Настоящее изобретение относится к новому способу биологического контроля наличия Pseudomonas и их размножения.
Pseudomonas представляет собой грамотрицательную бактерию, принадлежащую к семейству Pseudomonadaceae. У людей данная бактерия ответственна за различные кожные, висцеральные и легочные инфекции, в особенности, за кистозные фиброзы (4, 19). Эти бактерии устойчивы к многочисленным антисептикам и антибиотикам (2) (7), что, несомненно, частично объясняет их все возрастающее присутствие в больницах, где их можно выделить из влажной окружающей среды (раковины, сифоны, унитазы, полотенца и приспособления для мытья, контейнеры, содержащие воду, и т.д.). Некоторые виды также патогенны для растений (8), нематод (10) и амеб (1, 11, 16). Таким образом, мониторинг и контроль данной бактерии требует все возрастающего внимания.
В общем, известно, что в окружающей среде Pseudomonas распространена повсеместно (5), поскольку данную бактерию выделяют из почвы, из канализационных сетей или из промышленных сточных вод и из биопленок, характеристики, которые она разделяет со свободноживущими амебами. Некоторые потенциально патогенные бактерии (Legionella pneumophila, Mycobacterium spp. и Escherichia coli О157:H7) выработали механизмы выживания и воспроизведения внутри свободноживущих амеб (15). Кроме того, было показано, что бактерии, включая Pseudomonas, могут создавать различные стратегии, позволяющие им уклоняться от хищнического поведения свободноживущих амеб (12, 13, 18). В особенности, биопленка, которую образует Pseudomonas aeruginosa, представляет собой один из механизмов, позволяющих бактериям эффективно избегать истребления свободноживущими амебами, такими как Acanthamoeba polyphaga (18). Хотя известно, что некоторые свободноживущие амебы, такие как Acanthamoeba, способны вырабатывать хемотаксический ответ по отношению к Pseudomonas и питаться этими бактериями (17, 18), было также показано, что Pseudomonas aeruginosa быстро подавляет рост этих амеб и индуцирует их инцистирование и их гибель путем секреции токсинов (11-13, 17, 18). Токсическое действие Pseudomonas было также продемонстрировано в отношении ресничных простейших (9).
Следовательно, ясно видно, что свободноживущие простейшие и амебы представляют собой важный элемент экологии Pseudomonas. Кроме того, способность Pseudomonas инфицировать простейших и выживать внутриклеточно в простейших представляет собой мощный индикатор того, что данные простейшие служат факторами, которые усиливают устойчивость Pseudomonas к биоцидным обработкам, применяемым в настоящее время, как показано в работе Michel et al. (14).
В связи с этим авторы изобретения продемонстрировали совершенно неожиданно для всех, что амебы вида Willaertia magna уничтожают бактерии Pseudomonas. Этот биоцидный эффект служит дополнением к уже продемонстрированной способности Willaertia magna истреблять другие амебные агенты, которые могут служить в качестве вектора для Pseudomonas (3).
Следовательно, предмет настоящего изобретения, прежде всего, представляет собой способ контроля размножения Pseudomonas, в котором применяют простейших вида Willaertia magna. Способы в соответствии с изобретением не включают методы обработки, применяемые для организма человека или животного. В способе в соответствии с изобретением чаще всего простейшими рода Willaertia и, в особенности, вида Willaertia magna обрабатывают поток газа или жидкости.
В особенности, способ в соответствии с изобретением может быть применен для дезинфекции воды в канализационных системах или в распределительных сетях промышленных вод, охлаждающих контурах для промышленных предприятий или в сетях кондиционирования воздуха. Простейших можно непосредственно добавлять в воду или в жидкости, циркулирующие в подлежащих обработке трубах или в сетях. Кроме того, можно также распылить их, например, в виде водного раствора, как аэрозоль, в промышленных сетях, вытяжных трубах и в установках, и на производственных поверхностях, подлежащих дезинфекции.
Предпочтительно, простейшие, примененные в контексте изобретения, представляют собой штамм, депонированный 26 августа 2006, под номером РТА 7824 в АТСС, или представляют собой штамм, депонированный 26 августа 2006, под номером РТА 7825 в АТСС, эти два штамма, депонированные в пользу Centre National de la Recherche Scientifique (CNRS) [Французский национальный центр научных исследований] - 3 rue Michel Ange - 75794 Paris Cedex 16 / Франция - и Uinversite Lyon 1 Claude Bernard [Лионский университет I имени Клода Бернара] - 43 Boulevard du 11 Novembre 1918 - 69622 Villeurbanne Cedex / Франция.
Простейшие, принадлежащие к роду Willaertia, относящиеся к штамму, депонированному под номером РТА 7824 в АТСС, или к штамму, депонированному под номером РТА 7825 в АТСС, представляют собой неотъемлемую часть изобретения. Указанные депонированные штаммы РТА 7824 и РТА 7825 также описаны в публикации Международной заявки РСТ WO 2008/043969.
Следовательно, такие простейшие могут быть применены в дезинфицирующих средствах, в особенности, предназначенных для уничтожения бактерий Pseudomonas, и для контроля размножения и загрязнения Pseudomonas.
Кроме того, предмет изобретения представляет собой дезинфицирующее средство, содержащее простейшие рода Willaertia, и в особенности, вида Willaertia magna. Простейшие, представляющие собой штамм, депонированный под номером РТА 7824 в АТСС, или штамм, депонированный под номером РТА 7825 в АТСС, будут предпочтительными. Предпочтительно, дезинфицирующее средство в соответствии с изобретением находится в виде водного раствора или суспензии, например, в дистиллированной воде. Дезинфицирующее средство может находиться в распыляемой форме, например, в виде аэрозоля, или в виде любого другого средства для нанесения.
Ингибирующая размножение Pseudomonas активность простейших рода Willaertia, и, в особенности, вида Willaertia magna, была продемонстрирована авторами изобретения путем сравнения репликации Pseudomonas в родах Acanthamoeba и Hartmannella, примененными в качестве амебной модели, с репликацией, наблюдаемой в амебном роду Willaertia. Активность простейших рода Willaertia, и, в особенности, вида Willaertia magna, также была показана путем демонстрации уничтожения под действием Willaertia magna биопленок, сформированных Pseudomonas aeruginosas.
Предмет изобретения также заключается в применении дезинфицирующего средства или простейших, таких как описаны выше, в качестве биоцида на Pseudomonas.
С учетом существенной роли, которую играют амебы в размножении и поддержании Pseudomonas во внешней среде, способ и дезинфицирующее средство в соответствии с изобретением имеют многочисленные преимущества, в особенности, с точки зрения стоимости, эффективности и бережного отношение к окружающей среде.
Примеры, приведенные далее в настоящем документе, приведены для иллюстрации изобретения, но не имеют ограничительного характера.
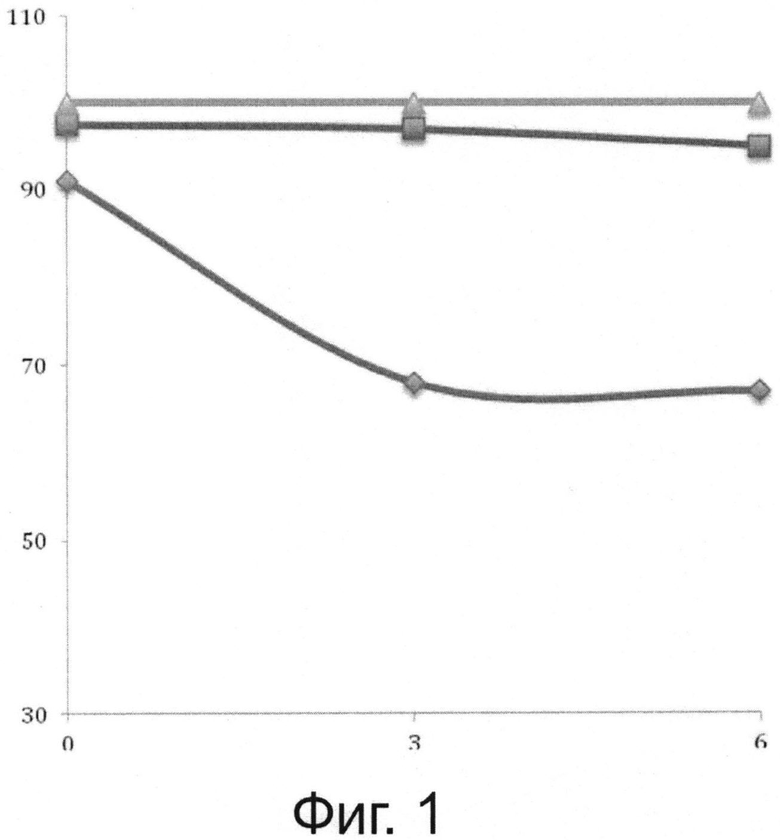
Фигура 1 показывает спонтанное развитие соответствующих популяций амеб Hartmannella vermiformis («квадратный» символ ■), Acanthamoeba castellanii («ромбовидный» символ ◆) и Willaertia {Willaertia magna - «треугольный» символ ▲) после помещения в совместную культуру с Pseudomonas при исходном отношении амеба/бактерия, равном 10.
Различные свободноживущие амебы помещали в совместные культуры (время 0 часов = Т0) с Pseudomonas в отношении, равном 10 (10 бактерий / одна амеба), как описано в разделе «Материалы и методы». Аликвоты суспензий совместных культур отбирали каждые 3 часа: т.е. Т0, T0+3h, T0+6h. Процент живых амеб определяли с помощью теста на невключение трипанового синего и исследование под микроскопом с применением камеры Малассе. Данные выражены в % от живых клеток в тесте на невключение трипанового синего.
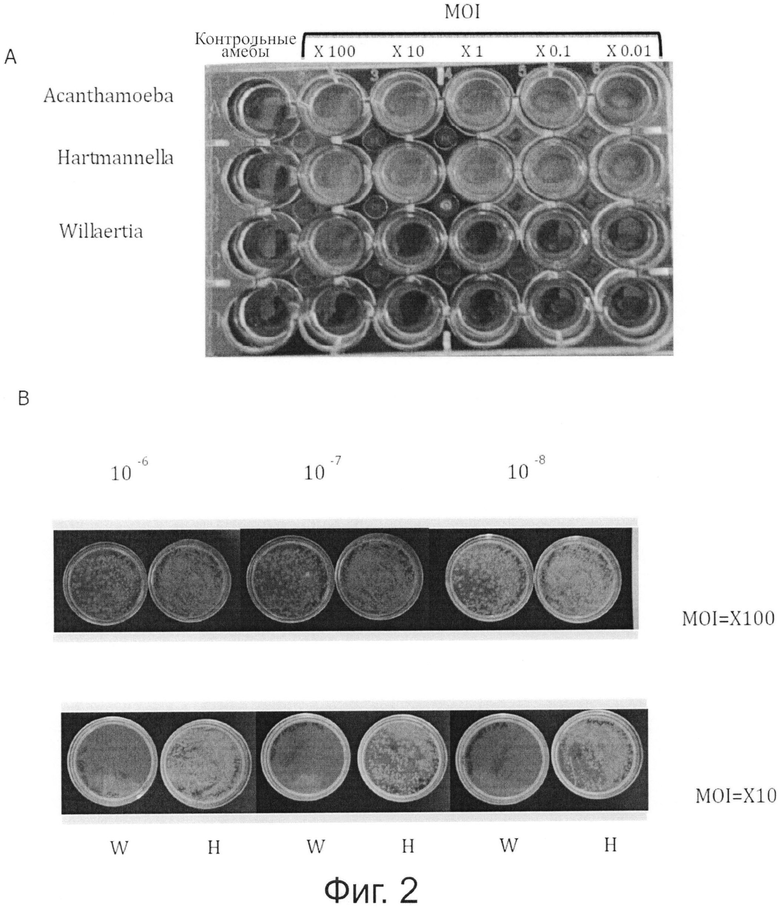
Фигура 2 показывает рост Pseudomonas в совместных культурах с Acanthamoeba и Hartmannella, но не с Willaertia.
Амебы (5×104) суспендировали в воде и инокулировали в лунки. Через 1 час Pseudomonas добавляли в лунки так, чтобы достичь различных MOI, как показано на панели А. Совместные культуры инкубировали при 30°С и анализировали через 24 и 48 часов. А. Обратите внимание на отсутствие бактериального размножения в лунке, содержавшей Willaertia с Pseudomonas. В. 100 мкл супернатанта из лунок, содержавших 100 бактерий / 1 амеба и 10 бактерии / 1 амеба (MOI 100 и MOI 10, соответственно) последовательно разводили (разведения в пределах от 10-6 до 10-8 стерильной деионизированной водой) и инокулировали в TSA-агар. Обратите внимание на отсутствие развития бактериальных колоний в супернатантах совместных культур с Willaertia magna (W) и, наоборот, интенсивное развитие Pseudomonas в присутствии Hartmanella (Н).
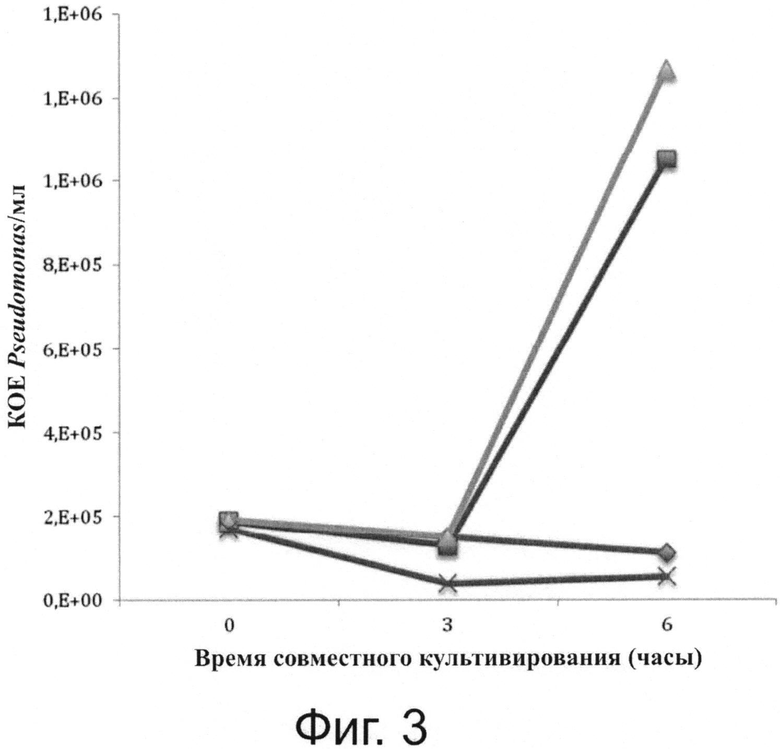
Фигура 3 показывает сравнительную кинетику рост Pseudomonas («ромбовидный» символ ◆), полученный в совместной культуре с различные родами амеб, включая род Willaertia (Willaertia magna - «крестообразный» символ ×).
Различные свободноживущие амебы по отдельности помещали в совместные культуры (время 0 часов = Т0) с Pseudomonas в отношении, равном 10 (10 бактерии / одна амеба). Аликвоты совместных культур суспензий отбирали каждые 3 часа: т.е. Т0, T0+3h, T0+6h, и концентрации Pseudomonas определяли, как описано в разделе «Материалы и методы». Положительный контроль, включавший только бактерии Pseudomonas в концентрации, равной концентрации в совместных культурах, будет служить в качестве контроля рост Pseudomonas в среде. Обратите внимание на бактериальное размножение в совместных культурах с Acanthamoeba castellanii («квадратный» символ ■) и Hartmannella vermiformis («треугольный» символ ▲).
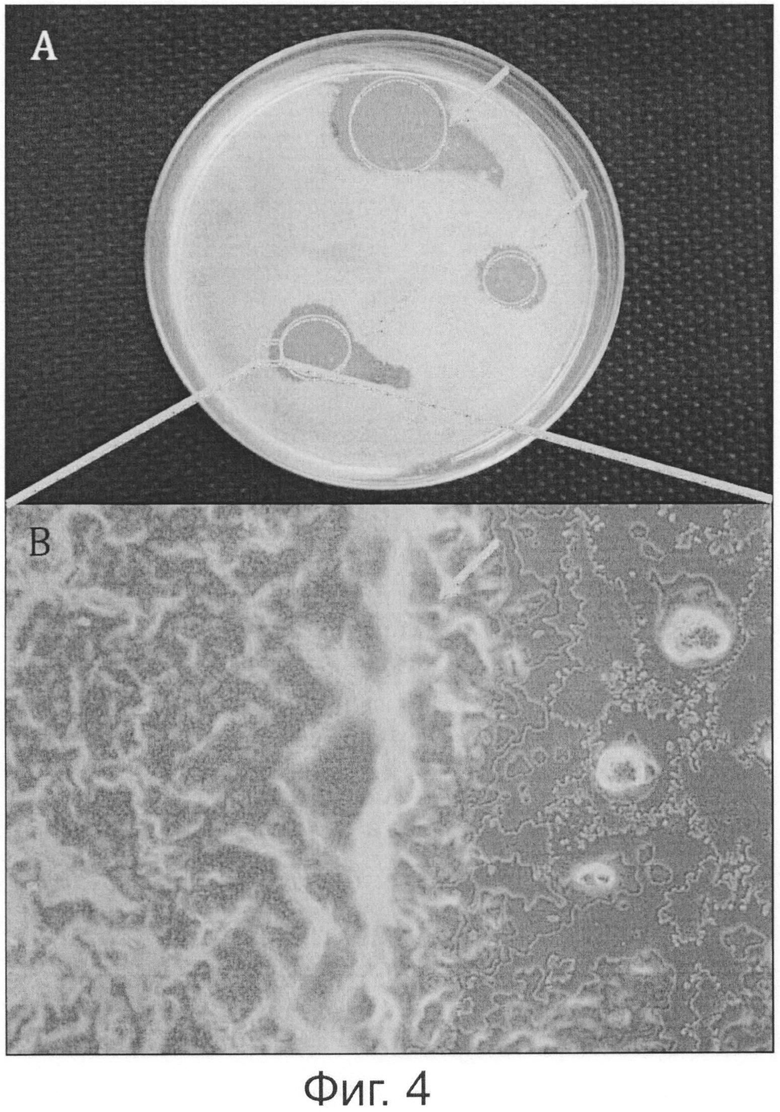
Фигура 4 показывает биоцидный эффект Willaertia magna на биопленку, сформированную Pseudomonas.
Pseudomonas и Willaertia magna инокулировали в TSA-агар, и инкубировали при 30°С в течение 24-х часов. А. На биопленке Pseudomonas в результате лизиса формируются бляшки и за пределами отложений Willaertia magna, как показано стрелками на фигуре 4А. В. Фронт сформированных на биопленке в результате лизиса бляшек указан стрелкой. Обратите внимание, справа расположено место отложения Willaertia magna в отсутствии бактериальной пленки. Обратите внимание, слева находится слой Pseudomonas.
1. МАТЕРИАЛЫ И МЕТОДЫ
1.1. Примененные штаммы
Pseudomonas: примененный штамм представлял собой штамм CL 5210 («Oxoid», Франция).
- Его поддерживали на TSA (триптон-соевый агар) (ref РО 5012, «Oxoid», Франция) в режиме один пересев в неделю. Штамм инокулировали широкими штрихами на чашку с TSA и инкубировали в течение 2 дней при 30°С.
- Амебы: примененные штаммы принадлежали к трем различным видам амеб:
- Hartmannella vermiformis,
- Acanthamoeba castellanii (АТСС 30010),
- Willaertia magna (штаммы, депонированные в АТСС под номерами РТА7824 и РТА 7825).
Данные три штамма культивировали стерильно, в присутствии 10%-ной эмбриональной телячьей сыворотки, на среде SCGYEM (среде с сывороткой, казеином, глюкозой и дрожжевым экстрактом), распределяли по пробиркам Falcon® (3033) в количестве 3 мл на пробирку. При поддержании вегетативные формы пересевали каждые 8-9 дней. Для совместных культур 3-4-дневные субкультуры применяли так, чтобы иметь трофозоиты прямо в экспоненциальной фазе роста.
Среду SCGYEM получали следующим образом
К 900 мл дистиллированной воды добавляли 2,5 мл NaOH (1N), а затем Na2HPO4 и КН2РО4. Смесь немного подогревали на нагревательной плитке и затем постепенно при перемешивании на магнитной мешалке добавляли казеин. После растворения казеина вводили глюкозу и дрожжевой экстракт.
После полного растворения смесь последовательно фильтровали через стекловолокно («Sartorius» SM 6513400) и затем через 1 мкм мембрану («Whatman» 7190 004). Среду затем разливали по аликвотам в стеклянные бутылки. Бутылки стерилизовали в автоклаве в течение 20-ти минут при 120°С. Перед окончательным применением и распределением среды в вытяжном шкафу с ламинарным потоком стерильно добавляли эмбриональную телячью сыворотку в количестве 10% от конечного объема.
1.2. Моноамебная совместная культура Pseudomonas
1.2.1. Получение бактериального инокулята
Суспензии Pseudomonas в стерильной дистиллированной воде получали из 2-дневной культуры на TSA, так чтобы получить 1 единицу оптической плотности при 550 нм, т.е. концентрацию, равную 109 КОЕ (колониеобразующих единиц)/мл.
1.2.2. Осуществление моноамебных совместных культур
Совместное культивирование осуществляли в пробирках (Falcon® 3033) для клеточных культур, содержавших 3 мл автоклавной стерилизованной воды. Инокуляцию пробирок осуществляли в соотношении 1×105 амеб/мл из аксенических амебных суспензий, подсчитанных заранее на гемоцитометре Малассе. Заражение амеб Pseudomonas осуществляли, фиксируя отношение Pseudomonas/амеба, равным 10, т.е. 1×106 бактерий/мл инкубационной среды. Сразу после заражения пробирки с совместной культурой центрифугировали на низкой скорости (760 g в течение 10 мин) с тем, чтобы содействовать связыванию между амебами и бактериями. Через 10 мин пробирки ресуспендировали вручную и инкубировали в наклонном положении в инкубаторе при 30°С.
Судьбу амеб и Pseudomonas, помещенных в совместную культуру, определяли следующим образом.
Совместные культуры отслеживали в течение 6-ти часов после бактериального заражения. В каждом временном интервале (каждые 3 часа) пробирки с совместной культурой отбирали и анализировали как с точки зрения амеб, так и с точки зрения бактерий, после энергичного перемешивания на Вортексе для того, чтобы отсоединить амеб от стенок. Для каждой пробирки проводили следующий анализ.
- Амеб считали непосредственно в счетной камере Малассе.
- Концентрацию Pseudomonas определяли с помощью непосредственного распределения культуральной среды на чашках с TSA после 10-кратного серийного разведения в стерильной дистиллированной воде, в микропробирках Eppendorf. Каждое разведение распределяли в трех повторностях на чашках с TSА в количестве 100 мкл на чашку. Чашки затем инкубировали 30°С в течение как минимум 48-ми часов. Первый анализ TSA осуществляли через 24 часа после распределения по чашкам путем подсчета колоний; затем проводили второй анализ на 2-й день для подтверждения. Концентрации Pseudomonas выражали в КОЕ/мл инкубационной среды, принимая во внимание коэффициент разбавления, и при условии, что каждая колония соответствует одной бактерии, исходно присутствовавшей в разбавленных суспензиях.
Для каждого рода амеб кривые роста Pseudomonas представлены как функция от времени.
Кроме того, возможный цитотоксический эффект Pseudomonas на различные виды амеб определяли следующим образом:
- путем подсчета количества амеб, положительных в тесте на невключение трипанового синего. Данный тест проводили под микроскопом путем подсчета, в счетной камере Малассе, числа клеток, положительных по трипановому синему / число всех клеток.
1.3. Влияние Willaertia magna на биопленки Pseudomonas
Willaertia осаждались на слое Pseudomonas, который был распределен по чашкам с TSA. Чашки с агаром помещали при 30°С в течение 24-х часов таким образом, чтобы позволить бактериальной пленке развиваться на поверхности агара. Агары затем анализировали под оптическим микроскопом (увеличение × 400) для обнаружения в нем возможного образование лизисных бляшек на бактериальном слое.
2. РЕЗУЛЬТАТЫ
2. 1. Willaertia magna демонстрирует устойчивость к Pseudomonas
Влияние Pseudomonas на выживание различных протестированных видов амеб определяли с помощью теста на невключение трипанового синего. Очень быстро после помещения Acanthamoeba castellanii в совместную с бактериями культуру в этом амебном виде развивается основной цитотоксический эффект, с падением в жизнеспособности, равным ~30%, после 3-х часов совместного культивирования (фигура 1). Наоборот, данное явление никогда не наблюдали, если Willaertia magna помещали на чашки в совместную культуру с Pseudomonas, включительно вплоть до 9-ти часов инкубации с жизнеспособностью, которая поддерживалась близкой к 100% (фигура 1). Подобно Willaertia magna, свободноживущая амеба Hartmanella vermiformis не проявляет никакого резкого снижения в терминах жизнеспособности, определенной с помощью теста на невключение трипанового синего (фигура 1). Все эти наблюдения (отсутствие инцистирования и отсутствие цитотоксичности, индуцированных под действием Pseudomonas) ясно показывают, что Willaertia magna и Hartmanella vermiformis, в отличие от другого амебного вида типа Acanthamoeba castellanii, обнаруживают первоначальную способность противостоять Pseudomonas.
2.2 Уничтожение Pseudomonas под действием Willaertia magna
Результаты совместного культивирования Pseudomonas, проведенного в присутствии амеб, принадлежащих к родам Hartmannella и Acanthamoeba, продемонстрировали значительное размножение бактерии в присутствии данных двух родов амеб, поскольку через 6 часов было отмечено увеличение в бактериальной концентрации (фигура 2). Наоборот (хотя совместное культивирование проводили в строго одинаковых условиях), отмечали снижение примерно на логарифм в обнаруживаемой концентрации Pseudomonas в присутствии амебы Willaertia magna, по сравнению с контролем, содержавшем только Pseudomonas (фигуры 2, 3). Измеренное падение концентрации Pseudomonas демонстрирует мощный уничтожающий эффект Willaertia magna в отношении Pseudomonas.
Willaertia magna и Hartmannella vermiformis выживают, но только Willaertia magna предупреждала бактериальное размножение. Данный эффект Willaertia magna на Pseudomonas дополнительно проиллюстрирован на фигурах 3 и 4. После инкубации в течение 48 часов в воде совместное культивирование Acanthamoeba и Hartmannella с бактериями продемонстрировало размножение Pseudomonas (обратите внимание на мутный вид из лунок, содержавших Pseudomonas и Hartmannella или Acanthamoeba, в результате бактериального размножения) (фигура 3, панель А). Концентрации Pseudomonas, определенные в супернатантах из лунок совместных культур, продемонстрировали отсутствие бактерий с Willaertia magna (фигура 3, панель В), если указанные бактерии инкубировали в MOI, равном 10 (10 бактерий / одна амеба).
Уничтожающее действие Willaertia magna в отношении Pseudomonas также было продемонстрировано на фигуре 4. Действительно, после 24 часов в присутствии Willaertia magna, поверхности агара, на котором бактериальный слой исчез, видны очень четко (данные зоны обозначали в настоящем документе как бактериальный слой/ лизисные бляшки на биопленке - фигура 4, панель А). Микроскопическое исследование агаров также показало, что Willaertia magna сосредоточена на границе этой лизисной бляшки; этот эффект также проиллюстрирован на фигуре 4, панель В, где исчезновение бактериального слоя под действием Willaertia magna отчетливо прослеживается. Все эти данные и наблюдения ясно показывают уничтожающее действие Willaertia magna в отношении патогенной бактерии Pseudomonas, развившейся в биопленку.
Литература
1. Abd Н, Wretlind В, Saeed A, Idsund Е, Hultenby К, and Sandstrom G. Pseudomonas aeruginosa utilises its type III secretion system to kill the free-living amoeba Acanthamoeba castellanii. JEukaryot Microbiol 55: -243.2008.
2. Ashish A, Shaw M, Winstanley C, Ledson MJ, and Walshaw MJ. Increasing resistance of the Liverpool Epidemic Strain (LES) of Pseudomonas aeruginosa (Psa) to antibiotics in cystic fibrosis (CF)-A cause for concern? J Cyst Fibros 2011.
3. Bodennec J, Dey R, and Pernin P. Novel method for biologically combating the proliferation of Legionella pneumophila, and novel disinfecting agent containing amoebic protozoa of the Willaertia genus, edited by University CBL. France: 2010.
4. Bodey GP, Bolivar R, Fainstein V, and Jadeja L. Infections caused by Pseudomonas aeruginosa. Rev Infect Dis 5: -313.1983.
5. Bredenbruch F, Geffers R, Nimtz M, Buer J, and Haussler S. The Pseudomonas aeruginosa quinolone signal (PQS) has an iron-chelating activity. Environ Microbiol 8: -1329.2006.
6. Davies B, Chattings LS, and Edwards SW. Superoxide generation during phagocytosis by Acanthamoeba castellanii: similarities to the respiratory burst of immune phagocytes. J Gen Microbiol 137: -710.1991.
7. Fernandez M, Conde S, de la Torre J, Molina-Santiago C, Ramos JL, and Duque E. Mechanisms of resistance to chloramphenicol by Pseudomonas putida KT2440. Antimicrob Agents Chemother 2011.
8. Fones H, and Preston GM. Reactive oxygen and oxydative stress tolerance in plant pathogenic pseudomonas. FEMS Microbiol Lett doi: 10.1111/j.1574-6968.2011.02449.x. [Epub ahead of print]: 2011.
9. Hahn MW, Moore ER, and Hofle MG. Role of Microcolony Formation in the Protistan Grazing Defense of the Aquatic Bacterium Pseudomonas sp.MWH1. Microb Ecol 39: 175-185, 2000.
10. Irazoqui JE, Troemel ER, Feinbaum RL, Luhachack LG, Cezairliyan BO, and Ausubel FM. Distinct pathogenesis and host responses during infection of C. elegans by P. aeruginosa and S. aureus. PloS Pathog 6: el000982, 2010.
11. Julia AG, and Morgan BM. The effects of selected strains of pigmented microorganisms on small free-living amoeba. Can J Microbiol 10: -584.1964.
12. Matz C, Bergfeld T, Rice SA, and Kjelleberg S. Microcolonies, quorum sensing and cytotoxicity determine the survival of Pseudomonas aeruginosa biofilms exposed to protozoan grazing. Environ Microbiol 6: -226.2004.
13. Matz C, Moreno AM, Alhede M, Manefleld M, Hauser AR, Givskov M, and Kjelleberg S. Pseudomonas aeruginosa uses type III secretion system to kill biofilm-associated amoebae. Isme J2: 843-852.2008.
14. Michel R, Burghardt H, and Bergmann H. Acanthamoeba, naturally intracellularly infected with pseudomonas aeruginosa, after their isolation from a microbiologically contaminated drinking water system in a hospital. Zentralbl Hyg Umweltmed 196: -544.1995.
15. Molmeret M, Horn M, Wagner M, Santic M, and Abu Kwaik Y. Amoebae as training grounds for intracellular bacterial pathogens. Appl Environ Microbiol 71: 20-28, 2005.
16. Qureshi MM, Perez AA, 2nd, Madayag RM, and Bottone EJ. Inhibition of Acanthamoeba species by Pseudomonas aeruginosa: rationale for their selective exclusion in corneal ulcers and contact lens care systems. J Clin Microbiol 31: -1910.1993.
17. Wang X, and Ahearn DG. Effect of bacteria on survival and growth of Acanthamoeba castellanii. Curr Microbiol 34: 212-215, 1997.
18. Weitere M, Bergfeld T, Rice SA, Matz C, and Kjelleberg S. Grazing resistance of Pseudomonas aeruginosa biofilms depends on type of protective mechanism, developmental stage and protozoan feeding mode. Environ Microbiol 7: -1601.2005.
19. Yang L, Jelsbak L, and Molin S. Microbial ecology and adaptation in cystic fibrosis airways. Environ Microbiol 13: 1682-1689.2011.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ БИОЛОГИЧЕСКОГО КОНТРОЛЯ ЛИСТЕРИЙ | 2012 |
|
RU2575998C1 |
| СПОСОБ БИОЛОГИЧЕСКОЙ БОРЬБЫ С NAEGLERIA FOWLERI И ДЕЗИНФИЦИРУЮЩЕЕ СРЕДСТВО, СОДЕРЖАЩЕЕ ПРОСТЕЙШИЕ ВИДОВ WILLAERTIA MAGNA | 2014 |
|
RU2649091C2 |
| НАБОР РЕКОМБИНАНТНЫХ ФЛУОРЕСЦЕНТНЫХ ШТАММОВ БАКТЕРИЙ ВИДА YERSINIA PESTIS АНТИЧНОГО БИОВАРА ОСНОВНОГО ПОДВИДА И АЛТАЙСКОГО БИОВАРА ЦЕНТРАЛЬНОАЗИАТСКОГО ПОДВИДА ДЛЯ ИНДИКАЦИИ ВОЗБУДИТЕЛЯ ЧУМЫ В ЭКСПЕРИМЕНТАЛЬНЫХ ОБРАЗЦАХ | 2020 |
|
RU2769790C1 |
| НЕТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ ПРОСТЕЙШИХ ИЗ РОДА WILLAERTIA В КАЧЕСТВЕ ФУНГИСТАТИКА И/ИЛИ ФУНГИЦИДА | 2018 |
|
RU2770017C2 |
| СПОСОБ ДЕСТРУКЦИИ БИОПЛЁНОК PSEUDOMONAS AERUGINOSA КОМБИНАЦИЕЙ ОЗОНА С ПЕРОКСИДОМ ВОДОРОДА | 2023 |
|
RU2802662C1 |
| ШТАММ БАКТЕРИЙ Lactobacillus rhamnosus, ОБЛАДАЮЩИЙ ШИРОКИМ СПЕКТРОМ АНТАГОНИСТИЧЕСКОЙ АКТИВНОСТИ ПО ОТНОШЕНИЮ К ПАТОГЕННЫМ И УСЛОВНО-ПАТОГЕННЫМ МИКРООРГАНИЗМАМ | 2012 |
|
RU2482176C1 |
| ШТАММ БАКТЕРИЙ Lactobacillus rhamnosus 7 дс, ОБЛАДАЮЩИЙ ШИРОКИМ СПЕКТРОМ АНТАГОНИСТИЧЕСКОЙ АКТИВНОСТИ ПО ОТНОШЕНИЮ К ПАТОГЕННЫМ И УСЛОВНО-ПАТОГЕННЫМ МИКРООРГАНИЗМАМ | 2016 |
|
RU2627165C1 |
| Штамм бактерий Lactobacillus rhamnosus, обладающий широким спектром антагонистической активности по отношению к патогенным и условно-патогенным микроорганизмам | 2016 |
|
RU2627166C1 |
| СРЕДСТВО ДЛЯ СЕЛЕКТИВНОГО ВЛИЯНИЯ НА БИОПЛЕНКООБРАЗОВАНИЕ МИКРООРГАНИЗМАМИ | 2016 |
|
RU2646488C2 |
| Штамм бактерий Bacillus toyonensis ВКПМ В-13249, обладающий выраженным антагонизмом по отношению к микроорганизмам Escherichia coli, Candida albicans, Staphylococcus aureus, St. epidermidis, Salmonella typhimurium, Shigella sonnei, Pseudomonas aeruginosa | 2018 |
|
RU2693439C1 |
Группа изобретений относится к области биохимии. Предложен способ контроля размножения Pseudomonas и применение дезинфицирующего средства. Способ включает добавление штамма Willaertia magna, депонированного под номером РТА 7824 в АТСС, или штамма Willaertia magna, депонированного под номером РТА 7825 в АТСС, к потоку газа, потоку жидкости или твердой поверхности. Дезинфицирующее средство содержит штамм Willaertia magna, депонированный под номером РТА 7824 в АТСС, или штамм Willaertia magna, депонированный под номером РТА 7825 в АТСС, применяемое в качестве биоцида на Pseudomonas для обработки потока газа, потока жидкости или твердой поверхности. Изобретения обеспечивают эффективную борьбу с размножением Pseudomonas. 2 н. и 3 з.п. ф-лы, 4 ил.
1. Способ контроля размножения Pseudomonas, включающий добавление штамма Willaertia magna, депонированного под номером РТА 7824 в АТСС, или штамма Willaertia magna, депонированного под номером РТА 7825 в АТСС, к потоку газа, потоку жидкости или твердой поверхности.
2. Способ по п. 1, характеризуемый тем, что он предназначен для дезинфекции воды в канализационных системах или промышленных вод в распределительных сетях, в охлаждающих контурах для промышленных предприятий или в сетях кондиционирования воздуха.
3. Способ по любому из пп. 1-2, характеризуемый тем, что он предназначен для контроля образования биопленок в водопроводных трубах или на поверхностях, возможно, находящихся в контакте с человеком или с пищевыми продуктами животного происхождения.
4. Применение дезинфицирующего средства, содержащего штамм Willaertia magna, депонированный под номером РТА 7824 в АТСС, или штамм Willaertia magna, депонированный под номером РТА 7825 в АТСС, в качестве биоцида на Pseudomonas для обработки потока газа, потока жидкости или твердой поверхности.
5. Применение по п. 4, характеризуемое тем, что дезинфицирующее средство находится в виде водного раствора или суспензии.
| US 20100119485 A1, 13.05.2010 | |||
| US 4762547, 09.08.1988 | |||
| СПОСОБ ПРОТИВОБАКТЕРИАЛЬНОЙ ОБРАБОТКИ ПОВЕРХНОСТИ ТУШ ЗАБИТЫХ ЖИВОТНЫХ, ПРИГОДНЫХ ДЛЯ ПИТАНИЯ ЧЕЛОВЕКА | 1991 |
|
RU2037301C1 |
| СПОСОБ И КОМПОЗИЦИЯ ДЛЯ АНТИМИКРОБНОЙ ОБРАБОТКИ ПИЩЕВОГО ПРОДУКТА | 2005 |
|
RU2390151C2 |

