ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к молекуле нуклеиновой кислоты и ее применению для модуляции иммунной системы.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ
Новой стратегией борьбы с болезнями иммунных комплексов, такими как рак, инфекционные заболевания, аллергия и астма, является использование иммунной системы пациента. Известно, что иммунная система или ее активность может модулироваться специфическими последовательностями ДНК. Самые известные иммуномодулирующие короткие последовательности ДНК содержат неметилированный мотив цитозин гуанин (CG мотив), который описан Krieg et al. (Nature 1995 374:6522 546-549). Проявление неметилированных CG мотивов по существу подавлено в геноме эукариот по сравнению с прокариотами или вирусами. Следовательно, молекулы ДНК, содержащие такой мотив, выступают как естественный «сигнал опасности» и запускают иммунную систему на борьбу с прокариотическими или вирусными патогенами. Это может быть использовано терапевтически или профилактически для лечения или профилактики инфекционных или неинфекционных заболеваний.
ДНК конструкты, содержащие неметилированые CG мотивы, способны оказывать значительное физиологическое воздействие за счет сильной стимуляции клеток-эффекторов врожденной иммунной системы, включающей дендритные клетки, макрофаги, естественные клетки-киллеры (NK) и NKT клетки. Неметилированные CG мотивы определяются Toll-подобными рецепторами 9 (TLR9) паттерн-распознающими рецепторами врожденной иммунной системы. Хотя точный механизм распознавания еще полностью не установлен, достигнут значительный прогресс в раскрытии основополагающих путей (A. Krieg, Nat. Rev. Drug Disc, 5:471-484, 2006). Предполагается, что при связывании ДНК конструктов, содержащих неметилированные CG, с рецептором в реактивных клетках активируется сложный сигнальный каскад. Адаптивный иммунитет индуцирован за счет повышенной регуляции специфических поверхностных молекул и по существу Th1 паттерна секреции цитокинов. Такие конструкты могут быть использованы в комбинации, например, с антителами, химиотерапией или радиационной терапией, вакцинами или цитокинами. Аллергические заболевания и астма являются главным образом Th2-опосредованными. За счет повышения соотношения Th1/Th2 Th2-опосредованные ответы ослабляются и, следовательно, эти типы заболеваний могут лечиться или может осуществляться их профилактика.
Поверхностные молекулы включают, например, CD40, CD69, CD80 или CD86 в зависимости от анализируемого типа специфических клеток. Секреция цитокинов также специфична для различных типов клеток; цитокины включают, например, воспалительный белок макрофагов (MIP)-1 альфа, MIP-1 бета, интерлейкин (IL)-6, IL-8, интерферон (IFN)-альфа, фактор некроза опухоли (TNF)-альфа, IFN-гамма, моноцитарный хемотаксический белок (МСР)-1 или IFN-гамма-индуцируемый белок 10 кДа (IP-10).
Вакцинация показала себя как очень эффективное средство для профилактики или лечения заболеваний. Для того чтобы гарантировать сильный и длительный иммунный ответ, как правило, вместе с антигеном вводят адъюванты, способные стимулировать антигенпрезентирующие клетки, такие как дендритные клетки, и для этой цели TLR9 агонисты проявили себя сильными иммуностимуляторами.
Независимо от любых объяснений основополагающих механизмов, за счет которых неметилированные CG мотивы влияют на иммунный ответ или модулируют иммунный ответ, было разработано множество подход для модуляции иммунной системы при использовании таких мотивов. В WO 1998/018810 описывается, что иммуностимулирующие последовательности, содержащие неметилированные CG мотивы, даже более эффективны, когда они являются частью одноцепочечной ДНК. Однако введение одноцепочечных с открытой цепью ДНК молекул практически не осуществимо из-за быстрой деградации одноцепочечных нуклеиновых кислот. Следовательно, были разработаны различные способы защиты одно- или двухцепочечных конструктов ДНК, содержащих неметилированный CG мотив.
Для достижения устойчивости к расщеплению ДНК нуклеазами фосфодиэфирные связи в скелете полимера нуклеиновой кислоты часто модифицируют до фосфоротиоатов. Помимо несколько меньшей стимулирующей активности такие фосфоротиоат-защищенные нуклеиновые кислоты в клинических исследованиях в последние годы показали, что токсичность фосфоротиоатной защиты исключает или очень сильно ограничивает применение таких нуклеиновых кислот в фармацевтических композициях или лекарственных средствах.
Другой подход для защиты ДНК последовательностей, содержащих CG мотив, описан, например, в EP 1196178. В этом документе описываются короткие молекулы дезоксирибонуклеиновой кислоты, содержащие частично одноцепочечные, гантелеобразные, ковалентно закрытые последовательности нуклеотидных остатков, содержащих CG мотивы («dSLIM»). Согласно описанию EP 1196178 CG мотивы расположены в одноцепочечных петлях на обоих концах двухцепочечного стебля описанной молекулы или в двухцепочечном стебле. Одноцепочечные шпилечные петли защищают двухцепочечный стебель от разрушения ДНК нуклеазами внутри или снаружи клетки.
В документе WO 2010/039137 описаны иммунорегулирующие олигонуклеотиды в качестве антагонистов TLR опосредованных заболеваний с одной или более химической модификацией в последовательности, фланкирующей иммуностимулирующий мотив, и/или в олигонуклеотидном мотиве, который был бы иммуностимулирующим, но для целей модификации. Следовательно, целью описанных в WO 2010/039137 олигонуклеотидов является подавление иммунного ответа, вызванного TLR.
В WO 2005/042018 описан новый, так называемый C-класс CpG олигонуклеотидов, где C-класс олигонуклеотидов характеризуется CpG последовательностями, как правило, расположенными на или вблизи 5′-конца или 3′-конца молекулы, и GC-мотив, богатый палиндромами, как правило, расположен на или вблизи другого конца молекулы. В документе описываются варианты палиндромной последовательности C-класса ДНК.
КРАТКОЕ ОПИСАНИЕ
Объект настоящего изобретения относится к альтернативным иммуномодулирующим конструктам ДНК, способным быть стабильными после переноса в эукариотические клетки и не имеющим побочных эффектов.
Настоящее изобретение относится к ДНК конструкту для иммунностимуляции, содержащему по меньшей мере одну последовательность мотива N1N2CGN3N4, где N1N2 и N3N4 представляют любую комбинацию C, G, A и T, и C представляет дезоксицитидин, G представляет дезоксигуанозин, A представляет дезоксиаденозин и T представляет дезокситимидин, и где конструкт представляет линейную одноцепочечную или двухцепочечную последовательность ДНК, содержащую по меньшей мере один нуклеотид в L-конфигурации. N1N2 может представлять элемент, выбранный из группы, состоящей из GT, GG, GA, AT или AA, N3N4 представляет элемент, выбранный из группы, состоящей из CT или TT.
В одном варианте воплощения настоящее изобретение относится к конструкту, где по меньшей мере один нуклеотид в L-конфигурации включен в пять терминальных нуклеотидов, расположенных на или вблизи 5′- и/или 3′-конца одноцепочечной ДНК.
Дополнительно, настоящее изобретение относится к конструкту ДНК по меньшей мере с одним G повтором из по меньшей мере этих последовательных дезоксигуанозинов, расположенных на или вблизи 5′- и/или 3′-конца, где G повтор может быть расположен между двумя мотивами согласно пункту 1 или 2.
Интервал между двумя мотивами по пункту 1 или 2 может составлять по меньшей мере пять оснований, в частности, когда дезоксигуанозин не является элементом последовательности.
Дополнительно, предполагается, что последовательность ДНК представляет линейный с открытой цепью ДНК конструкт, содержащий одноцепочечную или двухцепочечную ДНК, или представляет линейный ДНК конструкт, который включает по меньшей мере один конец с одноцепочечной петлей.
Указанный выше мотив последовательности N1N2CGN3N4 должен быть расположен в одноцепочечной и/или двухцепочечной области последовательности ДНК.
В одном варианте воплощения настоящего изобретения конструкт содержит меж- и/или внутримолекулярные пары оснований и по меньшей мере одну неспаренную одноцепочечную область.
Дополнительно, обеспечен мультимерный конструкт, где по меньшей мере два конструкта, содержащие меж- и/или внутримолекулярные пары оснований и по меньшей мере одну неспаренную одноцепочечную область, соединены друг с другом.
Дополнительно, конструкт может содержать по меньшей мере один нуклеотид в L- или D-конфигурации, модифицированный функциональной группой, выбранной из группы, содержащей карбоксильную группу, аминную, амидную, альдиминную, кетальную, ацетальную, сложного эфира, простого эфира, дисульфидную, тиоловую и альдегидные группы.
Модифицированный нуклеотид может быть связан с соединением, выбранным из группы, состоящей из пептидов, белков, углеводов, антител, синтетических молекул, полимеров, микрочастиц, частиц металлов, наночастиц, мицелл, липидных носителей или твердых фаз.
Настоящее изобретение относится к конструкту ДНК, имеющему первый G повтор на 5′-конце и три мотива последовательности по пунктам 1 или 2, где по меньшей мере пять оснований расположены между первым и вторым мотивами, исключая дезоксигуанозин, и G повтор, который расположен между вторым и третьим мотивами последовательности, и где два из трех 3′ терминальных дезоксинуклеотидов находятся в L-конфигурации.
Конструкты по настоящему изобретению могут быть использованы для лечения рака или аутоиммунных заболеваний или для модуляции иммунной системы.
В одном из вариантов воплощения настоящее изобретение относится к фармацевтической композиции, содержащей указанный выше ДНК конструкт. Фармацевтическая композиция также может содержать химиотерапевтический агент.
Дополнительно, настоящее изобретение относится к вакцине, содержащей указанный выше ДНК конструкт. В ней ДНК конструкт может содержаться в качестве адъювант.
ДЕТАЛЬНОЕ ОПИСАНИЕ
Настоящее изобретение относится к линейной с открытой цепью последовательности ДНК, полученной как ДНК конструкт. Указанная ДНК последовательность может представлять одноцепочечную или частично или полностью двухцепочечную. Используемый в описании настоящей патентной заявки термин ДНК конструкт не ограничивает длину соответствующей ДНК последовательности. Мономерные единицы ДНК конструктов представляют нуклеотиды.
ДНК конструкт может быть получен синтетически или частично или полностью может быть биологического происхождения, где биологическое происхождение относится к генным способам получения последовательностей ДНК.
L-ДНК или нуклеотиды в L-конфигурации относятся к нуклеотидам, содержащим L-дезоксирибозу в качестве сахарного остатка вместо естественным образом присутствующей D-дезоксирибозы. L-дезоксирибоза представляет энантиомер (зеркальное отражение) D-дезоксирибозы. ДНК конструкты, частично или полностью состоящие из нуклеотидов в L-конфигурации, могут представлять частично или полностью одноцепочечные или двухцепочечные; однако нуклеотиды в L-конфигурации не могут гибридизоваться с нуклеотидами в D-конфигурации (Hauser et al., Nucleic Acid Res. 2006 34:5101-11). L-ДНК является равно растворимой и селективной, как D-ДНК. При этом L-ДНК устойчива к расщеплению присутствующими естественным образом ферментами, в частности экзонуклеазами, таким образом, L-ДНК защищена от биологического разрушения (Urata et al., Nucleic Acids Res. 1992 20:3325-32). Следовательно, L-ДНК имеет очень широкое применение.
Используемый в описании термин «стебель» относится к двухцепочечной ДНК, образованной спариванием оснований в той же самой молекуле ДНК (которая затем частично самокомплементируется) или в других молекулах ДНК (которые частично или полностью комплементарны). Внутримолекулярное спаривание оснований означает спаривание оснований в тех же самых молекулах, а спаривание оснований между другими молекулами ДНК определяется как межмолекулярное спаривание оснований.
Используемый в описании термин «петля» относится к неспаренной, одноцепочечной области внутри или на конце структуры стебля. Используемый в описании настоящей патентной заявки термин «шпилька» относится к отличающейся комбинации стебля и петли, которая имеет место, когда две самокомлементирующиеся области одной и той же молекулы ДНК гибридизуются с образованием стебля с неспаренной петлей. Гантелеобразная форма описывает линейный конструкт ДНК со шпильками на обоих концах, фланкирующих область стебля. Таким образом, «линейный конструкт ДНК» в контексте настоящего изобретения описывается как линейный с открытой цепью конструкт ДНК, содержащий одно- или двухцепочечную ДНК, или как линейный гантелеобразный конструкт ДНК, содержащий одноцепочечные петли на обоих концах двухцепочечной ДНК стебля.
Используемый в описании термин «конец ДНК» обозначает 5′- или 3′-конец одноцепочечной ДНК и относится не только к терминальному нуклеотиду, но включает пять терминальных нуклеотидов соответствующего конца ДНК. Модификация конца ДНК относится по меньшей мере к одному из соответствующих нуклеотидов.
Используемый в описании термин «G повтор» относится к последовательности по меньшей мере из трех последовательных дезоксигуанозинов.
Используемый в описании термин «твердая фаза», к которой нуклеотиды ковалентно или не ковалентно прикреплены, относится без ограничения к колонке, матрице, гранулам, стеклу, включая модифицированное или функционализированное стекло, кремний или материалы на основе кремния, включая силикон и модифицированный силикон, пластики (включая полипропилен, полиэтилен и сополимеры стирола и другие материалы, акрилы, полибутилен, полиуританы и тому подобное), нейлон или нитроцеллюлозу, смолы, полисахариды, углерод, а также неорганическое стекло, металлы, наночастицы и пластики. Таким образом, микротитрационные планшеты также входят в объем твердой фазы по настоящему изобретению.
Иммуномодуляция по настоящему изобретению относится к иммуностимуляции и иммуносупрессии. Иммуностимуляция предпочтительно означает, что клетки-эффекторы иммунной системы стимулированы для пролиферации, миграции, дифференцировки или проявления активности в любой другой форме. Пролиферация клеток, например, может быть индуцирована без костимулирующих сигналов при использовании иммуностимулирующих молекул ДНК, которые в норме требуют костимулирующего сигнала от хелперных T-клеток.
С другой стороны, иммуносупрессию следует понимать как снижение активности или эффективности иммунной системы. Иммуносупрессия специально индуцирована для профилактики, например, отторжения трансплантированных органов, лечения реакции трансплантат против хозяина после трансплантации костного мозга или для лечения аутоиммунных заболеваний, например ревматоидных артритов или болезни Крона.
В этом контексте иммуностимуляция также может относиться к воздействию на природу или характер иммунной реакции, либо к воздействию на иммунную реакцию, которая все еще развивается или созревает, или к модуляции характера устойчивой иммунной реакции.
Используемый в описании термин «вакцинация» относится к введению антигенного материала (вакцина) с получением иммунитета к заболеванию. Вакцины могут предотвратить или улучшить последствия инфицирования множеством патогенов, таких как вирусы, грибки, простейшие паразиты, бактерии, а также аллергические заболевания и астма, наряду с опухолями. Как правило, вакцины содержат один или более адъювантов, например иммуностимулирующие нуклеиновые кислоты, используемые для стимуляции иммунного ответа. Общепринято, что вакцины являются самым эффективным и самым экономичным способом профилактики инфекций и заболеваний.
Вводимый материал может, например, представлять живые, но ослабленные формы патогенов (бактерии или вирусы), убитые или инактивированные формы этих патогенов, очищенный материал, такой как белки, нуклеиновые кислоты, кодирующие антигены, или клетки, такие как клетки опухоли или дендритные клетки. В частности, недавно была разработана ДНК вакцинация. ДНК вакцинация работает за счет вставки (и экспрессии, запуска распознавания иммунной системой) ДНК, кодирующей антигены, в клетки человека или животного. Некоторые клетки иммунной системы, которые распознают экспрессированные белки, атакуют эти белки и клетки, экспрессирующие их. Одним из преимуществ ДНК вакцин является простота их получения и хранения. Дополнительно, ДНК вакцины имеют множество преимуществ по сравнению с традиционными вакцинами, включая способность индуцировать широкий ряд типов иммунных ответов.
Вакцинация может быть использована в качестве профилактики, позволяющей создать иммунитет против антигена у вакцинированного здорового индивидуума после воздействия антигена. В качестве альтернативы, терапевтическая вакцинация может вызвать усиленный ответ иммунной системы вакцинированного больного индивидуума, направляя иммунную систему индивидуума против антигенов. Обе, и профилактическая, и терапевтическая вакцинации могут быть применены для людей и животных.
Используемый в описании термин «генная терапия» относится к временной или постоянной генетической модификации (например, вставка, изменение или удаление генов) клеток и/или биологических тканей индивидуума для лечения заболеваний, таких как опухоли или аутоиммунные заболевания. Самая распространенная форма генной терапии включает вставку функциональных генов в неспецифическую геномную локализацию для замещения мутированного гена, но другие формы включают непосредственное корректирование мутации или модифицирование нормального гена, которое создает условия для транскрипции вирусов или даже перемещения гена или фрагмента гена в клетку для его транскрипции.
Используемый в описании термин «аутогенная генная терапия» относится к применению тканей или клеток самого индивидуума. Выделенные клетки или ткани модифицируют при использовании генной терапии и повторно вводят донору. В противоположность, «аллогенная генная терапия» относится к применению клеток для генной терапии от индивидуума иного, чем индивидуум-акцептор. После генной модификации аллогенные клетки вводят акцептору.
Используемый в описании термин «ex-vivo генная терапия» относится к терапии, при которой клетки от индивидуума, например гематопоэтические стволовые клетки или гематопоэтические клетки-предшественники генетически модифицируют ex vivo и затем вводят лечимому индивидууму. Используемый в описании термин «in-vivo генная терапия» относится к терапии, при которой клетки от индивидуума, например гематопоэтические стволовые клетки или гематопоэтические клетки-предшественники, генетически модифицируют in vivo при использовании вирусных векторов или других конструктов экспрессии.
Генная терапия также может быть классифицирована как «генная терапия с использованием клеток зародышевой линии» и «генная терапия соматических клеток». В случае «генной терапии с использованием клеток зародышевой линии» зародышевые клетки, то есть сперма и яйцеклетки, генетически модифицируют. Генетические изменения, как правило, интегрированы в их геномы. Следовательно, изменения, вызванные терапией, будут наследственными и перейдут последующим поколениям. Этот подход используют для лечения генетических заболеваний и наследственных заболеваний. В случае «генной терапии соматических клеток» терапевтические гены перемещают в соматические клетки индивидуума. Любые модификации и эффекты будут ограниченны только индивидуумом и не будут унаследованы потомками или последующими поколениями.
Используемый в описании термин «рак» относится к раковым заболеваниям или опухолям, лечимым или предотвращаемым, которые выбраны из группы, состоящей без ограничения из рака молочной железы, меланомы, неоплазии кожи, лимфомы, лейкимии, опухоли желудочно-кишечного тракта, включая, карциному толстой кишки, карциному желудка, карциному поджелудочной железы, колоректальный рак, рак тонкой кишки, карциному яичников, карциному шейки матки, рак легких, рак простаты, почечно-клеточную карциному и/или метастазы в печени.
Аутоиммунные заболевания по настоящему изобретению включают ревматоидный артрит, болезнь Крона, системную красную волчанку (SLE), аутоиммунный тиреоидит, тиреоидит Хашимото, множественный склероз, диффузный токсический зоб, миастению гравис, целиакию и болезнь Аддисона.
Настоящее изобретение относится к линейной с открытой цепью последовательности ДНК, содержащей по меньшей мере один CG мотив и по меньшей мере один нуклеотид в L-конфигурации. За счет частичной/полной L-конфигурации ДНК не может выступать в роли естественного субстрата для специфических ДНК расщепляющих в D-конфигурации ферментов. Следовательно, ДНК конструкты по настоящему изобретению защищены от ферментативного расщепления без какого-либо использования фосфоротиоатного скелета, который продемонстрировал свою токсичность. Дополнительно, ДНК конструкты состоят только из минимального числа нуклеотидов, которые делают их маленькими и, следовательно, значительно улучшают их проникновение в клетки пациентов.
Эффект CG-содержащих ДНК конструктов зависит от взаимодействия с TLR9, а взаимодействие ДНК с белком зависит от конфигурации обоих, и ДНК, и белка. Поскольку хиральность одиночных молекул является решающей для конфигурации получаемого в результате полимера, не известно, способна ли молекула ДНК в частичной или полной L-конфигурации связываться с TLR9 и активировать TLR9. Данные экспериментов показали, что неожиданно такие защищенные молекулы ДНК подходят для индукции иммунного ответа. Как показано в Примерах и на Чертежах, по меньшей мере частичное изменение хиральности одиночных нуклеотидов очевидно все еще позволяет связывание с TLR9 и активацию TLR9. Следовательно, молекулы ДНК с CG мотивами и нуклеотидами в L-конфигурации могут быть использованы для иммуномодуляции.
Неожиданно индуцированный паттерн стимуляции отличается от паттерна стимуляции, индуцированного молекулой, описанной в EP 1196178, имеющей гантелеобразную форму, содержащей CG мотивы в одноцепочечных петлях на обоих концах молекулы или двухцепочечном стебле («dSLIM»), как видно на Чертежах, даже при использовании идентичных последовательностей нуклеотидов.
ДНК конструкт может представлять одноцепочечные или частично или полностью двухцепочечные. Это включает в себя спаривание оснований в одной и той же молекуле (внутримолекулярно), или с другими молекулами (межмолекулярно), или в любой их комбинации. Также возможно, чтобы конструкт содержал по меньшей мере одну неспаренную одноцепочечную область. Один из вариантов воплощения настоящего изобретения включает шпилечные структуры. Ввиду частичной или полной L-конфигурации гарантирован более длительный период полураспада конструкта, поскольку нуклеотиды в L-конфигурации не подвержены расщеплению.
Также предполагается, что по меньшей мере две молекулы, которые представляют одноцепочечные или частично или полностью двухцепоченые, могут быть соединены друг с другом с образованием мультимерных конструктов. Таким образом, эти мультимерные конструкты включают по меньшей мере так много CG мотивов, что они лигируют партнеров, плотно упакованных в одной молекуле и, следовательно, вызывают значительный иммунный ответ. Полученные в результате одноцепочечные или частично или полностью двухцепочечные мультимерные конструкты могут представлять как ковалентно закрытые, содержащие нуклеотиды в L-конфигурации в молекуле, так и открытые мультимерные конструкты, содержащие нуклеотиды в L-конфигурации на или вблизи 5′-конца, или 3′-конца молекулы для защиты от ферментативного расщепления.
Согласно настоящему изобретению CG мотив/ы расположен/ы в одноцепочечной и/или двухцепочечной области конструкта. Как описано в EP 1196178, CG мотивы способны вызвать иммунный ответ, не зависимо от того, включены они в одноцепочечную или в двухцепочечную область молекулы.
Дополнительно, настоящее изобретение включает химические модификации по меньшей мере одного нуклеотида в L- или D-конфигурации с функциональной группой, выбранной из группы, состоящей из карбоксильной группы, аминной, амидной, альдиминной, кетальной, ацетальной, сложного эфира, простого эфира, дисульфидной, тиоловой и альдегидной групп. Это позволяет связываться ДНК конструкту с соединением, выбранным из группы, состоящей из пептидов, белков, липидов, везикул, мицелл, углеводов, антител, синтетических молекул, полимеров, микрочастиц, частиц металлов, наночастиц или твердых фаз, посредством, например, адсорбции, ковалентного или ионного связывания. Модификация может быть подходящим образом выбрана для соответствующих целей. Следовательно, конструкт может быть использован, например, для переноски других молекул в специфические клетки, реагирующие на введенные CG мотив/ы. Дополнительно, возможно использование таких модификаций для связывания конструкта с микрочастицами, которые могут быть использованы для перемещения конструкта в клетку. Также конструкт может быть связан с твердой фазой, например с микротитрационным планшетом.
Активация на основе Th1 включает активацию NK клеток и цитотоксических T-клеток, и эти ответы могут быть использованы для лечения рака. Поскольку ДНК конструкты, содержащие неметилированные CG мотивы, предпочтительно приводят к Th1 активации, конструкты по настоящему изобретению могут быть использованы для лечения рака. Продолжаются многочисленные клинические исследования применения TLR9 агонистов для лечения рака. Такие молекулы эффективно вводились по отдельности или в комбинации, например с радиационной терапией, хирургией, химиотерапией и криотерапией (Krieg, J. Clin. Invest. 2007 117:1184-94). Предполагается, что ввиду сильной иммуномодуляции, малых размеров и стабильности конструкты по настоящему изобретению будут иметь в этом отношении огромные преимущества. Дополнительно, их отличающийся иммунологический профиль отличает их от других менее преимущественных TLR9 лигандов, и этот профиль может быть использован для рак-специфического лечения.
С другой стороны, TLR9 агонисты также участвуют в образовании регуляторных Т-клеток и, следовательно, могут быть использованы для лечения аутоиммунных заболеваний. Путь введения, по-видимому, представляет одну из переменных, определяющих воздействие конструктов ДНК, содержащих CG мотивы in vivo (Krieg, J. Clin. Invest. 2007 117:1184-94).
Иммуностимулирующий эффект таких молекул ДНК, содержащих CG-мотивы, показал улучшение эффективности стандартных терапевтических подходов, таких как химеотерапия, при терапии рака. Следовательно, фармацевтические композиции, содержащие конструкты по настоящему изобретению, также входят в объем притязаний настоящего изобретения. Опять же преимущественные признаки конструктов по настоящему изобретению сравнимы с TLR9 агонистами предшествующего уровня техники, что делает конструкты по настоящему изобретению перспективными инструментами для лечения таких заболеваний, как рак, инфекционные заболевания, аллергии и астмы. Таким образом, лечение аллергий и астмы (главным образом Th2-опосредованных) выигрывает от преимуществ Th1 активации.
Поскольку TLR9 агонисты показали себя как сильные адъюванты в вакцинах, вакцины, содержащие ДНК конструкт, также входят в объем притязаний настоящего изобретения. Конструкты по настоящему изобретению содержат только релевантные последовательности для стимуляции TLR9, и они стабильны благодаря L-модификации нуклеотида. Следовательно, отсутствуют побочные эффекты, вызываемые нерелеватными последовательностями. Более длительный период полураспада молекулы гарантирует эффективную стимуляцию, вызывая, таким образом, сильный иммунный ответ.
ДНК молекулы по настоящему изобретению получают при использовании колоночного метода синтеза и соответствующих нуклеотидов (Бета-L-дезокси «NT» (n-bz) CED фосфорамидит; «NT» цепочки для аденозина, цитидина, гуанозина или тимидина). Затем ДНК молекулы подвергают ВЭЖХ очистке.
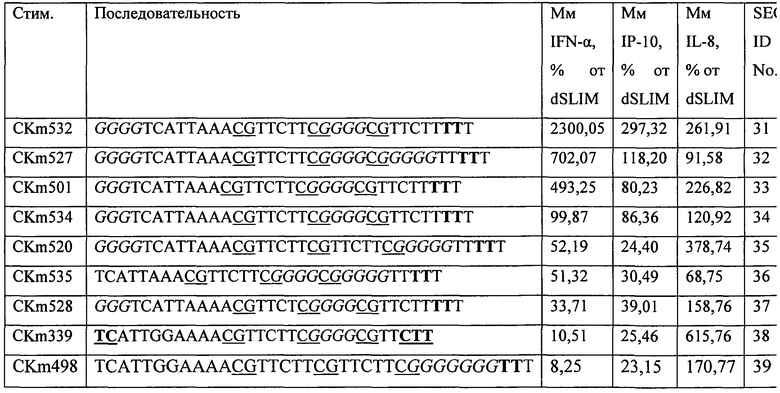
Для выявления эффекта использования ДНК с L-рибозой вместо D-рибозы были использованы следующие молекулы ДНК для приведенных в описании первоначальных экспериментов (Таблица 1).


Эксперименты с использованием последовательностей Таблицы 1 показали, что линейные последовательности, содержащие CG мотивы, защищенные L-рибозой, способны стимулировать иммунную систему и индуцировать иммунный явно выраженный ответ, отличающийся от иммунного ответа, индуцированного dSLIM, как описано в EP 1196178. Следовательно, модифицированную последовательность, названную ODN2216 (GGGGGACGATCGTCGGGGGG; SEQ ID 6) и ее модификацию использовали для исследования воздействия структурных различий, таких как влияние присутствия G-повторов, длина и положение, расстояние между CG-мотивами и расстояние между нуклеотидами L-рибозы и CG-мотивами соответствующих G-повторов.
В таблице 2 приведены последовательности и их воздействие на секрецию IFN-альфа и IP-10 по сравнению с ODN2216, имеющие первые два и последние шесть нуклеотидов, модифицированные фосфортиоатами, где выделенные жирным шрифтом буквы представляют нуклеотиды, содержащие L-рибозу, выделенные курсивом буквы относятся к G-повтору, а выделенные подчеркиванием буквы относятся к CG-мотиву. Пунктирная линия находится на месте расположения соответствующей последовательности в месте для сравнения с CKm508, но не указывает ни на структурную, ни на функциональную модификацию последовательности.


Хорошие результаты были получены с последовательностями, имеющими по меньшей мере один G повтор, в частности на или вблизи 3-′ конца. Дополнительно, стимуляция зависит от присутствия CG-мотивов (CKm477), что показывает, что стимуляция не является следствием воздействия L-рибозы, а является следствием воздействия CG мотивов.
Результаты, полученные при использовании модифицированных последовательностей ODN 2216, определяющих положительные структурные компоненты, перемещены в последовательность ДНК CKm374. В Таблице 3 приведены результаты использования модифицированных последовательностей по сравнению с гантелеобразной dSLIM, описанной в EP 1196178. Выделенные жирным шрифтом буквы представляют нуклеотиды, содержащие L-рибозу, выделенные курсивом буквы относятся к G-повтору, а выделенные и подчеркнутые буквы относятся к фосфортиоат модифицированным нуклеотидам и подчеркнутые буквы относятся к CG-мотиву.
Как видно из результатов, приведенных в Таблице 3, G-повтор, расположенный непосредственно на 5′-конце, по-видимому, дает преимущества (сравнение CKm532 и CKm499). Кроме того, использование четырех вместо трех дезоксигуанозинов на 5′-конце дополнительно увеличивает стимуляцию IFN-альфа и IP-10 (сравнение CKm501 и CKm532).
Добавление дополнительного G-повтора между CG-мотивами, по-видимому, также дает преимущества (сравнение CKm532 и CKm520). Расстояние между первым и вторым G-повтором дополнительно оказывает воздействие на эффективность ДНК молекулы. Дополнительно, использование дезоксинуклеотидов, содержащих L-рибозу, только на или вблизи 3′-конца, по-видимому, позволяет достичь достаточной степени стабилизации ДНК молекулы. Наблюдается предполагаемая хорошая стимуляция IFN-альфа и IP-10 (см. ниже). Поскольку IL-8 ответственен за индукцию неоангиогенеза, по-видимому, будет эффективно, чтобы секреция IL-8 индуцировалась только в малых количествах.
Ясно, что присутствие и тщательный выбор положения G-повторов в комбинации со стабилизирующим воздействием дезоксинуклеотидов, содержащих L-рибозу, позволяет получить молекулы ДНК, подавляющие стимулирующую эффективность dSLIM молекулы.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Далее описание будет приведено со ссылкой на иллюстрирующие Примеры, Чертежи и неограничивающие варианты воплощения настоящего изобретения.
Фигура 1 - электрофорез на агарозном геле ДНК конструктов после ферментативного расщепления.
Фигура 2 - интенсивность GFP после стимуляции клеточной линии макрофагов мыши.
Фигура 3 - концентрация MIP-1 альфа после стимуляции плазмацитоидных дендритных клеток (PDC).
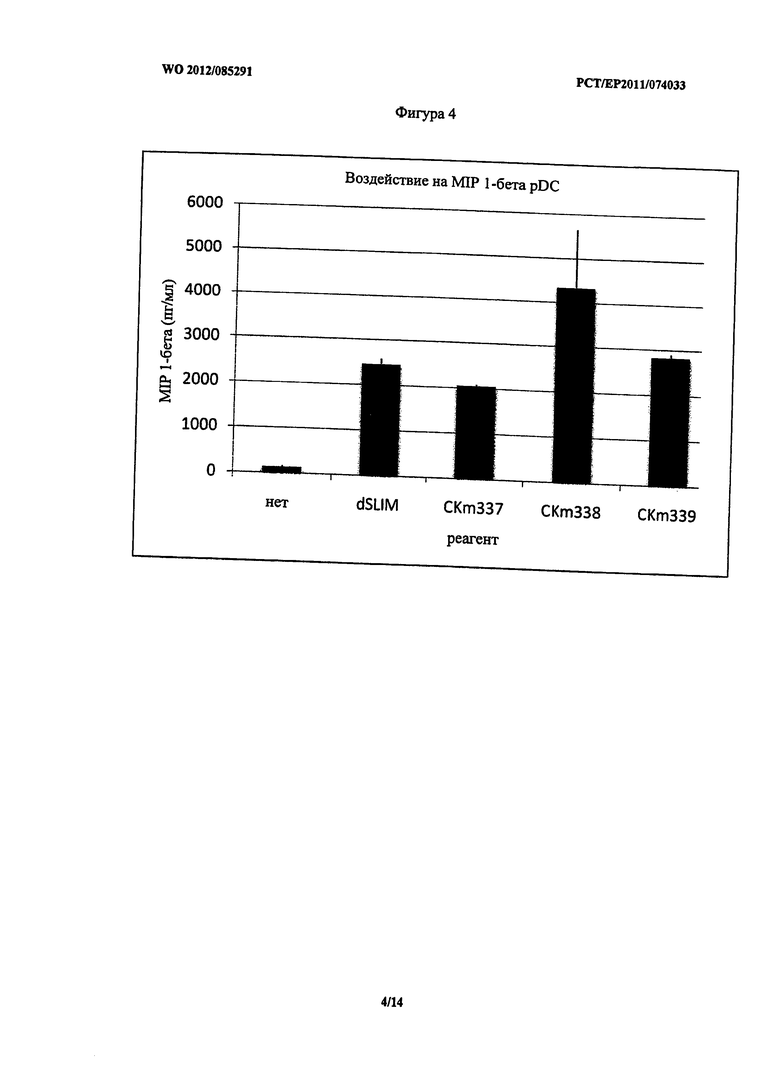
Фигура 4 - концентрация MIP-1 бета после стимуляции PDC.
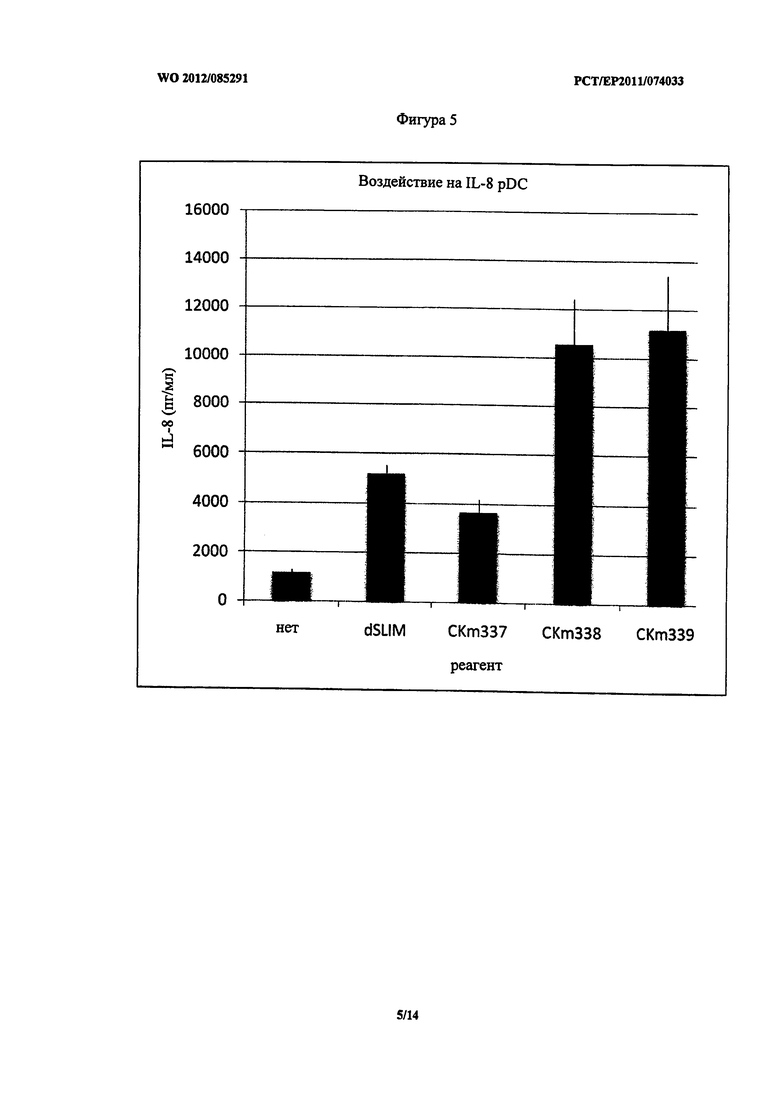
Фигура 5 - концентрация IL-8 после стимуляции PDC.
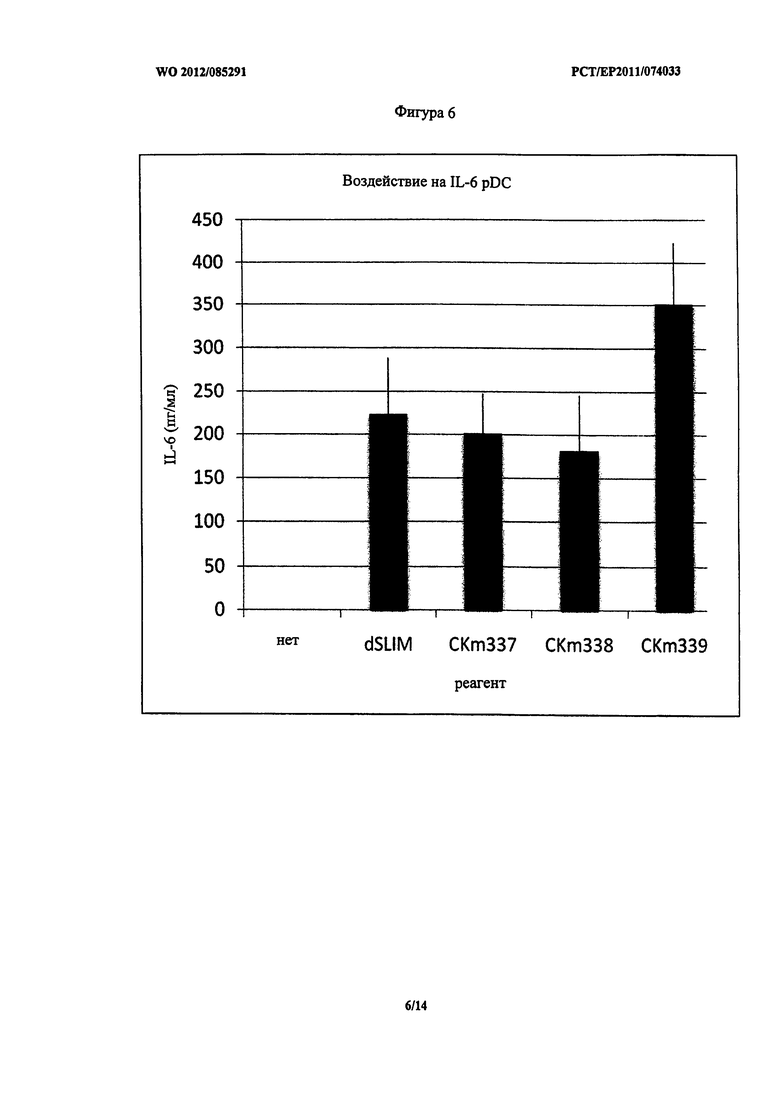
Фигура 6 - концентрация IL-6 после стимуляции PDC.
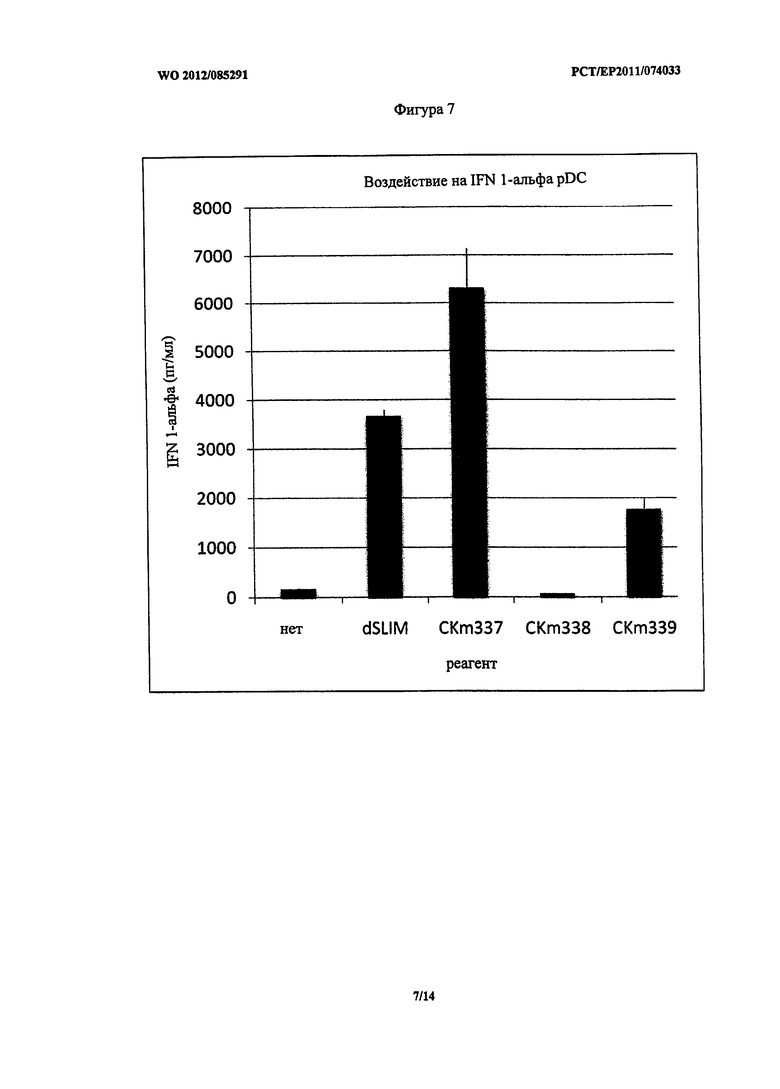
Фигура 7 - концентрация IFN-альфа после стимуляции PDC.
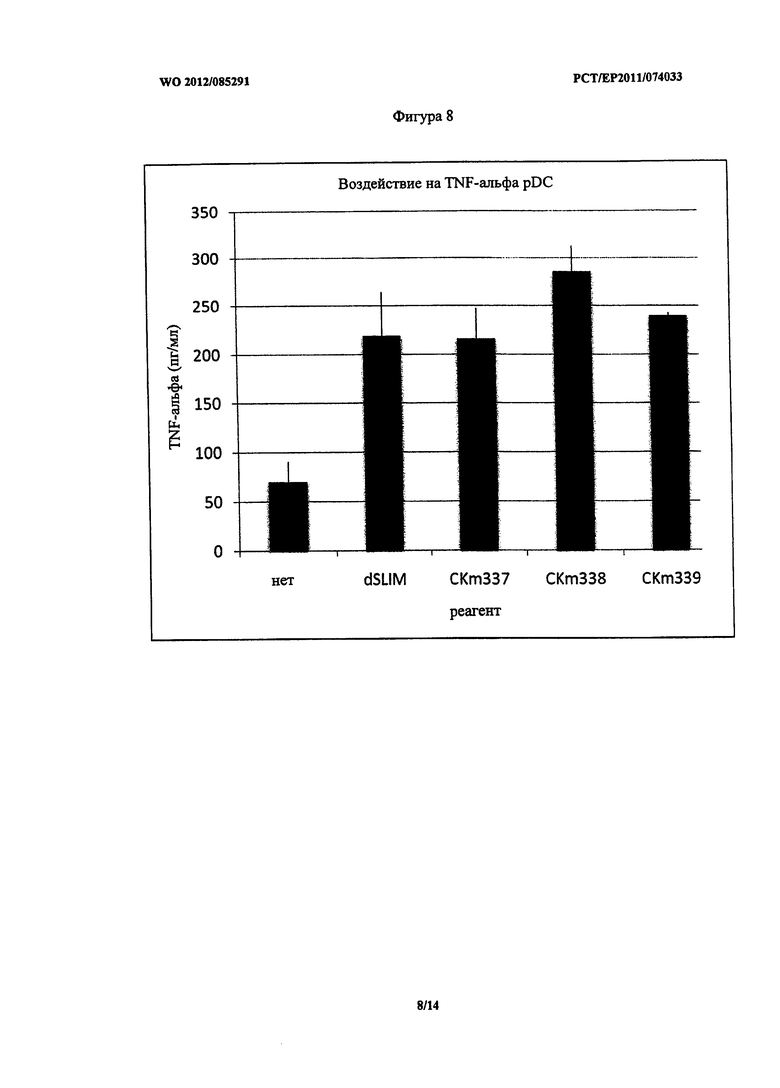
Фигура 8 - концентрация TNF-альфа после стимуляции PDC.
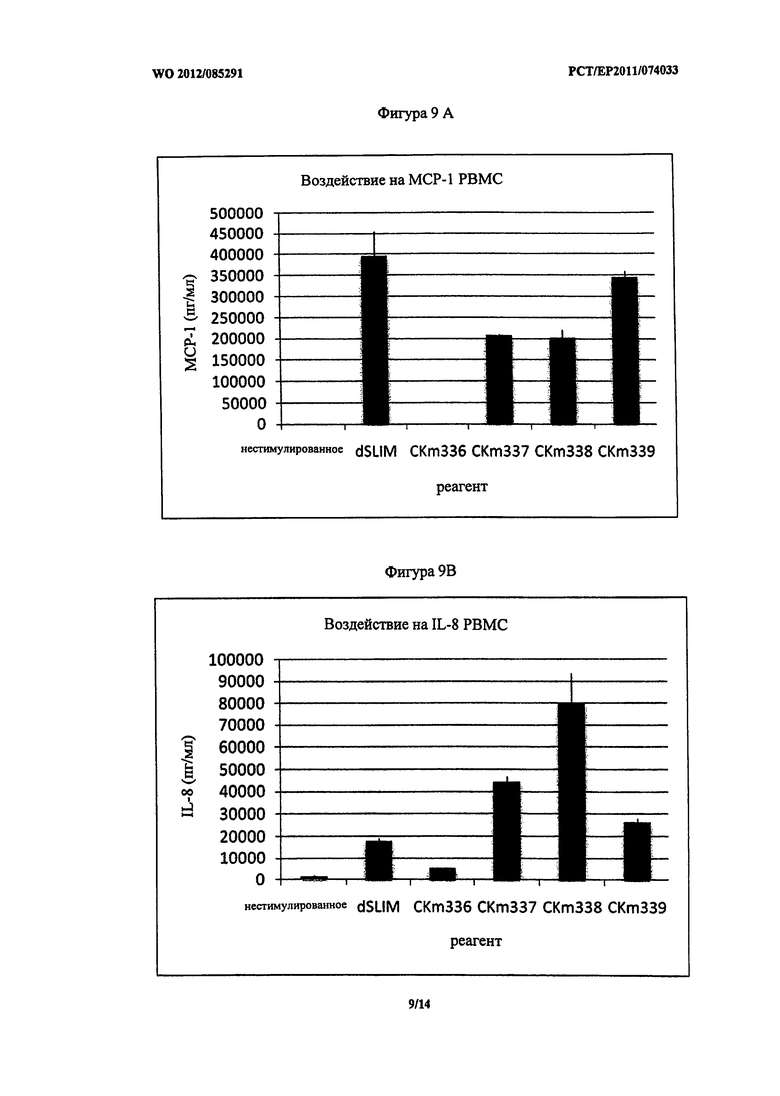
Фигура 9 - концентрация MCP-1 и IL-8 после стимуляции мононуклеарных клеток периферической крови (PBMC).
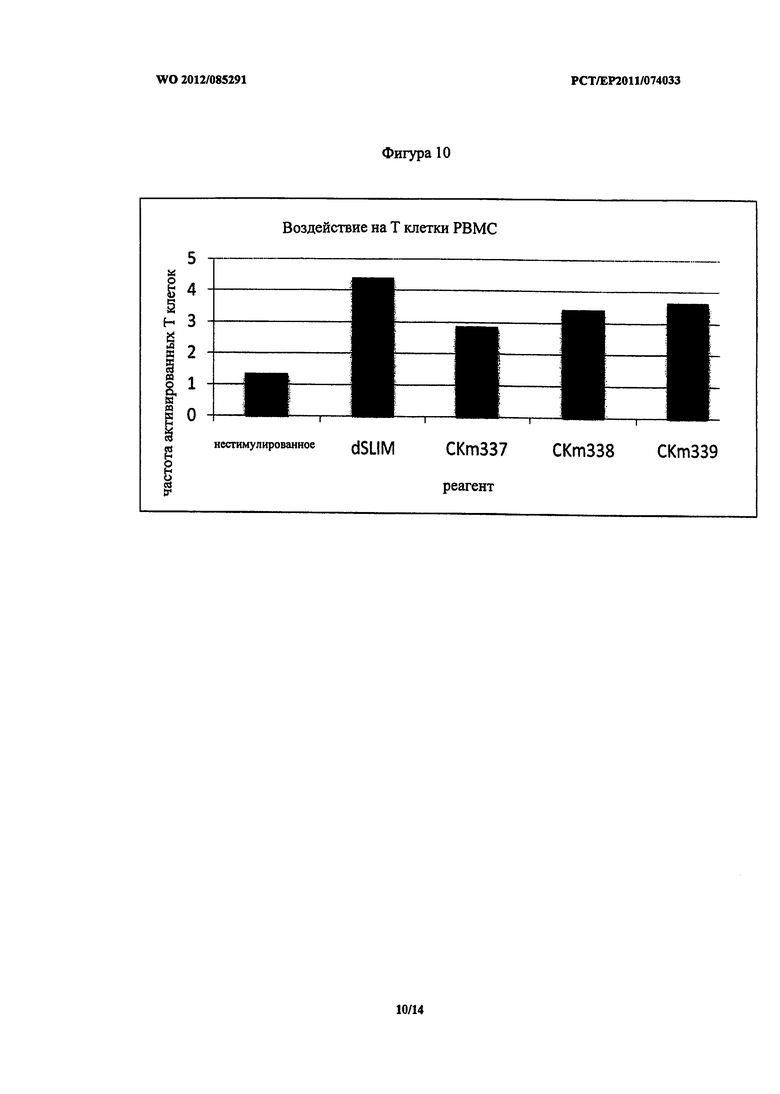
Фигура 10 - частота активированных T-клеток после стимуляции PBMC.
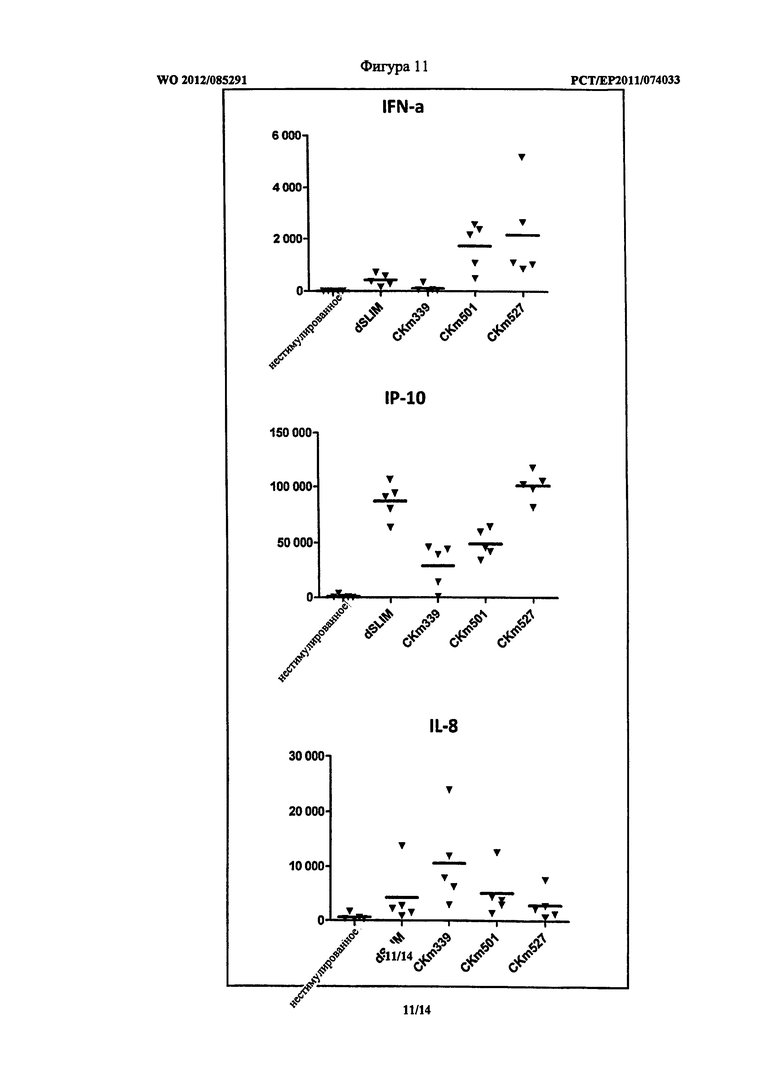
Фигура 11, 12 - секреция IFN-альфа, IP-10 и IL-8 клетками PBMC.
Фигура 13 - воздействие терминальных дезоксинуклеотидов, модифицированных L-рибозой, на стимуляцию ELAM9 клеток.
Фигура 14 - иммунная стимуляция B-клеток и PDC при использовании CKm532 и dSLIM по сравнению с нестимулированным состоянием.
ДЕТАЛЬНОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фигуре 1 приведен гель всех ДНК конструктов после расщепления T7-Полимеразой из Т7 бактериофага. 6 мкг каждого ДНК конструкта инкубировали с 10 единицами T7-Полимеразы (общий реакционный объем: 20 мкл). Через 0, 1, 2, 5, 30, и 1500 минут аликвоту 3 мл инкубируемой смеси удалили из образца и развели 5 мкл формамида, содержащего краситель Sanger. Все аликвоты были помещены в 3%-ный агарозный гель, через который в течение 40 минут пропускали ток 100 В.
Было обнаружено, что не модифицированная молекула ДНК линии 30L2 (дорожка 2) полностью расщеплена после 5 минут инкубации с Т7-Полимеразой, в то время как конструкт по настоящему изобретению (CKm337; дорожка 3), наряду с dSLIM (дорожка 1) и фосфортиоат модифицированными конструктами CKm338 (дорожка 4) и CKm339 (дорожка 5), сохранился в значительной степени даже после 1500 минут инкубации. Действительно, CKm337 показала высочайшую стабильность всех протестированных молекул. Ввиду недостаточной стабильности линия 30L2 была исключена из дальнейшего исследования.
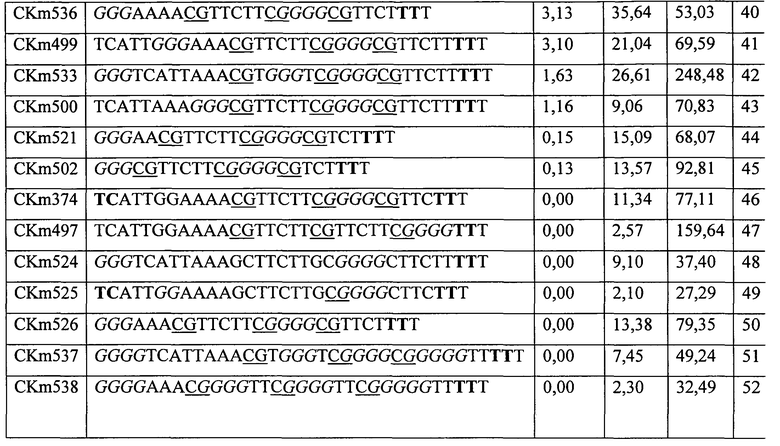
На Фигуре 2 приведена стимуляция ELAM9 клеток различными стимулирующими ДНК конструктами. ELAM9 клетки представляют TLR9-клетки положительных макрофагов мыши (RAW264), которые были стабильно трансфектированы d1-eGFP под контролем промотора человеческого эластина (hELAM), содержащего несколько NFκB элементов отклика. Через один день после высевания клеток их стимулировали описанными конструктами ДНК (3 мкМ) в течение 7 часов. Среднее Geo интенсивности GFP измерили при использовании проточной цитометрии.
ДНК конструкт со всеми нуклеотидами в L-конфигурации за исключением последнего T (CKm336) не имеет стимулирующей способности. Однако ДНК конструкт с нуклеотидами в L-конфигурации на обоих концах (CKm337) не стимулирует экспрессию GFP. Это было довольно неожиданно, поскольку не было известно, способны ли ДНК конструкты, содержащие CG мотивы с нуклеотидами в L-конфигурации, связываться и активировать TLR9. Дополнительно предполагается, что CKm337 будет поглощена клетками более легко, чем dSLIM (молекула, описанная в EP 1196178), и менее токсична, чем фосфортиоат модифицированные конструкты (CKm338 и CKm339).
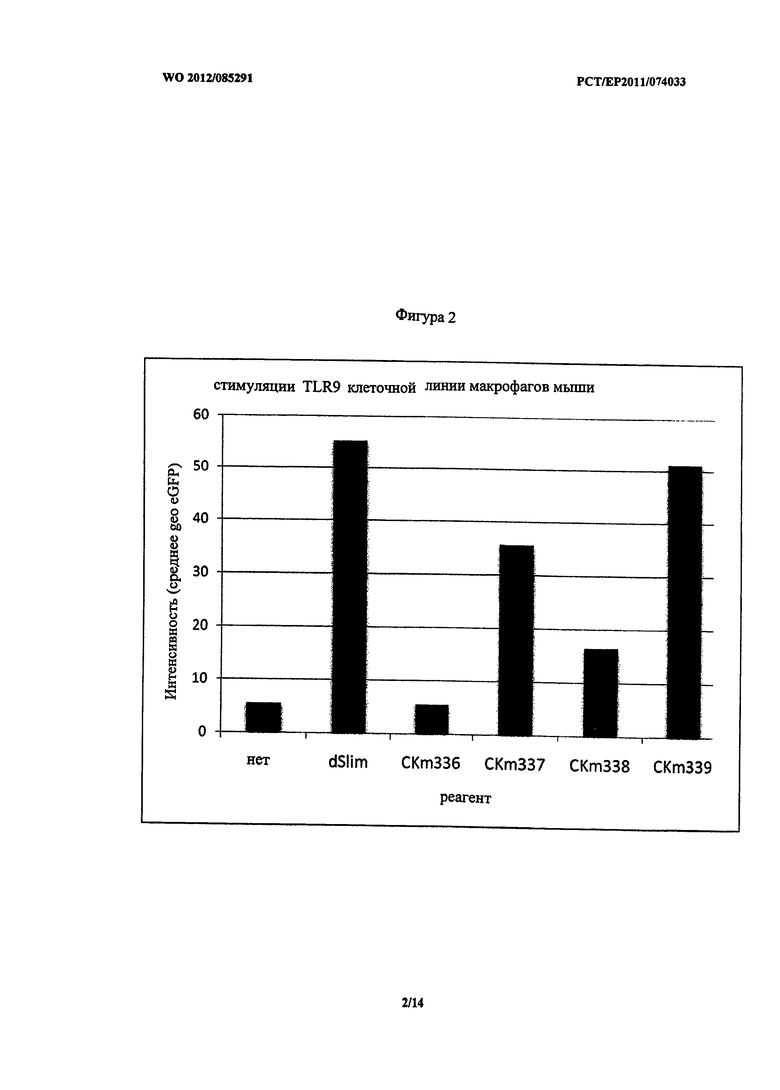
На Фигурах 3-8 показано воздействие ДНК конструктов на pDC в отношении секретированных хемокинов и цитокинов. pDC были обогащены PBMC, очищенной фиколлом, при использовании комбинированной процедуры сортировки с использованием набора Miltenyi, Diamond PDC Kit: сначала PBMC был обеднен не-pDC при использовании набора pDC Biotin-Antibody Cocktail от Miltenyi′s, затем проводили положительную сортировку клеток для pDC при использовании CD304 (BDCA-4) алмазных микрогранул от PDC Diamond Kit. PDC высевали 2,5×105/мл с 10 нг/мл IL-3 в среде (RPMI1640, 10%-ной фетальной телячьей сыворотки, 100 единиц/мл пенициллина, 100 единиц/мл стрептомицина 2 мМ глютамина, 37°C, 5% CO2) и стимулировали в течение 2 дней при использовании 3 мкм индивидуальных конструктов.
Для определения количества секретированных соединений при стимуляции клеток очищенный супернатант стимулированных клеток собирали и анализировали при использовании мультиплексной системы (FlowCytomix от eBioscience/Bender MedSystems) или ELISA.
Неожиданно pDC, стимулированные CKm337, показали аналогичное воздействие на секрецию MIP-1 альфа, 1 бета и IL-8 по сравнению со стимуляцией dSLIM. Секреция MIP-1 альфа, 1 бета и IL-8 при стимуляции lin CKm338 и CKm339 была немного выше (Фигуры 3, 4 и 5). Однако все фосфортиоат модифицированные конструкты унаследовали некоторые из указанных выше недостатков.
В отношении секреции IL-6 dSLIM, CKm337 и CKm338 имели аналогичное воздействие на pDC. CKm339 был немного более эффективен (Фигура 6).
Следует отметить, что CKm337 оказал неожиданно более сильное воздействие на секрецию IFN-альфа клетками pDC по сравнению со всеми другими линейными конструктами (Фигура 7).
dSLIM, CKm337, CKm338 и CKm339 все оказывали аналогичное воздействие на секрецию TNF-альфа клетками pDC. (Фигура 8).
PBMC выделили из лейкоцитарной пленки человека при использовании градиента плотности Фиколла. Для функционального анализа 106 клеток/мл в среде (RPMI1640, 10%-ной фетальной телячьей сыворотки, 100 единиц/мл пенициллина, 100 единиц/мл стрептомицина 2 мМ глютамина, 37°C, 5% CO2) стимулировали в течение 2 дней при использовании индивидуальных соединений, примененных в указанных концентрациях (2-3 мкм).
На Фигуре 9 (A и B) приведено воздействие описанных ДНК конструктов (3 мкМ каждого) на PBMC в отношении секреции МСР-1 и IL-8. Как ожидалось от экспериментов с pDC, ДНК конструкты со всеми нуклеотидами в L-конфигурации (CKm336) не имели стимулирующей способности при применении в PBMC. Однако CKm337 был эффективным в вызывании обоих, секреции МСР-1 и секреции IL-8. Неожиданно его воздействие в отношении секреции IL-8 было более сильным по сравнению с dSLIM и менее сильным в отношении секреции MCP-1.
Для определения клеточной субпопуляции и статуса активации характерные маркеры поверхности были помечены селективными флюорофорконъюгированными антителами. Окрашивание антителами проводили при 106 клетках/окрашивающий набор; каждый набор инкубировали с 4 различными антителами, соединенными с флюорофорными группами, наконец, ресуспендировали в 400 мкл FACS буфере и проводили анализ при использовании проточной цитометрии по меньшей мере на 100000 живых клеток. При использовании стратегии гейтов для определения T-клеток и активированных в них клеток оно составило CD3+/CD56- с маркером активации CD69.
На Фигуре 10 приведено воздействие описанных ДНК конструктов (2 мкМ каждый) на частоту активированных T-клеток в популяции PBMC. Все пять конструктов имели сравнимую стимулирующую способность. T-клетки не экспрессировали TLR9. Следовательно, при стимуляции ДНК конструктами популяции клеток PBMC происходит их активация, что в свою очередь вызывает активацию T-клеток.
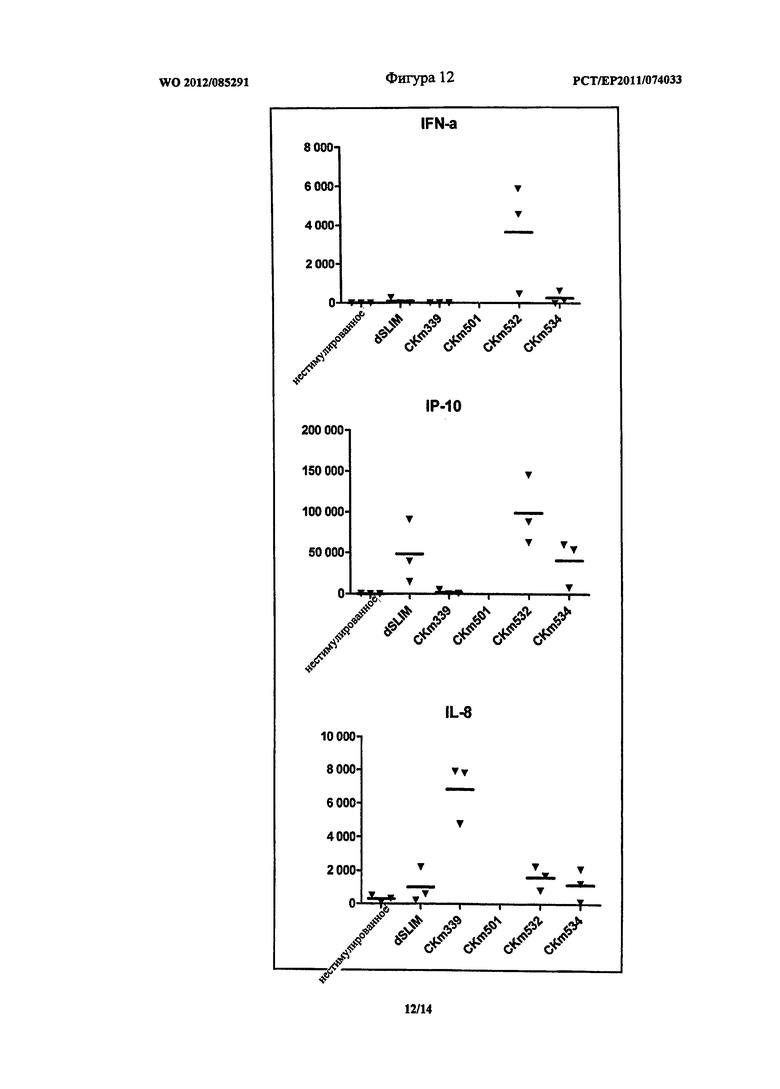
Оптимизация последовательностей показала, что введение G-повторов повышает эффективность олигонуклеотидов после трансфекции. Эффективность дополнительно зависит от расстояния между CG-мотивами. Линейные ДНК конструкции могут быть достаточно защищены от расщепления при использовании дезоксинуклеотидов, содержащих L-рибозу, на 3′-конце олигонуклеотида (сравнение Таблица 2 и 3). Олиго CKm501 (SEQ ID NO: 33), CKm527 (SEQ ID NO: 32), CK 532 (SEQ ID NO: 31) и CKm534 (SEQ ID NO: 34) показали неожиданно хорошие результаты, как можно видеть из Таблицы 3. На Фигурах 11 и 12 приведено воздействие описанных ДНК конструктов на секрецию цитокинов IFN-альфа (верхняя часть), IP-10 (средняя часть) и IL-8 (нижняя часть) в PBMC. Эксперименты проводили, как указано выше.
На Фигуре 11 показано, что CKm501 и CKm527 вызвали повышенные уровни секреции IFN-альфа и CKm527 и также повышение секреции IP-10 по сравнению с гантелеобразной dSLIM. Секреция IL-8 сравнительно низкая относительно dSLIM, но ниже по сравнению с CKm339, которая представляет последовательность одноцепочечных петель гантелеобразной dSLIM, защищенной с обоих концов фосфортиоат модифицированными дезоксинуклеотидами.
Как видно из Фигуры 12, CKm532 демонстрирует значительную и неожиданно высокую индукцию секреции IFN-альфа и IP-10, но сравнительно низкую индукцию секреции IL-8. Следовательно, CKm532 подтверждает, что структурный элемент G-повтора, расположенный непосредственно на 5′-конце, и дополнительный G-повтор, расположенный между двумя CG-мотивами (второй и третий GC-мотив), дает преимущества. Сравнение CKm520 и CKm532 в Таблице 3 указывает, что расположение G-повтора между вторым и третьим CG-мотивом в CKm532 отвечает за предположительное повышение секреции IFN-альфа и IP-10, при этом CKm520, главным образом, повышает секрецию IL-8. Дополнительно, считается достаточной защита олигонуклеотидов только двумя L-рибозами, содержащимися в дезоксинуклеотидах на 3′-конце.
Укорачивание G-повтора на 5′-конце в результате приводит к снижению эффективности по сравнению с CKm532 и CKm534 на Фигуре 12. Опять же CKm532 демонстрирует преимущества определенных структурных компонентов в отношении повышенной секреции IFN-альфа и IP-10 и пониженной секреции IL-8.
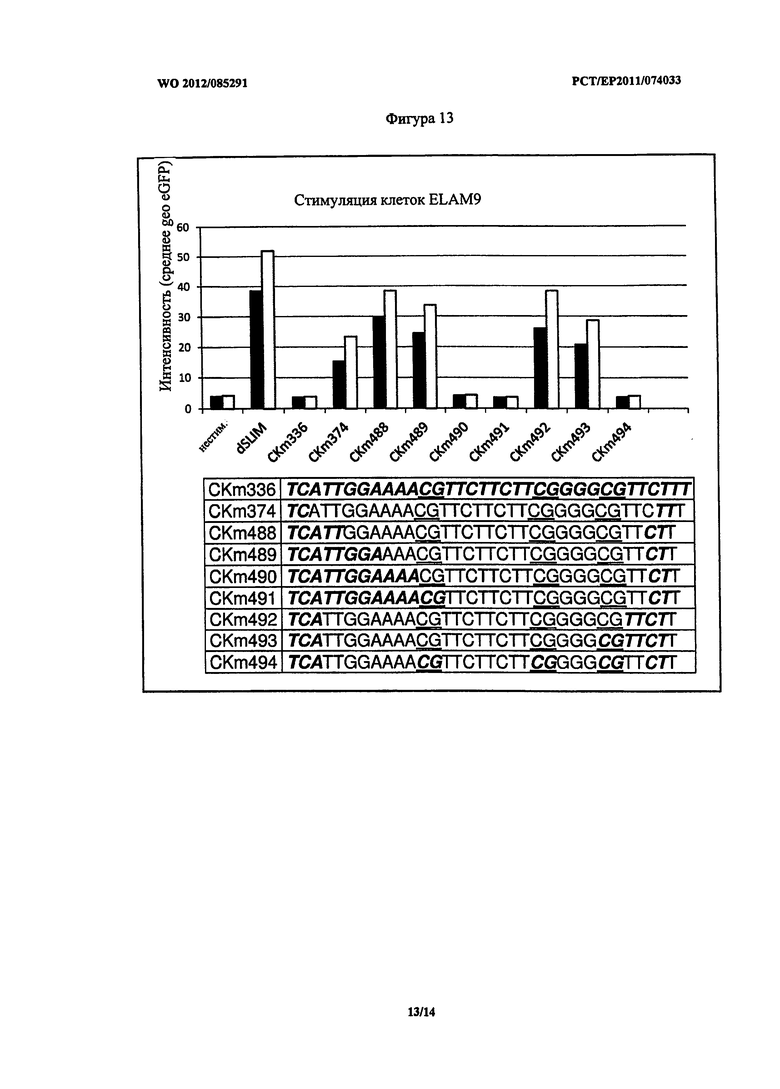
В верхней части Фигуры 13 приведены результаты стимуляции клеток ELAM9 описанными конструктами ДНК, которые содержат дезоксинуклеотиды с различной степенью модификации L-рибозой. В нижней части Фигуры 13 представлены последовательности нуклеотидов, нуклеотиды, содержащие L-рибозу, выделены жирным шрифтом. Эксперименты проводили дважды (L-dSLIM032 и L-dSLIM030).
Степень и положение дезоксинуклеотидов, содержащих L-рибозу, оказывают воздействие на стимуляцию клеток ELAM9. Полная последовательность в L-конфигурации (CKm336; SEQ ID NO: 2) вообще не оказывает никакого стимулирующего воздействия, как указано в описании WO 2010/039137. Хорошие эффекты достигнуты при использовании CG-мотива, содержащего олигонуклеотиды, защищенные дезоксинуклеотидами, содержащими L-рибозу, на 3′- и 5′-конце, при этом количество дезоксинуклеотидов, содержащих L-рибозу, на 3′- и 5′-концах, имеет обратный эффект (сравнение CKm489 и CKm490). Дополнительно, модификация CG-мотивов дезоксинуклеотидами, содержащими L-рибозу, приводит к потере воздействия. Следовательно, для достижения стимулирующего воздействия CG-мотивы не должны содержать L-рибозу, а количество дезокинуклеотидов, модифицированных L-рибозой, на обоих концах следует ограничивать, а именно не более чем восемь терминальных дезоксинуклеотидов на 5′- и максимально 3′-терминальных дезоксинуклеотидов, следующих за последним CG-мотивом.
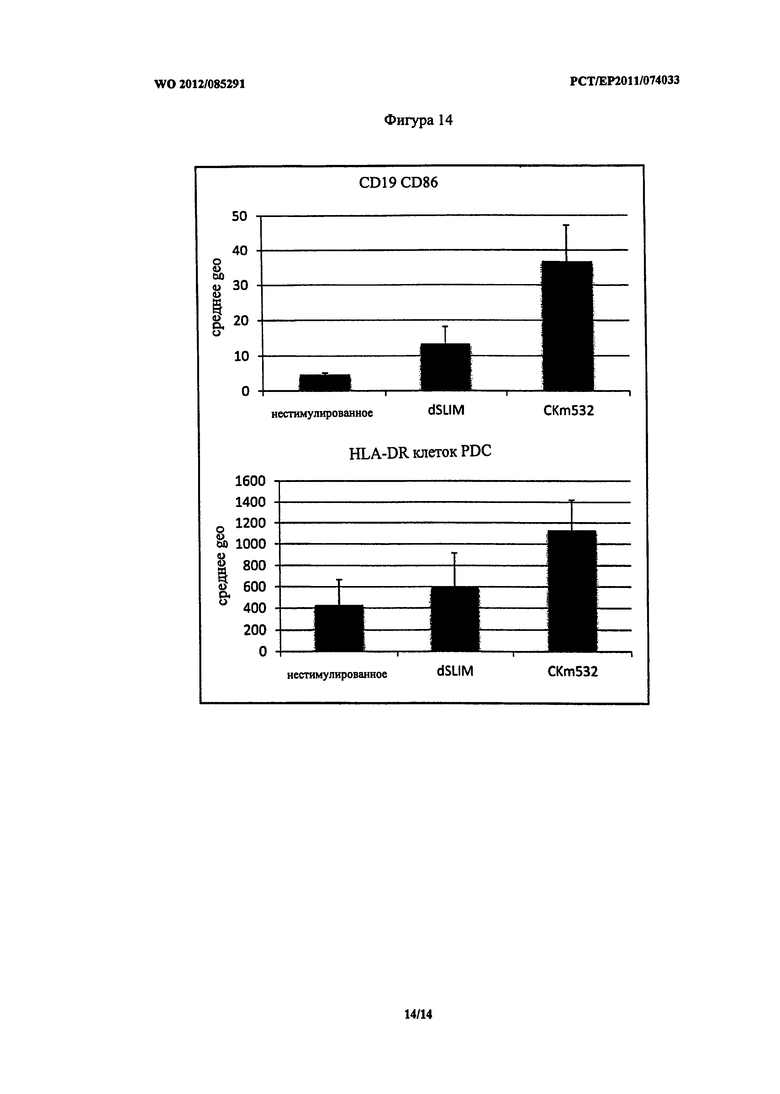
На Фигуре 14 приведена иммунная стимуляция при использовании CKm532 и dSLIM по сравнению с не стимулированным состоянием. FACS эксперименты были проведены по протоколу, использованному для экспериментов, приведенных на Фигуре 10, и адаптированы для B клеток (стратегия гейтов: CD 19 положительные, CD86 в качества маркера активации) и PDC (стратегия гейтов: клетки негативной линии дифференцировки, HLA-DR положительные, CD123 положительные клетки, CD40 и клетки негативной линии дифференцировки HLA-DR в качестве маркера активации) соответственно. Приведенные данные основаны на измерениях трех различных лейкоцитарных клеток человека.
В верхней части Фигуры 14 приведена стимуляция В клеток, что подтверждено маркером CD86. Очевидно, что CKm532 вызывает повышенную стимуляцию В клеток по сравнению с dSLIM и не стимулированным состоянием. Это показывает увеличение созревания В клеток, таких как клетки, продуцирующие антитела, что является очень важным признаком иммунной стимуляции.
В нижней части Фигуры 14 приведена стимуляция PDC, как определено при использовании маркера HLA-DR. HLA-DR является частью МНС молекул и, следовательно, частью антиген-презентирующих процессов иммунной системы. Опять же CKm532 вызывает большее проявление этого признака иммунной стимуляции по сравнению с dSLIM или не стимулированными клетками.
В заключение, CKm337 (D-ДНК конструкт с нуклеотидами в L-конфигурации на обоих концах) неожиданно продемонстрировал стимулирующее воздействие на оба, и на PBMC, и на выделенные pDC, в то время как ДНК конструкт со всеми нуклеотидами в L-конфигурации (CKm336) не оказывает такого воздействия. По-видимому, конфигурация CKm337 все еще позволяет соединение с TLR9, и CKm336 является неспособным стерически связываться с или стимулировать TLR9.
Неожиданно, паттерн стимуляции, индуцированный CKm337 по сравнению с гантелеобразной dSLIM и фосфортиоат модифицированными олиго, был уникальным по сравнению со всеми другими конструктами. CKm337 индуцировал самые высокие количества IFN-альфа, секретированных клетками pDC. Секреция IL-8 клетками PBMC слабее по сравнению с фосфортиоат модтифицированными молекулами, но сильнее по сравнению с dSLIM. В противоположность dSLIM индуцировал более высокое количество МСР-1, секретированных клетками PBMC, но Ckm337 сравнимо с фосфортиоат модифицированными молекулами.
Можно усилить воздействие, наблюдаемое у CKm337, за счет введения так называемых G-повторов непосредственно на 5′-конце линейной молекулы ДНК. Дополнительно, оказалось, что защиты от расщепления при использовании дезоксинуклеотидов, содержащих L-рибозу, на 3′-конце достаточно для стабилизации олигонуклеотидов. Определенные структурные признаки: G-повтор, CG-мотивы, расстояние между CG-мотивами и защита при использовании различных степеней и положений модифицированных L-рибозой дезоксинуклеотидов, позволяет модулировать иммуностимулирующее воздействие дезоксинуклеотидов, содержащих L-рибозу. Вполне очевидно, что настоящее изобретение предоставляет новые инструменты для конструирования иммунностимулирующих конструктов ДНК для целевой стимуляции клеток или иммунной системы.
IFN-альфа давно известен как антивирусный цитокин. Он стимулирует развитие Th1 клеток, следовательно, промотирует воздействие CG-содержащих ДНК молекул. Также IFN-альфа демонстрирует противоопухолевую активность при злокачественных опухолях у мышей и людей и способен снижать опухолегенность трансплантированных опухолевых клеток по существу за счет активации цитотоксичных T клеток и, следовательно, повышает вероятность цитолиза опухолевых клеток. Активность NK клеток и макрофагов, обоих, также важна для противоопухолевой цитотоксичности, также повышенной за счет IFN-альфа (Brassard et al., J. Leukoc. Biol. 2002 71:565-81). Следовательно, повышение количества IFN-альфа при стимуляции ДНК конструктами по настоящему изобретению предполагает положительный эффект для лечения рака.
Недавно было установлено, что IP-10 представляет сильный ангиостатический белок. Следовательно, индуцирование IP-10 по существу при лечении опухолевых заболеваний также может быть полезным.
IL-8 представляет провосполительный цитокин, который известен как медиатор активации и миграции нейрофилов в ткани из периферической крови. Возникшая в результате нейрофильная инфильтрация по существу может быть ответственна за ингибирование роста опухоли, как было показано при раке яичников (Lee et al., J. Immunol. 2000 164:2769-75). Дополнительно, IL-8 также является хемотаксическим для T-клеток и базофил. Следовательно, при лечении или профилактике по меньшей мере некоторых типов опухолей преимуществом обладает селективная повышающая регуляция IL-8 в ответ на CG-содержащие ДНК конструкты. С другой стороны, было установлено, что IL-8 запускает ангиогенез, таким образом, что индукция секреции IL-8 может быть контрпродуктивной. Следовательно, дифференцирование степеней индукции IL-8 за счет различных ДНК молекул по настоящему изобретению позволяет получить молекулу, адаптированную для заданного терапевтического воздействия.
Известно, что MCP-1 играет роль в рекрутинге моноцитов/макрофагов для мест повреждения и инфекции и, таким образом, возможно участвует в стимуляции антиопухолевого ответа хозяина. Было установлено, что MCP-1 может активировать моноциты, делая их более цитотоксичными против нескольких типов опухолевых клеток человека in vitro (Zachariae et al., J. Exp. Med. 1990 171:2177-82). Следовательно, аналогично IL-8 эффективно осуществлять модуляцию экспрессии MCP-1 в зависимости от конкретного типа опухоли.
Следовательно, специфический индуцированный паттерн цитокина является положительным для лечения и профилактики опухолей различных типов. Очевидно, что специфический контекст, в котором неметилированный CG-мотив представлен для TLR9, определяет индивидуальный соответствующий паттерн стимуляции, индуцированный в реактивных клетках.
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕКОДИРУЮЩИЙ ИММУНОМОДУЛИРУЮЩИЙ КОНСТРУКТ ДНК С КОВАЛЕНТНО ЗАКРЫТОЙ СТРУКТУРОЙ | 2015 |
|
RU2668387C2 |
| КОМБИНАЦИЯ, ВКЛЮЧАЮЩАЯ ИММУНОСТИМУЛИРУЮЩИЕ ОЛИГОНУКЛЕОТИДЫ | 2016 |
|
RU2766693C2 |
| ПРЕДИКТИВНЫЙ БИОМАРКЕР ДЛЯ ТЕРАПИИ РАКА | 2014 |
|
RU2672358C2 |
| КОНКАТЕМЕРЫ ДЛЯ ИММУНОМОДУЛЯЦИИ | 2010 |
|
RU2556119C2 |
| АЛЛОГЕННОЕ ПРОТИВООПУХОЛЕВОЕ ТЕРАПЕВТИЧЕСКОЕ СРЕДСТВО | 2004 |
|
RU2354694C2 |
| CPG-ОЛИГОНУКЛЕОТИДНЫЕ АНАЛОГИ, СОДЕРЖАЩИЕ ГИДРОФОБНЫЕ Т-АНАЛОГИ С УСИЛЕННОЙ ИММУНОСТИМУЛИРУЮЩЕЙ АКТИВНОСТЬЮ | 2007 |
|
RU2477315C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ ФОСФОРТИОАТНЫЕ CpG-ОЛИГОНУКЛЕОТИДЫ, СОДЕРЖАЩИЕ ФОСФОДИЭФИРНЫЕ СВЯЗИ, СПОСОБ ИММУНОМОДУЛЯЦИИ, СПОСОБ СТИМУЛИРОВАНИЯ ИММУННОГО ОТВЕТА | 2003 |
|
RU2338750C2 |
| Иммуногенная композиция, включающая ее вакцина, набор для приготовления вышеуказанной композиции и способ лечения заболеваний, связанных с патологией секреции гастрина | 2014 |
|
RU2664197C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ ОЛИГОНУКЛЕОТИДЫ | 2009 |
|
RU2477753C2 |
| Иммуностимулирующие олигонуклеотиды | 2012 |
|
RU2610690C2 |

Изобретение относится к биохимии. Описан некодирующий ДНК конструкт для иммунномодуляции, содержащий мотив последовательности N1N2CGN3N4, где N представляет собой нуклеотид, выбранный из группы, состоящей из С, G, А и Т, а С представляет дезоксицитидин, G представляет дезоксигуанозин, А представляет дезоксиаденозин, и Т представляет дезокситимидин, и, по меньшей мере, два из пяти терминальных нуклеотидов, расположенных на 5′- и/или 3′-конце ДНК конструкта, представлены в L-конфигурации, при этом в мотиве последовательности N1N2CGN3N4 отсутствуют нуклеотиды в L-конфигурации. Также представлена вакцина, содержащая описанный конструкт. Изобретение расширяет арсенал иммуномодулирующих средств. 4 н. и 12 з.п. ф-лы, 14 ил., 3 табл.
1. Некодирующий ДНК конструкт для иммунномодуляции, содержащий мотив последовательности N1N2CGN3N4, где N представляет собой нуклеотид, выбранный из группы, состоящей из С, G, А и Т, а С представляет дезоксицитидин, G представляет дезоксигуанозин, А представляет дезоксиаденозин, и Т представляет дезокситимидин, и, по меньшей мере, два из пяти терминальных нуклеотидов, расположенных на 5′- и/или 3′-конце ДНК конструкта, представлены в L-конфигурации, при этом в мотиве последовательности N1N2CGN3N4 отсутствуют нуклеотиды в L-конфигурации.
2. Конструкт по п. 1, где N1N2 представляет собой элемент, выбранный из группы, состоящей из GT, GG, GA, AT или АА, N3N4 представляет элемент, выбранный из группы, состоящей из СТ или ТТ.
3. Конструкт по любому из предшествующих пунктов, также включающий по меньшей мере один G повтор, включающий три последовательных дезоксигуанозина, расположенные вблизи 5′- и/или 3′-конца ДНК конструкта.
4. Конструкт по любому из предшествующих пунктов, где G повтор, включающий три последовательных дезоксигуанозина, расположен между CG нуклеотидами двух указанных мотивов последовательности N1N2CGN3N4.
5. Конструкт по любому из предшествующих пунктов, где по меньшей мере пять нуклеотидов, выбранных из группы, состоящей из С, Т, А и G, расположены между CG нуклеотидами двух указанных мотивов последовательности N1N2CGN3N4, исключая дезоксигуанозин.
6. Конструкт по любому из предшествующих пунктов, где ДНК конструкт выбирают из группы, состоящей из
а) линейного и с открытой цепью ДНК конструкта, содержащего одноцепочечную или двухцепочечную ДНК, и
б) линейного ДНК конструкта, включающего по меньшей мере один конец с одноцепочечной петлей.
7. Конструкт по любому из предшествующих пунктов, где мотив последовательности N1N2CGN3N4 расположен в одноцепочечной и/или двухцепочечной области последовательности ДНК.
8. Конструкт по любому из предшествующих пунктов, где конструкт содержит меж- и/или внутримолекулярные пары оснований и по меньшей мере одну неспаренную одноцепочечную область.
9. Конструкт по п. 8, также включающий дополнительные ДНК конструкты, где по меньшей мере два ДНК конструкта соединены друг с другом.
10. Конструкт по любому из предшествующих пунктов, где по меньшей мере один нуклеотид ДНК конструкта в L- или D-конфигурации вне мотива последовательности N1N2CGN3N4 модифицирован функциональной группой, выбранной из группы, содержащей карбоксильную группу, аминную, амидную, альдиминную, кетальную, ацетальную, сложного эфира, простого эфира, дисульфидную, тиоловую и альдегидные группы.
11. Конструкт по любому из предшествующих пунктов, содержащий первый G повтор на 5′ конце и три мотива последовательности N1N2CGN3N4, где по меньшей мере пять нуклеотидов, выбранных из группы, состоящей из С, Т, А и G, расположены между CG нуклеотидами первого и второго мотива, исключая дезоксигуанозин, и G повтор, который расположен между CG нуклеотидами второго и третьего мотива последовательности, и где два из трех 3′ терминальных дезоксинуклеотидов находятся в L-конфигурации.
12. Конструкт по п. 11, содержащий по меньшей мере один дополнительный G повтор.
13. Применение ДНК конструкта по любому из пп. 1-12 для лечения рака или аутоиммунных заболеваний.
14. Применение ДНК конструкта по любому из пп. 1-12 для модуляции иммунной системы.
| JOHNATHAN C LAI, et al "Comparison of D-G3139 and Its Enantiomer L-G3139 in Melanoma Cells Demonstrates Minimal In Vitro but Dramatic In Vivo Chiral Dependency", Molecular Therapy vol | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Приспособление для уменьшения дымовой тяги паровоза | 1920 |
|
SU270A1 |
| QIUVAN ZHAO, et al "SITE OF CHEMICAL MODIFICATIONS IN CpG CONTAINING PHOSPHOROTHIOATE OLIGODEOXYNUCLEOTIDE MODULATES ITS | |||

