Данное изобретение относится к новым антителам, в частности к мышиным моноклональным антителам, химерным и гуманизированным, которые способные специфически связываться с хемокиновыми рецепторами (CXCR), а также к аминокислотным и нуклеиновокислотным последовательностям, кодирующим такие антитела. В одном аспекте изобретение относится к новым антителам, производным соединениям или функциональным фрагментам, способным специфически связываться с CXCR4 и проявляющим высокую противоопухолевую активность. Изобретение также включает применение таких антител в качестве лекарственного препарата для профилактики и/или терапевтического лечения рака, а также в процедурах или наборах, связанных с диагностикой рака. Наконец, изобретение включает композиции, содержащие такие антитела в сочетании или конъюгации с другими противоопухолевыми соединениями, такими как антитела, токсины, цитотоксические/цитостатические агенты, и их применение для профилактики и/или лечения некоторых раков.
Хемокины представляют собой малые секретируемые пептиды, которые контролируют миграцию лейкоцитов по химическому градиенту лиганда, известному как хемокиновый градиент, особенно в иммунных реакциях (ZIotnick A. et al., 2000). Они разделены на два основных подсемейства, СС и СХС, в зависимости от положения их NH2-концевых остатков цистеина, и связываются с рецепторами, сопряженными с G-белком, два основных подсемейства которых обозначаются как CCR и CXCR. До настоящего времени было обнаружено более 50 человеческих хемокинов и 18 хемокиновых рецепторов.
Многие раки имеют сложную хемокиновую сеть, которая влияет на иммунную-клеточную инфильтрацию опухоли, а также на рост, выживание, миграцию опухолевых клеток и ангиогенез. Иммунные клетки, эндотелиальные клетки и опухолевые клетки сами экспрессируют хемокиновые рецепторы и могут реагировать на хемокиновые градиенты. Исследования образцов биопсии человеческого рака и мышиных моделей рака показали, что экспрессия хемокиновых рецепторов раковыми клетками связана с увеличением метастатического потенциала. Злокачественные клетки от различных типов рака имеют различные профили экспрессии хемокиновых рецепторов, но наиболее распространен хемокиновый рецептор-4 (CXCR4). Клетки по меньшей мере 23 различных типов человеческих раков эпителиального, мезенхимального и гемопоэтического происхождения экспрессируют СХСР4-рецептор (Balkwill F. et al., 2004).
Хемокиновый рецептор-4 (также известный как фузин, CD184, LESTR или HUMSTR) существует в виде двух изоформ, содержащих 352 или 360 аминокислот.Остаток Asn11 гликозилирован, остаток Туг21 модифицирован путем добавлением сульфатной группы, a Cys109 и Cys186 связаны дисульфидным мостиком на внеклеточной части рецептора (Juarez J. et al., 2004).
Этот рецептор экспрессируется различными видами нормальных тканей, наивными Т-клетки (не Т-клетками памяти), регуляторными Т-клетками, В-клетками, нейтрофилами, эндотелиальными клетками, первичными моноцитами, дендритными клетками, натуральными киллерами, CD34+гемопоэтическими стволовыми клетками и на низком уровне в сердце, кишечнике, печени, почках и головном мозге. CXCR4 играет ключевую роль в перемещении лейкоцитов, В-клеточном лимфопоэзе и миелопоэзе.
СХСР4-рецептор сверхэкспрессирован в большом числе раков, включая, но не ограничиваясь ими, рак толстой кишки (Ottaiano A. et al., 2004), молочной железы (Kato М. et al., 2003), предстательной железы (Sun Y.X. et al., 2003), легких [мелкоклеточная и немелкоклеточная карцинома (Phillips R.J. et al., 2003)], яичника (Scotton C.J. et al., 2002), поджелудочной железы (Koshiba Т. et al., 2000), почек и головного мозга (Barbero S et al., 2002), глиобластому и лимфомы.
Уникальным лигандом рецептора CXCR4, описанным до настоящего времени, является фактор стромальных клеток-1 (SDF-1) или CXCL12. SDF-1 секретируется в большом количестве в лимфатических узлах, костном мозге, печени, легких и в меньшей степени в почках, головном мозге и коже. CXCR4 также распознается антагонистическим хемокином, вирусным макрофагальным воспалительным белком II (vMIP-II), который кодируется человеческим вирусом герпеса III типа.
CXCR4/SDF-1 ось играет ключевую роль в раке и непосредственно участвует в миграции, инвазии, ведущей к метастазированию. Действительно, раковые клетки экспрессируют СХСР4-рецептор, они мигрируют и входят в системный кровоток. Затем раковые клетки задерживаются в сосудистом русле в органах, которые продуцируют высокие уровни SDF-1, где они пролиферируют, индуцируют ангиогенез и формируют метастатические опухоли (Murphy PM., 2001). Эта ось также участвует в пролиферации клеток через активацию пути внеклеточной сигнал-регулируемой киназы (ERK) (Barbero S. et al., 2003) и ангиогенезе (Romagnani P., 2004). Действительно, СХСР4-рецептор и его лиганд SDF-1 четко усиливает ангиогенез путем стимуляции экспрессии VEGF-A, который в свою очередь увеличивает экспрессию CXCR4/SDF-1 (Bachelder R.E. et al., 2002). Известно также, что опухоль-ассоциированные макрофаги (ТАМ) накапливаются в гипоксических областях опухолей и стимулируются к воздействию с опухолевыми клетками и усиливают ангиогенез. Было отмечено, что гипоксия селективно повышает экспрессию CXCR4 в различных типах клеток, включая ТАМ (Mantovani А. et al., 2004). Недавно было показано, что ось CXCR4/SDF-1 регулирует перемещение/хоминг CXCR4+гемопоэтических стволовых клеток/клеток-предшественников (HSC) и может играть роль в неоваскуляризации. Данные свидетельствуют о том, что помимо HSC функциональный CXCR4 также экспрессируется на стволовых клетках из других тканей (ранних тканевых стволовых клеток=TCSC), так что SDF-1 может играть ключевую роль в хемотаксисе CXCR4+TCSC, необходимом для регенерации органа/ткани, но эти TCSC могут также быть клеточным источником развития рака (теория раковых стволовых клеток). Происхождение рака из стволовых клеток было продемонстрировано для человеческой лейкемии и недавно для нескольких солидных опухолей, таких как опухоли головного мозга и молочной железы. Есть несколько примеров CXCR4+опухолей, которые могут происходить из нормальных CXCR4+ткане-Азрганоспецифичных стволовых клеток, например лейкемии, опухоли головного мозга, мелкоклеточный рак легких, рак молочной железы, гепатобластома, рак яичников и шейки матки (Kucia M. et al., 2005).
Влияние на метастазы рака путем воздействия на рецептор CXCR4 было продемонстрировано in vivo с помощью моноклональных антител, направленных против рецептора CXCR4 (Muller A. et al., 2001). Вкратце, было показано, что моноклональное антитело, направленное против СХСР4-рецептора (МКА 173 R&D Systems), значительно уменьшает количество метастазов в лимфатических узлах на модели ортотопического рака молочной железы (MDA-MB231) у мышей SCID. Другое исследование (Phillips R.J et al., 2003) также показало важную роль оси SDF-1/CXCR4 в метастазировании на модели ортотопической карциномы легких (А549) с помощью поликлональных антител против SDF-1, но в данном исследовании не было влияния ни на рост опухоли, ни на ангиогенез. Ряд других исследований также описывает ингибирование либо метастазов in vivo с помощью киРНК-дуплексов CXCR4 (Liang Z. et al., 2005), биоустойчивых антагонистов пептида CXCR4 (Tamamura H. et al., 2003), либо опухолевого роста in vivo с использованием низкомолекулярного антагониста CXCR4, такого как AMD 3100 (Rubin J.B. et al., 2003; De Faico V. et al., 2007), или МКА (патент W02004/059285 А2). Таким образом, CXCR4 является утвержденной терапевтической мишенью при раках.
Хемокиновый рецептор-2 (CXCR2), другой хемокиновый рецептор, также описывается как интересная мишень в онкологии. Действительно, CXCR2 передает аутокринный клеточный ростовой сигнал в нескольких типах опухолевых клеток и также может влиять на рост опухоли косвенным путем за счет усиления ангиогенеза (Tanaka Т. et al. 2005).
Хемокиновый рецептор CXCR2 включает 360 аминокислот. Он экспрессируется главным образом в эндотелиальных клетках и специфично во время неоваскуляризации. СХСР2-рецептор связывают несколько хемокинов: CXCL5, -6, -7, IL-8, GRO-α, -β и -γ, которые принадлежат к ERL+проангиогенным хемокинам. СХСР2-рецептор частично имеет гомологичную последовательность с СХСР4-рецептором: 37% идентичной последовательности и 48% гомологичной последовательности. Ось СХСР2/лиганды вовлечена в несколько механизмов опухолевого роста, таких как метастазирование (Singh RK. et аl., 1994), клеточная пролиферация (Owen J.D. et al., 1997) и ERL+хемокин-опосредованный ангиогенез (Stricter R.M. et al., 2004; Romagnani et al., 2004). Наконец, опухоль-ассоциированные макрофаги и нейтрофилы являются ключевыми элементами индуцированного воспалением опухолевого роста, и хемокины, такие как CXCL5, IL-8 и GRO-a, инициируют привлечение нейтрофилов.
Димеризация появилась как общий механизм регуляции функции рецепторов, связанных с G-белком, к числу которых относятся хемокиновые рецепторы (Wang J. and Norcross M., 2008). Было показано, что гомо- и гетеродимеризация в ответ на связывание хемокином необходима для инициирования и изменения передачи сигнала от большого количества хемокиновых рецепторов. Все больше фактов поддерживает концепцию, что рецепторные димеры или олигомеры, вероятно, являются основной функциональной единицей хемокиновых рецепторов, Димеры хемокиновых рецепторов присутствуют в отсутствие лигандов, а хемокины индуцируют конформационные изменения рецепторных димеров. CXCR4, как известно, формирует гомодимеры, а также гетеродимеры, например, с 6-опиоидным рецептором (DOR) (Hereld D., 2008) или CCR2 (Percherancier Y. et al., 2005). В последнем примере пептиды, полученные из трансмембранных доменов CXCR4, ингибировали активацию путем блокирования лиганд-индуцированных конформационных переходов димера (Percherancier Y. et al., 2005). Другое исследование показало, что пептид CXCR4-TM4, синтетический пептид трансмембранного участка CXCR4, уменьшает перенос энергии между протомерами гомодимеров CXCR4 и ингибирует SDF-1-индуцированную миграцию и полимеризацию актина в злокачественных клетках (Wang J. et al., 2006). Совсем недавно также было описано, что CXCR7 формирует функциональные гетеродимеры с CXCR4 и усиливает SDF-1-индуцированную передачу сигнала (Sierro F. et al., 2007). Другие примеры конститутивных гетеродимеров включают исследования, показывающие взаимодействие CXCR1 и CXCR2, а также формирование соответствующих гомодимеров. Для любого из них было отмечено отсутствие взаимодействий с другим GPCR (альфа(1А)-адренорецептором), что указывает на специфичность взаимодействия CXCR1 и CXCR2 (Wilson S. et al., 2005).
Как упоминалось ранее, рецепторы CXCR4 и CXCR2 являются интересными опухолевыми мишенями. Влияние на эти рецепторы должно ингибировать опухолевый рост и метастазирование очень эффективным образом путем уменьшения пролиферации опухолевых клеток, ангиогенеза, миграции и инвазии опухолевых клеток, привлечения нейтрофилов и макрофагов в опухоль и путем ингибирования СХСР4-раковых стволовых клеток.
Одним из предлагаемых аспектов данного изобретения является создание мышиного моноклонального антитела, индуцирующего конформационные изменения СХСР4-димеров. Изобретение охватывает CXCR4-MKA 414Н5 (или его фрагменты), способное связывать и индуцировать конформационные изменения как гомодимеров CXCR4, так и гетеродимеров CXCR4/CXCR2, и имеющее высокую противоопухолевую активность как на мышиной модели с ксенотрансплантатом, так и на моделях выживаемости. Изобретение также охватывает CXCR4-MKA 515Н7 (или его фрагменты), способное связывать и индуцировать конформационные изменения как гомодимеров CXCR4, так и гетеродимеров CXCR4/CXCR2, и имеющее высокую противоопухолевую активность. Анти-СХСР4-МКА 414Н5 ингибирует опухолевый рост на модели с ксенотрансплантатом MDA-МВ-231 и увеличивает выживаемость мышей на модели U937. Они индуцируют конформационные изменения в гомодимерах CXCR4, а также в гетеродимерах CXCR4/CXCR2. Это новое свойство должно представлять интерес для применения терапии рака с учетом важной роли этих двух хемокиновых рецепторов при раке.
Уже обнаружено, что влияние как гомо-, так и гетеродимеров рецепторов усиливает терапевтический эффект МКА. Действительно, было продемонстрировано, например, что МКА (h7C10), влияющее на оба гибридных рецептора IGF-1R и инсулин/IGF-1, более мощно ингибирует опухолевый рост in vivo, чем МКА, влияющее исключительно на IGF-1R (Pandini G., 2007).
Кроме того, анти-СХСР4-МКА 414Н5 и 515Н7 являются молчащими антагонистами для CXCR4, они не изменяют базальный сигнал в анализах in vitro, но ингибируют SDF-1-индуцированную передачу сигнала в различных анализах (GTPYS-связывание, высвобождение цАМФ), а также способны ингибировать SDF-1-индуцированную пролиферацию и миграцию опухолевых клеток in vitro.
Молекулы, действующие либо как частичные агонисты, либо как обратные агонисты, демонстрируют внутреннюю активность в отсутствие лигандов. Эти типы молекул стабилизируют, соответственно, высокоаффинное или низкоаффинное состояние GPCR, даже в отсутствие лиганда, тем самым активируя или ингибируя нисходящие сигнальные каскады (Galandin et al., 2007; Kenakin, 2004).
В случае МКА 414Н5 и 515Н7 эти молекулы ведут себя как молчащие антагонисты без какой-либо внутренней активности в отношении СХСР4-рецептора в отсутствие SDF-1. Эта фармакологическая черта может быть связана с менее неблагоприятными побочными эффектами по сравнению с частичными или обратными агонистами, как уже отмечалось для лигандов опиоидных рецепторов (Bosierand Hermans, 2007). Действительно, функциональная активность обоих МКА 414Н5 и 515Н7 полностью зависит от наличия SDF-1, и отсутствие модуляции активности рецептора CXCR4 будет наблюдаться в тканях и органах, в которых лиганд SDF-1 не экспрессируется, не секретируется или не доставляется кровотоком. Таким образом, МКА 414Н5 и 515Н7, вероятно, будут менее токсичны по сравнению с другими лигандами рецептора CXCR4 с положительной или отрицательной эффективностью. Кроме того, молчащие антагонисты составляют меньшую часть видов в фармакологической области (Wurch et al., 1999, Kenakin, 2004).
Неожиданно впервые изобретателям удалось создать антитела, которые способны связываться с CXCR4, а также способны вызывать конформационные изменения гомодимеров и/или гетеродимеров CXCR4. В частности, антитела изобретения способны вызывать конформационные изменения гомодимеров CXCR4, а также гетеродимеров CXCR4/CXCR2.
В последующей заявке выражение во множественном числе «димеры CXCR4» следует понимать как охватывающее гомодимеры CXCR4, а также гетеродимеры CXCR4/CXCR2.
На данном этапе следует отметить, что такие антитела никогда не были описаны в известном уровне техники. Кроме того, необходимо отметить, что существование гетеродимеров CXCR4/CXCR2 никогда не было описано.
Частью изобретения является открытие существования гетеродимера, образованного CXCR4 и CXCR2.
Так, в частном аспекте данное изобретение относится к изолированному комплексу, включающему или состоящему из гетеродимера CXCR4/CXCR2.
Предпочтительно соединение СХСР4-части указанного гетеродимерного комплекса CXCR4/CXCR2 является одной из двух человеческих изоформ CXCR4, выбранных из группы, состоящей из:
- хемокинового (С-Х-С-мотив) рецептора 4 изоформы b [Homo Sapiens], имеющего последовательность, указанную под регистрационным номером Genbank NP_003458 SEQ ID №29:

- хемокинового (С-Х-С-мотив) рецептора 4 изоформы a [Homo Sapiens], имеющего последовательность, указанную под регистрационным номером Genbank NP_001008540 SEQ ID №30:

- альтернативного транскрипционного варианта сплайсинга или природного варианта, имеющего по меньшей мере 95% идентичность с одной из этих изоформ b или а с SEQ ID №29 или 30; и
- его фрагмента, который может специфично распознаваться природным лигандом фактором стромальных клеток-1 (SDF-1) и предпочтительно имеет по меньшей мере 100, 150 и 200 аминокислот в длину.
Предпочтительно соединение СХСР2-части указанного гетеродимерного комплекса CXCR4/CXCR2 выбрано из группы, состоящей из:
- рецептора бета интерлейкина-8 [Homo Sapiens], имеющего последовательность, указанную под регистрационным номером Genbank NP_001548 SEQ ID №31:

- альтернативного транскрипционного варианта сплайсинга или природного варианта, имеющего по меньшей мере 95% идентичность с этим рецептором b интерлейкина-8 с SEQ ID №31; и
- его фрагмента, который может специфично распознаваться ИЛ-8 и предпочтительно имеет по меньшей мере 100, 150 и 200 аминокислот в длину.
В данном частном аспекте данное изобретение также включает изолированную РНК или ДНК, кодирующую полипептид, включающий указанный гетеродимерный комплекс CXCR4/CXCR2.
Это изобретение также включает нуклеиновую конструкцию, предпочтительно экспрессионный вектор, например плазмиду, кодирующую указанный гетеродимерный комплекс CXCR4/CXCR2.
Изобретение также включает композицию, включающую по меньшей мере одну нуклеиновую конструкцию, предпочтительно экспрессионный вектор, например плазмиду, кодирующую СХСР4-часть указанного гетеродимерного комплекса CXCR4/CXCR2, и вторую конструкцию, предпочтительно экспрессионный вектор, например плазмиду, кодирующую СХСР2-часть указанного гетеродимерного комплекса CXCR4/CXCR2.
В этом аспекте изобретение также включает способ получения рекомбинантной принимающей клетки (клетки-хозяина), экспрессирующей указанный гетеродимерный комплекс CXCR4/CXCR2, где этот способ включает этап трансформации указанной принимающей клетки:
а) нуклеиновой конструкцией, предпочтительно экспрессионным вектором, например плазмидой, кодирующей указанный гетеродимерный комплекс CXCR4/CXCR2; или
б) по меньшей мере одной нуклеиновой конструкцией, предпочтительно экспрессионным вектором, например плазмидой, кодирующей СХСР4-часть указанного гетеродимерного комплекса CXCR4/CXCR2, и второй конструкцией, предпочтительно экспрессионным вектором, например плазмидой, кодирующей СХСК2-часть указанного гетероди мерного комплекса CXCR4/CXCR2.
В предпочтительном воплощении указанная принимающая клетка является эукариотической клеткой, например клеткой млекопитающего.
В предпочтительном воплощении нуклеиновая конструкция (конструкции), кодирующая указанный гетеродимерный комплекс CXCR4/CXCR2, кодирует также первый маркер, который связан (в частности, с помощью ковалентной связи) с СХСР4-последовательностью, например маркер LUC (люцефераза), и второй маркер, который связан (в частности, с помощью ковалентной связи) с CXCR2-последовательностью, например маркер GFP (т.е. для анализа BRET).
Изобретение также включает способ выбора соединения, которое проявляет противораковую активность или которое может быть использовано для получения композиции для лечения рака, который характеризуется тем, что включает этап:
а) контактирования рекомбинантной принимающей клетки данного изобретения, которая экспрессирует указанный гетеродимерный комплекс CXCR4/CXCR2, с тестируемым соединением; и
б) определения того, способно ли это соединение модулировать, предпочтительно ингибировать, активность этого гетеродимерного комплекса CXCR4/CXCR2 в рекомбинантной принимающей клетке.
В первом аспекте предметом данного изобретения является способ создания и выбора антител в соответствии с изобретением.
Более конкретно, изобретение относится к способу выбора анти-СХСР4-антитела или одного из его функциональных фрагментов или производных, способных ингибировать как лиганд-зависимую, так и лиганд-независимую активацию CXCR4, при этом указанный способ включает следующие этапы:
i) скрининг созданных антител и выбор антител, способных специфически связывать CXCR4, а также для модулировать СХСР4-активацию;
ii) тестирование антител, выбранных на этапе i), и выбор антител, способных индуцировать конформационные изменения гомодимеров CXCR4, а затем
iii) тестирование антител, выбранных на этапе ii), и выбор антител, способных индуцировать конформационные изменения гетеродимеров CXCR4/CXCR2.
Под выражением «модулировать» следует понимать увеличение или ингибирование. Предпочтительно выбранные антитела изобретения должны ингибировать СХСР4-активацию.
Как было объяснено ранее, индукция конформационных изменений димеров CXCR4 является основным аспектом изобретения, так как такие антитела будут представлять реальный интерес для большего количества пациентов.
Создание антитела может быть реализовано любым способом, известным специалистам в данной области, таким как, например, слияние миеломной клетки с клетками селезенки от иммунизированных мышей или других видов, совместимых с выбранными миеломными клетками [Kohler & Milstein, 1975, Nature, 256:495-497]. Иммунизированные животные могут включать трансгенных мышей с человеческими иммуноглобулиновыми локусами, которые затем непосредственно продуцируют человеческие антитела. Другое возможное воплощение могло бы состоять в применении методики фагового дисплея для скрининга библиотек.
Этап скрининга i) может быть реализован любым способом или процессом, известным специалистам в данной области. В качестве неограничивающих примеров можно упомянуть ИФА, BIAcore, иммуногистохимию, FACS-анализ и функциональные скрининги. Предпочтительный способ состоит в скрининге с помощью FACS-анализа на СХСР4-трансфектантах и по меньшей мере на опухолевой клеточной линии, чтобы убедиться, что продуцируемые антитела также способны распознавать нативный рецептор на опухолевых клетках. Этот способ будет описан более точно в следующих примерах.
Под выражением «модулировать СХСР4-активацию» понимается модуляция по меньшей мере одной активности, указанной в примерах 4, 5, 7 и 13 ниже:
Предпочтительными для модуляции являются:
- специфическое связывание на клеточных мембранах лигандом SDF-1 рецептора CXCR4 (см. пример 4), в частности, путем конкурирования на мембране эукариотических трансформированных клеток, таких как мембраны СНО-К1, стабильно эксперессирующих человеческий рецептор CXCR4 дикого типа;
- специфическое связывание на клеточных мембран GTPyS рецептора CXCR4 (см. пример 5), в частности, на мембране эукариотических трансформированных клеток, например клеток NIH-3T3, стабильно и конститутивно эксперессирующих рецептор CXCR4 дикого типа;
- СХСР4-опосредованное ингибирование продукции цАМФ (см. пример 7); и
- СХСР4-рецептор-опосредованная мобилизация внутриклеточных депо кальция (см. пример 13).
Более предпочтительно эта модуляция по меньшей мере одной из этих видов активности представляет собой ингибирование активности.
В предпочтительном воплощении этапов выбора ii) и ill) способа изобретения указанные этапы ii) и ill) состоят в оценке антител с помощью BRET-анализа на клетках, экспрессирующих CXCR4-RLuc/CXCR4-YFP и CXCR4-Rluc/CXCR2-YFP, соответственно, и в выборе антител, способных ингибировать по меньшей мере 40%, предпочтительно 45%, 50%, 55% и наиболее предпочтительно 60% BRET-сигнала.
Методика BRET представляет собой методику, которая известна как репрезентативная для белковой димеризации [Angers et al., PNAS, 2000, 97:3684-89].
Методика BRET, используемая в этапах способа ii) и iii), хорошо известна специалистам в данной области и будет подробно описана в последующих примерах. Более конкретно BRET (bioluminescence resonance energy transfer, резонансный перенос энергии биолюминесценции) представляет собой нерадиационный перенос энергии, возникающий между биолюминесцентным донором (люциферазой Renilla (Rluc)) и флуоресцентным акцептором, мутантом GFP (green fluorescent protein, зеленый флуоресцентный белок) или YFP (yellow fluorescent protein, желтый флуоресцентный белок). В данном случае был использован EYFP (enhanced yellow fluorescent protein, усиленный желтый флуоресцентный белок). Эффективность переноса зависит от ориентации и расстояния между донором и акцептором. Кроме того, передача энергии может происходить только тогда, когда две молекулы находятся в непосредственной близости (1-10 нм). Это свойство используется для проведения анализа белково-белковых взаимодействий. Действительно, в целях изучения взаимодействия между двумя участниками первый из них генетически сливают с люциферазой Renilla, а второй с желтым мутантом GFP. Гибридные белки, как правило, но не обязательно, экспрессируются клетками млекопитающих. В присутствии своего мембранно-проницаемого субстрата (коэлентеразина) Rluc испускает синий свет.Если мутант GFP находится ближе 10 нм к Rluc, то может возникнуть передача энергии, и будет обнаружен дополнительный желтый сигнал. Сигнал BRET измеряется как соотношение между светом, излучаемым акцептором, и светом, излучаемым донором. Так, BRET-сигнал будет увеличиваться по мере сближения двух гибридных белков, или если конформационное изменение сблизит Rluc и GFP-мутант.
Если BRET-анализ включен в предпочтительное воплощение, то для измерения конформационных изменений димеров CXCR4 можно использовать любой способ, известный специалистам в данной области. Не ограничивая, можно отметить следующие методики: FRET (fluorescence resonance energy transfer, резонансный перенос энергии флуоресценции), HTRF (homogenous time resolved fluorescence, гомогенная флуоресценция с временным разрешением), FLIM (fluorescence lifetime imaging microscopy, флуоресцентная микроскопия с изображением по времени жизни флуоресценции) или SW-FCCS (single wavelength fluorescence cross-correlation spectroscopy, кросс-корреляционная спектроскопия флуоресценции одной длины волны).
Также могут быть использованы другие классические методики, например коиммунопреципитация, Alpha Screen, химическое сшивание, двойной гибрид, аффинная хроматография, ИФА и фар-вестерн-блоттинг.
В частном аспекте способа в соответствии с изобретением этап ii) состоит в оценке антител с помощью BRET-анализа на клетках, экспрессирующих оба CXCR4-RLuc/CXCR4-YFP, и в выборе антител, способных ингибировать по меньшей мере 40% BRET-сигнала.
В другом частном аспекте способа в соответствии изобретением этап iii) состоит в оценке антител с помощью BRET-анализа на клетках, экспрессирующих оба CXCR4-RLuc/CXCR4-YFP, и в выборе антител, способных ингибировать по меньшей мере 40% BRET-сигнала.
Во втором аспекте предметом изобретения является изолированное антитело или один из его функциональных фрагментов или производных, получаемых указанным способом. Указанное антитело или один из его указанных фрагментов или производных способны специфически связываться с человеческим CXCR4 и, более того, при необходимости предпочтительно способны ингибировать природное присоединение его лиганда; также указанное антитело способно индуцировать конформационные изменения димеров CXCR4.
Выражения «функциональные фрагменты и производные» подробно будут определены ниже в данном описании.
Здесь следует понимать, что изобретение не относится к антителам в природной форме, то есть они находятся не в их природной среде, но что они могут быть выделены или получены путем очистки из природных источников, либо получены путем генетической рекомбинации, либо путем химического синтеза, и что они могут содержать неприродные аминокислоты, как будет описано ниже.
В частности, в соответствии с другим аспектом изобретения заявлено антитело или один из его функциональных фрагментов или производных, при этом указанное антитело характеризуется тем, что оно содержит по меньшей мере один участок, определяющий комплементарность (CDR), выбранный среди CDR, содержащих аминокислотную последовательность SEQ ID №№1-12.
В частности, в соответствии с другим аспектом изобретения заявлено антитело или один из его функциональных фрагментов или производных, при этом указанное антитело характеризуется тем, что оно содержит по меньшей мере один участок, определяющий комплементарность (CDR), выбранный среди CDR, содержащих аминокислотную последовательность SEQ ID №№2, 5 или 40-49.
В соответствии с первым аспектом изобретение относится к изолированному антителу или производному соединению или его функциональному фрагменту, содержащему по меньшей мере один CDR, выбранный среди CDR последовательностей SEQ ID №1, 2, 3, 4, 5 или 6, или по меньшей мере один CDR последовательности, которая по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентична после оптимального выравнивания последовательностям SEQ ID №№1, 2, 3, 4, 5 или 6.
В соответствии с другим аспектом изобретение относится к изолированному антителу или производному соединению или его функциональному фрагменту, содержащему по меньшей мере один CDR, выбранный среди CDR последовательностей SEQ ID №№40, 2, 41, 42, 5 или 43, или по меньшей мере один CDR последовательности, которая по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентична после оптимального выравнивания последовательностям SEQ ID №№40, 2, 41, 42, 5 или 43.
Под «функциональным фрагментом» антитела понимается, в частности, фрагмент антитела, такой как фрагменты Fv, scFv (sc обозначает одну цепь), Fab, F(ab')2, Fab', scFv-Fc, или димерные антитела или любой фрагмент, время полужизни которого было увеличено. Такие функциональные фрагменты будут подробно описаны ниже в данном описании.
Под «производным соединением» или «производным» антитела понимается, в частности, связывающий белок, содержащий пептидную матрицу и по меньшей мере один из CDR исходного антитела для поддержания способности распознавать CXCR4. Такие производные соединения, хорошо известные специалистам в данной области, будут описаны более подробно в данном описании.
Более предпочтительно изобретение содержит антитела, их производные соединения или их функциональные фрагменты в соответствии с данным изобретением, особенно химерные или гуманизированные, полученные путем генетической рекомбинации или химического синтеза.
В соответствии с предпочтительным воплощением антитело в соответствии с изобретением или его производные соединения или функциональные фрагменты характеризуются тем, что они состоят из моноклональных антител.
Под «моноклональным антителом» понимается антитело, полученное из популяции существенно однородных антител. Более конкретно отдельные антитела популяции являются идентичными, за исключением возможных природных мутаций, которые могут присутствовать в минимальных пропорциях. Иными словами, моноклональное антитело представляет собой однородное антитело, полученное в результате роста одного клона клеток (например, гибридомной клетки, эукариотической принимающей клетки, трансфицированной молекулой ДНК, кодирующей однородное антитело, прокариотической принимающей клетки, трансфицированной молекулой ДНК, кодирующей однородное антитело и т.д.), и в целом характеризуется тяжелыми цепями одного и только одного класса и подкласса и легкими цепями только одного типа. Моноклональные антитела высоко специфичны и направлены против одного антигена. Кроме того, в отличие от получения поликлональных антител, которые обычно включают различные антитела, направленные против различных детерминант, или эпитопов, каждое моноклональное антитело направлено против одного эпитопа антигена.
Здесь следует понимать, что изобретение не относится к антителам в природной форме, то есть они взяты не из их природной среды, но что они выделены или получены путем очистки из природных источников, либо получены путем генетической рекомбинации, либо путем химического синтеза, и, таким образом, они могут содержать неприродные аминокислоты, как будет описано ниже.
В частности, в соответствии с предпочтительным воплощением изобретения антитело или его производные соединения или функциональные фрагменты характеризуются тем, что они содержат легкую цепь, содержащую по меньшей мере один CDR, выбранный среди CDR аминокислотных последовательностей SEQ ID №№1, 2 или 3, или по меньшей мере один CDR последовательности, которая по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентична после оптимального выравнивания последовательностям SEQ ID №№1, 2 или 3; или они содержат тяжелую цепь, содержащую по меньшей мере один CDR, выбранный среди CDR аминокислотных последовательностей SEQ ID №№4, 5 или 6, или по меньшей мере один CDR последовательности, которая по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентична после оптимального выравнивания последовательностям SEQ ID №№4, 5 или 6.
В соответствии с другим воплощением антитела изобретения или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат легкую цепь, содержащую по меньшей мере один из трех CDR последовательностей SEQ ID №№1, 2 или 3, или по меньшей мере одной последовательности, которая по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентична после оптимального выравнивания последовательностям SEQ ID №№1, 2 или 3.
Предпочтительным образом антитела изобретения или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат легкую цепь, содержащую следующие три CDR, соответственно CDR-L1, L2-CDR и CDR-L3, где:
- CDR-L1 содержит последовательность SEQ ID №1 или 9 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №1 или 9;
- CDR-L2 содержит последовательность SEQ ID №2 или 10 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №2 или 10, и
CDR-L3 содержит последовательность SEQ ID №3 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №3.
В соответствии с частным воплощением антитела или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат легкую цепь, содержащую CDR-L1 последовательности SEQ ID №1, CDR-L2 последовательности SEQ ID №2 и CDR-L3 последовательности SEQ ID №3.
В соответствии с другим частным воплощением антитела или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат легкую цепь, содержащую CDR-L1 последовательности SEQ ID №9, CDR-L2 последовательности SEQ ID №10 и CDR-L3 последовательности SEQ ID №3.
В частности, антитела изобретения или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат тяжелую цепь, содержащую по меньшей мере один из трех CDR последовательностей SEQ ID №№4, 5 или 6, или по меньшей мере одной последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательностям SEQ ID №№4, 5 или 6.
Еще более предпочтительно антитела изобретения или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат тяжелую цепь, содержащую следующие три CDR, соответственно CDR-H1, CDR-H2 и CDR-H3, где:
- CDR-H1 содержит последовательность SEQ ID №4, 7 или 11 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №4, 7 или 11;
- CDR-H2 содержит последовательность SEQ ID №5 или 12 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №5 или 12, и
- CDR-H3 содержит последовательность SEQ ID №6 или 8 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №6 или 8.
В соответствии с частным воплощением антитела или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат тяжелую цепь, содержащую CDR-H1 последовательности SEQ ID №7, CDR-H2 последовательности SEQ ID №5 и CDR-H3 последовательности SEQ ID №8.
В соответствии с другим частным воплощением антитела или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат тяжелую цепь, содержащую CDR-H1 последовательности SEQ ID №11, CDR-H2 последовательности SEQ ID №12 и CDR-H3 последовательности SEQ ID №6.
В частности, в соответствии с предпочтительным воплощением изобретения антитело или его производные соединения или функциональные фрагменты характеризуются тем, что они содержат легкую цепь, содержащую по меньшей мере один CDR, выбранный среди CDR аминокислотных последовательностей SEQ ID №№40, 2 или 41, или по меньшей мере один CDR последовательности, которая по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентична после оптимального выравнивания последовательностям SEQ ID №№40, 2 или 41; или они содержат тяжелую цепь, содержащую по меньшей мере один CDR, выбранный среди CDR аминокислотных последовательностей SEQ ID №№42, 5 или 43, или по меньшей мере один CDR последовательности, которая по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентична после оптимального выравнивания последовательностям SEQ ID №№42, 5 или 43.
В соответствии с другим воплощением антитела изобретения или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат легкую цепь, содержащую по меньшей мере один из трех CDR последовательностей SEQ ID №№40, 2 или 41, или по меньшей мере одной последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательностям SEQ ID №№40, 2 или 41.
Предпочтительным образом антитела изобретения или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат легкую цепь, содержащую следующие три CDR, соответственно CDR-L1, L2-CDR и CDR-L3, где:
- CDR-L1 содержит последовательность SEQ ID №40 или 46 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №40 или 46;
- CDR-L2 содержит последовательность SEQ ID №2 или 47 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №2 или 47; и
CDR-L3 содержит последовательность SEQ ID №41 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №41.
В соответствии с частным воплощением антитела или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат легкую цепь, содержащую CDR-L1 последовательности SEQ ID №40, CDR-L2 последовательности SEQ ID №2 и CDR-L3 последовательности SEQ ID №41.
В соответствии с другим частным воплощением антитела или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат легкую цепь, содержащую CDR-L1 последовательности SEQ ID №46, CDR-L2 последовательности SEQ ID №47 и CDR-L3 последовательности SEQ ID №41.
В частности, антитела изобретения или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат тяжелую цепь, содержащую по меньшей мере один из трех CDR последовательностей SEQ ID №№42, 5 или 43, или по меньшей мере одной последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательностям SEQ ID №№42, 5 или 43.
Еще более предпочтительно антитела изобретения или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат тяжелую цепь, содержащую следующие три CDR, соответственно CDR-H1, CDR-H2 и CDR-H3, где:
- CDR-H1 содержит последовательность SEQ ID №42, 44 или 48 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №42, 44 или 48;
- CDR-H2 содержит последовательность SEQ ID №5 или 49 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №5 или 49; и
- CDR-H3 содержит последовательность SEQ ID №45 или 43 или последовательность, по меньшей мере на 80% идентичную после оптимального выравнивания последовательности SEQ ID №45 или 43.
В соответствии с частным воплощением антитела или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат тяжелую цепь, содержащую CDR-H1 последовательности SEQ ID №44, CDR-H2 последовательности SEQ ID №5 и CDR-H3 последовательности SEQ ID №45.
В соответствии с другим частным воплощением антитела или одно из их производных соединений или функциональных фрагментов характеризуются тем, что они содержат тяжелую цепь, содержащую CDR-H1 последовательности SEQ ID №48, CDR-H2 последовательности SEQ ID №49 и CDR-H3 последовательности SEQ ID №43.
В данном описании термины «полипептиды», «полипептидные последовательности», «пептиды» и «белки», применяемые по отношению к соединениям антителам или их последовательностям, являются взаимозаменяемыми.
Здесь следует понимать, что изобретение не относится к антителам в природной форме, то есть они взяты не из их природной среды, но они выделены или получены путем очистки из природных источников, либо получены путем генетической рекомбинации, либо путем химического синтеза, и, таким образом, они могут содержать неприродные аминокислоты, как будет описано ниже.
В первом воплощении участок, определяющий комплементарность, или CDR, обозначает гипервариабельные участки тяжелых и легких цепей иммуноглобулинов в соответствии с определением Kabat et al. (Kabat et al., Sequences of proteins of immunological interest, 5th Ed., U.S. Department of Health and Human Services, NIH, 1991, и более поздние издания). Существует три CDR тяжелой цепи и три CDR легкой цепи. Термин «CDR» в единственном или множественном числе применяется здесь для обозначения в зависимости от случая одного или более или даже всех этих участков, которые содержат большинство аминокислотных остатков, ответственных за аффинное связывание антитела с антигеном или эпитопом, который оно распознает.
Во втором воплощении под CDR-участками, или CDR, понимаются гипервариабельные участки тяжелых и легких цепей иммуноглобулинов в соответствии с определением IMGT.
Уникальная нумерация IMGT была создана для сравнения вариабельных доменов независимо от антигенного рецептора, типа цепи или вида [Lefranc M.-P., Immunology Today 18, 509 (1997) / Lefranc M.-P., The Immunologist, 7, 132-136 (1999) / Lefranc, M.-P., Pommie, С., Ruiz, M., Giudicelli, V., Foulquier, E., Truong, L., Thouvenin-Contet, V. and Lefranc, Dev. Сотр. ImmunoL, 27, 55-77 (2003)]. В уникальной нумерации IMGT консервативные аминокислоты всегда занимают одну и ту же позицию, например цистеин 23 (1-CYS), триптофан 41 (CONSERVED-TRP), гидрофобная аминокислота 89, цистеин 104 (2-CYS), фенилаланин или триптофан 118 (J-PHE или J-TRP). Уникальная нумерация IMGT предусматривает стандартизированное разграничение каркасных участков (FR1-IMGT: позиции 1-26, FR2-IMGT: 39-55, FR3-IMGT: 66-104 и FR4-IMGT: 118-128) и участков, определяющих комплементарность: CDR1-IMGT: 27-38, CDR2-IMGT: 56-65 и CDR3-IMGT: 105-117. Т.к. разрывы представляют собой незанятые позиции, то длины CDR-IMGT (показаны в скобках и разделены точками, например [8.8.13]) становятся важной информацией. Уникальная нумерация IMGT используется в 2D-графическом представлении, обозначенном как IMGT Colliers de Perles [Ruiz, M. and Lefranc, M.-P., Immunogenetics, 53, 857-883 (2002) / Kaas, Q. and Lefranc, M.-P., Current Bioinformatics, 2, 21-30 (2007)], и в 3D-структурах в IMGT/3D структуре-DB [Kaas, Q., Ruiz, M. and Lefranc, M.-P., Т cell receptor and MHC structural data. Nucl. Acids. Res., 32, D208-D210 (2004)].
Существует три CDR тяжелой цепи и три CDR легкой цепи. Термин «CDR» в единственном или множественном числе используется здесь для обозначения в зависимости от случая одного из этих участков или нескольких, или даже всех этих участков, которые содержат большинство аминокислотных остатков, ответственных за аффинное связывание антитела с антигеном или эпитопом, который оно распознает.
Для большей ясности, необходимо понимать, что в последующем описании, и в частности в таблицах 2 и 3, CDR будут определены в соответствии с нумерацией IMGT, нумерацией Кабата и общей нумерацией.
Общая нумерация объединяет остаточные части каждого CDR, которые являются общими для CDR, определенного в соответствии с системами нумерации IMGT и Кабата.
Система нумерации IMGT определяет CDR в соответствии с IMGT-системой, как определено выше, в то время как система нумерации Кабата определяет CDR в соответствии с системой Кабата, как определено выше.
В частности, CDR-L1 содержит SEQ ID №1 (QSLYNSRTRKNY) в общей системе нумерации и системе нумерации IMGT и SEQ ID №9 (KSSQSLYNSRTRKNYLA) в системе нумерации Кабата.
Что касается CDR-L2, он содержит SEQ ID №2 (WAS) в общей системе нумерации и системе нумерации IMGT и SEQ ID №10 (WASTRES) в системе нумерации Кабата.
CDR-L3 содержит SEQ ID №3 (KQSYNLRT) в каждой из трех систем нумерации.
Для тяжелой цепи, CDR-H1 содержит SEQ ID №4 (TDYY) в общей системе нумерации, SEQ ID №7 (GFTFTDYY) в системе нумерации IMGT и SEQ ID №11 (TDYYMS) в системе нумерации Кабата.
CDR-H2 содержит SEQ ID №5 (IRNKANGYTT) в общей системе нумерации и системе нумерации IMGT и SEQ ID №12 (FIRNKANGYTTEYSASVKG) в системе нумерации Кабата.
Наконец, CDR-H3 содержит SEQ ID №6 (DIPGFAY) в общей системе нумерации и системе нумерации Кабата, в то время как в системе нумерации Кабата он содержит SEQ ID №8 (ARDIPGFAY).
В частности, CDR-L1 содержит SEQ ID №40 (QSLFNSRTRKNY) в общей системе нумерации и системе нумерации IMGT и SEQ ID №46 (KSSQSLFNSRTRKNYLA) в системе нумерации Кабата.
Что касается CDR-L2, он содержит SEQ ID №2 (WAS) в общей системе нумерации и системе нумерации IMGT и SEQ ID №47 (WASARDS) в системе нумерации Кабата.
CDR-L3 содержит SEQ ID №41 (MQSFNLRT) в каждой из трех систем нумерации.
Для тяжелой цепи, CDR-H1 содержит SEQ ID №42 (DNY) в общей системе нумерации, SEQ ID №44 (GFTFTDNY) системе нумерации IMGT и SEQ ID №48 (DNYMS) в системе нумерации Кабата.
CDR-H2 содержит SEQ ID №5 (IRNKANGYTT) в общей системе нумерации и SEQ ID №49 (FIRNKANGYTTDYSASVRG) в системе нумерации Кабата.
Наконец, CDR-H3 содержит SEQ ID №43 (DVGSNYFDY) в общей системе нумерации и системе нумерации Кабата, в то время как в системе нумерации IMGT он содержит SEQ ID №45 (ARDVGSNYFDY).
В контексте данного изобретения «процент идентичности» двух нуклеиновокислотных или аминокислотных последовательностей обозначает процент идентичных нуклеотидов или аминокислотных остатков в двух сравниваемых последовательностях, полученный после оптимального выравнивания; этот процент не чисто статистический, и различия между двумя последовательностями распределяются случайным образом по всей их длине. Сравнение двух нуклеиновокислотных или аминокислотных последовательностей традиционно проводится путем сравнения этих последовательностей после выравнивания их оптимальным образом; указанное сравнение может осуществляться по сегментам или с помощью «окна сравнения». Оптимальное выравнивание последовательностей для сравнения может быть осуществлено, в дополнение к ручному, с помощью алгоритма локальной гомологии Смита-Уотермана (1981) [Ad. App.Math. 2:482], с помощью алгоритма локальной гомологии Нидлмана-Вунша (1970) [J. Mol. Biol. 48: 443], с помощью способа поиска сходства Пирсона и Липмана (1988) [Proc. Natl. Acad. Sci. USA 85:2444) или с помощью компьютерного программного обеспечения с использованием этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wl, либо с помощью программного обеспечения для сравнения BLAST N или BLAST Р).
Процент идентичности двух нуклеиновокислотных или аминокислотных последовательностей определяют путем сравнения двух последовательностей, выравненных оптимальным образом, при этом сравниваемая нуклеиновокислотная или аминокислотная последовательность может включать добавления или делеции по сравнению с референсной последовательностью для оптимального выравнивания этих двух последовательностей. Процент идентичности рассчитывается путем определения числа позиций, в которых нуклеотид или аминокислотный остаток идентичен в двух последовательностях, предпочтительно в двух полных последовательностях, деления этого количества идентичных позиций на общее количество позиций в окне выравнивания и умножения полученного результата на 100, чтобы получить процент идентичности этих двух последовательностей.
Например, программа BLAST «BLAST 2 sequences» (Tatusova et al, "Blast 2 sequences - a new tool for comparing protein and nucleotide sequences", FEMS Microbiol, 1999, Lett. 174:247-250), доступная на сайте http://www.ncbi.nlm.nih.gov/gorf/bl2.html, может быть использована с параметрами по умолчанию (в частности, с параметрами «штраф за открытие делеции»: 5 и «штраф за удлинение делеции»: 2; выбранной матрицей будет, например, матрица «BLOSUM 62», предложенная в программе); процентная идентичность двух сравниваемых последовательностей рассчитывается непосредственно в программе.
Среди аминокислотных последовательностей, имеющих по меньшей мере 80%, предпочтительно 85%, 90%, 95% и 98% идентичности с референсной аминокислотной последовательностью, предпочтительными примерами являются те, которые содержат референсную последовательность, определенные модификации, в частности делецию, добавление или замену по меньшей мере одной аминокислоты, усечение или удлинение. В случае замены одной или более последовательных или непоследовательных аминокислот предпочтительными являются такие замены, в которых замененные аминокислоты заменены «эквивалентными» аминокислотами. Выражение «эквивалентные аминокислоты» используется здесь для обозначения любой аминокислоты, которая может быть заменена одной из структурных аминокислот, но без значительной модификации биологических активностей соответствующих антител и таких специфических примеров, какие будут определены позже.
Эквивалентные аминокислоты могут быть определены либо на основании их структурной гомологии с аминокислотами, которые они заменяют, либо по результатам сравнительных исследований биологической активности различных антител, на которую они способны.
В качестве неограничивающего примера приведенная ниже таблица 1 показывает замены, возможные без получения значительной модификации биологической активности соответствующего модифицированного антитела; обратные замены, разумеется, также возможны в тех же условиях.
Специалистам в данной области известно, что на текущем уровне техники наибольшая вариабельность (длины и состава) среди шести CDR находится в трех CDR тяжелой цепи, и, в частности, в CDR-H3 этой тяжелой цепи. Следовательно, будет очевидно, что предпочтительными характерными CDR антител изобретения или одного из их производных соединений или функциональных фрагментов будут три CDR тяжелой цепи, т.е. для 4141-15 CDR, кодируемые последовательностями SEQ ID №№7, 5, 8 и 11, 12, 6, определенные в соответствии с IMGT и Кабат соответственно, а для 515Н7 CDR, кодируемые последовательностями SEQ ID №№44, 5, 45 и 48, 49, 43, определенные в соответствии с IMGT и Кабат соответственно. Еще более предпочтителен CDR, соответствующий CDR-H3, кодируемый последовательностью SEQ ID №8 или 6 для 414Н5 и 45 или 43 для 515Н7.
В конкретном воплощении данное изобретение относится к мышиному антителу или его производным соединениям или функциональным фрагментам.
Другое воплощение изобретения раскрывает антитело или его производные соединения или функциональные фрагменты, содержащие легкую цепь с тремя следующими CDR:
CDR-L1 последовательности SEQ ID №1 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального соответствия последовательности SEQ ID №1;
CDR-L2 последовательности SEQ ID №2 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального соответствия последовательности SEQ ID №2; и
CDR-L3 последовательности SEQ ID №3 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального соответствия последовательности SEQ ID №3;
и тяжелую цепь с тремя следующими CDR:
CDR-H1 последовательности SEQ ID №4 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального соответствия последовательности SEQ ID №4;
CDR-H2 последовательности SEQ ID №5 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального соответствия последовательности SEQ ID №5; и
CDR-H3 последовательности SEQ ID №6 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального соответствия последовательности SEQ ID №6.
Еще одно воплощение изобретения раскрывает антитело или его производное соединение или функциональный фрагмент, содержащий легкую цепь с тремя следующими CDR:
- CDR-L1 последовательности SEQ ID №1 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №1;
- CDR-L2 последовательности SEQ ID №2 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №2; и
- CDR-L3 последовательности SEQ ID №3 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №3;
и тяжелую цепь с тремя следующими CDR:
- CDR-H1 последовательности SEQ ID №7 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №7;
- CDR-H2 последовательности SEQ ID №5 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №5; и
- CDR-H3 последовательности SEQ ID №8 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №8.
Еще одно воплощение изобретения раскрывает антитело или его производное соединение или функциональный фрагмент, содержащий легкую цепь с тремя следующими CDR:
- CDR-L1 последовательности SEQ ID №9 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №9;
- CDR-L2 последовательности SEQ ID №10 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №10; и
- CDR-L3 последовательности SEQ ID №3 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №3;
и тяжелую цепь с тремя следующими CDR:
- CDR-H1 последовательности SEQ ID №11 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №11;
- CDR-H2 последовательности SEQ ID №12 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №12; и
- CDR-H3 последовательности SEQ ID №6 или последовательности, по меньшей мере на 80% идентичной после оптимального соответствия последовательности SEQ ID №6.
Антитело или его производное соединение или функциональный фрагмент в соответствии с изобретением характеризуется тем, что содержит:
- легкую цепь, содержащую CDR-L1 последовательности SEQ ID №1, CDR-L2 последовательности SEQ ID №2 и CDR-L3 последовательности SEQ ID №3; и
- тяжелую цепь, содержащую CDR-H1 последовательности SEQ ID №7, CDR-H2 последовательности SEQ ID №5 и CDR-H3 последовательности SEQ ID №8.
В другом воплощении антитело или его производное соединение или функциональный фрагмент в соответствии с изобретением характеризуется тем, что содержит:
- легкую цепь, содержащую CDR-L1 последовательности SEQ ID №9, CDR-L2 последовательности SEQ ID №10 и CDR-L3 последовательности SEQ ID №3; и
- тяжелую цепь, содержащую CDR-H1 последовательности SEQ ID №11, CDR-H2 последовательности SEQ ID №12 и CDR-H3 последовательности SEQ ID №6.
В соответствии с еще одним воплощением антитело изобретения или его производные соединения или функциональные фрагменты характеризуются тем, что содержат последовательность легкой цепи с аминокислотной последовательностью SEQ ID №13 или последовательностью, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №13; и тем, что они содержат последовательность тяжелой цепи с аминокислотной последовательностью SEQ ID №14 или последовательностью, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №14.
Другое воплощение изобретения раскрывает антитело или его производные соединения или функциональные фрагменты, содержащие легкую цепь с тремя следующими CDR:
CDR-L1 последовательности SEQ ID №40 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №40;
CDR-L2 последовательности SEQ ID №2 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №2; и
CDR-L3 последовательности SEQ ID №41 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №41;
и тяжелую цепь с тремя следующими CDR:
CDR-H1 последовательности SEQ ID №42 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №42;
CDR-H2 последовательности SEQ ID №5 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №5; и
CDR-H3 последовательности SEQ ID №43 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №43.
Еще одно воплощение изобретения раскрывает антитело или его производное соединение или функциональный фрагмент, содержащий легкую цепь с тремя следующими CDR:
- CDR-L1 последовательности SEQ ID №40 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №40;
- CDR-L2 последовательности SEQ ID №2 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №2; и
- CDR-L3 последовательности SEQ ID №41 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №41;
и тяжелую цепь с тремя следующими CDR:
- CDR-H1 последовательности SEQ ID №44 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №44;
- CDR-H2 последовательности SEQ ID №5 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №5; и
- CDR-H3 последовательности SEQ ID №45 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №45.
Еще одно воплощение изобретения раскрывает антитело или его производное соединение или функциональный фрагмент, содержащий легкую цепь с тремя следующими CDR:
- CDR-L1 последовательности SEQ ID №46 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №46;
- CDR-L2 последовательности SEQ ID №47 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №47; и
- CDR-L3 последовательности SEQ ID №41 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №41;
и тяжелую цепь с тремя следующими CDR:
- CDR-H1 последовательности SEQ ID №48 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №48;
- CDR-H2 последовательности SEQ ID №49 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №49; и
- CDR-H3 последовательности SEQ ID №43 или последовательности, по меньшей мере на 80% идентичной после оптимального выравнивания последовательности SEQ ID №43.
Антитело или его производное соединение или функциональный фрагмент в соответствии с изобретением характеризуется тем, что содержит:
- легкую цепь, содержащую CDR-L1 последовательности SEQ ID №40, CDR-L2 последовательности SEQ ID №2 и CDR-L3 последовательности SEQ ID №41; и
- тяжелую цепь, содержащую CDR-H1 последовательности SEQ ID №44, CDR-H2 последовательности SEQ ID №5 и CDR-H3 последовательности SEQ ID №45.
В другом воплощении антитело или его производное соединение или функциональный фрагмент в соответствии с изобретением характеризуется тем, что содержит:
- легкую цепь, содержащую CDR-L1 последовательности SEQ ID №46, CDR-L2 последовательности SEQ ID №47 и CDR-L3 последовательности SEQ ID №41; и
- тяжелую цепь, содержащую CDR-H1 последовательности SEQ ID №48, CDR-H2 последовательности SEQ ID №49 и CDR-H3 последовательности SEQ ID №43.
В соответствии с еще одним воплощением антитело изобретения или его производные соединения или функциональные фрагменты характеризуются тем, что они содержат последовательность легкой цепи с аминокислотной последовательностью SEQ ID №50 или последовательностью, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №50; и тем, что они содержат последовательность тяжелой цепи с аминокислотной последовательностью SEQ ID №51 или последовательностью, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №51.
Как было сказано выше, изобретение также относится к любому соединению, полученному из антитела, описанного в изобретении.
В частности, антитело изобретения или его производные соединения или функциональные фрагменты характеризуются тем, что указанные производные соединения состоят из связывающего белка, содержащего пептидную матрицу, к которой привит по меньшей мере один CDR для сохранения целиком или частично свойств паратопа исходного антитела по распознаванию.
На различных иммуноглобулиновых белковых матрицах может присутствовать одна или более последовательность из числа шести CDR-последовательностей, описанных в данном изобретении. В этом случае белковая матрица делает возможным воспроизведение белкового скелета с соответствующим сворачиванием привитых CDR, тем самым позволяя их паратопу сохранять свойства по распознаванию антигена.
Как правило, специалистам в данной области известно, как выбирать тип белковой матрицы, на которую можно привить по меньшей мере один CDR, взятый из исходного антитела. В частности, известно, что такие матрицы, чтобы быть выбранными, должны соответствовать максимальному количеству из следующих критериев (Skerra A., J. Mol. Recogn., 13, 2000, 167-187):
- филогенетически хорошая сохранность;
- известная трехмерная структура (например, с помощью кристаллографии или ЯМР-спектроскопии или любых других методик, известных специалистам в данной области);
- малый размер;
- отсутствие или наличие только в малой степени посттрансляционных модификаций; и/или
- легкость продукции, экспрессии и очистки.
Такая белковая матрица может быть структурой, выбранной среди: фибронектина и предпочтительно десятого домена фибронектина III типа, липокалина, антикалина (Skerra A., J. Biotechnol., 2001, 74(4):257-75), белка Z, полученного из домена В белка A Staphylococcus aureus, тиоредоксина А или белков с повторяющимся мотивом, таким как «анкириновый повтор» (Kohl et al., PNAS, 2003, vol. 100, No. 4, 1700-1705), «повтор Армадилла», «повтор, богатый лейцином» или «тетратрикопептидный повтор», но не ограничиваясь ими.
Также можно упомянуть матрицы, полученные из токсинов, таких как, например, токсины скорпионов, насекомых, растений, моллюсков и т.д., или белковых ингибиторов нейронной синтазы оксида азота (PIN).
В качестве неограничивающего примера таких гибридных конструкций можно отметить вставку CDR-H1 (тяжелая цепь) анти-С04-антитела, а именно антитела 13 В8.2, в одну из петель PIN; связывающие свойства полученного таким образом нового связывающего белка остаются похожими на связывающие свойства исходного антитела (Bes et al., Biochem. Biophys. Res. Commun., 2006, 343(1), 334-344). Чисто иллюстративно можно также упомянуть прививку CDR-H3 (тяжелая цепь) анти-лизоцим-VHH антитела в петлю неокарциностатина (Nicaise et al.. Protein Science, 2004, 13(7): 1882-1891).
Наконец, как упоминалось выше, такие пептидные матрицы могут содержать от одного до шести CDR из исходного антитела. Предпочтительно, но без каких-либо ограничений, специалист в данной области может выбрать по меньшей мере один CDR из тяжелой цепи, которая, как известно, главным образом ответственна за специфичность антитела. Выбор одного или более релевантного CDR будет очевидным для специалиста в данной области, который также выберет подходящую из известных методик (Bes etal., FEBS letters 508, 2001, 67-74).
Специфический аспект данного изобретения относится к способу выбора соединения, полученного из антитела в соответствии с изобретением, где указанное производное соединение способно ингибировать in vitro и/или in vivo рост опухолевых клеток, и где указанное производное соединение содержит пептидную матрицу, к которой привит по меньшей мере один CDR антитела, характеризующегося тем, что он (способ) включает следующие этапы:
а) контактирование in vitro соединения, состоящего из пептидной матрицы, к которой привит по меньшей мере один CDR антитела, с биологическим образцом, содержащим опухолевые клетки, способные расти, в условиях, позволяющих этим клеткам расти; и
б) выбор указанного соединения, если указанное соединение способно ингибировать рост этих опухолевых клеток,
и характеризуется тем, что по меньшей мере один привитый CDR выбран среди следующих CDR:
- CDR последовательности SEQ ID №1, 9, 40, 46 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №1, 9, 40, 46;
- CDR последовательности SEQ ID №2, 10, 47 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №2, 10, 47;
- CDR последовательности SEQ ID №3, 41 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №3, 41;
- CDR последовательности SEQ ID №4, 7, 11, 42, 44, 48 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №4, 7, 11,42,44,48;
- CDR последовательности SEQ ID №5, 12, 49 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №5, 12, 49; и
- CDR последовательности SEQ ID №6, 8, 43, 45 или последовательности, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичной после оптимального выравнивания последовательности SEQ ID №6, 8, 43, 45.
В соответствии с предпочтительным режимом способ может включать в этапе а) контактирование in vitro соединения, содержащего пептидную матрицу, к которой привито по меньшей мере два или три CDR антитела.
В соответствии с еще более предпочтительным режимом этого способа пептидную матрицу выбирают среди матриц или связывающих белков, структуры которых были упомянуты выше.
Очевидно, что эти примеры ни в коей мере не являются ограничивающими, и любые другие структуры, известные или очевидные специалистам в данной области, следует рассматривать как находящиеся под защитой, предоставляемой данной патентной заявкой.
Данное изобретение также относится к антителу или его производным соединениям или функциональным фрагментам, характеризующимся тем, что пептидная матрица выбрана среди белков, которые обладают а) филогенетически хорошей сохранностью, б) надежной архитектурой, в) хорошо известной трехмерной организацией, г) малым размером и/или д) содержат области, которые можно модифицировать путем делеции и/или вставки без модификации свойств стабильности.
В соответствии с предпочтительным воплощением антитело изобретения или его производные соединения или функциональные фрагменты характеризуются тем, что указанная пептидная матрица выбрана среди i) матриц, полученных из фибронектина, предпочтительно десятого домена фибронектина III типа, липокалина, антикалина, белка Z, полученного из домена В белка А Staphylococcus aureus, тиоредоксина А или белков с повторяющимся мотивом, таким как «анкириновый повтор» (Kohl et al., PNAS, 2003, vol. 100, No. 4, 1700-1705), «повтор Армадилла», «повтор, богатый лейцином» и «тетратрикопептидный повтор», или ill) белковых ингибиторов нейронной синтазы оксида азота (PIN).
Другой аспект изобретения относится к функциональным фрагментам антитела, описанным выше.
Более конкретно, изобретение нацелено на антитело или его производные соединения или функциональные фрагменты, характеризующиеся тем, что указанный функциональный фрагмент выбран среди фрагментов Fv, Fab, F(ab')2, Fab', scFv, scFv-Fc или димерных антител или любых фрагментов, время полужизни которых было увеличено, например пегилированных фрагментов.
Такие функциональные фрагменты антитела в соответствии с изобретением состоят, например, из фрагментов Fv, scFv (sc обозначает одну цепь), Fab, F(ab')2, Fab', scFv-Fc, или димерных антител или любых фрагментов, время полужизни которых было увеличено в результате химической модификации, такой как добавление полиалкиленгликоля, такого как полиэтиленгликоль (пегилирование) (пегилированные фрагменты называются Fv-ПЭГ, ScFv-ПЭГ, Fab-ПЭГ, Р(аb')2-ПЭГ или Fab'-ПЭГ), либо путем включения в липосому; микросферу или PLGA (частицы на основе сополимеров молочной и гликолевой кислот), при этом указанный фрагмент имеет по меньшей мере один характерный CDR изобретения, который способен проявлять в общем виде даже частичную активность антитела, из которого он взят.
Предпочтительно указанные функциональные фрагменты будут состоять из или включать частичную последовательность вариабельной тяжелой или легкой цепи антитела, из которого они получены, при этом указанная частичная последовательность должна быть достаточной, чтобы сохранить такую же специфичность связывания, как у антитела, из которого она получена, и достаточную аффинность, предпочтительно равную по меньшей мере 1/100, более предпочтительно по меньшей мере 1/10 аффинности антитела, из которого она получена.
Такой функциональный фрагмент будет содержать как минимум 5 аминокислот, предпочтительно 6, 7, 8, 10, 15, 25, 50 или 100 последовательных аминокислот последовательности антитела, из которого он получен.
Предпочтительно этими функциональными фрагментами будут фрагменты типа Fv, scFv, Fab, F(аb')2, F(ab'), scFv-Fc или димерные антитела, которые в целом имеют такую же специфичность связывания, как и антитело, из которого они получены. В соответствии с данным изобретением фрагменты антитела изобретения могут быть получены из антител описанными выше способами, такими как расщепление ферментами, такими как пепсин или папаин, и/или расщепление дисульфидных мостиков путем химического восстановления. Фрагменты антитела также могут быть получены с помощью методик генетической рекомбинации, также известных специалистам в данной области, или путем пептидного синтеза, например с помощью автоматических пептидных синтезаторов, например поставляемых компанией Applied Biosystems, и т.д.
Для большей ясности в таблице 2 ниже представлены различные аминокислотные последовательности, соответствующие антителу изобретения.
Другой конкретный аспект данного изобретения относится к химерному антителу или его производным соединениям или функциональным фрагментам, характеризующимся тем, что указанное антитело также включает константные участки легкой цепи и тяжелой цепи, полученные из антитела видов, гетерологичных мыши, в частности, человека.
Еще один конкретный аспект данного изобретения относится к гуманизированному антителу или его производным соединениям или функциональным фрагментам, характеризующимся тем, что константные участки легкой цепи и тяжелой цепи, полученные из человеческого антитела, являются соответственно участками лямбда или каппа и гамма-1, гамма-2 или гамма-4.
В соответствии с другим аспектом изобретение относится к мышиной гибридоме, способной секретировать моноклональное антитело в соответствии с изобретением, в частности к гибридоме мышиного происхождения, зарегистрированной во Французской коллекции культур микроорганизмов (CNCM, Институт Пастера, Париж, Франция) 22 октября 2007 под номером 1-3860. Указанная гибридома была получена путем слияния спленоцитов иммунизированных мышей BALB/c и клеток миеломной линии Sp 2/0-Ag 14.
Моноклональное антитело, обозначаемое в данном документе как 414Н5, или его производные соединения или функциональные фрагменты, характеризующиеся тем, что указанное антитело секретируется гибридомой, зарегистрированной в CNCM 22 октября 2007 года под номером 1-3860, очевидно, является частью данного изобретения.
В соответствии с другим аспектом изобретение относится к мышиной гибридоме, способной секретировать моноклональное антитело в соответствии с изобретением, в частности к гибридоме мышиного происхождения, зарегистрированной во Французской коллекции культур микроорганизмов (CNCM, Институт Пастера, Париж, Франция) 25 июня 2008 под номером 1-4019. Указанная гибридома была получена путем слияния спленоцитов иммунизированных мышей BALB/c и клеток миеломной линии Sp 2/0-Ag 14.
Моноклональное антитело, обозначаемое в данном документе как 515Н7, или его производные соединения или функциональные фрагменты, характеризующиеся тем, что указанное антитело секретируется гибридомой, зарегистрированной в CNCM 25 июня 2008 года под номером 1-4019, очевидно, является частью данного изобретения.
Антитело изобретения также включает химерные или гуманизированные антитела.
Химерное антитело представляет собой антитело, которое содержит природный вариабельный участок (легкой цепи и тяжелой цепи), полученный из антитела данного вида, в сочетании с константными участками легкой цепи и тяжелой цепи антитела вида, гетерологичного указанному данному виду.
Антитела или их химерные фрагменты могут быть получены с помощью методик генетической рекомбинации. Например, химерное антитело может быть получено путем клонирования рекомбинантной ДНК, содержащей промотор и, последовательность, кодирующую вариабельный участок нечеловеческого, особенно мышиного, моноклонального антитела в соответствии с изобретением, и последовательность, кодирующую константный участок человеческого антитела. Химерное антитело в соответствии с изобретением, закодированное таким рекомбинантным геном, будет, например, химерой мыши и человека, при этом специфичность этого антитела будет определяться вариабельным участком, полученным из мышиной ДНК, а его изотип будет определяться константным участком, полученным из человеческой ДНК. Для способов получения химерных антител можно, например, сослаться на Verhoeyn etal. (BioEssays, 8:74, 1988).
В другом аспекте изобретение описывает антитело или его производное соединение или функциональный фрагмент, который состоит из химерного антитела.
В частном предпочтительном воплощении химерное антитело или его производное соединение или функциональный фрагмент изобретения содержит последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID №64, и последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID №65.
В другом предпочтительном воплощении химерное антитело или его производное соединение или функциональный фрагмент изобретения содержит последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID №66, и последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID №67.
Под «гуманизированными антителами» понимаются антитела, которые содержат CDR-участки, полученные из антитела нечеловеческого происхождения, при этом другие части молекулы антитела получены из одного (или нескольких) человеческих антител. Кроме того, некоторые остатки сегментов скелета (называемые FR, каркасные участки) могут быть изменены в целях сохранения аффинности связывания (Jones et al., Nature, 321:522-525, 1986; Verhoeyen et al., Science, 239:1534-1536, 1988; Riechmann etal., Nature, 332:323-327, 1988).
Гуманизированные антитела изобретения или их фрагменты могут быть получены с помощью методик, известных специалистам в данной области (например таких, которые описаны в документах Singer et al., J. Immun. 150:2844-2857, 1992; Mountain et al., Biotechnol. Genet. Eng. Rev., 10: 1-142, 1992; и Bebbington et al., Bio/Technology, 10:169-175, 1992). Такие гуманизированные антитела являются предпочтительными для их применения в способах, используемых в диагностике in vitro или профилактике и/или терапевтическом лечении in vivo. Другой методикой гуманизации, также известной специалистам в данной области, является, например, методика «CDR-прививки», описанная PDL в патентах ЕР 0 451261, ЕР 0 682 040, ЕР 0 9127, ЕР 0 566 647 или US 5530101, US 6180370, US 5585089 и US 5693761. Также можно отметить патенты США 5639641 или 6054297,5886152 и 5877293.
Кроме того, изобретение также относится к гуманизированным антителам, полученным из мышиных антител, описанных выше.
В предпочтительном виде константные участки легких цепей и тяжелых цепей, полученные из человеческого антитела, являются соответственно участками лямбда или каппа и гамма-1, гамма-2 или гамма-4.
В воплощении, соответствующем lgG1 изотипу lgG1 дополнительные характеристики антитела включают эффекторные функции, такие как антитело-зависимую клеточную цитотоксичность (antibody-dependent cell-mediated cytotoxicity, ADCC); и/или комплемент-зависимую цитотоксичность (complement dependent cytotoxicity, CDC).
Новый аспект данного изобретения относится к изолированной нуклеиновой кислоте, характеризующейся тем, что она выбрана среди следующих нуклеиновых кислот (включая любой вырожденный генетический код):
- нуклеиновой кислоты, ДНК или РНК, кодирующей антитело или его производное соединение или функциональный фрагмент в соответствии с изобретением;
нуклеиновой кислоты, комплементарной нуклеиновой кислоте, определенной в а);
- нуклеиновой кислоты не менее 18 нуклеотидов, способной к гибридизации в условиях высокой жесткости по меньшей мере с одним CDR нуклеиновокислотных последовательностей SEQ ID №№15-26 или SEQ ID №№52-61 или последовательностей, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичных после оптимального выравнивания последовательностям SEQ ID №№15-26 или SEQ ID №№52-61; и
- нуклеиновой кислоты не менее 18 нуклеотидов, способной к гибридизации в условиях высокой жесткости по меньшей мере с легкой цепью нуклеиновокислотной последовательности SEQ ID №27 или SEQ ID №62 или SEQ ID №68 или 70 и/или тяжелой цепью нуклеиновокислотной последовательности SEQ ID №28 или SEQ ID №63 или SEQ ID №69 или 71 или последовательностей, по меньшей мере на 80%, предпочтительно на 85%, 90%, 95% и 98% идентичных после оптимального выравнивания последовательностям SEQ ID №№27 и/или 28 или SEQ ID №62 и/или 63 или SEQ ID №№68 и/или 69 или SEQ ID №№70 и/или 71.
В таблице 3 приведены различные нуклеотидные последовательности, касающиеся антитела изобретения.
Термины «нуклеиновая кислота», «нуклеиновая последовательность» или «нуклеиновокислотная последовательность», «полинуклеотид», «олигонуклеотид», «полинуклеотидная последовательность» и «нуклеотидная последовательность», которые используются равнозначно в данном описании, обозначают четкую последовательность нуклеотидов, модифицированных или не модифицированных, определяющую фрагмент или участок нуклеиновой кислоты, содержащую или не содержащую неприродные нуклеотиды и являющуюся либо двуцепочечной ДНК, либо одноцепочечной ДНК, либо продуктами транскрипции указанных ДНК.
Здесь также следует упомянуть, что данное изобретение не относится к нуклеотидным последовательностям в их природной хромосомной среде, т.е. в природном состоянии. Последовательности данного изобретения были выделены и/или очищены, т.е. были взяты прямо или косвенно, например путем копирования, при этом их среда была по меньшей мере частично модифицирована. Таким образом, также здесь следует подразумевать изолированные нуклеиновые кислоты, полученные путем генетической рекомбинации, например с помощью принимающих клеток (клеток-хозяев), или полученные путем химического синтеза.
Под «нуклеиновыми последовательностями, демонстрирующими процент идентичности по меньшей мере 80%, предпочтительно 85%, 90%, 95% и 98%, после оптимального выравнивания с предпочтительной последовательностью» понимают нуклеиновые последовательности, демонстрирующие по сравнению с референсной нуклеотидной последовательностью определенные модификации, такие как, в частности, делецию, усечение, удлинение, химерное слияние и/или замещение, в частности, точечное. Предпочтительно это последовательности, которые кодируют те же самые аминокислотные последовательности, что и референсная последовательность, и связаны с вырождением генетического кода, или комплементарные последовательности, которые могут специфически гибридизироваться с референсными последовательностями, предпочтительно в условиях высокой жесткости, в частности таких, которые определены ниже.
Гибридизация в условиях высокой жесткости означает, что температурные условия и условия ионной силы выбирают таким образом, чтобы они позволяли сохранять гибридизацию двух комплементарных фрагментов ДНК. Исключительно в качестве примера, условия высокой жесткости этапа гибридизации для определения полинуклеотидных фрагментов, описанных выше, преимущественно являются следующими.
ДНК-ДНК- или ДНК-РНК-гибридизацию осуществляют в два этапа: (1) прегибридизация при 42°С в течение трех часов в фосфатном буфере (20 мМ, рН 7,5), содержащем 5х SSC (цитратно-солевой буфер) (1х SSC соответствует раствору 0,15 М NaCl+0,015 М цитрата натрия), 50% формамида, 7% додецилсульфата натрия (SDS), 10х раствор Денхардта, 5% сульфата декстрана и 1% ДНК спермы лосося; (2) собственно гибридизация в течение 20 часов при температуре в зависимости от длины зонда (например: 42°С для зонда длиной более 100 нуклеотидов), а затем два 20-минутных промывания при температуре 20°С в 2х SSC+2% SDS, одно 20-минутное промывание при 20°С в 0,1х SSC+0,1% SDS. Последнее промывание осуществляют в 0,1х SSC+0,1% SDS в течение 30 минут при температуре 60°С для зонда длиной более 100 нуклеотидов. Гибридизационные условия высокой жесткости, описанные выше для полинуклеотида определенного размера, специалист в данной области может адаптировать для олигонуклеотидов большей или меньшей длины в соответствии с процедурами, описанными в Sambrook, ef a/. (Molecular cloning: a laboratory manual, Cold Spring Harbor Laboratory; 3rd edition, 2001).
Изобретение также относится к вектору, содержащему нуклеиновую кислоту, описанную в изобретении.
Изобретение особенно нацелено на клонирующие и/или экспрессионные векторы, которые содержат такую нуклеотидную последовательность.
Векторы изобретения предпочтительно содержат элементы, которые позволяют экспрессировать и/или секретировать нуклеотидные последовательности в данной принимающей клетке (клетке-хозяине). Таким образом, вектор должен содержать промотор, сигналы инициации и терминации трансляции, а также подходящие участки регуляции транскрипции. Он должен быть способен поддерживать в стабильном состоянии принимающую клетку (клетку-хозяина) и дополнительно может иметь специфические сигналы, которые обуславливают секрецию транслированного белка. Специалисты в данной области выбирают и оптимизируют эти различные элементы в соответствии с используемой принимающей клеткой. С этой целью нуклеотидные последовательности могут быть вставлены в векторы автономной репликации в выбранном хозяине, или могут быть интегративными векторами выбранного хозяина.
Такие векторы получают с помощью способов, использующихся в настоящее время специалистами в данной области, и полученные клоны могут быть введены в подходящего хозяина с помощью стандартных способов, таких как липофекция, электропорация, тепловой шок или химические способы.
Векторы представляют собой, например, векторы плазмидного или вирусного происхождения. Они используются для трансформации принимающих клеток (клеток-хозяев), чтобы клонировать или экспрессировать нуклеотидные последовательности изобретения.
Изобретение также включает принимающие клетки, трансформированные с помощью вектора, описанного в данном изобретении, или содержащие его.
Принимающая клетка может быть выбрана из прокариотических или эукариотических систем, например бактериальных клеток, а также дрожжевых клеток или клеток животных, в частности клеток млекопитающих. Можно также использовать клетки насекомых или растений.
Изобретение также касается животных, за исключением человека, которые содержат по меньшей мере одну трансформированную клетку в соответствии с изобретением.
Другой аспект изобретения связан со способом продукции антитела в соответствии с изобретением или одного из его функциональных фрагментов, который характеризуется тем, что включает следующие этапы:
a) культивирование принимающей клетки в соответствии с изобретением в среде в подходящих культуральных условиях; и
b) восстановление указанного антитела или одного из его функциональных фрагментов, полученных таким образом, из культуральной среды или указанных культивируемых клеток.
Трансформированные клетки в соответствии с изобретением могут быть использованы в способах получения рекомбинантных полипептидов в соответствии с изобретением. Способы получения полипептида в соответствии с изобретением в рекомбинантной форме, характеризующиеся использованием вектора и/или клетки, трансформированной вектором в соответствии с изобретением, также входят в данное изобретение. Предпочтительно клетку, трансформированную вектором в соответствии с изобретением, культивируют в условиях, которые позволяют экспрессировать указанный полипептид, и указанный рекомбинантный пептид восстанавливают.
Как уже было сказано, принимающая клетка (клетка-хозяин) может быть выбрана из прокариотических или эукариотических систем. В частности, можно определить нуклеотидные последовательности изобретения, содействующие секреции в такой прокариотической или эукариотической системе. Вектор в соответствии с изобретением, несущий такую последовательность, таким образом, может быть выгодно использован для продукции рекомбинантных белков, предназначенных для секреции. Действительно, очистке этих целевых рекомбинантных белков будет способствовать тот факт, что они находятся в супернатанте клеточной культуры, а не внутри принимающих клеток.
Также можно получить полипептиды изобретения путем химического синтеза. Такой способ получения также является предметом изобретения. Специалисту в данной области известны способы химического синтеза, например методики с использованием твердых фаз [см. Steward et al., 1984, Solid phase peptide synthesis, Pierce Chem. Company, Rockford, 111, 2nd ed., (1984)] или методики с использованием частично твердых фаз, путем конденсации фрагментов или путем классического синтеза в растворе. Полипептиды, полученные путем химического синтеза и способные содержать соответствующие неприродные аминокислоты, также входят в изобретение.
Антитела или их производные соединения или функциональные фрагменты, которые могут быть получены с помощью способа изобретения, также входят в данное изобретение.
В соответствии с еще одним аспектом данное изобретение относится к антителу, описанному выше, которое характеризуется тем, что оно, кроме того, способно специфически связываться с рецептором семейства человеческих хемокинов и/или способно специфически ингибировать передачу сигнала от такого рецептора.
В соответствии с новым воплощением изобретение относится к антителу или его производным соединениям или функциональным фрагментам, состоящим из антитела, которое является биспецифическим в том смысле, что оно содержит второй мотив, способный взаимодействовать с каким-либо рецептором, вовлеченным в развитие опухолей, таким как, например, VEGFR, VEGF, EGFR, IGF-1R, HER2neu, HGF, СМЕТ, FGF, тетраспанины, интегрины, CXCR4 (кроме антитела данного изобретения, т.е. ориентированный на другой эпитоп), CXCR7 или CXCR2.
Биспецифические или бифункциональные антитела представляют собой второе поколение моноклональных антител, в которых два различных вариабельных участка объединены в одну молекулу (Hollinger and Bohlen, 1999, Cancer and metastasis, rev. 18:411-419). Как в диагностической, так и в терапевтической областях была продемонстрирована их полезность за счет их способности привлекать новые эффекторные функции или влиять на некоторые молекулы на поверхности опухолевых клеток; такие антитела могут быть получены с помощью химических способов (Glennie MJ et al., 1987, J. Immunol. 139, 2367-2375; Repp R. et al., 1995, J. Hemat., 377-382) или соматических способов (Staerz U.D. and Bevan M.J., 1986, PNAS 83, 1453-1457; Suresh M.R. et al., 1986, Method Enzymol., 121:210-228), а также, преимущественно, с помощью методик генной инженерии, которые делают возможной гетеродимеризацию и тем самым облегчают очистку антитела поиска (Merchand et al., 1998, Nature Biotech., 16:677-681).
Эти биспецифические антитела могут быть построены как целый IgG, биспецифический Fab'2, Раb'ПЭГ, димер антитела или биспецифический scFv, a также как четырехвалентное биспецифическое антитело, в котором присутствует два сайта связывания для каждого целевого антигена (Park et al., 2000, Mol. Immunol., 37(18): 1123-30) или его фрагментов, описанных выше.
В дополнение к экономическим преимуществам, учитывая, что продукция и введение биспецифического антитела дешевле, чем продукция двух специфических антител, применение таких биспецифических антител обладает преимуществом в плане снижения токсичности лечения. Действительно, применение биспецифического антитела позволяет уменьшить общее количество циркулирующих антител и, следовательно, возможную токсичность.
В предпочтительном воплощении изобретения биспецифическое антитело является двухвалентным или четырехвалентным антителом.
Наконец, данное изобретение относится к описанному выше антителу или его производным соединениям или функциональным фрагментам для применения их в качестве лекарственного средства.
Изобретение также относится к фармацевтической композиции, содержащей в качестве активного ингредиента соединение, включающей антитело изобретения или один из его производных соединений или функциональных фрагментов. Предпочтительно указанное антитело доставляется эксципиентом и/или фармацевтически приемлемым носителем.
Изобретение также относится к композиции, которая характеризуется тем, что она, в качестве комбинированного продукта для одновременного, раздельного или расширенного применения, содержит кроме того противоопухолевое антитело, отличное от антитела, направленного против CXCR4.
В соответствии с еще одним воплощением данное изобретение также относится к описанной выше фармацевтической композиции, которая содержит по меньшей мере второе противоопухолевое соединение, выбранное среди соединений, способных специфически ингибировать тирозинкиназную активность рецепторов, таких как IGF-IR, EGFR, HER2/neu, cMET, VEGFR или VEGF или любых других противоопухолевых соединений, известных специалистам в данной области.
Во втором предпочтительном аспекте изобретения указанное второе соединение может быть выбрано среди анти-EGFR, анти-IGF-IR, антиНЕR2/neu, антисМЕТ, VEGFR, VEGF и т.д., изолированных, или функциональных фрагментов или производных соединений, способных ингибировать пролиферативную и/или антиапоптотическую и/или ангиогенную и/или индукционную активность метастатической диссеминации, которую поддерживают указанные рецепторы.
Также подходящими для упоминания являются антиСD20-антитела, такие как ритуксимаб, ибритумомаб или тоситумомаб; антиСD33-антитела, такие как гемтузумаб или линтузумаб; антиСD22-антитела, такие как эпратузумаб; антиСD52-антитела, такие как алемтузумаб; антиЕрСАМ-антитела, такие как эдреколомаб, Ch 17-1A или IGN-101; антитела антиСТР21 или 16, такие как Xactin; антиДНК-Аg-антитела, такие как 131l-Cotara TNT-1; антиМuС1-антитела, такие как пемтумомаб или R1150; антиМUС18-антитела, такие как АВХ-МА1; антиСD3-антитела, такие как митумомаб; антиЕСА-антитела, такие как CeaVac или лабетузумаб; антиСА125-антитела, такие как OvaRex; антиHLA-DR-антитела, такие как аполизумаб; антиСТLА4-антитела, такие как MDX-010; антиРSМА-антитела, такие как MDX-070, 111In и 90Y-J591, 177Lu J591, J591-DM1; анти-Льюис Y-антитела (антитела к антигену Y Льюиса), такие как IGN311; антиангиогенные антитела, такие какА81405 и 90YmuBC1; антиТrail-R1-антитела, такие как МКА TRAIL R1 или МКА TRAIL R2.
Другое воплощение, дополняющее изобретение, состоит из композиции, описанной выше, которая в качестве комбинированного или конъюгированного продукта для одновременного, раздельного или расширенного применения содержит, помимо прочего, цитотоксический/цитостатический агент.
Под «одновременным применением» понимается введение обоих соединений композиции в единой фармацевтической форме.
Под «раздельным применением» понимается введение в одно и то же время обоих соединений композиции в различных фармацевтических формах.
Под «расширенным применением» понимается последовательное введение обоих соединений композиции, каждое в отдельной фармацевтической форме.
В целом, композиция в соответствии с изобретением значительно повышает эффективность лечения рака. Иными словами, терапевтический эффект антитела изобретения неожиданным образом усиливается при введении цитотоксического агента. Другое важное последующее преимущество, полученное с помощью композиции изобретения, касается возможности использования более низких эффективных доз активного ингредиента, что позволяет избежать или уменьшить риск проявления побочных эффектов, в частности влияний цитотоксического агента. Кроме того, эта композиция позволяет быстрее достигать ожидаемого терапевтического эффекта.
Под «терапевтическим противораковым агентом» или «цитотоксическим агентом» понимается вещество, которое при введении его пациенту лечит или предотвращает развитие рака у пациента. Неограничивающие примеры таких агентов включают «алкилирующие» агенты, антиметаболиты, противоопухолевые антибиотики, митотические ингибиторы, ингибиторы функционирования хроматина, антиангиогенные агенты, антиэстрогены, антиандрогены или иммуномодуляторы.
Такими агентами являются, например, агенты, упомянутые в издании VIDAL на странице, посвященной соединениям, применяемым в онкологии и гематологии, в колонке «Цитотоксические вещества»; цитотоксические соединения, цитируемые применительно к данному документу, приводятся здесь в качестве предпочтительных цитотоксических агентов.
Под «алкилирующим агентом» понимается любое вещество, которое может ковалентно связывать или алкилировать любую молекулу, предпочтительно нуклеиновую кислоту (например ДНК) в клетке. Примеры таких алкилирующих агентов включают азотные аналоги горчичного газа, такие как мехлоретамин, хлорамбуцил, мелфалан, хлоргидрат, пипоброман, преднимустин, гидрофосфат натрия или эстрамустин; оксазафосфорины, такие как циклофосфамид, алтретамин, трофосфамид, сульфофосфамид или ифосфамид; азиридины или этиленимины, такие как тиотепа, триэтиленамин или алтретамин; нитрозомочевины, такие как кармустин, стрептозоцин, фотемустин или ломустин; алкилсульфонаты, такие как бусульфан, треосульфан или импросульфан; триазены, такие как дакарбазин; или платиновые комплексы, такие как цисплатин, оксалиплатин или карбоплатин.
Под «антиметаболитом» понимается вещество, которое блокирует рост и/или клеточный метаболизм, вмешиваясь в определенные виды активности, как правило, в синтез ДНК. Примеры антиметаболитов включают метотрексат, 5-фторурацил, флоксуридин, 5-фтородезоксиуридин, капецитабин, цитарабин, флударабин, цитозинарабинозид, 6-меркаптопурин (6-МР), 6-тиогуанин (6-TG), хлородезоксиаденозин, 5-азацитидин, гемцитабин, кладрибин, дезоксикоформицин и пентостатин.
Под «противоопухолевым антибиотиком» понимается соединение, которое может предотвращать или ингибировать синтез ДНК, РНК и/или белков. Примеры таких противоопухолевых антибиотиков включают доксорубицин, даунорубицин, идарубицин, валрубицин, митоксантрон, дактиномицин, митрамицин, пликамицин, митомицин С, блеомицин и прокарбазин.
«Митотические ингибиторы» предотвращают нормальное развитие клеточного цикла и митоз. Как правило, ингибиторы микротрубочек, или «таксоиды», такие как паклитаксел и доцетаксел, способны ингибировать митоз.
Алкалоиды Винка, такие как винбластин, винкристин, виндезин и винорелбин, также способны ингибировать митоз.
К «ингибиторам хроматина» или «ингибиторам топоизомеразы» относятся вещества, которые ингибируют нормальное функционирование белков, моделирующих хроматин, таких как топоизомераза 1 или топоизомераза II. Примеры таких ингибиторов включают, для топоизомеразы I, камптотецин и его производные, такие как иринотекан или топотекан, и, для топоизомеразы II, этопозид, фосфат этопозида и тенипозид.
Под «антиангиогенным агентом» понимается любой лекарственный препарат, соединение, вещество или агент, который ингибирует рост кровеносных сосудов. Примеры антиангиогенных агентов включают, не ограничиваясь ими, разоксин, маримастат, батимастат, приномастат, таномастат, иломастат, CGS-27023А, галофугинон, COL-3, неовастат, BMS-275291, талидомид, CDC 501, DMXAA, L-651582, скваламин, эндостатин, SU 5416, SU6668, интерферон-альфа, EMD121974, интерлейкин-12, IM862, ангиостатин и витаксин.
Под «антиэстрогеном» или «антагонистом эстрогена» понимается любое вещество, которое уменьшает, противодействует или ингибирует действие эстрогена. Примерами таких агентов являются тамоксифен, торемифен, ралоксифен, дролоксифен, йодоксифен, анастрозол, летрозол и экземестан.
Под «антиандрогеном» или «антагонистом андрогена» понимается любое вещество, которое уменьшает, противодействует или ингибирует действие андрогена. Примеры антиандрогенов включают флутамид, нилутамид, бикалютамид, спироналоктон, ацетат ципротерона, финастерид и цимитидин.
Иммуномодуляторы представляют собой вещества, которые стимулируют иммунную систему. Примеры иммуномодуляторов включают интерферон, интерлейкины, такие как алдеслейкин, ОСТ-43, денилейкин дифтитокс и интерлейкин-2, факторы некроза опухоли, такие как тазонермин, или другие типы иммуномодуляторов, такие как лентинан, сизофиран, рохинимекс, пидотимод, пегадемаза, тимопентин, поли(1:С) (сополимер полиинозиновой и полицитидиловой кислот) или левамизол в комбинации с 5-фторурацилом.
За более подробной информацией специалист в данной области может обратиться к руководству, выпущенному под редакцией French Association of Therapeutic Chemistry Teachers и озаглавленному «Therapeutic chemistry, vol. 6, Antitumor drugs and perspectives in the treatment of cancer, TEC and DOC edition, 2003» (на французском языке).
В частном предпочтительном воплощении указанная композиция изобретения в качестве комбинированного продукта характеризуется тем, что указанный цитотоксический агент химически связан с указанным антителом для одновременного применения.
В частном предпочтительном воплощении указанная композиция характеризуется тем, что указанный цитотоксический/цитостатический агент выбран среди ингибиторов или стабилизаторов веретена деления, предпочтительно винорелбина и/или винфлунина и/или винкристина.
Для облегчения связывания указанного цитотоксического агента и антитела в соответствии с изобретением возможно введение между двумя соединениями, которые должны быть связаны, молекул-разделителей, например поли(алкилен)гликоля полиэтиленгликоля или аминокислот; или, в другом воплощении, использование активных производных указанных цитотоксических агентов, в которые были введены функциональные структуры, способные взаимодействовать с указанным антителом. Эти методики связывания хорошо известны специалистам в данной области и не будут более подробно рассматриваться в данном описании.
Другие ингибиторы EGFR включают, не ограничиваясь ими, моноклональные антитела С225 и антиЕСРР 22 (ImClone Systems Incorporated), ABX-EGF (Abgenix/Cell Genesys), EMD-7200 (Merck KgaA) или соединения ZD-1834, ZD-1838 и ZD-1839 (AstraZeneca), PKI-166 (Novartis), PKI-166/CGP-75166 (Novartis), PTK 787 (Novartis), CP 701 (Cephalon), флуномид (Pharmacia/Sugen), Cl-1033 (Warner Lambert Parke Davis), CI-1033/PD 183, 805 (Warner Lambert Parke Davis), CL-387, 785 (Wyeth-Ayerst), BBR-1611 (Boehringer Mannheim GMBH/Roche), Naamidine A (Bristol-board Myers Squibb), RC-3940-II (Pharmacia), BIBX-1382 (Boehringer Ingelheim), OLX-103 (Merck & Co), VRCTC-310 (Ventech Research), гибридный токсин EGF (Seragen Inc.), DAB-389 (Seragen/Lilgand), ZM-252808 (Imperial Cancer Research Fund), RG-50864 (INSERM), LFM-A12 (Parker Hughes Center Cancer), WHI-P97 (Parker Hughes Center Cancer), GW-282974 (Glaxo), KT-8391 (Kyowa Hakko) или «вакцину EGFR» (York Medical/Centro of Immunologia Molecular).
Другой аспект изобретения относится к композиции, которая характеризуется тем, что по меньшей мере одно из указанных антител или один из их производных соединений или функциональных фрагментов комбинирован или конъюгирован с клеточным токсином и/или радиоизотопом.
Предпочтительно указанный токсин или указанный радиоизотоп способен предотвращать рост или пролиферацию опухолевой клетки, особенно полностью инактивируя указанную опухолевую клетку.
Также предпочтительно указанный токсин представляет собой энтеробактериальный токсин, особенно экзотоксин А от Pseudomonas.
Радиоизотопы, предпочтительно конъюгированные с терапевтическими антителами, представляют собой радиоизотопы, которые излучают гамма-лучи, и предпочтительно иод131, иттрии90, золото199, палладии100, медь67, висмут217 и сурьму211. Радиоизотопы, которые излучают альфа- и бета-лучи, также могут быть использованы в терапии.
Под «токсином или радиоизотопом, комбинированным по меньшей мере с одним антителом изобретения или его функциональным фрагментом» понимают любое средство, позволяющее указанному токсину или указанному радиоизотопу связываться по меньшей мере с одним антителом, особенно путем ковалентного связывания двух соединений, с или без введения связывающей молекулы.
Примеры агентов, позволяющих связывать химическим (ковалентным), электростатическим или нековалентным образом все или часть элементов конъюгата, включают, в частности, бензохинон, карбодиимид и более предпочтительно EDC (1-этил-3-[3-диметил-аминопропил]-карбодиимида гидрохлорид), дималеимид, дитиобис-нитробензойную кислоту (DTNB), N-сукцинимидил-3-ацетилтиоацетат (SATA), связывающие агенты, имеющие одну или более фенилазидную группу, реагирующую с ультрафиолетовыми (УФ) лучами, наиболее предпочтительно N-[-4-(азидосалициламино)бутил]-3'-(2'-пиридилдитио)-пропионамид (APDP), N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) и 6-гидразино-никотинамид (HYNIC).
Другая форма связывания, особенно для радиоизотопов, может состоять в использовании бифункциональных ионных хелатирующих агентов.
Примеры таких хелатирующих агентов включают хелатирующие агенты, полученные из EDTA (этилендиаминтетрауксусной кислоты) или DTPA (диэтилентриаминпентауксусной кислоты), которые были разработаны для связывания металлов, в частности радиоактивных металлов, и иммуноглобулинов. Таким образом, DTPA и ее производные могут быть заменены различными группами в углеродной цепи в целях повышения стабильности и жесткости комплекса лиганд-металл (Krejcarek et al. (1977); Brechbiel et al. (1991); Gansow (1991); патент США 4831175).
Например, DTPA (диэтилентриаминпентауксусная кислота) и ее производные, которые широко используются в медицине и биологии в течение длительного времени либо в своей свободной форме, либо в комплексе с ионом металла, имеет замечательную особенность формировать стабильные хелаты с ионами металла, которые могут быть связаны с белками терапевтического или диагностического интереса, такими как антитела, для получения радиоиммуноконъюгатов для терапии рака (Meases et al., 1984; Gansow et al. 1990).
Также предпочтительно указанное по меньшей мере одно антитело изобретения, формирующее указанный конъюгат, выбирают среди его функциональных фрагментов, особенно фрагментов с удаленным Fc-компонентом, таких как scFv-фрагменты.
Данное изобретение также включает применение композиции для изготовления лекарственного средства, применяемого для профилактики или лечения рака.
Данное изобретение также относится к применению антитела или его производного соединения или функционального фрагмента, предпочтительно гуманизированного, и/или композиции в соответствии с изобретением для изготовления лекарственного средства, предназначенного для ингибирования роста опухолевых клеток. Главным образом, данное изобретение также относится к применению антитела или его производного соединения или функционального фрагмента, предпочтительно гуманизированного, и/или композиции для изготовления лекарственного средства для профилактики или лечения рака.
Предпочтительные раки, при которых можно проводить профилактику и/или лечение, включают рак предстательной железы, остеосаркому, рак легкого, рак молочной железы, рак эндометрия, рак толстой кишки, множественную миелому, рак яичников, рак поджелудочной железы или любой другой рак.
Изобретение также относится к применению антитела или его производного соединения или функционального фрагмента и/или композиции, описанной выше, для изготовления лекарственного препарата для модуляции СХСК4-активности в клетке.
Другой аспект данного изобретения относится к применению антитела в соответствии с описанием в способе диагностики, предпочтительно in vitro, заболеваний, связанных с уровнем экспрессии CXCR4. Предпочтительно указанными заболеваниями, связанными с белком CXCR4, в указанном диагностическом способе будут раки.
Таким образом, антитела изобретения или их производные соединения или функциональные фрагменты могут быть использованы в способы выявления и/или количественной оценки белка CXCR4 в биологическом образце in vitro, в частности, для диагностики заболеваний, связанных с ненормальной экспрессией этого белка, таких как рак, где указанный способ включает следующие этапы:
а) помещение биологического образца в контакт с антителом в соответствии с изобретением или его производным соединением или функциональным фрагментом;
б) демонстрация возможно сформированного комплекса антиген-антитело.
Таким образом, данное изобретение также включает наборы или приспособления для реализации описанного способа, включающие следующие элементы:
а) поликлональное или моноклональное антитело изобретения;
б) возможно, реагенты для составления среды, благоприятной для иммунологических реакций;
в) возможно, реагенты, которые выявляют комплексы антиген-антител, полученные в иммунологической реакции.
Преимущественно антитела или их функциональные фрагменты могут быть иммобилизованы на подложке, в частности на белковом чипе. Один из таких белковых чипов является объектом изобретения.
Преимущественно белковые чипы могут быть использованы в наборах или приспособлениях, необходимых для выявления и/или количественной оценки белка CXCR4 в биологическом образце.
Надо сказать, что термин «биологический образец» в данном документе относится к образцу, взятому у живого организма (в частности, кровь, ткань, орган или другие образцы, взятые у млекопитающих, особенно человека), или любому образцу, который может содержать один такой белок CXCR4 (такому как образцы клеток, трансформированные при необходимости).
Указанное антитело или его функциональный фрагмент может быть в форме иммуноконъюгата или меченого антитела для получения сигнала, который можно обнаружить и/или количественно оценить.
Меченые антитела изобретения или его функциональные фрагменты включают, например, конъюгаты антител (иммуноконъюгаты), которые могут быть комбинированы, например, с ферментами, такими как пероксидаза, щелочная фосфатаза, α-D-галактозидаза, глюкозооксидаза, глюкозоамилаза, карбоангидраза, ацетилхолинэстераза, лизоцим, малатдегидрогеназа или глюкозо- 6-фосфат-дегидрогеназа, или с молекулами, такими как биотин, дигоксигенин или 5-бромдезоксиуридин. Флуоресцентные метки также могут быть комбинированы с антителами изобретения или их функциональными фрагментами, особенно включая флуоресцеин и его производные, флуорохром, родамин и его производные, зеленый флуоресцентный белок (GFP), дансил, умбеллиферон и т.д. В таких конъюгатах антитела изобретения или их функциональные фрагменты могут быть получены с помощью способов, известных специалистам в данной области. Они могут быть связаны с ферментами или флуоресцентными метками непосредственно; посредством разделительной группы или связывающей группы, такой как полиальдегид, глутаральдегид, этилендиаминтетрауксусная кислота (EDTA) или диэтиленгликольтриаминпентауксусная кислота (DPTA); либо в присутствии связывающих агентов, таких как упомянутые выше для терапевтических конъюгатов. Конъюгаты, несущие флуоресцеиновые метки, могут быть получены путем реакции с изотиоцианатом.
Другие конъюгаты могут также включать хемилюминесцентные метки, такие как люминол и диоксетан, биолюминесцентные метки, такие как люцифераза и люциферин, либо радиоактивные метки, такие как йод123, йод125, йод126, йод133, бром77, технеций99m, индий111, индий113m, галий67, галий68, рутений95, рутений97, рутений103, рутений105, меркурий107, меркурий203, рений99m, рений101, рений105, скандий47, теллурий121m, теллурий122m, теллурий125m, тулий165, тулий167, тулий168, фтор18, иттрий199 и йод131. Способы, известные специалисту в данной области, существующие для связывания терапевтических радиоизотопов с антителами либо непосредственно, либо через хелатирующий агент, такой как упомянутые выше EDTA или DTPA, могут быть использованы для диагностических радиоизотопов. Также можно упомянуть метку с [I125Na с помощью хлорамина Т [Hunter W.M. and Greenwood F.C. (1962) Nature 194:495], либо также с технецием99m по описанию Crockford et al. (патент США 4424200), либо через DTPA, как описано у Hnatowich (патент США 4479930).
Изобретение также относится к применению антитела в соответствии с изобретением для получения лекарственного препарата для специфического воздействия соединения, которое является биологически активным по отношению к клеткам с экспрессией или сверхэкспрессией CXCR4.
В контексте данного описания «биологически активным соединением» является любое соединение, способное модулировать, в частности, ингибировать, клеточную активность, в частности рост, пролиферацию, транскрипцию и трансляцию гена.
Изобретение также относится к реагентам для диагностики in vivo, состоящим из антитела в соответствии с изобретением или его функционального фрагмента, предпочтительно меченого, в частности меченого радиоактивной меткой, и к их применению в медицинской визуализации, в частности для обнаружения рака, связанного с клеточной экспрессией или сверхэкспрессией CXCR4.
Изобретение относится также к композиции в виде комбинированного продукта или конъюгата антиСХСР4/токсин или радиоизотопа, в соответствии с изобретением, используемого в качестве лекарственного препарата.
Предпочтительно указанная композиция в качестве комбинированного продукта или указанного конъюгата в соответствии с изобретением будет смешана с эксципиентом и/или фармацевтическим носителем.
В данном описании под «фармацевтическим носителем» понимается соединение или комбинация соединений, входящих в фармацевтическую композицию, не провоцирующих вторичные реакции и позволяющих, например, облегчить введение активных соединений, увеличить время его жизни и/или его эффективность в организме, повысить его растворимость в растворе или улучшить его сохранность. Такие фармацевтические носители хорошо известны и будут адаптированы специалистом в данной области в зависимости от природы и способа введения выбранных активных соединений.
Предпочтительно такие соединения будут вводиться системным путем, в частности внутривенно, внутримышечно, внутрикожно, внутрибрюшинно, подкожно, или пероральным путем. Более предпочтительно композиция, содержащая антитело в соответствии с изобретением, будет вводиться в нескольких дозах через равные промежутки времени.
Способы введения, графики дозировок и оптимальные галеновые формы можно определить в соответствии с критериями, обычно принимаемыми при установлении лечения, адаптированного для пациентов, такими как, например, возраст или вес тела пациента, тяжесть его общего состояния, его толерантность к лечению и отмеченные побочные эффекты.
Таким образом, изобретение относится к применению антитела или одного из его функциональных фрагментов для изготовления лекарственного препарата для специфического воздействия соединения, которое является биологически активным по отношению к клеткам с экспрессией или сверхэкспрессией CXCR4.
Другие характеристики и преимущества изобретения появятся в последующем описании с примерами и фигурами, легенда к которым представлена ниже.
Легенда к графическим материалам
Фиг.1А и 1В показывают экспрессию соответственно CXCR4 и CXCR2 в раковых клетках с помощью анализа количественной ПЦР.
Фиг.2 показывает белковую экспрессию CXCR4 и CXCR2 в раковых клетках с помощью анализа FACS.
Фиг.3А и 3В показывают конкуренцию специфического [125I]SDF1-связывания немеченым SDF-1 (фиг.3А) и моноклональными антителами 414Н5 и 515Н7 (фиг.3В) на клеточных мембранах клеток СНО-К1, стабильно экспрессирующих человеческий CXCR4 дикого типа (Т: общее связывание; NS: неспецифическое связывание).
Фиг.4А и 4 В показывают модуляцию активации белка G моноклональными антителами 414Н5 (фиг.4А) и 515Н7 (фиг.4 В) путем мониторинга реакций [35S]GТРy3-связывания рецептора CXCR4 дикого типа, стабильно экспрессирующегося в клетках NIH-3T3.
Фиг.5 показывает модуляцию активации белка G моноклональными антителами 414Н5 (фиг.4А) и 515Н7 (фиг.4 В) путем мониторинга реакций [35S]GТРyS-связывания на человеческих опухолевых клетках HeLa, стимулированных SDF-1 (10 и 100 нМ).
Фиг.6A-6F показывают модуляцию ассоциации рецептора CXCR4 с различными взаимодействующими участниками SDF-1 и моноклональными антителами 414Н5 и 515Н7 с помощью подхода резонансного переноса энергии биолюминесценции (BRET) в клетках НЕК293. (Фиг.6А и 6В: CXCR4:CXCR4-гомодимеризация; фиг.6С и 6D:СХСР2:СХСР4-гетеродимеризация; и фиг.6Е и 6F: СХСР4-опосредованное привлечение (3-аррестина).
Фиг.7А и 7В показывают ингибирование форсколин-стимулированной продукции цАМФ SDF-1 и моноклональными антителами 414Н5 и 515Н7 в клетках NIH3T3, стабильно экспрессирующих рецептор CXCR4.
Фиг.8 показывает модуляцию активации белка G анти-СХСР4-моноклональными антителами 414Н5 и 515Н7 путем мониторинга реакций [35S]GTPYS-cвязывaния конститутивно активного мутантного рецептора Asn119Ser CXCR4, стабильно экспрессирующегося в клетках СНО-К1.
Фиг.9 иллюстрирует ингибирование SDF-1-индуцированной пролиферации клеток HeLa моноклональным антителом 414Н5 in vitro.
Фиг.10А и 10 В показывают ингибирование SDF-1-индуцированной миграции клеток U937 СХСР4-моноклональным антителом 414Н5 (фиг.10А) и моноклональным антителом 515Н7 (фиг.10 В) in vitro.
Фиг.11 показывает ингибирование роста ксенотрансплантатной опухоли MDA-MB-231 анти-СХСР4-моноклональными антителами 414Н5 (А) и 515Н7 (В) у мышей Nod/SCID.
Фиг.12 показывает активность анти-СХСР4-МКА 414Н5 на мышиных моделях выживания U937 Nod/Scid.
Фиг.13А-13С показывают ингибирование SDF-1-индуцированного высвобождения кальция анти-СХСР4-моноклональным антителом 515Н7 в клетках CHO-CXCR4 (фиг.13А) и MDA-MB-231 (фиг.13 В), раковых клетках U937 (фиг.13С).
Фиг.14 показывает ингибирование роста Т-клеточной ксенотрансплантатной опухоли KARPAS 299 у мышей Nod/ Scid моноклональным антителом 414Н5.
Фиг.15 показывает активность мышиного анти-СХСК4-МКА т515Н7 на мышиных моделях выживания U937 Nod/ Scid.
Фиг.16 показывает активность мышиного анти-СХСК4-МКА т515Н7 в ингибировании роста Т-клеточной ксенотрансплантатной опухоли KARPAS 299 у мышей Nod/ Scid.
Фиг.17 показывает конкуренцию специфического [125I]SDF1-связывания мышиными МКА m414Н5 и m515Н7 и химерными МКА с414Н5 и с515Н7 на клеточных мембранах клеток СНО-К1, стабильно экспрессирующих человеческий CXCR4 дикого типа (Т: общее связывание; NS: неспецифическое связывание).
Фиг.18 показывает модуляцию активации белка G мышиными МКА m414Н5 и m515Н7 и химерными МКА с414Н5 и с515Н7 путем реакций [35S]GTPyS-связывания рецептора CXCR4 дикого типа, стабильно экспрессирующегося в клетках NIH-3T3, стимулированных SDF-1 (10 нМ).
Фиг.19 показывает модуляцию активации белка G анти-СХСР4 мышиными МКА т414Н5 и т515Н7 и химерными МКА с414Н5 и с515Н7 путем мониторинга реакций [35S]GTPYS-cвязывaния на человеческих опухолевых клетках HeLa, стимулированных SDF-1 (10 нМ).
Фиг.20А-20С показывают модуляцию ассоциации рецептора CXCR4 с различными взаимодействующими участниками SDF-1 и моноклональными антителами m414Н5, с414Н5, m515Н7 и с515Н7 с помощью подхода резонансного переноса энергии биолюминесценции (BRET) в клетках НЕК293. (Фиг.20А: СХСР4:СХСР4-гомодимеризация; фиг.20 В: СХСР2:СХСР4-гетеродимеризация; и фиг.20С: СХСР4-опосредованное привлечение β-аррестина).
Фиг.21А и 21В показывают ингибирование SDF-1-индуцированного высвобождения кальция в клетках CHO-CXCR4 (фиг.21А) и клетках U937 (фиг.21 В).
Фиг.22А и 22 В показывают ингибирование SDF-1-индуцированной миграции клеток U937 СХСР4-моноклональными антителами m414Н5 и с414Н5 (фиг.22А) m515Н7 и c515Н7 (фиг.22 В) in vitro.
Фиг.23 показывает активность анти-СХСР4 химерных МКА с414Н5 и с515Н7 на мышиных моделях выживания U937Nod/ Scid.
ПРИМЕРЫ
Пример 1: Экспрессия CXCR4 и CXCR2 в раковых клетках
- количественный ПЦР-анализ (qPCR):
Для того, чтобы количественно оценить относительную экспрессию CXCR4 и CXCR2 в различных раковых клеточных линиях, используется ОТ-ПЦР в режиме реального времени(real time RT-PCR).
Образцы РНК выделяли из различных клеточных линий с помощью протоколов RNeasy Mini или Midi (Qiagen Corporation, Франция). Затем РНК-образцы проверяли с помощью автоматизированной системы электрофореза Experion (BIO-RAD Corporation, Франция), было показано хорошее качество/целостность. Один мкг каждого образца РНК переводили в кДНК-матрицу с помощью набора для синтеза кДНК iScript cDNA Syntesis (BIO-RAD Corporation, Франция). Уровни кДНК количественно оценивали с помощью количественной ПЦР либо с зондом TaqMan для CXCR2, либо с SYBERGreen для CXCR4. Сравниваемые образцы требовали нормализации, поэтому был введен внутренний референс RPLO. Зонды TaqMan (используемые для CXCR2) несли 5'-FAM-репортерную метку и 3'-ТАМРА-группу-гаситель. Фермент ПЦР активировали нагреванием в течение 2 мин при 50°С и 10 мин при 95°С. Использовали двухэтапную процедуру, 15 сек при 95°С и 1 мин при 62°С в течение 40 или 45 циклов с ПЦР-смесью, содержащей 5 мкл кДНК-матрицы (разведение 1/20), 1 х qPCR Mastermix (TaqMan Universal PCR Master Mix, Applied Biosystems corporation, Бранчбург, Нью-Джерси, США), 50-900 нМ каждого праймера и 50-100 нМ зонда в общем объеме 50 мкл. Все реакции проводили с помощью инструмента iCycler (BIO-RAD Corporation). Количественная ПЦР позволяла определить пороговый цикл (Ct, cycle threshold). Чем больше значений Ct являются низкими, тем больше тестируемых генов экспрессируется. Праймеры и зонды для большого человеческого рибосомального белка, РО, были следующими:
прямой праймер, 5'-GAAACTCTGCATTCTCGCTTCCTG-3 '(SEQ ID №32);
обратный праймер, 5'-AGGACTCGTTTGTACCCGTTGA-3 '(SEQ ID №33);
зонд, 5'-(FAM)-TGCAGATTGGCTACCCAACTGTTGCA-(TAMRA)-3' (SEQ ID №34).
Праймеры для человеческого CXCR4 (хемокинового рецептора-4) были следующими:
прямой праймер, 5'-CTCCTTCATCCTCCTGGAAATC-3'(SEQ ID №35);
обратный праймер, 5'-CCAAGGAAAGCATAGAGGATGG-3'(SEQ ID №36). Праймеры и зонд для человеческого CXCR2 (хемокинового рецептора-2) были следующими:
прямой праймер, 5'-GTGGTCATTATCTATGCCCTGG-3' (SEQ ID №37);
обратный праймер, 5'-CGACCCTGCTGTATAAGATGAC-3' (SEQ ID №38);
зонд, 5'-(FAM)-TATTCCTGCTGAGCCTGCTGGGAAA-(TAMRA)-3' (SEQ ID №39).
В нашем сравнительном исследовании экспрессию двух генов [тестируемого гена (CXCR4 или CXCR2) и RPLO] количественно оценивали в двух различных образцах: тестируемой клеточной линии и референсной клеточной линии.
Референсная клеточная линия соответствовала клеточной линии, содержащей самую низкую экспрессию оцениваемого гена. Сравнительный расчет экспрессии генов был сделан с использованием следующей формулы:
Относительная экспрессия гена=(1+Егена)-ΔCt(1)/(1+ERPL0)-ΔCt(2)
Егена=эффективность ПЦР при использовании праймеров/зонда для
оцениваемого гена
EERP0=эффективность ПЦР при использовании праймеров/зонда для RPL0
Ct=пороговый цикл
ΔCt(1)=Сtгена (тестируемая клеточная линия) - Сtгена (референсная клеточная линия)
ΔCt(2)=CtRPL0 (тестируемая клеточная линия) - CtppLo (референсная клеточная линия).
Для каждой серии ПЦР рассчитывали относительное количественное значение гена, и раковые клеточные линии классифицировали по группам с учетом уровня их экспрессии от максимальной до отрицательной. Все данные представлены на фиг.1А и 1В. Все тестируемые раковые клеточные линии экспрессировали CXCR4 (фиг.1А) и CXCR2, за исключением DU145 и U-87MG в отношении CXCR2 (фиг.1В).
- FACS анализ:
Раковые клеточные линии MDA-MB-231, РСЗ и U937 повергали пермеабилизации, а затем инкубировали либо с 10 мкг/мл анти-СХСР4-моноклональных антител [44717 (R&D Systems) в сравнении с изотипическим контролем lgG2b (SIGMA), либо с 10 мкг/мл анти-СХСР2-моноклональных антител (анти-П-СХСК2, клон 48311, R&D Systems, MKA 331) в сравнении с изотипическим контролем lgG2a. Затем клетки промывали 1% BSA/PBS/0,01% NaN3. Далее к клеткам добавляли Alexa-меченые вторичные антитела и инкубировали при 4°С в течение 20 мин. Затем клетки снова промывали два раза. После второго промывания проводили FACS-анализ. Результаты этих исследований связывания представлены на фиг.2. Таким образом, опухолевые клетки, такие как MDA-MB-231, РС3 и U937, экспрессируют как CXCR4-, так и СХСР2-белки.
Пример 2: Создание моноклональных антител (MKA) против человеческого CXCR4
Для создания моноклональных антител к CXCR4 мышей BALB/c иммунизировали рекомбинантными клетками NIH3T3-CXCR4 и/или пептидами, соответствующими внеклеточному М-концу и петлям CXCR4. Мышей 6-16-недельного возраста при первой иммунизации иммунизировали один раз антигеном в полном адъюванте Фрейнда подкожно (s.c.), а затем от 2 до 6 раз антигеном в неполном адъюванте Фрейнда подкожно. Иммунный ответ контролировали в крови из ретроорбитального синуса. Скрининг сывороток проводили с помощью ИФА (как описано ниже), и мышей с более высоким титром анти-СХСР4-антител использовали для слияния. Мышей бустировали внутривенно антигеном за два дня до умерщвления и забора селезенки.
-ИФА
Чтобы выбрать мышей, продуцирующих анти-СХСР4-антитела, сыворотки от иммунизированных мышей проверяли с помощью ИФА. Вкратце, микротитровальные планшеты покрывали очищенным [1-41] N-концевым пептидом, конъюгированным с BSA (бычьим сывороточным альбумином) в количестве 5 мкг эквивалентного пептида/мл, 100 мкл/лунка, инкубировали при 4°С в течение ночи, а затем блокировали 250 мкл/лунка 0,5% желатином в PBS (фосфатно-солевом буфере). В каждую лунку добавляли разведения плазмы от CXCR4-иммунизированных мышей и инкубировали 2 часа при 37°С. Платы промывали PBS, а затем инкубировали с козлиным противомышиным lgG-антителом, конъюгированным с HRP (Jackson Laboratories), в течение 1 часа при 37°С.После промывания в платы вносили субстрат ТМВ (тетраметилбензоидин), реакцию останавливали через 5 минут путем добавления 100 мкл/лунку 1М H2S04. Мышей, у которых развивались высокие титры анти-СХСК4-антител, использовали для создания антител.
- создание гибридом, продуцирующих МКА к CXCR4
Мышиные спленоциты, выделенные из мышей BALB/c, у которых развивались самые высокие титры анти-СХСК4-антител, сливали с помощью ПЭГ с мышиной миеломной клеточной линией Sр2/0. Клетки высевали в количестве примерно 1×105/лунка в микротитровальные платы, затем инкубировали две недели в селективной среде, содержащей ультра культуральную среду +2 мМ L-глутамин +1 мМ пируват натрия +1х HAT. Скрининг лунок проводили с помощью ИФА с анти-СХСК4-моноклональными антителами IgG. Затем гибридомы, секретирующие антитела, субклонировали по меньшей мере дважды путем предельного разведения, культивировали in vitro для создания антитела для дальнейшего анализа.
Пример 3: Характеризация с помощью FACS-анализа специфичности связывания анти-СХСР4-МКА 414Н5 и 515Н7 и распознавание раковых клеточных линий
В этом эксперименте специфическое связывание анти-СХСР4-МКА 414Н5 и 515Н7 с человеческим CXCR4 определяли с помощью FACS-анализа.
Трансфицированные клетки NIH3T3, NIH3T3-hCXCR4, раковые клеточные линии MDA-MB-231, Hela и U937 инкубировали с 10 мкг/мл моноклонального антитела 414Н5 и 515Н7. Затем клетки промывали 1% BSA/PBS/0,01% NaN3. Затем к клеткам добавляли Alexa-меченые вторичные антитела и инкубировали при 4°С в течение 20 мин. Затем клетки снова промывали два раза. После второго промывания проводили FACS-анализ. Результаты этих исследований связывания представлены в таб. 4, которая показывает [Средняя интенсивность флуоресценции (MFI), полученная при FACS], что анти-СХСК4-МКА 414Н5 и 515Н7 связывались специфично с человеческой трансфицированной клеточной линией CXCR4-NIH3T3, в то время на родительских клетках NIH3T3 распознавания не было. Эти МКА также были способны распознавать человеческие раковые клеточные линии, например клетки рака молочной железы MDA-MB-231, промиелоцитарные раковые клетки U937 и клетки рака шейки матки Hela.
Анти-СХСР4-МКА 414Н5 и 515Н7 распознавали трансфектант NIH3T3-hCXCR4, в то время как родительские клетки дикого типа NIH3T3 не распознавались. МКА 414Н5 и 515Н7 также были способны распознавать раковые клеточные линии.
Пример 4: Конкурентное связывание анти-СХСР4 МКА 414Н5 и 515Н7 с [125I]SDF-1 на мембранах СНО-К1, стабильно экспрессирующих человеческий рецептор CXCR4
Этот анализ позволяет оценить способность МКА 414Н5 и 515Н7 конкурировать за связывание меченого радиоактивной меткой [125I]SDF-I с человеческим рецептором CXCR4 либо на ортостерических, либо на аллостерических сайтах связывания.
Клетки СНО-К1, стабильно и конститутивно экспрессирующие человеческий рецептор CXCR4, были получены при трансфекции наивных клеток СНО-К1 (АТСС CCL-61) вектором для экспрессии в клетках млекопитающих, несущим всю кодирующую последовательность человеческого рецептора CXCR4 (RefSeq NM_003467). Клетки размножались в полной культуральной среде [DMEM-Хэма F12 с добавлением 5% эмбриональной телячьей сыворотки (FCS) и 500 мкг/мл генетицина]. Эксперименты со связыванием радиолиганда проводили на клеточных мембранах, полученных при механическом разрушении клеток CHO/CXCR4 в литическом буфере [Hepes 20 мМ, рН 7,4, NaCl 150 мМ] и последующем центрифугировании (10000 д, 15 мин). [125I]SDF-1-связывание (специфическая активность: 1500 Ки/ммоль) проводили с использованием методики SPA (сцинтилляционный анализ сближения - GE Healthcare). Вкратце, клеточные мембраны (30 мкг/лунка) инкубировали в связывающем буфере [Hepes 20 мМ, рН 7,4, CaCl2 1 мМ, МgСl2 5 мМ, NaCI 150 мМ, BSA 1%] вместе с оцениваемым соединением (SDF-1 или МКА), радиолигандом (1 нМ) и, наконец, частицами SPA-WGA-PVT (7,3 мг/лунка). Равновесие связывания достигали через 1 ч при 25°С. После центрифугирования [1000 g в течение 10 мин] измеряли уровень радиоактивности в сцинтилляционном счетчике (TopCount, Perkin Elmer).
Неспецифическое связывание оценивали в присутствии 10 мкМ немеченного SDF-1.
Немеченый SDF-1 дозозависимо ингибировал [125I]SDF-1-связывание со значением PKi (IC50=концентрации лиганда, дающая 50% ингибирование специфического [125I]SDF-1-связывания) 7,75±0,27 нМ (n=4) (фиг.3А). В тех же экспериментальных условиях наши анти-СХСР4-МКА (100 нМ) эффективно конкурировали за [125I]SDF-1-связывание со следующим порядком ранжирования эффективности конкуренции (% ингибирования [125I]SDF-I): 515H7 (64±3%) 414Н5 (43±4%) (фиг.3В).
Пример 5: Модуляция [35S]GTРyS-связывания на клеточных мембранах, экспрессирующих рецептор CXCR4 дикого типа, анти-СХСР4 моноклональными антителами 414Н5 и 515H7
Этот функциональный анализ позволяет контролировать активацию G-белка через человеческий рецептор CXCR4 дикого типа и его модуляцию CXCR4-лигандами и МКА 414Н5 и 515H7.
Клетки NIH-3T3, стабильно и конститутивно экспрессирующие рецептор CXCR4 дикого типа, были получены, как описано в примере выше для клеток СНО-К1. Клетки HeLa (человеческая карцинома шейки матки) размножались в полной культуральной среде [ЕМЕМ с добавлением 10% FCS, 1% L-глутамина, 2 µ бикарбоната натрия]. [35S]GTPyS-связывание выполняли на клеточных мембранах, полученных при механическом разрушении в литическом буфере [Hepes 20 мМ, рН 7,4, NaCl 150 мМ] и последующем центрифугировании (10000 д, 15 мин). Встраивание и выявление [35S]GTPyS (специфическая активность: 1000 Ки/ммоль) проводили с использованием методики SРА (сцинтилляционный анализ сближения - GE Healthcare). Вкратце, клеточные мембраны (10 мкг/лунка) инкубировали в связывающем буфере [Hepes 20 мМ, GDP 3 мкМ, МgСl2 10 мМ, NaCl 100 мМ, EDTA 1 мМ, рН=7,4] вместе с оцениваемым соединением (3DF-1 или представляющее интерес МКА), [35S]GTPyS (0,2-0,4 нМ) и, наконец, частицами 3PA-WGA-PVT (7,3 мг/лунка). Равновесие связывания достигали через 1 ч при 25°С. После центрифугирования [1000 g в течение 10 мин] измеряли уровень радиоактивности в сцинтилляционном счетчике (TopCount, Perkin Elmer). Силу антагониста рассчитывали с применением уравнения Cheng-Prussof:
КB=[конц. антагониста]/{(ЕС50'/ЕС50)-1}, где EC50 и EC50' соответственно являются силой 3DF-1 в отсутствие и в присутствии МКА.
SDF-1 индуцировал дозозависимое увеличение [35S]GTPyS-связывания в результате активации белка G рецептором CXCR4. Максимальная стимуляция [35S]GTPyS-связывания представляет соответственно 167% и 320% по сравнению с базальным [35S]GTPyS-связыванием для мембран клеток HeLa и NIH3T3/CXCR4. Сила 3DF-1 была одинакова для обеих клеточных линий и соответствовала 41,3±9,7 нМ (фиг.4А-4Б). В этих экспериментальных условиях сила антагониста МКА 414Н5 и 515Н7, определенная в клетках NIH3T3/CXCR4, составляла 51 нМ и 15 нМ, соответственно. Аналогичная эффективность антагониста наблюдалась для клеток HeLa (фиг.5).
Пример 6: Ассоциация CXCR4 с различными участниками взаимодействия: гомо- и гетеродимеризация, привлечение β-аррестина с помощью подхода резонансного переноса энергии биолюминесценции (ВРЕТ) и влияние МКА 414Н5 и 515Н7 на эти димеры
Этот функциональный анализ позволяет оценить конформационные изменения, вызванные 3DF-1 и/или связыванием МКА 414Н5 и 515Н7 с рецептором CXCR4 на уровне формирования гомодимера CXCR4 и гетеродимера CXCR2/CXCR4, а также привлечение сигнального белка Р-аррестина-2.
Экспрессионные векторы для каждого из исследуемых участников взаимодействия были построены как гибридные белки с соответствующей меткой (люцифераза Renilla reniformis, Rluc, и желтый флуоресцентный белок, YFP) с помощью обычных методик молекулярной биологии. За два дня до проведения BRET-экспериментов клетки НЕК293 транзиентно трансфицировали экспрессионными векторами, кодирующими соответствующих участников BRET: [CXCR4/Rluc+CXCR4/YFP] для изучения гомодимеризации CXCR4, [CXCR4/Rluc+CXCR2:YFP] для изучения гетеродимеризации CXCR4 и CXCR2 и [CXCR4/Rluc+β-arr2:YFP] для изучения СХСР4-опосредованного привлечения Р-аррестина-2. На следующий день клетки распределяли в полилизиновые предварительно покрытые (антителами) белые 96-луночные микротитровальные планшеты в полной культуральной среде [DMEM с добавлением 10% FB3]. Клетки вначале культивировали при 37°С с 5% СO2, с тем чтобы клетки прикрепились к планшету. Затем клетки оставляли в 200 мкл DMEM/лунка в течение ночи. Непосредственно перед BRET-экспериментом DMEM удаляли и клетки быстро промывали РВЗ. Затем клетки инкубировали в РВЗ в присутствии или в отсутствие антитела, 10 мин при 37°С, а затем добавляли 5 мкМ коэлентеразина Н с или без 300 нМ 3DF-1 в конечном объеме 50 мкл. После инкубации в течение еще 10 минут при 37°С начинали захват излучаемого света с помощью многофункционального анализатора Mithras LB940 при 485 нм и 530 нм (Berthold) (1 с/длина волны/лунка повторяется 15 раз при комнатной температуре).
Расчет отношения BRET проводили, как описано выше (Angers et al., 2000): [(эмиссия530 нм) - (эмиссия485 нм)×Cf] / эмиссия485 нм), где Cf=(эмиссия530 нм) / (эмиссия485 нм) для клеток, экспрессирующих только гибридный белок Rluc в тех же экспериментальных условиях. Упрощение этого уравнения показывает, что соотношение BRET соответствует отношению 530/485 нм, полученному, когда присутствуют два участника, скорректированному отношением 530/485 нм, полученным в тех же экспериментальных условиях, когда в анализе присутствует только участник, слитый с Rluc. Для удобочитаемости результаты выражаются в единицах миллиВРЕТ (mBU); mBU соответствует отношению BRET, умноженному на 1000.
SDF1 (300 нМ) увеличивал примерно на 20% сигнал BRET в результате пространственной близости адаптерных и акцепторных белков, слитых с рецептором CXCR4, что, вероятно, указывает на формирование гомодимеров CXCR4/CXCR4 или конформационных изменений уже существующих димеров (фиг.6А и 6В). Интересно, что SDF1 (300 нМ) уменьшал примерно на 24% сигнал BRET в результате пространственной близости адаптерных и акцепторных белков, слитых с CXCR2 и CXCR4, что, вероятно, указывает на формирование гетеродимеров CXCR2/CXCR4 или конформационных изменений уже существующих димеров (фиг.6С и 6D). В этом последнем случае SDF-1 -активированная конформация CXCR4/CXCR2 представляется менее благоприятной для переноса энергии BRET. В обоих случаях МКА 414Н5 и 515Н7 были способны модулировать SDF-1-индуцированные конформационные изменения гомодимеров CXCR4 (63% ингибирование SDF-1-индуцированного увеличения BRET для 414Н5 и 69% ингибирование SDF-1-индуцированного увеличения BRET для 515Н7, фиг.6А и 6 В, соответственно), а также формирование гетеродимеров CXCR2/CXCR4 (50% ингибирование SDF-1 -индуцированного увеличения BRET для 414Н5 и 90% ингибирование SDF-1-индуцированного увеличения BRET для 515Н7, фиг.6С и 6D, соответственно). МКА 414Н5 и 515Н7 сами по себе также способны модулировать пространственную близость CXCR4/CXCR4 и CXCR2/CXCR4 соответственно, что указывает на влияние МКА 414Н5 и 515Н7 на конформацию и гомодимера CXCR4/CXCR4, и гетеродимера CXCR2/CXCR4 (фиг.6А, 6В, 6С и 6D).
Активация CXCR4 с помощью SDF-1 (300 нМ) привела к мощному привлечению внутриклеточных сигнальных молекул (3-аррестина, о чем говорит повышение BRET-сигнала на 233% (фиг.6Е и 6F). Это привлечение частично ингибировалось МКА 414Н5 и 515Н7 (примерно 20% ингибирование для 414Н5 и 95% для 515Н7, фиг.6Е и 6F, соответственно), показывая влияние МКА 414Н5 и 515Н7 на передачу сигнала.
Пример 7: СХСР4-опосредованное ингибирование продукции цАМФ
Этот функциональный анализ предназначен для мониторинга передачи сигнала от рецептора CXCR4 на уровне аденилатциклаз через ингибиторные белки Gi/o.
Процедуру cAMP LANCE (Perkin Elmer) применяли в соответствии с описанием производителя. Вкратце, клетки NIH3T3, стабильно и конститутивно экспрессирующие рецептор CXCR4 дикого типа, получали и размножали, как описано выше. Клетки собирали с помощью свободного от трипсина агента Versene и ресуспендировали в концентрации 106 клеток/мл в растворе, содержащем AlexaFluor-связанное анти-цАМФ МКА (разведение 1/100) и соединение (форсколин, SDF-1 и/или МКА 414Н5 и 515Н7). После инкубации при комнатной температуре в течение 30 мин добавляли смесь для обнаружения, содержащую комплексы европий-стрептавидин (разведение 1/125) и биотин-цАМФ (разведение 1/125). После инкубации при комнатной температуре в течение 1 часа полученный FRET-сигнал измеряли с помощью многофункционального анализатора Mithras LB940 (Berthold). Данные выражали либо в виде условных люминесцентных значений, либо в виде относительной стимуляции в сравнении с SDF-1-ответом после вычитании влияния FK.
Форсколин (FK) дозозависимо стимулировал продукцию цАМФ с силой примерно 0,3 мкМ в клетках NIH3T3/CXCR4 (фиг.7А). В присутствии SDF-1 внутриклеточный уровень цАМФ снижался в результате активации ингибиторного белка Gi/o рецептором CXCR4. Сила SDF-1 составляла 5,0±3,1 нМ (фиг.7А). МКА 414Н5 и 515Н7 эффективно ингибировали форсколин-стимулированное влияние SDF-1 (100 нМ) более чем на 60% для 414Н5 и более чем на 80% для 515Н7 (фиг.7В).
Пример 8: Модуляция [35S]GTPyS-связывания на клеточных мембранах, конститутивно экспрессирующих мутантный Asn191Ser СХСР4-рецептор, моноклональными антителами 414Н5 и 515Н7
Этот функциональный анализ позволяет контролировать активацию G-белка через конститутивно активный мутантный (САМ) Asn119Ser СХСР4-рецептор (см. Zhang et al., 2002). Это чувствительный анализ позволяет различать лиганды CXCR4 на основании их внутренней активности (частичный агонист, молчащий антагонист или обратный агонист). Как описано ранее Zhang и коллегами, CXCR4-лиганды, такие как AMD3100 или Т140, вели себя соответственно как частичный агонист и обратный агонист САМ-рецептора CXCR4. Идентификация молчащего антагониста может быть затруднена, так как этот класс молекул должен демонстрировать похожую аффинность и к активному, и к неактивному состоянию CXCR4 (Wurchetal., 1999).
Введение мутации Asn119Ser в кодирующую последовательность рецептора CXCR4 выполняли с помощью обычных методик молекулярной биологии (набор для сайт-направленного мутагенеза QuickChange, Stratagene, США). Клетки СНО-К1, стабильно и конститутивно экспрессирующие САМ-рецептор CXCR4, получали, как описано в примере выше. [35S]GTPyS-связывание выполняли на клеточных мембранах, полученных при механических разрушении в литическом буфере [Hepes 20 мМ, рН 7,4, 150 мМ NaCl] и дальнейшем центрифугировании (10000 g, 15 мин). Встраивание [35S]GTPyS (специфическая активность: 1000 Ки/ммоль) проводили с использованием методики SPA (сцинтилляционный анализ сближения - GE Healthcare). Вкратце, клеточные мембраны (10 мкг/лунка) инкубировали в связывающем буфере [Hepes 20 мМ, GDP 3 мкМ, МдСЬ 10 мМ, NaCI 100 мМ, EDTA 1 мМ, рН=7,4] вместе с оцениваемым соединением (SDF-1 или МКА), [35S]GTPyS (0,2-0,4 нМ) и, наконец, частицами SPA-WGA-PVT (7,3 мг/лунка). Реакцию связывания проводили в течение 1 ч при 25°С. После центрифугирования [1000 g в течение 10 мин] измеряли уровень радиоактивности в сцинтилляционном счетчике (TopCount, Perkin Elmer).
SDF-1 (100 нМ) стимулировал [35S]GTPYS-cвязывaниe на 130%. Обратный агонист Т140 ингибировал как базальное (-17%), так и SDF-1-стимулированное (-159%) [35S]GTPYS-cвязывaниe. Напротив, МКА 414Н5 и 515Н7 проявляли себя как молчащие антагонисты САМ CXCR4, не изменяя базальное [35S]GTPYS-cвязывaниe (фиг.8), но ингибируя SDF-1-индуцированное [35S]GTPYS-cвязывaниe (фиг.8).
Пример 9: Ингибирование SDF-1-индуцированной пролиферации клеток Hela CXCR4-MKA 414Н5 in vitro
Клетки HeLa из АТСС стандартно культивировали в среде ЕМЕМ (Lonza Corporation, Вервье, Бельгия), 10% FCS (Sigma Corporation. Сент-Луис, США), 1% L- глутамин (Invitrogen Corporation, Шотландия, Великобритания), 2% бикарбонат натрия 7,5% раствор (Invitrogen Corporation, Шотландия, Великобритания). Клетки разделяли за 3 дня до анализа пролиферации, чтобы они были конфлюэнтными.
- SDF-1-индуцированная пролиферация клеток Неlа
Клетки HeLa помещали в 96-луночные планшеты для культивирования с плотностью 1×104 клеток/лунка в 200 мкл бессывороточной среды (среда ЕМЕМ плюс 1% L-глутамин, 2% натрия бикарбоната 7,5% раствор). Через двадцать четыре часа после посева к клеткам HeLa добавляли соответствующие разведения SDF-1. Через 76 часов культивирования в общей сложности на клетки воздействовали импульсом 0,25 мкКи [3H]тимидина (Amersham Biosciences AB, Упсала, Швеция) в течение 16 часов. Значение включенного в ДНК [3H]тимидина количественно оценивали с помощью жидкостного сцинтилляционного счета.
Результаты выражали как индекс пролиферации=[среднее срm клеток+SDF-1/среднее срm клеток - SDF-1].
Клетки HeLa инкубировали с SDF-1 (от 0 до 1000 нг/мл). SDF-1 увеличивал пролиферацию клеток HeLa in vitro в 1,5-2 раза. Концентрация SDF-1 для получения наиболее высокого и воспроизводимого индекса пролиферации составляла 200 нг/мл (25 нМ).
- ингибирование SDF-1-индуцированной пролиферации клеток in vitro СХСР4-моноклональным антителом 414Н5
Клетки HeLa помещали в 96-луночные планшеты для культивирования с плотностью 1×104 клеток/лунка в 200 мкл бессывороточной среды (среда ЕМЕМ плюс 1% L-глутамин. 2% натрия бикарбоната 7,5% раствор). Через двадцать четыре часа после посева к клеткам HeLa в трех экземплярах добавляли соответствующие разведения анти-СХСК4 МКА 414Н5, разводя среду в отсутствие или в присутствии SDF-1 в конечной концентрации 200 нг/мл (25 нМ). Через 76 часов культивирования в общей сложности на клетки воздействовали импульсом 0,25 мкКи [3H]тимидина (Amersham Biosciences AB, Упсала, Швеция) в течение 16 часов. Значение включенного в ДНК [3H]тимидина количественно оценивали с помощью жидкостного сцинтилляционного счета.
Результаты выражали как фракцию, находящуюся под влиянием (Fa), рассчитанную по формуле:
Fa=[1-[среднее срm клеток, инкубированных с МКА+SDF-1 / среднее срт клеток, инкубированных с разведенными средами+SDF-1)]×100.
Было охарактеризовано влияние in vitro анти-СХСР4-МКА 414Н5 на SDF-1-индуцированную пролиферацию клеток HeLa. Клетки HeLa инкубировали либо с МКА 414Н5, либо с контролем с или без SDF-1 (200 нг/мл). SDF-1 увеличивал рост клеток HeLa in vitro (в 1,5-2 раза). Дозозависимую кривую для МКА 414Н5 получали путем обработки клеток серийными двукратными разведениями МКА от 0 до 1500 нМ через 24 ч после посева клеток. Так как пролиферацию клеток оценивали в течение 76 часов после посева, каждое тестируемое условие соответствовало времени экспозиции 48 ч для МКА или контроля. Результаты выражали как фракцию, находящуюся под влиянием (Fa), рассчитанную по описанной выше формуле. Результаты (представлены на фиг.9) показали, что CXCR4-MKA 414Н5 ингибирует SDF-1-индуцированную пролиферацию клеток HeLa in vitro.
Пример 10: Влияние анти-СХСК4-МКА 414Н5 и 515Н7 на SDF-1-индуцированную миграцию клеток U937
Для оценки ингибирующего влияния анти-СХСК4-моноклональных антител 414Н5 и 515Н7 на процесс миграции, 100000 клеток U-937 в среде RPMI 1640 с добавлением 2% FCS высевали в верхнюю камеру миграционных камер (24-луночные планшеты с размером пор 8 мкм) в присутствии или в отсутствие SDF-1 в нижней части лунок с или без МКА 414Н5 и 515Н7 в верхней камере. В этом тесте мышиные lgG2a и lgG2b вводили в качестве изотипических контролей. Через два часа после посева подсчитывали мигрировавшие клетки. Результаты, представленные на фиг.10А для 414Н5 и 10 В для 515Н7 показали, что, как и ожидалось, SDF-1 мог вызывать значительное увеличение миграции клеток U-937. Влияния не наблюдалось, когда клетки инкубировали с изотипическим контролем lgG2. Напротив, для клеток, инкубированных с МКА 414Н5 и 515Н7, наблюдалось значительное и воспроизводимое уменьшение SDF-1-индуцированной миграции клеток U937: 50% с МКА 414Н5 и более 80% с МКА 515Н7.
Пример 11: Ингибирование роста ксенотрансплантатной опухоли MDA-МВ-231 у мышей Nod/Scid моноклональным анти-СХСР4-антителом 414Н5
Цель этих экспериментов заключалась в оценке способности анти-СХСК4-МКА 414Н5 и 515Н7 ингибировать рост ксенотрансплантата MDB-MB-231 у мышей Nod/Scid.
Клетки MDA-MB-231 из ЕСАСС стандартно культивировали в среде DMEM (Invitrogen Corporation, Шотландия, Великобритания), 10% FCS (Sigma, Сент-Луис, Мэриленд, США). Клетки разделяли за 48 часов до введения трансплантата, так что они были в экспоненциальной фазе роста. Десять миллионов клеток MDA-MB-231 прививали в PBS 7-недельным мышам Nod/Scid (Charles River, Франция).
Через пять дней после имплантации опухоли измеряли (34 mm^v^o мм3) и животных разделяли на группы по 6 мышей с сопоставимым размером опухоли. Мышам вводили внутрибрюшинно (i.p.) нагрузочную дозу 2 мг/мышь МКА 414Н5 и МКА 515Н7, соответственно.
Затем мышам инъекционно вводили 1 мг/доза/мышь МКА 414Н5 два раза в неделю или 0,5 мг/доза/мышь МКА 515Н7 три раза в неделю. Группа PBS в этом эксперименте была введена в качестве контрольной группы. Объем опухоли измеряли два раза в неделю и рассчитывали по формуле: π/6 × длина × ширина × высота. Статистический анализ проводили для каждого измерения с помощью критерия Манна-Уитни.
В этих экспериментах во время лечения смертности не наблюдалось. По сравнению с группой PBS наблюдалось значительное ингибирование роста опухоли между 7 и 39 днями (р≤0,002) для 415Н5 МКА 1 мг/доза или 515Н7 0,5 мг/доза, а средний объем опухоли через 5 недель лечения уменьшился на 82% и 50% по сравнению с PBS для МКА 415Н5 и 515Н7, соответственно (фиг.11А и 11В).
Пример 12: Активность анти-СХСР4-МКА 414Н5 на мышиной модели выживаемости U937
Клетки U937 из АТСС культивировали в среде RPMI 1640, 10% FCS, 1% L-глутамин. Клетки разделяли за два дня до приживления, так что они находились в экспоненциальной фазе роста. Десять миллионов клеток U937 вводили внутрибрюшинно самкам мышей NOD/Scid. Через два дня после имплантации мышам подкожно вводили нагрузочную дозу 2 мг/мышь МКА 414Н5, а затем два раза в неделю по 1 мг антитела/мышь. Контрольные мыши получали инъекции PBS, и как было указано в предыдущих исследованиях, не наблюдалось никакой разницы в выживаемости между мышами, которым вводили PBS, и мышами, которым вводили изотипический контроль мышиный IgG. Мышиную выживаемость контролировали каждый день.
Результаты описаны на фиг.12, показывающей, что у мышей, которым вводили МКА 414Н5, наблюдалось резкое и значительное увеличение продолжительности жизни с Т/С% примерно 343.
Пример 13: СХСР4-рецептор-опосредованная мобилизация внутриклеточных депо кальция
Этот функциональный анализ был предназначен для наблюдения за СХСК4-рецепторной передачей сигнала через стимуляцию пути фосфолипазы С, вызывающей высвобождение кальция из внутриклеточных депо в эндоплазматическом ретикулуме.
Клетки СНО-К1, стабильно и конститутивно экспрессирующие CXCR4-рецептор дикого типа, были получены, как описано в примере выше. Клетки MDA-МВ-231 (человеческая аденокарцинома молочной железы) и U937 (человеческая лимфома) размножались в полной культуральной среде, соответственно [DMEM с добавлением 10% FCS] и [RPMI 1640 с добавлением 10% FCS, 20 мМ HEPES, 1% заменимых аминокислот, 1% пирувата натрия, 1% L-глутамина, 4,5 г/л глюкозы]. Все типы клеток высевали в черные 96-луночные микротитровальные планшеты с плотностью 100000 клеток/лунка в соответствующей культуральной среде. Клетки оставляли на ночь перед проведением экспериментов. Клетки загружали с флуоресцентной кальциевой меткой (Fluo-4 No Wash, Invitrogen, США) в загрузочный буфер [HBSS 1х, HEPES 20 мМ, кислота пробенецид 25 мМ] на 30 мин при 37°С, затем на 30 мин при 25°С. Стимуляцию SDF-1 выполняли путем прямого введения в каждую лунку. Для антагонистических экспериментов 10 мкл раствора МКА добавляли непосредственно в загрузочный буфер по меньшей мере за 10 мин до SDF-1. Кинетические измерения флуоресценции выполняли на многорежимном планшетном флуоресцентном анализаторе Mithras LB940 (Berthold), используя следующие параметры: возбуждение при 485 нм, излучение при 535 нм, энергия возбуждения 10000 условных единиц. Флуоресценцию в каждой лунке регистрировали в течение 0,1 секунды каждую секунду и в течение 20 сек до введения SDF-1 (базальный сигнал). Затем вводили 20 мкл SDF-1 и записывали данные в течение 2 мин. Каждое экспериментальное условие повторяли дважды. Значения в каждой лунке вначале корректировали путем вычитания базальной флуоресценции и флуоресценции, испускаемой контрольными лунками без клеток. Относительные данные выражали в процентах от максимальной стимуляции, полученной SDF-1 (100 нМ).
SDF1 (100 нМ) индуцировал быстрый и сильный выход внутриклеточного кальция в рекомбинантных CHO/CXCR4, в то время как в наивных клетках СНО-К1 сигнала флуоресценции обнаружено не было. Максимальная интенсивность достигала >160% по сравнению с базальной флуоресценцией и наблюдалась примерно 30 сек при стимуляции SDF-1; подобные кинетические кривые наблюдались и для MDA-MB-231, и для U-937 (фиг.13А, 13 В, 13С), хотя максимальная интенсивность флуоресценции SDF-1 (100 нМ) была ниже (130-140% по сравнению с базальной). Антитело 515Н7 (133 нМ) давало сильное и почти полное ингибирование SDF-1 (100 нМ)-индуцированного кальциевого сигнала во всех трех исследуемых клеточных линиях.
Пример 14: Ингибирование роста Т-клеточной ксенотрансплантатной опухоли KARPAS 299 у мышей Nod/Scid моноклональным анти-СХСК4 антителом 414Н5
Цель этого эксперимента заключалась в оценке способности анти-СХСР4-МКА 414Н5 ингибировать рост ксенотрансплантата KARPAS 299 у мышей Nod/Scid.
Клетки KARPAS 299 из ЕСАСС стандартно культивировали в среде RPMI, 1% L-Glu и 10% FCS (Sigma, Сент-Луис, Мэриленд, США). Клетки разделяли за 48 часов до введения трансплантата, так что они находились в экспоненциальной фазе роста. Пять миллионов клеток KARPAS 299 в PBS прививали 7-недельным мышам Nod/Scid (Charles River, Франция). Через пять дней после имплантации опухоли измеряли (32 мм3<V3<49 мм3) и животных разделяли на группы по 6 мышей с сопоставимыми размерами опухолей. Мышам вводили внутрибрюшинно нагрузочную дозу 2 мг/мышь МКА 414Н5.
Затем мышам инъекционно вводили 1 мг/доза/мышь МКА 414Н5 два раза в неделю. Группа PBS была введена в этот эксперимент в качестве контрольной группы. Объем опухоли измеряли два раза в неделю и рассчитывали по формуле: π/6 × длина × ширина × высота. Статистический анализ проводили для каждого измерения с помощью критерия Манна-Уитни.
В этом эксперименте во время лечения смертности не наблюдалось. По сравнению с группой PBS наблюдалось значительное ингибирование роста опухоли между 7 и 33 днями (р≤0,002) для 415Н5 МКА 1 мг/доза, а средний объем опухоли через 5 недель лечения уменьшился на 73% для МКА 415Н5 по сравнению с PBS (фиг.14).
Пример 15: Активность анти-СХСК4-МКА 515Н7 на мышиной модели выживаемости U937
Клетки U937 из АТСС культивировали в среде RPMI 1640, 10% FCS, 1% L-глутамин. Клетки разделяли за два дня до приживления, так что они находились в экспоненциальной фазе роста. Десять миллионов клеток U937 вводили внутрибрюшинно самкам мышей NOD/Scid. Через два дня после имплантации мышам подкожно вводили нагрузочную дозу 2 мг/мышь МКА 515Н7, а затем два раза в неделю 1 мг антитела/мышь. Контрольные мыши получали инъекции PBS, и как было указано в предыдущих исследованиях, не наблюдалось никакой разницы в выживаемости между мышами, которым вводили PBS, и мышами, которым вводили изотипический контроль мышиный IgG. Мышиную выживаемость контролировали каждый день.
Результаты описаны на фиг.15, показывающей, что у мышей, которым вводили МКА 515Н7, наблюдалось резкое и значительное увеличение продолжительности жизни с Т/С% примерно 280 для МКА 515Н7 (фиг.15).
Пример 16: Ингибирование роста Т-клеточной ксенотрансплантатной опухоли KARPAS 299 у мышей Nod/Scid моноклональным анти-СХСРД-антителом 515Н7
Цель этого эксперимента заключалась в оценке способности анти-СХСР4-МКА 515Н7 ингибировать рост ксенотрансплантата KARPAS 299 у мышей Nod/Scid.
Клетки KARPAS 299 из ЕСАСС стандартно культивировали в среде RPMI, 1% L-Glu и 10% FCS (Sigma, Сент-Луис, Мэриленд, США). Клетки разделяли за 48 часов до введения трансплантата, так что они находились в экспоненциальной фазе роста. Пять миллионов клеток KARPAS 299 в PBS прививали 7-недельным мышам Nod/Scid (Charles River, Франция). Через пять дней после имплантации опухоли измеряли (32 мм3<V3<49 мм3) и животных разделяли на группы по 6 мышей с сопоставимыми размерами опухолей. Мышам вводили внутрибрюшинно нагрузочную дозу 2 мг/мышь МКА 515Н7.
Затем мышам инъекционно вводили 1 мг/доза/мышь МКА 515Н7 два раза в неделю. Группа PBS была введена в этот эксперимент в качестве контрольной группы. Объем опухоли измеряли два раза в неделю и рассчитывали по формуле: π/6 × длина × ширина × высота. Статистический анализ проводили для каждого измерения с помощью критерия Манна-Уитни.
В этих экспериментах во время лечения смертности не наблюдалось. По сравнению с группой PBS наблюдалось значительное ингибирование роста опухоли между 7 и 33 днями (р≤0,002) для МКА 515Н7 1 мг/доза, а средний объем опухоли через 5 недель лечения уменьшился на 63% для МКА 515Н7 по сравнению с PBS (фиг.16).
Пример 17: Продукция анти-СХСР4 химерных МКА с414Н5 и с515Н7
Были разработаны химерные формы мышиных МКА 414Н5 и 515Н7: они соответствуют вариабельным доменам легкой и тяжелой цепи мышиных антител интереса, генетически слитым с человеческим константаными доменами Скаппа и lgG1. Все рекомбинантные МКА были получены путем транзиентной трансфекции с помощью системы HEK293/EBNA с экспрессионным вектором рСЕР4 (Invitrogen, США).
Целые нуклеотидные последовательности, соответствующие вариабельным доменам легкой и тяжелой цепей МКА 414Н5 и 515Н7, синтезировали путем глобального синтеза генов (Genecust, Люксембург). Их субклонировали в вектор рСЕР4 (Invitrogen, США), несущий целую кодирующую последовательность константного домена либо легкой [Скаппа], либо тяжелой [СН1-шарнир-СН2-СН3] цепи человеческого иммуноглобулина lgG1. Все этапы клонирования выполняли в соответствии с обычными методиками молекулярной биологии, описанными в Laboratory manual (Sambrook and Russel, 2001), или в соответствии с инструкциями производителя. Каждую генетическую конструкцию полностью подтверждали путем секвенирования с использованием набора для секвенирования Big Dye terminator cycle sequencing kit (Applied Biosystems, США) и анализировали с помощью анализатора 3100 Genetic Analyzer (Applied Biosystems, США).
Адаптированные к суспензии клетки НЕК293 EBNA (Invitrogen, США) выращивали стандартным способом в 250 мл колбах в 50 мл бессывороточной среды Excel 293 (SAFC Biosciences) с добавлением 6 мМ глутамина на орбитальном шейкере (скорость вращения 110 об/мин). Транзиентную трансфекцию проводили на 2х106 клеток/мл с помощью линейного 25 кДа полиэтиленимина (PEI) (Polysciences), приготовленного в воде в конечной концентрации 1 мг/мл, и плазмидной ДНК (конечная концентрация 1,25 мкг/мл с плазмидным отношением тяжелой и легкой цепи 1:1). Через 4 часа после трансфекции культуры разводили одним объемом свежей питательной среды для достижения конечной плотности клеток 106/мл. Процесс культивации осуществляли на основе жизнеспособности клеток и продукции МКА. Как правило, культуры поддерживали в течение 4-5 дней. МКА очищали с помощью обычной хроматографии на смоле Protein A resin (GE Healthcare, США). Все разные МКА были получены на уровне, подходящем для функциональной оценки. Уровни производительности, как правило, находились в диапазоне между 6 и 15 мг/л очищенных МКА.
Пример 18: Характеристика с помощью FACS-анализа специфичности связывания анти-СХСР4 химерных МКА с414Н5 и с515Н7 и распознавание раковой клеточной линии
В этом эксперименте с помощью FACS-анализа проверяли специфическое связывание анти-СХСР4 химерных МКА с414Н5 и с515Н7.
Трансфицированные клетки NIH3T3, NIH3T3-hCXCR4 и раковую клеточную линию MDA-MB-231 линии инкубировали с 10 мкг/мл моноклонального антитела с414Н5 и с515Н7. Затем клетки промывали 1% BSA/PBS/0,01% NaN3. Далее к клеткам добавляли Alexa-меченые вторичные антитела и инкубировали при 4°С в течение 20 мин. Затем клетки снова промывали два раза. После второго промывания проводили FACS-анализ. Результаты этих исследований связывания представлены в следующей таблице 5, которая показывает [средняя интенсивность флуоресценции (MFI), полученная в FACS], что анти-СХСК4 химерные МКА с414Н5 и с515Н7 специфически связываются с человеческой трансфицированной клеточной линией CXCR4-NIH3T3, а также распознают человеческие раковые клеточные линии, например клетки рака молочной железы MDA-MB-231.
Пример 19: Конкурентное связывание анти-СХСК4 МКА т414Н5 и т515Н7 и химерных МКА с414Н5 и с515Н7 с [125I]SDF-1 на мембранах СНО-К1, стабильно экспрессирующих человеческий рецептор CXCR4
Этот анализ позволяет оценить способность МКА т414Н5 и т515Н7 и химерных МКА с414Н5 и с515Н7 конкурировать за связывание меченого радиоактивной меткой [^IJSDF-I с человеческим рецептором CXCR4 либо на ортостерических, либо на аллостерических сайтах связывания.
Клетки СНО-К1, стабильно и конститутивно экспрессирующие человеческий рецептор CXCR4, получали путем трансфекции наивных клеток СНО-К1 (АТСС CCL-61) вектором для экспрессии в клетках млекопитающих, несущим всю кодирующую последовательность человеческого рецептора CXCR4 (RefSeq NM_003467). Клетки размножались в полной культуральной среде [DMEM-Хэма F12 с добавлением 5% эмбриональной телячьей сыворотки (FCS) и 500 мкг/мл генетицина]. Эксперименты со связыванием радиолиганда проводили на клеточных мембранах, полученных при механическом разрушении клеток CHO/CXCR4 в литическом буфере [Hepes 20 мМ, рН 7,4, NaCl150 мМ] и последующем центрифугировании (10000 д, 15 мин). [125I]SDF-1-связывание (специфическая активность: 1500 Ки/ммоль) проводили с использованием методики SPA (сцинтилляционный анализ сближения - GE Healthcare). Вкратце, клеточные мембраны (30 мкг/лунка) инкубировали в связывающем буфере [Hepes 20 мМ, рН 7,4, CaCl2 1 мМ, МgСl2 5 мМ, NaCl 150 мМ, BSA 1%] вместе с оцениваемым соединением (SDF-1 или МКА), радиолигандом (1 нМ) и, наконец, частицами SPA-WGA-PVT (7,3 мг/лунка). Равновесие связывания достигали через 1 ч при 25°С.После центрифугирования [1000 g в течение 10 мин] измеряли уровень радиоактивности на сцинтилляционном счетчике (TopCount, Perkin Elmer). Неспецифическое связывание оценивали в присутствии 10 мкМ немеченного SDF-1.
AHTH-CXCR4-MKA (100 нМ) эффективно конкурировали за [125I]SDF-1-связывание со следующим порядком ранжирования эффективности конкуренции (% ингибирования [125I]SDF-1):m515Н7 (62±10%), с515Н7 (55±4%), m414Н5 (30±5%) и с414Н5 (21±10%) (фиг.17).
Пример 20: Модуляция [35S]GTPyS-связывания на клеточных мембранах, экспрессирующих рецептор CXCR4 дикого типа, анти-СХСК4 МКА m414Н5 и m515Н7 и химерными МКА с414Н5 и с515Н7.
Этот функциональный анализ позволяет контролировать активацию G-белка через человеческий рецептор CXCR4 дикого типа и его модуляцию анти-СХСР4 мышиными МКА m414Н5 и m515Н7 и химерными МКА с414Н5 и с515Н7.
Клетки NIH-3T3, стабильно и конститутивно экспрессирующие рецептор CXCR4 дикого типа, получали, как описано в примере выше для клеток СНО-К1. Клетки HeLa (человеческая карцинома шейки матки) размножались в полной культуральной среде [ЕМЕМ с добавлением 10% FCS, 1% L-глутамина, 2 мкМ бикарбоната натрия]. [35S]GTPyS-связывание выполняли на клеточных мембранах, полученных при механическом разрушении в литическом буфере [Hepes 20 мМ, рН 7,4, NaCl 150 мМ] и последующем центрифугировании (10000 д, 15 мин). Встраивание и выявление [35S]GTPyS (специфическая активность: 1000 Ки/ммоль) проводили с использованием методики SPA (сцинтилляционный анализ сближения - GE Healthcare). Вкратце, клеточные мембраны (10 мкг/лунка) инкубировали в связывающем буфере [Hepes 20 мМ, GDP 3 мкМ, MgCl2 10 мМ, NaCl 100 мМ, EDTA 1 мМ, рН=7,4] вместе с оцениваемым соединением (SDF-1 и МКА интереса), [35S]GTPyS (0,2-0,4 нМ) и, наконец, частицами SPA-WGA-PVT (7,3 мг/лунка). Равновесие связывания достигали через 1 ч при 25°С. После центрифугирования [1000 g в течение 10 мин] измеряли уровень радиоактивности на сцинтилляционном счетчике (TopCount, Perkin Elmer). IC5o рассчитывали для каждого МКА.
В этих экспериментальных условиях IC50 МКА m414Н5, с414Н5, m515Н7 и с515Н7, определенная в клетках NIH3T3/CXCR4, составляла 1,6 нМ, 1,1 нМ, 1,9 нМ и 1,5 нМ, соответственно (фиг.18). IC50 МКА m414Н5, с414Н5, m515Н7 и с515Н7, определяемая с помощью клеток HeLa в тех же экспериментальных условиях, составляла 0,5 нМ, 0,3 нМ, 0,2 нМ и 0,6 нМ, соответственно (фиг.19).
Пример 21: Ассоциация CXCR4 с различными участниками взаимодействия: гомо- и гетеродимеризация, привлечение р-аррестина с помощью подхода резонансного переноса энергии биолюминесценции (ВРЕТ) и влияние мышиных МКА m414Н5 и m515Н7 и химерных с414Н5 и с515Н7 на эти димеры
Этот функциональный анализ позволяет оценить конформационные изменения, вызванные SDF-1 и/или связыванием мышиных МКА т414Н5 и т515Н7 и химерных с414Н5 и с515Н7 с рецептором CXCR4, на уровне формирования гомодимера CXCR4 и гетеродимера CXCR2/CXCR4, а также привлечение сигнального белка β-аррестина-2.
Экспрессионные векторы для каждого из исследуемых участников взаимодействия были построены как гибридные белки с соответствующей меткой (люцифераза Renilla reniformis, Rluc, и желтый флуоресцентный белок, YFP) с помощью обычных методик молекулярной биологии. За два дня до проведения экспериментов BRET клетки НЕК293 транзиентно трансфицировали экспрессионными векторами, кодирующими соответствующих участников BRET: [CXCR4/Rluc+CXCR4/YFP] для изучения гомодимеризации CXCR4, [CXCR4-Rluc+CXCR2-YFP] для изучения гетеродимеризации CXCR4 и CXCR2 и [CXCR4-Rluc+β-arr2-YFP] для изучения СХСР4-опосредованного привлечения (3-аррестина-2. На следующий день клетки распределяли в полилизиновые предварительно покрытые (антителами) белые 96-луночные микротитровальные планшеты в полной культуральной среде [DMEM с добавлением 10% FBS]. Клетки вначале культивировали при 37°С с 5% С02, с тем чтобы клетки прикрепились к планшету. Затем клетки оставляли в 200 мкл DMEM/лунка в течение ночи. Непосредственно перед BRET-экспериментом DMEM удаляли и клетки быстро промывали PBS. Затем клетки инкубировали в PBS в присутствии или в отсутствие антитела 15 мин при 37°С, а затем добавляли 5 мкМ коэлентеразина Н с или без 100 нМ SDF-1 в конечном объеме 50 мкл. После инкубации в течение 5 минут при 37°С, а также инкубации в течение 20 минут при комнатной температуре только для гомо- и гетеродимеров, начинали захват излучаемого света с помощью многофункционального анализатора Mithras LB940 (Berthold) при 485 нм и 530 нм (1 с/длина волны/лунка повторяется 15 раз при комнатной температуре).
Расчет отношения BRET проводили, как описано выше (Angers et al., 2000): [(эмиссия530 нм) - (эмиссия485 нм) × Cf] / (эмиссия485 нм), где Cf=(эмиссия530 нм) / (эмиссия485 нм) для клеток, экспрессирующих только гибридный белок Rluc в тех же экспериментальных условиях. Упрощение этого уравнения показывает, что соотношение BRET соответствует отношению 530/485 нм, полученному, когда присутствуют два участника BRET, скорректированному отношением 530/485 нм, полученным в тех же экспериментальных условиях, когда в анализе присутствует только участник, слитый с Rluc. Для удобочитаемости результаты выражаются в единицах миллиВРЕТ (mbu); mbu соответствует отношению BRET, умноженному на 1000.
SDF1 (300 нМ) увеличивал примерно на 10% сигнал BRET, полученный из пространственной близости донорных и акцепторных белков, слитых с рецептором CXCR4, что, вероятно, указывает на формирование гомодимеров CXCR4/CXCR4 или конформационные изменения уже существующих димеров (фиг.20А).
Интересно, что SDF1 (100 нМ) уменьшал примерно на 17% сигнал BRET полученный из пространственной близости донорных и акцепторных белков, слитых с CXCR4 и CXCR2, что, вероятно, указывает на формирование гетеродимеров CXCR2/CXCR4 или конформационные изменения уже существующих димеров (фиг.20 В). В этом последнем случае SDF-1 -активированная конформация CXCR4/CXCR2 представляется менее благоприятной для переноса энергии BRET. В обоих случаях МКА т414Н5, с414Н5, т515Н7 и с515Н7 были способны модулировать SDF-1-индуцированные конформационные изменения гомодимеров CXCR4 (75% ингибирование SDF-1-индуцированного увеличения BRET для с414Н5 и 96% ингибирование SDF-1-индуцированного увеличения BRET для с515Н7, фиг.20А), а также формирование гетеродимеров CXCR2/CXCR4 (77% ингибирование SDF-1-индуцированного уменьшения BRET для с414Н5 и 98% ингибирование SDF-1-индуцированного уменьшения BRET для с515Н7, фиг.20 В). МКА т414Н5, с414Н5, т515Н7 и с515Н7 сами по себе также были способны модулировать пространственную близость CXCR4/CXCR4 и CXCR2/CXCR4 соответственно, что указывает на влияние этих МКА на конформацию и гомодимера CXCR4/CXCR4, и гетеродимера CXCR2/CXCR4 (фиг.20А и 20В).
Активация CXCR4 с помощью SDF-1 (100 нМ) привела к мощному привлечению внутриклеточных сигнальных молекул β-аррестина, о чем говорит повышение BRET-сигнала на 400% (фиг.20С). Это привлечение частично ингибируется МКА с414Н5 и с515Н7 (примерно 63% ингибирование для с414Н5 и 93% для с515Н7, фиг.20С), показывая влияние этих МКА на передачу сигнала.
Пример 22. СХСР4-рецептор-опосредованная мобилизация внутриклеточных депо кальция
Этот функциональный анализ был предназначен для наблюдения за CXCR4-рецепторной передачей сигнала через стимуляцию пути фосфолипазы С, вызывающей высвобождение кальция из внутриклеточных депо в эндоплазматическом ретикулуме.
Клетки СНО-К1, стабильно и конститутивно экспрессирующие рецептор CXCR4 дикого типа, были получены, как описано в примере выше. Клетки U-937 (человеческая лимфома) размножались в полной культуральной среде, соответственно [DMEM с добавлением 10% FCS] и [RPMI 1640 с добавлением 10% FCS, 20 мМ HEPES, 1% заменимых аминокислот, 1% пирувата натрия, 1% L-глутамина, 4,5 г/л глюкозы]. Все типы клеток высевали в черные 96-луночные микротитровальные планшеты с плотностью 100000 клеток/лунка в соответствующей культуральной среде. Клетки оставляли на ночь перед проведением экспериментов. Клетки загружали с флуоресцентной кальциевой меткой (Fluo-4 No Wash, Invitrogen, США) в загрузочный буфер [HBSS 1х, HEPES 20 мМ, кислота пробенецид 25 мМ] на 30 мин при 37°С, затем на 30 мин при 25°С. Стимуляцию SDF-1 выполняли путем прямого введения в каждую лунку. Для антагонистических экспериментов 10 мкл раствора МКА добавляли непосредственно в загрузочный буфер по меньшей мере за 10 мин до SDF-1. Кинетические измерения флуоресценции выполняли на многорежимном планшетном анализаторе Mithras LB940 (Berthold), используя следующие параметры: возбуждение при 485 нм, излучение при 535 нм, энергия возбуждения 10000 условных единиц. Флуоресценцию в каждой лунке регистрировали в течение 0,1 секунды каждую секунду и в течение 20 сек до введения SDF-1 (базальный сигнал). Затем вводили 20 мкл SDF-1 и записывали данные в течение 2 мин. Каждое экспериментальное условие повторяли дважды. Значения в каждой лунке вначале корректировали путем вычитания базальной флуоресценции и флуоресценции, испускаемой контрольными лунками без клеток. Относительные данные выражали в процентах от максимальной стимуляции, полученной с SDF-1 (100 нМ).
SDF1 (100 нМ) индуцировал быстрый и сильный выход внутриклеточного кальция в рекомбинантных CHO/CXCR4, в то время как в наивных клетках СНО-К1 сигнала флуоресценции обнаружено не было. Максимальная интенсивность достигала более 140% по сравнению с базальной флуоресценцией и наблюдалась примерно 40 сек при стимуляции SDF-1; подобные кинетические кривые наблюдались для клеток U-937 (фиг.21А, 21В). Химерное антитело с515Н7 (133 нМ) давало сильное и почти полное ингибирование SDF-1 (100 нМ)-индуцированного кальциевого сигнала во обеих исследуемых клеточных линиях.
Пример 23. Влияние анти-СХСК4 мышиных МКА т414Н5 и т515Н7 и химерных МКА с414Н5 и с515Н7 на SDF-1-индуцированную миграцию клеток U937
Для оценки ингибирующего влияния анти-СХСР4-МКА т414Н5, т515Н7, с414Н5 и с515Н7 на процесс миграции 100000 клеток U-937 в среде RPMI 1640 с добавлением 2% FCS высевали в верхнюю камеру миграционных камер (24-луночные планшеты с размером пор 8 мкм) в присутствии или в отсутствие SDF-1 в нижней части лунки и с или без МКА с414Н5, т414Н5, с515Н7 и m515Н7 в верхней камере. В этом тесте мышиные lgG2a и lgG2b вводили в качестве изотипического контроля. Через два часа после посева подсчитывали мигрировавшие клетки. Результаты, представленные на фиг.22А (для с414Н5 против т414Н5) и 22 В (для с515Н7 против m515Н7) показали, что, как и ожидалось, SDF-1 мог вызывать значительное увеличение миграции клеток U-937. Влияния не наблюдалось, когда клетки инкубировали с изотипическим контролем lgG2. Напротив, для клеток, инкубированных с МКА с414Н5, m414Н5, с515Н7 и m515Н7, наблюдалось значительное и воспроизводимое уменьшение SDF-1-индуцированной миграции клеток U937: примерно 50% с МКА с414Н5 и m414Н5 и более 80% с МКА с515Н7 и m515Н7.
Пример 24. активность анти-СХСР4 химерных МКА с414Н5 и с515Н7 на мышиной модели выживаемости U937
Клетки U937 из АТСС культивировали в среде RPMI 1640, 10% FCS, 1% L-глутамин. Клетки разделяли за два дня до приживления, так что они находились в экспоненциальной фазе роста. Десять миллионов клеток U937 вводили внутрибрюшинно самкам мышей NOD/Scid. Через два дня после имплантации мышам подкожно вводили нагрузочную дозу 2 мг МКА с414Н5 или с515Н7/мышь, а затем два раза в неделю 1 мг антитела/мышь. Контрольные мыши получали инъекции PBS, и как было указано в предыдущих исследованиях, не наблюдалось никакой разницы в выживаемости между мышами, которым вводили PBS, и мышами, которым вводили изотипический контроль мышиный IgG. Мышиную выживаемость контролировали каждый день.
Результаты, описанные на фиг.23, показывают, что у мышей, которым вводили МКА с414Н5 и с515Н7, наблюдалось резкое и значительное увеличение продолжительности жизни с Т/С% примерно 210 и 180 для с414Н5 и с515Н7, соответственно.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CXCR4 ДЛЯ ЛЕЧЕНИЯ РАКА | 2011 |
|
RU2595394C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА I-3859 ДЛЯ ВЫЯВЛЕНИЯ И ДИАГНОСТИКИ РАКА | 2012 |
|
RU2636032C2 |
| АНТИТЕЛА ПРОТИВ CXCR4 ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИИ ВИЧ | 2010 |
|
RU2573902C2 |
| НОВОЕ АНТИТЕЛО К CXCR4 И ЕГО ПРИМЕНЕНИЕ ДЛЯ ВЫЯВЛЕНИЯ И ДИАГНОСТИКИ РАКА | 2012 |
|
RU2636345C2 |
| СПОСОБ МОДУЛИРОВАНИЯ АНТАГОНИСТИЧЕСКОЙ АКТИВНОСТИ МОНОКЛОНАЛЬНОГО АНТИТЕЛА | 2009 |
|
RU2575600C2 |
| АНТИТЕЛО ПРОТИВ cMet И ЕГО ПРИМЕНЕНИЕ ДЛЯ ОПРЕДЕЛЕНИЯ И ДИАГНОСТИКИ РАКА | 2010 |
|
RU2582265C2 |
| АНТИТЕЛО ПРОТИВ c-Met | 2009 |
|
RU2560257C2 |
| НОВЫЕ АНТИТЕЛА, ИНГИБИРУЮЩИЕ ДИМЕРИЗАЦИЮ С-МЕТ, И ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2552161C2 |
| ANTI-CXCR4 АНТИТЕЛА И КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВО | 2014 |
|
RU2675516C2 |
| НОВОЕ АНТИТЕЛО ПРОТИВ с-МЕТ | 2011 |
|
RU2607377C2 |






















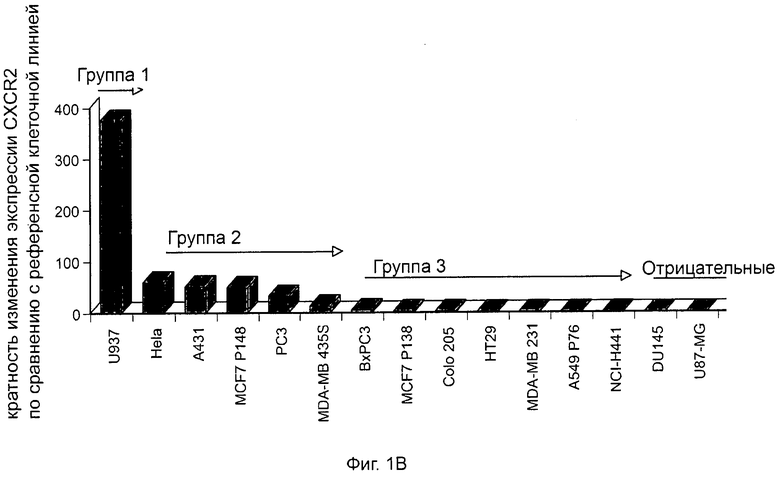
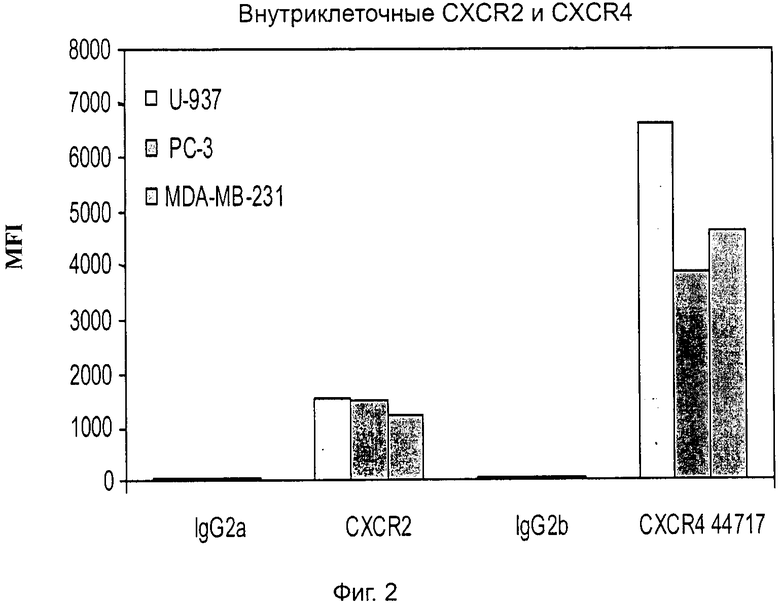
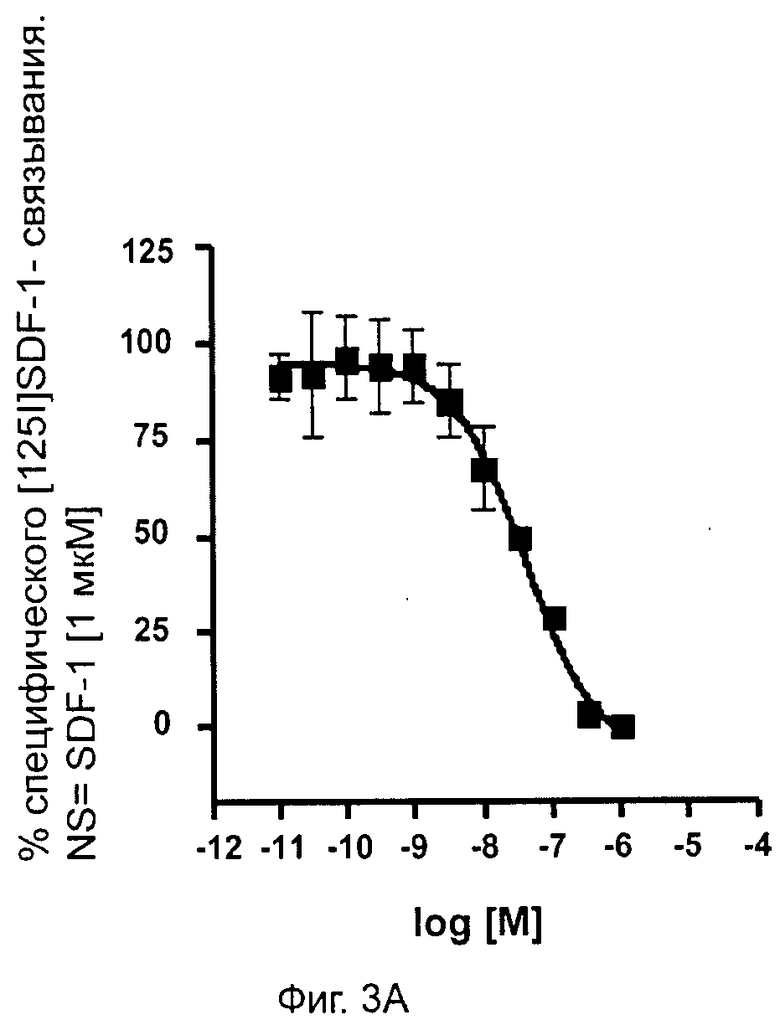
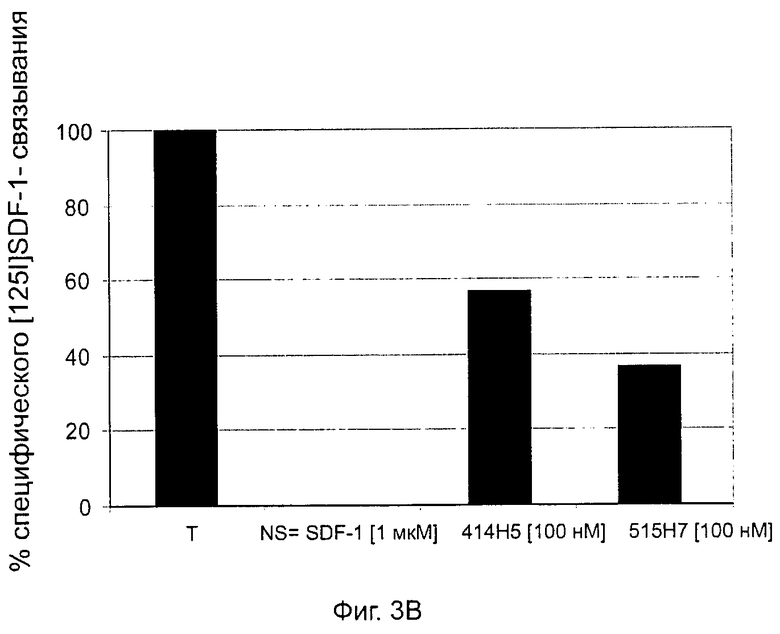
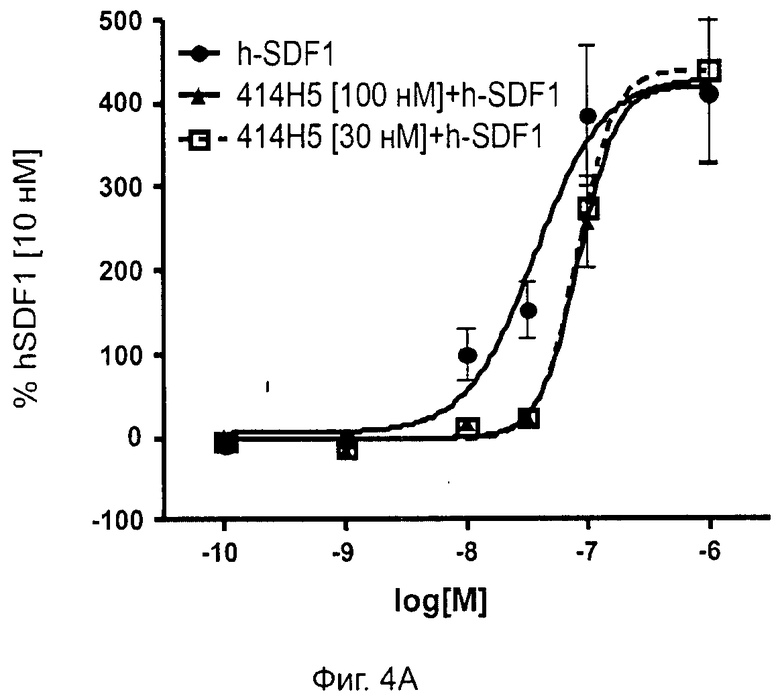
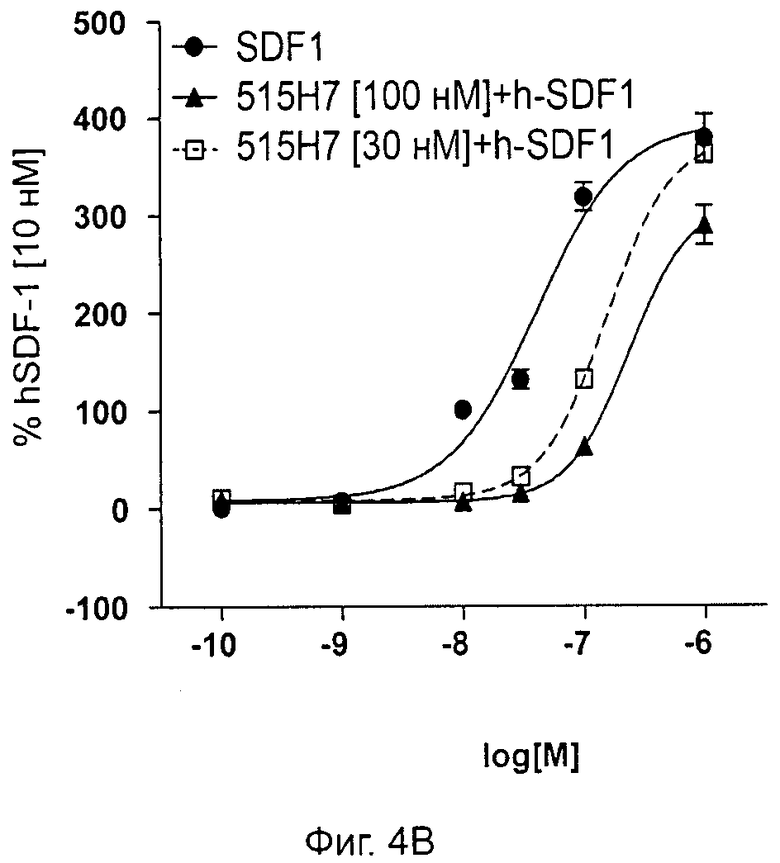
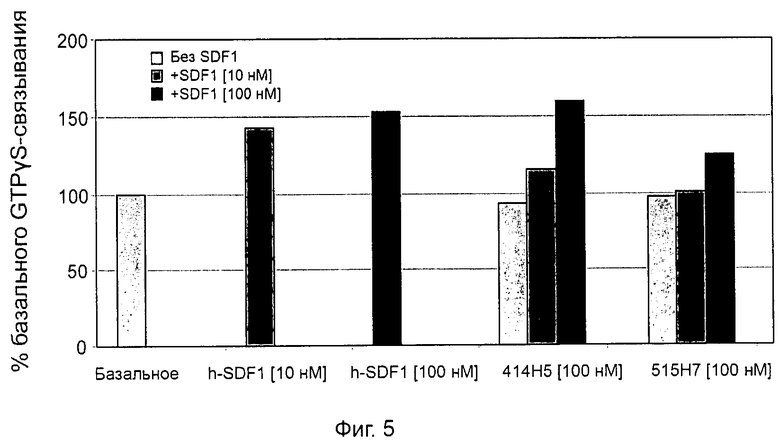
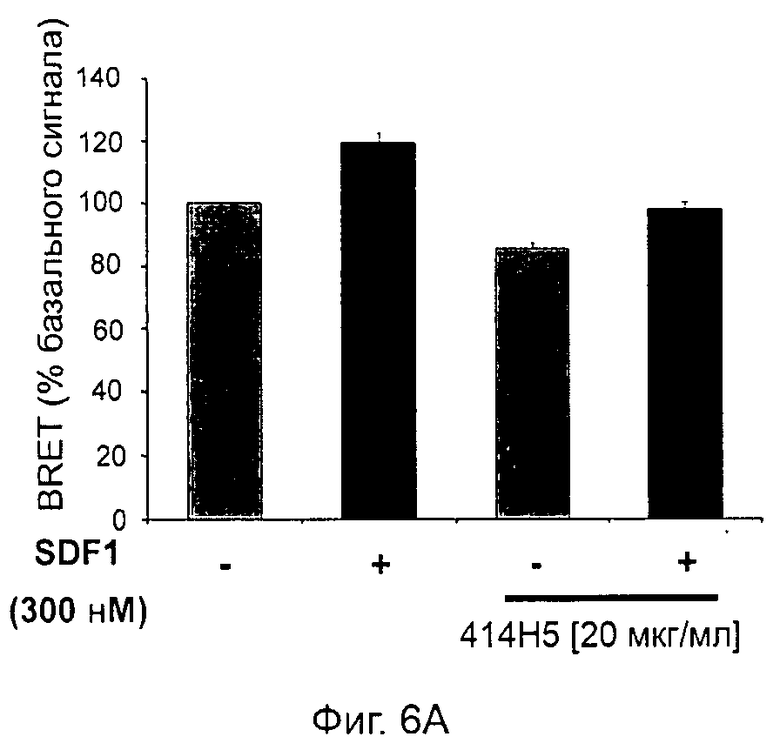
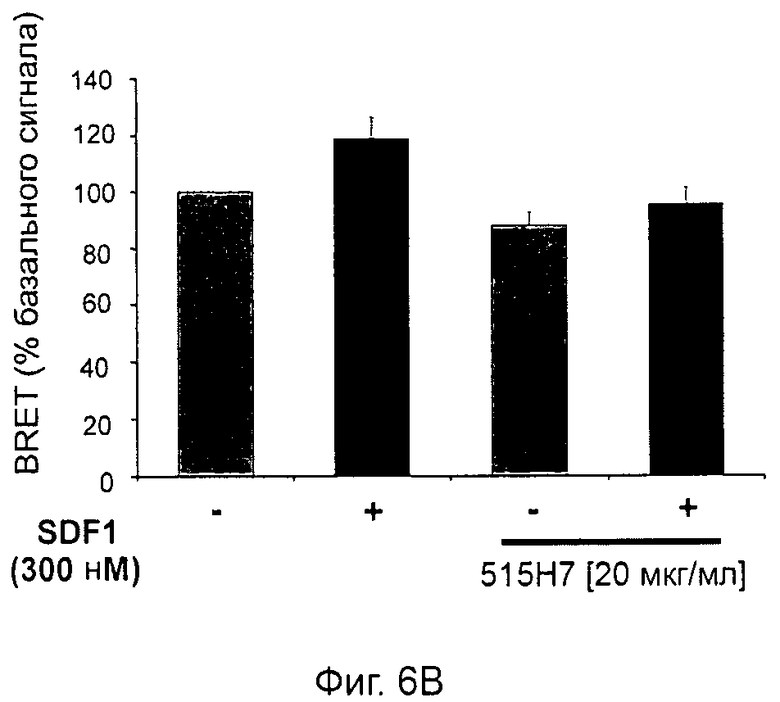
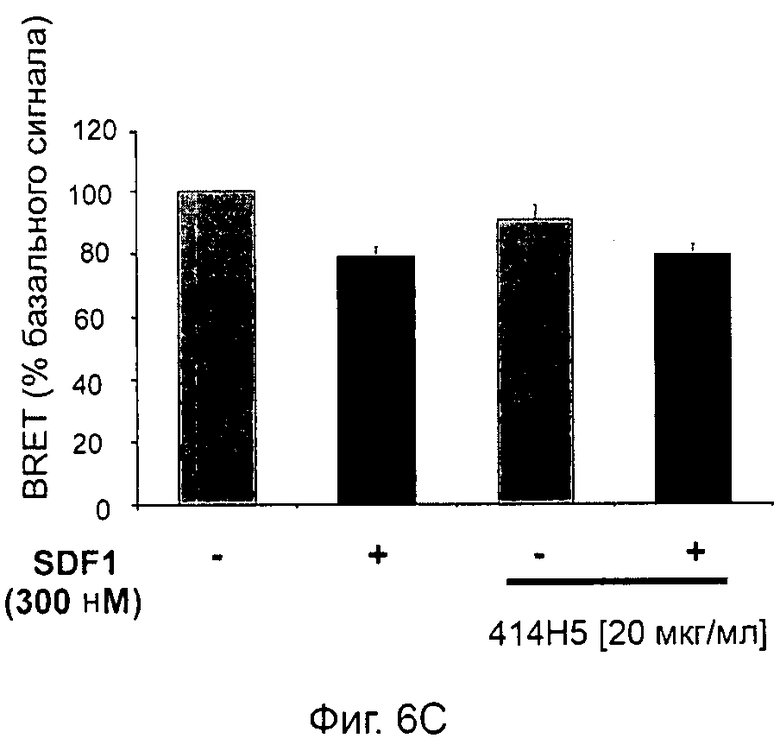
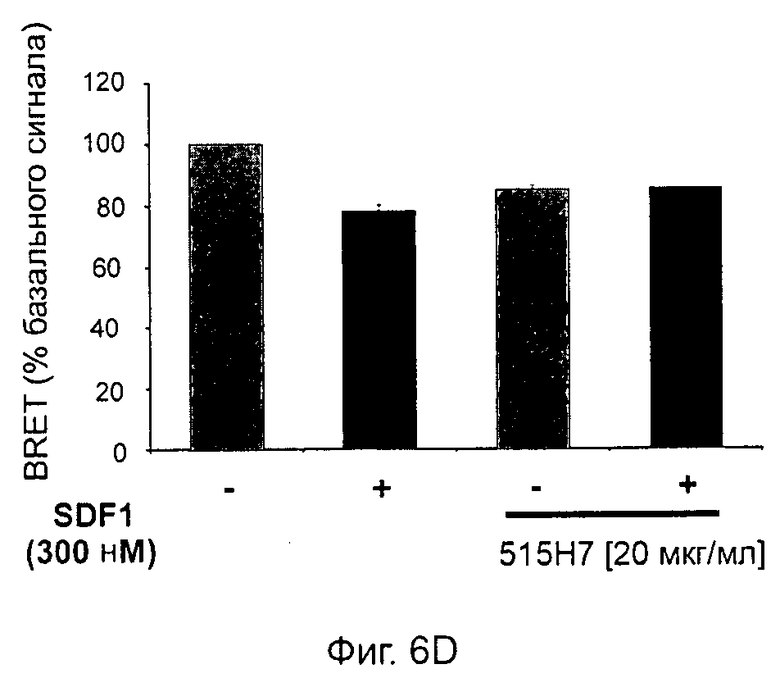
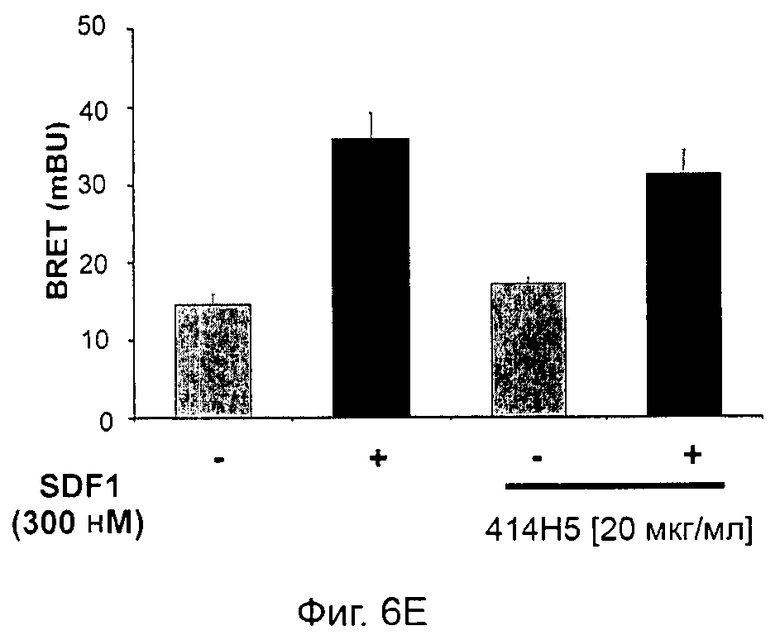
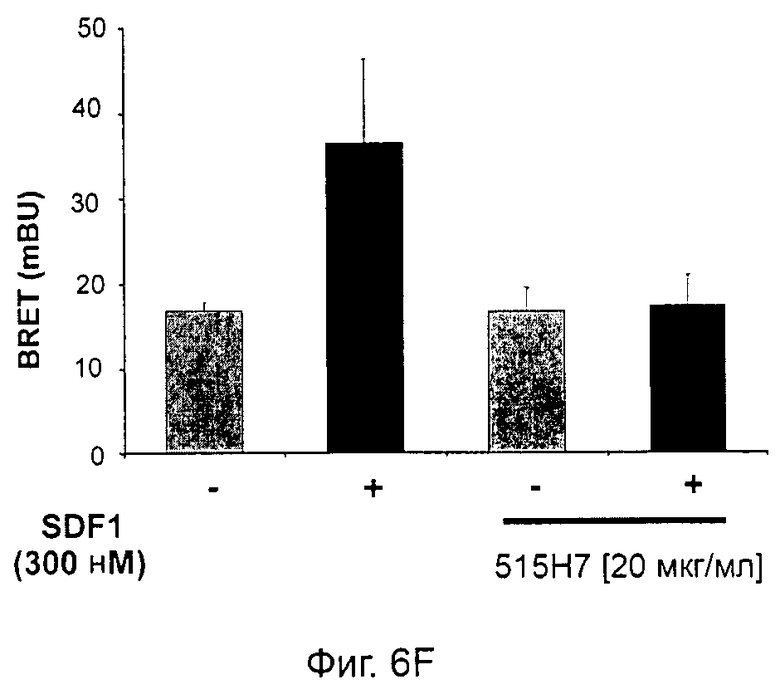
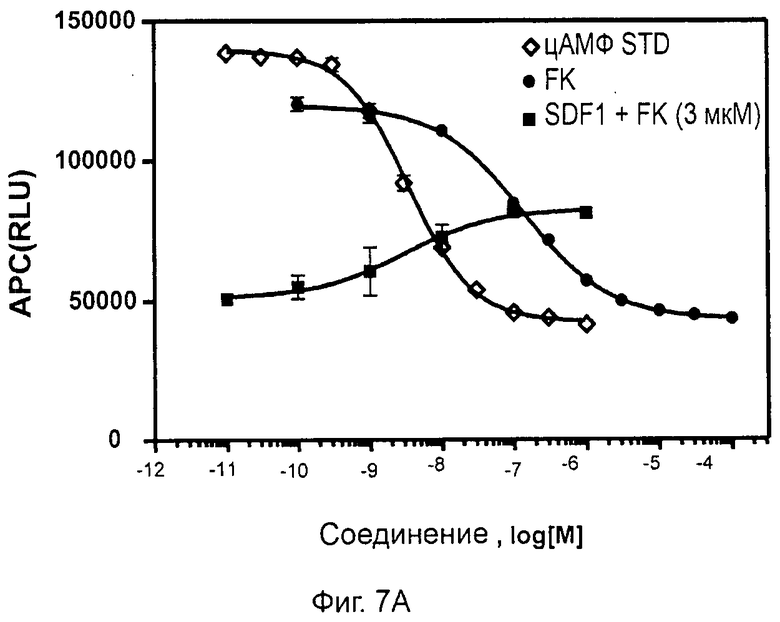

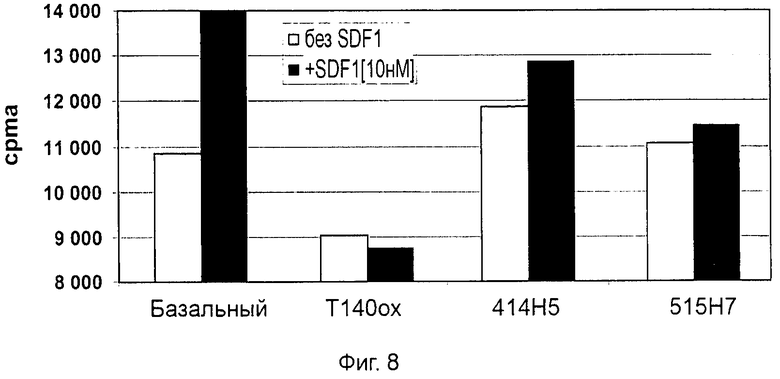
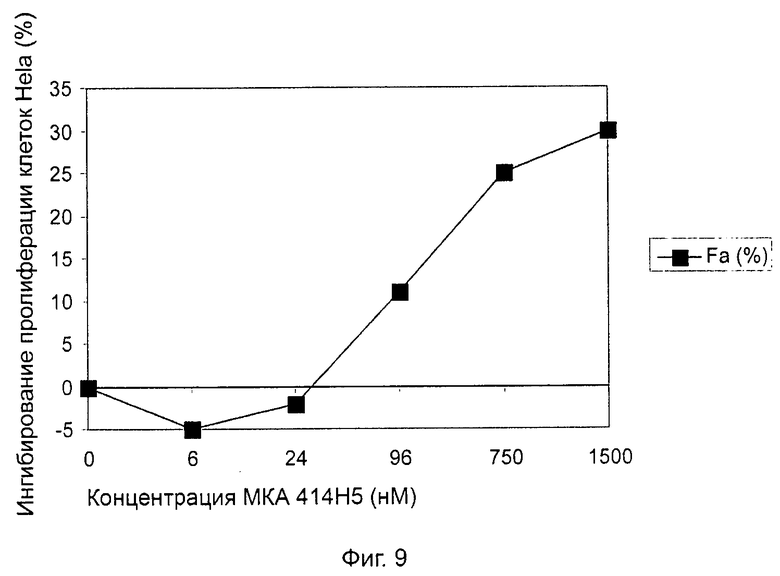
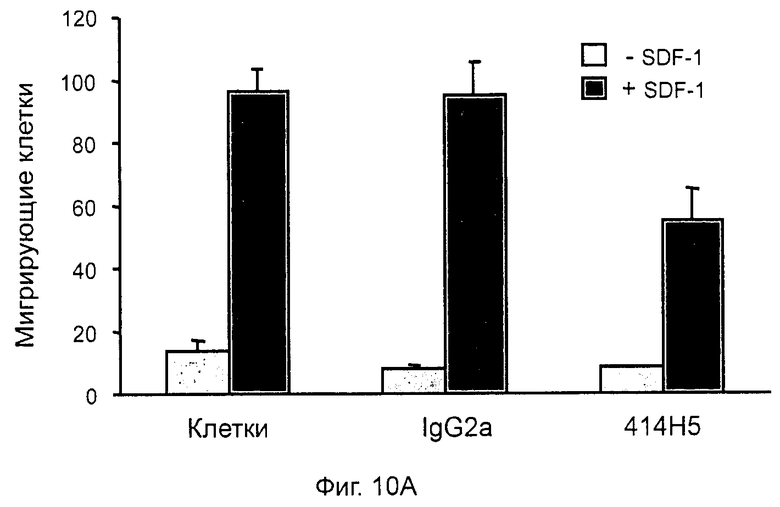
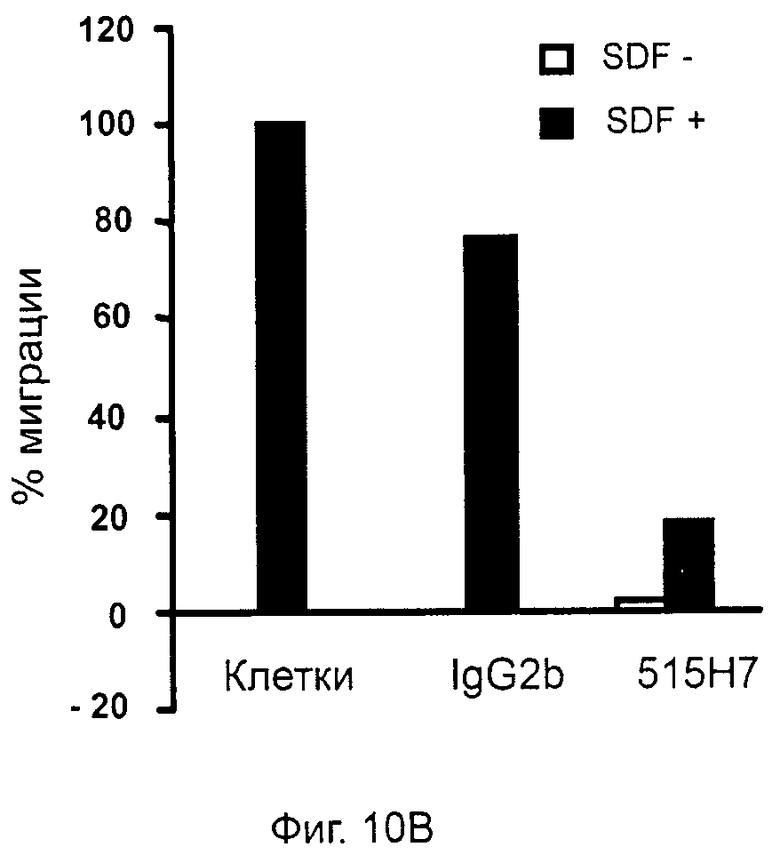
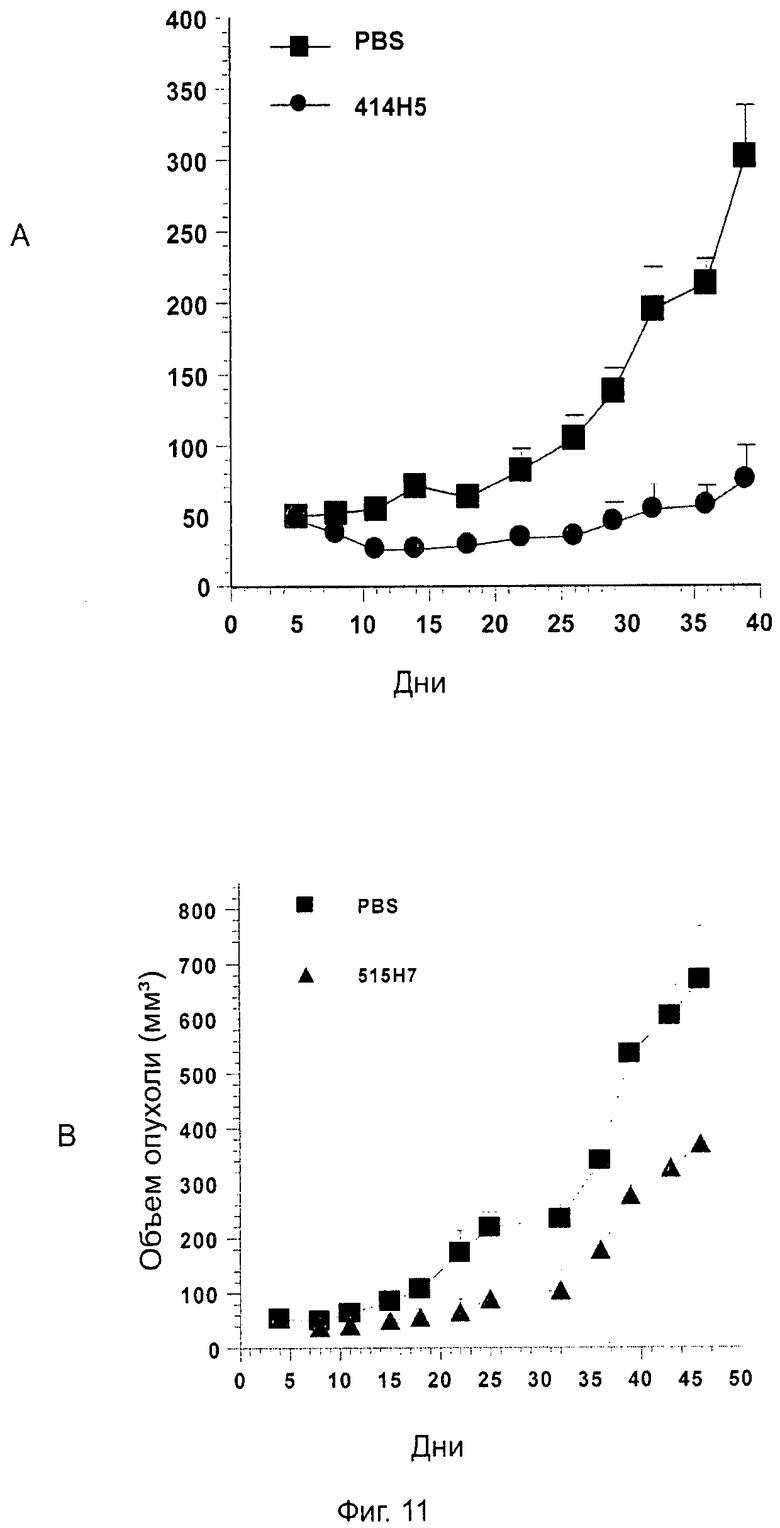
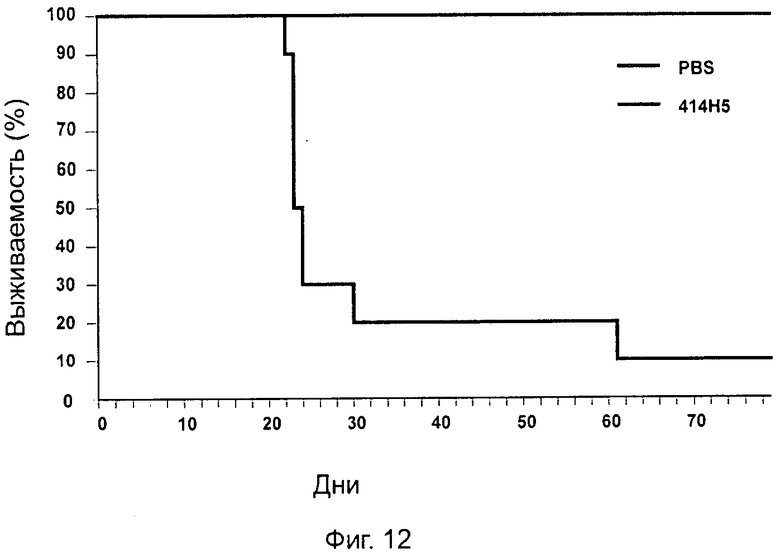
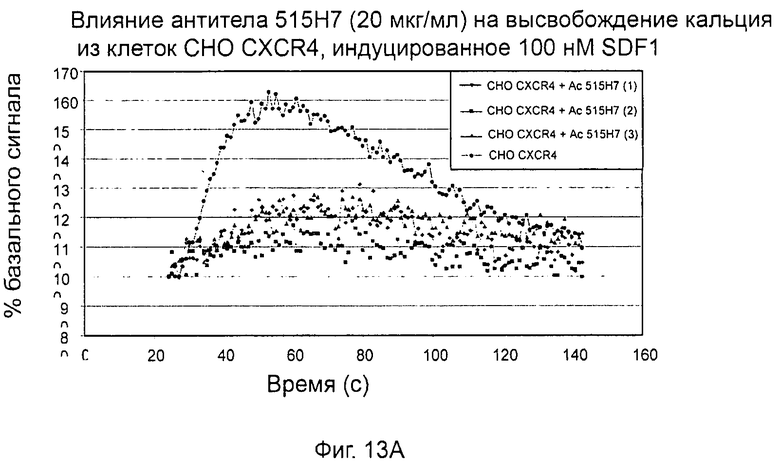
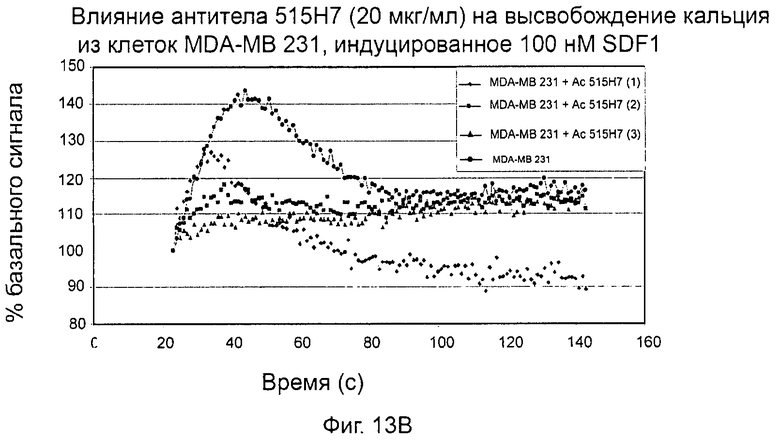
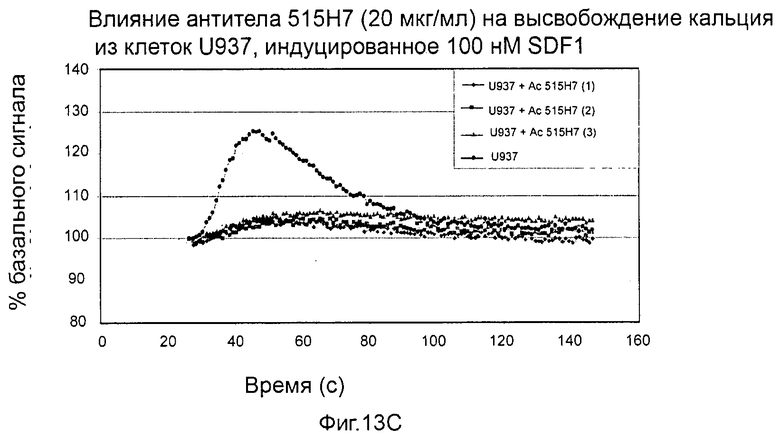
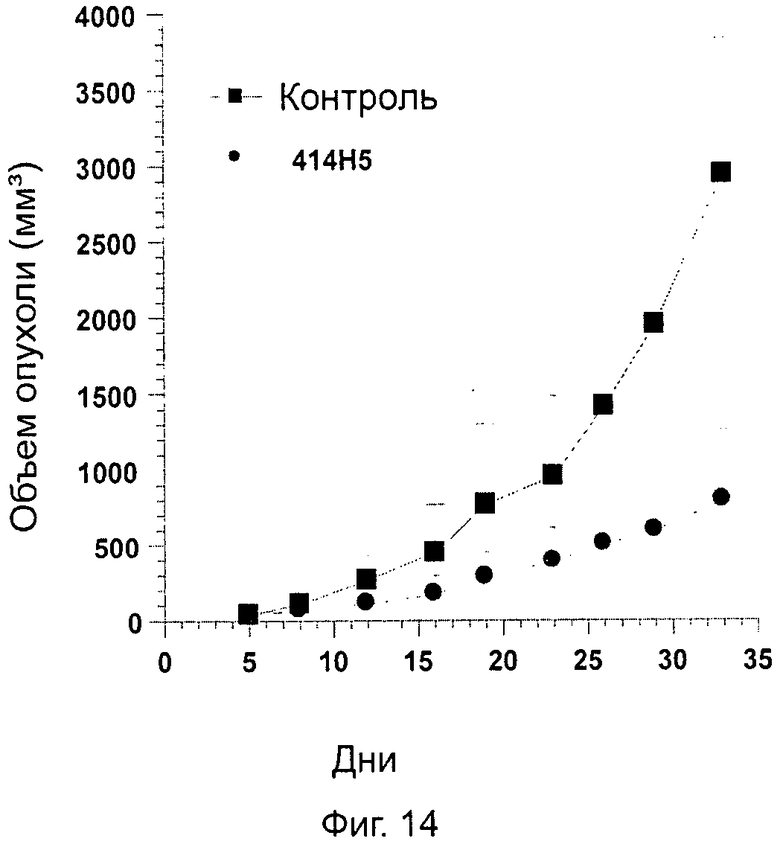
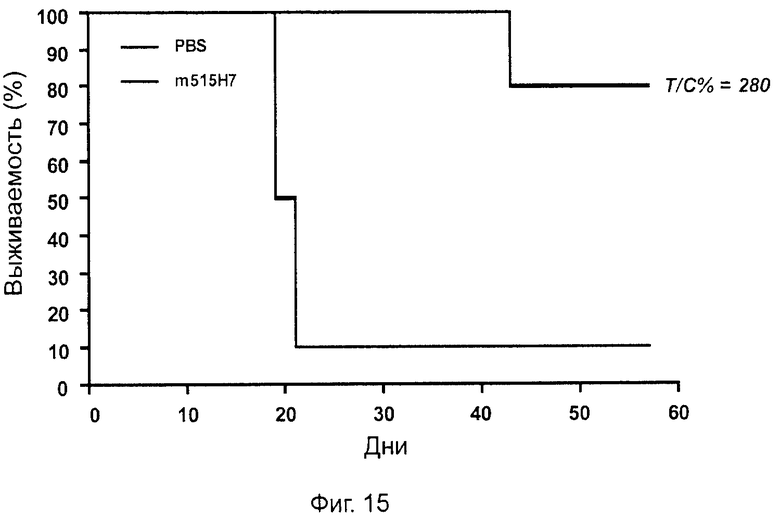
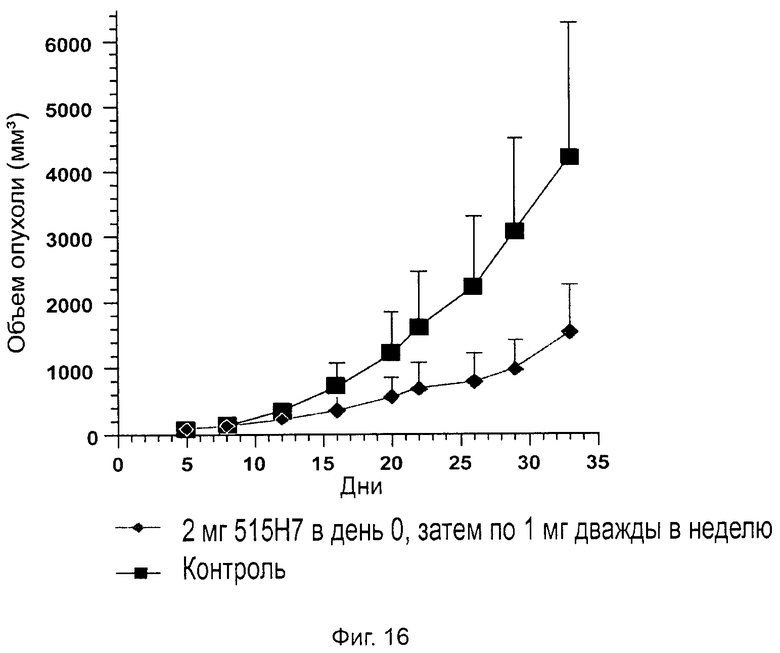
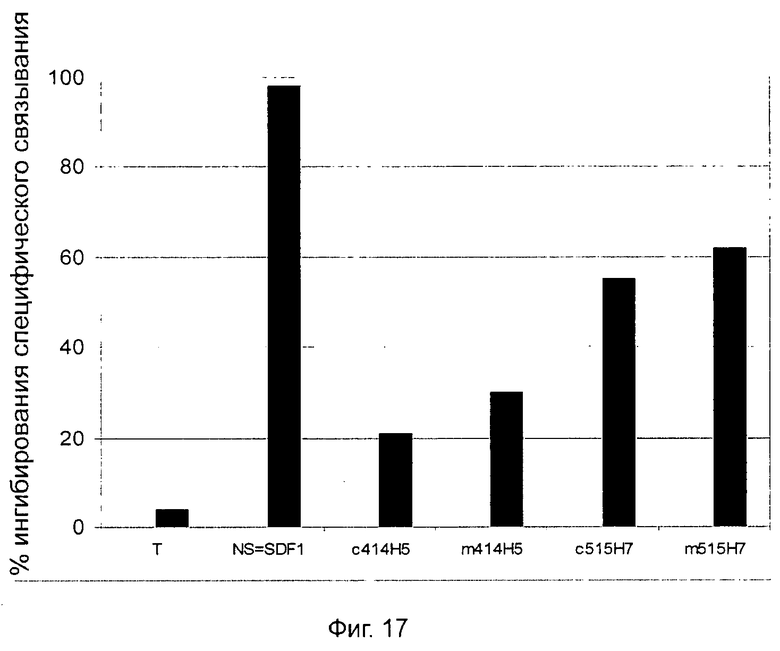
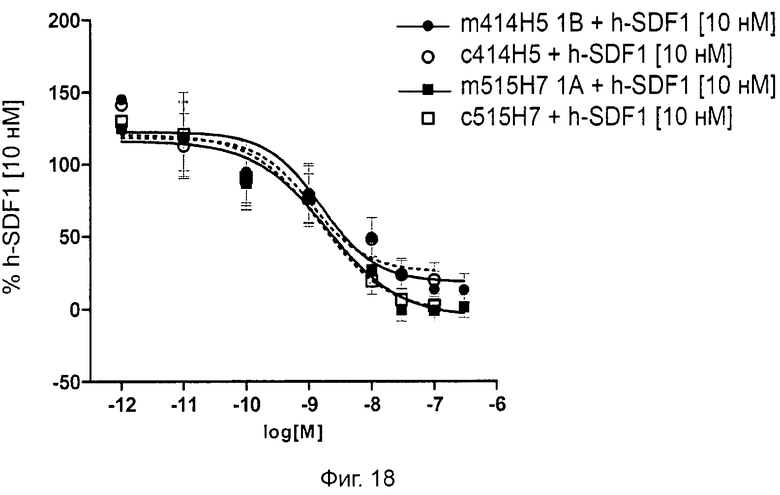
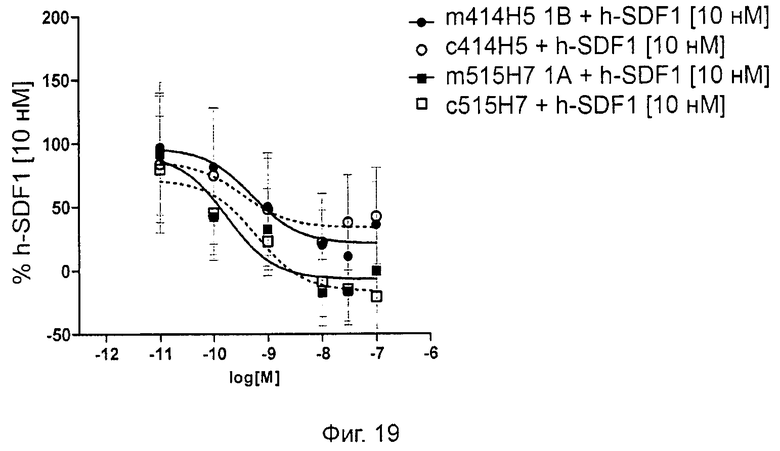
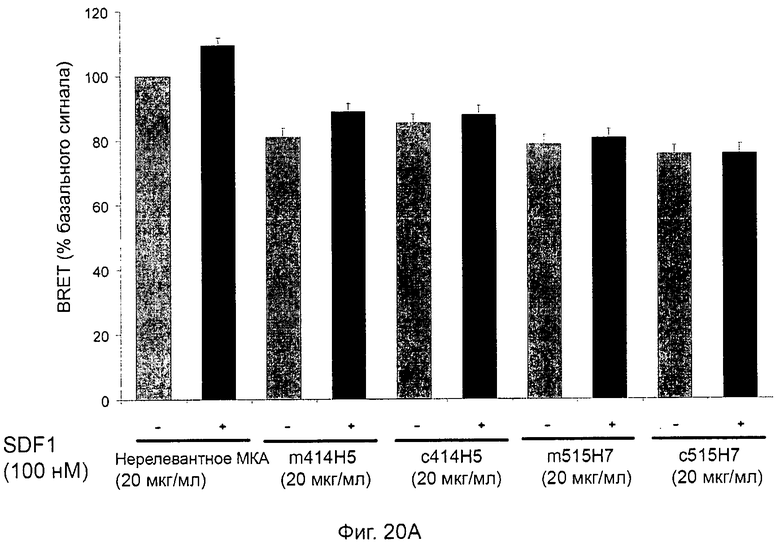
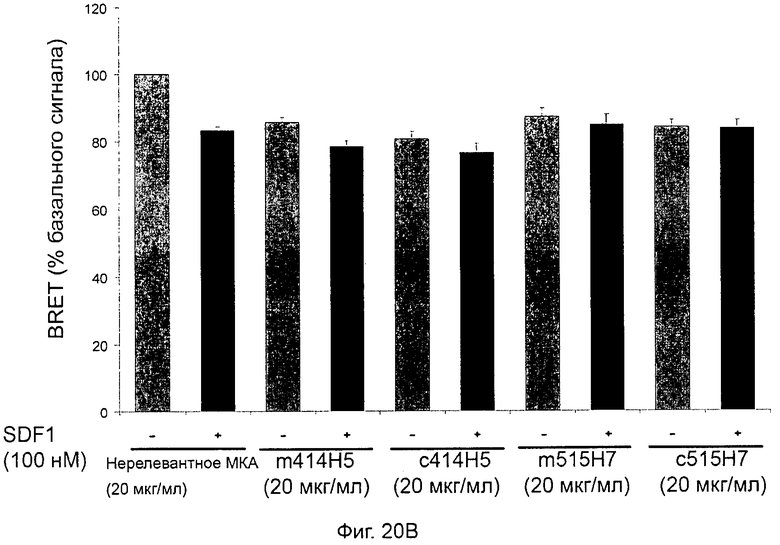
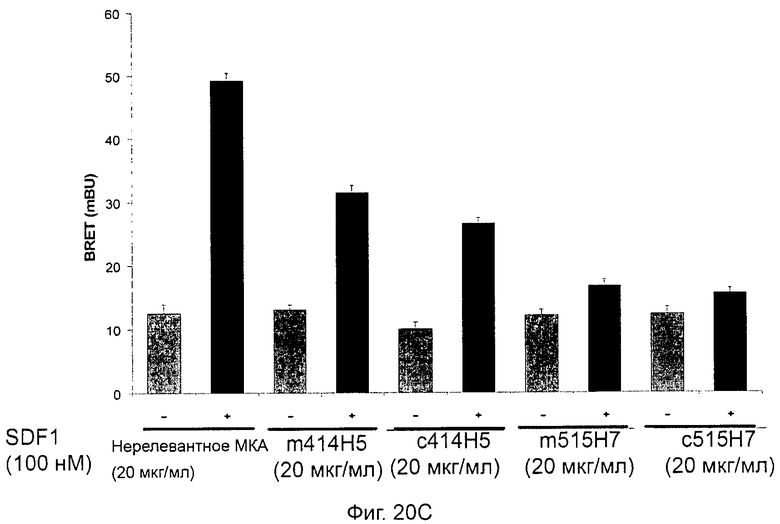
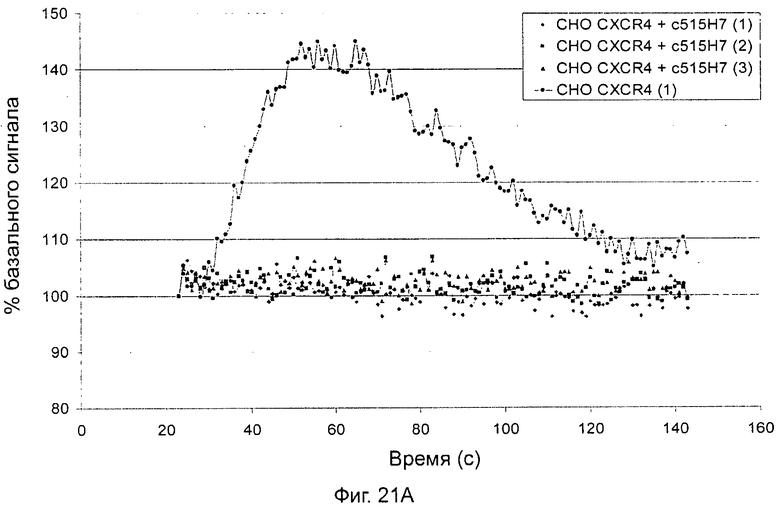
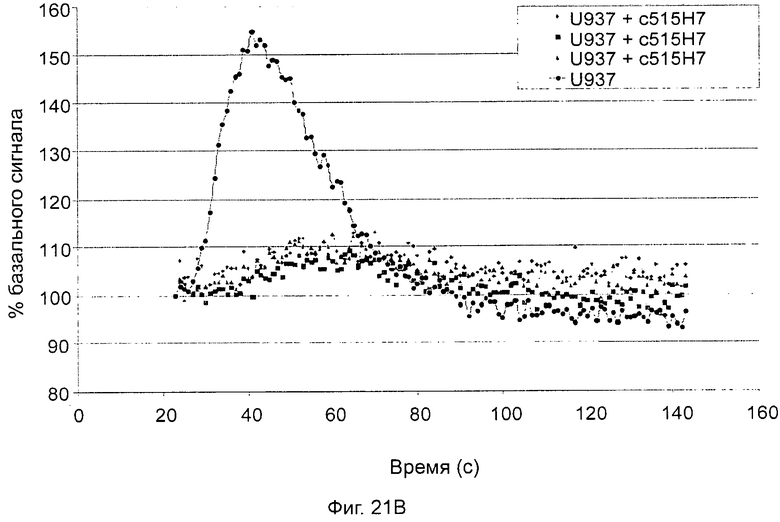
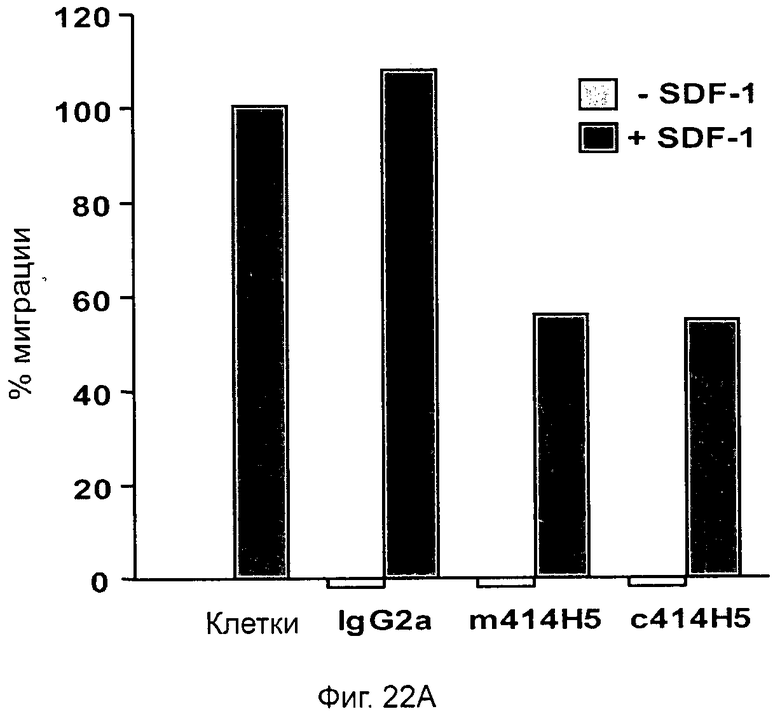
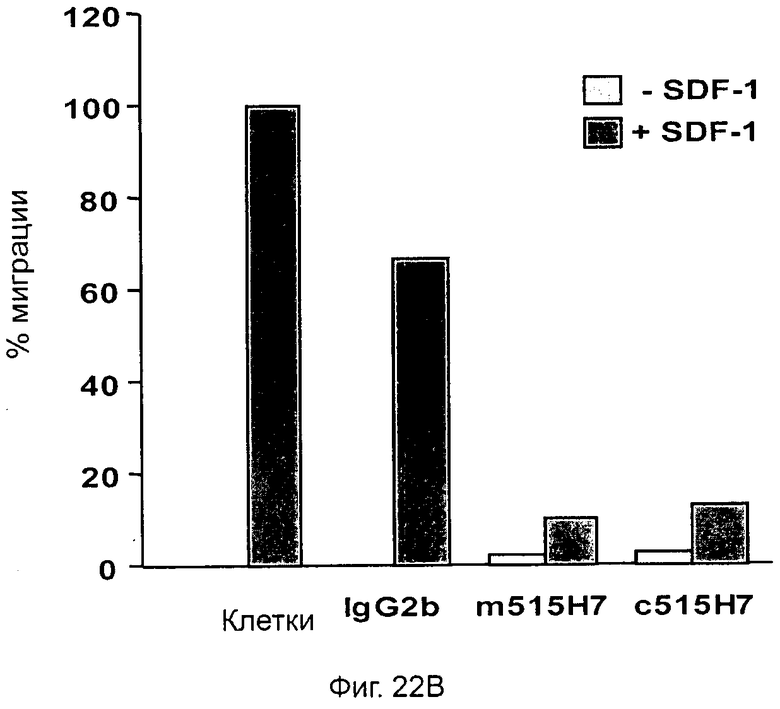
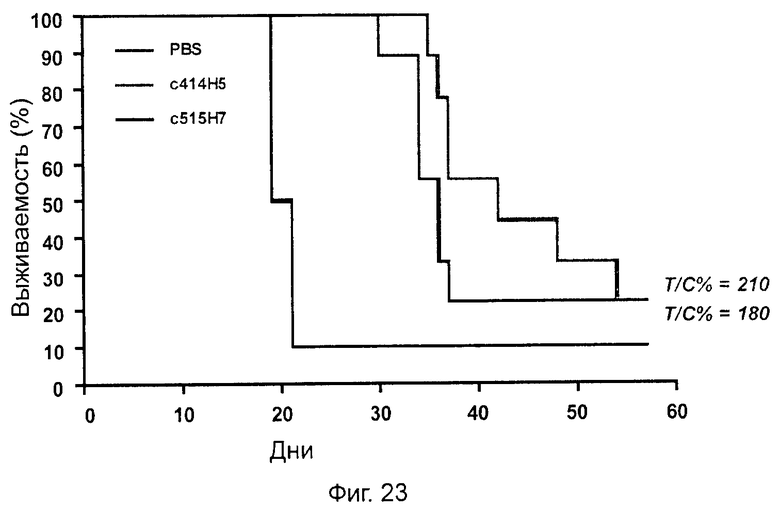
Изобретение относится к области иммунологии. Представлены вариантные выделенные анти-CXCR4-антитела 414H5, 515H7 и их антигенсвязывающие фрагменты, способные ингибировать активацию CXCR4, где каждое антитело или его антигенсвязывающий фрагмент содержит набор из 6 CDR, последовательности, которых представлены в описании. Описаны: нуклеиновая кислота, кодирующая указанное антитело, вектор для получения анти-CXCR4-антитела и клетка-хозяин, продуцирующая указанное антитело против CXCR4, трансфицированная указанным вектором. Предложен способ получения анти-CXCR4-антитела или его функционального фрагмента по настоящему изобретению, включающий: культивирование указанной клетки-хозяина в среде в соответствующих культуральных условиях; и выделение указанного антитела или одного из его функциональных фрагментов. Раскрыта фармацевтическая композиция, содержащая в качестве активного ингредиента указанное антитело или его функциональный фрагмент в эффективном количестве, а также эксципиент и/или фармацевтически приемлемый носитель. Изобретение позволяет получить антитела против CXCR4, модулирующие пространственную конформацию димеров CXCR4/CXCR4 и CXCR2/CXCR4 при раковых заболеваниях. 9 н. и 14 з.п. ф-лы, 23 ил., 5 табл., 24 пр.
1. Анти-CXCR4-антитело или его функциональный фрагмент, способные ингибировать активацию CXCR4, где указанное антитело или его функциональный фрагмент выбраны из группы, состоящей из:
(а) антитела или его функционального фрагмента, которые содержат:
- легкую цепь, содержащую, согласно IMGT, CDR-L1 последовательности SEQ ID NO:1, CDR-L2 последовательности SEQ ID NO:2 и CDR-L3 последовательности SEQ ID NO:3; и
- тяжелую цепь, содержащую, согласно IMGT, CDR-H1 последовательности SEQ ID NO:7, CDR-H2 последовательности SEQ ID NO:5 и CDR-H3 последовательности SEQ ID NO:8:
(б) антитела или его функционального фрагмента, содержащих:
- легкую цепь, содержащую, согласно Kabat, CDR-L1 последовательности SEQ ID NO:9, CDR-L2 последовательности SEQ ID NO:10 и CDR-L3 последовательности SEQ ID NO:3; и
- тяжелую цепь, содержащую, согласно Kabat, CDR-H1 последовательности SEQ ID NO:11, CDR-H2 последовательности SEQ ID NO:12 и CDR-H3 последовательности SEQ ID NO:6:
(в) антитела или его функционального фрагмента, содержащих:
- легкую цепь, содержащую, согласно IMGT, CDR-L1 последовательности SEQ ID NO:40, CDR-L2 последовательности SEQ ID NO:2 и CDR-L3 последовательности SEQ ID NO:41; и
- тяжелую цепь, содержащую, согласно IMGT, CDR-H1 последовательности SEQ ID NO:44, CDR-H2 последовательности SEQ ID NO:5 и CDR-H3 последовательности SEQ ID NO:45; и
(г) антитела или его функционального фрагмента, содержащих:
- легкую цепь, содержащую, согласно Kabat, CDR-L1 последовательности SEQ ID NO:46, CDR-L2 последовательности SEQ ID NO:47 и CDR-L3 последовательности SEQ ID NO:41; и
- тяжелую цепь, содержащую, согласно Kabat, CDR-H1 последовательности SEQ ID NO:48, CDR-H2 последовательности SEQ ID NO:49 и CDR-H3 последовательности SEQ ID NO:43.
2. Анти-CXCR4-антитело или его функциональный фрагмент по п. 1, способные ингибировать по меньшей мере 40% сигнала BRET на клетках, экспрессирующих оба CXCR4-RLuc/CXCR4-YFP и/или CXCR4-RLuc/CXCR4-YFP.
3. Анти-CXCR4-антитело или его функциональный фрагмент по п. 1 или п. 2, характеризующиеся тем, что указанное антитело или его функциональный фрагмент выбраны из группы, состоящей из
(а) антитела или его функционального фрагмента, содержащих последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:13, и последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:14; и
(б) антитела или его функционального фрагмента, содержащих последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:50, и последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:51.
4. Мышиная гибридома, секретирующая анти-CXCR4-антитело по п. 1, депонированная в CNCM, Институт Пастера, Париж, под номером I-3860.
5. Мышиная гибридома, секретирующая анти-CXCR4-антитело по п. 1, депонированная в CNCM, Институт Пастера, Париж, под номером I-4019.
6. Анти-CXCR4-антитело, секретируемое гибридомой по п. 4 или 5.
7. Анти-CXCR4-антитело или его функциональный фрагмент по п. 1, характеризующееся тем, что оно является химерным антителом.
8. Анти-CXCR4-антитело или его функциональный фрагмент по п. 7, характеризующиеся тем, что указанное антитело или его функциональный фрагмент выбраны из группы, состоящей из:
(а) химерного антитела, содержащего последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:64, и последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:65; и
(б) химерного антитела или его функционального фрагмента, содержащих последовательность легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:66, и последовательность тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:67.
9. Изолированная нуклеиновая кислота, кодирующая анти-CXCR4-антитело, характеризующаяся тем, что она представляет собой нуклеиновую кислоту, ДНК или РНК, кодирующую антитело или его функциональный фрагмент по одному из пп. 1, 2, 3, 6, 7 или 8.
10. Вектор для получения анти-CXCR4-антитела, содержащий нуклеиновую кислоту по п. 9.
11. Клетка-хозяин для получения анти-CXCR4-антитела, содержащая вектор по п. 10.
12. Способ получения анти-CXCR4-антитела или его функционального фрагмента по одному из пп. 1, 2, 3, 6, 7 или 8, характеризующийся тем, что он включает следующие этапы:
- культивирование клетки-хозяина по п. 11 в среде в соответствующих культуральных условиях; и
- выделение указанного антитела или одного из его функциональных фрагментов, полученных таким образом, из культуральной среды или указанных культивируемых клеток.
13. Анти-CXCR4-антитело или его функциональный фрагмент по п. 1 для применения в качестве лекарственного средства.
14. Фармацевтическая композиция, связывающая CXCR4, содержащая в качестве активного ингредиента соединение, представляющее собой антитело, или его функциональный фрагмент по п. 1 в эффективном количестве, а также эксципиент и/или фармацевтически приемлемый носитель.
15. Фармацевтическая композиция по п. 14, характеризующаяся тем, что она дополнительно содержит противоопухолевое антитело, отличное от антитела, направленного против CXCR4.
16. Фармацевтическая композиция по п. 14, характеризующаяся тем, что она дополнительно содержит цитотоксический/цитостатический агент, клеточный токсин и (или) радиоизотоп.
17. Фармацевтическая композиция по п. 14 для применения в качестве лекарственного средства.
18. Антитело или его функциональный фрагмент по п. 1 в качестве лекарственного средства для модуляции активности CXCR4 в клетке.
19. Антитело или его функциональный фрагмент по п. 1 для профилактики или лечения рака.
20. Антитело или его функциональный фрагмент по п. 19, где указанный рак является раком, выбранным среди рака предстательной железы, остеосаркомы, рака легкого, рака молочной железы, рака эндометрия, множественной миеломы, рака яичников, рака поджелудочной железы и рака толстой кишки.
21. Фармацевтическая композиция по п. 14 в качестве лекарственного средства для модуляции активности CXCR4 в клетке.
22. Фармацевтическая композиция по п. 14 для профилактики или лечения рака.
23. Фармацевтическая композиция по п. 22, характеризующаяся тем, что указанный рак является раком, выбранным среди рака предстательной железы, остеосаркомы, рака легкого, рака молочной железы, рака эндометрия, множественной миеломы, рака яичников, рака поджелудочной железы и рака толстой кишки.
| WO 2006089141 А2, 24.08.2006 | |||
| US 2003206909 А1, 06.11.2003 | |||
| US 2005123563 А1, 09.06.2005 | |||
| А.С | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ образования коричневых окрасок на волокне из кашу кубической и подобных производных кашевого ряда | 1922 |
|
SU32A1 |

