В соответствии с настоящим изобретением предложены продукты и композиции, а также варианты их применения. В частности, настоящее изобретение относится к производным нуклеиновых кислот, которые препятствуют экспрессии гена LPA или ингибируют его экспрессию. Такая лечебная терапия, направленная на снижение Lp(a), служит для предотвращения и уменьшения риска развития инсульта, атеросклероза, тромбоза и сердечно-сосудистых заболеваний, таких как ишемическая болезнь сердца и стеноз аорты, или любого другого нарушения, патологии или синдрома, связанных с повышенными уровнями Lp(a)-содержащих частиц.
Уровень техники
Было показано, что двухцепочечная РНК (дцРНК), способная комплементарно связывать экспрессированную мРНК, способна блокировать экспрессию генов (Fire et a.l, 1998, Nature. 1998 Feb 19; 391(6669):806-11 и Elbashir et al., 2001, Nature. 2001 May 24; 411(6836):494-8) с помощью механизма, который был назван РНК-интерференцией (РНКи). Короткие дцРНК направляют геноспецифичное посттранскрипционное подавление («silencing») у многих организмов, включая позвоночных, и такие молекулы стали полезным инструментом для изучения функции генов. РНКи опосредуется РНК-индуцированным комплексом подавления (RISC), специфичной в отношении последовательности многокомпонентной нуклеазой, которая разрушает информационные РНК, гомологичные инициирующему фактору подавления, загруженному в комплекс RISC. Интерферирующие РНК (называемые в настоящей заявке иРНК), такие как миРНК, направленная против второй цепи РНК, и микроРНК, представляют собой олигонуклеотиды, которые предотвращают образование белков путем подавления генов, т.е. ингибируют трансляцию гена белка за счет разрушения молекул мРНК. Агенты, подавляющие гены, приобретают все большее значение для терапевтических приложений в медицине.
Согласно Watts and Corey в Journal of Pathology (2012; Vol 226, p 365-379), существуют алгоритмы, которые можно применять для конструирования инициирующих факторов подавления нуклеиновых кислот, однако все они имеют серьезные ограничения. Для идентификации эффективных иРНК могут потребоваться различные экспериментальные способы, поскольку алгоритмы не учитывают такие факторы как третичная структура мРНК-мишени или участие РНК-связывающих белков. Таким образом, обнаружение эффективного инициирующего фактора подавления нуклеиновой кислоты с минимальными нецелевыми эффектами является сложным процессом. Для фармацевтической разработки этих очень потенциально опасных молекул необходимо, чтобы их синтез был экономически выгодным, они распределялись по тканям-мишеням, проникали в клетки и функционировали в приемлемых пределах токсичности.
Lp(a) представляет собой гетерогенную частицу липопротеина низкой плотности, экспрессируемую преимущественно в печени (Witztum and Ginsberg, J Lipid Res. 2016 Mar; 57(3):336-9). Она состоит из аполипопротеина (a) (Apo(a), кодируемого геном LPA), соединенного с ЛПНП за счет полипептида ApoB. Высокие уровни Lp(a) в сыворотке, обусловленные генетически, не зависят от рациона и физических упражнений и ассоциированы с повышенным риском развития сердечно-сосудистого заболевания за счет ассоциированного потенциала к развитию атеросклероза (Alonso et al., Journal of the American College of Cardiology Vol. 63, No. 19, 2014). С точки зрения диагностики и превентивной медицины уровень Lp(a) в сыворотке крови пациента является широко распространенным, независимым генетическим фактором риска развития ишемической болезни сердца и стеноза аорты (Saeedi and Frohlich Clinical Diabetes and Endocrinology (2016) 2:7). В настоящее время не существует одобренной специфической терапии, направленной на уменьшение уровня Lp(a), помимо косвенных стандартных общих мер по снижению ЛПНП. Соответственно, в настоящее время необходимы способы эффективного лечения, предотвращения и уменьшения риска развития нарушений, таких как инсульт, атеросклероз, тромбоз и сердечно-сосудистые заболевания или ассоциированных с ними, таких как ишемическая болезнь сердца, стеноз аорты и другие пока не идентифицированные ассоциированные нарушения, патологии или синдромы. Настоящее изобретение направлено на решение этой неудовлетворенной медицинской потребности.
Первый аспект изобретения относится к нуклеиновой кислоте для ингибирования экспрессии LPA в клетке, содержащей по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, при этом указанная первая цепь содержит нуклеотидную последовательность, выбранную из следующих последовательностей: SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73.
Согласно одному варианту реализации настоящего изобретения, первая цепь нуклеиновой кислоты содержит последовательность из по меньшей мере 15, предпочтительно по меньшей мере 16, более предпочтительно по меньшей мере 17, еще более предпочтительно по меньшей мере 18 и наиболее предпочтительно по меньшей мере 19 нуклеотидов любой одной из эталонных («reference») последовательностей SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73.
Согласно одному варианту реализации настоящего изобретения, количество несовпадений отдельных нуклеотидов в последовательности первой цепи относительно части эталонной последовательности, которая содержится в последовательности первой цепи, составляет не более трех, предпочтительно не более двух, более предпочтительно не более одного и наиболее предпочтительно ноль.
Вторая цепь может содержать нуклеотидную последовательность SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 38, 40, 42, 44, 64, 66, 68, 70, 72 или 74.
Первая цепь может содержать нуклеотидную последовательность SEQ ID NO: 5 и SEQ ID NO: 9, и/или вторая цепь может содержать нуклеотидную последовательность SEQ ID NO: 6 и SEQ ID NO: 10.
Каждая из указанных первой цепи и/или второй цепей может содержать 17-35 нуклеотидов в длину, и по меньшей мере одна дуплексная область может содержать 10-25 нуклеотидов в длину. Дуплекс может содержать две отдельные цепи или он может содержать одну цепь, которая содержит первую цепь и вторую цепь.
Нуклеиновая кислота может: a) иметь тупые концы на обоих концах; b) содержать липкий конец на одном конце и тупой конец на другом; или c) содержать липкие концы на обоих концах.
Один или более нуклеотидов на первой и/или второй цепях могут быть модифицированы с образованием модифицированных нуклеотидов. Один или более из нуклеотидов в нечетных положениях первой цепи могут быть модифицированы. Один или более из нуклеотидов в четных положениях первой цепи могут быть модифицированы с применением по меньшей мере второй модификации, причем указанная по меньшей мере вторая модификация отличается от модификации на одном или более нечетных нуклеотидах. По меньшей мере один из одного или более модифицированных нуклеотидов в четных положениях может быть смежным с по меньшей мере одним или более модифицированными нечетными нуклеотидами.
Множество нуклеотидов в нечетных положениях последовательности первой цепи могут быть модифицированы в нуклеиновой кислоте согласно настоящему изобретению. Множество нуклеотидов в четных положениях последовательности первой цепи могут быть модифицированы с применением второй модификации. Первая цепь может содержать смежные нуклеотиды, которые модифицированы с применением общей модификации. Первая цепь также может содержать смежные нуклеотиды, которые модифицированы с применением второй отличающейся модификации.
Один или более из нуклеотидов в нечетных положениях последовательности второй цепи могут быть модифицированы с применением модификации, которая отличается от модификации нуклеотидов в нечетных положениях первой цепи, и/или один или более из нуклеотидов в четных положениях второй цепи могут быть модифицированы с применением модификации, которая аналогична таковой для нуклеотидов в нечетных положениях первой цепи. По меньшей мере один из одного или более модифицированных нуклеотидов в четных положениях второй цепи может быть смежным с одним или более модифицированными нуклеотидами в нечетных положениях. Множество нуклеотидов в нечетных положениях второй цепи могут быть модифицированы с применением общей модификации, и/или множество нуклеотидов в четных положениях могут быть модифицированы с применением модификации, аналогичной той, которая присутствует на нечетных нуклеотидах первой цепи. Множество нуклеотидов в нечетных положениях второй цепи могут быть модифицированы с применением второй модификации, причем указанная вторая модификация отличается от модификации нуклеотидов в нечетных положениях первой цепи.
Вторая цепь может содержать смежные нуклеотиды, которые модифицированы с применением общей модификации, которая может представлять собой вторую модификацию, которая отличается от модификации нуклеотидов в нечетных положениях первой цепи.
В нуклеиновой кислоте согласно настоящему изобретению каждый из нуклеотидов в нечетных положениях первой цепи и каждый из нуклеотидов в четных положениях второй цепи может быть модифицирован с применением общей модификации, а каждый из нуклеотидов в четных положениях первой цепи может быть модифицирован с применением второй модификации, и каждый из нуклеотидов в нечетных положениях второй цепи может быть модифицирован с применением второй отличающейся модификации.
В нуклеиновой кислоте согласно настоящему изобретению модифицированные нуклеотиды первой цепи могут быть сдвинуты по меньшей мере на один нуклеотид относительно немодифицированных или по-разному модифицированных нуклеотидов второй цепи.
Модификация и/или модификации могут быть выбраны по отдельности и независимо из группы, состоящей из 3'-концевого дезокситимина, 2'-O-метила, 2'-дезокси-модификации, 2'-амино-модификации, 2'-алкил-модификации, морфолино-модификации, фосфоамидатной модификации, модификации 5'-фосфотиоатной группой, модификации 5'-фосфатом или миметиком 5'-фосфата, а также модификации производным холестерина или группой бисдециламида додекановой кислоты, и/или модифицированный нуклеотид может представлять собой любой из закрытого нуклеотида, лишенного азотистого основания нуклеотида или содержащего неприродное основание нуклеотида. По меньшей мере одна модификация может представлять собой 2'-O-метил и/или по меньшей мере одна модификация может представлять собой 2'-F.
Согласно настоящему изобретению также предложена, в качестве второго аспекта, нуклеиновая кислота для ингибирования экспрессии LPA в клетке, содержащая по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, причем указанная первая цепь содержит нуклеотидную последовательность, выбранную из следующих последовательностей: SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27,29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73, при этом нуклеотиды первой цепи модифицированы с применением первой модификации для нуклеотидов в нечетных положениях и модифицированы с применением второй модификации для нуклеотидов в четных положениях, а нуклеотиды второй цепи модифицированы с применением третьей модификации для нуклеотидов в четных положениях и модифицированы с применением четвертой модификации для нуклеотидов в нечетных положениях, при этом по меньшей мере первая модификация отличается от второй модификации, и третья модификация отличается от четвертой модификации. Вторая цепь может содержать нуклеотидную последовательность SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28 30, 32, 34, 36, 38, 40, 42, 44, 64, 66, 68, 70, 72 или 74. Третья модификация и первая модификация могут быть одинаковыми и/или вторая модификация и четвертая модификация могут быть одинаковыми.
Первая модификация может представлять собой 2'-OMe, а вторая модификация может представлять собой 2'-F.
В нуклеиновой кислоте согласно второму аспекту первая цепь может содержать нуклеотидную последовательность SEQ ID NO: 5 и SEQ ID NO: 9, и вторая цепь может содержать нуклеотидную последовательность SEQ ID NO: 6 и SEQ ID NO: 10.
Последовательность и модификации могут представлять собой те, которые показаны в таблице ниже; где представлены предпочтительные последовательности на основе выдержки из таблицы 1, представленной в настоящей заявке:
где конкретные модификации обозначены следующими номерами
1=2'-F-dU,
2=2'-F-dA,
3=2'-F-dC,
4=2'-F-dG,
5=2'-OMe-rU;
6=2'-OMe-rA;
7=2'-OMe-rC;
8=2'-OMe-rG.
Нуклеиновая кислота согласно настоящему изобретению может содержать фосфотиоатную связь между одним, двумя или тремя концевыми 3'-нуклеотидами и/или одним, двумя или тремя 5'-нуклеотидами первой цепи и/или второй цепи. Она может содержать две фосфотиоатные связи между каждым из трех концевых 3'-нуклеотидов и между каждым из трех концевых 5'-нуклеотидов на первой цепи и две фосфотиоатные связи между тремя концевыми нуклеотидами 3'-конца второй цепи.
Такая нуклеиновая кислота может быть конъюгирована с лигандом.
Согласно настоящему изобретению также предложена, в качестве третьего аспекта, нуклеиновая кислота для ингибирования экспрессии LPA в клетке, содержащая по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, причем указанная первая цепь содержит нуклеотидную последовательность, выбранную из следующих последовательностей: SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73, и при этом указанная нуклеиновая кислота конъюгирована с лигандом.
Лиганд может содержать (i) одну или более групп N-ацетилгалактозамина (GalNAc) и его производных и (ii) линкер, причем с помощью указанного линкера конъюгируют группы GalNAc с последовательностью, определенной в любых предыдущих аспектах. Линкер может иметь двухвалентную, трехвалентную или четырехвалентную разветвленную структуру. Нуклеотиды могут быть модифицированы, как определено в настоящей заявке.
Лиганд может содержать формулу I:
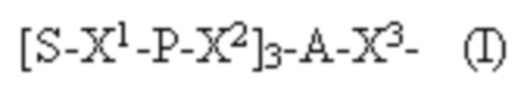
в которой:
S представляет собой сахарид, причем указанный сахарид представляет собой N-ацетилгалактозамин;
X1 представляет собой C3-C6 алкилен или (-CH2-CH2-O)m(-CH2)2-, где m равно 1, 2 или 3;
P представляет собой фосфат или модифицированный фосфат (предпочтительно тиофосфат);
X2 представляет собой алкилен или простой алкиленовый эфир формулы (-CH2)n-O-CH2-, где n=1-6;
А представляет собой блок разветвления;
X3 представляет собой мостиковый блок;
при этом нуклеиновая кислота в соответствии с настоящим изобретением конъюгирована с X3 за счет фосфата или модифицированного фосфата (предпочтительно тиофосфата).
Таким образом, согласно настоящему изобретению дополнительно предложена конъюгированная нуклеиновая кислота, имеющая одну из следующих структур

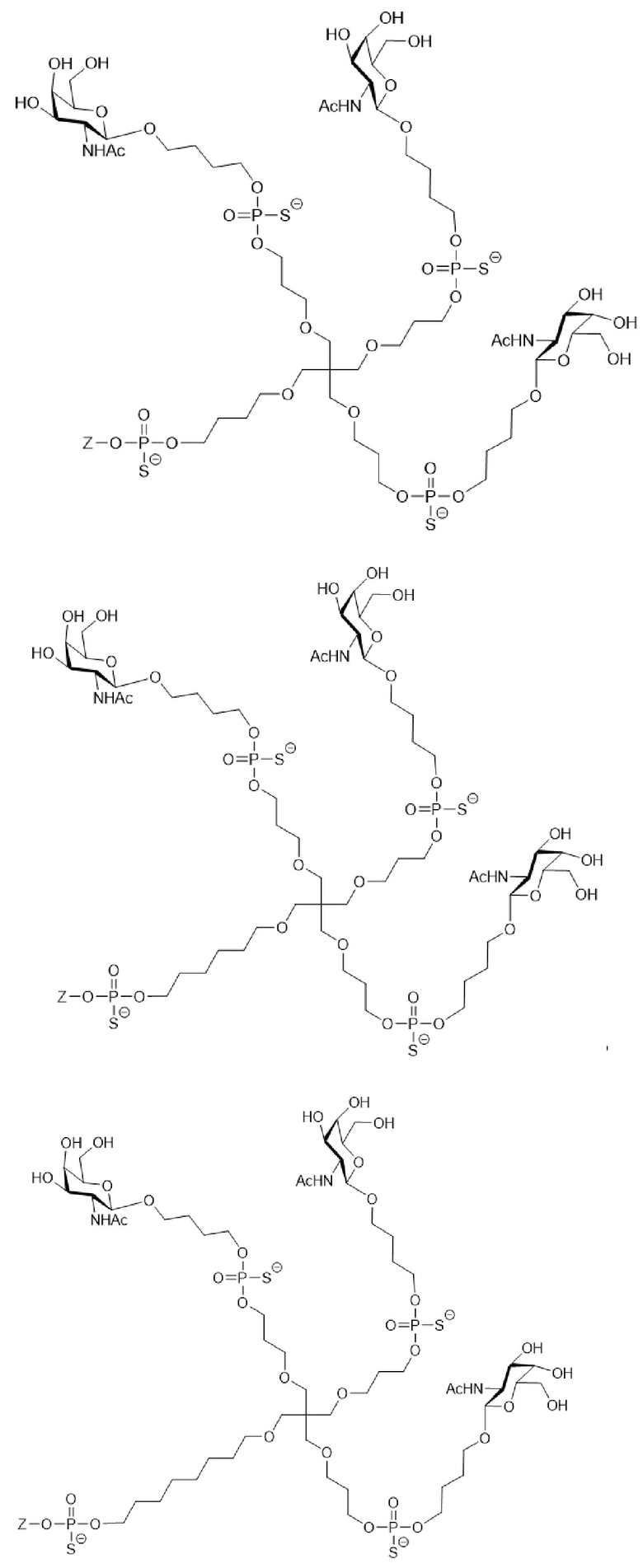
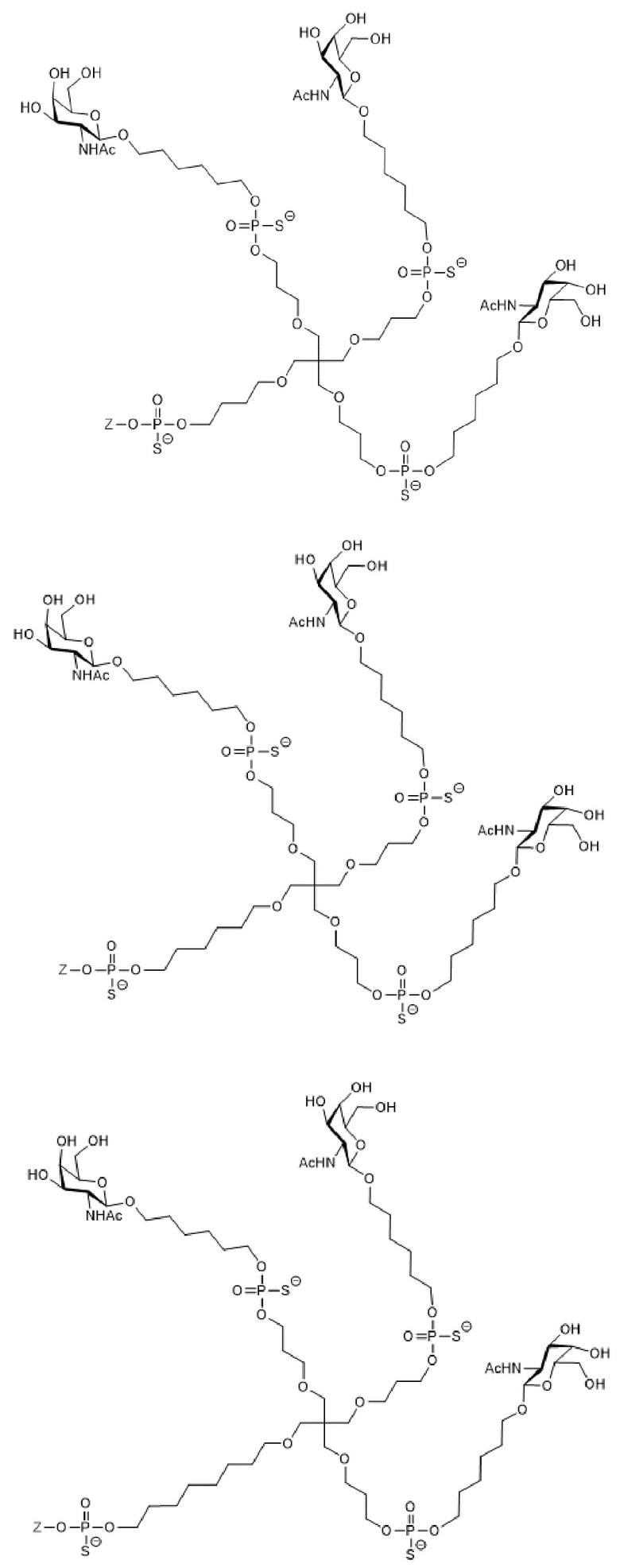
где Z представляет собой нуклеиновую кислоту, определенную в настоящей заявке ранее.
Согласно другому варианту нуклеиновая кислота в соответствии с настоящим изобретением может быть конъюгирована с лигандом, имеющим следующую структуру
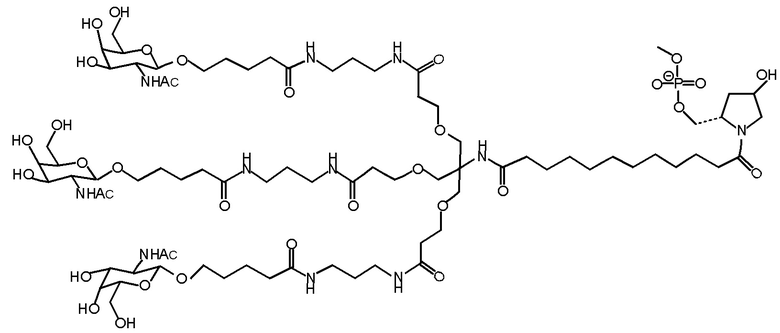
Настоящее изобретение также относится к конъюгату для ингибирования экспрессии гена LPA в клетке, причем указанный конъюгат содержит часть нуклеиновой кислоты, содержащую нуклеиновую кислоту согласно любому аспекту настоящего изобретения, и части лиганда, причем указанная часть нуклеиновой кислоты содержит по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи РНК и по меньшей мере часть второй цепи РНК, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с указанного гена LPA, при этом указанные части лиганда содержат линкерную группу, предпочтительно линкерную группу, происходящую из серинола, а также нацеливающий лиганд для нацеливания в условиях in vivo на клетки и конъюгированный исключительно с 3' и/или 5'-концами одной или обеих цепей РНК, причем 5'-конец первой цепи РНК не конъюгирован, при этом:
(i) вторая цепь РНК конъюгирована на 5'-конце с нацеливающим лигандом, и при этом (a) вторая цепь РНК также конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец первой цепи РНК не конъюгирован; или (b) первая цепь РНК конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец второй цепи РНК не конъюгирован; или (c) обе вторая цепь РНК и первая цепь РНК также конъюгированы на 3'-концах с нацеливающим лигандом; или
(ii) обе вторая цепь РНК и первая цепь РНК конъюгированы на 3'-концах с нацеливающим лигандом и 5'-конец второй цепи РНК не конъюгирован, или
настоящее изобретение относится к конъюгату для ингибирования экспрессии гена LPA в клетке, причем указанный конъюгат содержит часть нуклеиновой кислоты и части лиганда, причем указанная часть нуклеиновой кислоты содержит по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи РНК и по меньшей мере часть второй цепи РНК, которая по меньшей мере частично комплементарна первой цепи, при этом указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с указанного гена LPA, при этом указанные части лиганда содержат линкерную группу и нацеливающий лиганд для нацеливания в условиях in vivo на клетки и конъюгированный исключительно с 3' и/или 5'-концами одной или обеих цепей РНК, причем 5'-конец первой цепи РНК не конъюгирован, при этом:
(i) вторая цепь РНК конъюгирована на 5'-конце с нацеливающим лигандом, и при этом (a) вторая цепь РНК также конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец первой цепи РНК не конъюгирован; или (b) первая цепь РНК конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец второй цепи РНК не конъюгирован; или (c) обе вторая цепь РНК и первая цепь РНК также конъюгированы на 3'-концах с нацеливающим лигандом; или
(ii) обе вторая цепь РНК и первая цепь РНК конъюгированы на 3'-концах с нацеливающим лигандом и 5'-конец второй цепи РНК не конъюгирован.
Линкерная группа может представлять собой, например, линкерную группу, происходящую из серинола, или один из других типов линкеров, описанных в настоящей заявке.
Согласно настоящему изобретению также предложена композиция, содержащая нуклеиновую кислоту или конъюгированную нуклеиновую кислоту согласно любому аспекту настоящего изобретения и физиологически приемлемое вспомогательное вещество.
Также предложена нуклеиновая кислота или конъюгированная нуклеиновая кислота в соответствии с любым аспектом настоящего изобретения для применения при лечении заболевания, нарушения или синдрома и/или при изготовлении лекарственного средства для лечения заболевания, нарушения или синдрома.
Согласно настоящему изобретению предложен способ лечения или предотвращения заболевания, нарушения или синдрома, включающий введение композиции, содержащей нуклеиновую кислоту или конъюгированную нуклеиновую кислоту в соответствии с любым аспектом настоящего изобретения индивидууму, нуждающемуся в лечении. Нуклеиновую кислоту или конъюгированную нуклеиновую кислоту можно вводить субъекту подкожно, внутривенно или с помощью любых других путей применения, таких как пероральный, ректальный или внутрибрюшинный.
После подкожного применения объект изобретения может быть доставлен тканеспецифичным способом в печень (гепатоциты) и нацелен конкретно на LPA, чтобы уменьшить нежелательные побочные эффекты и достичь более низкой терапевтической дозы, необходимой для достижения целевого эффекта.
Настоящее изобретение или фармацевтическую композицию, содержащую нуклеиновую кислоту или конъюгированную нуклеиновую кислоту согласно настоящему изобретению, можно применять при лечении заболевания, нарушения или синдрома. Лечение может заключаться в предотвращении и уменьшении риска развития инсульта, атеросклероза, тромбоза или сердечно-сосудистых заболеваний, таких как ишемическая болезнь сердца или стеноз аорты, и любого другого заболевания или патологии, ассоциированных с повышенными уровнями частиц, содержащих Lp(a).
Также предусмотрен способ получения нуклеиновой кислоты или конъюгированной нуклеиновой кислоты в соответствии с настоящим изобретением.
Подробное описание изобретения
Настоящее изобретение относится к нуклеиновой кислоте, которая является двухцепочечной и направлена к экспрессированному транскрипту РНК LPA, и ее композициям. Эти нуклеиновые кислоты или конъюгированные нуклеиновые кислоты можно применять при лечении и предотвращении различных заболеваний, нарушений и синдромов, при которых желательна уменьшенная экспрессия продукта гена LPA.
Первый аспект настоящего изобретения относится к нуклеиновой кислоте для ингибирования экспрессии LPA в клетке, содержащей по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, при этом указанная первая цепь содержит нуклеотидную последовательность, выбранную из следующих последовательностей: SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73.
Нуклеиновая кислота может содержать первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 9, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 10; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 5, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 6; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 1, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 2; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 4, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 4; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 7, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 8; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 11, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 12; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 13, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 14; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 15, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 16; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 17, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 18; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 19, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 20; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 21, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 22; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 23, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 24; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 25, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 26; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 27, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 28; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 29, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 30; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 31, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 32; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 33, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 34; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 35, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 36; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 37, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 38; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 39, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 40; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 41, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 42; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 43, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 44; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 63, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 64; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 65, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 66; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 67, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 68; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 69, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 70; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 71, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 72; или первую цепь, которая содержит нуклеотидную последовательность SEQ ID NO: 73, и при этом необязательно вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 74.
Ген LPA содержит последовательности с очень большим количеством повторов. Следовательно, нуклеиновые кислоты первой цепи с очень сходными последовательностями могут иметь идеальную комплементарность последовательностей с очень разными целевыми областями мРНК.
Родственным аспектом настоящего изобретения является нуклеиновая кислота для ингибирования экспрессии LPA в клетке, причем указанная нуклеиновая кислота содержит по меньшей мере одну дуплексную область, которая содержит: первую цепь; и вторую цепь, причем указанная вторая цепь по меньшей мере частично комплементарна первой цепи, при этом указанная первая цепь содержит последовательность из по меньшей мере 15, предпочтительно по меньшей мере 16, более предпочтительно по меньшей мере 17, еще более предпочтительно по меньшей мере 18 и наиболее предпочтительно по меньшей мере 19 нуклеотидов любой из эталонных последовательностей SEQ ID NO: 9, 5, 1, 3, 7, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73, и при этом количество несовпадений отдельных нуклеотидов и/или делеций и/или вставок в последовательности первой цепи относительно части эталонной последовательности, которая содержится в последовательности первой цепи, составляет не более трех, предпочтительно не более двух, более предпочтительно не более одного и наиболее предпочтительно ноль.
Согласно одному аспекту первая цепь нуклеиновой кислоты содержит последовательность из по меньшей мере 18 нуклеотидов любой из эталонных последовательностей, предпочтительно любой из эталонных последовательностей SEQ ID NO: 9 и 5, и при этом количество несоответствий отдельных нуклеотидов и/или делеций и/или вставок в последовательности первой цепи относительно части эталонной последовательности, которая содержится в последовательности первой цепи, составляет не более одного, а предпочтительно ноль.
Согласно одному аспекту первая цепь нуклеиновой кислоты содержит последовательность из по меньшей мере 19 нуклеотидов любой из эталонных последовательностей SEQ ID NO: 9 и 5.
Определенное количество несовпадений, делеций или вставок между первой (антисмысловой) цепью и последовательностью-мишенью или между первой цепью и второй (смысловой) цепью может быть допустимым применительно к миРНК и в некоторых случаях даже может увеличивать активность.
Под нуклеиновой кислотой подразумевают нуклеиновую кислоту, содержащую две цепи, содержащие нуклеотиды, которая способна препятствовать экспрессии генов. Ингибирование может быть полным или частичным и приводит к снижению экспрессии генов нацеленным образом. Нуклеиновая кислота содержит две отдельные полинуклеотидные цепи; первую цепь, которая также может быть направляющей цепью; и вторую цепь, которая также может быть пассажирской цепью. Первая цепь и вторая цепь могут быть частью одной и той же полинуклеотидной молекулы, которая является самокомплементарной и «складывается», разворачиваясь назад, с образованием двухцепочечной молекулы. Нуклеиновая кислота может представлять собой молекулу миРНК.
Нуклеиновая кислота может содержать рибонуклеотиды, модифицированные рибонуклеотиды, дезоксинуклеотиды, дезоксирибонуклеотиды или ненуклеотидные аналоги нуклеотидов, которые способны имитировать нуклеотиды так, что они могут «спариваться» с соответствующим основанием на последовательности-мишени или комплементарной цепи. Нуклеиновая кислота может дополнительно содержать двухцепочечную часть нуклеиновой кислоты или дуплексную область, образованную всей первой цепью или ее частью (также известной в данной области техники как направляющая цепь) и всей второй цепью или ее частью (также известной в данной области техники как пассажирская цепь). Дуплексная область определяется как область, начинающаяся с первой пары оснований, образованной между первой цепью и второй цепью, и заканчивающаяся последней парой оснований, образованной между первой цепью и второй цепью, включительно.
Под дуплексной областью подразумевают область в двух комплементарных или по существу комплементарных олигонуклеотидах, которые образуют пары оснований друг с другом, либо путем спаривания оснований по Уотсону-Крику, либо любым другим способом, который обеспечивает образование дуплекса между олигонуклеотидными цепями, которые являются комплементарными или по существу комплементарными. Например, олигонуклеотидная цепь, содержащая 21 нуклеотидный блок, может спариваться с другим олигонуклеотидом из 21 нуклеотидного блока, но только 19 нуклеотидов на каждой цепи являются комплементарными или по существу комплементарными так, что «дуплексная область» состоит из 19 пар оснований. Оставшиеся пары оснований могут существовать в виде 5'- и 3'-липких концов или в виде одноцепочечных областей. Кроме того, в дуплексной области не требуется 100% комплементарность; в дуплексной области допустима существенная комплементарность. Существенная комплементарность относится к такой комплементарности между цепями, что они способны ренатурировать в биологических условиях. Методики, позволяющие эмпирически определить, способны ли две цепи ренатурировать в биологических условиях, хорошо известны в данной области техники. Согласно другому варианту две цепи могут быть синтезированы и добавлены вместе в биологических условиях, чтобы определить, ренатурируют ли они друг с другом.
Части первой цепи и второй цепи, которые образуют по меньшей мере одну дуплексную область, могут быть полностью комплементарны и по меньшей мере частично комплементарны друг другу. В зависимости от длины нуклеиновой кислоты не обязательно требуется идеальное соответствие между первой цепью и второй цепью с точки зрения комплементарности оснований. Однако первая и вторая цепи должны быть способны гибридизоваться в физиологических условиях.
Комплементарность между первой цепью и второй цепью по меньшей мере в одной дуплексной области может быть идеальной в том, что в любой цепи отсутствуют несоответствия нуклеотидов или дополнительные/удаленные нуклеотиды. Согласно другому варианту комплементарность может быть неидеальной. Комплементарность может составлять от приблизительно 70% до приблизительно 100%. Более конкретно, комплементарность может составлять по меньшей мере 70%, 75%, 80%, 85%, 90% или 95% и промежуточные значения.
Применительно к настоящему изобретению «часть», например, в «одной дуплексной области, которая содержит по меньшей мере часть первой цепи», следует понимать как то, что дуплексная область содержит по меньшей мере 10, предпочтительно по меньшей мере 12, более предпочтительно по меньшей мере 14, еще более предпочтительно по меньшей мере 16, даже более предпочтительно по меньшей мере 18 и наиболее предпочтительно все из нуклеотидов данной эталонной последовательности цепи. Часть эталонной последовательности в дуплексной области по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90%, еще более предпочтительно по меньшей мере на 95% и наиболее предпочтительно на 100% идентична соответствующей части эталонной последовательности. Согласно другому варианту количество несоответствий отдельных нуклеотидов относительно части эталонной последовательности составляет не более трех, предпочтительно не более двух, более предпочтительно не более одного и наиболее предпочтительно ноль.
Первая цепь и вторая цепь могут каждая содержать область комплементарности, которая содержит по меньшей мере 15 смежных нуклеотидов, отличающуюся не более чем 3 нуклеотидами от любой из последовательностей, перечисленных в таблице 1.
Нуклеиновая кислота может содержать вторую последовательность, содержащую нуклеотидную последовательность SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28 30, 32, 34, 36, 38, 40, 42, 44, 64, 66, 68, 70, 72 или 74.
Применение нуклеиновой кислоты в соответствии с настоящим изобретением включает образование дуплексной области между всей первой цепь или ее частью и частью нуклеиновой кислоты-мишени. Часть нуклеиновой кислоты-мишени, которая образует дуплексную область с первой цепью, определяемую как область, которая начинается с первой пары оснований, образованной между первой цепью и последовательностью-мишенью, и заканчивается последней парой оснований, образованной между первой цепью и последовательностью-мишенью, включительно, представляет собой последовательность нуклеиновой кислоты-мишени или просто последовательность-мишень. Дуплексная область, образованная между первой цепью и второй цепью, необязательно должна быть аналогична дуплексной области, образованной между первой цепью и последовательностью-мишенью. Это означает, что вторая цепь может иметь последовательность, отличающуюся от последовательности-мишени; однако первая цепь должна быть способна образовывать дуплексную структуру с обеими второй цепью и последовательностью-мишенью, по меньшей мере в физиологических условиях.
Комплементарность между первой цепью и последовательностью-мишенью может быть идеальной (в каждой нуклеиновой кислоте отсутствуют несовпадения нуклеотидов или дополнительные/удаленные нуклеотиды).
Комплементарность между первой цепью и последовательностью-мишенью может быть неидеальной. Комплементарность может составлять от приблизительно 70% до приблизительно 100%. Более конкретно, комплементарность может составлять по меньшей мере 70%, 80%, 85%, 90% или 95% и промежуточные значения.
Идентичность между первой цепью и комплементарной последовательностью последовательности-мишени может варьироваться от приблизительно 75% до приблизительно 100%. Более конкретно, комплементарность может составлять по меньшей мере 75%, 80%, 85%, 90% или 95% и промежуточные значения, при условии, что нуклеиновая кислота способна уменьшать или ингибировать экспрессию LPA.
Нуклеиновая кислота, имеющая менее чем 100% комплементарность между первой цепью и последовательностью-мишенью, может быть способна уменьшить экспрессию LPA до того же уровня, как и нуклеиновая кислота, имеющая идеальную комплементарность между первой цепью и последовательностью-мишенью. Согласно другому варианту она может быть способна уменьшить экспрессию LPA до уровня, который составляет 15-100% от уровня уменьшения, достигнутого с помощью нуклеиновой кислоты с идеальной комплементарностью.
Нуклеиновая кислота может содержать первую цепь и вторую цепь, каждая из которых содержит 19-25 нуклеотидов в длину. Первая цепь и вторая цепь могут иметь разную длину.
Нуклеиновая кислота может содержать 15-25 пар нуклеотидов в длину. Нуклеиновая кислота может содержать 17-23 пары нуклеотидов в длину. Нуклеиновая кислота может содержать 17-25 пар нуклеотидов в длину. Нуклеиновая кислота может содержать 23-24 пары нуклеотидов в длину. Нуклеиновая кислота может содержать 19-21 пару нуклеотидов в длину. Нуклеиновая кислота может содержать 21-23 пары нуклеотидов в длину.
Нуклеиновая кислота может содержать дуплексную область, которая состоит из 19-25 пар нуклеотидных оснований. Дуплексная область может состоять из 17, 18, 19, 20, 21, 22, 23, 24 или 25 пар оснований, которые могут быть смежными.
Нуклеиновая кислота может содержать последовательность первой цепи SEQ ID NO: 5 или SEQ ID NO: 9. Нуклеиновая кислота может содержать последовательность второй цепи SEQ ID NO: 6 или SEQ ID NO: 10.
Предпочтительно нуклеиновая кислота опосредует РНК-интерференцию.
Согласно одному варианту реализации настоящего изобретения нуклеиновая кислота для ингибирования экспрессии LPA в клетке содержит по меньшей мере одну дуплексную область, которая содержит первую цепь и вторую цепь, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь содержит последовательность из по меньшей мере 15, предпочтительно по меньшей мере 16, более предпочтительно по меньшей мере 17, еще более предпочтительно по меньшей мере 18 и наиболее предпочтительно по меньшей мере 19 нуклеотидов, при этом последовательность по меньшей мере на 70%, предпочтительно по меньшей мере на 80%, более предпочтительно по меньшей мере на 90%, еще более предпочтительно по меньшей мере на 95% и наиболее предпочтительно на 100% идентична любой из последовательностей SEQ ID NO: 9, 5, 1, 3, 7, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73.
Согласно дополнительному аспекту описанная нуклеиновая кислота или конъюгированная нуклеиновая кислота может уменьшать экспрессию LPA по меньшей мере на 15% по сравнению с экспрессией, наблюдаемой в отсутствие нуклеиновой кислоты или конъюгированной нуклеиновой кислоты. Все предпочтительные признаки любого из предыдущих аспектов также применимы к этому аспекту. В частности, экспрессия LPA может быть уменьшена по меньшей мере до следующих заданных процентных значений (%) или до менее чем 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 15% или менее и промежуточных значений относительно экспрессии, которая наблюдалась в отсутствие нуклеиновой кислоты или конъюгированной нуклеиновой кислоты или в присутствии неподавляющего контроля.
Нуклеиновая кислота может содержать тупые концы на обоих концах; может содержать липкий конец на одном конце и тупой конец на другом конце; или содержать липкий конец на обоих концах.
В настоящей заявке «липкий конец (“overhang”)» имеет свое обычное и принятое значение в данной области техники, т.е. одноцепочечная часть нуклеиновой кислоты, которая простирается за пределы концевого нуклеотида комплементарной цепи в двухцепочечной нуклеиновой кислоте. Термин «тупой конец» включает двухцепочечную нуклеиновую кислоту, в которой обе цепи оканчиваются в одном и том же положении, независимо от того, спарен (ы) ли концевой (ые) нуклеотид (ы). Концевой нуклеотид первой цепи и второй цепи на тупом конце может быть спаренным. Концевой нуклеотид первой цепи и второй цепи на тупом конце может быть неспаренным. Два концевых нуклеотида первой цепи и второй цепи на тупом конце могут быть спаренными. Два концевых нуклеотида первой цепи и второй цепи на тупом конце могут быть неспаренными.
Нуклеиновая кислота может содержать липкий конец на одном конце и тупой конец на другом. Нуклеиновая кислота может содержать липкий конец на обоих концах. Нуклеиновая кислота может содержать тупые концы на обоих концах. Нуклеиновая кислота может иметь тупой конец на конце с 5'-концом первой цепи и 3'-концом второй цепи или на 3'-конце первой цепи и 5'-конце второй цепи.
Нуклеиновая кислота может содержать липкий конец на 3'- или 5'-конце. Нуклеиновая кислота может содержать 3'-липкий конец на первой цепи. Нуклеиновая кислота может содержать 3'-липкий конец на второй цепи. Нуклеиновая кислота может содержать 5'-липкий конец на первой цепи. Нуклеиновая кислота может содержать 5'-липкий конец на второй цепи. Нуклеиновая кислота может содержать липкий конец как на 5'-конце, так и на 3'-конце первой цепи. Нуклеиновая кислота может содержать липкий конец как на 5'-конце, так и на 3'-конце второй цепи. Нуклеиновая кислота может содержать 5'-липкий конец на первой цепи и 3'-липкий конец на второй цепи. Нуклеиновая кислота может содержать 3'-липкий конец на первой цепи и 5'-липкий конец на второй цепи. Нуклеиновая кислота может содержать 3'-липкий конец на первой цепи и 3'-липкий конец на второй цепи. Нуклеиновая кислота может содержать 5'-липкий конец на первой цепи и 5'-липкий конец на второй цепи.
Липкий конец на 3'-конце или 5'-конце второй цепи или первой цепи может быть выбран из 1, 2, 3, 4 и 5 нуклеотидов в длину. Необязательно липкий конец может состоять из 1 или 2 нуклеотидов, которые могут быть модифицированы, но необязательно.
Немодифицированные полинуклеотиды, в частности, рибонуклеотиды, могут быть подвержены разрушению клеточными нуклеазами, и, как таковые, модификации/модифицированные нуклеотиды могут быть включены в нуклеиновую кислоту согласно настоящему изобретению. Такие модификации могут помочь стабилизировать нуклеиновую кислоту, делая ее более устойчивой к нуклеазам. Улучшенная устойчивость обеспечивает активность нуклеиновых кислот в качестве посредников РНК-интерференции в течение более длительных периодов времени и особенно желательна, если нуклеиновые кислоты должны применяться для лечения.
Один или более нуклеотидов на второй цепи и/или первой цепи нуклеиновой кислоты согласно настоящему изобретению могут быть модифицированы.
Модификации нуклеиновой кислоты согласно настоящему изобретению обычно обеспечивают эффективный инструмент для преодоления потенциальных ограничений, включая, но не ограничиваясь этим, стабильность и биодоступность в условиях in vitro и в условиях in vivo, присущие природным молекулам РНК. Нуклеиновая кислота в соответствии с настоящим изобретением может быть модифицирована с помощью химических модификаций. Модифицированная нуклеиновая кислота также может свести к минимуму возможность индукции активности интерферона у человека. Модификация может дополнительно усиливать функциональную доставку нуклеиновой кислоты к клетке-мишени. Модифицированная нуклеиновая кислота согласно настоящему изобретению может содержать один или более химически модифицированных рибонуклеотидов любой или обеих из первой цепи или второй цепи. Рибонуклеотид может содержать химическую модификацию основания, сахара или фосфатных групп. Рибонуклеиновая кислота может быть модифицирована путем замены или вставки аналогов нуклеиновых кислот или оснований.
Один или более нуклеотидов нуклеиновой кислоты согласно настоящему изобретению могут быть модифицированы. Нуклеиновая кислота может содержать по меньшей мере один модифицированный нуклеотид. Модифицированный нуклеотид может находиться в первой цепи. Модифицированный нуклеотид может находиться во второй цепи. Модифицированный нуклеотид может находиться в дуплексной области. Модифицированный нуклеотид может находиться вне дуплексной области, т.е. в одноцепочечной области. Модифицированный нуклеотид может находиться на первой цепи и может находиться за пределами дуплексной области. Модифицированный нуклеотид может находиться на второй цепи и может находиться за пределами дуплексной области. 3'-концевой нуклеотид первой цепи может представлять собой модифицированный нуклеотид. 3'-концевой нуклеотид второй цепи может представлять собой модифицированный нуклеотид. 5'-концевой нуклеотид первой цепи может представлять собой модифицированный нуклеотид. 5'-концевой нуклеотид второй цепи может представлять собой модифицированный нуклеотид.
Нуклеиновая кислота согласно настоящему изобретению может содержать 1 модифицированный нуклеотид или нуклеиновая кислота согласно настоящему изобретению может содержать приблизительно 2-4 модифицированных нуклеотида, или нуклеиновая кислота может содержать приблизительно 4-6 модифицированных нуклеотидов, приблизительно 6-8 модифицированных нуклеотидов, приблизительно 8-10 модифицированных нуклеотидов, приблизительно 10-12 модифицированных нуклеотидов, приблизительно 12-14 модифицированных нуклеотидов, приблизительно 14-16 модифицированных нуклеотидов, приблизительно 16-18 модифицированных нуклеотидов, приблизительно 18-20 модифицированных нуклеотидов, приблизительно 20-22 модифицированных нуклеотида, приблизительно 22-24 модифицированных нуклеотида, 24-26 модифицированных нуклеотидов или приблизительно 26-28 модифицированных нуклеотидов. В каждом случае нуклеиновая кислота, содержащая указанные модифицированные нуклеотиды, сохраняет по меньшей мере 50% своей активности по сравнению с этой же нуклеиновой кислотой, но без указанных модифицированных нуклеотидов или наоборот. Нуклеиновая кислота может сохранять 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100%, включая промежуточные значения, своей активности по сравнению с этой же нуклеиновой кислотой, но без указанных модифицированных нуклеотидов, или может иметь более 100% активности этой же нуклеиновой кислоты без указанных модифицированных нуклеотидов.
Модифицированный нуклеотид может представлять собой пурин или пиримидин. По меньшей мере половина пуринов может быть модифицирована. По меньшей мере половина пиримидинов может быть модифицирована. Все из пуринов могут быть модифицированы. Все из пиримидинов могут быть модифицированы. Модифицированные нуклеотиды могут быть выбраны из группы, состоящей из 3'-концевого дезокситиминового (dT) нуклеотида, 2'-О-метил-модифицированного нуклеотида, 2'-модифицированного нуклеотида, 2'-дезокси-модифицированного нуклеотида, закрытого нуклеотида, лишенного азотистого основания нуклеотида, 2'-амино-модифицированного нуклеотида, 2'-алкил-модифицированного нуклеотида, морфолино-нуклеотида, фосфоамидата, содержащего неприродное основание нуклеотида, нуклеотида, содержащего 5'-фосфотиоатную группу, нуклеотида, содержащего 5'-фосфат или миметик 5'-фосфата, и концевого нуклеотида, соединенного с производным холестерина или группой бисдециламида додекановой кислоты.
Нуклеиновая кислота может содержать нуклеотид, содержащий модифицированный нуклеотид, причем основание выбрано из 2-аминоаденозина, 2,6-диаминопурина, инозина, пиридин-4-она, пиридин-2-она, фенила, псевдоурацила, 2,4,6-триметоксибензола, 3-метилурацила, дигидроуридина, нафтила, аминофенила, 5-алкилцитидина (например, 5-метилцитидина), 5-алкилуридина (например, риботимидина), 5-галоуридина (например, 5-бромуридина), 6-азапиримидина, 6-алкилпиримидина (например, 6-метилуридина), пропина, квеуозина, 2-тиоуридина, 4-тиоуридина, вайбутозина, вайбутоксозина, 4-ацетилцитидина, 5-(карбоксигидроксиметил)уридина, 5'-карбоксиметиламинометил-2-тиоуридина, 5-карбоксиметиламинометилуридина, бета-D-галактозилквеуозина, 1-метиладенозина, 1-метилинозина, 2,2-диметилгуанозина, 3-метилцитидина, 2-метиладенозина, 2-метилгуанозина, N6-метиладенозина, 7-метилгуанозина, 5-метоксиаминометил-2-тиоуридина, 5-метиламинометилуридина, 5-метилкарбонилметилуридина, 5-метилоксиуридина, 5-метил-2-тиоуридина, 2-метилтио-N6-изопентениладенозина, бета-D-маннозилквеуозина, уридин-5-оксиуксусной кислоты и 2-тиоцитидина.
Нуклеиновые кислоты, обсуждаемые в настоящей заявке, включают немодифицированную РНК, а также РНК, которая была модифицирована, например, для улучшения эффективности, и полимеры нуклеозидных суррогатов. Немодифицированная РНК относится к молекуле, в которой компоненты нуклеиновой кислоты, а именно сахара, основания и фосфатные группы, являются такими же или по существу такими же, как и те, которые встречаются в природе, например, которые встречаются в природе в организме человека. В настоящей заявке модифицированный нуклеотид относится к нуклеотиду, в котором один или более из компонентов нуклеотидов, а именно сахара, основания и фосфатные группы, отличаются от тех, которые встречаются в природе. Несмотря на то, что модифицированные нуклеотиды так называются из-за своей модификации, термин также включает молекулы, которые не являются нуклеотидами, например, полинуклеотидную молекулу, в которой рибофосфатный остов заменен нерибофосфатной конструкцией, которая обеспечивает гибридизацию между цепями, т.е. модифицированные нуклеотиды имитируют рибофосфатный остов.
Многие из описанных ниже модификаций, которые встречаются в нуклеиновой кислоте, будут повторены в полинуклеотидной молекуле, например, модификация основания или фосфатной группы, или несоединяющего О из фосфатной группы. В некоторых случаях модификация будет встречаться во всех из возможных положений/нуклеотидов в полинуклеотиде, но во многих случаях не будет встречаться. Модификация может встречаться только в 3'- или 5'-концевом положении, может встречаться только в концевых областях, например, в положении на концевом нуклеотиде или в последних 2, 3, 4, 5 или 10 нуклеотидах цепи. Модификация может встречаться в двухцепочечной области, одноцепочечной области или в обеих. Модификация может встречаться только в двухцепочечной области нуклеиновой кислоты согласно настоящему изобретению или может встречаться только в одноцепочечной области нуклеиновой кислоты согласно настоящему изобретению. Фосфотиоатная модификация в положении несоединяющего O может встречаться только на одном или обоих концах, может встречаться только в концевой области, например, в положении на концевом нуклеотиде или в последних 2, 3, 4 или 5 нуклеотидах или может встречаться в дуплексе и/или в одноцепочечных областях, в частности, на концах. 5'-Конец или 3'-конец могут быть фосфорилированы.
Стабильность нуклеиновой кислоты согласно настоящему изобретению может быть повышена путем включения конкретных оснований в липких концах или включения модифицированных нуклеотидов, в одноцепочечных липких концах, например, в 5' или 3'-липком конце или в обоих. В липкие концы могут быть включены пуриновые нуклеотиды. Все или некоторые из оснований в 3'- или 5'-липком конце могут быть модифицированы. Модификации могут включать применение модификаций в группе 2'-OH рибозного сахара, применение дезоксирибонуклеотидов вместо рибонуклеотидов и модификации в фосфатной группе, такие как фосфотиоатные модификации. Липкие концы необязательно должны быть гомологичны последовательности-мишени.
Нуклеазы могут гидролизовать фосфодиэфирные связи нуклеиновых кислот. Однако химические модификации нуклеиновых кислот могут придавать улучшенные свойства и могут сделать олигорибонуклеотиды более устойчивыми к нуклеазам.
В настоящей заявке модифицированные нуклеиновые кислоты могут включать одно или более из следующего:
(i) изменение, например, замещение одного или обоих из несоединяющих атомов кислорода фосфата и/или одного или более из соединяющих атомов кислорода фосфата (упоминается как событие соединения, даже если происходит на 5'- и 3'-конце нуклеиновой кислоты согласно настоящему изобретению);
(ii) изменение, например, замещение компонента рибозного сахара, например, 2'-гидроксила на рибозном сахаре;
(iii) замещение фосфатной группы «дефосфо» линкерами;
(iv) модификация или замещение природного основания;
(v) замещение или модификация рибозофосфатного остова;
(vi) модификация 3'-конца или 5'-конца РНК, например, удаление, модификация или замещение концевой фосфатной группы или конъюгация группы, например, флуоресцентномеченой группы, с 3'- или 5'-концом РНК.
Термины замещение, модификация, изменение указывают на отличие от природной молекулы.
Конкретные модификации более подробно обсуждаются ниже.
Примеры модифицированных фосфатных групп включают фосфотиоат, фосфоселенаты, боранофосфаты, сложные эфиры боранофосфата, фосфонаты водорода, фосфоамидаты, алкил или арилфосфонаты и фосфотриэфиры. В фосфодитиоатах оба несоединяющих атома кислорода замещены серой. Один, каждый или оба несоединяющих атома кислорода в фосфатной группе могут быть независимо замещены любым из S, Se, B, C, H, N или OR (R представляет собой алкил или арил).
Фосфатный линкер также можно модифицировать путем замещения соединяющего атома кислорода азотом (мостиковые фосфоамидаты), серой (мостиковые фосфотиоаты) и углеродом (мостиковые метиленфосфонаты). Замещение может происходить у концевого кислорода. Возможно замещение несоединяющих атомов кислорода азотом.
Модифицированный нуклеотид может содержать модификацию сахарных групп. 2'-гидроксильная группа (OH) может быть модифицирована или замещена рядом различных «окси» или «дезокси» заместителей.
Примеры модификаций «окси»-2'-гидроксильная группа включают алкокси или арилокси (OR, например, R=H, алкил, циклоалкил, арил, аралкил, гетероарил или сахар); полиэтиленгликоли (ПЭГ), O(CH2CH2O)nCH2CH2OR; «закрытые» нуклеиновые кислоты (LNA), в которых 2'-гидроксил соединен, например, метиленовым мостиком, с 4'-углеродом этого же рибозного сахара; O-АМИН (АМИН=NH2; алкиламино, диалкиламино, гетероциклил, ариламино, диариламино, гетероариламино или дигетероариламино, этилендиамин, полиамино) и аминоалкокси, O(CH2)nАМИН, (например, АМИН=NH2; алкиламино, диалкиламино, гетероциклил, ариламино, диариламино, гетероариламино или дигетероариламино, этилендиамин, полиамино).
«Дезокси» модификации включают водород, галоген, амино (например, NH2; алкиламино, диалкиламино, гетероциклил, ариламино, диариламино, гетероариламино, дигетероариламино или аминокислоту); NH(CH2CH2NH)nCH2CH2-АМИН (АМИН=NH2; алкиламино, диалкиламино, гетероциклил, ариламино, диариламино, гетероариламино или дигетероариламино), -NHC(O)R (R = алкил, циклоалкил, арил, аралкил, гетероарил или сахар), циано; меркапто; алкил-тио-алкил; тиоалкокси; и алкил, циклоалкил, арил, алкенил и алкинил, которые могут быть необязательно замещены, например, функциональной аминогруппой. Другие заместители из определенных вариантов реализации включают 2'-метоксиэтил, 2'-OCH3, 2'-O-аллил, 2'-C-аллил и 2'-фтор. Сахарная группа также может содержать один или более атомов углерода, которые имеют стереохимическую конфигурацию, противоположную той, которая имеется у соответствующего углерода в рибозе. Таким образом, модифицированные нуклеотиды могут содержать сахар, такой как арабиноза.
Модифицированные нуклеотиды также могут содержать «лишенные азотистого основания» сахара, в которых отсутствует нуклеиновое основание у С-1'. Эти лишенные азотистого основания сахара могут дополнительно содержать модификации у одного или более атомов, составляющих сахар.
2'-модификации можно применять в комбинации с одной или более модификациями фосфатного линкера (например, фосфотиоат).
Фосфатные группы могут быть индивидуально замещены соединителями, не содержащими фосфор.
Примеры групп, которые могут замещать фосфатную группу, включают силоксан, карбонат, карбоксиметил, карбамат, амид, простой тиоэфир, этиленоксидный линкер, сульфонат, сульфонамид, тиоформацеталь, формацеталь, оксим, метиленимино, метиленметилимино, метиленгидразо, метилендиметилгидразо и метиленоксиметилимино. Согласно определенным вариантам реализации настоящего изобретения замещения могут включать метиленкарбониламино- и метиленметилиминогруппы.
Фосфатный линкер и рибозный сахар могут быть замещены устойчивыми к нуклеазе нуклеотидами.
Примеры включают такие суррогаты нуклеозидов как морфолино, циклобутил, пирролидин и пептидная нуклеиновая кислота (ПНК). В определенных вариантах реализации могут применяться ПНК-суррогаты.
3'- и 5'-концы олигонуклеотида могут быть модифицированы. Такие модификации могут быть на 3'-конце или 5'-конце или на обоих концах молекулы. Они могут включать модификацию или замещение всего концевого фосфата или одного или более атомов фосфатной группы. Например, 3'- и 5'-концы олигонуклеотида могут быть конъюгированы с другими функциональными молекулярными группами, такими как группы, представляющие собой метки, например, флуорофоры (например, пирен, TAMRA, флуоресцеин, красители Cy3 или Cy5), или защитные группы (например, на основе серы, кремния, бора или сложного эфира). Функциональные молекулярные группы могут быть присоединены к сахару за счет фосфатной группы и/или линкера. Концевой атом линкера может соединяться или замещать соединяющий атом фосфатной группы или C-3' или C-5' O, N, S или C группы сахара. Согласно другому варианту линкер может соединяться с концевым атомом суррогата нуклеотида (например, ПНК) или замещать его. Эти спейсеры или линкеры могут включать, например, -(CH2)n-, -(CH2)nN-, -(CH2)nO-, -(CH2)nS-, -(CH2CH2O)nCH2CH2O- (например, n=3 или 6), лишенные азотистого основания сахара, амид, карбокси, амин, оксиамин, оксиимин, тиоэфир, дисульфид, тиомочевину, сульфонамид или морфолино, или биотин и флуоресцеиновые реагенты. 3'-конец может представлять собой группу -OH.
Другие примеры концевых модификаций включают красители, интеркалирующие агенты (например, акридины), сшивающие агенты (например, псорален, митомицин C), порфирины (TPPC4, тексафирин, сапфирин), полициклические ароматические углеводороды (например, феназин, дигидрофеназин), искусственные эндонуклеазы, ЭДТА, липофильные носители (например, холестерин, желчную кислоту, адамантануксусную кислоту, 1-пиренмасляную кислоту, дигидротестостерон, 1,3-бис-O-(гексадецил)глицерин, геранилоксигексильную группу, гексадецилглицерин, борнеол, ментол, 1,3-пропандиол, гептадецильную группу, пальмитиновую кислоту, миристиновую кислоту, O3-(олеоил)литохолевую кислоту, O3-(олеоил)холеновую кислоту, диметокситритил или феноксазин) и пептидные конъюгаты (например, пептид Antennapedia, пептид Tat), алкилирующие агенты, фосфат, амино, меркапто, ПЭГ (например, ПЭГ-40К), MPEG, [MPEG]2, полиамино, алкил, замещенный алкил, радиомеченые маркеры, ферменты, гаптены (например, биотин), стимуляторы транспорта/абсорбции (например, аспирин, витамин Е, фолиевую кислоту), синтетические рибонуклеазы (например, имидазол, бисимидазол, гистамин, кластеры имидазола, конъюгаты акридин-имидазол, комплексы Eu3+ и тетраазамакроциклов).
Концевые модификации могут быть добавлены по ряду причин, включая модуляцию активности или модуляцию устойчивости к разрушению. Концевые модификации, которые можно применять для модулирования активности, включают модификацию 5'-конца фосфатом или аналогами фосфата. Нуклеиновые кислоты согласно настоящему изобретению могут быть 5'-фосфорилированы на первой или второй цепях или могут содержать фосфорильный аналог на 5'-конце. Модификации 5'-фосфатом включают модификации, которые совместимы с RISC-опосредуемым подавлением генов. Подходящие модификации включают: 5'-монофосфат ((HO)2(O)P-O-5′); 5'-дифосфат ((HO)2(O)P-O-P(HO)(O)-O-5′); 5'-трифосфат ((HO)2(O)P-O-(HO)(O)P-O-P(HO)(O)-O-5'); 5'-гуанозиновый кэп (7-метилированный или неметилированный) (7m-G-O-5'-(HO)(O)P-O-(HO)(O)P-O-P(HO)(O)-O-5'); 5'-аденозиновый кэп (Appp) и любую кэп-структуру модифицированного или немодифицированного нуклеотида (N-O-5'-(HO)(O)P-O-(HO)(O)P-O-P(HO)(O)-O-5'); 5'-монотиофосфат (фосфотиоат; (HO)2(S)P-O-5'); 5'-монодитиофосфат (фосфодитиоат; (HO)(HS)(S)P-O-5'), 5'-фосфотиолат ((HO)2(O)P-S-5'); любую дополнительную комбинацию монофосфата, дифосфата и трифосфатов, в которых кислород замещен серой (например, 5'-альфа-тиотрифосфат, 5'-гамма-тиотрифосфат и т.д.), 5'-фосфоамидаты ((HO)2(O)P-NH-5', (HO)(NH2)(O)P-O-5'), 5'-алкилфосфонаты (R = алкил = метил, этил, изопропил, пропил и т.д., например, RP(OH)(O)-O-5′-, (OH)2(O)P-5′-CH2-), 5'-винилфосфонат, 5'-алкилэфирфосфонаты (R = алкилэфир = метоксиметил (MeOCH2-), этоксиметил и т.д., например, RP(OH)(O)-O-5'-).
Нуклеиновая кислота согласно настоящему изобретению может содержать одну или более фосфотиоатных модификаций на одном или более концевых участках первой и/или второй цепей. Необязательно каждый или любой конец первой цепи может содержать один, два или три модифицированных фосфотиоатом нуклеотида. Необязательно каждый или любой конец второй цепи может содержать один, два или три модифицированных фосфотиоатом нуклеотида.
Концевые модификации также можно применять для мониторинга распределения, и в таких случаях группы, которые будут добавлены, могут включать флуорофоры, например, флуоресцеин или краситель Alexa. Концевые модификации также можно применять для усиления поглощения, подходящие для этого модификации включают холестерин. Концевые модификации также можно применять для поперечной сшивки РНК-агента с другой группой.
Аденин, гуанин, цитозин и урацил являются наиболее распространенными основаниями, обнаруженными в РНК. Эти основания могут быть модифицированы или замещены для обеспечения РНК, имеющих улучшенные свойства. Например, устойчивые к нуклеазе олигорибонуклеотиды могут быть получены с применением этих оснований или синтетических и природных нуклеиновых оснований (например, инозина, тимина, ксантина, гипоксантина, нубуларина, изогуанизина или туберцидина) и любой одной из указанных выше модификаций. Согласно другому варианту можно применять замещенные или модифицированные аналоги любого из вышеуказанных оснований и «универсальных оснований». Примеры включают 2-аминоаденин, 6-метил и другие алкильные производные аденина и гуанина, 2-пропил и другие алкильные производные аденина и гуанина, 5-галоурацил и цитозин, 5-пропинилурацил и цитозин, 6-азоурацил, цитозин и тимин, 5-урацил (псевдоурацил), 4-тиоурацил, 5-галоурацил, 5-(2-аминопропил)урацил, 5-аминоаллилурацил, 8-галоген, амино, тиол, тиоалкил, гидроксил и другие 8-замещенные аденины и гуанины, 5-трифторметил и другие 5-замещенные урацилы и цитозины, 7-метилгуанин, 5-замещенные пиримидины, 6-азапиримидины и N-2, N-6 и O-6-замещенные пурины, включая 2-аминопропиладенин, 5-пропинилурацил и 5-пропинилцитозин, дигидроурацил, 3-деаза-5-азацитозин, 2-аминопурин, 5-алкилурацил, 7-алкилгуанин, 5-алкилцитозин, 7-деазааденин, N6,N6-диметиладенин, 2,6-диаминопурин, 5-аминоаллилурацил, N3-метилурацил, замещенные 1,2,4-триазолы, 2-пиридинон, 5-нитроиндол, 3-нитропиррол, 5-метоксиурацил, урацил-5-оксиуксусную кислоту, 5-метоксикарбонилметилурацил, 5-метил-2-тиоурацил, 5-метоксикарбонилметил-2-тиоурацил, 5-метиламинометил-2-тиоурацил, 3-(3-амино-3-карбоксипропил)урацил, 3-метилцитозин, 5-метилцитозин, N<4>-ацетилцитозин, 2-тиоцитозин, N6-метиладенин, N6-изопентиладенин, 2-метилтио-N6-изопентениладенин, N-метилгуанины или O-алкилированные основания.
В настоящей заявке термин «неспаривающийся аналог нуклеотида» означает аналог нуклеотида, который содержит неспаривающуюся с основанием группу, включая, но не ограничиваясь ими: 6-дезаминоаденозин (небуларин), 4-Me-индол, 3-нитропиррол, 5-нитроиндол, Ds, Pa, N3-Me-рибо-U, N3-Me-рибо-T, N3-Me dC, N3-Me-dT, N1-Me-dG, N1-Me-dA, N3-этил-dC, N3-Me dC. Согласно определенным вариантам реализации настоящего изобретения неспаривающийся с основанием аналог нуклеотида представляет собой рибонуклеотид. Согласно другим вариантам реализации настоящего изобретения он представляет собой дезоксирибонуклеотид.
В настоящей заявке термин «концевая функциональная группа» включает, но не ограничивается ими, галогеновые, спиртовые, аминные, карбоновые, сложноэфирные, амидные, альдегидные, кетоновые, простые эфирные группы.
Определенные группы могут быть соединены с 5'-концом первой цепи или второй цепи. Они включают лишенную азотистого основания рибозную группу, лишенную азотистого основания дезоксирибозную группу, модификации лишенного азотистого основания рибозной группы и лишенной азотистого основания дезоксирибозной группы, включая 2'O-алкильные модификации; инвертированные лишенные азотистого основания рибозные группы и лишенные азотистого основания дезоксирибозные группы и их модификации, C6-имино-Pi; зеркальный нуклеотид, включая L-ДНК и L-РНК; 5'-OMe-нуклеотид; и аналоги нуклеотидов, включая 4',5'-метиленнуклеотид; 1-β-D-эритрофуранозилнуклеотид; 4'-тионуклеотид, карбоциклический нуклеотид; 5'-аминоалкилфосфат; 1,3-диамино-2-пропилфосфат, 3-аминопропилфосфат; 6-аминогексилфосфат; 12-аминододецилфосфат; гидроксипропилфосфат; 1,5-ангидрогекситолнуклеотид; альфа-нуклеотид; треопентофуранозилнуклеотид; ациклический 3',4'-секонуклеотид; 3,4-дигидроксибутилнуклеотид; 3,5-дигидроксипентилнуклеотид, 5'-5'-инвертированная лишенная азотистого основания группа; 1,4-бутандиолфосфат; 5'-амино; и мостиковые или немостиковые метилфосфонатные и 5'-меркапто группы.
Нуклеиновые кислоты согласно настоящему изобретению могут содержать один или более инвертированных нуклеотидов, например, инвертированный тимидин или инвертированный аденин (например, см. Takei, et al., 2002. JBC 277 (26):23800-06).
В настоящей заявке термин «ингибировать», «подавлять» или «уменьшать» в отношении экспрессии гена означает, что экспрессия гена или уровень молекул РНК или эквивалентных молекул РНК, кодирующих один или более белков или субъединиц белков (например, мРНК), или активность одного или более белков или субъединиц белков уменьшается ниже уровня, наблюдаемого в отсутствие нуклеиновой кислоты или конъюгированной нуклеиновой кислоты согласно настоящему изобретению, или по отношению к молекуле миРНК без известной гомологии с транскриптами человека (в настоящей заявке называется неподавляющий контроль). Такой контроль может быть конъюгирован и модифицирован аналогичным образом, как и молекула согласно настоящему изобретению, и доставлен в клетку-мишень аналогичным путем; например, экспрессия может уменьшиться до 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 15% или до промежуточных значений, или до меньшего значения, чем то, которое наблюдается в отсутствие нуклеиновой кислоты или конъюгированной нуклеиновой кислоты или в присутствии неподавляющего контроля.
Нуклеиновая кислота согласно настоящему изобретению может содержать лишенный азотистого основания нуклеотид. В настоящей заявке термин «лишенный азотистого основания» относится к группам, не содержащим основание или имеющим другие химические группы вместо основания в положении 1', например, 3',3'-соединенное или 5',5'-соединенное производное лишенной азотистого основания дезоксирибозы.
Нуклеиновая кислота может содержать один или более нуклеотидов на второй и/или первой цепях, которые модифицированы. Чередующиеся нуклеотиды могут быть модифицированы с образованием модифицированных нуклеотидов.
Чередование, описанное в настоящей заявке, означает, что оно происходит один за другим на регулярной основе. Другими словами, чередование означает повторение по очереди. Например, если один нуклеотид модифицирован, следующий смежный нуклеотид не модифицирован, а следующий смежный нуклеотид модифицирован и так далее. Один нуклеотид может быть модифицирован с применением первой модификации, следующий смежный нуклеотид может быть модифицирован с применением второй модификации, а следующий смежный нуклеотид модифицирован с применением первой модификации и так далее, при этом первая и вторая модификации отличаются.
Один или более из нуклеотидов в нечетных положениях первой цепи нуклеиновой кислоты согласно настоящему изобретению могут быть модифицированы, причем первая цепь пронумерована от 5' к 3', самый 5'-крайний нуклеотид представляет собой нуклеотид номер 1 первой цепи. Термин «нечетный», описанный в настоящей заявке, означает число, неделимое на два. Примерами нечетных чисел являются 1, 3, 5, 7, 9, 11 и так далее. Один или более из нуклеотидов в четных положениях первой цепи нуклеиновой кислоты согласно настоящему изобретению могут быть модифицированы, причем первая цепь пронумерована от 5' к 3'. Термин «четный», описанный в настоящей заявке, означает число, которое поровну делится на два. Примерами четных чисел являются 2, 4, 6, 8, 10, 12, 14 и так далее. Один или более из нуклеотидов в нечетных положениях второй цепи нуклеиновой кислоты согласно настоящему изобретению могут быть модифицированы, при этом вторая цепь пронумерована от 3' к 5', причем наиболее 3'-концевой нуклеотид представляет собой нуклеотид номер 1 второй цепи. Один или более из нуклеотидов в четных положениях второй цепи нуклеиновой кислоты согласно настоящему изобретению могут быть модифицированы, причем вторая цепь пронумерована от 3' к 5'.
Один или более нуклеотидов на первой и/или второй цепях могут быть модифицированы с образованием модифицированных нуклеотидов. Один или более из нуклеотидов в нечетных положениях первой цепи могут быть модифицированы. Один или более из нуклеотидов в четных положениях первой цепи могут быть модифицированы с применением по меньшей мере второй модификации, причем по меньшей мере вторая модификация отличается от модификации одного или более нуклеотидов в нечетных положениях. По меньшей мере один из одного или более модифицированных нуклеотидов в четных положениях может быть смежным с по меньшей мере одним или более модифицированными нуклеотидами в нечетных положениях.
Множество нуклеотидов в нечетных положениях первой цепи могут быть модифицированы в нуклеиновой кислоте согласно настоящему изобретению. Множество нуклеотидов в четных положениях первой цепи могут быть модифицированы с применением второй модификации. Первая цепь может содержать смежные нуклеотиды, которые модифицированы с применением общей модификации. Первая цепь также может содержать смежные нуклеотиды, которые модифицированы с применением второй отличающейся модификации.
Один или более из нуклеотидов в нечетных положениях второй цепи могут быть модифицированы с применением модификации, которая отличается от модификации нуклеотидов в нечетных положенияхн первой цепи, и/или один или более из нуклеотидов в четных положениях второй цепи могут быть модифицированы с применением модификации, аналогичной таковой для нуклеотидов в нечетных положениях первой цепи. По меньшей мере один из одного или более модифицированных нуклеотидов в четных положениях второй цепи может быть смежным с одним или более модифицированными нечетными нуклеотидами. Множество нуклеотидов в нечетных положениях второй цепи могут быть модифицированы с применением общей модификации, и/или множество нуклеотидов в четных положениях могут быть модифицированы с применением модификации, которая аналогична присутствующей на нечетных нуклеотидах первой цепи. Множество нуклеотидов в нечетных положениях второй цепи могут быть модифицированы с применением второй модификации, причем вторая модификация отличается от модификации нуклеотидов в нечетных положениях первой цепи.
Вторая цепь может содержать смежные нуклеотиды, которые модифицированы с применением общей модификации, которая может представлять собой вторую модификацию, которая отличается от модификации нуклеотидов в нечетных положениях первой цепи.
В нуклеиновой кислоте согласно настоящему изобретению каждый из нуклеотидов в нечетных положениях в первой цепи и каждый из нуклеотидов в четных положениях второй цепи может быть модифицирован с применением общей модификации, и каждый из нуклеотидов в четных положениях первой цепи может быть модифицирован с применением второй модификации, и каждый из нуклеотидов в нечетных положениях во второй цепи может быть модифицирован с применением второй модификации.
Модифицированные нуклеотиды первой цепи нуклеиновой кислоты согласно настоящему изобретению могут быть сдвинуты по меньшей мере на один нуклеотид относительно немодифицированных или по-другому модифицированных нуклеотидов второй цепи.
Один или более или каждый из нуклеотидов в нечетных положениях могут быть модифицированы в первой цепи, и один или более или каждый из нуклеотидов в четных положениях в четных положениях могут быть модифицированы во второй цепи. Один или более или каждый из чередующихся нуклеотидов на любой или обеих цепях могут быть модифицированы с применением второй модификации. Один или более или каждый из нуклеотидов в четных положениях могут быть модифицированы в первой цепи, и один или более или каждый из нуклеотидов в четных положениях могут быть модифицированы во второй цепи. Один или более или каждый из чередующихся нуклеотидов на любой или обеих цепях могут быть модифицированы с применением второй модификации. Один или более или каждый из нуклеотидов в нечетных положениях в первой цепи могут быть модифицированы, и один или более из нуклеотидов в нечетных положениях во второй цепи могут быть модифицированы с применением общей модификации. Один или более или каждый из чередующихся нуклеотидов на любой или обеих цепях могут быть модифицированы с применением второй модификации. Один или более или каждый из нуклеотидов в четных положениях в первой цепи могут быть модифицированы, и один или более или каждый из нуклеотидов в нечетных положениях во второй цепи могут быть модифицированы с применением общей модификации. Один или более или каждый из чередующихся нуклеотидов на любой или обеих цепях могут быть модифицированы с применением второй модификации.
Нуклеиновая кислота согласно настоящему изобретению может содержать одноцепочечные или двухцепочечные конструкции, которые содержат по меньшей мере две области чередующихся модификаций в одной или обеих цепях. Эти чередующиеся области могут содержать до приблизительно 12 нуклеотидов, но предпочтительно содержат от приблизительно 3 до приблизительно 10 нуклеотидов. Области чередующихся нуклеотидов могут быть расположены на концах одной или обеих цепей нуклеиновой кислоты согласно настоящему изобретению. Нуклеиновая кислота может содержать от 4 до приблизительно 10 чередующихся нуклеотидов на каждом конце (3' и 5'), и эти области могут быть отделены с помощью от приблизительно 5 до приблизительно 12 смежных немодифицированных или по-разному модифицированных или модифицированных с применением общей модификации нуклеотидов.
Нечетные нуклеотиды первой цепи могут быть модифицированы и четные нуклеотиды могут быть модифицированы с применением второй модификации. Вторая цепь может содержать смежные нуклеотиды, которые модифицированы с применением общей модификации, которая может быть аналогична модификации нуклеотидов в нечетных положениях первой цепи. Один или более нуклеотидов второй цепи также могут быть модифицированы с применением второй модификации. Один или более нуклеотидов со второй модификацией могут быть смежными друг с другом и с нуклеотидами, имеющими модификацию, которая аналогична модификации нуклеотидов в нечетных положениях первой цепи. Первая цепь также может содержать фосфотиоатные связи между двумя нуклеотидами на 3'-конце и на 5'-конце. Вторая цепь может содержать фосфотиоатную связь между двумя нуклеотидами на 5'-конце. Вторая цепь также может быть конъюгирована с лигандом на 5'-конце.
Нуклеиновая кислота согласно настоящему изобретению может содержать первую цепь, содержащую смежные нуклеотиды, которые модифицированы с применением общей модификации. Один или более таких нуклеотидов могут быть смежными с одним или более нуклеотидами, которые могут быть модифицированы с применением второй модификации. Один или более нуклеотидов со второй модификацией могут быть смежными. Вторая цепь может содержать смежные нуклеотиды, которые модифицированы с применением общей модификации, которая может быть аналогична одной из модификаций одного или более нуклеотидов первой цепи. Один или более нуклеотидов второй цепи также могут быть модифицированы с применением второй модификации. Один или более нуклеотидов со второй модификацией могут быть смежными. Первая цепь также может содержать фосфотиоатные связи между двумя нуклеотидами на 5'-конце и на 3'-конце. Вторая цепь может содержать фосфотиоатную связь между двумя нуклеотидами на 3'-конце. Вторая цепь также может быть конъюгирована с лигандом на 5'-конце.
Нуклеотиды, пронумерованные от 5' к 3' на первой цепи и от 3' к 5' на второй цепи 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23 и 25, могут быть модифицированы с применением модификации на первой цепи. Нуклеотиды, пронумерованные 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 и 24, могут быть модифицированы с применением второй модификации на первой цепи. Нуклеотиды, пронумерованные 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, могут быть модифицированы с применением модификации на второй цепи. Нуклеотиды, пронумерованные 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 и 24, могут быть модифицированы с применением второй модификации на второй цепи. В отношении нуклеиновой кислоты согласно настоящему изобретению нуклеотиды пронумерованы от 5' к 3' на первой цепи и от 3' к 5' на второй цепи.
Нуклеотиды, пронумерованные 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 и 24, могут быть модифицированы с применением модификации на первой цепи. Нуклеотиды, пронумерованные 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, могут быть модифицированы с применением второй модификации на первой цепи. Нуклеотиды, пронумерованные 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, могут быть модифицированы с применением модификации на второй цепи. Нуклеотиды, пронумерованные 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22 и 24, могут быть модифицированы с применением второй модификации на второй цепи.
Понятно, что если первая и/или вторая цепи короче чем 25 нуклеотидов в длину, например, 19 нуклеотидов в длину, то нуклеотиды с номерами 20, 21, 22, 23, 24 и 25, подлежащие модификации, отсутствуют. Специалист поймет приведенное выше описание для применения к более коротким цепям, соответственно.
Один или более модифицированных нуклеотидов на первой цепи могут быть спарены с модифицированными нуклеотидами на второй цепи, имеющими общую модификацию. Один или более модифицированных нуклеотидов на первой цепи могут быть спарены с модифицированными нуклеотидами на второй цепи, имеющими другую модификацию. Один или более модифицированных нуклеотидов на первой цепи могут быть спарены с немодифицированными нуклеотидами на второй цепи. Один или более модифицированных нуклеотидов на второй цепи могут быть спарены с немодифицированными нуклеотидами на первой цепи. Другими словами, чередующиеся нуклеотиды могут быть выравнены на двух цепях таким образом, что, например, все модификации в чередующихся областях второй цепи спарены с идентичными модификациями в первой цепи или, согласно другому варианту, модификации могут быть компенсированы одним нуклеотидом с общими модификациями в чередующихся областях одной цепи, спаренным с отличающимися модификациями (т.е. второй или дополнительной модификацией) в другой цепи. Другой вариант заключается в наличии разных модификаций в каждой из цепей.
Модификации на первой цепи могут быть сдвинуты на один нуклеотид относительно модифицированных нуклеотидов на второй цепи так, что нуклеотиды с общей модификацией не спариваются друг с другом.
Модификация и/или модификации могут быть по отдельности и независимо выбраны из группы, состоящей из 3'-концевого дезокситимина, 2'-O-метила, 2'-дезокси-модификации, 2'-амино-модификации, 2'-алкил-модификации, морфолино-модификации, модификации фосфоамидатом, модификации 5'-фосфотиоатной группой, модификации 5'-фосфатом или миметиком 5'-фосфата, а также модификации производным холестерина или группой бисдециламида додекановой кислоты, и/или модифицированный нуклеотид может представлять собой любой из закрытого нуклеотида, лишенного азотистого основания нуклеотида или содержащего неприродное основание нуклеотида.
По меньшей мере одна модификация может представлять собой 2'-O-метил и/или по меньшей мере одна модификация может представлять собой 2'-F. Дополнительные модификации, описанные в настоящей заявке, могут присутствовать на первой и/или второй цепях.
Нуклеиновая кислота согласно настоящему изобретению может содержать инвертированный РНК-нуклеотид на одном или нескольких концах цепи. Такие инвертированные нуклеотиды обеспечивают стабильность нуклеиновой кислоты. Предпочтительно нуклеиновая кислота содержит по меньшей мере инвертированный нуклеотид на одном или нескольких из 3'-конца по меньшей мере одной из цепей и/или на 5'-конце второй цепи. Более предпочтительно нуклеиновая кислота содержит инвертированный нуклеотид на 3'-конце второй цепи. Наиболее предпочтительно нуклеиновая кислота содержит инвертированный РНК-нуклеотид на 3'-конце второй цепи, и этот нуклеотид предпочтительно представляет собой инвертированный А. Инвертированный нуклеотид предпочтительно присутствует на конце цепи не в виде липкого конца, а напротив соответствующего нуклеотида в другой цепи. Нуклеиновая кислота с такой модификацией является стабильной и легко синтезируется.
В описании настоящего изобретения «аналогичная или общая модификация» означает одну и ту же модификацию для любого нуклеотида, будь то A, G, С или U, модифицированного с применением группы, такой как метильная группа или фторгруппа. Это не подразумевает аналогичное добавление на одном и том же нуклеотиде. Например, 2'-F-dU, 2'-F-dA, 2'-F-dC, 2'-F-dG все считаются аналогичными или общими модификациями, так же как и 2'-OMe-rU, 2'-OMe-rA; 2'-OMe-rC; 2'-OMe-rG. Модификация 2'-F представляет собой модификацию, отличную от модификации 2'-OMe.
Некоторые типичные модифицированные последовательности нуклеиновых кислот согласно настоящему изобретению показаны в примерах. Подразумевается, что эти примеры являются типичными, а не ограничивающими.
Предпочтительно нуклеиновая кислота может содержать модификацию и вторую или дополнительную модификацию, которые по отдельности и независимо выбраны из группы, включающей 2'-O-метил-модификацию и 2'-F-модификацию. Нуклеиновая кислота может содержать модификацию, которая представляет собой 2'-O-метил (2'-OMe), который может представлять собой первую модификацию, и вторую модификацию, которая представляет собой 2'-F. Нуклеиновая кислота согласно настоящему изобретению также может содержать фосфотиоатную модификацию и/или дезокси-модификацию, которая может присутствовать на 1, 2 или 3 концевых нуклеотидах или между ними каждого или любого конца каждой или обеих цепей.
Согласно настоящему изобретению предложена, в качестве дополнительного аспекта, нуклеиновая кислота для ингибирования экспрессии LPA в клетке, содержащая нуклеотидную последовательность SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73, в которой нуклеотиды первой цепи модифицированы с применением первой модификации на нечетных нуклеотидах и модифицированы с применением второй модификации на четных нуклеотидах, и нуклеотиды второй цепи модифицированы с применением третьей модификации на четных нуклеотидах и модифицированы с применением четвертой модификации на нечетных нуклеотидах, причем по меньшей мере первая модификация отличается от второй модификации, а третья модификация отличается от четвертой модификации. Третья и первая модификации могут быть одинаковыми или различными, вторая и четвертая модификации могут быть одинаковыми или различными. Первая и вторая модификации могут отличаться друг от друга, и третья и четвертая модификации могут отличаться друг от друга.
Вторая цепь может содержать нуклеотидную последовательность SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 64, 66, 68, 70, 72 или 74. Нуклеотиды первой цепи могут быть модифицированы с применением первой модификации на нечетных нуклеотидах и модифицированы с применением второй модификации на четных нуклеотидах, и вторая цепь может быть модифицирована на нечетных нуклеотидах с применением второй модификации и модифицирована с применением первой модификации на четных нуклеотидах. Первая модификация может представлять собой 2'-OMe, а вторая модификация может представлять собой 2'-F. Первая цепь может содержать нуклеотидную последовательность SEQ ID NO: 5 или SEQ ID NO: 9, и/или вторая цепь может содержать нуклеотидную последовательность SEQ ID NO: 6 или SEQ ID NO: 10. Модификации могут представлять собой те, которые представлены в таблице 1.
Нуклеиновая кислота согласно настоящему изобретению может быть конъюгирована с лигандом. Эффективная доставка олигонуклеотидов, в частности, двухцепочечных нуклеиновых кислот согласно настоящему изобретению, в клетки в условиях in vivo является важной и требует специфичного нацеливания и существенной защиты от внеклеточной среды, в частности, от сывороточных белков. Одним из способов достижения специфичного нацеливания является конъюгирование лиганда с нуклеиновой кислотой. Лиганд помогает нацеливать нуклеиновую кислоту на требуемый сайт-мишень. Существует потребность в конъюгации соответствующих лигандов для целевых рецепторных молекул для того чтобы клетки-мишени поглощали конъюгированные молекулы с помощью таких механизмов как различные опосредуемые рецепторами пути эндоцитоза или функционально аналогичные процессы.
Одним из примеров является комплекс рецептора асиалогликопротеина (ASGP-R), состоящий из варьирующихся соотношений мультимеров мембранных рецепторов ASGR1 и ASGR2, который очень распространен на гепатоцитах и обладает высокой аффинностью в отношении группы GalNAc, описанной в настоящей заявке. В патенте США №5885968 представлено одно из первых раскрытий применения трехантенных кластерных гликозидов в качестве конъюгированных лигандов. Конъюгаты, имеющие три лиганда GalNAc и содержащие фосфатные группы, известны и описаны в Dubber et al. (Bioconjug. Chem. 2003 Jan-Feb; 14(1):239-46.). Комплекс ASGP-R проявляет в 50 раз более высокую аффинность в отношении N-ацетил-D-галактозиламина (GalNAc) по сравнению с D-Gal.
Комплекс рецептора асиалогликопротеина (ASGP-R), который специфично распознает концевые β-галактозильные субъединицы гликозилированных белков или других олигосахаридов (Weigel, P.H. et. al., Biochim. Biophys. Acta. 2002 Sep 19; 1572(2-3):341-63), можно применять для доставки лекарственного средства в гепатоциты печени, экспрессирующие рецепторный комплекс, путем ковалентного связывания галактозы или галактозамина с лекарственным веществом (Ishibashi,S.; et. al., J Biol. Chem. 1994 Nov 11;269(45):27803-6). Кроме того, аффинность связывания может быть значительно повышена с помощью поливалентного эффекта, который может быть достигнут путем повторения нацеливающей группы (Biessen EA, et al., J Med Chem. 1995 Apr 28; 38(9):1538-46.).
Комплекс ASGP-R выступает в качестве посредника для активного поглощения гликопротеинов, содержащих концевой β-галактозил, в эндосомы клетки. Таким образом, ASGPR очень подходит для нацеленной доставки лекарственных средств-кандидатов, конъюгированных с такими лигандами, например, нуклеиновых кислот, в экспрессирующие рецептор клетки (Akinc et al., Mol Ther. 2010 Jul; 18(7):1357-64).
В более общем случае лиганд может содержать сахарид, который выбран так, чтобы обладать аффинностью в отношении по меньшей мере одного типа рецептора на клетке-мишени. В частности, рецептор находится на поверхности клетки печени млекопитающего, например, печеночный комплекс рецептора асиалогликопротеина, описанный ранее (ASGP-R).
Сахарид может быть выбран из N-ацетилгалактозамина, маннозы, галактозы, глюкозы, глюкозамина и фукозы. Сахарид может представлять собой N-ацетилгалактозамин (GalNAc).
Таким образом, лиганд для применения в настоящем изобретении может содержать (i) одну или более групп N-ацетилгалактозамина (GalNAc) и его производных, и (ii) линкер, причем указанный линкер конъюгирует группы GalNAc с последовательностью, определенной в любых предыдущих аспектах. Линкер может иметь двухвалентную, трехвалентную или четырехвалентную разветвленную структуру. Нуклеотиды могут быть модифицированы, как определено в настоящей заявке.
«GalNAc» относится к 2-ацетиламино-2-дезокси-D-галактопиранозе, обычно называемой в литературе N-ацетилгалактозамином. Ссылка на «GalNAc» или «N-ацетилгалактозамин» включает обе β-форму, 2-ацетиламино-2-дезокси-β-D-галактопиранозу, и α-форму, 2-ацетиламино-2-дезокси-α-D-галактопиранозу. Обе β-форма, 2-ацетиламино-2-дезокси-β-D-галактопираноза, и α-форма, 2-ацетиламино-2-дезокси-α-D-галактопираноза, могут использоваться взаимозаменяемо. Предпочтительно соединения согласно настоящему изобретению содержат β-форму, 2-ацетиламино-2-дезокси-β-D-галактопиранозу.
Таким образом, лиганд может содержать GalNAc.
Лиганд может содержать соединение формулы I:
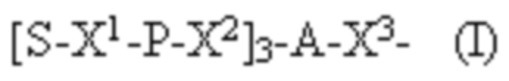
в которой:
S представляет собой сахарид, причем указанный сахарид представляет собой N-ацетилгалактозамин;
X1 представляет собой C3-C6 алкилен или (-CH2-CH2-O)m(-CH2)2-, где m равно 1, 2 или 3;
P представляет собой фосфат или модифицированный фосфат (предпочтительно тиофосфат);
X2 представляет собой алкилен или простой алкиленовый эфир формулы (-CH2)n-O-CH2-, где n=1-6;
А представляет собой блок разветвления;
X3 представляет собой мостиковый блок;
при этом нуклеиновая кислота в соответствии с настоящим изобретением конъюгирована с X3 за счет фосфата или модифицированного фосфата (предпочтительно тиофосфата).
В формуле I блок разветвления «А» разветвляется на три для размещения трех сахаридных лигандов. Блок разветвления ковалентно присоединен к оставшимся закрепленным частям лиганда и нуклеиновой кислоты. Блок разветвления может содержать разветвленную алифатическую группу, содержащую группы, выбранные из алкильной, амидной, дисульфидной, полиэтиленгликолевой, простой эфирной, простой тиоэфирной и гидроксиаминной групп. Блок разветвления может содержать группы, выбранные из алкильной и простой эфирной групп.
Блок разветвления A может иметь структуру, выбранную из:
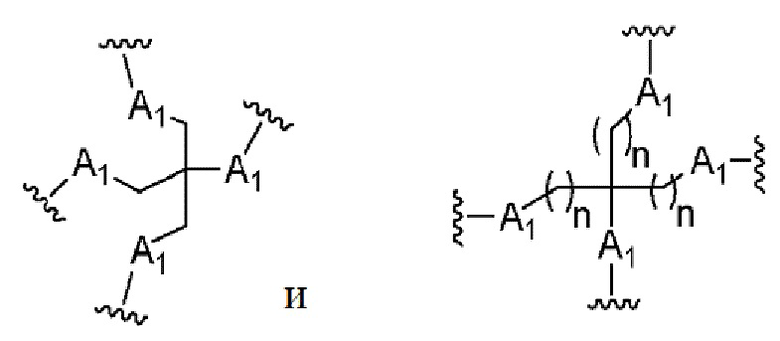
в которой каждый A1 независимо представляет собой O, S, C=O или NH; и
каждый n независимо представляет собой целое число от 1 до 20.
Блок разветвления может иметь структуру, выбранную из:

в которой каждый A1 независимо представляет собой O, S, C=O или NH; и
каждый n независимо представляет собой целое число от 1 до 20.
Блок разветвления может иметь структуру, выбранную из:
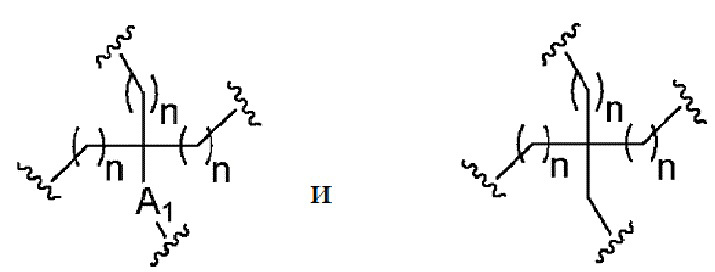
в которой A1 представляет собой O, S, C=O или NH; и
каждый n независимо представляет собой целое число от 1 до 20.
Блок разветвления может иметь структуру:
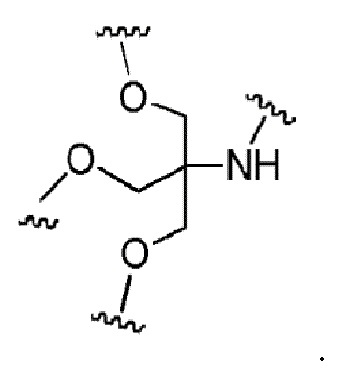
Блок разветвления может иметь структуру:
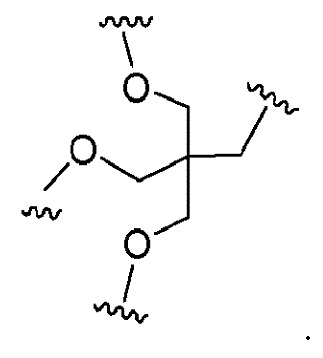
Блок разветвления может иметь структуру:
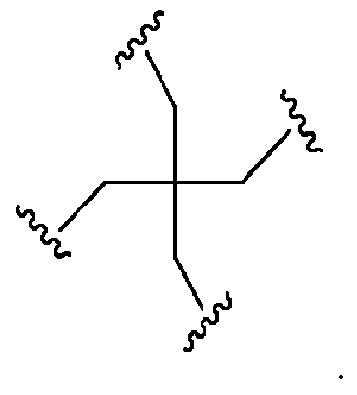
Необязательно блок разветвления состоит только из атома углерода.
Часть «X3» представляет собой мостиковый блок. Мостиковый блок является линейным и ковалентно связан с блоком разветвления и нуклеиновой кислотой.
X3 может быть выбран из -C1-C20 алкилен-, -C2-C20 алкенилен-, простого алкиленового эфира формулы -(C1-C20 алкилен)-O-(C1-C20 алкилен)-, -C(O)-C1-C20 алкилен-, -C0-C4 алкилен(Cy)C0-C4 алкилен-, где Cy представляет собой замещенное или незамещенное 5 или 6-членное циклоалкиленовое, ариленовое, гетероциклиленовое или гетероариленовое кольцо, -C1-C4 алкилен-NHC(O)-C1-C4-алкилен-, -C1-C4 алкилен-C(O)NH-C1-C4-алкилен-, -C1-C4 алкилен-SC(O)-C1-C4-алкилен-, -C1-C4 алкилен-C(O)S-C1-C4 алкилен-, -C1-C4 алкилен-OC(O)-C1-C4 алкилен-, -C1-C4 алкилен-C(O)O-C1-C4 алкилен- и -C1-C6 алкилен-S-S-C1-C6 алкилен-.
X3 может представлять собой простой алкиленовый эфир формулы -(C1-C20 алкилен)-O-(C1-C20 алкилен)-. X3 может представлять собой простой алкиленовый эфир формулы -(C1-C20 алкилен)-O-(C4-C20 алкилен)-, причем указанный (C4-C20 алкилен) связан с Z. X3 может быть выбран из группы, состоящей из -CH2-O-C3H6-, -CH2-O-C4H8-, -CH2-O-C6H12- и -CH2-O-C8H16-, в частности, -CH2-O-C4H8-, -CH2-O-C6H12- и -CH2-O-C8H16-, где в каждом случае группа -CH2- соединена с A.
Лиганд может содержать соединение формулы (II):
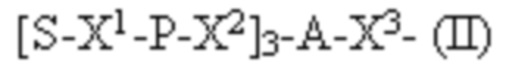
в которой:
S представляет собой сахарид;
X1 представляет собой C3-C6 алкилен или (-CH2-CH2-O)m(-CH2)2-, где m равно 1, 2 или 3;
P представляет собой фосфат или модифицированный фосфат (предпочтительно тиофосфат);
X2 представляет собой C1-C8 алкилен;
A представляет собой блок разветвления, выбранный из:

X3 представляет собой мостиковый блок;
при этом нуклеиновая кислота в соответствии с настоящим изобретением конъюгирована с X3 за счет фосфата или модифицированного фосфата (предпочтительно тиофосфата)
Блок разветвления A может иметь структуру:
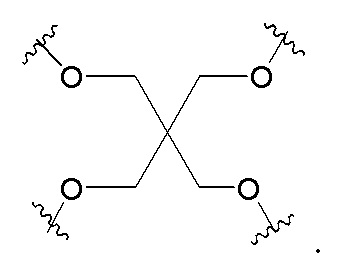
Блок разветвления A может иметь структуру:
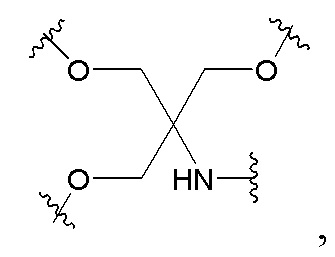
где X3 присоединен к атому азота.
X3 может представлять собой C1-C20 алкилен. Предпочтительно X3 выбран из группы, состоящей из -C3H6-, -C4H8-, -C6H12- и -C8H16-, в частности, -C4H8-, -C6H12- и -C8H16-.
Лиганд может содержать соединение формулы (III):

в которой:
S представляет собой сахарид;
X1 представляет собой C3-C6 алкилен или (-CH2-CH2-O)m(-CH2)2-, где m равно 1, 2 или 3;
P представляет собой фосфат или модифицированный фосфат (предпочтительно тиофосфат);
X2 представляет собой простой алкиленовый эфир формулы -C3H6-O-CH2-;
A представляет собой блок разветвления;
X3 представляет собой простой алкиленовый эфир формулы, выбранной из группы, состоящей из -CH2-O-CH2-, -CH2-O-C2H4-, -CH2-O-C3H6-, -CH2-O-C4H8-, -CH2-O-C5H10-, -CH2-O-C6H12-, -CH2-O-C7H14- и -CH2-O-C8H16-, при этом в каждом случае группа -CH2- соединена с A,
и при этом X3 конъюгирован с нуклеиновой кислотой в соответствии с настоящим изобретением с помощью фосфата или модифицированного фосфата (предпочтительно тиофосфата).
Блок разветвления может содержать углерод. Предпочтительно блок разветвления представляет собой углерод.
X3 может быть выбран из группы, состоящей из -CH2-O-C4H8-, -CH2-O-C5H10-, -CH2-O-C6H12-, -CH2-O-C7H14- и -CH2-O-C8H16-. Предпочтительно X3 выбран из группы, состоящей из -CH2-O-C4H8-, -CH2-O-C6H12- и -CH2-O-C8H16.
Для любого из вышеуказанных аспектов, если P представляет собой модифицированную фосфатную группу, P может быть представлен как:
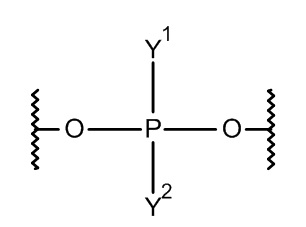
где Y1 и Y2 каждый независимо представляют собой =O, =S, -O-, -OH, -SH, -BH3, -OCH2CO2, -OCH2CO2Rx, -OCH2C(S)ORx и –ORx, где Rx представляет собой C1-C6 алкил и где  указывает на присоединение к остальной части соединения.
указывает на присоединение к остальной части соединения.
Под модифицированным фосфатом подразумевается фосфатная группа, в которой один или более из несоединяющих атомов кислорода замещены. Примеры модифицированных фосфатных групп включают фосфотиоат, фосфоселенаты, боранофосфаты, сложные эфиры боранофосфата, фосфонаты водорода, фосфоамидаты, алкил или арилфосфонаты и сложные фосфотриэфиры. В фосфодитиоатах оба несоединяющих атома кислорода замещены серой. Один, каждый или оба несоединяющих атома кислорода в фосфатной группе могут независимо представлять собой любой из S, Se, B, C, H, N или OR (R представляет собой алкил или арил).
Фосфат также может быть модифицирован путем замещения соединяющего атома кислорода азотом (мостиковые фосфоамидаты), серой (мостиковые фосфотиоаты) и углеродом (мостиковые метиленфосфонаты). Замещение может происходить на концевом атоме кислорода. Возможно замещение несоединяющих атомов кислорода азотом.
Например, Y1 может представлять -OH, а Y2 может представлять =O или =S; или
Y1 может представлять -O-, а Y2 может представлять =O или =S;
Y1 может представлять =O, а Y2 может представлять -CH3, -SH, -ORx или -BH3;
Y1 может представлять =S, а Y2 может представлять -CH3, ORx или -SH.
Специалист в данной области техники поймет, что в определенных случаях будет делокализация между Y1 и Y2.
Предпочтительно модифицированная фосфатная группа представляет собой тиофосфатную группу. Тиофосфатные группы включают битиофосфат (т.е. где Y1 представляет =S и Y2 представляет -S-) и монотиофосфат (т.е. где Y1 представляет -O- и Y2 представляет =S, или где Y1 представляет =O и Y2 представляет -S-). Предпочтительно P представляет собой монотиофосфат. Авторы настоящего изобретения обнаружили, что конъюгаты, имеющие тиофосфатные группы вместо фосфатных групп, обладают улучшенной активностью и продолжительностью действия в условиях in vivo.
P также может представлять собой этилфосфат (т.е. где Y1 представляет собой =O и Y2 представляет собой OCH2CH3).
Сахарид может быть выбран так, чтобы обладать аффинностью в отношении по меньшей мере одного типа рецептора на клетке-мишени. В частности, рецептор находится на поверхности клетки печени млекопитающего, например, печеночный комплекс рецептора асиалогликопротеина (ASGP-R).
Для любого из вышеуказанных аспектов сахарид может быть выбран из N-ацетила с одним или более из галактозамина, маннозы, галактозы, глюкозы, глюкозамина и фруктозы. Как правило, лиганд для применения в настоящем изобретении может содержать N-ацетилгалактозамин (GalNAc). Предпочтительно соединения согласно настоящему изобретению могут содержать 3 лиганда, каждый из которых будет предпочтительно содержать N-ацетилгалактозамин.
«GalNAc» относится к 2-ацетиламино-2-дезокси-D-галактопиранозе, обычно называемой в литературе N-ацетилгалактозамином. Ссылка на «GalNAc» или «N-ацетилгалактозамин» включает обе β-форму, 2-ацетиламино-2-дезокси-β-D-галактопиранозу, и α-форму, 2-ацетиламино-2-дезокси-α-D-галактопиранозу. Согласно определенным вариантам реализации настоящего изобретения β-форма, 2-ацетиламино-2-дезокси-β-D-галактопираноза, и α-форма, 2-ацетиламино-2-дезокси-α-D-галактопираноза, могут использоваться взаимозаменяемо. Предпочтительно соединения согласно настоящему изобретению содержат β-форму, 2-ацетиламино-2-дезокси-β-D-галактопиранозу.
2-ацетиламино-2-дезокси-D-галактопираноза
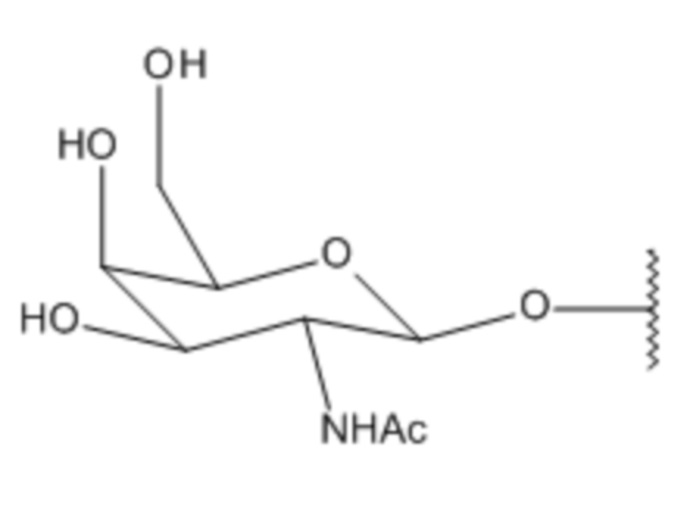
2-ацетиламино-2-дезокси-β-D-галактопираноза
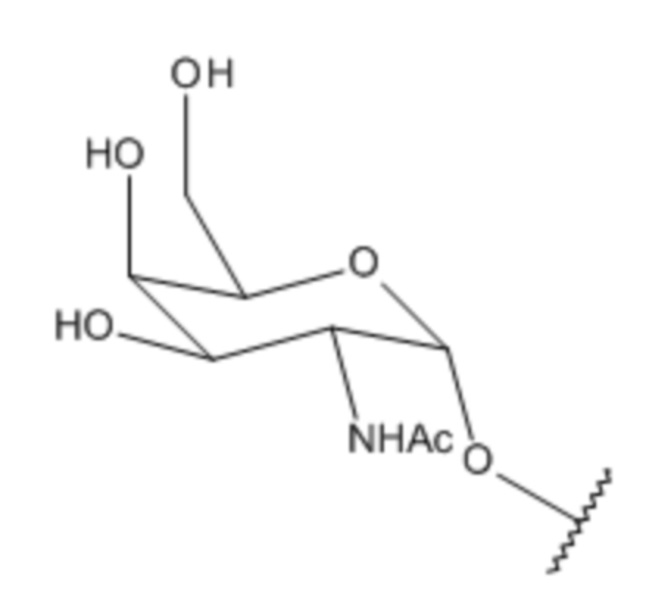
2-ацетиламино-2-дезокси-α-D-галактопираноза.
Для любого из вышеуказанных соединений формулы (III) X1 может представлять собой (-CH2-CH2-O)(-CH2)2-. X1 может представлять собой (-CH2-CH2-O)2(-CH2)2-. X1 может представлять собой (-CH2-CH2-O)3(-CH2)2-. Предпочтительно X1 представляет собой (-CH2-CH2-O)2(-CH2)2-. Согласно другому варианту X1 представляет собой C3-C6 алкилен. X1 может представлять собой пропилен. X1 может представлять собой бутилен. X1 может представлять собой пентилен. X1 может представлять собой гексилен. Предпочтительно алкил представляет собой линейный алкилен. В частности, X1 может представлять собой бутилен.
Для соединений формулы (III) X2 представляет собой простой алкиленовый эфир формулы -C3H6-O-CH2-, т.е. C3 алкоксиметилен, или –CH2CH2CH2OCH2-.
Согласно настоящему изобретению предложена конъюгированная нуклеиновая кислота, имеющая одну из следующих структур:
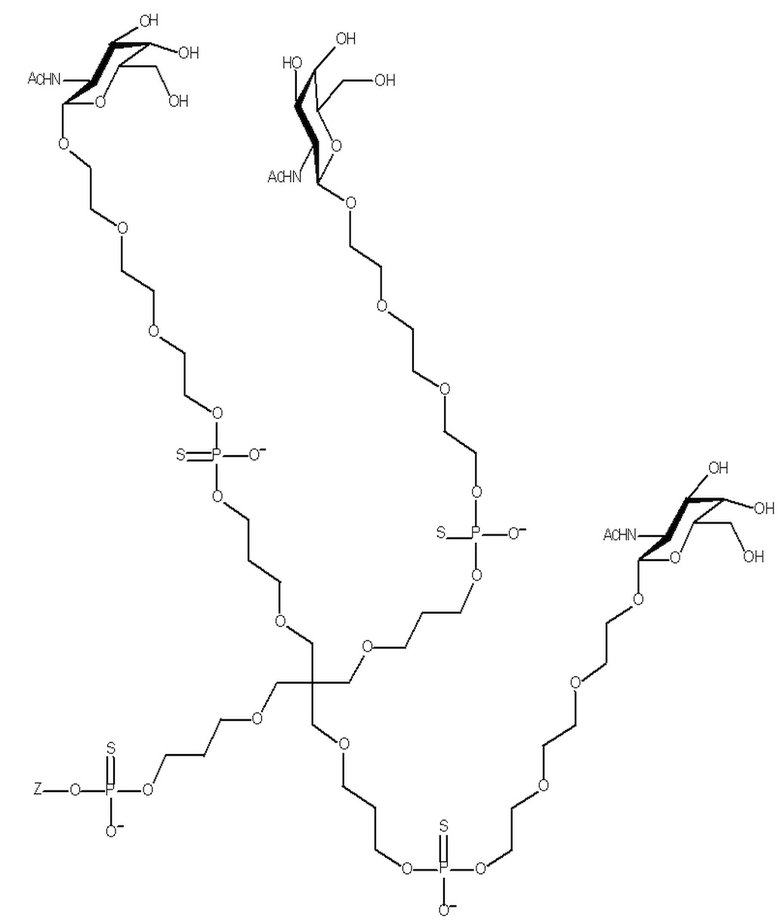

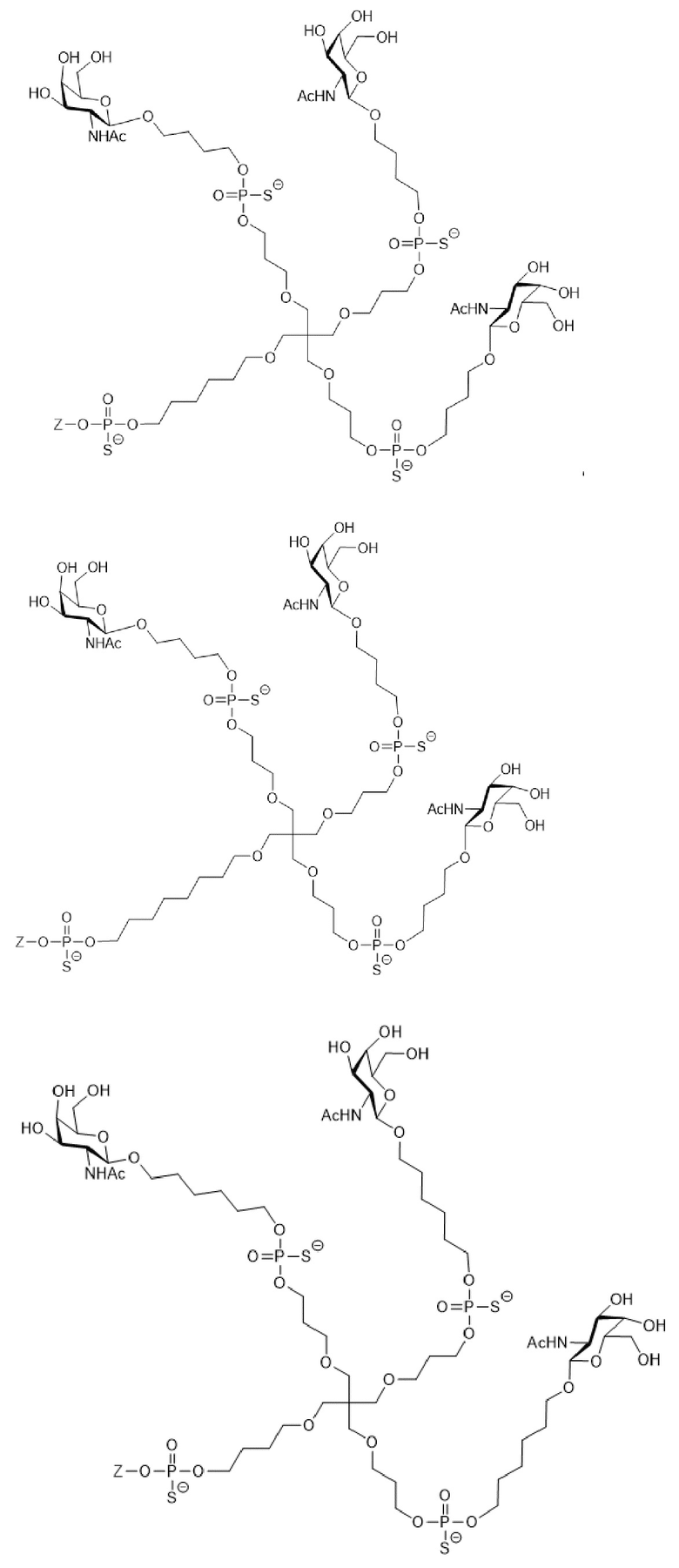

где Z представляет собой нуклеиновую кислоту, определенную в настоящей заявке ранее.
Лиганд формулы (I), (II) или (III) может быть присоединен на 3'-конце первой (антисмысловой) цепи и/или на любом из 3'- и/или 5'-конца второй (смысловой) цепи. Нуклеиновая кислота может содержать более одного лиганда формулы (I), (II) или (III). Однако предпочтительным является один лиганд формул (I), (II) или (III), поскольку одного такого лиганда достаточно для эффективного нацеливания нуклеиновой кислоты на клетки-мишени.
Предпочтительно 5'-конец первой (антисмысловой) цепи не присоединен к лиганду формулы (I), (II) или (III), поскольку лиганд в этом положении потенциально может препятствовать биологической активности нуклеиновой кислоты.
Нуклеиновая кислота с одним лигандом формулы (I), (II) или (III) на 5'-конце цепи имеет более простую структуру и, следовательно, ее дешевле синтезировать, чем аналогичную нуклеиновую кислоту с аналогичным лигандом на 3'-конце. Таким образом, предпочтительно один лиганд любой из формул (I), (II) или (III) ковалентно присоединен (конъюгирован с ним) к 5'-концу второй цепи нуклеиновой кислоты.
Согласно одному варианту реализации настоящего изобретения нуклеиновая кислота конъюгирована с лигандом, который содержит липид, и более предпочтительно с лигандом, который содержит холестерин.
Конъюгат согласно настоящему изобретению может содержать любую нуклеиновую кислоту, описанную в настоящей заявке, конъюгированную с любым лигандом или лигандами, описанными в настоящей заявке.
Настоящее изобретение также относится к конъюгату для ингибирования экспрессии гена LPA в клетке, причем указанный конъюгат содержит часть нуклеиновой кислоты, содержащую нуклеиновую кислоту согласно любому аспекту настоящего изобретения, и по меньшей мере одну часть лиганда, причем указанная часть нуклеиновой кислоты содержит по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи РНК и по меньшей мере часть второй цепи РНК, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с указанного гена LPA, указанная по меньшей мере одна часть лиганда содержит линкерную группу, предпочтительно линкерную группу, происходящую из серинола, а также нацеливающий лиганд для нацеливания в условиях in vivo на клетки и конъюгированный исключительно с 3' и/или 5'-концами одной или обеих цепей РНК, причем 5'-конец первой цепи РНК не конъюгирован, при этом:
(i) вторая цепь РНК конъюгирована на 5'-конце с нацеливающим лигандом, и при этом (a) вторая цепь РНК также конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец первой цепи РНК не конъюгирован; или (b) первая цепь РНК конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец второй цепи РНК не конъюгирован; или (c) обе вторая цепь РНК и первая цепь РНК также конъюгированы на 3'-концах с нацеливающим лигандом; или
(ii) обе вторая цепь РНК и первая цепь РНК конъюгированы на 3'-концах с нацеливающим лигандом и 5'-конец второй цепи РНК не конъюгирован.
Согласно варианту реализации настоящего изобретения вторая цепь РНК (т.е. смысловая цепь) конъюгирована на 5'-конце с нацеливающим лигандом, первая цепь РНК (т.е. антисмысловая цепь) конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец второй цепи РНК (т. е. смысловой цепи) не конъюгирован, так, что образуется конъюгат со схематической структурой, представленной на Фигуре 40А.
Согласно варианту реализации настоящего изобретения вторая цепь РНК (т.е. смысловая цепь) конъюгирована на 5'-конце с нацеливающим лигандом, вторая цепь РНК (т.е. смысловая цепь) также конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец первой цепи РНК (т.е. антисмысловой цепи) не конъюгирован, так, что образуется конъюгат со схематической структурой, представленной на Фигуре 40B.
Согласно варианту реализации настоящего изобретения обе вторая цепь РНК (т.е. смысловая цепь) и первая цепь РНК (т.е. антисмысловая цепь) конъюгированы на 3'-концах с нацеливающим лигандом, и 5'-конец второй цепи РНК (т.е. смысловая цепь) не конъюгирован, так, что образуется конъюгат со схематической структурой, представленной на Фигуре 40C.
Согласно варианту реализации настоящего изобретения вторая цепь РНК (т.е. смысловая цепь) конъюгирована на 5'-конце с нацеливающим лигандом, и обе вторая цепь РНК (т.е. смысловая цепь) и первая цепь РНК (т.е. антисмысловая цепь) также конъюгированы на 3'-концах с нацеливающим лигандом, так, что образуется конъюгат со схематической структурой, представленной на Фигуре 40D.
В любом из приведенных выше вариантов реализации  указывает на линкер, с помощью которого лиганд конъюгирован с концами части нуклеиновой кислоты; лиганд может представлять собой группу GalNAc, такую как GalNAc; и схематическая структура, представленная на Фигуре 40Е, представляет часть нуклеиновой кислоты.
указывает на линкер, с помощью которого лиганд конъюгирован с концами части нуклеиновой кислоты; лиганд может представлять собой группу GalNAc, такую как GalNAc; и схематическая структура, представленная на Фигуре 40Е, представляет часть нуклеиновой кислоты.
Эти схематические диаграммы не предназначены для ограничения количества нуклеотидов в первой или второй цепях, а также диаграммы не представляют собой какое-либо ограничение комплементарности оснований или какое-либо другое ограничение.
Лиганды могут быть мономерными или мультимерными (например, димерными, тримерными и т.д.).
Является приемлемым, когда лиганды являются мономерными и, соответственно, содержат одну группу нацеливающего лиганда, например, одну группу GalNAc.
Согласно другому варианту лиганды могут представлять собой димерные лиганды, в которых части лигандов содержат две линкерные группы, такие как линкерные группы, происходящие из серинола, или линкерные группы, отличные от серинола, каждая из которых соединена с одной группой нацеливающего лиганда.
Лиганды могут представлять собой тримерные лиганды, в которых части лигандов содержат три линкерные группы, такие как линкерные группы, происходящие из серинола, или линкерные группы, отличные от серинола, каждая из которых соединена с одной группой нацеливающего лиганда.
Две или три линкерные группы, происходящие из серинола, могут быть соединены последовательно, например, как показано ниже:

где n равно 1 или 2, а Y представляет собой S или O.
Предпочтительно лиганды являются мономерными.
Является приемлемым, когда конъюгированные цепи РНК конъюгируют с нацеливающим лигандом за счет линкерной группы, включая дополнительный линкер, причем указанный дополнительный линкер представляет собой или содержит насыщенную, неразветвленную или разветвленную C1-15 алкильную цепь, при этом необязательно один или более атомов углерода (например, 1, 2 или 3 атома углерода, является приемлемым, когда 1 или 2, в частности, 1), замещены гетероатомом, выбранным из O, N, S(O)p, где p равно 0, 1 или 2 (например, группа CH2 замещена O или NH, или S, или SO2, или группа -CH3 на конце цепи или на ветви замещена OH или NH2), причем указанная цепь необязательно замещена одной или более оксогруппами (например, 1-3, например, 1 группой).
Является приемлемым, когда линкерная группа представляет собой линкерную группу, происходящую из серинола.
Является более приемлемым, когда дополнительный линкер содержит насыщенную неразветвленную C1-15 алкильную цепь, в которой один или более атомов углерода (например, 1, 2 или 3 атома углерода, является приемлемым, когда 1 или 2, в частности, 1) замещен (ы) атомом кислорода.
Является более приемлемым, когда дополнительный линкер содержит ПЭГ-цепь.
Является более приемлемым, когда дополнительный линкер содержит насыщенную неразветвленную C1-15 алкильную цепь.
Является более приемлемым, когда дополнительный линкер содержит насыщенную неразветвленную C1-6 алкильную цепь.
Является более приемлемым, когда дополнительный линкер содержит насыщенную неразветвленную C4 или C6 алкильную цепь, например, C4 алкильную цепь.
Согласно варианту реализации настоящего изобретения представляет собой соединяющую группу формулы (V):

в которой n, Y и L1 определены ниже, и O фосфогруппы присоединен к концевому олигонуклеозиду цепей РНК.
Таким образом, в варианте реализации часть нацеливающего лиганда представляет собой соединяющую группу формулы (VI):
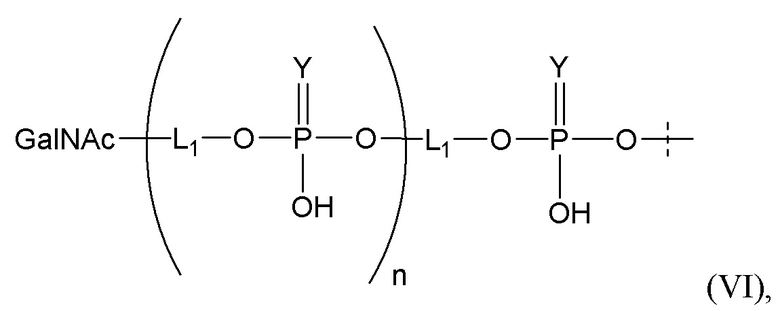
в которой n, Y и L1 определены ниже, и O фосфогруппы присоединен к концевому олигонуклеозиду цепей РНК.
Является приемлемым, когда представляет собой соединяющую группу формулы (XIV):
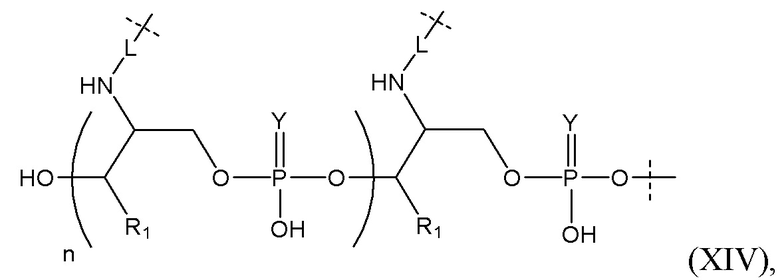
в которой n, Y, R1 и L определены ниже, L соединен с нацеливающим лигандом, например, GalNAc, и O фосфогруппы присоединен к концевому олигонуклеозиду цепей РНК.
Является приемлемым, когда часть нацеливающего лиганда представляет собой соединяющую группу формулы (IV):
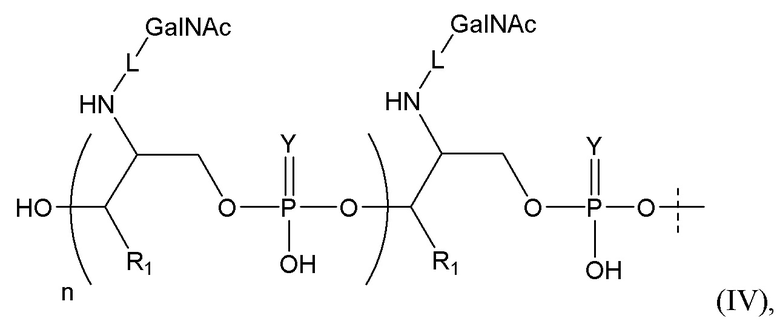
в которой n, Y, R1 и L определены ниже, и O фосфогруппы присоединен к концевому олигонуклеозиду цепей РНК.
Является приемлемым, когда представляет собой соединяющую группу формулы (VII):

в которой n, Y и L2 определены ниже, и O фосфогруппы присоединен к концевому олигонуклеозиду цепей РНК.
Является приемлемым, когда часть нацеливающего лиганда представляет собой соединяющую группу формулы (VIII):
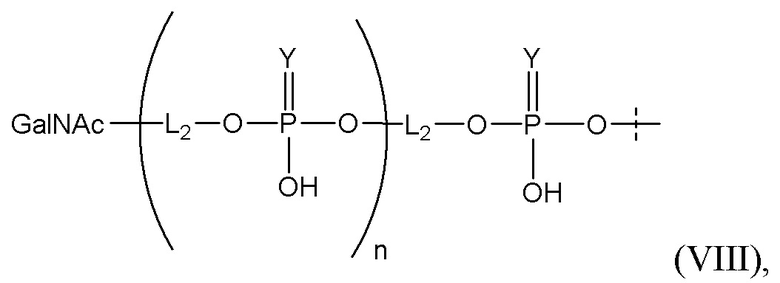
в которой n, Y и L2 определены ниже, и O фосфогруппы присоединен к концевому олигонуклеозиду цепей РНК.
Является приемлемым, когда представляет собой соединяющую группу формулы (IX):
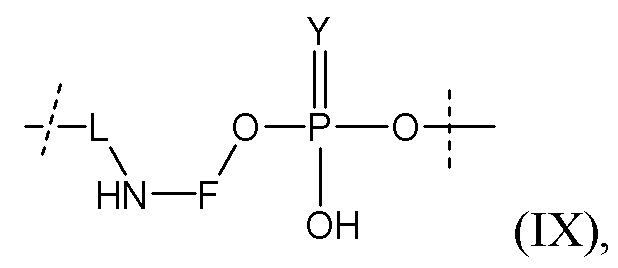
в которой F, Y и L определены ниже, и O фосфогруппы присоединен к концевому олигонуклеозиду цепей РНК.
Является приемлемым, когда часть нацеливающего лиганда представляет собой соединяющую группу формулы (IXa):

в которой F, Y и L определены ниже, и O фосфогруппы присоединен к концевому олигонуклеозиду цепей РНК.
Является приемлемым, когда L представляет собой:

В любой из вышеуказанных структур приемлемо, когда лиганды выбраны из групп GalNAc и галактозы, в частности, групп GalNAc. Согласно другому варианту GalNac может быть замещен другим нацеливающим лигандом, например, сахаридом.
Согласно варианту реализации настоящего изобретения первая цепь РНК представляет собой соединение формулы (X):
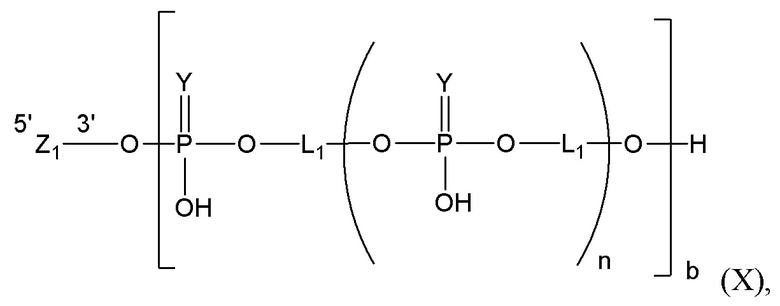
в которой b равно 0 или 1; и
вторая цепь РНК представляет собой соединение формулы (XI):
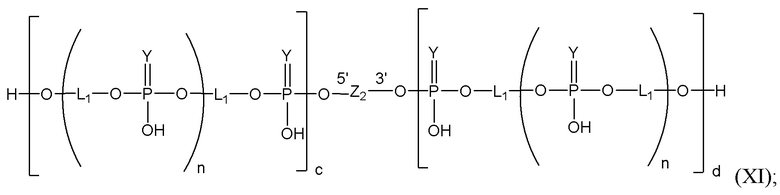
в которой:
с и d независимо равны 0 или 1;
Z1 и Z2 представляют собой части РНК первой и второй цепей РНК, соответственно;
Y представляет собой O или S;
n равно 0, 1, 2 или 3; и
L1 представляет собой линкер, к которому присоединен лиганд;
и при этом b + c + d равно 2 или 3.
Является приемлемым, когда первая цепь РНК представляет собой соединение формулы (XV)

в которой b равно 0 или 1; и
вторая цепь РНК представляет собой соединение формулы (XVI):

в которой с и d независимо равны 0 или 1;
в которой:
Z1 и Z2 представляют собой части РНК первой и второй цепей РНК, соответственно;
Y представляет собой O или S;
R1 представляет собой H или метил;
n равно 0, 1, 2 или 3; и
L является одинаковым или различным в формулах (XV) и (XVId и выбран из группы, состоящей из:
-(CH2)r-C(O)-, где r=2-12;
-(CH2-CH2-O)s-CH2-C(O)-, где s=1-5;
-(CH2)t-CO-NH-(CH2)t-NH-C(O)-, где t независимо составляет 1-5;
-(CH2)u-CO-NH-(CH2)u-C(O)-, где u независимо составляет 1-5; и
-(CH2)v-NH-C(O)-, где v составляет 2-12; и
причем концевой C(O) (если он присутствует) присоединен к группе NH;
и при этом b + c + d равно 2 или 3.
Является приемлемым, когда первая цепь РНК представляет собой соединение формулы (XII):
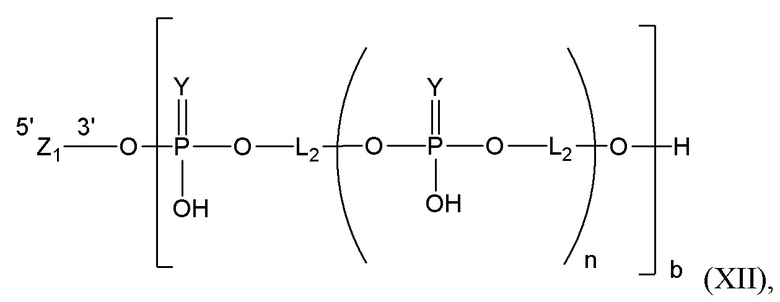
в которой b равно 0 или 1; и
вторая цепь РНК представляет собой соединение формулы (XIII):

в которой:
с и d независимо равны 0 или 1;
Z1 и Z2 представляют собой части РНК первой и второй цепей РНК, соответственно;
Y представляет собой O или S;
n равно 0, 1, 2 или 3; и
L2 является одинаковым или различным в формулах (XII) и (XIII) и одинаковым или различным в группах, заключенных в скобки с индексом b, c и d, и выбран из группы, состоящей из:
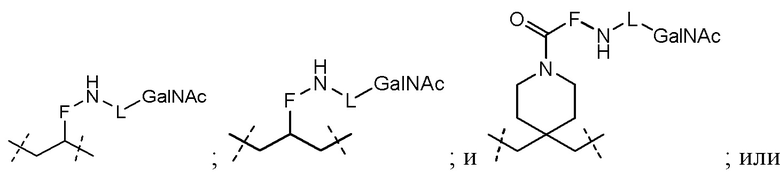
n равно 0 и L2 представляет собой:
и концевая группа OH отсутствует, так, что образуется следующая группа:

где
F представляет собой насыщенную разветвленную или неразветвленную (например, неразветвленную) C1-8 алкильную (например, C1-6 алкильную) цепь, в которой один из атомов углерода необязательно замещен атомом кислорода, при условии, что указанный атом кислорода отделен от другого гетероатома (например, атома O или N) по меньшей мере 2 атомами углерода;
L является одинаковым или различным в формулах (I) и (II) и выбран из группы, состоящей из:
-(CH2)r-C(O)-, где r=2-12;
-(CH2-CH2-O)s-CH2-C(O)-, где s=1-5;
-(CH2)t-CO-NH-(CH2)t-NH-C(O)-, где t независимо составляет 1-5;
-(CH2)u-CO-NH-(CH2)u-C(O)-, где u независимо составляет 1-5; и
-(CH2)v-NH-C(O)-, где v составляет 2-12; и
причем концевой C(O) (если он присутствует) присоединен к группе NH;
и при этом b + c + d равно 2 или 3.
В любой из приведенных выше формул, в которых присутствует GalNAc, GalNAc может быть заменен любым другим нацеливающим лигандом, таким как те, которые упомянуты в настоящей заявке.
Является приемлемым, когда b равно 0, с равно 1 и d равно 1; b равно 1, с равно 0 и d равно 1; b равно 1, с равно 1 и d равно 0; или b равно 1, с равно 1 и d равно 1.
Является более приемлемым, когда b равно 0, с равно 1 и d равно 1; b равно 1, с равно 0 и d равно 1; или b равно 1, с равно 1 и d равно 1.
Является наиболее приемлемым, когда b равно 0, с равно 1 и d равно 1.
Согласно одному варианту реализации настоящего изобретения Y представляет собой O. В другом варианте Y представляет собой S.
Согласно одному варианту реализации настоящего изобретения R1 представляет собой H или метил. Согласно одному варианту реализации настоящего изобретения R1 представляет собой H. Согласно другому варианту реализации настоящего изобретения R1 представляет собой метил.
Согласно одному варианту реализации настоящего изобретения n равно 0, 1, 2 или 3. Является приемлемым, когда n равно 0.
Согласно одному варианту реализации настоящего изобретения L выбран из группы, состоящей из:
-(CH2)r-C(O)-, где r=2-12;
-(CH2-CH2-O)s-CH2-C(O)-, где s=1-5;
-(CH2)t-CO-NH-(CH2)t-NH-C(O)-, где t независимо составляет 1-5;
-(CH2)u-CO-NH-(CH2)u-C(O)-, где u независимо составляет 1-5; и
-(CH2)v-NH-C(O)-, где v составляет 2-12;
при этом концевой C(O) присоединен к группе NH.
Является приемлемым, когда L представляет собой -(CH2)r-C(O)-, где r=2-12. Является приемлемым, когда r=2-6. Является более приемлемым, когда r=4 или 6, например, 4.
Является приемлемым, когда L представляет собой:
Примерные группы F включают (CH2)1-6, например, (CH2)1-4, например, CH2, (CH2)4, (CH2)5 или (CH2)6, или CH2O(CH2)2-3, например, CH2O(CH2)CH3.
Является приемлемым, когда L2 представляет собой:

Является приемлемым, когда L2 представляет собой:
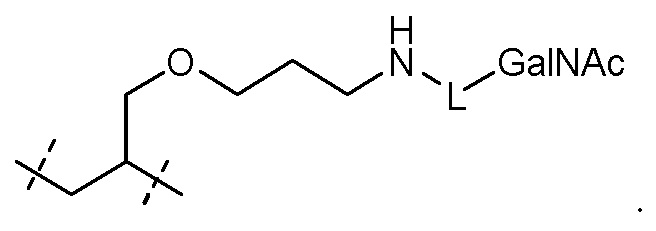
Является приемлемым, когда L2 представляет собой:
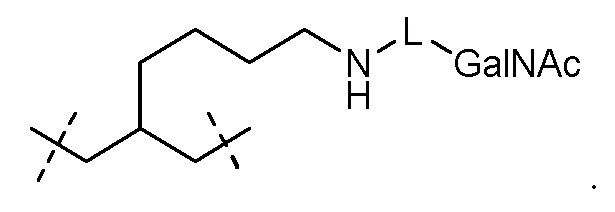
Является приемлемым, когда L2 представляет собой:
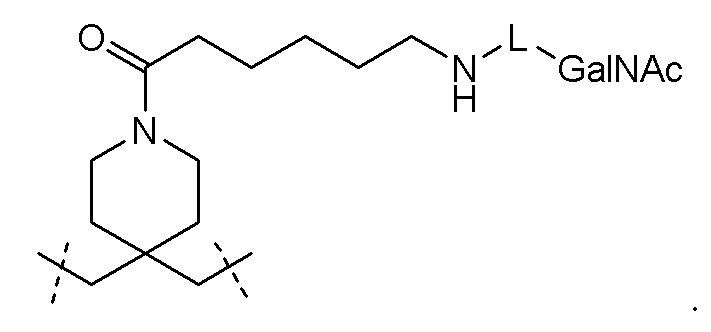
Является приемлемым, когда n равно 0 и L2 представляет собой:

и концевая группа OH отсутствует, так, что образуется следующая группа:
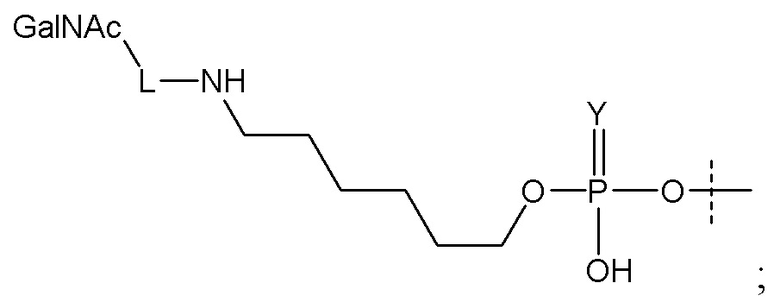 ;
;
где Y определен в другом месте в настоящей заявке.
Как правило, L2 внутри группы, заключенной в скобки с индексом b, c и d, является одинаковым. L2 может быть одинаковым или различным в группах, заключенных в скобки с индексами b, c и d. В варианте реализации L2 в группе, заключенной в скобки с индексом c, аналогична L2 в группе, заключенной в скобки с индексом d. Согласно одному варианту реализации настоящего изобретения L2 в группе, заключенной в скобки с индексом с, отличается от L2 в группе, заключенной в скобки с индексом d. Согласно одному варианту реализации настоящего изобретения L2 в группе, заключенной в скобки с индексами b, c и d, является одинаковой, например, когда линкерная группа представляет собой линкерную группу, происходящую из серинола.
Линкерные группы, происходящие из серинола, могут быть основаны на сериноле с любой стереохимией, т.е. происходят из изомера L-серина, изомера D-серина, рацемического серина или другой комбинации изомеров. В предпочтительном аспекте настоящего изобретения группа серинол-GalNAc (SerGN) имеет следующую стереохимию:
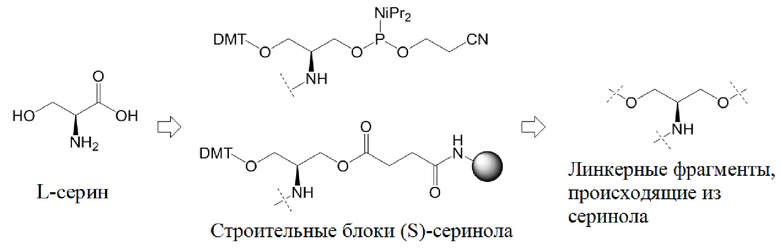
т.е. основан на (S)-сериноламидитном или (S)-серинолсукцинатном твердом поддерживаемом строительном блоке, полученном из изомера L-серина.
Согласно одному варианту реализации настоящего изобретения клетки-мишени представляют собой гепатоциты.
Общие схемы синтеза: 1
Примерные соединения могут быть синтезированы в соответствии со способами, описанными ниже и известными специалисту в данной области техники. Несмотря на то, что схемы иллюстрируют синтез конкретных конъюгатов, следует понимать, что другие заявленные конъюгаты могут быть получены с помощью аналогичных способов. Сборку олигонуклеотидной цепи и линкерных строительных блоков можно выполнять, например, с помощью твердофазного синтеза с применением фосфоамидитной методологии. Твердофазный синтез можно начинать с основания или модифицированного строительного блока, загруженного на lcaa CPG. Цикл связывания при фосфоамидитном синтезе состоит из 1) удаления DMT, 2) удлинения цепи с применением необходимого DMT-замаскированного фосфоамидита и активатора, который может представлять собой бензилтиотетразол (БТТ), 3) кэппирования неудлиненных олигонуклеотидных цепей с последующим окислением P(III) до P(V) либо с помощью йода (если требуется фосфодиэфирная связь), либо EDITH (если требуется фосфотиоатная связь) и затем повторного кэппирования (Cap/Ox/Cap или Cap/Thio/Cap). Конъюгацию GalNAc можно осуществлять путем образования пептидной связи между строительным блоком GalNAc-карбоновая кислота и предварительно собранным и очищенным олигонуклеотидом, имеющим необходимое количество присоединенных амино-модифицированных линкерных строительных блоков. Необходимые строительные блоки либо доступны коммерчески, либо их синтез описан ниже. Все готовые одноцепочечные продукты были проанализированы с помощью АХ-ВЭЖХ для подтверждения их чистоты. Чистота представлена как % FLP (% полноразмерного продукта), который представляет собой процент УФ-площади под сигналом заданного продукта на УФ-кривой анализа конечного продукта методом АХ-ВЭЖХ. Идентичность соответствующих одноцепочечных продуктов была подтверждена с помощью анализа методом ЖХ/МС.
Синтез синтонов
Схема 1: Синтез синтонов с линкером DMT-серинол(TFA)

i) этилтрифторацетат, NEt3, MeOH, 0°C, 16 ч, 2: 86% 5: 90%, ii) DMTCl, пиридин, 0°C, 16 ч, 74%, iii) LiBH4, EtOH/THF (1/1, об./об.), 0°C, 1 ч, 76%, iv) 2-цианоэтил-N,N-диизопропилхлорфосфоамидит, EtNiPr2, CH2Cl2, 56%, v) сукциновый ангидрид, DMAP, пиридин, RT, 16 ч, 38%, vi) HBTU, DIEA, амино-Icaa CPG (500 A), к.т., 18 ч, 29% (загрузка 26 мкмоль/г).
(S)-DMT-серинол(TFA)-фосфоамидит 7 можно синтезировать из производного сложного метилового эфира (L)-серина 1 в соответствии с опубликованными в литературе способами (Hoevelmann et al. Chem. Sci., 2016,7, 128-135).
(S)-DMT-серинол(TFA)-сукцинат 6 может быть получен путем превращения промежуточного соединения 5 с сукциновым ангидридом в присутствии катализатора, такого как DMAP.
Загрузку 6 в твердую подложку, такую как подложка из стекла с заданным размером пор (CPG), можно осуществлять путем образования пептидной связи с твердой подложкой, такой как амино-модифицированная нативная подложка CPG (500 A), с применением связывающего реагента, такого как HBTU. (S)-DMT-серинол(TFA)-сукцинат 6 и связывающий реагент, такой как HBTU, растворяют в растворителе, таком как CH3CN. Основание, такое как диизопропилэтиламин, добавляют к раствору, и реакционную смесь перемешивают в течение 2 мин. К реакционной смеси добавляют твердую подложку, такую как нативная подложка амино-Icaa-CPG (500 A, 3 г, содержание амина: 136 мкмоль/г) с образованием суспензии. Суспензию осторожно встряхивают при комнатной температуре на шейкере, имитирующем движение запястья, в течение 16 ч, затем фильтруют и промывают растворителем, таким как ДХМ и EtOH. Подложку высушивают в вакууме в течение 2 ч. Не вступившие в реакцию амины на подложке могут быть кэппированы путем перемешивания с уксусным ангидридом/лутидином/N-метилимидазолом при комнатной температуре. Промывку подложки можно повторять, как описано выше. Твердую подложку высушивают в вакууме с получением твердой подложки 10.
Схема 2: Синтез GalNAc синтона 9
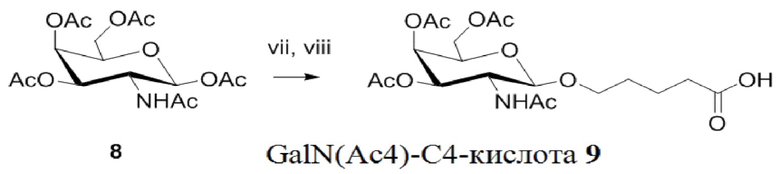
(vii) TMSOTf, ДХМ, гексенол, viii) RuCl3, NaIO4, ДХМ, CH3CN, H2O, 46% в течение двух этапов.
Синтез GalNAc синтона 9 можно выполнять в соответствии со способами, описанными в Nair et al. (2014), начиная с коммерчески доступного пер-ацетилированного галактозамина 8.
Синтез одноцепочечных GalNAc-конъюгатов, происходящих из серинола
Схема 3: Общая процедура синтеза олигонуклеотидов для линкеров, происходящих из серинола
Синтез олигонуклеотидов для 3'-моно-GalNAc-конъюгированных олигонуклеотидов (таких как соединение A0264) показан на Фигуре 13 и обобщен на схеме 3. Синтез начинается с применением (S)-DMT-серинол(TFA)-сукцинат-Icaa-CPG 10, как в примерном соединении A0264. В случае потребности в дополнительных серинольных строительных блоках (S)-DMT-серинол(TFA)амидит (7) применяют в соответствующем цикле твердофазного синтеза. Например, чтобы получить соединение A0329, сборку цепи завершают с применением связывания дополнительного сериноламидита после того, как последовательность оснований полностью собрана. Кроме того, синтез олигонуклеотидов для 5'-моно-GalNAc-конъюгированных олигонуклеотидов можно начинать с твердой подложки, загруженной соответствующим нуклеозидом из его соответствующей последовательности. В примерном соединении A0220 это может быть 2'-fA. Олигонуклеотидную цепь собирают в соответствии с ее последовательностью и, в случае необходимости, применяют строительный блок (S)-DMT-серинол(TFA)-амидит (7). После завершения удлинения цепи удаляют защитную группу DMT последнего связанного амидитного строительного блока, как на этапе 1) цикла фосфоамидитного синтеза.
После завершения последнего этапа в синтезаторе отдельные цепи могут быть отщеплены от твердой подложки путем обработки амином, таким как обработка 40% водн. метиламином. На этом этапе также удаляют любые оставшиеся защитные группы, и обработка метиламином также высвобождает функциональную аминогруппу серинола. Затем каждый из неочищенных продуктов очищали с помощью АХ-ВЭЖХ и гель-проникающей хроматографии (SEC) с получением предшественника олигонуклеотида для дальнейшей конъюгации с GalNAc.
Схема 4: GalNAc-конъюгационный синтез предшественников олигонуклеотидов, происходящих из серинола
После твердофазного синтеза конъюгацию с GalNAc осуществляли путем предварительной активации GalN(Ac4)-C4-кислоты (9) с помощью связывающего реагента для образования пептидной связи, такого как HBTU. Предварительно активированную кислоту 9 затем подвергали реакции с аминогруппами в 11 (например, A0264) с образованием промежуточных GalN(Ac4)-конъюгатов. Ацетильные группы, защищающие гидроксильные группы в группах GalNAc, отщепляли с помощью обработки метиламином с получением целевых примерных соединений 12 (например, A0268), которые дополнительно очищали с помощью АХ-ВЭЖХ и SEC.
Синтез одноцепочечных GalNAc-конъюгатов, происходящих не из серинола
Амино-модифицированные строительные блоки, отличные от серинола, коммерчески доступны от различных поставщиков и могут применяться вместо серинола, чтобы обеспечить реакционноспособные аминогруппы, которые обеспечивают конъюгацию GalNAc. Например, коммерчески доступные строительные блоки, показанные в таблице 6 ниже, можно применять для того чтобы обеспечить амино-модифицированные предшественники олигонуклеотидов, происходящие не из серинола, 14 (схема 5A) с применением загруженного амино-модификатором CPG, например, от 10-1 до 10-3, с последующей сборкой последовательности, как описано выше, и в завершение связыванием фосфоамидитов амино-модификаторов, таких как 13-1, 13-2 или 13-4.
Например, для получения 14 (A0653) GlyC3Am-CPG (10-2) применяли в комбинации с GlyC3Am-амидитом 13-2. Структурно отличающиеся модификаторы можно применять для получения 14, например, для получения A0651 C7Am-CPG применяли в комбинации с C6Am-амидитом в качестве второй амино-модификации. Аналогичным образом коммерчески доступный загруженный амино-модификатором CPG 10-5 и амино-модифицированный фосфоамидит 13-5 можно применять для синтеза амино-модифицированных молекул-предшественников 14 (A0655).
Схема 5: Общая процедура синтеза олигонуклеотидов
Полученные предшественники олигонуклеотидов 14 затем могут быть конъюгированы с GalN(Ac4)-C4-кислотой (9) с получением целевых примерных соединений 15 (схема 6).
Схема 6: GalNAc-конъюгационный синтез предшественников олигонуклеотидов
Синтез одноцепочечных трехантенных GalNAc-конъюгатов в эталонных конъюгатах 3-4
Синтез олигонуклеотидов для конъюгированной с трехантенным GalNAc-кластером миРНК показан на Фигуре 14. Сборку олигонуклеотидной цепи начинали с применением загруженной основанием подложки, например, 5'DMT-2'-FdA(bz)-сукцинат-lcaa-CPG, как в примерном соединении A0006. Цикл связывания фосфоамидитного синтеза, состоящий из 1) удаления DMT, 2) удлинения цепи с применением необходимого DMT-замаскированного фосфоамидита, 3) кэппирования неудлиненных олигонуклеотидных цепей с последующим окислением P (III) до P (V) либо с помощью йода, либо EDITH (если требуется фосфотиоатная связь) и в завершение кэппирования (Cap/Ox/Cap или Cap/Thio/Cap), повторяют до тех пор, пока не будет достигнута полная длина продукта. Для конъюгации на колонке трехвалентного трехантенного GalNAc-кластера применяли аналогичный цикл синтеза с применением необходимого трехвалентного разветвляющего амидита C4XLT-phos, за которым следовал другой раунд цикла синтеза с применением GalNAc-амидита ST23-phos. После завершения этого последнего этапа в синтезаторе олигонуклеотид отщепляли от твердой подложки, и дополнительные защитные группы могут быть удалены с помощью обработки метиламином. Затем каждый из неочищенных продуктов очищали с помощью АХ-ВЭЖХ и SEC.
Общая процедура образования двойной цепи
Чтобы получить двухцепочечные конъюгаты, отдельные одиночные цепи растворяют в концентрации 60 OD/мл в H2O. Оба раствора отдельных олигонуклеотидов могут быть добавлены вместе в реакционный сосуд. Для мониторинга реакции можно выполнять титрование. Первую цепь добавляют с 25% избытком по сравнению со второй цепью, что определяется по УФ-поглощению при 260 нм. Реакционную смесь нагревают, например, до 80°C в течение 5 минут, а затем медленно охлаждают до комнатной температуры. Образование двойной цепи можно контролировать по образованию ионных пар с помощью ВЭЖХ с обращенной фазой. На основании УФ-области остаточной одиночной цепи можно рассчитать необходимое количество второй цепи и добавить его в реакционную смесь. Реакционную смесь снова нагревают, например, до 80°C, и медленно охлаждают до комнатной температуры. Эту процедуру можно повторять до тех пор, пока не будет обнаружено менее 10% остаточной одиночной цепи.
Вышеуказанный способ (включая схемы 1-6 и Фигуры 13 и 14) можно легко адаптировать для замены GalNAc другим нацеливающим лигандом, например, сахаридом.
В любом из вышеупомянутых аспектов вместо конъюгации после твердофазного синтеза можно получить заранее образованный серинол(GN)-фосфоамидит и применять его для конъюгации на колонке.
Общие схемы синтеза: 2
Примерные соединения могут быть синтезированы в соответствии со способами, описанными ниже и известными специалисту в данной области техники. Сборку олигонуклеотидной цепи и линкерных строительных блоков можно выполнять, например, с помощью твердофазного синтеза с применением фосфоамидитной методологии. Конъюгацию GalNAc можно осуществлять путем образования пептидной связи между строительным блоком GalNAc-карбоновой кислоты и предварительно собранным и очищенным олигонуклеотидом, содержащим необходимое количество присоединенных амино-модифицированных линкерных строительных блоков.
Схема 1: Синтез синтонов с линкером DMT-серинол(TFA)

i) этилтрифторацетат, NEt3, MeOH, 0°C, 16 ч, 2: 86% 5: 90%, ii) DMTCl, пиридин, 0°C, 16 ч, 74%, iii) LiBH4, EtOH/THF (1/1, об./об.), 0°C, 1 ч, 76%, iv) 2-цианоэтил-N,N-диизопропилхлорфосфоамидит, EtNiPr2, CH2Cl2, 56%, v) сукциновый ангидрид, DMAP, пиридин, к.т., 16 ч, 38%, vi) HBTU, DIEA, амино-lcaa CPG (500A), к.т., 18 ч, 29%.
DMT-серинол(TFA)-фосфоамидит 7 можно синтезировать из производного серинола 1 в соответствии с опубликованными в литературе способами (Hoevelmann et al. Chem. Sci., 2016,7, 128-135).
DMT-серинол(TFA)-сукцинат 6 может быть получен путем превращения промежуточного соединения 5 с сукциновым ангидридом в присутствии катализатора, такого как DMAP.
Загрузку 6 на твердую подложку, такую как подложка CPG, можно осуществлять путем образования пептидной связи с твердой подложкой, такой как амино-модифицированная нативная подложка CPG (500A), с применением связывающего реагента, такого как HBTU. DMT-серинол(TFA)-сукцинат 6 и связывающий реагент, такой как HBTU, растворяют в растворителе, таком как CH3CN. Основание, такое как диизопропилэтиламин, добавляют к раствору и реакционную смесь перемешивают в течение 2 мин. К реакционной смеси добавляют твердую подложку, такую как нативная подложка амино-Icaa-CPG (500 A, 3 г, содержание амина: 136 мкмоль/г), с образованием суспензии. Суспензию осторожно встряхивают при комнатной температуре на шейкере, имитирующем движение запястья, в течение 16 ч, затем фильтруют и промывают растворителем, таким как ДХМ и EtOH. Подложку высушивают в вакууме в течение 2 ч. Не вступившие в реакцию амины на подложке могут быть кэппированы путем перемешивания с уксусным ангидридом/лутидином/N-метилимидазолом при комнатной температуре. Промывку подложки можно повторять, как описано выше. Твердую подложку высушивают в вакууме с получением твердой подложки 10.
Схема 2: Синтез GalNAc синтона 9

(vii) TMSOTf, ДХМ, гексенол, viii) RuCl3, NaIO4, ДХМ, CH3CN, H2O, 46% в течение двух этапов.
Синтез GalNAc синтона 9 можно осуществлять в соответствии со способами, описанными в Nair et al. J. Am. Chem. Soc., 2014, 136 (49), pp 16958-16961, начиная с коммерчески доступного пер-ацетилированного галактозамина 8.
Схема 3: Общая процедура синтеза олигонуклеотидов
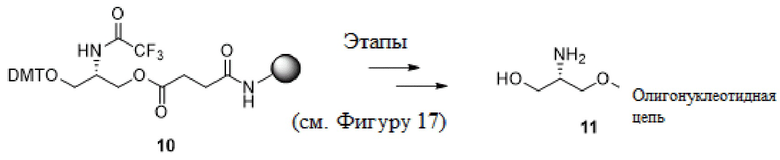
Все олигонуклеотиды могут быть синтезированы на синтезаторе AKTA oligopilot 10 с применением стандартных фосфоамидитных реакций, которые подробно описаны ниже.
Синтез олигонуклеотидов из 3'- и 5'-конъюгированных с GalNAc предшественников олигонуклеотидов (таких как соединение X0385B-prec) представлен на Фигуре 17 и обобщен на схеме 3. Синтез начинают с применением DMT-серинол(TFA)-сукцинат-lcaa-CPG 10. Цикл фосфоамидитного синтеза применяют до тех пор, пока не будет достигнута полная длина продукта. После завершения удлинения цепи защитная группа DMT последнего связанного амидитного строительного блока может быть удалена таким же образом, как и в каждом отдельном цикле синтеза. Чтобы завершить синтез примерного соединения X0385B-prec (которое содержит линкерную группу, происходящую из серинола, на 3' и 5'-концах второй цепи), сборку цепи заканчивали с применением связывания дополнительного сериноламидита после того, как последовательность оснований была полностью собрана.
После завершения последнего этапа в синтезаторе одиночные цепи могут быть отщеплены от твердой подложки путем обработки амином, таким как 40% водн. метиламин. На этом этапе также удаляют любые оставшиеся защитные группы, и обработка метиламином также высвобождает функциональную аминогруппу серинола. Полученный неочищенный олигонуклеотид может быть очищен с помощью ионообменной хроматографии (Resource Q, 6 мл, GE Healthcare) на системе AKTA Pure HPLC с применением градиента, такого как градиент хлорида натрия. Избыток соли после очистки с помощью ионообменной хроматографии (ИХ) можно удалить методом SEC, чтобы получить амино-модифицированный предшественник олигонуклеотида 11. Содержащие продукт фракции объединяют, обессоливают на колонке для гель-проникающей хроматографии (Zetadex, EMP Biotech) и лиофилизируют.
Схема 4: Общая процедура для конъюгации GalNAc

(i) 9, HBTU, DIPEA, ДМСО; 11, H2O, ДМСО, DIPEA; затем активирован 9, 11; затем 40% MeNH2, H2O.
Конъюгацию GalNAc синтона 9, как описано выше, можно осуществлять путем связывания 9 с функциональной аминогруппой серинола соответствующей олигонуклеотидной цепи 11 с применением стандартных условий связывания пептидов, известных специалисту в данной области техники. Например, соответствующую амино-модифицированную молекулу-предшественника 11 растворяют в H2O и добавляют полярный растворитель, такой как ДМСО (например, ДМСО/H2O, 2/1, об./об.), а затем основание, такое как DIPEA (например, 2,5% от общего объема). В отдельном реакционном сосуде можно выполнять предварительную активацию GalNAc синтона 9 с помощью взаимодействия 2 экв. (на функциональную аминогруппу в амино-модифицированном предшественнике олигонуклеотида) компонента карбоновой кислоты с 2 экв. связывающего реагента, такого как HBTU, в присутствии 8 экв. основания, такого как DIPEA, в полярном растворителе, таком как ДМСО. Через 2 мин активированное соединение 9 добавляют к раствору соответствующей амино-модифицированной молекулы-предшественника 11. Ход реакции можно контролировать с помощью ЖХ/МС или АХ-ВЭЖХ. После завершения реакции конъюгирования (например, 30 мин) неочищенный продукт может быть осажден путем добавления 10× iPrOH 0,1×2 М NaCl и собран с помощью центрифугирования и декантации. Ацетилгидрокси-защитные группы удаляют в основных условиях, таких как 40% MeNH2 (1 мл на 500 OD). Через 15 мин при комнатной температуре добавляют H2O (1:10 об./об.) и выделяют соединение 12 (такое как X0385B, показанное на Фигуре 17), повторно очищают с помощью анионообменной и гель-проникающей хроматографии и затем лиофилизируют.
Общая процедура образования двойной цепи
Отдельные одиночные цепи растворяют в концентрации 60 OD/мл в H2O. Оба раствора отдельных олигонуклеотидов могут быть добавлены вместе в реакционный сосуд. Для мониторинга реакции можно выполнять титрование. Первую цепь добавляют с 25% избытком по сравнению со второй цепью, что определяется по УФ-поглощению при 260 нм. Реакционную смесь нагревают, например, до 80°C, в течение 5 минут, а затем медленно охлаждают до комнатной температуры. Образование двойной цепи можно контролировать по образованию ионных пар с помощью ВЭЖХ с обращенной фазой. На основании УФ-площади остаточной одиночной цепи можно рассчитать необходимое количество второй цепи и добавить его в реакционную смесь. Реакционную смесь снова нагревают, например, до 80°C, и медленно охлаждают до комнатной температуры. Эту процедуру можно повторять до тех пор, пока не будет обнаружено менее 10% остаточной одиночной цепи.
Вышеупомянутый способ (включая схемы 1-4 и Фигуру 17) можно легко адаптировать для замены GalNac другим нацеливающим лигандом, например, сахаридом.
Настоящее изобретение относится к конъюгату для ингибирования экспрессии гена LPA в клетке, причем указанный конъюгат содержит часть нуклеиновой кислоты и части лиганда, причем указанная часть нуклеиновой кислоты содержит по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи РНК и по меньшей мере часть второй цепи РНК, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с указанного гена LPA, при этом указанные части лиганда содержат линкерную группу и нацеливающий лиганд для нацеливания в условиях in vivo на клетки и конъюгированный исключительно с 3'- и/или 5'-концами одной или обеих цепей РНК, причем 5'-конец первой цепи РНК не конъюгирован, при этом:
(i) вторая цепь РНК конъюгирована на 5'-конце с нацеливающим лигандом, и при этом (a) вторая цепь РНК также конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец первой цепи РНК не конъюгирован; или (b) первая цепь РНК конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец второй цепи РНК не конъюгирован; или (c) обе вторая цепь РНК и первая цепь РНК также конъюгированы на 3'-концах с нацеливающим лигандом; или
(ii) обе вторая цепь РНК и первая цепь РНК конъюгированы на 3'-концах с нацеливающим лигандом, и 5'-конец второй цепи РНК не конъюгирован.
Линкерная группа может представлять собой, например, линкерную группу, происходящую из серинола, или один из других типов линкеров, описанных в настоящей заявке.
Согласно настоящему изобретению предложена, в качестве другого аспекта, нуклеиновая кислота для ингибирования экспрессии LPA в клетке, содержащая по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая является по меньшей мере частично комплементарной первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, причем указанная первая цепь содержит нуклеотидную последовательность, выбранную из следующих последовательностей: SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73, при этом нуклеиновая кислота конъюгирована с лигандом. Вторая цепь может содержать нуклеотидную последовательность SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 64, 66, 68, 70, 72 или 74. Нуклеотиды первой и/или второй цепей могут быть модифицированы, как описано в настоящей заявке.
Предпочтительно нуклеиновая кислота содержит SEQ ID NO: 5 или SEQ ID NO: 9 и SEQ ID NO: 6 или SEQ ID NO: 10, конъюгированные с лигандом формулы I (представленной выше), причем указанный лиганд конъюгирован с описанной нуклеиновой кислотой, и при этом первая цепь модифицирована с применением модификации 2'-OMe на нечетных нуклеотидах и модифицирована с применением 2'-F на четных нуклеотидах, а вторая цепь модифицирована с применением 2'-OMe на четных нуклеотидах и модифицирована с применением 2'-F на нечетных нуклеотидах.
Более предпочтительно нуклеиновая кислота содержит SEQ ID NO: 5 или SEQ ID NO: 9 и SEQ ID NO: 6 или SEQ ID NO: 10, причем указанная нуклеиновая кислота конъюгирована с лигандом формулы I (представленной выше), и, кроме того, нуклеиновая кислота имеет профиль модификации, представленный ниже, который представляет собой выдержку из таблицы 1, представленной в настоящей заявке.
где конкретные модификации обозначены номерами
1=2'-F-dU,
2=2'-F-dA,
3=2'-F-dC,
4=2'-F-dG,
5=2'-OMe-rU;
6=2'-OMe-rA;
7=2'-OMe-rC;
8=2'-OMe-rG.
Лиганд может содержать GalNAc, и на Фигуре 4А или Фигуре 4В дополнительно проиллюстрированы примеры настоящего изобретения.
Согласно настоящему изобретению также предложены фармацевтические композиции, содержащие нуклеиновую кислоту или конъюгированную нуклеиновую кислоту согласно настоящему изобретению. Фармацевтические композиции можно применять в качестве лекарственных средств или в качестве диагностических агентов, отдельно или в комбинации с другими агентами. Например, один или более конъюгатов нуклеиновой кислоты согласно настоящему изобретению можно комбинировать со средством для доставки (например, липосомами) и/или вспомогательными веществами, такими как носители, разбавители. Также могут быть добавлены другие агенты, такие как консерванты и стабилизаторы. Способы доставки нуклеиновых кислот известны в данной области техники и известны специалисту в данной области техники.
Нуклеиновую кислоту или конъюгированную нуклеиновую кислоту согласно настоящему изобретению также можно вводить в комбинации с другими терапевтическими соединениями, либо вводить по отдельности, либо одновременно, например, в виде комбинированной стандартной дозы. Настоящее изобретение также включает фармацевтическую композицию, содержащую одну или более нуклеиновых кислот или конъюгированных нуклеиновых кислот в соответствии с настоящим изобретением в физиологически/фармацевтически приемлемом вспомогательном веществе, таком как стабилизатор, консервант, разбавитель, буфер и тому подобное.
Уровни дозировки для лекарственного средства и фармацевтических композиций согласно настоящему изобретению могут быть определены специалистами в данной области техники с помощью стандартных экспериментов. Согласно одному варианту реализации настоящего изобретения стандартная доза может содержать от приблизительно 0,01 мг/кг до приблизительно 100 мг/кг массы тела нуклеиновой кислоты или конъюгированной нуклеиновой кислоты. Согласно другому варианту доза может составлять от 10 мг/кг до 25 мг/кг массы тела или от 1 мг/кг до 10 мг/кг массы тела, или от 0,05 мг/кг до 5 мг/кг массы тела, или от 0,1 мг/кг до 5 мг/кг массы тела, или от 0,1 мг/кг до 1 мг/кг массы тела, или от 0,1 мг/кг до 0,5 мг/кг массы тела, или от 0,5 мг/кг до 1 мг/кг массы тела. Уровни дозировки также могут быть рассчитаны с помощью других параметров, таких как, например, площадь поверхности тела.
Фармацевтическая композиция может представлять собой стерильную инъецируемую водную суспензию или раствор, или может быть в лиофилизированной форме.
Фармацевтические композиции и лекарственные средства согласно настоящему изобретению могут быть введены субъекту-млекопитающему в фармацевтически эффективной дозе. Млекопитающее может быть выбрано из человека, примата, отличного от человека, обезьяны или древесного примата, собаки, кошки, лошади, крупного рогатого скота, свиньи, козы, овцы, мыши, крысы, хомяка, ежа и морской свинки или других целесообразных видов. Исходя из этого, в настоящей заявке формулировка «LPA» или «LPA» обозначает нуклеиновую кислоту или белок в любом из вышеупомянутых видов, если они экспрессируются у него естественным или искусственным образом, но предпочтительно эта формулировка обозначает нуклеиновые кислоты или белки человека.
Дополнительный аспект настоящего изобретения относится к нуклеиновой кислоте или конъюгированной нуклеиновой кислоте согласно настоящему изобретению или фармацевтической композиции, содержащей нуклеиновую кислоту или конъюгированную нуклеиновую кислоту согласно настоящему изобретению, для применения при лечении заболевания, нарушения или синдрома. Лечение может заключаться в предотвращении и уменьшении риска развития инсульта, атеросклероза, тромбоза или сердечно-сосудистых заболеваний, таких как ишемическая болезнь сердца или стеноз аорты, и любого другого заболевания или патологии, ассоциированных с повышенными уровнями частиц, содержащих Lp(a). Настоящее изобретение включает фармацевтическую композицию, содержащую одну или более нуклеиновых кислот или конъюгированных нуклеиновых кислот в соответствии с настоящим изобретением в физиологически/фармацевтически приемлемом вспомогательном веществе, таком как стабилизатор, консервант, разбавитель, буфер и тому подобное.
Фармацевтическая композиция может представлять собой стерильную инъецируемую водную суспензию или раствор, или может быть в лиофилизированной форме, или может быть прикреплена, абсорбирована или включена в любое другое подходящее галеновое транспортирующее вещество или внутрь его, такое как драже, таблетки, капсулы, наночастицы, гели, таблетки, гранулы или сходные структуры.
Нуклеиновая кислота, описанная в настоящей заявке, может быть способна ингибировать экспрессию LPA. Нуклеиновая кислота, описанная в настоящей заявке, может быть способна частично ингибировать экспрессию LPA. Ингибирование может быть полным, т.е. 0% по сравнению с уровнем экспрессии LPA в отсутствие нуклеиновой кислоты согласно настоящему изобретению. Ингибирование экспрессии LPA может быть частичным, т.е. оно может составлять 15%, 20%, 30%, 40%, 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95% или промежуточные значения от экспрессии LPA в отсутствие нуклеиновой кислоты согласно настоящему изобретению. Ингибирование может длиться 2 недели, 3 недели, 4 недели, 5 недель, 6 недель, 7 недель, 8 недель, 10 недель, 11 недель, 12 недель, 13 недель, 14 недель или до 3 месяцев при применении у субъекта, такого как пациент-человек. Нуклеиновую кислоту или конъюгированную нуклеиновую кислоту согласно настоящему изобретению или композиции, содержащие ее, можно применять в схеме, включающей обработки один или два раза в неделю, каждую неделю, каждые две недели, каждые три недели, каждые четыре недели, каждые пять недель, каждые шесть недель, каждые семь недель или каждые восемь недель, или в схемах с различной частотой дозирования, таких как комбинации вышеупомянутых интервалов. Нуклеиновую кислоту можно применять для подкожного, внутривенного или любого другого пути применения, такого как пероральный, ректальный или внутрибрюшинный.
В клетках и/или у субъектов, обработанных или получающих нуклеиновую кислоту или конъюгированную нуклеиновую кислоту согласно настоящему изобретению, экспрессия LPA может ингибироваться по сравнению с необработанными клетками и/или субъектами на величину в диапазоне от 15% до 100%, но по меньшей мере приблизительно 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 98% или 100% или промежуточные значения. Уровень ингибирования может обеспечить лечение заболевания, ассоциированного с экспрессией или сверхэкспрессией LPA, или может служить для дальнейшего изучения функций и физиологических ролей продукта гена LPA.
Дополнительный аспект настоящего изобретения относится к нуклеиновой кислоте или конъюгированной нуклеиновой кислоте согласно настоящему изобретению при изготовлении лекарственного средства для лечения заболевания, нарушения или синдромов, таких как те, которые перечислены выше, или дополнительных патологий, ассоциированных с повышенными уровнями Lp(a), или при дополнительных терапевтических подходах, когда желательным является ингибирование экспрессии LPA.
Настоящее изобретение также включает способ лечения или предотвращения заболевания, нарушения или синдрома, таких как те, которые перечислены выше, включающий введение фармацевтической композиции, содержащей нуклеиновую кислоту или конъюгированную нуклеиновую кислоту, описанные в настоящей заявке, индивидууму, нуждающемуся в лечении (чтобы улучшить такие патологии). Композицию нуклеиновой кислоты можно вводить в схеме, включающей обработки два раза в неделю, один раз в неделю, каждые две недели, каждые три недели, каждые четыре недели, каждые пять недель, каждые шесть недель, каждые семь недель или каждые восемь недель, или в схемах с различной частотой дозирования, таких как комбинации вышеупомянутых интервалов. Нуклеиновая кислота или конъюгированная нуклеиновая кислота могут быть предназначены для подкожного или внутривенного применения или других путей применения, таких как пероральный, ректальный или внутрибрюшинный.
Нуклеиновую кислоту или конъюгированную нуклеиновую кислоту согласно настоящему изобретению также можно вводить для применения в комбинации с другими терапевтическими соединениями, либо вводить по отдельности, либо одновременно, например, в виде комбинированной стандартной дозы. Также возможна молекулярная конъюгация с другими биологически активными молекулярными частицами, такими как пептиды, клеточные или искусственные лиганды или малые и большие молекулы.
Нуклеиновую кислоту или конъюгированную нуклеиновую кислоту согласно настоящему изобретению можно получить с применением обычных способов в данной области техники, включая химический синтез или экспрессию нуклеиновой кислоты как в условиях in vitro (например, прерванная транскрипция), так и в условиях in vivo. Например, с применением твердофазного химического синтеза или с применением вектора экспрессии на основе нуклеиновой кислоты, включая вирусные производные или частично или полностью синтетические системы экспрессии. Согласно одному варианту реализации настоящего изобретения вектор экспрессии можно применять для получения нуклеиновой кислоты согласно настоящему изобретению в условиях in vitro, внутри промежуточного организма-хозяина или типа клетки, внутри промежуточного или конечного организма или внутри желаемой клетки-мишени. Способы получения (синтеза или ферментативной транскрипции) нуклеиновой кислоты, описанные в настоящей заявке, известны специалистам в данной области техники.
Объект изобретения состоит из химических молекулярных частиц, которые опосредуют разрушение мРНК LPA путем связывания с транскриптами гена LPA посредством механизмов клеточной РНК-интерференции. Молекулярные соединения согласно настоящему изобретению можно применять в качестве конъюгатов с сахарной группой N-ацетилгалактозамина (GalNAc), но не ограничиваясь им, который обеспечивает специфичное для гепатоцитов клеточное поглощение за счет специфичного связывания с комплексом рецептора асиалогликопротеина (ASGPR). Объект изобретения может быть соединен с различными другими химическими структурами, придающими разные свойства, как указано ниже. Применение профиля химической модификации нуклеиновых кислот придает устойчивость к нуклеазам в сыворотке и делает возможным, например, путь подкожного применения.
Объект изобретения характеризуется высокой специфичностью направленной доставки на молекулярном уровне и тканевом уровне, что потенциально обеспечивает лучший профиль безопасности, чем доступные в настоящее время способы лечения.
Согласно настоящему изобретению также предложена нуклеиновая кислота в соответствии с любым аспектом настоящего изобретения, описанным в настоящей заявке, в которой первая цепь РНК имеет концевой 5'-(Е)-винилфосфонатный нуклеотид, и концевой 5'-(Е)-винилфосфонатный нуклеотид соединен со вторым нуклеотидом в первой цепи с помощью фосфодиэфирной связи.
Согласно одному варианту реализации настоящего изобретения первая цепь может содержать более 1 фосфодиэфирной связи.
Согласно одному варианту реализации настоящего изобретения первая цепь может содержать фосфодиэфирные связи между по меньшей мере тремя концевыми 5'-нуклеотидами.
Согласно одному варианту реализации настоящего изобретения первая цепь может содержать фосфодиэфирные связи между по меньшей мере четырьмя концевыми 5'-нуклеотидами.
Согласно одному варианту реализации настоящего изобретения первая цепь может содержать формулу (XVII):
(vp)-N(po)[N(po)]n-
(XVII)
в которой «(vp)-» представляет собой 5'-(E)-винилфосфонат, «N» представляет собой нуклеотид, «po» представляет собой фосфодиэфирную связь, а n составляет от 1 до (общее количество нуклеотидов в первой цепи -2), при этом предпочтительно n составляет от 1 до (общее количество нуклеотидов в первой цепи -3), при этом более предпочтительно n составляет от 1 до (общее количество нуклеотидов в первой цепи -4).
Согласно одному варианту реализации настоящего изобретения первая цепь может содержать по меньшей мере одну фосфотиоатную (ps) связь.
Согласно одному варианту реализации настоящего изобретения первая цепь может дополнительно содержать фосфотиоатную связь между двумя концевыми 3'-нуклеотидами или фосфотиоатные связи между тремя концевыми 3'-нуклеотидами.
Согласно одному варианту реализации настоящего изобретения связи между другими нуклеотидами в первой цепи представляют собой фосфодиэфирные связи.
Согласно одному варианту реализации настоящего изобретения первая цепь может содержать более 1 фосфотиоатной связи.
Согласно дополнительному варианту реализации вторая цепь может содержать фосфотиоатную связь между двумя концевыми 3'-нуклеотидами или фосфотиоатные связи между тремя концевыми 3'-нуклеотидами.
Согласно другому дополнительному варианту реализации вторая цепь может содержать фосфотиоатную связь между двумя концевыми 5'-нуклеотидами или фосфотиоатные связи между тремя концевыми 5'-нуклеотидами.
Согласно варианту реализации настоящего изобретения концевой 5'-(E)-винилфосфонатный нуклеотид представляет собой РНК-нуклеотид.
Концевой 5'-(E)-винилфосфонатный нуклеотид представляет собой нуклеотид, в котором природная фосфатная группа на 5'-конце замещена E-винилфосфонатом, в котором мостиковый 5'-атом кислорода концевого нуклеотида 5'-фосфорилированной цепи замещен метинильной (-CH=) группой:
Нуклеотиды с природным фосфатом на 5'-конце

Нуклеотид с E-винилфосфонатом на 5'-конце
5'-(E)-винилфосфонат представляет собой имитатор 5'-фосфата. Биологический имитатор представляет собой молекулу, которая способна выполнять ту же функцию, как и исходная молекула, которую она имитирует, и структурно очень сходна с ней. Применительно к настоящему изобретению 5'-(E)-винилфосфонат имитирует функцию нормального 5'-фосфата, например, обеспечивая эффективную загрузку RISC. Кроме того, благодаря своей незначительно измененной структуре 5'-(E)-винилфосфонат способен стабилизировать 5'-концевой нуклеотид, защищая его от дефосфорилирования ферментами, такими как фосфатазы.
Один аспект настоящего изобретения представляет собой нуклеиновую кислоту, раскрытую в настоящей заявке, для ингибирования экспрессии целевого гена в клетке, содержащую по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с указанного гена-мишени, причем указанная первая цепь содержит модифицированные нуклеотиды или немодифицированные нуклеотиды во множестве положений для облегчения процессинга нуклеиновой кислоты под действием RISC.
Согласно одному аспекту «облегчает процессинг под действием RISC» означает, что нуклеиновая кислота может подвергаться процессингу под действием RISC, например, любая присутствующая модификация делает возможным процессинг нуклеиновой кислоты под действием RISC приемлемо таким образом, чтобы могла реализовываться активность миРНК.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, а нуклеотид на второй цепи, который соответствует положению 13 первой цепи, не модифицирован с применением 2'-O-метильной модификации.
Нуклеотид на второй цепи, который «соответствует» положению на первой цепи, приемлемо, является нуклеотидом, который спаривается с указанным нуклеотидом на первой цепи.
Согласно одному аспекту нуклеотид на второй цепи, который соответствует положению 13 первой цепи, представляет собой нуклеотид, который образует пару оснований с положением 13 первой цепи.
Согласно одному аспекту нуклеотид на второй цепи, который соответствует положению 11 первой цепи, представляет собой нуклеотид, который образует пару оснований с положением 11 первой цепи.
Согласно одному аспекту нуклеотид на второй цепи, который соответствует положению 12 первой цепи, представляет собой нуклеотид, который образует пару оснований с положением 12 первой цепи.
Эту номенклатуру можно применять к другим положениям второй цепи. Например, в 19-членной нуклеиновой кислоте, которая является двухцепочечной и имеет тупые концы, положение 13 первой цепи будет спариваться с положением 7 второй цепи. Положение 11 первой цепи будет спариваться с положением 9 второй цепи. Эту номенклатуру можно применять к другим положениям второй цепи.
Нуклеотид, который соответствует положению 13 первой цепи, приемлемо представляет собой положение 13 второй цепи, при отсчете от 3'-второй цепи, начиная с первого нуклеотида двухцепочечной области. Аналогичным образом, положение 11 второй цепи приемлемо представляет собой 11 нуклеотид с 3'-второй цепи, начиная с первого нуклеотида двухцепочечной области. Эту номенклатуру можно применять к другим положениям второй цепи.
Согласно одному аспекту, в случае частичной комплементарности первой и второй цепей, нуклеотид на второй цепи, который «соответствует» положению на первой цепи, необязательно может образовывать пару оснований, если это положение представляет собой положение, в котором присутствует несоответствие, однако принцип номенклатуры все еще применим.
Предпочтительной является первая и вторая цепи, которые полностью комплементарны на протяжении дуплексной области (не учитывая любые выступающие области), и в двухцепочечной области нуклеиновой кислоты отсутствуют несоответствия.
Также предпочтительными являются:
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, а нуклеотид на второй цепи, который соответствует положению 11 первой цепи, не модифицирован с применением 2'-O-метильной модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, а нуклеотиды на второй цепи, которые соответствуют положениям 11 и 13 первой цепи, не модифицированы с применением 2'-O-метильной модификации.
Согласно одному аспекту нуклеотид на второй цепи, который соответствует положению 12 первой цепи, не модифицирован с применением 2'-O-метильной модификации. Это ограничение на нуклеиновой кислоте может наблюдаться с любым другим ограничением, описанным в настоящей заявке.
Таким образом, другим аспектом настоящего изобретения является нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, а нуклеотиды на второй цепи, которые соответствуют положению 11-13 первой цепи, не модифицированы с применением 2'-O-метильной модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, а нуклеотиды на второй цепи, которые соответствуют положению 11 или 13, или 11 и 13, или 11-13 первой цепи, модифицированы с применением 2'-фтор-модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи модифицированы с применением 2'-фтор-модификации, а нуклеотиды на второй цепи, которые соответствуют положениям 11 или 13, или 11 и 13, или 11-13 первой цепи, не модифицирован с применением 2'-O-метильной модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи модифицированы с применением 2'-фтор-модификации, а нуклеотиды на второй цепи, которые соответствуют положениям 11 или 13, или 11 и 13, или 11-13 первой цепи, модифицированы с применением 2'-фтор-модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой более 50% нуклеотидов первой и/или второй цепей содержат 2'-O-метильную модификацию, например, более 55%, 60%, 65%, 70%, 75%, 80% или 85% или более первой и/или второй цепей содержат 2'-O-метильную модификацию, предпочтительно как измерено в процентах от общего количества нуклеотидов в обеих первой и второй цепях.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой более 50% нуклеотидов первой и/или второй цепей содержат природную модификацию РНК, например, в которой более 55%, 60%, 65%, 70%, 75%, 80% или 85% или более первой и/или второй цепей содержат такую модификацию, предпочтительно как измерено в процентах от общего количества нуклеотидов обеих первой и второй цепей. Подходящие природные модификации включают, наряду с 2-O'-метилом, другие 2'-модификации сахара, в частности, 2'-H-модификацию, приводящую к получению ДНК-нуклеотида.
Нуклеиновая кислота, раскрытая в настоящей заявке, содержащая не более 20%, например, не более 15%, например, более 10%, нуклеотидов, которые имеют 2'-модификации, которые не являются 2'-O-метильными модификациями, на первой и/или второй цепях, предпочтительно как измерено в процентах от общего количества нуклеотидов обеих первой и второй цепей.
Нуклеиновая кислота, раскрытая в настоящей заявке, содержащая не более 20% (например, не более 15% или не более 10%) 2'-фтор-модификаций на первой и/или второй цепях, предпочтительно как измерено в процентах от общего количества нуклеотидов обеих цепей.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой все нуклеотиды модифицированы с применением 2'-O-метильной модификации, за исключением положений 2 и 14 с 5'-конца первой цепи и нуклеотидов на второй цепи, которые соответствуют положению 11 или 13, или 11 и 13, или 11-13 первой цепи. Предпочтительно нуклеотиды, которые не модифицированы с применением 2'-O-метила, модифицированы с применением фтора в положении 2'.
Предпочтительной является нуклеиновая кислота, раскрытая в настоящей заявке, в которой все нуклеотиды нуклеиновой кислоты модифицированы в 2'-положении сахара. Предпочтительно эти нуклеотиды модифицированы с применением 2'-фтор-модификации, причем указанная модификация не является 2'-O-метильной модификацией.
Нуклеиновые кислоты согласно настоящему изобретению могут содержать один или более нуклеотидов, модифицированных в 2'-положении с применением 2'-H, и, следовательно, содержащие ДНК-нуклеотид внутри нуклеиновой кислоты. Нуклеиновые кислоты согласно настоящему изобретению могут содержать ДНК-нуклеотиды в положениях 2 и/или 14 первой цепи, при отсчете от 5'-конца первой цепи. Нуклеиновые кислоты могут содержать ДНК-нуклеотиды на второй цепи, которые соответствуют положению 11 или 13, или 11 и 13, или 11-13 первой цепи.
Согласно одному аспекту присутствует не более одной ДНК на нуклеиновую кислоту согласно настоящему изобретению.
Нуклеиновые кислоты согласно настоящему изобретению могут содержать один или более LNA-нуклеотидов. Нуклеиновые кислоты согласно настоящему изобретению могут содержать LNA-нуклеотиды в положениях 2 и/или 14 первой цепи, при отсчете от 5'-конца первой цепи. Нуклеиновые кислоты могут содержать LNA на второй цепи, которые соответствуют положению 11 или 13, или 11 и 13, или 11-13 первой цепи.
Согласно одному аспекту нуклеиновая кислота модифицирована на первой цепи с применением чередующихся 2-O-метильных модификаций и 2-фтор-модификаций, и положения 2 и 14 (начиная с 5'-конца) модифицированы с применением 2'-фтор. Предпочтительно вторая цепь модифицирована с применением 2'-фтор-модификаций в нуклеотидах на второй цепи, которые соответствуют положению 11 или 13, или 11 и 13, или 11-13 первой цепи. Предпочтительно вторая цепь модифицирована с применением 2'-фтор-модификаций в положениях 11-13, при отсчете от 3'-конца, начиная с первого положения комплементарной (двухцепочечной) области, а остальные модификации представляют собой природные модификации, предпочтительно 2'-O-метил.
Согласно одному аспекту нуклеиновая кислота согласно настоящему изобретению содержит один или более инвертированных рибонуклеотидов, предпочтительно инвертированный аденин, с применением 5'-5'-связи или 3'-3'-связи, предпочтительно 3'-3'-связи на 3'-конце второй цепи.
Согласно одному аспекту нуклеиновая кислота содержит одну или более фосфодитиоатных связей, таких как 1, 2, 3 или 4 фосфодитиоатные связи. Предпочтительно присутствует до 4 фосфодитиоатных связей, по одной на 5' и 3'-концах первой и второй цепей.
Все признаки нуклеиновых кислот могут быть объединены со всеми другими аспектами настоящего изобретения, раскрытыми в настоящей заявке.
В частности, предпочтительными являются нуклеиновые кислоты, которые представляют собой молекулы миРНК, в которых нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, и нуклеиновая кислота содержит одно или более или все из:
(i) инвертированного нуклеотида, предпочтительно 3'-3'-связь на 3'-конце второй цепи;
(ii) одной или более фосфодитиоатных связей;
(iii) нуклеотид второй цепи, соответствующий положению 11 или 13 первой цепи, не модифицирован с применением 2'-O-метильной модификации, при этом предпочтительно одно или оба из этих положений содержат 2'-фтор-модификацию;
(iv) нуклеиновая кислота содержит по меньшей мере 80% от всех нуклеотидов, содержащих 2'-O-метильную модификацию;
(v) нуклеиновая кислота содержит не более 20% нуклеотидов, которые содержат 2'-фтор-модификации.
Согласно настоящему изобретению также предложена нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи и нуклеотиды в положениях 7 и/или 9 или 7-9 с 5'-конца второй цепи модифицированы с применением 2'-фтор-модификации, и по меньшей мере 90% оставшихся нуклеотидов являются 2'-O-метил-модифицированными или содержат другую природную 2'-модификацию.
Конкретными предпочтительными примерами для 19-членной двухцепочечной нуклеиновой кислоты с тупыми концами и без липких концов, являются:
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, и нуклеотид в положении 7 с 5'-конца второй цепи не модифицирован с применением 2'-O-метильной модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, и нуклеотид в положении 9 с 5'-конца второй цепи не модифицирован с применением 2'-O-метильной модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, и нуклеотиды в положениях 7 и 9 с 5'-конца второй цепи не модифицированы с применением 2'-O-метильной модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, и нуклеотиды в положениях 7-9 с 5'-конца второй цепи не модифицированы с применением 2'-O-метильной модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи не модифицированы с применением 2'-O-метильной модификации, и нуклеотиды в положениях 7 и/или 9, или 7-9 с 5'-конца второй цепи модифицированы с применением 2'-фтор-модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи модифицированы с применением 2'-фтор-модификации, и нуклеотиды в положениях 7 и/или 9, или 7-9 с 5'-конца второй цепи не модифицированы с применением 2'-O-метильной модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой нуклеотиды в положениях 2 и 14 с 5'-конца первой цепи модифицированы с применением 2'-фтор-модификации, и нуклеотиды в положениях 7 и/или 9, или 7-9 с 5'-конца второй цепи модифицированы с применением 2'-фтор-модификации.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой более 50% нуклеотидов первой и/или второй цепей содержат 2'-O-метильную модификацию, например, более 55%, 60%, 65%, 70%, 75%, 80% или 85% или более первой и/или второй цепей содержат 2'-O-метильную модификацию, предпочтительно как измерено в процентах от общего количества нуклеотидов обеих первой и второй цепей.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой более 50% нуклеотидов первой и/или второй цепей содержат природную модификацию РНК, например, более 55%, 60%, 65%, 70%, 75%, 80% или 85% или более первой и/или второй цепей содержат такую модификацию, предпочтительно как измерено в процентах от общего количества нуклеотидов обеих первой и второй цепей. Подходящие природные модификации включают, наряду с 2-O'-метилом, другие 2'-модификации сахара, в частности, 2'-H-модификацию, приводящую к получению ДНК-нуклеотида.
Нуклеиновая кислота, раскрытая в настоящей заявке, содержащая не более 20%, например, не более 15%, например, более 10%, нуклеотидов, которые содержат 2'-модификации, которые не являются 2'-O-метильными модификациями, на первой и/или второй цепях, предпочтительно как измерено в процентах от общего количества нуклеотидов обеих первой и второй цепей.
Нуклеиновая кислота, раскрытая в настоящей заявке, содержащая не более 20% (например, не более 15% или не более 10%) 2'-фтор-модификаций на первой и/или второй цепях, предпочтительно как измерено в процентах от общего количества нуклеотидов обеих цепей.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой все нуклеотиды модифицированы с применением 2'-O-метильной модификации, за исключением положений 2 и 14 с 5'-конца первой цепи и нуклеотидов в положениях 7 и/или 9 с 5'-конца второй цепи. Предпочтительно нуклеотиды, которые не модифицированы с применением 2'-O-метила, модифицированы с применением фтора в положении 2'.
Нуклеиновая кислота, раскрытая в настоящей заявке, в которой все нуклеотиды модифицированы с применением 2'-O-метильной модификации, за исключением положений 2 и 14 с 5'-конца первой цепи и нуклеотидов в положениях 7-9 с 5'-конца второй цепи. Предпочтительно нуклеотиды, которые не модифицированы с применением 2'-O-метила, модифицированы с применением фтора в положении 2'.
Для нуклеиновой кислоты, содержащей дуплексную область из 20 пар оснований, вторая цепь предпочтительно не содержит 2'-O-метильную группу в нуклеотидах 8, 9 или 10, при отсчете от 5'-конца дуплекса, соответствующих положениям 13, 12 и 11 первой цепи, соответственно.
Для нуклеиновой кислоты, содержащей дуплексную область из 21 пары оснований, вторая цепь предпочтительно не содержит 2'-O-метильную группу в нуклеотидах 9 или 10, или 11, при отсчете от 5'-конца дуплекса, соответствующих положениям 13, 12 и 11 первой цепи, соответственно.
Настоящее изобретение также относится к немодифицированным последовательностям всех модифицированных последовательностей, раскрытых в настоящей заявке.
Настоящее изобретение будет далее описано со ссылкой на следующие неограничивающие чертежи и примеры.
Чертежи
На Фигуре 1 показаны результаты скрининга неконъюгированной молекулы миРНК в отношении ингибирования экспрессии мРНК LPA в клетках RT-4 человека.
На Фигурах 2А и 2В показан дозозависимый ответ неконъюгированных нацеленных на LPA молекул миРНК на экспрессию мРНК LPA в клетках RT-4 человека.
На Фигуре 3 показано ингибирование экспрессии мРНК LPA в первичных гепатоцитах человека и яванских макак различными дозами GalNAc-L1 LPA-1038-конъюгированных молекул миРНК, доставленных путем опосредуемого рецепторами поглощения.
На Фигурах 4А и 4В показаны примеры структуры лигандов GalNAc с различными линкерами L1 и L6, соответственно, с которыми были конъюгированы олигонуклеотиды, представленные в качестве примера.
На Фигуре 5 показаны типичные примеры выключения мРНК LPA с помощью указанных L6-конъюгированных GalNAc миРНК в первичных гепатоцитах человека, доставленных путем опосредуемого рецепторами поглощения.
На Фигуре 6 изображен конъюгат 1.
На Фигуре 7 изображен конъюгат 2.
На Фигуре 8 изображен конъюгат 3.
На Фигуре 9 изображен эталонный конъюгат 1.
На Фигуре 10 изображен эталонный конъюгат 2.
На Фигуре 11 изображен эталонный конъюгат 3.
На Фигуре 12 изображен эталонный конъюгат 4.
На каждой из Фигур 6-12 и 19-30 верхняя цепь представляет собой антисмысловую цепь, а нижняя цепь представляет собой смысловую цепь. Кроме того, чтобы более ясно показать связь между нуклеиновой кислотой и частями лиганда, нуклеотид на конце соответствующих конъюгированных цепей нарисован полностью.
На Фигуре 13 показан синтез A0268, который представляет собой одноцепочечный олигонуклеотид, конъюгированный на 3'-конце с моно-GalNAc, и является исходным материалом при синтезе конъюгата 1 и конъюгата 3. (ps) обозначает фосфотиоатную связь.
На Фигуре 14 показан синтез A0006, который представляет собой одноцепочечный олигонуклеотид, конъюгированный на 5'-конце с трехантенным GalNAc, применяемый для синтеза эталонного конъюгата 4. (ps) обозначает фосфотиоатную связь.
На Фигуре 15 проиллюстрировано определение выключения TTR в условиях in vitro. В частности, на Фигуре 15А показано определение выключения TTR в условиях in vitro с помощью эталонных конъюгатов (RC) 1 и 3, а также необработанного контроля «UT»; на Фигуре 15B показано определение выключения TTR в условиях in vitro с помощью эталонных конъюгатов (RC) 2 и 3, а также необработанного контроля «UT»; и на Фигуре 15C показано определение выключения TTR в условиях in vitro с помощью конъюгатов 1, 2 и 3, а также с помощью RC3 и необработанного контроля «UT». Эталонные конъюгаты 1 и 2 представляют собой конъюгаты для сравнения. Эталонный конъюгат 3 представляет собой ненацеливающую GalNAc миРНК, а «необработанный» («UT») представляет необработанные клетки. Оба RC3 и UT являются отрицательными контролями. Уровни мРНК были нормированы против PtenII.
На Фигуре 16 показана временная динамика сывороточного TTR в когортах мышей c57BL/6 с n=4 через 7, 14 и 27 дней после обработки путем подкожного введения 1 мг/кг - конъюгаты 1-3, эталонные конъюгаты (RC) 1, 2 и 4 и индивидуумы с имитированной обработкой (ФСБ).
На Фигуре 17 показан синтез олигонуклеотидов из предшественников олигонуклеотидов, конъюгированных с GalNAc на 3' и 5'-концах (таких как соединение X0385B-prec).
На Фигуре 18 показан равный дозозависимый ответ выключения для миРНК, нацеленной на LPA, с двумя отдельными блоками GalNAc, конъюгированными со второй цепью, по сравнению с трехантенным блоком GalNAc на 5' второй цепи, в первичных гепатоцитах яванской макаки.
На Фигуре 19А изображен конъюгат 4. Последние три нуклеотида на 5' и 3'-концах антисмысловой и смысловой цепей соединены фосфотиоатным линкером между каждым нуклеотидом. Серинол-GalNAc-линкеры конъюгированы за счет фосфодиэфирной связи с 3'-концом и 5'-концом смысловой цепи.
На Фигуре 19В изображен конъюгат 5. Последние три нуклеотида на 5' и 3'-концах антисмысловой цепи соединены фосфотиоатным линкером между каждым нуклеотидом. Серинол-GalNAc-линкеры конъюгированы за счет фосфотиоатной связи с 3'-концом и 5'-концом смысловой цепи.
На Фигуре 20 изображен конъюгат 6. Последние три нуклеотида на 5' и 3'-концах антисмысловой цепи соединены фосфотиоатным линкером между каждым нуклеотидом. Серинол-GalNAc-линкеры конъюгированы за счет фосфодиэфирной связи с 3'-концом и 5'-концом смысловой цепи.
На Фигуре 21 изображен конъюгат 7. Последние три нуклеотида на 5' и 3'-концах антисмысловой и смысловой цепей соединены фосфотиоатным линкером между каждым нуклеотидом. Серинол-GalNAc-линкеры конъюгированы за счет фосфотиоатной связи с 3'-концом и 5'-концом смысловой цепи. Серинол-GalNAc-линкеры соединены друг с другом за счет фосфотиоатной связи.
На Фигуре 22 изображен конъюгат 8. Последние три нуклеотида на 5' и 3'-концах антисмысловой и смысловой цепей соединены фосфотиоатным линкером между каждым нуклеотидом. Линкер GalNAc-C6-амино-модификатор конъюгирован на 5'-конце смысловой цепи, и линкер GalNAc-C7-амино-модификатор конъюгирован на 3'-конце смысловой цепи.
На Фигуре 23 изображен конъюгат 9. Последние три нуклеотида на 5' и 3'-концах антисмысловой и смысловой цепей соединены фосфотиоатным линкером между каждым нуклеотидом. Линкер GalNAc-GlyC3-амино-модификатор конъюгирован на 5' и 3'-концах смысловой цепи.
На Фигуре 24 изображен конъюгат 10. Последние три нуклеотида на 5' и 3'-концах антисмысловой и смысловой цепей соединены фосфотиоатным линкером между каждым нуклеотидом, и смысловые цепи соединены фосфотиоатным линкером между каждым нуклеотидом. Линкер GalNAc-пиперидил-амино-модификатор конъюгирован на 5' и 3'-концах смысловой цепи.
На Фигуре 25 изображен конъюгат 11. Последние три нуклеотида на 5' и 3'-концах антисмысловой и смысловой цепей соединены фосфотиоатным линкером между каждым нуклеотидом. Линкер GalNAc-C3-амино-модификатор конъюгирован на 5' и 3'-концах смысловой цепи.
На Фигуре 26 изображен конъюгат 12. Последние три нуклеотида на 5' и 3'-концах антисмысловой и смысловой цепей соединены фосфотиоатным линкером между каждым нуклеотидом. Линкер GalNAc-C6-амино-модификатор конъюгирован на 5'-конце смысловой цепи, и линкер GalNAc-GlyC3-амино-модификатор конъюгирован на 3'-конце смысловой цепи.
На Фигуре 27 изображены конъюгаты 15, 16, 18 и 19, которые отличаются только своими последовательностями РНК. Последние три нуклеотида на 5' и 3'-концах антисмысловой и смысловой цепей соединены фосфотиоатным линкером между каждым нуклеотидом в каждом конъюгате. Серинол-GalNAc-линкеры конъюгированы за счет фосфотиоатной связи с 3'-концом и 5'-концом смысловой цепи.
На Фигуре 28 изображен эталонный конъюгат 5 и эталонный конъюгат 9, которые отличаются только своими последовательностями РНК. Последние три нуклеотида на 5' и 3'-концах антисмысловой цепи и 3'-конце смысловой цепи соединены фосфотиоатным линкером между каждым нуклеотидом в обоих конъюгатах. Тримерный GalNAc-линкер конъюгирован за счет фосфотиоатной связи с 5'-концом смысловой цепи в обоих конъюгатах.
На Фигуре 29 изображен эталонный конъюгат 6 и эталонный конъюгат 7, которые отличаются только своими последовательностями РНК. Последние три нуклеотида на 5' и 3'-концах антисмысловой цепи и 3'-конце смысловой цепи соединены фосфотиоатным линкером между каждым нуклеотидом в обоих конъюгатах. Тримерный GalNAc-линкер конъюгирован за счет фосфотиоатной связи с 5'-концом смысловой цепи в обоих конъюгатах.
На Фигуре 30 изображен эталонный конъюгат 8. Последние три нуклеотида на 5' и 3'-концах антисмысловой цепи и 3'-конце смысловой цепи соединены фосфотиоатным линкером между каждым нуклеотидом. Тримерный GalNAc-линкер конъюгирован за счет фосфотиоатной связи с 5'-концом смысловой цепи.
На Фигуре 31 проиллюстрировано определение выключения TTR в условиях in vitro. В частности, на Фигуре 31A показано определение выключения TTR в условиях in vitro с помощью конъюгатов 4, 5, 6 и 2 по сравнению с «Luc» (эталонный конъюгат 3), а также необработанным контролем «UT»; на Фигуре 31B показано определение выключения TTR в условиях in vitro с помощью конъюгатов 7 и 2 по сравнению с «Luc» (эталонный конъюгат 3), а также необработанным контролем «UT». Luc или эталонный конъюгат 3 (RC3) представляет собой ненацеливающую GalNAc миРНК, а «необработанный» («UT») представляет необработанные клетки. Оба RC3 и UT являются отрицательными контролями. Уровень мРНК был нормирован против PtenII.
На Фигуре 32 проиллюстрировано определение выключения TTR в условиях in vitro. В частности, на Фигуре 32A показано определение выключения TTR в условиях in vitro с помощью конъюгатов 8, 9, 10, 11 и 2 по сравнению с «Luc» (эталонный конъюгат 3), а также необработанным контролем «UT»; на Фигуре 32B показано определение выключения TTR в условиях in vitro с помощью конъюгатов 12 и 2 по сравнению с «Luc» (эталонный конъюгат 3), а также необработанным контролем «UT». Luc или эталонный конъюгат 3 представляет ненацеливающую GalNAc миРНК, а «необработанный» («UT») представляет необработанные клетки. Оба RC3 и UT являются отрицательными контролями. Уровень мРНК был нормирован против PtenII.
На Фигуре 33 проиллюстрировано определение выключения мРНК LPA в условиях in vitro с помощью конъюгата 19 по сравнению с контролями. Ctr представляет ненацеливающую GalNAc миРНК, а «необработанный» («UT») представляет необработанные клетки. Оба Ctr и UT являются отрицательными контролями. Уровень мРНК был нормирован против ACTB.
На Фигуре 34 показана временная динамика уровней мРНК Aldh2 печени в когортах мышей c57BL/6 с n=6 через 14, 28 и 42 дня после обработки путем подкожного введения 1 мг/кг - конъюгат 15, эталонный конъюгат (RC) 6 и индивидуумы с имитированной обработкой (ФСБ). Уровень мРНК нормировали против Pten.
На Фигуре 35 показана временная динамика уровней мРНК Aldh2 печени в когортах мышей c57BL/6 с n=6 через 14, 28 и 42 дня после обработки путем подкожного введения 1 мг/кг - конъюгат 16, эталонный конъюгат (RC) 7 и индивидуумы с имитированной обработкой (ФСБ). Уровень мРНК нормировали против Pten.
На Фигуре 36 показана временная динамика уровней мРНК Tmprss6 печени в когортах мышей c57BL/6 с n=6 через 14, 28 и 42 дня после обработки путем подкожного введения 1 мг/кг - конъюгат 18, эталонный конъюгат (RC) 8 и индивидуумы с имитированной обработкой (ФСБ). Уровень мРНК нормировали против Pten.
На Фигуре 37 показана стабильность в сыворотке конъюгатов 4, 5, 6, 7 и 2, а также необработанного контроля (UT) при 37°C в течение 3 дней.
На Фигуре 38 показана стабильность в сыворотке конъюгатов 8, 9, 10, 11, 12 и 2, а также необработанного контроля (UT) при 37°C в течение 3 дней.
На Фигуре 39 показано ингибирование экспрессии мРНК LPA в первичных гепатоцитах человека при разных дозах GalNAc-L6-связанных миРНК, доставленных путем опосредуемого рецепторами поглощения.
На Фигуре 40 схематически представлены различные варианты реализации нуклеиновых кислот, конъюгированных с лигандами за счет линкеров.
Примеры
Нумерация, упомянутая в каждом примере, является конкретной для упомянутого примера.
Пример 1
Модифицированные и конъюгированные молекулы миРНК, применяемые для функциональных примеров в настоящей заявке.
Производные LPA-1038:
GalNAc-LPA-1038-L1
Первая цепь (SEQ ID NO: 119, на основании SEQ ID NO: 5)
OMeA-(ps)-FU-(ps)-OMeA-FA-OMeC-FU-OMeC-FU-OMeG-FU-OMeC-FC-OMeA-FU-OMeU-FA-OMeC-(ps)-FC-(ps)-OMeA 3'
Вторая цепь (SEQ ID NO: 120, на основании SEQ ID NO SEQ ID NO: 6)
5'[ST23 (ps)]3 длинный утроитель (ps)FU-OMeG-FG-OMeU-FA-OMeA-FU-OMeG-FG-OMeA-FC-OMeA-FG-OMeA-FG-OMeU-FU-(ps)-OMeA-(ps)-FU 3'
GalNAc-LPA-1038-L6
Первая цепь (SEQ ID NO: 121, на основании SEQ ID NO: 5)
OMeA-(ps)-FU-(ps)-OMeA-FA-OMeC-FU-OMeC-FU-OMeG-FU-OMeC-FC-OMeA-FU-OMeU-FA-OMeC-(ps)-FC-(ps)-OMeA 3'
Вторая цепь (SEQ ID NO: 122, на основании SEQ ID NO: 6)
5'[ST23 (ps)]3 ST43 (ps)FU-OMeG-FG-OMeU-FA-OMeA-FU-OMeG-FG-OMeA-FC-OMeA-FG-OMeA-FG-OMeU-FU-(ps)-OMeA-(ps)-FU 3'
FN (N=A, C, G, U) обозначает 2'-фтор, 2'-дезоксинуклеозиды
OMeN (N=A, C, G, U) обозначает 2'-O-метилнуклеозиды
(ps) обозначает фосфотиоатную связь
ST23 и ST43 представлены ниже.
Дополнительным примером являются производные LPA 1041:
GalNAc-LPA-1041-L1
Первая цепь (SEQ ID NO: 123, на основании SEQ ID NO: 9)
5' OMeA-(ps)-FU-(ps)-OMeA-FA-OMeC-FU-OMeC-FU-OMeG-FU-OMeC-FC-OMeA-FU-OMeU-FA-OMeC-(ps)-FC-(ps)-OMeG 3'
Вторая цепь (SEQ ID NO: 124, на основании SEQ ID NO: 10)
5'[ST23 (ps)]3 длинный утроитель (ps) FC-OMeG-FG-OMeU-FA-OMeA-FU-OMeG-FG-OMeA-FC-OMeA-FG-OMeA-FG-OMeU-FU-(ps)-OMeA-(ps)-FU 3'
GalNAc-LPA-1041-L6
Первая цепь (SEQ ID NO: 125, на основании SEQ ID NO: 9)
5' OMeA-(ps)-FU-(ps)-OMeA-FA-OMeC-FU-OMeC-FU-OMeG-FU-OMeC-FC-OMeA-FU-OMeU-FA-OMeC-(ps)-FC-(ps)-OMeG 3'
Вторая цепь (SEQ ID NO: 126, на основании SEQ ID NO: 10)
5'[ST23 (ps)]3 ST43 (ps) FC-OMeG-FG-OMeU-FA-OMeA-FU-OMeG-FG-OMeA-FC-OMeA-FG-OMeA-FG-OMeU-FU-(ps)-OMeA-(ps)-FU 3'
FN (N=A, C, G, U) обозначает 2'-фтор, 2'-дезоксинуклеозиды
OMeN (N=A, C, G, U) обозначает 2'-O-метилнуклеозиды
(ps) обозначает фосфотиоатную связь
ST23 и ST43 представлены ниже.
ST23 представляет собой GalNAc-C4-фосфоамидит (структурные компоненты представлены ниже)
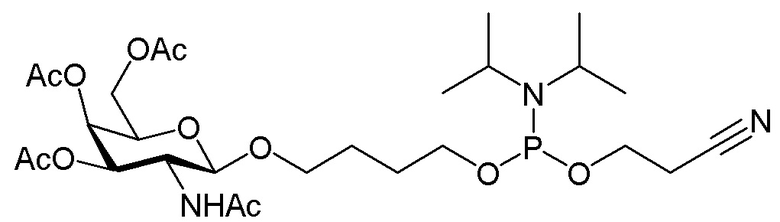
ST23
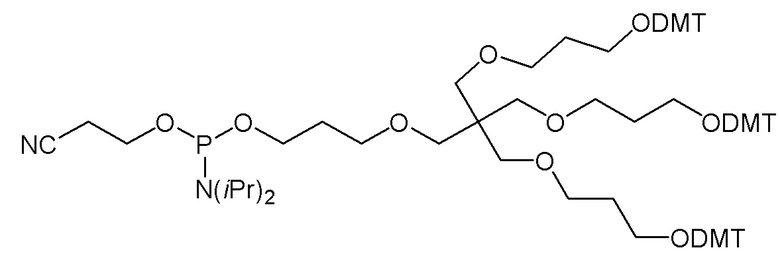
Длинный утроитель (STKS)
ST41 представлен ниже:
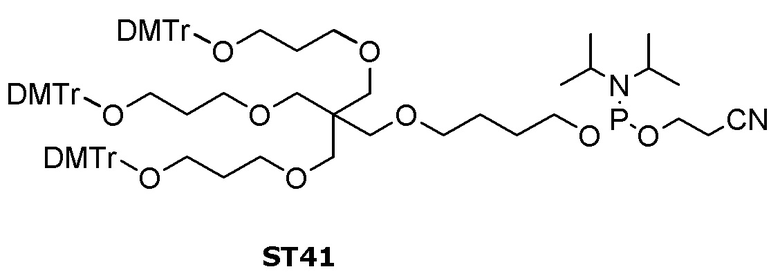
ST43 представлен ниже и описан в WO2017/174657:
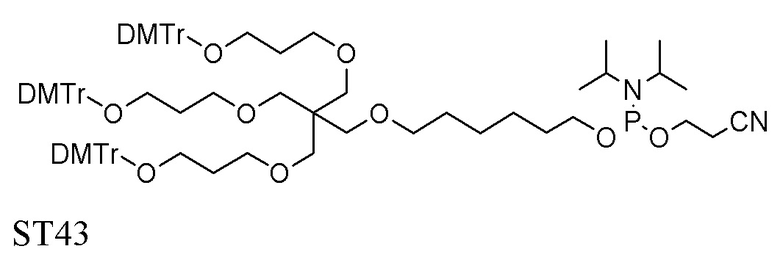
Все олигонуклеотиды либо получали от коммерческих производителей олигонуклеотидов (Biospring, Франкфурт, Германия, или RiboBio, Гуанчжоу, Гуандун, КНР), либо синтезировали на синтезаторе AKTA oligopilot (самостоятельно) с применением стандартных фосфоамидитных реакций. Применяли коммерчески доступную твердую подложку и 2'-O-метил-РНК-фосфоамидиты, 2'-фтор-ДНК-фосфоамидиты (все со стандартной защитой) и коммерчески доступный длинный утроительный фосфоамидит (Glen research). Синтез выполняли с применением 0,1 М растворов фосфоамидита в сухом ацетонитриле, а в качестве активатора применяли бензилтиотетразол (БТТ) (0,3 М в ацетонитриле). Все остальные реагенты представляли собой коммерчески доступные стандартные реагенты.
Конъюгацию соответствующего синтона GalNac (например, ST23, ST41 или ST43) осуществляли путем связывания соответствующего фосфоамидита с 5'-концом олигоцепи в стандартных условиях связывания фосфоамидита. Фосфотиоаты вводили с применением стандартных коммерчески доступных реагентов для тиолирования (EDITH, Link technologies).
Одиночные цепи отщепляли от CPG с применением метиламина (40% водн.) и полученный неочищенный олигонуклеотид очищали с помощью ионообменной хроматографии (Resource Q, 6 мл, GE Healthcare) на системе для ВЭЖХ AKTA Pure с применением градиента хлорида натрия. Содержащие продукт фракции объединяли, обессоливали на колонке для гель-проникающей хроматографии (Zetadex, EMP Biotech) и лиофилизировали.
Для ренатурации эквимолярные количества соответствующих одиночных цепей растворяли в воде и нагревали до 80°C в течение 5 минут. После постепенного охлаждения до комнатной температуры полученный дуплекс лиофилизировали.
Последовательности полученных нуклеиновых кислот (миРНК) приведены в таблице 1 ниже.
Таблица 1: последовательности неконъюгированных нуклеиновых кислот, испытанные в отношении ингибирования экспрессии мРНК LPA. Указаны последовательности и применяемый профиль модификации
Таблица 1
Таблица 1: модификации нуклеотидов обозначены следующими номерами (колонка 4), 1=2'-F-dU, 2=2'-F-dA, 3=2'-F-dC, 4=2'-F-dG, 5=2'-OMe-rU; 6=2'-OMe-rA; 7=2'-OMe-rC; 8=2'-OMe-rG.
Таблица 2: последовательности наборов ампликонов кПЦР LPA, APOB, бета-актина и Pten, которые применяли для измерения уровней мРНК, показаны ниже.
Таблица 2
Пример 2
Скрининг неконъюгированных молекул миРНК (таблица 1) в отношении ингибирования экспрессии мРНК LPA в клетках RT-4 человека.
Липосомальные трансфекционные комплексы готовили с тремя повторами в соотношении 1,5 мкл РНКиMax (ThermoFisher)/80 пмоль указанных молекул миРНК. Комплекс разбавляли до указанных концентраций 2,5 нМ и 25 нМ, соответственно (значения представлены попарно в виде светлых и более темных серых столбиков). Клетки переходноклеточной папилломы мочевого пузыря человека RT4, эндогенно экспрессирующие LPA, высевали при плотности 125000 клеток на лунку в 24-луночном формате поверх предварительно нанесенных трансфекционных комплексов (обратная трансфекция) в указанной концентрации. Через 24 часа после трансфекции общую РНК выделяли с применением набора Spin Cell Mini Kit 250 (Stratec). Уровни мРНК LPA определяли с помощью количественной ПЦР в режиме реального времени относительно экспрессии мРНК PPIB в соответствующих образцах в качестве транскрипта конститутивного гена. Значения нормировали к количеству мРНК LPA, обнаруженному в необработанных клетках (интрапланшет). Неподавляющее соединение миРНК трансфицировали в качестве дополнительного контроля. Представлены средние значения и стандартные отклонения (SD) нормированных трехкратных значений. Результаты показаны на Фигуре 1.
Пример 3
Дозозависимый ответ неконъюгированных LPA-нацеленных соединений миРНК на экспрессию мРНК LPA в клетках RT-4 человека.
Клетки переходноклеточной папилломы мочевого пузыря человека RT4 подвергали обратной трансфекции, как описано выше (пример 2), и обрабатывали в указанной концентрации (в диапазоне от 100 нМ до 0,2 нМ) с применением различных неконъюгированных соединений миРНК (таблица 1), как отмечено. Через 24 ч после трансфекции общую РНК выделяли с применением набора Spin Cell Mini Kit 250 (Stratec). Уровни мРНК LPA определяли с помощью кПЦР в режиме реального времени относительно экспрессии мРНК PPIB в соответствующих образцах в качестве транскрипта конститутивного гена. Значения нормировали к количеству мРНК LPA, обнаруженному в необработанных клетках. Столбики представляют остаточную экспрессию мРНК LPA для каждой точки данных. Результаты показаны на Фигуре 2.
Пример 4
Ингибирование экспрессии мРНК LPA в первичных гепатоцитах человека и яванской макаки под действием различных доз GalNAc-L1 LPA-1038-конъюгированной молекулы миРНК, доставленной путем опосредуемого рецепторами поглощения.
Первичные гепатоциты (ThermoFisher) высевали на покрытые коллагеном 96-луночные планшеты при плотности 45000 клеток на лунку (яванская макака) и 30000 клеток на лунку (человек). GalNAc-L1-конъюгированный LPA-1038 добавляли в указанных концентрациях (нМ) сразу после посева. Через 24 часа после обработки миРНК общую РНК выделяли с применением набора InviTrap RNA cell HTS (Stratec) для 96-луночного планшета. Уровни мРНК LPA определяли с помощью кПЦР в режиме реального времени относительно уровней мРНК актина (яванская макака) или APOB (человека) в соответствующих образцах в качестве транскрипта конститутивного гена. Значения нормировали к экспрессии LPA в необработанных клетках. Средние значения и стандартные отклонения нормированных трехкратных значений остаточных уровней мРНК LPA показаны в виде черных столбиков. Результаты показаны на Фигуре 3.
Пример 5
Выключение мРНК LPA в первичных гепатоцитах человека под действием различных указанных L6-GalNAc-конъюгированных миРНК в первичных гепатоцитах человека после опосредуемой рецепторами доставки.
Первичные гепатоциты человека (ThermoFisher) высевали на покрытые коллагеном 96-луночные планшеты в количестве 30000 клеток на лунку (96-луночный формат). GalNAc-L6-конъюгированные миРНК, включая неподавляющий контроль, добавляли в двух указанных концентрациях сразу после посева клеток. Через 24 часа после обработки миРНК общую РНК выделяли с применением набора InviTrap RNA cell HTS (Stratec) для 96-луночного планшета. Уровни экспрессии мРНК LPA определяли с помощью кПЦР в режиме реального времени относительно мРНК APOB в качестве транскрипта конститутивного гена. Значения нормировали к экспрессии мРНК LPA в необработанных клетках, а остаточные уровни мРНК LPA представлены попарно в виде столбиков (100 нМ черные столбики, 20 нМ серые столбики). Средние значения и стандартные отклонения нормированных трехкратных значений показаны на Фигуре 5.
Пример 6 - Синтез конъюгатов
Примерные соединения синтезировали в соответствии со способами, описанными ниже, и способами, известными специалисту в данной области техники. Сборку олигонуклеотидной цепи и линкерных строительных блоков выполняли с помощью твердофазного синтеза с применением фосфоамидитной методологии. Конъюгацию GalNAc осуществляли путем образования пептидной связи между строительным блоком GalNAc-карбоновой кислоты и предварительно собранным и очищенным олигонуклеотидом, содержащим необходимое количество присоединенных амино-модифицированных линкерных строительных блоков.
Синтез, удаление защитных групп и очистку олигонуклеотидов проводили в соответствии со стандартными процедурами, известными в данной области техники.
Все олигонуклеотиды синтезировали на синтезаторе AKTA oligopilot с применением стандартных фосфоамидитных реакций. Применяли коммерчески доступную твердую подложку и 2'-O-метил-РНК-фосфоамидиты, 2'-фтор, 2'-дезокси-РНК-фосфоамидиты (все со стандартной защитой, ChemGenes, LinkTech) и коммерчески доступный 3'-амино-модификатор TFA-амино-C-6-lcaa CPG 500 Å (Chemgenes). Пер-ацетилированный галактозамин 8 доступен коммерчески.
Вспомогательные реагенты приобретали у EMP Biotech. Синтез выполняли с применением 0,1 М раствора фосфоамидита в сухом ацетонитриле, и в качестве активатора применяли бензилтиотетразол (БТТ) (0,3 М в ацетонитриле). Время связывания составляло 15 мин. Применяли цикл Cap/OX/Cap или Cap/Thio/Cap (Cap: Ac2O/NMI/лутидин/ацетонитрил, окислитель: 0,1 М I2 в пиридине/H2O). Фосфотиоаты вводили с применением стандартного коммерчески доступного реагента для тиолирования (EDITH, Link technologies). Отщепление DMT осуществляли с помощью обработки 3% дихлоруксусной кислотой в толуоле. После завершения запрограммированных циклов синтеза выполняли промывку диэтиламином (DEA). Все олигонуклеотиды синтезировали в режиме DMT-off.
Присоединение линкерной группы, происходящей из серинола, осуществляли с применением либо (S)-DMT-серинол(TFA)-сукцинат-lcaa-CPG 10, загруженного основанием, либо (S)-DMT-серинол(TFA)фосфоамидита 7 (синтез выполняли, как описано в Hoevelmann et al. (Chem. Sci., 2016, 7, 128-135)). Трехантенные кластеры GalNAc (ST23/C4XLT) вводили путем последовательного связывания соответствующих утроительных амидитных производных (C4XLT-phos) с последующим добавлением амидита GalNAc (ST23-phos).
Одиночные цепи отщепляли от CPG с помощью обработки 40% водн. метиламином. Полученный неочищенный олигонуклеотид очищали с помощью ионообменной хроматографии (Resource Q, 6 мл, GE Healthcare) на системе для ВЭЖХ AKTA Pure с применением градиента хлорида натрия. Содержащие продукт фракции объединяли, обессоливали на колонке для гель-проникающей хроматографии (Zetadex, EMP Biotech) и лиофилизировали.
Отдельные одиночные цепи растворяли в концентрации 60 OD/мл в H2O. Оба раствора отдельных олигонуклеотидов добавляли вместе в реакционный сосуд. Для облегчения мониторинга реакции выполняли титрование. Первую цепь добавляли с 25% избытком по сравнению со второй цепью, что определяли по УФ-поглощению при 260 нм. Реакционную смесь нагревали до 80°C в течение 5 минут и затем медленно охлаждали до комнатной температуры. Образование двойной цепи контролировали по образованию ионных пар с помощью ВЭЖХ с обращенной фазой. На основании УФ-площади остаточной одиночной цепи рассчитывали необходимое количество второй цепи и добавляли его к реакционной смеси. Реакционную смесь снова нагревали до 80°C и медленно охлаждали до комнатной температуры. Эту процедуру повторяли до тех пор, пока не было обнаружено менее 10% остаточной одиночной цепи.
Синтез соединений 2-10
Соединения 2-5 и (S)-DMT-серинол(TFA)-фосфоамидит 7 синтезировали в соответствии с опубликованными в литературе методами (Hoevelmann et al. Chem. Sci., 2016,7, 128-135).
(S)-4-(3-(бис(4-метоксифенил)(фенил)метокси)-2-(2,2,2-трифторацетамидо)пропокси)-4-оксобутановая кислота (6).
К раствору 5 в пиридине добавляли сукциновый ангидрид, а затем DMAP. Полученную смесь перемешивали при комнатной температуре в течение ночи. Весь исходный материал был израсходован согласно оценке методом ТСХ. Реакцию концентрировали. Неочищенный материал подвергали хроматографии на силикагеле, используя градиент от 0% до 5% метанола в ДХМ (+ 1% триэтиламина), с получением 1,33 г 6 (выход = 38%). m/z (ESI-): 588,2 (100%), (рассчитано для C30H29F3NO8- [M-H]- 588,6). 1H-ЯМР: (400 МГц, CDCl3)δ[ppm]=7,94 (d, 1H, NH), 7,39-7,36 (m, 2H, СНарил), 7,29-7,25 (m, 7H, СНарил), 6,82-6,79 (m, 4H, СНарил), 4,51-4,47 (m, 1H), 4,31-4,24 (m, 2H), 3,77 (s, 6H, 2xDMTr-OMe), 3,66-3,60 (m, 16H, HNEt3+), 3,26-3,25 (m, 2H), 2,97-2,81 (m, 20H, NEt3), 2,50-2,41 (4H, m), 1,48-1,45 (m, 26H, HNEt3+), 1,24-1,18 (m, 29H, NEt3).
(S)-DMT-серинол(TFA)-сукцинат-Icaa-CPG (10)
(S)-DMT-серинол(TFA)-сукцинат (159 мг, 270 мкмоль) и HBTU (113 мг, 299 мкмоль) растворяли в CH3CN (10 мл). К раствору добавляли диизопропилэтиламин (DIPEA, 94 мкл, 540 мкмоль), и смесь перемешивали в течение 2 мин с последующим добавлением нативного амино-Icaa-CPG (500 A, 3 г, содержание амина: 136 мкмоль/г). Суспензию осторожно встряхивали при комнатной температуре на шейкере, имитирующем движение запястья, в течение 16 ч, затем фильтровали и промывали ДХМ и EtOH. Твердую подложку высушивали в вакууме в течение 2 ч. Не вступившие в реакцию амины на подложке кэппировали путем перемешивания с уксусным ангидридом/лутидином/N-метилимидазолом при комнатной температуре. Промывку подложки повторяли, как описано выше. Твердое вещество высушивали в вакууме с получением твердой подложки 10 (3 г, загрузка 26 мкмоль/г).
GalNAc синтон (9)
Синтез GalNAc синтона 9 выполняли, как описано в Nair et al. J. Am. Chem. Soc., 2014, 136 (49), pp 16958-16961, с 46% выходом в течение двух этапов.
Характеристические данные соответствуют опубликованным данным.
Синтез олигонуклеотидов
Все одноцепочечные олигонуклеотиды синтезировали в соответствии с условиями реакции, описанными выше и на Фигуре 13 и 14.
Все готовые одноцепочечные продукты анализировали с помощью АХ-ВЭЖХ для подтверждения их чистоты. Чистота представлена как % FLP (% полноразмерного продукта), который представляет собой процент УФ-площади под сигналом заданного продукта на УФ-кривой анализа готового продукта методом АХ-ВЭЖХ. Идентичность соответствующих одноцепочечных продуктов (немодифицированных, амино-модифицированных предшественников или GalNAc-конъюгированных олигонуклеотидов) подтверждали с помощью анализа методом ЖХ/МС.
Таблица 3: Одноцепочечные неконъюгированные олигонуклеотиды
(АХ-ВЭЖХ)
5'1×NH2 означает положение (5'-конец) и количество (1×NH2) свободных аминогрупп, происходящих из серинола, которые доступны для конъюгации. Например, 1×3'NH2 на A0264 означает, что имеется свободная аминогруппа, которая может реагировать с GalNAc синтоном 9 на 3'-конце цепи A0264. 3'5'1×NH2 означает, что имеется одна свободная аминогруппа, происходящая из серинола, которая может реагировать с линкером GalNAc 9 на 3'-конце и 5'-конце цепи.
Синтез конъюгатов 1-3 и эталонных конъюгатов 1-2
Конъюгированные одиночные цепи для конъюгатов 1-2 и эталонных конъюгатов 1-2
Конъюгацию GalNAc синтона (9) осуществляли путем связывания с функциональной аминогруппой серинола соответствующей олигонуклеотидной цепи 11 с применением связывающего реагента для образования пептидной связи. Таким образом, соответствующую амино-модифицированную молекулу-предшественника 11 растворяли в H2O (500 OD/мл) и добавляли ДМСО (ДМСО/H2O, 2/1, об./об.), а затем DIPEA (2,5% от общего объема). В отдельном реакционном сосуде выполняли предварительную активацию GalN(Ac4)-C4-кислоты (9) с помощью взаимодействия 2 экв. (на функциональную аминогруппу в амино-модифицированном предшественнике олигонуклеотида 11) компонента карбоновой кислоты с 2 экв. HBTU в присутствии 8 экв. DIPEA в ДМСО. Через 2 мин предварительно активированное соединение 9 добавляли к раствору соответствующей амино-модифицированной молекулы-предшественника. Через 30 мин ход реакции контролировали с помощью ЖХ/МС или АХ-ВЭЖХ. После завершения реакции конъюгирования неочищенный продукт осаждали добавлением 10× iPrOH и 0,1×2 М NaCl и собирали с помощью центрифугирования и декантации. Чтобы высвободить ацетилированные гидроксильные группы в группах GalNAc, полученный осадок растворяли в 40% MeNH2 (1 мл на 500 OD) и через 15 минут при комнатной температуре разбавляли в H2O (1:10), в завершение повторно очищали с помощью анионного обмена и гель-проникающей хроматографии и лиофилизировали с получением конечного продукта 12.
Таблица 4: Одноцепочечные GalNAc-конъюгированные олигонуклеотиды
Образование двойной цепи
Образование двойной цепи выполняли в соответствии со способами, описанными выше.
Чистота двойной цепи представлена в % двойной цепи, который представляет собой процент УФ-площади под сигналом заданного продукта на УФ-кривой анализа образования ионных пар методом ВЭЖХ с обращенной фазой (ИП-ОФ-ВЭЖХ).
Таблица 5: Конъюгаты нуклеиновых кислот
Последовательности
Обозначения модификаций для следующих последовательностей:
f обозначает 2'-фтор-2'-дезоксирибонуклеотид или 2'-фторрибонуклеотид (термины являются взаимозаменяемыми)
m обозначает 2'-O-метилрибонуклеотид
(ps) обозначает фосфотиоатную связь
Ser(GN) представляет собой строительный блок GalNAc-C4, присоединенный к линкерной группе, происходящей из серинола:
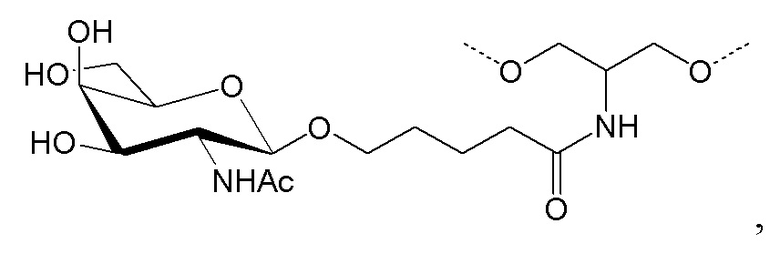
где O--- представляет собой связь между атомом кислорода и, например, H, фосфодиэфирную связь или фосфотиоатную связь.
C4XLT представляет собой:
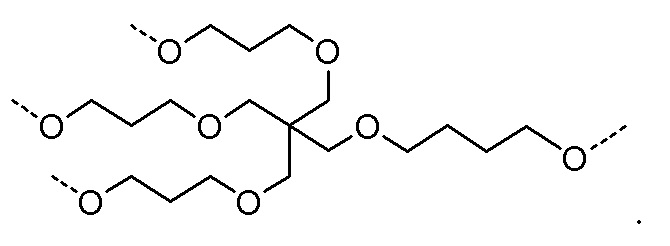
ST23 представляет собой:
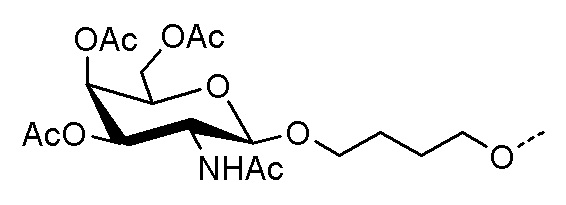
Синтез фосфоамидитных производных C4XLT (C4XLT-phos), а также ST23 (ST23-phos) можно выполнять, как описано в WO2017/174657.
C4XLT-phos:
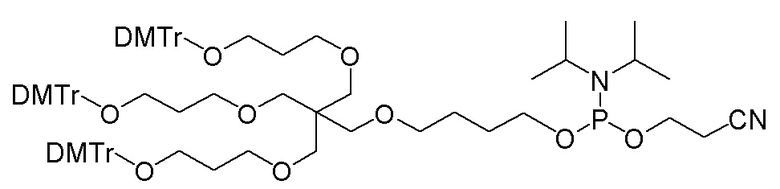
ST23-phos:
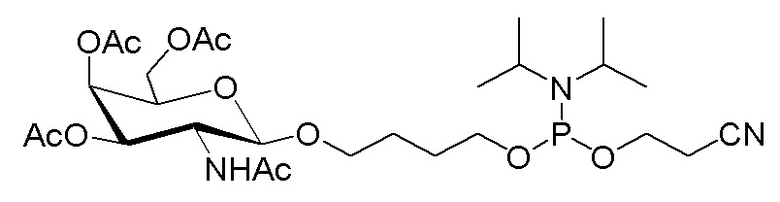
Конъюгат 1
Антисмысловая цепь-STS16001AL33 (SEQ ID NO: 127)
5' mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU (ps) Ser(GN) 3'
Смысловая цепь-STS16001BL20 (SEQ ID NO: 128)
5' Ser(GN) (ps) fA mA fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA 3'
Конъюгат 2
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU
Смысловая цепь-STS16001BV1L42 (SEQ ID NO: 130)
Ser(GN) (ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA (ps) Ser(GN)
Конъюгат 3
Антисмысловая цепь-STS16001AL33 (SEQ ID NO: 127)
5' mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU (ps) Ser(GN) 3'
Смысловая цепь-STS16001BV1L42 (SEQ ID NO: 130)
5' Ser(GN) (ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA (ps) Ser(GN) 3'
Эталонный конъюгат 1
Антисмысловая цепь–STS16001A (SEQ ID NO: 129)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU
Смысловая цепь-STS16001BL20 (SEQ ID NO: 128)
Ser(GN) (ps) fA mA fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA
Эталонный конъюгат 2
Антисмысловая цепь-STS16001AL33 (SEQ ID NO: 127)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU (ps) Ser(GN)
Смысловая цепь-STS16001V1B (SEQ ID NO: 131)
fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA
Эталонный конъюгат 3
Антисмысловая цепь-STS18001A (A0130; SEQ ID NO: 132)
mU (ps) fC (ps) mG fA mA fG mU fA mU fU mC fC mG fC mG fU mA (ps) fC (ps) mG
Смысловая цепь-STS18001BL4 (A0131, SEQ ID NO: 133)
[(ST23) (ps)]3 C4XLT (ps) fC mG fU mA fC mG fC mG fG mA fA mU fA mC fU mU fC (ps) mG (ps) fA
Эталонный конъюгат 4
Антисмысловая цепь-STS16001AL33 (SEQ ID NO: 127)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU
Смысловая цепь-STS16001BL4 (SEQ ID NO: 134)
5'[(ST23) (ps)]3 C4XLT(ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA
Пример 7 - Определение выключения TTR в условиях in vitro для различных GalNAc-конъюгатов миРНК TTR
Первичные гепатоциты мыши высевали в предварительно покрытые коллагеном 96-луночные планшеты (Thermo Fisher Scientific, каталожный номер A1142803) при плотности клеток 30000 клеток на лунку и обрабатывали миРНК-конъюгатами в концентрациях, которые находятся в пределах диапазона от 10 нМ до 0,0001 нМ. Через 24 часа после обработки клетки лизировали и РНК экстрагировали с помощью препаративных наборов InviTrap® RNA Cell HTS 96/C24×96 (Stratec, каталожный номер 7061300400) в соответствии с протоколом производителя. Уровни транскриптов TTR и мРНК конститутивных генов (PtenII) определяли количественно с помощью анализа TaqMan.
Экспрессия гена-мишени в первичных гепатоцитах мыши через 24 ч после обработки конъюгатами согласно настоящему изобретению, конъюгатами 1-3, показала, что экспрессия гена-мишени снижается с увеличением дозы конъюгата по сравнению с отрицательными контролями (см. столбик «UT» и эталонный конъюгат 3), как показано на Фигуре 15. Это свидетельствует о том, что первая цепь связывается с геном-мишенью, что снижает экспрессию гена. На Фигуре 15 также показаны уровни экспрессии гена-мишени для эталонных конъюгатов 1 и 2, которые действуют как конъюгаты для сравнения. Как следует из сравнения данных, представленных на Фигуре 15A и 15C, а также 15B и 15C, конъюгаты согласно настоящему изобретению (конъюгаты 1-3) снижают экспрессию гена-мишени по сравнению с эталонными конъюгатами 1 и 2. Наиболее эффективным конъюгатом при 0,01 нМ, по-видимому, является конъюгат 2. Наиболее эффективным конъюгатом при 0,1 нМ, 0,5 нМ, 1 нМ и 10 нМ, по-видимому, является конъюгат 3.
Пример 8 - Временная динамика сывороточного TTR у мышей в условиях in vivo
Мышей c57BL/6 обрабатывали с помощью подкожного введения 1 мг/кг миРНК-конъюгатов в 0 день. Образцы сыворотки отбирали на 7, 14 и 27 день путем сбора крови из орбитального синуса и хранили при -20°C до анализа. Количественное определение TTR в сыворотке выполняли с помощью ИФА для мышиного преальбумина (ALPCO, 41-PALMS/партия 22, 2008003B) в соответствии с протоколом производителя (разведение образца 1:8000 или 1:800).
Результаты для временной динамики сывороточного TTR в когортах мышей c57BL/6 с n=4 через 7, 14 и 27 дней после обработки с помощью подкожного введения 1 мг/кг конъюгатов 1-3, эталонных конъюгатов 1, 2 и 4 и у индивидуумов с имитированной обработкой (ФСБ) показаны на Фигуре 16. Как свидетельствуют данные на Фигуре 16, конъюгаты согласно настоящему изобретению особенно эффективны в уменьшении экспрессии гена-мишени по сравнению с отрицательным контролем (ФСБ) и эталонными конъюгатами 1, 2 и, в частности, эталонным конъюгатом 4. Конъюгаты 2 и 3 также более эффективны, чем эталонные конъюгаты 1, 2 и 4. Наиболее эффективным конъюгатом является конъюгат 2. Таким образом, можно ожидать, что уровень дозирования конъюгата 3 будет приблизительно в три раза ниже для достижения аналогичного начального выключения и также приведет к большей продолжительности выключения по сравнению с эталонным конъюгатом 4.
Пример 9 - Синтез конъюгатов 2
Примерные соединения синтезировали в соответствии со способами, описанными ниже, и способами, известными специалисту в данной области техники. Сборку олигонуклеотидной цепи и линкерных строительных блоков выполняли с помощью твердофазного синтеза с применением фосфоамидитной методологии. Конъюгацию GalNAc осуществляли путем образования пептидной связи между строительным блоком GalNAc-карбоновая кислота и предварительно собранным и очищенным олигонуклеотидом, содержащим необходимое количество присоединенных амино-модифицированных линкерных строительных блоков.
Синтез, удаление защитных групп и очистку олигонуклеотидов проводили в соответствии со стандартными процедурами, известными в данной области техники.
Все олигонуклеотиды синтезировали на синтезаторе AKTA oligopilot с применением стандартных фосфоамидитных реакций. Применяли коммерчески доступную твердую подложку и 2'-O-метил-РНК-фосфоамидиты, 2'-фтор, 2'-дезокси-РНК-фосфоамидиты (все со стандартной защитой, ChemGenes, LinkTech) и коммерчески доступный 3'-амино-модификатор TFA-амино-C-6-lcaa-CPG 500 Å (Chemgenes). Пер-ацетилированный галактозамин 8 доступен коммерчески.
Вспомогательные реагенты приобретали у EMP Biotech. Синтез выполняли с применением 0,1 М раствора фосфоамидита в сухом ацетонитриле, и в качестве активатора применяли бензилтиотетразол (БТТ) (0,3 М в ацетонитриле). Время связывания составляло 15 мин. Применяли цикл Cap/OX/Cap или Cap/Thio/Cap (Cap: Ac2O/NMI/лутидин/ацетонитрил, окислитель: 0,1 М I2 в пиридине/H2O). Фосфотиоаты вводили с применением стандартного коммерчески доступного реагента для тиолирования (EDITH, Link technologies). Отщепление DMT осуществляли с помощью обработки 3% дихлоруксусной кислотой в толуоле. После завершения запрограммированных циклов синтеза выполняли промывку диэтиламином (DEA). Все олигонуклеотиды синтезировали в режиме DMT-off.
Присоединение линкерной группы, происходящей из серинола, осуществляли с применением либо (S)-DMT-серинол(TFA)-сукцинат-lcaa-CPG 10, загруженного основанием, либо (S)-DMT-серинол(TFA)-фосфоамидита 7 (синтез выполняли, как описано в литературе Hoevelmann et al. Chem. Sci., 2016,7, 128-135). Трехантенные кластеры GalNAc (ST23/C4XLT или ST23/C6XLT) вводили путем последовательного связывания соответствующих утроительных амидитных производных (C4XLT-phos или C6XLT-phos) с последующим добавлением амидита GalNAc (ST23-phos).
Одиночные цепи отщепляли от CPG с помощью обработки 40% водн. метиламином. Полученный неочищенный олигонуклеотид очищали с помощью ионообменной хроматографии (Resource Q, 6 мл, GE Healthcare) на системе для ВЭЖХ AKTA Pure с применением градиента хлорида натрия. Содержащие продукт фракции объединяли, обессоливали на колонке для гель-проникающей хроматографии (Zetadex, EMP Biotech) и лиофилизировали.
Отдельные одиночные цепи растворяли в концентрации 60 OD/мл в H2O. Оба раствора отдельных олигонуклеотидов добавляли вместе в реакционный сосуд. Для облегчения мониторинга реакции выполняли титрование. Первую цепь добавляли с 25% избытком по сравнению со второй цепью, что определяли по УФ-поглощению при 260 нм. Реакционную смесь нагревали до 80°C в течение 5 минут и затем медленно охлаждали до комнатной температуры. Образование двойной цепи контролировали по образованию ионных пар методом ВЭЖХ с обращенной фазой. На основании УФ-площади остаточной одиночной цепи рассчитывали необходимое количество второй цепи и добавляли его к реакционной смеси. Реакционную смесь снова нагревали до 80°C и медленно охлаждали до комнатной температуры. Эту процедуру повторяли до тех пор, пока не было обнаружено менее 10% остаточной одиночной цепи.
Синтез соединений 2-10
Соединения 2-5 и (S)-DMT-серинол(TFA)-фосфоамидит 7 синтезировали в соответствии с опубликованными в литературе способами (Hoevelmann et al. Chem. Sci., 2016,7, 128-135).
(S)-4-(3-(бис(4-метоксифенил)(фенил)метокси)-2-(2,2,2-трифторацетамидо)пропокси)-4-оксобутановая кислота (6).
К раствору 5 в пиридине добавляли сукциновый ангидрид, а затем DMAP. Полученную смесь перемешивали при комнатной температуре в течение ночи. Весь исходный материал был израсходован согласно оценке методом ТСХ. Реакцию концентрировали. Неочищенный материал подвергали хроматографии на силикагеле, используя градиент от 0% до 5% метанола в ДХМ (+ 1% триэтиламина), с получением 1,33 г 6 (выход=38%). m/z (ESI-): 588,2 (100%), (рассчитано для C30H29F3NO8- [M-H]- 588,6), 1H-ЯМР: (400 МГц, CDCl3) δ [ppm]=7,94 (d, 1H, NH), 7,39-7,36 (m, 2H, СНарил), 7,29-7,25 (m, 7H, СНарил), 6,82-6,79 (m, 4H, СНарил), 4,51-4,47 (m, 1H), 4,31-4,24 (m, 2H), 3,77 (s, 6H, 2×DMTr‐OMe), 3,66–3,60 (m, 16H, HNEt3+), 3,26-3,25 (m, 2H), 2,97-2,81 (m, 20H, NEt3), 2,50-2,41 (4H, m), 1,48–1,45 (m, 26H, HNEt3+), 1,24-1,18 (m, 29H, NEt3).
(S)-DMT-серинол(TFA)-сукцинат-Icaa-CPG (10)
(S)-DMT-серинол(TFA)-сукцинат (159 мг, 270 мкмоль) и HBTU (113 мг, 299 мкмоль) растворяли в CH3CN (10 мл). К раствору добавляли диизопропилэтиламин (DIPEA, 94 мкл, 540 мкмоль) и смесь перемешивали в течение 2 мин с последующим добавлением нативного амино-Icaa-CPG (500 A, 3 г, содержание амина: 136 мкмоль/г). Суспензию осторожно встряхивали при комнатной температуре на шейкере, имитирующем движение запястья, в течение 16 ч, затем фильтровали и промывали ДХМ и EtOH. Твердую подложку высушивали в вакууме в течение 2 ч. Не вступившие в реакцию амины на подложке кэппировали путем перемешивания с уксусным ангидридом/лутидином/N-метилимидазолом при комнатной температуре. Промывку подложки повторяли, как описано выше. Твердое вещество высушивали в вакууме с получением твердой подложки 10 (3 г, загрузка 26 мкмоль/г).
GalNAc синтон (9)
Синтез GalNAc синтона 9 выполняли, как описано в Nair et al. J. Am. Chem. Soc., 2014, 136 (49), pp 16958-16961, с 46% выходом в течение двух этапов.
Характеристические данные соответствуют опубликованным данным.
Синтез олигонуклеотидов
Все одноцепочечные олигонуклеотиды синтезировали в соответствии с условиями реакции, описанными выше и на Фигуре 13 и 14.
Все готовые одноцепочечные продукты анализировали с помощью АХ-ВЭЖХ для подтверждения их чистоты. Чистота представлена в % FLP (% полноразмерного продукта), который представляет собой процент УФ-площади под сигналом заданного продукта на УФ-кривой анализа конечного продукта методом АХ-ВЭЖХ. Идентичность соответствующих одноцепочечных продуктов (немодифицированных, амино-модифицированных предшественников, C4XLT/ST23 или C6XLT/ST23 GalNAc конъюгированных олигонуклеотидов) подтверждали с помощью анализа методом ЖХ/МС.
Таблица 7: Одноцепочечные неконъюгированные и конъюгированные на колонке олигонуклеотиды
Синтез конъюгатов 1-3 и эталонных конъюгатов 1-2
Конъюгированные одиночные цепи для конъюгатов 1-2 и эталонных конъюгатов 1-2
Конъюгацию GalNAc синтона (9) осуществляли путем связывания с функциональной аминогруппой серинола соответствующей олигонуклеотидной цепи 11 с применением связывающего реагента для образования пептидной связи. Таким образом, соответствующую амино-модифицированную молекулу-предшественника 11 растворяли в H2O (500 OD/мл) и добавляли ДМСО (ДМСО/H2O, 2/1, об./об.), а затем DIPEA (2,5% от общего объема). В отдельном реакционном сосуде выполняли предварительную активацию GalN(Ac4)-C4-кислоты (9) с помощью взаимодействия 2 экв. (на функциональную аминогруппу в амино-модифицированном предшественнике олигонуклеотида 11) компонента карбоновой кислоты с 2 экв. HBTU в присутствии 8 экв. DIPEA в ДМСО. Через 2 мин предварительно активированное соединение 9 добавляли к раствору соответствующей амино-модифицированной молекулы-предшественника. Через 30 мин ход реакции контролировали с помощью ЖХ/МС или АХ-ВЭЖХ. После завершения реакции конъюгирования неочищенный продукт осаждали добавлением 10× iPrOH и 0,1× 2 М NaCl и собирали с помощью центрифугирования и декантации. Чтобы высвободить ацетилированные гидроксильные группы в группах GalNAc, полученный осадок растворяли в 40% MeNH2 (1 мл на 500 OD) и через 15 минут при комнатной температуре разбавляли в H2O (1:10), и в завершение повторно очищали с помощью анионного обмена и гель-проникающей хроматографии и лиофилизировали с получением конечного продукта 12.
Таблица 8: Одноцепочечные GalNAc-конъюгированные олигонуклеотиды
(АХ-ВЭЖХ)
Образование двойной цепи
Двойную цепь получали в соответствии со способами, описанными выше.
Чистота двойной цепи представлена в % двойной цепи, который представляет собой процент УФ-площади под сигналом заданного продукта на УФ-кривой анализа ИП-ОФ-ВЭЖХ.
Таблица 9: Конъюгаты нуклеиновых кислот
Последовательности
Обозначения модификаций для следующих последовательностей:
f обозначает 2'-фтор-2'-дезоксирибонуклеотид или 2'-фторрибонуклеотид (термины являются взаимозаменяемыми)
m обозначает 2'-O-метилрибонуклеотид
(ps) обозначает фосфотиоатную связь
Ser(GN) представляет собой строительный блок GalNAc-C4, присоединенный к линкерной группе, происходящей из серинола:

где O--- представляет собой связь между атомом кислорода и, например, H, фосфодиэфирную связь или фосфотиоатную связь.
C4XLT представляет собой:
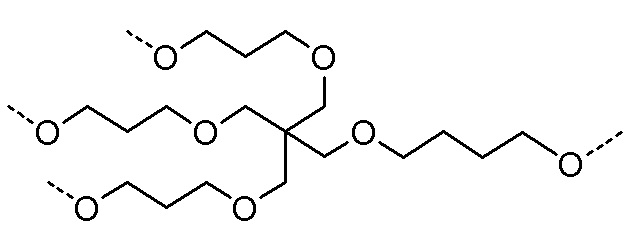
C6XLT представляет собой:
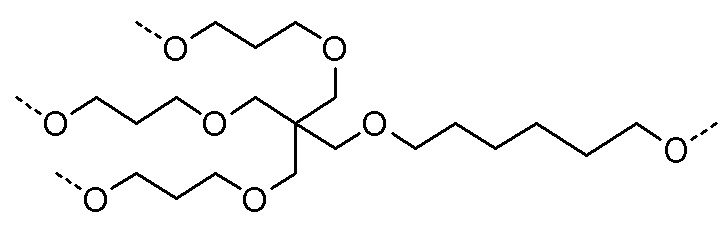
ST23 представляет собой:

Синтез фосфоамидитных производных C4XLT (C4XLT-phos), C6XLT (C6XLT-phos), а также ST23 (ST23-phos) можно выполнять, как описано в WO2017/174657.

Пример 10
Равное дозозависимое выключение для миРНК, нацеленной на LPA, с двумя отдельными блоками GalNAc, конъюгированными со второй цепью, по сравнению с трехантенным блоком GalNAc на 5' второй цепи в первичных гепатоцитах яванской макаки.
МиРНК модифицированы с применением чередующихся 2'-OMe/2'-F и каждая из них содержит две фосфотиоатные (PS) межнуклеотидные связи в своих двух 5' и 3'-концевых межнуклеотидных связях. В конъюгате 19 каждый блок серинол-GalNAc присоединен за счет PS-связи к 5' и 3' второй цепи. В конъюгате 20 два концевых 5'-внутренних нуклеотида второй цепи представляют собой фосфодиэфиры, и трехантенный GalNAc-линкер присоединен за счет PS-связи к этому концу.
Дозозависимое выключение LPA в первичных гепатоцитах яванской макаки оценивали через 24 ч после обработки 100, 20, 4, 0,8, 0,16, 0,032 и 0,006 нМ миРНК. Эталонный контроль представляет собой конструкцию 2, ненацеливающий контроль назван Cte. Ct-значение транскрипта для каждой группы обработки нормировали к значению ct транскрипта конститутивного гена ACTB (Δct) и к необработанным гепатоцитам, названным ut (ΔΔct).
Данные показаны на Фигуре 18
Материал и методы:
миРНК
Подпись
(ps) - фосфотиоат
(po) - фосфодиэфир
Праймер:
Общие методы
Эксперименты в условиях in vitro
Первичные мышиные гепатоциты (Thermo Scientific: GIBCO партия № MC798) размораживали и заменяли среду для криоконсервации средой Williams E с добавлением 5% фетальной бычьей сыворотки (ФБС), 1 мкМ дексаметазона, 2 мМ GlutaMax, 1% PenStrep, 4 мг/мл человеческого рекомбинантного инсулина, 15 мМ Hepes. Плотность клеток доводили до 250000 клеток на 1 мл. По 100 мкл на лунку этой клеточной суспензии высевали в предварительно покрытые коллагеном 96-луночные планшеты. Тестируемое изделие предварительно разводили в одной и той же среде (5-кратно концентрированной) для каждой концентрации, и 25 мкл этой предварительно разведенной миРНК или только среды добавляли к клеткам. Клетки культивировали при 37°C и 5% CO2. Через 24 ч после обработки супернатант удаляли, а клетки промывали в холодном ФСБ и добавляли 250 мкл РНК-лизисного буфера S (Stratec). После 15 мин инкубации при комнатной температуре планшеты хранили при -80°C до выделения РНК в соответствии с протоколом производителя.
Анализ TaqMan
Для анализа mTTR&PTEN MultiPlex TaqMan по 10 мкл выделенной РНК для каждой группы обработки смешивали с 10 мкл общей реакционной смеси для ПЦР (TAKYON low Rox), содержащей 600 нМ праймера mTTR, 400 нМ праймера ApoB и 200 нМ каждого зонда, а также 0,5 единицы полимеразы Euroscript II RT с 0,2 ед. ингибитора РНКазы. Анализ TaqMan проводили в 384-луночном планшете с 10 мин RT-этапом при 48°C, 3 мин начальной денатурацией при 95°C и 40 циклами при 95°C в течение 10 с и 60°C в течение 1 минуты. Праймеры содержат два из BHQ1, FAM и YY, по одному на каждом конце последовательности.
Для анализа TMPRSS6&ApoB MultiPlex TaqMan по 10 мкл выделенной РНК для каждой группы обработки смешивали с 10 мкл общей реакционной смеси для ПЦР (TAKYON low Rox), содержащей 800 нМ праймера TMPRSS6, 100 нМ праймера ApoB и 200 нМ каждого зонда, а также 0,5 единицы полимеразы Euroscript II RT с 0,2 ед. ингибитора РНКазы. Анализ TaqMan проводили в 384-луночном планшете с 10 мин RT-этапом при 48°C, 3 мин начальной денатурацией при 95°C и 40 циклами при 95°C в течение 10 с и 60°C в течение 1 мин.
Эксперименты в условиях in vivo
Для сравнения эффективности в условиях in vivo различных конъюгатов миРНК 1 мг/кг миРНК, растворенной в ФСБ, вводили подкожно в область лопатки мышей c57BL/6. Когорты с n=6 обрабатывали миРНК, нацеленной на Aldh2 или Tmprss6, в 1 день и умерщвляли в выбранные моменты времени после обработки. Образцы печени быстро замораживали в жидком азоте и хранили при -80°C до экстракции РНК с помощью мини-набора InviTrap Spin Tissue RNA (Stratec) в соответствии с инструкцией производителя. После этого уровень транскриптов Aldh2, Tmprss6 и Pten определяли количественно, как описано выше.
Анализ стабильности в тритосомах
Чтобы исследовать стабильность к РНКазе в эндосомальном/лизосомальном компартменте печеночных клеток в условиях in vitro, миРНК инкубировали в течение 0 ч, 4 ч, 24 ч или 72 ч в тритосомах печени крыс Sprague Dawley (Tebu-Bio, каталожный номер R0610.LT, партия: 1610405, pH: 7,4, 2,827 единиц/мл). Для имитации закисленной среды тритосомы смешивали в соотношении 1:10 с буфером с низким рН (1,5 М уксусная кислота, 1,5 М ацетат натрия, рН=4,75). 30 мкл подкисленных тритосом. Затем 10 мкл миРНК (20 мкМ) смешивали и инкубировали в течение указанных моментов времени при 37°C. После инкубации РНК выделяли с помощью картриджей для стартового набора Clarity OTX (Phenomenex, каталожный номер KSO-8494) в соответствии с протоколом производителя для биологических жидкостей. Лиофилизированную РНК восстанавливали в 30 мкл H2O, смешивали с 4-кратным буфером для загрузки и 5 мкл загружали в 20% TBE-полиакриламидный гель для электрофореза (PAGE) для разделения в качественном полуколичественном анализе. PAGE проводили при 120 В в течение 2 ч, и РНК визуализировали с помощью окрашивания бромидом этидия с последующей цифровой визуализацией с применением системы визуализации Biorad.
Пример 11 - Синтез конъюгатов 3
Примерные соединения синтезировали в соответствии со способами, описанными ниже, и способами, известными специалисту в данной области техники. Сборку олигонуклеотидной цепи и линкерных строительных блоков выполняли с помощью твердофазного синтеза с применением фосфоамидитной методологии. Конъюгацию GalNAc осуществляли путем образования пептидной связи между строительным блоком GalNAc-карбоновая кислота и предварительно собранным и очищенным олигонуклеотидом, содержащим необходимое количество присоединенных амино-модифицированных линкерных строительных блоков.
Синтез, удаление защитных групп и очистку олигонуклеотидов проводили в соответствии со стандартными процедурами, известными в данной области техники.
Все олигонуклеотиды синтезировали на синтезаторе AKTA oligopilot с применением стандартных фосфоамидитных реакций. Применяли коммерчески доступную твердую подложку и 2'-O-метил-РНК-фосфоамидиты, 2'-фтор, 2'-дезокси-РНК-фосфоамидиты (все со стандартной защитой, ChemGenes, LinkTech) и коммерчески доступный 3'-амино-модификатор TFA-амино-C-6-lcaa-CPG 500 Å (Chemgenes), Fmoc-амино-DMT-C-7-CE-фосфоамидит (GlyC3Am), 3'-амино-модификатор C-3-lcaa-CPG 500 Å (C3Am), Fmoc-амино-DMT-C-3-CED-фосфоамидит (C3Am) и TFA-амино-C-6-CED-фосфоамидит (C6Am) (Chemgenes), 3'-амино-модификатор C7 CPG (C7Am) (Glen Research), ненуклеозидный TFA-аминофосфоамидит (Pip), ненуклеозидную TFA-амино-твердую подложку (PipAm) (AM Chemicals). Пер-ацетилированный галактозамин 8 доступен коммерчески.
Вспомогательные реагенты приобретали у EMP Biotech. Синтез выполняли с применением 0,1 М раствора фосфоамидита в сухом ацетонитриле, и в качестве активатора применяли бензилтиотетразол (БТТ) (0,3 М в ацетонитриле). Время связывания составляло 15 мин. Применяли цикл Cap/OX/Cap или Cap/Thio/Cap (Cap: Ac2O/NMI/лутидин/ацетонитрил, окислитель: 0,1 М I2 в пиридине/H2O). Фосфотиоаты вводили с применением стандартного коммерчески доступного реагента для тиолирования (EDITH, Link technologies). Отщепление DMT осуществляли с помощью обработки 3% дихлоруксусной кислотой в толуоле. После завершения запрограммированных циклов синтеза выполняли промывку диэтиламином (DEA). Все олигонуклеотиды синтезировали в режиме DMT-off.
Присоединение линкерной группы, происходящей из серинола, осуществляли с применением либо (S)-DMT-серинол(TFA)-сукцинат-lcaa-CPG 10, загруженного основанием, либо (S)-DMT-серинол(TFA)фосфоамидита 7 (синтез выполняли, как описано в Hoevelmann et al. (2016)). Трехантенные кластеры GalNAc (ST23/C4XLT) вводили путем последовательного связывания соответствующих утроительных амидитных производных (C4XLT-phos) с последующим добавлением амидита GalNAc (ST23-phos).
Присоединение амино-модифицированных групп (линкеров, происходящих не из серинола) осуществляли путем применения либо соответствующего коммерчески доступного амино-модифицированного строительного блока CPG, либо амидита.
Одиночные цепи отщепляли от CPG с помощью обработки 40% водн. метиламином. Полученный неочищенный олигонуклеотид очищали с помощью ионообменной хроматографии (Resource Q, 6 мл, GE Healthcare) на системе для ВЭЖХ AKTA Pure с применением градиента хлорида натрия. Содержащие продукт фракции объединяли, обессоливали на колонке для гель-проникающей хроматографии (Zetadex, EMP Biotech) и лиофилизировали.
Отдельные одиночные цепи растворяли в концентрации 60 OD/мл в H2O. Оба раствора отдельных олигонуклеотидов добавляли вместе в реакционный сосуд. Для облегчения мониторинга реакции выполняли титрование. Первую цепь добавляли с 25% избытком по сравнению со второй цепью, что определяли по УФ-поглощению при 260 нм. Реакционную смесь нагревали до 80°C в течение 5 минут и затем медленно охлаждали до комнатной температуры. Образование двойной цепи контролировали по образованию ионных пар методом ВЭЖХ с обращенной фазой. На основании УФ-площади остаточной одиночной цепи рассчитывали необходимое количество второй цепи и добавляли его к реакционной смеси. Реакционную смесь снова нагревали до 80°C и медленно охлаждали до комнатной температуры. Эту процедуру повторяли до тех пор, пока не было обнаружено менее 10% остаточной одиночной цепи.
Синтез соединений 2-10
Соединения 2-5 и (S)-DMT-серинол(TFA)-фосфоамидит 7 синтезировали в соответствии с опубликованными в литературе способами (Hoevelmann et al. Chem. Sci., 2016,7, 128-135).
(S)-4-(3-(бис(4-метоксифенил)(фенил)метокси)-2-(2,2,2-трифторацетамидо)пропокси)-4-оксобутановая кислота (6).
К раствору 5 в пиридине добавляли сукциновый ангидрид, а затем DMAP. Полученную смесь перемешивали при комнатной температуре в течение ночи. Весь исходный материал был израсходован согласно оценке методом ТСХ. Реакцию концентрировали. Неочищенный материал подвергали хроматографии на силикагеле, используя градиент от 0% до 5% метанола в ДХМ (+ 1% триэтиламина), с получением 1,33 г 6 (выход = 38%). m/z (ESI-): 588,2 (100%), (рассчитано для C30H29F3NO8- [M-H]- 588,6), 1H-ЯМР: (400 МГц, CDCl3) δ [ppm]=7,94 (d, 1H, NH), 7,39-7,36 (m, 2H, СНарил), 7,29-7,25 (m, 7H, СНарил), 6,82-6,79 (m, 4H, СНарил), 4,51-4,47 (m, 1H), 4,31-4,24 (m, 2H), 3,77 (s, 6H, 2xDMTr-OMe), 3,66-3,60 (m, 16H, HNEt3+), 3,26-3,25 (m, 2H), 2,97-2,81 (m, 20H, NEt3), 2,50-2,41 (4H, m), 1,48-1,45 (m, 26H, HNEt3+), 1,24-1,18 (m, 29H, NEt3).
(S)-DMT-серинол(TFA)-сукцинат-Icaa-CPG (10)
(S)-DMT-серинол(TFA)-сукцинат (159 мг, 270 мкмоль) и HBTU (113 мг, 299 мкмоль) растворяли в CH3CN (10 мл). К раствору добавляли диизопропилэтиламин (DIPEA, 94 мкл, 540 мкмоль) и смесь перемешивали в течение 2 минут с последующим добавлением нативного амино-Icaa-CPG (500 A, 3 г, содержание амина: 136 мкмоль/г). Суспензию осторожно встряхивали при комнатной температуре на шейкере, имитирующем движение запястья, в течение 16 ч, затем фильтровали и промывали ДХМ и EtOH. Твердую подложку высушивали в вакууме в течение 2 ч. Не вступившие в реакцию амины на носителе кэппировали с помощью перемешивания с уксусным ангидридом/лутидином/N-метилимидазолом при комнатной температуре. Промывку подложки повторяли, как описано выше. Твердое вещество высушивали в вакууме с получением твердой подложки 10 (3 г, загрузка 26 мкмоль/г).
GalNAc синтон (9)
Синтез GalNAc синтона 9 выполняли, как описано в Nair et al. J. Am. Chem. Soc., 2014, 136 (49), pp 16958-16961, с 46% выходом в течение двух этапов.
Характеристические данные соответствуют опубликованным данным.
Синтез олигонуклеотидов
Все одноцепочечные олигонуклеотиды синтезировали в соответствии с условиями реакции, описанными выше и на Фигуре 13 и 14, и представили в таблицах 10 и 11.
Все готовые одноцепочечные продукты анализировали с помощью АХ-ВЭЖХ для подтверждения их чистоты. Чистота представлена в % FLP (% полноразмерного продукта), который представляет собой процент УФ-площади под сигналом заданного продукта на УФ-кривой анализа готового продукта методом АХ-ВЭЖХ. Идентичность соответствующих одноцепочечных продуктов (немодифицированных, амино-модифицированных предшественников или GalNAc-конъюгированных олигонуклеотидов) подтверждали с помощью анализа методом ЖХ/МС.
Таблица 10: Одноцепочечные неконъюгированные олигонуклеотиды
5'1×NH2 означает положение (5'-конец) и количество (1×NH2) свободных аминогрупп, происходящих из серинола, которые доступны для конъюгации. Например, 1×3'NH2 на A0264 означает, что имеется свободная аминогруппа, которая может реагировать с GalNAc синтоном 9 на 3'-конце цепи A0264. 3'5'1×NH2 означает, что имеется одна свободная аминогруппа, происходящая из серинола, которая может реагировать с GalNAc линкером 9 на 3'-конце и 5'-конце цепи.
Таблица 11: Одноцепочечные олигонуклеотиды с 5' и 3' модификациями
Аналогичным образом, 3'5'1×NH2 относится к положению (3' и 5'-конец) и количеству (1×NH2 каждая) свободных аминогрупп, которые доступны для конъюгации. Например, 3'5'1×NH2 на A0561 означает наличие 2 свободных аминогрупп (1 на 3'-конце и 1 на 5'-конце), которые могут реагировать с GalNAc синтоном 9 на 3'-конце цепи A0561.
Синтез некоторых конъюгатов согласно настоящему изобретению и эталонных конъюгатов 1-2
Конъюгацию GalNAc синтона (9) осуществляли путем связывания с функциональной аминогруппой серинола соответствующей олигонуклеотидной цепи 11 с применением связывающего реагента для образования пептидной связи. Таким образом, соответствующую амино-модифицированную молекулу-предшественника 11 растворяли в H2O (500 OD/мл) и добавляли ДМСО (ДМСО/H2O, 2/1, об./об.), а затем DIPEA (2,5% от общего объема). В отдельном реакционном сосуде выполняли предварительную активацию GalN(Ac4)-C4-кислоты (9) с помощью взаимодействия 2 экв. (на функциональную аминогруппу в амино-модифицированном предшественнике олигонуклеотида 11) компонента карбоновой кислоты с 2 экв. HBTU в присутствии 8 экв. DIPEA в ДМСО. Через 2 мин предварительно активированное соединение 9 добавляли к раствору соответствующей амино-модифицированной молекулы-предшественника. Через 30 мин ход реакции контролировали с помощью ЖХ/МС или АХ-ВЭЖХ. После завершения реакции конъюгирования неочищенный продукт осаждали добавлением 10× iPrOH и 0,1× 2 М NaCl и собирали с помощью центрифугирования и декантации. Чтобы высвободить ацетилированные гидроксильные группы в группах GalNAc, полученный осадок растворяли в 40% MeNH2 (1 мл на 500 OD) и через 15 минут при комнатной температуре разбавляли в H2O (1:10), а в завершение повторно очищали с помощью анионного обмена и гель-проникающей хроматографии и лиофилизировали с получением готового продукта 12 (таблица 12).
Таблица 12: Одноцепочечные GalNAc-конъюгированные олигонуклеотиды
Синтез некоторых конъюгатов согласно настоящему изобретению
Конъюгацию GalNAc синтона (9) осуществляли путем связывания с функциональной аминогруппой соответствующей олигонуклеотидной цепи 14 с применением связывающего реагента для образования пептидной связи. Таким образом, соответствующую амино-модифицированную молекулу-предшественника 14 растворяли в H2O (500 OD/мл) и добавляли ДМСО (ДМСО/H2O, 2/1, об./об.), а затем DIPEA (2,5% от общего объема). В отдельном реакционном сосуде выполняли предварительную активацию GalN(Ac4)-C4-кислоты (9) с помощью взаимодействия 2 экв. (на функциональную аминогруппу в амино-модифицированном предшественнике олигонуклеотида 14) компонента карбоновой кислоты с 2 экв. HBTU в присутствии 8 экв. DIPEA в ДМСО. Через 2 мин предварительно активированное соединение 9 добавляли к раствору соответствующей амино-модифицированной молекулы-предшественника. Через 30 мин ход реакции контролировали с помощью ЖХ/МС или АХ-ВЭЖХ. После завершения реакции конъюгирования неочищенный продукт осаждали добавлением 10× iPrOH и 0,1× 2 М NaCl и собирали с помощью центрифугирования и декантации. Чтобы высвободить ацетилированные гидроксильные группы в группах GalNAc, полученный осадок растворяли в 40% MeNH2 (1 мл на 500 OD) и через 15 минут при комнатной температуре разбавляли в H2O (1:10), а в завершение повторно очищали с помощью анионного обмена и гель-проникающей хроматографии и лиофилизировали с получением готового продукта 15 (таблица 13).
Таблица 13: Одноцепочечные GalNAc-конъюгированные олигонуклеотиды
Образование двойной цепи
Двойные цепи получали в соответствии со способами, описанными выше.
Чистота двойной цепи представлена в % двойной цепи, который представляет собой процент УФ-площади под сигналом заданного продукта на УФ-кривой анализа методом ИП-ОФ-ВЭЖХ (таблица 14).
Таблица 14: Конъюгаты нуклеиновых кислот
Последовательности
Обозначения модификаций для следующих последовательностей:
f обозначает 2'-фтор-2'-дезоксирибонуклеотид или 2'-фторрибонуклеотид (термины являются взаимозаменяемыми)
m обозначает 2'-O-метилрибонуклеотид
(ps) обозначает фосфотиоатную связь
FAM=6-карбоксифлуоресцеин
BHQ = тушитель «Черная дыра» 1
YY = Yakima Yellow
Определения
Ser(GN) представляет собой строительный блок GalNAc-C4, присоединенный к линкерной группе, происходящей из серинола:
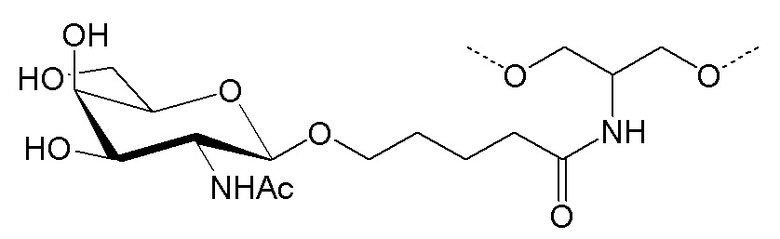
где O--- представляет собой связь между атомом кислорода и, например, H, фосфодиэфирную связь или фосфотиоатную связь.
GN представляет собой:

C4XLT (также известный как ST41) представляет собой:
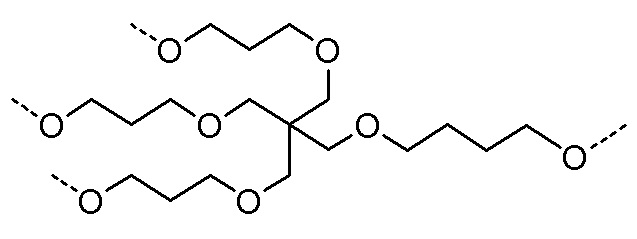
ST23 представляет собой:
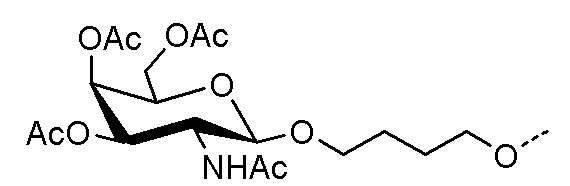
Синтез фосфоамидитных производных C4XLT (C4XLT-phos), а также ST23 (ST23-phos) можно выполнять, как описано в WO2017/174657.
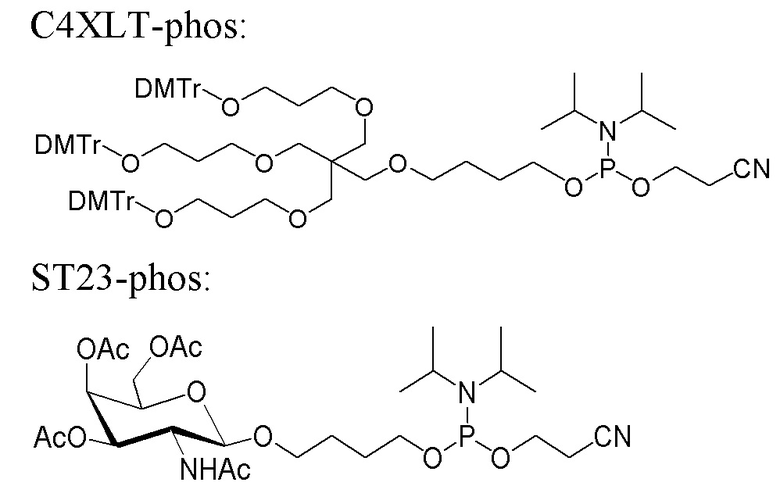
где G=H (до конъюгации) или G=GN (после конъюгации).
Конъюгат 1
Антисмысловая цепь-STS16001AL33 (SEQ ID NO: 127)
5' mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU (ps) Ser(GN) 3'
Смысловая цепь-STS16001BL20 (SEQ ID NO: 128)
5' Ser(GN) (ps) fA mA fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA 3'
Конъюгат 2
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU
Смысловая цепь-STS16001BV1L42 (SEQ ID NO: 130)
Ser(GN) (ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA (ps) Ser(GN)
Конъюгат 3
Антисмысловая цепь-STS16001AL33 (SEQ ID NO: 127)
5' mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU (ps) Ser(GN) 3'
Смысловая цепь-STS16001BV1L42 (SEQ ID NO: 130)
5' Ser(GN) (ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA (ps) Ser(GN) 3'
Конъюгат 4
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU
Смысловая цепь-STS16001BV1L75 (SEQ ID NO: 142)
5' Ser(GN) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA Ser(GN) 3'
Конъюгат 5
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU
Смысловая цепь-STS16001BV16L42 (SEQ ID NO: 143)
5' Ser(GN) (ps) fA mA fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU mA fA (ps) Ser(GN) 3'
Конъюгат 6
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU
Смысловая цепь-STS16001BV20L75 (SEQ ID NO: 144)
5' Ser(GN) fA mA fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU mA fA Ser(GN) 3'
Конъюгат 7
Антисмысловая цепь-(SEQ ID NO: 129)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU
Смысловая цепь-STS16001BV1L94 (SEQ ID NO: 145)
5' Ser(GN) (ps) Ser(GN) (ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA (ps) Ser(GN) (ps) Ser(GN) 3'
Конъюгат 8
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
5' mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU 3'
Смысловая цепь-STS16001V1BL96 (SEQ ID NO: 146)
5' C6Am(GN) (ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA (ps) C7Am(GN) 3'
Конъюгат 9
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
5' mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU 3'
Смысловая цепь-STS16001V1BL97 (SEQ ID NO: 147)
5' GlyC3Am(GN) (ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA (ps) GlyC3Am(GN) 3'
Конъюгат 10
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
5' mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU 3'
Смысловая цепь (SEQ ID NO: 148)
5' PipAm(GN) (ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA (ps) PipAm(GN) 3'
Конъюгат 11
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
5' mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU 3'
Смысловая цепь-STS16001V1BL88 (SEQ ID NO: 149)
5' C3Am(GN) (ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA (ps) C3Am(GN) 3'
Конъюгат 12
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
5' mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU 3'
Смысловая цепь-STS16001V1BL87 (SEQ ID NO: 150)
5' C6Am(GN) (ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA (ps) GlyC3Am(GN) 3'
Конъюгат 15
Антисмысловая цепь (SEQ ID NO: 151)
mU (ps) fC (ps) mU fU mC fU mU fA mA fA mC fU mG fA mG fU mU (ps) fU (ps) mC
Смысловая цепь (SEQ ID NO: 152)
Ser(GN) (ps) fG (ps) mA (ps) fA mA fC mU fC mA fG mU fU mU fA mA fG mA fA (ps) mG (ps) fA (ps) Ser(GN)
Конъюгат 16
Антисмысловая цепь (SEQ ID NO: 153)
mA (ps) fU (ps) mG fU mA fG mC fC mG fA mG fG mA fU mC fU mU (ps) fC (ps) mU
Смысловая цепь (SEQ ID NO: 154)
Ser(GN) (ps) fA (ps) mG (ps) fA mA fG mA fU mC fC mU fC mG fG mC fU mA fC (ps) mA (ps) fU (ps) Ser(GN)
Конъюгат 18
Антисмысловая цепь (SEQ ID NO: 155)
mA (ps) fA (ps) mC fC mA fG mA fA mG fA mA fG mC fA mG fG mU (ps) fG (ps) mA
Смысловая цепь (SEQ ID NO: 156)
Ser(GN) (ps) fU (ps) mC (ps) fA mC fC mU fG mC fU mU fC mU fU mC fU mG fG (ps) mU (ps) fU (ps) Ser(GN)
Конъюгат 19
Антисмысловая цепь (SEQ ID NO: 135)
mA (ps) fU (ps) mA fA mC fU mC fU mG fU mC fC mA fU mU fA mC (ps) fC (ps) mG
Смысловая цепь (SEQ ID NO: 136)
Ser(GN) (ps) fC (ps) mG (ps) fG mU fA mA fU mG fG mA fC mA fG mA fG mU fU (ps) mA (ps) fU (ps) Ser(GN)
Эталонный конъюгат 1
Антисмысловая цепь-STS16001A (SEQ ID NO: 129)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU
Смысловая цепь-STS16001BL20 (SEQ ID NO: 128)
Ser(GN) (ps) fA mA fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA
Эталонный конъюгат 2
Антисмысловая цепь-STS16001AL33 (SEQ ID NO: 127)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU (ps) Ser(GN)
Смысловая цепь-STS16001BV1 (SEQ ID NO: 157)
fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA
Эталонный конъюгат 3 - “Luc”
Антисмысловая цепь-STS18001A (A0130, SEQ ID NO: 132)
mU (ps) fC (ps) mG fA mA fG mU fA mU fU mC fC mG fC mG fU mA (ps) fC (ps) mG
Смысловая цепь-STS18001BL4 (A0131, SEQ ID NO: 133)
[(ST23) (ps)]3 C4XLT (ps) fC mG fU mA fC mG fC mG fG mA fA mU fA mC fU mU fC (ps) mG (ps) fA
Эталонный конъюгат 4
Антисмысловая цепь-STS16001AL33 (SEQ ID NO: 127)
mU (ps) fU (ps) mA fU mA fG mA fG mC fA mA fG mA fA mC fA mC fU mG (ps) fU (ps) mU
Смысловая цепь-STS16001BL4 (SEQ ID NO: 134)
5'[(ST23) (ps)]3 C4XLT(ps) fA (ps) mA (ps) fC mA fG mU fG mU fU mC fU mU fG mC fU mC fU mA fU (ps) mA (ps) fA
Эталонный конъюгат 5 - “Ctr”
Антисмысловая цепь (SEQ ID NO: 138)
mC (ps) fU (ps) mU fA mC fU mC fU mC fG mC fC mC fA mA fG mC (ps) fG (ps) mA
Смысловая цепь (SEQ ID NO: 139)
[(ST23) (ps)]3 (C6XLT) (ps) fU mC fG mC fU mU fG mG fG mC fG mA fG mA fG mU fA (ps) mA (ps) fG
Эталонный конъюгат 6
Антисмысловая цепь (SEQ ID NO: 151)
mU (ps) fC (ps) mU fU mC fU mU fA mA fA mC fU mG fA mG fU mU (ps) fU (ps) mC
Смысловая цепь (SEQ ID NO: 158)
[ST23 (ps)]3 ltrb (ps) fG mA fA mA fC mU fC mA fG mU fU mU fA mA fG mA fA (ps) mG (ps) fA
Эталонный конъюгат 7
Антисмысловая цепь (SEQ ID NO: 153)
mA (ps) fU (ps) mG fU mA fG mC fC mG fA mG fG mA fU mC fU mU (ps) fC (ps) mU
Смысловая цепь (SEQ ID NO: 159)
[ST23 (ps)]3 ltrb (ps) fA mG fA mA fG mA fU mC fC mU fC mG fG mC fU mA fC (ps) mA (ps) fU
Эталонный конъюгат 8
Антисмысловая цепь (SEQ ID NO: 160)
mU (ps) fA (ps) mC fC mA fG mA fA mG fA mA fG mC fA mG fG mU (ps) fG (ps) mA
Смысловая цепь (SEQ ID NO: 161)
[ST23 (ps)]3 ST41 (ps)fU mC fA mC fC mU fG mC fU mU fC mU fU mC fU mG fG (ps) mU (ps) fA
Эталонный конъюгат 9
Антисмысловая цепь (SEQ ID NO: 135)
mA (ps) fU (ps) mA fA mC fU mC fU mG fU mC fC mA fU mU fA mC (ps) fC (ps) mG
Смысловая цепь (SEQ ID NO: 162)
[ST23 (ps)]3 C6XLT (ps) fC mG fG mU fA mA fU mG fG mA fC mA fG mA fG mU fU (ps) mA (ps) fU
Пример 12 - Определение выключения TTR в условиях in vitro для различных GalNAc-конъюгатов миРНК TTR
Конъюгаты 1-3
Первичные гепатоциты мыши высевали в предварительно покрытые коллагеном 96-луночные планшеты (Thermo Fisher Scientific, каталожный номер A1142803) при плотности клеток 30000 клеток на лунку и обрабатывали миРНК-конъюгатами в концентрациях, варьирующихся от 10 нМ до 0,0001 нМ. Через 24 часа после обработки клетки лизировали и РНК экстрагировали с помощью препаративных наборов InviTrap® RNA Cell HTS 96/C24×96 (Stratec, каталожный номер 7061300400) в соответствии с протоколом производителя. Уровни транскриптов мРНК TTR и конститутивного гена (PtenII) определяли количественно с помощью анализа TaqMan.
Экспрессия гена-мишени в первичных мышиных гепатоцитах через 24 ч после обработки 0,01 нМ, 0,1 нМ, 0,5 нМ, 1 нМ и 10 нМ конъюгатов согласно настоящему изобретению, конъюгатов 1-3, показала, что экспрессия гена-мишени снижается с увеличением дозы конъюгата по сравнению с отрицательными контролями (см. столбик «UT» и эталонный конъюгат 3), как показано на Фигуре 15. Это свидетельствует о том, что первая цепь связывается с геном-мишенью, что снижает экспрессию гена. На Фигуре 15 также показаны уровни экспрессии гена-мишени для эталонных конъюгатов 1 и 2, которые действуют как конъюгаты для сравнения. Как следует из сопоставления данных, представленных на Фигуре 15А и 15С, а также 15В и 15С, конъюгаты согласно настоящему изобретению (конъюгаты 1-3) снижают экспрессию гена-мишени по сравнению с эталонными конъюгатами 1 и 2. Наиболее эффективным конъюгатом при 0,01 нМ, по-видимому, является конъюгат 2. Наиболее эффективным конъюгатом при 0,1 нМ, 0,5 нМ, 1 нМ и 10 нМ, по-видимому, является конъюгат 3.
Конъюгаты 4-7
Применяли способ, описанный выше под заголовком «Эксперименты в условиях in vitro» в разделе «Общие методы».
Экспрессия гена-мишени в первичных мышиных гепатоцитах через 24 ч после обработки 0,01 нМ, 0,1 нМ, 0,5 нМ, 1 нМ и 10 нМ конъюгатов согласно настоящему изобретению, конъюгатов 4-7, показала, что экспрессия гена-мишени снижается с увеличением дозы конъюгата по сравнению с отрицательными контролями (см. столбик «UT» и Luc [эталонный конъюгат 3]), как показано на Фигуре 31. Это свидетельствует о том, что первая цепь связывается с геном-мишенью, снижая экспрессию гена.
Данные, полученные в условиях in vitro, показывают, что в случае одной или двух линкерных групп, происходящих из серинола, обеспеченных на 5' и 3'-концах смысловой цепи в конъюгатах 4-7, количество фосфотиоатных (PS) связей между концевым нуклеотидом и линкером, и/или между тремя концевыми нуклеотидами в смысловой цепи можно изменять, сохраняя эффективность снижения экспрессии гена-мишени.
Конъюгаты 8-12 и 19
Применяли способ, описанный выше под заголовком «Эксперименты в условиях in vitro» в разделе «Общие методы».
Экспрессия гена-мишени в первичных мышиных гепатоцитах через 24 ч после обработки 0,01 нМ, 0,1 нМ, 0,5 нМ, 1 нМ и 10 нМ конъюгатов согласно настоящему изобретению, конъюгатов 8-12, показала, что экспрессия гена-мишени снижается с увеличением дозы конъюгата по сравнению с отрицательными контролями (см. столбик «UT» и Luc [эталонный конъюгат 3]), как показано на Фигуре 32. Это свидетельствует о том, что первая цепь связывается с геном-мишенью, что снижает экспрессию гена. В частности, конъюгаты 8, 9, 10 и 11, по-видимому, сопоставимы с конъюгатом 2 или превосходят его, который, как показано ранее, был наиболее эффективным конъюгатом при 0,01 нМ.
Также было показано, что конъюгат 19 снижает экспрессию гена-мишени по сравнению с отрицательными контролями (см. столбик «UT» и Ctr, который представляет собой ненацеливающую миРНК и также называется эталонным конъюгатом 5), как показано на Фигуре 33. Это свидетельствует о том, что первая цепь связывается с геном-мишенью, что снижает экспрессию гена.
Данные, полученные в условиях in vitro, для конъюгатов 8-12 и 19, показывают, что ряд линкеров, которые отличаются по структуре и которые конъюгированы на обоих концах смысловой цепи, эффективно снижают экспрессию гена-мишени. Конъюгаты 8-12 и 19 снижают экспрессию гена-мишени более эффективно, чем «Luc», который представляет собой эталонный конъюгат 3 (для конъюгатов 8-12), «Ctr», который представляет собой эталонный конъюгат 5 (для конъюгата 19), и необработанный контроль.
Пример 13 - Временная динамика сывороточных Ttr, Aldh2 и Tmprss6 у мышей в условиях in vivo
Конъюгаты 1-3
Мышей c57BL/6 обрабатывали путем подкожного введения 1 мг/кг миРНК-конъюгатов в 0 день. Образцы сыворотки отбирали на 7, 14 и 27 день путем сбора крови из орбитального синуса и хранили при -20°C до анализа. Количественное определение TTR в сыворотке проводили с помощью ИФА для мышиного преальбумина (ALPCO, 41-PALMS/партия 22, 2008003B) в соответствии с протоколом производителя (разведение образца 1:8000 или 1:800).
Результаты для временной динамики сывороточного TTR у мышей c57BL/6 с n=4 на 7, 14 и 27 день после обработки путем подкожного введения 1 мг/кг конъюгатов 1-3, эталонных конъюгатов 1, 2 и 4 и у индивидуумов с имитированной обработкой (ФСБ) показаны на Фигуре 16. Данные на Фигуре 16 свидетельствуют о том, что конъюгаты согласно настоящему изобретению особенно эффективны в уменьшении экспрессии гена-мишени по сравнению с отрицательным контролем (ФСБ) и эталонными конъюгатами 1, 2 и, в частности, эталонным конъюгатом 4. Конъюгаты 2 и 3 также более эффективны, чем эталонные конъюгаты 1, 2 и 4. Наиболее эффективным конъюгатом является конъюгат 2. Таким образом, можно ожидать, что уровень дозирования конъюгата 2 может быть приблизительно в три раза ниже, чтобы достичь сходного первоначального выключения, и также приведет к большей продолжительности выключения по сравнению с эталонным конъюгатом 4.
Более конкретно, у мышей дикого типа конъюгат 2 приводил к 3-кратному снижению уровня целевого белка в сыворотке на седьмой день и к 4-кратному снижению уровня целевого белка в сыворотке на 27 день по сравнению с эталонным конъюгатом 4 в эквимолярной дозе. Кроме того, конъюгат 2 приводил к 85% уменьшению уровня целевого белка в сыворотке на 27 день после однократной инъекции по сравнению с 36% уменьшением при применении эквимолярного количества эталонного конъюгата 4.
Конъюгаты 15-18
Применяли способ, описанный выше под заголовком «Эксперименты в условиях in vivo» в разделе «Общие методы».
Результаты для временной динамики сывороточного Aldh2 в когортах мышей c57BL/6 с n=6 на 14, 28 и 42 день после обработки с помощью подкожного введения 1 мг/кг конъюгатов 15 и 16, эталонных конъюгатов 6 и 7 и у индивидуумов с имитированной обработкой (ФСБ) показаны на Фигуре 34 и 35. Данные на Фигуре 34 и 35 свидетельствуют о том, конъюгаты согласно настоящему изобретению особенно эффективно уменьшают экспрессию гена-мишени по сравнению с отрицательным контролем (ФСБ) и эталонными конъюгатами 6 и 7, соответственно.
Результаты для временной динамики сывороточного Tmprss6 в когортах мышей c57BL/6 с n=6 на 14, 28 и 42 день после обработки с помощью подкожного введения 1 мг/кг конъюгата 18, эталонного конъюгата 8 и у индивидуумов с имитированной обработкой (ФСБ) показаны на Фигуре 36. Данные на Фигуре 36 свидетельствуют о том, что конъюгаты согласно настоящему изобретению особенно эффективно уменьшают экспрессию гена-мишени по сравнению с отрицательным контролем (ФСБ) и эталонным конъюгатом 8.
В целом, данные, полученные в условиях in vivo, показывают, что различные примерные линкеры, которые конъюгированы на обоих концах второй цепи, эффективно снижают экспрессию гена-мишени в условиях in vivo. Расположение линкера улучшает эффективность конъюгатов в условиях in vivo по сравнению с контрольным трехантенным GalNAc-линкером на 5'-конце второй цепи (эталонные конъюгаты 6, 7 и 8).
Пример 14 - Исследования стабильности в сыворотке
Применяли способ, описанный выше под заголовком «Анализ стабильности в тритосомах» в разделе «Общие методы».
На Фигуре 37 показаны результаты исследований стабильности в сыворотке в отношении конъюгатов 2, 4, 5, 6 и 7. На Фигуре 38 показана стабильность в сыворотке конъюгатов 2, 8, 9, 10, 11 и 12.
Все испытанные конъюгаты согласно настоящему изобретению более стабильны в сыворотке по сравнению с контролем.
Каждый из всех протестированных конъюгатов содержит один блок GalNAc-линкер на 5'-конце и другой на 3'-конце второй цепи. МиРНК модифицированы с применением чередующихся 2'-OMe/2'-F и каждая содержит две фосфотиоатные (PS) межнуклеотидные связи на своих 5' и 3'-концевых двух межнуклеотидных связях, если не указано иное.
В конъюгате 4 блоки серинол-GalNAc присоединены за счет фосфодиэфирной связи. В конъюгате 5 блоки серинол-GalNAc конъюгированы за счет PS, тогда как все межнуклеотидные связи во второй цепи являются фосфодиэфирными. В конъюгате 6 вторая цепь не содержит PS. В конъюгате 7 два блока серинол-GalNAc присоединены к каждому концу второй цепи и друг к другу за счет PS-связей на соответствующих концах. В конъюгате 8 для присоединения лиганда применяли C6-амино-модификатор на 5' и C7-амино-модификатор на 3'-конце второй цепи. В конъюгате 9 в качестве линкеров для конъюгации с обоими концами второй цепи применяли Gly-C3-амино-модификаторы, в конъюгате 10 применяли пиперидил-амино-модификаторы, в конъюгате 11 применяли C3-амино-модификаторы и в конъюгате 2 применяли блоки серинол-GalNAc. В конъюгате 2 обе концевые межнуклеотидные связи, а также нуклеотид-серинольные связи представляют собой PS. В конъюгате 12 для присоединения лиганда применяли C6-амино-модификатор на 5' и GlyC3-амино-модификатор на 3'-конце второй цепи. «Ut» обозначает необработанный образец, к которому нормировали другие образцы. «Luc» обозначает нацеленную на люциферазу миРНК (эталонный конъюгат 3), которую применяли в качестве ненацеливающего контроля и которая не уменьшает уровни мРНК-мишени.
Данные показывают, что в случае линкерной группы, происходящей из серинола, обеспеченной на 5' и 3'-концах смысловой цепи, количество фосфотиоатных (PS) связей между концевым нуклеотидом и линкером, и/или между тремя концевыми нуклеотидами в смысловой цепи можно изменять, сохраняя стабильность в сыворотке.
Пример 15
Выключение мРНК LPA в первичных гепатоцитах человека под действием различных указанных L6-GalNAc-конъюгированных миРНК в первичных гепатоцитах человека после опосредуемой рецепторами доставки.
Таблица 15: Дополнительные последовательности конъюгированных нуклеиновых кислот, протестированные в отношении ингибирования экспрессии мРНК LPA. Указаны последовательности и применяемый профиль модификации
Таблица 15: Модификации нуклеотидов обозначены следующими цифрами (столбик 4), 1=2'-F-dU, 2=2'-F-dA, 3=2'-F-dC, 4=2'-F-dG, 5=2'-OMe-rU; 6=2'-OMe-rA; 7=2'-OMe-rC; 8=2'-OMe-rG.
Первичные гепатоциты человека (ThermoFisher) высевали на покрытые коллагеном 96-луночные планшеты по 30000 клеток на лунку (96-луночный формат). GalNAc-L6-конъюгированные миРНК добавляли в указанных концентрациях сразу после посева клеток. Через 24 часа после обработки миРНК общую РНК выделяли с применением набора InviTrap RNA cell HTS (Stratec) для 96-луночного планшета. Уровни экспрессии мРНК LPA определяли с помощью кПЦР в режиме реального времени относительно мРНК APOB в качестве транскрипта конститутивного гена. Значения нормировали к экспрессии мРНК LPA в необработанных клетках. Средние значения и стандартные отклонения нормированных трехкратных значений показаны на Фигуре 39. Значения ИК50 и максимальное ингибирование оценивали путем аппроксимации кривой с применением четырехпараметрической нелинейной регрессии.
Изложение сущности изобретения
1. Нуклеиновая кислота для ингибирования экспрессии LPA в клетке, содержащая по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, при этом указанная первая цепь содержит нуклеотидную последовательность, выбранную из следующих последовательностей: SEQ ID NO: 9, 1, 3, 5, 7, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73 или любой последовательности, раскрытой в настоящей заявке.
2. Нуклеиновая кислота по утверждению 1, в которой указанная вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 10, 2, 4, 6, 8, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 64, 66, 68, 70, 72 или 74 или любую последовательность, раскрытую в настоящей заявке.
3. Нуклеиновая кислота по утверждению 1 или утверждению 2, в которой указанная первая цепь содержит нуклеотидную последовательность SEQ ID NO: 9 или SEQ ID NO: 5.
4. Нуклеиновая кислота по любому из утверждений 1-3, в которой указанная вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 10 или SEQ ID NO: 6.
5. Нуклеиновая кислота по любому из утверждений 1-4, в которой каждая из указанной первой цепи и/или указанной второй цепи содержит 17-35 нуклеотидов в длину.
6. Нуклеиновая кислота по любому из утверждений 1-5, в которой указанная по меньшей мере одна дуплексная область состоит из 19-25 последовательных пар нуклеотидных оснований.
7. Нуклеиновая кислота по любому из предшествующих утверждений, которая
a) имеет тупые концы на обоих концах; или
b) имеет липкий конец на одном конце и тупой конец на другом; или
c) имеет липкий конец на обоих концах.
8. Нуклеиновая кислота по любому из предшествующих утверждений, в которой один или более нуклеотидов на указанной первой и/или второй цепи модифицированы с образованием модифицированных нуклеотидов.
9. Нуклеиновая кислота по утверждению 8, в которой один или более из нуклеотидов в нечетных положениях первой цепи модифицированы.
10. Нуклеиновая кислота по утверждению 9, в которой один или более из нуклеотидов в четных положениях первой цепи модифицированы с применением по меньшей мере второй модификации, при этом указанная по меньшей мере вторая модификация отличается от модификации по утверждению 9.
11. Нуклеиновая кислота по утверждению 10, в которой по меньшей мере один из одного или более модифицированных нуклеотидов в четных положениях является смежным с по меньшей мере одним из одного или более модифицированных нуклеотидов в нечетных положениях.
12. Нуклеиновая кислота по любому из утверждений 9-11, в которой множество нуклеотидов в нечетных положениях являются модифицированными.
13. Нуклеиновая кислота по утверждению 10-12, в которой множество нуклеотидов в четных положениях модифицировано с применением второй модификации.
14. Нуклеиновая кислота по любому из утверждений 8-13, в которой указанная первая цепь содержит смежные нуклеотиды, которые модифицированы с применением общей модификации.
15. Нуклеиновая кислота по любому из утверждений 9-14, в которой указанная первая цепь содержит смежные нуклеотиды, которые модифицированы с применением второй модификации, которая отличается от модификации по утверждению 9.
16. Нуклеиновая кислота по любому из утверждений 9-15, в которой один или более из нуклеотидов в нечетных положениях второй цепи модифицированы с применением модификации, которая отличается от модификации по утверждению 9.
17. Нуклеиновая кислота по любому из утверждений 9-15, в которой один или более из нуклеотидов в четных положениях второй цепи модифицированы с применением модификации по утверждению 9.
18. Нуклеиновая кислота по утверждению 16 или 17, в которой по меньшей мере один из одного или более модифицированных нуклеотидов в четных положениях второй цепи является смежным с одним или более модифицированными нечетными нуклеотидами второй цепи.
19. Нуклеиновая кислота по любому из утверждений 16-18, в которой множество нуклеотидов в нечетных положениях второй цепи модифицированы с применением общей модификации.
20. Нуклеиновая кислота по любому из утверждений 16-19, в которой множество нуклеотидов в четных положениях модифицированы с применением модификации по утверждению 9.
21. Нуклеиновая кислота по любому из утверждений 16-20, в которой множество нуклеотидов в нечетных положениях на второй цепи модифицированы с применением второй модификации, причем указанная вторая модификация отличается от модификации по утверждению 9.
22. Нуклеиновая кислота по любому из утверждений 16-21, в которой указанная вторая цепь содержит смежные нуклеотиды, которые модифицированы с применением общей модификации.
23. Нуклеиновая кислота по любому из утверждений 16-22, в которой указанная вторая цепь содержит смежные нуклеотиды, которые модифицированы с применением второй модификации, которая отличается от модификации по утверждению 9.
24. Нуклеиновая кислота по любому из утверждений 8-23, в которой каждый из нуклеотидов в нечетных положениях в первой цепи и каждый из нуклеотидов в четных положениях во второй цепи модифицирован с применением общей модификации.
25. Нуклеиновая кислота по утверждению 24, в которой каждый из нуклеотидов в четных положениях в первой цепи модифицирован с применением второй модификации, и каждый из нуклеотидов в нечетных положениях во второй цепи модифицирован с применением второй модификации.
26. Нуклеиновая кислота по любому из утверждений 8-25, в которой модифицированные нуклеотиды первой цепи сдвинуты по меньшей мере на один нуклеотид относительно немодифицированных или по-другому модифицированных нуклеотидов второй цепи.
27. Нуклеиновая кислота по любому из утверждений 8-26, в которой указанная модификация и/или модификации по отдельности и независимо выбраны из группы, состоящей из 3'-концевого дезокситимина, 2'-O-метил, 2'-дезокси-модификации, 2'-амино-модификации, 2'-алкил-модификации, морфолино-модификации, модификации фосфоамидатом, модификации 5'-фосфотиоатной группой, модификации 5'-фосфатом или имитатором 5'-фосфата, а также модификации производным холестерина или группой бисдециламида додекановой кислоты.
28. Нуклеиновая кислота по любому из утверждений 8-27, в которой указанная модификация представляет собой любой из закрытого нуклеотида, лишенного азотистого основания нуклеотида или нуклеотида, содержащего неприродное основание.
29. Нуклеиновая кислота по любому из утверждений 8-28, в которой по меньшей мере одна модификация представляет собой 2'-O-метил.
30. Нуклеиновая кислота по любому из утверждений 8-29, в которой по меньшей мере одна модификация представляет собой 2'-F.
31. Нуклеиновая кислота для ингибирования экспрессии LPA в клетке, содержащая по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, при этом указанная первая цепь содержит нуклеотидную последовательность, выбранную из следующих последовательностей: SEQ ID NO: 9, 1, 3, 5, 7, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73, при этом нуклеотиды первой цепи модифицированы с применением первой модификации на нечетных нуклеотидах и модифицированы с применением второй модификации на четных нуклеотидах, и нуклеотиды второй цепи модифицированы с применением третьей модификации на четных нуклеотидах и модифицированы с применением четвертой модификации на нечетных нуклеотидах, при этом по меньшей мере указанная первая модификация отличается от указанной второй модификации, и указанная третья модификация отличается от указанной четвертой модификации.
32. Нуклеиновая кислота по утверждению 31, в которой вторая последовательность содержит нуклеотидную последовательность SEQ ID NO: 10, 2, 4, 6, 8, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 64, 66, 68, 70, 72 или 74.
33. Нуклеиновая кислота по утверждению 31 или 32, в которой указанная четвертая модификация и указанная вторая модификация являются одинаковыми.
34. Нуклеиновая кислота по любому из утверждений 31-33, в которой указанная первая модификация и указанная третья модификация являются одинаковыми.
35. Нуклеиновая кислота по любому из утверждений 31-34, в которой указанная первая модификация представляет собой 2'-O-Me, и указанная вторая модификация представляет собой 2'-F.
36. Нуклеиновая кислота по любому из утверждений 31-35, в которой указанная первая цепь содержит нуклеотидную последовательность SEQ ID NO: 9 или SEQ ID NO: 5, и указанная вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 10 или SEQ ID NO: 6
37. Нуклеиновая кислота по любому из утверждений 31-36, содержащая последовательность и модификации, представленные ниже:
где конкретные модификации обозначены номерами
1=2'-F-dU,
2=2'-F-dA,
3=2'-F-dC,
4=2'-F-dG,
5=2'-OMe-rU;
6=2'-OMe-rA;
7=2'-OMe-rC;
8=2'-OMe-rG.
38. Нуклеиновая кислота по любому из утверждений 1-37, конъюгированная с лигандом.
39. Нуклеиновая кислота по любому из утверждений 1-38, содержащая фосфотиоатную связь между одним, двумя или тремя концевыми 3'-нуклеотидами и/или 5'-нуклеотидами первой и/или второй цепей.
40. Нуклеиновая кислота по любому из утверждений 1-39, содержащая две фосфотиоатные связи между каждым из трех концевых 3'- и между каждым из трех концевых 5'-нуклеотидов на первой цепи и две фосфотиоатные связи между тремя концевыми нуклеотидами 3'-конца второй цепи.
41. Нуклеиновая кислота для ингибирования экспрессии LPA в клетке, содержащая по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, при этом указанная первая цепь содержит нуклеотидную последовательность, выбранную из следующих последовательностей: SEQ ID NO: 9, 1, 3, 5, 7, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73, и при этом указанная нуклеиновая кислота конъюгирована с лигандом.
42. Нуклеиновая кислота по утверждению 41, в которой указанный лиганд содержит (i) одну или более групп N-ацетилгалактозамина (GalNac) и их производных, и (ii) линкер, причем с помощью указанного линкера конъюгируют группы GalNac с нуклеиновой кислотой по утверждению 41.
43. Нуклеиновая кислота по любому из утверждений 41 или 42, в которой линкер может иметь двухвалентную или трехвалентную, или четырехвалентную разветвленную структуру.
44. Нуклеиновая кислота по любому из утверждений 41-43, в которой указанные нуклеотиды модифицированы по любому из предшествующих утверждений.
45. Нуклеиновая кислота по любому из предшествующих утверждений, которая конъюгирована с лигандом, содержащим формулу I:
[S-X1-P-X2]3-A-X3-, (I)
в которой:
S представляет собой сахарид, причем указанный сахарид представляет собой N-ацетилгалактозамин;
X1 представляет собой C3-C6 алкилен или (-CH2-CH2-O)m(-CH2)2-, где m равно 1, 2 или 3;
P представляет собой фосфат или модифицированный фосфат (предпочтительно тиофосфат);
X2 представляет собой алкилен или простой алкиленовый эфир формулы (-CH2)n-O-CH2-, где n=1-6;
А представляет собой блок разветвления;
X3 представляет собой мостиковый блок;
при этом нуклеиновая кислота по любому из утверждений 1-40 конъюгирована с X3 за счет фосфата или модифицированного фосфата (предпочтительно тиофосфата).
46. Конъюгированная нуклеиновая кислота, имеющая одну из следующих структур
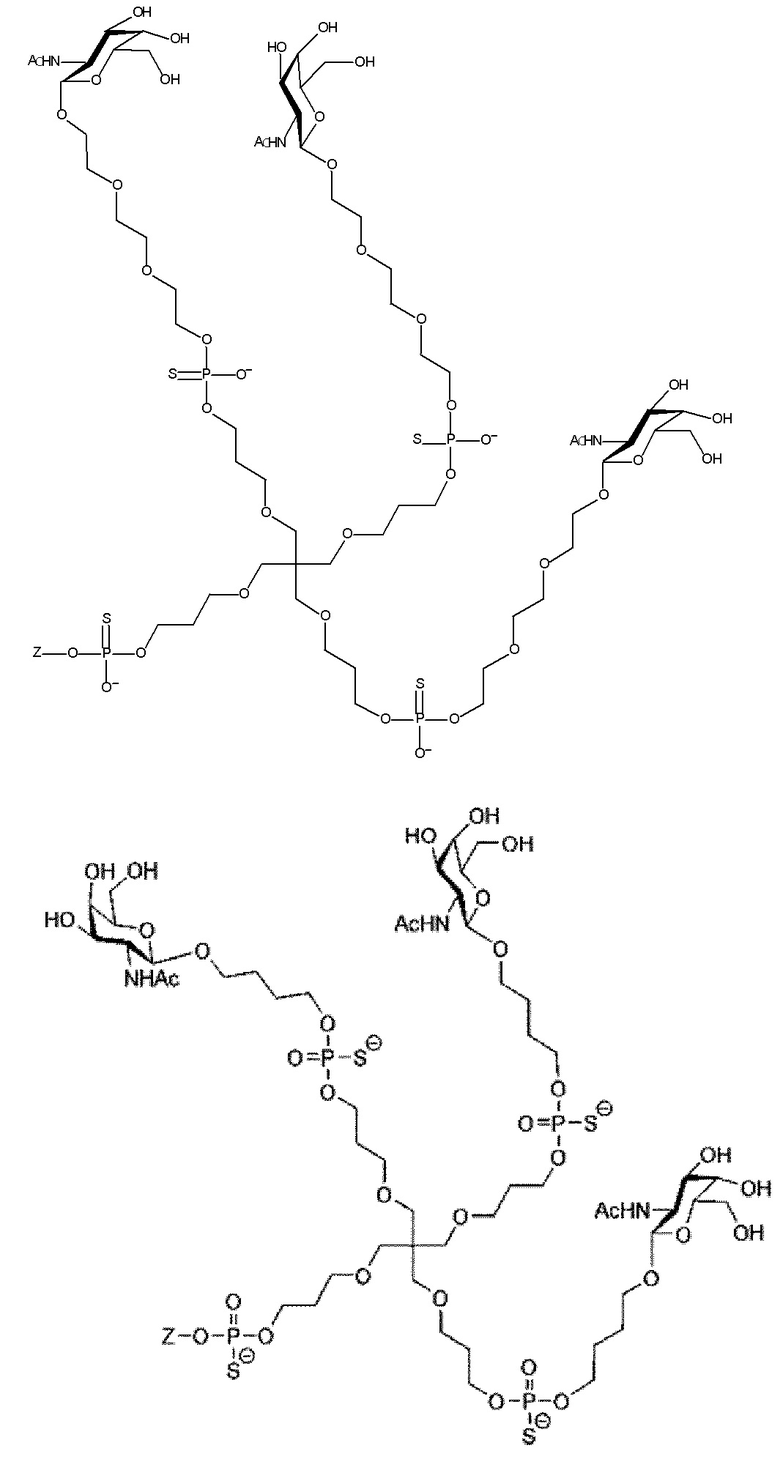
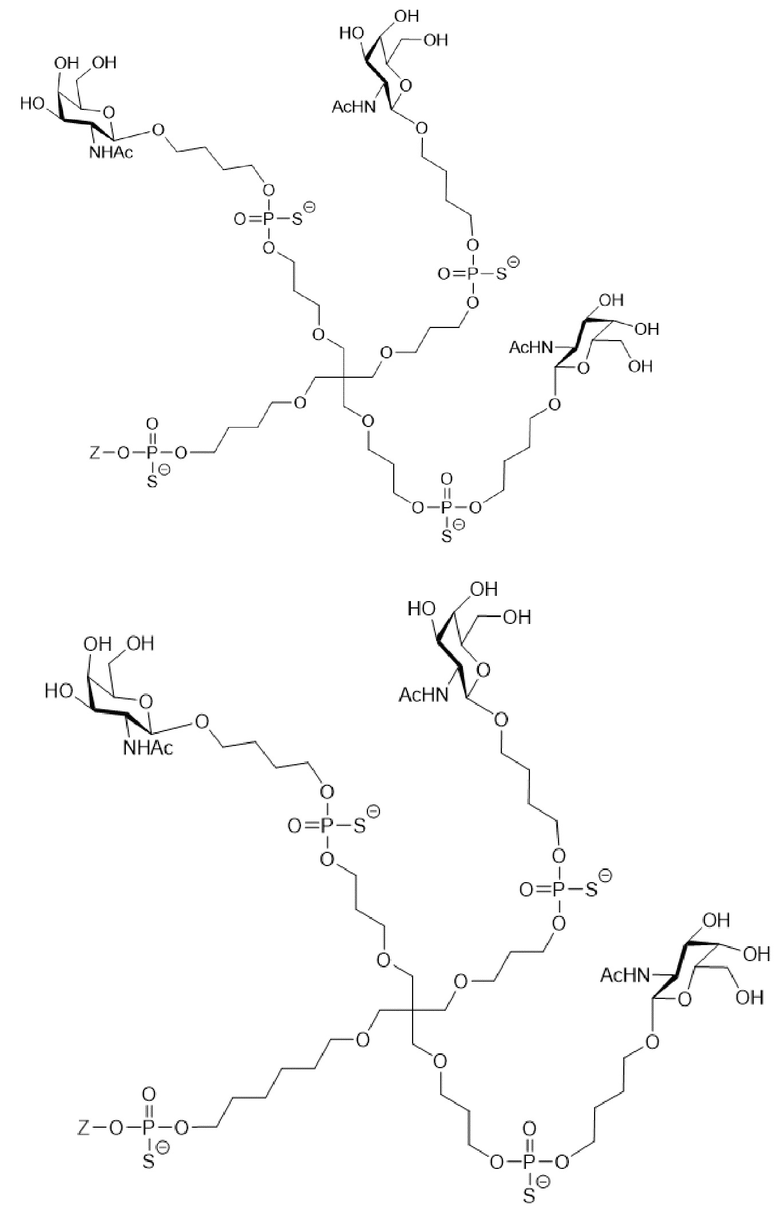
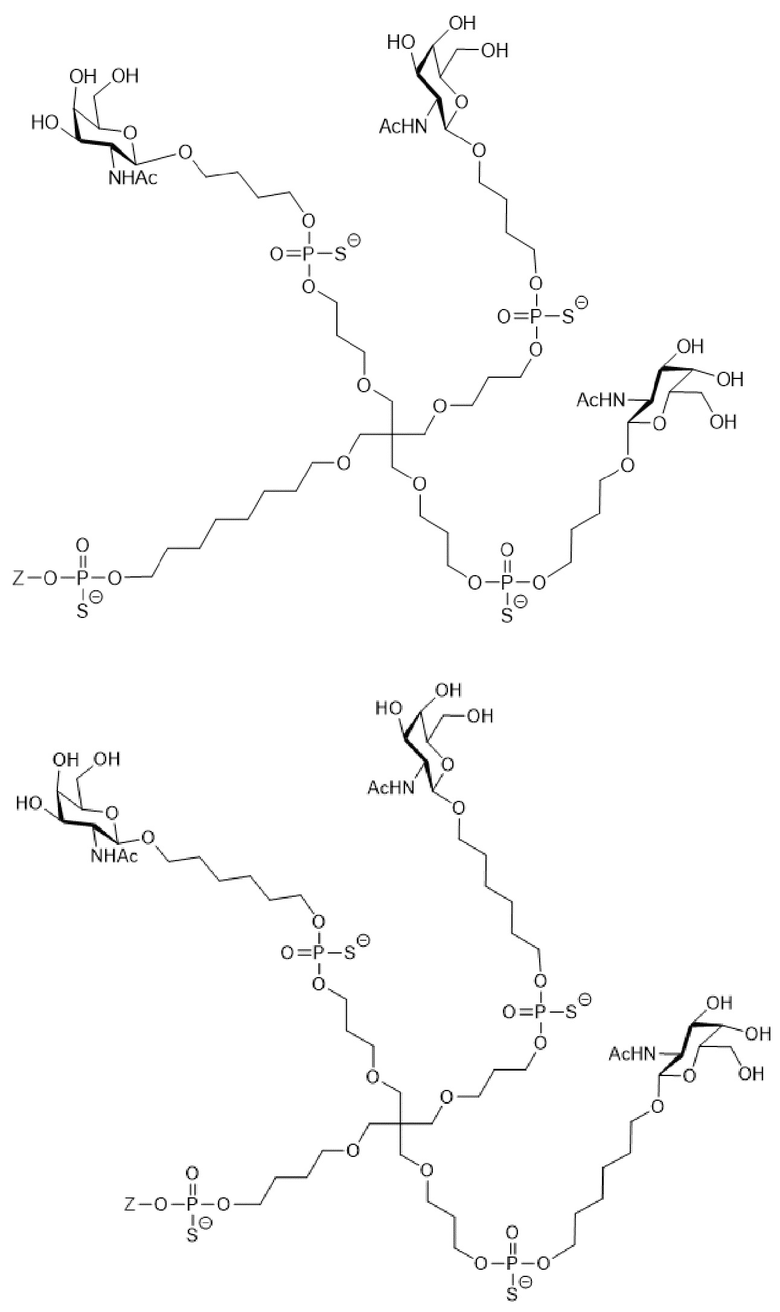
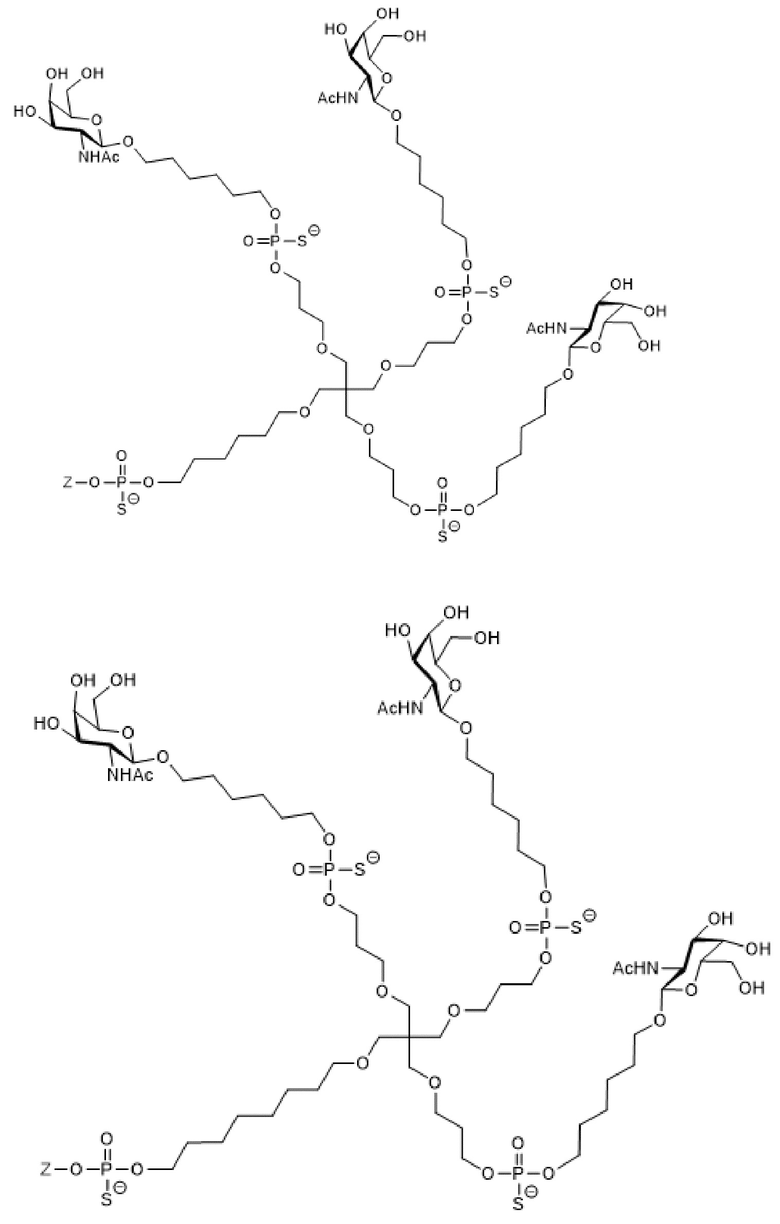
где Z представляет собой нуклеиновую кислоту по любому из утверждений 1-40.
47. Нуклеиновая кислота по любому из утверждений 1-40, которая конъюгирована с лигандом следующей структуры
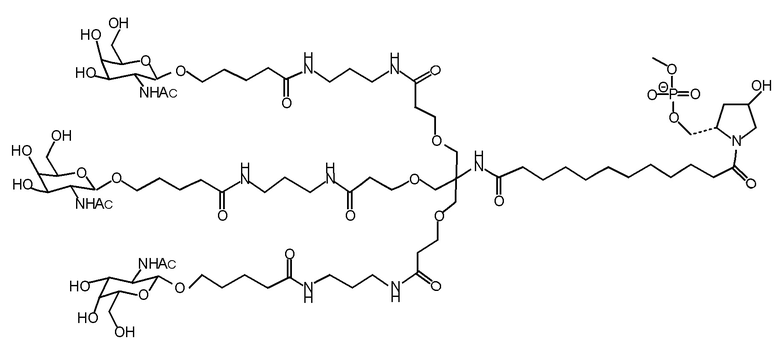
48. Нуклеиновая кислота или конъюгированная нуклеиновая кислота по любому из предшествующих утверждений, в которых указанный дуплекс содержит отдельные цепи.
49. Нуклеиновая кислота или конъюгированная нуклеиновая кислота по любому из предшествующих утверждений, в которых указанный дуплекс содержит одиночную цепь, содержащую первую цепь и вторую цепь.
50. Композиция, содержащая нуклеиновую кислоту или конъюгированную нуклеиновую кислоту по любому из предшествующих утверждений и физиологически приемлемое вспомогательное вещество.
51. Нуклеиновая кислота или конъюгированная нуклеиновая кислота по любому из предшествующих утверждений для применения при предотвращении или лечении, или уменьшении риска заболевания или патологии.
52. Применение нуклеиновой кислоты или конъюгированной нуклеиновой кислоты по любому из предшествующих утверждений при изготовлении лекарственного средства для предотвращения или лечения заболевания, нарушения или синдрома.
53. Способ предотвращения или лечения заболевания, нарушения или синдрома, включающий введение композиции, содержащей нуклеиновую кислоту или конъюгированную нуклеиновую кислоту по любому из предшествующих утверждений, индивидууму, нуждающемуся в лечении.
54. Способ по утверждению 53, в котором указанную нуклеиновую кислоту или конъюгированную нуклеиновую кислоту вводят субъекту подкожно, внутривенно или с применением любых других путей применения, таких как пероральный, ректальный или внутрибрюшинный.
55. Применение или способ по утверждениям 52-54, в которых указанное заболевание или патология представляет собой сердечно-сосудистое заболевание, инсульт, атеросклероз, тромбоз или сердечно-сосудистые заболевания, такие как ишемическая болезнь сердца или стеноз аорты, и любое другое заболевание или патологию, ассоциированные с повышенными уровнями частиц, содержащих Lp(a).
56. Применение или способ по утверждению 55, в которых указанное сердечно-сосудистое заболевание представляет собой инсульт, атеросклероз, тромбоз, ишемическую болезнь сердца или стеноз аорты и любое другое заболевание или патологию, ассоциированные с повышенными уровнями частиц, содержащих Lp(a).
57. Способ получения нуклеиновой кислоты или конъюгированной нуклеиновой кислоты по любому из утверждений 1-49.
58. Нуклеиновая кислота для ингибирования экспрессии LPA, содержащая по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, при этом экспрессия LPA уменьшена до уровней, которые по меньшей мере на 15% ниже уровней экспрессии, наблюдаемых в аналогичных условиях испытаний, но в отсутствие указанной нуклеиновой кислоты или конъюгированной нуклеиновой кислоты или в присутствии неподавляющего контроля.
59. Нуклеиновая кислота для ингибирования экспрессии LPA, содержащая по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, при этом указанная первая цепь содержит нуклеотидную последовательность, выбранную из следующих последовательностей: SEQ ID NO: 9, 1, 3, 5, 7, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29 31, 33, 35, 37, 39, 41, 43, 63, 65, 67, 69, 71 или 73, при этом экспрессия LPA уменьшена до уровней, которые по меньшей мере на 15% ниже уровней экспрессии, наблюдаемых в аналогичных условиях испытаний, но в отсутствие указанной нуклеиновой кислоты или конъюгированной нуклеиновой кислоты или в присутствии неподавляющего контроля.
60. Нуклеиновая кислота для ингибирования экспрессии LPA в клетке, содержащая по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, при этом указанная первая цепь содержит нуклеотидную последовательность, выбранную из следующих последовательностей: SEQ ID NO: 63, 65, 67 и 73, при этом указанная первая цепь РНК содержит концевой 5'-(E)-винилфосфонатный нуклеотид.
Другие положения настоящего изобретения включают:
Конъюгат для ингибирования экспрессии гена LPA в клетке, причем указанный конъюгат содержит часть нуклеиновой кислоты и части лиганда, причем указанная часть нуклеиновой кислоты содержит по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи РНК и по меньшей мере часть второй цепи РНК, которая по меньшей мере частично комплементарна первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с указанного гена LPA, при этом указанные части лиганда содержат линкерную группу, происходящую из серинола, и нацеливающий лиганд для нацеливания в условиях in vivo на клетки и конъюгированный исключительно с 3' и/или 5'-концами одной или обеих цепей РНК, причем 5'-конец первой цепи РНК не конъюгирован, при этом:
вторая цепь РНК конъюгирована на 5'-конце с нацеливающим лигандом, и при этом (a) вторая цепь РНК также конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец первой цепи РНК не конъюгирован; или (b) первая цепь РНК конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец второй цепи РНК не конъюгирован; или (c) обе вторая цепь РНК и первая цепь РНК также конъюгированы на 3'-концах с нацеливающим лигандом; или
обе вторая цепь РНК и первая цепь РНК конъюгированы на 3'-концах с нацеливающим лигандом, и 5'-конец второй цепи РНК не конъюгирован,
или
конъюгат для ингибирования экспрессии гена LPA в клетке, причем указанный конъюгат содержит часть нуклеиновой кислоты и части лиганда, причем указанная часть нуклеиновой кислоты содержит по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи РНК и по меньшей мере часть второй цепи РНК, которая по меньшей мере частично комплементарна первой цепи, при этом указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с указанного гена LPA, при этом указанные части лиганда содержат линкерную группу и нацеливающий лиганд для нацеливания в условиях in vivo на клетки и конъюгированный исключительно с 3' и/или 5'-концами одной или обеих цепей РНК, причем 5'-конец первой цепи РНК не конъюгирован, при этом:
вторая цепь РНК конъюгирована на 5'-конце с нацеливающим лигандом, и при этом (a) вторая цепь РНК также конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец первой цепи РНК не конъюгирован; или (b) первая цепь РНК конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец второй цепи РНК не конъюгирован; или (c) обе вторая цепь РНК и первая цепь РНК также конъюгированы на 3'-концах с нацеливающим лигандом; или
обе вторая цепь РНК и первая цепь РНК конъюгированы на 3'-концах с нацеливающим лигандом, и 5'-конец второй цепи РНК не конъюгирован, необязательно
при этом указанная линкерная группа представляет собой линкерную группу, происходящую из серинола, или один из других типов линкеров, описанных в настоящей заявке.
Конъюгат по положению 1, в котором указанная вторая цепь РНК конъюгирована на 5'-конце с нацеливающим лигандом, указанная вторая цепь РНК также конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец указанной первой цепи РНК не конъюгирован.
Конъюгат по положению 1, в котором указанная вторая цепь РНК конъюгирована на 5'-конце с нацеливающим лигандом, указанная первая цепь РНК конъюгирована на 3'-конце с нацеливающим лигандом и 3'-конец указанной второй цепи РНК не конъюгирован.
Конъюгат по положению 1, в котором указанная вторая цепь РНК конъюгирована на 5'-конце с нацеливающим лигандом, и обе указанные вторая цепь РНК и первая цепь РНК также конъюгированы на 3'-концах с нацеливающим лигандом.
Конъюгат по положению 1, в котором обе указанные вторая цепь РНК и первая цепь РНК конъюгированы на 3'-концах с нацеливающим лигандом, и 5'-конец указанной второй цепи РНК не конъюгирован.
Конъюгат по любому из положений 1-5, в котором указанные лиганды представляют собой мономерные лиганды.
Конъюгат по любому из положений 1-6, в котором указанные лиганды выбраны из групп GalNAc и галактозы, в частности, групп GalNAc.
Конъюгат по любому из положений 1-7, в котором указанные конъюгированные цепи РНК конъюгированы с нацеливающим лигандом за счет линкерной группы, происходящей из серинола, включая дополнительный линкер, причем указанный дополнительный линкер представляет собой или содержит насыщенную, неразветвленную или разветвленную C1-15 алкильную цепь, при этом необязательно один или более атомов углерода (например, 1, 2 или 3 атома углерода, является приемлемым, когда 1 или 2, в частности, 1) замещен (ы) гетероатомом, выбранным из O, N, S(O)p, где p равен 0, 1 или 2 (например, группа CH2 замещена O или NH, или S, или SO2, или группа –CH3 на конце цепи или на ветви замещена OH или NH2), причем указанная цепь необязательно замещена одной или более оксогруппами (например, 1-3, например, 1 группой).
Конъюгат по положению 8, в котором указанный дополнительный линкер содержит насыщенную неразветвленную C1-15 алкильную цепь, в которой один или более атомов углерода (например, 1, 2 или 3 атома углерода, является приемлемым, когда 1 или 2, в частности, 1) замещен (ы) атомом кислорода.
Конъюгат по положению 9, в котором указанный дополнительный линкер содержит цепь ПЭГ.
Конъюгат по положению 8, в котором указанный дополнительный линкер содержит насыщенную неразветвленную C1-15 алкильную цепь.
Конъюгат по положению 11, в котором указанный дополнительный линкер содержит насыщенную неразветвленную C1-6 алкильную цепь.
Конъюгат по положению 12, в котором указанный дополнительный линкер содержит насыщенную неразветвленную C4 или C6 алкильную цепь, например, C4 алкильную цепь.
Конъюгат по положению 1 или утверждению 1, в котором указанная первая цепь РНК представляет собой соединение формулы (XV):
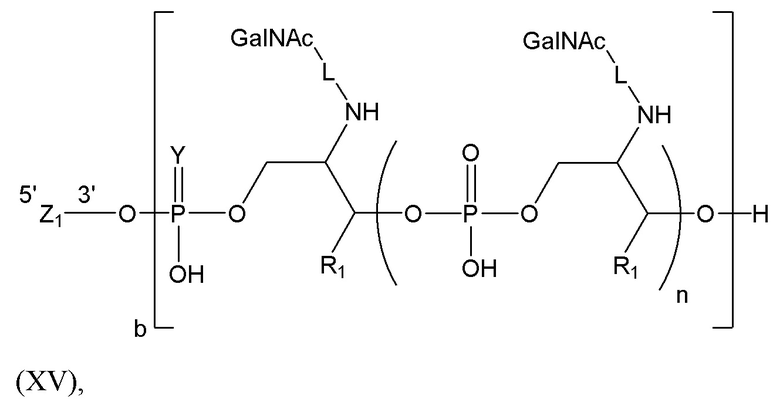
в которой b равно 0 или 1; и
указанная вторая цепь РНК представляет собой соединение формулы (XVI):
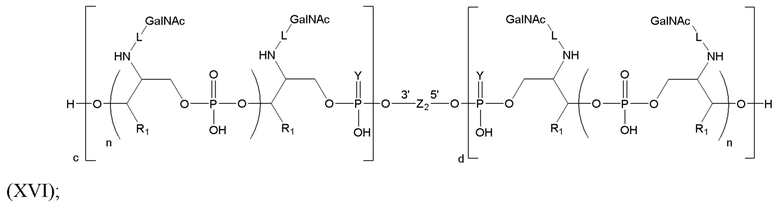
в которой с и d независимо равны 0 или 1;
в которой:
Z1 и Z2 представляют собой части РНК указанных первой и второй цепей РНК, соответственно;
Y представляет собой O или S;
R1 представляет собой H или метил;
n равно 0, 1, 2 или 3; и
L является одинаковым или различным в формулах (XV) и (XVI) и выбран из группы, состоящей из:
-(CH2)q-, где q=2-12;
-(CH2)r-C(O)-, где r=2-12;
-(CH2-CH2-O)s-CH2-C(O)-, где s=1-5;
-(CH2)t-CO-NH-(CH2)t-NH-C(O)-, где t независимо составляет 1-5;
-(CH2)u-CO-NH-(CH2)u-C(O)-, где u независимо составляет 1-5; и
-(CH2)v-NH-C(O)-, где v составляет 2-12; и
при этом концевой C(O) (если он присутствует) присоединен к группе NH;
и при этом b + c + d равно 2 или 3.
Конъюгат по положению 14, в котором b равно 0, c равно 1 и d равно 1.
Конъюгат по положению 14, в котором b равно 1, c равно 0 и d равно 1.
Конъюгат по положению 14, в котором b равно 1, c равно 1 и d равно 0.
Конъюгат по положению 14, в котором b равно 1, c равно 1 и d равно 1.
Конъюгат по любому из положений 14-18, в котором Y представляет собой O.
Конъюгат по любому из положений 14-18, в котором Y представляет собой S.
Конъюгат по любому из положений 14-20, в котором R1 представляет собой H.
Конъюгат по любому из положений 14-20, в котором R1 представляет собой метил.
Конъюгат по любому из положений 14-22, в котором n равно 0.
Конъюгат по любому из положений 14-23, в котором L представляет собой -(CH2)r-C(O)-, где r=2-12.
Конъюгат по положению 24, в котором r=2-6.
Конъюгат по положению 25, в котором r=4 или 6, например, 4.
Конъюгат по любому из предшествующих положений с любым признаком или комбинацией признаков, раскрытых в настоящей заявке.
Обозначение

Одна последовательность может иметь более одного названия. В этих случаях одно из этих названий приведено в сводной таблице последовательностей.
Если в последовательности РНК указаны конкретные линкеры и/или модифицированные связи, такие как PS и [ST23 (ps)]3 ST41 (ps) и т.д., они представляют собой необязательные части последовательности, но являются предпочтительным вариантом реализации этой последовательности.
Могут быть использованы следующие сокращения:
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Silence Therapeutics GmbH
<120> НУКЛЕИНОВЫЕ КИСЛОТЫ ДЛЯ ИНГИБИРОВАНИЯ ЭКСПРЕССИИ LPA В КЛЕТКЕ
<130> S109PCT
<150> EP17201449.0
<151> 2017-11-13
<150> EP18179175.7
<151> 2018-06-21
<150> GB1815915.2
<151> 2018-09-28
<160> 162
<170> PatentIn version 3.5
<210> 1
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 1
ucguauaaca auaaggggc 19
<210> 2
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 2
gccccuuauu guuauacga 19
<210> 3
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 3
gauaacucug uccauuacc 19
<210> 4
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 4
gguaauggac agaguuauc 19
<210> 5
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 5
auaacucugu ccauuacca 19
<210> 6
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 6
ugguaaugga cagaguuau 19
<210> 7
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 7
uaacucuguc cauuaccgu 19
<210> 8
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 8
acgguaaugg acagaguua 19
<210> 9
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 9
auaacucugu ccauuaccg 19
<210> 10
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 10
cgguaaugga cagaguuau 19
<210> 11
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 11
agaaugugcc ucgauaacu 19
<210> 12
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 12
aguuaucgag gcacauucu 19
<210> 13
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 13
auaacucugu ccaucacca 19
<210> 14
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 14
uggugaugga cagaguuau 19
<210> 15
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 15
auaacucugu ccaucaccu 19
<210> 16
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 16
aggugaugga cagaguuau 19
<210> 17
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 17
uaacucuguc cauuaccau 19
<210> 18
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 18
augguaaugg acagaguua 19
<210> 19
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 19
augugccuug auaacucug 19
<210> 20
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 20
cagaguuauc aaggcacau 19
<210> 21
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 21
aguuggugcu gcuucagaa 19
<210> 22
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 22
uucugaagca gcaccaacu 19
<210> 23
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 23
aauaaggggc ugccacagg 19
<210> 24
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 24
ccuguggcag ccccuuauu 19
<210> 25
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 25
uaacucuguc caucaccau 19
<210> 26
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 26
auggugaugg acagaguua 19
<210> 27
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 27
augagccucg auaacucug 19
<210> 28
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 28
cagaguuauc gaggcucau 19
<210> 29
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 29
aaugagccuc gauaacucu 19
<210> 30
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 30
agaguuaucg aggcucauu 19
<210> 31
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 31
aaugcuucca ggacauuuc 19
<210> 32
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 32
gaaauguccu ggaagcauu 19
<210> 33
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 33
acaguggugg agaaugugc 19
<210> 34
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 34
gcacauucuc caccacugu 19
<210> 35
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 35
guaugugccu cgauaacuc 19
<210> 36
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 36
gaguuaucga ggcacauac 19
<210> 37
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 37
ucgauaacuc uguccauca 19
<210> 38
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 38
ugauggacag aguuaucga 19
<210> 39
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 39
ugucacugga cauuguguc 19
<210> 40
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 40
gacacaaugu ccagugaca 19
<210> 41
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 41
cugggaucca ugguguaac 19
<210> 42
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 42
guuacaccau ggaucccag 19
<210> 43
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 43
agaugaccaa gcuuggcag 19
<210> 44
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 44
cugccaagcu uggucaucu 19
<210> 45
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 45
aagtgtcctt gcgacgtcc 19
<210> 46
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 46
cctggactgt ggggcttt 18
<210> 47
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд
<400> 47
ctgtttctga acaagcacca acggagc 27
<210> 48
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 48
gtgtcctcgc aacgtcca 18
<210> 49
<211> 15
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 49
gaccccgggg ctttg 15
<210> 50
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд
<400> 50
tggctgtttc tgaacaagca ccaatgg 27
<210> 51
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 51
tcattccttc cccaaagaga cc 22
<210> 52
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 52
cacctccgtt ttggtggtag ag 22
<210> 53
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд
<400> 53
caagctgctc agtggaggca acacatta 28
<210> 54
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 54
gcatgggtca gaaggattcc tat 23
<210> 55
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 55
tgtagaaggt gtggtgccag att 23
<210> 56
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд
<400> 56
tcgagcacgg catcgtcacc aa 22
<210> 57
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 57
aaggccaacc gcgagaag 18
<210> 58
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 58
agaggcgtac agggacagca 20
<210> 59
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд
<400> 59
tgagaccttc aacaccccag ccatgtac 28
<210> 60
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 60
agatgtaggc cgggtgatct tt 22
<210> 61
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 61
gtagccaaat cctttctctc ctgt 24
<210> 62
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд
<400> 62
tgttccaaaa acagtggata attttgtggc c 31
<210> 63
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 63
uuaacucugu ccauuaccg 19
<210> 64
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 64
cgguaaugga cagaguuaa 19
<210> 65
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 65
uuaacucugu ccauuaccc 19
<210> 66
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 66
ggguaaugga cagaguuaa 19
<210> 67
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 67
uuaacucugu ccauuaccu 19
<210> 68
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 68
agguaaugga cagaguuaa 19
<210> 69
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 69
auaacucugu ccauuaccc 19
<210> 70
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 70
ggguaaugga cagaguuau 19
<210> 71
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 71
auaacucugu ccauuaccu 19
<210> 72
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 72
agguaaugga cagaguuau 19
<210> 73
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 73
uuaacucugu ccauuacca 19
<210> 74
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК
<400> 74
ugguaaugga cagaguuaa 19
<210> 75
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК – модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 75
ucguauaaca auaaggggc 19
<210> 76
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 76
gccccuuauu guuauacga 19
<210> 77
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 77
gauaacucug uccauuacc 19
<210> 78
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 78
gguaauggac agaguuauc 19
<210> 79
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 79
auaacucugu ccauuacca 19
<210> 80
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 80
ugguaaugga cagaguuau 19
<210> 81
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 81
uaacucuguc cauuaccgu 19
<210> 82
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 82
acgguaaugg acagaguua 19
<210> 83
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 83
auaacucugu ccauuaccg 19
<210> 84
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 84
cgguaaugga cagaguuau 19
<210> 85
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 85
agaaugugcc ucgauaacu 19
<210> 86
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 86
aguuaucgag gcacauucu 19
<210> 87
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 87
auaacucugu ccaucacca 19
<210> 88
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 88
uggugaugga cagaguuau 19
<210> 89
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 89
auaacucugu ccaucaccu 19
<210> 90
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 90
aggugaugga cagaguuau 19
<210> 91
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 91
uaacucuguc cauuaccau 19
<210> 92
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 92
augguaaugg acagaguua 19
<210> 93
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 93
augugccuug auaacucug 19
<210> 94
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 94
cagaguuauc aaggcacau 19
<210> 95
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 95
aguuggugcu gcuucagaa 19
<210> 96
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 96
uucugaagca gcaccaacu 19
<210> 97
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 97
aauaaggggc ugccacagg 19
<210> 98
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 98
ccuguggcag ccccuuauu 19
<210> 99
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 99
uaacucuguc caucaccau 19
<210> 100
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 100
auggugaugg acagaguua 19
<210> 101
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 101
augagccucg auaacucug 19
<210> 102
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 102
cagaguuauc gaggcucau 19
<210> 103
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 103
aaugagccuc gauaacucu 19
<210> 104
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 104
agaguuaucg aggcucauu 19
<210> 105
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 105
aaugcuucca ggacauuuc 19
<210> 106
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 106
gaaauguccu ggaagcauu 19
<210> 107
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 107
acaguggugg agaaugugc 19
<210> 108
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 108
gcacauucuc caccacugu 19
<210> 109
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 109
guaugugccu cgauaacuc 19
<210> 110
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 110
gaguuaucga ggcacauac 19
<210> 111
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 111
ucgauaacuc uguccauca 19
<210> 112
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 112
ugauggacag aguuaucga 19
<210> 113
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 113
ugucacugga cauuguguc 19
<210> 114
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 114
gacacaaugu ccagugaca 19
<210> 115
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 115
cugggaucca ugguguaac 19
<210> 116
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 116
guuacaccau ggaucccag 19
<210> 117
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 117
agaugaccaa gcuuggcag 19
<210> 118
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 118
cugccaagcu uggucaucu 19
<210> 119
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 119
auaacucugu ccauuacca 19
<210> 120
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 120
ugguaaugga cagaguuau 19
<210> 121
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 121
auaacucugu ccauuacca 19
<210> 122
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 122
ugguaaugga cagaguuau 19
<210> 123
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 123
auaacucugu ccauuaccg 19
<210> 124
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 124
cgguaaugga cagaguuau 19
<210> 125
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 125
auaacucugu ccauuaccg 19
<210> 126
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 126
cgguaaugga cagaguuau 19
<210> 127
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 127
uuauagagca agaacacugu u 21
<210> 128
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 128
aacaguguuc uugcucuaua a 21
<210> 129
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 129
uuauagagca agaacacugu u 21
<210> 130
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 130
aacaguguuc uugcucuaua a 21
<210> 131
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 131
aacaguguuc uugcucuaua a 21
<210> 132
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 132
ucgaaguauu ccgcguacg 19
<210> 133
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 133
cguacgcgga auacuucga 19
<210> 134
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 134
aacaguguuc uugcucuaua a 21
<210> 135
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 135
auaacucugu ccauuaccg 19
<210> 136
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 136
cgguaaugga cagaguuau 19
<210> 137
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 137
cgguaaugga cagaguuau 19
<210> 138
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 138
cuuacucucg cccaagcga 19
<210> 139
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 139
ucgcuugggc gagaguaag 19
<210> 140
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 140
tggctgtttc tgaacaagca ccaatgg 27
<210> 141
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Зонд - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 141
tcgagcacgg catcgtcacc aa 22
<210> 142
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 142
aacaguguuc uugcucuaua a 21
<210> 143
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 143
aacaguguuc uugcucuaua a 21
<210> 144
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 144
aacaguguuc uugcucuaua a 21
<210> 145
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 145
aacaguguuc uugcucuaua a 21
<210> 146
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 146
aacaguguuc uugcucuaua a 21
<210> 147
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 147
aacaguguuc uugcucuaua a 21
<210> 148
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 148
aacaguguuc uugcucuaua a 21
<210> 149
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 149
aacaguguuc uugcucuaua a 21
<210> 150
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 150
aacaguguuc uugcucuaua a 21
<210> 151
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 151
ucuucuuaaa cugaguuuc 19
<210> 152
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 152
gaaacucagu uuaagaaga 19
<210> 153
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 153
auguagccga ggaucuucu 19
<210> 154
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 154
agaagauccu cggcuacau 19
<210> 155
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 155
aaccagaaga agcagguga 19
<210> 156
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 156
ucaccugcuu cuucugguu 19
<210> 157
<211> 21
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 157
aacaguguuc uugcucuaua a 21
<210> 158
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 158
gaaacucagu uuaagaaga 19
<210> 159
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 159
agaagauccu cggcuacau 19
<210> 160
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 160
uaccagaaga agcagguga 19
<210> 161
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 161
ucaccugcuu cuucuggua 19
<210> 162
<211> 19
<212> РНК
<213> Искусственная последовательность
<220>
<223> миРНК - модифицированное_основание, модифицированное согласно
сводной таблице последовательностей в конце описания
<400> 162
cgguaaugga cagaguuau 19
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ЛЕЧЕНИЯ ИНФЕКЦИИ ГЕПАТИТА В | 2018 |
|
RU2780021C2 |
| НУКЛЕИНОВЫЕ КИСЛОТЫ ДЛЯ ИНГИБИРОВАНИЯ ЭКСПРЕССИИ ГЕНА-МИШЕНИ, СОДЕРЖАЩИЕ ФОСФОДИТИОАТНЫЕ СВЯЗИ | 2018 |
|
RU2812806C2 |
| МОДУЛЯЦИЯ ЭКСПРЕССИИ HSP47 | 2010 |
|
RU2575056C2 |
| НИЗКОМОЛЕКУЛЯРНЫЕ КОНЪЮГАТЫ ДЛЯ ВНУТРИКЛЕТОЧНОЙ ДОСТАВКИ НУКЛЕИНОВЫХ КИСЛОТ | 2011 |
|
RU2582235C2 |
| КОНЪЮГАТЫ УГЛЕВОДА И LNA-ОЛИГОНУКЛЕОТИДА | 2014 |
|
RU2649367C2 |
| КОМПОЗИЦИИ И СПОСОБЫ МОДУЛИРОВАНИЯ ЭКСПРЕССИИ HBV И TTR | 2014 |
|
RU2670614C9 |
| КОМПОЗИЦИИ И СПОСОБЫ | 2014 |
|
RU2686080C2 |
| КОМПОЗИЦИИ И СПОСОБЫ МОДУЛИРОВАНИЯ ЭКСПРЕССИИ АПОЛИПОПРОТЕИНА (A) | 2014 |
|
RU2824214C1 |
| КОМПОЗИЦИИ И СПОСОБЫ МОДУЛИРОВАНИЯ ЭКСПРЕССИИ АПОЛИПОПРОТЕИНА C-III | 2014 |
|
RU2650510C2 |
| КОМПОЗИЦИИ И СПОСОБЫ МОДУЛИРОВАНИЯ ЭКСПРЕССИИ HBV И TTR | 2014 |
|
RU2782034C2 |
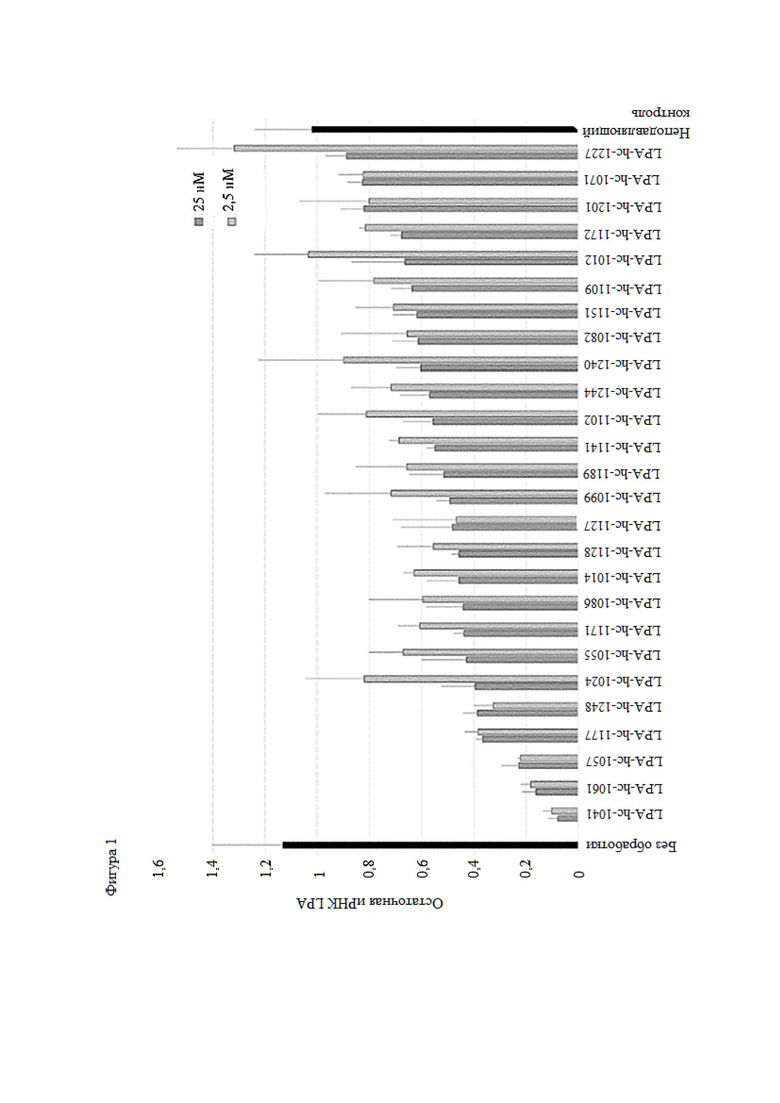
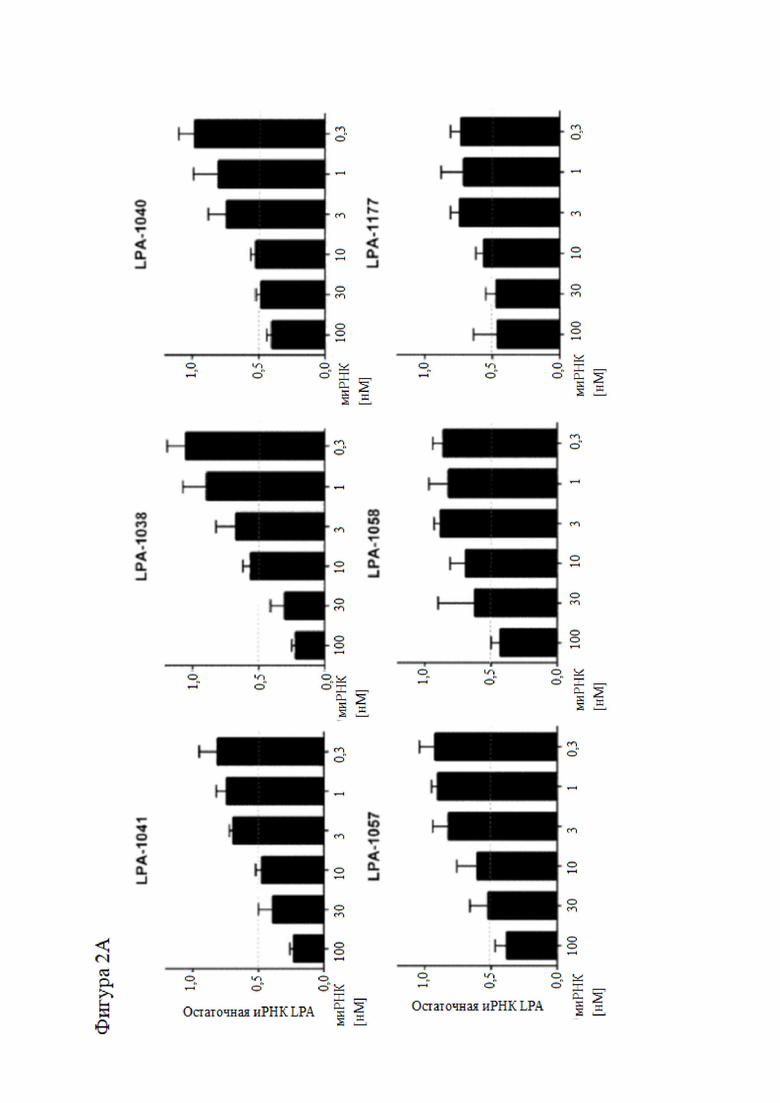

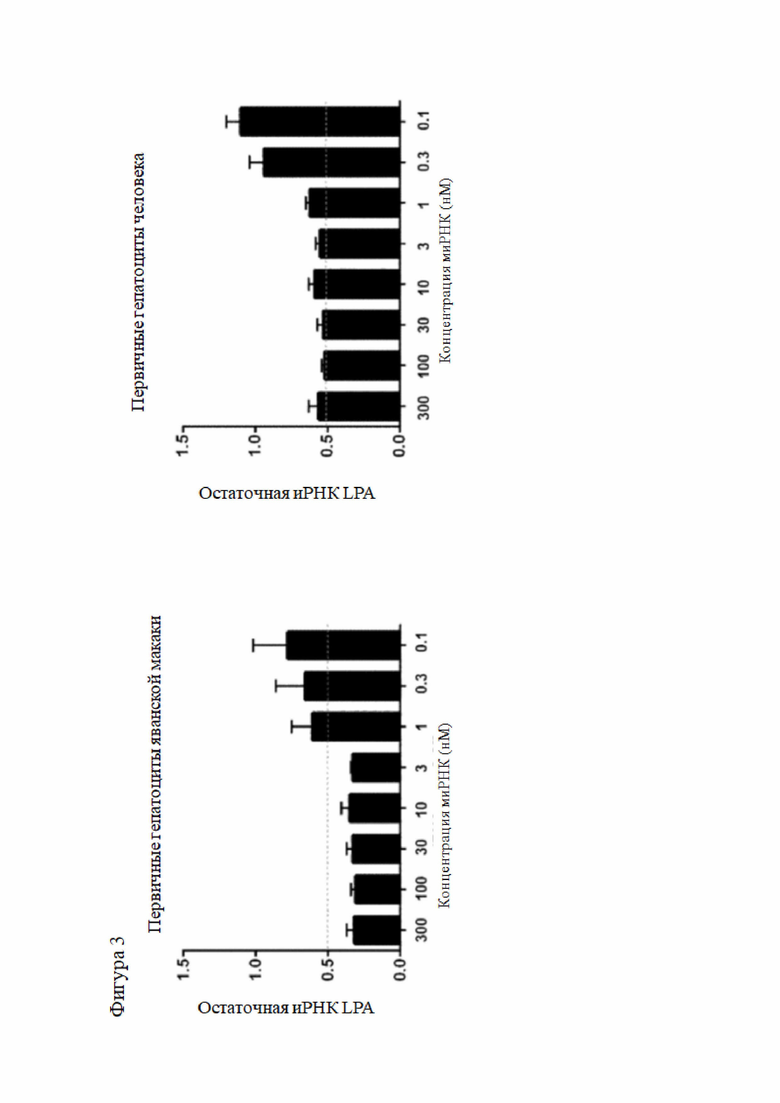
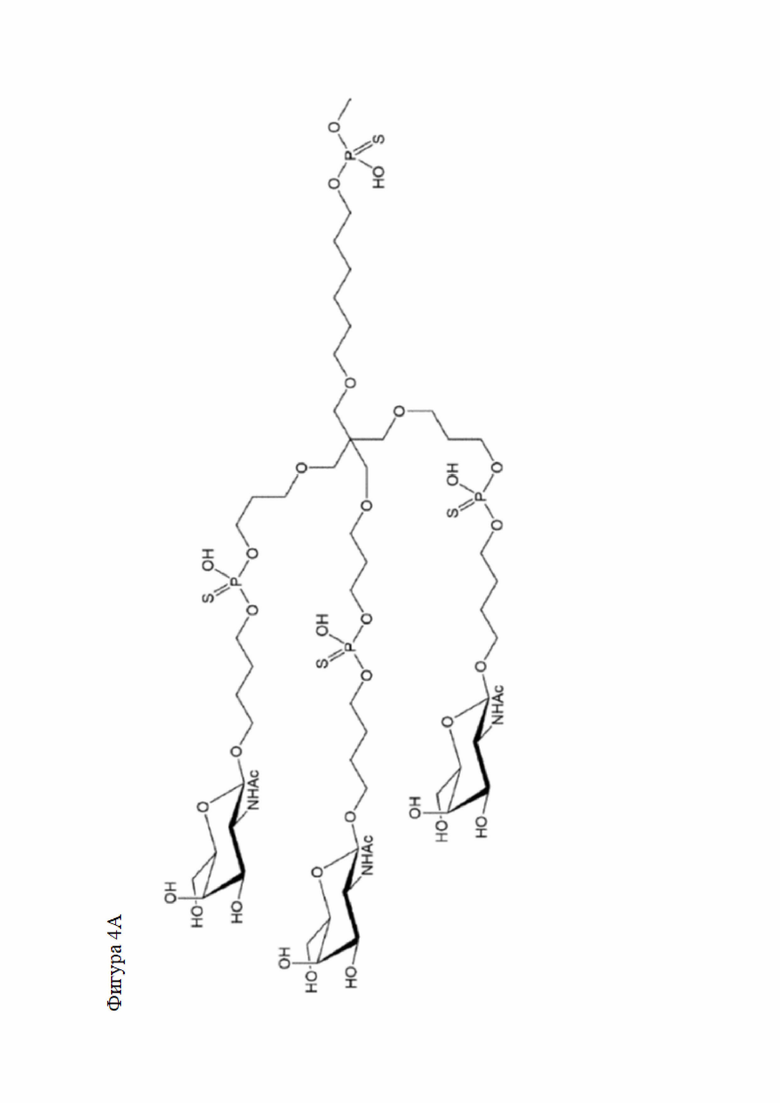
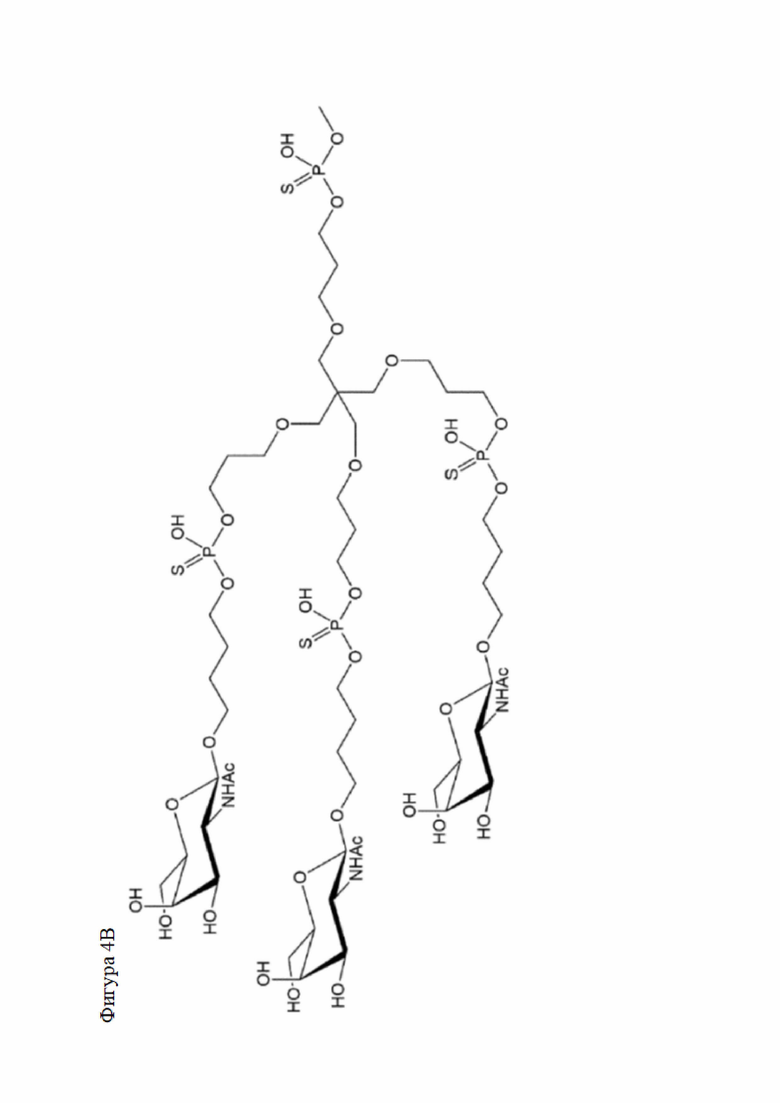

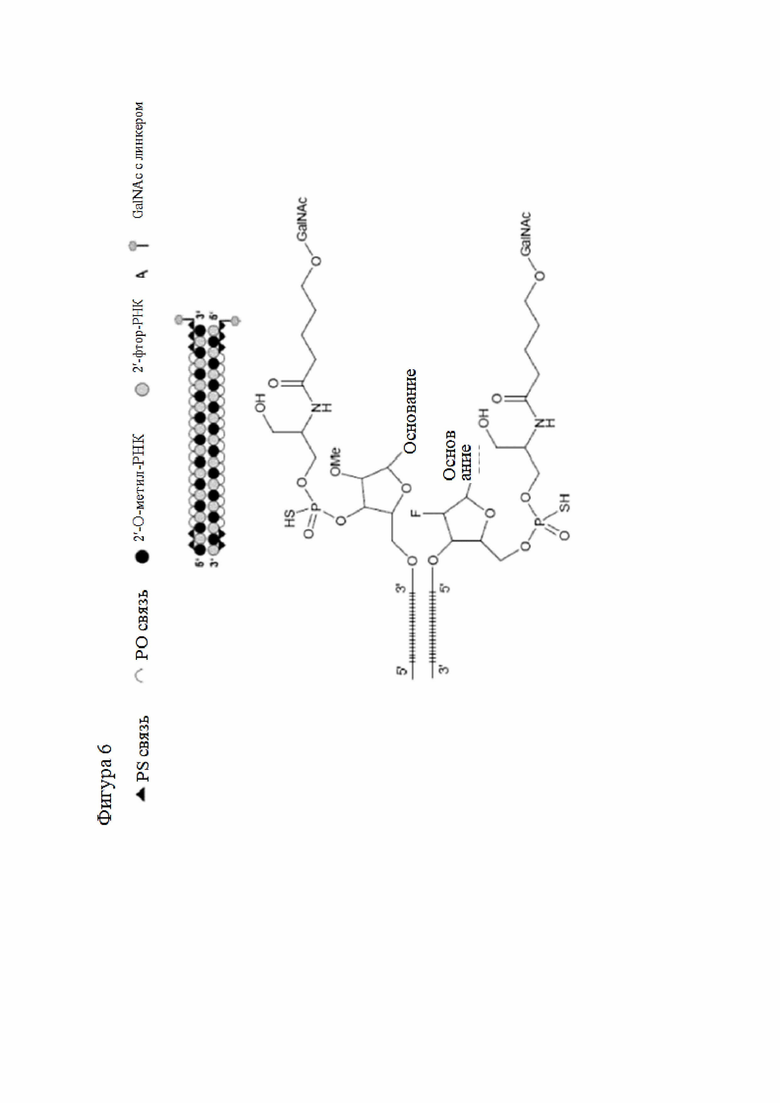
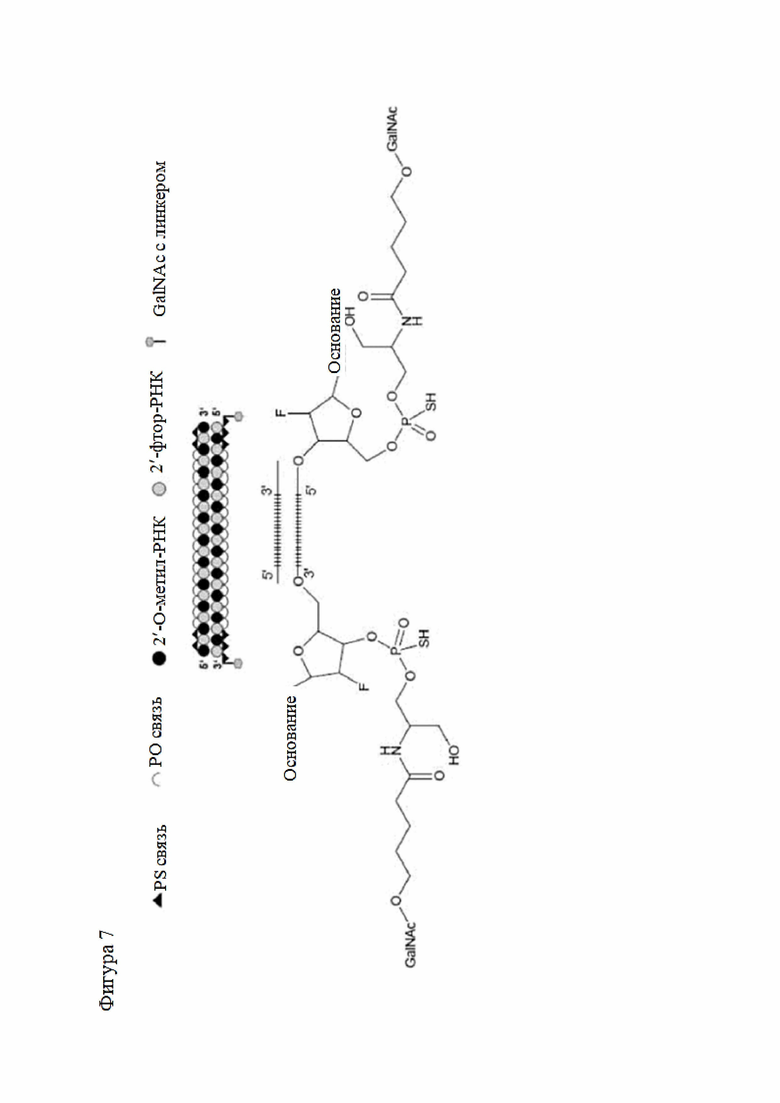
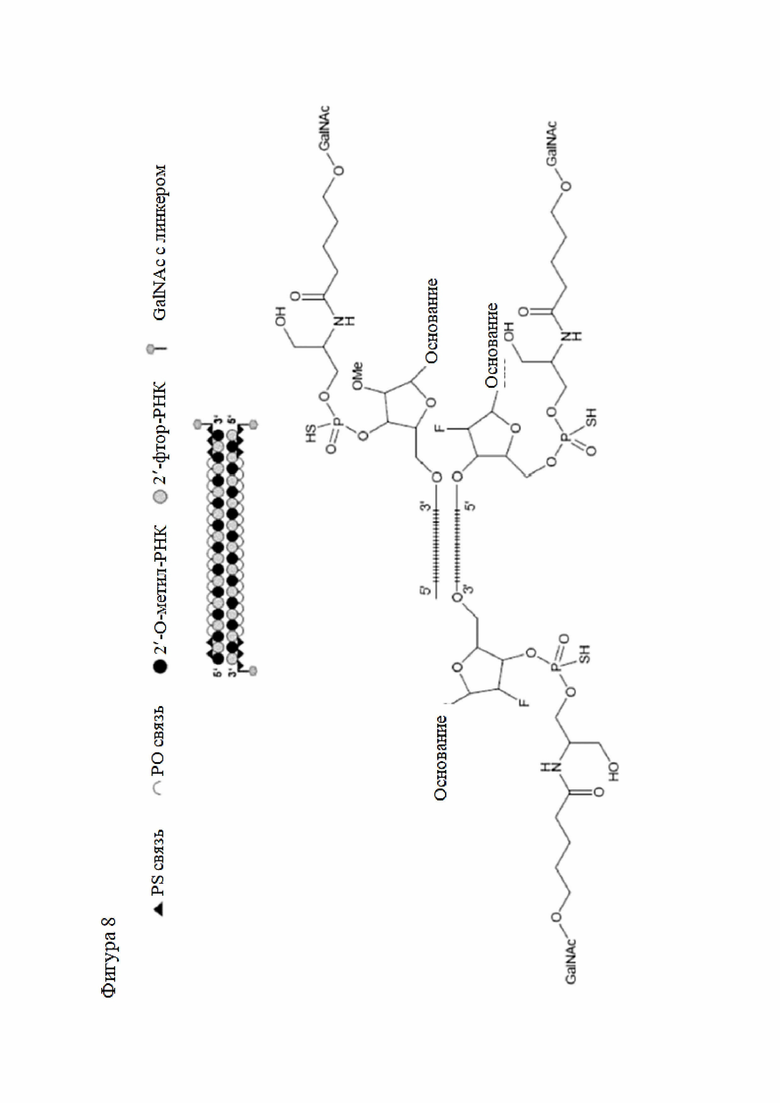
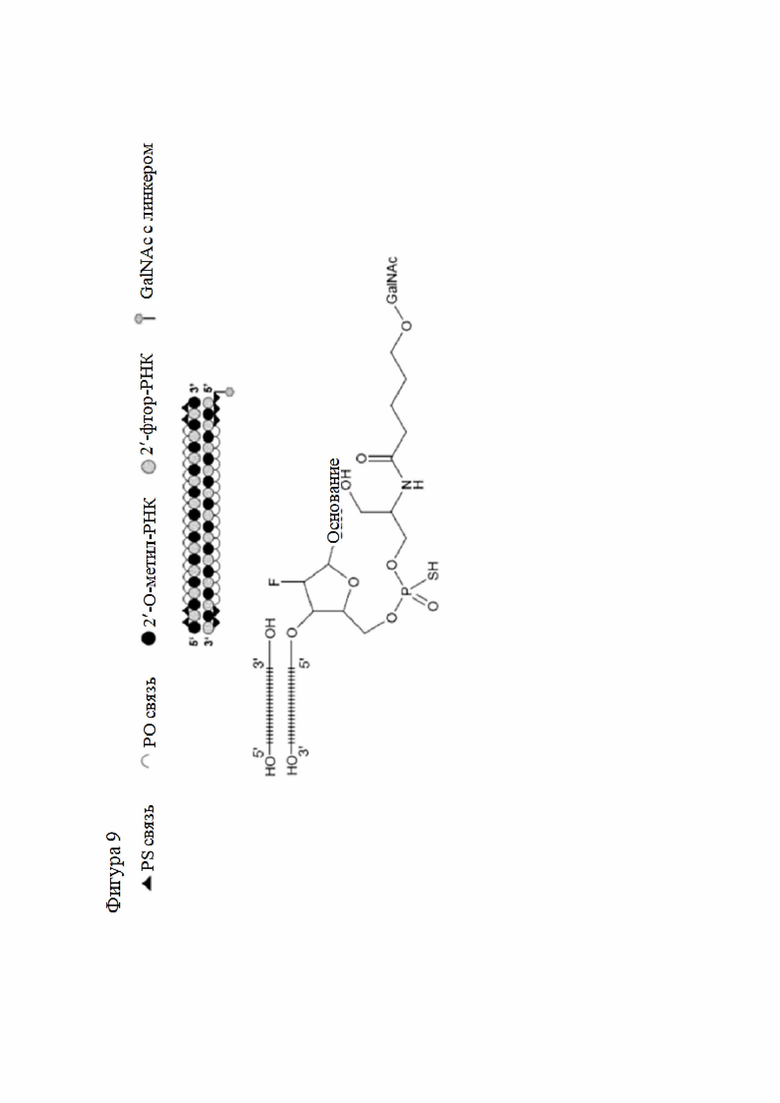

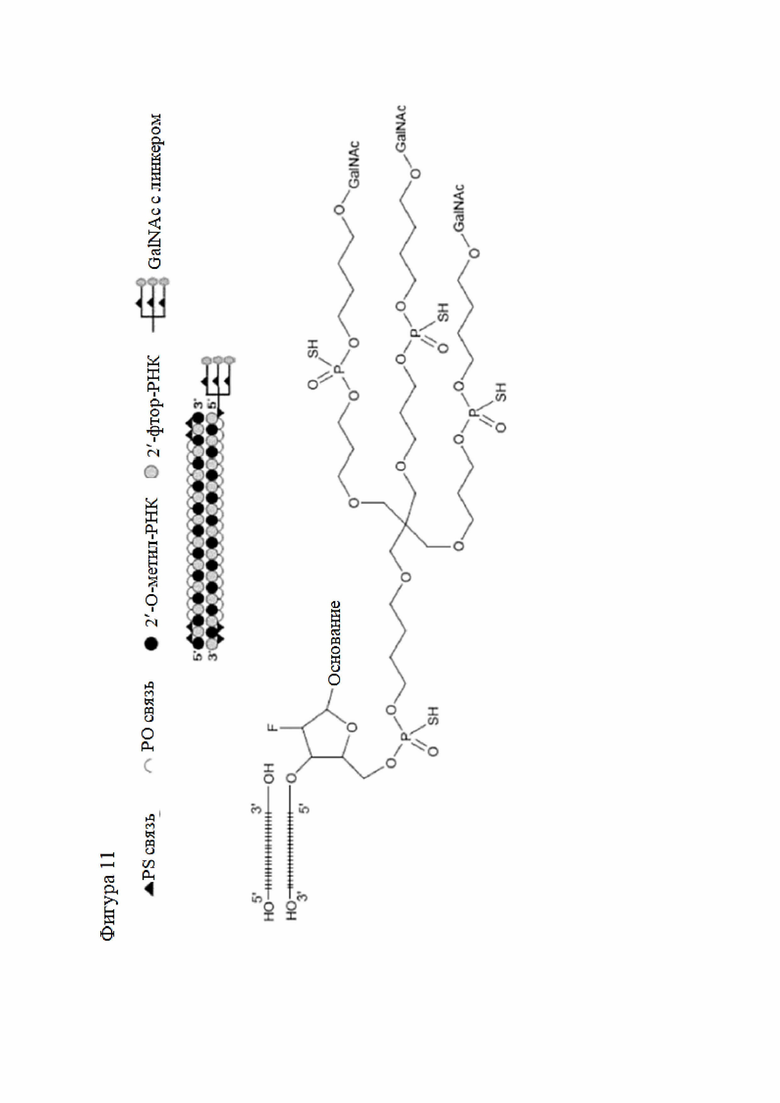
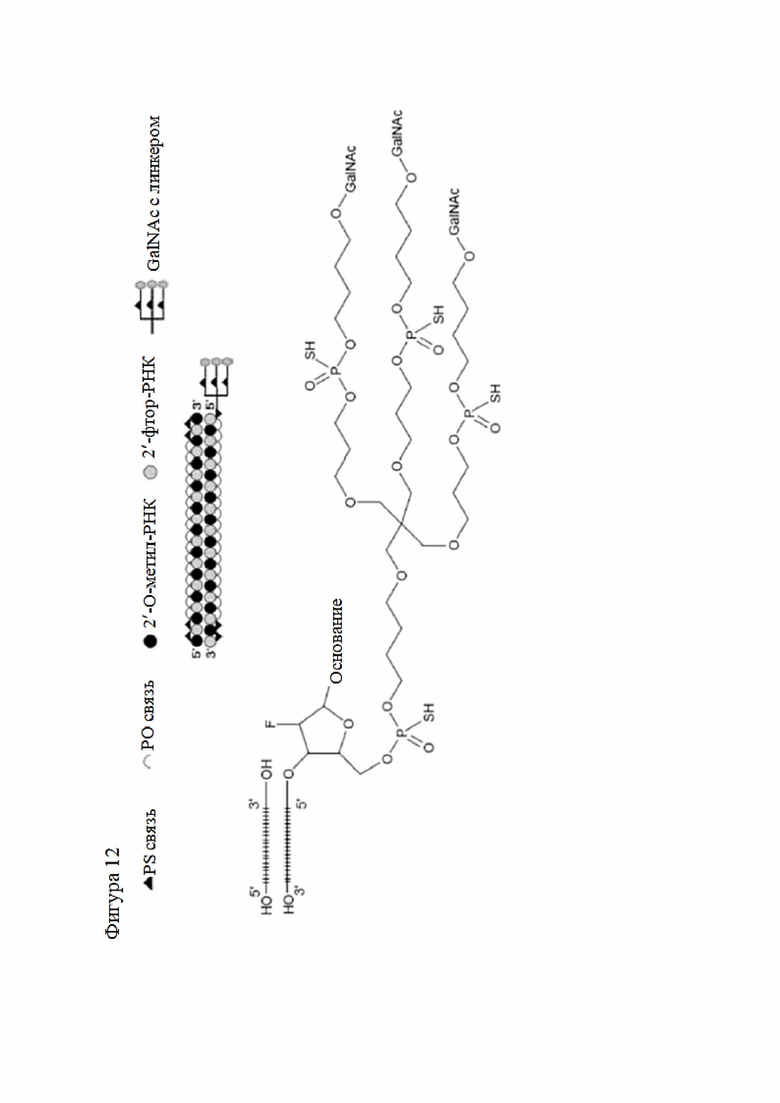


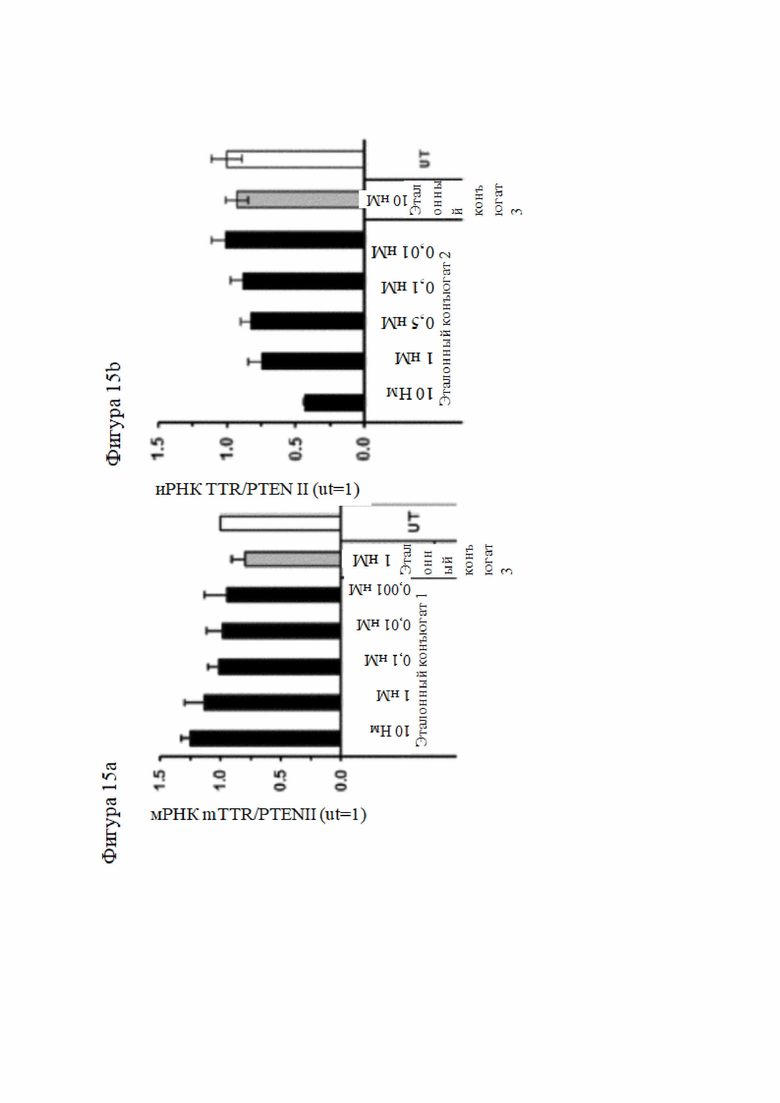
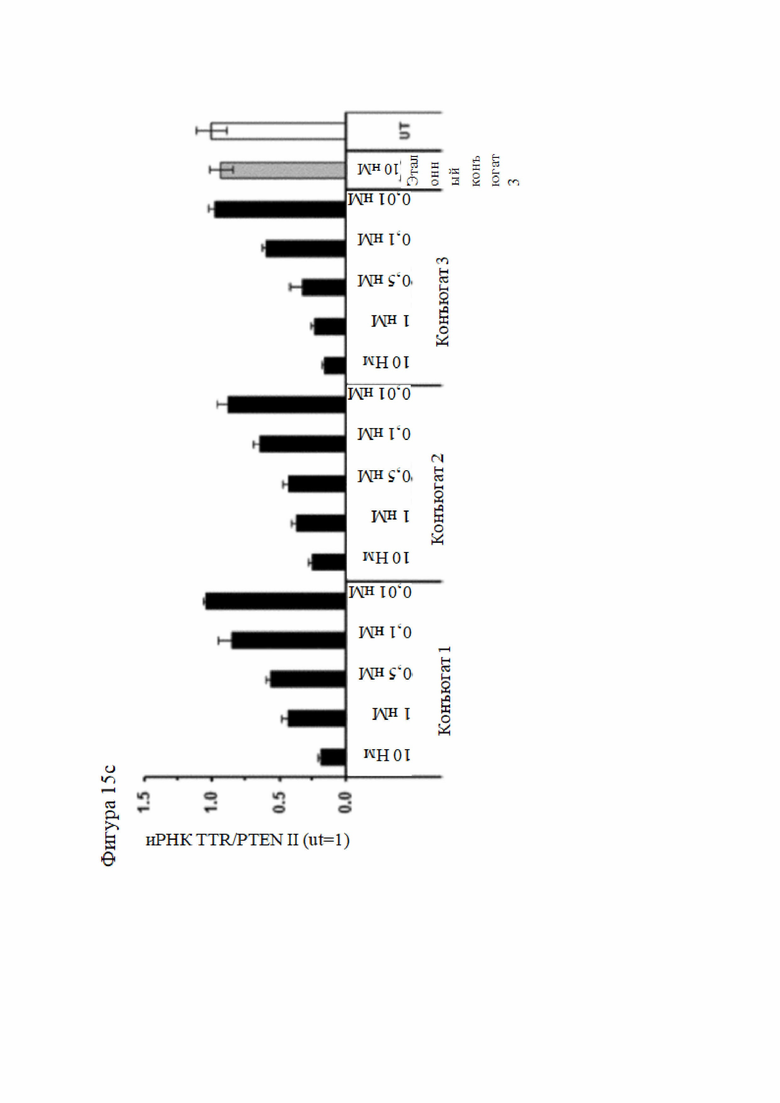





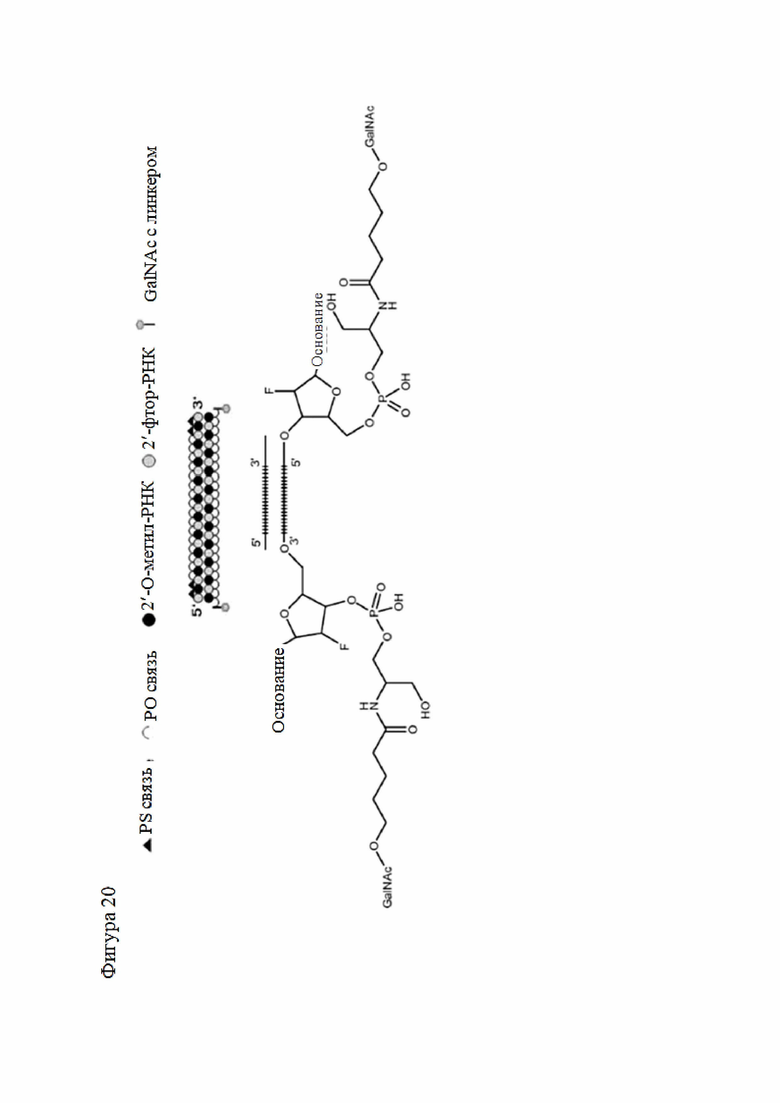
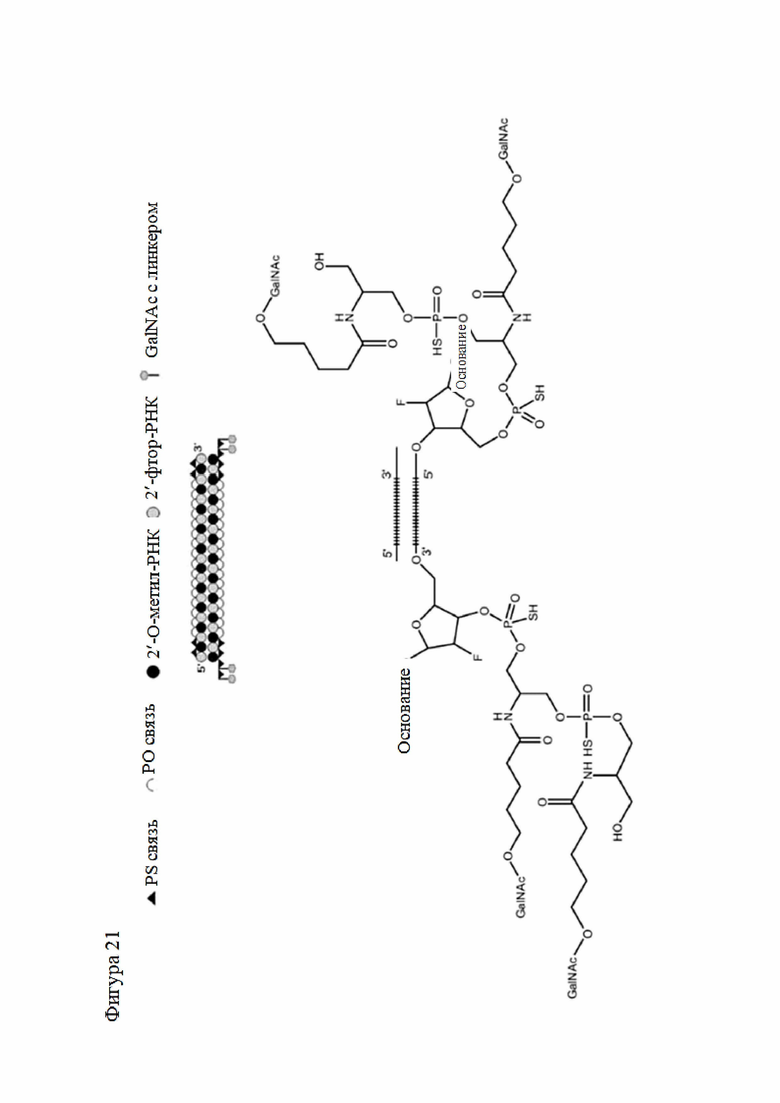
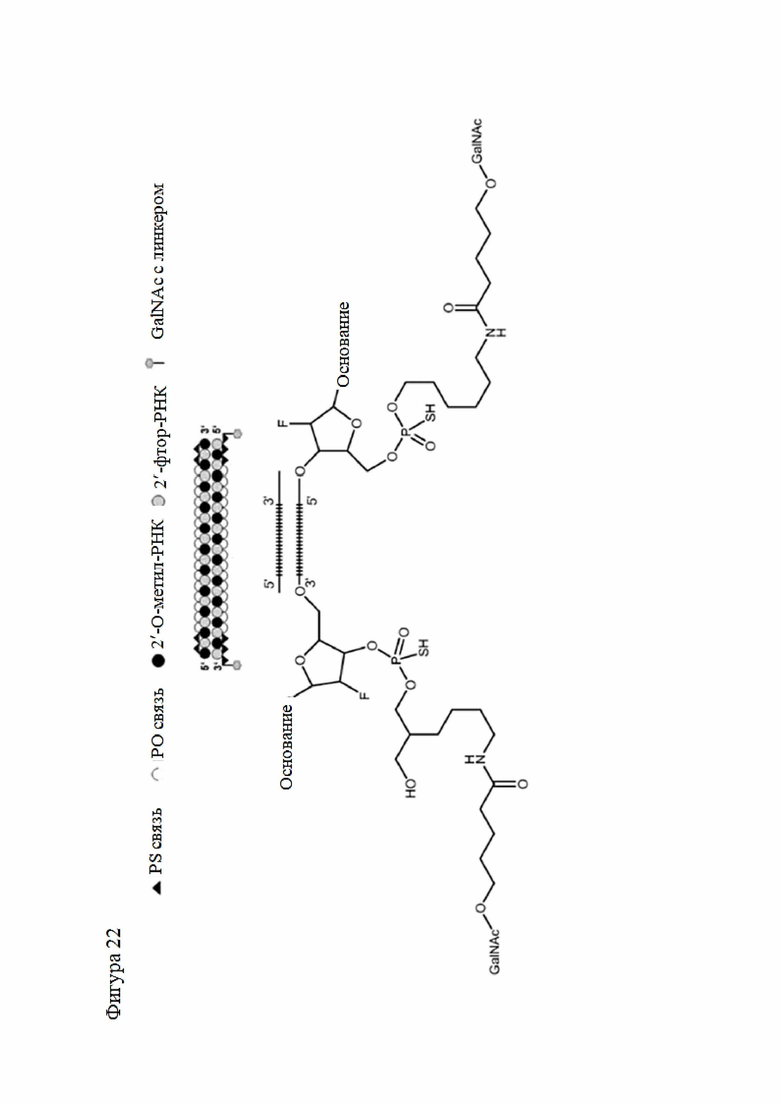
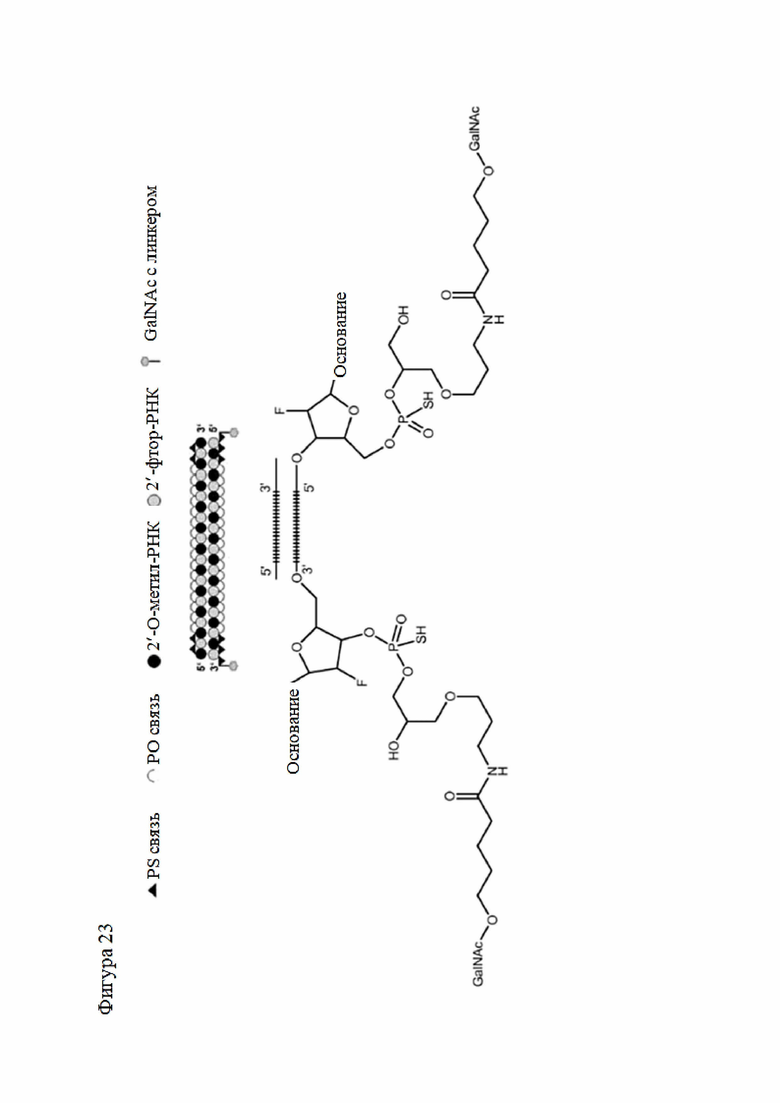

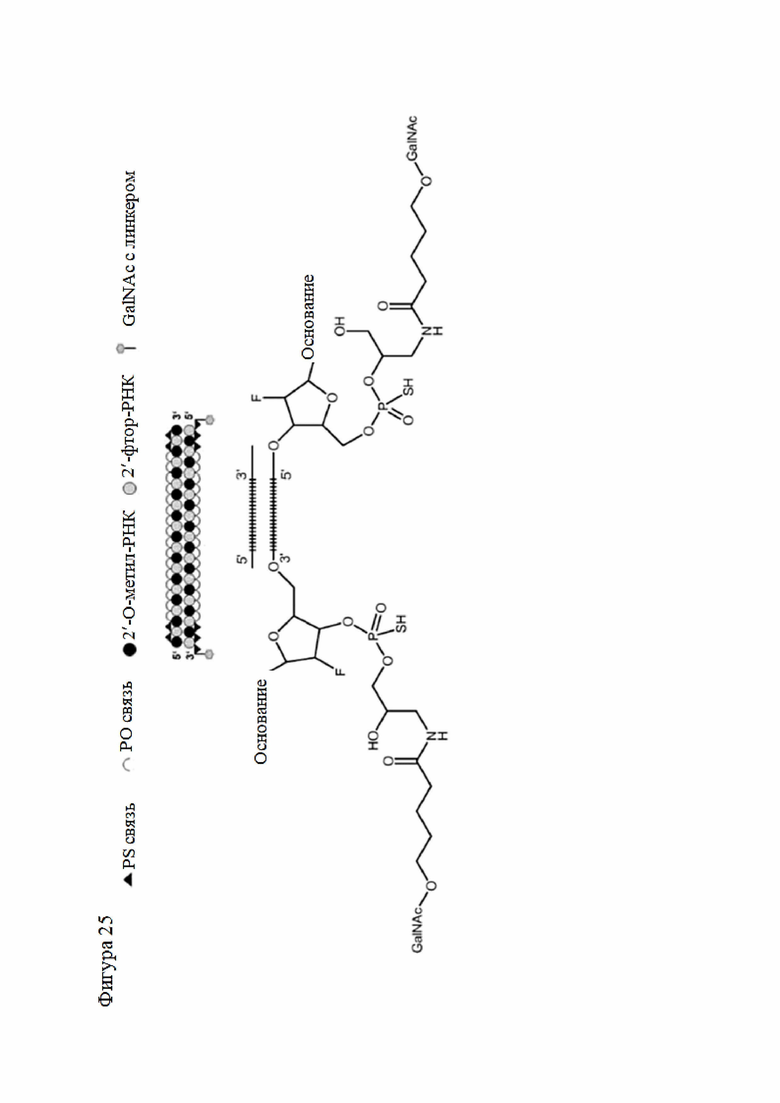
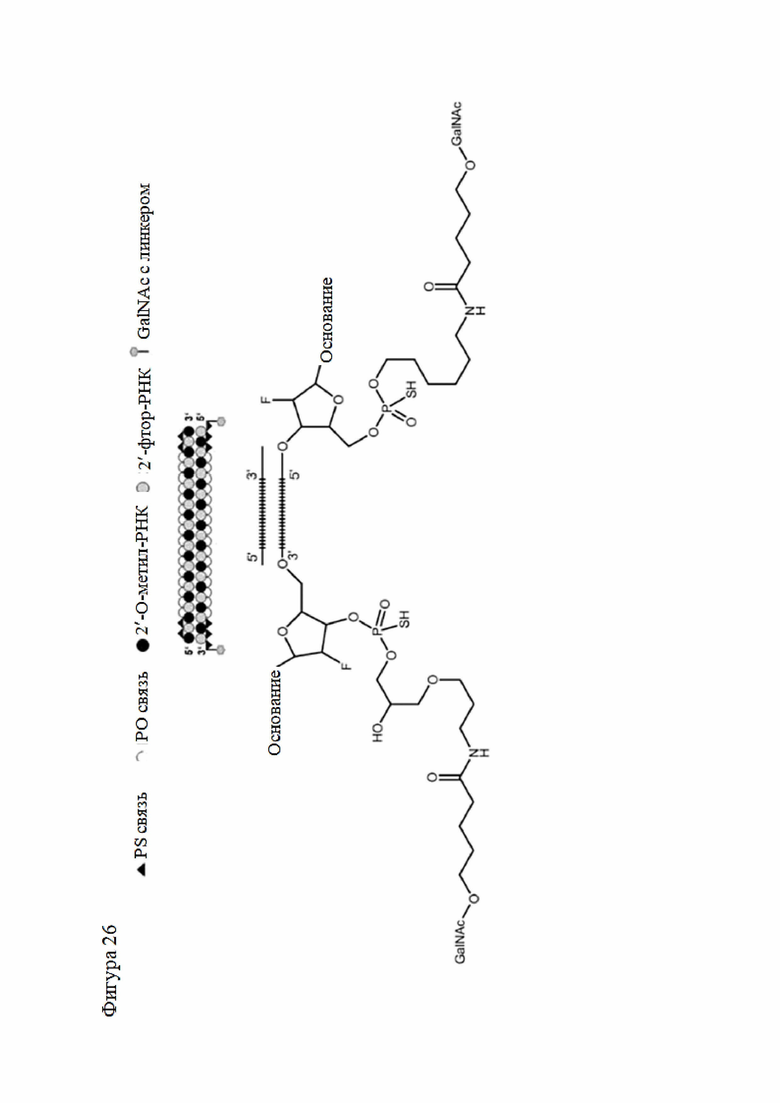
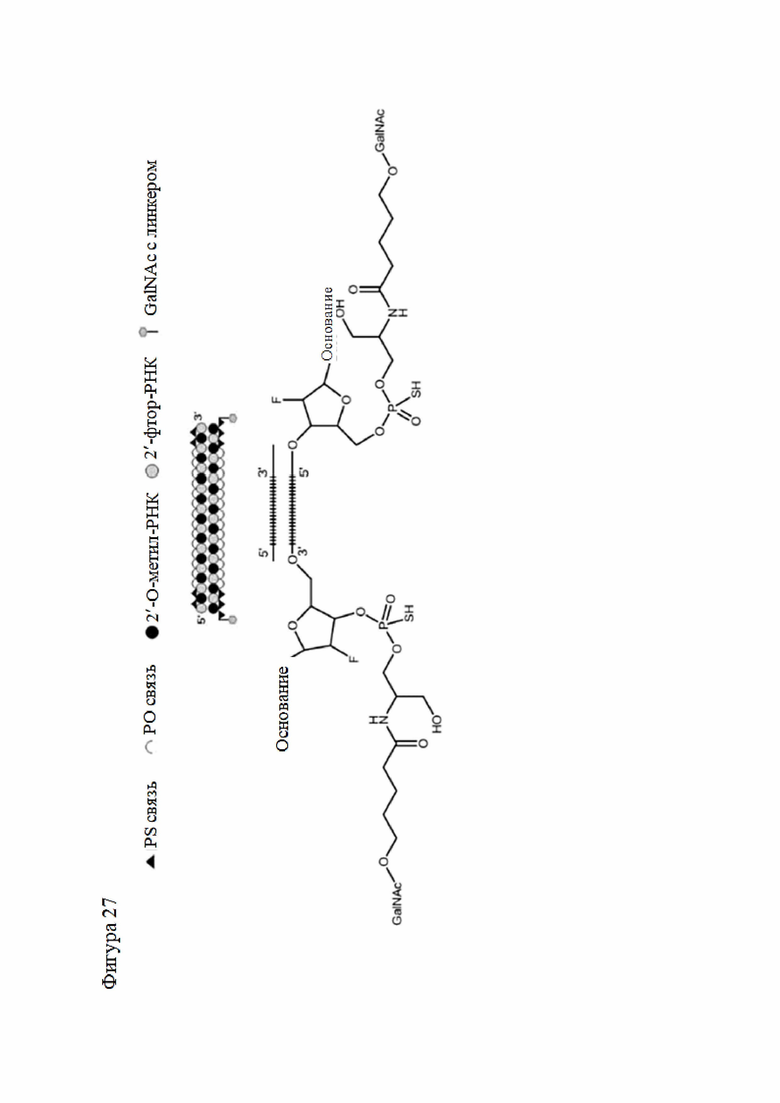
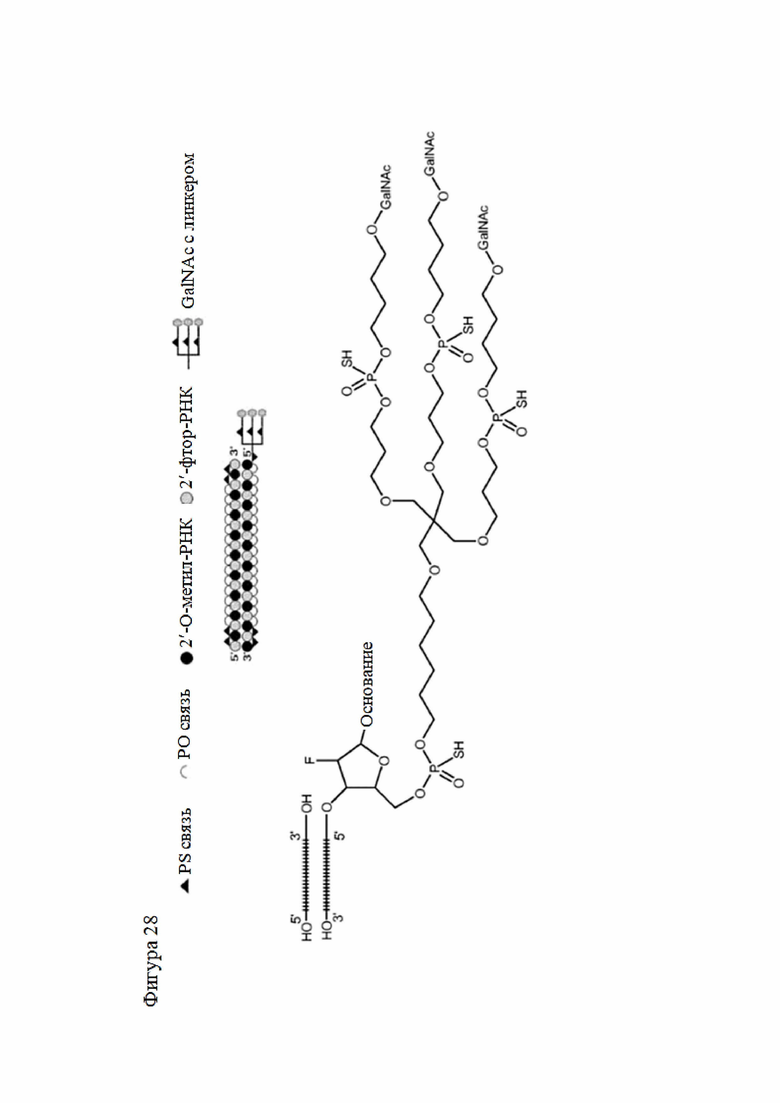
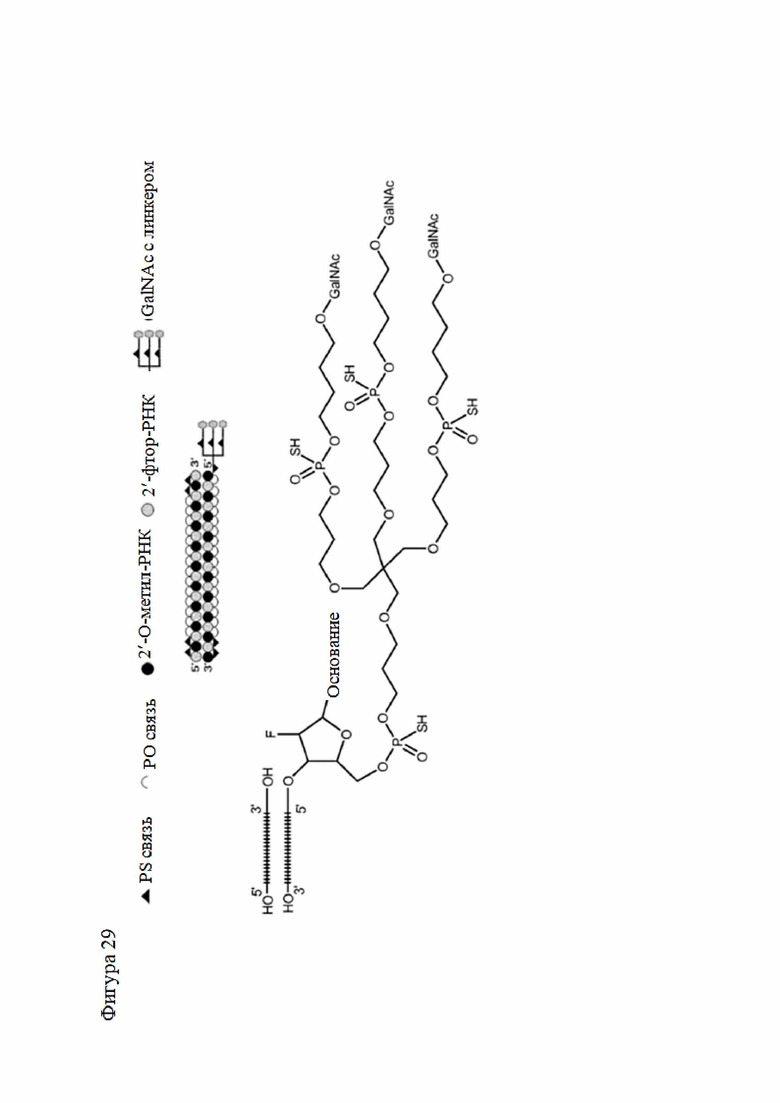
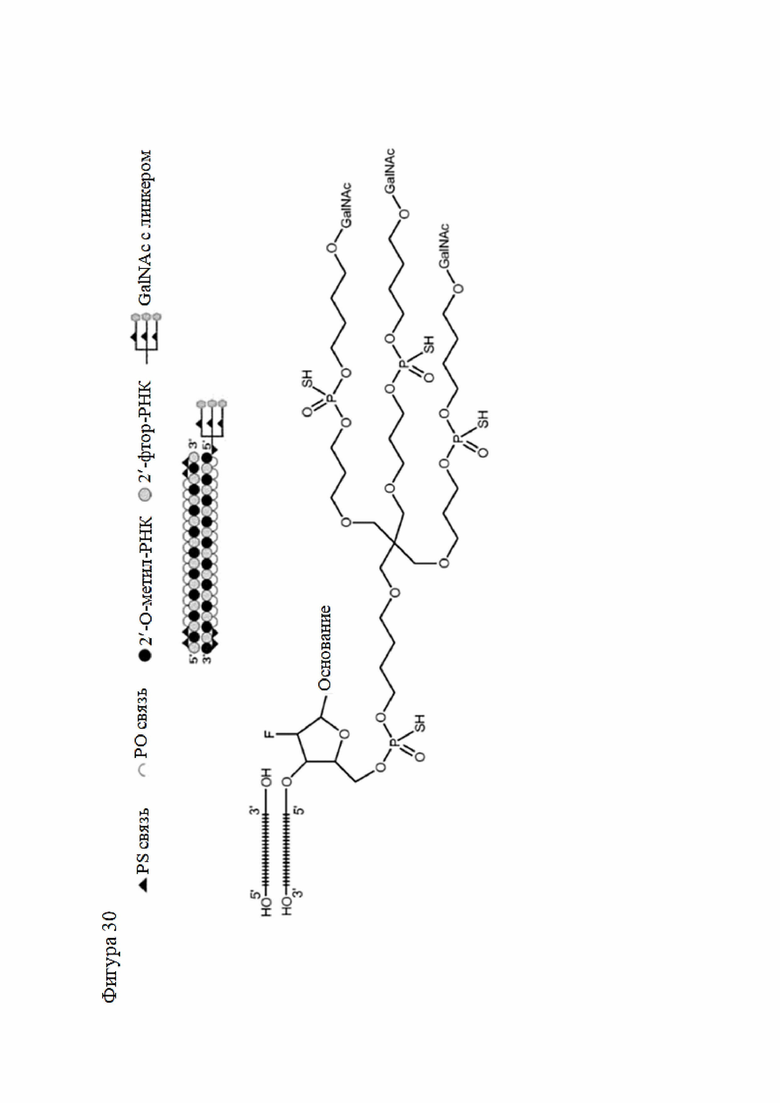

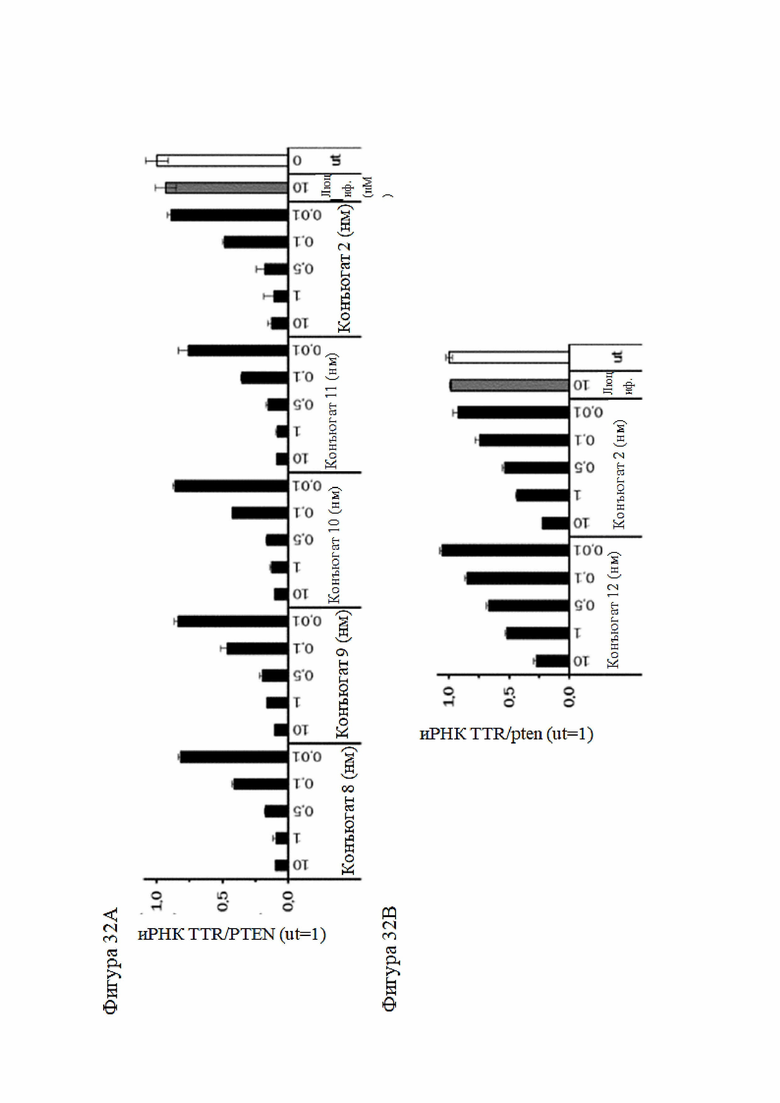

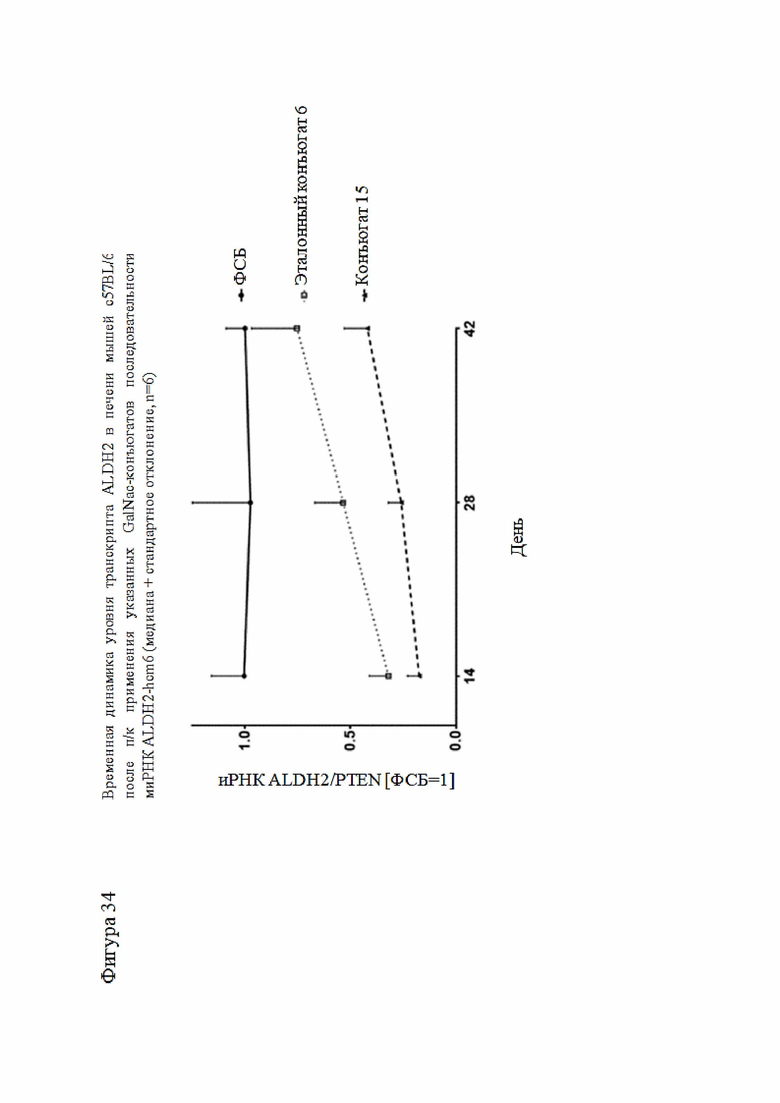
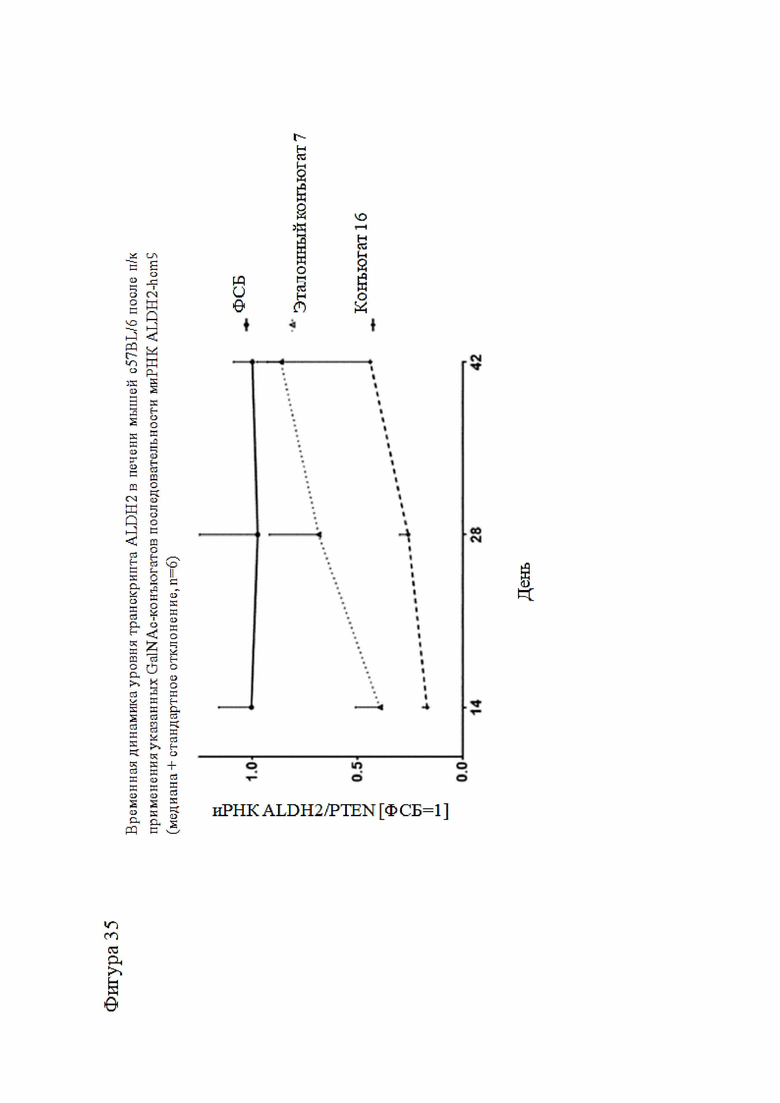
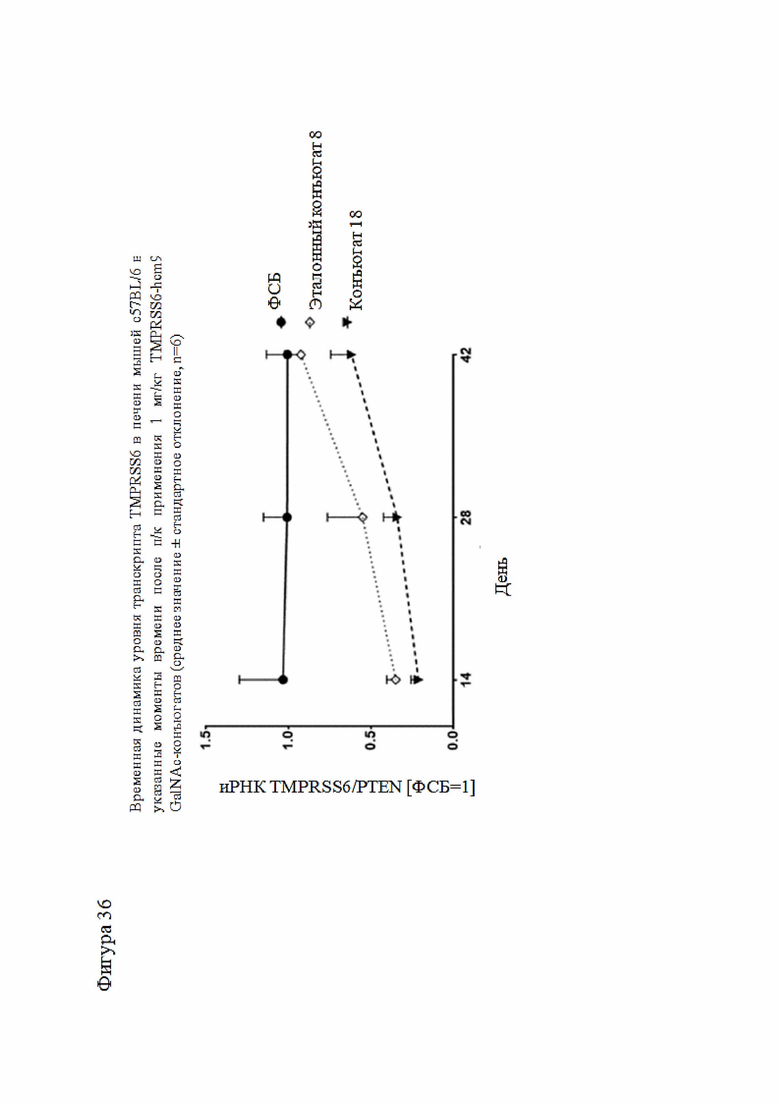

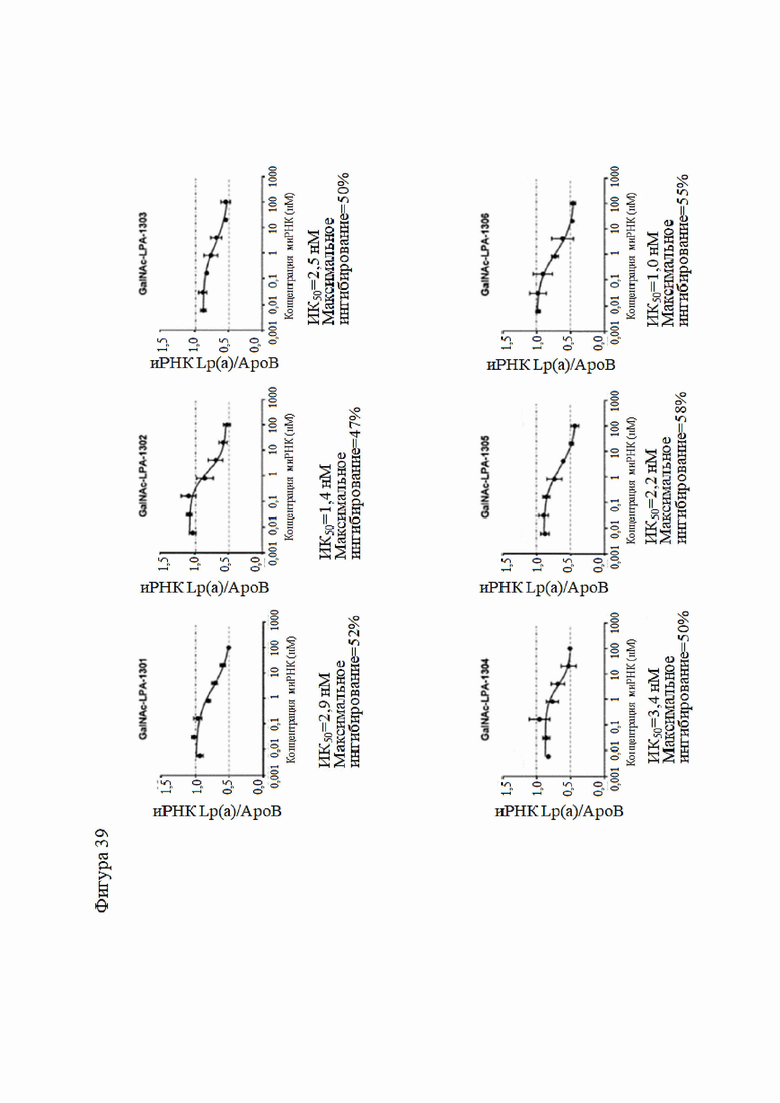
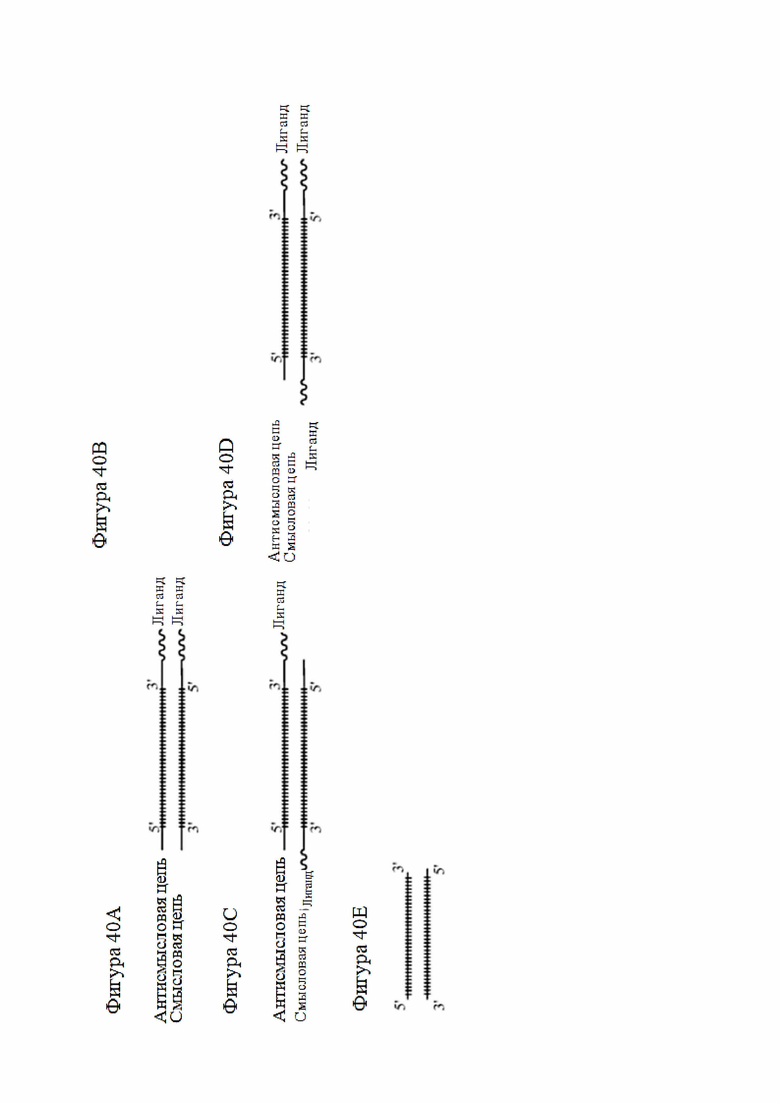
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая нуклеиновую кислоту для ингибирования экспрессии LPA в клетке, конъюгат для ингибирования экспрессии LPA в клетке (варианты), композицию для предотвращения, или лечения, или уменьшения риска развития сердечно-сосудистого заболевания и применение нуклеиновой кислоты, или конюгата, или композиции для предотвращения, или лечения, или уменьшения риска развития сердечно-сосудистого заболевания. В одном варианте реализации нуклеиновая кислота содержит по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна указанной первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA. Изобретение расширяет арсенал средств для ингибирования экспрессии LPA в клетке. 5 н. и 21 з.п. ф-лы, 40 ил., 15 табл., 15 пр.
1. Нуклеиновая кислота для ингибирования экспрессии LPA в клетке, содержащая по меньшей мере одну дуплексную область, которая содержит по меньшей мере часть первой цепи и по меньшей мере часть второй цепи, которая по меньшей мере частично комплементарна указанной первой цепи, причем указанная первая цепь по меньшей мере частично комплементарна по меньшей мере части РНК, транскрибированной с гена LPA, при этом указанная первая цепь содержит нуклеотидную последовательность SEQ ID NO: 9,
где часть первой цепи в дуплексной области содержит по меньшей мере 10 нуклеотидов SEQ ID NO: 9 и где часть второй цепи комплементарна на 70-100% части первой цепи.
2. Нуклеиновая кислота по п. 1, где указанная часть первой цепи в дуплексной области содержит по меньшей мере 12 нуклеотидов SEQ ID NO: 9, предпочтительно по меньшей мере 14 нуклеотидов SEQ ID NO: 9.
3. Нуклеиновая кислота по п. 1 или 2, где указанная часть первой цепи в дуплексной области содержит по меньшей мере 16 нуклеотидов SEQ ID NO: 9, предпочтительно по меньшей мере 18 нуклеотидов SEQ ID NO: 9.
4. Нуклеиновая кислота по любому из предшествующих пунктов, где указанная часть первой цепи в дуплексной области содержит все нуклеотиды SEQ ID NO: 9.
5. Нуклеиновая кислота по любому из предшествующих пунктов, отличающаяся тем, что указанная вторая цепь содержит нуклеотидную последовательность SEQ ID NO: 10.
6. Нуклеиновая кислота по любому из предшествующих пунктов, отличающаяся тем, что указанная первая цепь составляет 19-35 нуклеотидов в длину и/или указанная вторая цепь составляет 17-35 нуклеотидов в длину.
7. Нуклеиновая кислота по любому из предшествующих пунктов, отличающаяся тем, что указанная по меньшей мере одна дуплексная область состоит из 17-25, предпочтительно 19-25, последовательных пар нуклеотидных оснований.
8. Нуклеиновая кислота по любому из предшествующих пунктов, отличающаяся тем, что указанная нуклеиновая кислота:
a) имеет тупые концы на обоих концах; или
b) имеет липкий конец на одном конце и тупой конец на другом; или
c) имеет липкий конец на обоих концах.
9. Нуклеиновая кислота по любому из предшествующих пунктов, отличающаяся тем, что один или более нуклеотидов на указанных первой и/или второй цепях модифицированы с образованием модифицированных нуклеотидов.
10. Нуклеиновая кислота по любому из предшествующих пунктов, отличающаяся тем, что указанная нуклеиновая кислота содержит фосфотиоатную связь между одним, двумя или тремя концевыми 3'-нуклеотидами и/или 5'-нуклеотидами одного или обоих концов указанных первой и/или второй цепей.
11. Конъюгат для ингибирования экспрессии LPA в клетке, содержащий нуклеиновую кислоту по любому из предшествующих пунктов, конъюгированную с лигандом.
12. Конъюгат по п. 11, отличающийся тем, что указанный лиганд содержит (i) одну или более групп N-ацетилгалактозамина (GalNAc) или их производных и (ii) линкер, причем с помощью указанного линкера по меньшей мере одна группа GalNAc или его производное конъюгированы с указанной нуклеиновой кислотой.
13. Конъюгат по любому из пп. 11, 12, отличающийся тем, что лиганд содержит соединение формулы (I)
[S-X1-P-X2]3-A-X3-, (I)
в которой S представляет собой сахарид, при этом предпочтительно указанный сахарид представляет собой N-ацетилгалактозамин;
X1 представляет собой C3-C6 алкилен или (-CH2-CH2-O)m(-CH2)2-, где m равно 1, 2 или 3;
P представляет собой фосфат или модифицированный фосфат, предпочтительно тиофосфат;
X2 представляет собой алкилен или простой алкиленовый эфир формулы (-CH2)n-O-CH2-, где n=1-6;
А представляет собой блок разветвления;
X3 представляет собой мостиковый блок;
при этом нуклеиновая кислота по любому из пп. 1-8 конъюгирована с X3 за счет фосфата или модифицированного фосфата, предпочтительно тиофосфата.
14. Конъюгат по п. 11 или 12, отличающийся тем, что указанная первая цепь представляет собой РНК и представляет собой соединение формулы (X)
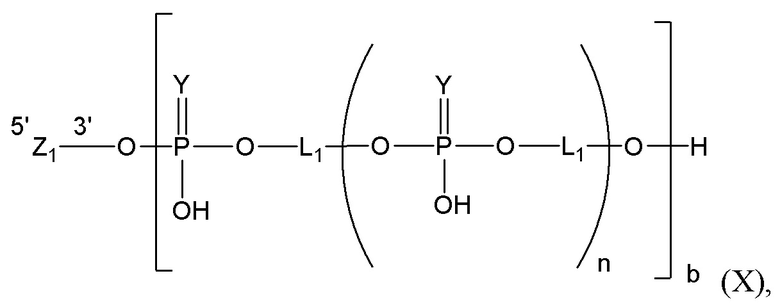
в которой b равно 0 или 1; и
указанная вторая цепь представляет собой РНК и представляет собой соединение формулы (XI)
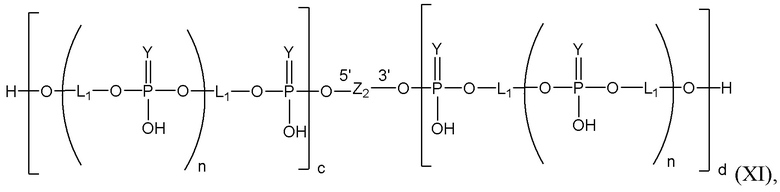
в которой с и d независимо равны 0 или 1;
Z1 и Z2 представляют собой части РНК указанных первой и второй цепей РНК соответственно;
Y представляет собой O или S;
n равно 0, 1, 2 или 3; и
L1 представляет собой линкер, к которому присоединен лиганд; и
при этом b+c+d равно 2 или 3.
15. Нуклеиновая кислота по п. 1, где указанная первая цепь состоит из нуклеотидной последовательности SEQ ID NO: 9, и при этом необязательно указанная вторая цепь содержит и предпочтительно состоит из нуклеотидной последовательности SEQ ID NO: 10.
16. Конъюгат для ингибирования экспрессии LPA в клетке, содержащий нуклеиновую кислоту по п. 15, конъюгированную с лигандом.
17. Конъюгат по п. 11 или 16, отличающийся тем, что конъюгат имеет следующую структуру:

где Z представляет собой нуклеиновую кислоту по п. 1 или 15.
18. Конъюгат по п. 17, отличающийся тем, что указанная нуклеиновая кислота содержит две фосфотиоатные связи между каждым из трех 3'-концевых и между каждым из трех 5'-концевых нуклеотидов на указанной первой цепи и две фосфотиоатные связи между тремя концевыми нуклеотидами 3'-конца указанной второй цепи, и при этом указанный лиганд конъюгирован с 5'-концом указанной второй цепи.
19. Конъюгат по п. 17, отличающийся тем, что первая цепь представляет собой РНК и представляет собой соединение формулы (XV)
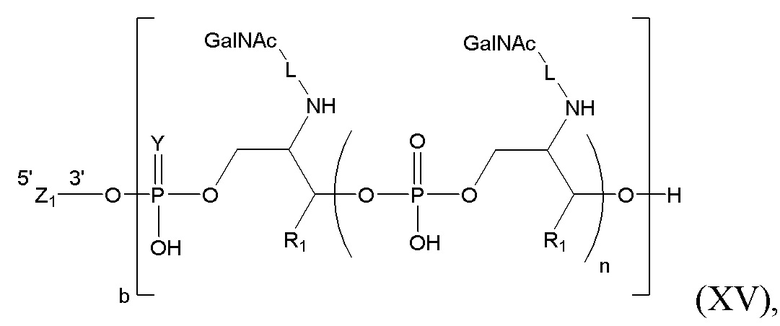
в которой b равно 0 или 1; и
указанная вторая цепь представляет собой РНК и представляет собой соединение формулы (XVI)
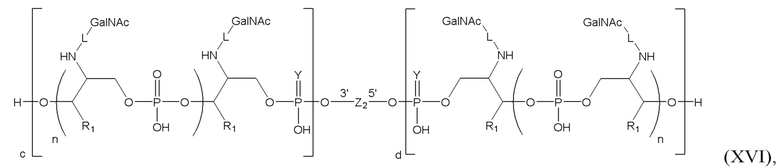
в которой с и d независимо равны 0 или 1;
при этом Z1 и Z2 представляют собой части РНК указанных первой и второй цепей РНК соответственно;
Y представляет собой O или S;
R1 представляет собой H или метил;
n равно 0, 1, 2 или 3; и
L является одинаковым или различным в формулах (XV) и (XVI) и выбран из группы, состоящей из:
-(CH2)q, где q=2-12;
-(CH2)r-C(O)-, где r=2-12;
-(CH2-CH2-O)s-CH2-C(O)-, где s=1-5;
-(CH2)t-CO-NH-(CH2)t-NH-C(O)-, где t независимо представляет собой 1-5;
-(CH2)u-CO-NH-(CH2)u-C(O)-, где u независимо представляет собой 1-5; и
-(CH2)v-NH-C(O)-, где v представляет собой 2-12; и
при этом концевой C(O), если он присутствует, присоединен к группе NH;
и при этом b+c+d равно 2 или 3.
20. Нуклеиновая кислота по любому из пп. 1-10 или 15 или конъюгат по любому из пп. 11-14 или 16-19, содержащие последовательность и модификации, представленные ниже:
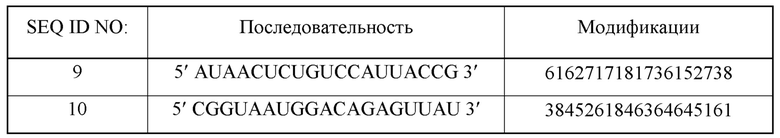
где конкретные модификации обозначены номерами
1=2'-F-dU,
2=2'-F-dA,
3=2'-F-dC,
4=2'-F-dG,
5=2'-OMe-rU;
6=2'-OMe-rA;
7=2'-OMe-rC;
8=2'-OMe-rG.
21. Нуклеиновая кислота по любому из пп. 1-10 или 15 или конъюгат по любому из пп. 11-14 или 16-19, отличающиеся тем, что нуклеотиды в положениях 2 и 14 с 5'-конца указанной первой цепи модифицированы с применением 2'-фтор-модификации и нуклеотиды на указанной второй цепи, которые соответствуют положению 11 или 13, или 11 и 13, или 11-13 указанной первой цепи модифицированы с применением 2'-фтор-модификации.
22. Композиция для предотвращения, или лечения, или уменьшения риска развития сердечно-сосудистого заболевания, где указанная композиция содержит фармацевтически эффективную дозу нуклеиновой кислоты или конъюгата по любому из пп. 1-21.
23. Композиция по п. 22, дополнительно содержащая средство для доставки, предпочтительно липосомы, и/или физиологически приемлемое вспомогательное вещество, и/или носитель, и/или разбавитель, и/или буфер, и/или консервант.
24. Композиция по п. 22 или 23, где указанное сердечно-сосудистое заболевание представляет собой инсульт, атеросклероз, тромбоз, ишемическую болезнь сердца или стеноз аорты и/или любое другое заболевание или патологию, ассоциированные с повышенными уровнями частиц, содержащих Lp(a).
25. Применение нуклеиновой кислоты или конюгата по любому из пп. 1-21 или композиции по любому из пп. 22-24 для предотвращения, или лечения, или уменьшения риска развития сердечно-сосудистого заболевания.
26. Применение по п. 25, где указанное сердечно-сосудистое заболевание представляет собой инсульт, атеросклероз, тромбоз, ишемическую болезнь сердца или стеноз аорты и любое другое заболевание или патологию, ассоциированные с повышенными уровнями частиц, содержащих Lp(a).
| WO 2017156012 A1, 14.09.2017 | |||
| WO 2005000201 A3, 23.03.2006 | |||
| EP 1417351 A4, 01.06.2005 | |||
| WO 2017059223 A2, 06.04.2017 | |||
| НОВАЯ ЛИЗОФОСФОЛИПИД-АЦИЛТРАНСФЕРАЗА | 2010 |
|
RU2534559C2 |

