'-5' рибонуклеотид в положении 7, и С3Р1-С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом; и смысловая цепь представляет собой SEQ ID NO:60 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 7, 13, 16 и 18, 2'5' рибонуклеотид в положении 9, С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу без основания, ковалентно соединенную с 5'-концом.
[0154] Согласно некоторым вариантам реализации (таким как, например, Соединение_2 и Соединение_7 согласно настоящему описанию) обеспечиваются 19 мерные двухцепочечные молекулы нуклеиновой кислоты, в которых смысловая цепь представляет собой SEQ ID NO:63, и антисмысловая цепь представляет собой SEQ ID NO:130. Согласно некоторым вариантам реализации обеспечиваются 19 мерные двухцепочечные молекулы нуклеиновой кислоты, в которых смысловая цепь представляет собой SEQ ID NO:63 и включает модифицированные пиримидиновые рибонуклеотиды с 2'ОМе сахаром; не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом; и блокирующую группу, ковалентно соединенную с 5'-концом; и антисмысловая цепь представляет собой SEQ ID NO:130 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром; 2'-5' рибонуклеотид в положении 7; и не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом. Согласно некоторым вариантам реализации обеспечиваются 19 мерные двухцепочечные молекулы нуклеиновой кислоты, в которых смысловая цепь представляет собой SEQ ID NO:63 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром, не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и блокирующую группу, ковалентно соединенную с 5'-концом; и антисмысловая цепь представляет собой SEQ ID NO:130 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром; 2'-5' рибонуклеотид в по меньшей мере одном из положений 5, 6 или 7; и не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом.
[0155] Согласно одному варианту реализации, обеспечивается Соединение_2, которое представляет собой 19 мерную двухцепочечную молекулу нуклеиновой кислоты, в которой смысловая цепь представляет собой SEQ ID NO:63 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 14 и 18; С3ОН группу, ковалентно соединенную с 3'-концом; и инвертированную без основания группу дезоксирибонуклеотида, ковалентно соединенную с 5'-концом; и антисмысловая цепь представляет собой SEQ ID NO:130 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 12, 13 и 17; 2'-5' рибонуклеотид в по меньшей мере одном из положений 5, 6 или 7; и C3Pi-С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом.
[0156] Согласно одному варианту реализации, обеспечивается Соединение_7, которое представляет собой 19 мерную двухцепочечную молекулу нуклеиновой кислоты, в которой смысловая цепь представляет собой SEQ ID NO:63 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 14 и 18; С3ОН группу, ковалентно соединенную с 3'-концом; и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь представляет собой SEQ ID NO:130 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 3, 5, 9, 11, 13 и 17; 2'-5' рибонуклеотид в положении 7; и C3Pi-C3OH не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом.
[0157] Согласно некоторым вариантам реализации (таким как, например, Соединение_3 согласно настоящему описанию) обеспечиваются 19 мерные двухцепочечные молекулы нуклеиновой кислоты, в которых смысловая цепь представляет собой SEQ ID NO:98 и антисмысловая цепь представляет собой SEQ ID NO:165. Согласно некоторым вариантам реализации обеспечиваются 19 мерные двухцепочечные молекулы нуклеиновой кислоты, в которых смысловая цепь представляет собой SEQ ID NO:98 и включает 2'-5' рибонуклеотиды в положениях на 3'-конце: не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и блокирующую группу, ковалентно соединенную с 5'-концом; и антисмысловая цепь представляет собой SEQ ID NO:165 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром; 2'-5' рибонуклеотид в по меньшей мере одном из положений 5, 6 или 7 и не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом. Согласно одному варианту реализации, обеспечивается Соединение_3, которое представляет собой 19 мерную двухцепочечную молекулу нуклеиновой кислоты, в которой смысловая цепь представляет собой SEQ ID NO:98 и включает 2'-5' рибонуклеотиды в положениях (5'>3') 15, 16, 17, 18 и 19: С3-ОН 3' группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь представляет собой SEQ ID NO:165 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 4, 6, 8, 11, 13, 15, 17 и 19; 2'-51 рибонуклеотид в положении 7 и C3Pi-C3OH, ковалентно соединенную с 3'-концом.
[0158] Согласно некоторым вариантам реализации (таким как, например, Соединение_4, Соединение_8 и Соединение_9 согласно настоящему описанию) обеспечиваются 19 мерные двухцепочечные молекулы нуклеиновой кислоты, в которых смысловая цепь представляет собой SEQ ID NO:101, и антисмысловая цепь представляет собой SEQ ID NO:168. Согласно некоторым вариантам реализации обеспечиваются 19 мерные двухцепочечные молекулы нуклеиновой кислоты, в которых смысловая цепь представляет собой SEQ ID NO:101 и включает модифицированные пиримидиновые рибонуклеотиды с 2'ОМе сахаром и возможно, 2'-5' рибонуклеотид в одном из положений 9 или 10, не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и блокирующую группу, ковалентно соединенную с 5'-концом; и антисмысловая цепь представляет собой SEQ ID NO:168 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром, 2'5' рибонуклеотид в по меньшей мере одном из положений 5, 6 или 7; и не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом.
[0159] Согласно одному варианту реализации, обеспечивается Соединение_4, которое представляет собой 19 мерную двухцепочечную молекулу нуклеиновой кислоты в которой смысловая цепь представляет собой SEQ ID NO:101 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13, и 17, 2'-5' рибонуклеотид в положении 9, С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь представляет собой SEQ ID NO:168 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 4, 8, 11 и 15, 2'5' рибонуклеотид в положении 6; группу C3Pi-C3OH, образующую выступающий конец, ковалентно соединенную с 3'-концом.
[0160] Согласно одному варианту реализации, обеспечивается Соединение_8, которое представляет собой 19 мерную двухцепочечную молекулу нуклеиновой кислоты, в которой смысловая цепь представляет собой SEQ ID NO:101 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 4, 11, 13 и 17, С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь представляет собой SEQ ID NO:168 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 4, 8, 13 и 15, 2'5' рибонуклеотид в положении 6; и группу C3Pi-C3OH, образующую выступающий конец, ковалентно соединенный с 3'-концом.
[0161] Согласно одному варианту реализации, обеспечивается Соединение_9, которое представляет собой 19 мерную двухцепочечную молекулу нуклеиновой кислоты, в которой смысловая цепь представляет собой SEQ ID NO:101 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 2, 4, 11, 13 и 17, С3ОН не являющуюся нуклеотидом группу, ковалентно соединенную с 3'-концом, и инвертированную группу дезоксирибонуклеотида без основания, ковалентно соединенную с 5'-концом; и антисмысловая цепь представляет собой SEQ ID NO:168 и включает модифицированные рибонуклеотиды с 2'ОМе сахаром в положениях (5'>3') 1, 4, 8, 11 и 15, 2'5' рибонуклеотид в положении 6; не являющуюся нуклеотидом группу C3Pi-C3OH, ковалентно соединенную с 3'-концом.
[0162] Согласно другому аспекту обеспечиваются способы снижения экспресссии hsp47 в клетке посредством введения в клетку молекулы нуклеиновой кислоты согласно настоящему описанию в количестве, достаточном для снижения экспрессии hsp47. Согласно одному варианту реализации клетка представляет собой звездчатую клетку печени. Согласно другому варианту реализации клетка представляет собой звездчатую клетку ткани почек или легких. Согласно некоторым вариантам реализации способы осуществляют in vitro, согласно другим вариантам реализации способ осуществляют in vivo.
[0163] Согласно другому аспекту обеспечиваются способы лечения пациента, страдающего от заболевания, вызванного hsp47. Способы включают введение пациенту молекулы нуклеиновой кислоты, например, согласно настоящему описанию, в количестве, достаточном для снижения экспрессии hsp47. Согласно некоторым вариантам реализации, заболевание, вызванное hsp47, представляет собой заболевание, выбранное из группы, состоящей из фиброза печени, цирроза, фиброза легких, включая фиброз легких (включая ИЛФ), любое состояние, вызывающее фиброз почек (например, ХБП, включая ТХПН), перитонеального фиброза, хронического поражения печени, фибриллогенеза, фибротических заболеваний в других органах, ненормального рубцевания (келоидные рубцы), ассоциированные со всеми возможными типами травмы кожи: в результате несчастного случая и ятрогенными (операции); склеродермии, кардиофироза, неудачного исхода фильтрующей операции при глаукоме и кишечных спаек. Согласно некоторым вариантам реализации соединение можно использовать для лечения заболеваний органов, например, показаний, включая представленные в Таблице 2 ниже:
[0164] Согласно некоторым вариантам реализации предпочтительные показания включают цирроз печени, вызванный гепатитом С, после трансплантации печени; цирроз печени по причине неалкогольного стеатогепатита (НАСГ); идиопатический фиброз легких; лучевой пневмонит, вызывающий фиброз легких; диабетическую нефропатию; перитонеальный склероз, вызванный хроническим перитонеальным диализом в амбулаторных условиях (CAPD), и пузырчатку глаз.
[0165] Фибротические заболевания печени включают алкогольный цирроз, цирроз, вызванный гепатитом В, цирроз, вызванный гепатитом С, цирроз, вызванный гепатитом С (Геп С) после ортопической трансплантации печени (OLTX), НАСГ/НЖБП, первичный билиарный цирроз печени (ПБЦП), первичный склерозирующий холангит (PSC), атрезию желчных протоков, дефицит альфа 1-антитрипсина (A1AD), болезни накопления меди (болезнь Вильсона), фруктоземию, галактоземию, болезни накопления гликогена (в частности, типов III, IV, VI, IX и X), синдромы перегрузки железом (гемохроматозы), нарушения липидного обмена (например, болезнь Гоше), пероксисомальные болезни (например, синдром Целлвегера), тирозинемию, врожденный фиброз печени, бактериальные инфекции (например, бруцеллез), паразитов (эхинококкоз), болезнь Бадда-Киари (облитерирующий эндофлебит печеночных вен).
[0166] Заболевания легких включают идиопатический фиброз легких, силикоз, пневмокониоз, бронхолегочную лисплазию новорожденных после синдрома острой дыхательной недостаточности, повреждение легких в результате применение блеомицина/химических агентов, облитерирующй бронхиолит (ОБ) после трансплантации легких, хроническую обструктивную болезнь легких (ХОБЛ), муковисцидоз, астму.
[0167] Заболевания сердца включают кардиомиопатию, атеросклероз (заболевание Бергенра и т.д.), фиброз эндомиокарда, фибрилляцию предсердий, рубцевание после инфаркта миокарда (ИМ)
[0168] Другие заболевания торакальной области включают индуцированные облучением реакции ткани капсулы вокруг структурированных грудных имплантатов и субмукозальный фиброз полости рта.
[0169] Заболевания почек включают аутосомно-доминантный поликистоз почек (АДПП), диабетическую нефропатию (диабетический гломерулосклероз), ФСГС (коллапсныйпротив других гистологических вариантов), IgA нефропатию (болезнь Бергера), волчаночный нефрит, остеохондрит Вегнера, склеродермию, синдром Гудпасчера, тубулоинтерстициальный фиброз: вызванную лекарствами (защитными) пенициллинами, сергалоспоринами, анальгетиками нефропатию, мембранозно-пролиферативный гломерулонефрит (МПГН), болезнь Шенлейн-Геноха, врожденные нефропатии: ювенильный нефронофтиз, наследственный онихоартроз и синдром Альпорта.
[0170] Болезни костного мозга включают лимфангиолиомиоматоз (ЛАМ), хроническую реакцию трансплантат-против-хозяина, полицитемия, идиопатическую тромбоцитемию, миелофиброз.
[0171] Болезни глаз включают ретинопатию ретролентальную фиброплазию (РЛФ), пузырчатку глаз, фиброз слезных желез, операции по прикреплению сетчатки, помутнение роговицы, герпетический кератит, птеригиум, глаукому, возрастную макулярную дистрофию (ВДМ), ретиноплатию, вызванную фиброзом сетчатки при сахарном диабете (СД).
[0172] Гинекологические заболевания включают эндометриоз при гормональной терапии для предотвращения рубцевания, фиброз/сальпингит после ИПП.
[0173] Системные заболевания включают синдром Дюпюитрена, контрактуру ладонного апоневроза, болезнь Пейрони, Болезнь Леддерхозе, келоидные рубцы, множественный фибросклероз, нефрогенный системный фиброз, нефрогенный миелофиброз (анемия).
[0174] Фибротические заболевания, вызванные травмами, включают образование рубцов и контрактур на коже и мягких тканях в результате ожогов (включая химические), вызванное облучением образование шрамов в коже и органах после применения облучения при терапии рака, келоидные рубцы (кожа).
[0175] Показания после операций включают перитонеальный фиброз из-за катетера перитонеального диализа брюшины, имплантата роговицы, кохлеарного имплантата, других имплантов, силиконовых имплантов молочных желез, хронический синусит; спайки, псевдоинтимальная гиперплазия трансплантатов диализа.
[0176] Другие состояния включают хронический панкреатит.
[0177] Согласно некоторым вариантам реализации обеспечивается способ лечения пациента, страдающего от фиброза печени, включающий введение пациенту эффективного количества молекулы нуклеиновой кислоты согласно настоящему описанию, благодаря чему лечится фиброз печени. Согласно некоторым вариантам реализации пациент страдает от цирроза печени из-за гепатита. Согласно некоторым вариантам реализации пациент страдает от цирроза печени из-за НАСГ.
[0178] Согласно некоторым вариантам реализации обеспечивается применение молекулы нуклеиновой кислоты согласно настоящему описанию для изготовления лекарственного средства для лечения фиброза печени. Согласно некоторым вариантам реализации фиброз печени вызван гепатитом. Согласно некоторым вариантам реализации фиброз печени вызван НАСГ.
[0179] Согласно некоторым вариантам реализации обеспечивается способ ремоделирования рубцовой ткани, включающий введение пациенту, который в этом нуждается, эффективного количества молекулы нуклеиновой кислоты согласно настоящему описанию, влияя, таким образом, на ремоделирование рубцовой ткани. Согласно некоторым вариантам реализации рубцовая ткань находится в печени. Согласно некоторым вариантам реализации пациент страдает от цирроза печени из-за гепатита. Согласно некоторым вариантам реализации пациент страдает от цирроза печени из-за НАСГ.
[0180] Согласно некоторым вариантам реализации обеспечивается способ влияния на регрессию фиброза, включающий введение пациенту, который в этом нуждается, эффективного количества молекулы нуклеиновой кислоты согласно настоящему описанию, влияя, таким образом, на регрессию фиброза.
[0181] Согласно некоторым вариантам реализации обеспечивается способ уменьшения рубцовой ткани у пациента, включающий этап введения пациенту эффективного количества молекулы нуклеиновой кислоты согласно настоящему описанию для уменьшения рубцовой ткани. Согласно некоторым вариантам реализации обеспечивается способ уменьшения рубцовой ткани у пациента, включающий этап топического нанесения на рубцовую ткань эффективного количества молекулы нуклеиновой кислоты согласно настоящему описанию для уменьшения рубцовой ткани.
[0182] Согласно некоторым вариантам реализации обеспечивается способ улучшения внешнего вида рубцовой ткани, включающий этап топического нанесения на рубцовую ткань эффективного количества молекулы нуклеиновой кислоты согласно настоящему описанию для улучшения внешнего вида рубцовой ткани.
[0183] Согласно некоторым вариантам реализации обеспечивается способ лечения пациента, страдающего от фиброза легких, включающий введение пациенту эффективного количества молекулы нуклеиновой кислоты согласно настоящему описанию, благодаря чему лечится фиброз легких. Согласно некоторым вариантам реализации пациент страдает от интерстициального фиброза легких (ИЛФ). Согласно некоторым вариантам реализации пациент страдает от лучевого пневмонита, вызывающего фиброз легких. Согласно некоторым вариантам реализации пациент страдает от фиброза легких, вызванного лекарственными средствами.
[0184] Согласно некоторым вариантам реализации обеспечивается применение молекулы нуклеиновой кислоты согласно настоящему описанию для изготовления лекарственного средства для лечения фиброза легких. Согласно некоторым вариантам реализации фиброз легких представляет собой ИЛФ. Согласно некоторым вариантам реализации фиброз легких представляет собой фиброз легких, вызванный лекарственными средствами или облучением.
[0185] Согласно одному аспекту обеспечиваются фармацевтические композиции, которые включают молекулу нуклеиновых кислот (например, молекулу siНК) согласно настоящему описанию в фармацевтически приемлемом носителе. Согласно некоторым вариантам реализации лекарственная форма включает или содержит систему доставки, подходящую для доставки молекул нуклеиновых кислот (например, молекулы siНК) пациенту; примеры систем доставки описаны более подробно ниже.
[0186] Согласно родственному аспекту обеспечиваются композиции или наборы, которые включают молекулу нуклеиновой кислоты (например, молекулу siНК), упакованную для использования пациентом. Упаковка может иметь этикетку или включать бирку места или вкладыш, на котором указано содержимое упаковки и дана конкретная информация относительно того, как пациент должен или может использовать молекулу нуклеиновой кислоты (например, молекулу siНК), например, этикетка может содержать информацию о дозе и/или показания к применению. Согласно некоторым вариантам реализации содержание этикетки включает сообщение в форме, предписанной государственным учреждением, например. Управлением по контролю качества пищевых продуктов и лекарственных препаратов. Согласно некоторым вариантам реализации на этикетке может быть указано, какая молекула нуклеиновой кислоты (например, молекула siНК) подходит для применения для лечения пациента, страдающего от заболевания, ассоциированного с hsp47; например, на этикетке может быть указано, какая молекула нуклеиновой кислоты (например, молекула siНК) подходит для применения для лечения фиброза; или, например, на этикетке может быть указано, какая молекула нуклеиновой кислоты (например, молекула siНК) подходит для применения для лечения заболевания, выбранного из группы, состоящей из фиброза, фиброза печени, цирроза, фиброза легких, фиброза почек, перитонеального фиброза, хронического поражения печени и фибриллогенеза.
[0187] В настоящем описании термины «белок теплового шока 47» или «hsp47» или «HSP47» используются взаимозаменяемо и обозначают любой белок теплового шока 47, пептид или полипептид, обладающий любым действием белка hsp47. Белок теплового шока 47 представляет собой ингибитор сериновой протеазы (серпин), также обозначаемый термином, например, ингибитор пептидазы серпина, клада Н, 1 член (SERPINH1), SERPINH2, белок, связывающий коллаген 1 (СВР1), СВР2, gp46; мышьяк-трансактивированный белок 3 (AsTP3); HSP47; ген, вызывающий пролиферацию 14 (PIG14); PPROM; антиген ревматоидного артрита А-47 (RA-A47); коллигин-1 и коллигин-2. Согласно некоторым предпочтительным вариантам реализации «hsp47» обозначает hsp47 человека. Белок теплового шока 47 (или более точно hsp47 человека) может иметь аминокислотную последовательность, которая совпадает или существенно совпадает с SEQ ID NO. 2 (Фигура 7).
[0188] В настоящем описании термин «нуклеотидная последовательность, кодирующая hsp47» обозначает последовательность нуклеотидов, которая кодирует белок hsp47 человека или его часть. Термин «нуклеотидная последовательность, кодирующая hsp47» также включает кодирующие последовательности hsp47, такие как изоформы hsp47, мутантные гены hsp47, варианты, полученные при сплайсинге генов hsp47, и полиморфизмы гена hsp47. Последовательность нуклеиновой кислоты, кодирующая hsp47, включает последовательности мРНК, кодирующие hsp47, которые также обозначаются термином «мРНК hsp47». Примером последовательностей мРНК hsp47 человека является SEQ ID. NO. 1.
[0189] В настоящем описании термины «молекула нуклеиновой кислоты» или «нуклеиновая кислота» используются взаимозаменяемо и обозначают олигонуклеотид, нуклеотид или полинуклеотид. Варианты «молекулы нуклеиновой кислоты» более подробно описаны в настоящем описании. Молекула нуклеиновой кислоты включает как модифицированные молекулы нуклеиновой кислоты, так и немодифицированные молекулы нуклеиновой кислоты согласно настоящему описанию. Молекула нуклеиновой кислоты может включать дезоксирибонуклеотиды, рибонуклеотиды, модифицированные нуклеотиды или аналоги нуклеотидов в любой комбинации.
[0190] В настоящем описании термин «нуклеотид» обозначает химическую группу, содержащую сахар (или его аналог или модифицированный сахар), нуклеиновое основание (или его аналог или модифицированное основание), фосфатную группу (или ее аналог или модифицированную фосфатную группу). Нуклеотид включает как модифицированные нуклеотиды, так и немодифицированные нуклеотиды согласно настоящему описанию. В настоящем описании нуклеотиды могут включать дезоксирибонуклеотиды (например, немодифицированные дезоксирибонуклеотиды), рибонуклеотиды (например, немодифицированные рибонуклеоитды) и аналоги модифицированных нуклеотидов, включая, между прочим, замкнутую нуклеиновую кислоту, пептидо-нуклеиновые кислоты, L-нуклеотиды (также обозначаемые термином зеркальные нуклеотиды), нуклеиновые кислоты с этиленовым мостиком (ENA), арабинозиды, РАСЕ, нуклеотиды с 6-углеродным сахаром, а также аналоги нуклеотидвов (включая нуклеотиды, лишенные основания), которые зачастую считают ненуклеотидами. Согласно некоторым вариантам реализации нуклеотиды могут быть модифицированы по остатку сахара, нуклеинового основания и/или фосфатной группе с помощью любых модификаций, известных в данной области техники, например, модификаций согласно настоящему описанию. Термины «полинуклеотид» или «олигонуклеотид» в настоящем описании обозначают цепь связанных нуклеотидов; полинуклеотиды и олигонуклеотиды также могут иметь модификации в сахаре нуклеотида, нуклеиновых основаниях и фосфатных остовах, примеры которых хорошо известны в данной области техники и/или указаны в настоящем описании.
[0191] В настоящем описании термины «малая интерферирующая нуклеиновая кислота» или «siНК» или «молекула малой интерферирующей нуклеиновой кислоты» обозначают любую молекулу нуклеиновой кислоты, способную изменять экспрессию гена или репликацию вируса. Предпочтительно siНК подавляет или снижает экспрессию гена или репликацию вируса. siНК включает, но не ограничивается, молекулы нуклеиновой кислоты, которые способны опосредовать РНКи, специфичную в отношении последовательностей, например, малые интерферирующие РНК (siРНК), двухцепочечные РНК (dsPHK), микро-РНК (miPHK), малые шпилечные РНК (shPHK), малый интерферирующий олигонуклеотид, малую интерферирующую нуклеиновую кислоту, малый интерферирующий модифицированный олигонуклеотид, химически модифицированную siРНК, РНК постранскрипционного сайленсинга генов (ptgsPHK) и другие. В настоящем описании термин «малая интерферирующая нуклеиновая кислота», «siНК» или «молекула малой интерферирующей нуклеиновой кислоты» имеют значение, которое более подробно описано в настоящем документе.
[0192] В настоящем описании термин «комплементарность» означает, что нуклеиновая кислота может образовывать водородную связь (связи) с другой последовательностью нуклеиновой кислоты посредством традиционных связей по Уотсону-Крику или других нетрадиционных связей. Для молекул нуклеиновых кислот согласно настоящему описанию свободная энергия связывания молекулы нуклеиновой кислоты с комплементарной ей последовательностью является достаточной для того, чтобы соответствующая функция нуклеиновой кислоты превратилась, например, в активность РНКи. Определение свободных энергий связывания для молекул нуклеиновых кислот хорошо известно в данной области техники (смотрите, например, публикацию Turner et al., 1987, CSH Symp. Quant. Biol. LII pp.123-133; Frier et al., 1986, Proc. Nat. Acad. Sci. USA 83:9373-9377; Turner et al., 1987, J. Am. Chem. Soc. 109:3783-3785). Процент комплементарности указывает долю смежных остатков в молекуле нуклеиновой кислоты, которые могут образовывать водородные связи (например, спаривание оснований по Уотсону-Крику) с последовательностью второй нуклеиновой кислоты (например, если 5, 6, 7, 8, 9 или 10 нуклеотидов из 10 нуклеотидов первого олигонуклеотида спариваются с основаниями второй последовательности нуклеиновой кислоты, имеющей 10 нуклеотидов, это означает 50%, 60%, 70%, 80%, 90% и 100% комплементарности, соответственно). «Полная комплементарность» означает, что все смежные остатки последовательности нуклеиновой кислоты образуют водородные связи с тем же количеством смежных остатков в последовательности второй нуклеиновой кислоты. Согласно одному варианту реализации молекула нуклеиновой кислоты согласно настоящему описанию включает приблизительно от 15 до приблизительно 35 или больше (например, приблизительно 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34 или 35 или больше) нуклеотидов, которые комплементарны одной или нескольким молекулам нуклеиновых кислот-мишеней или их частям.
[0193] В настоящем описании термин «смысловая область» обозначает последовательность нуклеотидов молекулы siНК, комплементарную (частично или полностью) антисмысловой области молекулы siНК. Смысловая цепь молекулы siНК может включать последовательность нуклеиновой кислоты, обладающую гомологией с последовательностью нуклеиновой кислоты-мишени. В настоящем описании «смысловая цепь» обозначает молекулу нуклеиновой кислоты, которая включает смысловую область и также может включать дополнительные нуклеотиды.
[0194] В настоящем описании термин «антисмысловая область» обозначает последовательность нуклеотидов молекулы siНК, комплементарную (частично или полностью) последовательности нуклеиновой кислоты-мишени. Антисмысловая цепь молекулы siНК может в некоторых случаях включать последовательность нуклеиновой кислоты, комплементарную смысловой области молекулы siНК. В настоящем описании термин «антисмысловая цепь» обозначает молекулу нуклеиновой кислоты, которая включает антисмысловую область и также может включать дополнительные нуклеотиды.
[0195] В настоящем описании термин «РНК» обозначает молекулу, которая включает по меньшей мере один остаток рибонуклеотида.
[0196] В настоящем описании термин «дуплексная область» обозначает участок в двух комплементарных или существенно комплементарных олигонуклеотидах, которые образуют пары оснований друг с другом как с помощью спаривания оснований по Уотсону-Крику, так и другим образом, при котором образуется дуплекс между цепями олигонуклеотидов, которые являются комплементарными или существенно комплементарными. Например, олигонуклеотид, имеющий 21 нуклеотид, может образовывать пары оснований с другим олигонуклеотидов из 21 нуклеотидов, при этом только 19 оснований каждой цепи являются комплементарными или существенно комплементарными, таким образом, «дуплексная область» состоит из 19 пар оснований. Остальные пары оснований могут, например, существовать в виде 5' и 3'-выступающих концов. Также в пределах дуплексной области не требуется 100% комплементарность; допускается существенная комплементарность в пределах дуплексной области. Существенная комплементарность обозначет такую комплементарность между цепями, при которой они могут гибридизоваться при биологических условиях. Методики эмперического определения способности к гибридизации двух цепей при биологических условиях хорошо известны в данной области техники. В другом случае можно синтезировать и добавить и объединить при биологических условиях цепи для того, чтобы определить, гибридизуются ли они друг с другом.
[0197] В настоящем описании термин «не образующий пару аналог нуклеотида» обозначает аналог нуклеоитида, который включает не образующую пару группу, включая, но неограничиваясь ими: 6-дезамино аденозин (небуларин), 4-Ме-индол, 3-нитропиррол, 5-нитроиндол, Ds, Pa, N3-Me рибо U, N3-Me рибо Т, N3-Me dC, N3-Me-dT, N1-Me-dG, N1-Me-dA, N3-этил-dC, N3-Me dC. Согласно некоторым вариантам реализации не образующий пару аналог нуклеотида представляет собой рибонуклеотид. Согласно другим вариантам реализации он представляет собой дезоксирибонуклеотид.
[0198] В настоящем описании термин «концевая функциональная группа» включает, но не ограничивается, группу галогена, спирта, амина, карбокси, сложного эфира, амида, альдегида, кетона, эфира.
[0199] «Нуклеотид без основания» или «аналог нуклеотида без основания» в настоящем описании и в данной области техники может обозначать псевдо-нуклеотид или нетрадиционную группу. В отличие от нуклеотида, который представляет собой мономерную единицу нуклеиновой кислоты, обычно состоящую из остатка сахара рибозы или дезоксирибозы, фосфата и основания (аденина, гуанина, тимина или цитозина в ДНК; аденина, гуанина, урацила или цитозина в РНК), у нуклеотида без основания или псевдо-нуклеотида отсутствует основание, и поэтому он не является нуклеотидом, в том смысле, в котором термин обычно понимают в данной области техники. Группы дезоксирибозы без основания включают, например, лишенные основания дезоксирибоза-3'-фосфат; 1,2-дидезокси-D-рибофураноза-3-фосфат; 1,4-ангидро-2-дезокси-D-рибитол-3-фосфат. Инвертированные группы дезоксирибозы без основания, включают инвертированную дезоксирибозу без основания; 3',5' инвертированный дезокси-5'-фосфат без основания.
[0200] Термин «блокирующая группа» (z") в настоящем описании включает группу, которая может быть ковалентно связана с 5'-концом (N')y, и включает группу рибозы без основания, группу дезоксирибозы без основания, модификации групп рибозы без основания и групп дезоксирибозы без основания, включая, модификации 2' О алкил; инвертированные группы рибозы без основания и дезоксирибозы без основания, и их модификации; С6-имино-Pi; зеркальный нуклеотид, включая L-ДНК и L-PHK; 5'ОМе нуклеотид; и аналоги нуклеотидов, включая, 4',5'-метилен нуклеотид; 1-(β-D-эритрофуранозил)нуклеотид; 4'-тио нуклеотид, карбоциклический нуклеотид; 5'-амино-алкил фосфат; 1,3-диамино-2-пропил фосфат, 3-аминопропил фосфат; 6-аминогексил фосфат; 12-аминододецил фосфат; гидроксипропил фосфат; 1,5-ангидрогексит нуклеотид; альфа-нуклеотид; трео-пентофуранозил нуклеотид; ациклический 3',4'-нуклеотид; 3,4-дигидроксибутил нуклеотид; 3,5-дигидроксипентил нуклеотид, 5'-5'-инвертированная группа без основания; 1,4-бутандиол фосфат; 5'-амино; и соединенные или не соединенные мостиком метилфосфонат и 5'-меркапто группы.
[0201] Известными блокирующими группами могут быть группы рибозы или дезоксирибозы без основания; инвертированные группы рибозы или дезоксирибозы без основания; С6-амино-Pi; зеркальный нуклеотид, включая L-ДНК и L-PHK. Молекулы нуклеиновой кислоты согласно настоящему описанию можно синтезировать с использованием одного или более инвертированных нуклеотидов, например, инвертированного тимидина или инвертированного аденина (например, смотрите публикацию Takei, et al., 2002. JBC 277(26):23800-06).
[0202] Термин «нетрадиционная группа» в настоящем описании обозначает ненуклеотидные группы, включая группу без основания, инвертированную группу без основания, углеводородную группу (алкил) и их производные, и также включает дезоксирибонуклеотид, модифицированный дезоксирибонуклеотид, зеркальный нуклеотид (L-ДНК или L-PHK), аналог нуклеотида, не образующий пару, и нуклеотид, соединенный со смежным нуклеотидом посредством 2'-5' межнуклеотидной фосфатной связи; мостиковые нуклеиновые кислоты, LHK и нуклеиновые кислоты, соединенные мостиком из этилена, нуклеотиды с модифицированными связями (например, РАСЕ) и нуклеотиды с модифицированными основаниями, а также дополнительные группы, обозначенные в настоящем описании как нетрадиционные группы.
[0203] В настоящем описании термин «подавлять», «снижать» или «уменьшать» в отношении экспрессии генов обозначает, что экспрессия гена или уровень молекул РНК или соответствующих молекул РНК, кодирующих один или несколько белков или субъединиц белков (например, мРНК) или активность одного или более белков или белковых единиц уменьшена ниже уровня, который наблюдается в отсутствии подавляющего фактора (такого как молекула нуклеиновой кислоты, например, siНК, например, имеющей структурные особенности согласно настоящему описанию); например, экспрессия может быть снижена на 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 10%, 5% или меньше, чем наблюдается в отсутствии ингибитора.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0204] На Фигуре 1 представлена столбцовая диаграмма, показывающая действие siНК GFP на различные репортерные клеточные линии. Клеточные линии получали с помощью введения лентивирусной конструкцией кДНК-GFP HSP47 человека или кДНК-GFP GP46 крысы в HEK293, клеточную линию фибросаркомы человека НТ1080, линию HSC человека hTERT или клеточную линию NRK. siНК или siНК против GFP отрицательного контроля вводили в клетки и измеряли флуоресценцию GFP. Результаты показали, что siНК против GFP снижает флуоресценцию в различной степени в разных клеточных линиях. Клеточные линии 293_HSP47-GFP и 293_GP46-GFP выбирали для скрининга siHsp47 по причине легкости, с которой они поддаются трансфекции и чувствительности к снижению флуоресценции.
[0205] На Фигуре 2 представлен набор столбцовых диаграмм, показывающих цитотоксичность и эффективность снижения различных siHsp47 в клеточных линиях 293_HSP47-GFP и 293_GP46-GFP. Результаты показали, что siHsp47-C, siHsp47-2 и siHsp47-2d эффективно понижают как HSP47 человека, так и GP46 крысы (гомолог hsp47 человека) без существенной цитотоксичности. siGp46A против GP46 не снижает HSP47 человека. Также заово созданные siHsp47 превосходили siGp46A в снижении GP46 крысы.
[0206] На Фигуре 3 представлена столбцовая диаграмма, показывающая эффект снижения различных siHsp47s на мРНК hsp47, измеренный методом TaqMan® количественной ПЦР с использованием линии HSC клеток человека hTERT. На оси Y представлен остаточный уровень мРНК hsp47. HSP47-C была самой эффективной среди всех исследованных siНК hsp47.
[0207] На Фигуре 4 представлена столбцовая диаграмма, показывающая влияние разных siНК hsp47 на экспрессию коллагена I в клетках hTERT. Уровень мРНК коллагена I определяли методом количественной ПЦР в режиме реального времени с использованием зонда TaqMan®. На оси Y представлен остаточный уровень мРНК экспрессии коллагена I. Результат показал, что уровень мРНК коллагена I значительно снижен в клетках, обработанных некоторьми кандидатами (siHsp47-2, siHsp47-2d и их комбинацией с siHsp47-1).
[0208] На Фигуре 5 показано снижение площади фиброза в печени у животных, обработанных siHSP47.
[0209] На Фигуре 6 представлен пример последовательности нуклеиновой кислоты кДНК мРНК hsp47 человека (SEQ ID NO:1; основана на кДНК, описанной под номером доступа в GenBank: NM_001235).
[0210] На Фигуре 7 представлен пример аминокислотной последовательности hsp47 человека (SEQ ID NO:2).
[0211] На Фигуре 8 представляет собой последовательность нуклеиновой кислоты, кодирующую белок, кДНК hsp47 человека (SEQ ID NO:59), которая соответствует нуклеотидам 230-1486 SEQ ID NO:1.
[0212] На Фигурах 9А-9I показана стабильность в плазме Соединения_1, Соединения_2, Соединения_3, Соединения_4, Соединения_5, Соединения_6, Соединения_7, Соединения_8 и Соединения_9, соответственно, по результатам окрашивания бромистым этидием.
[0213] На Фигурах 10A-10I показана направленная/ненаправленная активность Соединения_1, Соединения_2, Соединения_3, Соединения_4, Соединения_5, Соединения_6, Соединения_7, Соединения_8 и Соединения_9, соответственно. AS_CM показывает активность антисмысловой цепи соединения в отношении плазмиды, содержащей вставку с полным соответствием; AS_SM показывает активность антисмысловой цепи соединения в отношении плазмиды, содержащей вставку последовательности затравки; S_CM показывает активность смысловой цепи соединения в отношении плазмиды, содержащей вставку с полным соответствием. Все анализы проводили на клетках человека, кроме данных, представленных на Фигуре 10F, которые проводили на клетках REF52 крысы.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
РНК интерференция и молекулы нуклеиновой кислоты siНК
[0214] Термин РНК интерференция обозначает процесс специфичного в отношении последовательностей пострансляционного сайленсинга генов у животных, регулируемый короткими интерферирующими РНК (siРНК) (Zamore et al., 2000, Cell, 101, 25-33; Fire et al., 1998, Nature, 391, 806; Hamilton et al., 1999, Science, 286, 950-951; Lin et al., 1999, Nature, 402, 128-129; Sharp, 1999, Genes & Dev., 13:139-141 и Strauss, 1999, Science, 286, 886). Соответствующий процесс у растений (Heifetz et al., международная публикация РСТ № WO 99/61631) часто обозначают термином посттранскрипционный сайленсинг генов (PTGS) или РНК сайленсинг. Считается, что процесс посттранскрипционного сайленсинга генов является эволюционно консервативным механизмом защиты клетки, используемым для предотвращения экспрессии чужих генов (Fire et al., 1999, Trends Genet., 15, 358). Такая защита от экспрессии чужих генов могла появиться в ответ на образование двухцепочечных РНК (dsPHK), полученных в результате вирусной инфекции или от случайного встраивания транспозонов в геном хозяина, через клеточный ответ, который специфично разрушает гомологичную одноцепочечную РНК или РНК генома вируса. Наличие dsPHK в клетке запускает ответ РНКи посредством механизма, который на сегодняшний день полностью не описан. По-видимому, этот механизм отличается от других известных механизмов, в которых участвуют рибонуклеазы, чувствительные по отношению к двухцепочечным РНК, таких как ответ интерферонов, который возникает в результате активации, опосредованной dsPHK, протеинкиназы PKR и 2',5'-олигоаденилатциклазы, приводящий к неспецифичному расщеплению мРНК рибонуклеазой L (смотрите, например, Патенты США №6,107,094; 5,898,031; публикации Clemens et al., 1997, J. Interferon & Cytokine Res., 17, 503-524; Adah et al., 2001, Curr. Med. Chem., 8, 1189).
[0215] Присутствие длинных dsPHK в клетках стимулирует активность фермента рибонуклеазы III, обозначаемого как Dicer (Bass, 2000, Cell, 101, 235; Zamore et al., 2000, Cell, 101, 25-33; Hammond et al., 2000, Nature, 404, 293). Dicer участвует в процессинге dsPHK на короткие куски dsPHK, известные как малые интерферирующие РНК (siРНК) (Zamore et al., 2000, Cell, 101, 25-33; Bass, 2000, Cell, 101, 235; Berstein et al., 2001, Nature, 409, 363). Малые интерферирующие РНК, полученные в результате активности Dicer, обычно имеют приблизительно от 21 до 23 нуклеотидов в длину и включают приблизительно 19 дуплексов пар нуклеотидов (Zamore et al., 2000, Cell, 101, 25-33; Elbashir et al., 2001, Genes Dev., 15, 188). Также Dicer участвует в вырезании малых временных РНК длиной 21 и 22 нуклеотида (stPHK) из РНК-предшественника консервативной структуры, которые участвуют в контроле трансляции (Hutvagner et al., 2001, Science, 293, 834). Ответ РНКи также включает комплекс эндонуклеазы, который обычно обозначают термином РНК-индуцированный комплекс сайленсинга (RISC), который опосредует расщепление одноцепочечной РНК, имеющей последовательность, комплементарную антисмысловой цепи дуплекса siРНК. Расщепление РНК-мишени происходит посередине участка, комплементарного антисмысловой цепи дуплекса siРНК (Elbashir et al., 2001, Genes Dev., 15, 188).
[0216] РНКи исследовали в различных системах. Fire et al., 1998, Nature, 391, 806 впервые наблюдали РНКи у С.elegans. В публикациях Bahramian и Zarbl, 1999, Molecular and Cellular Biology, 19, 274-283 и Wianny и Goetz, 1999, Nature Cell Biol., 2, 70, описывается РНКи, опосредованная dsPHK, у млекопитающих. Hammond et al., 2000, Nature, 404, 293, описывает РНКи в клетках дрозофилы, трансфицированных dsPHK. Elbashir et al., 2001, Nature, 411,494 и Tuschi et al., международная РСТ публикация № WO 01/75164, описывали РНКи, индуцированную при введении дуплексов синтетических РНК из 21 нуклеотида, в культурах клеток млекопитающих, включая клетки печени эмбриона человека и клетки HeLa. Недавняя работа с использованием лизатов эмбрионов дрозофилы (Elbashir et al., 2001, EMBO J., 20, 6877 и Tuschi et al., международная РСТ публикация № WO 01/75164) открыла несколько требований к структуре, длине, химическому составу и последовательности siРНК, которые необходимы для опосредования эффективной активности РНКи.
[0217] Молекулы нуклеиновых кислот (например, имеющие структурные особенности согласно настоящему описанию) могут подавлять или отрицательно регулировать экспрессию генов или репликацию вирусов посрердством опосредования РНК интерференции «РНКи» или сайленсинга генов специфичным в отношении последовательности образом, (см., например, публикацию Zamore et al., 2000, Cell, 101, 25-33; Bass, 2001, Nature, 411, 428-429; Elbashir et al., 2001, Nature, 411, 494-498; и Kreutzer et al., Международная публикация PCT № WO 00/44895; Zernicka-Goetz et al., Международная публикация PCT № WO 01/36646; Fire, Международная публикация PCT № WO 99/32619; Plaetinck et al., Международная публикация PCT № WO 00/01846; Mello and Fire, Международная публикация PCT № WO 01/29058; Deschamps-Depaillette, Международная публикация PCT № WO 99/07409; и Li et al., Международная публикация PCT № WO 00/44914; Allshire, 2002, Science, 297, 1818-1819; Volpe et al., 2002, Science, 297, 1833-1837; Jenuwein, 2002, Science, 297, 2215-2218; and Hall et al., 2002, Science, 297, 2232-2237; Hutvagner и Zamore, 2002, Science, 297, 2056-60; McManus et al., 2002, РНК, 8, 842-850; Reinhart et al., 2002, Gene & Dev., 16, 1616-1626; и Reinhart & Bartel, 2002, Science, 297, 1831).
[0218] Молекула нуклеиновой кислоты siHK может быть образована двумя отдельными полинуклеотидными цепями, при этом одна цепь является смысловой цепью, и вторая является антисмысловой, при этом антисмысловая и смысловая цепи являются комлементарными друг другу (т.е. каждая цепь включает последовательность нуклеотидов, которая комплементарна последовательности нуклеотидов во второй цепи); таким образом, что антисмысловая цепь и смысловая цепь образуют дуплекс или двухцепочечную структуру любой длины и структуры согласно настоящему описанию для молекул нуклеиновых кислот, например, при этом двухцепочечный участок (дуплексный участок) составляет от приблизительно 15 до приблизительно 49 (например, приблизительно 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48 или 49 пар оснований); антисмысловая цепь включает последовательность нуклеотидов, которая комплементарна последовательности нуклеотидов молекулы нуклеиновой кислоты-мишени (т.е. мРНК hsp47) или ее части, и смысловая цепь включает последовательность нуклеотидов, соответствующую последовательности нуклеиновой кислоты-мишени или ее части (например, приблизительно от 17 до 49 или больше нуклеотидов молекул нуклеиновых кислот в настоящем описании комплементарны нуклеиновой кислоте-мишени или ее части).
[0219] Согласно определенным аспектам и вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК), обеспечиваемая в настоящем описании, может представлять собой молекулу «длины RISC» или может являться субстратом Dicer, как более подробно описано ниже.
[0220] Молекула нуклеиновой кислоты siНК может включать отдельные смысловые и антисмысловые последовательности или участки, при этом смысловые и антисмысловые участки ковалентно соединены с помощью нуклеотидных или ненуклеотидных линкерных молекул, известных в данной области техники, или в другом случае не ковалентно соединены посредством ионных связей, водородных связей, Вад-дер-Ваальсовых связей, гидрофобных связей и/или стэкинг-взаимодействий. Молекулы нуклеиновых кислот могут включать последовательность нуклеотидов, которая комплементарна последовательности нуклеотидов гена-мишени. Молекулы нуклеиновых кислот могут взаимодействовать с нуклеотидной последовательностью гена-мишени таким образом, что это вызывает подавление экспрессии гена-мишени.
[0221] В другом случае молекула нуклеиновой кислоты siНК состоит из отдельного полинуклеотида, при этом комплементарные друг другу смысловой и антисмысловой участки молекулы нуклеиновой кислоты соединены посредством линкеров на основе нуклеиновой кислоты или не на основе нуклеиновой кислоты, т.е. антисмысловая цепь и смысловая цепь являются частью одного полинуклеотида, имеющего антисмысловой участок и смысловой участок, которые укладываются для образования дуплексного участка (например, для образования структуры «шпильки», как хорошо известно в данной области). Такие молекулы нуклеиновой кислоты siНК могут представлять собой полинуклеотид с ассимметричной дуплексной, шпилечной или ассимметричной вторичной шпилечной структурой, имеющей комлементарные друг другу смысловые и антисмысловые участки, при этом антисмысловой участок включает последовательность нуклеотидов, которая комплементарна последовательности нуклеотидов в отдельной молекуле нуклеиновой кислоты-мишени или ее части, и смысловой участок имеет последовательность нуклеотидов, соответствующую последовательности нуклеиновой кислоты - мишени (например, последовательности мРНК hsp47). Такие молекулы нуклеиновых кислот siНК могут представлять собой кольцевой одноцепочечный полинуклеотид, имеющий две или больше петельных структур и ствол, содержащий комплементарные смысловые и антисмысловые участки, при этом антисмысловой участок включает последовательность нуклеотидов, которая комплементарна последовательности нуклеотидов в молекуле нуклеиновой кислоты-мишени или ее части, и смысловой участок имеет последовательность нуклеотидов, соответствующую последовательности нуклеиновой кислоты - мишени или ее части, и при этом кольцевой полинуклеотид может подвергаться процессингу in vivo или in vitro с образованием активной молекулы нуклеиновой кислоты, способной опосредовать РНКи.
[0222] Следующую номенклатуру часто используют в данной области для описания длины и выступающих концов молекул siНК, и она используется в настоящем описании и Примерах. Названия, присваиваемые дуплексам, указывают длину олигомеров и наличие или отсутствие выступающих концов. Например, дуплекс «21+2» содержит две цепи нуклеиновых кислот, обе из которых имеют 21 нуклеотид в длину, также обозначают термином 21-мерный дуплекс siРНК или 21-мерная нуклеиновая кислота, и имеет 3'-выступающий конец из 2 нуклеотидов. «21-2» обозначает 21-мерный дуплекс нуклеиновых кислот с 5'-выступающим концом из 2 нуклеотидов. «21-02» представляет собой 21-мерный дуплекс нуклеиновых кислот без выступающих концов («тупые» концы). «21+2UU» представляет собой 21-мерный дуплекс с 3'-выступающим концом из 2 нуклеотидов, и оба 2 концевых нуклеотида на 3'-концах являются остатками U (что может приводить к несоответствию последовательности-мишени). Указанную выше номенклатуру можно применять к молекулам siНК с различной длиной цепей, дуплексов и выступающих концов (например, 19-0, 21+2, 27+2 и т.п). Согласно другой, но похожей номенклатуре «25/27» означает ассимметричный дуплекс, имеющий смысловую цепь из 25 оснований и антисмысловую цепь из 27 оснований с 3'-выступающим концом из 2 нуклеотидов. «27/25» означает ассимметричный дуплекс, имеющий смысловую цепь из 27 оснований и антисмысловую цепь из 25 оснований.
Химические модификации
[0223] Согласно определенным аспектам и вариантам реализации молекулы нуклеиновых кислот (например, молекулы siНК) согласно настоящему описанию включают одну или несколько модификаций (или химических модификаций). Согласно некоторым вариантам реализации такие модификации включают любые изменения в молекуле нуклеиновой кислоты или полинуклеотида, которые сделают молекулу отличной от стандартного рибонуклеотида или молекулы РНК (т.е. которая включает стандартные группы аденозина, цитозина, урацила или гуанозина); которую можно обозначать термином «немодифицированный» рибонуклеотид или немодифицированная рибонуклеиновая кислота. Традиционные основания ДНК и полинуклеотиды, имеющие 2'-дезоксисахар, представленные группами аденозина, цитозина, тимина или гуанозина, можно обозначать термином «немодифицированный дезоксирибонуклеотид» или «немодифицированная дезоксирибонуклеиновая кислота»; соответственно, термин «немодифицированный нуклеотид» или «немодифицированная нуклеиновая кислота» в настоящем описании обозначает «немодифицированный рибонуклеотид» или «немодифицированную рибонуклеиновую кислоту», если не указано иначе. Такие модификации могут происходить в сахаре нуклеотида, нуклеиновом основании, фосфатной группе нуклеотида и/или фосфатном остове полинуклеотида.
[0224] Согласно некоторьм вариантам реализации модификации согласно настоящему описанию можно использовать для повышения активности РНКи молекулы и/или для повышения in vivo стабильности молекул, в частности стабильности в сыворотке, и/или для повышения биологической доступности молекул. Примеры модификаций включают, но не ограничиваются, межнуклеотидные или межнуклеозидные связи; дезоксинуклеотиды или дидезоксирибонуклеотиды в любом положении и цепи молекулы нуклеиновой кислоты; нуклеиновую кислоту (например, рибонуклеиновую кислоту) с модификацией в 2'-положении, предпочтительно выбранную из амино, фтор, метокси, алкокси и алкила; 2'-дезоксирибонуклеотиды, 2'-O-метил рибонуклеотиды, 2'-дезокси-2'-фтор рибонуклеотиды, нуклеотиды с «универсальным основанием», «ациклические нуклеотиды», 5-С-метил нуклеотиды, биотиновую группу и вставки концевых глицериловых и/или инвертированных дезокси остатков без основания, пространственно затрудненные молекулы, такие как флуоресцентные молекулы и т.п. Другие модификаторы нуклеотидов могут включать 3-дезоксиаденозин (кордицепин), 3'-азидо-3'-дезокситимидин (AZT), 2',3'-дидезоксиинозин (ddI), 2',3'-дидезокси-3'-тиаацитидин (3ТС), 2',3'-дидегидро-2',3'-дидезокситимидин (d4T) и монофосфатные нуклеотиды 3'-азидо-3'-дезокситимидин (AZT), 2',3'-дидезокси-3'-тиаацитидин (3ТС) и 2',3'-дидегидро-2',3'-диде-окситимидин (d4T). Другие подробности различных модификаций более подробно описаны ниже.
[0225] Модифицированные нуклеотиды включают нуклеотиды, имеющие нозерн-конформацию (например, цикл псевдовращения по Нозерну, смотрите, например, Saenger, Principles of Nucleic Acid Structure, Springer-Verlag ed., 1984). Примеры нуклеотидов, имеющих нозерн-конформацию, включают, но не ограничиваются ими, нуклеотиды закрытой нуклеиновой кислоты (LNA) (например, нуклеотиды 2'-O, 4'-С-метилен-(D-рибофуранозил)); нуклеотиды 2'-метоксиэтокси (МОЕ); нуклеотиды 2'-метил-тио-этил, 2'-дезокси-2'-фтор, нуклеотиды 2'-дезокси-2'-хлор, нуклеотиды 2'-азидо и нуклеотиды 2'-O-метил. Закрытые нуклеиновые кислоты или LNA описаны, например, в публикации Elman et al., 2005; Kurreck et al., 2002; Crinelli et al., 2002; Braasch and Corey, 2001; Bondensgaard et al., 2000; Wahlestedt et al., 2000; в публикациях международных патентов № WO 00/47599, WO 99/14226, WO 98/39352 и WO 2004/083430. Согласно одному варианту реализации LNA встроена на 5'-конце смысловой цепи.
[0226] Химические модификации также включают незакрытые нуклеиновые кислоты или UNA, которые представляют собой ациклические ненуклеотидные аналоги, в которых отсутствует связь С2'-С3' (несмотря на то, что UNA не являются настоящими нуклеотидами, подчеркивается, что они включены в масштаб определения «модифицированных» нуклеотидов или модифицированных нуклеиновых кислот согласно настоящему описанию). Согласно определенным вариантам реализации молекулы нуклеиновой кислоты с выступающими концами могут быть модифицированы таким образом, что они содержат UNA в положениях выступающих концов (т.е. выступающий конец из 2 нуклеотидов). Согласно другим вариантам реализации UNA включены на 3'- или 5'-концах. UNA может быть расположена в любом месте цепи нуклеиновой кислоты, т.е. в положении 7. Молекулы нуклеиновой кислоты могут содержать одну или больше UNA. Примеры UNA описаны в публикации Nucleic Acids Symposium Series No. 52 p.133-134 (2008). Согласно некоторым вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию включает одну или более UNA или одну UNA. Согласно некоторым вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию, которая имеет выступающий 3'-конец, включает одну или более UNA в составе выступающего 3'-конца. Согласно некоторым вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию включает UNA (например, одну UNA) в антисмысловой цепи; например, в положении 6 или положении 7 антисмысловой цепи. Химические модификации также включают аналоги нуклеотидов, не образующие пару, например, согласно настоящему описанию. Также химические модификации включают нетрадиционные группы согласно настоящему описанию.
[0227] Также химические модификации включают терминальные модификации в 5' и/или 3' части олигонуклеотидов и их обозначают термином блокирующие группы. Такие терминальные модификации выбирают из нуклеотида, модифицированного нуклеотида, липида, пептида и сахара.
[0228] Также химические модификации включают шестичленные «аналоги шестичленных кольцевых нуклеотидов». Примеры аналогов шестичленных кольцевых нуклеотидов описаны в публикации Allart, et al (Nucleosides & Nucleotides, 1998, 17:1523-1526,; и Perez-Perez, et al., 1996, Bioorg. and Medicinal Chem Letters 6:1457-1460). Олигонуклеотиды, включающие аналоги шестичленных кольцевых нуклеотидов, включая мономеры нуклеотидов с гекситолом и альтриолом, описаны в публикации международной заявки на получение патента № WO 2006/047842.
[0229] Также химические модификации включают «зеркальные» нуклеотиды, которые имеют обращенную хиральность по сравнению с обычным природным нуклеотидом; т.е. зеркальный нуклеотид может представлять собой «L-нуклеотидный» аналог природного D-нуклеотида (смотрите. Патент США №6,602,858). Зеркальные нулеотиды могут также включать по меньшей мере одну модификацию сахара или основания и/или каркарса, например, согласно настоящему описанию, такую как группу фосфотиоата или фосфоната. В патенте США №6,602,858 описаны катализаторы нуклеиновых кислот, включающие замену по меньшей мере на один L-нуклеотид. Зеркальные нуклеотиды включают, например, L-ДНК (L-дезоксирибоаденозин-3'-фосфат (зеркальный dA); L-дезоксирибоцитидин-3'-фосфат (зеркальный dC); L-дезоксирибогуанозин-3'-фосфат (зеркальный dG); L-дезоксириботимидин-3'-фосфат (зеркальный dT)) и L-PHK (L-рибоаденозин-3'-фосфат (зеркальный rA); L-рибоцитидин-3'-фосфат (зеркальный rC); L-рибогуанозин-3'-фосфат (зеркальный rG); L-рибоурацил-3'-фосфат (зеркальный dU).
[0230] Согласно некоторым вариантам реализации модифицированные рибонуклеотиды включают модифицированные дезоксирибонуклеотиды, например, ДНК 5'ОМе (5-метил-дезоксирибогуанозин-3'-фосфат), который можно использовать в качестве нуклеотида в 5'-концевом положении (положение номер 1); РАСЕ (дезоксирибоаденозин 3' фосфоноацетат, дезоксирибоцитидин 3' фосфоноацетат, дезоксирибогуанозин 3' фосфоноацетат, дезоксириботимидин 3' фосфоноацетат).
[0231] Модификации могут присутствовать в одной или нескольких цепях молекулы нуклеиновой кислоты согласно настоящему описанию, например, в смысловой цепи, в антисмысловой цепи или в обеих цепях. Согласно некоторым вариантам реализации антисмысловая цепь может включать модификации, и смысловая цепь может включать только немодифицированную РНК.
Нуклеиновые основания
[0232] Нуклеиновые основания нуклеиновой кислоты согласно настоящему описанию могут включать немодифицированные рибонуклеотиды (пурины и пиримидины), такие как аденин, гуанин, цитозин, урацил. Нуклеиновые основания в одной или обеих цепях могут быть модифицированы природными и синтетическими нуклеиновыми основаниями, такими как тимин, ксантин, гипоксантин, инозин, 2-аминоаденин, 6-метил и другие алкильные производные аденина и гуанина, любые нуклеотиды с «универсальным основанием»; 2-пропил и другие алкильные производные аденина и гуанина, 5-галоурацил и цитозин, 5-пропинилурацил и цитозин, 6-азо урацил, цитозин и тимин, 5-урацил (псевдоурацил), 4-тиоурацил, 8-гало, амино, тиол, тиоалкил, гидроксил и другие 8-замещенные аденины и гуанины, 5-трифторметил и другие 5-замещенные урацилы и цитозины, 7-метилгуанин, деазапурины, гетероциклические замещенные аналоги пуринов и пиримидинов, например, аминоэтокси фенооксазин, производные пуринов и пиримидинов (например, 1-алкил-, 1-алкенил-, гетероароматические- и 1-алкинил производные) и их таутомеры, 8-оксо-N6-метиладенин, 7-диазаксантин, 5-метилцитозин, 5-метилурацил, 5-(1-пропинил)урацил, 5-(1-пропинил) цитозин и 4,4-этанцитозин). Другие примеры подходящих оснований включают непуриниловые и непиримидиловые основания, например, 2-аминопиридин и триазины.
Группы сахара
[0233] Сахарные группы в нуклеиновых кислоты согласно настоящему описанию могут включать группу сахара 2'-гидроксил-пентофуранозил без модификаций. В другом случае группы сахара могут быть модифицированы, например, 2'-дезокси-пентофуранозильная сахарная группа, D-рибоза, гексоза, модификации в 2' положении пентофуранозильной сахарной группы, такая как 2'-O-алкил (включая 2'-O-метил и 2*-O-этил), т.е. 2'-алкокси, 2'-амино, 2'-O-аллил, 2'-8-алкил, 2'-галоген (включая 2'-фтор, хлор и бром), 2'-метоксиэтокси, 2'-O-метоксиэтил, 2'-O-2-метоксиэтил, 2'-аллилокси (-ОСН2СН=СН2), 2'-пропаргил, 2'-пропил, этинил, этенил, пропенил, CF, циано, имидазол, карбоксилат, тиоат, С1-С10 низший алкил, замещенный низший алкил, алкиларил или аралкил, OCF3, OCN, О-, S- или N- алкил; О-, S или N-алкенил; SOCH3; S02CH3; ONO2; NO2, N3; гетероциклоалкил; гетероциклоалкиларил; аминоалкиламино; полиалкиламино или замещенный силили, как среди прочих, например, согласно описанию в европейских патентах ЕР 0586520 В1 или ЕР 0618925 B1.
[0234] Алкильные группы включают насыщенные алифатические группы, включая неразветвленные алкильные группы (например, метил, этил, пропил, бутил, пентил, гексил, гептил, октил, нонил, децил и т.д.), разветвленные алкильные группы (изопропил, трет-бутил, изобутил и т.д.), циклоалкильные группы (алициклические) (никлопропил, циклопентил, циклогексил, циклогептил, циклооктил), алкил-замещенные циклоалкильные группы и циклоалкил-замещенные алкильные группы. Согласно некоторым вариантам реализации неразветвленная или разветвленная алкильная цепь имеет 6 или меньше атомов углерода в своем остове (например, С1-С6 для неразветвленной цепи, С3-С6 для разветвленной цепи) и более предпочтительно 4 или меньше. Аналогично предпочтительные циклоалкилы могут иметь от 3-8 атомов углерода в структуре кольца и более предпочтительно имеют 5 или 6 атомов углерода в структуре кольца. Термин С1-С6 включает алкильные группы, содержащие 1-6 атомов углерода. Алкильную группу можно заместить на алкильную группу, например, алкильную группу, имеющую заместитель водорода у одного или более атомов углерода остова углероводорода. Такие заместители могут включать, например, алкенил, алкинил, галоген, гидроксил, алкилкарбонилокси, арилкарбонилокси, алкоксикарбонилокси, арилоксикарбонилокси, карбоксилат, алкилкарбонил, арилкарбонил, алкоксикарбонил, аминокарбонил, алкиламинокарбонил, диалкиламинокарбонил, алкилтиокарбонил, алкокси, фосфат, фосфонат, фосфинат, циано, амино (включая алкиламино, диалкилалкиламино, ариламино, диариламино и алкилариламино), ациламино (включая алкилкарбониламино, арилкарбониламино, карбомил и уреидо), амидино, имино, сульфургидрил, алкилтио, арилтио, тиокарбоксилат, сульфаты, алкилсульфинил, сульфонато, сульфамоил, сульфонамидо, нитро, трифторметил, циано, азидо, гетероциклическую, алкиларил или ароматическую или гетероароматическую группу.
[0235] Алкокси-группа включает замещенные и незамещенные алкильную, алкенильную и алкинильную группу, ковалентно связанную с атомом кислорода. Примеры алкокси-групп включают метокси, этокси, изопропилокси, пропокси, бутокси и пентокси-группы. Примеры замещенных алкокси-групп включают галогенированные алкокси-группы. Алкокси-группы могут быть замещены на группы, такие как алкенил, алкинил, галоген, гидроксил, алкилкарбонилокси, арилкарбонилокси, алкоксикарбонилокси, арилоксикарбонилокси, карбоксилат, алкилкарбонил, арилкарбонил, алкоксикарбонил, аминокарбонил, алкиламинокарбонил, диалкиламинокарбонил, алкилтиокарбонил, алкокси, фосфат, фосфонат, фосфинат, циано, амино (включая алкил амино, диалкиламино, ариламино, диариламино и алкилариламино), ациламино (включая алкилкарбониламино, арилкарбониламино, карбомоил и уреидо), амидино, имино, сульфгидрил, алкилтио, арилтио, тиокарбоксилат, сульфат, алкилсульфинил, сульфонат, сульфамоил, сульфонамидо, нитро, трифторметил, циано, азидо, гетероциклическую, алкиларил или ароматическую или гетероароматическую группы. Примеры галоген-замещенных алкокси-групп включают, но не ограничиваются, фторметокси, дифторметокси, трифторметокси, хлорметокси, дихлорметокси, трихлорметокси и т.д.
[0236] Согласно некоторым вариантам реализации пентафуранозильное кольцо может быть замещено на ациклические производные, в которых отсутствует С2'-С3'-связь пентафуранозильного кольца. Например, ациклические нуклеотиды могут замещать 2-гидроксиэтоксиметильную группу на 2'-дезоксирибофуранозильный сахар, обычно представленный в dNMP.
[0237] Галогены включают фториды, бромиды, хлориды, ийодиды.
Остов
[0238] Нуклеозиды нуклеиновой кислоты согласно настоящему описанию могут быть соединены друг с другом с помощью фосфодиэфирной связи. Фосфодиэфирная связь может быть заменена на другие связи. Например, фосфотиоат, тиофосфат-D-рибоза, триэфир, тиоат, остов, соединенный 2'-5' мостиками (также можно обозначать как 5'-2' или 2'5' нуклеотид или 2'5' рибонуклеотид), РАСЕ, 3'-(или-5')дезокси-3'-(или-5')тио-фосфотиоат, фосфодитиоат, фосфоселенаты, 3'-(или -5')дезоксифосфинаты, боранофосфаты, 3'-(или -5')дезокси-3'-(или 5'-)аминофосфоамидаты, гидроген фосфонаты, фосфонаты, боранофосфатные эфиры, фосфоамидаты, алкил или арил фосфонаты и фосфотриэфирные модификации, такие как алкилфосфотриэфиры, фосфотриэфирные фосфорные связи, 5'-этоксифосфодиэфир, Р-алкилоксифосфотриэфир, метилфосфонат и связи, не содержащие фосфор, например, карбонат, карбамат, силили, сульфур, сульфонат, сульфонамид, формацетал, тиоформацетил, оксим, метиленимино, метиленметилимино, метиленгидразо, метилендиметилгидразо и метиленоксиметилимино связи.
[0239] Молекулы нуклеиновой кислоты согласно настоящему описанию могут включать остов пептидной нуклеиновой кислоты (ПНК). Остов ПНК включает повторяющиеся единицы N-(2-аминоэтил)-глицина, соединенные пептидными связями. Различные основания, такие как пурин, пиримидин, природные и синттические основания соединены в остове посредством метилен-карбонильных связей.
Терминальные (концевые) фосфаты
[0240] Модификации можно осуществлять в концевых фосфатных группах. Примеры различных стабилизаторов, которые можно использовать, например, для того, чтобы стабилизировать 3′-конец последовательностей нуклеиновых кислот, включают, но не ограничиваются, (1) [3′-3′]-инвертированную дезоксирибозу; (2) дезоксирибонуклотид; (3) [5′-3′]-3′-дезоксирибонуклеотид; (4) [5′-3′]-рибонуклеотид; (5) [5′-3′]-3′-O-метил рибонуклеотид; (6) 3′-глицерил; (7) [3′-5′]-3′-дезоксирибонуклеотид; (8) [3′-3′]-дезоксирибонуклеотид; (9) [5′-2′]-дезоксирибонуклеотид и (10) [5′-3′]-дидезоксирибонуклеотид. Кроме немодифицированного остова стабилизаторы можно сочетать с одной или несколькими различными модификациями остова согласно настоящему описанию.
[0241] Примеры химически модифицированных концевых фосфатных групп включают указанные ниже:
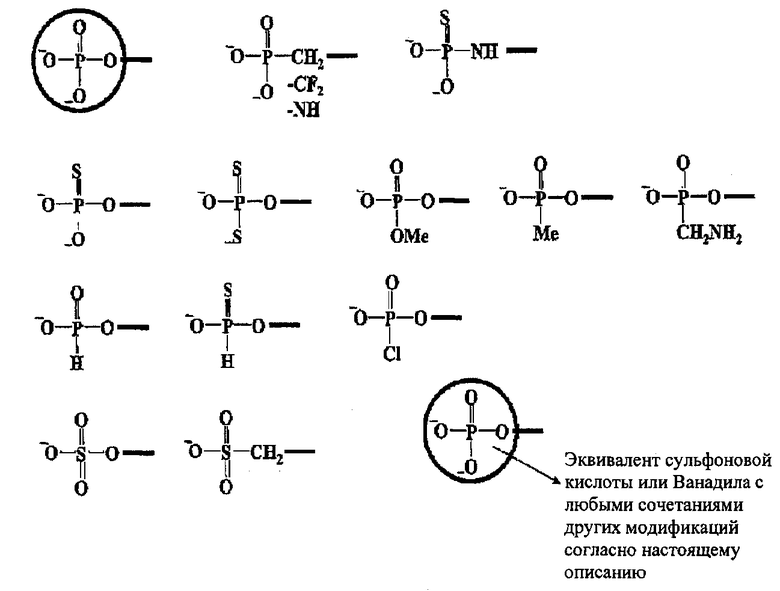
Конъюгаты
[0242] Модифицированные нулеотиды и молекулы нуклеиновых кислот (например, молекулы siHK) согласно настоящему описанию могут включать конъюгаты, например, конъюгат, ковалентно соединенный с химически модифицированной молекулой нуклеиновой кислоты. Примеры конъюгатов включают, но не ограничиваются, конъюгаты и лиганды согласно описанию в публикации Vargeese et al., U.S. Ser. No. 10/427,160. конъюгат может быть ковалентно соединен с молекулой нуклеиновой кислоты (например, молекулой siHK) с помощью биодеградируемого линкера. Молекула конъюгата может быть соединена с 3′-концом смысловой цепи и/или антисмысловой цепи химически модифицированной молекулы нуклеиновой кислоты. Молекула конъюгата может быть соединена с 5′- концом смысловой цепи и/или антисмысловой цепи химически модифицированной молекулы нуклеиновой кислоты. Молекула конъюгата может быть соединена как 3′-концом, так и 5′-концом смысловой цепи и/или антисмысловой цепи химически модифицированной молекулы нуклеиновой кислоты или любой их комбинации. Согласно одному варианту реализации молекула конъюгата может включать молекулу, которая облегчает доставку химически модифицированной нуклеиновой кислоты в биологическую систему, например, в клетку. Согласно другому варианту реализации молекула конъюгата, прикрепленная к химически модифицированной молекуле нуклеиновой кислоты, представляет собой полиэтиленгликоль, альбумин сыворотки человека или лиганд рецептора клетки, который опосредует поглощение клеткой. Примеры определенных молекул конъюгата согласно настоящему изобретению, которые могут быть присоединены к химически модифицированной молекуле нуклеиновой кислоты, описаны в публикации Vargeese et al., U.S. Ser. No. 10/201,394.
Линкеры
[0243] Молекула нуклеиновой кислоты согласно настоящему описанию (например, siНК) может содержать нуклеотидный, не состоящий из нуклеотидов или смешанный нуклеотидный/не состоящий из нуклеотидов линкер, который соединяет смысловой участок нуклеиновой кислоты и антисмысловой участок нуклеиновой кислоты. Нуклеотидный линкер может представлять собой линкер длиной ≥2 нуклеотидов, например, приблизительно 3, 4, 5, 6, 7, 8, 9 или 10 нуклеотидов в длину. Нуклеотидный линкер может представлять собой «аптамер» нуклеиновой кислоты. Термин «аптамер» или «аптамер нуклеиновой кислоты» в настоящем описании обозначает молекулу нуклеиновой кислоты, которая специфично связывается с молекулой мишенью, при этом молекула нуклеиновой кислоты имеет последовательность, которая включает последовательность, распознаваемую молекулой мишенью в естественных условиях. В другом случае аптамер может представлять собой молекулу нуклеиновой кислоты, которая связывается с молекулой-мишенью (например, мРНК hsp47), при этом молекула мишень не связывается с нуклеиновой кислотой в естественных условиях. Например, аптамер можно использовать для связывания с лиганд-связывающим доменом белка, предотвращая при этом взаимодействие природного лиганда с белком. Это является неограничивающим примером, и специалистам в данной области техники будет ясно, что другие варианты реализации можно будет с легкостью получить с использованием методик, обычно известных в данной области техники. Смотрите, например, публикации Gold et al.; 1995, Annu. Rev. Biochem., 64, 763; Brody and Gold, 2000, J. Biotechnol., 74, 5; Sun, 2000, Curr. Opin. Mol. Ther., 2, 100; Kusser, 2000, J. Biotechnol., 74, 27; Hermann and Patel, 2000, Science, 287, 820; и Jayasena, 1999, Clinical Chemistry, 45, 1628.
[0244] Не состоящий из нуклеотидов линкер может включать молекулу без основания, полиэфир, полиамин, полиамид, пептид, углевод, липид, полиуглевод или другие полимерные соединения (например, полиэтилен гликоль, например, имеющие от 2 до 100 групп этиленгликоля). Некоторые примеры включают примеры согласно описанию в публикации Seela and Kaiser, Nucleic Acids Res. 1990, 18:6353 и Nucleic Acids Res. 1987, 15:3113; Cload and Schepartz, J. Am. Chem. Soc. 1991, 113:6324; Richardson and Schepartz, J. Am. Chem. Soc. 1991, 113:5109; Ma et al., Nucleic Acids Res. 1993, 21:2585 and Biochemistry 1993, 32:1751; Durand et al., Nucleic Acids Res. 1990, 18:6353; McCurdy et al., Nucleosides & Nucleotides 1991, 10:287; Jschke et al., Tetrahedron Lett. 1993, 34:301; Ono et al., Biochemistry 1991, 30:9914; Arnold et al., Международная публикация № WO 89/02439; Usman et al., Международная публикация № WO 95/06731; Dudycz et al., Международная публикация № WO 95/11910 и Ferentz and Verdine, J. Am. Chem. Soc. 1991, 113:4000.
5'-концы, 3'-концы и выступающие концы
[0245] Молекулы нуклеиновой кислоты согласно настоящему описанию (например, молекулы siНК) могут иметь тупые концы с двух сторон, иметь выступающие («липкие») концы с обеих сторон или комбинацию тупых и липких концов. Выступающие концы могут быть как на 5'-, так и 3'-конце смысловой или антисмысловой цепи.
[0246] 5'- и/или 3'-концы двухцепочечной молекулы нуклеиновой кислоты (например, siНК) могут иметь тупые концы или выступающие концы. 5'-конец может быть тупым, и 3'-конец может иметь выступающий конец на смысловой или антисмысловой цепи. Согласно другим вариантам реализации 3'-конец может быть тупым, и 5'-конец может иметь выступающий конец на смысловой или антисмысловой цепи. Согласно другому варианту реализации как 5'-, так и 3'-конец являются тупыми, или как 5'-, так и 3'-концы имеют выступающие концы.
[0247] 5'- и/или 3'-конец одной или обеих цепей нуклеиновой кислоты могут включать свободную гидроксильную группу. 5'- и/или 3'-конец любой цепи молекулы нуклеиновой кислоты может быть модифицирован для включения химической модификации. Такая модификация может стабилизировать молекулы нуклеиновой кислоты, например, 3'-конец может иметь повышенную стабильность из-за наличия модификации молекулы нуклеиновой кислоты. Примеры модификаций конца (например, терминальные блокирующие группы) включают, но не ограничиваются, отсутствие основания (без основания), дезокси без основания, инвертированные (дезокси) без основания, глицерил, динуклеотид, ациклический нуклеотид, амино, фтор, хлор, бром, CN, CF, метокси, имидазол, карбоксилат, тиоат, C1-С10 низший алкил, замещенный низший алкил или арилалкил, OCF3, OCN, Ο-, S- или Ν-алкил; Ο-, S- или Ν-алкенил; SOCH3; SO2CH3; ONO2; NO2, N3; гетероциклоалкил; гетероциклоалкиларил; аминоалкиламино; полиалкиламино или замещенный силил, в том числе, описанные в Европейских патентах ЕР 586,520 и ЕР 618,925, и другие модификации согласно настоящему описанию.
[0248] Молекулы нуклеиновой кислоты включают молекулы с тупыми концами, т.е. концами, которые не включают свисающие нуклеотиды. Молекула нуклеиновой кислоты может включать один или несколько тупых концов. Молекула нуклеиновой кислоты с тупыми концами имеет некоторое количество пар нуклеотидов, равное числу нуклеотидов в каждой цепи молекулы нуклеиновой кислоты. Молекула нуклеиновой кислоты может включать один тупой конец, например, при этом 5′-конец антисмысловой цепи и 3′-конец смысловой цепи не имеет свисающих нуклеотидов. Молекула нуклеиновой кислоты может включать один тупой конец, например, при этом 3′-конец антисмысловой цепи и 5′-конец смысловой цепи не имеет свисающих нуклеотидов. Молекула нуклеиновой кислоты может включать два тупых конца, например, при этом 3′- конец антисмысловой цепи и 5′-конец смысловой цепи, а также 5′- конец антисмысловой цепи и 3′-конец смысловой цепи не имеет свисающих нуклеотидов. Другие нуклеотиды в молекулах нуклеиновых кислот с тупыми концами могут включать, например, ошибочно спаренные основания, петли, петли или неоднозначные пары оснований для модуляции активности молекулы нуклеиновой кислоты для опосредования РНК-интерференции.
[0249] Согласно некоторым вариантам реализации молекул нуклеиновой кислоты (например, молекул siHK) согласно настоящему описанию по меньшей мере один конец молекулы имеет выступающий конец длиной по меньшей мере один нуклеотид (например, 1-8 свисающих нуклеотидов). Например, одна или обе цепи двухцепочечной молекулы нуклеиновой кислоты согласно настоящему описанию могут иметь выступающий конец на 5′-конце и/или на 3′-конце. Выступающий конец может быть как на одной, так и на обеих смысловой и антисмысловой цепях молекулы нуклеиновой кислоты. Длина выступающего конца может составлять всего лишь один нуклеотид или целых 1-8 или больше нуклеотидов (например, 1, 2, 3, 4, 5, 6, 7 или 8 нуклеотидов); согласно некоторым предпочтительным вариантам реализации выступающий конец составляет 2, 3, 4, 5, 6, 7 или 8 нуклеотидов; например, выступающий конец может составлять всего 2 нуклеотида. Нуклеотид (нуклеотиды), образующий выступающий конец, может представлять собой дезоксирибонуклеотид(ы), рибонуклеотид(ы), природные и неприродные нуклеиновые основания или любой нуклеотид с модифицированным сахаром, основанием или фосфатной группой, например, согласно настоящему описанию. Двухцепочечная молекула нуклеиновой кислоты может иметь как 5'-, так и 3'-выступающие концы. Выступающие концы на 5'- и 3'-конце могут иметь разную длину. Выступающий конец может включать по меньшей мере одну модификацию нуклеиновой кислоты, которая может представлять собой дезоксирибонуклеотид. Один или несколько дезоксирибонуклеотидов могут быть расположены на 5'-конце. 3'-конец соответствующей противоположно направленной цепи молекулы нуклеиновой кислоты может не иметь выступающий конец, более предпочтительно не выступающий конец из дезоксирибонуклеотидов. Один или несколько дезоксирибонуклеотидов могут быть расположены на 3'-конце. 5'-конец соответствующей противоположно направленной цепи dsPHK может не иметь выступающий конец, более предпочтительно не выступающий конец из дезоксирибонуклеотидов. Выступающий конец на 5'- или 3'-конце цепи может составлять 1-8 (например, приблизительно 1, 2, 3, 4, 5, 6, 7 или 8) неспаренных нуклеотидов, предпочтительно, выступающий конец составляет 2-3 неспаренных нуклеотида; более предпочтительно 2 неспаренных нуклеотида. Молекулы нуклеиновой кислоты могут включать двуспиральные молекулы нуклеиновой кислоты с выступающими концами длиной приблизительно 1-20 (например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 1, 15, 16, 17, 18, 19 или 20); предпочтительно 1-8 (например, приблизительно 1, 2, 3, 4, 5, 6, 7 или 8) нуклеотидов, например, приблизительно дуплексы из 21 нуклеотида приблизительно с 19 парами оснований и 3'-выступающими концами из одного, двух или трех нуклеотидов. Молекулы нуклеиновой кислоты согласно настоящему изобретению могут включать дуплекс молекул нуклеиновой кислоты с тупыми концами, при этом оба конца являются тупыми или в другом случае один из концов является тупым. Молекулы нуклеиновой кислоты согласно настоящему описанию могут включать один или несколько тупых концов, т.е. при этом тупой конец не имеет свисающих нуклеотидов. Согласно одному варианту реализации молекула нуклеиновой кислоты с тупыми концами имеет число пар нуклеотидов, равное числу нуклеотидов в каждой цепи молекулы нуклеиновой кислоты. Молекула нуклеиновой кислоты может включать один тупой конец, например, при этом 5'-конец антисмысловой цепи и 3'-конец смысловой цепи не имеет свисающих нуклеотидов. Молекула нуклеиновой кислоты может включать один тупой конец, например, при этом 3'-конец антисмысловой цепи и 5'-конец смысловой цепи не имеет свисающих нуклеотидов. Молекула нуклеиновой кислоты может включать два тупых конца, например, при этом 3'-конец антисмысловой цепи и 5'-конец смысловой цепи, а также 5′- конец антисмысловой цепи и 3′-конец смысловой цепи не имеет свисающих нуклеотидов. Согласно некоторым предпочтительным вариантам реализации соединения нуклеиновой кислоты имеют тупые концы. Другие нуклеотиды молекулы siHK с тупыми концами могут включать, например, ошибочно спаренные основания, петли, петли или неоднозначные пары оснований для модуляции активности молекулы нуклеиновой кислоты для опосредования РНК-интерференции.
[0250] Во многих вариантах реализации один или несколько или все нуклеотиды выступающего конца молекулы нуклеиновой кислоты (например, молекулы siHK) согласно настоящему описанию модифицированы согласно настоящему описанию; например, один или несколько или все нуклеотиды могут представлять собой 2′-дезоксинуклеотиды.
Количество, расположение и распределение расположение (рисунок) модификаций.
[0251] Молекулы нуклеиновой кислоты (например, молекулы siHK) согласно настоящему описанию могут включать модифицированные нуклеотиды в процентном отношении от общего числа нуклеотидов в молекуле нуклеиновой кислоты. Поэтому молекула нуклеиновой кислоты может включать приблизительно от 5% до приблизительно 100% модифицированных нуклеотидов (например, приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% модифицированных нуклеотидов). Точная доля модифицированных нуклеотидов в данной молекуле нуклеиновой кислоты будет зависеть от общего числа нуклеотидов в нуклеиновой кислоте. Если молекула нуклеиновой кислоты является одноцепочечной, доля модификаций может зависеть от общего числа нуклеотидов в одноцепочечной молекуле нуклеиновой кислоты. Аналогично, если молекула нуклеиновой кислоты является двухцепочечной, доля модификаций зависит от общего числа нуклеотидов в смысловой цепи, антисмысловой цепи или как в смысловой, так и в антисмысловой цепях.
[0252] Молекулы нуклеиновой кислотысогласно настоящему описанию могут включать немодифицированные РНК в процентном отношении от общего числа нуклеотидов в молекуле нуклеиновой кислоты. Поэтому молекула нуклеиновой кислоты может включать приблизительно от 5% до приблизительно 100% модифицированных нуклеотидов (например, приблизительно 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% от общего числа нуклеотидов в молекуле нуклеиновой кислоты).
[0253] Молекула нуклеиновой кислоты (например, молекула siHK) может включать смысловую цепь, которая включает приблизительно от 1 до приблизительно 5, более точно приблизительно 1, 2, 3, 4 или 5 фосфотиоатных межнуклеотидных связей и/или один или несколько (например, приблизительно 1, 2, 3, 4, 5 или более) 2′-дезокси, 2′-O-метил, 2′-дезокси-2′-фтор и/или один или более (например, приблизительно 1, 2, 3, 4, 5, или более) модифицированных нуклеотидов с универсальным основанием, и, возможно, концевую блокирующую молекулу на 3′-конце или 5′-конце или как на 3′-, так и на 5′-конце смысловой цепи; и при этом антисмысловая цепь включает приблизительно от 1 до приблизительно 5 или более, более точно приблизительно 1, 2, 3, 4, 5 или более фосфотиоатных межнуклеотидных связей и/или одну или несколько (например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более) 2′-дезокси, 2′-O-метил, 2′-дезокси-2′-фтор и/или одну или более (например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более) модифицированных нуклеотидов с универсальным основанием, и, возможно, концевую блокирующую молекулу на 3′-конце, 5′-конце или как 3′-, так и 5′-конце антисмысловой цепи. Молекула нуклеиновой кислоты может включать приблизительно 1,2, 3, 4, 5, 6, 7, 8, 9, 10 или более пиримидиновых нуклеотидов смысловой и/или антисмысловой цепи нуклеиновой кислоты химически модифицированы 2′-дезокси, 2′-O-метил и/или 2′-дезокси-2′-фтор нуклеотидами с или без от приблизительно 1 до приблизительно 5 или более, например, приблизительно 1, 2, 3, 4, 5 или более фосфотиоатных межнуклеотидных связей и/или терминальной блокирующей молекулы на 3′-конце, 5′-конце или как 3′-, так и 5′-конце, находящихся на одной или разных цепях.
[0254] Молекула нуклеиновой кислоты может включать приблизительно от 1 приблизительно до 5 или более (в частности приблизительно 1, 2, 3, 4, 5 или более) фосфотиоатных межнуклеотидных связей в каждой цепи молекулы нуклеиновой кислоты.
[0255] Молекула нуклеиновой кислоты может включать 2′-5′ межнуклеотидные связи, например, на 3′-конце, 5′-конце или как на 3′-, так и на 5′-конце одной или обеих цепях нуклеиновой кислоты. Также в других различных положениях в пределах одной или обеих цепей последовательности нуклеиновой кислоты могут находиться 2′-5′ межнуклеотидные связи, например, приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более, включая каждую межнуклеотидную связь пиримидинового нуклеотида в одной или обеих цепях молекулы siHK, может включать 2′-5′ межнуклеотидную связь или приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более, включая каждую межнуклеотидную связь пуринового нуклеотида в одной или обеих цепях молекулы siНК, может включать 2'-5' межнуклеотидную связь.
[0256] Химически модифицированная молекула короткой интерферирующей нуклеиновой кислоты (siНК) может включать антисмысловую область, при этом любые (например, один или более или все) пиримидиновые нуклеотиды в антисмысловой области представляют собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды (например, при этом все пиримидиновые нуклеотиды представляют собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды или в другом случае множество пиримидиновых нуклеотидов представляют собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды), и при этом любые (например, один или более или все) пуриновые нуклеотиды в антисмысловой области представляют собой 2'-дезокси пуриновые нуклеотиды (например, при этом все пуриновые нуклеотиды представляют собой 2'-дезокси пуриновые нуклеотиды или в другом случае множество пуриновых нуклеотидов представляют собой 2'-дезокси пуриновые нуклеотиды).
[0257] Химически модифицированная молекула короткой интерферирующей нуклеиновой кислоты (siНК) может включать антисмысловую область, при этом любые (например, один или более или все) пиримидиновые нуклеотиды в антисмысловой области представляют собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды (например, при этом все пиримидиновые нуклеотиды представляют собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды или в другом случае множество пиримидиновых нуклеотидов представляет собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды), и при этом любые (например, один или более или все) пуриновые нуклеотиды в антисмысловой области представляют собой 2'-O-метил пуриновые нуклеотиды (например, при этом все пуриновые нуклеотиды представляют собой 2'-O-метил пуриновые нуклеотиды или в другом случае множество пуриновых нуклеотидов представляет собой 2'-O-метил пуриновые нуклеотиды).
[0258] Химически модифицированная молекула короткой интерферирующей нуклеиновой кислоты (siНК), способная опосредовать РНК-интерференцию (РНКи) против hsp47 внутри клетки или в системе in vitro, может включать смысловую область, при этом один или несколько пиримидиновых нуклеотидов в смысловой области представляют собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды (например, при этом все пиримидиновые нуклеотиды представляют собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды, или в другом случае множество пиримидиновых нуклеотидов представляет собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды), и один или несколько пуриновых нуклеотидов в смысловой области представляют собой 2'-дезокси пуриновые нуклеотиды (например, при этом все пуриновые нуклеотиды представляют собой 2'-дезокси пуриновые нуклеотиды или в другом случае множество пуриновых нуклеотидов представляет собой 2'-дезокси пуриновые нуклеотиды), и антисмысловую область, при этом один или более пиримидиновых нуклеотидов антисмысловой области представляют собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды (например, при этом все пиримидиновые нуклеотиды представляют собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды, или в другом случае множество пиримидиновых нуклеотидов представляет собой 2'-дезокси-2'-фтор пиримидиновые нуклеотиды), и один или более пуриновых нуклеотидов в антисмысловой области представляют собой 2'-O-метил пуриновые нуклеотиды (например, при этом пуриновые нуклеотиды представляют собой 2'-O-метил пуриновые нуклеотиды, или в другом случае множество пуриновых нуклеотидов представляет собой 2'-O-метил пуриновые нуклеотиды). Смысловая область и/или антисмысловая область могут иметь модификацию терминальной защитной группы, например, любую модификацию, которая по желанию находится на 3'-конце, 5'- конце или 3'- и 5'-концах смысловой и/или антисмысловой последовательности. Также дополнительно смысловая область и/или антисмысловая область могут включать 3'-концевой выступающий конец из нуклеотидов приблизительно от 1 до 4 (например, приблизительно 1, 2, 3 или 4) 2'-дезоксинуклеотидов. Также нуклеотиды выступающего конца могут включать одну или несколько (например, приблизительно 1,2, 3,4 или более) фосфотиоатную, фосфоноацетатную и/или тиофосфоноацетатную межнуклеотидные связи. Пуриновые нуклеотиды в смысловой области в другом случае могут представлять собой 2'-O-метил пуриновые нуклеотиды (например, при этом все пуриновые нуклеотиды представляют собой 2'-O-метил пуриновые нуклеотиды, или в другом случае множество пуриновых нуклеотидов представлет собой 2'-O-метил пуриновые нуклеотиды) и один или несколько пуриновых нуклеотидов антисмысловой области представляют собой 2'-O-метил пуриновые нуклеотиды (например, при этом все пуриновые нуклеотиды представляют собой 2'-O-метил пуриновые нуклеотиды, или в другом случае множество пуриновых нуклеотидов представляет собой 2'-O-метил пуриновые нуклеотиды). Один или несколько пуриновых нуклеотидов смысловой области могут в другом случае представлять собой пуриновые рибонуклеотиды (например, при этом все пуриновые нуклеотиды представляют собой пуриновые рибонуклеотиды или в другом случае множество пуриновых нуклеотидов представляют собой пуриновые рибонуклеотиды) и любые пуриновые нуклеотиды в антисмысловой области представляют собой 2'-O-метил пуриновые нуклеотиды (например, при этом все пуриновые нуклеотиды представляют собой 2'-O-метил пуриновые нуклеотиды, или в другом случае множество пуриновых нуклеотидов представляет собой 2'-O-метил пуриновые нуклеотиды). В другом случае один или несколько пуриновых нуклеотидов в смысловой области и/или антисмысловой области можно выбрать из группы, состоящей из 2'-дезоксинуклеотидов, нуклеотидов закрытой нуклеиновой кислоты (LNA), 2'-метоксиэтилнуклеотидов, 4'-тионуклеотидов и 2'-O-метилнуклеотидов (например, при этом все пуриновые нуклеотиды выбирают из группы, состоящей из 2'-дезоксинуклеотидов, нуклеотидов закрытой нуклеиновой кислоты (LNA), 2'-метоксиэтилнуклеотидов, 4'-тионуклеотидов и 2'-O-метилнуклеотидов, или в другом случае множество пуриновых нуклеотидов выбирают из группы, состоящей из 2'-дезоксинуклеотидов, нуклеотидов закрытой нуклеиновой кислоты (LNA), 2'-метоксиэтилнуклеотидов, 4'-тионуклеотидов и 2'-O-метилнуклеотидов).
[0259] Согласно некоторым вариантам реализации молекула нуклеиновой кислоты (например, молекула siНК) согласно настоящему описанию включает модифицированный нуклеотид (например, один модифицированный нуклеотид) в антисмысловой цепи; например, в положении 6 или положении 7 антисмысловой цепи.
[0260] Рисунки модификаций и чередующиеся модификации
[0261] Молекулы нуклеиновых кислот (например, молекулы siНК) согласно настоящему описанию могут иметь последовательности модифицированных и немодифицированных нуклеиновых кислот. Последовательность модификаций нуклеотидов в протяженной цепи нуклеотидов может представлять собой модификацию, заключенную в пределах одного нуклеотида или группы нуклеотидов, которые ковалентно соединены друг с другом с помощью стандартных фосфодиэфирных связей или по меньшей мере частично с помощью фосфотиоатных связей. Соответственно, «последовательность» (паттерн) согласно настоящему описанию может, но необязательно должен включать повторяющиеся единицы. Примеры рисунков модификаций, которые могут быть использованы в отношении молекул нуклеиновых кислот (например, молекул siНК) согласно настоящему описанию, включают рисунки, описанные в Giese, патент США №7,452,987. Например, молекулы нуклеиновых кислот (например, молекулы siНК) согласно настоящему описанию включают молекулы, имеющие рисунки модификаций, такие как, похожие или такие же, как рисунки показанные схематично на Фигуре 2 в Giese Патент США №7,452,987.
[0262] Модифицированный нуклеотид или группа модифицированных нуклеотидов может находиться на 5'-конце или 3'-конце смысловой или антисмысловой цепи, фланкирующий нуклеотид или группа нуклеотидов находится с обеих сторон модифицированного нуклеотида или группы, при этом фланкирующий нуклеотид или группа может быть немодифицирована или не имеет ту же модификацию, что и предшествующий нуклеотид или группа нуклеотидов. Фланкирующий нуклеотид или группа нуклеотидов, тем не менее, может иметь другую модификацию. Эта последовательность модифицированного нуклеотида или группы модифицированных нуклеотидов, соответственно, и немодифицированного или иначе модифицированного нуклеотида или группы немодифицированных или иначе модифицированных нуклеотидов могут повторяться один или несколько раз.
[0263] В некоторых последовательностях 5'-концевой нуклеотид цепи представляет собой модифицированный нуклеотид, тогда как в других последовательностях 5'-концевой нуклеотид цепи представляет собой немодифицированный нуклеотид. В некоторых последовательностях 5'-конце цепи начинается с группы модифицированных нуклеотидов, тогда как в других последовательностях 5'-концевой конец представляет собой немодифицированную группу нуклеотидов. Этот рисунок модификаций может находиться на первой цепи и/или на второй цепи молекулы нуклеиновой кислоты.
[0264] Модифицированные нуклеотиды одной цепи молекулы нуклеиновой кислоты могут быть комплементарны в положении модифицированным или не модифицированным нуклеотидам или группам нуклеотидов другой цепи.
[0265] Может присутствовать фазовый сдвиг между модификациями или последовательностями модификаций на одной цепи относительно рисунка модификации другой цепи, при котором последовательности модификаций не перекрываются. Например, сдвиг происходит таким образом, что модифицированная группа нуклеотидов смысловой цепи соответствует немодифицированной группе нуклеотидов антисмысловой цепи и наоборот.
[0266] Может происходить частичный сдвиг рисунка модификаций, при котором модифицированные группы перекрываются. Группы модифицированных нуклеотидов в любой данной цепи могут по желанию иметь одинаковую длину, но могут иметь и разную длину. Аналогично, группы модифицированных нуклеотидов любой данной цепи могут иметь одинаковую длину или разную длину.
[0267] В некоторых последовательностях второй (предпоследний) нуклеотид на конце цепи представляет собой немодифицированный нуклеотид или начало группы немодифицированных нуклеотидов. Предпочтительно, этот немодифицированный нуклеотид или немодифицированная группа нуклеотидов располагается на 5′-конце смысловой и/или антисмысловой цепи и даже более предпочтительно на конце смысловой цепи. Немодифицированный нуклеотид или немодифицированная группа нуклеотидов может быть расположена на 5′-конце смысловой цепи. Согласно предпочтительному варианту реализации рисунок модификаций состоит из отдельных чередующихся модифицированных и немодифицированных нуклеотидов.
[0268] В некоторых случаях двухцепочечные молекулы нуклеиновых кислот включают 2′-O-метил модифицированный нуклеотид и немодифицированный нуклеотид, предпочтительно, нуклеотид, который не является 2′-O-метил модифицированным, встроены в обе цепи чередующимся образом, что приводит к последовательности чередующихся 2′-O-метил модифицированных нуклеотидов и нуклеотидов, которые могут быть немодифицированными или по меньшей мере не включают модификацию 2′-О-метил. Согласно некоторым вариантам реализации та же последовательность модификации 2′-O-метил и не модификации существует во второй цепи; согласно другим вариантам реализации чередующиеся 2′-O-метил модифицированные нуклеотиды находятся только в смысловой цепи и не представлены в антисмысловой цепи, и согласно другим вариантам реализации чередующиеся 2′-O-метил модифицированные нуклеотиды находятся только в антисмысловой цепи и не находятся в смысловой цепи. Согласно некоторым вариантам реализации существует фазовый сдвиг между двумя цепями, при котором 2′-O-метил модифицированный нуклеотид на первой цепи образует пары оснований с немодифицированным (ми) нуклеотидом (ми) на второй цепи и наоборот. Данное соглашение, т.е. спаривание оснований 2′-O-метил модифицированного и немодифицированного нуклеотида (дов) на обеих цепях является особенно предпочтительным согласно некоторым вариантам реализации. Согласно некоторым вариантам реализации последовательность чередующихся 2′-O-метил модифицированных нуклеотидов существует на протяжении всей молекулы нуклеиновой кислоты или всего дуплексного участка. Согласно другим вариантам реализации последовательность чередующихся 2′-O-метил модифицированных нуклеотидов существует только в части нуклеиновой кислоты или всего дуплексного участка.
[0269] При рисунке модификаций с «фазовым сдвигом» может быть предпочтительно, если антисмысловая цепь начинается с 2′-O-метил модифицированного нуклеотида на 5′-конце, в результате чего второй нуклеотид является немодифицированным, третий, пятый, седьмой и т.к. нуклеотиды опять, таким образом, являются 2′-O-метил модифицированными, тогда как второй, четвертый, шестой, восьмой и т.п. нуклеотиды являются немодифицированными нуклеотидами.
Примеры расположений и рисунков модификаций
[0270] В то время как примеры рисунков модификаций представлены ниже более подробно, предусматриваются все комбинации рисунков модификаций со всеми возможными характеристиками молекул нуклеиновых кислот согласно настоящему описанию и известными в данной области техники (например, характеристики включают, но не ограничиваются ими, длину смысловой цепи, длину антисмысловой цепи, длину дуплексного участка, длину выступающего одноцепочечного конца, при этом один или оба конца двухцепочечной молекулы нуклеиновой кислоты могут быть тупыми или «липкими» (выступающими), расположение модифицированной нуклеиновой кислоты, число модифицированных нуклеиновых кислот, типы модификаций, при этом двойная молекула нуклеиновой кислоты с выступающими концами может иметь одинаковое или разное количество нуклеотидов в выступающих концах с каждой стороны, при этом в молекуле нуклеиновой кислоты используется один или больше одного типа модификаций, и число смежных модифицированных/немодифицированных нуклеотидов). По отношению ко всем подробным примерам ниже, несмотря на то, что показано, что дуплексный участок составляет 19 нуклеотидов, молекулы нуклеиновой кислоты согласно настоящему описанию могут иметь дуплексный участок в пределах от 1 до 49 нуклеотидов в длину, тогда как каждая цепь дуплексного участка может независимо иметь 17-49 нуклеотидов в длину. Примеры рисунков модификаций приведены в настоящем описании.
[0271] Молекулы нуклеиновой кислоты могут иметь тупой конец (если n равно 0) с обоих концов, который включает один или непрерывный набор модифицированных нуклеиновых кислот. Модифицированная нуклеиновая кислота может находиться в любом положении на всем протяжении смысловой или антисмысловой цепи. Молекулы нуклеиновой кислоты могут включать группу приблизительно из 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48 или 49 смежных модифицированных нуклеотидов. Модифицированные нуклеиновые кислоты могут составлять до 1%, 2%, 3%, 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98% или 100% от цепи нуклеиновой кислоты. Модифицированные нуклеиновые кислоты в примерах непосредственно ниже могут находиться только в смысловой цепи, только в антисмысловой цепи или как в смысловой, так и в антисмысловой цепи.
[0272] Общие примеры строения нуклеиновой кислоты показаны ниже, где Х = нуклеотид смысловой цепи в дуплексном участке; Ха = нуклеотид выступающего 5'-конца в смысловой цепи; Xb = нуклеотид выступающего 3'-конца в смысловой цепи; Y = нуклеотид антисмысловой цепи в дуплексном участке; Ya = нуклеотид выступающего 3'-конца в антисмысловой цепи; Yb = нуклеотид выступающего 5'-конца в антисмысловой цепи; и М = модифицированный нуклеотид в дуплексном участке. А и b независимо друг от друга составляют 0-8 (например, 0, 1, 2, 3, 4, 5, 6, 7 или 8). X, Y, а и b независимо друг от друга модифицированы или немодифицированы. Смысловая и антисмысловая цепи независимо могут содержать каждая 17-49 нуклеотидов в длину. Примеры, представленные ниже, имеют дуплексный участок из 19 нуклеотидов; однако молекулы нуклеиновых кислот согласно настоящему описанию могут иметь дуплексный участок длиной нечто среднее между 17 и 49 нуклеотидами, и при этом каждая цепь независимо друг от друга имеет от 17 до 49 нуклеотидов в длину.
[0273] Другие примеры строения молекулы нуклеиновой кислоты показаны ниже, где Х = немодифицированные нуклеотиды смысловой цепи; x = немодифицированный нуклеотид выступающего конца смысловой цепи; Y = немодифицированные нуклеотиды антисмысловой цепи; y = немодифицированный нуклеотид выступающего конца антисмысловой цепи; и М = модифицированный нуклеотид. Смысловая и антисмысловая цепи независимо могут иметь 17-49 нуклеотидов в длину. Примеры, представленные ниже, имеют дуплексный участок из 19 нуклеотидов; однако молекулы нуклеиновой кислоты согласно настоящему описанию могут иметь дуплексный участок длиной нечто среднее между 17 и 49 нуклеотидами, и при этом каждая цепь независимо друг от друга имеет от 17 до 49 нуклеотидов в длину.

[0274] Молекулы нуклеиновых кислот могут иметь тупые концы с обеих сторон с чередующимися модифицированными нуклеиновыми кислотами. Модифицированные нуклеиновые кислоты могут быть расположены в любом положении на всем протяжении смысловой или антисмысловой цепи.

[0275] Молекулы нуклеиновых кислот с тупым 5'-концом и выступающим 3'-концом оканчиваются одной модифицированной нуклеиновой кислотой.
[0276] Молекулы нуклеиновой кислоты с выступающим 5'-концом и тупым 3'-концом оканчиваются одной модифицированной нуклеиновой кислотой.
[0277] Молекулы нуклеиновых кислот с выступающими концами с обеих сторон и все выступающие концы представляют собой модифицированные нуклеиновые кислоты. В структуре непосредственной ниже М представляет собой количество модифицированных нуклеиновых кислот, где n представляет собой целое число от 0 до 8 (т.е. 0, 1, 2, 3, 4, 5, 6, 7 и 8).
[0278] Молекулы нуклеиновых кислот с выступающими концами с обоих концов и некоторые нуклеотиды на выступающих концах представляют собой модифицированные нуклеотиды. В структурах непосредственно ниже М представляет собой число модифицированных нуклеотидов, x представляет собой число немодифицированных нуклеотидов выступающего конца в смысловой цепи, у представляет собой число немодифицированных нуклеотидов выступающего конца в антисмысловой цепи, при этом каждое n независимо представляет собой целое число от 0 до 8 (т.е. 0, 1, 2, 3, 4, 5, 6, 7 и 8), и каждый выступающий конец составляет максимально 20 нуклеотидов; предпочтительно максимум 8 нуклеотидов (модифицированных и/или немодифицированных).


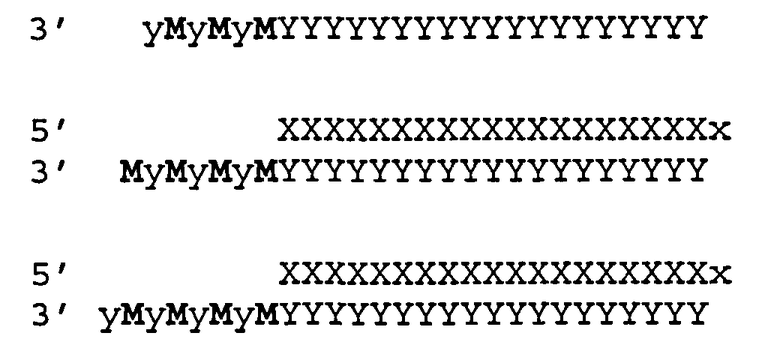

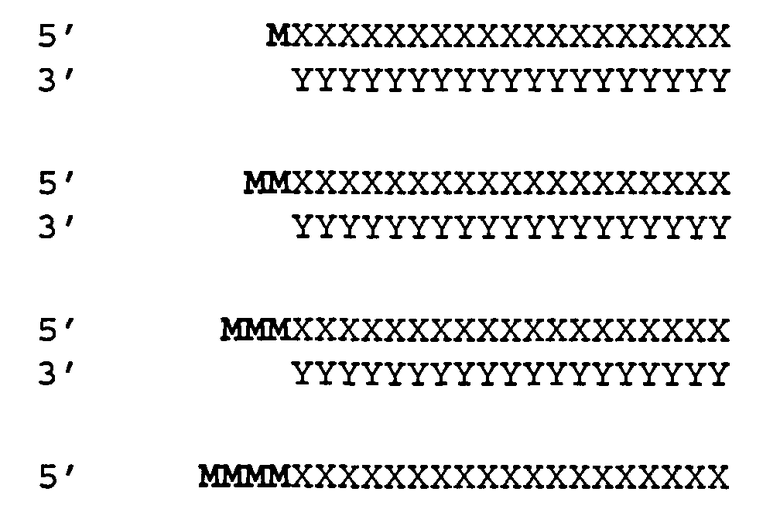


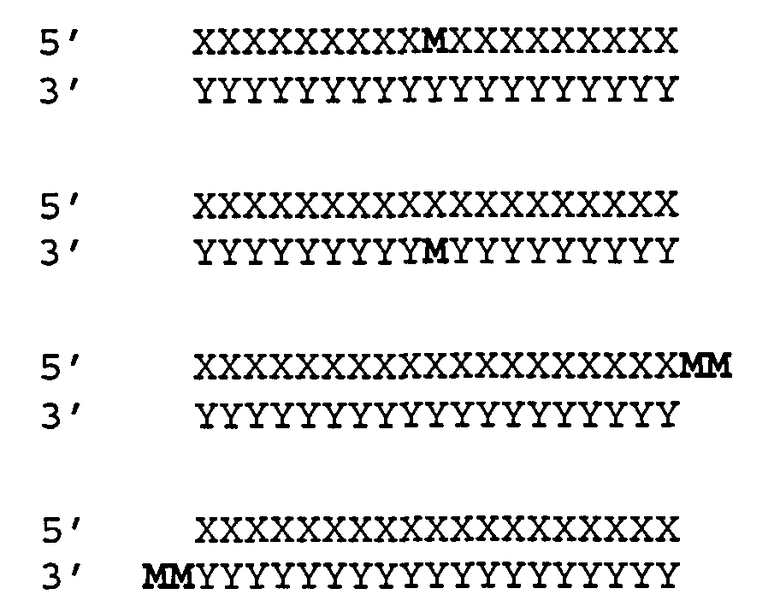
[0283] Примеры молекул нуклеиновых кислот представлены ниже вместе с эквивалентной общей структурой в соответствии с символами, использованными выше:
[0284] siHSP47-C siРНК к hsp47 человека и крысы, имеющая 19 нуклеотидный (т.е. 19 мерный) дуплексный участок и выступающие концы из 2 модифицированных нуклеотидов (т.е. дезоксинуклеотидов) на 3'-концах смысловой и антисмысловой цепи.
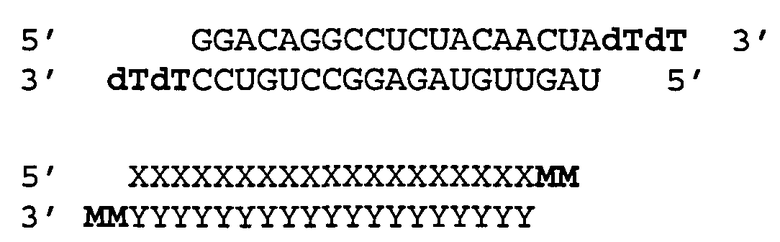
[0285] siHSP47-Cd siРНК к hsp47 человека и крысы, имеющая 25-мерный дуплексный участок, выступающий конец из 2 нуклеотидов на 3'-конце антисмысловой цепи и 2 модифицированных нуклеотиды в 5'-концевом и предпоследнем положениях смысловой цепи.
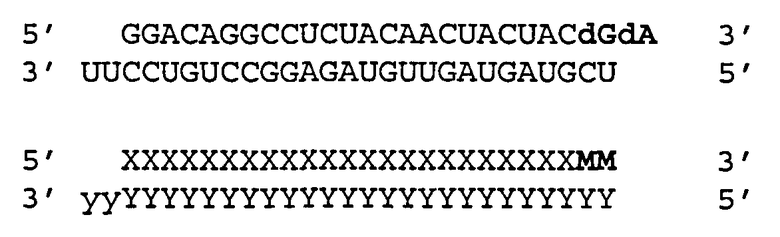
[0286] siHSP47-1 siРНК к кДНК 719-737 hsp47 человека и крысы, имеющая 19-мерный дуплексный участок и выступающие концы из 2 модифицированных нуклеотидов (т.е. дезоксинуклеотидов) на 3'-концах смысловой и антисмысловой цепи.
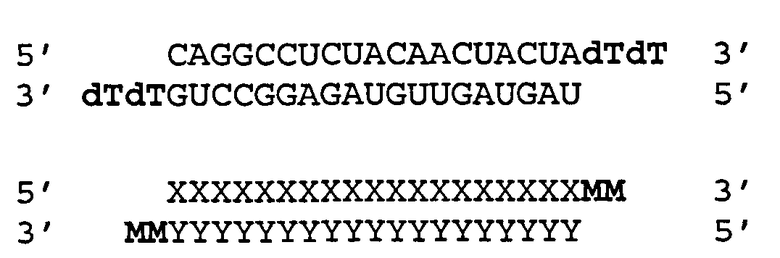
[0287] siHSP47-1d siРНК к 719-737 кДНК hsp47 человека и крысы, имеющая 25-мер с тупым концом на 3'-конце смысловой цепи и выступающий конец из 2 нуклеотидов на 3'-конце антисмысловой цепи и 2 модифицированных нуклеотида в 5'-концевом и предпоследнем положениях смысловой цепи.
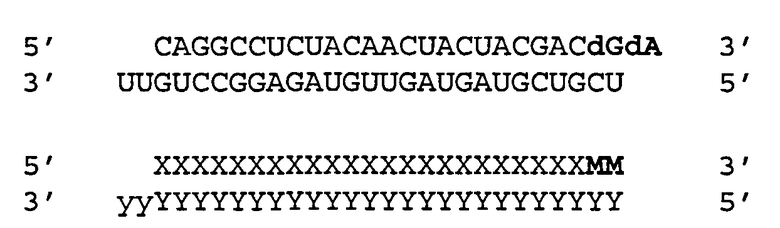
[0288] siHSP47-2 siРНК к 469-487 кДНК hsp47 человека, имеющая 19-мерный дуплексный участок и выступающие концы из 2 модифицированных нуклеотидов (т.е. дезоксинуклеотидов) на 3'-концах смысловой и антисмысловой цепи.
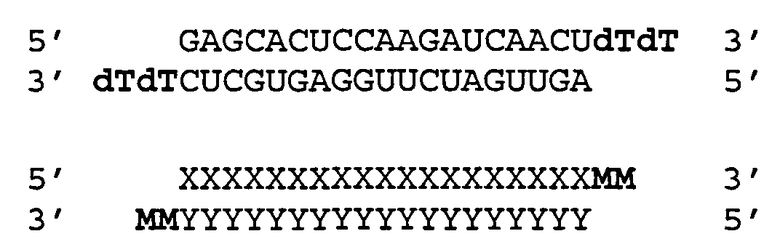
[0289] siHSP47-2d siРНК к 469-493 кДНК hsp47 человека, имеющая 25-мерный дуплексный участок с тупым концом на 3'-конце смысловой цепи и выступающий конец из 2 нуклеотидов на 3'-конце антисмысловой цепи и 2 модифицированных нуклеотида в 5'-концевом и предпоследнем положениях смысловой цепи.
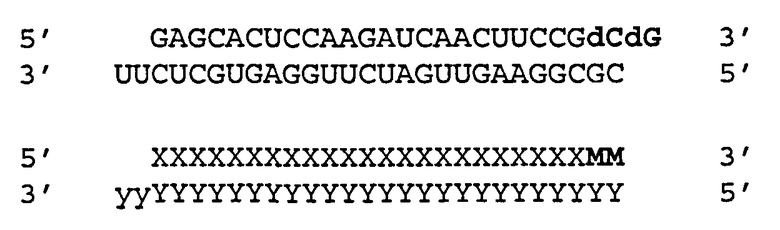
[0290] siHSP47-2d siРНК крысы к 466-490 кДНК Gp46 крысы, имеющая 25-мерный дуплексный участок с тупым концом на 3'-конце смысловой цепи и выступающий конец из 2 нуклеотидов на 3'-конце антисмысловой цепи и 2 модифицированных нуклеотида в 5'-концевом и предпоследнем положениях смысловой цепи.
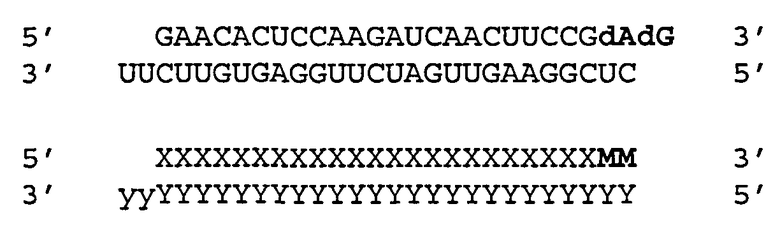
[0291] siHSP47-3 siРНК к 980-998 кДНК hsp47 человека, имеющая 19-мерный дуплексный участок и выступающие концы из 2 модифицированных нуклеотидов (т.е. дезоксинуклеотидов) на 3'-концах смысловой и антисмысловой цепи.
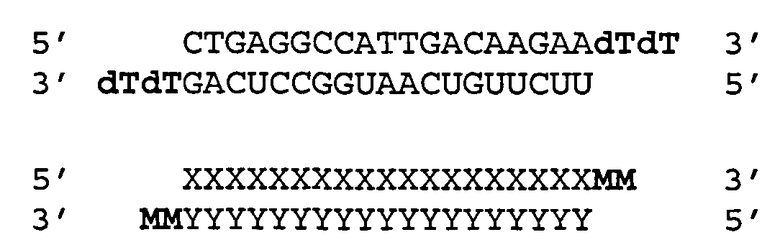
[0292] siHSP47-3d siРНК к 980-1004 кДНК hsp47 человека, имеющая 25-мерный дуплексный участок с тупым концом на 3'-конце смысловой цепи и выступающий конец из 2 нуклеотидов на 3'-конце антисмысловой цепи и 2 модифицированных нуклеотида в 5'-концевом и предпоследнем положениях смысловой цепи.
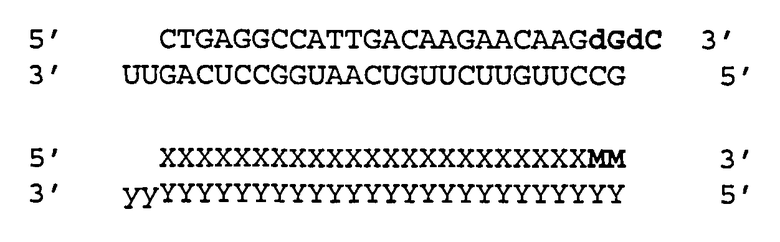
[0293] siHSP47-4 siРНК к 735-753 кДНК hsp47 человека, имеющая 19-мерный дуплексный участок и выступающие концы из 2 модифицированных нуклеотидов (т.е. дезоксинуклеотидов) на 3'-концах смысловой и антисмысловой цепи.
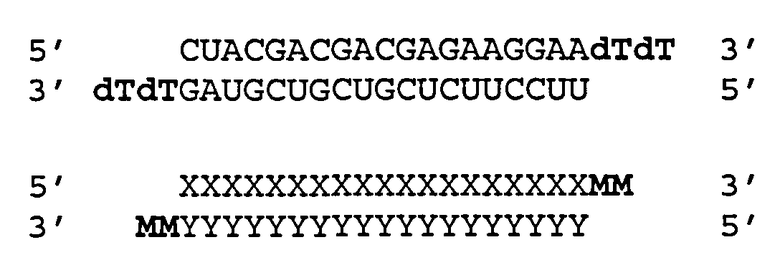
[0294] siHSP47-4d siРНК к 735-759 кДНК hsp47 человека, имеющая 25-мерный дуплексный участок с тупым концом на 3'-конце смысловой цепи и выступающим концом из 2 нуклеотидов на 3'-конце антисмысловой цепи и 2 модифицированных нуклеотида в 5'-концевом и предпоследнем положениях смысловой цепи.
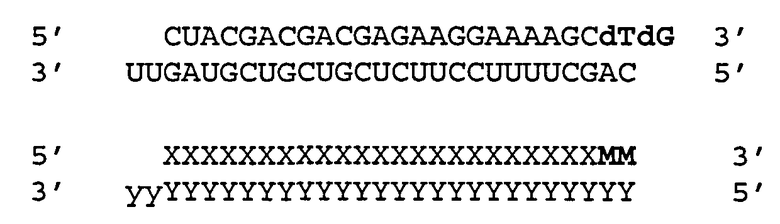
[0295] siHSP47-5 siРНК к 621-639 кДНК hsp47 человека, имеющая 19-мерный дуплексный участок и выступающие концы из 2 модифицированных нуклеотидов (т.е. дезоксинуклеотидов) на 3'-концах смысловой и анти смысловой цепи.
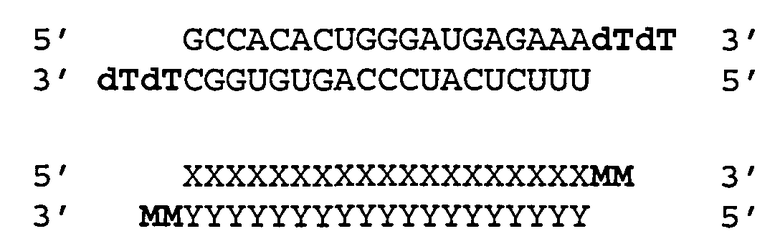
[0296] siHSP47-6 siРНК к 446-464 кДНК hsp47 человека, имеющая 19-мерный дуплексный участок и выступающие концы из 2 модифицированных нуклеотидов (т.е. дезоксинуклеотидов) на 3'-концах смысловой и антисмысловой цепи
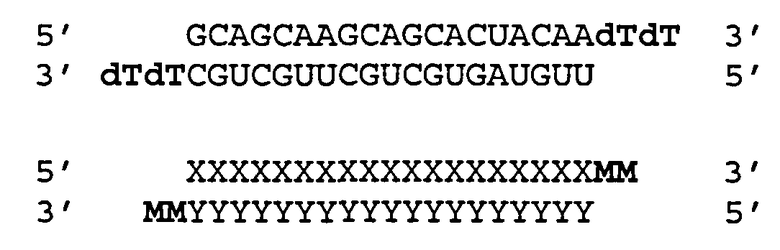
[0297] siHSP47-7 siРНК к 692-710 кДНК hsp47 человека, имеющая 19-мерный дуплексный участок и выступающие концы из 2 модифицированных нуклеотидов (т.е. дезоксинуклеотидов) на 3'-концах смысловой и антисмысловой цепи.
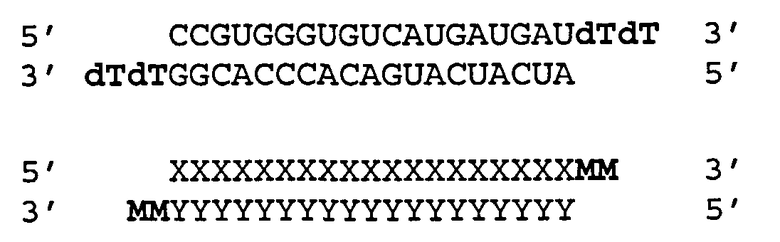
Разрывы и пробелы в цепях нуклеиновой кислоты
[0298] Молекулы нуклеиновой кислоты (например, молекулы siНК), обеспечиваемые в настоящем описании, могут иметь цепь, предпочтительно смысловую цепь, которая имеет разрывы или пробелы. В связи с этим молекулы нуклеиновой кислоты могут иметь три или более цепей, например, меродуплекс РНК (mdPHK), описанный в международной заявке на получение патента № PCT/US07/081836. Молекулы нуклеиновой кислоты с цепью, имеющей разрывы или пробелы, могут иметь от 1-49 нуклеотидов вдлину или иметь длину RISC (например, приблизительно 15-25 нуклеотидов) или длину субстрата Dicer (например, приблизительно 25-30 нуклеотидов), например, согласно настоящему описанию.
[0299] Молекулы нуклеиновой кислоты с тремя или более цепями включают, например, «А» (антисмысловую) цепь, «S1» (вторую) цепь и «S2» (третью) цепь, при этом «S1» и «S2» являются комплементарными и образуют пары оснований с неперекрывающимися участками цепи «А» (например, mdPHK может иметь структуру A:S1S2). Вместе S1, S2 или более цепей образуют то, что по существу похоже на смысловую цепь для антисмысловой «А» цепи. Двухцепочечный участок, образованный при гибридизации цепей «S1» и «А» отличается от и не перекрывается с двухцепочечным участком, образованным при гибридизации цепей «S2» и «А». Молекула нуклеиновой кислоты (например, молекула siНК) может представлять собой молекулу с «пробелами», что означает, что «пробел» составляет от 0 нуклеотидов до 10 нуклеотидов (например. О, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 нуклеотидов). Предпочтительно смысловая цепь имеет пробелы. Согласно некоторым вариантам реализации дуплекс A:S1 отделен от дуплекса A:S2 пробелом, возникающим из-за по меньшей мере одного неспаренного нуклеотида (приблизительно до 10 неспаренных нуклеотидов) в «А» цепи, который располагается между дуплексом A:S1 и дуплексом A:S2, и который отличается от любого одного или более неспаренного нуклеотида на 3'-конце одной или нескольких «A», «S1» или «S2» цепей. Дуплекс A:S1 может быть отделен от дуплекса А:В2 пробелом из 0 нуклеотидов (т.е. разрывом, при котором в молекуле полинуклеотида отсутствует или разорвана только фосфодиэфирная связь между двумя нуклеотидами) между дуплексом A:S1 и дуплексом A:S2, который также можно обозначать термином dsPHK с разрывами (ndsPHK). Например, A:S1S2 может включать dsPHK, имеющую по меньшей мере два двухцепочечных участка, которые содержат в общем от приблизительно 14 пар оснований до приблизительно 40 пар оснований, и двухцепочечные участки разделены пробелом из приблизительно от 0 до приблизительно 10 нуклеотидов, по желанию имеющую тупые концы, или A:S1S2 может включать dsPHK, имеющую по меньшей мере два двухцепочечных участка, разделенные пробелом до 10 нуклеотидов, при этом по меньшей мере один из двухцепочечных участков включает от приблизительно 5 пар нуклеотидов до 13 пар оснований.
Субстраты Dicer
[0300] Согласно некоторым вариантам реализации молекулы нуклеиновых кислот (например, молекулы siНК) согласно настоящему описанию могут представлять собой молекулу-предшественник «субстрата Dicer», например, двухцепочечную нуклеиновую кислоту, обработанную in vivo, для получения активных молекул нуклеиновых кислот, например, согласно описанию в публикации Rossi, Заявка на патент США №20050244858. В определенных условиях и ситуациях оказалось, что эти относительно более длинные типы siНК dsPHK, например, от приблизительно 25 до приблизительно 30 нуклеотидов, могут давать неожиданно эффективные результаты от отношении активности и продолжительности действия. Не желая быть связанными конкретной теорией, считается, что более длинные типы dsPHK служат субстратом для фермента Dicer в цитоплазме клетки. Кроме расщепления двухцепочечной нуклеиновой кислоты на более короткие сегменты, Dicer может облегчать встраивание одноцепочечных продуктов расщепления, полученных от расщепленных dsPHK в комплекс сайленсинга, вызванный РНК (комплекс RISC), который отвечает за разрушение цитоплазматической РНК, полученной от гена-мишени.
[0301] Субстраты Dicer могут иметь некоторые свойства, которые усиливают их процессинг Dicer. Субстраты Dicer имеют длину, достаточную для того, чтобы Dicer выполнял их процессинг, для получения активной молекулы нуклеиновой кислоты, и также могут включать одно или несколько из следующих свойств: (i) dsPHK является ассимметричной, например, имеет выступающий 3′-конец на первой цепи (антисмысловой цепи), и (ii) dsPHK имеет модифицированный 3′-конец на второй цепи (смысловой цепи), для направления ориентации связывания Dicer и процессинга dsPHK в активную siPHK. Согласно некоторым вариантам реализации самая длинная цепь в субстате Dicer может составлять 24-30 нуклеотидов.
[0302] Субстраты Dicer могут быть симметричными или ассимметричными. Субстрат Dicer может иметь смысловую цепь, которая включает 22-28 нуклеотидов, и антисмысловая цепь может включать 24-30 нуклеотидов, таким образом, согласно некоторым вариантам реализации полученный субстрат Dicer может иметь выступающий конец на 3′-конце антисмысловой цепи. Субстрат Dicer может иметь смысловую цепь из 25 нуклеотидов в длину, и антисмысловую цепь, имеющую 27 нуклеотидов в длину с выступающим концом из 2 оснований на 3′-конце. Выступающий конец может составлять 1-3 нуклеотида, например, 2 нуклеотида. Смысловая цепь также может иметь 5′ фосфат.
[0303] Также ассимметричный субстрат Dicer может содержать два дезоксинуклеотида на 3′-конце смысловой цепи вместо двух рибонуклеотидов. Некоторые примеры длин и структур субстратов Dicer представляют собой 21+0, 21+2, 21-2, 22+0, 22+1, 22-1, 23+0, 23+2, 23-2, 24+0, 24+2, 24-2, 25+0, 25+2, 25-2, 26+0, 26+2, 26-2, 27+0, 27+2 и 27-2.
[0304] Смысловая цепь субстрата Dicer может составлять приблизительно от 22 до приблизительно 30 (например, приблизительно 22, 23, 24, 25, 26, 27, 28, 29 или 30); приблизительно от 22 до приблизительно 28; приблизительно от 24 до приблизительно 30; приблизительно от 25 до приблизительно 30; приблизительно от 26 до приблизительно 30; приблизительно от 26 до приблизительно 29; или приблизительно от 27 до приблизительно 28 нуклеотидов в длину. Согласно некоторым предпочтительным вариантам реализации субстраты Dicer содержат смысловую и антисмысловую цепи, которые имеют по меньшей мере приблизительно 25 нуклеотидов в длину и не более, чем приблизительно 30 нуклеотидов; приблизительно от 26 до 29 нуклеотидов или 27 нуклеотидов в длину. Смысловая и антисмысловая цепи могут иметь одинаковую длину (с тупыми концами), различные длины (имеют выступающие концы) или их комбинацию. Смысловая и антисмысловая цепи могут находиться на одном полинуклеотиде или на разных полинуклеотидах. Субстрат Dicer может иметь дуплексный участок из приблизительно 19, 20, 21, 22, 23, 24, 25 или 27 нуклеотидов.
[0305] Как и другие молекулы siНК согласно настоящему описанию антисмысловая цепь субстрата Dicer может иметь любую последовательность, которая гибридизуется с антисмысловой цепью при биологических условиях, например, в цитоплазме эукариотической клетки.
[0306] Субстраты Dicer могут иметь любые модификации в нуклеиновом основании, сахаре или фосфатном остове, известные в данной области техники и/или согласно настоящему описанию для других молекул нуклеиновых кислот (например, молекул siНК). Согласно некоторым вариантам реализации субстраты Dicer могут иметь смысловую цепь, модифицированную для процессинга Dicer с помощью приемлемых модификаторов, расположенных на 3'-конце смысловой цепи, т.е. dsPHK создана для направления ориентации связывания и процессинга Dicer. Приемлемые модификаторы включают нуклеотиды, например, дезоксирибонуклеотиды, дидезоксирибонуклеотиды, ациклические нуклеотиды и т.п. и стерически затрудненные молекулы, такие как флуоресцентные молекулы и т.п. Ациклические нуклеотиды замещают 2-гидроксиэтоксиметильную группу на 2'-дезоксирибофуранозил сахар, обычно представленный в dNMP. Другие нуклеотидные модификаторы, которые могут быть использованы в качестве молекул siНК субстратов Dicer включают 3'-дезоксиаденозин (кордицепин), 3'-азидо-3'-дезокситимидин (AZT), 2',3'-дидезоксиинозин (ddI), 2',3'-дидезокси-3'-тиацитидин (3ТС), 2',3'-дидегидро-2',3'-дидезокситимидин (d4T) и монофосфатные нуклеотиды 3'-азидо-3'-дезокситимидина (AZT), 2',3'-дидезокси-3'-тиацитидина (3ТС) и 2',3'-дидегидро-2',3'-дидезокситимидина (d4T). Согласно одному варианту реализации дезоксинуклеотиды используют в качестве модификаторов. Если используют модификаторы нуклеотидов, они могут заменить рибонуклеотиды (например, 1-3 нуклеотидные модификаторы или 2 нуклеотидные модификаторы замещаются на рибонуклеотиды на 3'-конце смысловой цепи), при этом длина субстратов Dicer не меняется. Если используют стерически затрудненные молекулы, они могут быть прикреплены к рибонуклеотиду на 3'-конце антисмысловой цепи. Таким образом, согласно некоторым вариантам реализации длина цепи не меняется при встраивании модификаторов. Согласно некоторым вариантам реализации два основания ДНК в dsPHK являются замещенными для направления ориентации процессинга Dicer антисмысловой цепи. Согласно другому варианту реализации два концевых ДНК основания замещены на два рибонуклеотида на 3'-конце смысловой цепи, образующей тупой конец дуплекса на 3'-конце смысловой цепи и 5'-конце антисмысловой цепи, и выступающий конец из двух нуклеотидов РНК расположен на 3'-конце антисмысловой цепи. Эта композиция является ассимметричной с ДНК на тупом конце и РНК основаниями на выступающем (липком) конце.
[0307] Согласно некоторым вариантам реализации модификаторы включены в субстрат Dicer таким образом, что модификация не мешает молекуле нуклеиновой кислоты служить субстратом для Dicer. Согласно одному варианту реализации производят одну или несколько модификаций, которые усиливают процессинг Dicer субстрата Dicer. Может быть сделана одна или несколько модификаций, которые приводят к получению более эффективной РНКи. Может быть сделана одна или несколько модификаций, которые поддерживают больший эффект РНКи. Может быть сделана одна или несколько модификаций, которые приводят к большей мощности на каждый субстрат Dicer, который необходимо доставить в клетку. Модификации могут быть встроены в 3′-концевом участке, 5′-концевом участке, как в 3′-концевом, так и в 5′-концевом участке или в различных положениях в последовательности. Любое число и комбинация модификаций может быть встроена в субстрат Dicer до тех пор, пока модификация не мешает молекуле нуклеиновой кислоты служить субстратом для Dicer. Если присутствуют множественные модификации, они могут быть одинаковыми или различными. Предусматриваются модификации оснований, групп сахара, фосфатного остова и их комбинации. Любой 5′-конце может быть фосфорилирован.
[0308] Примеры модификаций фосфатного остова субстрата Dicer включают фосфонаты, включая метилфосфонат, фосфотиоат и фосфотриэфирные модификации, такие как алкилфосфотриэфиры и т.п. Примеры модификаций сахарных групп субстрата Dicer включают 2′-алкилпиримидин, например, 2′-O-метил, 2′-фтор, амино и дезокси модификации и т.п. (смотрите, например, Amarzguioui et al., 2003). Примеры модификаций групп оснований субстрата Dicer включают сахара без оснований, 2′-O-алкил модифицированные пиримидины, 4-тиоурацил, 5-бромурацил, 5-йодурацил и 5-(3-аминоаллил)-урацил и т.п. Закрытые нуклеиновые кислоты, или LHK, также могут быть включены.
[0309] Смысловая цепь может быть модифицирована для процессинга Dicer приемлемыми модификаторами, расположенными на 3′-конце смысловой цепи, т.е. субстрат Dicer разработан для направления ориентации связывания и процессинга Dicer. Приемлемые модификаторы включают нуклеотиды, такие как дезоксирибонуклеотиды, дидезоксирибонуклеотиды, ациклические нуклеотиды и т.п. и стерически затрудненные молекулы, например, флуоресцентные молекулы и т.п. Ациклические нуклеотиды замещают 2-гидроксиэтоксиметильную группу в 2'-дезоксирибофуранозном сахаре, обычно представленную в dNMP. Другие модификаторы нуклеотидов могут включать 3'-дезоксиаденозин (кордицепин), 3'-азидо-3'-дезокситимидин (AZT), 2',3'-дидезоксиинозин (ddI), 2',3'-дидезокси-3'-тиаацитидин (3ТС), 2',3'-дидегидро-2',3'-дидезокситимидин (d4T) и монофосфатные нуклеотиды 3'-азидо-3'-дезокситимидин (AZT), 2',3'-дидезокси-3'-тиацитидин (3ТС) и 2',3'-дидегидро-2',3'-дидезокситимидин (d4T). Согласно одному варианту реализации, дезоксинуклеотиды используются в качестве модификаторов. Если используются нуклеотидные модификаторы, 1-3 нуклеотидные модификаторы или 2-нуклеотидные модификаторы замещены на рибонуклеотиды на 3'-конце смысловой цепи. Если используют стерически затрудненные молекулы, их присоединяют к рибонуклеотиду на 3'-конце антисмысловой цепи. Таким образом, длина цепей не меняется при встраивании модификаторов. Согласно другому варианту реализации изобретение включает замещение двух оснований ДНК в субстрате Dicer для ориентации процессинга Dicer антисмысловой цепи. Согласно другому варианту реализации настоящего изобретения два концевых (терминальных) основания ДНК замещены на два рибонуклеотида на 3'-конце смысловой цепи, образуя тупой конец дуплекса на 3'-конце смысловой цепи и 5'-конце антисмысловой цепи, и выступающий конец из 2 нуклеотидов РНК расположен на 3'-конце антисмысловой цепи. Эта композиция является ассимметричной с ДНК на тупом конце и РНК основаниями на липком конце.
[0310] Антисмысловая цепь может быть модифицирована для процессиинга Dicer с помощью подходящих модификаторов, расположенных на 3'-конце антисмысловой цепи, т.е. dsPHK сконструирована для направления ориентации связывания и процессинга Dicer. Приемлемые модификаторы включают нуклеотиды, такие как дезоксирибонуклеотиды, дидезоксирибонуклеотиды, ациклические нуклеотиды и т.п. и стерически затрудненные молекулы, такие как флуоресцентные молекулы и т.п. Ациклические нуклеотиды замещают 2-гидроксиэтоксиметильную группу на 2'-дезоксирибофуранозный сахар, обычно представленный в dNMP. Другие модификаторы нуклеотидов могут включать 3'-дезоксиаденозин (кордицепин), 3'-азидо-3'-дезокситимидин (AZT), 2',3'-дидезоксиинозин (ddI), 2',3'-дидезокси-3'-тиаацитидин (3ТС), 2',3'-дидегидро-2',3'-дидезокситимидин (d4T) и монофосфатные нуклеотиды 3'-азидо-3'-дезокситимидин (AZT), 2',3'-дидезокси-3'-тиацитидин (3ТС) и 2',3'-дидегидро-2',3'-дидезокситимидин (d4T). Согласно одному варианту реализации, дезоксинуклеотиды используются в качестве модификаторов. Если используются нуклеотидные модификаторы, 1-3 нуклеотидные модификаторы или 2-нуклеотидные модификаторы замещаются на рибонуклеотиды на 3'-конце смысловой цепи. Если используются стерически затрудненные молекулы, они присоединены к рибонуклеотиду на 3'-конце антисмысловой цепи. Таким образом, длина цепи не меняется при встраивании модификаторов. Согласно другому варианту реализации изобретение включает замещение двух оснований ДНК в dsPHK для ориентации процессинга Dicer антисмысловой цепи. Согласно другому варианту реализации настоящего изобретения два концевых ДНК основания расположены на 3'-конце антисмысловой цепи вместо двух рибонуклеотидов, образуя тупой конец дуплекса на 5'-конце смысловой цепи и 3'-конце антисмысловой цепи, и выступающий конец из 2 нуклеотидов РНК расположен на 3'-конце смысловой цепи. Эта композиция является ассимметричной с ДНК на тупом конце и РНК основаниями на выступающем (липком) конце.
[0311] Субстраты Dicer со смысловой и антисмысловой цепью могут быть соединены с помощью третьей структуры. Третья структура не блокирует активность Dicer на субстрате Dicer и не мешает направленному разрушению РНК, транскрибированной с гена-мишени. Третья структура может представлять собой химическую связывающую группу. Приемлемые химические связывающые группы известны в данной области техники и могут быть использованы. В другом случае, третья структура может представлять собой олигонуклеотид, который связывает два олигонуклеотида dsPHK таким образом, что образуется структура шпильки при гибридизации двух олигонуклеотидов, составляющих субстрат Dicer. Структура шпильки предпочтительно не блокирует активность Dicer на субстрате Dicer или не мешает направленному разрушению РНК, транскрибированной с гена-мишени.
[0312] Не требуется, чтобы смысловая и антисмысловая цепи субстрата Dicer были полностью комплементарными. Нужно, чтобы они были существенно комплементарными для гибридизации при биологических условиях и для обеспечения субстрата для Dicer, который дает siРНК, достаточно комплементарную последовательности-мишени.
[0313] Субстрат Dicer может иметь некоторые свойства, которые усиливают его процессинг Dicer. Субстрат Dicer может иметь длину достаточную такую, что он процесируется Dicer с получением активной молекулы нуклеиновой кислоты (например, siРНК), и может иметь одно или несколько следующих свойств: (i) субстрат Dicer является ассимметрическим, например, имеет выступающий 3'-конец на первой цепи (антисмысловая цепь) и (ii) субстрат Dicer имеет модифицированный 3'-конец на второй цепи (смысловая цепь) для направления ориентации связывания и процессинга Dicer для получения активной siРНК. Субстрат Dicer может быть ассимметричным, так что смысловая цепь включает 22-28 нуклеотидов, и антисмысловая цепь включает 24-30 нуклеотидов. Таким образом, получаемый субстрат Dicer имеет выступающий 3'-конец антисмысловой цепи. Выступающий конец состоит из 1-3 нуклеотидов, например, 2 нуклеотидов. Смысловая цепь также может иметь 5'-фосфат.
[0314] Субстрат Dicer может иметь выступающий 3'-конец антисмысловой цепи, и смысловая цепь модифицирована для процессинга Dicer. 5'-конец смысловой цепи может иметь фосфат. Смысловая и антисмысловая цепи могут гибридизоваться при биологических условиях, например, условиях, встречающихся в цитоплазме клетки. Участок одной из цепей, в частности, антисмысловой цепи, субстрата Dicer может иметь последовательность длиной по меньшей мере 19 нуклеотидов, при этом указанные нуклеотиды находятся в 21-нуклеотидном участке, прилегающем к 3'-концу антисмысловой цепи, и достаточно комплементарны нуклеотидной последовательности РНК, синтезированной с гена-мишени. Субстрат Dicer также может иметь одно или больше из следующих дополнительных свойтсв: (а) антисмысловая цепь имеет сдвиг вправо от соответствующего 21-мера (т.е. антисмысловая цепь включает нуклеотиды с правой стороны молекулы, если сравнивать с соответствующим 21-мером), (b) цепи могут не быть полностью комплементарными, т.е. цепи могут содержать простые ошибочные спаренности оснований, и (с) модификации оснований, такие как закрытая нуклеиновая кислота (ы) могут быть включены на 5'-конце смысловой цепи.
[0315] Антисмысловая цепь молекулы нуклеиновой кислоты субстрата Dicer может быть модифицирована для включения 1-9 рибонуклеотидов на 5'-конце для получения длины 22-28 нуклеотидов. Если антисмысловая цепь имеет длину 21 нуклеотид, то 1-7 рибонуклеотидов или 2-5 рибонуклеотидов или 4 рибонуклеотида могут быть добавлены к 3'-концу. Добавленные рибонуклеотиды могут иметь любую последовательность. Несмотря на то, что добавленные рибонуклеотиды могут быть комплементарны последовательности гена-мишени, полной комлементарности между последовательностью-мишенью и антисмысловыми цепями не требуется. То есть, полученная антисмысловая цепь достаточно комплементарна последовательности-мишени. Затем смысловая цепь может иметь 24-30 нуклеотидов. Смысловая цепь быть достаточно комплементарна антисмысловой цепи для гибридизации с антисмысловой цепью при биологических условиях. Согласно одному варианту реализации антисмысловая цепь может быть синтезирована для включения модифицированного 3'-конца для направления процессинга Dicer. Смысловая цепь может иметь выступающий 3'-конец. Антисмысловая цепь может быть синтезирована для включения модифицированного 3'-конца для направления процессинга и связывания Dicer, и смысловая цепь имеет выступающий 3'-конец.
Белок теплового шока 47
[0316] Белок теплового шока 47 (HSP47) представляет собой молекулярный шаперон, специфичный в отношении коллагена, и располагается в эндоплазматическом ретикулуме. Он взаимодействует с коллагеном во время сворачивания, сбоки и транспортировки из эндоплазматического ретикулума (Nagata Trends Biochem Sci 1996; 21:22-6; Razzaque et al. 2005; Contrib Nephrol 2005; 148:57-69; Koide et al. 2006 J. Biol. Chem.; 281:3432-38; Leivo et al. Dev. Biol. 1980; 76:100-114; Masuda et al. J. Clin. Invest. 1994; 94:2481-2488; Masuda et al. Cell Stress Chaperones 1998; 3:256-264). Сообщали, что HSP47 имеет повышенную экспрессию при фиброзе различных тканей (Koide et al. J Biol Chem 1999; 274:34523-26), таких как цирроз печени (Masuda et al. J Clin Invest 1994; 94:2481-8), фиброз легких (Razzaque et al. Virchows Arch 1998; 432:455-60; Kakugawa et al. Eur Respir J 2004; 24:57-65) и гломерулосклероз (Moriyama et al. Kidney Int 1998; 54:110-19). Примеры последовтельности нуклеиновой кислоты кДНК-мишени человека hsp47 описываются под номером доступа GenBank: NM_001235 и соответствующей последовательности мРНК, например, представленной на SEQ ID NO:1. Специалисту в данной области техники будет ясно, что данная последовательность может меняться со временем и включать любые изменения, необходимые в молекулах нуклеиновых кислот в настоящем описании, соответственно.
[0317] Специфическая ассоциация HSP47 со спектром различных типов коллагена делает HSP47 возможной мишенью для лечения фиброза. Подавление экспрессии hsp47 может предотвращать секрецию внеклеточного коллагена I. Sato et al. (Nat Biotechnol 2008; 26:431-442) исследовал эту возможность посредством использования РНК для подавления экспрессии hsp47 и предотвращения развития фиброза печени у крыс. Аналогично, Chen et al. (Br J Dermatol 2007; 156:1188-1195) и Wang et al. (Plast. Reconstr Surg 2003; 111:1980-7) исследовали подавление экспрессии hsp47 с помощью технлогии РНК интерференции.
[0318] Способы и композиции для подавления hsp47
[0319] Обеспечиваются композиции и способы для подавления экспрессии HSP47 с использованием малых молекул нуклеиновых кислот, таких как молекулы малых интерферирующих нуклеиновых кислот (siНК), интерферирующих РНК (РНКи), коротких интерферирующих РНК (siРНК), двухцепочечных РНК (dsPHK), микро-РНК (miPHK) и малых шпильковых РНК (shPHK), способных опосредовать или которые опосредуют интерференцию РНК, против экспрессии гена hsp47. Композиции и способы согласно настоящему описанию также является полезными при лечении различного фиброза, например, фиброза печени, фиброза легких и фиброза почек.
[0320] Молекула (молекулы) нуклеиновых кислот и/или способы согласно настоящему изобретению используются для подавления экспрессии гена (генов), которые кодируют РНК, обозначаемые, например, номером доступа Genbank NM _001235.
[0321] Композиции, способы и наборы согласно настоящему описанию могут включать одну или несколько молекул нуклеиновых кислот (например, siНК) и способов, которые независимо или в комбинации модулируют (например, подавляют) экспрессию белка hsp47 и/или генов, кодирующих белки hsp47, белков и/или генов или нарушений, связанных с поддержанием и/или развитием заболеваний, состояний или нарушений, связанных с hsp47, таких как фиброз печени, цирроз, фиброз легких, фиброз почек, перитонеальный фиброз, хроническое поражение печени и фибриллогенез (например, гены, кодирующие последовательности, содержащие последовательности под номером доступа GenBank NM_001235), или члена семейства генов hsp47, при этом гены или семейство генов имеют гомологию последовательностей. Описание различных аспектов и вариантов реализации обеспечивается со ссылкой на примеры гена hsp47. Однако различные аспекты и варианты реализации также направлены на другие родственные гены hsp47, например, гены-гомологи и варианты транскриптов и полиморфизмы (например, полиморфизм отдельных нуклеотидов (SNP)), ассоциированные с определенными генами hsp47. По этой причине различные аспекты и варианты реализации также направлены на другие гены, которые участвуют в путях передачи сигнала или экспрессии генов, опосредуемых hsp47, которые вовлечены, например, в сохранение или развитие заболеваний, признаков или состояний согласно настоящему описанию. Можно провести анализ указанных дополнительных генов на предмет сайтов-мишеней, используя способы, описанные в настоящей публикации для гена hsp47. Таким образом, модуляцию других генов и эффекты такой модуляции других генов можно производить, определять и измерять согласно настоящему описанию.
[0322] Согласно одному варианту реализации композиции и способы согласно настоящему описанию включают молекулу двухцепочечной короткой интерферирующей нуклеиновой кислоты (siНК), которая подавляет экспрессию гена hsp47 (например, hsp47 человека, примером которого является SEQ ID NO:1), при этом молекула нуклеиновой кислоты включает от приблизительно 15 до приблизительно 49 пар оснований.
[0323] Согласно одному варианту реализации нуклеиновую кислоту согласно настоящему описанию можно использовать для подавления экспрессии гена hsp47 или семейства гена hsp47, при этом последовательность гена или семейства генов имеют гомологию последовательностей. Такие гомологичные последовательности можно идентифицировать способами, известным в данной области техники, например, с использованием выравнивания последовательностей. Молекулы нуклеиновой кислоты можно разработать для нацеливания на такие гомологичные последовательности, например, с использованием идеально комплементраных последовательностей или посредством встраивания неканонических пар оснований, например, ошибочных пар оснований и/или неоднозначных пар оснований, что может давать дополнительные последовательности-мишени. Если идентифицируют ошибочные спаривания, можно использовать неканонические пары оснований (например, ошибочные спаривания и/или неоднозначно спаренные основания) для получения молекул нуклеиновых кислот, которые нацелены более чем на одну последовательность гена. Например, но не ограничиваясь, неканонические пары оснований, такие как UU и СС пары оснований используются для получения молекул нуклеиновых кислот, которые способны нацеливаться на последовательности для различения hsp47-мишеней, которые имеют гомологию последовательностей. Поэтому одним из преимуществ использования siНК согласно настоящему описанию является то, что можно разработать одноцепочечную нуклеиновую кислоту, чтобы она включала последовательность нуклеиновой кислоты, которая комплементарна последовательности нуклеотидов, которая является консервативной среди гомологичных генов. При таком подходе отдельную нуклеиновую кислоту можно использовать для подавления экспрессии более чем одного гена вместо использования более чем одной молекулы нуклеиновой кислоты для воздействия на различные гены.
[0324] Молекулы нуклеиновых кислот можно использовать для нацеливания на консервативные последовательности, соответствующие семейству гена или семействам генов, таким как семейство генов hsp47. Поэтому молекулы нуклеиновой кислоты, нацеленные на различные мишени hsp47, могут обеспечивать повышенный терапевтический эффект. Кроме этого, нуклеиновую кислоту можно использовать для описания путей функции генов для различных применений. Например, молекулы нуклеиновых кислот можно использовать для подавления активности гена (генов)-мишени в путях для определения функции неохарактеризованного гена (генов) в анализе функции гена (генов), анализе функции мРНК или анализе трансляции. Молекулы нуклеиновых кислот можно использовать для определения возможных путей гена-мишени, участвующих в различных заболеваниях и состояниях для разработки лекарственных средств. Молекулы нуклеиновых кислот можно использовать для понимания путей экспрессии генов, вовлеченных, например, в фиброзы, такие как фиброз печени, почек или фиброз легких, и/или воспалительные и пролиферативные признаки, заболевания, нарушения и/или состояния.
[0325] Согласно одному варианту реализации композиции и способы согласно настоящему описанию включают молекулу нуклеиновой кислоты, имеющую активность РНКи против РНК hsp47, при этом молекула нуклеиновой кислоты включает последовательность, комплементарную любой РНК, имеющей кодирующую последовательность hsp47, такую как последовательности, имеющие последовательности, показанные в Таблице I. Согласно другому варианту реализации молекула нуклеиновой кислоты может иметь активность РНКи против РНК hsp47, при этом молекула нуклеиновой кислоты включает последовательность, комплементарную РНК, имеющей вариант кодирующей последовательности hsp47, например, другие мутантные гены hsp47, не показанные в Таблице I, но для которых в данной области техники известно, что они связаны с поддержанием и/или развитием фиброза. Химические модификации, показанные в Таблице I, или иные согласно настоящему описанию можно применять по отношению к любой конструкции нуклеиновой кислоты согласно настоящему описанию. Согласно другому варианту реализации молекула нуклеиновой кислоты согласно настоящему описанию включает нуклеотидную последовательность, которая может взаимодействовать с нуклеотидной последовательностью гена hsp47, и таким образом опосредовать сайленсинг экспрессии гена hsp47, например, при этом молекула нуклеиновой кислоты опосредует регуляцию экспрессии гена hsp47 через клеточные процессы, которые модулируют структуру хроматина или рисунки метилирования гена hsp47 и предотвращают транскрипцию гена hsp47.
[0326] Молекулы нуклеиновых кислот согласно настоящему описанию могут обладать активностью РНКи против РНК hsp47, при этом молекула нуклеиновой кислоты включает последовательность, комплементарную любой РНК, имеющей последовательность, кодирующую hsp47, например, последовательност под номером доступа GenBank NM_001235. Молекулы нуклеиновых кислот могут обладать активностью РНКи против РНК hsp47, при этом молекула нуклеиновой кислоты включает последовательность, комплементарную РНК, имеющей вариант кодирующей последовательности hsp47, например, другие мутантные гены hsp47, которые, как известно в данной области техники, связаны с поддержанием и/или развитием фиброза. Молекулы нуклеиновых кислот согласно настоящему описанию включают послдовательность нуклеотидов, которая может взаимодействовать с нуклеотидной последовательностью гена hsp47, и, таки образом, опосредовать сайленсинг экспрессии гена hsp47, например, если молекула нуклеиновой кислоты опосредует регуляцию экспрессии гена hsp47 через клеточные процессы, которые модулируют структуру хроматина или рисунки метилирования гена hsp47 и предотвращают транскрипцию гена hsp47.
Способы лечения
[0327] Специфическая ассоциация HSP47 со спектром различных типов коллагена делает hsp47 возможной мишенью для лечения фиброза. Подавление экспрессии hsp47 может предотвращать секрецию внеклеточного коллагена I. Sato et al. (Nat Biotechnol 2008; 26:431-442) исследовал эту возможность посредством использования siРНК для ингибирования экспрессии hsp47 и предотвращения развития фиброза печени у крыс. Аналогично, Chen et al. (Br J Dermatol 2007; 156:1188-1195) и Wang et al. (Plast. Reconstr Surg 2003; 111:1980-7) исследовали подавление экспрессии hsp47 с помощью теънологии РНК интерференции.
[0328] Согласно одному варианту реализации молекулы нуклеиновых кислот можно использовать для снижения или подавления экспрессии hsp47 и/или белков hsp47, происходящих от hsp47 и/или полиморфизмов гаплотипа hsp47, которые ассоциированы с заболеванием или состоянием (например, фиброзом). Анализ уровней hsp47 и/или генов hsp47, hsp47 и/или белка hsp47 или уровня РНК можно использовать для идентификации пациентов с такими полиморфизмами или пациентов, у которых есть риск развития признаков, состояний или заболеваний согласно настоящему описанию. Указанные пациенты поддаются лечению, например, лечению с помощью молекул нуклеиновых кислот согласно настоящему описанию и любой другой композиции, полезной для лечения заболеваний, связанных с hsp47 и/или экспрессией гена hsp47. Поэтому анализ уровней hsp47 и/или белка hsp47 или РНК можно использовать для определения типа и курса лечения для лечения пациента. Мониторинг уровней hsp47 и/или белка hsp47 или РНК можно использовать для предсказания клинического исхода и для определения эффективности соединений и композиций, которые модулируют уровень и/или активность определенных hsp47 и/или белков hsp47, ассоциированных с признаком, состоянием или заболеванием.
[0329] Обеспечиваются способы и композиции для подавления экспрессии hsp47 посредством использования малых молекул нуклеиновых кислот согласно настоящему описанию, таких как молекулы коротких интерферирующих нуклеиновых кислот (siНК), интерферирующих РНК (РНКи), коротких интерферирующих РНК (siРНК), двухцепочечных РНК (dsPHK), микро-РНК (miPHK) и коротких шпильковых РНК (shPHK), способных опосредовать или которые опосредуют РНК интерференцию, против экспрессии гена hsp47. Композиция и способы согласно настоящему описанию также являются полезными при лечении различных форм фиброза, таких как фиброз печени, фиброз легких и фиброз почек.
[0330] Молекулы нуклеиновых кислот согласно настоящему описанию по-отдельности или в комбинации или в соединении с другими лекарственными средствами можно использовать для предотвращения или лечения заболеваний, признаков, состояний и/или нарушений, ассоциированных с hsp47, таких как фиброз печени, цирроз, фиброз легких, фиброз почек, перитонеальный фиброз, хроническое поражение печени и фибриллогенез.
[0331] Молекулы нуклеиновой кислоты согласно настоящему описанию способны подавлять экспрессию hsp47 специфичным в отношении последовательности образом. Молекулы нуклеиновых кислот могут включать смысловую цепь и антисмысловую цепь, которые включают смежные нуклеотиды, которые являются по меньшей мере частично комплементарными (антисмысловыми) мРНК hsp47.
[0332] Согласно некоторым вариантам реализации dsPHK, специфичную в отношении hsp47, можно использовать в сочетании с другими dsPHK, специфичными для других молекулярных шаперонов, которые принимают участие в сворачивании вновь синтезированных белков, таких как кальнексин, кальретикулин, BiP (Bergeron et al. Trends Biochem. Sci. 1994; 19:124-128; Herbert et al. 1995; Cold Spring Harb. Symp. Quant. Biol. 60:405-415)
[0333] Фиброз можно вылечить с помощью РНК интерференции с использованием молекул нуклеиновых кислот согласно настоящему описанию. Примеры фиброза включают фиброз печени, перитонеальный фиброз, фиброз легких, фиброз почек. Молекулы нуклеиновых кислот согласно настоящему описанию могут подавлять экспрессию hsp47 специфичным в отношении последовательности образом.
[0334] Можно проводить мониторинг лечения фиброза по определению уровня внеклеточного коллагена с использованием приемлемых методик, известных в данной области, например, с использованием антител к коллагену I. Также можно проводить мониторинг лечения фиброза по определению уровня мРНК hsp47 или уровня белка HSP47 в клетках пораженной ткани. Также можно проводить мониторинг лечения фиброза с помощью неинвазивного сканирования пораженного органа или ткани, например, компьютерной томографии, магнитно-резонансной эластографии.
[0335] Способ лечения или предотвращения у пациента или организма заболевания или состояния, ассоциированного с hsp47, может включать обеспечение контакта пациента или организма с молекулой нуклеиновой кислоты согласно настоящему описанию при условиях, подходящих для модуляции экспрессии гена hsp47 у пациента или организма.
[0336] Способ лечения или предотвращения фиброза у пациента или организма может включать обеспечение контакта пациента или организма с молекулой нуклеиновой кислоты при условиях, подходящих для модуляции экспрессии гена hsp47 у пациента или организма.
[0337] Способ лечения или предотвращения одного или нескольких типов фиброза, выбранных из группы, состоящей из фиброза печени, фиброза почек и фиброза легких у пациента или организма, может включать обеспечение контакта пациента или организма с молекулой нуклеиновой кислоты при условиях, подходящих для модуляции экспрессии гена hsp47 у пациента или организма.
Фибротические заболевания
[0338] Фибротические заболевания обычно характеризуются избыточным отложением волокнистого материала во внеклеточном матриксе, которое способствуют анормальным изменениям архитектуры тканей и нарушает нормальную работу органов.
[0339] Все ткани, нарушенные при травме, отвечают посредством запуска программы заживления раны. Фиброз, тип заболевания, характеризующийся избыточным рубцеванием, возникает, если нарушается нормальный самоорганичивающийся процесс ответа заживления раны, и вызывает избыточную выработку и отложение коллагена. В результате нормальная ткань органа замещается рубцовой тканью, которая в итоге приводит к функциональному отказу органа.
[0340] Фиброз может быть вызван различными причинами и в разных органах. Цирроз печени, фиброз легких, саркоидоз, келоидные рубцы и фиброз почек - все являются хроническими заболеваниями, ассоциированными с прогрессивным фиброзом, благодаря чему происходит постоянная утрата нормальной функции ткани.
[0341] Острый фиброз (обычно с внезапным и серьезным началом и быстрым течением) возникает как общий ответ на различные формы травм, включая травмы в результате несчастного случая (в частности травмы спинного мозга и центральной нервной системы), инфекций, хирургического вмешательства, ишемических заболеваний (например, образование рубцов в сердце после сердечного приступа), ожогов, загрязнения окружающей среды, алкоголя и других типов токсинов, синдрома острой дыхательной недостаточности, лучевой терапии и химиотерапии.
[0342] Фиброз, патология, вызванная фиброзом, или патология, вызванная неправильным образованием поперечных связей клеточных белков, все могут быть вылечены с помощью siРНК согласно настоящему описанию. Фибротические заболевания или заболевания, при которых очевиден фиброз (патология, вызванная фиброзом) включают как острые, так и хронические формы фиброза органов, включая все следующие этиологические варианты: фиброз легких, включая интерстициальную болезнь легких и фибротическую болезнь легких, фиброз печени, фиброз сердца, включая фиброз миокарда, фиброз почек, включая хроническую почечную недостаточность, фиброз кожи, включая склеродермию, келоидные и гипертрофические рубцы; миелофиброз (фиброз костного мозга); все типы рубцев глаз, включая пролиферативную витреоретинопатию (ПВР) и рубцы в результате хирургического вмешательства для лечения катаракты или глаукомы; воспалительная болезнь кишечника различной этиологии, макулярная дегенерация, офтальмопатия Грейвса, эрготизм, вызванный лекарственными средствами, келоидные рубцы, склеродермия, псориаз, глиобластома при синдроме Ли-Фраумени, спорадическая глиобластома, миелоидная лейкемия, острая миелогенная лейкемия, миелодиспластический синдром, миелопродиферативный синдром, гинекологическая онкология, саркома Капоши, болезнь Гансена и коллагенозный колит.
[0343] Согласно различным вариантам реализации соединения (молекулы нуклеиновых кислот) согласно настоящему описанию можно использовать для лечения фибротических заболеваний, например, согласно настоящему описанию, а также множества других заболеваний и состояний, кроме фибротических заболеваний, например, таких как согласно настоящему описанию. Другие заболевания, которые необходимо лечить, включают фибротические заболевания в других органах - фиброз почек любой причины (ХБП, включая ТХПН); фиброз легких (включая ИЛФ); миелофиброз, ненормальное рубцевание (келоидные рубцы), вызванные всевозможными типами травм кожи - случайными и ятрогенными (операции); склеродермию, кардиофиброз, неудачная фильтрующая операция при глаукоме; кишечные спайки.
Офтальмологическая хирургия и фибротические осложнения
[0344] Часто может происходить контрактура рубцовой ткани в результате операции на глазах. Хирургическое вмешательство при глаукоме для создания новых дренажных каналов часто заканчивается неудачно из-за образования рубцов и контрактуры тканей, и созданная дренажная система может быть блокирована, что требует дополнительного хирургического вмешательства. Существующие схемы лечения против рубцевания (митомицин С или 5FU) ограничены сопуствующими осложнениями (например, слепотой), например, смотрите публикацию Cordeiro MF, et al., Human anti-transforming growth factor-beta2 antibody: a new glaucoma anti-scarring agent Invest Ophthalmol Vis Sci. 1999 Sep;40(10):2225-34. Также могут происходить контрактура рубцовой ткани, образованной после травмы роговицы или операции на роговице, например, лазерного или хирургического лечения миопии или рефракционной аномалии, при которой контрактура тканей может привести к неверным результатам. Рубцовая ткань может быть образована, например, на/в стекловидном теле или сетчатке и в итоге может вызвать слепоту у некоторых пациентов с диабетом, и может быть образована после операции отслоения, имеющей название пролиферативная витреоретинапатия (ПВР). ПВР является обычным осложнением после отслойки сетчатки и связана с дырой или разрывом сетчатки. ПВР обозначает рост клеточных мембран в полости стекловидного тела и на передней и задней поверхности сетчатки, содержащей клетки пигментного эпителия сетчатки (RPE). Эти мембраны, которые по существу представляют собой рубцовые ткани, растягиваются на сетчатке и могут привести к повторению отслоения сетчатки, даже после первоначально успешной процедуры отслоения сетчатки.
[0345] Рубцовая ткань может образоваться в орбите или на глазу и мышцах век после хирургических операций при косоглазии, операциях на орбите или веках или тиреоидной болезни глаза, и при которых происходит образование рубцов на конъюктве, что может произойти после операции при глаукоме или при руьцовом заболевании, воспалительном заболевании, например, пемфигоиде или инфекционном заболевании, например, трахоме. Также глазным заболеванием, связанным с контрактурой тканей, содержащих коллаген, является помутнение и контрактура капсулы хрусталика после удаления катаракты. Была признана важная роль ММР в офтальмологических заболеваниях, включая заживление ран, сухой глаз, незаживающее изъязвление роговицы, возвратную эрозию эпителия, неоваскуляризацию роговицы, птеригий, халазию конъюктивы, глаукому, ПВР и окулярный фиброз.
Фиброз печени
[0346] Фиброз печени (ФП) является обычно необратимым последствием поражения печени нескольких этиологии. В Западном Мире главными этиологическими категориями являются: алкогольная болезнь печени (30-50%), вирусный гепатит (30%), болезнь желчных протоков (5-10%), первичный гемохроматоз (5%) и лекарственный и криптогенный цирроз неизвестной этиологии (10-15%). Также фиброз печени является одним из симптомов болезни Вильсона, недостаточности α1-антитрипсина и других редких заболеваний. При циррозе печени - последней стадии фиброза печени - часто необходима трансплантация печени, и цирроз печени входит в десять самых распространенных причин смертности в Западном Мире.
Фиброз почек и связанные состояния
Хроническая почечная недостаточность (ХПН)
[0347] Хроническая почечная недостаточность представляет собой постепенную и прогрессирующую потерю способности почек выводить отходы, концентрировать мочу и сохранять электролиты. ХПН является медленно прогрессирующей. Наиболее часто она возникает в результате любого заболевания, которое вызывает постепенную потерю функции почек, и фиброз является основной патологией, при которой развивается ХПН.
Диабетическая нефропатия
[0348] Диабетическая нефропатия, отличительным признаком которой является гломерулосклероз и тубулоинтерстициальный фиброз, является отдельной наиболее преобладающей причиной терминальной стадии почечной недостаточности в современном мире, и пациенты с диабетом составляют самую большую группу на диализе. Такой способ лечения является дорогостоящим и далек от оптимального. Трансплантация предлагает лучший исход, но страдает от серьезного недостатка доноров.
Хроническая болезнь почек
[0349] Хроническая болезнь почек (ХБП) является мировой проблемой общественного здоровья и считается рядовым состоянием, которое связано с повышенным риском развития сердечно-сосудистых заболеваний и хронической почечной недостаточностью (ХПН).
[0350] Инициатива Качества Лечения Заболевания Почек (K/DOQI) Национального Почечного Фонда (NKF) определяет хроническую болезнь почек как повреждение почек или сниженную скорость клубочковой фильтрации (СКФ) в течение трех или более месяцев. Другие маркеры ХБП также известны и используются для диагностики. В общем, разрушение тела почки с необратимым склерозом и утратой нефронов приводит к погрессивному ухудшению СКФ. Недавно K/DOQI опубликовала следующую классификацию стадий ХБП:
Стадия 1: повреждение почек с нормальной или повышенной СКФ (>90 мл/мин/1,73 м2)
Стадия 2: слабое снижение СКФ (60-89 мл/мин/1,73 м2)
Стадия 3: умеренное снижение СКФ (30-59 мл/мин/1,73 м2)
Стадия 4: серьезное снижение СКФ (15-29 мл/мин/1,73 м2)
Стадия 5: почечная недостаточность (СКФ <15 мл/мин/1,73 мл2 или диализ)
[0351] На 1 и 2 стадии ХБП СКФ в отдельности не подтверждает диагноз. Можно полагаться на другие маркеры повреждения почек, включая нарушения в составе крови или мочи или отклонения в рентрегологических исследованиях.
Патофизиология ХБП
[0352] Приблизительно 1 миллион нефронов находится в каждой почке, каждый из которых вносит вклад в общую СКФ. Независимо от этиологии повреждения почки с прогрессивным разрушением нефронов, почка может поддерживать СКФ с помощью гиперфильтрации и компенсаторной гипертрофии оставшихся здоровых нефронов. Эта приспособляемость нефронов способствует длительному нормальному клиренсу растворенных веществ плазмы, вследствие чего такие вещества, как мочевина и креатинин начинают показывать значительное повышение уровней в плазме только после того, как общая СКФ снизилась до 50%, когда почечный резерв исчерпан. Уровень креатинина в плазме приблизительно удвоится при снижении СКФ на 50%. Поэтому повышение уровня креатинина в плазме в два раза по сравнению с исходным значением 0,6 мг/дл до 1,2 мг/дл у пациента фактически представляет потерю 50% массы функционирующих нефронов.
[0353] Считается, что остаточная гиперфильтрация и гипертрофия нефронов, несмотря на ее полезность в силу отмеченых причин, представляет основную причину прогрессирующей дисфункции почек. Считается, что это происходит из-за повышенного капиллярного давления в клубочках, которое повреждает капилляры и сначала приводит к местному и сегментному гломерулосклерозу, а в итоге к глобальному гломерулосклерозу. Эта гипотеза основывается на исследованиях крыс с 5/6 нефроэктомией, у которых развились поражения, которые были идентичны поражениям, наблюдаемым у людей с ХБП.
[0354] Двумя наиболее распространенными причинами хронической болезни почек являются диабет и гипертензия. Другие факторы включают острые инсульты в результате нефротоксинов, включая контрастирующие агенты или сниженную перфузию; протеинурии; повышенного почечного аммониагенеза с интерстициальной травмой, гиперлипидемии, гиперфосфатемии с отложением фосфата кальция, пониженных уровней оксида азота и курения.
[0355] В Соединенных Штатах Америки частота и распространенность ХБП повышается с плохими исходами и высокой стоимостью для системы здравоохранения. Болезнь почек является девятой ведущей причиной смерти в США. Высокая степень смертности заставила включить в приказ Министра здравоохранения США для граждан Америки, Здоровые люди 2010, главу, посвященную ХБП. Целями этой главы является сформулировать задачи и обеспечить стратегии для снижения распространенности, заболеваемости, смертности и расходов здравоохранения на хроническую болезнь почек в США.
[0356] Встречаемость терминальной стадии хронической почечной недостаточности (ТХПН) в мире также постоянно повышается с 1989. Соединенные Штаты Америки имеют самую высокую степень встречаемости ТХПН, после них следует Япония. Япония имеет самую высокую распространенность на миллион популяции, после нее следуют США.
[0357] Смертность, вызванная гемодиализом, является поразительной и указывает, что ожидаемая продолжительности жизни пациентов, которым начинают проводить гемодиализ, заметно сокращается. В любом возрасте пациенты с ТХПН на диализе имеют значительно повышенную смертность по сравнению с пациентами без диализа и людьми без болезни почек. В возрасте 60 лет здоровый человек может предполагать прожить еще 20 лет, тогда как ожидаемая продолжительность жизни 60-летнего пациента, начинающего гемодиализ, приближается к 4 годам (Aurora and Verelli, May 21, 2009. Chronic Renal Failure: Treatment & Medication. Emedicine. http://emedicine.medscape.com/article/238798-treatment).
Фиброз легких
[0358] Интерстициальный фиброз легких (ИФЛ) представляет собой образование рубцов в почках, вызванное различными вдыхаемыми агентами, включая минеральные частицы, органическую пыль и окислительные газы или по неизвестным причинам (идиопатический фиброз легких). Заболевание поражает миллионы человек в мире, и эффективные способы лечения отсутствуют. Главной причиной отсутствия полезного лечения является то, что мало молекулярных механизмов исследовали в достаточной степени для разработки соответствующих мишеней терапии (Lasky JA., Brody AR. (2000), "Interstitial fibrosis and growth factors". Environ Health Perspect.; 108 Suppl 4:751-62).
Фиброз сердца
[0359] Сердечная недостаточность занимает уникальное место среди главных сердечно-сосудистых заболеваний, поскольку только у него распространенность повышается, в то время как происходит заметное снижение других заболеваний. Частично это может объясняться старением населения Соединенных Штатов Америки и Европы. Возможность спасать пациентов с повреждением миокарда также является главным фактором, поскольку у этих пациентов может развиваться прогрессия дисфункции левого желудочка из-за разрушительного ремоделирования сердца.
[0360] Нормальный миокард состоит из разных клеток, кардиомиоцитов и не кардиомиоцитов, которые включают эндотелиальные и гладкомышечные клетки сосудов и фибробласты.
[0361] Ремоделирование структуры стенки желудочка является ключевой детерминантой клинического исхода болезни сердца. Такое ремоделирование включает выработку и деградацию белков внеклеточного матрикса, пролиферацию и миграцию клеток и апоптотическую или некротическую гибель клеток. Фибробласты сердца принципиально вовлечены в эти процессы, вырабатывая факторы роста и цитокины, которые действуют как аутокринные и паракринные факторы, а также белки и протеиназы внеклеточного матрикса. Недавние исследования показали, что взаимодействия между фибробластами сердца и кардиомиоцитами абсолютно необходимы для прогрессии ремоделирования сердца, совокупным эффектом которого является нарушение работы сердца и начало сердечной недостаточности (Manabe I, et al., (2002), "Gene expression in fibroblasts and fibrosis: involvement in cardiac hypertrophy", Circ Res. 13;91(12):1103-13).
Ожоги и рубцы
[0362] Отдельной проблемой, которая может возникнуть, в частности, при фибротическом заболевании, является контрактура тканей, например, контрактура рубцов. Контрактура тканей, включая компоненты внеклеточного матрикса, в частности, тканей, содержащих коллаген, может происходить в связи со многими различными патологическими состояниями и хирургическими или косметическими процедурами. Контрактура, например, рубцов может вызывать физические проблемы, которые могут приводить к необходимости медицинского вмешательства, или она может вызывать проблемы чисто косметической природы. Коллаген является главным компонентом рубца и поэтому является наиболее важным структурным компонентом для рассмотрения. Тем не менее, рубцы и другие контрактированные ткани также включают другие структурные компоненты, в частности другие компоненты внеклеточного матрикса, например эластин, который также может способствовать контрактуре ткани.
[0363] Контрактура ткани, содержащей коллаген, которая также может включать другие компоненты внеклеточного матрикса, часто встречается при заживлении ожогов. Ожоги могут быть химическими, термическими или радиационными ожогами, и могут быть на глазах, поверхности кожи или на коже и лежащих ниже тканях. Также в некоторых случаях ожоги могут быть на внутренних тканях, например, вызванные действием облучения. Контрактура Ожеговых тканей часто является проблемой и может приводить к физическим и/или косметическим проблемам, например, потере подвижности и/или обезображиванию.
[0364] Кожные трансплантаты могут применять по различным причинам, и они могут претерпевать контрактуру после нанесения. При заживлении обожженных тканей контрактура может приводить к физическим и косметическим проблемам. Это является особенно серьезной проблемой, если необходимо много кожных трансплантатов, например, в случае серьезных ожогов.
[0365] Контрактура также является проблемой при создании искусственной кожи. Для получения искусственной кожи необходимо иметь эпидермис, состоящий из эпителиальных клеток (кератиноцитов), и дермис, состоящий из коллагена, заселенного фибробластами. Важно иметь оба типа клеток, т.к. они запускают сигналлинг и стимулируют друг друга с использованием факторов роста. Коллагеновый компонент искусственной кожи часто сжимается менее чем до десятой части от его исходной площади при заселении фибробластами.
[0366] Контрактура рубцов, контрактура из-за сжимания фиброзной ткани рубца, является обычной. В некоторых случаях рубец может превратиться в деформирующий рубец, рубец, в котором контракция вызывает серьезное уродство. Желудок пациента может быть фактически разделен на две отдельные камеры контрактурой песочных часов при сокращении рубцовой ткани, образованной при заживлении язвы желудка. Закупорка проходов и протоков, рубцовый стеноз могут возникнуть при сокращении рубцовой ткани. Контрактура кровеносных сосудов может быть связана с первичной обструкцией или хирургической травмой, например, после операции или ангиопластики. Может также произойти стеноз других полых органов, например, мочеточников. Проблемы могут возникнуть, если происходит любая форма образования рубцов, как в результате случайного ранения, так и после операции. Заболевания кожи и сухожилий, которые включают сокращение тканей, включающих коллаген, включают посттравматические состояния, вызванные операцией или несчастным случаем, например, травмы сухожилий рук или ног, состояния после трансплантации и патологические состояния, такие как склеродермия, контрактура Дюпюитрена и буллезный эпидермолиз. Рубцевание и контрактура тканей в глазах может встречаться при различных состояниях, например, последствиях отслойки сетчатки или диабетического поражения глаз (как упоминалось выше). Контрактура глазницы, обнаруженная в черепе глазных яблок, и связанных с ней структур, в том числе внеглазных мышц и век, может возникать, если существует травма или воспалительное повреждение. Ткани сокращаются в глазнице, вызывая целый ряд проблем, включая удвоение в глазах и неприглядный внешний вид.
[0367] Для получения большей информации о различных типах фиброза смотрите публикации: Molina V, et al., (2002), "Fibrotic diseases", Harefuah, 141(11):973-8, 1009; Yu L, et al., (2002), "Therapeutic strategies to halt renal fibrosis", Curr Opin Pharmacol. 2(2):177-81; Keane WF and Lyle PA. (2003), "Recent advances in management of type 2 diabetes and nephropathy: lessons from the RENAAL study". Am J Kidney Dis. 41(3 Suppl 2): S22-5; Bohle A, et al., (1989), "The pathogenesis of chronic renal failure", Pathol Res Pract. 185(4):421-40; Kikkawa R, et al., (1997), "Mechanism of the progression of diabetic nephropathy to renal failure". Kidney Int Suppl. 62:S39-40; Bataller R, and Brenner DA. (2001), "Hepatic stellate cells as a target for the treatment of liver fibrosis", Semin Liver Dis. 21(3):437-51; Gross TJ and Hunninghake GW, (2001) "Idiopathic pulmonary fibrosis", N Engl J Med. 345(7):517-25; Frohlich ED. (2001) "Fibrosis and ischemia: the real risks in hypertensive heart disease". Am J Hypertens;14(6 Pt 2):194S-199S; Friedman SL. (2003), "Liver fibrosis - from bench to bedside", J Hepatol. 38 Suppl 1:S38-53; Albanis E, et al., (2003), "Treatment of hepatic fibrosis: almost there", Curr Gastroenterol Rep. 5(1):48-56; (Weber KT. (2000), "Fibrosis and hypertensive heart disease", Curr Opin Cardiol. 15(4):264-72).
Доставка молекул нуклеиновых кислот и лекарственные формы
[0368] Молекулы нуклеиновых кислот могут быть адаптированы для использования для предотвращения или лечения фиброзных заболеваний (например, фиброза печени, почек, перитонеального или легочного), признаков, состояний и/нарушений, которые связаны или будут отвечать на уровни hsp47 в клетке или ткани по-отдельности или в комбинации с другими способами лечения. Молекула нуклеиновой кислоты может включать носитель для доставки, включая липосомы, для введения пациенту, носители и разбавитель и их соли, и/или могут быть представлены в фармацевтически приемлемых лекарственных формах.
[0369] Молекулы нуклеиновых кислот согласно настоящему описанию могут быть доставлены или вводиться непосредственно с носителем или разбавителем, но не с любым носителем для доставки, который помогает, способствует или облегчает вход в клетку, включаявирусные векторы, вирусные частицы, лекарственные формы липосом, липофектин или осаждающие агенты и т.п.
[0370] Молекулы нуклеиновых кислот можно доставлять или вводить пациенту посредством прямого нанесения молекул нуклеиновых кислот с носителем или разбавителем или любых другим носителем для доставки, который может действовать для облегчения, способствования или облегчения входа в клетку, включая вирусные последовательности, вирусные частицы, лекарственные формы липосом, липофектин или осаждающие агенты и т.п. Полипептиды, которые облегчают введение нуклеиновой кислоты желаемому пациенту, такие как описанные в публикации заявки США №20070155658 (например, производное меламина, такое как 2,4,6-тригуанидино триазин и 2,4,6-триамидосаркозил меламин, полиаргининовый полипептид и полипептид, включающий чередующиеся остатки глутамина и аспарагина).
[0371] Способы доставки молекул нуклеиновых кислот описаны в публикациях Akhtar et al., Trends Cell Bio., 2: 139 (1992); Delivery Strategies for Antisense Oligonucleotide Therapeutics, ed. Akhtar, (1995), Maurer et al., Mol. Membr. Biol., 16: 129-140 (1999); Hofland and Huang, Handb. Exp. Pharmacol., 137: 165-192 (1999); и Lee et al., ACS Symp. Ser., 752: 184-192 (2000); патентах США №6,395,713; 6,235,310; 5,225,182; 5,169,383; 5,167,616; 4,959217; 4.925,678; 4,487,603 и 4,486,194 и публикации Sullivan et al., PCT WO 94/02595; PCT WO 00/03683 и PCT WO 02/08754; и публикации заявки на патент США №2003077829. Указанные протоколы можно использовать для доставки буквально любой молекулы нуклеиновой кислоты. Молекулы нуклеиновых кислот можно вводить в клетки с помощью различных способов, известных специалистам в данной области техники, включая, но не ограничиваясь ими, инкапсуляцию в липосомы, метод ионтофореза, или с помощью встраивания в другие носители, такие как биодеградируемые полимеры, гидрогели, циклодекстрины (смотрите, например, Gonzalez et al., Bioconjugate Chem., 10: 1068-1074 (1999); Wang et al., международная публикация PCT № WO 03/47518 и WO 03/46185), поли(молочная-ко-гликолевая) кислота (PLGA) и микросферы PLCA (смотрите, например, патент США №6,447,796 и публикацию заявки на патент США №2002130430), биодеградируемые нанокапсулы и биоадгезивные микросферы или белковые векторы (О′Наге и Normand, международная публикация PCT №WO 00/53722). В другом случае комбинацию нуклеиновой кислоты/носителя доставляют местно с помощью прямой инъекции или с использованием инфузионной помпы. Прямая инъекция молекул нуклеиновых кислот согласно настоящему изобретению, подкожные, внутримышечные, чрезкожные можно осуществлять с использованием стандартных методов с использованием иглы и шприца или с использованием безигольных технологий, например, описанных в публикации Conry et al., Clin. Cancer Res., 5:2330-2337 (1999) и Barry et al., международная публикация РСТ № WO 99/31262. Молекулы настоящего изобретения можно использовать в качестве фармацевтических агентов. Фармацевтические агенты предотвращают, модулируют встречаемость или лечат (облегчают симптом до некоторой степени, предпочтительно все симптомы) заболевания у пациента.
[0372] Молекулы нуклеиновых кислот можно включать в комплексы с катионными липидами, можно упаковывать в липосомы или иначе доставлять в клетки или ткани-мишени. Нуклеиновую кислоту или комплексы нуклеиновой кислоты можно вводить местно в соответствующие ткани ex vivo, или in vivo посредством прямого дермального введения, трансдермального введения или инъекции, или без встраивания их в биополимеры. Молекулы нуклеиновых кислот согласно настоящему изобретению могут включать последовательности, показанные в Таблицах I. Примеры таких молекул нуклеиновых кислот по существу состоят из последовательностей, представленных в Таблице I.
[0373] Системы доставки включают липосомы с измененной поверхностью, содержащие поли(этиленгликоль) липиды (PEG-модифицированные или долго циркулирующие липосомы или малозаметные липосомы). Указанные лекарственные формы предлагают способ повышения накопления лекарственных средств в тканях-мишенях. Данный класс носителей лекарственных средств устойчив к опсонизации и уничтожению системой фагоцитирующих мононуклеаров (MPS или RES), способствуя, таким образом, более длительному сроку циркуляции в крови и усиленному воздействию на ткани инкапсулированного лекарственного средства (Lasic et al. Chem. Rev. 1995, 95, 2601-2627; Ishiwata et al., Chem. Pharm. Bull. 1995, 43, 1005-1011).
[0374] Молекулы нуклеиновых кислот можно включать в состав или в комплекс с полиэтиленимином (например, линейным или разветвленным PEI) и/или производными полиэтиленимина, включая, например, полиэтиленимин-полиэтиленгликоль-N-ацетилгалактозамин (PEI-PEG-GAL) или производные полиэтиленимин-полиэтиленгликоль-три-N-ацетилгалактозамина (PEI-PEG-triGAL), привитые PEI, такие как галактоза PEI, холестерол PEI, производное PEI с антителами и их производные с полиэтилен гликолем PEI (PEG-PEI) (смотрите, например, публикации Ogris et al., 2001, AAPA PharmSci, 3, 1-11; Furgeson et al., 2003, Bioconjugate Chem., 14, 840-847; Kunath et al., 2002, Pharmaceutical Research, 19, 810-817; Choi et al., 2001, Bull. Korean Chem. Soc., 22, 46-52; Bettinger et al., 1999, Bioconjugate Chem., 10, 558-561; Peterson et al., 2002, Bioconjugate Chem., 13, 845-854; Erbacher et al., 1999, Journal of Gene Medicine Preprint, 1, 1-18; Godbey et al., 1999., PNAS USA, 96, 5177-5181; Godbey et al., 1999, Journal of Controlled Release, 60, 149-160; Diebold et al., 1999, Journal of Biological Chemistry, 274, 19087-19094; Thomas and Klibanov, 2002, PNAS USA, 99, 14640-14645; Sagara, патент США №6,586,524 и публикацию заявки на патент США №20030077829).
[0375] Молекулы нуклеиновых кислот можно включать в состав комлексов с агентами, разрушающими мембраны, например, описанными в публикации заявки на патент США №20010007666. Агент или агенты, разрушающие мембраны, и молекулу нуклеиновой кислоты также можно включать в состав комплексов с катионным липидом или вспомогательной липидной молекулой, такими как липиды, например, согласно описанию в патенте США №6,235,310.
[0376] Молекулы нуклеиновых кислот можно вводить с помощью введения в легкие, например, посредством вдыхания высушенной лекарственной формы в виде аэрозоля или спрея, вводимой с помощью устройства для ингаляции или распылителя, обеспечивающего быстрое местное поглощение молекул нуклеиновых кислот соответствующими тканями легких. Твердые дисперсные композиции, содержащие подходящие для вдыхания сухие частицы тонко измельченных композиций нуклеиновых кислот, можно изготовить посредством измельчения высушенных или лиофилизированных композиций нуклеиновых кислот и затем пропускания измельченных композиций, например, через сито с размером ячеек 400, чтобы разбить или отделить крупные аггломераты. Твердая дисперсная композиция, содержащая композиции нуклеиновых кислот согласно настоящему изобретению, может дополнительно содержать диспергатор, который служит для облегчения образования аэрозоля, а также другие терапевтические соединения. Приемлемым диспергатором является лактоза, которую можно смешивать с соединением нуклеиновой кислоты в любом приемлемом соотношении, например, соотношении 1 к 1 по массе.
[0377] Аэрозоли жидких частиц могут включать молекулы нуклеиновых кислот согласно настоящему описанию и могут быть получены любыми приемлемыми способами, например, с помощью распылителя (смотрите, например, патент США №4,501,729). Распылители представляют собой коммерчески доступные устройства, которые превращают растворы или суспензии активного ингредиента в терапевтический аэрозоль, например, с помощью ускорения сжатого газа, обычно воздуха или кислорода, через узкие отверстия трубки Вентури или с помощью ультразвукового перемешивания. Приемлемые лекарственные формы для использования в распылителях включают активный ингредиент в жидком носителе в количестве до 40% w/w предпочтительно меньше, чем 20% w/w лекарственной формы. Обычно носитель представляет собой воду или разбавленный водно-спиртовой раствор, предпочтительно сделанный изотоничным жидкостям тела посредством добавления, например, хлорида натрия или других приемлемых солей. Возможные добавки включают консерванты, если лекарственная форма изготавливается нестерильной, например, метил гидроксибензоат, антиоксиданты, ароматизаторы, эфиные масла, буферные вещества и эмульгаторы и другие поверхностно активные вещества. Аэрозоли твердых частиц, включая активную композицию и поверхностно активное вещество, также можно изготовить с помощью любого твердодисперсного генератора аэрозолей. Генераторы аэрозолей для введения твердодисперсного лекарственного средства пациенту производят частицы, которые можно вдыхать, как описано выше, и генерируют объем аэрозоля, содержащий предварительно измеренную дозу терапевтической композиции со скоростью, подходящей для введения человеку. Одни примером типа твердодисперсного генератора аэрозолей является инжектор. Приемлемые лекарственные формы для введения посредством вдувания включают тонко измельченные порошки, которые можно вводить с помощью инжектора. В инжекторе порошок, например, его измеренная доза, эффективная для осуществления лечения согласно настоящему описанию, заключена в капсулы или картриджи, обычно сделанные из желатина или пластика, которые прокалывают или открывают in situ, и порошок, доставляемый воздухом, выпускаемым из устройства при вдыхании или с помощью ручной помпы. Порошок, используемый в инжекторе, состоит только из активного ингредиента или из смеси порошков, содержащей активный ингредиент, приемлемый разбавитель порошка, такой как лактоза, и, возможно, поверхностно активное вещество. Активный ингредиент обычно составляет от 0,1 до 100% w/w лекарственной формы. Второй тип примеров генераторов аэрозолей включает дозирующий ингалятор. Дозирующие ингаляторы представляют собой аэрозольные распылители под давлением, обычно содержащие суспензию или раствор активного ингредиента в сжиженном пропелленте. В процессе применения указанные устройства выбрасывают лекарственную форму через клапан для доставки измеренного объема для производства тонкодисперсного спрея, содержащего активный ингридиент. Приемлемые пропелленты включают некоторые соединения хлорфторуглеродов, например, дихлордифторметан, трихлорфторметан, дихлортетрафторэтан и их смеси. Дополнительно лекарственная форма может содержать один или несколько сорастворителей, например, этанол, эмульгаторов и других поверхностно активных веществ, таких как масляная кислота или сорбитан триолеат, антиоксидантов и приемлемых ароматизаторов. Другие способы для доставки в легкие описываются, например, в заявке на патент США №20040037780 и патентах США №№6,592,904; 6,582,728; 6,565,885. РСТ публикация патента № WO2008/132723 относится к аэрозольной доставке олигонуклеотидов в общем и siРНК в частности в дыхательную систему.
[0378] Молекулы нуклеиновых кислот можно вводить в центральную нервную систему (ЦНС) или периферическую нервную систему (ПНС). Эксперименты показали эффективное поглощение in vivo нуклеиновых кислот нейронами. Смотрите, например, публикации Sommer et al., 1998, Antisense Nuc. Acid Drug Dev., 8, 75; Epa et al., 2000, Antisense Nuc. Acid Drug Dev., 10, 469; Broaddus et al., 1998, J. Neurosurg., 88(4), 734; Karle et al., 1997, Eur. J. Pharmocol., 340(2/3), 153; Bannai et al., 1998, Brain Research, 784(1,2), 304; Rajakumar et al., 1997, Synapse, 26(3), 199; Wu-pong et al., 1999, BioPharm, 12(1), 32; Bannai et al., 1998, Brain Res. Protoc., 3(1), 83; и Simantov et al., 1996, Neuroscience, 74(1), 39. Молекулы нуклеиновых кислот, таким образом, подходят для доставки и поглощения клетками в ЦНС и/или ПНС.
[0379] Доставка молекул нуклеиновых кислот в ЦНС обеспечивается большим количеством различных стратегий. Традиционные подходы к доставке в ЦНС, которые могут быть использованы, включают, но не ограничиваются, интратекальное и интрацеребровентрикулярное введение, имплантацию катетеров и помп, непосредственную инъекцию или перфузию в место поражения или травмы, инъекцию в систему мозговых артерий или введение посредством химического или осмотического открытия гематоэнцефалического барьера. Другие подходы могут включать использование различных транспортных систем и систем носителей, например, посредством использования конъюгатов и биодеградируемых полимеров. Более того, методы генной терапии, например, согласно описанию Kaplitt et al., патент США №6,180,613 и Davidson, WO 04/013280, могут быть использованы для экспрессии молекул нуклеиновых кислот в ЦНС.
[0380] Системы доставки могут включать, например, водные и безводные гели, кремы, различные эмульсии, микроэмульсии, липосомы, мази, водные и безводные растворы, лосьоны, аэрозоли, углеводородные основы и порошки, и могут включать наполнители, такие как растворители, усилители проникновения (например, жирные кислоты, эфиры жирных кислот, жирные спирты и аминокислоты), и гидрофильные полимеры (например, поликарбофил и поливинилпирролидон). Согласно одному варианту реализации фармацевтически приемлемый носитель представляет собой липосому или усилитель чрезкожного проникновения. Примеры липосом, которые могут быть использованы в настоящем изобретении, включают следующие: (1) CellFectin, 1:1.5 (М/М) липосомная форма катионного липида N,NI,NII,NIII-тетраметил-N,NI,NII,NIII-тетрапальмит-y-спермина и диолеилфосфатидилэтаноламина (DOPE) (GIBCO BRL); (2) Cytofectin GSV, 2:1 (М/М) липосомная форма катионного липида и DOPE (Glen Research); (3) DOTAP (N-[1-(2,3-диолеилокси)-N,N,N-три-метил-аммонийметилсульфат) (Boehringer Manheim); и (4) Lipofectamine, 3:1 (М/М) липосомная форма поликатионного липида DOSPA, нейтрального липида DOPE (GIBCO BRL) и ди-алкилированной аминокислоты (D1LA2).
[0381] Системы доставки могут включать пластыри, таблетки, суппозитории, свечи, гели и кремы, а также могут содержать наполнители, такие как растворители и усилители (например, пропиленгликоль, соли желчных кислот и аминокислоты), и другие носители (например, полиэтиленгликоль, эфиры жирных кислот и их производные, а также гидрофильные полимеры, такие как гидроксипропилметилцелюлоза и гиалуроновая кислота).
[0382] Молекулы нуклеиновых кислот можно включать в композиции или комплексы с полиэтиленимином (например, линейным или разветвленным PEI) и/или производными полиэтиленимина, включая, например, пересаженные PEI, такие как производные галактоза PEI, холестерол PEI, производное PEI с антителом и полиэтиленгликоль PEI (PEG-PEI) (смотрите, например, Ogris et al., 2001, AAPA PharmSci, 3, 1-11; Furgeson et al., 2003, Bioconjugate Chem., 14, 840-847; Kunath et al., 2002, Pharmaceutical Research, 19, 810-817; Choi et al., 2001, Bull. Korean Chem. Soc., 22, 46-52; Bettinger et al., 1999, Bioconjugate Chem., 10, 558-561; Peterson et al., 2002, Bioconjugate Chem., 13, 845-854; Erbacher et al., 1999, Journal of Gene Medicine Preprint, 1, 1-18; Godbey et al., 1999., PNAS USA, 96, 5177-5181; Godbey et al., 1999, Journal of Controlled Release, 60, 149-160; Diebold et al., 1999, Journal of Biological Chemistry, 274, 19087-19094; Thomas and Klibanov, 2002, PNAS USA, 99,14640-14645; и Sagara, патент США №6,586,524.
[0383] Молекулы нуклеиновых кислот могут включать биоконъюгат, например, конъюгат нуклеиновой кислоты согласно описанию Vargeese et al., номер п/п США 10/427,160; патент США №6,528,631; патент США №6,335,434; патент США №6,235,886; патент США№6,153,737; патент США №5,214,136; патент США №5,138,045.
[0384] Композиции, способы и наборы согласно настоящему описанию могут включать вектор экспрессии, который включает последовательность нуклеиновой кислоты, кодирующую по меньшей мере одну молекулу нуклеиновой кислоты согласно настоящему изобретению, таким образом, что это способствует экспрессии молекулы нуклеиновой кислоты. Способы введения молекул нуклеиновых кислот или одного или нескольких векторов, способных экспрессировать цепи dsPHK в среде клетки, будут зависеть от типа клетки и состав ее среды. Молекулу нуклеиновой кислоты или векторную конструкцию можно вводить непосредственно в клетку (т.е. внутриклеточно) или вводить внеклеточно в полость, интерстициальное пространство, в кровообращение организма, вводить перорально или можно вводить посредством погружения организма или клетки в раствор, содержащий dsPHK. Клетка предпочтительно представляет собой клетку млекопитающего, более предпочтительно клетку человека. Молекула нуклеиновой кислоты вектора экспрессии может включать смысловую область и антисмысловую область. Антисмысловая область может включать последовательность, комплементарную последовательности РНК или ДНК, кодирующей hsp47, и смысловая область может включать последовательность, комплементарную антисмысловой области. Молекула нуклеиновой кислоты может включать две отдельные цепи, имеющие комплементарные смысловую и антисмысловую области. Молекула нуклеиновой кислоты может включать одну цепь, имеющую комплементарные смысловую и антисмысловую области.
[0385] Молекулы нуклеиновых кислот, которые взаимодействуют с молекулами РНК-мишенями и подавляют ген, кодирующий молекулы РНК-мишени (например, молекулы РНК-мишени, обозначаемые в настоящем описании номерами доступа Genbank), могут экспрессироваться с единиц транскрипции, вставленных в векторы ДНК или РНК. Рекомбинантные векторы могут представлять собой ДНК плазмиды и вирусные векторы. Молекула нуклеиновой кислоты, экспрессирующая вирусные векторы, может быть создана на основе, но не ограничиваясь ими, аденассоциированного вируса, ретровируса, аденовируса или альфавируса. Рекомбинантные векторы, способные к экспрессии молекул нуклеиновых кислот, могут быть доставлены согласно настоящему описанию и сохраняться в клетках-мишенях. В другом случае, можно использовать вирусные векторы, которые способствуют временной экспрессии молекул нуклеиновых кислот. Такие векторы при необходимости можно вводить повторно. После экспрессии молекулы нуклеиновых кислот связываются с и подавляют функцию или экспрессию гена посредством РНК интерференции (РНКи). Доставка молекулы нуклеиновой кислоты, экспрессирующей векторы, может быть системной, например, посредством внутривенного или внутримышечного введения, посредством введения в клетки-мишени эксплантированные из пациента, с последующим обратным введением пациенту, или другими способами, которые позволяют вводить в желательную клетку-мишень.
[0386] Векторы экспрессии могут включать последовательность нуклеиновой кислоты, кодирующую по меньшей мере одну молекулу нуклеиновой кислоты согласно настоящему описанию, таким образом, который способствует экспрессии молекулы нуклеиновой кислоты. Например, вектор может содержать последовательность(ти), кодирующие обе цепи молекулы нуклеиновой кислоты, которые включают дуплекс. Также вектор может содержать последовательность (последовательности), кодирующую одну молекулу нуклеиновой кислоты, которая является комплементарной самой себе и, таким образом, образует молекулу нуклеиновой кислоты. Не ограничивающие примеры таких векторов экспрессии описаны в публикациях Paul et al., 2002, Nature Biotechnology, 19, 505; Miyagishi and Taira, 2002, Nature Biotechnology, 19, 497; Lee et al., 2002, Nature Biotechnology, 19, 500; and Novina et al., 2002, Nature Medicine, предварительная публикация Интернете doi:10.1038/nm725. Векторы экспрессии также могут быть включены в клетку млекопитающего (например, человека).
[0387] Вектор экспрессии может включать последовательность нуклеиновой кислоты, кодирующую две или несколько молекул нуклеиновых кислот, которые могут быть одинаковыми или разными. Векторы экспрессии могут включать последовательность молекулы нуклеиновой кислоты, комплементарную молекуле нуклеиновой кислоты под номером доступа Genbank NM_001235, например, показанную в Таблице I.
[0388] Вектор экспрессии может кодировать одну или обе цепи дуплекса нуклеиновых кислот, или одну комплементарную себе самой цепь, которая гибридизуется сама с собой с образованием дуплекса нуклеиновой кислоты. Последовательности нуклеиновых кислот, кодирующие молекулы нуклеиновых кислот, могут быть функционально соединены таким образом, который способствует экспрессии молекулы нуклеиновой кислоты (смотрите, например, Paul et al., 2002, Nature Biotechnology, 19, 505; Miyagishi and Taira, 2002, Nature Biotechnology, 19,497; Lee et al., 2002, Nature Biotechnology, 19, 500; и Novina et al., 2002, Nature Medicine, предварительная публикация Интернете doi:10.1038/nm725).
[0389] Вектор экспрессии может включать одно или несколько из следующих: а) участок инициации транскрипции (например, участок инициации полимеразы I, II или III эукариот); b) участок терминации транскрипции (например, участок терминации полимеразы I, II или III эукариот); с) интрон и d) последовательность нуклеиновой кислоты, кодирующую по меньшей мере одну молекулу нуклеиновой кислоты, при этом указанная последовательность функционально связана с участком инициации и участком терминации таким образом, который способствует экспрессии и/или доставке молекулы нуклеиновой кислоты. Вектор может включать открытую рамку считывания (ORF) для белка, функционально соединенную с 5' стороной или 3'-стороной последовательности, кодирующей молекулу нуклеиновой кислоты; и/или интрон (промежуточные последовательности).
[0390] Транскрипция последовательностей молекулы нуклеиновой кислоты может запускаться с промотора эукариотической РНК полимеразы I (pol I), РНК полимеразы (pol II) или РНК полимеразы III (pol III). Транскрипты промоторов pol II или pol III экспрессируются в клетке на высоком уровне; уровни данного промотора pol II в данном типе клеток зависит от природы регуляторных последовательностей гена (энхансеры, сайленсеры и т.п.), расположенных поблизости. Также используют промоторы прокариотической РНК полимеразы, если фермент прокариотической РНК полимеразы экспрессирутся в соответствующих клетках (Elroy-Stein and Moss, 1990, Proc. Natl. Acad. Sci. USA, 87, 6743-7; Gao and Huang 1993, Nucleic Acids Res., 21, 2867-72; Lieber et al., 1993, Methods Enzymol., 217, 47-66; Zhou et al., 1990, Mol. Cell. Biol., 10, 4529-37). Несколько исследователей показали, что молекулы нуклеиновой кислоты, экспрессированные с таких промоторов, могут функционировать в клетках млекопитающих (например, Kashani-Sabet et al., 1992, Antisense Res. Dev., 2, 3-15; Ojwang et al., 1992, Proc. Natl. Acad. Sci. USA, 89, 10802-6; Chen et al., 1992, Nucleic Acids Res., 20, 4581-9; Yu et al., 1993, Proc. Natl. Acad. Sci. USA, 90, 6340-4; L'Huillier et al., 1992, EMBO J., 11, 4411-8; Lisziewicz et al., 1993, Proc. Natl. Acad. Sci. U.S.A. 90, 8000-4; Thompson et al., 1995, Nucleic Acids Res., 23, 2259; Sullenger & Cech, 1993, Science, 262, 1566). Более конкретно, единицы транскрипции, например, полученные от генов, кодирующих U6 малые ядерные (snPHK), транспортные РНК (tPHK) и аденовирусные VA РНК, являются полезными при получении высоких концентраций желаемых молекул РНК, таких как siНК, в клетках (Thompson et al., ранее; Couture and Stinchcomb, 1996, ранее; Noonberg et al., 1994, Nucleic Acid Res., 22, 2830; Noonberg et al., U.S. Pat. No. 5,624,803; Good et al., 1997, Gene Ther., 4, 45; Beigelman et al., международная публикация РСТ № WO 96/18736. Указанные выше транскрипты нуклеиновых кислот могут быть встроены в различные векторы для введения в клетки млекопитающих, включая, но не ограничиваясь ими, векторы плазмид ДНК, вирусные ДНК векторы (например, векторы на основе аденовируса или адено ассоциированного вируса) или вирусные РНК векторы (например, ретровирусные или альфавирусные векторы) (смотрите. Couture and Stinchcomb, 1996 ранее).
[0391] Молекула нуклеиновой кислоты может экспрессироваться в клетке с эукариотических промоторов (например, Izant and Weintraub, 1985, Science, 229, 345; McGarry and Lindquist, 1986, Proc. Natl. Acad. Sci., USA 83, 399; Scanlon et al., 1991, Proc. Natl. Acad. Sci. USA, 88, 10591-5; Kashani-Sabet et al., 1992, Antisense Res. Dev., 2, 3-15; Dropulic et al., 1992, J. Virol., 66, 1432-41; Weerasinghe et al., 1991, J. Virol., 65, 5531-4; Ojwang et al., 1992, Proc. Natl. Acad. Sci. USA, 89, 10802-6; Chen et al., 1992, Nucleic Acids Res., 20, 4581-9; Sarver et al., 1990 Science, 247, 1222-1225; Thompson et al., 1995, Nucleic Acids Res., 23,2259; Good et al., 1997, Gene Therapy, 4, 45. Специалистам в данной области техники будет понятно, что любая нуклеиновая кислота может экспрессироваться в эукариотической клетке с соответствующего ДНК/РНК вектора. Активность таких нуклеиновых кислоты может быть повышена при их высвобождении от первичного транскрипта с помощью ферментативной нуклеиновой кислоты (Draper et al., РСТ WO 93/23569, and Sullivan et al., РСТ WO 94/02595; Ohkawa et al., 1992, Nucleic Acids Symp. Ser., 27, 15-6; Taira et al., 1991, Nucleic Acids Res., 19, 5125-30; Ventura et al., 1993, Nucleic Acids Res., 21,3249-55; Chowrira et al., 1994, J. Biol. Chem., 269, 25856.
[0392] Вирусная конструкция, упакованная в вирусную частицу, будет осуществлять как эффективное внедрение конструкции экспрессии в клетку, так и транскрипцию конструкции dsPHK, кодируемой конструкцией экспрессии.
[0393] Способы перорального введения включают непосредственное смешивание РНК с пищей организма, а также методы генной инженерии, при которых виды, которые используют в качестве пищи, модифицированы методами инженерии для экспрессии РНК, затем скармиливают организму, на который необходимо подействовать. Физические способы можно использовать для введения раствора молекулы нуклеиновой кислоты в клетку. Физические способы введения нуклеиновых кислот включают инъекцию раствора, содержащего молекулу нуклеиновой кислоты, бомбардировку частицами, покрытыми молекулой нуклеиновой кислоты, выдерживание клетки или организма в растворе РНК или электропорацию мембран клеток в присутствии молекулы нуклеиновой кислоты.
[0394] Другие способы, известные в данной области техники, для введения нуклеиновых кислот в клетки могут быть использованы, такие как опосредованный липидами перенос носителей, опосредованный химически транспорт, например, фосфатом кальция и т.п. Таким образом, молекулы нуклеиновых кислот можно вводить вместе с компонентами, которые выполняют одну или более из следующих действий: усиливают поглощение РНК клеткой, обеспечивают гибридизацию цепей дуплекса, стабилизируют гибридизованные цепи или иным образом усиливают подавление гена-мишени.
[0395] Молекулы нуклеиновых кислот или векторную конструкцию можно вводить в клетку с использованием приемлемых лекарственных форм. Одной предпочтительной лекарственной формой является форма с липидом, например, липосомы, соединенные с Lipofectamine™ 2000 (Invitrogen, CA, USA), витамином A (Sato et al. Nat Biotechnol 2008; 26:431-442, PCT публикация патента № WO 2006/068232). Также липидные лекарственные формы можно вводить животным, например, посредством внутривенной, внутримышечной или интраперитонеальной инъекции или перорально или посредством ингаляции или другими способами, известными в данной области техники. Если лекарственная форма подходит для введения животным, например, млекопитающим и более конкретно людям, лекарственная форма также является фармацевтически приемлемой. Фармацевтически приемлемые лекарственные формы для введения олигонуклеотидов известны и могут быть использованы. В некоторых случаях может быть предпочтительно готовить лекарственные формы dsPHK в буферном или физиологическом растворе и непосредственно вводить с помощью инъекции лекарственную форму dsPHK в клетки, например, как в исследованиях на ооцитах. Также можно проводить непосредственную инъекцию дуплексов dsPHK. Для приемлемых способов введения dsPHK, смотрите опубликованную заявку на получение патента США №2004/0203145, 20070265220, которая включена в настоящее описание посредством ссылки.
[0396] Полимерные нанокапсулы или микрокапсулы ускоряют транспорт и высвобождение инкапсулированной или связанной dsPHK в клетку. Они включают полимерные и мономерные материалы, особенно включая полибутилцианоакрилат. Обзор материалов и технологий изготовления были опубликованы (смотрите, Kreuter, 1991). Полимерные материалы, которые образованы из мономерных и/или олигомерных предшественников на этапе полимеризации/получения наночастиц, сами по себе известны из предыдущего уровня техники, также как молекулярная масса и распределение молекулярной массы полимерного материала, который специалист в области изготовления наночастиц, может выбрать в соответствии с обычными навыками.
[0397] Молекулы нуклеиновых кислот могут быть в виде лекарственной формы микроэмульсии. Микроэмульсия представляет собой систему воды, масла и амфифильного вещества, которое представляет собой отдельный раствор оптически изотропной и термодинамически стабильной жидкости. Обычно микроэмульсии изготавливают посредством сначала диспергирования масла в водном растворе поверхностно активного вещества и затем добавления достаточного количества четвертого компонента, обычно спирта с цепью промежуточной длины для образования прозрачной системы.
[0398] Поверхностно активные вещества, которые можно использовать при приготовлении микроэмульсии, включают, но не ограничиваются ими, ионные поверхностно активные вещества, неионные поверхностно активные вещества, Brij 96, эфиры полиоксиэтилена олеиновой кислоты, эфиры полиглицерина и жирных кислот, тетраглицерол монолаурат (ML310), тетраглицерол моноолеат (MO310), гексаглицерол моноолеат (РО310), гексаглицерол пентаолеат (PO500), декаглицерол монокарпат (МСА750), декаглицерол моноолеат (MO750), декаглицерол секвиолеат (SO750), декаглицерол декаолеат (DA0750), по отдельности или в комбинации с вторичными поверхностно активными веществами. Вторичное поверхностно активное вещество, обычно коротко цепочечный спирт, например, этанол, 1-пропанол и 1-бутанол, служит для повышения текучести на границе раздла фаз посредством проникания в пленку поверхностно активного вещества и создания неупорядоченной пленки из-за вакуума, созданного между молекулами поверхностно активного вещества.
[0399] Водорастворимые полимеры с поперечными сшивками
[0400] Лекарственные формы для доставки могут включать водорастворимые деградирующие полимеры с поперечными сшивками, которые включают одну или несколько разлагаемых поперечно связанных липидных групп, одну или несколько групп ΡΕΙ и/или одну или несколько mPEG (метальных эфирных производных PEG (метоксиполи(этиленгликоль)).
[0401] Разлагаемые липидные группы предпочтительно включают соединения, имеющие следующий структурный мотив:
[0402] В формуле выше сложноэфирные связи являются биоразлагаемыми группами, R представляет собой относительно гидрофобную «липо» группу, и показанный структурный мотив возникает в случае, если m находится в пределах от приблизительно 1 до приблизительно 30. Например, согласно некоторым вариантам реализации R выбирают из группы, состоящей из С2-С50 алкила, С2-С50 гетероалкил, С2-С50 алкенила, С2-С50 гетероалкенила, С5-С50 арила; С2-С50 гетероарила; С2-С50 алкинила, С2-С50 гетероалкинила, С2-С50 карбоксиалкенила и С2-С50 карбоксигетероалкенила. Согласно предпочтительным вариантам реализации, R представляет собой насыщенный или ненасыщенный алкил, имеющий 4-30 атомов углерода, более предпочтительно 8-24 атомов углерода или стерол, предпочтительно группу холестерола. Согласно предпочтительным вариантам реализации, R является олеиновой, лауриновой, миристиновой, пальмитиновой, маргариновой, стеариновой, арахидоновой, бегеновой или лигноцериновой, В наиболее предпочтительном варианте R является олеиновой.
[0403] N в формуле (А) может иметь пару электронов или связь с атомом водорода. Если N имеет пару электроном, повторяющаяся единица может быть катионной при низком pH.
[0404] Разлагаемая поперечно связанная липидная группа может вступать в реакцию с полиэтиленимином (PEI), как показано на Схеме А ниже:
[0405] На Схеме (A) R имеет те же значения, как описано выше. ΡΕΙ может содержать повторяющиеся единицы формулы (В), в которой x представляет собой целое число в пределах приблизительно от 1 до приблизительно 100, и у представляет собой целое число в пределах приблизительно от 1 до приблизительно 100.
[0406] Реакцию, изображенную на Схеме А, можно проводить посредством смешивания ΡΕΙ с диакрилатом (I) во взаимном растворителе, таком как этанол, метанол или дихлорметан, при перемешивании предпочтительно при комнатной температуре в течение нескольких часов, затем испарения растворителя для выделения полученного полимера. Не желая быть связанным какой-либо определенной теорией, считается, что реакция между ΡΕΙ и диакрилатом (I) включает реакцию Михаэля между одним или несколькими аминами ΡΕΙ с двойной связью (связями) диакрилата (смотрите J. March, Advanced Organic Chemistry 3rd Ed., pp.711-712 (1985)). Диакрилат, показанный на Схеме А, можно приготовить согласно описанию в заявке США №11/216,986 (публикация США №2006/0258751).
[0407] Молекулярная масса PEI предпочтительно находится в пределах от приблизительно 200 до 25,000 Дальтон, более предпочтительно 400-5,000 Дальтон, еще более предпочтительно 600-2000 Дальтон. PEI может быть как разветвленным, так и линейным.
[0408] Молярное соотношение PEI к диакрилату предпочтительно находится в пределах от приблизительно 1:2 до приблизительно 1:20. Средная молекулярная масса катионного липополимера может находиться в пределах от приблизительно 500 Дальтон до приблизительно 1,000,000 Дальтон, предпочтительно в пределах от приблизительно 2,000 Дальтон до приблизительно 200,000 Дальтон. Молекулярную массу можно определить с помощью гель-хроматографии с использованием стандартов PEG или методом электрофореза в агарозном геле.
[0409] Катионный липополимер предпочтительно является разлагаемым, более предпочтительно биоразлагаемым, например, разлагаемым по механизму, выбранному из группы, состоящей из гидрозила, ферментативного расщепления, восстановления, фото расщепления и разрушения ультразвуком. Не желая быть связанным определенной теорией, но считается, что после разрушения катионного липополимера формулы (II) в клетке происходит ферментативное расщепление и/или гидролиз сложноэфирных связей.
[0410] Синтез можно проводить посредством взаимодействия разлагаемой липидной группы с группой PEI согласно описанию выше. Затем добавляют mPEG (метилэфирное производное PEG (метоксиполи(этиленгликоль)) для образования деградирующего поперечно связанного полимера. Согласно предпочтительным вариантам реализации реакцию проводят при комнатной температуре. Продукты реакции можно выделять любыми способами, известными в данной области техники, включая методы хроматографии. Согласно предпочтительному варианту реализации продукт можно удалять с помощью преципитации с последующим центрифугированием.
Дозировки
[0411] Полезные дозы для введения и определенный способ введения будут различаться в зависимости от таких факторов, как тип клетки, или для использования in vivo, возраста, веса и определенного животного и части его тела, которую надо лечить, определенной нуклеиновой кислоты и использованного способа доставки, предполагаемого терапевтического или диагностического применения и формы лекарственной формы, например, суспензии, эмульсии, мицеллы или липосомы, что будет очевидно для специалистов в данной области техники. Обычно дозу вводят на более низких уровнях и увеличивают до тех пор, пока достигают желаемый эффект.
[0412] Если используют липиды для доставки нуклеиновой кислоты, количество липидного соединения, которое вводят, может варьировать и обычно зависит от количества нуклеиновой кислоты, которое вводят. Например, соотношение по массе липидного соединения к нуклеиновой кислоте составляет предпочтительно приблизительно от 1:1 до приблизительно 30:1, при этом соотношение по массе приблизительно 5:1 до приблизительно 10:1 является более предпочтительным.
[0413] Приемлемые единицы дозирования молекул нуклеиновых кислот могут находится в пределах 0,001-0,25 милиграмм на килограмм массы тела реципиента в сутки, или в пределах 0,01-20 микрограмм на килограмм массы тела в сутки или в пределах 0,01-10 микрограмм на килограмм массы тела в сутки или в пределах 0,10-5 микрограмм на килограмм массы тела в сутки или в пределах 0,1-2,5 микрограмм на килограмм массы тела в сутки.
[0414] Приемлемые количества молекул нуклеиновых кислот можно вводить и эти количества можно определить эмпирически с использованием стандартных методов. Эффективные концентрации отдельных типов нуклеиновых кислот в среде клетки могут оставлять приблизительно 1 фемтомолярную, приблизительно 50 фемтомолярную, 100 фемтомолярную, 1 пикомолярную, 1,5 пикомолярную, 2,5 пикомолярную, 5 пикомолярную, 10 пикомолярную, 25 пикомолярную, 50 пикомолярную, 100 пикомолярную, 500 пикомолярную, 1 наномолярную, 2,5 наномолярную, 5 наномолярную, 10 наномолярную, 25 наномолярную, 50 наномолярную, 100 наномолярную, 500 наномолярную, 1 микромолярную, 2,5 микромолярную, 5 микромолярную, 10 микромолярную, 100 микромолярную или больше.
[0415] Дозировка может составлять от 0,01 мкг до 1 г на кг массы тела (например, 0,1 мкг, 0,25 мкг, 0,5 мкг, 0,75 мкг, 1 мкг, 2,5 мкг, 5 мкг, 10 мкг, 25 мкг, 50 мкг, 100 мкг, 250 мкг, 500 мкг, 1 мг, 2,5 мг, 5 мг, 10 мг, 25 мг, 50 мг, 100 мг, 250 мг или 500 мг на кг).
[0416] Уровни доз порядка от приблизительно 0,1 мг до приблизительно 140 мг на кг массы тела в сутки являются полезными для лечения перечисленных выше состояний (приблизительно 0,5 мг до приблизительно 7 г для пациента в сутки). Количество активного ингридиента, который можно сочетать с носителем для получения единичной лекарственной формы, меняется в зависимости от хозяина, которого лечат, и конкретного способа введения. Единичные лекарственные формы обычно содержат от приблизительно 1 мг до приблизительно 500 мг активного ингридиента.
[0417] Будет понятно, что определенный уровень дозы для любого конкретного пациента зависит от многих факторов, включая активность использованного определеного соединения, возраст, массу тела, общее состояние здоровья, пол, питание, время введения, способ введения и скорость выведения, комбинацию лекарственного средства и тяжесть конкретного заболевания, которое лечат.
[0418] Фармацевтические композиции, которые включают молекулу нуклеиновой кислоты согласно настоящему описанию можно вводить один раз ежедневно, четыре раза в день, три раза в день, два раза в день, ежедневно или с любым интервалом и в течение любого времени, которое приемлемо с медицинской точки зрения. Тем не менее, терапевтический агент также можно включать в дозу в единицы дозирования, содержащие две, три, четыре, пять, шесть или более частей дозы, вводимых с соответствующими интервалами в течение дня. В таком случае молекулы нуклеиновых кислот, заключенные в каждой части дозы, могут быть соответственно меньше для того, чтобы обеспечить полную ежедневную единицу дозирования. Также единица дозирования может быть составлена для одной дозыв течение нескольких дней, например, с использованием традиционной лекарственной формы с замедленным высвобождением, которая обеспечивает замедленное и последовательное высвобождение dsPHK в течение нескольких дней. Лекарственные формы с замедленным высвобождением хорошо известны в данной области техники. Единица дозирования может содержать соответствующее кратное количество ежедневной дозы. Композиция может быть составлена таким образом, что сумма множественных единиц нуклеиновой кислоты вместе содержат достаточную дозу.
Фармацевтические композиции, наборы и контейнеры
[0419] Также обеспечиваются композиции, наборы, контейнеры и лекарственные формы, которые включают молекулу нуклеиновой кислоты (например, молекулу siНК) согласно настоящему описанию для снижения экспрессии hsp47 для введения или распространения молекулы нуклеиновой кислоты пациенту. Набор может включать по меньшей мере один контейнер и по меньшей мере одну этикетку. Приемлемые контейнеры включают, например, бутылки, ампулы, шприцы и пробирки. Контейнеры могут быть образованы из различных материалов, таких как стекло, металл или пластик. Контейнер может содержать аминокислотную последовательность (последовательности), малую молекулу (молекулы), последовательность (последовательности) нуклеиновой кислоты, популяцию (популяции) клеток и/или антитело (антитела). Согласно одному варианту реализации контейнер содержит полинуклеотид для использования для изучения профиля экспрессии мРНК клетки вместе с реагентами, используемыми для этой цели. Согласно другому варианту реализации контейнер содержит антитело, его связывающий фрагмент или специфичный связывающий белок для применения для оценки экспрессии белка hsp47 в клетках и тканях или для соответствующих лабораторных, прогностических, диагностических, профилактических и терапевтических целей; показания и/или указания для таких применений могут быть включены на или вместе с таким контейнеров, как и реагенты и другие композиции или инструменты, используемые для таких целей. Набор также может включать ассоциированные показания и/или указания, реагенты и другие композиции или инструменты, используемые для таких целей, также могут быть включены.
[0420] В другом случае контейнер может содержать композицию, которая является эффективной для лечения, диагностики, прогнозирования или профилактики состояния, и может иметь порт для стерильного доступа (например, контейнер может представлять собой пакет внутривенного раствора или ампулу, имеющую пробку, поддающуюся прокалыванию гиподермической иглой для инъекций). Активные агенты в композиции могут представлять собой молекулу нуклеиновой кислоты, способную к специфическому связыванию hsp47 и/или модуляции функции hsp47.
[0421] Также набор может включать второй контейнер, который содержит фармацевтически приемлемый буфер, например, фосфатно-солевой буфер, раствор Рингера и/или раствор декстрозы. Также он может включать другие материалы, желательные с коммерческой или потребительской точки зрения, включая другие буферы, разбавители, фильтры, мешалки, шприцы и/или вкладыши с показаниями и/или инструкциями для применения.
[0422] Ампулы с единичной дозой или многодозовые контейнеры, в которых молекулы нуклеиновых кислот упакованы перед использованием, могут включать герметически запаянный контейнер, содержащий количество полинуклеотида или раствор, содержащий полинуклеотид, подходящий для его фармацевтически эффективной дозы или нескольких эффективных доз. Полинуклеотид упакован в виде стерильной лекарственной формы, и герметически запаянный контейнер разработан для сохранения стерильности лекарственной формы до применения.
[0423] Контейнер, в котором находится полинуклеотид, включающий последовательность, кодирующую элемент клеточного иммунного ответа или его фрагмент, может включать упаковку, которая имеет этикетку, и этикетка может нести уведомление в форме, предписанной государственным органом, например. Управлением по контролю качества пищевых продуктов и лекарственных препаратов, которое отражает одобрение органом в рамках фередального закона изготовления, применения или распространения материала полинуклеотида для применения для человека.
[0424] Федральные законы требуют, чтобы применение фармацевтических композиций для лечения людей было одобрено агентством Федерального правительства. В Соединенных Штатах контроль является обязанностью Управления по контролю качества пищевых продуктов и лекарственных препаратов, которое выпускает соответствующие нормативные акты для гарантии такого одобрения, подробно описанные в 21 U.S.C. § 301-392. Нормативные акты для биологического материала, включая продукты, изготовленные из тканей животных, обеспечиваются 42 U.S.C. § 262. Аналогичное одобрение необходимо в большинстве зарубежных стран. Нормативные акты различаются в странах, но отдельные процедуры хорошо известны специалистам в данной области, и композиции и способы согласно настоящему описанию предпочтительно соответствуют им.
[0425] Доза для введения в большой степени зависит от состояния и размера пациента, которого необходимо лечить, а также от частоты лечения и способа введения. Схемы для продолжения терапии, включая дозу и частоту, можно установить по начальному ответу и клинической оценке. Парентеральный путь введения в интерстициальное пространство ткани является предпочтительным, но также другие парентеральные способы введения, например, вдыхание аэрозольной лекарственной формы, могут потребоваться для конкретного введения, например, на слизистые оболочки носа, горла, ткани бронхов или легкие.
[0426] Поэтому согласно настоящему описанию обеспечивается фармацевтический продукт, который может включать полинуклеотид, включающий последовательность, кодирующую элемент клеточного иммунного ответа или его фрагмент, в растворе в фармацевтически приемлемом инъекционном носителе и подходящий для введения интерстициально в ткань для того, чтобы клетки ткани экспрессировали элемент клеточного иммунного ответа, контейнер содержит раствор и уведомление вместе с контейнером в форме, предписанной правительственным агенством, регулирующим изготовление, применение или распространение лекарственных средств, которое отражает одобрение органом изготовления, применения или распространения раствора полинуклеотида для применения для человека.
Показания
[0427] Молекулы нуклеиновых кислот согласно настоящему описанию можно использовать для лечения заболеваний, состояний или нарушений, ассоциированных с hsp47, таких как фиброз печени, цирроз, фиброз легких, фиброз почек, перитонеальный фиброз, хроническое поражение печени и фибриллогенез, и любого другого заболевания или состояния, которые связаны или отвечают на уровни hsp47 в клетке или ткани, по отдельности или в комбинации с другими способами лечения. По этой причине композиции, наборы и методы согласно настоящему описанию могут включать упаковку молекулы нуклеиновой кислоты согласно настоящему описанию, которая включает этикету или аннотацию. Этикета может включать показания к применению молекул нуклеиновых кислот, например, для лечения или предотвращения фиброза печени, перитонеального фиброза, фиброза почек и фиброза легких и любого другого заболевания или состояний, которые связаны или ответят на уровни hsp47 в клетке или ткани, по отдельности или в комбинации с другими способами лечения. Этикетка может включать показание к применению для снижения экспрессии hsp47. «Аннотация» обозначает инструкции, обычно вложенные в коммерческие упаковки лекарственных продуктов, которые содержат информацию о показаниях, применении, дозировке, введении, противопоказаниях, других терапевтических средствах, которые можно сочетать с упакованным продуктом, и/или предупреждения, касающиеся применения таких терапевтических продуктов и т.д.
[0428] Специалистам в данной области техники будет ясно, что другие способы лечения фиброза, лекарственные средства и способы лечения, известные в данной области техники можно с легкостью сочетать с молекулами нуклеиновых кислот согласно настоящему описанию (например, молекулами siНК), и, таким образом, это также предусматривается настоящим изобретением.
[0429] Способы и композиции согласно настоящему описанию будут описаны более подробно с помощью следующих не ограничивающих примеров.
ПРИМЕР 1
Отбор последовательностей нуклеиновой кислоты hsp47:
[0430] Молекулы нуклеиновых кислоты (например, siНК≤25 нуклеотидов) против Hsp47 получали с использованием нескольких компьютерных программ, включая siRNA в Институте Whitehead (Whitehead Institute for Biomedical Research), IDT siРНК Design (Integrated DNA Technologies), BLOCK-iT RNAi Designer (Invitrogen), siDESIGN Center (Dharmacon) и BIOPREDsi (Friedrich Miescher Institute for Biomedical Research, часть Novartis Research Foundation, ссылка http://www.biopredsi.org/start.html). Последовательности самых верхних siРНК из этих программ сравнивали и отбирали (смотрите Таблица 1) по алгоритмам, а также гомологии последовательностей между человеком и крысой. Кандидатные последовательности подтверждали с помощью in vitro анализов нокаута.
[0431] При отборе последовательностей молекул нуклеиновых кислот (например, 21-мерной siРНК) учитывали несколько параметров. Примеры параметров включают:
1) термодинамическую стабильность (RISC благоприятствует цепи с менее стабильным 5'-концом)
2) содержание пар GC 30-52%
3) предпочтительное расположение нуклеотидов:
(C/G)NNNNNNNN(A/U)10NNNNNNNN(A/U), при этом N представляет собой любой нуклеотид
4) лишенный предполагаемых иммуностимуляторных мотивов
5) 3'-выступающий конец из 2 нуклеотидов
6) расположение siРНК в пределах транскрипта (предпочтительно в пределах области кДНК)
7) специфичность последовательностей (проверено с использованием BLAST)
8) различия по отдельным нуклеотидам при проверке базы данных SNP
[0432] Разработали последовательности siРНК, содержащие ≤25 нуклеотидов, на основе изложенных методов. Получили соответствующую siРНК - субстрат для Dicer (например, ≥26 нуклеотидов), основанный на меньших последовательностях, и продлили сайт-мишень siНК≤25 нуклеотидов посредством добавления 4 оснований к 3'-концу смысловой цепи и 6 оснований к 5'-концу антисмысловой цепи. Субстраты для Dicer, которые были получены, обычно имели смысловую цепь из 25 оснований, антисмысловую цепь из 27 оснований с ассимметричной молекулой, имеющей тупой конец и выступающий 3'-конец. Последовательности смысловой и антисмысловой цепи без модификации основания (последовательности основания) и с модификациями (экспериментальная последовательность) указаны в Таблице 1.
ПРИМЕР 2
[0433] Для скрининга эактивности различных молекул siHK в отношении генов hsp47 крысы и человека получали разнообразные репортерные клеточные линии посредством введения кДНК HSP47 человека, меченую зеленым флуоресцентным белком (GFP) или конструкции кДНК GP46 крысы с GFP в клеточную линию 293, НТ1080, линию HSC человека hTERT или линию NRK. Затем в указанных клеточных линиях определяли siPHK против GFP. Измеряли сигнал остаточной флуоресценции и нормировали его относительно случайной siPHK (Ambion) и затем нормировали по жизнеспособности клеток. Результаты показали, что siPHK против GFP подавляет флуоресценцию в различной степени в разных клеточных линиях (Фиг. 1). Клеточные линии 293_HSP47-GFP и 293_GP46-GFP выбирали с помощью скрининга siHsp47 по причине легкости их трансфекции и чувствительности к подавлению флуоресценции.
Трансфекция siPHK:
[0434] Клетки трансфицировали 1,5 пмоль на лунку siHK против GFP в 96-луночных планшетах для культур клеток с использованием Lipofectamine РНКиМАХ (Invitrogen) методом обратной трансфекции. Клетки высеивали в плотности 6,000 клеток на лунку и смешивали с комплексами siHK. Флуоресценцию считывали через 72 часа инкубации на ридере микропланшетов Synergy 2 Multi-Mode (BioTek).
Анализ жизнеспособности клеток:
[0435] Определяли жизнеспособность клеток, обработанных или не обработанных siHK, через 72 часа инкубации с использованием набора CellTiter-Glo Luminescent Cell Viability Assay Kit в соответствии с инструкцией (Promega). Результаты считывания нормировали относительно образцов, обработанных случайными молекулами siHK.
ПРИМЕР 3
Оценка эффективности подавления siHsp47 экспрессии hsp47 в репортерных клеточных линиях
[0436] Оценивали эффективность подавления siHK hsp47 в клеточных линиях 293 HSP47-GFP и 293_GP46-GFP посредством регистрации изменений сигнала флуоресценции репортерного GFP. Эксперименты проводили согласно описанию в Примере 2. Сигналы флуоресеценции нормировали относительно сигналов флуоресценции от клеток, обработанных случайными siPHK (Ambion), которые служили контролем. Результаты показали, что исследованные молекулы siНК hsp47 были эффективными в отношении подавления мРНК hsp47 в обеих клеточных линиях. Однако siНК против GP46 мРНК (согласно публикации 2008 Sato et al) была эффективна только в отношении клеточной линии 293_GP46-GFP. Результаты показаны на Фигуре 2А-В.
[0437] Определяли жизнеспособность клеточных линий 293_HSP47-GFP и 293_GP46-GFP, обработанных siРНК против hsp47 и gp46, с использованием способов, описанных в Примере 2. Жизнеспособность клеток нормировали относительно клеток, обработанных случайными siРНК (Ambion). Результаты показали, что обработка молекулами siНК не оказывала существенного действия на жизнеспособность клеток. Тем не менее, жизнеспособность клеток клеточной линии 293_HSP47-GFP, обработанных различными молекулами siНК hsp47, различалась в зависимости от использованных молекул siНК, тогда как жинеспособность клеточных линий 293_GP46-GFP была похожа. Жинеспособность клеток 293_HSP47-GFP была ниже у клеток, обработанных siHsp47-6 и Hsp47-7, чем у остальных клеток. Результаты показаны на Фигуре 2C-D.
ПРИМЕР 4
Оценка подавляющего действия siHsp47 на мРНК hsp47 методом количественной ПЦР TaqMan®
[0438] В Примере 3 эффективность подавления siHsp47s в репортерных клеточных линиях оценивали по изменению сигнала флуоресценции. Для того, чтобы обосновать результат уровня мРНК, siРНК, направленные на эндогенный hsp47, трансфицировали стволовые клетки человека линии hTERT с использованием Lipofectamine РНКиМАХ (Invitrogen) методом обратной трансфекции согласно описанию в Примере 2.
[0439] Уровень мРНК hsp47 оценивали по эффективности подавления различных исследованных молекул siНК siHsp47. Вкратце, мРНК выделяли из клеток hTERT через 72 часа после трансфекции с использованием мининабора RNeasy (Qiagen). Уровень мРНК hsp47 определяли методом количественной ПЦР с обратной транскрипцией с использованием зондов TaqMan®. Вкратце, синтез кДНК проводили с использованием набора High-Capacity cDNA Reverse Transcription (ABI) согласно инструкции производителя, и проводили ее анализ методом TaqMan Gene Expression Assay (ABI, hsp47 анализ ID Hs01060395_gl). Уровень мРНК hsp47 нормировали относительно уровня мРНК GAPDH согласно инструкции производителя (ABI). Результаты показали, что siHsp47-C была наиболее эффективной среди всех siРНК hsp47, siHsp47-2 и siHsp47-2d находились на втором месте по эффективности. Комбинации siHsp47-1 с siHsp47-2 или siHsp47-1 с siHsp47-2d были более эффективны, чем siHsp47-1 по отдельности. Результаты показаны на Фигуре 3.
ПРИМЕР 5
Подтверждение действия подавления siHsp47 на уровне белка
[0440] Подавляющее действие различных молекул siНК Hsp47 (siHsp47) на экспрессию мРНК hsp47 подтверждали на уровне белка с помощью измерения HSP47 в клетках hTERT, трансфицированных различными siHsp47. Трансфекцию клеток hTERT различными siHsp47 осуществляли согласно описанию в Примере 2. Трансфицированные клетки hTERT лизировали и лизаты клеток осаждали центрифугированием. Белки в очищенном лизате клеток разделяли с помощью электрофореза в полиакриламидном геле SDS. Уровень белка HSP47 в лизате клеток определяли с использованием антитела к HSP47 (Assay Designs) в качестве первичного антитела, антитела козы против IgG мыши, конъюгированных с HRP (Millipore) в качестве вторичного антитела с последующей детекцией с использованием набора Supersignal West Pico Chemiluminescence (Pierce). Антитело к антину (Abeam) использовали в качестве контроля загрузки белка. Результаты показали значительное снижение уровня белка Hsp47 в клетках, обработанных siHsp47-C, siHsp47-2d, по отдельности или комбинацией siHsp47-1 с siHsp47-2d.
ПРИМЕР 6
Подавление экспрессии коллагена I siРНК hsp47
[0441] Для определения влияния siHsp47 на уровень экспрессии коллагена I определяли уровень мРНК коллагена I в клетках hTERT, обработанных различными siРНК против hsp47. Вкратце, клетки hTERT трансфицировали различными siHsp47 согласно описанию в Примере 2. Клетки лизировали через 72 часа и выделяли мРНК с использованием мининабора RNeasy согласно инструкции (Qiagen). Уровень мРНК коллагена I определяли методом количественной ПЦР с обратной транскрипцией с использованием зондов TaqMan®. Вкратце, синтез кДНК проводили с использованием набора High-Capacity cDNA Reverse Transcription (ABI) согласно инструкции и проводили ее анализ методом TaqMan Gene Expression Assay (ABI, COL1A1 ID набора Hs01076780_g1). Уровень мРНК коллагена I нормировали относительно уровня мРНК GAPDH согласно инструкции производителя (ABI). Сигналы нормировали относительно сигнала, полученного от клеток, трансфицированных случайными siНК. Результаты показали, что уровень мРНК коллагена I значительно снижен в клетках, обработанных некоторыми кандидатными siHsp47-2, siHsp47-2d и их комбинациями с siHsp47-1, как показано на Фигуре 4.
ПРИМЕР 7
Иммунофлуоресцентное окрашивание клеток hTERT, обработанных siРНК hsp47
[0442] Для визуализации экспрессии двух маркеров фиброза - коллагена I и альфа-актина гладких мышц (SMA) - в клетках hTERT, трансфицированных или не трансфицированных siHsp47, клетки окрашивали антителом кролика против коллагена I (Abeam) и мышиным антителом против альфа-SMA (Sigma). Антитела козы Alexa Fluor 594 против IgG мыши и антитела козы Alexa Fluor 488 против IgG кролика (Invitrogen (Molecular Probes)) использовали в качестве вторичных антител для визуализации коллагена I (зеленый) и альфа-SMA (красный). Hoescht использовали для визуализации ядра (голубой). Результаты показали наличие корреляции между подавлением siРНК некоторых генов-мишеней и экспресией коллагена/SMA.
ПРИМЕР 8
Исследование SJHSP47 in vivo на животных моделях фиброза печени
[0443] siРНК для лечения цирроза печени крыс.
Последовательность дуплекса siРНК HSP47 (siHSP47C) представлена ниже.
Смысловая (5'->3') ggacaggccucuacaacuaTT
Антисмысловая (5'->3') uaguuguagaggccuguccTT
[0444] Маточный раствор siРНК 10 мг/мл готовили посредством, растворяя в воде, не содержащей нуклеаз (Ambion). Для лечения крыс с циррозом готовили лекарственную форму siРНК с липосомой, соединенной с витамином А, согласно описанию в публикации Sato et al (Sato Y. et al. Nature Biotechnology 2008. Vol.26, p431) для того, чтобы воздействовать на активированные звездчатые клетки печени, которые продуцируют коллаген. Композиция витамин A (VA)-липосома-siРНК состояла из 0,33 мкМоль/мл VA, 0,33 мкМоль/мл липосомы (Coatsome EL-01-D, NOF Corporation) и 0,5 мкг/мкл siРНК в 5% растворе глюкозы.
[0445] Сообщение о животной модели цирроза печени приводится в публикации Sato et al (Sato Y. et al. Nature Biotechnology 2008. Vol.26, p431). Цирроз печени у 4-недельных самцов мышей SD вызывали 0,5% диметилнитрозамином (DMN) (Wako Chemicals, Japan) в фосфатном буфере (PBS). Дозу 2 мл/кг массы тела вводили интраперитонеально в течение 3 последовательных дней в неделю на 0, 2, 4, 7, 9, 11, 14, 16, 18, 21, 23, 25, 28, 30, 32, 34, 36, 38 и 40 сутки.
[0446] Обработка siРНК: Обработку siРНК проводили, начиная с 32 дня и в течение 5 внутривенных инъекций. Более подробно, крыс обрабатывали siРНК на 32, 34, 36, 38 и 40 сутки. Затем крыс забивали на 42 или 43 сутки. Исследовали 3 различных дозы siРНК (1,5 мг siРНК на кг массы тела, 2,25 мг siРНК на кг массы тела, 3,0 мг siРНК на кг массы тела). Подробное описание групп исследования и количество животных в каждой группе представлено ниже:
1) Цирроз вызывали инъекцией DMN, затем вводили 5% глюкозу вместо siРНК) (n=10)
2) VA-Lip-siHSP47C 1,5 мг/кг (n=10)
3) VA-Lip-siHSP47C 2,25 мг/кг (n=10)
4) VA-Lip-siHSP47C 3,0 мг/кг (n=10)
5) Ложно обработанные животные (вместо DMN вводили PBS. 5% глюкозу вводили вместо siРНК) (n=6)
6) Контроль без обработки (интактный) (n=6)
VA-Lip обозначает комплекс витамин А - липосома.
[0447] Оценка терапевтической эффективности: На 43 день 2 из 10 животных в группе «заболевшие крысы» и 1 из 10 животных в группе «VA-Lip-siHSP47C siРНК 1,5 мг/кг» умерли из-за развития цирроза печени. Оставшиеся животные выжили. После того, как животных забили, ткани печени фиксировали в 10% формалине. Затем левую долю каждой печени заключали в парафин для гистологического исследования. Срезы ткани окрашивали Сириус красным и гематоксилином и эозином (ГЭ). Окрашивание Сириус красным использовали для визуализации отложений коллагена для определения степени цирроза. Окрашивание ГЭ использовали для контрастной окраски ядер и цитоплазмы. Каждое стекло рассматривали под микроскопом (BZ-8000, Keyence Corp.Japan) определяли и процент площади на срезе, окрашенной Сириус красным, с помощью программного обеспечения анализа изображений, прилагаемого к микроскопу. Из каждой печени приготовили по меньшей мере 4 стекла для анализа изображений, и всю площадь каждого стекла (среза печени) фотографировали камерой и анализировали. Статистическую обработку проводили методом t-критерия Стюдента.
[0448] Результаты: На Фигуре 5 показаны области фиброза. Площадь фиброза в группе «заболевших крыс» была больше, чем в «ложно обработанных» или «не обработанных» группах. Таким образом, обработка DMN вызывала отложение коллагена в печени, что обычно наблюдается при фиброзе печени. Тем не менее, площадь фиброза была значительно снижена при обработке siРНК, направленной на ген HSP47, по сравнению с группой «заболевших крыс» (Фигура 5). Полученные результаты показали, что siРНК согласно настоящему описанию обладает терапевтической эффективностью при данном заболевании.
[0449] Дополнительные соединения siРНК исследовали на животной модели фиброза печени и показали, что они снижают площадь фиброза в печени.
ПРИМЕР 9
Получение последовательностей активных соединений двухцепочечной РНК к HSP47/SERPINH1 и получение siРНК, представленных в Таблицах А-18, А-19 и В-Е.
[0450] Используя собственные алгоритмы и известные последовательности генов-мишеней, синтезировали последовательности многих сильных siРНКs. Эти последовательности, которые синтезировали с использованием данного способа, являлись комплементарными соответствующей последовательности мРНК.
[0451] Дуплексы получали посредством гибридизации комплементарных одноцепочечных олигонуклеотидов. В ламинарном боксе готовили приблизительно 500 мкМоль маточного раствора одноцепочечных олигонуклеотидов посредством разведения в WFI (вода для инъекций, Norbrook). Фактические концентрации ssPHK определяли, разбавляя каждые 500 мкМоль ssPHK в отношении 1:200 с использованием WFI и измеряя OD с использованием Nano Drop. Эту процедуру повторяли 3 раза и рассчитывали среднюю концентрацию. Затем разбавляли маточный раствор до конечной концентрации 250 мкМоль. Комплементарные отдельные цепи гибридизовали при нагревании до 85°С и охлаждении до комнатной температуры в течение по меньшей мере 45 минут. Проверяли полную гибридизацию дуплексов посредством нанесения 5 мкл на 20% полиакриламидный гель и окрашивания. Образцы хранили при -80°С.
[0452] В Таблицах А-18, А-19 и В-Е представлены siРНКs для HSP47/SERPINH1. Для каждого гена существует отдельный список 19-мерных последовательностей siРНК, которые являются приоритетными на основе количества баллов у них в собственном алгоритме в качестве наилучших последовательностей для нацеливания на экспрессию гена человека.
[0453] Следующие сокращения используются в Таблицах А-18, А-19 и В-Е в настоящем описании: «другие виды или Sp» обозначает межвидовую идентичность с другими животными: D-собака, Rt-крыса, Rb-кролик, Rh-макака Резуса, Р- свинья, М-мышь; ORF: открытая рамка считывания. 19-меры и 18+1-меры обозначают олигомеры длиной 19 и 18+1 (U в положении 1 антисмысловой цепи, А в положении 19 смысловой цепи) рибонуклеиновых кислот, соответственно.
In vitro исследования нацеливания соединений siРНК на гены
Скрининг с низкой пропускной способностью (LTS) олигов siРНК, направленных на ген SERPINH1 человека и крысы.
[0454] Клеточные линии: клетки аденокарциномы простаты человека РСЗ (АТСС, Cat# CRL-1435) культивировали в среде RPMI с добавлением 10% FBS и 2 мМоль L-глутамина, и клетки рака шейки матки человека HeLa (ATCC, Cat#CCL-2) культивировали в среде Игла в модификации Дульбекко (DMEM) с добавлением 10% FBS, 2 мМоль L-глутамина. Клетки культивировали при 37°С и 5% CO2.
[0455] Приблизительно 2х105 клеток РС-3 человека, эндогенно экспрессирующих ген SERPINH1, вносили в 1,5 мл питательной среды для достижения 30-50% конфлюэнтности через 24 часа. Клетки трансфицировали реагентом Lipofectamine™2000 в конечной концентрации 0,01-5 нМоль на трансфицированные клетки. Клетки инкубировали при 37±1°С, 5% СО2 в течение 48 часов. Получали клетки, трансфицированные siРНК, и выделяли РНК с использованием набора EZ-PHK [Biological Industries (#20-410-100)].
[0456] Обратную транскрипцию проводили следующим образом: Синтезировали кДНК и методом ПЦР в режиме реального времени определяли уровни мРНК SERPINH1 и нормировали их относительно уровней мРНК циклофилина A (CYNA, PPIA) для каждого образца. Определяли активность siРНК на основе соотношения количества мРНК SERPINH1 в образцах, обработанных siРНК, и в нетрансфицированных контрольных образцах.
[0457] Самые активные последовательности отобрали для дальнейших анализов. В качестве предпочтительных соединений из Таблицы А-18 были выбраны соединения siРНК SERPINH1_2, SERPINH1_6, SERPINH1_13, SERPINH1_45 SERPINH1_45a, SERPINH1_51, SERPINH1_51a, SERPINH1_52 и SERPrNH1_86. В качестве предпочтительных соединений из Таблицы А-19 были выбраны соединения siРНК SERPINH1_4, SERPINH1_12, SERPINH1_18, SERPINH1_30, SERPINH1_58 и SERPINH1_88.
[0458] Другие предпочтительные соединения включали SERPINH1_50, SERPINH1_67, SERPINH1_73, SERPINH1_74.
Значения IC50 для выбраных LTS олигов siРНК SERPINH1
[0459] Приблизительно 2х105 клеток РС-3 человека или 0,9х105 клеток REF52 крысы, эндогенно экспрессирующих ген SERPINH1, инокулировали в 1,5 мл питательной среды для достижения 30-50% конфлюэнтности. Клетки трансфицировали соединениями двухцепочечных РНК SERPINH1 (т.е. SERPINH1_2, 4, 6, 12, 13, 18, 45, 51, 58, 88) реагентом Lipofectamine™2000 для достижения конечной концентрации трансфекции в пределах 0,0029-100 нМоль. В качестве отрицательного контроля клетки обрабатывали реагентом Lipofectamme™2000 или синтетическими siРНК со случайной последовательностью, не специфичными для мишени в конечных концентрациях 20-100 нМоль. В качестве положительного контроля эффективности трансфекции использовали клетки, трансфицированные siРНК, меченые Су3.
[0460] Клетки инкубировали при 37±1°С, 5% CO2 в течение 48 часов. Получали клетки, трансфицированные siРНК, и выделяли РНК с использованием набора EZ-RNA [Biological Industries (#20-410-100). Обратная транскрипция: проводили синтез кДНК и определяли уровни мРНК SERPINH1 человека методом ПЦР в режиме реального времени и нормировали их относительно уровня мРНК циклофилина A (CYNA, PPIA) в каждом образце.
[0461] Значения IC50 активности исследованной РНКи определяли с помощью построения кривой доза-эффект с использованием результатов активности, полученных с различными конечными концентрациями siРНК. Кривую доза-эффект строили, нанося на диаграмму относительное количество остаточной мРНК SERPINH1 в зависимости от логарифма концентрации трансфицирующей siРНК. Кривую рассчитывали, вычерчивая лучшую сигмоидную кривую по измеренным данным. Метод построения сигмоидной кривой также известен как построение кривой по трем точкам.
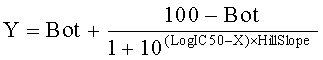
где У обозначает остаточный ответ мРНК SERPINH1, Х представляет логарифм концентраций трансфицированных siРНК, Bot представляет собой значение Y по нижнему плато, LogIC50 представляет собой значение X, когда Y находится посередине между нижним и верхним плато, и HillSlope представляет крутизну кривой.
[0462] Процент подавления экспрессии гена с использованием специфичных siРНК определяли с использованием анализа гена-мишени в клетках, экспрессировавших эндогенный ген, методом количественной ПЦР. Другие соединения siРНК из Таблиц А-18, А-19 и В-Е исследовали in vitro, при этом оказалось, что указанные соединения siРНК подавляют экспрессию генов. Активность показана как доля остаточной мРНК; соответственно, чем меньше значение, тем выше активность.
[0463] Для проверки стабильности соединений siРНК в сыворотке определенные молекулы siРНК инкубировали в четырех различных партиях сыворотки человека (100% концентрация) при 37°С в течение 24 часов. Образцы собирали через 0,5, 1, 3, 6, 8, 10, 16 и 24 часа. Параметры миграции, как показатели, определяли в каждый момент сбора с помощью электрофореза в полиакриламидном геле (PAGE).
[0464] В таблице 3 показана IC50 (или активность, если не рассчитана IC50) в линии клеток человека для соединений немодифицированной двухцепочечной нуклеиновой кислоты (смысловая и антисмысловая цепи немодифицированы, dTdT выступающие концы на 3'-конце), выбранных из Таблиц А-18 и А-19.
Подавляющая активность siРНК:
[0465] Приблизительно 2х105 клеток PC-3 человека, эндогенно экспрессирующих ген SERPINH1, высеивали в лунку 6 луночных планшетов и оставляли расти в течение 24 часов до 30-70% конфлюэнтности. Клетки трансфицировали исследуемыми siРНК в различных концентрациях с использованием реагента Lipofectamine™2000 (Invitrogen). Клетки инкубировали при 37°С в инкубаторе при 5% CO2 в течение 48 часов или 72 часов. Через 48-72 часа после трансфекции собирали клетки и из клеток выделяли РНК. Меченые Су3 дуплексы siРНК использовали в качестве положительного контроля эффективности трансфекции. Ложно трансфицированные клетки, обработанные реагентом Lipofectamine™2000, обозначали как «контрольные не активные образцы» («отрицательный контроль»), и клетки, обработанные siРНК (HSP47-C) с известной активностью в конечной концентрации 5 нМоль, обозначали как «контрольные активные образцы» («положительный контроль»). Z' и порядки контролей {порядок=среднее(отрицательный)/средний(положительный)} представляют собой средства для описания эффективности анализа.
[0466] Относительное подавление экспрессии гена-мишени каждой исследованной siРНК определяли методом количественной ПЦР мРНК мишени из клеток. Проводили обратную транскрипцию посредством синтезирования кДНК из клеток и определения уровней мРНК гена-мишени методом ПЦР в режиме реального времени. Измеренные уровни мРНК в клетке нормировали относительно уровней мРНК циклофилина A (CYNA, PPIA) для каждого образца. Активность siРНК определяли по соотношению количества мРНК гена-мишени в образцах, обработанных siРНК, и в контрольных не трансфицированных образцах. Z' и порядки контролей {порядок=среднее(отрицательный)/средний(положительный)} представляют собой средства для описания эффективности анализа.
[0467] Результатами количественной ПЦР считали те, которые удовлетворяли стандартам QC, т.е. величина уклона стандартной кривой находилась в интервале [-4, -3], R2>0.99, без димеров праймеров. Результаты, которые не соответствовали требованиям QC, исключали из анализа.
[0468] Значение IC50 активности исследованной РНКи определяли с помощью построения кривой доза-эффект с использованием результатов активности, полученных при различных конечных концентрациях siРНК. Кривую доза-эффект строили, нанося на диаграмму относительное количество остаточной мРНК SERPINH1 в зависимости от логарифма концентрации трансфицирующей siРНК, как описано выше.
Проверка направленности и ненаправленности молекул двухцепочечных РНК:
[0469] Система psiCHECK позволяет оценить способность «ведущей цепи» (GS) (антисмысловой) и цепи-спутника (PS) (смысловой цепи) вызывать направленные (целевые) и ненаправленные эффекты посредством отслеживания изменений уровней экспрессии их последовательностей-мишеней. Приготовили четыре конструкции с использованием psiCHECK™-2 (Promega) для оценки направленной активности и возможной ненаправленной активности каждой исследованной GS и PS цепи молекулы. В каждой конструкции одну копию или три копии полной последовательности или последовательности затравки исследуемой молекулы PS или GS клонировали в сайт множественного клонирования, расположенный даунстрим от стоп-кодона трансляции люциферазы Renilla в 3'-UTR области.
[0470] Полученные векторы были названы:
[0471] 1-GS-CM (ведущая цепь, полное соответствие) вектор, содержащий одну копию полной последовательности мишени (последовательности нуклеотидов, полностью комплементарной целой последовательности из 19 оснований GS исследуемой молекулы);
[0472] 2-PS-CM (цепь-спутник, полное соответствие) вектор, содержащий одну копию полной последовательности мишени (последовательности нуклеотидов, полностью комплементарной целой последовательности из 19 оснований PS исследуемой молекулы);
[0473] 3-GS-SM (ведущая цепь, соответствие затравки) вектор, содержащий одну копию или три копии участка затравки последовательности-мишени (последовательности, комплементарной нуклеотидам 1-8 GS исследуемой молекулы);
[0474] 4-PS-SM (сопровождающая цепь, соответствие затравки) вектор, содержащий одну копию участка затравки последовательности-мишени (последовательности, комплементарной нуклеотидам 1-8 PS исследуемой молекулы).
Номенклатура:
[0475] Ведущая цепь: цепь siРНК, которая проникает в комплекс RISC и направляет расщепление/сайленсинг комплементарной последовательности РНК
[0476] Последовательность затравки: Нуклеотиды 2-8 от 5'-конца руководящей цепи.
[0477] cm (полное соответствие): Фрагмент ДНК, полностью комплементарный руководящей цепи siРНК. Этот фрагмент ДНК клонировали в 3'UTR репортерного гена и он служит мишенью для непосредственного сайленсина РНК.
[0478] sm (соответствие затравки): 19-мерный фрагмент ДНК с нулеотидами ns 12-18 полностью комплементарными ns 2-8 руководящей цепи siРНК. Этот фрагмент ДНК клонировали в 3'UTR репортерного гена, и он служит мишенью для «ненаправленного» сайленсинга.
[0479] X1: одна копия cm или sm, клонированная в 3'UTR репортерного гена.
[0480] Х3 Три копии cm или sm, клонированные в 3'UTR репортерного гена, отделенные друг от друга 4 нуклеотидами.
[0481] Последовательности-мишени клонировали с использованием сайтов рестрикции, совместимых с ферментами XhoI и NotI. Смеси для гибридизации готовили в плотно закрытых пробирках типа эппендорф 0,5 мл, нагревали на водяной бане до 85°С, погружили в кипящую водяную баню и в конце постепенно охлаждали до комнатной температуры.
[0482] Лигирование: Двухцепочечные олигонуклеотиды, полученные с помощью процедуры гибридизации, лигировали с линеаризированной (с помощью XhoI и NotI) psiCHECK™-2 и трансфицировали ими клетки с использованием стандартных методов. Определяли положительные колонии и секвенировали для подтверждения ставленной последовательности. В Таблице 5 показаны нуклеотидные последовательности вставленных олигонуклеотидов.
[0483] Соответствующие цепи, как описано выше, клонировали в 3'UTR репортерной мРНК люциферазы Renilla в вектор psiCHECK™-2 (Promega). XhoI и NotI использовали в качестве сайтов клонирования с использованием стандартных олекулярно-биологических методов. Каждую цепь синтезировали химически и гибридизовали посредством нагревания до 100°С и охлаждали до комнатной температуры. Лигирование проводили в течение 3 часов с использованием стандартных молекулярнобиологических методов и трансформировали клетки Е.coli DH5a. Проводили скрининг полученных колоний на К заявке №2012122470 наличие конструкций плазмиды методом ПЦР колоний с использованием соответствующих праймеров. Каждую плазмиду (вектор) выделяли из одной положительной колонии и подтверждали ее последовательность.
[0484] Приблизительно 1,3х106 клеток HeLa человека инокулировали в чашку Петри диаметром 10 см. Затем клетки инкубировали при 37±1°С, 5% CO2 в инкубаторе в течение 24 часов. Через день после инокуляции меняли питательную среду на 8 мл свежей питательной среды, и каждую чашку трансфицировали одной из плазмид, указанных выше, с использованием реагента Lipofectamine™2000 по протоколу изготовителя и инкубировали в течение 5 часов при 37±1°С и 5% CO2. После инкубации клетки высеивали в 96-луночный планшет в конечной концентрации 5х103 клеток на лунку в 80 мкл питательной среды. Через 16 часов клетки трансфицировали молекулами siРНК SERPINH1 с использованием реагента Lipofectamme™2000 в различных концентрациях в пределах от 0,001 нМоль до 5 нМоль в конечном объеме 100 мкл. Ложно трансфицированные клетки, обработанные реагентом Lipofectamine™2000 с соответствующей плазмидой psiCHECK™-2, обозначали как «контрольные неактивные образцы» (отрицательный контроль), и клетки, обработанные известной активной siРНК (HSP47-C) в конечной концентрации 5 нМоль, обозначали как «контрольные активные образцы» (положительный контроль). Z' и порядки контролей {порядок=среднее(отрицательный)/средний(положительный)} представляют собой средства для описания эффективности анализа.
[0485] Затем клетки инкубировали в течение 48 часов при 37±1°С и определяли активности люциферазы Renilla и FireFly в каждом трансфицированном siРНК образце с использованием набора Dual-Luciferase® (Promega, Cat#E1960) согласно инструкции производителя. Активность синтетической siРНК в отношении указанной последовательности-мишени приводила к расщеплению и последующему разрушению гибридных мРНК или подавлению трансляции кодируемого белка. Таким образом, измерение снижения активности люциферазы Renilla обеспечивает удобный способ наблюдения за действием siРНК, тогда как люцифераза Firefly позволяет нормировать экспрессию люциферазы Renilla. Значение активности Renilla делили на значение активности люциферазы Firefly для каждого образца (нормирование). В конечном итоге активность люциферазы Renilla выражали в виде доли от нормированного значения активности в исследованном образце по сравнению с «контрольными неактивными образцами».
[0486] Результаты уровней TNFα и IL-6 в мононуклеарных клетках периферической крови (МКПК), подвергнутых действию немодифицированных или модифицированных siРНК/Lipofectamine™2000. Результаты представлены в пг/мл, рассчитанных по стандартной кривой. «Контроль Lipofec2000» обозначает уровень секреции цитокинов, вызванной реагентом трансфекции Lipofectamine™2000. Ни один из модифицированных соединений уровни цитокинов TNFa или IL6 выше контрольного реагента трансфекции.
[0487] Данные индукции генов, отвечающих на интерферон (IFN), MX1 и IFIT1, немодифицированными и модифицированными соединениями двухцепочечных нуклеиновых кислот. Представленные результаты являются остаточными генами IFIT1 и MX1 человека (порядок от контрольных клеток, обработанных Lipofectamine2000) по исследованиям на МКПК человека. Данные показывают, что все модифицированные соединения вызывали незначительные уровни генов, контролируемых IFN, по сравнению с немодифицированными соединениями (_S709).
[0488] В Таблицах ниже показана активность Соединения_1, Соединения_2, Соединения_3 и Соединения_4 по сравнению с немодифицированными (_S709) соединениями в клетках крысы. Результаты показаны в остаточной мишени (% от контрольных клеток, обработанных Lipofectamine™2000) гена SERPINH1 крысы в клетках REF52. Показаны результаты двух отдельных экспериментов. Подавление гена-мишени в клетках крысы соотносится с исследованию соединений на животных моделях заболеваний человека.
Анализ стабильности в сыворотке
[0489] Модифицированные соединения исследовали на стабильность дуплекса согласно настоящему изобретению в сыворотке человека или экстракте ткани человека, как описано ниже:
[0490] Молекулы siРНК в конечной концентрации 7 мкМоль инкубировали при 37°С в 100% сыворотке человека (Sigma Cat# H4522). (Маточный раствор siРНК 100 мкМоль, разведенный в сыворотке человека 1:14,29 или экстракте ткани человека из различных типов ткани). Пять мкл (5 мкл) добавляли к 15 мкл 1,5хТВЕ-загрузочного буфера в различные моменты времени (например, 0, 30 минут, 1 час, 3 часа, 6 часа, 8 часов, 10 часов, 16 часов и 24 часа). Образцы незамедлительно замораживали в жидком азоте и хранили при -20°С.
[0491] Каждый образец вносили в неденатурирующий 20% акриламидный гель, приготовленный методами, известными в данной области техники. Олигонуклеотиды визуализировали бромистым этидием в УФ свете.
Анализ экзонуклеазной стабильности
[0492] Для исследования стабилизирующего действия 3' ненуклеотидных групп на смысловую цепь молекулы нуклеиновой кислоты, антисмысловую цепь и гибридизованный дуплекс siРНК инкубировали с экстрактами цитозоля, приготовленными из клеток различных типов.
Экстракт: экстракт цитозоля НСТ116 (12 мг/мл).
Буфер для экстрактции: свежий раствор 25 мМоль Hepes pH=7,3 при 37°С; 8 мМоль MgCl; 150 мМоль NaCl с 1 мМоль DTT добавили непосредственно перед использованием.
[0493] Метод: 3,5 мл исследуемой siРНК (100 мМоль) смешали с 46,5 мл, содержащими 120 мг экстракта цитозоля НСТ116. 46,5 мл состояли из 12 мл экстракта НСТ116 и 34,5 мл буфера для экстракции с добавлением DTT и коктейля ингибиторов протеаз/100 (Calbiochem, setIII-539134). Конечная концентрация siРНК в пробирке для инкубации составляла 7 мМоль. Образец инкубировали при 37°С, и в указанный момент времени 5 мл переносили в новую пробирку, смешивали с 15 мл буфера для внесения 1ХТВЕ-50% глицерола и быстро замораживали в жидком N2. Конечная концентрация siРНК в буфере для внесения составляла 1,75 мМоль (21 нг siРНК/мл). Для анализа методом нативного PAGE и окрашивания EtBr 50 нг вносили в дорожку. Для Нозерн-блоттинга 1 нг исследуемой siРНК вносили в дорожку.
Врожденный иммунный ответ на молекулы siРНК SERPINH1:
[0494] Свежую кровь человека (при комнатной температуре) смешивали в соотношении 1:1 со стерильным 0,9% NaCl при комнатной температуре (RT) и аккуратно наслаивали (в соотношении 1:2) на Фиколл (Lymphoprep, Axis-Shield cat# 1114547). Образец центрифугировали при RT (22°C, 800g) на раскачивающейся центрифуге в течение 30 минут, промывали средой RPMI1640 и центрифугировали (RT, 250 g) в течение 10 минут. Считали количество клеток и высеивали их в конечной концентрации 1,5Х106 клеток/мл питательной среды (RPMI1640+10%FBS+2 мМоль L-глутамина + 1% Pen-Strep) и инкубировали в течение 1 часа при 37°С перед обработкой siРНК.
[0495] Затем клетки обрабатывали исследуемыми siРНК в различных концентрациях с использованием реагента Lipofectamine™2000 (Invitrogen) согласно инструкциям производителя и инкубировали при 37°С в инкубаторе при 5% CO2 в течение 24 часов.
[0496] В качестве положительного контроля ответа IFN клетки обрабатывали поли(I:С), синтетическим аналогом двухцепочечной РНК (dsPHK), который является лигандом TLR3 (InvivoGen Cat# tlrl-pic) в конечных концентрациях 0,25-5,0 мкг/мл или тиазолхинолоном (CLO75), лигандом TLR 7/8 (InvivoGen Cat# tlrl-c75) в конечных концентрациях 0,075-2 мкг/мл. Клетки, обработанные реагентом Lipofectamine™2000, использовали в качестве отрицательного (референс) контроля IFN ответа.
[0497] Приблизительно через 24 часа после инкубации собирали клетки и переносили супернатант в новые пробирки. Незамедлительно замораживали образцы в жидком азоте и проверяли секрецию цитокинов IL-6 и TNF-α с использованием набора IL-6, DuoSet ELISA (R&D System DY2060) и TNF-α, DuoSet ELISA kit (R&D System DY210) согласно инструкции производителя. РНК выделяли из осадка клеток и методом количественной ПЦР определяли уровни мРНК генов IFIT1 (индуцированный интерфероном белок с тетратрикопептидными повторами 1) и МХ1 (устойчивость к миксовирусам (вирус гриппа) 1, интерферон-индуцибельный белок р78) человека. Измеренные количества мРНК нормировали относительно количества мРНК контрольного гена пептидилпролилизомеразы А (циклофилин A; CycloA). Оценивали индукцию сигнальных путей IFN посредством сравнения количества мРНК от генов IFIT1 и МХ1 из обработанных клеток с их количествами в необработанных клетках. За результаты количественной ПЦР принимали те, которые проходили стандарты QC, т.е. величина уклона стандартной кривой находилась в пределах [-4, -3], R2>0.99, без димеров праймеров. Результаты, которые не отвечали требованиям QC, исключали из анализа.
[0498] В Таблице 6 показаны соединения siSERPINH1. Данные о стабильности и активности некоторых соединений представлены в Таблице 6. Код структур смысловой и антисмысловой цепи представлены ниже в Таблице 7.
[0499] Олигонуклеотиды siРНК, которые являются полезными для создания молекул двухцепочечных РНК, описаны ниже в Таблицах А-18, А-19 В-Е.
Олигонуклеотидные последовательности SERPINHL полезные для приготовления соединений siРНК
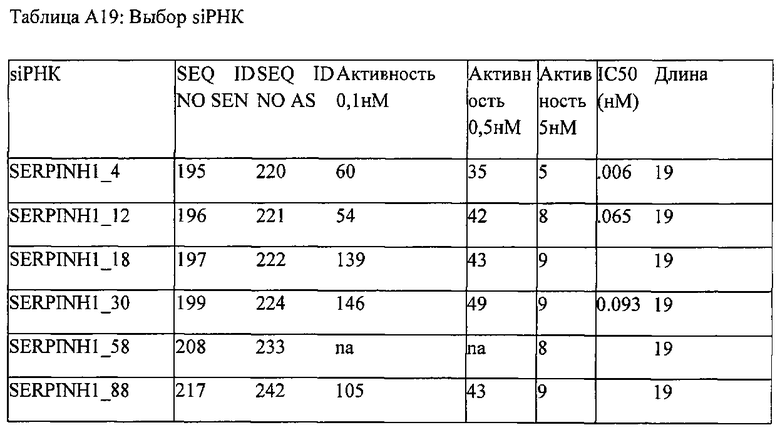
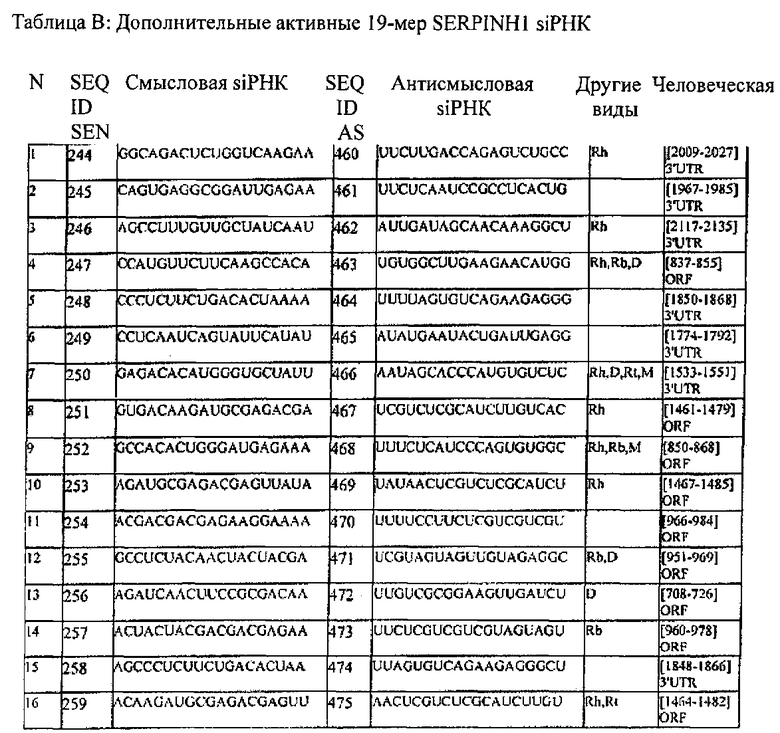
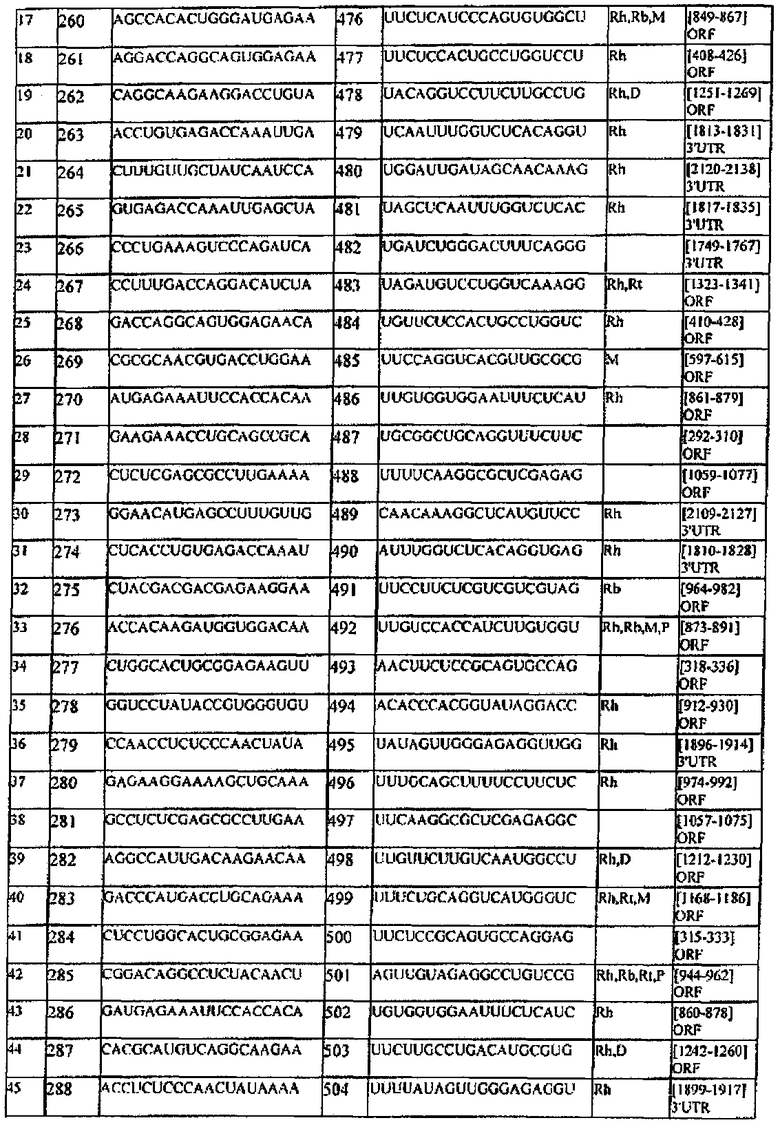
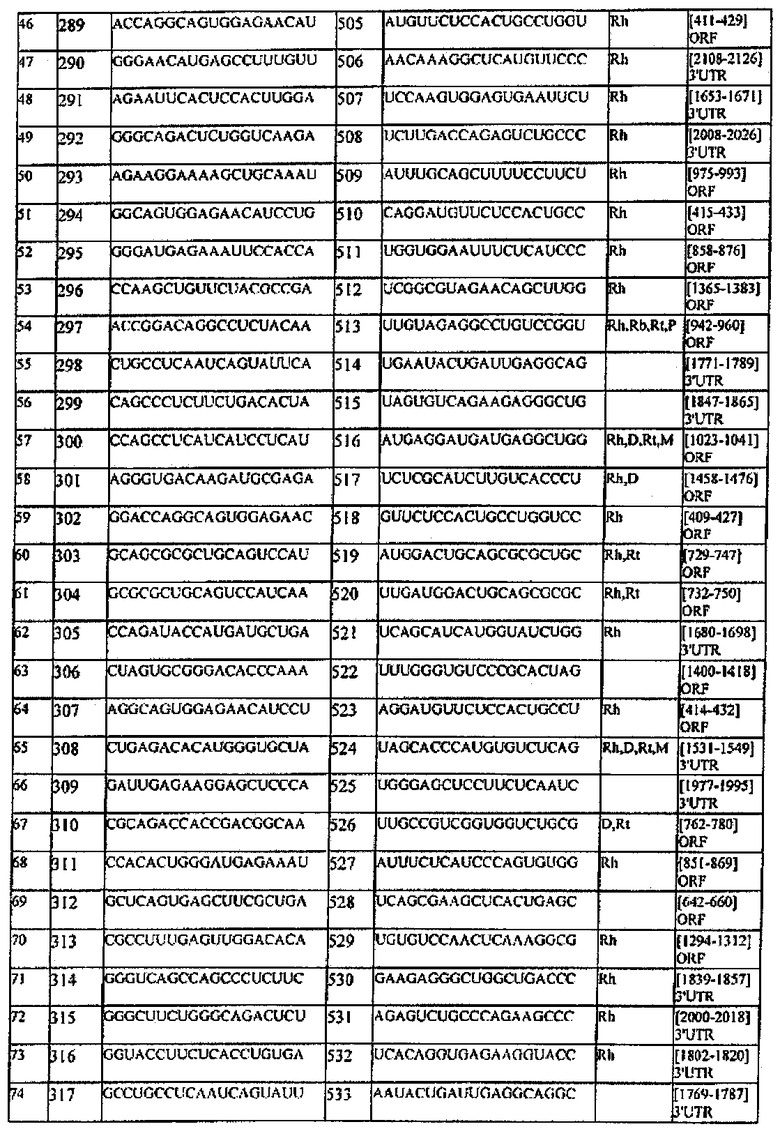
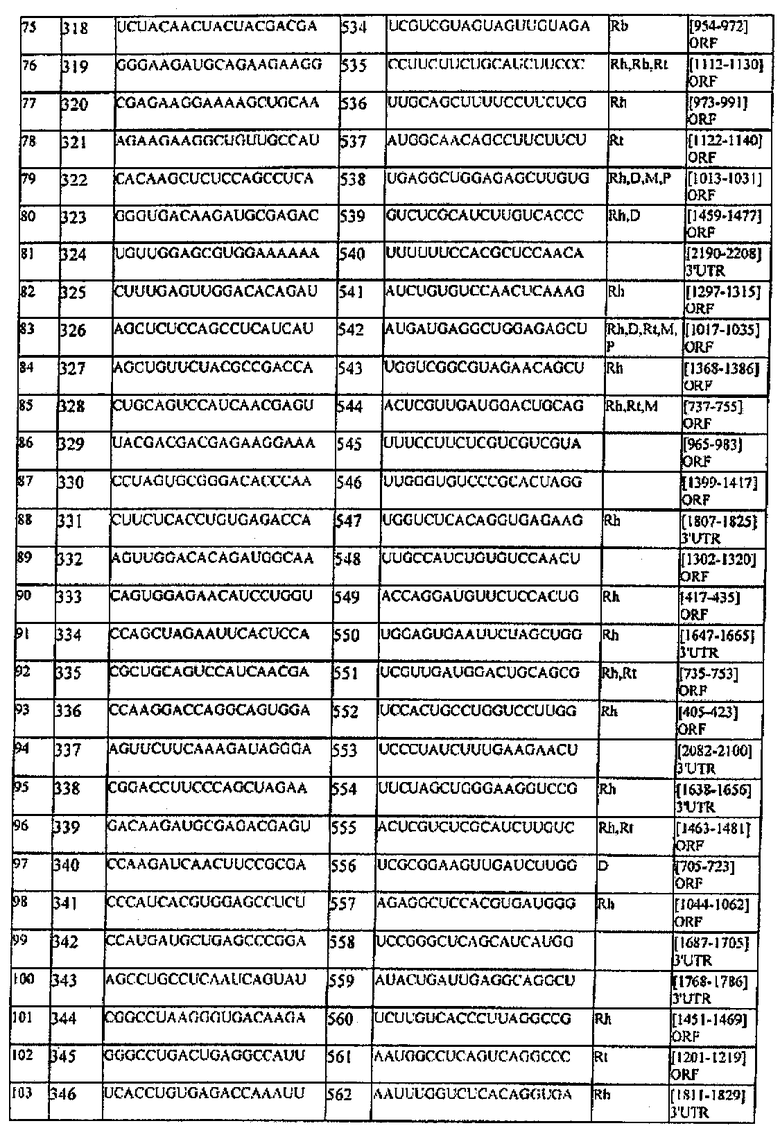
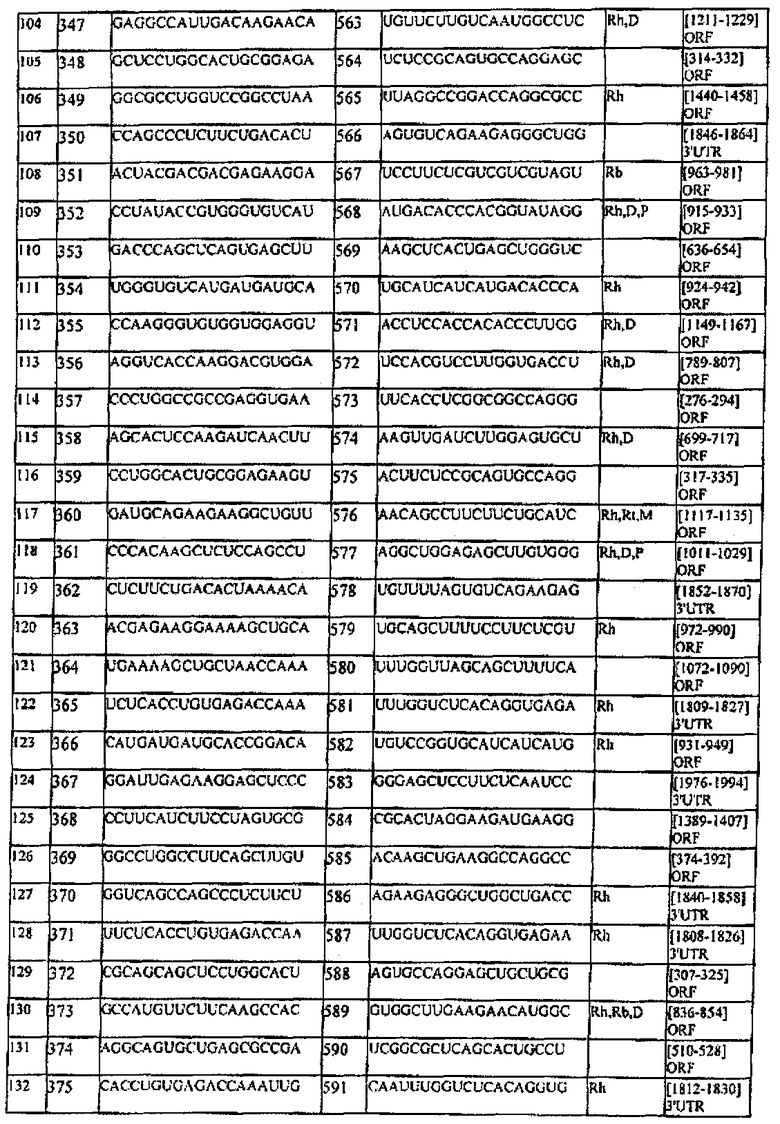

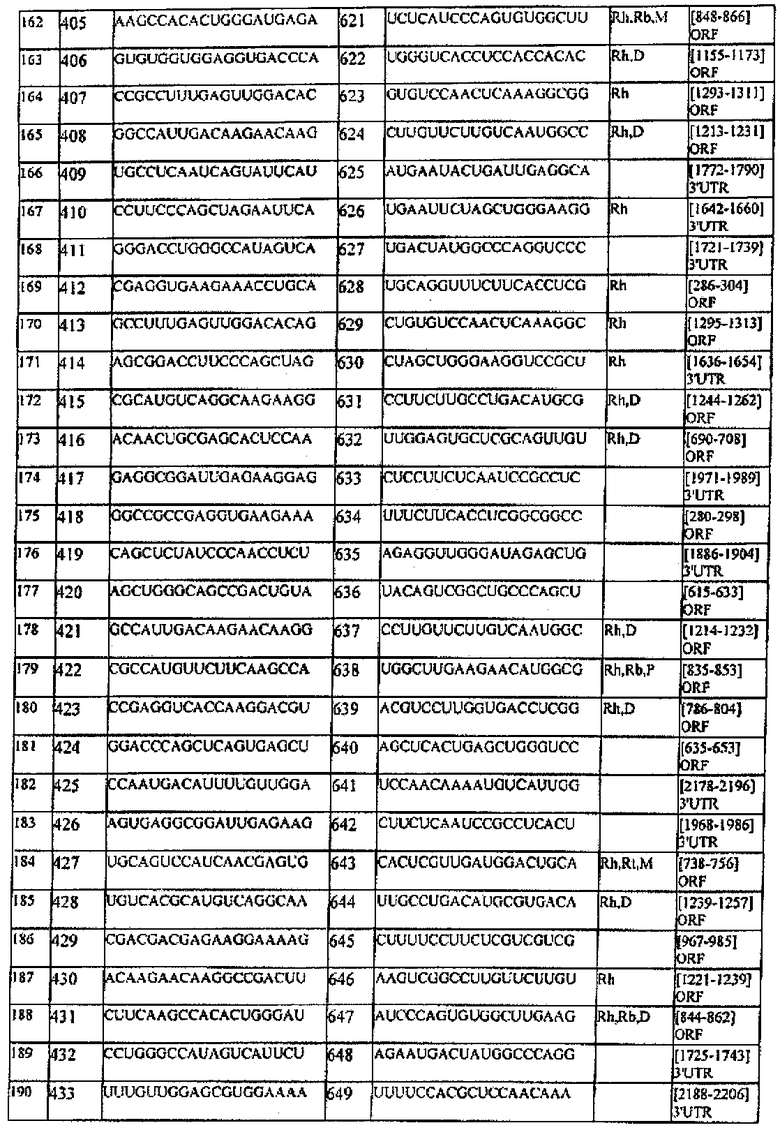
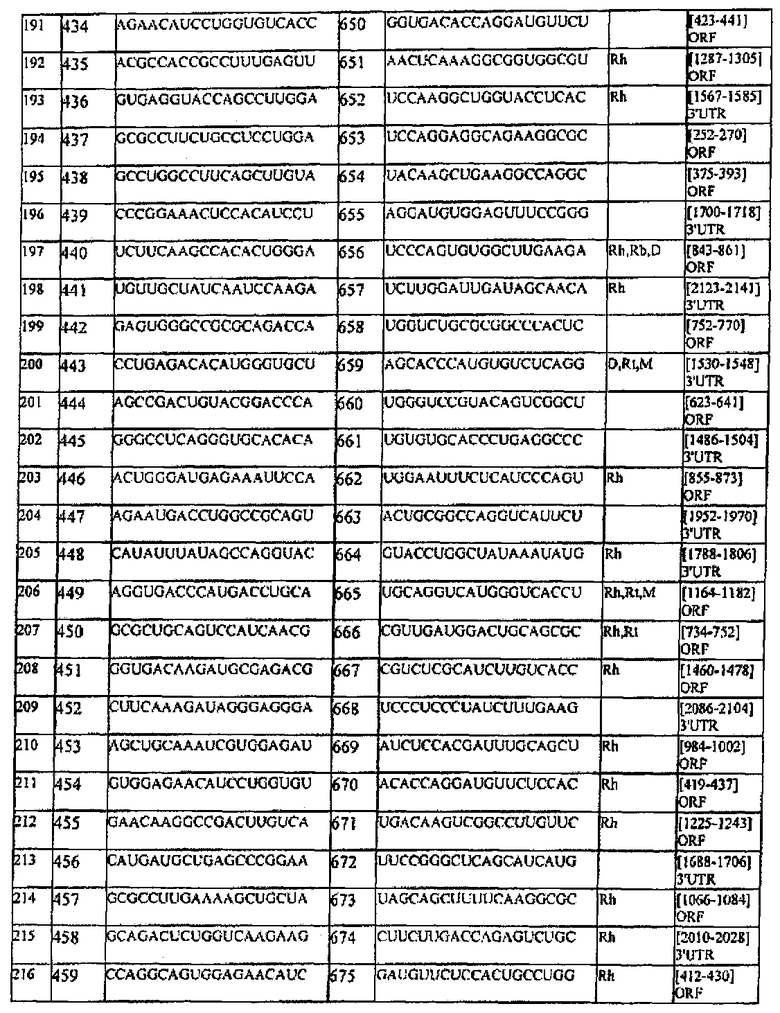
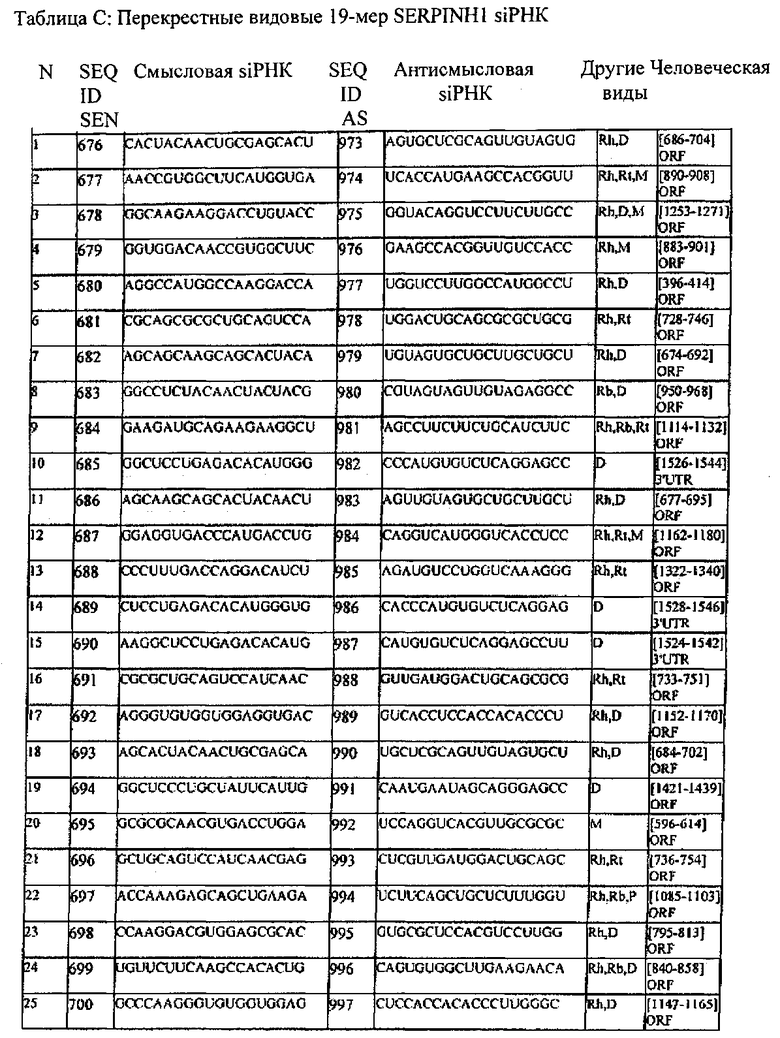
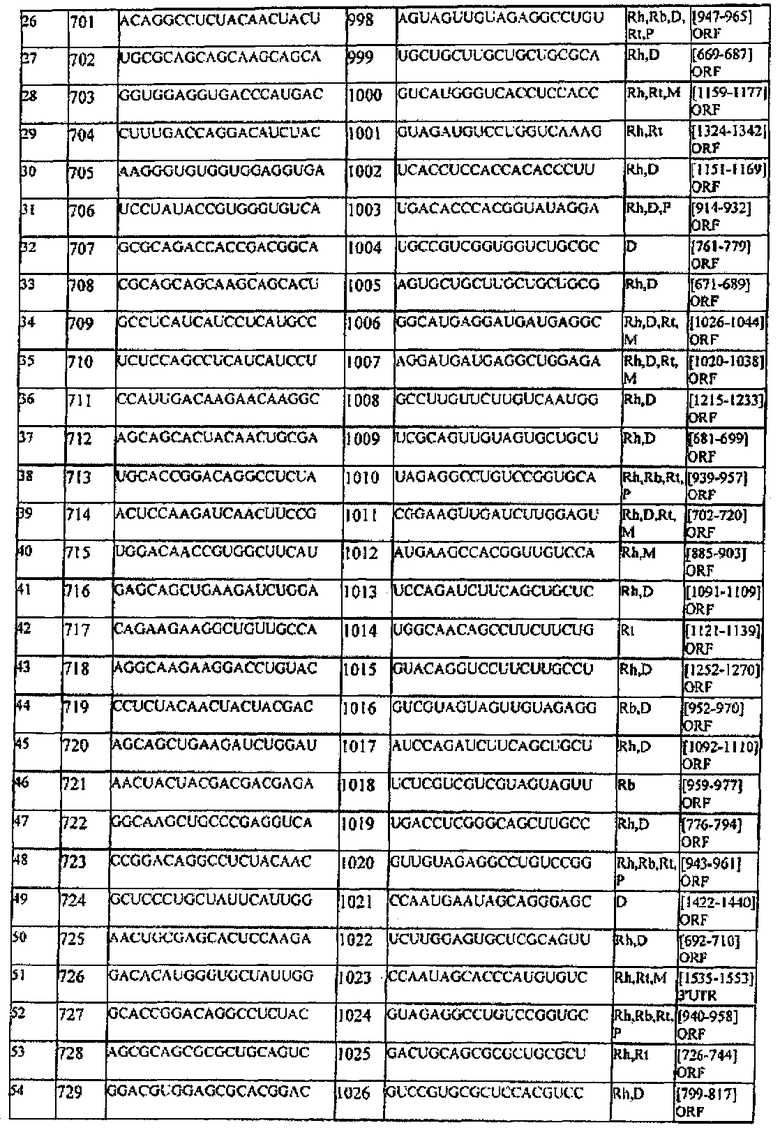
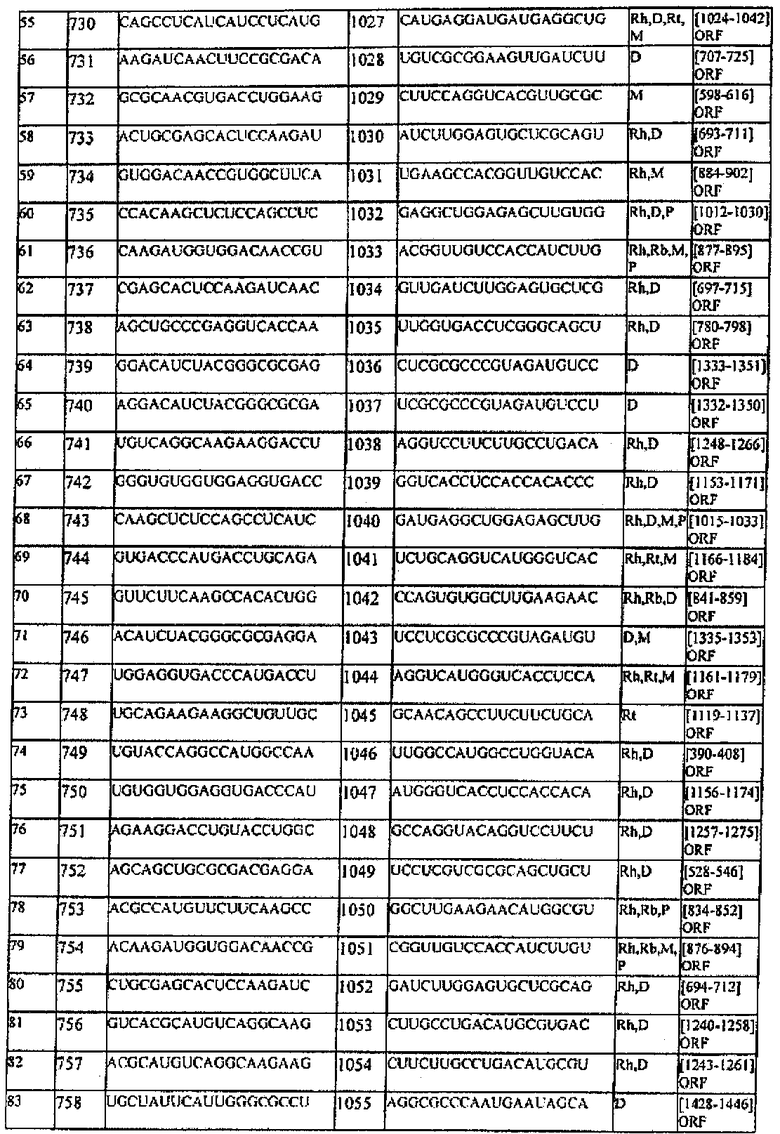
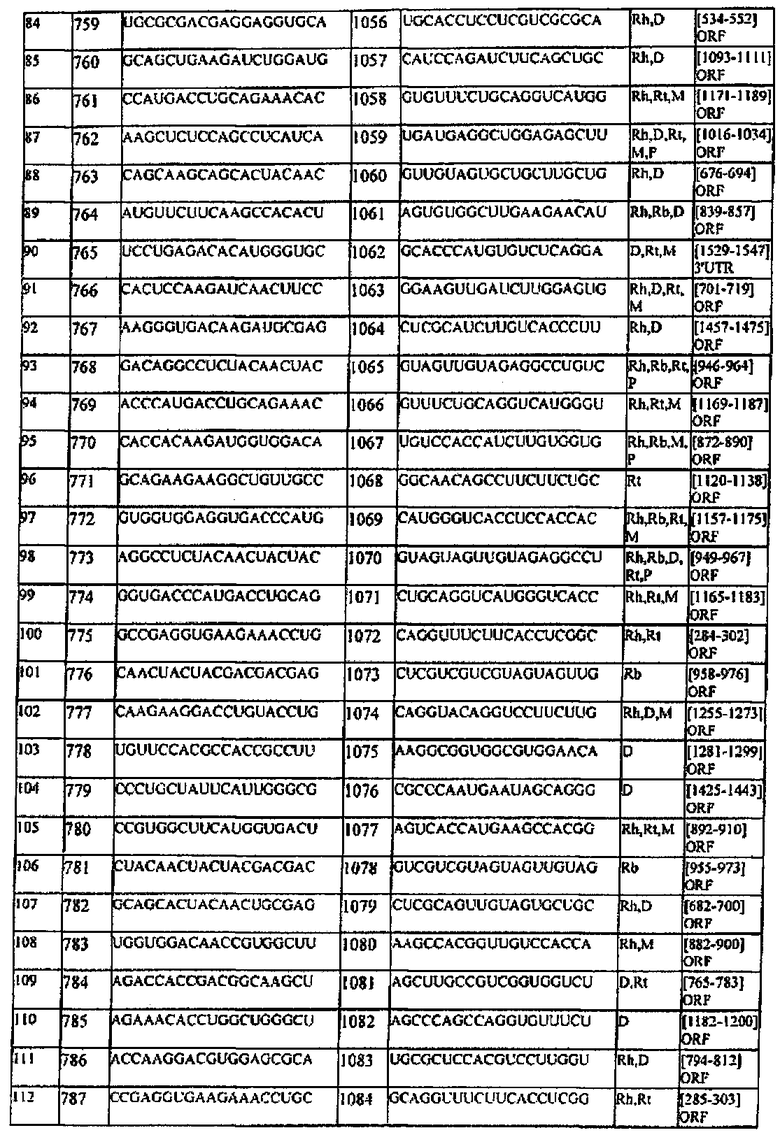
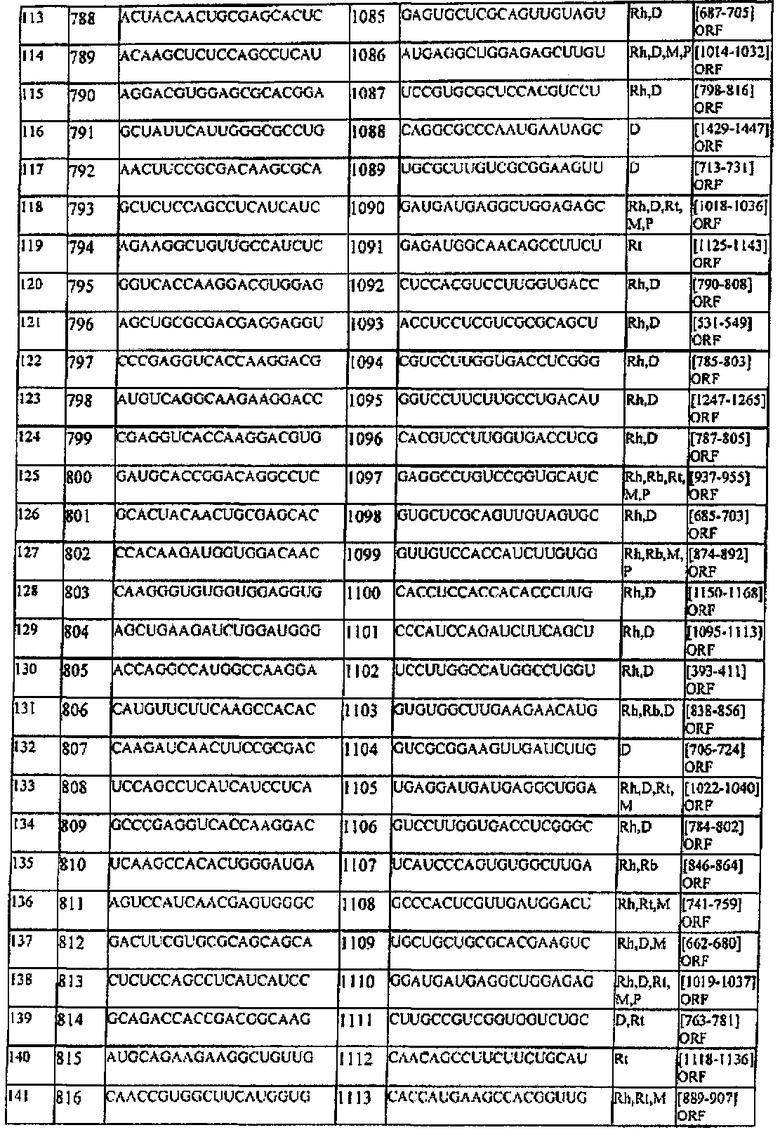

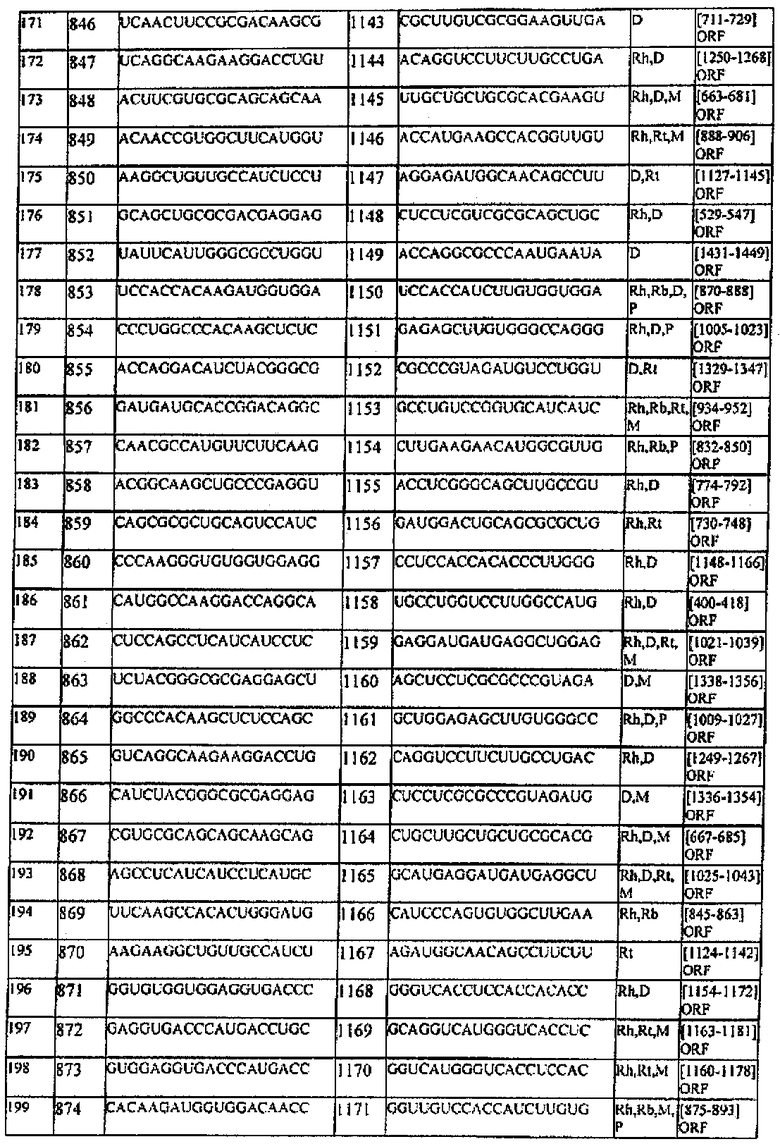
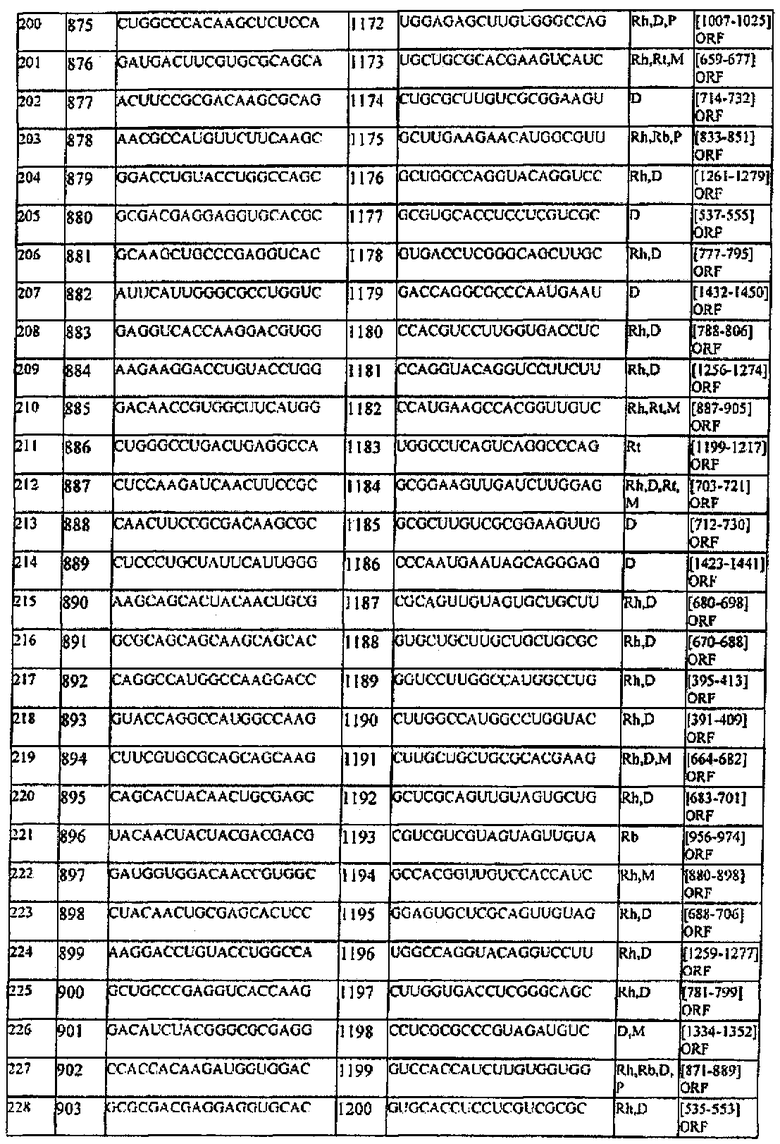
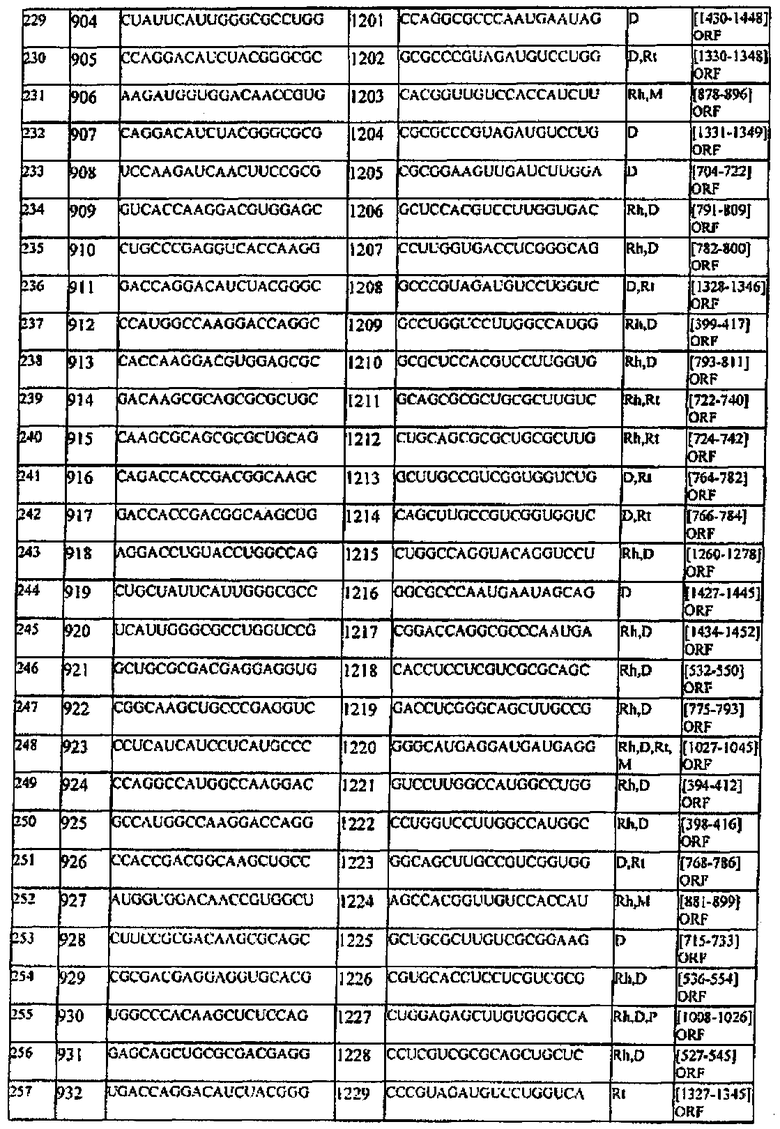
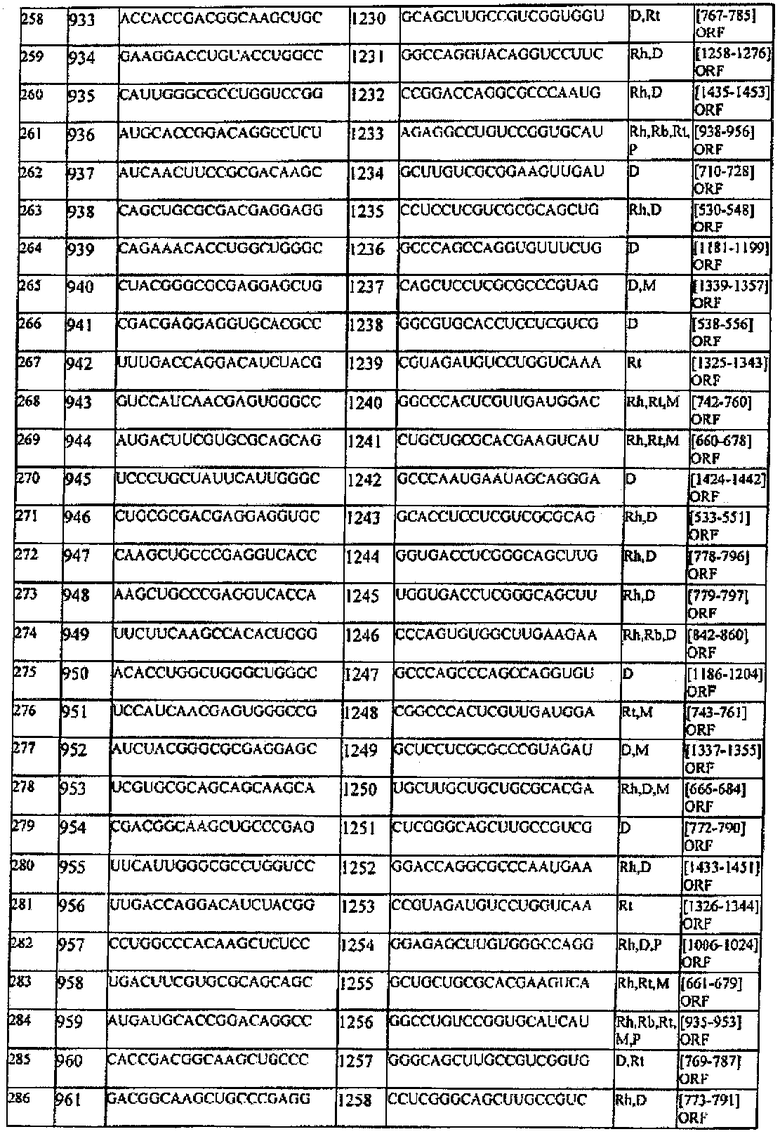

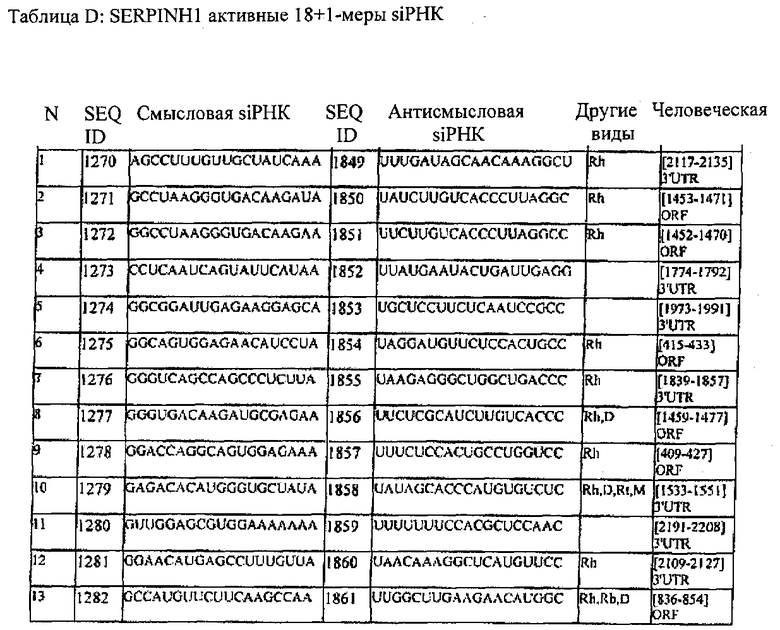
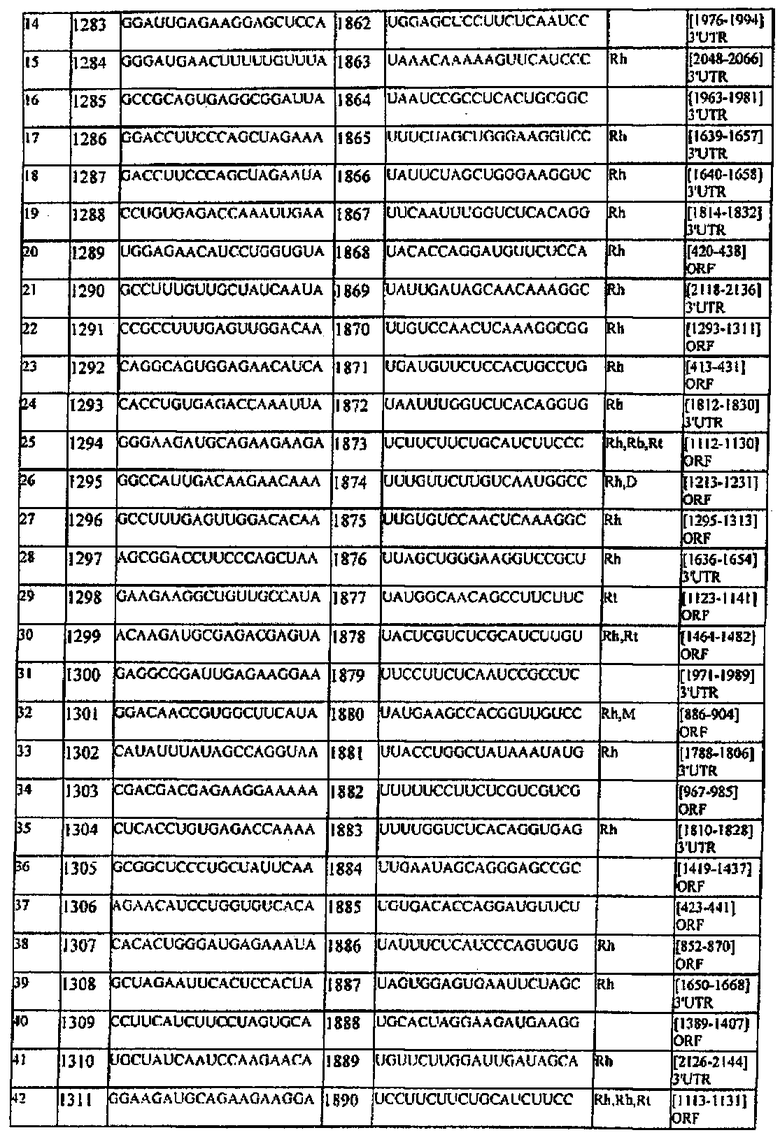
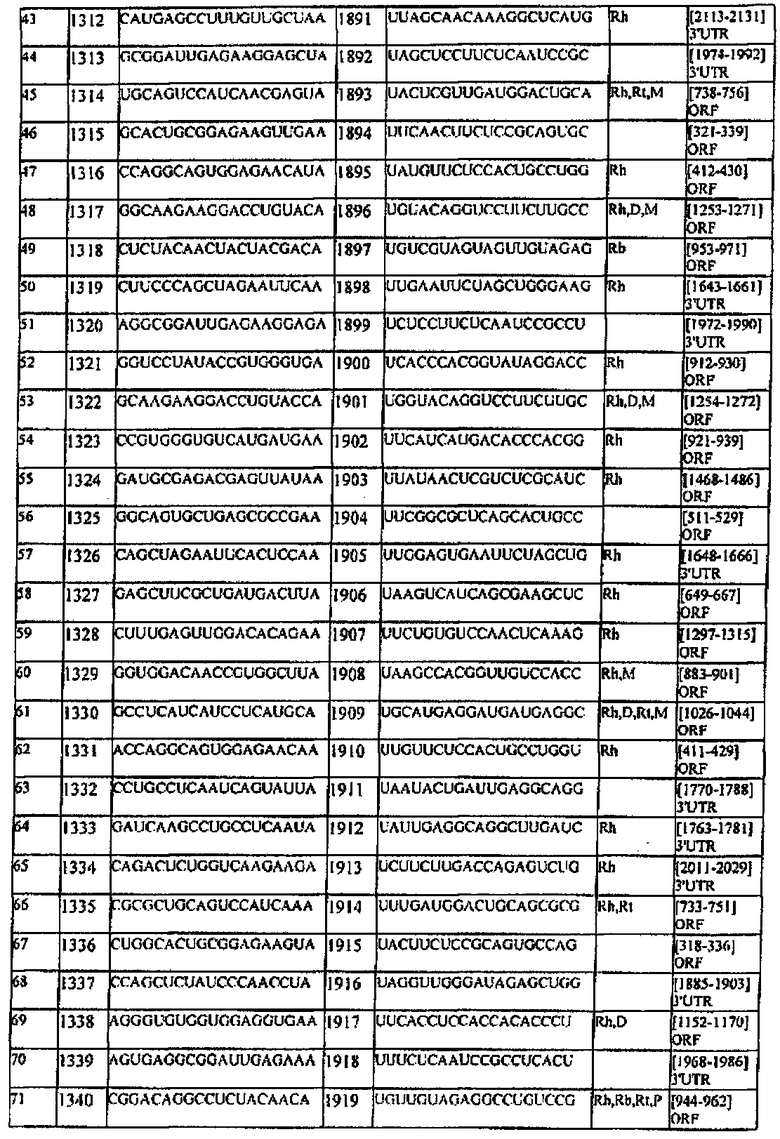
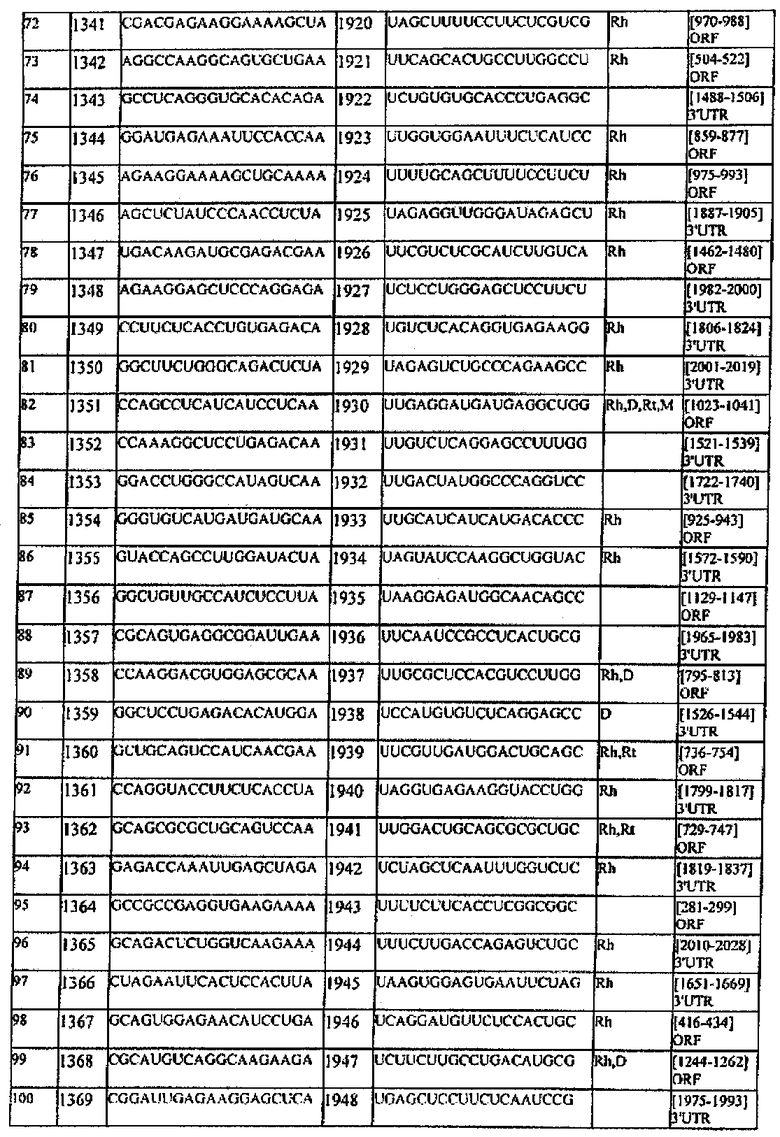
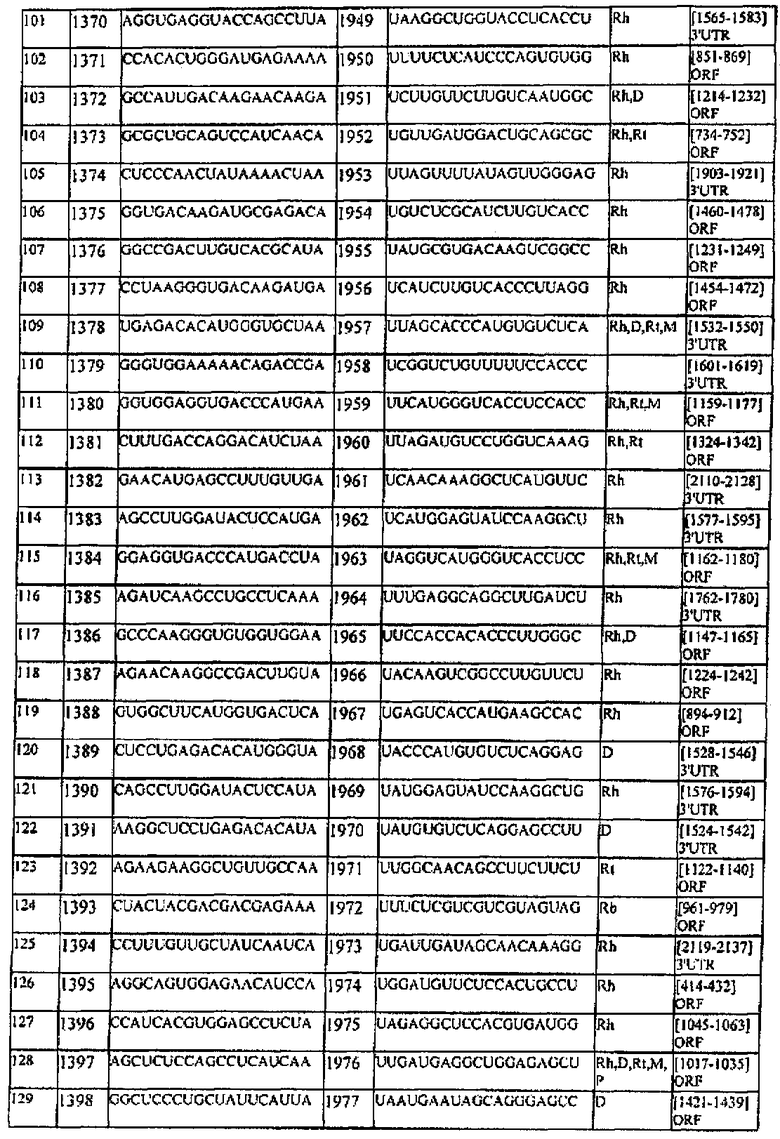
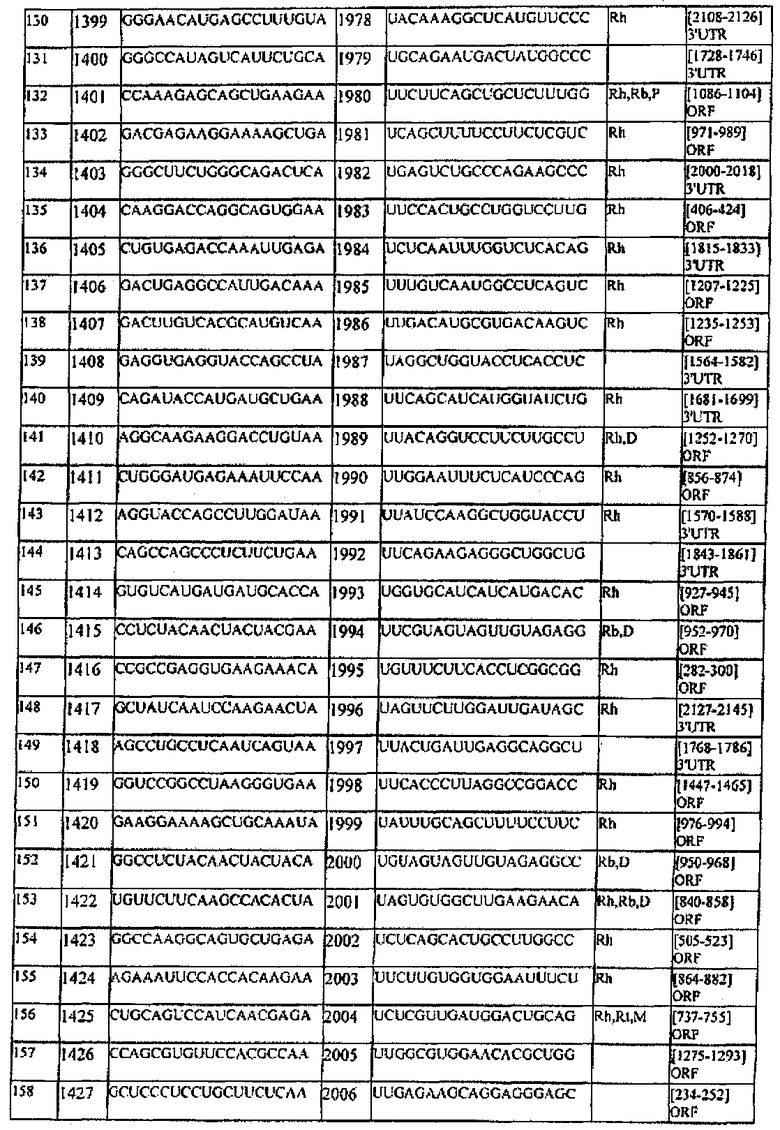
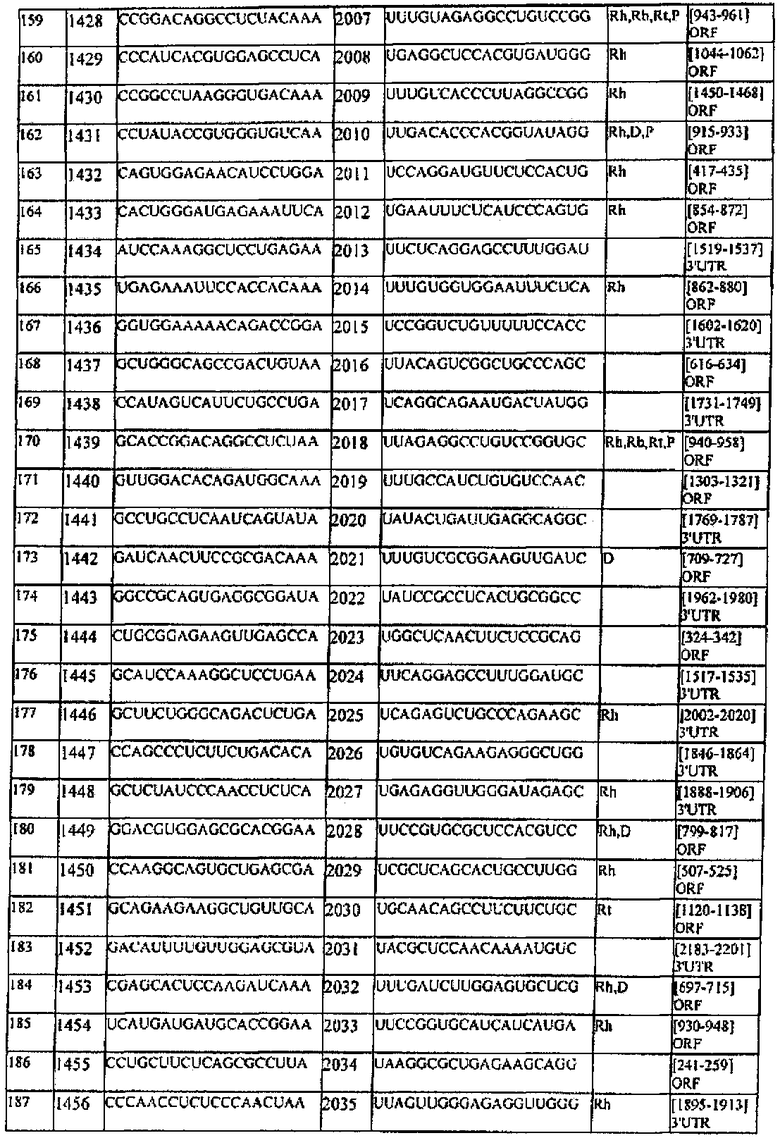
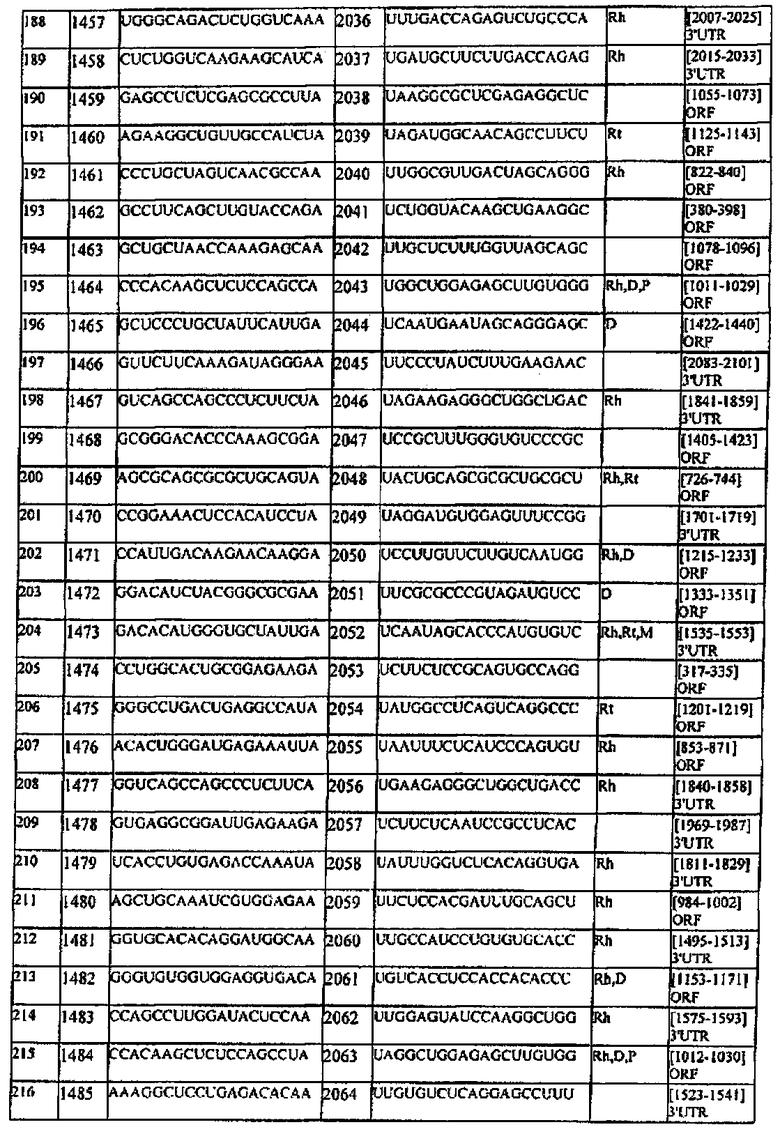
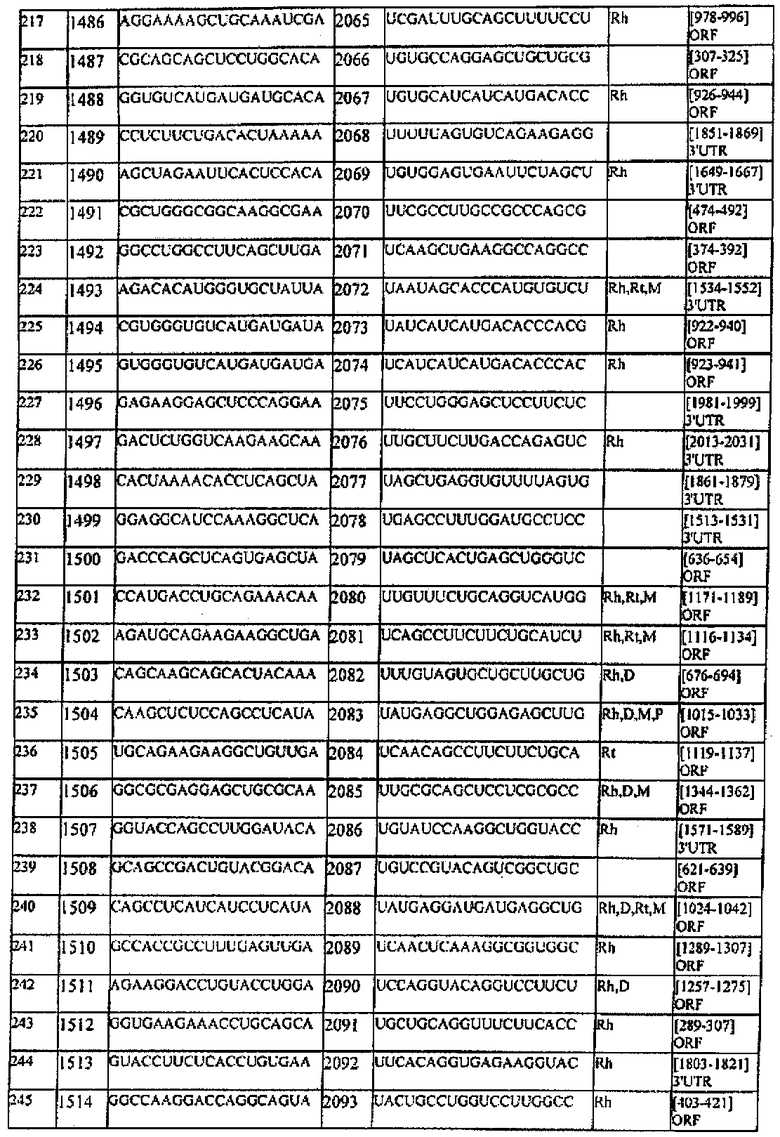
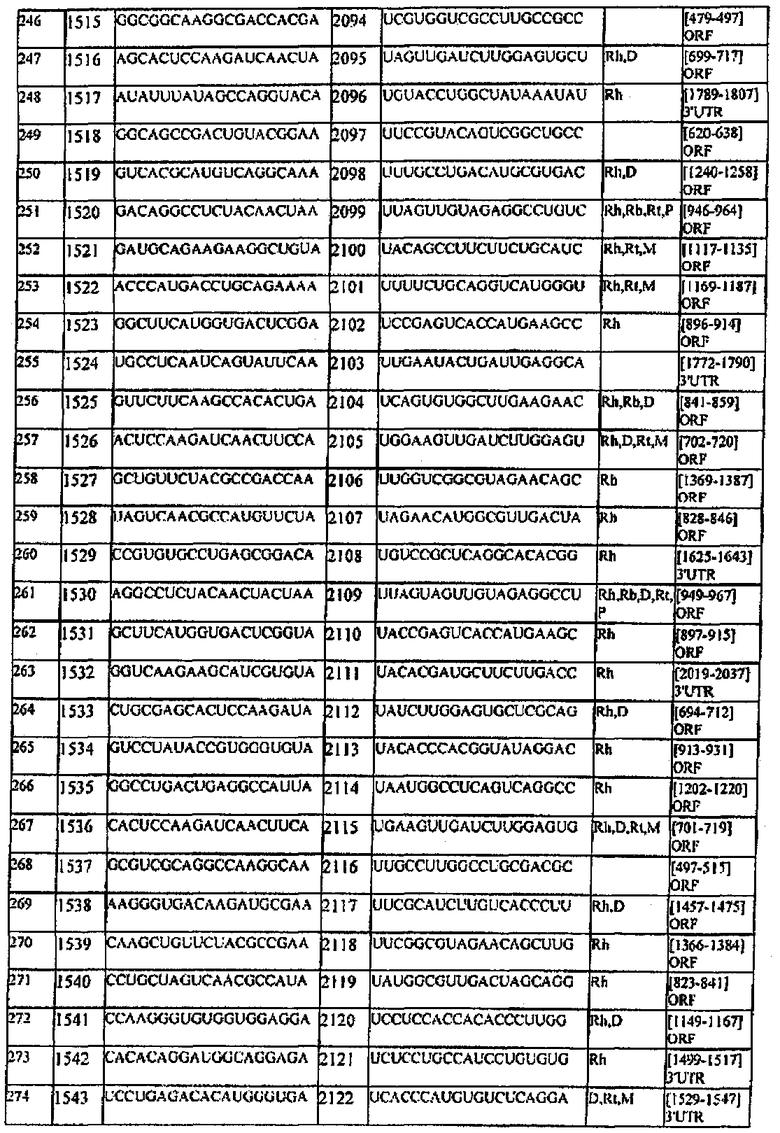
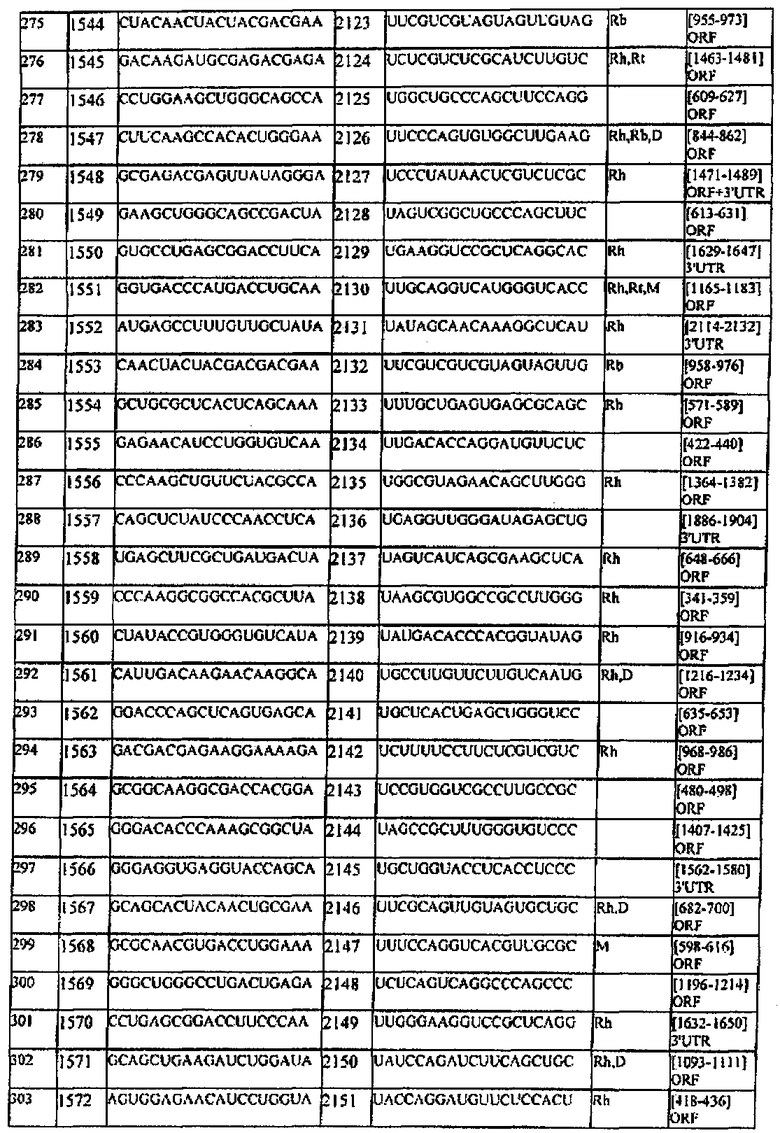
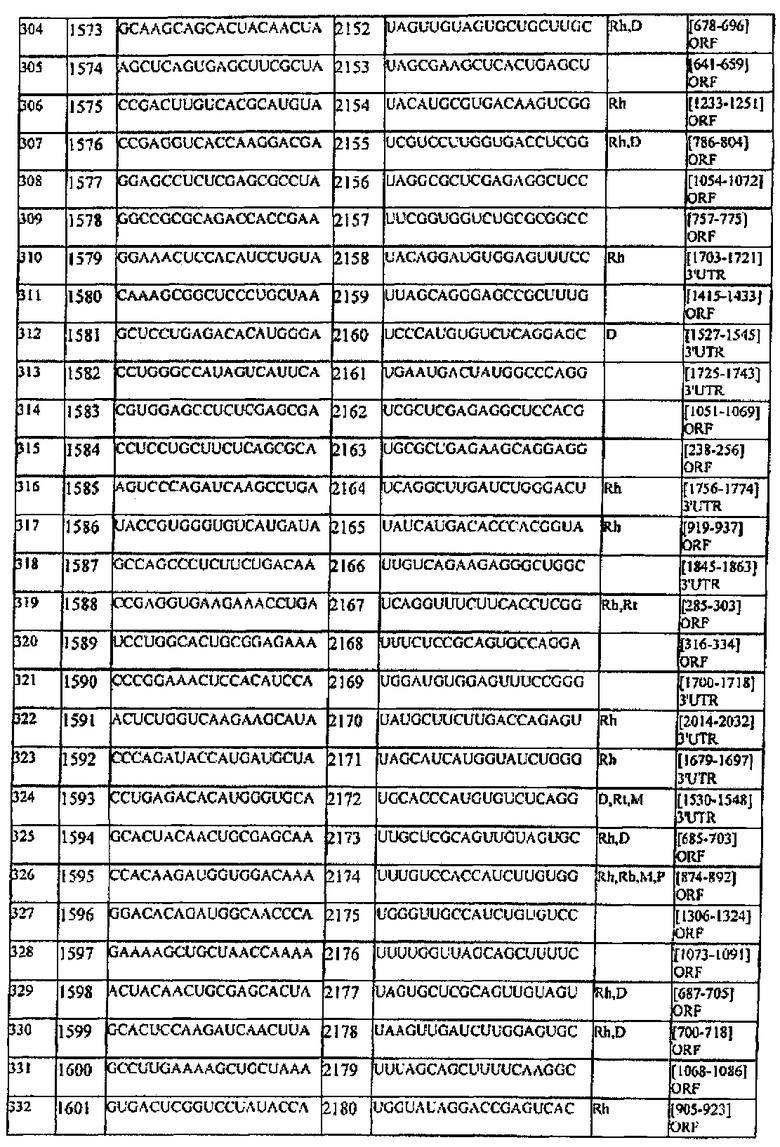
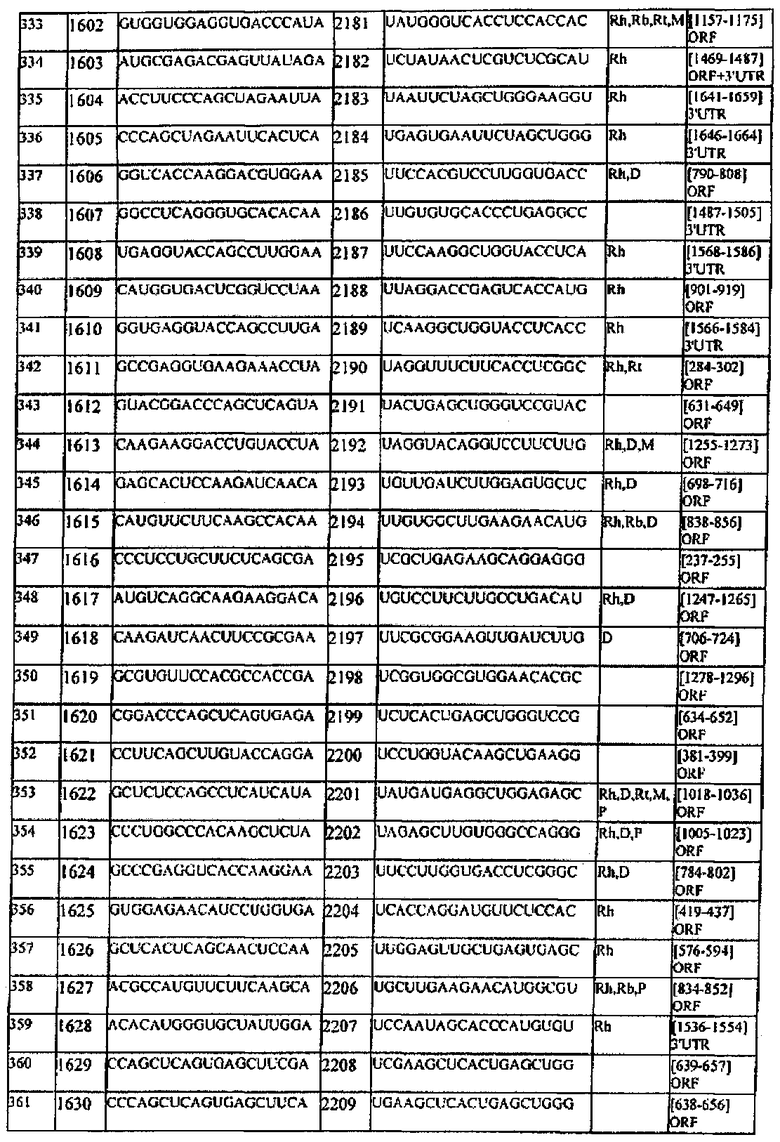
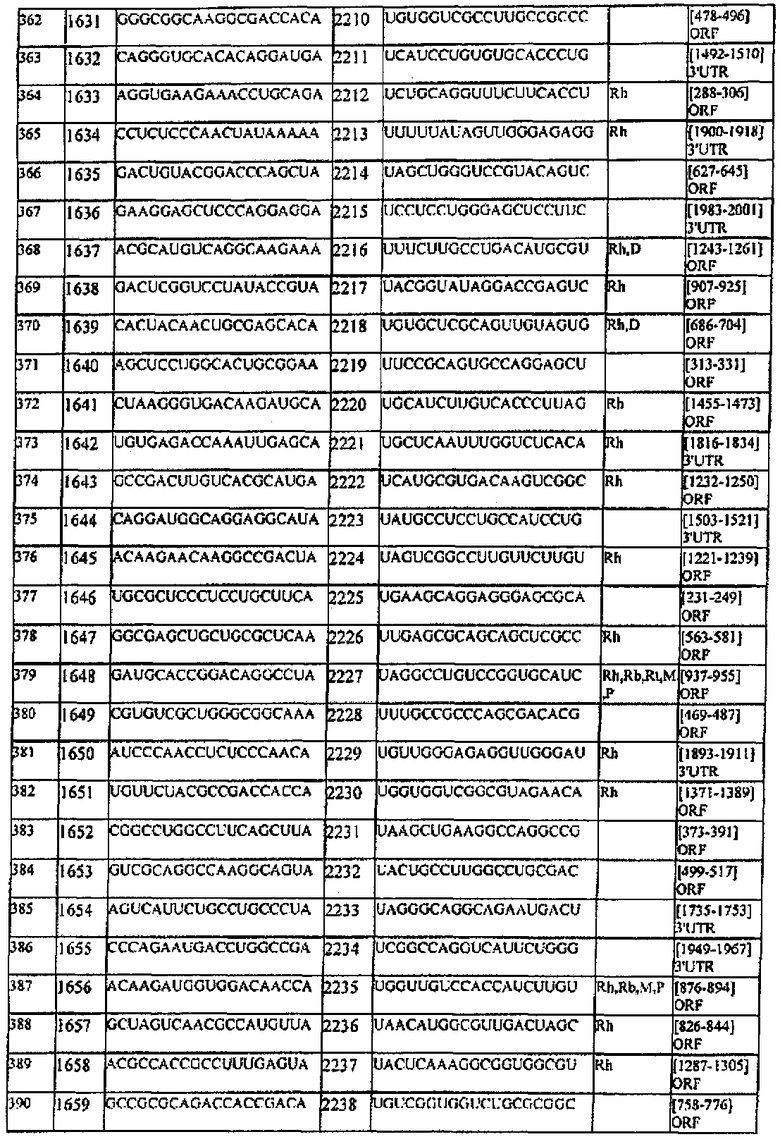
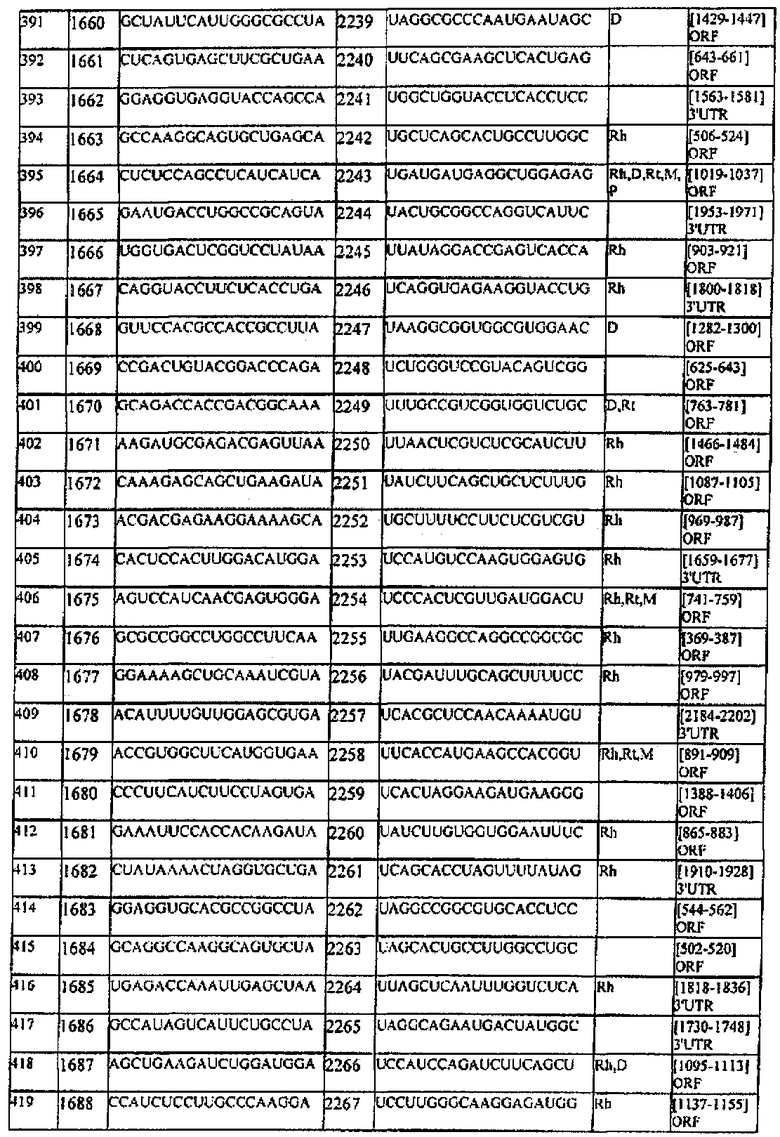
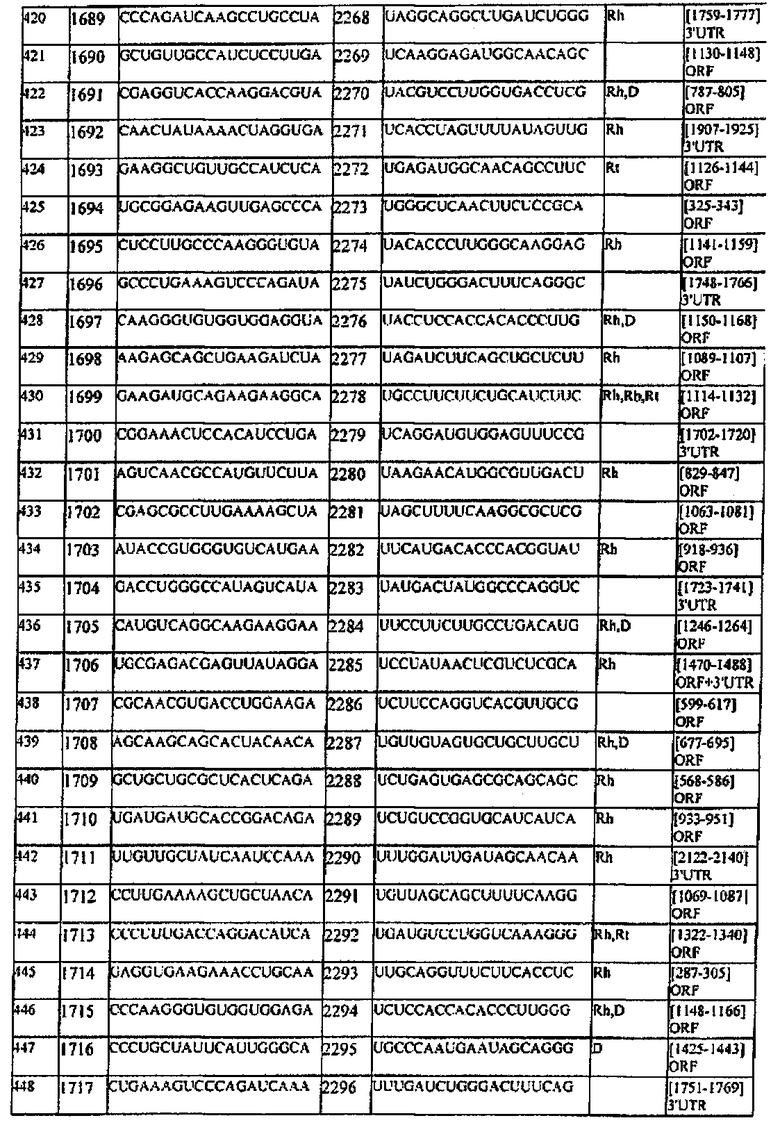
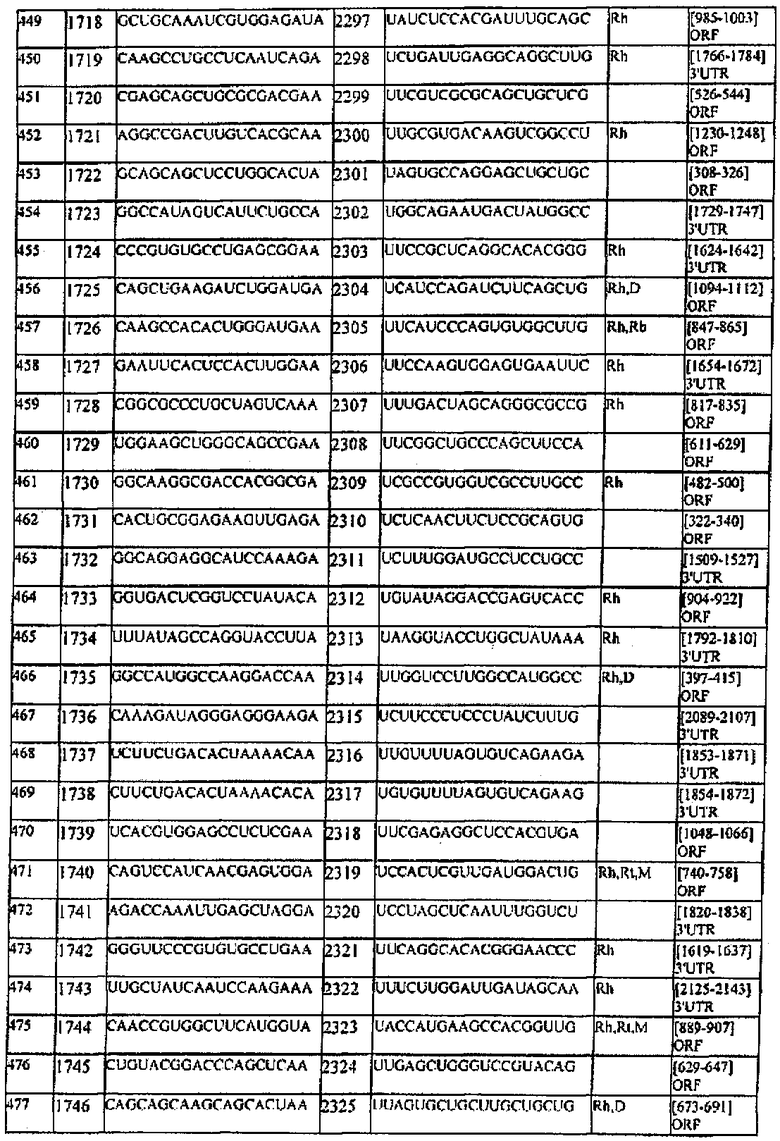
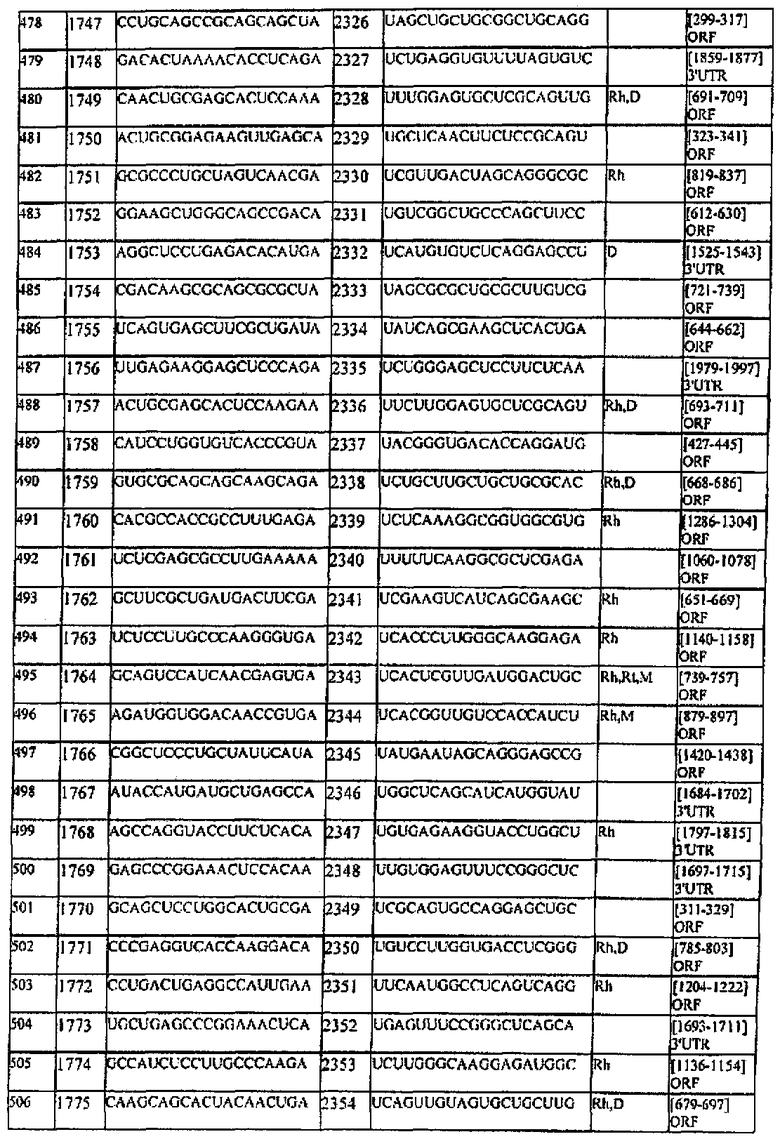
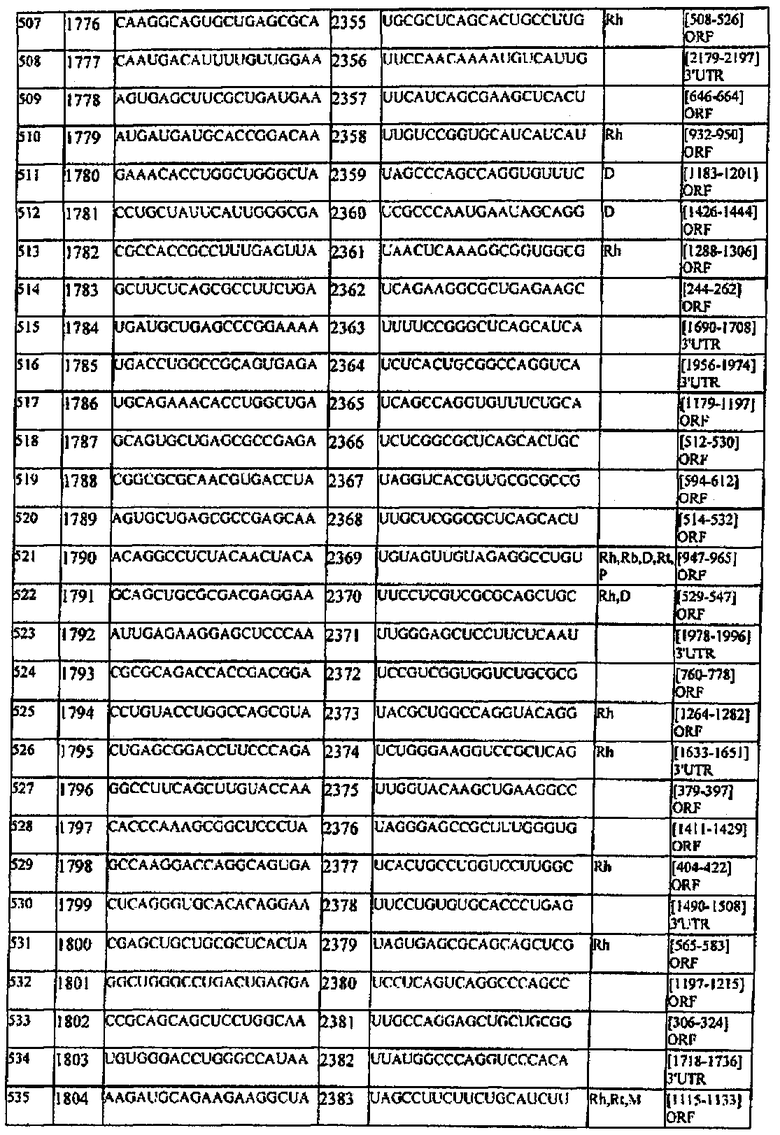
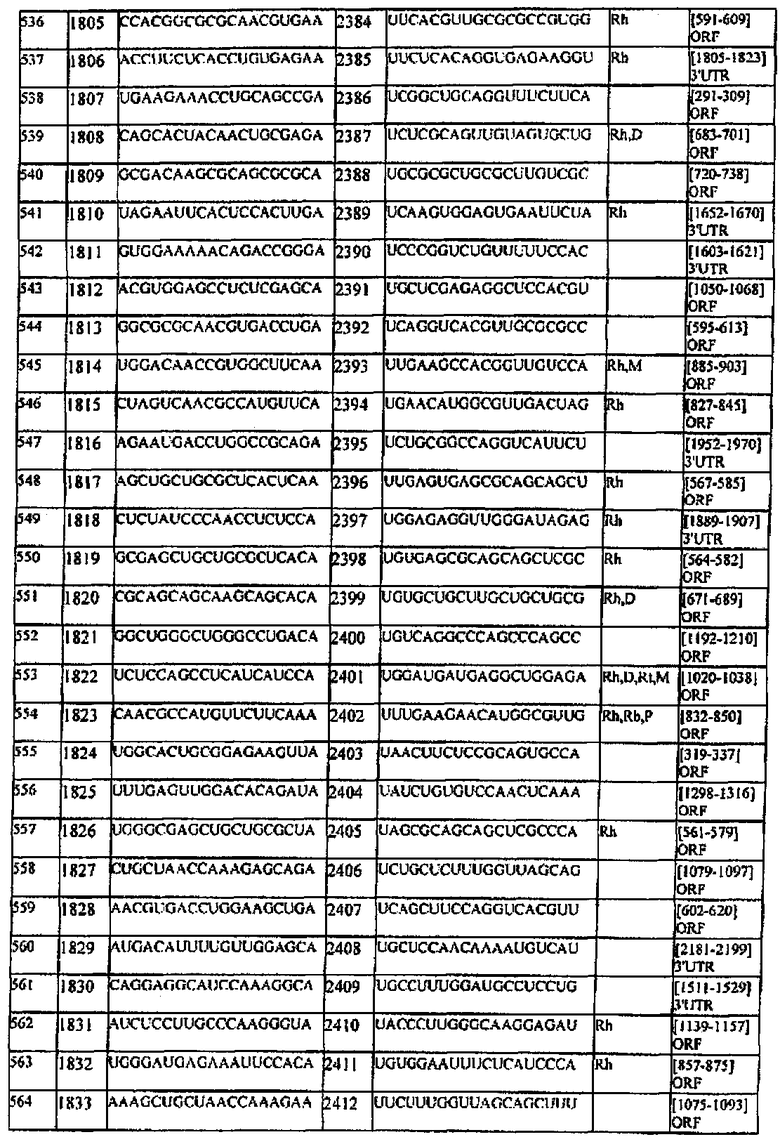
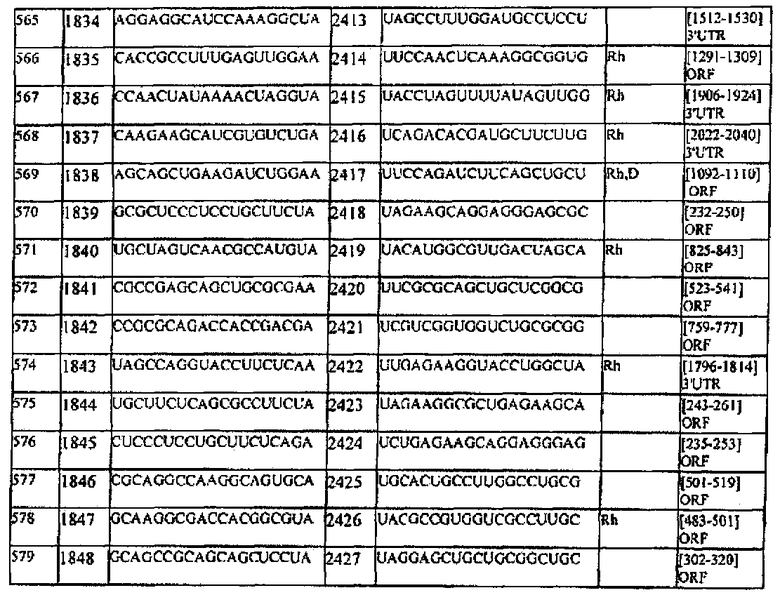

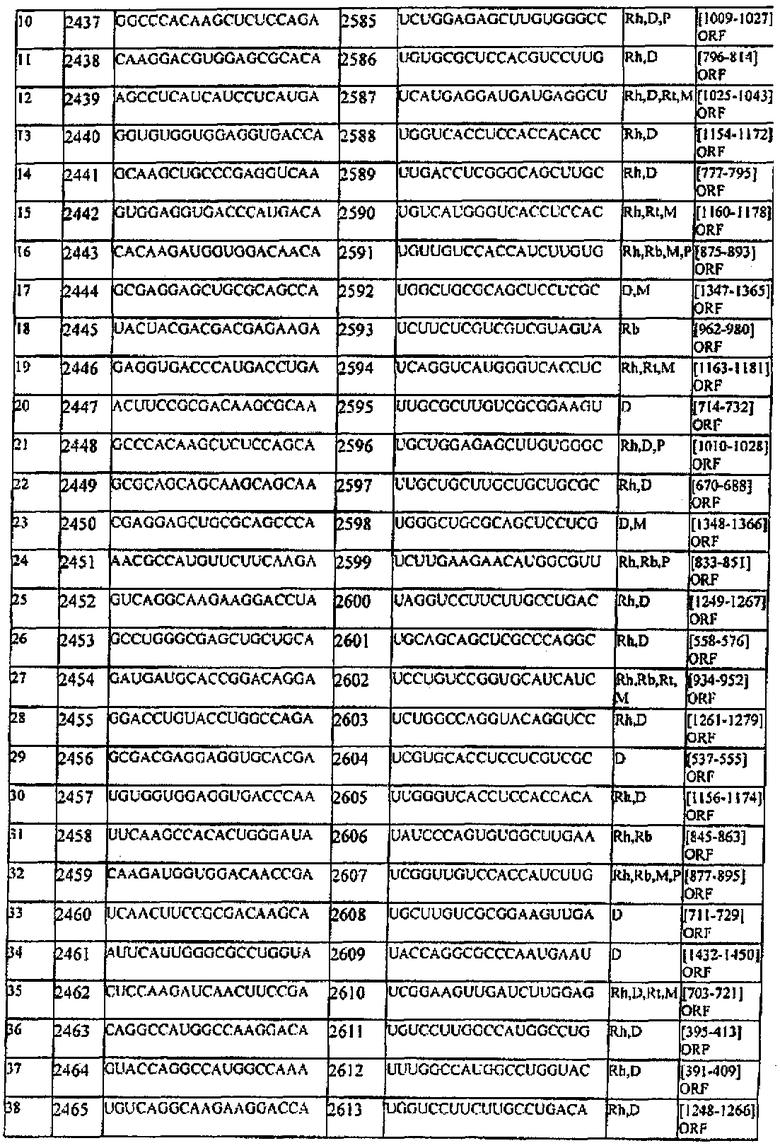

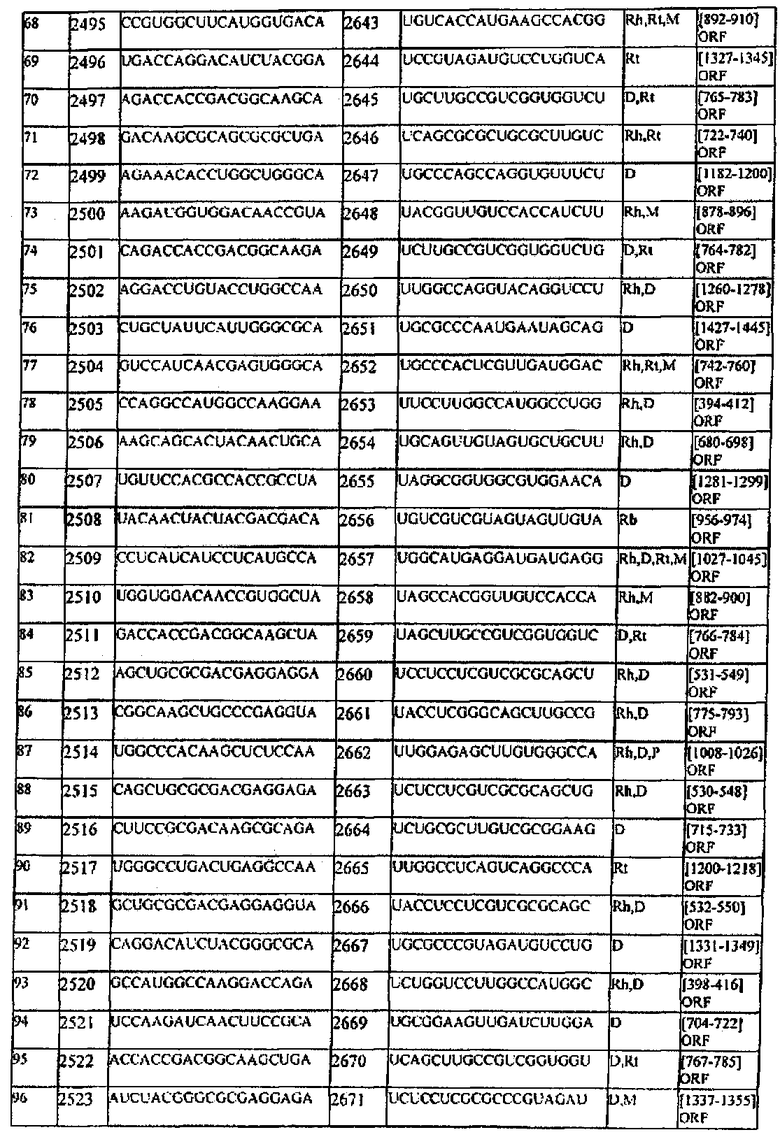
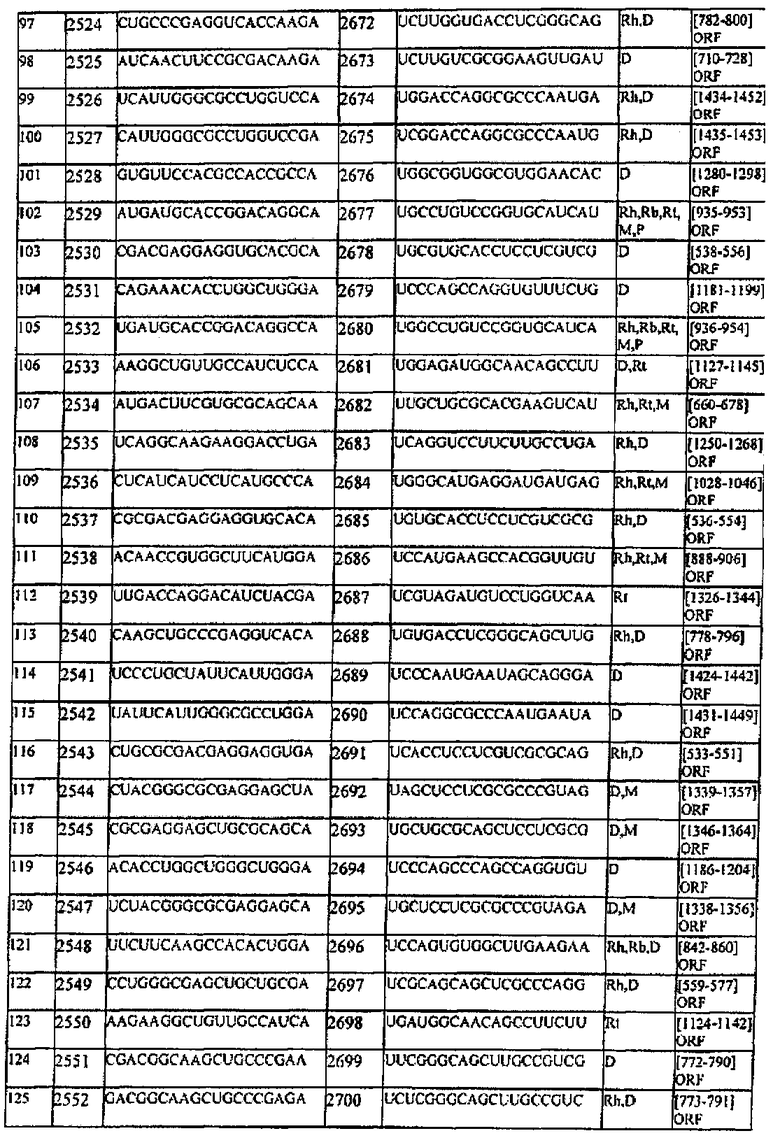
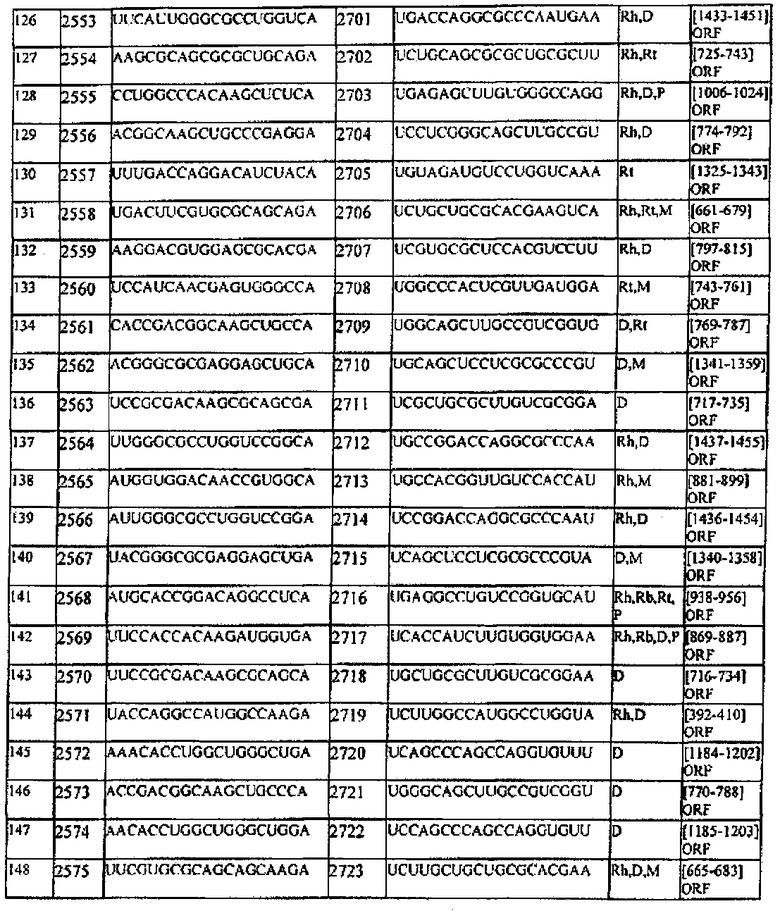
ПРИМЕР 10:
Животные модели
Модельные системы фиброзных заболеваний
[0500] Анализ активных siPHK согласно настоящему изобретению можно было выполнять на предиктивных животных моделях. Крысиные диабетические и стареющие модели фиброза почек включали крыс Цукера, страдающих ожирением и диабетом (ZDF), старых fa/fa крыс Цукера с ожирением, старых мышей Спрейг-Доули (SD) и крыс Гото Какизаки (GK); крысы GK представляют собой инбредный штамм, полученный от крыс линии Вистар, отобранных по спонтанному развитию NIDDM (диабета типа II). Индуцированные модели фиброза почек включали модель перманентной односторонней обструкции мочеточника (UUO), которая представляет собой модель острого фиброза кишечника, встречающегося у здоровых не диабетических животных; фиброз почек развивается черехз несколько дней после обструкции. Другой индуцированной моделью фиброза почек является нефреэктомия на 5/6.
[0501] Две модели фиброза печени на крысах представляли собой Перевязку желчных протоков (BDL) с имитацией операции в качестве контролей и отравление СС14 с животными, содержавшихся на диете из оливкового масла, в качестве контролей, как описано в следующих публикациях: Lotersztajn S, et al Hepatic Fibrosis: Molecular Mechanisms and Drug Targets. Annu Rev Pharmacol Toxicol. 2004 Oct 07; Uchio K, et al., Down-regulation of connective tissue growth factor and type I collagen мРНК expression by connective tissue growth factor antisense oligonucleotide during experimental liver fibrosis. Wound Repair Regen. 2004 Jan-Feb; 12(1):60-6; Xu XQ, et al., Molecular classification of liver fibrosis in a rat model by proteomics and bioinformatics Proteomics. 2004 Oct; 4(10):3235-45.
[0502] Модели образования шрамов на глазах хорошо известны в данной области техники, например, в публикации Sherwood MB et al., J Glaucoma. 2004 Oct; 13(5):407-12. A new model of glaucoma filtering surgery in the rat; Miller MH et al., Ophthalmic Surg. 1989 May; 20(5):350-7. Wound healing in an animal model of glaucoma fistulizing surgery in the Rb; vanBockxmeer FM et al., Retina. 1985 Fall-Winter; 5(4): 239-52. Models for assessing scar tissue inhibitors; Wiedemann Ρ et al., J Pharmacol Methods. 1984 Aug; 12(1): 69-78. Proliferative vitreoretinopathy: the Rb cell injection model for screening of antiproliferative drugs.
[0503] Модели катаракт описываются в следующих публикациях: The role of Src family kinases in cortical cataract formation. Zhou J, Menko AS.Invest Ophthalmol Vis Sci. 2002 Jul; 43(7):2293-300; Bioavailability and anticataract effects of a topical ocular drug delivery system containing disulfiram and hydrохупропил-beta-cyclodextrin on selenite-treated rats. Wang S, et al. http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=15370367 Curr Eye Res. 2004 Jul; 29(1):51-8; and Long-term organ culture system to study the effects of UV-A irradiation on lens transglutaminase. Weinreb O, Dovrat Α.; Curr Eye Res. 2004 Jul; 29(1):51-8.
[0504] Соединения Таблицы A-18 и Таблицы A-19 исследовали на указанных моделях фиброзных заболеваний, на которых обнаружили, что они являются эффективными при лечении фиброза печени и других фиброзных заболеваниях.
Модельные системы глаукомы
[0505] Исследование активных siPHK согласно настоящему изобретению для лечения или профилактики глаукомы проводили на крысиных животных моделях повреждения зрительного нерва, описанных, например, в публикациях: Maeda, K. et al., "A Novel
Neuroprotectant against Retinal Ganglion Cell Damage in a Glaucoma Model and an Optic Nerve Crush Model in the rat". Investigative Ophthalmology and visual Science (IOVS), March 2004, 45(3)851. В частности, для поперечного разреза зрительного нерва обнажали глазничную часть зрительного нерва (ON) крыс под анастезией с заходом через надглазничную часть, разрывали мягкие оболочки мозга и все аксоны в ON рассекали с помощью шипцов в течение 10 секунд, 2 мм от решетчатой пластинки.
[0506] Молекулы нуклеиновых кислот согласно настоящему описанию исследовали на данной животной модели, и результаты показали, что указанные соединения siРНК являются полезными при лечении и/или предотвращении глаукомы.
Крысиная модель повреждения зрительно нерва (ONC): интравитриальная доставка siРНК и доставка в виде глазных капель
[0507] Для поперечного разреза зрительного нерва обнажали глазничную часть зрительного нерва (ON) крыс под анастезией с заходом через надглазничную часть, разрывали мягкие оболочки мозга и все аксоны в ON рассекали с помощью шипцов в течение 10 секунд, 2 мм от решетчатой пластинки.
[0508] Содинения siРНК доставляли по-отдельности или в комбинации в объеме 5 мкл (10 мкг/мкл) в виде глазных капель. Непосредственно после разрушения оптического нерва (ONC) вводили 20 мкг/10 мкл исследуемых siРНК или 10 мкл PBS в один или оба глаза взрослым крысам Вистар и определяли уровни siРНК, впитавшихся в рассеченные и быстрозамороженные цельные сетчатки через 5 часов и 1 день и позжена 2, 4, 7, 14 и 21 день после инъекции. Похожие эксперименты проводили для проверки активности и эффективности siРНК, введенных в виде глазных капель.
Модельные системы травмы реперфузии ишемии после трансплантации легких у крыс
[0509] Получали травму ишемия/реперфузия легких на крысиной модели согласно описанию в публикации Mizobuchi et al., The Journal of Heart and Lung Transplantation, Vol 23 No. 7 (2004) и публикации Kazuhiro Yasufaku et al., Am. J. Respir. Cell Mol Biol, Vol 25, pp 26-34 (2001).
[0510] В частности, после выполнения анестезии изофлураном в трахею вставляли тефлоновый катетер 14 размера и крысе осуществляли механическую вентиляцию с помощью вентилятора для грызунов с использованием 100% кислорода, со скоростью 70 вдохов в минуту и положительным давлением в конце выдоха в 2 см H2O. Левую легочную артерию, вены и главный ствол бронха перевязывали с помощью зажима Кастанеды. Во время операции легкое увлажняли физиологическим раствором и разрез закрывали для сведения к минимуму потерь от испарения. Продолжительность периода ишемии составила 60 минут. В конце периода ишемии снимали зажим и обеспечивали вентиляцию и реперфузию легкого в течение следующих 4, 24 часов и 5 дней после стимуляции ишемии легких. В конце указанного эксперимента аккуратно изымали легкие и замораживали для выделения РНК или фиксировали в коктейле глутарового альдегида для последующего гистологического исследования.
Животная модель блеомицина в качестве модели идиопатического фиброза легких (IPF).
[0511] Исследование осуществимости доставки в легкие и печень siРНК в составе лекарственной формы с витамином A-Coatsome, введенной посредством внутривенной инъекции, и интратрахеальное введение компекса siРНК-витамин A-Coatsome здоровым мышам и мышам, получавшим блеомицин.
[0512] Цель: Проверить два способа введения на осуществимость доставки лекарственной формы siРНК с витамином A-Coatsome в нормальные легкие и фиброзные легкие мышей. Главная проверяемая гипотеза настоящего исследования- обеспечивает ли системное введение лекарственной формы модифицированной siРНК с витамином А-Coatsome полезное поглощение и специфичное в отношении клеток распределение в фиброзных и нормальных легких мышей. Одновременно исследовали интратрахеальный способ введения лекарственной формы модифицированной siРНК с витамином А-Coatsome. Определение siРНК и специфичное в отношении клеток распределение в легких и печени осущесвляли методом гибридизации in situ (ISH)
[0513] Уровень техники: Модель фиброза легких, вызванного блеомицином, была успешно разработана и охарактеризована в течение последних трех десятилетий (Moeller, et al. Int J Biochem Cell Biol, 40:362-382, 2008; Chua et al., Am J Respir Cell Mol Biol 33:9-13, 2005). Гистологические признаки, такие как внутриальвеолярные утолщения, встаивание в стенку коллагена и облитерация пространства альвеол, присутствуют у животных BLM, как и у пациентов с IPF. Ранние исследования показали, что мыши С57/В1 были систематически склонны к вызванному BLM фиброзу легких, тогда как мыши Balb/C были по своей природе устойчивы. В зависимости от способа введения развиваются различные картины фиброза. Интратрахеальное введение BLM приводит к акцентированному бронхиоцентрическому фиброзу, тогда как внутривенное или интраперитонеальное введение вызывает образование рубцов под плеврой, как и в случае болезни у человека (Chua et al. ibid). Использовали мышиную модель обычной интерстициальной пневмонии (UIP). Данная модель проявляет гетерогенное распределение накопления фибрилл (фибропроилиферации), в основном расположенное субплеврально, что приводит к повреждениям, похожим на наблюдаемые в легких пациентов с идиопатическим фиброзом легких (IPF) (Onuma, et al., Tohoku J Exp Med 194:147-156, 2001 and Yamaguchi and Ruoslahti, Nature 336:244-246, 1988). UIP вызывают интраперитонеальной инъекцией блеомицина через день в течение 7 суток для постоянного поддержания субплевральной фибропролиферации в легких мышей (Swiderski et al. Am J Pathol 152:821-828,1998 и Shimizukawa et al., Am J Physiol Lung Cell Mol Physiol 284: L526-L532,2003).
[0514] Как было показано ранее, siРНК, содержащие липосомы, нагруженные витамином А, взаимодействуют с ретинолсвязывающим белком (RBP) и обеспечивают эффективную доставку в звездчатые клетки печени через рецептор RBP (Sato et al. Nat Biotechnol 26:431-442, 2008). Данное исследование предполагалось для проверки того, будет ли комплекс siРНК с витамином A-Coatsome эффективно поглощаться активированными миофибробластами, экспрессирующими рецептор RBP, в легких мышей обработанных блеомицином. Кроме этого проверяли способ местного введения (интратрахеальное введение).
[0515] Общий дизайн исследования
Мыши - С57 В1 самцы
Исходное N (BLM I.P.) - 40 (6 для первой пилотной группы, 34 для исследования, принимая во внимание ожидаемую смертность 25%)
Исходное N (Общее) - 60
Исследуемые siРНК: соединения SERPINHI согласно настоящему описанию.
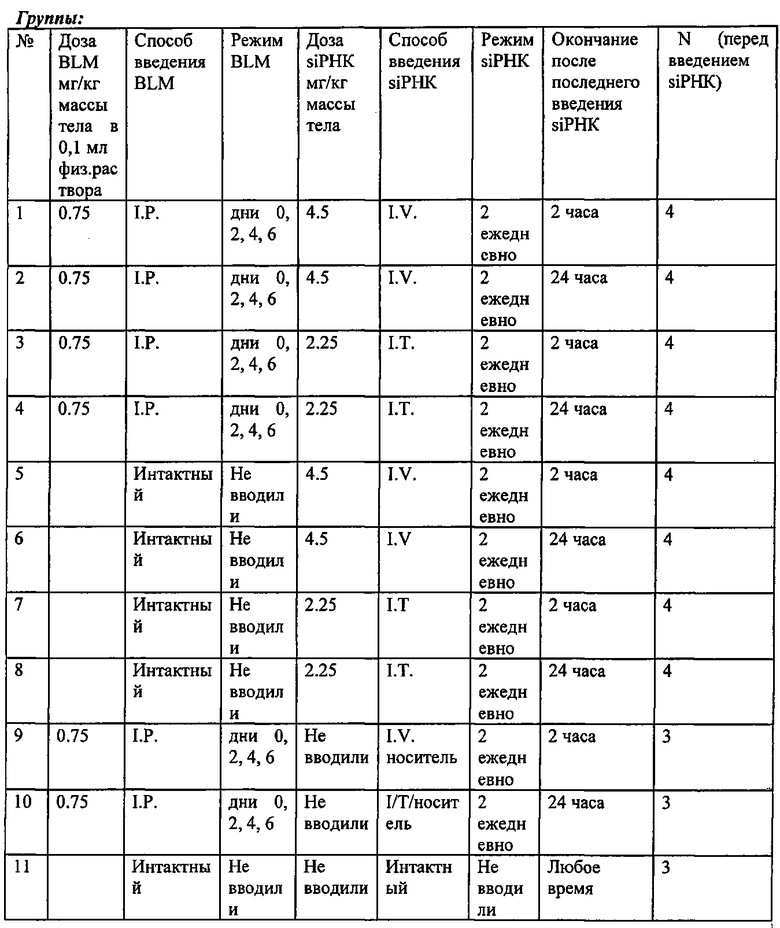
[0516] Фиброз легких, вызванный блеомицином. Фиброз легких у женских особей мышей в возрасте 12 недель C57BL/6 вызывали интраперитонеальным введением гидросульфата блеомицина: 0.75 мг/кг массы тела, растворенный в 0,1 мл физиологического раствора, через день в течение 7 дней, на 0, 2, 4 и 6 сутки.
[0517] Повторный визит и мониторинг. Мышей взвешивали перед началом введения BLM и два раза в неделю в течение исследования.
[0518] Пилотная оценка установления фиброза. Мышей (N=30) подвергали действию BLM в группах с интервалом в одну неделю между первой обработанной группой (N=5) и остальными животными. На 14 день забивали двух мышей из первой группы и забирали легкие для быстрой окраски ГЭ и гистопатологической оценки фиброза. Если подтверждали диагноз фиброза легких, оставшихся крыс распределяли по группам и обрабатывали siPHK на 14 сутки после первого введения BLM. Если на 14 день у животных не развивался достаточный фиброз в легких, остальных мышей из первой обработанной группы забивали на 21 день с последующей быстрой гистопатологической оценкой фиброза. Остальных животных обрабатывали комплексом исследуемой siPHK, начиная с 21 дня после введения BLM.
[0519] Введение siPHK. На 14 или 21 день после первого введения BLM (TBD во время исследования, на основе пилотных оценок установления фиброза) животных распределяли по группам по массе тела. Животным из 1 и 2 группы вводили внутривенно (инъекция в хвостовую вену) комплекс siPHK/витамин A/Coatsome с концентрацией siPHK 4,5 мг/кг массы тела. Интактных животных того же возраста (группы 5 и 6) обрабатывали таким же образом. Мышей, обработанных BLM (группа 9), использовали как контроль с носителем витамин A-coatsome. Через 24 часа инъекцию повторяли всем животным согласно описанию выше.
[0520] Животным BLM из группы 3 и 4 и интактным мышам из 7 и 8 группы проводили анестезию изофураном и подвергали их интратекальному введению 2,25 мг/кг массы тела siPHK в составе лекарственной формы с липосомами, загруженными витамином А. Мышам из группы 10 BLM вводили только носитель витамин A/Coatsome. Интратекальное введение повторяли через 24 часа.
[0521] Завершение исследования. Животных из групп 1, 3, 5, 7, 9 забивали через 2 часа после второй инъекции или введения комплекса siPHK. Животных из групп 2,4,6,8,10 забивали через 24 часа после второй инъекции или введения комплекса siPHK.
[0522] После умерщвления мышам транскардиально вводили 10% нейтральный забуференный формалин. Легкие наполняли 0,8-1,0 мл 10% NBF и перевязывали трахею.
Вырезали легкие и фиксировали их в течение 24 часов в 10% NBF. Печень получали у каждого животного и фиксировали ее в 10% NBF в течение 24 часов.
[0523] Приготовление срезов и оценка. Полученные срезы были приготовлены из легких и печени. Сначала полученные срезы окрашивали гематоксилином и эозином для определения морфологии легких и печени, затем срезы окрашивали Сириус Красным (трехцветный) для идентификации коллагена. Затем полученные срезы подвергали гибридизации in situ (ISH) для детекции siPHK.
[0524] Соединения согласно настоящему описанию исследовали на данной животной модели, и результаты показали, что данные соединения siPHK являются полезными при лечении и/или предотвращении фиброза легких.
[0525] Содержание статей, патентов и заявок на патент и все другие документы и сведения в электронном виде, которые указаны или цитируются в настоящем документе, приведены в качестве ссылки во всей их полноте в той же степени, как если было бы отдельно указано, что каждая публикация должна быть включена посредством ссылки.
[0526] Заявители оставляют за собой право физически включить в публикацию все без исключения материалы и информацию из любых таких статей, патентов, заявок на патент или других физических и электронных документов.
[0527] Специалисту в данной области будет очевидно, что различные замены и модификации могут быть внесены в изобретение согласно настоящему описанию без отклонения от объема и сущности изобретения. Таким образом, такие дополнительные варианты реализации находятся в рамках настоящего изобретения и формулы изобретения. Настоящее изобретение учит специалистов в данной области проверять различные сочетания и/или замены химических модификаций согласно настоящему описанию для получения конструкций нуклеиновых кислот с улучшенной активностью для опосредования активности РНКи. Такая улучшенная активность может включать в себя улучшенную стабильность, улучшенную биодоступность и/или улучшенную активацию клеточных ответов, опосредующих РНКи. Таким образом, конкретные варианты согласно настоящему описанию не являются ограничивающими, и специалисты в данной области с легкостью поймут, что можно исследовать определенные комбинации изменений согласно настоящему описанию без неоправданных экспериментов для идентификации молекул нуклеиновых кислот с улучшенной активностью РНКи.
[0528] Изобретения наглядно согласно настоящему описанию можно практиковать в отсутствии любого элемента или элементов, ограничения или ограничений, не конкретно согласно настоящему описанию. Таким образом, например, неопределенный артикль и определенный артикль и похожие определения в контексте изобретения (особенно в контексте следующих пунктов формулы) следует толковать как единственное и множественное число, если не указано иначе или из контекста не следует противоположное. Термины «содержащий», «имеющий», «включающий» и т.п. следует понимать в широком смысле и без ограничения (например, означающими «включая, но не ограничиваясь ими»). Перечисление пределов значений в настоящем описании направлено только на то, чтобы служить сокращенным способом обозначения отдельного значения, попадающего в предел, если не указано иное, и каждое отдельное значение включено в описание, как если бы было отдельно перечислено. Все способы согласно настоящему описанию можно осуществлять в любом порядке, если не указано иное, или не следует иначе из контекста. Использование любых примеров (например, «такие как») согласно настоящему описанию предназначено только для лучшего освещения изобретения и не создает ограничений объема изобретения, если не указано иначе. Ни одно выражение в описании не должно быть истолковано как указание, что любой не заявленный элемент является важным для практики изобретения. Кроме того, термины и выражения, используемые в настоящем описании, были использованы в качестве условия описания, а не ограничения, и указанные термины и выражения не нацелены на исключение любых эквивалентов показанных и описанных функций или их частей, наоборот, признается, что в рамках заявленного изобретения возможны различные модификации. Таким образом, следует понимать, что, хотя изобретение было конкретно раскрыто в предпочтительных вариантах и дополнительных функциях, специалисты в данной области могут прибегать к модификациям и изменениям изобретений, изложенных в настоящей публикации, и считается, что такие изменения и вариации находятся в рамках данного изобретения.
[0529] Изобретение было описано в широком смысле и общем документе. Каждый узкий вид и подродовая группа, входящие в общее раскрытие информации, также являются частью изобретения. Это включает обобщенное описание изобретения с оговоркой или отрицательным ограничением, удаляющим любой предмет из рода, вне зависимости от того, изложен ли вырезанный материал в настоящем описании. Другие варианты находятся в пределах следующей формулы. Кроме того, если функции и аспекты изобретения, описанного в терминах групп Маркуша, специалистам в данной области будет понятно, что изобретение также описывается в терминах отдельных членов или подгруппы членов группы Маркуша.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕРАПЕВТИЧЕСКИЕ КОМПОЗИЦИИ И СПОСОБЫ ПРОТИВ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ С МОЛЕКУЛАМИ РНКИ, НАПРАВЛЕННЫМИ ПРОТИВ HSP47 | 2015 |
|
RU2756253C2 |
| ЛИПОСОМЫ С РЕТИНОИДОМ ДЛЯ УСИЛЕНИЯ МОДУЛЯЦИИ ЭКСПРЕССИИ HSP47 | 2012 |
|
RU2628694C2 |
| НУКЛЕИНОВЫЕ КИСЛОТЫ ДЛЯ ИНГИБИРОВАНИЯ ЭКСПРЕССИИ LPA В КЛЕТКЕ | 2018 |
|
RU2822093C1 |
| СТРУКТУРЫ МИРНК С ВЫСОКОЙ АКТИВНОСТЬЮ И СНИЖЕННЫМ ВОЗДЕЙСТВИЕМ ВНЕ МИШЕНИ | 2016 |
|
RU2788030C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ИНФЕКЦИИ ГЕПАТИТА В | 2018 |
|
RU2780021C2 |
| МИМЕТИКИ MIR-29 И ПУТИ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2712511C2 |
| iPHK-ОПОСРЕДОВАННОЕ ИНГИБИРОВАНИЕ СВЯЗАННЫХ С ФАКТОРОМ НЕКРОЗА ОПУХОЛИ АЛЬФА СОСТОЯНИЙ | 2007 |
|
RU2469090C2 |
| ОЛИГОРИБОНУКЛЕОТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ОСТРОЙ ПОЧЕЧНОЙ НЕДОСТАТОЧНОСТИ | 2005 |
|
RU2584609C2 |
| ЛЕЧЕНИЕ НАРУШЕНИЙ ЦНС | 2007 |
|
RU2426544C2 |
| ОЛИГОРИБОНУКЛЕОТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ОСТРОЙ ПОЧЕЧНОЙ НЕДОСТАТОЧНОСТИ И ДРУГИХ ЗАБОЛЕВАНИЙ | 2005 |
|
RU2434942C2 |






























































































































































































































































































































































































































































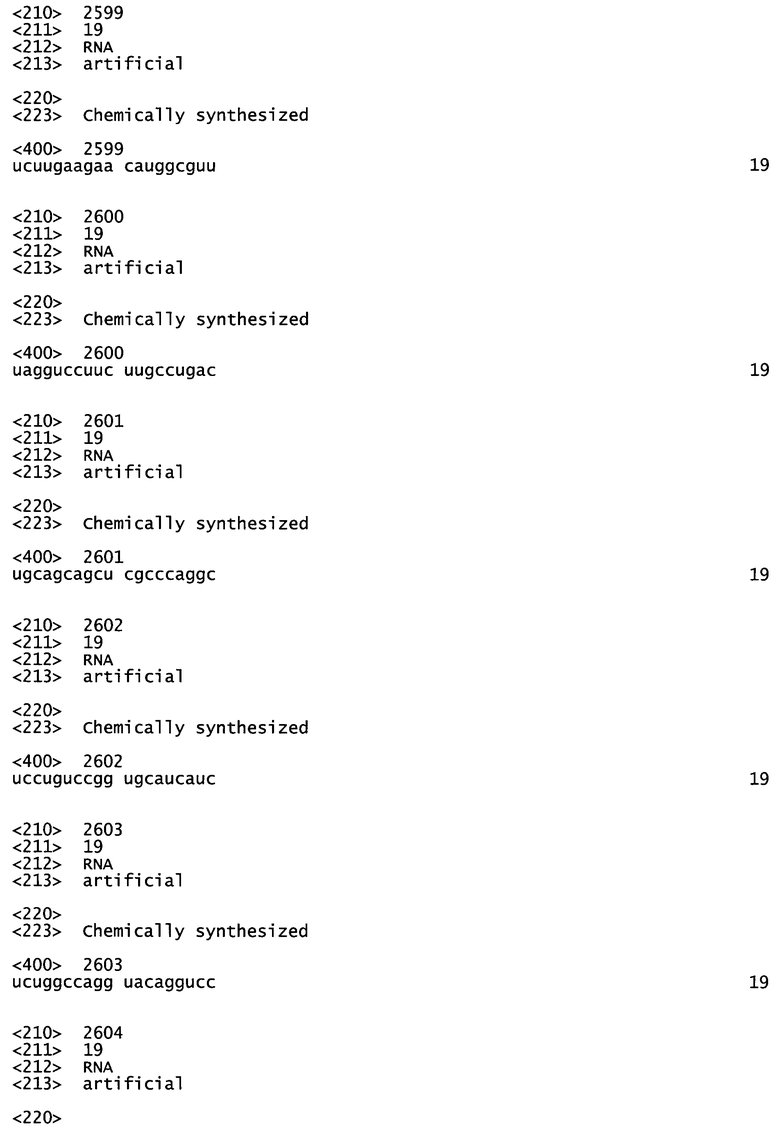








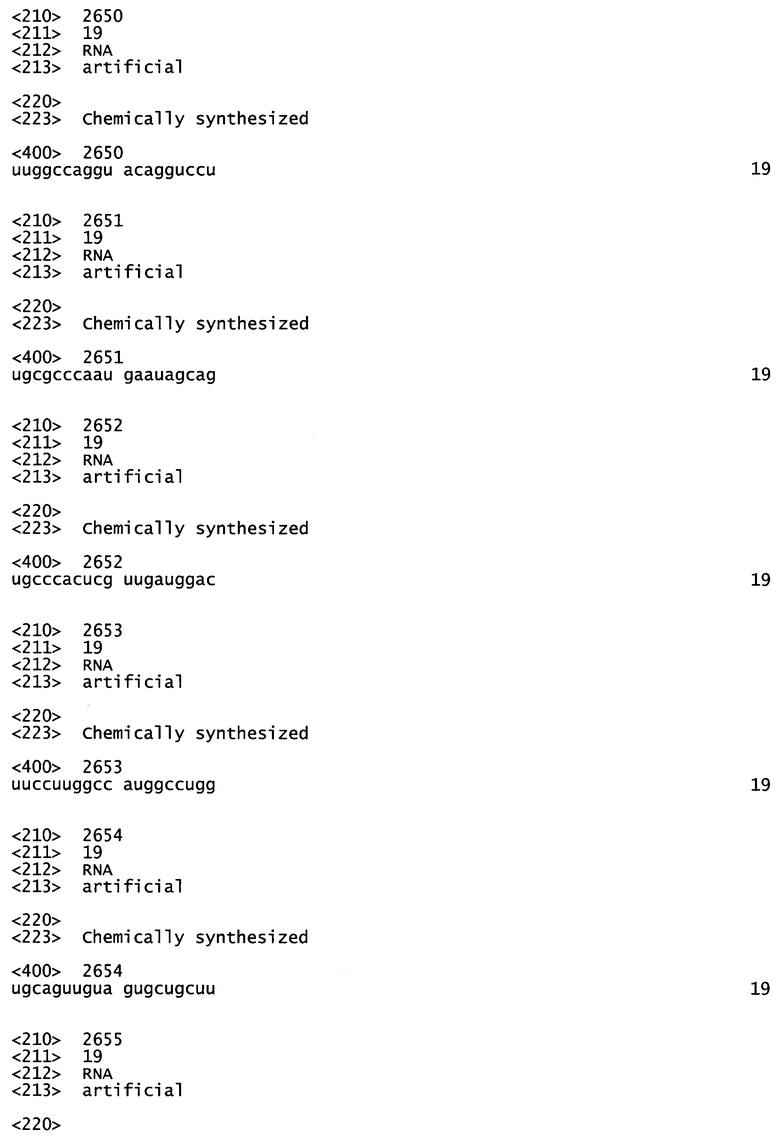



















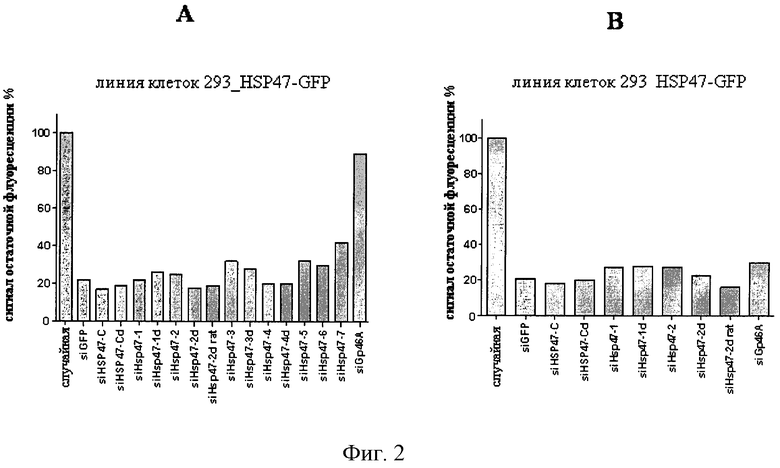
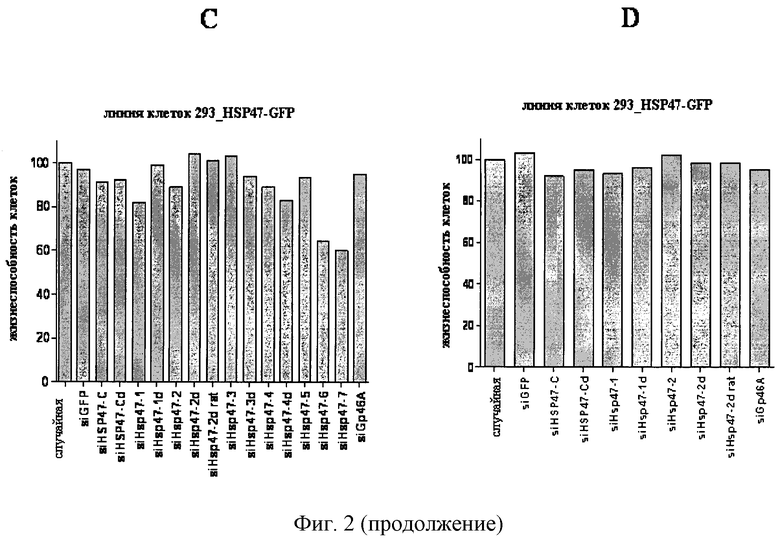
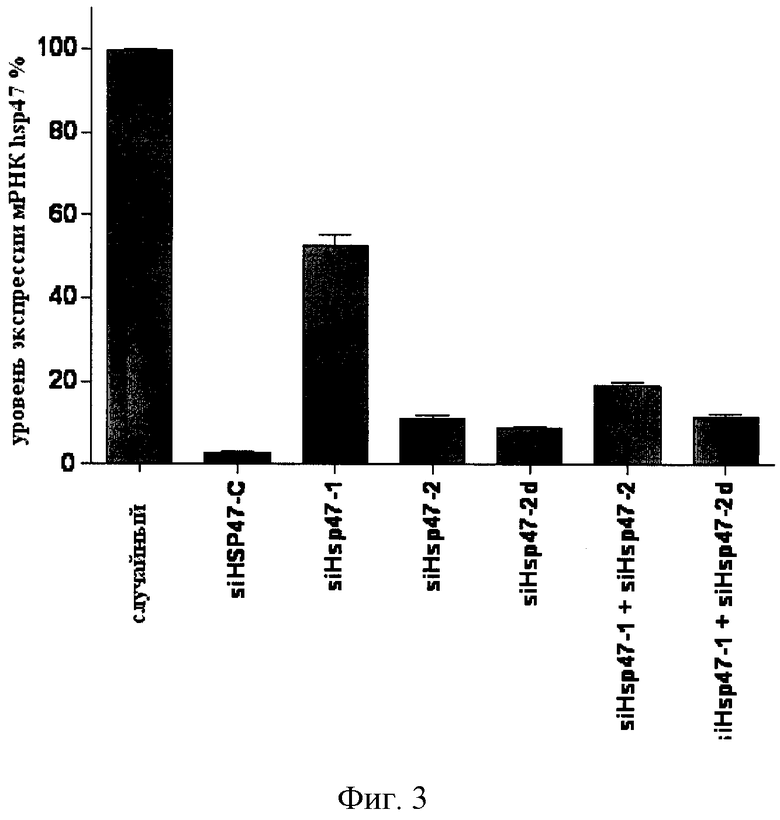
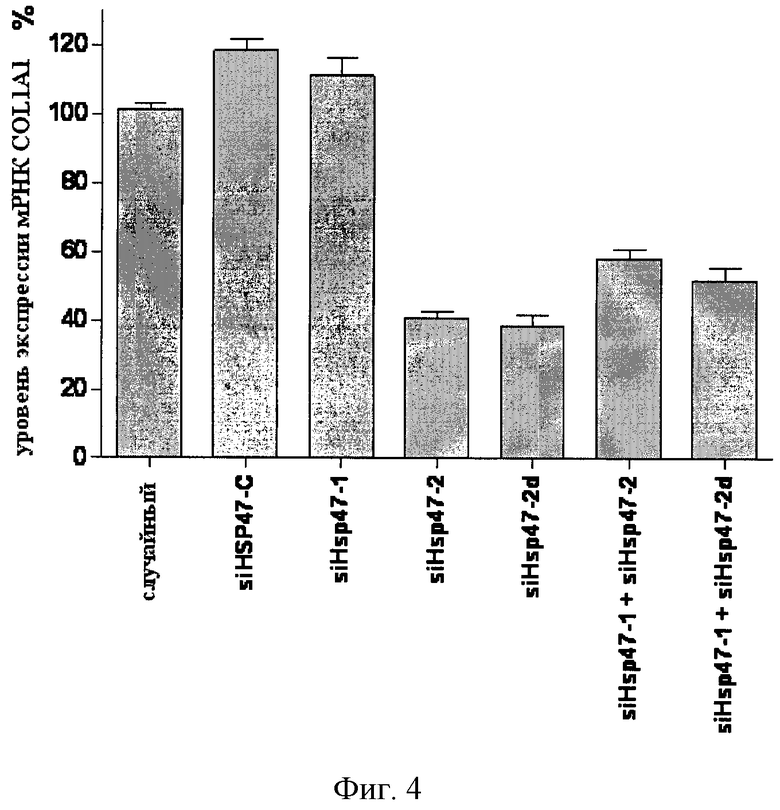
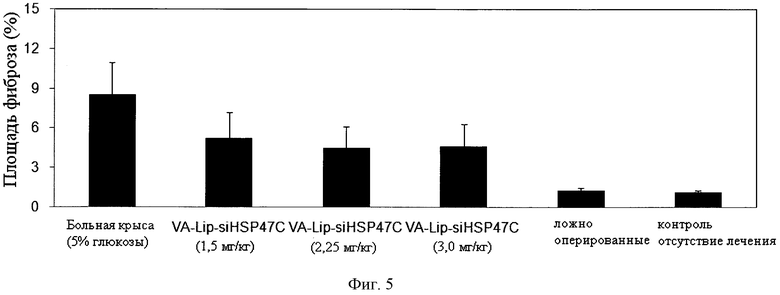



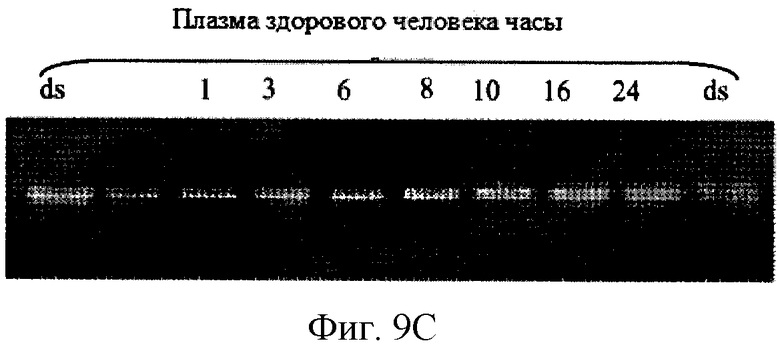

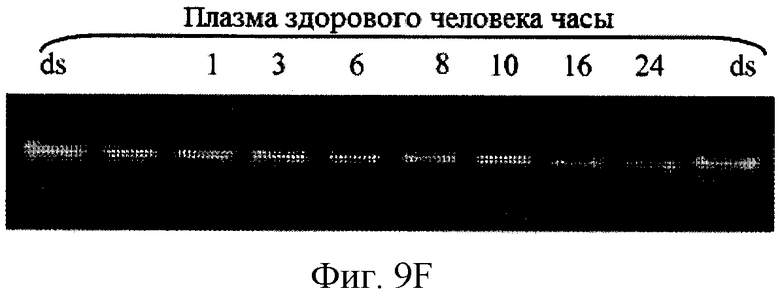
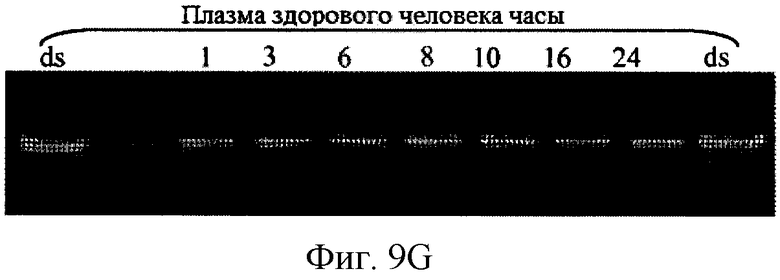

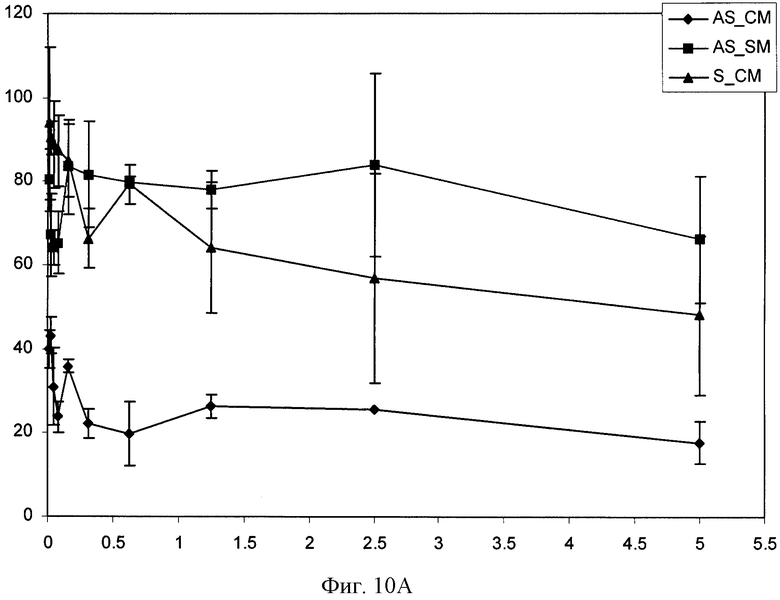
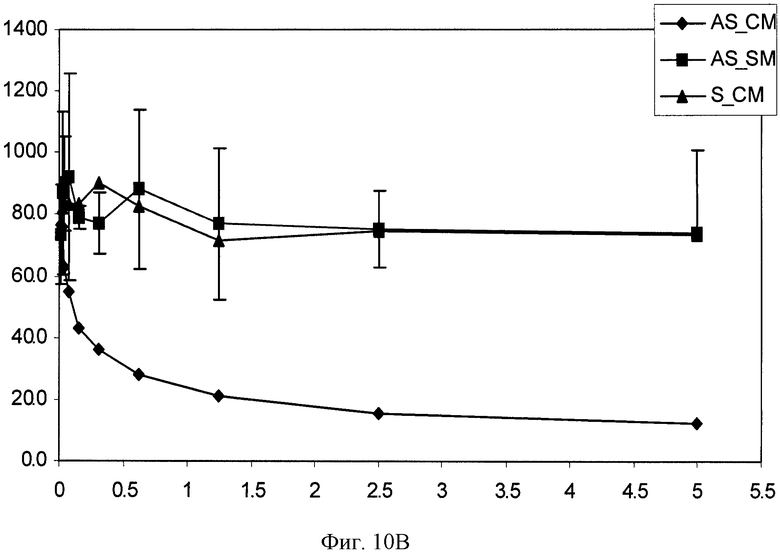
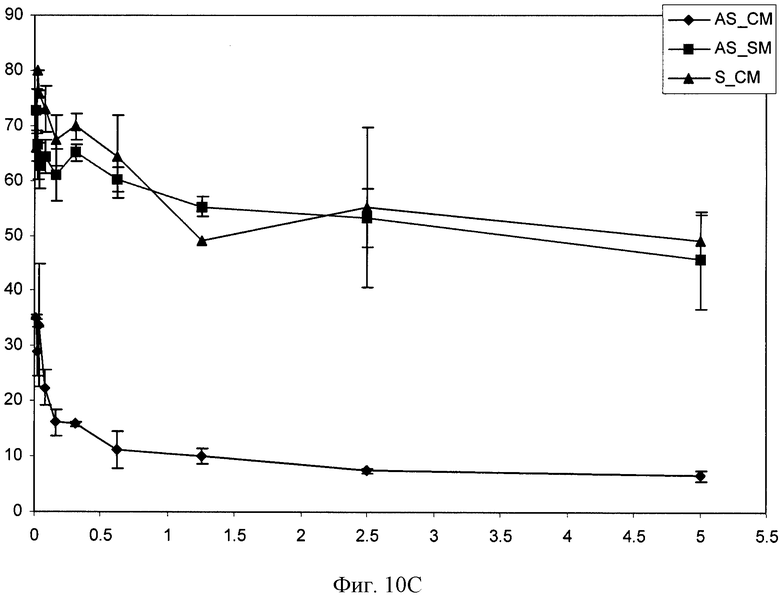
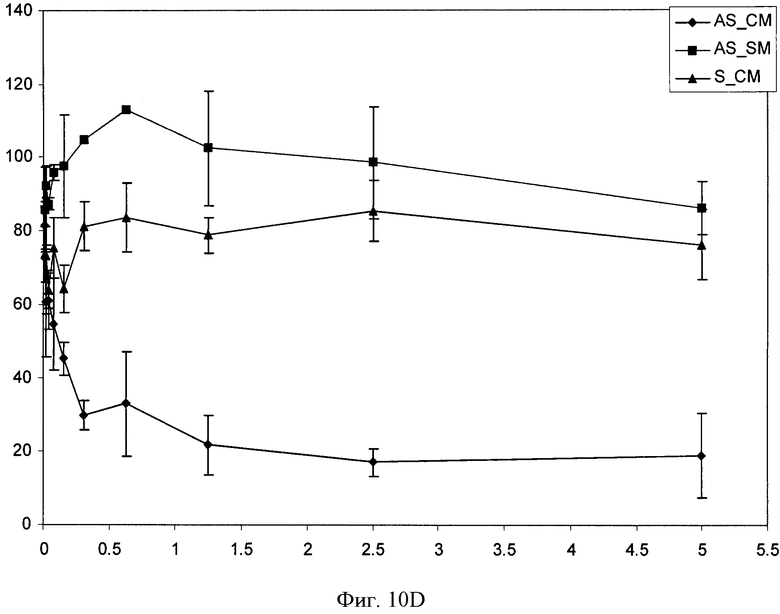
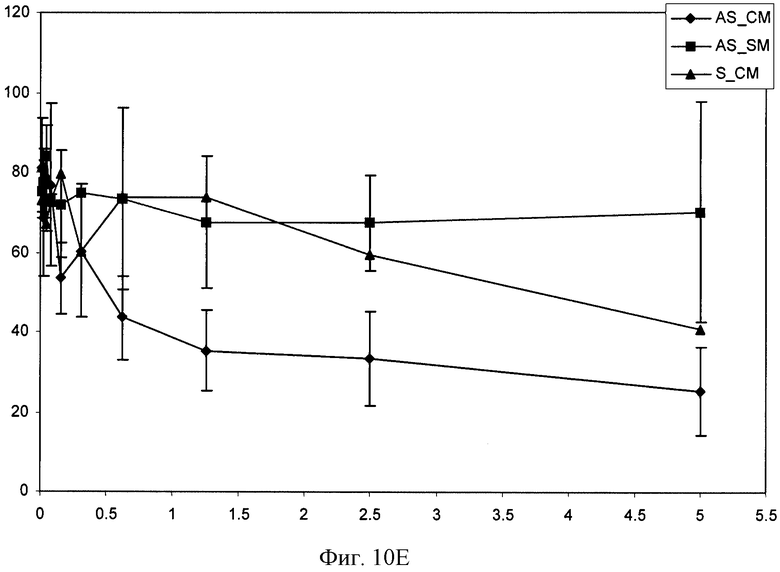
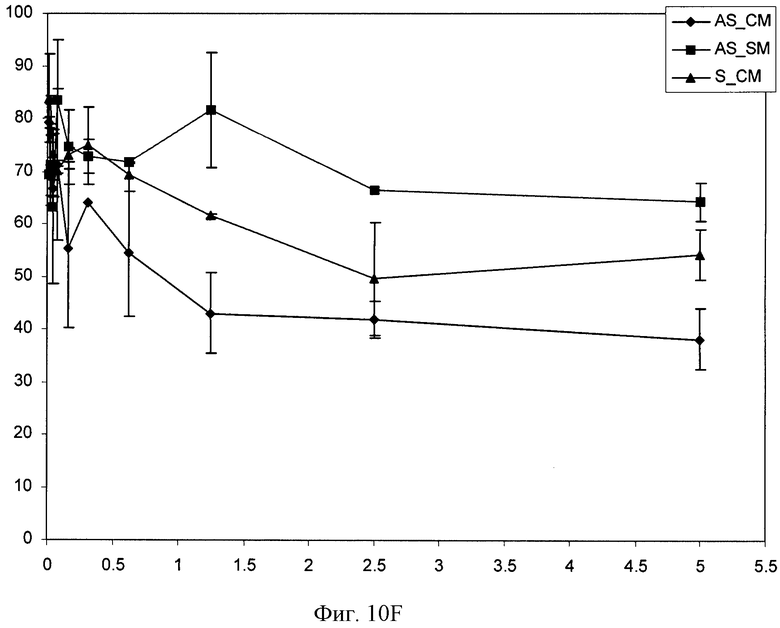
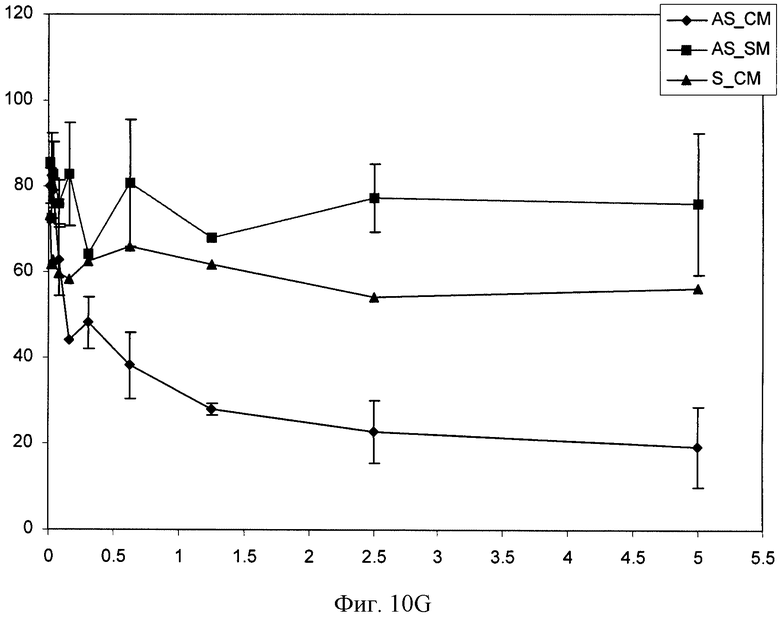
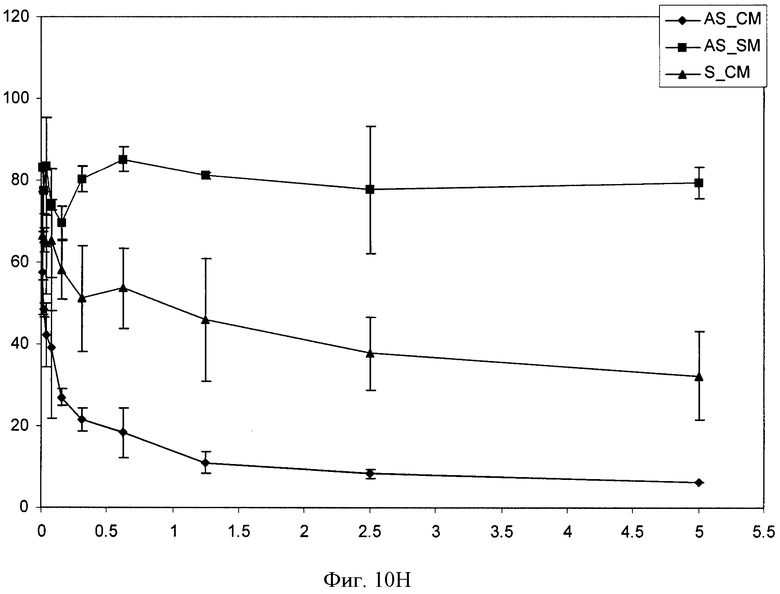
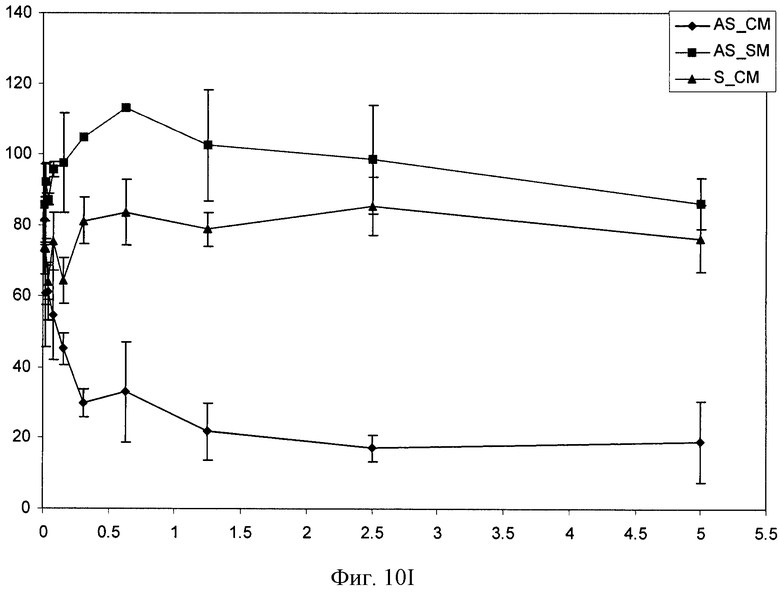
Изобретение относится к области биохимии, в частности к молекулам нуклеиновой кислоты для модуляции экспрессии генов-мишеней. Заявлена двухцепочечная молекула нуклеиновой кислоты для ингибирования экспрессии гена белка теплового шока 47 (hsp47). Также представлены композиция, включающая указанную молекулу нуклеиновой кислоты, и способ лечения пациента, страдающего от заболевания, связанного с hsp47. Изобретение может быть использовано для лечения ассоциированных с hsp47 состояний и нарушений, таких как фиброз печени, фиброз легких, фиброз брюшины и фиброз почек. 3 н. и 22 з.п. ф-лы, 27 ил., 31 табл., 10 пр. 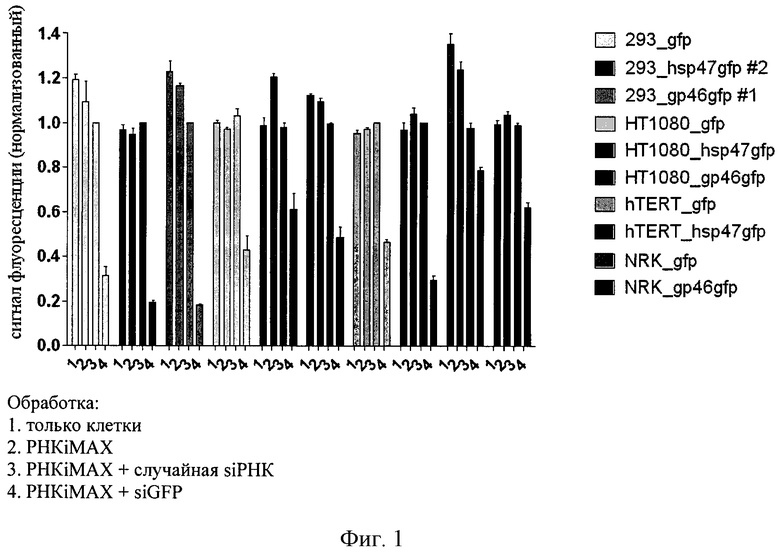
1. Двухцепочечная молекула нуклеиновой кислоты для ингибирования экспрессии гена hsp47, имеющая структуру (А2), изображенную ниже:
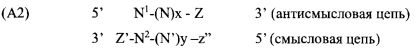
в которой каждый из N2, N и N′ представляет собой немодифицированный или модифицированный рибонуклеотид или нетрадиционную группу;
в которой каждый из (N)x и (N′)y представляет собой олигонуклеотид, в котором каждый последующий N или N′ соединен с соседним N или N′ с помощью ковалентной связи;
в которой каждый из x и y независимо представляет собой целое число от 17 до 39;
в которой последовательность (N′)y имеет комплементарность последовательности (N)x, и последовательность (N)x имеет комплементарность непрерывной последовательности в мРНК, кодирующей hsp47;
в которой N1 ковалентно связана с (N)x и не комплементарна мРНК, кодирующей hsp47, или представляет собой ДНК группу, комплементарную мРНК, кодирующей hsp47;
при этом N1 представляет собой группу, выбранную из группы, состоящей из природного или модифицированного уридина, дезоксиуридина, риботимидина, дезоксириботимидина, аденозина или дезоксиаденозина;
в которой z″ может присутствовать или отсутствовать, но при наличии представляет собой защитную группу, ковалентно присоединенную к 5′-концу N2-(N′)y;
в которой каждый из Z и Z′ может независимо присутствовать или отсутствовать, но при наличии независимо представляет собой 1-5 последовательных нуклеотидов, последовательных ненуклеотидных групп или сочетание нуклеотидов и ненуклеотидных групп, ковалентно присоединенное к 3′-концу цепи, в которой она присутствует, и
при этом указанная двухцепочечная молекула нуклеиновой кислоты включает олигонуклеотид, обозначенный как SERPINH1_2 (SEQ ID NO: 60 и SEQ ID NO: 127), SERPINH1_45a (SEQ ID NO: 98 и SEQ ID NO: 165), или SERPINH1_51 (SEQ ID NO: 101 и SEQ ID NO: 168).
2. Двухцепочечная молекула нуклеиновой кислоты по п.1, отличающаяся тем, что x=y=18.
3. Двухцепочечная молекула нуклеиновой кислоты по п.1, отличающаяся тем, что смысловая цепь и антисмысловая цепь выбраны из олигонуклеотида, представленного SERPINH1_2 (SEQ ID NO: 60 и SEQ ID NO: 127), SERPINH1_45a (SEQ ID NO: 98 и SEQ ID NO: 165) и SERPINH1_51 (SEQ ID NO: 101 и SEQ ID NO: 168).
4. Двухцепочечная молекула нуклеиновой кислоты по п.3, представленная SERPINH1_2, которая имеет структуру

в которой каждый знак «|» обозначает спаривание оснований между рибонуклеотидами;
в которой каждый из А, С, G и U независимо представляет собой немодифицированный или модифицированный рибонуклеотид или нетрадиционную группу:
в которой каждый из Z и Z′ независимо может присутствовать или отсутствовать, но при наличии независимо представляет собой 1-5 последовательных нуклеотидных или ненуклеотидных групп или сочетание нуклеотидов и ненуклеотидных групп, ковалентно присоединенное к 3′-концу цепи, в которой оно присутствует; и
в которой z″ может присутствовать или отсутствовать, но при наличии представляет собой защитную группу, ковалентно присоединенную к 5′-концу смысловой цепи.
5. Двухцепочечная молекула нуклеиновой кислоты по п.4, отличающаяся тем, что каждый из А, С, G и U представляет собой немодифицированный рибонуклеотид, и z″, Z и Z′ отсутствуют.
6. Двухцепочечная молекула нуклеиновой кислоты по п.4, отличающаяся тем, что Z и Z′ присутствуют.
7. Двухцепочечная молекула нуклеиновой кислоты по п.6, отличающаяся тем, что Z и Z′ независимо содержит выступающий конец из нуклеотидов или не являющиеся нуклеотидами группы.
8. Двухцепочечная молекула нуклеиновой кислоты по п.7, отличающаяся тем, что Z и Z′ содержит не являющиеся нуклеотидами группы.
9. Двухцепочечная молекула нуклеиновой кислоты по п.8, отличающаяся тем, что Z′ содержит C3Pi формулы:
 ; и
; и
при этом Z содержит C3Pi-C3OH формулы:
 .
.
10. Двухцепочечная молекула нуклеиновой кислоты по п.4, отличающаяся тем, что антисмысловая цепь содержит рибонуклеотиды с модифицированным 2′-O-метил остатком сахара.
11. Двухцепочечная молекула нуклеиновой кислоты по п.10, отличающаяся тем, что рибонуклеотиды с модифицированным 2′-O-метил остатком сахара находятся в положениях (5′>3′)3, 5, 9, 11, 13, 15, 17 и 19.
12. Двухцепочечная молекула нуклеиновой кислоты по п.11, отличающаяся тем, что антисмысловая цепь также содержит рибонуклеотид с модифицированным 2′-O-метил остатком сахара в положении (5′>3′) 1.
13. Двухцепочечная молекула нуклеиновой кислоты по п.12, отличающаяся тем, что антисмысловая цепь содержит 2′5′рибонуклеотид в положении (5'>3′)7.
14. Двухцепочечная молекула нуклеиновой кислоты по п.4 или 13, отличающаяся тем, что смысловая цепь содержит 4 или 5 последовательных 2′5′ рибонуклеотида в 3′-терминальном или предпоследнем положениях.
15. Двухцепочечная молекула нуклеиновой кислоты по п.14, отличающаяся тем, что смысловая цепь содержит 2′5′ рибонуклеотиды в положениях (5′>3′) 15, 16, 17 и 18.
16. Двухцепочечная молекула нуклеиновой кислоты по п.4, отличающаяся тем, что z″ присутствует и содержит защитную группу, ковалентно присоединенную к 5′-концу смысловой цепи.
17. Двухцепочечная молекула нуклеиновой кислоты по п.16, отличающаяся тем, что защитная группа выбрана из группы, состоящей из группы рибозы без основания, инвертированной группы рибозы без основания, инвертированной группы дезоксирибозы без основания, группы дезоксирибозы без основания и их модификаций; С6-имино-Pi; инвертированного нуклеотида; 5′ОМе нуклеотида; 4′,5′-метилен нуклеотида; 1-(β-D-эритрофуранозил)нуклеотида; 4′-тио нуклеотида, карбоциклического нуклеотида; 5′-амино-алкил фосфата, 1,3-диамино-2-пропил-фосфата, 3-аминопропил фосфата, 6-фосфат аминогексила; 12-аминододецил фосфата; гидроксипропил фосфата; 1,5-ангидрогекситол нуклеотида; альфа-нуклеотида; трео-пентофуранозил нуклеотида; ациклического 3′,4′-секо нуклеотида; 3,4-дигидроксибутил нуклеотида; 3,5-дигидроксипентил нуклеотида, 5′-5′ инвертированной группы без основания; 1,4-бутандиол фосфата; 5′-амино; и мостиковой или немостиковой метилфосфонатной и 5′-меркапто группы.
18. Двухцепочечная молекула нуклеиновой кислоты по п.17, отличающаяся тем, что защитная группа содержит инвертированную группу дезоксирибозы без основания.
19. Двухцепочечная молекула нуклеиновой кислоты по п.1, отличающаяся тем, что антисмысловая цепь содержит рибонуклеотиды с модифицированным 2′-O-метил остатком сахара, 2′5′ рибонуклеотид по меньшей мере в одном из положений (5′>3′) 1, 5, 6 или 7, и 3′-терминальную не являющуюся нуклеотидом группу, дополнительно C3Pi-C3OH не являющуюся нуклеотидом группу, ковалентно присоединенную к 3′-концу; и смысловая цепь содержит по меньшей мере один 2′5′ рибонуклеотид и/или рибонуклеотид с модифицированным 2′-O-метил остатком сахара, не являющуюся нуклеотидом группу, ковалентно присоединенную к 3′-концу и защитную группу, ковалентно присоединенную к 5′-концу.
20. Двухцепочечная молекула нуклеиновой кислоты по п.2 или 4, отличающаяся тем, что антисмысловая цепь содержит рибонуклеотиды с модифицированным 2′-O-метил остатком сахара в положениях (5′>3′) 1, 3, 5, 9, 11, 13, 15, 17 и 19, 2′5′ рибонуклеотид в положении 7, и C3Pi-C3OH не являющуюся нуклеотидом группу, ковалентно присоединенную к 3′-концу; и смысловая цепь содержит 5 последовательных 2′5′ рибонуклеотидов в 3′-терминальных положениях (5′>3′) 15, 16, 17, 18 и 19, C3Pi не являющуюся нуклеотидом группу, ковалентно присоединенную к 3′-концу, и инвертированную группу, лишенную основания, ковалентно присоединенную к 5′-концу.
21. Двухцепочечная молекула нуклеиновой кислоты по п.2 или 4, отличающаяся тем, что антисмысловая цепь содержит рибонуклеотиды с модифицированным 2′-O-метил остатком сахара в положениях (5′>3′) 3, 5, 9, 11, 13, 15, 17 и 19, 2′5′ рибонуклеотид в положениях 1 и 7, и C3Pi-C30H не являющуюся нуклеотидом группу, ковалентно присоединенную к 3′-концу; и смысловая цепь содержит 5 последовательных 2′5′ рибонуклеотидов в 3′-терминальных положениях (5′>3′) 15, 16, 17, 18 и 19, C3Pi не являющуюся нуклеотидом группу, ковалентно присоединенную к 3′-концу, и инвертированную группу, лишенную основания, ковалентно присоединенную к 5′-концу.
22. Двухцепочечная молекула нуклеиновой кислоты по п.2 или 4, отличающаяся тем, что антисмысловая цепь содержит рибонуклеотиды с модифицированным 2′-O-метил остатком сахара в положениях (5′>3′) 1, 3, 5, 9, 11, 13, 15, 17 и 19, 2′5′ рибонуклеотид в положении 7, C3Pi- С3ОН не являющуюся нуклеотидом группу, ковалентно присоединенную к 3′-концу; и смысловая цепь содержит рибонуклеотиды с модифицированным 2′-O-метил остатком сахара в положениях (5′>3′) 7, 13, 16 и 18, 2′5′ рибонуклеотид в положении 9, С3 не являющуюся нуклеотидом группу, ковалентно присоединенную к 3′-концу, и инвертированную группу, лишенную основания, ковалентно присоединенную к 5′-концу.
23. Композиция, содержащая двухцепочечную молекулу нуклеиновой кислоты по любому из пп.1-22 и фармацевтически приемлемый носитель для ингибирования экспрессии гена hsp47.
24. Способ лечения пациента, страдающего от заболевания, связанного с hsp47, включающий введение указанному пациенту двухцепочечной молекулы нуклеиновой кислоты по любому из пп.1-22 в количестве, достаточном для снижения экспрессии hsp47, при этом указанная двухцепочечная молекула нуклеиновой кислоты подходит для применения для лечения пациента.
25. Способ по п.24, отличающийся тем, что указанное состояние представляет собой фиброз.
| US 20080193512 A1, 14.08.2008 | |||
| US 20090192104 A1, 30.07.2009 | |||
| US 20060216722 A1, 28.09.2006 | |||
| US 20060134663 A1, 22.06.2006 | |||
| US 20070042982 A1, 22.02.2007 | |||
| JP 2003159087 A, 03.06.2003 | |||
| JP 2001031588 A, 06.02.2001 | |||
| WO 1999060142 A2, 25.11.1999 | |||
| ДИАМИНОТРИАЗОЛЫ, ПРИГОДНЫЕ В КАЧЕСТВЕ ИНГИБИТОРОВ ПРОТЕИНКИНАЗ | 2003 |
|
RU2350606C2 |

