Область техники, к которой относится изобретение
Настоящее изобретение относится к моноклональным антителам, способным распознавать и связывать белок РВР2а и другие белки, презентирующие последовательности, гомологичные РВР2а, включая патогены устойчивого к метициллину золотистого стафилококка (Staphylococcus aureus) - MRSA (МРЗС), коагулазонегативного стафилококка, стафилококка Staphylococcus sciuri, энтерококка (Enterococcus spp.) и любых других бактерий, обладающих РВР2а или последовательности, гомологичные этому белку.
Изобретение также относится к применению моноклональных антител, способных распознавать и связывать белок РВР2а и другие белки, презентирующие последовательности, гомологичные РВР2а, в иммунодиагностике комплементарных структур для обнаружения устойчивости к бета-лактамам.
Уровень техники
Инфекции, причиной которых является метициллин-резистентный штамм Staphylococcus aureus (MRSA), представляют главную проблему для врачей-клиницистов, так как вызываемая им заболеваемость и смертность выше, чем в результате инфицирования метиллин-чувствительными стафилококками Staphylococci (1). Помимо этого эти инфекции служат причиной более продолжительного пребывания в больнице и более высоких трат на антибактериальные препараты, что ведет к более высокой стоимости лечения пациентов, инфицированных этим патогеном (1).
Ванкомицин был противомикробным препаратом первого выбора для лечения инфекций, вызванных MRSA. Однако с учетом растущего числа выделенных штаммов MRSA в группах населения в Соединенных Штатах и в Австралии (2, 3, 4), наряду с идентификацией штаммов MRSA с промежуточным уровнем устойчивости к ванкомицину в Японии, Соединенных Штатах (5) и Бразилии (6), являются причиной того, что картина современного состояния становится более серьезной. Описание в 2004 году MRSA штаммов, полностью резистентных (устойчивых) к ванкомицину (7), вызвало колоссальное беспокойство в научно- медицинском сообществе. В настоящее время MRSA является серьезным кандидатом на звание внушающего страх "суперинфекта или супербактерии" - патогена, резистентного ко всем имеющимся в настоящее время лекарствам.
В целом в последние десятилетия коэффициент распространенности MRSA (отношение инфекций, вызываемых MRSA, к общему числу инфекций, вызываемых S. aureus) среди больничных инфекций постепенно увеличивается. В исследовании, проведенном Jarvis et al., включающем 1268 ICU (отделений интенсивной терапии) в 337 госпиталях в Соединенных Штатах, указывается, что число инфицированных MRSA в ICU увеличилось с 660 до 2184, а коэффициент распространенности с 35% повысился до 64.4% (8). В Японии повышение коэффициента распространенности больничных (госпитальных) инфекций (HI), вызванных MRSA, от 60% до 90% (9) может вызвать тревогу. В исследованиях, проведенных в Соединенных Штатах, изменение значения перцентили составляет от 2% в 1974 до 50% в 1997 (10, 11), а в некоторых госпиталях в Соединенных Штатах более 80% HI вызвано MRSA (12). В Англии между 1989 и 1995 годом коэффициент распространенности увеличился с 1.5% до 15.2% и в настоящее время (2004) оценивается в 41.5% (13).
Полагают, что помимо высоких коэффициентов распространенности, в особенности в обучающих и в больших госпиталях, MRSA является главным патогеном, вызывающим эпидемические вспышки в больницах Бразилии (14). В 1986 году более 50% штаммов S. aureus больничного происхождения, выделенных у больных университетского госпиталя в Сан Паулу, были резистентны к метициллину, а в 1993 году частота новых случаев MRSA инфекции в Детском госпитале Медицинской школы Паулиста (Paulista) составляла 70% (15). В исследовании, проведенном в госпиталях Белу-Оризонте (Belo Horizonte), Resende et al. (16) указывают распространенность MRSA 71%.
Штаммы MRSA презентируют пенициллин - связывающий белок с очень низкой аффинностью к противомикробным препаратам класса бета-лактамов, таким как РВР2а (17). В присутствии этого фермента, который кодируется геном mecA, бактерия успешно синтезирует пептидогликан даже в присутствии бета-лактамов. Этот фермент может также находиться в коагулазонегативном штамме Staphylococcus и в Staphylococcus sciuri - бактерии, присутствующей в нормальной флоре собак. Помимо резистентности к бета-лактамам, больничные штаммы MRSA проявляют резистентность к большинству других существующих классов противомикробных препаратов, при этом гликопептиды (ванкомицин и тейкопланин) продолжают оставаться препаратами первого выбора.
Два исследования с применением ДНК-вакцины против РВР2а показали, что этот белок является иммуногенным и что вырабатываемая иммунная реакция способна обеспечить защиту против MRSA в анализах, проводившихся на мышиной модели (18, 19). Известно, однако, что при больничных инфекциях большинство больных являются иммунодепрессированными (20). В этих случаях вакцина не способна обеспечить своевременную выработку организмом защитных антител для контроля бактериальной инфекции.
Иммуногенность РВР2а
Согласно классификации Goffin and Ghuysen's (40) РВР2а представляет собой мультимодульный фермент класса II. Этот фермент с молекулярной массой 76 килодальтон составляют мембраносвязанную область, не-транспептидазный домен и транспептидазный домен, содержащий активное ядро из 4 аминокислот (STQK), отвечающее за реакции транспептидации в бактериях (20 bis. Ryfell, 1990).
Исследования с применением ДНК-вакцины против РВР2а в предыдущем уровне техники показывают, что результаты бактериального восстановления (количественные показатели почек) у иммунизированных животных, подвергнувшихся контрольному заражению системным инфицированном, составляли от 3 до 4 раз в работе Ohwada et al. и 1000 раз в работе Senna et al. Авторы этих исследований применяли полную последовательность (за исключением области фиксации мембран) гена mecA и внутренний фрагмент транспептидазного домена, соответственно.
Но, в соответствии с вышеуказанным, вакцина не способна обеспечить своевременную выработку антител для контроля бактериальной инфекции. Таким образом, в случае инфицирования с помощью MRSA применение моноклональных антител против РВР2а является наиболее походящей терапией для лечения этих инфекций.
Раскрытие изобретения
Основной задачей настоящего изобретения является предоставление моноклональных антител, способных узнавать (распознавать) и связывать белок РВР2а (SEQ ID NO:1) и другие белки, представляющие последовательности, гомологичные РВР2а, включая патогены устойчивого к метициллину золотистого стафилококка (Staphylococcus aureus) - MRSA (MP3C), коагулазонегативного стафилококка, стафилококка Staphylococcus sciuri, энтерококка (Enterococcus spp.) и некоторых других бактерий, продуцирующих РВР2а или последовательности, гомологичные этому белку.
Другой задачей изобретения является применение моноклональных антител, способных узнавать (распознавать) и связывать белок РВР2а и другие белки, презентирующие последовательности, гомологичные РВР2а, в иммунодиагностике комплементарных структур для обнаружения устойчивости к бета-лактамам.
Моноклональные антитела по настоящему изобретению представлены последовательностями SEQ ID NO:6, SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:15, SEQ ID NO:16 и SEQ ID NO:17.
Краткое описание фигур
На Фигуре 1 показаны результаты иммуноферментного анализа (ИФА, ELISA) сыворотки животных, иммунизированных с целью продуцирования антител против РВР2а.
На Фигуре 2 показаны результаты электрофореза в полиакриламидном геле сырых (неочищенных) образцов и образцов после очистки.
На Фигуре 3 показаны результаты иммуноблоттинга лизатов MRSA и MSSA в сопоставлении с супернатантом, содержащим моноклональные антитела против РВР2а.
На Фигуре 4 представлены результаты проточной цитометрии для бактерий MRSA (СЕВ) и MSSA, инкубированных с моноклональным антителом против РВР2а и помеченных фикоэритрином (РЕ).
На Фигуре 5 показан тест in vitro на защитные свойства (иммунизацию) (MIC (МИК) - минимальная ингибирующая концентрация), сообщаемые очищенным моноклональным антителом против РВР2а и ванкомицином, против инокулума (инокулята) различных штаммов MRSA.
На Фигуре 6А показаны результаты почечной пробы для пролеченных и не пролеченных моноклональным антителом против РВР2а животных, подвергнутых системному заражению сублетальной дозой штамма MRSA СЕВ.
На Фигуре 6B показаны результаты почечной пробы для пролеченных и не пролеченных моноклональным антителом против РВР2а животных, подвергнутых системному заражению сублетальной дозой Иберийского штамма MRSA (Европейский эпидемический клон).
На Фигуре 6С показаны результаты почечной пробы для пролеченных и не пролеченных моноклональным антителом против РВР2а животных, подвергнутых системному заражению сублетальной дозой штамма WB79 СА- MRSA (Бразильский штамм).
На Фигуре 7А показана кривая выживаемости пролеченных (иммунизированных) и непролеченных (контрольных) животных после заражения летальной дозой бактерий (MRSA СЕВ).
На Фигуре 7B показана кривая выживаемости пролеченных (иммунизированных) и непролеченных (контрольных) животных после заражения летальной дозой бактерий (Иберийский штамм MRSA).
На Фигуре 7С показана кривая выживаемости пролеченных (иммунизированных) и непролеченных (контрольных) животных после заражения летальной дозой бактерий (WB79 СА-MRSA).
На Фигуре 8А показаны результаты количественного определения бактерий в почках животных, пролеченных моноклональным антителом против РВР2а, ванкомицином и антителом + ванкомицином, и непролеченных животных после заражения их бактериями (MRSA СЕВ).
На Фигуре 8B показаны результаты количественного определения бактерий в почках животных, пролеченных моноклональным антителом против РВР2а.
На Фигурах 9 показано взаимодействие между рекомбинантным РВР2а (антиген) и моноклональными антителами клон 38 (Фигура 9А) и клон 10 (Фигура 9B).
На Фигуре 10 представлен график (кривая) второго анализа методом проточной цитометрии образцов MRSA в присутствии FITC- меченного антитела против РВР2а.
На Фигуре 11 представлены результаты in vitro иммунизации (защиты), обеспечиваемой антителом против штамма энтерококков VRE (устойчивого к ванкомицину).
На Фигуре 12 показана LD50 и летальная доза для определения интраперитонеальной инфекции.
На Фигуре 13 показаны данные по эффективности in vivo иммунизации моноклональным антителом к РВР2а и РВР5 для защиты против системного заражения энтерококками.
На Фигуре 14 показаны количественные результаты почечной пробы в тесте in vivo иммунизации - системное инфицирование интраперитонеальным путем мышиной модели ванкомицин-резистентным штаммом Enterococcus faecium.
Осуществление изобретения
Увеличение числа инфекций, вызываемых бактериями, мультирезистентными к противомикробным препаратам, вызывает тревогу. Распространенность больничных инфекций, вызванных бактериями MRSA, повысила заболеваемость во всем мире и является причиной высоких расходов вследствие интенсивного применения антибиотиков и более продолжительного периода пребывания пациента в больнице (24). Химиотерапевтическое лечение проявляет признаки исчерпания, так как на рынке появляется очень мало действительно новых и эффективных лекарств (25).
В связи с этим появляется (возвращается) пассивная иммунотерапия как весьма перспективная альтернатива, имеющая ряд предпочтительных особенностей (признаков) по сравнению с обычной химиотерапией: среди них пониженная токсичность, повышенный период полужизни в плазме (что облегчает лечение с применением пониженной дозы и, возможно, обеспечивает более низкую конечную стоимость лечения), и, в особенности, в случае представленного в данном описании продукта, селективная токсичность, рекомендуемая Paul Ehrlich (когда лекарство селективно устраняет вышеупомянутый патоген) (25а). Этот последний признак особенно применим для предупреждения так называемого давления отбора, приводимого в действие антимикробными препаратами широкого спектра действия - который вызывает появление мультирезистентных бактерий (26).
Для разработки этого продукта заявители в качестве мишени выбрали РВР2а на основании предварительных исследований с применением ДНК-вакцины на мышиной модели (18, 19, 21) и анализов иммуноструктурных свойств модели. Структура РВР2а была изучена в 2002 году (29) и депонирована в PDB (Банке данных белков) под кодовым названием Iwmr.
В отличие от других подходов, заявители работали с внутренней областью молекулы из 76 аминокислот, которые фланкируют активное ядро фермента (SxxK), в транспептидазном домене. Анализы по идентификации эпитопа проводили на молекуле и ее презентации различным класса II СНМ аллелям (программа Tepitope), с последующей проверкой (верификацией) их локализации в молекуле (программа SPDB-Viewer). Такой подход позволяет нам оценить легкость доступа антитела к этим мишеням в нативной молекуле.
Этот компьютерный анализ показал наличие эпитопов близ активного ядра (центра) фермента с высокой степенью узнавания СНМ аллелями класса II, локализованными на поверхности РВР2а (данные не показаны).
Этот предварительные данные компьютерного моделирования (in silico) были подтверждены с помощью тестов по распознаванию мишени (иммуноблоттинга и проточной цитометрии), и антитело, связывающееся с этой областью РВР2а, показало свою способность предоставить высокую степень защиты (иммунизации), продемонстрированную результатами, полученными в проведенных in vitro и in vivo тестах. Было показано, что антитела, образуемые с применением фрагмента из 76 аминокислот, сообщают более высокую степень защиты (иммунизации), чем антитела, вырабатываемые при использовании полного РВР2а, как ранее сообщалось в работах Ohwada и Senna и как подтверждено результатами, полученными заявителями.
Эпидемиология инфекций, вызванных MRSA, показывает присутствие преобладающих клональных типов, ответственных за вспышки и эпидемии в больницах по всему миру. Эти клоны показывают повышенную способность к колонизации и вирулентность по сравнению с неэпидемическими штаммами MRSA (30).
Для подтверждения полученных результатов мы запланировали работу с различными MRSA клонами (эпидемическими клонами), характерными для большинства инфекций (особенно больничных), вызванных MRSA.
Штамм СЕВ (Бразильский эпидемический клон), отвечающий за большинство инфекций, вызываемых MRSA в больницах Бразилии (31), был также идентифицирован в других странах Южной Америки (32, 33), в Португалии и Чехословакии (34). Европейский эпидемический клон (Иберийский MRSA) обнаружен в Европейских странах и в Соединенных Штатах (35).
В связи с увеличением числа инфекций в локальных сообществах, вызванных MRSA, один из этих штаммов (WB79 СА-MRSA), выделенный в Бразилии, был включен в настоящие испытания. Эти испытания выявляют дифференцированные признаки по сравнению с признаками, обнаруженными в госпиталях, с более высокой вирулентностью (наличие лейкоцидина Пантона-Валентина (Panton-Valentine)) и профилем резистентности к противомикробным препаратам, отличным от профиля больничных MRSA штаммов (36). Недавно в Соединенных Штатах была идентифицирована вспышка инфекции СА-MRSA в популяции мужчин, практикующих секс с другими мужчинами (37). С этого момента мы можем говорить, что инфекции, вызываемые MRSA, приобретают характер STD (характер заболевания, передаваемого половым путем).
Результаты иммунизации (защиты) путем введения антитела против РВР2а показали эффективность против различных тестированных клонов, которая заставила нас поверить в их применимость против инфекций (локальной (общинной) или больничной), вызванной MRSA любого типа.
Некоторые важные свойства (признаки, особенности) продукта по настоящему изобретению необходимо прокомментировать.
(i) Результаты in vitro иммунизации заставляют поверить, что предусматриваемый механизм действия-блокада области близ активного ядра молекулы-достаточен для ингибирования роста и размножения бактерий. Аналогично механизму действия бета-лактамов, этот механизм является динамическим (функцией времени), нуждающимся в том, чтобы бактерия в фазе размножения подставляла, открывала мишень для действия лекарства.
(ii) Этому свойству способствуют такие фармакокинетические свойства антител, как продолжительный период полужизни в плазме (38). Это означает введение меньшего числа доз по сравнению с лечением антибиотиками, что сделает лечение более простым для пациента и, как следствие, возможно, уменьшит конечную стоимость. Эти предположения были подтверждены на практике в сравнительном анализе иммунизации ванкомицином (см. ниже).
(iii) Ингибирование in vitro роста бактерий также означает, что антителу не требуются традиционные дополнительные механизмы реакции антиген-антитело, такие как опсонизация и активация комплемента. С учетом того, что большинство больничных инфекций поражает больных в состоянии иммунодепрессии (иммуносупрессии, состоянии угнетения иммунитета), это свойство продукта является чрезвычайно важным.
(iv) Известно, что золотистый стафилококк (Staphylococcus aureus) представляет (являет) белок А на своей поверхности. Эта молекула характеристически связывается с областью Fc иммуноглобулинов, препятствуя опсонизирующей активности иммунной системы (38). На основании результатов наших испытаний мы полагаем, что концентрация введенного антитела могла подавить белок А, имеющийся на поверхности бактерий, не предотвращая блокаду РВР2а вводимыми антителами.
Системные инфекции, вызываемые MRSA, лечили гликопептидами, в особенности ванкомицином. Однако это лекарство имеет ряд побочных эффектов, среди которых иммунотоксичность (42), ототоксичность, нефротоксичность и транзиторная нейтропения (43), и требует применения в течение продолжительных периодов времени, что делает окончательную стоимость лечения высокой.
Используя мышиную модель инфицирования (заражения), мы могли сравнить лечение ванкомицином с лечением моноклональным антителом и комбинацией этих двух лекарств. Полученные результаты показали, что доза антитела оказывает действие, аналогичное или более сильное, чем 5 доз ванкомицина, и что одновременное введение двух лекарств намного более эффективно устраняет бактерии, чем изолированная терапия. Эти результаты являются чрезвычайно важными, так как позволяют предложить более дешевое лечение антителом и более эффективное выздоровление (восстановление) пациентов в тяжелом состоянии в результате введения ванкомицина и антитела.
Помимо применения моноклональных антител против РВР2а для лечения инфекций, вызванных MRSA, настоящее изобретение рассматривает еще следующие области применения:
(i) результаты распознавания моноклональным антителом против РВР2а белка с молекулярной массой, близкой молекулярной массе РВР5 штамма Enterococcus sp., указывают на то, что с помощью введения этого антитела можно осуществить иммунизацию (защитную реакцию) против этого патогена. РВР5 гомологичен РВР2а, причем оба белка относятся к мультимодульным РВР класса В с низкой аффинностью к бета-лактамам (40). Энтерококки представляют собой бактерии, которые вызывают тяжелые больничные инфекции, проявляющие высокую имманентную (собственную) и приобретенную устойчивость (резистентность) к противомикробным препаратам. Штаммы, резистентные к ванкомицину (VRE), представляют собой хранилище (источник) генов резистентности к гликопептидам и могут быть перенесены на другие патогены (41).
(ii) эти антитела могут найти применение для идентификации РВР2а с помощью иммунодиагностических тестов. Например, применение иммунных тестов с использованием агглютинации частиц латекса, связанных с антителом, через несколько (немного) часов может дать результат, который позволит предсказать резистентность ко всем бета-лактамам. В обычных тестах на чувствительность к противомикробным препаратам (таким как тест на чувствительность к антибиотикам) эти результаты можно получить только через 12-24 часов.
(iii) если моноклональные антитела успешно используются для местного применения против хронических язв (инфликсимаб, см. ссылку Streit et al. и Международную заявку на патент WO 1999041285 A1), эти антитела можно применять топически (местно) для того, чтобы вызвать деколонизацию назальных MRSA, и для лечения повреждений кожи, вызванных этим патогеном.
С учетом этих сведений и результатов, обнаруженных в уровне техники, заявители (авторы настоящего изобретения) осуществляли иммунизацию животных ДНК-вакциной с конструкциями, соответствующими гену mecA (SEQ ID NO:2) за исключением области фиксации мембран, и иммунизацию содержащей 76 аминокислот внутренней областью транспептидазного домена (SED ID NO:3), содержащей активное ядро фермента.
Иммунизацию проводили в тех же самых условиях, которые описаны в работах Ohwada et al. и Senna et al, разработавших ДНК-вакцину против РВР2а, используя полную последовательность (за исключением области фиксации мембран) гена mecA и, и внутренний фрагмент транспептидазного домена. Иммунизацию проводили четырьмя дозами, и иммунизированных животных, и неиммунизированную контрольную группу заражали (контрольное заражение) путем введения системной инфекции MRSA и определяли число бактерий в почках животных через два отдельных периода. Полученные результаты показывают, что иммунизация транспептидазным фрагментом вызвала большее снижение числа бактерий в почках животных, чем иммунизация геном mecA.
Следовательно, мы видим, что антитела, вырабатываемые против транспептидазного фрагмента, обеспечивают лучшую защиту (иммунизацию) в применяемых условиях (ДНК-вакцина на мышиной модели), чем антитела против полной последовательности РВР2а. Транспептидазный фрагмент включает активное ядро фермента STQK (SEQ ID NO:4).
Поэтому на основании полученных результатов заявители создали изобретение, направленное на получение моноклональных антител, способных распознавать и связывать РВР2а белок и другие белки, имеющие последовательности, гомологичные белку РВР2а, и на использование транспептидазного фрагмента, который включает активное ядро фермента.
Далее изобретение описывается в примерах, которые не следует рассматривать как ограничивающие.
Пример 1
МАТЕРИАЛЫ И МЕТОДЫ
1. Бактерии
Использовались следующие метициллин-резистентные штаммы Staphylococcus aureus (золотистого стафилококка): berian-MRSA, COL-MRSA (предоставленный Unite des Agents Antimicrobiens - Institut Pasteur [Отделением противомикробных агентов - Институт Пастера], Dr. Patrice Courvalin), WB79 CA-MRSA, и СЕВ-MRSA (предоставленные Dr. Agnes Figueiredo, Institute de Microbiologic [Институт микробиологии] UFRJ); ванкомицин-резистентный штамм Enterococcus faecalis; и чувствительный к метициллину штамм Staphylococcus aureus (MSSA). Также в качестве контрольных использовали штаммы Escherichia coli BL21 DE3 (Novagen) и TOP 10 (Invitrogen).
2. Животные
Для иммунизации и для in vivo анализа иммунитета брали самок мышей Balb/C, в возрасте 4-8 недель, полученных из Центра CECAL-FIOCRUZ и помещенных в лабораторию LAEAN-BioManguinhos.
3. Иммунизация
Мыши (4 животных) получали начальную дозу 100 микрограмм pCI-Neo плазмиды: фрагмента mecA гена MRSA (18), а затем, через 14 дней, дозу 10 микрограмм - очищенного рекомбинантного белка, соответствующего внутренней области MRSA РВР2а (21), эмульгированного в неполном адъюванте Фрейнда. Через четырнадцать дней животные, у которых обнаружен наилучший иммунный ответ (по оценке иммуноферментным анализом - ELISA (ИФА)), получали внутривенно дозу 10 микрограмм очищенного белка, разведенного в PBS (фосфатно-солевом буферном растворе). Через три дня после IV инъекции животных подвергали эвтаназии (асфиксия в атмосфере CO2) (CEUA L0009-07 Protocol - FIOCRUZ), и в асептических условиях удаляли селезенку, чтобы использовать для слияния клеток. Сыворотку животного с наилучшим иммунным ответом использовали в качестве позитивного контроля для других иммунологических тестов.
4. Получение моноклональных антител
Лимфоциты, удаленные из селезенки, подвергали слиянию с миеломными клетками SP2/0-Ag14 (ATCC 1581), используя для слияния полиэтиленгликоль, и культивировали в среде, содержащей гипоксантин-аминоптерин-тимидин, при 37°С в атмосфере 10% СО2, в соответствии с протоколом получения моноклональных антител в Current Protocols in Immunology (22). Полученные гибридомы проверяли методом ИФА (тест ELISA) через 14 дней, используя в качестве антигена очищенный рекомбинантный белок, как описано ниже. Лучшие гибридомы клонировали, отбирая лучшие клоны методом ELISA, и эти клоны хранили в жидком азоте.
5. Иммуноферментный анализ - ELISA (ИФА)
98-луночные пластиковые планшеты Maxisorb сенсибилизировали, добавляя 500 нанограмм/лунка рекомбинированного белка (фрагмента РВР2а) в карбонат/бикарбонатном буфере и инкубировали при 4°С в течение ночи. На следующий день планшеты трижды отмывали в PBS, содержащем 0.05% Tween (Твин) 20 и блокировали в PBS и 5% обезжиренном молоке в течение двух часов при 37°С. Использовали образцы для анализа: сыворотку иммунизированных животных, разведенную 1:100, или супернатанты культур клеток, инкубированные в течение 2 часов при 37°С. Планшет снова трижды отмывали в PBS и Tween 20 (0.05%), и добавляли конъюгат антитела против Ig (конъюгированное с HRP антитело к Ig мыши SIGMA А 0412) в разведении 1:5000, а затем инкубировали при 37°С в течение 90 минут. После этого планшет трижды отмывали в PBS и Tween 20 (0.05%), добавляли ТМВ-субстрат для проявления цвета пероксидазы хрена (BioRad) и в течение 15 минут инкубировали в темноте. Реакцию прерывали, добавляя 0.5 N H2SO4, считывали показания при 450 нм. В качестве положительного контроля использовали поликлональную гипериммунную сыворотку в разведении 1:200.
5.1. Анализы на авидность на основе иммуноферментного анализа
5.1.1. Анализ на авидность с мочевиной (Niederhouser et al. - 5.1)
Протокол аналогичен иммуноанализу один (5), за исключением следующих модификаций: после инкубации образца (100 нг клона 10 очищенного моноклонального антитела и 2.0 нг клона 38 очищенного моноклонального антитела) в течение двух часов при 37°С образцы трижды отмывали в 8 М растворе мочевины в PBS и Tween 20 (0.05%), а затем четырежды отмывали в PBS и Tween 20 (0.05%), в качестве контроля использовали тот же самый образец, обработанный в нормальных условиях (без мочевины). После считывания оптической плотности образца коэффициент авидности рассчитывали как отношение оптической плотности после обработки мочевиной к оптической плотности без обработки мочевиной, умноженное на 100 (результат в перцентилях).
5.1.2. Анализ на авидность с тиоцианатом аммония (Goldblat et al. - 5.2)
Протокол аналогичен иммуноанализу один (5), за исключением следующих модификаций: после инкубации образца (100 нг клона 10 очищенного моноклонального антитела и 2.0 нг клона 38 очищенного моноклонального антитела) в течение двух часов при 37°С образцы в течение 30 минут при 37°С обрабатывали тиоцианатом аммония в следующих концентрациях: 3 М; 1.5 М; 1.0 М; 0.75 М; 0.50 М; 0.25 М и 0.125 М. Образец, не обработанный тиоцианатом аммония, использовали в качестве контрольного для каждого клона.
После считывания оптической плотности коэффициент авидности рассчитывали по следующей формуле: AR (коэффициент авидности) = [(log 50 - log А) × (В - A)/log В - log А] + А; где log 50 = 1.70. А обозначает наименьшую концентрацию тиоцианата аммония, которая приводит к уменьшению оптической плотности ниже, чем на 50%, а В обозначает наивысшую концентрацию тиоцианата аммония, которая приводит к уменьшению оптической плотности больше, чем на 50%.
6. Получение и очистка моноклонального антитела
Образец предварительно отобранных клонов 10 и 38 выращивали в бессывороточной среде (GIBCO VP-SFM) с добавлением 1% BSA в виалах на 100 мл в инкубаторе (сушильном шкафу) в атмосфере 10% СО2. Супернатанты центрифугировали, а затем фильтровали через фильтры с диаметром пор 0.22 микрометра и очищали высокоэффективной жидкостной хроматографией (ВЭЖХ) на колонке с белком А (МАВ SelectSure (GE Healthcare)). Антитела нейтрализовали при рН 7.0 с помощью 1 М Tris, pH 10.0, подвергали диализу против PBS 0.5х в деионизированной воде. Образцы лиофилизировали, ресуспендировали в деионизированной воде, концентрацию белка определяли методом Лоури и оценивали электрофорезом в полиакриламидном геле.
7. Распознавание мишени
7.1. In vitro РВР2а распознавание - иммуноблоттинг
Штамм MRSA (СЕВ), чувствительный к метициллину штамм Staphylococcus aureus (MSSA), резистентный к ванкомицину штамм Enterococcus fascism и штамм BL-21 DE3 Escherichia coli выращивали в экспоненциальной фазе роста. Один мл каждого образца центрифугировали и лизировали при перемешивании со стеклянными гранулами в приборе mini-Bead Beater (Biospect Products), 3 три раза по 30 секунд. Аликвоту каждого образца подвергали электрофорезу в 12% денатурирующем акриламидном геле (SDS-PAGE), а затем белки переносили на нейлоновую мембрану (Hybond N-BioRad). Мембрану блокировали при слабом перемешивании в течение двух часов в буфере PBS, содержащем 10% обезжиренного молока и 1% BSA (альбумина бычьей сыворотки). Мембрану три раза отмывали в PBS и Tween 20 (0.05%) и три раза в PBS. Затем мембрану помещали для инкубации на два часа с супернатантом моноклонального антитела против РВР2а, разведенным PBS в соотношении 1:1. После инкубации мембрану отмывали как описано ранее и в течение девяноста минут инкубировали с конъюгатом щелочной фосфатазы (мышиное антитело против-IgG-Sigma A3688) в соотношении 1: 15000. После этого снова отмывали в PBS и обрабатывали субстратом Western Blue для щелочной фосфатазы (Promega).
7.2. Распознавание РВР2а на поверхности бактерии - проточная цитометрия
Штамм MRSA (СЕВ) выращивали в стационарной (ON) или экспоненциальной фазе роста. Образцы отмывали в PBS 1х и ресуспендировали при OD600 0.6 (~108 бактерий/мл). Затем центрифугировали в объеме 1 мл (108 бактерий) и ресуспендировали в 0.5% BSA и разведении 1:10 нормальной сыворотки (мышиной/человеческой). Затем образцы инкубировали в течение 30 мин при 4°С, и концентраты (пеллеты, осадок) дважды отмывали в PBS и ресуспендировали в 100 микролитрах PBS, содержащего 0.5% BSA и 1:10000 разведение моноклонального антитела против РВР2а, и инкубировали в течение 30 минут при 4°С. После этой стадии образцы снова отмывали как описано ранее и ресуспендировали в 100 микролитрах PBS и 0.5% BSA и разведении (1:1000) конъюгата РЕ (фикоэритрина) с мышиным антителом против Ig, затем инкубировали в темноте в течение 30 мин при 4°С. Затем образцы снова отмывали и фиксировали 15 минут при 4°С в PBS, содержащем 2% параформальдегида. После этого образцы анализировали проточной цитометрией (Becton and Dickinson - FACScalibur).
8. In vitro анализ иммунизации (защиты) - определение минимальной ингибирующей концентрации
Штаммы MSRA (CED, Иберийский, COL и СА) выращивали в экспоненциальной фазе роста до оптической плотности 0.5 при длине волны 600 нм. Содержание бактерий в применяемом инокулуме (инокуляте) доводят, примерно, до 100000. Бульон Мюллера-Хинтона, бактериальный инокулум и увеличивающиеся (растущие) количества очищенного моноклонального антитела против РВР2а добавляли в пробирки или 24-луночные планшеты. Планшеты или пробирки инкубировали при 37°С в течение 12 часов. После этого в образцах проверяли наличие или отсутствие помутнения. Под минимальной ингибирующей концентрацией понимают наименьшее количество антитела, способное ингибировать рост бактериального инокулума (100000 бактерий).
9. In vivo анализы иммунизации (защиты)
9.1. Определение летальной дозы и LD50 для штаммов MRSA
Определение летальной дозы и LD50 проводят по методу Рида-Мюнха (23) для MRSA штаммов (СЕВ, Иберийского, WB79 СА и COL). Группам самок Balb/C мышей в возрасте 8 недель инокулировали интраперитонеально увеличивающиеся дозы бактерий и наблюдали в течение 7 дней. Животных, оставшихся в живых к этому времени, умерщвляли (эвтаназия) в соответствии с установленными правилами обращения с животными.
9.2. Системное заражение (инфицирование) и определение количественных показателей почек (почечные пробы)
Штаммы MSRA (CED, Иберийский и WB79 СА) выращивали в экспоненциальной фазе роста (DO600 ~0.6), отмывали и ресуспендировали в стерильном PBS 1х при DO600 ~0.5, что соответствует, примерно, 2×108 бактерий. Эту концентрацию рассчитывали, используя разведение и засевая в планшеты с BHI агаром (агаром с сердечно- мозговым экстрактом), содержащим 10 микрограмм оксациллина/мл. 8-недельные самки мышей Balb/C в первый день получали интраперитонеально одну дозу 400 микрограмм очищенного моноклонального антитела против РВР2а. На шестой день животных умерщвляли (эвтаназия) и почки удаляли в асептических условиях. Затем почки гомогенизировали в 1 мл стерильного бульона Лурия и серийно разводили 1:10. Сто микролитров каждого разведения засевали в планшетах с BHI агаром, содержащим 10 микрограмм/мл оксациллина, и инкубировали в течение 24 часов при 37°С, подсчитывали полученные в результате колонии и делали вычисления для тотальных разведении.
9.3. Анализ выживаемости
Штаммы MRSA выращивали в вышеописанных условиях, но регулирование (корректировку) инокулума проводили по заранее установленной дозе LD50. 8-недельные самки мышей Balb/C в первый день получали интраперитонеально одну дозу 500 микрограмм очищенного моноклонального антитела против РВР2а. На следующий день этих животных плюс контрольную группу инфицировали, интраперитонеально вводя одну дозу, около 2.5-6.0×108 бактерий в соответствии с величиной LD50 для каждого штамма. За животными наблюдали в течение 10 дней, а выживших подвергали эвтаназии.
10. Сравнительное in vivo исследование иммунизации (защиты) моноклональным антителом против РВР2а и ванкомицином - количественные показатели почек
Анализ I
Четыре группы 8-недельных самок Balb/C мышей (по 4 животных в группе) получали инфицирующую дозу 6.0×107 бактерий (СЕВ-MRSA), вводимую интраперитонеально. Животные, в зависимости от группы, в которую они входят (см. ниже), получали дозы очищенного моноклонального антитела (МАВ), ванкомицина, МАВ + ванкомицина и отрицательного контроля:
группа 1: МАВ (400 микрограмм) (первый день),
группа 2: ванкомицин (150 микрограмм, внутримышечно, 12/12 часов),
группа 3: ванкомицин + МАВ (350 микрограмм) (1 день после заражения),
группа 4: контрольная.
Первые дозы антитела и ванкомицина вводили через 4 часа после введения инфицирующей дозы. Животных подвергали эвтаназии на четвертый день, почки удаляли в асептических условиях и определяли количественные показатели почек согласно описанному выше.
Анализ II
Этот анализ проводили так же, как предыдущий, но с более низкой инфицирующей дозой (7.0×106 бактерий), причем группе 1 вводили 500 микрограмм очищенного моноклонального антитела; группе 2 вводили ванкомицин (150 микрограмм, внутримышечно, 12/12 часов; 5 доз); группе 3 вводили ванкомицин + 500 микрограмм моноклонального антитела; и группа 4 являлась контрольной (непролеченные животные).
11. Идентификация гипервариабельных областей (CDR) легкой и тяжелой цепей моноклонального антитела против РВР2а
11.1. Экстракция мРНК из гибридомных клеток
10 мл центрифугата клеточной культуры гибридом, продуцирующих моноклональное антитело, обрабатывали, используя набор для выделения (экстракции) мРНК RNeasy Mini kit (Qiagen).
11.2. Получение кДНК
Реакция с обратной транскриптазой: Для получения комплементарной ДНК использовали набор с М-MLV обратной транскриптазой (Invitrogen) в соответствии с инструкциями производителя.
11.3. Амплификация VH и VL цепей с применением полимеразной цепной реакции (ПЦР)
Реакции проводили, используя пусковые последовательности (праймеры), приведенные ниже.
11.4. Секвенирование легкой и тяжелой цепей моноклонального антитела против РВР2а
Стадии секвенирования:
I. Амплификация
Амплификацию проводили, используя вышеприведенные пусковые последовательности (праймеры) от SEQ ID NO:18 до SEQ ID NO:39.
II. Секвенирование
Использовали автоматический секвенатор ABI Prism 3100, Genetic Analyser (Hitachi).
11.5. Анализ и идентификация последовательностей CDR тяжелой и легкой цепи
Полученные последовательности ДНК анализировали с помощью программы DNA Star и транслировали в аминокислотные последовательности (сайт ExPASy - программа Translate) для дальнейшего анализа с помощью алгоритмов Kabat (24) и Chotia (25) с целью идентификации областей CDR легкой и тяжелой цепи.
12. Определение констант ассоциации и диссоциации моноклональных антител (клоны 10 и 38 методом поверхностного плазмонного резонанса [SPR, ППР] [Biacore])
SPR измерения проводили, используя сенсорный чип СМ-5 на приборе Biacore X® (Biacore AB, Uppsala, Sweden). Реагенты для связывания и HBS-ЕР буферы (10 мМ hepes, 150 мМ NaCl, 3 мМ EDTA, 0.005% P20 [Tween 0, рН 7.4]) получали от компании GE Healthcare.
Пример 2
1. Получение мышиных моноклональных антител против РВР2а
Группу животных иммунизировали в соответствии с вышеописанным протоколом. Результаты, полученные методом ELISA (ИФА), представлены на Фигуре 1.
После процесса слияния (слияние номер 90-LATAM) супернатант из 96 лунок (гибридомы) анализировали методом ELISA (ИФА). Из этого общего числа выбрали пять лучших образцов и клетки размножали (клонирование). Затем полученный супернатант снова анализировали с помощью ELISA. Результаты, полученные методом ELISA для позитивных образцов, подтверждали иммуноблоттингом (с антителами) против очищенного рекомбинантного белка (РВР2а). Конечный результат показан в Таблице I.
На Фигуре 1 показаны результаты иммуноферментного анализа (ELISA, ИФА) сыворотки иммунизированных животных на продуцирование антител против РВР2а. Каждая штриховая линия соответствует 1: 100 разведенной сыворотке иммунизированных животных. Позитивная контрольная сыворотка обозначена угловой штриховкой [ ]. Первый столбец в каждой группе: преиммунная сыворотка; второй столбец: сыворотка после четвертой иммунизации; и третий столбец: сыворотка после пятой иммунизации.
]. Первый столбец в каждой группе: преиммунная сыворотка; второй столбец: сыворотка после четвертой иммунизации; и третий столбец: сыворотка после пятой иммунизации.
Из выбранных клонов клон 77-38 подвергали реклонированию с целью проверки клеточной стабильности в отношении секретирования моноклональных антител. Из 50 проанализированных лунок все показали положительный результат при анализе методом ELISA. Этот клон размножали и хранили в жидком азоте в оборудовании LATAM (Лаборатории моноклональных антител).
2. Рост, продуцирование и очистка моноклональных антител
Процесс проводили в стандартных условиях, описанных выше. Выход составлял около 4 миллиграмм моноклонального антитела на каждые 100 мл очищаемого супернатанта. Полученные результаты можно видеть ниже.
На Фигуре 2 мы можем видеть полиакриламидный гель с неочищенными образцами и образцами после очистки. На этой Фигуре 2 показан акриламидный гель после электрофореза образцов супернатанта в неденатурирующих условиях до (дорожка 1) и после очистки (2) на ВЭЖХ колонке с MAb Selectsure смолой. Дорожки 3-9 соответствуют полученным фракциям очищенного образца. Стрелкой указан примерный размер (молекулярная масса) 150 кДа.
3. Функциональная характеристика моноклональных антител
3.1. In vitro PBP2a распознавание - иммуноблоттинг
Чтобы изучить способность распознавания антителом мишени в патогене (РВР2а-аналогичные последовательности), осуществляли иммуноблоттинг с использованием бактерий, вырабатывающих PBP2a (СЕВ и COL MRSA), штамма MSSA (метициллин-чувствительного штамма золотистого стафилококка, Staphylococcus aureus) (не вырабатывающего PBP2a), ванкомицин-резистентного штамма Enterococcus sp. (вырабатывающего РВР5), транспептидазы с низкой аффинностью к бета-лактамным антибиотикам (гомологичной PBP2a) и штамма Escherichia coli (в котором вырабатывается рекомбинантный белок) в качестве негативного (отрицательного) контроля. Полученные результаты показывают, что антитело способно распознавать белок с молекулярной массой около 76 кДа в штаммах MRSA и Enterococcus sp., соответствующих размеру PBP2a и РВР5 соответственно. Не наблюдается никакой реакционной способности моноклонального антитела как в отношении белков метициллин-чувствительного штамма Staphylococcus aureus (РВР2а-негативного), так и штамма Escherichia coli. Результаты показаны на Фигуре 3 (иммуноблоттинг с использованием штаммов MRSA и MSSA).
Результат иммуноблоттинга лизатов MRSA и MSSA по сравнению с супернатантом, содержащим моноклональные антитела против PBP2a, показаны на Фигуре 3.
1 - маркер молекулярной массы (Kaleidoscope);
2 - MSSA образец, выращенный в экспоненциальной фазе;
3 и 4 - MRSA образец, выращенный в экспоненциальной фазе;
5 - MSSA образец, выращенный в стационарной фазе (в течение ночи); и
6 и 7 - MRSA образцы, выращенные в стационарной фазе. Левой стрелкой показана молекулярная масса PBP2a.
3.2. Распознавание PBP2a на поверхности бактерий - проточная цитометрия
Целью теста, проведенного методом проточной цитометрии, являлось подтверждение способности моноклонального антитела распознавать мишень на бактерии в ее нативной форме. В предыдущих тестах (иммуноблоттинг) мы наблюдали распознавание мишени на белках после процесса денатурации, который происходит в ходе разделения белков электрофорезом в денатурирующем полиакриламидном геле. Опять же, мы анализировали негативный контрольный штамм (MSSA, РВР2а-негативный) и MRSA штамм (СЕВ), выращенные в экспоненциальной и стационарной фазах роста. Полученные результаты показывают, что антитело способно распознавать мишень на бактериальной поверхности в обоих состояниях. Присутствие белка А на поверхности Staphylococcus не способно ингибировать связывание антитела с РВР2а. Полученные результаты можно видеть на Фигуре 4.
На Фигуре 4 показаны результаты анализа методом проточной цитометрии для бактерий MRSA (СЕВ) и MSSA, инкубированных с моноклональным антителом против РВР2а и помеченных фикоэритрином (РЕ). (1) бактерии MSSA; (2) бактерии MRSA, выращенные в стационарной фазе; (3) бактерии MRSA, выращенные в экспоненциальной фазе. В MRSA популяциях наблюдается сдвиг вправо, соответствующий увеличению в помеченных клетках за счет конъюгата с флуоресцентным веществом.
4. Оценка иммунитета, обеспечиваемого моноклональным антителом против РВР2а
4.1. In vitro анализы
Определение минимальной ингибирующей концентрации (MIC, МИК)
Целью этих анализов являлась оценка способности антитела непосредственно связываться с мишенью в замкнутой системе. Для моноклональных антител с конечным терапевтическим применением положительные результаты имеют чрезвычайное значение, так как они означают, что антитело способно распознавать мишень и блокировать рост бактерий без участия иммунной системы хозяина, такого как механизмы активации комплемента и опсонизации, являющиеся результатом комбинированного действия врожденной и адаптивной иммунных систем хозяина. Оценивали СЕВ, COL и Иберийский MRSA штаммы, которые в оцениваемых условиях имеют сходные значения MIC (МИК) (около 500 микрограмм). Эти данные показывают, что вне зависимости от генетического фона различных штаммов MRSA дозы антитела, необходимые для блокады роста, не одинаковы. Эти результаты показаны на Фигуре 5.
На Фигуре 5 показаны результаты теста in vitro иммунизации (MIC - минимальная ингибирующая концентрация, МИК), обеспечиваемой очищенным моноклональным антителом против РВР2а и ванкомицином, против инокулума 105 клеток из различных штаммов MRSA. Отсутствие помутнения указывает на то, что в условиях анализа бактериальный рост не происходит.
1А: СЕВ MRSA (Бразильский эпидемический клон) + 250 мкг антитела;
2А: СЕВ MRSA + 500 мкг антитела;
3А: СЕВ MRSA + 750 мкг антитела;
4А и 6А: негативный контроль штамма СЕВ MRSA.
1В: COL MRSA + 250 мкг антитела;
2В: COL MRSA + 500 мкг антитела;
3В: COL MRSA + 750 мкг антитела;
4В и 6В: негативный контроль COL MRSA штамма.
1C: Иберийский MRSA (Европейский эпидемический клон - EEC) + 250 мкг антитела;
2С: EEC MRSA + 500 мкг антитела;
3С: EEC MRSA + 750 мкг антитела;
4С и 6С: негативный контроль EEC MRSA штамма.
1D: СЕВ MRSA + 150 мкг ванкомицина;
2D: СЕВ MRSA + 300 мкг антитела;
3D: СЕВ MRSA + 500 мкг антитела;
4D: СЕВ MRSA + 750 мкг антитела;
5D и 6D: негативный контроль.
4.2. In vivo тесты по иммунизации
4.2.1. Определение летальных и сублетальных доз для СЕВ, Иберийского, WB79 СА и COL MRSA штаммов
Эти анализы были необходимы для того, чтобы определить, что in vivo иммунизация на двух используемых моделях квантификации, введения количественных характеристик для почек с применением сублетальной дозы один и теста на выживаемость после системного заражения большим числом бактерий в инокулуме-способна вызвать гибель около 50% животных (LD50). Был выбран интраперитонеальный путь введения вследствие простоты и отсутствия потерь. Анализ проводили с применением модифицированного метода Рида-Мюнха, по два животных на состояние (инфицирующая доза бактерий в увеличивающихся концентрациях), для определения LD50 и сублетальных доз, и тест проводили для трех различных доз, так как мы предварительно знали средние летальные дозы для Staphylococcus aureus. Для исследований на этом патогене использовали штамм COL MRSA, первый клон MRSA, геном которого секвенирован. Однако он проявил слабую вирулентность для того, чтобы инфицировать животное, требовались высокие инфицирующие дозы по сравнению с другими MRSA клонами. Поэтому его не использовали в анализах иммунизации (защиты).
4.2.2. Почечные показатели иммунитета после системного заражения сублетальной дозой
Использование модели квантификации (количественного выражения) изменения функции почек в этих анализах позволяет оценить способность антитела уменьшать количество бактерий в жизненно важных органах (почках) после системного заражения (инфицирования). Уменьшение более чем в 3 log (1000 раз) достигалось в трех независимых анализах с вирулентными штаммами MRSA с разным генетическим фоном. В этих анализах животные получали ранее применявшуюся дозу 500 микрограмм антитела. Иммунитет (защиту), обеспечиваемый меньшей дозой антитела (250 микрограмм), оценивали в анализе на штамме СА-MRSA, в котором также наблюдалось уменьшение количества бактерий, но меньшее, чем при введении дозы 500 микрограмм. Результаты показаны на Фигурах 6А, 6В и 6С.
На Фигуре 6А показано количество бактерий (в логарифмическом виде) в почках животных, пролеченных и непролеченных моноклональным антителом против РВР2а и подвергшихся системному заражению сублетальной дозой штамма MRSA СЕВ. Столбцы с горизонтальными полосами показывают log концентрации бактерий, выделенных из почек каждого непролеченного животного. Столбцы с шахматным рисунком изображают log количества бактерий, выделенных из почек каждого животного, пролеченного антителом. Количество (квантификация) бактерий: контрольные: С1: 2000 бактерий; С2: 29000 бактерий; С3: 220000 бактерий; С4: 52000 бактерий (среднее 75750 бактерий). Пролеченные (иммунизированные) животные: Р1: 20 бактерий; Р2, Р3 и Р4: 10 бактерий (среднее 12.5 бактерий). Уменьшение числа бактерий, выделенных из пролеченных животных, по сравнению с непролеченными в 6060 раз.
На Фигуре 6В представлено количество бактерий (в логарифмическом виде) в почках животных, пролеченных и непролеченных моноклональным антителом против РВР2а и подвергшихся системному заражению сублетальной дозой Иберийского штамма MRSA (Европейский эпидемический клон). Столбцы с горизонтальными полосами показывают log концентрации бактерий, выделенных из почек каждого непролеченного животного. Столбцы с шахматным рисунком в крупную клетку изображают log количества бактерий, выделенных из почек каждого животного, пролеченного антителом. Столбцы с шахматным рисунком в мелкую клетку изображают соответствующее среднее значение. Количество (квантификация) бактерий: контрольные: С1: 210000 бактерий; С2: 44000 бактерий; С3: 300000 бактерий; С4: 290000 бактерий (среднее 211000 бактерий). Пролеченные (иммунизированные) животные: Р1: 80 бактерий; Р2: 200 бактерий; Р3: 10 бактерий; и Р4: 60 бактерий (среднее 87.5 бактерий). Уменьшение числа бактерий, выделенных из пролеченных животных, по сравнению с непролеченными в 2420 раз.
На Фигуре 6С показано количество (в логарифмическом виде) бактерий в почках животных, пролеченных и непролеченных моноклональным антителом против РВР2а и подвергшихся системному заражению сублетальной дозой штамма WB79 CA-MRSA (Бразильского локального штамма). Первые пять столбцов (в хх, горизонтальная штриховка и шахматный рисунок в крупную клетку): log концентрации бактерий, выделенных из почек каждого непролеченного животного. Первый столбец (в хх): оценка по сравнению с животным, умершим до эвтаназии. Столбцы 6, 7, 8 и 9 (горизонтальная штриховка и шахматный рисунок): log количества бактерий, выделенных из почек животных, которым вводили 250 мкг моноклонального антитела против РВР2а. Столбцы 10, 11, 12 и 13 (треугольники): log количества бактерий, выделенных из почек каждого животного, которому вводили 500 мкг антитела. Столбцы в клетку (шахматный рисунок) (5ый, 9ый и 13ый столбцы) показывают соответствующее среднее значение. Количество бактерий: контрольные: С1: 650000 бактерий; С2: 26000 бактерий; С3: 17000 бактерий; С4: 500000 бактерий (оценка умерших животных) (среднее 231000 бактерий). Животные, получавшие 250 мкг антитела: Р1: ноль; Р2: 5400 бактерий; Р3: 830 бактерий; и Р4: 10 бактерий (среднее 1560 бактерий). Животные, получавшие 500 мкг антитела: Р1: 80; Р2: ноль; Р3: 210; Р4: 80 бактерий (среднее 92.5). Уменьшение числа бактерий, выделенных из животных, пролеченных дозой 250 мкг, по сравнению с непролеченными: 149 раз. Пролеченных дозой 500 мкг по сравнению с непролеченными: 2497 раз.
4.2.3. Анализы на выживаемость
В анализах этого типа мы оценивали иммунитет, обеспечиваемый антителом, у животного после инфицирования с помощью бактериальной нагрузки, способной убить 50% животных (LD50) или более. Оценивали иммунитет против трех штаммов, используемых в анализах по определению количества бактерий в почках. Значительное уменьшение (i) времени жизни (выживания) и (ii) абсолютного (истинного) выживания животных, обработанных моноклональными антителами против MRSA, наблюдали в трех независимых анализах.
В анализе на штамме СЕВ MRSA 70% животных, получавших лечение, выжили после заражения по сравнению всего лишь с 10% выживших в контрольной группе (непролеченных). В анализе на Иберийском штамме MRSA результаты были аналогичны, 60% выживших иммунизированных животных и 100% умерших в контрольной группе. В анализе на штамме СА MRSA наблюдалась защита 100% (100% выживших) по сравнению с 70% в группе контрольных животных. Эти результаты показаны на Фигурах 7А, 7В и 7С.
На Фигуре 7А представлена кривая выживаемости пролеченных (иммунизированных) и непролеченных (контрольных) животных после заражения (инфицирования) дозой 2.3×108 бактерий (СЕВ MRSA), вводимой интраперитонеально (LD50).
На Фигуре 7В представлена кривая выживаемости пролеченных (иммунизированных) и непролеченных (контрольных) животных после заражения (инфицирования) дозой 4.2×108 бактерий (Иберийский штамм MRSA), вводимой интраперитонеально (LD50).
На Фигуре 7С представлена кривая выживаемости пролеченных (иммунизированных) и непролеченных (контрольных) животных после заражения (инфицирования) дозой 1.1×109 бактерий (WB79 СА-MRSA), вводимой интраперитонеально (LD50).
4.2.4. Сравнительное исследование иммунитета (защиты), обеспечиваемого моноклональным антителом и ванкомицином
Так как ванкомицин представляет собой антимикробный препарат первого выбора для лечения тяжелых инфекций бактериями MRSA, было предпринято сравнительное исследование иммунизации (защиты) на модели, отличной от предыдущих моделей. В этом исследовании животных инфицировали и введение антимикробных препаратов или моноклональных антител начинали сразу через четыре часа после заражения. Исследование проводили на трех отдельных группах: в одной группе лечили ванкомицином, в другой группе лечили моноклональными антителами, а в третьей группе одновременно вводили антимикробные препараты + антитела. Дозы ванкомицина корректировали и вводили аналогично введению в случае инфицирования людей (500 мг каждые 12 часов).
Полученные результаты показывают, что через три дня после заражения наблюдалось почти 15-кратное снижение количества бактерий в почках животных, получавших антимикробный препарат или антитела. Однако в группе, которой вводили антимикробный препарат плюс антитела, наблюдалось уменьшение количества бактерий в 4617.
На основании этих результатов мы можем заключить, что иммунитет, обеспечиваемый дозой антитела, соответствовал иммунитету, вызываемому пятью дозами ванкомицина, и что одновременное введение ванкомицина и антител очень эффективно снижало бактериальную нагрузку в почках инфицированных животных. Это исследование было повторено с более низкой инфицирующей дозой, чтобы можно было лучше наблюдать защитное (иммунизирующее) действие моноклинальных антител против MRSA, как выделенных (отдельных), так и в сочетании с ванкомицином (См. Фигуры 8).
Результаты второго анализа, проведенного с уменьшенной инфицирующей дозой, подтвердили первоначальные результаты. Иммунитет (защита), обеспечиваемый моноклональным антителом, вызывал снижение бактериальной нагрузки в 89 раз, что выше, чем при иммунизации введением 5 доз ванкомицина (снижение в 35 раз). Однако наиболее значительное уменьшение (бактериальной нагрузки) наблюдалось в группе, получавшей антитело + ванкомицин, которые вызывали уменьшение (бактериальной нагрузки) в 450 раз.
На фигуре 8А даны количественные показатели в почках животных, пролеченных моноклональным антителом против РВР2а, ванкомицином и совместно антителом + ванкомицином, и непролеченных животных после инфицирования с помощью 6.0×107 бактерий (СЕВ MRSA). Лечение начинали через 4 часа после заражения. Ванкомицин вводили каждые 12 часов (5 доз). Столбцы 1, 2, 3, 4 и 5 (штриховка): log концентрации бактерий, выделенных из почек непролеченных животных (контрольных). С1: 7000000; С2: 295000; С3: 380000; С4: 3200000 (среднее: 2718750 бактерий). Столбцы 6, 7, 8, 9 и 10 (шахматный рисунок): log концентрации бактерий, выделенных из почек животных, получавших 400 мкг моноклонального антитела против РВР2а. Р1: 4200; Р2: 310000; Р3: 330000; Р4: 90000 (среднее 183550 бактерий). Столбцы 11, 12, 13, 14 и 15 (сферы): животные, пролеченные ванкомицином. Р1: 110000; Р2: 58000; Р3: 500000; Р4: 21000 (среднее 172250 бактерий). Столбцы 16, 17, 18, 19 и 20 (треугольники): log концентрации бактерий, выделенных из почек животных, получавших (300 мкг) + ванкомицин. Р1: 1100; Р2: 700; Р3: 450; Р4: 90 (среднее 585 бактерий).
На фигуре 8В даны количественные показатели в почках животных, пролеченных моноклональным антителом против РВР2а (столбцы 7-12), ванкомицином (столбцы 13-18), совместно антителом + ванкомицин (19-24), и непролеченных животных (столбцы 1-6) после заражения (инфицирования) с помощью 7.0×106 бактерий (СЕВ MRSA). Лечение начинали через 4 часа после заражения. Ванкомицин вводили каждые 12 часов (5 доз). Столбцы 1-6: log концентрации бактерий, выделенных из почек непролеченных животных (контрольных). С1: 6000; С2: 1000; С3: 500; С4: 118000; и С5: 1000 (среднее: 25220 бактерий). Столбцы 7-12: log концентрации бактерий, выделенных из почек животных, получавших 500 мкг моноклонального антитела против РВР2а. МВ1: 450; МВ2: 200; МВЗ: 100; МВ4: 20; МВ5: ноль (среднее 284 бактерий). Столбцы 13-18: животные, пролеченные ванкомицином. VC1: 100; VC2: 700; VC3: ноль; VC4: ноль; VC5: 2800 (среднее 720 бактерий). Столбцы 19-24: log концентрации бактерий, выделенных из почек животных, получавших антитело (500 мкг) + ванкомицин. MBV1: 130; MBV2: 20; MBV3: 10; MBV4: 80; MBV5: 20 (среднее 56 бактерий).
5. Анализы на авидность
Результаты тестов на авидность клонов 10 и 38 моноклональных антител
Протокол анализа на авидность с мочевиной:
клон 10: 1.03/1.46 (считывание DO при наличии/отсутствии обработки) = 70.5%
клон 38: 1.00/1.21 = 82.6%
Протокол анализа на авидность с тиоцианатом:
Клон 10 (DO):
Контроль: 1.12
Образцы, обработанные тиоцианатом:
2 М=0.046; 1.5 М=0,047; 1 М=0.107; 0.75 М=0.483; 0.5 М=0.602; 0.375 М=0.684
Коэффициент авидности: 2.47
Клон 38 (DO):
Контроль: 1.22
Образцы, обработанные тиоцианатом:
2 М=0.056; 1.5 М=0.062; 1 М=0.129; 0.75 М=0.648; 0.5 М=0.758; 0.375 М=0.793
Коэффициент авидности: 4.40
Видно, что в обоих анализах коэффициенты авидности клона 38 выше коэффициентов авидности клона 10. Помимо этого подтверждено, что в соответствии с обоими протоколами для того, чтобы достичь DO около 1.0, необходимо в 50 раз больше антитела из клона 10, чем из клона 38.
6. Определение констант ассоциации и диссоциации моноклональных антител (клоны 10 и 38 методом поверхностного плазменного резонанса [SPR, ППР] [BIAcore])
Результаты, полученные методом SPR, подтвердили предварительные результаты по авидности антител, клон 38 снова показал более высокие результаты, чем клон 10. Согласно данным, приведенным в Таблице II, аффинность клона 38 в 450 раз выше, чем аффинность клона 10.
Эта аффинность обуславливается, главным образом, более высокой константой ассоциации, которая, примерно, в 100 раз выше, чем константа ассоциации клона 10. Благодаря очень высокой аффинности клона 38, его показатели близки показателям, предельным для обнаружения с помощью данного оборудования. Все же благодаря тщательному планированию и внимательному проведению испытания мы добились отличной корректировки экспериментальных данных, используя модель Ленгмюра, это показывает, что полученные данные надежны.
На Фигуре 9 показано взаимодействие между рекомбинантным РВР2а (антиген) и моноклональными антителами клона 38 (Фигура 9А) и клона 10 (Фигура 9В). Изображенные пульсирующей, колеблющейся (smoky) линией кривые представляют результаты ППР (SPR) в концентрациях, указанных справа. Все образцы анализировали в двойном повторе, а теоретическая модель Ленгмюра 1:1 для каждой кривой показана черным под каждой кривой. Относительные единицы (отклика) показаны на вертикальной оси, а время представлено в секундах по горизонтальной оси. Линии, ближайшие к горизонтальной оси, представляют собой фон для каждого образца (негативный контроль).
7. Идентификация гипервариабельных областей (областей, определяющих комплементарность, CDR) легкой и тяжелой цепей моноклонального антитела против РВР2а
После процесса экстракции мРНК из гибридомных клеток клона, продуцирующего используемые антитела (клона 38), получали кДНК и проводили ПЦР (полимеразные цепную реакции) с этим материалом для различных аллелей легкой и тяжелой цепей. Полученный материал секвенировали, используя те же самые пусковые последовательности (праймеры) (определяемые последовательностями с SEQ ID NO:18 по SEQ ID NO:39), применяемые в реакциях ПЦР. Легкую цепь 391 и тяжелую цепь 310 идентифицировали в трех различных секвенированиях. Применяя алгоритмы Kabat и Chotia, провели идентификацию областей CDR легкой и тяжелой цепей, которые являются целью, изложенной в прилагаемой Формуле изобретения.
Ниже мы представляем последовательности областей CDR легкой и тяжелой цепей.
SEQ ID NO:6 - аминокислотная последовательность CDR 1 легкой цепи.
RSSQSIGHSNGNTYLE
SEQ ID NO:7 - аминокислотная последовательность CDR 2 легкой цепи.
KVSNRFS
SEQ ID NO:8 - аминокислотная последовательность CDR 3 легкой цепи.
FQGSYVPLT
SEQ ID NO:9 - ДНК CDR 1 легкой цепи.
cgcagcagccagagcattggccatagcaacggcaacacctatctggaa
SEQ ID NO:10 - ДНК CDR 2 легкой цепи.
aaagtgagcaaccgctttagc
SEQ ID NO:11 - ДНК CDR 3 легкой цепи.
tttcagggcagctatgtgccgctgacc
SEQ ID NO:12 - аминокислотная последовательность CDR 1 тяжелой цепи.
GFSITSSSSCWH
SEQ ID NO:13 - аминокислотная последовательность CDR 2 тяжелой цепи.
RICYEGSISYSPSLKS
SEQ ID NO:14 - аминокислотная последовательность CDR 3 тяжелой цепи.
ENHDWFFDV
SEQ ID NO:15 - ДНК CDR 1 тяжелой цепи.
ggctttagcattaccagcagcagcagctgctggcat
SEQ ID NO:16 - ДНК CDR 2 легкой цепи.
cgcatttgctatgaaggcagcattagctatagcccgagcctgaaaagc
SEQ ID NO:17 - ДНК CDR 3 легкой цепи.
gaaaaccatgattggttttttgatgtg
Дополнительные данные
Продолжая разработку изобретения, проводили другие, дополнительные, анализы. Эти анализы описаны в нижеприведенных примерах.
Пример 3
Второе исследование с применением штамма СЕВ MRSA проводили в соответствии с протоколом, описанным в Примере 2, пункт 7.2, с добавлением двукратного, по 15 секунд, перемешивания на вортексе, после каждой отмывки; целью этих операций являлось разрушение агломератов Staphylococcus aureus и увеличение числа молекул РВР2а, экспонированных с антителами.
Тестировали маркеры FITC (флуоресцеин изотиоцианат) и РЕ (фикоэритрин) с контрольными образцами (i. чистые бактерии, без контакта с моноклональным антителом; и ii. чистые бактерии плюс маркер FITC или РЕ) и анализировали образец, обработанный моноклональным антителом и помеченный с помощью маркера РЕ или FITC. Считывание на приборе FACsalibur проводили в линейном виде.
На Фигуре 10 представлен график, полученный при анализе методом проточной цитометрии образцов MRSA в присутствии помеченного FITC антитела против РВР2а. Кривая (x) соответствует образцу без маркера а кривая (y) соответствует образцу с маркером.
Полученные результаты показали, что прибор обнаруживает около 22% помеченной популяции, что подтверждает распознавание антителами против РВР2а мишени (РВР2а) на бактериальной поверхности (Фигура 10).
Пример 4
Кроме того, заявители изучали иммунитет, обеспечиваемый моноклональным антителом против РВР2а к метициллин-резистентному Staphylococcus aureus, против энтерококка. Согласно приведенному выше, антитело распознает белки в штаммах Enterococcus sp., по всей вероятности, РВР5 - транспептидазу с низкой аффинностью к бета-лактамам, присутствующую во всех штамма энтерококка, с молекулярной массой около 76 кДа (237 аминокислот). Этот фермент гомологичен РВР2а бактерий MRSA, согласно выравниванию (ClustalW), приведенному ниже:

Выравнивание осуществляли с применением последовательностей, соответствующих РВР5 из энтерококков Е. faecalis (Efas) и Е. faecium (Efam), с MRSA РВР2а. Выделенные последовательности (жирным шрифтом - РВР5; подчеркнутая - РВР2а) соответствуют области РВР2а, используемой для выработки моноклональных антител. Аминокислоты, соответствующие активному ядру фермента, выделены курсивом.
Были проведены анализы иммунизации (защиты) in vitro, определение летальной дозы и LD50 и in vivo анализы (сублетальной дозы с количественным определением почечных показателей и анализ на выживаемость с летальной дозой) с использованием штаммов энтерококка на мышиной модели (мыши Balb/C) аналогично анализам, проводившимся с MRSA. Эти результаты можно видеть в соответствующих сообщениях.
1.1. In vitro анализы иммунизации (защиты)
Целью данного анализа было оценить in vitro иммунитет, сообщаемый антителом против VRE штамма энтерококка.
In vitro тест иммунизации (иммунитета, защиты) (MIC, МИК, минимальной ингибирующей концентрации), очищенное моноклональное антитело клон 38 (90/DA5/CB5/AA3 hib 77) против клинического штамма Enterococcus f. (VRE), лаборатория Richet.
Условия
Образец очищали от супернатанта методом ВЭЖХ, смола SelecSure MAB, подвергали диализу, лиофилизировали.
Концентрация антител (метод Лоури): 3.5 мг/мл
Инокулум (инокулят): VRE штамм
Преинокулум (прединокулят): 1 VRE колония в 20 мл Lb-бульона и ванкомицин (10 мг/мл), ON 37°C, 160 об/мин
Инокулум: 400 мл преинокулума в 20 мл Lb, колба Эрленмейера на 200 мл, 37°С, 160 об/мин
DC600 нм, считывание через 7 часов: 0.7
Количественное определение: 5.5×108 бактерий/мл
Условия теста:
Инокулум: 5.5×105 бактерий
Концентрация антитела: 300, 400, 500, 600 и 700 мг антитела
Культуральная среда: 1 мл бульона Лурия
Планшет для клеточных культур, 24-луночный
Позитивный контроль: Бульон Лурия + бактериальный инокулум (инокулят)
Негативный контроль: Бульон Лурия
Инкубация: 18 часов, 37°С
На Фигуре 11 показаны результаты оценки иммунизации, сообщаемой антителом. На Фигуре 11 показано:
А. 300 мг антитела
В. 400 мг антитела
С. 500 мг антитела
D. 600 мг антитела
Е. Негативный контроль
F. Позитивный контроль
G. 700 мг антитела
1.2. Определение LD50 и летальной дозы при интраперитонеальном способе введения - ванкомицин резистентный Enterococcus faecium
Протокол
Самки мышей Balb/C в возрасте 7 недель, средняя масса 20 грамм
День 01 - преинокулум: 1 колония VRE штамма в 10 мл бульона Lb и ванкомицин (10 мг/мл), пробирка Фалькон (Falcon) на 50 мл, выращивание ON 37°C, 160 об/мин
День 02 - инокулум: 1 мл преинокулума в 50 мл бульона Lb/ванкомицин (колба Эрленмейера на 250 мл) - 4 виалы, 37°С, 160 об/мин, выращивание до OD600 нм=0.80, центрифугирование 10 мин, 4000 об/мин, ресуспендирование в PBS 1х стерильном, OD 1.2
Количественное определение: 2.1×108 bacteria/mL
А: 60 микролитров (1.5×107)
В. 300 микролитров (6.5×107)
С. 900 микролитров (уменьшено до 300 микролитров/доза) (1.5×108)
D. 4.5 мл (уменьшено до 300 микролитров/доза) (6.5×108)
Е. 9.0 мл (1.2×109 бактерий) (уменьшено до 300 микролитров/доза)
F. 45.0 мл (6.5×109 бактерий) (уменьшено до 300 микролитров/доза)
Наблюдение за животными проводили начиная с дня 02 до десятого дня анализа. Результат показан на Фигуре 12, и сделан вывод, что летальная доза составляет 1.2×109 бактерий, a LD50 составляет 6.5×108 бактерий.
Определение количественных показателей в почках животных, выживших на седьмой день:
А (1.5×107): рост бактерий отсутствует
В (6.5×107): рост бактерий отсутствует
С (1.5×108): 3100 бактерий
D (6.5×108): 2.8×104 бактерий
1.3. In vivo тест на иммунитет - тест на выживаемость - летальная доза, системное инфицирование (заражение) интраперитонеальным путем на мышиной модели
Целью данного анализа была оценка эффективности in vivo иммунизации моноклональным антителом к РВР2а против системного заражения летальной дозой (VRE) штамма Enterococcus faecium.
1. Антитело (очищенное от супернатанта клеточной культуры в сывороточной среде)
- Диализовали и лиофилизировали очищенный образец (ВЭЖХ SelecSure MAB), ресуспендировали и фильтровали перед употреблением.
- Квантификация (получение количественных характеристик) (метод Лоури): 1.0 мг/мл.
2. Мышиная модель: самки мышей Balb/C, возраст 8 недель, масса от 23 до 25 грамм.
3. Протокол:
группа А (6 животных): 650 мг антитела (350 мг + 300 мг)
группа В (6 животных): контроль (введение физиологического раствора)
4. Приготовление бактериального инокулума (инокулята)
VRE штамм:
преинокулум, день 01, 10 мл бульона BHI и ванкомицин 10 мг/мл ON, 37°C, 160 об/мин
инокулум, день 03: 300 мл преинокулума в 30 мл BHI и ванкомицин, DO600 1.31, центрифугировали 10 мин, 4000 об/мин, ресуспендировали в стерильном PBS 0.5х, корректировка при OD=1.10, разведения и планшеты для квантификации (2.0×108 бактерий/мл); инокулум: 12 мл, центрифугат ресуспендировали в 300 мл, IP (и.п.) путь (~2.2×109 бактерий)
График исследования
День 01: IP инокуляция антитела (350 мг).
День 02: IP инокуляция антитела (300 мг), системное инфицирование после полудня (IP, 250 мл бактериального раствора - 2.2×109 бактерий).
День 02 - День 13: наблюдение за животными.
Результаты показаны на Фигуре 13. Только 2 иммунизированных животных умерли (защита 66.6%). Все контрольные животные умерли на второй день.
1.4. Тест in vivo иммунизации - системное инфицирование интраперитонеальным путем мышиной модели ванкомицин-резистентным штаммом Enterococcus faecium
Целью являлась оценка эффективности иммунизации (защиты) in vivo моноклональным антителом против РВР2а и РВР5 от системного заражения (инфекции) Е. faecium (VRE) штаммом.
Тест:
1. Антитело (очищенное от супернатанта клеточной культуры в сывороточной среде)
- Диализовали, лиофилизировали и ресуспендировали очищенный образец (AffiPrep Protein (Белок) А Biorad/ВЭЖХ SelecSure MAB).
- Квантификация (метод Лоури): 1.5 мг/мл
2. Мышиная модель: самки мышей Balb/C, возраст 8 недель, масса от 19 до 23 грамм.
3. Протокол
группа А (4 животных): 500 микрограмм антитела (в 2 месяца, d01, d02)
группа В (4 животных): неиммунизированная контрольная
4. Приготовление бактериального инокулума (инокулята)
Иберийский штамм MRSA:
преинокулум, 10 мл бульона Lb ON, 37°C, 120 об/мин
инокулум: 200 преинокулума в 20 мл Lb, DO600 0.80, центрифугировали 10 мин, 4000 об/мин, ресуспендировали в стерильном PBS 0.5х, корректировка при OD=0.51, разведения и планшеты для квантификации (2.4×108 бактерий/мл); инокулум: 500 микролитров, IP способ (путь) (2.4×108 бактерий)
График исследования:
День 01: инокуляция 250 микрограмм антитела интраперитонеально.
День 02: инокуляция 250 микрограмм антитела интраперитонеально и системное инфицирование (заражение) (интраперитонеально, 500 микролитров бактериального раствора).
День 06: эвтаназия, определение концентрации бактерий (квантификация) в почках.
На основании полученных выше результатов мы можем отметить, что:
Анализы in vitro иммунизации (иммунитета, защиты) - определение минимальной ингибирующей концентрации
Общее количество 700 микрограмм антитела способно блокировать рост 550000 бактерий. Эти значения выше, чем МИК (MIC), полученные для MRSA штаммов, которые составляют около 500 микрограмм.
Анализы in vivo иммунизации (защиты)
Тест in vivo иммунизации (защиты) - системное инфицирование (заражение) интраперитонеальным путем мышиной модели сублетальной дозой ванкомицин-резистентного Enterococcus faecium.
Животные получали 500 микрограмм моноклонального антитела, вводимого интраперитонеально (IP), и подвергались системному инфицированию (заражению), IP, введением 2.4×108 бактерий. Через четыре дня осуществляли эвтаназию и извлекали почки для количественного определения (квантификации) бактерий. Иммунизированные животные показали результат в среднем 87.5 бактерий/животное, тогда как результат для контрольных животных (неиммунизированных (непролеченных) инфицированных животных составлял в среднем 211000 бактерий/животное.
Тест на выживаемость после введения летальной дозы
Животные получали 650 микрограмм моноклонального антитела (IP) и подвергались системному инфицированию (заражению) (IP) и наблюдались ежедневно в течение 10 дней. Контрольные (неиммунизированные) животные умерли ранее второго дня после заражения, два из иммунизированных животных (6) умерли на второй день; остальные оставались живыми до конца испытания. Коэффициент выживаемости составлял 66.6%.
Настоящее изобретение предусматривает антитело по определению выше, характеризующееся тем, что это антитело представляет собой антитело изотипа IgG1, IgG2, IgG3, IgG4, IgD, IgA, IgE или IgM.
В одном варианте настоящего изобретения предусматривается фрагмент антитела, который содержит по меньшей мере участок вариабельного домена тяжелой цепи, включающий одну или более VH CDR (гипервариабельных областей, областей, определяющих комплементарность) антитела по настоящему изобретению, а также, необязательно, вариабельный домен легкой цепи, который включает одну или более VL CDR антитела по настоящему изобретению, причем вариабельный домен тяжелой цепи и, необязательно, вариабельный домен легкой цепи соединен (соединены) с дополнительным участком, таким как константный домен иммуноглобулина. Последовательности константного домена можно присоединять к последовательности(-ям) тяжелой и/или легкой цепи (цепям) для получения типа с парциальными последовательностями тяжелой и/или легкой цепей. Для этой цели можно привлечь (использовать) константные области антитела каждого изотипа или их участки, включая константные области IgG, IgM, IgA, IgD и IgE.
Если не указано иначе или определенно не противоречит контексту, термин "антитело" по настоящему изобретению включает также функционально эквивалентные фрагменты антитела, т.е. фрагменты, которые сохраняют способность специфически связываться с антигеном. Было показано, что антиген - связывающая функция антитела может осуществляться фрагментами антитела. Примеры связывающих фрагментов, охватываемых термином "антитело", включают (i) фрагмент Fab, одновалентный фрагмент, состоящий из доменов VL, CL и CH1; (ii) фрагменты F(ab)2 и F(ab')2, двухвалентные (бивалентные) фрагменты, содержащие два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fd, который по существу состоит из доменов VH и CHI; (iv) фрагмент Fv, который состоит по существу из доменов VL и VH одной (единой) цепи антитела, (v) фрагмент dAb (Ward et al.f Nature 341, 544-546 (1989)), который состоит практически (по существу) из VH домена; (vi) выделенную гипервариабельную область (CDR) и (vii) комбинацию из двух или более выделенных CDR, которые, необязательно, могут быть связаны связью, образованной посредством химического синтеза. Кроме того, хотя два домена фрагмента Fv, VL и VH могут кодироваться отдельными (разными) генами, они могут связываться методами рекомбинантной ДНК, связью, образованной посредством химического синтеза, которая позволяет получать этот фрагмент, так как единичные белковые цепи в областях VL и VH спариваются с образованием одновалентных молекул (известных как одноцепочечные антитела или одноцепочечные Fv's (scFv), (см., например, Bird et al, Science 242, 423-426 Р988) и Huston et al, PNAS USA 85, 5879-5883 (1988)). Такие одноцепочечные антитела охватываются термином "антитело", если в контексте не устанавливается, не оговаривается или четко не указывается иначе. Термином "антитела" охватываются другие формы одноцепочечных антител, такие как биспецифические рекомбинантные одноцепочечные фрагменты антител (диабоди, diabodies). Хотя такие фрагменты обычно входят в понятие "антитело", они, совместно и независимо, являются уникальными признаками настоящего изобретения, проявляющими биологические свойства и имеющими различное применение.
Настоящее изобретение предусматривает фармацевтическую композицию, которая содержит терапевтически эффективное количество антитела по настоящему изобретению. Фармацевтические композиции можно приготовить с фармацевтически приемлемыми носителями или разбавителями, а также с любыми другими адъювантами и эксципиентами из уровня техники в соответствии с обычными методами, такими как методы, раскрываемые в Remington: The Science and Practice of Pharmacy, 19th Edition, Gennaro, Ed., Mack Publishing Co., Easton. PA, 1995.
Фармацевтически приемлемые носители или разбавители, а также любые другие адъюванты и эксципиенты из уровня техники должны быть пригодными для антитела по настоящему изобретению и для метода применения. Пригодность для носителей и других компонентов фармацевтических композиций определяется наименьшим выраженным (существенным) воздействием на заданные биологические свойства антитела или фармацевтической композиции по настоящему изобретению при связывании с антигеном.
Фармацевтическая композиция по настоящему изобретению может также включать разбавители, соли, буферы, детергенты (например, неионный детергент, такой как Твин80), стабилизаторы (например, не содержащие белков сахара и аминокислоты), консерванты, солюбилизаторы и/или другие материалы, пригодные для включения в фармацевтическую композицию.
Фактические уровни доз активных ингредиентов в фармацевтических композициях по настоящему изобретению можно изменять с целью получения некоторого количества активного ингредиента, эффективного для достижения нужного терапевтического ответа пациента, при использовании конкретной композиции и конкретного метода введения, но не являющегося токсичным для пациента. Выбранный уровень дозы зависит от различных фармакокинетических факторов, включая активность конкретных применяемых композиций по настоящему изобретению, метод введения, время введения, скорость выведения конкретного применяемого соединения из организма, продолжительность лечения, другие лекарственные средства, соединения и/или материалы, применяемые в сочетании с конкретными применяемыми композициями, возраст, пол, вес, заболевание, общее состояние здоровья и анамнез пациента, проходящего лечение, и другие факторы, хорошо известные специалисту в данной области техники.
Фармацевтическую композицию можно вводить с помощью любых подходящих средств и методов. Подходящие средства введения соединения по настоящему изобретению in vivo или in vitro общеизвестны из уровня техники и могут быть выбраны специалистами в данной области техники.
Композицию по настоящему изобретению можно вводить любым подходящим методом, таким как пероральный, назальный, в виде ингаляций, топический (местный, включая трансбуккальный, трансдермальный и подъязычный (сублингвальный), ректальный, вагинальный и/или парентеральный.
Таким образом, мы еще раз подтвердили, что моноклональное антитело против MRSA РВРа обеспечивает перекрестный иммунитет (перекрестную защиту) от энтерококков. Однако дозы, необходимые для выработки иммунитета, выше, чем дозы, применяемые против MRSA в аналогичных условиях. Возможно, это вызвано пониженной способностью антитела распознавать РВР5 с такой же эффективностью, что и РВР2а.
Таким образом, предмет настоящего изобретения - моноклональные антитела против РВР2а, способные специфически связываться с РВР2а и гомологичными последовательностями, применимы при инфекциях, вызываемых бактериями, содержащими этот белок или аналогичные вещества (MRSA, MRSE и Enterococcus spp., и любой другой патоген, который содержит белок, гомологичный РВР2а).
Важно отметить, что поскольку эти инфекции представляют собой общемировую (всемирную) проблему и поскольку анализы проводились против основных известных эпидемических MRSA клонов, то продукт по настоящему изобретению применим в любом месте, где наблюдается инфицирование этим патогеном.
Ниже перечислены документы (материалы), относящиеся к уровню в области техники, на которые ссылаются заявители в описании.
1. KOPP. BJ. NIX DE, ARMSTRONG EP. Clinical and economic analysis of methicillin-resistant Staphylococcus aureus infections. The Annals of Pharmacotherapy; 38:1377-82. 2004.
2. BOYCE, JM. Are the epidemiology and microbiology of methicillin-resistant Staphylococcus aureus changing? JAMA; 279(8):623-4. 1998.
3. HUNT, С; DIONNE M, et. al. Four pediatric deaths from community-acquired methicillin-resistant Staphylococcus aureus in Minnesota and North Dakota, 1997-1999. Morbidity and Mortality weekly report-CDC USA. 48(32):707-10. 1999.
4. O'BRIEN, FG; PEARMAN, JW; GRACEY, M; RILEY, TV. GRUBB, WB. Community strain of methicillin-resistant Staphylococcus aureus involved in a hospital outbreak. Journal of Clin Microbiol. 37(9):2858-62. 1999.
5. TENOVER, FC; LANCASTER, MV; HILL. ВС; STEDWARD, CD; STOCKER, SA; HANCOCK, GA; el. al. Characterization of staphylococci with reduced susceptibilities to vancomycin and other glycopeptides. Journal of Clin Microbiol. 36(4):1020-27. 1998.
6. LUTZ, L; MATOS, SB; KUPLICH, N; MACHADO A; BARTH AL. Caracteristicas laboratoriais de Staphylococcus aureus isolado de paciente que nao respondeu ao tratamento com vancomicina. Primeiro Encontro de Controle de Infeccao Hospitalar do MERCOSUL. 1999.
7. COSGROVE, SE; CARROLL, KC; PERL, TM. Staphylococcus aureus with reduced susceptibility to vancomycin. Clin Infect Diseases. 39(4):539-45. 2004.
8. JARVIS, WR; SCHLOSSER JMA, CHINN RY, TWETTEN S, JACSON M. National prevalence of methicillin-resistant Staphylococcus aureus in patients at US health care facilities, 2006. Am J Infect Control. 35(10):631-7. 2007.
9. YAMAUCHI, M. Japan struck by resistant Staphylococcus aureus. British Medical Journal. 306:740. 1993.
10. PANLILIO, AL; CULVER, DH; GAYNES, RP. Methicillin-resistant Staphylococcus aureus in US hospitals. 1975-1991. Infection Control and Hospital Epid. 13:582-86. 1992.
11. LOWRY, F. Staphylococcus aureus infections. New England Journal of Medicine. 339:520-32. 1998.
12. FARR, BM. Prevention and control of methicillin-resistant Staphylococcus aureus infections. Current Opinion Infectious Diseases. 17:317-22. 2004.
13. TIEMERSMA, EW; BRONZWA, ER; SLAM, et. al. Methicillin-resistant Staphylococcus aureus in Europe, 1999-2002. Emerging Infectious Diseases. 10:1627-34, 2004.
14. BERETTA, ALRZ; TRABASSO. P.; STUCCHI, RB; MORETTI, ML. Use of molecular epidemiology to monitor the nosocomial dissemination of methicillin-resistant Staphylococcus aureus in an University hospital from 1991-2001. Braz. Journal of Medical and Biological Research. 37:1345-51. 2004.
15. PANNUTI. CS; GRINBAUM, RS. An overview of nosocomial infection control in Brazil. Infection Control and Hospital Epidemiology. 16(3):170-74. 1995.
16. RESENDE, EM; COUTO, BRGM; STARLING, CEF; MODENA, CM. Prevalence of nosocomial infections in general hospitals in Belo Horizonte. Infection Control and Hospital Epidemiology. 19(11):872-76. 1998.
16a. Grundmann H., Aires de Souza M., Boyce J. and Tiemersma J. Emergence and resurgence of Methicillinn-resistant Staphylococcus aureus as a public threat health. Lancet. 2006. 368:874- 85.
17. GUIGNARD, В; ENTENZA, JM; MOREILLON P. Beta-lactams against methicillin-resistant Staphylococcus aureus. Curr. Opin Pharmacol. 5(5):479-89. 2005.
18. SENNA, JP; ROTH, DM; OLIVEIRA, JS; MACHADO, DC, SANTOS DS. Protective immune response against methicillin resistant Staphylococcus aureus in a murine model using a DNA vaccine approach. Vaccine. 21:2661-66. 2003.
19. OHWADA, A; SEKIYA, M; HANAKI, H; ARAI, KK; HIRAMATSU, K, FUKUCHI, Y. DNA vaccination by mecA sequence evokes an antibacterial immune response against methicillin-resistant Staphylococcus aureus. Antimicrob Agents and Chemother. 44:767-74. 1999.
20. DOMBROWSKI, JC; WINSTON, LG. Clinical failures of appropriately-treated methicillin-resistant Staphylococcus aureus infections. J. Infect. 57(2):110-5. 2008.
20bis. RYFFEL, С; TESCH, W; BICH-MACHIN, I.; REYNOLDS, PE; BARBERIS-MAINO, L; KAYSER, FH; BERGER-BACHI, B. Sequence comparison of mecA genes isolated from methicillin-resistant S. aureus and S. epidermidis. Gene. 94:137-38. 1990.
21. ROTH, DM; SENNA, JP; MACHADO, DC. Evaluation of the humoral immune response in BALB/c mice immunized with a naked DNA vaccine anti-methicillin-resistant Staphylococcus aureus. Genetics Molecular Research. 5(3):503-12. 2006.
22. YOKOYAMA, W. Production of monoclonal antibodies: induction of immune responses, p.2.5.4-2.5.8. In: J.E. Coligan, A.M. Kruisbeek, D.H. Margulies, E.M. Shevach and W. Strober (ed). Current Protocols in Immunology, vol. 1. Wiley and Sons, Hoboken, N.J. 1995.
23. REED, LJ AND MUENCH H. Am. J. Hyg. 27:493-97. 1938.
24. KABAT, EA; WU, TT; BILOFSKY, H; M REID-MULLER, PERRY, H. Sequence of proteins of immunological interest. National Institutes of Health, Bethesda. 1993.
25. CHOTIA, С; LESK, AM; TRAMONTANO, A; LEVITT, M; SMITH-GILL, PADLAN, EA; DA VIES, D; TULIP, WR. Conformations of immunoglobulin hyper-variable regions. Nature, 342:877-883. 1989.
26. GOULD IM. The clinical significance of methicillin-resistant Staphylococcus aureus. J. Hospital Infections, 61(4):277- 82. 2005.
27. ANDREMONT, A; TIBON-CORNILLOT. M. Le triomphe des bacteries, la fin des antibiotiques? Ed Max Milo, 1 ed, 2007.
28. Silverstein, Arthur M. Paul Ehrlich's receptor immunology. San Diego: Academic Press, 2002.
29. LIM, D; STRINADKA NC. Structural basis for the beta-lactam resistance of PBP2a from methicillin-resistant Staphylococus aureus. Nat Struct Biol. 9(11):870-6. 2002.
30. PAPAKYIRIAKOU, H; VAZ, D; SIMOR, A; LOUIE M; MCGAVIN MJ. Molecular analysis of the accessory gene regulator (agr) locus and balance of virulence factor expression in epidemic methicillin-resistant Staphylococcus aureus. J Infect Dis. 181(3):2400-4.2000.
31. TEIXEIRA, LA; RESENDE, CA; ORMONDE, LR; ROSENBAUM, R; FIGUEIREDO AM, DE LENCASTRE H, TOMASZ A. Geographic spread of epidemic multiresistant Staphylococcus aureus clone in Brazil. J Clin Microbiol. 33(9):2400-4.1995.
32. DA SILVA, COIMBRA MV; TEIXEIRA, LA; RAMOS, RL; PREDARI, SC; CASTELLO, L; FAMIGLIETTI A, VA, C; KLAN, L; FIGUEIREDO AM. Spread of the Brazilian epidemic clone of a multiresistant MRSA in two cities in Argentina. J Med Microbiol. 49(2):187-92.2000.
33. SENNA, JP; PINTO, CA; MATEOS, S; QUINTANA, A; SANTOS DS. Spread of a dominant methicillin-resistant Staphylococcus aureus (MRSA) clone between Uruguayan and South Brazil Hospitals. J Hospital Infect. 53(2):15607. 2003.
34. MELTER, O; SANTOS SANCHES, I; SCHINDLER, J; AIRES DE SOUZA, M; KOVAROVA, V; ZEMLICKOVA, H; DE LENCASTRE H. Methicillin-resistant Staphylococcus aureus clonal types in the Czech Republic. J Clin Microbiol, 37(9):2798-803. 1999.
35. DA SILVA COIMBRA MV, SILVA-CARVALHO MC; WISPLINGHOFF H; HALL GO; TALLENT S; WALLACE, S; EDMOND, MB; FIGUEIREDO, AM; WEINZEL, RP. Clonal spread of methicillin-resistant Staphylococcus aureus in a large geographic area of the United States. J Hosp Infect. 53(2):103-10. 2003.
36. NYGAARD, TK; DELEO, FR; VOYICH, JM. Community-associated methicillin-resistant Staphylococcus aureus skin infections: advances toward identifying the key virulence factors. Curr Op Infect Dis. 21(2):147-52. 2008.
37. DIEP, BA; CHAMBERS, HF; GRABER, CJ; SZUMOWSKI, JD; MILLER, LG; HAN, LL; CHEN, JH; LIN, F; et. al. Emergence of multidrug-resistant, community-associated, methicillin-resistant Staphylococcus aureus clone USA300 in men who have sex with men. Ann Intern Med. 148(4):249-57. 2008.
38. REICHERT, JM; DEWITZ, MC. Anti-infective monoclonal antibodies: perils and promise of development. Nat Rev Drug Discov. 5(3):191-5. 2006.
39. GAO, J; STEWART, GC. Regulatory elements of the Staphylococcus aureus protein A (Spa) promoter. J Bacteriol. 186(12):3738-48. 2004.
40. GOFFIN, С; GHUYSEN JM. Multimodular penicillin-binding proteins: an enigmatic family of orthologs and parologs. Microbiol Mol Biol Rev. 62(4):1079-93. 1998.
41. COURVALIN P. Vancomycin resistance in gram-positive cocci. Clin Infect Dis. 1; 42 Suppl 1: S25-34.2006.
42. BAILIE GR, NEAL D. Vancomycin ototoxicity and nephrotoxicity. Med Toxicol Adverse Drug Exp. 3:376-86. 1988.
43. SALGUERO E; PLAZA D; MARINO A; MORENO C; DELGADO G. Characterising vancomycin's immunotoxic profile using Swiss and CFW mice as na experimental model. Biomed Pharmacother. Sept 16. 2008.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНАЦИИ ЛИЗИНА БАКТЕРИОФАГА И АНТИБИОТИКА ПРОТИВ ГРАМ-ПОЛОЖИТЕЛЬНЫХ БАКТЕРИЙ | 2013 |
|
RU2659208C2 |
| ПРЕДОТВРАЩЕНИЕ, РАЗРУШЕНИЕ И ОБРАБОТКА БИОПЛЕНКИ ЛИЗИНОМ БАКТЕРИОФАГА | 2013 |
|
RU2646102C2 |
| ПРЕДОТВРАЩЕНИЕ, РАЗРУШЕНИЕ И ОБРАБОТКА БИОПЛЕНКИ ЛИЗИНОМ БАКТЕРИОФАГА | 2013 |
|
RU2735103C2 |
| АНТИТЕЛА ПРОТИВ СТЕНОЧНОЙ ТЕЙХОЕВОЙ КИСЛОТЫ И ИХ КОНЪЮГАТЫ | 2014 |
|
RU2687044C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ ИНФЕКЦИЙ, ВЫЗВАННЫХ Staphylococcus aureus | 2015 |
|
RU2636780C1 |
| СПОСОБЫ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ ИНФЕКЦИЙ СТАФИЛОКОККА ЗОЛОТИСТОГО И СВЯЗАННЫХ С НИМИ НАРУШЕНИЙ | 2012 |
|
RU2635462C2 |
| ЛЕЧЕНИЕ ИНФЕКЦИИ, ВЫЗЫВАЕМОЙ МИКРООРГАНИЗМАМИ | 2002 |
|
RU2303460C2 |
| АНТИТЕЛА ИЛИ ИХ ФРАГМЕНТЫ, НАПРАВЛЕННЫЕ ПРОТИВ ЭПИТОПА СТАФИЛОКОККА ЗОЛОТИСТОГО (STAPHYLOCOCCUS AUREUS) | 2010 |
|
RU2560415C2 |
| АНТИТЕЛА, КОТОРЫЕ СПЕЦИФИЧНО СВЯЗЫВАЮТ АЛЬФА-ТОКСИН STAPHYLOCOCCUS AUREUS, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2620065C2 |
| Фармацевтическая композиция для лечения глазных инфекций, вызванных метициллин-устойчивыми штаммами Staphylococcus aureus, включающая в качестве активного начала N-концевой CHAP-домен эндолизина бактериофага K Staphylococcus aureus | 2019 |
|
RU2715694C1 |























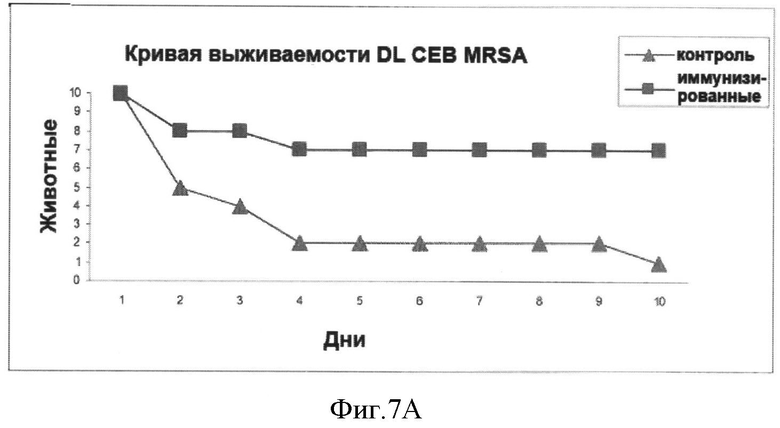
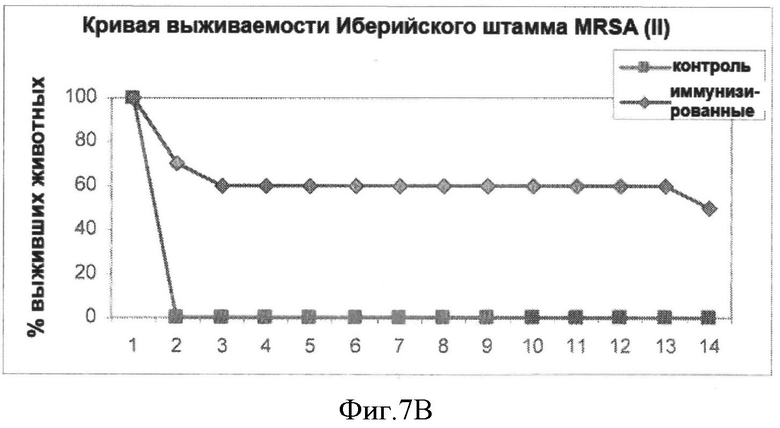










Группа изобретений относится к области биохимии. Предложено выделенное моноклональное антитело, которое связывается с белком РВР2а или РВР5. Указанное антитело содержит вариабельную область тяжелой цепи, имеющую область CDR1, которая включает SEQ ID NO: 12, область CDR2, которая включает SEQ ID NO: 13, область CDR3, которая включает SEQ ID NO: 14, и вариабельную область легкой цепи, имеющую область CDR1, которая включает SEQ ID NO: 6, область CDR2, которая включает SEQ ID NO: 7, и область CDR3, которая включает SEQ ID NO: 8. Также предложена фармацевтическая композиция, содержащая вышеуказанное антитело. Группа изобретений оказывает иммунизирующее действие против инфекции, вызванной бактериями, обладающими белком РВР2а или РВР5. 2 н. и 2 з.п. ф-лы, 20 ил., 3 табл., 4 пр.
1. Выделенное моноклональное антитело, которое связывается с белком РВР2а или белком РВР5, характеризующееся тем, что указанное антитело содержит вариабельную область тяжелой цепи, имеющую область CDR1, которая включает SEQ ID NO: 12, область CDR2, которая включает SEQ ID NO: 13, область CDR3, которая включает SEQ ID NO: 14, и вариабельную область легкой цепи, имеющую область CDR1, которая включает SEQ ID NO: 6, область CDR2, которая включает SEQ ID NO: 7, и область CDR3, которая включает SEQ ID NO: 8.
2. Выделенное моноклональное антитело по п. 1, характеризующееся тем, что указанное антитело содержит вариабельную область тяжелой цепи, имеющую область CDR1, которая включает аминокислотную последовательность, которая кодируется SEQ ID NO: 15, область CDR2, которая включает аминокислотную последовательность, которая кодируется SEQ ID NO: 16, область CDR3, которая включает аминокислотную последовательность, которая кодируется SEQ ID NO: 17, и вариабельную область легкой цепи, имеющую область CDR1, которая включает аминокислотную последовательность, которая кодируется SEQ ID NO: 9, область CDR2, которая включает аминокислотную последовательность, которая кодируется SEQ ID NO: 10, и область CDR3, которая включает аминокислотную последовательность, которая кодируется SEQ ID NO: 11.
3. Моноклональное антитело по п. 1, характеризующееся тем, что представляет собой антитело изотипа IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgD или IgE.
4. Фармацевтическая композиция для лечения или предупреждения инфекции, вызванной бактериями, обладающими белком РВР2а или белком РВР5, содержащая некоторое количество выделенного моноклонального антитела по пп. 1-2 и фармацевтический наполнитель, носитель или эксципиент.
| WO00/71585 A1, 30.11.2000 | |||
| US20030224000 A1, 04.12.2003 | |||
| СПОСОБ ЛЕЧЕНИЯ ИНФЕКЦИИ, ВЫЗЫВАЕМОЙ ШТАММАМИ РОДА STAPHYLOCOCCUS, STREPTOCOCCUS ИЛИ ENTEROCOCCUS С МНОЖЕСТВЕННОЙ ЛЕКАРСТВЕННОЙ УСТОЙЧИВОСТЬЮ | 1996 |
|
RU2202360C2 |

