Предпосылки создания изобретения
Иммунные связующие вещества, в том числе антитела, их конъюгаты и производные, чрезвычайно важны с коммерческой точки зрения в качестве терапевтических и диагностических средств. В общепринятых способах получения или скрининга антител обычно используют растворимые антигены. Однако у определенных мембраносвязанных белковых антигенов конформационные антигенные эпитопы изменяются, в том числе, если антигены солюбилизированы из мембраны, что препятствует получению и скринингу антител. Кроме того, одной из основных проблем способов иммуноблоттинга и аффинной хроматографии является отбор антител со средним уровнем аффинности к антигену. Это подразумевает включение большого числа перекрестнореагирующих или липких антител, которые осложняют методику последовательного скрининга. Несмотря на то, что клетки, экспрессирующие мембраносвязанные антигены, уже были использованы непосредственно для получения антител, эффективный способ скрининга, способный выявлять и увеличивать количество антител с высокой степенью аффинности к поверхностным антигенам клетки, до сих пор отсутствует.
Краткое описание изобретения
Изобретение относится к способам идентификации иммунных связующих веществ, таких как антитела scFv, способных специфически связываться с поверхностными антигенами клетки. Способы по изобретению в основном включают взаимодействие меченых клеток, экспрессирующих антиген, с мечеными клетками, экспрессирующими иммунное связующее вещество, и выделение клеток, экспрессирующих иммунное связующее вещество, которые связываются с антиген-экспрессирующими клетками, используя клеточный сортер. Эти способы особенно подходят для быстрой и эффективной идентификации иммунных связующих веществ против конформационных эпитопов, присутствующих в интегральных мембранных белках, таких как GPCR. Изобретение также относится к выделенным иммунным связующим веществам и нуклеиновым кислотам, кодирующим иммунные связующие вещества, идентифицированные с использованием способов по изобретению.
В одном из аспектов изобретение относится к способу идентификации иммунного связующего вещества, которое специфически связывается с мишеневым поверхностным антигеном клетки. Способ предусматривает наличие множества клеток, экспрессирующих иммунные связующие вещества, функционально связанных с первой сортируемой меткой; наличие множества антиген-экспрессирующих клеток, функционально связанных со второй сортируемой меткой, где мишеневый антиген презентирован на поверхности экспрессирующей антиген клетки; взаимодействие антиген-экспрессирующих клеток с клетками, экспрессирующими иммунные связующие вещества; отделение от множества клеток, экспрессирующих иммунные связующие вещества, одной или нескольких клеток, экспрессирующих иммунные связующие вещества, которые могут специфически связываться с антиген-экспрессирующими клетками, используя клеточный сортер (например, клеточный сортер с возбуждением флуоресценции), где присутствие первой и второй сортируемой метки в одном клеточном комплексе (например, комплексе, образованном между антигеном и рецептором В-клетки) является индикатором связывания клетки, экспрессирующей иммунное связующее вещество, с антиген-экспрессирующией клеткой, тем самым идентифицируя иммунное связующее вещество, которое связывается с мишеневым антигеном.
В некоторых вариантах осуществления изобретения отобранные клетки, экспрессирующие иммунное связующее вещество, клонально выделяют. В некоторых вариантах осуществления изобретения клетки, экспрессирующие иммунное связующее вещество, подвергают клональной экспансии. В других вариантах осуществления изобретения последовательность нуклеиновой кислоты, кодирующей иммунное связующее вещество, выделяют из клеток, экспрессирующих иммунное связующее вещество. Подходящие способы выделения последовательности нуклеиновой кислоты, кодирующей иммунное связующее вещество, включают ПЦР, например, ПЦР одной клетки. Последовательность нуклеиновой кислоты, кодирующая иммунное связующее вещество, может быть выделена после клонального выделения клеток и/или после клональной экспансии.
В некоторых вариантах осуществления изобретения клетки, экспрессирующие иммунное связующее вещество, выделенные с использованием способов по изобретению, подвергают клеточному анализу для функциональной характеристики иммунного связующего вещества. Подходящий клеточный анализ включает CELISA.
В некоторых вариантах осуществления изобретения иммунным связующим веществом является антитело. Такие антитела включают мышиные, кроличьи, окрольчаченные, куриные, верблюжьи, оверблюженные, человеческие, гуманизированные и химерные антитела. Подходящие структуры антитела включают, помимо прочего, Fab, Dab, нанотело и scFv.
В некоторых вариантах осуществления изобретения мишеневый антиген экспрессируется с экзогенного гена. В других вариантах осуществления изобретения мишеневый антиген является созданным способами генной инженерии антигеном. В других вариантах осуществления изобретения мишеневый антиген является интегральным мембранным белком. Подходящие интегральные мембранные белки включают, помимо прочего, GPCR (например, CXCR2) или ионные каналы.
В некоторых вариантах осуществления изобретения первая и вторая сортируемая метка является флуоресцентной меткой. Подходящие флуоресцентные метки включают, помимо прочего, флуоресцентные белки, конъюгаты антитело/фтор и флуоресцентные клеточные метки.
В некоторых вариантах осуществления изобретения антиген-экспрессирующие клетки представляют собой дрожжевые клетки или клетки млекопитающих (например, клетки человека). В некоторых вариантах осуществления изобретения антиген-экспрессирующие клетки экспрессируют экзогенный антиген. В некоторых вариантах осуществления изобретения антиген-экспрессирующие клетки трансфицируют экспрессионным вектором.
В некоторых вариантах осуществления изобретения клетки, экспрессирующие иммунное связующее вещество, представляют собой дрожжевые клетки или клетки млекопитающих. Подходящие клетки млекопитающих включают, помимо прочего, В-клетки, например, кроличьи В-клетки. В некоторых вариантах осуществления изобретения В-клетки выделяют из иммунизированного животного, например, животного, иммунизированного вакцинацией ДНК. В некоторых вариантах осуществления изобретения клетки, экспрессирующие иммунные связующие вещества, содержат иммунное связующее вещество, экспрессирующееся с экспрессионного вектора.
В другом аспекте изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей иммунное связующее вещество, идентифицированное способами по изобретения.
В другом аспекте изобретение относится к способу получения иммунного связующего вещества, способного связывать мишеневый антиген, включающий введение последовательности нуклеиновой кислоты, кодирующей иммунное связующее вещество, идентифицированное способами по изобретению, в экспрессионную среду так, чтобы происходил синтез кодируемого иммунного связующего вещества.
В другом аспекте изобретение представляет иммунное связующее вещество, получаемое способами по изобретению.
В другом аспекте изобретение также относится к способу идентификации В-клеточного клона, который специфически связывается с поверхностным мишеневым антигеном клетки, предусматривающий: иммунизирование животного ДНК, кодирующей поверхностный антиген клетки; выделение В-клеток из иммунизированного животного; мечение В-клеток первой сортируемой меткой; наличие множества антиген-экспрессирующих клеток, функционально связанных со второй сортируемой меткой, где мишеневый антиген презентирован на поверхности экспрессирующей антиген клетки; взаимодействие антиген-экспрессирующих клеток с В-клетками; и отделение от множества В-клеток одной или нескольких В-клеток, которые могут специфически связываться с антиген-экспрессирующими клетками, используя клеточный сортер, где присутствие первой и второй сортируемой метки в одном клеточном комплексе является индикатором связывания В-клетки с антиген-экспрессирующей клеткой, тем самым идентифицируя клон В-клетки, который связывается с мишеневым антигеном.
Краткое описание чертежей
Фиг.1 схематически демонстрирует В-клетку 1, меченную флуоресцентным антителом 2, взаимодействующую с мишень-экспрессирующей клеткой, окрашенной внутриклеточным красителем 3. (4: мишень отбора/антиген; 5: В-клеточный рецептор (BCR)).
Фиг.2: процесс отбора способом FACS кроличьих В-клеток, связывающихся с растворимой мишенью ESBA903. Фиг.2А: Лимфоциты были отобраны в соответствии с прямым и боковым светорассеянием. Фиг.2В: Среди них IgG+ IgM- клетки (вероятно, В-клетки памяти) были выделены (отмечены кругом). Фиг.2С: Ожидалось, что клетки, дважды окрашенные ESBA903-PE и ESBA903-PerCP (отмечены кругом), кодируют высоко аффинные IgG к ESBA903. Клетки, демонстрирующие самую яркую флуоресценцию (не отмечены кругом), были отсортированы в 96-луночных плашках.
Фиг.3: Бусы, покрытые антителами анти-TNFальфа (меченые РЕ), связываются с TNFальфа-трансфицированными клетками СНО (верхняя панель). Контрольные бусы, покрытые антителами анти-CD19 (меченые АРС), не связывали TNFальфа-трансфицированные клетки СНО (средняя панель). Бусы, покрытые антителами анти-TNFальфа (меченые РЕ), не связывали клетки СНО дикого типа (wt) (нижняя панель). Точечные диаграммы слева демонстрируют прямое и боковое светорассеяние, что указывает, соответственно, на размер и зернистость объектов. Популяция обособленных бус (~3 мкм) была отобрана в Р2. Клетки СНО, в конечном итоге связавшиеся с бусами (~30 мкм), были отобраны в Р1. Точечные диаграммы посередине демонстрируют объекты Р1 (клетки СНО) применительно к их окрашиванию РЕ или АРС. Таким образом, клетки, взаимодействующие с анти-TNFальфа-бусами, окажутся в Р3, и клетки, взаимодействующие с анти-CD19-бусами, окажутся в Р4. Справа, статистические данные для каждого образца детализированы.
Фиг.4: Бусы, покрытые анти-TNFальфа-РЕ, и бусы, покрытые анти-CD19-АРС, были смешаны вместе с TNFальфа-трансфицированными клетками СНО. Клетки СНО были отобраны (Р1), и среди них клетки, связывающиеся либо с бусами, покрытыми анти-TNFальфаРЕ, либо с бусами, покрытыми анти-CD19-АРС, показаны в группах Р3 и Р4, соответственно. Несвязанные бусы видны в группе Р2.
Фиг.5а: FACS-анализ 3 различных суспензий СНО-TNFα - В-клеток памяти. В верхнем левом углу: точечная диаграмма, демонстрирующая прямое и боковое светорассеяние клеточной суспензии. Жизнеспособные клетки, включающие большую популяцию трансгенных клеток СНО и небольшую популяцию В-клеток памяти, были отобраны. В нижнем левом углу: точечная диаграмма, демонстрирующая флуоресценцию АРС и FITC. В данном случае, В-клетки памяти (IgG+/IgM-) были отобраны. Эти две точечные диаграммы были идентичными для всех трех образцов; ввиду этого они представлены только единожды.
Фиг.5b: гистограммы и иерархия популяций 3 образцов: сверху: CHO-TNFα клетки + ESBA105 + В-клетки памяти кролика, иммунизированного ESBA105; посередине: CHO-TNFα клетки + ESBA105 + В-клетки памяти неиммунизированного кролика; внизу: CHO-TNFα клетки + В-клетки памяти кролика, иммунизированного ESBA105. На гистограммах В-клетки памяти, связавшиеся с клетками СНО, были отобраны.
Фиг.6: FACS-анализ суспензии, состоящей из иммунизированных лимфоцитов, смешанных с трансгенными по TNFα клетками СНО, «покрытых» ESBA105. Фиг.6а: точечная диаграмма, демонстрирующая прямое и боковое светорассеяние клеточной суспензии. Жизнеспособные клетки, включающие большую популяцию трансгенных клеток СНО и небольшую популяцию лимфоцитов, были отобраны. Фиг.6b: точечная диаграмма, демонстрирующая флуоресценцию APC и FITC. В данном случае, В-клетки памяти (IgG+/IgM-) были отобраны. Фиг.6с: гистограмма, демонстрирующая флуоресценцию кальцеина отсортированных В-клеток памяти. Отобранная популяция была отсортирована (В-клетки памяти, связывающиеся с комплексом СНО-TNFα-ESBA105).
Фиг.7: Изображения светлопольной микроскопии бус, покрытых IgG анти-TNFальфа, которые взаимодействуют с трансгенными клетками СНО-TNFальфа (B220).
Фиг.8: Изображения светлопольной микроскопии (левая колонка) и изображения флуоресцентной микроскопии (правая колонка) клеток СНО-TNFальфа/ESBA105 (большие клетки), связанных с В-клетками, которые имеют антитела анти-ESBA105 на поверхности (клетки меньшего размера).
Подробное описание изобретения
Изобретение относится к способам идентификации иммунных связующих веществ, таких как антитела scFv, способных специфически связываться с поверхностными антигенами клетки. Способы по изобретению в основном включают взаимодействие меченых антиген-экспрессирующих клеток с мечеными клетками, экспрессирующими иммунное связующее вещество, и выделение клеток, экспрессирующих иммунное связующее вещество, которые связываются с экспрессирующими антиген клетками, используя клеточный сортер. Эти способы, в частности, подходят для быстрой и эффективной идентификации иммунных связующих веществ против конформационных эпитопов, представленных в интегральных белках мембраны, таких как GPCR. Изобретение также относится к выделенным иммунным связующим веществам и нуклеиновым кислотам, кодирующим иммунные связующие вещества, идентифицированные с использованием способов по изобретению.
В одном из аспектов изобретение относится к способу идентификации иммунного связующего вещества, которое специфически связывается с мишеневым поверхностным антигеном клетки. Способ предусматривает: наличие множества клеток, экспрессирующих иммунные связующие вещества, функционально связанных с первой сортируемой меткой; наличие множества антиген-экспрессирующих клеток, функционально связанных со второй сортируемой меткой, где мишеневый антиген презентирован на поверхности экспрессирующей антиген клетки; взаимодействие антиген-экспрессирующих клеток с клетками, экспрессирующими иммунные связующие вещества; отделение от множества клеток, экспрессирующих иммунные связующие вещества, одной или нескольких клеток, экспрессирующих иммунные связующие вещества, которые могут специфически связываться с антиген-экспрессирующими клетками, используя клеточный сортер (например, клеточный сортер с возбуждением флуоресценции), где присутствие первой и второй сортируемой метки в одном клеточном комплексе (например, комплексе, образованном между антигеном и рецептором В-клетки) является индикатором связывания клетки, экспрессирующей иммунное связующее вещество, с антиген-экспрессирующей клеткой, тем самым идентифицируя иммунное связующее вещество, которое связывается с мишеневым антигеном.
В некоторых вариантах осуществления изобретения отобранные клетки, экспрессирующие иммунное связующее вещество, клонально выделяют.
В определенных вариантах осуществления изобретения клонально выделенные клетки, экспрессирующие иммунное связующее вещество, подвергаются клональной экспансии, используя способы, хорошо известные специалистам в данной области техники.
В других вариантах осуществления изобретения последовательность нуклеиновой кислоты, кодирующей иммунное связующее вещество, выделяют из клеток, экспрессирующих иммунное связующее вещество. Выделение последовательности нуклеиновой кислоты можно проводить после клонального выделения или после клональной экспансии. Подходящие способы выделения последовательности нуклеиновой кислоты, кодирующей иммунное связующее вещество, включают ПЦР, например, ПЦР одной клетки.
В некоторых вариантах осуществления изобретения клетки, экспрессирующие иммунное связующее вещество, выделенные с использованием способов по изобретению, подвергают клеточному анализу для функциональной характеристики иммунного связующего вещества. Подходящий клеточный анализ включает CELISA.
В некоторых вариантах осуществления изобретения иммунным связующим веществом является антитело. Такие антитела включают мышиные, кроличьи, окрольчаченные, куриные, верблюжьи, оверблюженные, человеческие, гуманизированные и химерные антитела. Подходящие структуры антитела включают, помимо прочего, Fab, Dab, нанотело и scFv.
В некоторых вариантах осуществления изобретения мишеневый антиген экспрессируется с экзогенного гена. В других вариантах осуществления изобретения мишеневый антиген является созданным способами генной инженерии антигеном. В других вариантах осуществления изобретения мишеневый антиген является интегральным мембранным белком. Подходящие интегральные мембранные белки включают, помимо прочего, рецепторы, сопряженные с G-белком (GPCR, такие как CXCR2), или ионные каналы.
В некоторых вариантах осуществления изобретения первая и вторая сортируемая метка является флуоресцентной меткой. Подходящие флуоресцентные метки включают, помимо прочего, флуоресцентные белки, конъюгаты антитело/фтор и флуоресцентные клеточные метки.
В некоторых вариантах осуществления изобретения антиген-экспрессирующие клетки представляют собой дрожжевые клетки или клетки млекопитающих (например, клетки человека). В некоторых вариантах осуществления изобретения антиген-экспрессирующие клетки экспрессируют экзогенный антиген. В некоторых вариантах осуществления изобретения антиген-экспрессирующие клетки трансфицируют экспрессионным вектором.
В некоторых вариантах осуществления изобретения клетки, экспрессирующие иммунное связующее вещество, представляют собой дрожжевые клетки или клетки млекопитающих. Подходящие клетки млекопитающих включают, помимо прочего, В-клетки, например, кроличьи В-клетки. В некоторых вариантах осуществления изобретения В-клетки выделяют из иммунизированного животного, например, животного, иммунизированного вакцинацией ДНК. В некоторых вариантах осуществления изобретения клетки, экспрессирующие иммунные связующие вещества, содержат иммунное связующее вещество, экспрессирующееся с экспрессионного вектора.
В другом аспекте изобретение относится к выделенной молекуле нуклеиновой кислоты, кодирующей иммунное связующее вещество, идентифицированное способами по изобретению.
В другом аспекте изобретение относится к способу получения иммунного связующего вещества, способного связывать мишеневый антиген, включающий введение последовательности нуклеиновой кислоты, кодирующей иммунное связующее вещество, идентифицированное способами по изобретению, в экспрессионную среду так, чтобы происходил синтез кодируемого иммунного связующего вещества.
В другом аспекте изобретение относится к иммунному связующему веществу, получаемому способами по изобретению.
В другом аспекте изобретение также относится к способу идентификации В-клеточного клона, который специфически связывается с поверхностным мишеневым антигеном клетки, предусматриывающий: иммунизирование животного ДНК, кодирующей поверхностный антиген клетки; выделение В-клеток из иммунизированного животного; мечение В-клеток первой сортируемой меткой; наличие множества антиген-экспрессирующих клеток, функционально связанных со второй сортируемой меткой, где мишеневый антиген представлен на поверхности экспрессирующей антиген клетки; взаимодействие антиген-экспрессирующих клеток с В-клетками; и отделение от множества В-клеток одной или нескольких В-клеток, которые могут специфически связываться с антиген-экспрессирующими клетками, используя клеточный сортер, где присутствие первой и второй сортируемой метки в одном клеточном комплексе является индикатором связывания В-клетки с антиген-экспрессирующей клеткой, тем самым идентифицируя В-клеточный клон, который связывается с мишеневым антигеном.
Определения
Для большей ясности настоящего изобретения, отдельные термины определены, как указано ниже. Дополнительные определения изложены на всем протяжении подробного описания.
Термин «антитело» относится к целым антителам или любому антиген-связывающему фрагменту (т.е. «антиген-связывающая часть», «антиген-связывающий полипептид» или «иммунное связующее вещество») или одиночная цепь вышеперечисленного. «Антитело» относится к гликопротеину, включающему, по меньшей мере, две тяжелые (Н) цепи и две легкие (L) цепи, сцепленные друг с другом дисульфидными мостиками, или его антиген-связывающей части. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно в настоящем документе как VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, CH1, СН2 и СН3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно в настоящем документе как VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL могут быть дополнительно подразделены на гипервариабельные области, названные определяющими комплементарность областями (CDR), вперемежку с областями, являющимися более консервативными, названными каркасными участками (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца до карбоксильного конца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат домен связывания, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента.
Термин «химерное антитело» относится к молекуле антитела, в которой (а) константная область или ее часть изменена, замещена или заменена таким образом, что сайт связывания антигена (вариабельная область) соединен с константной областью другого или видоизмененного класса, эффекторной функции и/или вида, или совершенно другой молекулой, которая придает новые свойства химерному антителу, например, ферментом, токсином, гормоном, ростовым фактором, лекарственным средством, и т.д.; или (b) вариабельная область или ее часть изменена, замещена или заменена вариабельной областью, имеющей другую или измененную антигенную специфичность.
Термин «антиген-связывающая часть» антитела (или просто «часть антитела») относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфически связываться с антигеном (например, TNF). Было показано, что антиген-связывающая функция антитела может осуществляться фрагментами полноразмерного антитела. Примеры фрагментов связывания, подразумеваемые под термином «антиген-связывающая часть» антитела, включают (i) фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) фрагмент F(ab')2, бивалентный фрагмент, включающий два фрагмента Fab, соединенных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fd, состоящий из доменов VH и CH1; (iv) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела, (v) одиночный домен, такой как фрагмент dAb (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH; и (vi) выделенную определяющую комплементарность область (CDR) или (vii) комбинацию двух или более выделенных CDR, которые могут дополнительно быть объединены синтетическим линкером. Более того, хотя два домена фрагмента Fv, VL и VH, кодируются отдельными генами, они могут быть объединены, используя способы рекомбинации, синтетическим линкером, который дает им возможность быть синтезированными в виде одной белковой цепи, в которой области VL и VH составляют пару с образованием моновалентных молекул (известных как одноцепочечный Fv (scFv); см. например, Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Такие одноцепочечные антитела также подразумевают под термином «антиген-связывающая часть» антитела. Эти фрагменты антитела получают, используя общепринятые способы, известные специалистам в данной области техники, и фрагменты отбирают на пригодность таким же образом, как интактные антитела. Антиген-связывающие части могут быть получены способами рекомбинантных ДНК или ферментативным или химическим расщеплением интактных иммуноглобулинов. Антитела могут быть разных изотипов, например, антитело IgG (например, подтип IgG1, IgG2, IgG3 или IgG4), IgA1, IgA2, IgD, IgE или IgM.
Термин «иммунное связующее вещество» относится к молекуле, которая содержит весь антиген-связывающий участок антитела или его часть, например, весь вариабельный домен тяжелой и/или легкой цепи или его часть таким образом, что иммунное связующее вещество специфически узнает мишеневый антиген. Неограниченные примеры иммунных связывающих веществ включают полноразмерные иммуноглобулиновые молекулы и scFv, а также фрагменты антитела, включая, помимо прочего, (i) фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) фрагмент F(ab')2, бивалентный фрагмент, включающий два фрагмента Fab, соединенных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fab', который по сути является Fab с частью шарнирной области (см., FUNDAMENTAL IMMUNOLOGY (Paul ed., 3.sup.rd ed. 1993); (iv) фрагмент Fd, состоящий из доменов VH и CH1; (v) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела, (vi) однодоменное антитело, такое как фрагмент Dab (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH или VL, антитело Camelid (см. Hamers-Casterman, et al., Nature 363:446-448 (1993) и Dumoulin, et al., Protein Science 11:500-515 (2002)) или Shark (например, акульи Ig-NAR Nanobodies®); и (vii) нанотело, вариабельную область тяжелой цепи, содержащую один вариабельный домен и два константных домена.
Термин «функциональная характеристика» в том смысле, в котором он используется в настоящем описании, означает характеристику полипептида (например, иммунного связующего вещества), для которого усовершенствования (например, относительно обычного полипептида) желательны и/или предпочтительны одному из специалистов в данной области техники, например, для улучшения производственных характеристик или терапевтической эффективности полипептида. В одном из вариантов осуществления функциональной характеристикой является улучшенная стабильность (например, улучшенная термостабильность). В еще одном варианте осуществления изобретения функциональной характеристикой является улучшенная растворимость (например, в клеточных условиях). В еще одном варианте осуществления изобретения функциональной характеристикой является отсутствие агрегации. В еще одном варианте осуществления изобретения функциональной характеристикой является усовершенствование в экспрессии (например, в прокариотической клетке). В еще одном варианте осуществления изобретения функциональной характеристикой является усовершенствование в выраженности рефолдинга вслед за процессом очистки телец включения. В определенных вариантах осуществления изобретения функциональной характеристикой не является улучшение аффинности связывания антигена.
Термин «каркасные участки» относится к хорошо известным в области техники частям вариабельной области антитела, которые находятся между более отличающимися областями CDR. Такие каркасные области обычно называются каркасными областями с 1 по 4 (FR1, FR2, FR3 и FR4) и обеспечивают область фиксации, в трехмерном пространстве, трех CDR, обнаруженных в вариабельной области тяжелой или легкой цепи антитела, таким образом, что CDR могут образовывать антигенсвязывающую поверхность. Такие каркасные области также могут называться подложками, так как они обеспечивают представление более отличающихся CDR. Другие CDR и каркасные области суперсемейства иммуноглобулинов, такие как анкириновые повторы и фибронектин, могут быть использованы в качестве антигенсвязывающих молекул (см. также, например, патенты США №№ 6300064, 6815540 и публикации США № 20040132028).
Термин «эпитоп» или «антигенная детерминанта» относится к сайту на антигене, с которым специфически связывается иммуноглобулин или антитело. Эпитоп, как правило, включает, по меньшей мере, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислот в уникальной пространственной конформации. См., например, Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, G. E. Morris, Ed. (1996).
Термины «специфическое связывание», «селективное связывание», «селективно связывается» и «специфически связывается» относятся к связыванию антитела с эпитопом на заранее определенном антигене. Как правило, антитело связывается с аффинностью (KD), приблизительно, менее чем 10-7 М, как, например, приблизительно, менее чем 10-8 М, 10-9 М или 10-10 М или даже ниже.
Термин «KD» относится к равновесной константе диссоциации определенного взаимодействия антитело-антиген. Как правило, антитела по изобретению связываются с антигеном с равновесной константой диссоциации (KD), равной менее чем приблизительно 10-7 М, как, например, приблизительно менее чем 10-8 М, 10-9 М или 10-10 М или даже ниже, например, как определяется с использованием поверхностной плазмонной резонансной (SPR) технологии в аппарате BIACORE.
Термин «идентичность» в том смысле, в котором он используется в настоящем описании, относится к совпадению последовательностей двух полипептидов, молекул или двух нуклеиновых кислот. Если в двух сравниваемых последовательностях в определенном положении расположены одинаковые основания или аминокислотные мономерные субъединицы (например, если в каждой из двух молекул ДНК в определенной положении расположен аденин или в каждом из двух полипептидов в определенной положении расположен лизин), тогда соответствующие молекулы идентичны в данном положении. «Процент идентичности» между двумя последовательностями является функцией числа положений совпадения, принадлежащих двум последовательностям, деленного на число сравниваемых положений, × 100. Например, если 6 из 10 положений в двух последовательностях совпадают, тогда две последовательности имеют 60% идентичность. В качестве примера, последовательности ДНК CTGACT и CAGGTT имеют 50% идентичность (3 из всех 6 положений совпадают). Как правило, сравнительный анализ проводят, если выравнивают две последовательности для достижения максимальной идентичности. Такое выравнивание может быть обеспечено, используя, например, способ Needleman et al. (1970) J. Mol. Biol. 48: 443-453, удобно реализуемый с помощью компьютерных программ, таких как программа Align (DNAstar, Inc.). Процент идентичности двух аминокислотных последовательностей может также быть определен, используя алгоритм E. Meyers and W. Miller (Comput. Appl. Biosci., 4:11-17 (1988)), который был включен в программу ALIGN (версия 2.0), используя таблицу веса остатков PAM120, штраф за длину бреши размером 12 и штраф за брешь размером 4. Кроме того, процент идентичности между двумя аминокислотными последовательностями может быть определен, используя алгоритм Needleman and Wunsch (J. Mol. Biol. 48:444-453 (1970)), который был включен в программу GAP в программном обеспечении GCG (имеется в наличии на www.gcg.com), используя либо матрицу Blossum 62, либо матрицу PAM250, и вес бреши размером 16, 14, 12, 10, 8, 6 или 4 и вес длины размером 1, 2, 3, 4, 5 или 6.
«Сходные» последовательности - это последовательности, которые при выравнивании, имеют идентичные и сходные аминокислотные остатки, где сходные остатки являются консервативными заменами соответствующих аминокислотных остатков в выравниваемой эталонной последовательности. В связи с этим «консервативная замена» остатка в эталонной последовательности является заменой остатком, который физически или функционально сходен с соответствующим эталонным остатком, например, который имеет сходный размер, форму, электрический заряд, химические свойства, включая способность образовывать ковалентные или водородные связи или тому подобное. Таким образом, «последовательность, модифицированная консервативной заменой» - это последовательность, которая отличается от эталонной последовательности или последовательности дикого типа в том, что присутствует одна или несколько консервативных замен. «Процент сходства» двух последовательностей является функцией числа положений, которые содержат совпадающие остатки или консервативные замены, принадлежащие двум последовательностям, деленного на число сравниваемых положений, × 100. Например, если 6 из 10 положений в двух последовательностях совпадают и 2 из 10 положений содержат консервативные замены, тогда две последовательности имеют 80% положительное сходство.
Термин «модификации консервативной последовательности» в том смысле, в котором он используется в настоящем описании, предназначен для обозначения аминокислотных модификаций, которые не оказывают негативного влияния или не изменяют характеристик связывания антитела, содержащего аминокислотную последовательность. Такие модификации консервативной последовательности включают нуклеотидные и аминокислотные замены, присоединения и делеции. Например, модификации могут быть введены стандартными методиками, известными в данной области техники, такими как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Консервативные аминокислотные замены включают таковые, в которых аминокислотный остаток заменен аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, были определены в данной области техники. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, теоретически найденный несущественный аминокислотный остаток может быть заменен другим аминокислотным остатком из того же самого семейства боковых цепей. Способы идентификации нуклеотидных и аминокислотных консервативных замен, которые не нарушают связывание антигена, хорошо известны в данной области техники (см., например, Brummell et al., Biochem. 32:1180-1187 (1993); Kobayashi et al. Protein Eng. 12(10):879-884 (1999); и Burks et al. Proc. Natl. Acad. Sci. USA 94:412-417 (1997)).
Термин «аминокислотная консенсусная последовательность» в том смысле, в котором он используется в настоящем описании, относится к аминокислотной последовательности, которая может быть получена, используя матрицу, по меньшей мере, двух и предпочтительно большего количества выравниваемых аминокислотных последовательностей, и допуская бреши при выравнивании таким образом, что оказывается возможным определить наиболее часто встречаемый аминокислотный остаток в каждом положении. Консенсусная последовательность - это та последовательность, которая включает аминокислоты, которые наиболее часто представлены в каждом положении. В том случае, если две или более аминокислоты равнозначно представлены в одном положении, консенсусная последовательность включает обе или все эти аминокислоты.
Аминокислотная последовательность белка может быть проанализирована на различных уровнях. Например, консервативность или вариабельность может быть обнаружена на уровне одного остатка, уровне множественных остатков, множественных остатков с брешами, и т.д. Остатки могут демонстрировать консервативность идентичного остатка или могут быть консервативными на уровне класса. Примеры классов аминокислот включают полярные, но незаряженные группы R (серин, треонин, аспарагин и глутамин); положительно заряженные группы R (лизин, аргинин и гистидин); отрицательно заряженные группы R (глутаминовая кислота и аспарагиновая кислота); гидрофобные группы R (аланин, изолейцин, лейцин, метионин, фенилаланин, триптофан, валин и тирозин); и особые аминокислоты (цистеин, глицин и пролин). Другие классы известны специалистам в данной области техники и могут быть определены, используя структурный анализ или другие данные для оценки взаимозаменяемости. В этом смысле взаимозаменяемая аминокислота может относиться к любой аминокислоте, которая может быть заменена и сохранять функциональную консервативность в данном положении.
Общепризнанно, однако, что аминокислоты одного и того же класса могут изменяться по степени их биофизических свойств. Например, общепризнанно, что определенные гидрофобные группы R (например, аланин) более гидрофильны (т.е. имеют более высокую гидрофильность или более низкую гидрофобность), чем другие гидрофобные группы R (например, валин или лейцин). Относительная гидрофильность или гидрофобность может быть определена, используя общепризнанные в данной области техники способы (см., например, Rose et al., Science, 229:834-838 (1985) и Cornette et al., J. Mol. Biol., 195: 659-685 (1987)).
Как используется в настоящем описании, если одна аминокислотная последовательность (например, первая последовательность VH или VL) выравнивается с одной или несколькими дополнительными аминокислотными последовательностями (например, одной или несколькими последовательностями VH или VL в базе данных), положение аминокислоты в одной последовательности (например, первой последовательности VH или VL) может быть сравнена с «соответствующим положением» в одной или нескольких дополнительных аминокислотных последовательностях. В том смысле, в котором используется в настоящем описании, «соответствующее положение» представляет эквивалентное положение в подлежащей(их) сравнению последовательности(ях), когда последовательности оптимально выровнены, т.е. когда последовательности выравниваются для достижения наивысшего процента идентичности или процента сходства.
Термин «молекула нуклеиновой кислоты» относится к молекулам ДНК и молекулам РНК. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, но, предпочтительно, двухцепочечной ДНК. Нуклеиновая кислота является «функционально связанной», когда она находится в функциональной связи с другой последовательностью нуклеиновой кислоты. Например, промотор или энхансер является функционально связанным с кодирующей последовательностью, если он оказывает влияние на транскрипцию данной последовательности.
Термин «вектор» относится к молекуле нуклеиновой кислоты, способной участвовать в переносе другой нуклеиновой кислоты, с которой она была связана. Один тип вектора представляет собой «плазмиду», которая относится к кольцевой двухцепочечной замкнутой цепи ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другой тип вектора представляет собой вирусный вектор, где дополнительные ДНК сегменты могут быть лигированы в вирусный геном. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальную точку начала репликации и эписомальные векторы млекопитающих). Другие векторы (например, отличные от эписомальных векторы млекопитающих) могут быть вставлены в геном клетки-хозяина после введения в клетку-хозяина, и вследствие этого реплицируются вместе с хозяйским геномом.
Термин «клетка-хозяин» относится к клетке, в которую был введен вектор эксперссии. Клетки-хозяева могут включать бактериальные, микробные, растительные или животные клетки. Бактерии, которые поддаются трансформации, включают представителей энтеробактерий, таких как штаммы Escherichia coli или Salmonella; Bacillaceae, таких как Bacillus subtilis; Pneumococcus; Streptococcus и Haemophilus influenzae. Подходящие микробы включают Saccharomyces cerevisiae и Pichia pastoris. Подходящие животные линии клеток-хозяев включают CHO (линии яичника китайского хомячка) и клетки NS0.
Термины «лечить» и «лечение» относятся к терапевтическим или профилактическим мерам, описанным в настоящем документе. Способы «лечения» используют введение индивиду, при необходимости, антитела по настоящего изобретению, например, индивиду, имеющему GPCR-опосредованное нарушение, или индивиду, у которого в конечном счете может возникнуть такое нарушение, для профилактики, устранения, задержки, уменьшения тяжести или улучшения одного или нескольких симптомов заболевания, или повторение заболевания, или для увеличения продолжительности жизни индивида выше таковой, ожидаемой в отсутствие такого лечения.
Термин «эффективная доза» или «эффективная дозировка» относится к количеству, достаточному для достижения или, по меньшей мере, частичного достижения желаемого эффекта. Термин «терапевтически эффективная доза» определяется как количество, достаточное для устранения или, по меньшей мере, частичного подавления заболевания и его осложнений у пациента, уже страдающего заболеванием. Количества, эффективные для этого использования, будут зависеть от тяжести заболевания, подлежащего лечению, и общего состояния собственной иммунной системы пациента.
Термин «индивид» относится к любому человеку или животному, отличному от человека. Например, способы и композиции по настоящему изобретению могут быть использованы для лечения индивида с GPCR-опосредованным нарушением.
Термин «кролик» в том смысле, в котором он используется в настоящем описании, относится к животному, принадлежащему семейству leporidae.
Термин «клеточный сортер» относится к любому способу разделения клеток, основанному на присутствии детектируемой «сортируемой» метки. Такие способы включают, помимо прочего, клеточный сортер с возбуждением флуоресценции. Любые клеточные метки могут быть использованы в качестве сортируемых меток, включая, помимо прочего, флуоресцентные белки, например, зеленый флуоресцентный белок, конъюгаты антитело/фтор и флуоресцентные клеточные метки, например, флуоресцентные кальциевые ионофоры.
Термин «клеточный комплекс» относится к одной или нескольким антиген-экспрессирующим клеткам, связанным с одной или несколькими клетками, экспрессирующими иммунное связующее вещество, где связывание опосредовано антигеном на поверхности антиген-экспрессирующей клетки. В некоторых вариантах осуществления изобретения связывание антиген-экспрессирующей клетки с клеткой, экспрессирующей иммунное связующее вещество, состоит из прямого взаимодействия между антигеном на поверхности антиген-экспрессирующей клетки и иммунным связующим веществом на поверхности клетки, экспрессирующей иммунное связующее вещество.
Термин «клональное выделение» относится к любому способу выделения индивидуальных клеточных клонов из клеточной популяции. Подходящие способы включают, помимо прочего, предельное разведение и перенос клеток в мультилуночные плашки так, что каждая лунка содержит не более чем одну клетку.
Термин «получение последовательности нуклеиновой кислоты, кодирующей иммунное связующее вещество» относится к любому способу получения последовательности нуклеиновой кислоты иммунного связующего вещества, экспрессируемого клеткой, экспрессирующей иммунное связующее вещество. Подходящие способы включают, помимо прочего, выделение нуклеиновой кислоты, ПЦР-амплификацию и ДНК-секвенирование последовательности нуклеиновой кислоты, кодирующей иммунное связующее вещество, из клетки, экспрессирующей иммунное связующее вещество. В некоторых вариантах осуществления изобретения последовательности нуклеиновых кислот, кодирующих иммунные связующие вещества, амплифицируют с помощью ПЦР единичных клеток, т.е. «ПЦР одной клетки».
Термин «экзогенный антиген» относится к антигену, обычно не экспрессирующегося в определенной клетке-хозяине. Например, экзогенный антиген может быть из другого царства, типа класса, отряда, рода или вида из клетки-хозяина, например, человеческий антиген, экспрессируемый в дрожжевой клетке. Дополнительно или альтернативно экзогенный антиген может быть из того же самого вида, но неуместно экспрессирующийся в этой клетке-хозяине, например, специфичный для легкого антиген, экспрессирующийся в клетке мозга. «Экзогенный антиген» также относится к мутантному антигену, обычно не обнаруживаемому в нормальной клетке, например, специфичный для рака мутантный антиген, экспрессирующийся в клетке легкого.
Термин «полученный способами генной инженерии антиген» относится к любому антигену, который был получен способами рекомбинантных ДНК, и включает антигены, которые являются химерами или содержат точечную мутацию, делеции и/или вставки. Если не указано иное, все технические и научные термины, используемые в настоящем описании, имеют то же самое значение, как обычно понимается одним из специалистов в данной области техники, которому принадлежит данное изобретение. Хотя способы и материалы, сходные или эквивалентные тем, что описаны в настоящем документе, могут быть использованы на практике или при тестировании настоящего изобретения, подходящие способы и материалы описаны ниже. В случае наличия расхождений, настоящее описание, включая определения, будет обеспечивать контроль. Кроме того, материалы, способы и примеры являются только иллюстративными и не предназначены для ограничения притязаний.
Различные аспекты изобретения описаны более подробно в приведенных ниже подразделах. Понятно, что различные варианты осуществления изобретения, предпочтения и диапазоны могут быть объединены по желанию. Более того, в зависимости от конкретного варианта осуществления изобретения, выбранные определения, варианты осуществления или диапазоны можно не применять.
Настоящее изобретение относится к способу скрининга, используя FACS, для идентификации и фракционирования клеток, экспрессирующих иммунное связующее вещество, в плотном соединении с клетками, экспрессирующими соответствующий антиген. В частном варианте осуществления изобретения, иммунным связующим веществом является антитело.
Экспрессия антигена
Мишеневым антигеном для получения антитела может быть любой белок, пептид, нуклеотид, углевод, липид и другие молекулы, которые являются растворимыми или экспрессируются на клеточной поверхности или встроены в плазматическую мембрану. Антигены могут быть нативными или синтетическими. Предпочтительно мишеневый антиген является белком или пептидом. Неограниченные примеры мишеневого антигена включают CXCR1, CXCR2, CXCR3, CXCR4, CXCR6, CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR8, CFTR, CIC-1, CIC-2, CIC-4, CIC-5, CIC-7, CIC-Ka, CIC-Kb, бестрофины, TMEM16A, рецептор GABA, рецептор глицина, АВС переносчики, NAV1.1, NAV1.2, NAV1.3, NAV1.4, NAV1.5, NAV1.6, NAV1.7, NAV1.8, NAV1.9, рецептор сфингозин-1-фосфата (S1P1R), канал NMDA и т.д. В одном варианте осуществления мишеневый антиген представляет собой трансмембранный белок. В другом варианте осуществления изобретения мишеневый антиген представляет собой многократно пронизывающие мембрану трансмембранные белки, например, сопряженные с G-белком рецепторы (GPCR), ионные каналы и т.д.
Семейство GPCR состоит, по меньшей мере, из 250 членов (Strader et al. FASEB J., 9:745-754, 1995; Strader et al. Annu. Rev. Biochem., 63:101-32, 1994). На данный момент установлено, что один процент генов человека может кодировать GPCR. GPCR связываются с большим разнообразием лигандов, начиная от фотонов, малых биогенных аминов (т.е. эпинефрина и гистамина), пептидов (т.е. IL-8) до больших гликопротеиновых гормонов (т.е. паратиреоидного гормона). При связывании с лигандом GPCR регулируют внутриклеточные сигнальные пути путем активации гуанин нуклеотид-связывающих белков (G-белков). Примечательно, что GPCR имеет функциональных гомологов в цитомегаловирусе и герпесвирусе человека, предполагая, что GPCR появились во время эволюции для вирусного патогенеза (Strader et al., FASEB J., 9:745-754, 1995; Arvanitakis et al. Nature, 385:347-350, 1997; Murphy, Annu. Rev. Immunol. 12:593-633, 1994).
Характерным свойством большинства GPCR, известных на сегодняшний день, является то, что семь кластеров гидрофобных аминокислотных остатков расположены в первичной структуре и проходят сквозь (прошивают) клеточную мембрану в каждой их области. Считается, что домены представляют собой трансмембранные альфа-спирали, соединенные тремя внутриклеточными петлями, тремя внеклеточными петлями, и амино- и карбоксил-концевые домены (K. Palczewski et al., Science 289, 739-45 (2000)). Большинство GPCR имеет по одному консервативному цистеиновому остатку в каждой из первых двух внеклеточных петель, которые образуют дисульфидные связи, которые, как полагают, стабилизируют функциональную структуру белка. 7 трансмембранных участков обозначают как ТМ1, ТМ2, ТМ3, ТМ4, ТМ5, ТМ6 и ТМ7. Хорошо известно, что эти структуры, описанные выше, являются общими среди рецепторных белков, сопряженных с G-белком, и что аминокислотные последовательности, соответствующие участку, где белок проходит сквозь мембрану (охватывающий мембрану участок или трансмембранный участок), и аминокислотные последовательности вблизи трансмембранного участка, как правило, являются высоко консервативными среди рецепторов. Таким образом, из-за высокой степени гомологии в GPCR идентификация новых GPCR, а также идентификация как внутриклеточной, так и внеклеточной частей таких новых представителей семейства, легко осуществляется специалистами в данной области техники. В качестве примера, книга Watson and Arkinstall (1994), включенная в настоящее описание в качестве ссылки, представляет последовательности свыше 50 GPCR. Книга также описывает для каждой последовательности точные остатки, входящие в состав каждого из трансмембранных доменов.
Считается, что сайты связывания для малых лигандов в рецепторах, сопряженных с G-белком, включают гидрофильное углубление, расположенное вблизи внеклеточной поверхности и образованное трансмембранными доменами нескольких сопряженных с G-белком рецепторов, где углубление окружено гидрофобными остатками сопряженных с G-белком рецепторов. Считается, что гидрофильная сторона каждой трансмембранной спирали сопряженного с G-белком рецептора обращена вовнутрь и образует полярный сайт связывания лиганда. ТМ3 задействован в нескольких сопряженных с G-белком рецепторах, имея лиганд-связывающий сайт, в том числе такой, как остаток аспартата ТМ3. Кроме того, серины ТМ5, аспарагин ТМ6 и фенилаланины или тирозины ТМ6 или ТМ7 также вовлечены в связывание лиганда. Сайт связывания лиганда для рецепторов пептидных гормонов и рецепторов с другими большего размера лигандами, такими как гликопротеины (LH, FSH, hCG, TSH), и классов рецепторов Ca2+/глутамат/GABA, по-видимому, находится во внеклеточных доменах и петлях.
Основным событием при переключении из неактивного в активный рецептор является лиганд-индуцированные конформационные изменения трансмембранных спиралей 3 (ТМ3) и 6 (ТМ6) GPCR, которые имеют 7 пронизывающих мембрану трансмембранных спиралей (U. Gether and B. K. Kolbilka, J. Biol. Chem. 273, 17979-17982 (1998)). Эти изменения спиралей, в свою очередь, изменяют конформацию внутриклеточных петель рецептора для инициации активации ассоциированных гетеротримерных G-белков. Исследования мутагенеза (S. Cotecchia, J. Ostrowski, M. A. Kjelsberg, M. G. Caron and R. J. Lefkowitz, J. Biol. Chem. 267, 1633-1639 (1992); E. Kostenis, B. R. Conklin and J. Wess, Biochemistry 36, 1487-1495 (1997); M. A. Kjelsberg, S. Coteechia, J. Ostrowski, M. G. Caron, and R. J. Lefkowitz, J. Biol. Chem. 267, 1430-1433 (1992)) показали, что третья внутриклеточная петля (i3) опосредует большую часть сопряжения между рецептором и G-белком. Также было показано, что петли i3, экспрессирующиеся как минигены, непосредственно конкурируют с адренергическими рецепторами за связывание Gq (L. M. Luttrell, J. Ostrowski, S. Cotecchia, H. Kendal and R. J. Lefkowitz, Science 259, 1453-1457 (1993)), или могут активировать G-белки как растворимые пептиды в бесклеточных условиях (T. Okamoto et al., Cell 67, 723-730 (1991)).
Мишеневый антиген может быть эндогенного происхождения в клетке-мишени (в ряде случаев называемый антиген-экспрессирующей клеткой). Альтернативно, экзогенные молекулы могут быть введены в клетки для экспрессии антигена. Введение антигена в клетки может быть осуществлено любым способом, известным специалисту в данной области техники. В одном из вариантов осуществления изобретения полинуклеотид, кодирующий антиген в качестве полипептида, может быть вставлен in vitro в вектор, который может быть затем введен в клетки-мишени для экспрессии. Полинуклеотид может содержать последовательность кДНК, последовательность ДНК или другие последовательности, известные в данной области техники, мишеневого антигена. Вектор может представлять собой плазмиду, космиду, липосому или другие природные или искусственные вектора, известные в данной области техники. Введение может представлять собой процесс трансфекции, трансформации, инфекции, прямой микроинъекции веществ, биолистическую доставку частиц, электропорацию и другие способы, известные в данной области техники. Клетка-мишень, экспрессирующая антиген, может быть любой клеткой, известной в данной области техники, включая, например, клетки, напрямую полученные из животных, например, раковые клетки, нераковые клетки, эмбриональные клетки и т.д., или клетки, полученные молекулярно-инженерным способом, например, клетки культуры (например, клетки яичника китайского хомячка (СНО), клетки НЕК293 и т.д.), иммортализованные клетки, трансфицированные/инфецированные клетки, Т-клетки и т.д. Альтернативно, клетка-мишень может быть неживотного происхождения, например, бактериального, из насекомых и т.д. В одном из вариантов осуществления изобретения клетки, экспрессирующие мишеневый антиген, представляют собой дрожжевые клетки, предпочтительно, дрожжевые сферобласты. Альтернативно, «клетка»-мишень может быть искусственным клеточноподобным образованием или структурой, например, липосомой, однослойным мембранным образованием и т.д. Экспрессия антигена в клетке-мишени или клеточноподобных образованиях может быть кратковременной, т.е. экспрессия будет ослабевать или прекращаться после сравнительно короткого периода времени (например, в диапазоне от минут до нескольких дней), или стабильной, т.е. экспрессия будет поддерживаться на относительно стабильном уровне сравнительно продолжительное время (например, после нескольких дней или нескольких поколений клеток). В одном из предпочтительных вариантов осуществления изобретения антиген экспрессируется на внеклеточной поверхности плазматической мембраны клетки-мишени. В другом предпочтительном варианте осуществления изобретения антиген является интегральным или многократно пронизывающим мембрану антигеном. Для того чтобы добраться до его местоположения на плазматической мембране, антиген может быть экспрессирован непосредственно в данных местах или может быть транслоцирован в эти местоположения после его экспрессии в цитоплазме клетки-мишени. Эта транслокация может быть естественным процессом в клетке-мишени или сконструированным процессом, например, путем прикрепления сигнальной/маркерной молекулы (например, сигналы сортинга в аппарате Гольджи, антитела к определенным мембраносвязанным молекулам и т.д.), якорного трансплантата (например, якорного гликозилфосфатидилинозитола (GPI)), или химической сшивки с антигеном, видоизменяя антиген, или другими способами, известными в данной области техники, до или после экспрессии антигена, которая приводит к его транслокации.
Клетки, экспрессирующие иммунное связующее вещество, и иммунизация
В одном из вариантов осуществления изобретения экспрессирующие иммунное связующее вещество клетки, подлежащие отбору способом, описанным в настоящем документе, представляют собой В-клетки, предпочтительно В-клетки кролика.
В предпочтительном варианте осуществления изобретения В-клетки происходят из животного, которое было иммунизировано мишеневой молекулой. Иммунизация животного может быть проведена любым способом, известным специалисту в данной области техники. Как правило, В-клетки выделяют из лимфатических органов иммунизированного животного (таких как селезенка и лимфатические узлы).
В одном из предпочтительных вариантов осуществления изобретения иммунизация мишеневым антигеном выполняется ДНК-иммунизацией/вакцинацией. Альтернативно, клетки, экспрессирующие мишеневый антиген, инъецируют в животных (например, кролика, крысу, мышь, хомячка, овцу, козу, курицу и т.д.) для иммунизации. Предпочтительным животным для данного этапа иммунизации является кролик. ДНК-иммунизация/вакцинация индуцирует быстрый иммунный ответ и создает возможность для нативной экспрессии, и, как правило, только нативной экспрессии, мишеневых антигенов. Поскольку она не включает экспрессию и использование рекомбинантных белков, этот процесс более эффективный и экономически выгодный, нежели общепринятая иммунизация рекомбинантными белками. Кроме того, что еще более важно, экспрессированный in vivo антиген обладает такой же вторичной структурой и может даже обладать теми же посттрансляционными модификациями, что и мишеневый белок в естественных условиях, что улучшает точность узнавания полученными антителами против мишеневого антигена. Иллюстративная ДНК-иммунизация показана в канадской патентной заявке СА2350078 и в WO04/087216. В частности, ДНК, кодирующая полипептид в виде мишеневого антигена, вводится непосредственно в животное способом генной пушки, приводя к экспрессии полипептида в животном, экспрессия которого вызывает образование антител против полипептида. Для достижения более интенсивного образования антител, так называемые, генетические адъюванты применяются одновременно с кодирующей полипептид ДНК. Эти генетические адъюванты представляют собой плазмиды, которые экспрессируют цитокины (например, GM-CSF, IL-4 и IL-10) и которые стимулируют гуморальный иммунный ответ в лабораторных животных. В одном из предпочтительных вариантов осуществления изобретения антитело против мишеневого антигена экспрессируется на клеточной поверхности В-клеток в виде В-клеточного рецептора (BCR). В другом предпочтительном варианте осуществления изобретения экспрессирующая антитело клетка представляет собой В-клетку памяти, характеризующуюся и отличающуюся от обычных В-клеток отсутствием любого IgM на ее поверхности.
В одном из вариантов осуществления изобретения клетки, экспрессирующие иммунное связующее вещество, представляют собой дрожжевые клетки, и иммунные связующие вещества являются предпочтительно фрагментами антитела, предпочтительнее scFv.
Скрининг, используя сортировку клеток с возбуждением флуоресценции (FACS)
После этапа иммунизации мембраносвязанное антитело на В-клетках, которое специфически связывается с мишеневым антигеном, необходимо отделить от других клеток, экспрессирующих неспецифические антитела. В одном из предпочтительных вариантов осуществления изобретения посредством связывания антиген-антитело В-клетки, экспрессирующие специфическое антитело на их плазматической мембране, присоединяются к клеткам-мишеням, экспрессирующим антиген. В другом предпочтительном варианте осуществления изобретения другие или большее количество взаимодействий (например, те же или другие взаимодействия антиген-антитело, химические сшивки, лиганд-рецепторные взаимодействия и т.д.) могут происходить между В-клетками и клетками-мишенями. В-клетки могут находиться в пуле В-клеток, экспрессирующих различные антитела, или в сочетании с другими иммунными клетками, собранными непосредственно из иммунизированного животного, из пула иммунизированных/неиммунизированных животных или в результате генно-инженерных процессов in vitro, например, библиотеки В-клеток, экспрессирующих различные антитела способом генетической рекомбинации V(D)J.
Фракционирование В-клеток, экспрессирующих антитела, специфичные к мишеневому антигену, может быть выполнено любым способом, известным в данной области техники. Они включают, помимо прочего, пэннинг на антигене, предельное разведение, аффинную очистку или другие способы, использующие характеристики экспрессируемых антител или продуцирующих антитело В-клеток.
В одном из предпочтительных вариантов осуществления изобретения антиген-экспрессирующие клетки метят маркером, подходящим для дальнейшего выявления и/или выделения присоединившихся В-клеток. Маркером может быть сшивка, антиген/антитело, небольшие молекулы (например, глутатион (GSH), биотин/авидин и т.д.), магнитные частицы, флуоресцентный маркер и т.д. В одном из предпочтительных вариантов осуществления изобретения антиген-экспрессирующие клетки метят флуоресцентным белком/пептидом. В другом варианте осуществления изобретения продуцирующие антитело В-клетки также метят маркером, предпочтительно другим флуоресцентным белком/пептидом. Еще в одном варианте осуществления изобретения В-клетки в плотном соединении с антиген-экспрессирующими клетками могут быть выявлены путем эмиссии флуоресценции из флуоресцентно меченого белка/пептида на В-клетках. Еще в одном варианте осуществления изобретения В-клетка и плотно связанная с ней антиген-экспрессирующая клетка могут быть выявлены путем эмиссии флуоресценции из двух различных флуоресцентно меченых белков/пептидов на них. В одном предпочтительном варианте осуществления изобретения В-клетки, экспрессирующие антитела, специфичные к мишеневому антигену, могут быть выявлены и затем отделены от других продуцирующих антитело В-клеток путем эмиссии обеих флуоресценций из флуоресцентно меченых белков/пептидов на них и на присоединенных к ним АРС. Предпочтительно данное выявление и фракционирование могут быть выполнены способом сортировки клеток с возбуждением флуоресценции (FACS).
Аббревиатура FACS официально зарегистрирована в качестве товарного знака и принадлежит на правах собственности Becton Dickinson (Franklin Lakes, NJ). Термин FACS в том смысле, в котором он используется в настоящем описании, означает любой вид клеточной сортировки, основанной на проточной цитометрии.
Сортировка клеток с возбуждением флуоресценции представляет собой особый тип проточной цитометрии. Она представляет способ сортировки гетерогенной смеси биологических клеток в два или большее количество контейнеров, одна клетка за раз, основанный на специфическом светорассеянии и флуоресцентных характеристиках каждой клетки. Это эффективное инструментальное средство для научных исследований, поскольку оно обеспечивает быструю, объективную и количественную регистрацию флуоресцентных сигналов из индивидуальных клеток, а также физическое фракционирование клеток, представляющих особый интерес.
В стандартной системе FACS клеточная суспензия находится в центре узкого, быстро текущего потока жидкости. Поток организован таким образом, что существует значительное расстояние между клетками относительно их диаметра. Вибрирующий механизм заставляет поток клеток распадаться на индивидуальные капли. Система отрегулирована таким образом, чтобы имела место низкая вероятность нахождения в капле более чем одной клетки. Перед тем как поток распадается на капли, поток проходит сквозь измеряющую флуоресценцию станцию, где измеряется флуоресцентный представляющий интерес параметр каждой клетки. Электрическое зарядное кольцо расположено непосредственно в том месте, где поток распадается на капли. Кольцо заряжают на основании выполненных, непосредственно до этого, измерений интенсивности флуоресценции, и противоположный заряд улавливается на капле по мере ее образования из потока. Заряженные капли затем проходят сквозь систему электростатического отклонения, которая направляет капли в контейнеры на основании их заряда. В некоторых системах заряд наносят непосредственно на поток, и образующиеся из потока капли сохраняют заряд того же знака, что и поток. Поток затем снова становится нейтральным после распада на капли.
Флуоресцентные метки для способа FACS зависят от лампы или лазера, используемых для возбуждения флуорохромов, и от имеющихся в наличии детекторов. Наиболее общепринятыми, годными для использования лазерами на одномодовых лазерных установках являются голубые аргоновые лазеры (488 нм). Флуоресцентные метки, применяемые для лазеров этого вида, включают помимо прочего 1) для зеленой флуоресценции (обычно обозначенной FL1): FITC, Alexa Fluor 488, GFP, CFSE, CFDA-SE и DyLight 488; 2) для оранжевой флуоресценции (обычно FL2): PE и PI; 3) для красной флуоресценции (обычно FL3): PerCP, PE-Alexa Fluor 700, PE-Cy5 (TRI-COLOR), и PE-Cy5.5; и 4) для инфракрасной флуоресценции (обычно FL4; в некоторых установках FACS): PE-Alexa Fluor 750 и PE-Cy7. Другие лазеры и их соответствующие флуоресцентные метки включают, помимо прочего, 1) красные полупроводниковые лазеры (635 нм): аллофикоцианин (АРС), АРС-Cy7, Alexa Fluor 700, Cy5 и Draq-5; и 2) фиолетовые лазеры (405 нм): Pacific Orange, Amine Aqua, Pacific Blue, 4',6-диамидино-2-фенилиндол (DAPI) и Alexa Fluor 405.
В предпочтительном варианте осуществления изобретения В-клетки окрашивают мечеными анти-IgG и анти-IgM антителами, и В-клетки памяти, будучи только положительно окрашены анти-IgG антителами, но не анти-IgM антителами, предпочтительно отбираются. IgG, как правило, обладают более высокой аффинностью, нежели IgM; положительные В-клетки, экспрессирующие IgG, но не IgM, на своей поверхности (что является характеристикой В-клеток памяти) посредством этого отбираются. Для указанной цели предпочтительно используется многоцветное окрашивание, где антитела, специфичные для IgG и IgM, дифференциально мечены, например, с использованием APC и FITC, соответственно. Предпочтительно антиген-мишень и/или клетку-мишень, экспрессирующую мишеневый антиген, также метят. В одном варианте осуществления изобретения антиген-мишень окрашивают опосредованно путем окрашивания клетки, которая экспрессирует антиген-мишень с внутриклеточным флуоресцентным красителем.
Настоящее изобретение представляет способ использования FACS для отбора пулов В-клеток, в которых В-клетки могут находиться в плотном соединении с клетками, экспрессирующими антигены-мишени, для идентификации и дальнейшего выделения В-клеток, продуцирующих антитела, специфически связывающих представляющий интерес антиген-мишень. Предпочтительно В-клетки метят флуоресцентной меткой, и клетки, экспрессирующие антиген-мишень, отдельно метят другой флуоресцентной меткой. Эти метки могут быть либо внутриклеточными, внеклеточными, либо встроенными в плазматическую мембрану. После иммунизации и продукции антител все В-клетки объединяют вместе и пропускают через систему FACS. Только те В-клетки, которые продуцируют анитела, специфичные к антигену-мишени, будут плотно соединяться с антиген-экспрессирующими клетками. Их плотное взаимодействие укорачивает расстояние между этими двумя клетками в потоке по сравнению со значительным расстоянием между другими индивидуальными клетками, приводя к «двухцветному явлению», детектируемому во время их совместного прохождения сквозь сканирующий лазерный луч. Таким образом, В-клетки, продуцирующие мишеневые антитела, могут быть идентифицированы и затем отсортированы от других неспецифичных В-клеток в другую пробирку для сбора образцов.
В другом предпочтительном варианте осуществления изобретения, если взаимодействие между В-клеткой и соответствующей антиген-экспрессирующей клеткой приводит к определенным модификациям клеточных характеристик, например, деполяризации, резонансному переносу энергии флуоресценции (FRET) и др., больше флуоресцентных меток может быть добавлено к клетке, способной к такой модификации. Таким образом, В-клетки и антиген-экспрессирующие клетки в контакте друг с другом будут выдавать «трехцветное явление» или явление, включающее даже более трех цветов единовременно.
Альтернативно, идентификация и сортировка В-клеток в плотном контакте не требует наличия флуоресцентной метки. В одном варианте осуществления изобретения межклеточное взаимодействие приводит к функциональным изменениям в любой из клеток. В другом варианте осуществления изобретения эти функциональные изменения могут быть использованы для идентификации и дальнейшего фракционирования В-клеток, продуцирующих антитела, специфически связывающих антиген-мишень. Например, взаимодействие клетка-клетка может функционально блокировать или активировать передачу сигнала через рецептор в любой из клеток, приводя к клеточным изменениям, например, изменениям выброса Ca2+ и т.д., детектируемым системой FACS. Таким образом, путем мониторинга этих детектируемых функциональных изменений мишеневые В-клетки могут также быть идентифицированы и фракционированы. Один частный вариант осуществления изобретения функционального блокирования или активации передачи сигнала через рецептор включает инкубацию В-клеток с клетками, которые функционально экспрессируют GPCR (сопряженный с G-белком рецептор). Агонист, который передает сигнал через GPCR, может быть добавлен в смесь для индукции опосредованного через GPCR выброса Ca2+ из эндоплазматического ретикулума. В случае если антитело, представленное на В-клетке, функционально будет блокировать передачу сигнала агонистом, выброс Ca2+ будет в результате также блокирован путем этого межклеточного взаимодействия. Выброс Ca2+ может, например, быть количественно измерен проточной цитометрией. Ввиду этого, только конгломераты В-клетка/клетка-мишень, которые либо демонстрируют увеличение или уменьшение выброса Ca2+, будут отобраны.
Анализ аффинности антител, продуцируемых выделенными В-клетками
В определенных вариантах осуществления изобретения В-клетки культивируют в подходящих условиях так, чтобы происходила секреция антител в культуральную среду. Продуцируемые антитела представляют собой, например, моноклональные антитела. Культивирование может включать использование хелперной клеточной линии, такой как хелперная клеточная линия тимомы (например, EL4-B5, см. Zubler et al, 1985, J. Immunol., 134 (6): 3662-3668).
Факультативно, дополнительный анализ аффинности может быть выполнен перед дальнейшей обработкой с целью оценки селективности и способности антител, продуцируемых выделенными В-клетками, конкурировать с лигандом. Эти анализы включают, помимо прочего, клеточные анализы (например, ELISA клетки (CELISA), который является модификацией процесса ELISA, где клетки целиком используются для покрытия). Как указано в СА2350078, CELISA может проводиться, как описано ниже в Примерах. Стадию проверки проводят для тестирования образованных антител на специфическое связывание с мишенью, например, для исключения антител, которые направлены против белка, экспрессируемого на клеточной поверхности, отличного от белка-мишени.
Альтернативно, идентифицированные и выделенные мишеневые В-клетки могут быть напрямую исследованы на аффинность антитела, и В-клетки могут быть отделены от плотно связанных с ними антиген-экспрессирующих клеток до исследования.
Другие исследования выделенных В-клеток на продукцию антител
Идентифицированные и выделенные В-клетки, дополнительно протестированные с помощью анализа аффинности (например, CELISA), могут быть также обработаны для продукции мишеневых иммунных связующих веществ. Может быть использован, например, стандартный гибридомный способ. Он может включать стадии, такие как очистка иммунных связующих веществ, выяснение их аминокислотной последовательности и/или последовательности нуклеиновой кислоты.
Альтернативно, характеристика связующих веществ выполняется в виде их scFv. Для этого способа последовательности CDR связующих веществ, экспрессирующихся на отобранных В-клетках, воспроизводят способом ОТ-ПЦР из любых культивируемых отобранных клеток или из единичных клеток напрямую. Объединение двух пулов частично перекрывающихся олигонуклеотидов, в которых один олигонуклеотидный пул кодирует CDR и второй пул кодирует каркасные области пригодной scFv-подложки, позволит получить гуманизированный scFv в ходе одностадийной процедуры ПЦР. HT секвенирование, клонирование и продукция позволят выполнить отбор клона, основанный на рабочих характеристиках очищенного гуманизированного scFv вместо получения характеристик секретируемых IgG в супернатанте клеточной культуры. Подложка scFv, пригодная для акцептирования CDR из любого кроличьего антитела, была идентифицирована и охарактеризована («окрольчаченный» человеческий FW или кроличий актептор RabTor; см. WO09/155726, который включен в настоящее описание полностью в качестве ссылки). Было продемонстрировано доказательство концепции о различных CDR, которые в некоторых случаях даже содержат кроличьи специфические дисульфидные связи между CDR.
Основное описание создания окрольчаченного антитела описано ниже.
Пересадка иммунных связующих веществ
Области связывания антигена или CDR иммунных связующих веществ, идентифицированных способами по изобретению, могут быть пересажены в акцепторные каркасные участки антитела. Такая пересадка может, например, снизить иммуногенность иммунного связующего вещества или улучшить его функциональные свойства, например, улучшить термодинамическую стабильность.
Основные способы пересадки CDR в человеческие акцепторные каркасные участки были описаны Winter в патенте США № 5225539, который включен в настоящее описание полностью в качестве ссылки.
Характерные принципы пересадки CDR из кроличьих моноклональных антител описаны в предварительных патентных заявках США №№ 61/075697 и 61/155041, которые включены в настоящее описание полностью в качестве ссылки. Эти принципы относятся к таковым Winter, но отличаются тем, что акцепторные каркасные участки антитела особенно пригодны в качестве универсального акцептора для всех человеческих и не имеющих отношение к человеку донорских антител. В частности, было показано, что человеческий одноцепочечный каркасный участок FW1.4 (комбинация SEQ ID NO: 1 (именуемая а43 в WO03/097697) и SEQ ID NO: 2 (именуемая KI27 в WO03/097697)) в высокой степени соответствует антиген-связывающим сайтам кроличьих антител. Таким образом, FW1.4 представляет подходящую подложку для создания стабильных гуманизированных фрагментов антитела scFv, полученных при пересадке кроличьих петель.
Более того, было обнаружено, что FW1.4 может быть оптимизирован путем замещения 5 или 6 положений остатков в тяжелой цепи FW1.4 и/или путем замещения 1 положения в легкой цепи FW1.4. Таким образом, было неожиданно обнаружено, что конформация петли богатого ассортимента кроличьих CDR в VH может быть полностью сохранена в значительной мере независимо от последовательности донорского каркасного участка. Указанные 5 или 6 остатки в тяжелой цепи, а также 1 положение в легкой цепи FW1.4 являются консервативными в кроличьих антителах. Консенсусный остаток для 5 или 6 положения в тяжелой цепи, а также одном положении в легкой цепи, был предсказан из кроличьего репертуара и введен в последовательность человеческого акцепторного каркасного участка. В результате модифицированный каркасный участок 1.4 (именуемый в настоящем описании rFW1.4) соответствует практически любому кроличьему CDR. В отличие от одиночных цепей дикого типа кролика, rFW1.4, содержащий различные кроличьи CDR, хорошо экспрессируется и практически полностью сохраняет аффинность исходных донорских кроличьих антител.
В связи с этим иллюстративные акцепторные каркасные участки иммунного связующего вещества включают
(i) каркасный участок вариабельной тяжелой цепи, имеющей, по меньшей мере, 70% идентичности, предпочтительно, по меньшей мере, 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере, 95% идентичности, последовательности SEQ ID NO: 1; и/или
(ii) каркасный участок вариабельной легкой цепи, имеющей, по меньшей мере, 70% идентичности, предпочтительно, по меньшей мере, 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере, 95% идентичности, последовательности SEQ ID NO: 2.
В предпочтительном варианте осуществления изобретения вариабельная легкая цепь содержит треонин (Т) в положении 87 (нумерация АНо).
В предпочтительном варианте осуществления изобретения указанный акцепторный каркасный участок иммунного связующего вещества включает
(i) каркасный участок вариабельной тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO: 1, SEQ ID NO: 4 и SEQ ID NO: 6; и/или
(ii) каркасный участок вариабельной легкой цепи SEQ ID NO: 2 или SEQ ID NO: 9.
В предпочтительном варианте осуществления изобретения каркасный участок вариабельной тяжелой цепи соединен с каркасным участком вариабельной легкой цепи через линкер. Линкером может быть любой подходящий линкер, например, линкер, включающий от 1 до 4 повторов последовательности GGGGS (SEQ ID NO: 10), предпочтительно пептид (GGGGS)4 (SEQ ID NO: 8), или линкер, как описано в Alfthan et al. (1995) Protein Eng. 8:725-731.
В наиболее предпочтительном варианте осуществления изобретения акцепторный каркасный участок иммунного связующего вещества представляет собой последовательность, имеющую, по меньшей мере, 70%, 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере, 95% идентичности SEQ ID NO: 3. Более предпочтительно акцепторный каркасный участок иммунного связующего вещества включает или представляет собой SEQ ID NO: 3.
В другом предпочтительном варианте осуществления изобретения акцепторный каркасный участок иммунного связующего вещества представляет собой последовательность, имеющую, по меньшей мере, 70%, 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере, 95% идентичности SEQ ID NO: 5. Более предпочтительно, акцепторный каркасный участок иммунного связующего вещества включает или представляет собой SEQ ID NO: 5.
В другом предпочтительном варианте осуществления изобретения акцепторный каркасный участок иммунного связующего вещества представляет собой последовательность, имеющую, по меньшей мере, 70%, 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере, 95% идентичности SEQ ID NO: 7. Более предпочтительно, акцепторный каркасный участок иммунного связующего вещества включает или представляет собой SEQ ID NO: 7.
Более того, может применяться типичный каркасный участок вариабельной тяжелой цепи SEQ ID NO: 1, дополнительно включающий один или несколько аминокислотных остатков, которые в целом поддерживают конформацию CDR, полученных из кроличьего иммунного связующего вещества. В частности, указанные остатки находятся в одном или нескольких аминокислотных положениях, выбранных из группы, состоящей из 24Н, 25Н, 56Н, 82Н, 84Н, 89Н и 108Н (нумерация АНо). Доказано, что эти положения влияют на конформацию CDR и поэтому рассматриваются для мутаций, чтобы вместить донорские CDR. Предпочтительно указанные один или несколько остатков выбирают из группы, состоящей из: треонина (Т) в положении 24, валина (V) в положении 25, глицина или аланина (G/A) в положении 56, лизина (К) в положении 82, треонина (Т) в положении 84, валина (V) в положении 89 и аргинина (R) в положении 108 (нумерация АНо).
В предпочтительном варианте осуществления изобретения указанный каркасный участок вариабельной тяжелой цепи представляет собой или включает SEQ ID NO: 4 или SEQ ID NO: 6. Оба каркасных участка вариабельной тяжелой цепи можно, например, комбинировать с любым подходящим каркасным участком легкой цепи.
Последовательности, рассмотренные выше, указаны ниже (остатки Х представляют собой сайты вставки CDR):
SEQ ID NO: 1: каркасный участок вариабельной тяжелой цепи FW1.4 (a43)
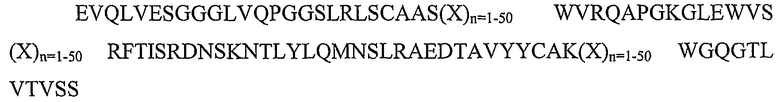
SEQ ID NO: 2: каркасный участок вариабельной легкой цепи FW1.4 (KI27)
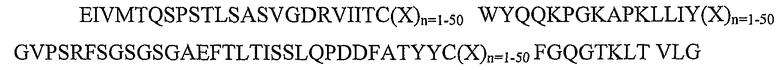
SEQ ID NO: 3: каркасный участок FW1.4

SEQ ID NO: 4: каркасный участок вариабельной тяжелой цепи rFW1.4
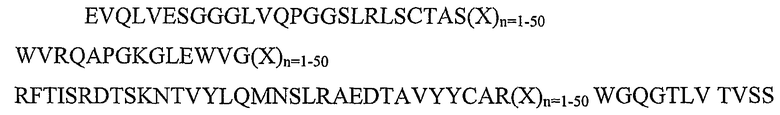
SEQ ID NO: 5: каркасный участок rFW1.4

SEQ ID NO: 6: каркасный участок вариабельной тяжелой цепи rFW1.4(V2)
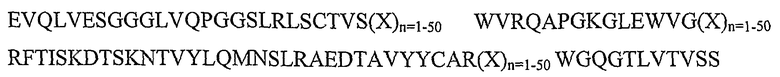
SEQ ID NO: 7: каркасный участок rFW1.4(V2)

SEQ ID NO: 8: линкер
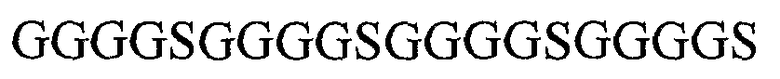
SEQ ID NO: 9: замещенный каркасный участок вариабельной легкой цепи FW1.4
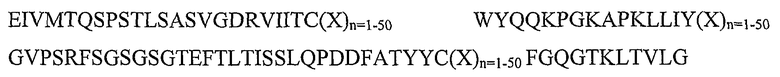
Таким образом, в отличие от основного способа Winter, последовательность каркасного участка, используемая для способов гуманизации по изобретению, не обязательно является последовательностью каркасного участка, которая проявляет наибольшее сходство с последовательностью не имеющего отношения к человеку (например, кроличьего) антитела, из которого получают донорские CDR. Кроме того, пересадка остатка каркасного участка из донорской последовательности для поддержания конформации CDR не требуется.
Каркасные участки акцепторного антитела могут также включать одну или несколько повышающих стабильность мутаций, описанных в предварительной патентной заявке США серийного № 61/075692, которая включена полностью в настоящее описание в качестве ссылки. Типичные повышающие растворимость замены в каркасном участке тяжелой цепи обнаружены в положениях 12, 103 и 144 (нумерация АНо). Более предпочтительно, если каркасный участок тяжелой цепи включает (а) серин (S) в положении 12; (b) серин (S) или треонин (Т) в положении 103 и/или (с) серин (S) или треонин (Т) в положении 144. Более того, повышающие стабильность аминокислоты могут быть расположены в одном или нескольких положениях 1, 3, 4, 10, 47, 57, 91 и 103 каркасного участка вариабельной легкой цепи (нумерация АНо). Более предпочтительно, когда каркасный участок вариабельной легкой цепи включает глутаминовую кислоту (Е) в положении 1, валин (V) в положении 3, лейцин (L) в положении 4, серин (S) в положении 10; аргинин (R) в положении 47, серин (S) в положении 57, фенилаланин (F) в положении 91 и/или валин (V) в положении 103.
ПРИМЕРЫ
Для всех без исключения примеров были использованы нижеприведенные материалы и способы, если не указано иное.
Материалы и способы
В целом, для осуществления на практике настоящего изобретения применяются, если не указано иное, стандартные способы химии, молекулярной биологии, способы рекомбинантных ДНК, иммунологии (в частности, например, технология антител) и стандартные способы получения полипептидов. См., например, Sambrook, Fritsch and Maniatis, Molecular Cloning: Cold Spring Harbor Laboratory Press (1989); Antibody Engineering Protocols (Methods in Molecular Biology), 510, Paul, S., Humana Pr (1996); Antibody Engineering: A Practical Approach (Practical Approach Series, 169), McCafferty, Ed., Irl Pr (1996); Antibodies: A Laboratory Manual, Harlow et al, C.S.H.L. Press, Pub. (1999); и Current Protocols in Molecular Biology, eds. Ausubel et al, John Wiley & Sons (1992).
ELISA клеток (CELISA)
Приведенный ниже пример описывает использование CELISA для анализа клеток, экспрессирующих CXCR2:
Клетки СНО, экспрессирующие CXCR2, могут быть засеяны плотностью 50000 клеток/лунка на половине площади 96-луночных плашек. После ночной инкубации при 37°С клетки могут быть зафиксированы 1% формальдегидом в PBS в течение 30 минут при комнатной температуре. Клеточные слои могут быть промыты три раза, и сайты неспецифического связывания могут быть заблокированы клеточной культуральной средой в течение часа при комнатной температуре. Далее, после трех стадий промывки супернатанты могут быть разведены 1:1 в культуральной среде и добавлены в лунки. В три контрольные лунки коммерческие кроличьи антитела анти-CXCR2 могут быть внесены в супернатант. Супернатанты могут быть затем инкубированы на клеточных слоях в течение полутора часов при комнатной температуре. В заключение, кроличьи IgG детектируют с помощью козьего антикроличьего антитела IgG Fc, соединенного с HRP. При добавлении пероксидазного субстрата (субстрат Blue POD от Roche) развивается колориметрическая реакция, которая может быть остановлена после 25 минут 1М HCl. Оптическая плотность может быть измерена при 450 нм.
Пример 1
Система скрининга В-клеток с использованием растворимых антигенов
Основанная на FACS (сортировка клеток с возбуждением флуоресценции) система скрининга, описанная в настоящем изобретении, приведена здесь в качестве примера и способна отбирать В-клетки, которые связываются с представляющей интерес мишенью, в частности, растворимым белком, посредством их В-клеточных рецепторов (BCR). В этом примере мишенью было одноцепочечное анти-VEGF антитело ESBA903, меченное флуоресцентным красителем (PE и PerCP). Суспензия лимфоцитов была приготовлена из селезенки кроликов, иммунизированных рекомбинантной мишенью. Клетки затем были инкубированы с scFv, мечеными PE и PerCP, а также с антителами, специфичными к IgG (АРС-мечеными) или IgM (мечеными FITC). ESBA903-положительные В-клетки, которые экспрессируют IgG, но не IgM на своей поверхности, были фракционированы и отобраны в 96-луночные плашки (фиг.2). Как показано на панели А фиг.2, лимфоциты были отобраны в соответствии с прямым и боковым светорассеянием. Среди них, IgG+ IgM- клетки (вероятно В-клетки памяти) были отобраны (панель В). Предполагалось, что клетки, окрашенные дважды scFv-PE и scFv-PerCP, кодируют высоко аффинные IgG против scFv (панель С). Клетки, демонстрирующие самую яркую флуоресценцию, были отобраны в 96-луночные плашки со статистическими данными отбора, перечисленными в панели D. Посредством хелперной клеточной линии тимомы (EL4-B5: см. Zubler et al, 1985, J. Immunol. 134(6):3662-3668), отобранные В-клетки пролиферировали, дифференцировались в плазматические клетки и затем секретировали антитела. Сродство этих молекул IgG к белку-мишени было проверено измерениями с помощью ELISA и Biacore. Кинетические параметры показаны в Таблице 1 для семи отобранных клонов. Эти клоны, из пула в ~200 отобранных клеток, демонстрируют высокие аффинности связывания в низком диапазоне значений от наномолярных до пикомолярных. В заключение, мРНК секретируемых молекул IgG была выделена из 6 мишеневых клонов, и CDR были перенесены в RabTor одноцепочечный каркасный участок ESBATech, также называемый каркасным участком rFW1.4.
Кинетические значения для супернатантов 7 В-клеточных культур
Статистические данные отбора для фиг.2
Выбранная литература:
Zuber et al. Mutant EL-4 thymoma cells polyclonally activate murine and human B cells via direct cell interaction. J Immunol. 1985;134(6):3662-3668.
Трансмембранные мишени
Системы скрининга, описанные выше, работают эффективно, когда мишень является растворимой, и когда имеется в наличии рекомбинантный белок. Однако некоторые представляющие интерес мишени являются многократно пронизывающими мембрану трансмембранными белками (например, GPCR и ионные каналы). Стандартная иммунизация рекомбинантным белком в этих случаях нецелесообразна и невозможна. Также, отбор способом FACS В-клеток не может быть выполнен, основываясь на связывании очищенных, меченых белков, если антиген представляет собой интегральный мембранный белок. В целях решения этих вопросов, были реализованы следующие усовершенствования вышеупомянутой методики.
1) Иммунизация с помощью ДНК вместо рекомбинантного белка:
ДНК-вакцинация индуцирует быстрый иммунный ответ против нативного антигена. Поскольку не требуется никакого рекомбинантного белка, эта технология, с одной стороны, экономически выгодна, с другой стороны, и что более важно, этот способ создает возможность для нативной экспрессии интегральных мембранных комплексов и/или многократно пронизывающих мембрану мембранных белков.
2) Отбор способом FACS В-клеток, которые связываются с клетками, экспрессирующими интегральный мембранный белок-мишень:
Проточная цитометрия обычно измеряет флуоресценцию, испускаемую единичными клетками, когда они пересекают лазерный луч. Однако некоторые исследователи уже использовали цитометры для исследования межклеточных взаимодействий, например, адгезии, опосредованной кадгеринами (Panorchan et al, 2006, J. Cell Science, 119, 66-74; Leong et Hibma, 2005, J. Immunol. Methods, 302, 116-124) или интегринами (Gawaz et al, 1991, J. Clin. Invest, 88, 1128-1134). Однако такие исследования не выявляют того, остаются ли такие межклеточные взаимодействия интактными во время физической стадии клеточной сортировки. Более того, никогда не было показано, что связывание В-клеточного рецептора с его мишенью, представленной на поверхности другой клетки, является достаточно плотным для создания возможности проведения такой физической сортировки.
С целью осуществления отбора В-клеток, которые связываются с трансмембранными мишенями, клетки (например, клетки СНО или HEK293) могут быть случайно или предпочтительно стабильно трансфицированы выбранной мишенью, или могут быть использованы клетки, которые естественным образом экспрессируют выбранную мишень. Такие клетки-мишени окрашивают внутриклеточным флуоресцентным красителем (например, кальцеином) и инкубируют с В-лимфоцитами памяти иммунизированного кролика. В-лимфоциты окрашивают флуоресцентными антителами, связывающимися со специфичными маркерами на клеточной поверхности. Таким образом, отбор двухцветных «явлений», происходящих в двух клетках, плотно взаимодействующих друг с другом посредством взаимодействий BCR-мишени (см. фиг.1), может быть осуществлен.
Дополнительная обработка этих В-клеток выполняется, как описано выше, которая приводит к продукции моноклональных антител, например, в виде IgG или scFv. Для количественной оценки сродства этих антител к мишени, выполняют CELISA (ELISA, где стадию поверхностного покрытия выполняют на целых клетках). С помощью этого способа можно оценить селективность и способность антител конкурировать с лигандом. Наконец, CDR представляющих интерес клонов клонируют в созданный авторами изобретения окрольчаченный каркасный участок (RabTor) путем генного синтеза способом удлинения олигонуклеотида.
Считывание данных по отбору В-клеток не обязательно ограничивается межклеточным взаимодействием, но может также быть использовано для отбора по способности этого взаимодействия функционально блокировать/активировать передачу сигнала через рецептор. Например, В-клетки могут быть инкубированы с клетками, которые функционально экспрессируют GPCR. Агонист, который проводит сигнал через GPCR, может быть добавлен в смесь для индукции GPCR-опосредованного выброса Ca2+ из эндоплазматического ретикулума. В случае, если антитело, представленное на В-клетке, функционально блокирует передачу сигнала агонистом, выброс Ca2+ будет в результате также блокирован этим межклеточным взаимодействием. Выброс Ca2+ может быть количественно измерен проточной цитометрией. Таким образом, будут отобраны только конгломераты В-клетка/клетка-мишень, которые не демонстрируют никакого выброса Ca2+.
Пример 2
Выявление взаимодействия между бусами, покрытыми антителами анти-TNFα, и клетками СНО, экспрессирующими мембраносвязанный TNFα
Перед началом скрининга В-клеток по отношению к трансмембранному белку, должно быть показано, что межклеточные взаимодействия (и, в частности, взаимодействия между BCR и трансмембранным белком на клетке-мишени) могут быть положительным образом выявлены способом FACS. Чтобы определить, разрушает ли высокое давление в потоке проточной цитометрии нековалентное связывание между двумя клетками, был выполнен нижеприведенный эксперимент.
Клетки СНО, стабильно трансфицированные мембраносвязанным TNFα (клетки В-220) (содержащие мутантный мембраносвязанный TNFα, который содержит точечную мутацию в сайте расщепления ТАСЕ, который предотвращает расщепление и нарушение прочности прикрепления лиганда TNFα; см., например, Scallon et al. J Pharmacol Exp Ther 2002; 301:418-26), были инкубированы с бусами, покрытыми РЕ-мечеными антителами анти-TNFα. В этой системе бусы имитируют В-клетки памяти. В качестве отрицательного контроля были использованы клетки СНО, а также бусы, покрытые не имеющими к этому отношения АРС-мечеными антителами (анти-CD19). После 2 часов инкубации при 4°С с перемешиванием суспензия клетка-бусина была проанализирована способом FACS (используя отверстие размером 130 мкм). Фиг.3 демонстрирует, что специфическое связывание между анти-TNFα бусами и TNFα-трансфицированными клетками СНО было отчетливо выявлено способом FACS. Действительно, в этом образце (верхняя панель) приблизительно две трети бус были связаны с клетками (585 связанных на фоне 267 несвязанных). Напротив, в контрольных образцах (средняя и нижняя панели) практически ни одной бусины не было связано с клетками СНО. Также, обе популяции бус (анти-TNFα-РЕ и анти-CD19-АРС) были смешаны вместе с TNFα-трансфицированными клетками СНО. Фиг.4 демонстрирует, что приблизительно половина анти-TNFα бус связана с клетками СНО, в то время как подавляющее большинство анти-CD19 бус остались несвязанными. Процент бус, связывающихся с клеткой в каждом образце, подробно описан в Таблице 2. Таким образом, наглядно показано, что специфический отбор единичных В-клеток, которые связываются с интегральным мембранным белком-мишенью посредством их В-клеточного рецептора, был возможен с использованием проточной цитометрии.
Процент бус, связанных с клетками СНО, в каждом образце
Статистические данные по сортировке для верхней панели фиг.3; связывание бус, покрытых антителами анти-TNFα, с TNFα-трансфицированными клетками СНО
Статистические данные по сортировке для средней панели фиг.3; в отсутствие связывания бус, покрытых антителами анти-CD19, с TNFα-трансфицированными клетками СНО
Статистические данные по сортировке для нижней панели фиг.3; в отсутствие связывания бус, покрытых антителами анти-TNFα, с клетками СНО дикого типа
Статистические данные по сортировке для фиг.4
Пример 3
Выявление взаимодействия между В-клеткой, выделенной из кролика, иммунизированного антителом анти-TNFα, и клетками СНО, экспрессирующими мембраносвязанный TNFα, которые были насыщены антителом анти-TNFα
Для эксперимента, показанного на фиг.5, лимфоциты были выделены либо из селезенки кролика, иммунизированного антителом анти-TNFα (ESBA105, собственного производства) или из селезенки неиммунизированного кролика. Они были окрашены антикроличьими IgG-APC и антикроличьими IgM-FITC (AbD serotec) и впоследствии предварительно отобраны для получения чистых популяций В-клеток памяти (IgG+/IgM-). Параллельно, TNFα-экспрессирующие клетки СНО (даны в пользование Dr. P Scheurich, Univ. of Stuttgart) были погружены в раствор 1 мкг/мл кальцеинового красного (Invitrogen), цитоплазматического красителя, который флуоресцентно окрашивает живые клетки. Эти клетки были затем однократно промыты и инкубированы в присутствии (в отсутствие, для отрицательного контроля) 100 мкг/мл ESBA105, и в заключение снова промыты 3 раза PBS. В-клетки памяти были в конечном итоге смешаны в соотношении приблизительно 1:10 с клетками СНО и инкубированы в течение 2 часов при 4°С на вращающемся планшете (концентрация: 3*107 клеток/мл). Были приготовлены следующие образцы:
1) клетки CHO-TNFα + ESBA105 + В-клетки памяти кролика, иммунизированного ESBA105
2) клетки CHO-TNFα + ESBA105 + В-клетки памяти неиммунизированного кролика
3) клетки CHO-TNFα + В-клетки памяти кролика, иммунизированного ESBA105
После 2 часов инкубации 3 образца были подвергнуты измерениям способом FACS, используя отверстие в 70 мкм. В соответствии с иерархией популяций, показанной в Таблице 3а, 5% иммунизированных ESBA105 клеток связываются с «покрытыми ESBA105» трансгенными по TNFα клетками СНО. Для сравнения, только 0,5% неиммунизированных В-клеток связываются с этими «покрытыми ESBA105» трансгенными по TNFα клетками СНО (Таблица 3b), и 0,6% иммунизированных В-клеток связываются в отсутствие ESBA105 на поверхности клеток СНО (Таблица 3с). Эти результаты демонстрируют, что специфическое взаимодействие между BCR (В-клеточным рецептором) и трансмембранной мишенью может быть выявлено способом FACS.
Статистические данные по сортировке для верхней панели фиг.5b
Статистические данные по сортировке для средней панели фиг.5b
Статистические данные по сортировке для нижней панели фиг.5b
Пример 4
Выявление взаимодействия между В-клеткой, выделенной из кролика, иммунизированного ESBA105, и клетками СНО, экспрессирующими мембраносвязанный TNFα, которые были насыщены ESBA105
В дополнительном эксперименте не было проведено никакого предварительного отбора В-клеток памяти. Вся популяция лимфоцитов была инкубирована с окрашенными клетками СНО-TNFα-ESBA105. Трансгенные клетки СНО были приготовлены, как описано выше. Однако, чтобы предотвратить их пролиферацию в В-клеточной культуральной среде после сортировки в 96-луночных плашках, у клеток был остановлен клеточный цикл действием митомицина С (M4287-2MG) перед окрашиванием кальцеином. Лимфоциты кролика, иммунизированного ESBA105, были смешаны в соотношении 3:1 с окрашенными клетками СНО (концентрация клеточной суспензии: ≈3-107 клеток/мл) и инкубированы в течение 2 часов при 4°С на вращающемся планшете. После этого клеточная суспензия была проанализирована способом FACS, и В-клетки памяти, связавшиеся с CHO-TNFα-ESBA105, были сортированы в соответствии с выделенной областью, показанной на фиг.6, с 1, 10 или 100 клетками/лунка, как показано в Таблице 3. Отобранные клетки представляли собой 5,5% популяции В-клеток памяти, соответственно 0,2% от суммарного количества событий.
Отобранные клетки были собраны в 96-луночные плашки и культивированы в течение 13 дней при 37°С в условиях 5% CO2. После этого супернатанты культуры клеток были собраны и проанализированы прямым ELISA с целью проверки наличия ESBA105-связывающих IgG. Результаты ELISA (Таблица 3) показывают, что ESBA105-специфичные антитела могли быть обнаружены во многих лунках, и также в лунках, где единичные В-клетки были отобраны. Анализ Biacore (GE Healthcare) (Таблица 4) этих супернатантов подтвердил, что эти антитела действительно связываются с ESBA105-мишенью.
ELISA-анализ образцов супернатантов культуры В-клеток, взятых спустя 13 дней после сортировки. Ни одна В-клетка не была сортирована в лунках ряда А с целью проверки специфичности сигналов OD450. В лунках А11 и А12 поликлональное кроличье антитело анти-ESBA105 (AK3A; 2 мкг/мл) было внесено в супернатант в качестве положительного контроля. Лунки, где OD450 значительно превышает фон, выделены жирным шрифтом
Кинетические значения и концентрации, определенные с помощью Biacore, для супернатантов культуры В-клеток. Была проведена количественная оценка только тех супернатантов, где была отобрана одна клетка в лунке. BLQ: ниже предела количественного определения
Пример 5
Скрининг лимфоцитов кроликов, иммунизированных CXCR2 (типы 27/29)
Три кролика были иммунизированы вектором экспрессии CXCR2. После нескольких внутрикожных применений CXCR2-кДНК, была отобрана сыворотка и проверена на CXCR2-трансфицированных клетках на наличие специфических антител. Клетки лимфатических узлов были извлечены, заморожены в виде пяти аликвот с содержанием в каждой по 1,6×107 клеток и хранились в емкости с жидким азотом.
Аликвоту затем оттаивали и окрашивали антителами, специфичными для IgG (АРС-меченых) или IgM (меченых FITC). Параллельно, CXCR2-экспрессирующие клетки СНО были обработаны митомицином С для предотвращения дальнейшего роста без цитолиза клеток и погружены в раствор 1 мкг/мл кальцеинового красного. Оба образца клеток были затем смешаны с конечной клеточной концентрацией 107 клеток/мл, где лимфоциты были вдвойне многочисленнее, как и CXCR2-трансфицированные клетки СНО. После 2 часов инкубации с легким перемешиванием при 4°С клеточная суспензия была отфильтрована через 50-мкм фильтр и загружена в FACS. Отбор клеток был выполнен, как описано на фиг.6. Одно «событие» (В-клетки памяти, связанные с CXCR2-трансфицированной клеткой СНО) было отобрано в лунку в общей сложности 10×96-луночной плашки (всего 900 событий). Отобранные события составляли 3,1% от популяции В-клеток памяти, соответственно 0,035% от общего числа клеток в образце.
Отобранные лимфоциты были культивированы в течение 21 дня при 37°С в инкубаторе. Каждые 2-3 дня 100 мкл супернатанта клеточной культуры отбирали из лунок и заменяли свежей средой. В течение этого времени культивирования В-клетки пролиферировали, дифференцировались в плазматические клетки и секретировали антитела. Для визуализации того, какие супернатанты содержали CXCR2-специфические антитела, был проведен CELISA. Для этого, клетки СНО, экспрессирующие CXCR2, были засеяны с плотностью 50000 клеток/лунка на половине области 96-луночных плашек. После ночной инкубации при 37°С клетки были фиксированы 1% формальдегидом в PBS в течение 30 минут при комнатной температуре. Клеточные слои были затем промыты три раза, и места неспецифического связывания были блокированы клеточной культуральной средой в течение одного часа при комнатной температуре. Далее, после стадии трех промывок супернатанты были разведены 1:1 в культуральной среде и добавлены в лунки. В трех контрольных лунках коммерческие кроличьи антитела анти-CXCR2 были внесены в супернатант. Супернатанты инкубировали на клеточных слоях в течение полутора часов при комнатной температуре. В завершение, кроличьи IgG определяли козьим анти-кроличьим IGg Fc антителом, соединенным с HRP. При добавлении субстрата пероксидазы (субстрат Blue POD от Roche) развивалась колориметрическая реакция, которую останавливали спустя 25 минут 1М HCl. Оптическую плотность измеряли при 450 нм.
Результатом проведения CELISA было появление положительных сигналов в 1,8% лунок (16/900). Все положительные сигналы были подтверждены во втором CELISA. Супернатанты были также протестированы на других клеточных линиях: СНО-К1 (дикий тип) для выявления клонов возможного неспецифического связывания, СНО-человеческий CXCR1 и СНО-мышиный CXCR2 для демонстрации перекрестной активности по отношению к тесно связанному варианту рецептора или вида. В завершение, супернатанты были протестированы в прямом ELISA на предмет связывания с пептидом, состоящим из 48 N-концевых аминокислот CXCR2. Результаты отображены в Таблице 5. Все отобранные супернатанты порождали интенсивные сигналы OD450 против человеческого CXCR2 в CELISA. Некоторые из них были также слабо положительными в контрольном эксперименте с СНО клетками дикого типа, означая, что они могут связываться неспецифическим образом. Ни один из клонов не проявлял перекрестную реактивность с человеческим CXCR1 или мышиным CXCR2. Наконец, некоторые клоны, но не все из них, связывались с N-концевым пептидом CXCR2, указывая на возможный альтернативный сайт связывания на CXCR2. Принимая во внимание невозможность иммобилизации клеток целиком на чипе Biacore, в настоящее время невозможно количественно измерить сродство отобранных антител к рецептору CXCR2. Однако собранные данные указывают на то, что антитела, специфичные к человеческому CXCR2, были отобраны, используя систему сортировки межклеточного взаимодействия, описанную выше.
Сводные результаты CELISA по клонам анти-CXCR2, выделенным во время сортировки 27 и 29
ЭКВИВАЛЕНТЫ
Многочисленные модификации и альтернативные варианты осуществления настоящего изобретения очевидны специалистам в данной области техники в свете вышеизложенного описания. Соответственно, это описание следует составлять в качестве только иллюстративного материала и с целью обучения специалистов в данной области техники лучшему способу исполнения настоящего изобретения. Детали структуры могут существенно варьировать без отклонения от сущности изобретения, и исключительное использование всех модификаций, которые подпадают под объем прилагаемых пунктов формулы изобретения, сохраняется за автором изобретения. Предполагается, что настоящее изобретение ограничено только в той степени, требуемой прилагаемыми пунктами формулы изобретения и применимыми нормами закона.
Вся литература и подобный материал, цитированный в настоящей заявке, включая патенты, патентные заявки, статьи, книги, трактаты, диссертации, веб-страницы, фигуры и/или приложения, независимо от формата такой литературы и подобных материалов, безоговорочно включены в качестве ссылки полностью. В том случае, если один или более включенных литературных источников и подобных материалов отличается от этой заявки или ей противоречит, включая определяемые термины, использование терминов, описываемые методики или т.п., находится под контролем этой заявки.

















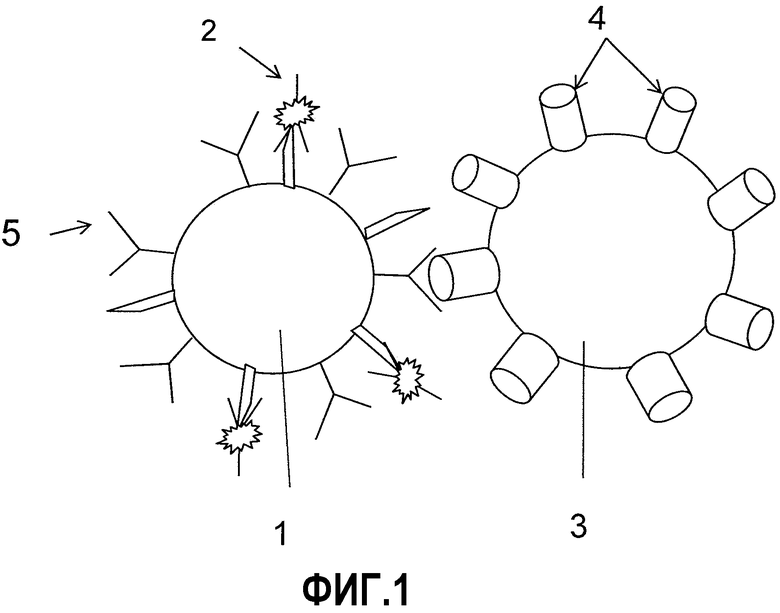
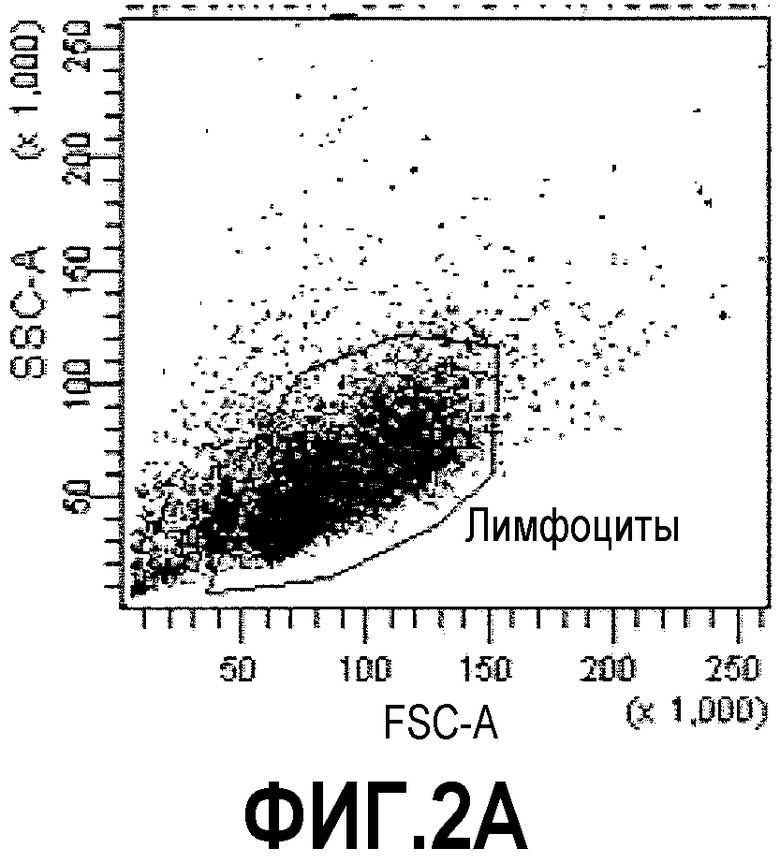
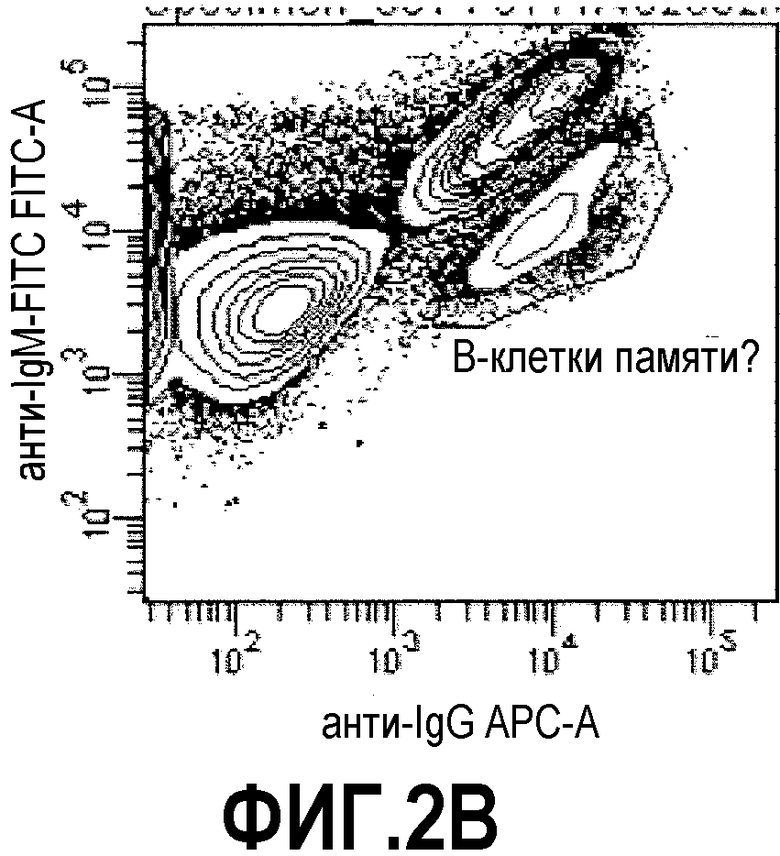
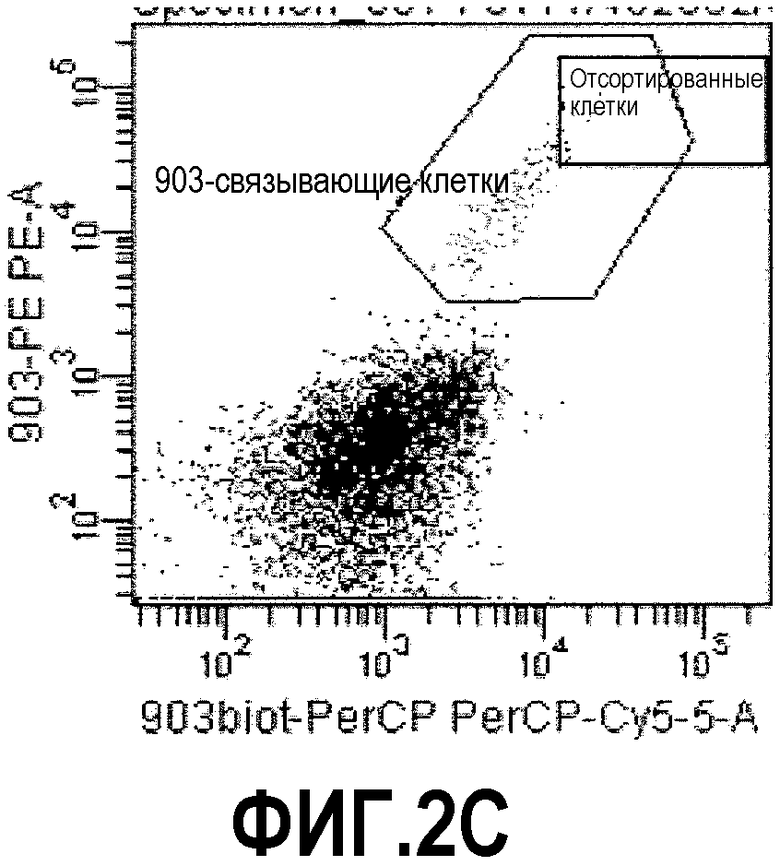
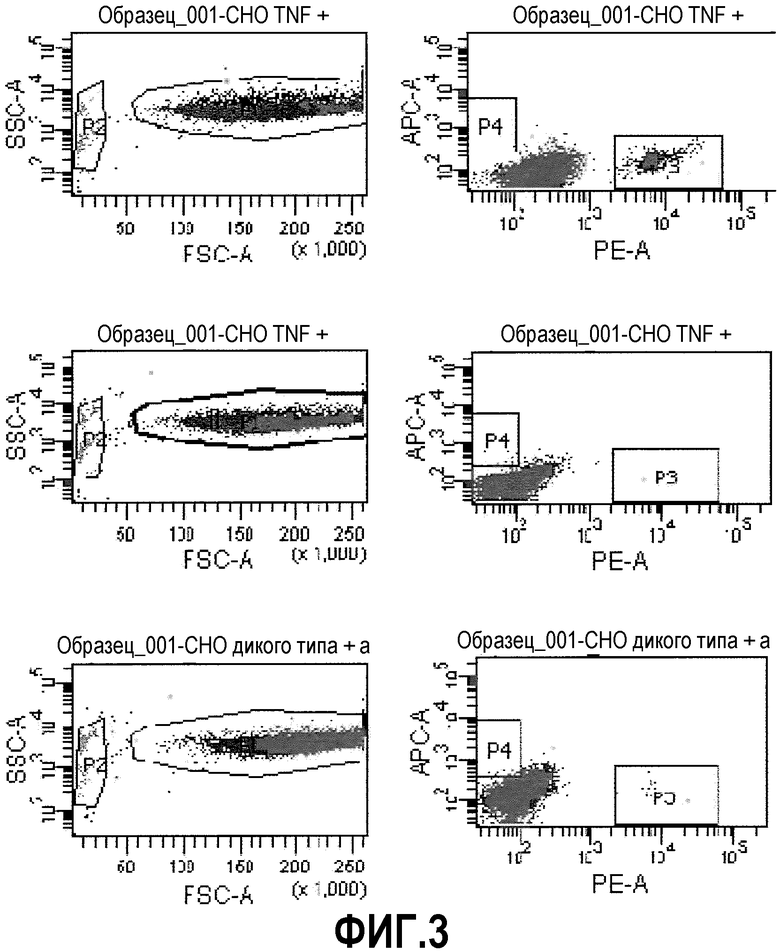
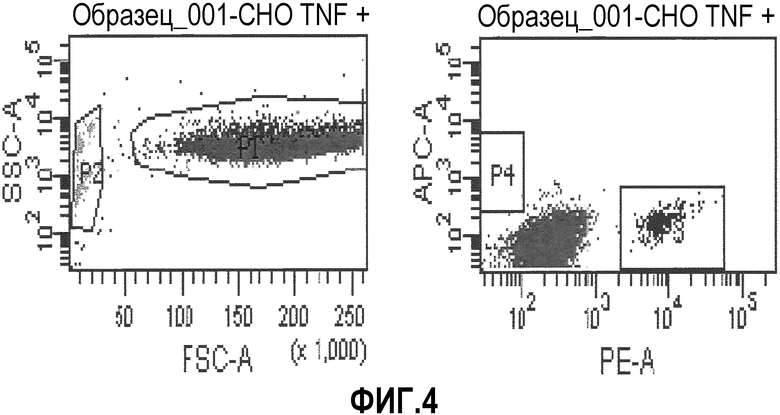
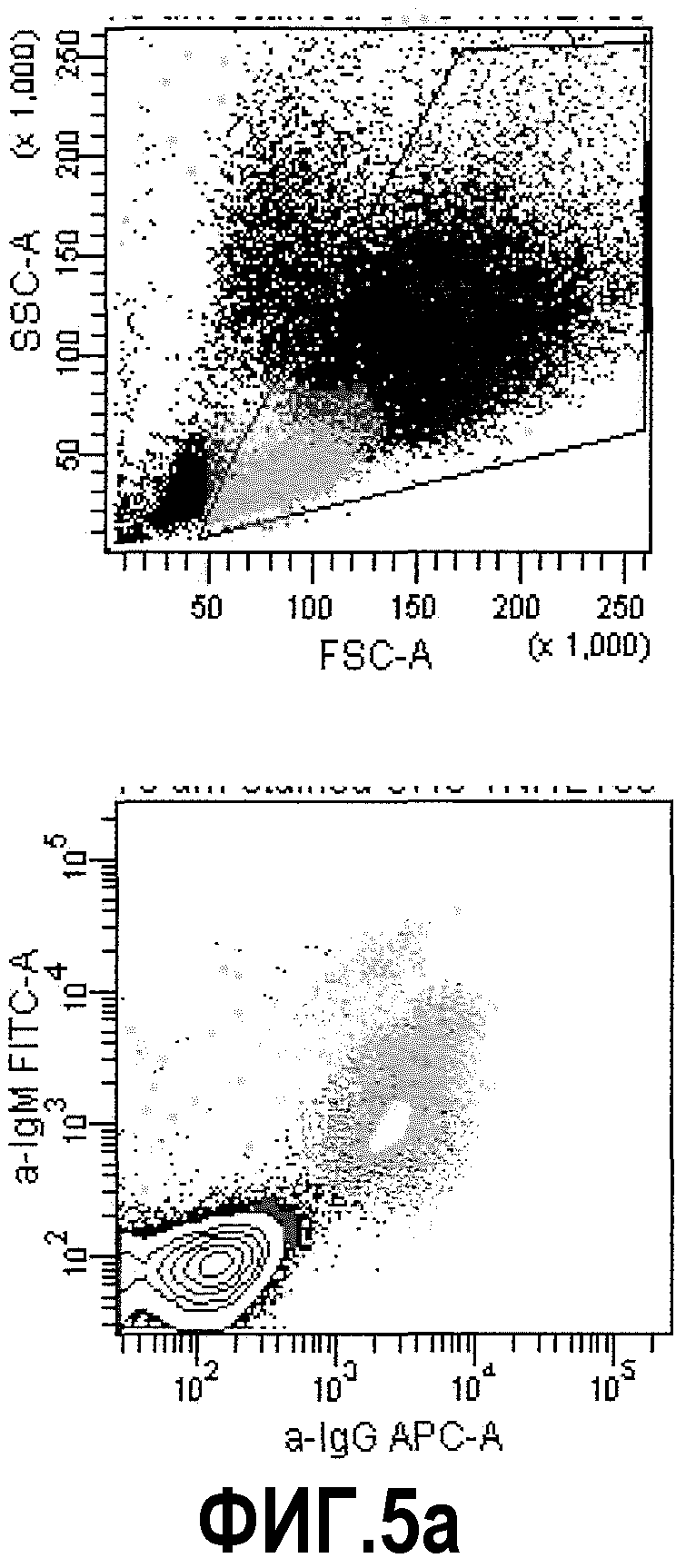
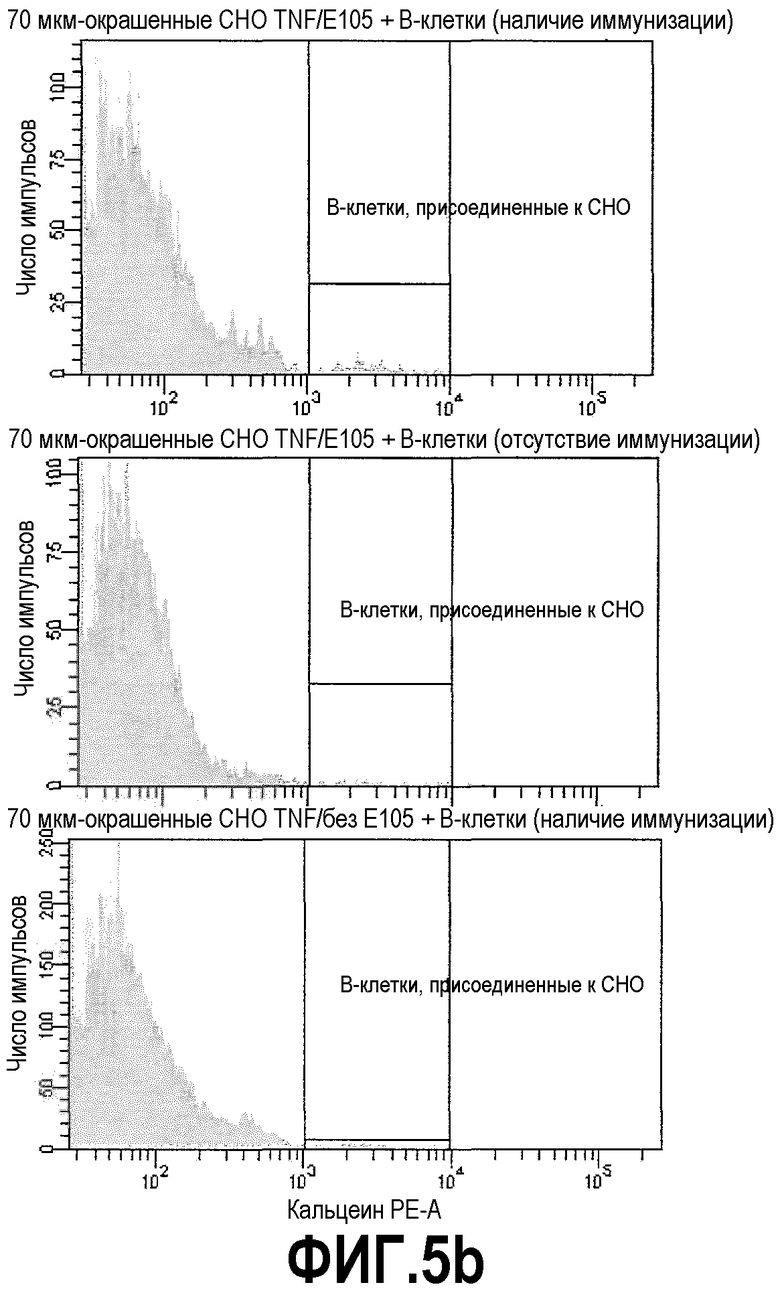
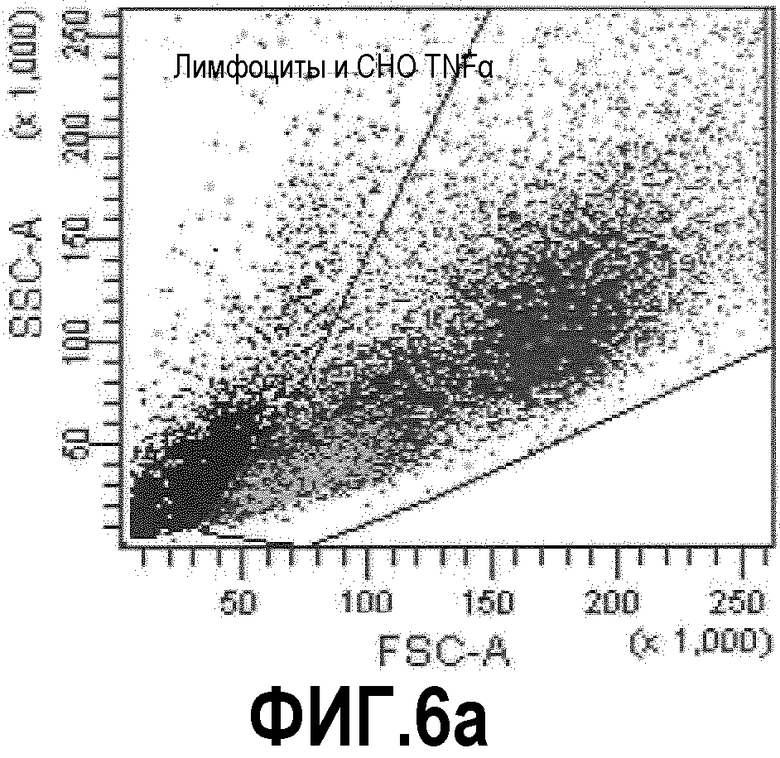
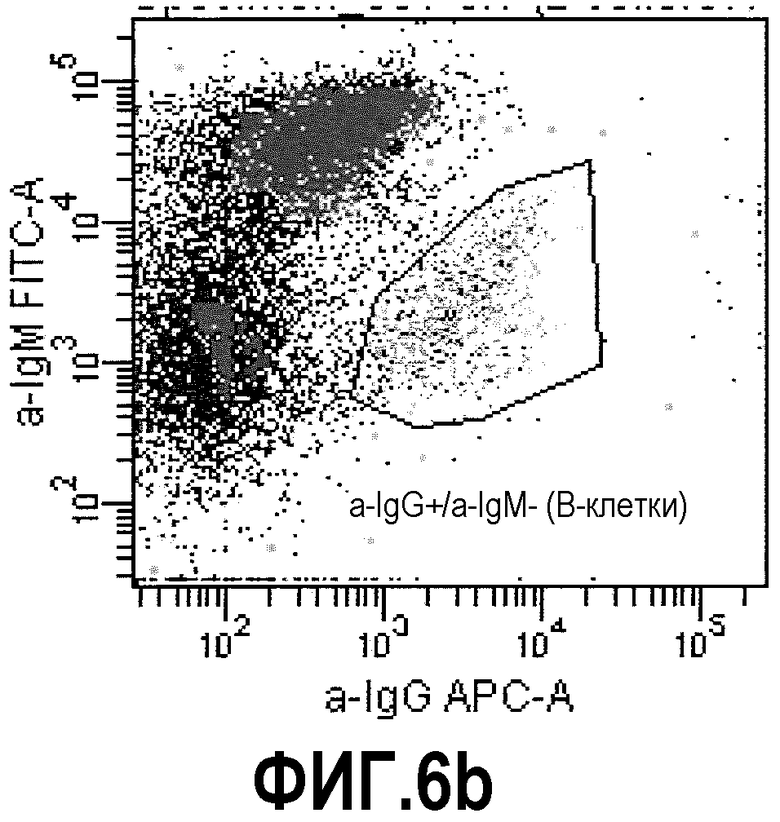
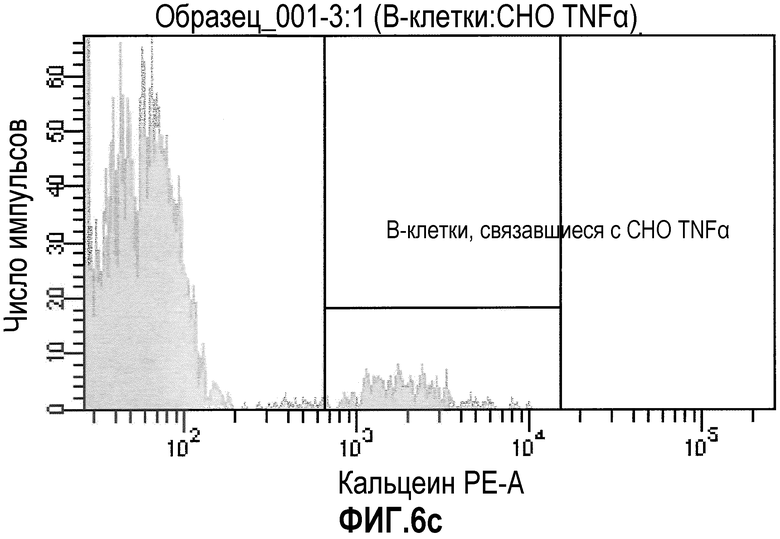
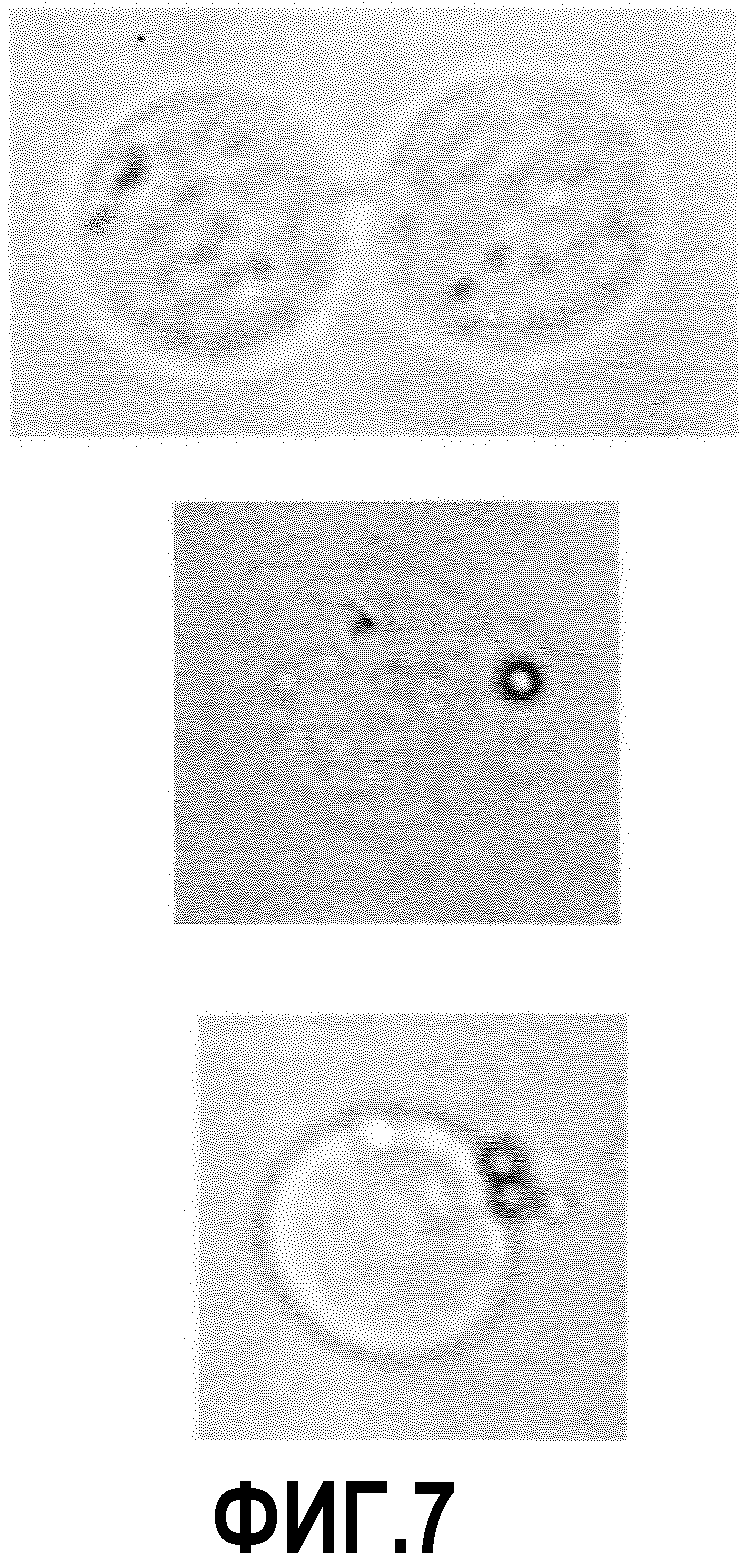
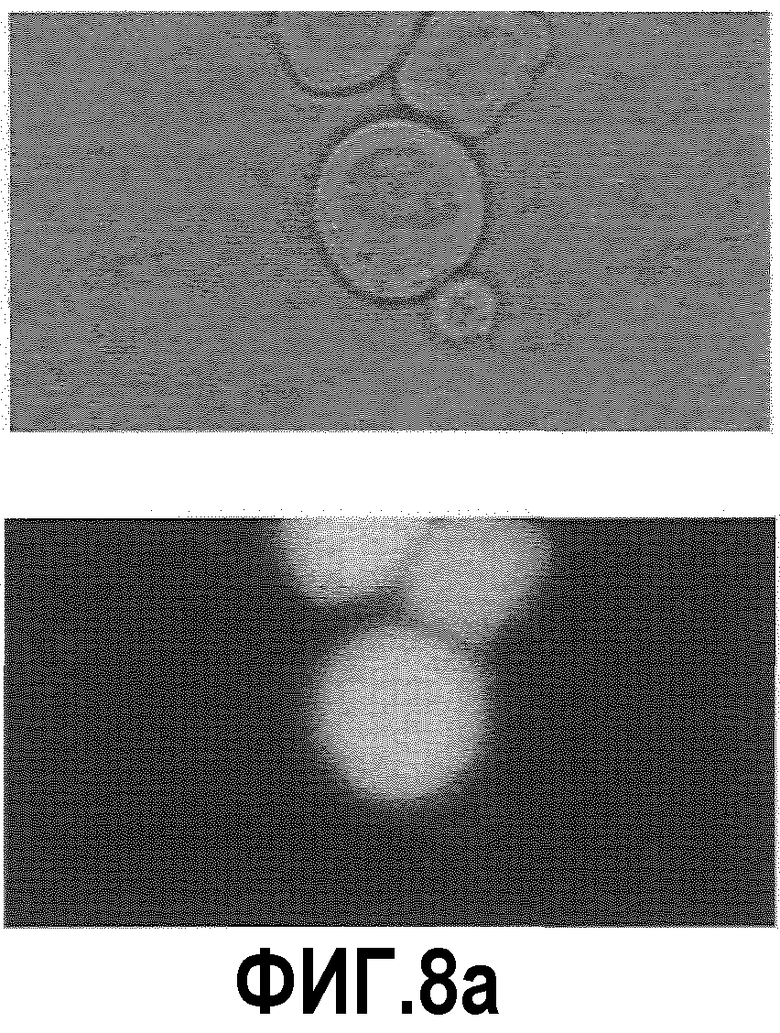
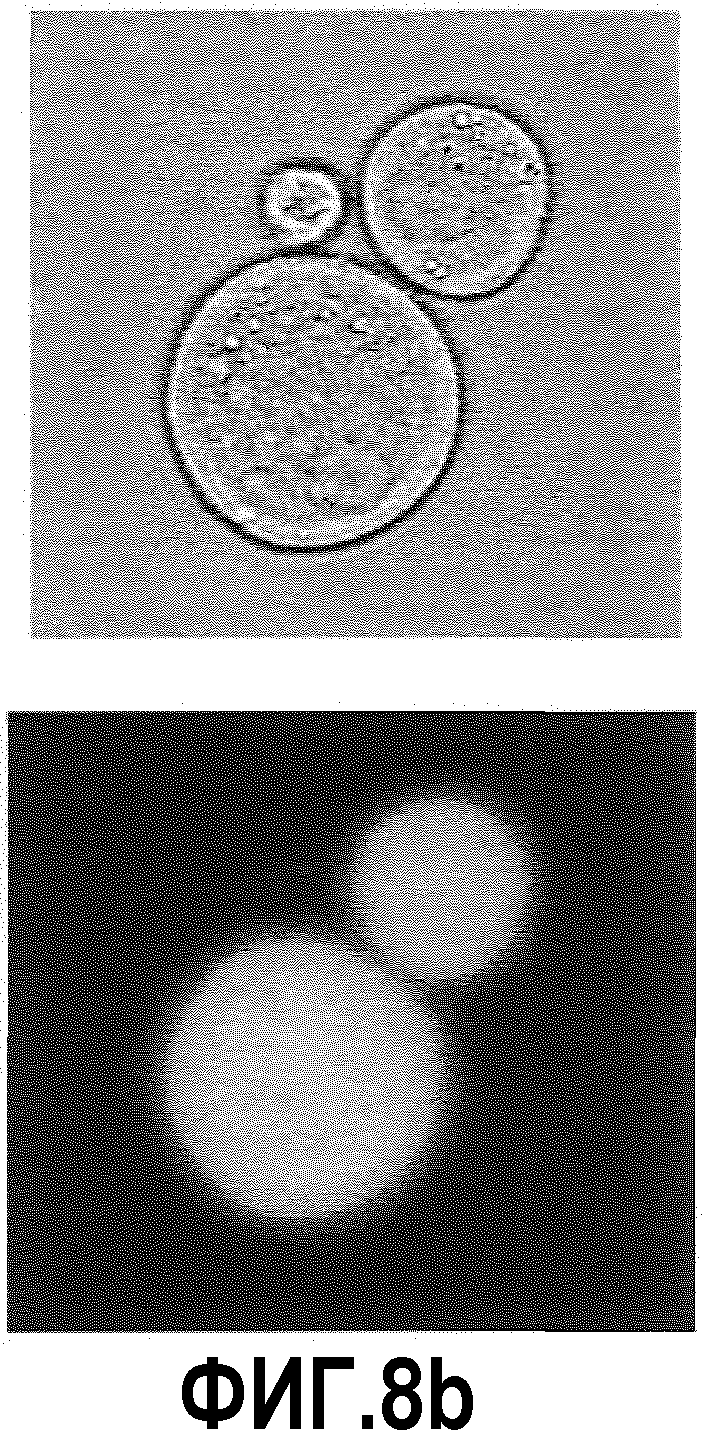
Настоящее изобретение относится к области биотехнологии. Предложен способ идентификации антитела, которое специфически связывается с интересующим антигеном на клеточной поверхности, предусматривающий иммунизацию животного вектором экспрессии, кодирующим антиген клеточной поверхности; приведение в контакт клеток, содержащих на своей поверхности антитела и выделенных из животного, подвергнутого иммунизации, причем антитела связаны с первой сортируемой меткой, с клетками, экспрессирующими антиген, связанный со второй сортируемой меткой; определение специфического связывания с использованием клеточного сортера по наличию первой и второй сортируемой метки в клеточном комплексе; а также идентификацию участков, определяющих комплементарность с антигеном (CDR), у идентифицированного антитела и прививку CDR на каркасную область акцепторного антитела. Данное изобретение предполагает скрининг антител, специфичных в отношении мембраносвязанных антигенов, что может найти дальнейшее применение в разработке терапевтических и диагностических средств. 3 з.п. ф-лы, 8 ил., 5 табл., 5 пр.
1. Способ идентификации антитела, которое специфически связывается с интересующим антигеном на клеточной поверхности, включающий:
(a) иммунизацию животного вектором экспрессии, кодирующим антиген клеточной поверхности;
(b) выделение и наличие множества клеток, экспрессирующих антитела, из животного стадии (а), где у клеток, экспрессирующих антитела, антитела находятся на поверхности клеток и функционально связаны с первой сортируемой меткой;
(c) наличие множества клеток, экспрессирующих антиген, функционально связанных со второй сортируемой меткой, где интересующий антиген находится на поверхности клетки, экспрессирующей антиген;
(d) приведение клеток, экспрессирующих антиген, в контакт с клетками, экспрессирующими антитела; и
(e) отделение от множества клеток, экспрессирующих антитела, одной или нескольких клеток, экспрессирующих антитела, которые могут специфически связываться с клетками, экспрессирующими антиген, используя клеточный сортер,
где наличие первой и второй сортируемой метки в одноклеточном комплексе, образованном между клеткой, экспрессирующей антитело, и клеткой, экспрессирующей антиген, указывает на связывание клетки, экспрессирующей антитело, с клеткой, экспрессирующей антиген, тем самым идентифицируя антитело, которое связывается с интересующим антигеном;
(f) идентификацию CDR указанного антитела и прививку CDR на каркасную область акцепторного антитела.
2. Способ по п.1, где каркасная область акцепторного антитела содержит
(i) каркасный участок вариабельной области тяжелой цепи, который по меньшей мере на 70% идентичен последовательности SEQ ID NO: 1; и/или
(ii) каркасный участок вариабельной области легкой цепи, который по меньшей мере на 70% идентичен последовательности SEQ ID NO: 2.
3. Способ по п.1 или 2, где каркасные участки акцепторного антитела содержат
(i) каркасный участок вариабельной области тяжелой цепи с последовательностью SEQ ID NO: 1, SEQ ID NO: 4 или SEQ ID NO: 6; и/или
(ii) каркасный участок вариабельной области легкой цепи с последовательностью SEQ ID NO: 2 или SEQ ID NO: 9.
4. Способ по п.2, дополнительно включающий стадию введения
(a) серина (S) в положение 12 тяжелой цепи (нумерация по АНо);
(b) серина (S) или треонина (Т) в положение 103 тяжелой цепи (нумерация по АНо); и/или
(c) серина (S) или треонина (Т) в положение 144 тяжелой цепи (нумерация по АНо).
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| BURKE J., MANN | |||
| C | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| ERDILE L | |||
| F | |||
| et al., "Whole cell ELISA for detection of tumor antigen expression in tumor samples." Journal of immunological | |||

