Настоящее изобретение относится к олигонуклеотидным ингибиторам активности или экспрессии TSC22D4 и их применению для профилактики, лечения и/или регуляции резистентности к инсулину, метаболического синдрома и/или сахарного диабета и/или для усиления чувствительности к инсулину у млекопитающего.
Уровень техники
У людей комбинация избыточного накопления липидов и уменьшения удаления приводит к избыточному весу и связанным с ним сопутствующим заболеваниям, включающим в себя резистентность к инсулину, сердечно-сосудистые осложнения и дислипидемию (Langin D. In and out: adipose tissue lipid turnover in obesity and dyslipidemia. Cell Metab. 2011 Nov 2; 14(5):569-70), которые в настоящее время затрагивают более 1,5 миллиарда человек во всем мире (Finucane MM, et al. National, regional, and global trends in body-mass index since 1980: systematic analysis of health examination surveys and epidemiological studies with 960 country-years and 91 million participants. Lancet. 2011 Feb 12; 377(9765):557-67). Действительно, резистентность к инсулину представляет собой основной компонент так называемого метаболического синдрома, который в конечном итоге приводит к развитию метаболической дисфункции, такой как непереносимость глюкозы, панкреатическая бета-клеточная недостаточность и, в конечном счете, сахарный диабет 2 типа.
Нарушенная секреция инсулина (бета-клетка), увеличенное производство глюкозы в печени (печень) и сниженное периферическое использование глюкозы (мышца) составляют традиционные первичные дефекты, ответственные за развитие и прогрессирование сахарного диабета 2 типа. В настоящее время известно, что бета-клеточная недостаточность, в конечном счете приводящая к уменьшению секреции инсулина, происходит намного раньше в естественном анамнезе сахарного диабета 2 типа, чем предполагалось первоначально. Кроме того, лучшее понимание патофизиологии сахарного диабета 2 типа выявляет другие этиологические механизмы за пределами классической триады, теперь называемой зловещим октетом. В дополнение к бета-клетке, печени и мышцам другие патогенные механизмы включают в себя резистентность адипоцитов к инсулину (повышенный липолиз), сниженную секрецию/чувствительность к инкретину (желудочно-кишечный тракт), повышенную секрецию глюкагона (альфа-клетка), усиленную реабсорбцию глюкозы (почка) и инсулинорезистентность центральной нервной системы, возникающую в результате дисфункции нейромедиатора (мозг). В настоящее время управление сахарным диабетом 2 типа сосредоточено на контроле глюкозы путем снижения содержания глюкозы в крови (при голодании и после приема пищи) и гемоглобина А(1с). Однако цель терапии должна заключаться в том, чтобы задержать прогрессирование заболевания и возможную неэффективность лечения. Лечение должно быть нацелено на известные патогенные нарушения заболевания (т.е. снижение ухудшения функции бета-клеток и повышение чувствительности к инсулину). В последние годы стратегии лечения были сфокусированы на разработке новых терапевтических вариантов, которые влияют на многие из дефектов, способствующих сахарному диабету 2 типа, и которые обеспечивают прочный контроль глюкозы посредством притупления прогрессирования заболевания. Оптимальное лечение сахарного диабета 2 типа должно предусматривать раннее начало терапии с использованием комбинации нескольких лекарственных средств с различными механизмами действия (DeFronzo RA. (Current issues in the treatment of type 2 diabetes. Overview of newer agents: where treatment is going. Am J Med. 2010 Mar; 123(3 Suppl):S38-48).
Особенно нечувствительность основных метаболических органов к действию инсулина, включая в себя печень, скелетную мышцу и жировую ткань, существенно способствует прогрессированию заболевания и конечной потребности в фармакологическом вмешательстве для предотвращения диабетических поздних осложнений. Таким образом, эффективная и безопасная сенсибилизация к инсулину остается привлекательной целью и направлена на противодиабетическую терапию.
Транскрипционные кофакторные комплексы были идентифицированы как важные контрольные точки в координации метаболических программ в различных тканях, включая в себя печень и белую жировую ткань (WAT) (для обзора смотрите Sommerfeld A, Krones-Herzig A, Herzig S. Transcriptional co-factors and hepatic energy metabolism. Mol Cell Endocrinol. 2011 Jan 30; 332(1-2):21-31).
Kester HA, et al. (в: Transforming growth factor-beta-stimulated clone-22 is a member of a family of leucine zipper proteins that can homo- and heterodimerize and has transcriptional repressor activity. J Biol Chem. 1999 Sep 24; 274(39):27439-47) описывают, что стимулированный TGF-бета клон-22 (TSC-22) кодирует содержащий лейциновую «молнию» белок, который представляет собой высококонсервативный во время эволюции.
Кроме того, Jones et al. (в Jones, A., et al., Transforming growth factor-betal Stimulated Clone-22 D4 is a molecular output of hepatic wasting metabolism. EMBO Mol Med. 2013 Feb; 5(2):294-308) описывают это как путь выхода молекулярной кахексии, причем содержание в печени стимулированного трансформирующим фактором роста бета-1 клона транскрипционного фактора (TSC) 22 D4 было увеличено при раковой кахексии. Подражание высоким деградационным уровням TSC22D4 в здоровой печени приводило к ингибированию печеночного высвобождения VLDL и липогенным генам и снижению системных уровней VLDL как в нормальных условиях, так и при диете с высоким содержанием жира. Таким образом, печеночная активность TSC22D4 может представлять собой молекулярное обоснование депривации периферической энергии у пациентов с заболеваниями метаболического истощения, включая в себя раковую кахексию.
Kulozik, Ph., et al. (Hepatic deficiency in transcriptional co-factor TBLI promotes liver steatosis and hypertriglyceridemia. 2011 Cell Metab. 13: 389-400) описывают, что нарушенная печеночная экспрессия транскрипционного кофактора трансдуцин бета-подобного (TBL) 1 белка представляет собой общую черту моно- и мультигенньгх мышиных моделей с жировым перерождением печени. Специфическая для печени абляция экспрессии гена TBL1 у здоровых мышей способствовала гипертриглицеридемии и стеатозу печени при нормальных условиях и диете с большим содержанием жиров. Поскольку было обнаружено, что уровни экспрессии TBL1 также обратно коррелировали с содержанием жира в печени у пациентов-людей, отсутствие активности кофактора TBL1/TBLR1 печени может представлять собой молекулярное обоснование стеатоза печени у субъектов с ожирением и метаболическим синдромом.
Berriel Diaz, М., et al. (Nuclear receptor co-factor RIP140 controls lipid metabolism during wasting in mice. 2008. Hepatology 48: 782-791) описывают, что посредством предотвращения мобилизации печеночных хранилищ TG, индукция RIP 140 в печени обеспечивает молекулярное обоснование стеатоза печени при голодании, сепсисе или истощении при злокачественной опухоли. Таким образом, ингибирование печеночной транскрипционной активности RIP 140 может обеспечить привлекательную вспомогательную схему при лечении этих состояний.
Farese et al. (in: The problem of establishing relationships between hepatic steatosis and hepatic insulin resistance. Cell Metab. 2012 May 2; 15(5):570-3) описывают, что чрезмерное отложение жира в печени (печеночный стеатоз) часто сопровождается печеночной резистентностью к инсулину.
Основные классы противодиабетических и/или сенсибилизирующих к инсулину лекарственных средств включают в себя сульфонилмочевины, метформин, тиазолидиновые дионы, ингибиторы альфа-глюкозидазы, инкретиновые миметики и ингибиторы дипептидилпептидазы 4, все из которых связаны с серьезными ограничениями (для обзора смотрите Moller, Metabolic disease drug discovery- "hitting the target" is easier said than done. Cell Metab. 2012 Jan 4; 15(1): 19-24).
Несмотря на ключевую роль инсулинорезистентности в патогенезе сахарного диабета 2 типа, эффективных и безопасных сенсибилизаторов к инсулину все еще не хватает. Действительно, применяемые в настоящее время лекарственные средства семейства тиазолидиндионов проявляют умеренный профиль эффективности и сопровождаются существенными побочными эффектами, включающими в себя увеличение массы, повышенный риск сердечной недостаточности, возможный повышенный риск развития злокачественной опухоли мочевого пузыря и повышенный риск инфаркта миокарда, например, что привело к недавнему выводу с рынка росиглитазона.
В публикации международной заявки WO 2013/076501 раскрыт способ скрининга для идентификации средств, применимых при лечении и/или профилактике заболевания, связанного с резистентностью к инсулину и/или непереносимостью глюкозы, который предусматривает стадию исследования способности тестируемого средства ингибировать сигнальный путь Vps34 и/или сигнальный путь Rholota3Kappa-02beta. Аналогично, в публикации международной заявки WO 2005/059564 раскрыт способ сканирования молекул, которые модулируют активность ретинол-связывающего белка 4 (RBP4), а также описано их применение для лечения резистентности к инсулину. Также описаны способы диагностики резистентности к инсулину и связанные с ней состояния путем обнаружения модуляции активности RBP4.
Публикация международной заявки WO 2012/158123 относится к способу лечения или предотвращения синдрома резистентности к инсулину в организме животного путем введения ингибитора гена подобной РНК-зависимой проитеникинзе киназы эндоплазматического ретикулума (PERK) или его функционального варианта, или ингибитора белка PERK или его функционального варианта, или способу снижения активности транскрипционных факторов семейства FOXO (Foxo 1, 3а, 4 и 6) путем введения ингибитора гена подобной РНК-зависимой проитеникинзе киназы эндоплазматического ретикулума (PERK) или его функционального варианта или ингибитора белка PERK или его функционального варианта.
Публикация международной заявки WO 2014/202602, как правило, относится к модуляторам, в частности ингибиторам, активности или экспрессии TSC22D4 и их применению для профилактики, лечения и/или регуляции резистентности к инсулину, метаболического синдрома и/или сахарного диабета и/или для усиления чувствительности к инсулину у млекопитающего. Публикация международной заявки WO 2014/202602 относится также к способам скрининга для идентификации этих модуляторов.
Было доказано, что экспериментальный нокдаун с помощью вирусной доставки направленных на TSC22D4 конструкций кшРНК или миРНК, эффективно улучшает метаболический статус диабетических животных, конструкции миРНК, подходящие для эффективного и специфического нокдауна TSC22D4 у разных видов при доставке различными технологиями, еще не идентифицированы.
Принимая во внимание вышеописанные недостатки предшествующего уровня техники, цель настоящего изобретения заключается в предоставлении новой терапевтической стратегии для предотвращения, лечения и/или регулирования резистентности к инсулину, метаболического синдрома и/или сахарного диабета и/или для усиления чувствительности к инсулину.
Согласно первому аспекту настоящего изобретения вышеуказанная цель решается путем обеспечения ингибитора экспрессии и/или биологической активности TSC22D4, выбранного из олигонуклеотида, который представляет собой интерферирующую рибонуклеиновую кислоту, PNA (белковая нуклеиновая кислота) или LNA (закрытая нуклеиновая кислота), содержащую по меньшей мере одну из следующих последовательность: 5'-GGACGUGUGUGGAUGUUUAdTdT-3' (SEQ ID No. 1); 5'-GGAUGUUUACGAGAGAGAUdTdT-3' (SEQ ID No. 2); 5'-AGUCCCACCUCAUGUUUGCdTdT-3' (SEQ ID No. 3); их антисмысловую последовательность или их функциональные варианты.
mhD4-siRNAl: (NM_ 030935.3_ миРНК_ 1024; открытая рамка считывания)
Смысловая: 5'- GGACGUGUGUGGAUGUUUAdTdT-3' (SEQ ID No. 1);
Антисмысловая: 5'- UAAACAUCCACACACGUCCdTdT-3' (SEQ ID No. 4);
GC: 47% (без TT-липкого конца)


Недавно исследователи показали, что стимулированный трансформирующим фактором роста бета1 клон транскрипционного регулятора 22 D4 (TSC22D4), контролирует печеночную и системную чувствительность к инсулину. Специфическая для печени потеря TSC22D4 значительно улучшала толерантность к глюкозе и чувствительность к инсулину и противодействовала гиперинсулинемии у мышей дикого типа. Анализ ChlP-Seq цистрона TSC22D4 в сочетании с высокопроизводительными исследованиями транскриптома TSC22D4 у здоровых животных показал, что основные узлы пути передачи сигналов инсулина были прямо или косвенно нацелены на TSC22D4, в первую очередь на липокалин 13.
Действительно, понижающая регуляция или сверхэкспрессия TSC22D4 в первичных мышиных гепатоцитах, а также у мышей дикого типа приводила к активации или деактиванции пути сигнализации инсулина, как определено фосфорилированием киназы Akt/PKB на Ser473 и GSK3beta в Ser9, в ответ на острое воздействие инсулина, соответственно. Интригующе, инактивация в печени TSC22D4 у мышей db/db с сахарным диабетом улучшала непереносимость глюкозы и резистентность к инсулину у этих животных и нормализовала содержание глюкозы в крови до почти здорового уровня. В сопоставлении с общим улучшением метаболического статуса у диабетических животных циркулирующие уровни провоспалительных цитокинов и резистина были значительно ниже у мышей со специфическим для печени дефицитом TSC22D4.
Хотя было доказано, что экспериментальный нокдаун с помощью вирусной доставки направленных на TSC22D4 конструкций кшРНК или миРНК эффективно улучшает метаболический статус диабетических животных, конструкции миРНК, подходящие для эффективного и специфического нокдауна TSC22D4 у разных видов при доставке различными технологиями еще не идентифицированы.
Инактивация TSC22D4 в клетках гепатомы не увеличивала клеточный рост, а скорее уменьшала пролиферацию, указывая на то, что функция сенсибилизации к инсулину TSC22D4 не приводит к повышенной восприимчивости к злокачественной опухоли в пораженных клетках/органах. Кроме того, инактивация в печени TSC22D4 также не вызывала гипогликемии.
«Ингибитор» представляет собой вещество, которое может снижать эффективность катализатора в катализируемой реакции (либо небиологического катализатора, либо фермента). Указанный в настоящем документе ингибитор может снижать эффективность активности фермента; также указанный в настоящем документе ингибитор может снижать эффективность экспрессии фермента. В контексте настоящего изобретения предпочтительный ингибитор представляет собой олигонуклеотид.
Термин «олигонуклеотид», как правило, относится к интерферирующей рибонуклеиновой кислоте (иРНК) или белковой нуклеиновой кислоте (PNA), или к закрытой нуклеиновой кислоте (LNA). Термин «олигонуклеотид», как правило, относится к одноцепочечному нуклеотидному полимеру, состоящему более чем из 19 нуклеотидных субъединиц, ковалентно соединенных вместе. Предпочтительно присутствуют от 19 до 100 нуклеотидных единиц, наиболее предпочтительно от 19 до 50 единиц нуклеотидов соединены вместе, как также объясняется дополнительно ниже.
Сахарные группы нуклеотидных субъединиц могут представлять собой рибозу, дезоксирибозу или модифицированные производные, такие как 2'-O-метилрибоза. Нуклеотидные субъединицы олигонуклеотида могут быть соединены фосфодиэфирными связями, фосфоротиоатными связями, метилфосфонатными связями или другими редкими или неприродными связями, которые не препятствуют гибридизации олигонуклеотида. Кроме того, олигонуклеотид может содержать редкие нуклеотиды или ненуклеотидные фрагменты.
Термин «олигонуклеотид» также может относиться, в контексте спецификации, к аналогу известных в настоящей области техники нуклеиновых кислот, например, закрытой нуклеиновой кислоте (LNA), или их смеси. Термин «олигонуклеотид» включает в себя олигонуклеотиды, состоящие из встречающихся в природе нуклеотидных оснований, сахаров и межнуклеозидных (остовных) связей, а также олигонуклеотидов, содержащих не встречающиеся в природе участки, которые функционируют аналогично или с определенными улучшенными функциями. Полностью или частично модифицированный или замещенный олигонуклеотид часто предпочтителен по сравнению с природными формами из-за нескольких желательных свойств таких олигонуклеотидов, таких как, например, способность проникать в клеточную мембрану, хорошая устойчивость к вне- и внутриклеточным нуклеазам, высокая аффинность и специфичность к мишени нуклеиновой кислоты. Способы модификации олигонуклеотидов, таким образом, известны в настоящей области техники.
В некоторых олигонуклеотидах, иногда называемых олигонуклеотидными миметиками, как сахара, так и межнуклеозидная связь, то есть остов, нуклеотидных единиц заменяются новыми группами. Единицы оснований поддерживаются для гибридизации с соответствующим соединением-мишенью нуклеиновой кислоты. Одно такое олигомерное соединение, олигонуклеотидный миметик, который, как было показано, обладает превосходными свойствами гибридизации, упоминается как белковая нуклеиновая кислота (PNA). В соединениях PNA сахарный остов олигонуклеотида заменяют амидосодержащим остовом, в частности аминоэтилглициновым остовом. Нуклеотидные основания сохраняются и непосредственно или косвенно связываются с атомами азота амидной части остова.
Дополнительная модификация включает в себя закрытые нуклеиновые кислоты (LNA), в которых 2'-гидроксильная группа связана с 3' или 4' атомом углерода сахарного кольца, тем самым образуя бициклический сахарный фрагмент. Связывание представляет собой предпочтительно метиленовую (-СН2-)n-группу, соединяющую 2' атом кислорода и 4' атом углерода, где n равно 1 или 2. Термин «LNA», как правило, относится к нуклеотиду, содержащему один бициклический нуклеозидный аналог, также называемый мономером LNA, или олигонуклеотиду, содержащему один или несколько бициклических нуклеозидных аналогов.
Предпочтительным является ингибитор в соответствии с настоящим изобретением, где интерферирующая рибонуклеиновая кислота представляет собой малую интерферирующую рибонуклеиновую кислоту (миРНК) или короткую шпилечную рибонуклеиновую кислоту (кшРНК) или микрорибонуклеиновую кислоту (микроРНК) или их комбинации.
Кроме того, предпочтительным является ингибитор в соответствии с настоящим изобретением, причем миРНК составляет в длину от 19 до 30 нуклеотидов.
Согласно одному аспекту биоактивное средство использует «РНК-интерференцию (RNAi)». RNAi представляет собой процесс специфического к последовательности посттранскрипционного сайленсинга гена, инициированного двухцепочечной РНК (дцРНК) или миРНК. RNAi наблюдается у ряда организмов, таких как дрозофила, нематоды, грибы и растения и, как полагают, участвует в противовирусной защите, модуляции активности транспонов и регуляции экспрессии генов. Во время RNAi дцРНК или миРНК индуцирует деградирование мРНК-мишени с последующим специфическим к последовательности ингибированием экспрессии гена. Используемый в настоящем документе термин «малая интерферирующая РНК» (миРНК) представляет собой РНК-дуплекс нуклеотидов, который нацелен на ген TSC22D4. «РНК-дуплекс» относится к структуре, образованной комплементарным спариванием между двумя областями молекулы РНК. миРНК «нацеливается» на ген в том плане, что нуклеотидная последовательность дуплексной части миРНК комплементарна нуклеотидной последовательности гена-мишени. Согласно некоторым вариантам осуществления длина дуплекса миРНК составляет менее 30 нуклеотидов. Согласно некоторым вариантам осуществления дуплекс может составлять в длину 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11 или 10 нуклеотидов. Согласно некоторым вариантам осуществления длина дуплекса составляет 19-25 нуклеотидов. РНК-дуплексная часть миРНК может быть частью структуры шпильки. В дополнение к дуплексной части структура шпильки может содержать петлевую часть, расположенную между двумя последовательностями, которые образуют дуплекс. Петля может варьировать по длине. Согласно некоторым вариантам осуществления петля составляет в длину 5, 6, 7, 8, 9, 10, 11, 12 или 13 нуклеотидов. Структура шпильки также может содержать 3' и/или 5' липкие концы. Согласно некоторым вариантам осуществления липкий конец представляют собой 3' и/или 5' липкий конец длиной 0, 1, 2, 3, 4 или 5 нуклеотидов. миРНК может кодироваться последовательностью нуклеиновой кислоты, а последовательность нуклеиновой кислоты может также включать в себя промотор. Последовательность нуклеиновой кислоты может также включать в себя сигнал полиаденилирования. Согласно некоторым вариантам осуществления сигнал полиаденилирования представляет собой синтетический минимальный сигнал полиаденилирования.
Используемый в настоящем документе термин «миРНК» относится к рибонуклеиновой кислоте (РНК) или аналогу РНК, содержащему от приблизительно 19 до 50 нуклеотидов (или нуклеотидных аналогов), способных направлять или опосредовать путь интерференции РНК. Эти молекулы могут варьировать по длине и могут содержать разные степени комплементарности к их целевой информационной РНК (мРНК) в антисмысловой цепи. Термин «миРНК» включает в себя дуплексы двух отдельных цепей, то есть двухцепочечную РНК, а также отдельные цепи, которые могут образовывать структуры шпильки, состоящие из дуплексной области. миРНК может составлять в длину от приблизительно 19 до 50 нуклеотидов или от приблизительно 25 до 50 нуклеотидов, или от приблизительно 30 до 50 нуклеотидов, или от приблизительно 35 до 50 нуклеотидов, или от приблизительно 40 до 50 нуклеотидов. Согласно одному варианту осуществления миРНК составляет в длину от 19 до 30 нуклеотидов.
Применение миРНК для снижения активности своей целевой мРНК известно в настоящей области техники. Согласно некоторым вариантам осуществления деградация мРНК происходит, когда антисмысловая цепь или направляющая цепь миРНК направляет индуцированный РНК комплекс сайленсинга (RISC), который содержит РНК-эндонуклеазу Ago2, чтобы расщепить свою мРНК-мишень, несущую комплементарную последовательность. Соответственно, миРНК может быть комплементарной любой части разной длины на гене PERK. миРНК также может быть комплементарной смысловой цепи и/или антисмысловой цепи гена TSC22D4. Соответственно, обработка миРНК может быть использована для сайленсинга гена TSC22D4, тем самым истощая белок TSC22D4 на последующих стадиях.
Используемый в настоящем документе термин «кшРНК» относится к мономолекулярной РНК, которая способна к выполнению RNAi, и которая содержит «сопровождающую» цепь, петлю и направляющую цепь. Сопровождающая и направляющая цепи могут быть по существу комплементарны друг другу. Термин «кшРНК» может также включать в себя нуклеиновые кислоты, которые содержат фрагменты, отличные от рибонуклеотидных фрагментов, включая в себя, без ограничения, модифицированные нуклеотиды, модифицированные межнуклеотидные связи, ненуклеотиды, дезоксинуклеотиды и аналоги нуклеотидов.
миРНК снижают активность их целевых мРНК. Термин «миРНК», как правило, относится к одноцепочечной молекуле, но согласно конкретным вариантам осуществления может также охватывать область или дополнительную цепь, которая частично (от 10% до 50% комплементарной по длине цепи), по существу (более 50%, но менее 100% комплементарной по длине цепи) или полностью комплементарна другой области одной и той же одноцепочечной молекулы или другой нуклеиновой кислоты. Таким образом, нуклеиновые кислоты могут охватывать молекулу, которая содержит одну или несколько комплементарных или самокомплементарных цепей или «комплементов» определенной последовательности, содержащей молекулу. Например, миРНК-предшественник может содержать самокомплементарную область, которая комплементарна до 100%. Зонды миРНК или нуклеиновые кислоты по настоящему изобретению могут включать в себя или могут быть комплементарны полностью или по меньшей мере на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% по отношению к своей мишени.
Наиболее предпочтительным является ингибитор в соответствии с настоящим изобретением, в котором миРНК состоит из последовательности: в соответствии с ее антисмысловой последовательностью SEQ ID No. 1-3.
Также предпочтительным является ингибитор в соответствии с настоящим изобретением, причем его функциональный вариант содержит по меньшей мере один модифицированный или замещенный нуклеотид. Термин «функциональный вариант» также включает в себя фрагмент, вариант, основанный на коде дегенеративной нуклеиновой кислоты или химическом производном. Функциональный вариант может содержать консервативные изменения, причем замещенная нуклеиновая кислота характеризуется сходными структурными или химическими свойствами с замещенной нуклеиновой кислотой. Функциональный вариант может также содержать делецию и/или вставку одной или нескольких нуклеиновых кислот. Понятно, что функциональный вариант по меньшей мере частично сохраняет свою биологическую активность, например, функции гена TSC22D4, или даже проявляет улучшенную биологическую активность.
Примеры модифицированных олигонуклеотидов включают в себя, без ограничения, олигонуклеотиды с фосфоротиоатными остовами (смотрите выше) и олигонуклеозиды с гетероатомными остовами и, в частности, CH2-NH-O-CH2-, -CH2-N(CH3)-O-CH2-, -CH2-O-N(CH3)-CH2-, -CH2-N(CH3)-N(CH3)-CH2- и -O-N(CH3)-CH2-СН2- [причем нативный фосфодиэфирный остов представлен в виде -О-Р-О-СН2-]. Также применимы олигонуклеотиды, содержащие морфолиновые остовные структуры. Модифицированные олигонуклеотиды, используемые в качестве интерферирующих рибонуклеиновых кислот, также могут содержать один или несколько замещенных фрагментов сахара. Предпочтительные олигонуклеотиды содержат одно из следующего в положении 2': ОН; F; О-, S- или N-алкил; О-, S- или N-алкенил; О-, S- или N-алкинил; или О-алкил-О-алкил, где алкил, алкенил и алкинил могут быть замещенными или незамещенными C1-С10-алкилом или С2-С10-алкенилом и алкинилом. Конкретные примеры включают в себя, без ограничения, O[(СН2)nO]mCH3, O(СН2)nOCH3, O(CH2)nNH2, O(СН2)nCH3, O(CH2)nONH2 и O(CH2)nON[(CH2)nCH3)]2, где n и m составляют от 1 до 10. Другие иллюстративные олигонуклеотиды содержат одно из следующего в положении 2': C1-С10 низший алкил, замещенный низший алкил, алкенил, алкинил, алкарил, аралкил, О-алкарил или О-аралкил, SH, SCH3, OCN, Cl, Br, CN, CF3, OCF3, SOCH3, SO2CH3, ONO2, NO2, N3, NH2, гетероциклоалкенил, гетероциклоалкарил, аминоалкиламино, полиалкиламино, замещенный силил, расщеплющую РНК группу, репортерную группу, интеркалатор, группу для улучшения фармакокинетических свойств олигонуклеотида или группу для улучшения фармакодинамических свойств олигонуклеотида и других заместителей, характеризующихся сходными свойствами. Одна иллюстративная модификация включает в себя 2'-метоксиэтокси (2'-O-СН2СН2ОСН3, также известный как 2'-O-(2-метоксиэтил) или 2'-МОЕ), то есть алкоксиалкоксигруппу.
Другой аспект затем относится к рекомбинантному вектору, содержащему олигонуклеотид в соответствии с настоящим изобретением. Как правило, олигонуклеотид вводят в вектор экспрессии, такой как плазмида, для экспрессии. Если необходимо, олигонуклеотид может быть связан с соответствующими транскрипционными и трансляционными регуляторными контрольными нуклеотидными последовательностями, распознаваемыми желаемым хозяином, хотя такие контроли, как правило, доступны в векторе экспрессии. Затем вектор вводится в хозяина посредством стандартных техник.
Векторы, которые экспрессируют миРНК в клетках млекопитающих, как правило, используют промотор РНК-полимеразы III, чтобы управлять экспрессией короткой шпилечной РНК, которая имитирует структуру миРНК. Вставка, которая кодирует эту шпильку, предназначена для двух инвертированных повторов, разделенных короткой спейсерной последовательностью. Один инвертированный повтор комплементарен мРНК, на которую нацелена миРНК. Строка тимидинов, добавленных к 3'-концу, служит в качестве сайта терминации транскрипции pol III. Сразу внутри клетки вектор конститутивно экспрессирует РНК шпильки, которая индуцирует сайленсинг гена-мишени.
Другие подходящие векторы включают в себя вирусные векторы, такие как аденовирусные, ретровирусные и лентивирусные векторы или соответствующие системы экспрессии (смотрите, например, Catanotto, D. et al. (2002) Functional siRNA expression from transfected PCR products. RNA 8, 1454-1460; Barton, G.M. et al. (2002) Retroviral delivery of small interfering RNA into primary cells. Proc Natl Acad Sci USA. 99(23): 14943-5. Abbas-Terki, T. et al.(2002) Lentiviral-mediated RNA interference. Hum.Gene Ther. 13, 2197-2201, and Xia, H. et al. (2002) siRNA-mediate gene silencing in vitro and in vivo. Nat. Biotechnol. 20, 1006-1010).
Как правило, не все хозяева будут преобразованы вектором. Следовательно, необходимо будет выбрать трансформированные клетки-хозяева. Одна техника выбора предусматривает включение в вектор экспрессии последовательности ДНК с любыми необходимыми контрольными элементами, которая кодирует селективный признак в трансформированной клетке, такой как устойчивость к антибиотикам. Альтернативно, ген для такого селективного признака может быть на другом векторе, который используется для совместной трансформации желаемой клетки-хозяина. Затем клетки-хозяева, которые были трансформированы олигонуклеотидом по настоящему изобретению, культивируют в течение достаточного времени и в подходящих условиях, известных специалистам в настоящей области техники, с учетом раскрытых в настоящем документе принципов, позволяющих экспрессию полипептида, который затем может быть восстановлен.
Другие примеры можно найти в литературе, например, в публикации Yang J. et al. (Design, preparation and application of nucleic acid delivery carriers." Biotechnol Adv. 2014 Jul-Aug; 32(4):804-17).
Каждый из классов нуклеиновых кислот, как описано в настоящем документе (например, олигонуклеотиды и/или векторы), может быть введен в клетки несколькими способами. При опосредованной липидом трансфекции клетки захватывают нековалентные комплексы между нуклеиновой кислотой и липидным или полимерным реагентом посредством эндоцитоза. Электропорация использует короткий электрический импульс, чтобы вызвать разрушения или отверстия в плазматической мембране клеток, через которые поступает нуклеиновая кислота. Оба этих способа успешно доставляют любую из нуклеиновых кислот RNAi, кроме вирусных векторов. Доставка вирусных векторов происходит только путем заражения клеток соответствующим вирусом, как правило, использующим вспомогательные вирусы. Инфекция нужной клеточной линии вирусом вводит миРНК или кшРНК и производит нокдаун генной экспрессии.
Другой аспект настоящего изобретения затем относится к рекомбинантной клетке, предпочтительно к рекомбинантной гепатоцитарной клетке, содержащей олигонуклеотид в соответствии с настоящим изобретением или рекомбинантный вектор в соответствии с настоящим изобретением. «Клетка» согласно настоящему изобретению может представлять собой прокариотическую или эукариотическую клетку. «Клетку» согласно настоящему изобретению предпочтительно и без ограничения выбирают из клеток печени. Клетки млекопитающих могут быть предпочтительно выбраны из клеток человека, кролика, мыши или крысы. Предпочтительно клетка представляет собой клетку человека, например, гепатоцитарную клетку. Термин «клетка» также включает в себя клетки животной модели. Кроме того, клетка может представлять собой часть тканевой культуры.
Задача настоящего изобретения также решается с помощью способа получения фармацевтической композиции, содержащей стадии составления указанного по меньшей мере одного ингибитора в соответствии с настоящим изобретением по меньшей мере с одним фармацевтически приемлемым вспомогательным веществом. Носитель и/или вспомогательное вещество фармацевтической композиции должны быть «приемлемыми» в том смысле, что они должны быть совместимы с другими ингредиентами состава и не быть вредными для реципиента.
Другой аспект настоящего изобретения затем относится к фармацевтической композиции, содержащей по меньшей мере один из ингибиторов в соответствии с настоящим изобретением, рекомбинантный вектор в соответствии с настоящим изобретением и рекомбинантную клетку в соответствии с настоящим изобретением вместе с фармацевтически приемлемым носителем. Предпочтительной является фармацевтическая композиция в соответствии с настоящим изобретением, причем указанная фармацевтическая композиция предназначена для введения пероральным, ректальным, трансмукозальным, трансдермальным, внутрикишечным, парентеральным, внутримышечным, интратекальным, прямо интравентрикулярным, внутривенным, внутрибрюшинным, интраназальным, внутриглазным или подкожным способом.
Другой аспект настоящего изобретения затем относится к ингибитору в соответствии с настоящим изобретением, вектору экспрессии в соответствии с настоящим изобретением, рекомбинантной клетке в соответствии с настоящим изобретением или фармацевтической композиции в соответствии с настоящим изобретением для применения при профилактике, регуляции и/или лечения заболеваний.
Синдром резистентности к инсулину составляет широкий клинический спектр и определяется как любые аномалии, связанные с резистентностью к инсулину. Нарушения, такие как резистентность к инсулину, сахарный диабет, гипертония, дислипидемия и сердечно-сосудистые заболевания, представляют собой синдром инсулинорезистентности.
Синдром инсулинорезистентности может представлять собой индуцированную диетой инсулинорезистентность и/или индуцированную ожирением инсулинорезистентность. Индуцированная диетой инсулинорезистентность означает, что резистентность к инсулину индуцируется диетой с высоким содержанием насыщенных жиров и углеводов. Индуцированная ожирением инсулинорезистентность означает, что резистентность к инсулину индуцируется генетической предрасположенностью к ожирению или ожирением, которое обусловлено диетическими привычками.
Таким образом, другой аспект настоящего изобретения относится к ингибитору в соответствии с настоящим изобретением, вектору экспрессии в соответствии с настоящим изобретением, рекомбинантной клетке в соответствии с настоящим изобретением или фармацевтической композиции в соответствии с настоящим изобретением для применения в профилактике, регуляции и/или лечении заболевания, которое выбрано из резистентности к инсулину, гипертонии, дислипидемии, заболевания коронарной артерии, метаболического синдрома и/или сахарного диабета 1 или 2 типа и/или для улучшения чувствительности к инсулину, такого как, например, чувствительность к инсулину в контексте опухолевого заболевания. Предпочтительно синдром инсулинорезистентности представляет собой индуцированную диетой инсулинорезистентность и/или индуцированную ожирением инсулинорезистентность.
Объект дополнительно решается с помощью способа лечения и/или профилактики заболевания, выбранного из резистентности к инсулину, метаболического синдрома и/или сахарного диабета у нуждающегося в этом субъекта, предусматривающего стадию введения эффективного количества ингибитора в соответствии с настоящим изобретением или фармацевтической композиции в соответствии с настоящим изобретением нуждающемуся в этом пациенту.
Раскрытые способы могут быть использованы для лечения любого из следующих состояний, которые вызваны синдромом инсулинорезистентности: резистентность к инсулину, гипертония, дислипидемия, сахарный диабет 2 типа или заболевание коронарной артерии.
Термин «профилактика» в контексте настоящего изобретения следует понимать как медицинское вмешательство, целью которого является предотвращение возникновения негативного события, которое, скорее всего, приведет к ухудшению состояния пациента, характеризующегося наличием заболевания, или к травме или смерти здорового и/или больного субъекта. «Нуждающийся в этом пациент» может представлять собой, без ограничения, любое животное или человека, страдающего от заболевания, связанного с синдромом инсулинорезистентности, особенно резистентности к инсулину, гипертонии, дислипидемии, сахарного диабета 2 типа или ишемической болезни сердца. Предпочтительно, нуждающийся в этом пациент представляет собой человека.
Объект дополнительно решается терапевтическим набором, содержащим ингибитор в соответствии с настоящим изобретением, рекомбинантный вектор в соответствии с настоящим изобретением, рекомбинантную клетку в соответствии с настоящим изобретением или фармацевтическую композицию в соответствии с настоящим изобретением, необязательно вместе с подходящими буферами и вспомогательными веществами, а также инструкции по использованию.
Объект дополнительно решается терапевтическим набором в соответствии с настоящим изобретением для применения в профилактике, регуляции и/или лечении заболевания, причем указанное заболевание выбрано из резистентности к инсулину, гипертонии, дислипидемии, заболевания коронарной артерии, метаболического синдрома и/или сахарного диабета 1 или 2 типа и/или для улучшения чувствительности к инсулину, например, чувствительности к инсулину в контексте опухолевого заболевания.
Недавно исследователи показали, что стимулированный трансформирующим фактором роста бета 1 клон транскрипционного регулятора 22 D4 (TSC22D4) контролирует печеночную и системную чувствительность к инсулину. Специфическая потеря в печени TSC22D4 значительно улучшала толерантность к глюкозе и чувствительность к инсулину и противодействовала гиперинсулинемии у мышей дикого типа. Анализ ChlP-Seq цистрона TSC22D4 в сочетании с исследованиями транскриптома-мишени TSC22D4 с высокой пропускной способностью у здоровых животных показал, что основные узлы пути сигнализации инсулина были прямо или косвенно нацелены на TSC22D4, в первую очередь на липокалин 13. Действительно, понижающая регуляция или сверхэкспрессия TSC22D4 в первичных мышиных гепатоцитах, а также у мышей дикого типа приводили к активации или инактивации внутриклеточного пути сигнализации инсулина, как определено фосфорилированием киназы Akt/PKB в Ser473 и GSK3beta в Ser9, в ответ на острое воздействие инсулином, соответственно.
Интригующе, инактивация в печени TSC22D4 у мышей db/db с сахарным диабетом улучшала интолерантность к глюкозе и резистентность к инсулину у этих животных и нормализовала содержание глюкозы в крови до почти здорового уровня. В сопоставлении с общим улучшением метаболического статуса у диабетических животных циркулирующие уровни провоспалительных цитокинов и резистина были значительно ниже у мышей со специфичным для печени дефицитом TSC22D4.
Инактивация TSC22D4 в клетках гепатомы не увеличивала клеточный рост, а скорее уменьшала пролиферацию, указывая на то, что функция сенсибилизации TSC22D4 к инсулину не приводит к повышенной восприимчивости к злокачественной опухоли в пораженных клетках. Кроме того, инактивация в печени TSC22D4 также не вызывала гипогликемии.
Хотя было доказано, что экспериментальный нокдаун с помощью вирусной доставки направленных TSC22D4 конструкций кшРНК или миРНК эффективно улучшает метаболический статус диабетических животных, конструкции миРНК, подходящие для эффективного и специфического нокдауна TSC22D4 у разных видов при доставке различными технологиями еще не определены. Чтобы преодолеть эту проблему, исследователи идентифицировали, функционально исследовали и проверяли различные миРНК, направленные против последовательности мРНК TSC22D4, в исследованиях нокдауна in vitro с использованием мышиных Нера1.6, а также человеческих клеток гепатомы Huh7, как описано в настоящем документе.
Следующие фигуры, последовательности и примеры служат просто для иллюстрации настоящего изобретения и не должны толковаться как ограничивающие объем настоящего изобретения конкретными вариантами осуществления настоящего изобретения, описанными в примерах. Для целей настоящего изобретения все приведенные в тексте ссылки полностью включены в настоящий документ.
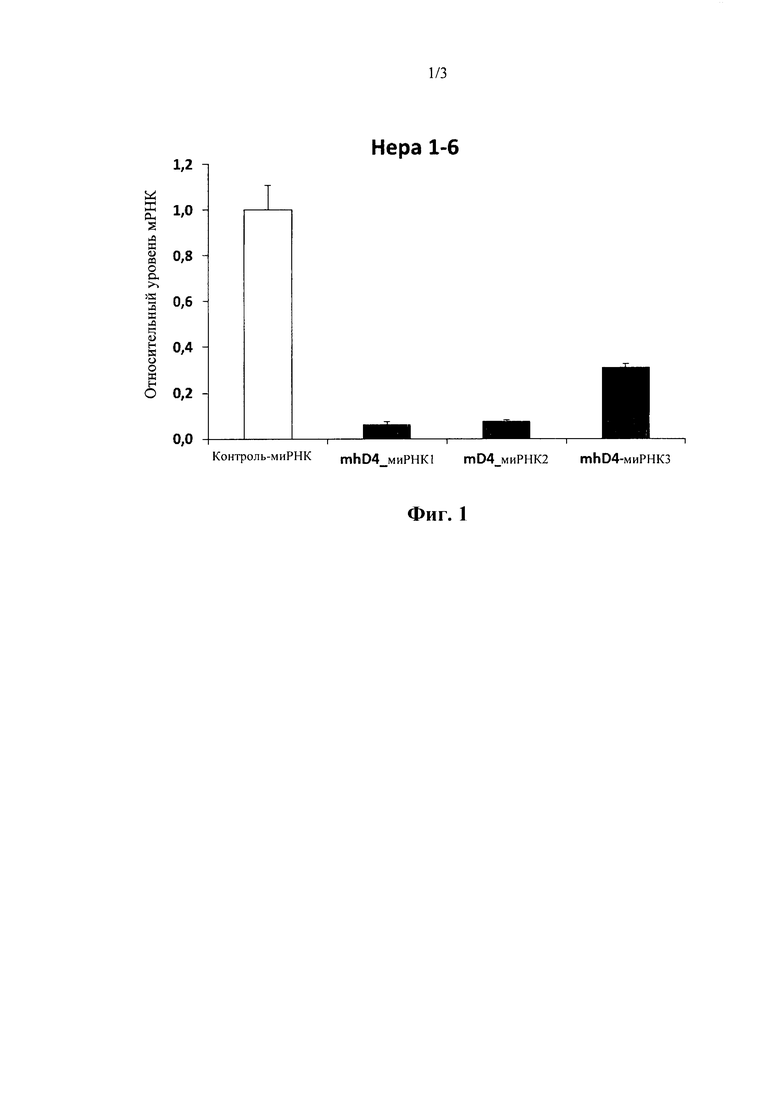
На фиг. 1 показана эффективность нокдауна выбранных направленных на TSC22D4 миРНК с цветовой индикацией в клетках мышиной гепатомы. Показано относительное содержание мРНК. Все остальные исследуемые последовательности миРНК не показали значительного нокдауна TSC22D4 в этих экспериментах (не показаны).
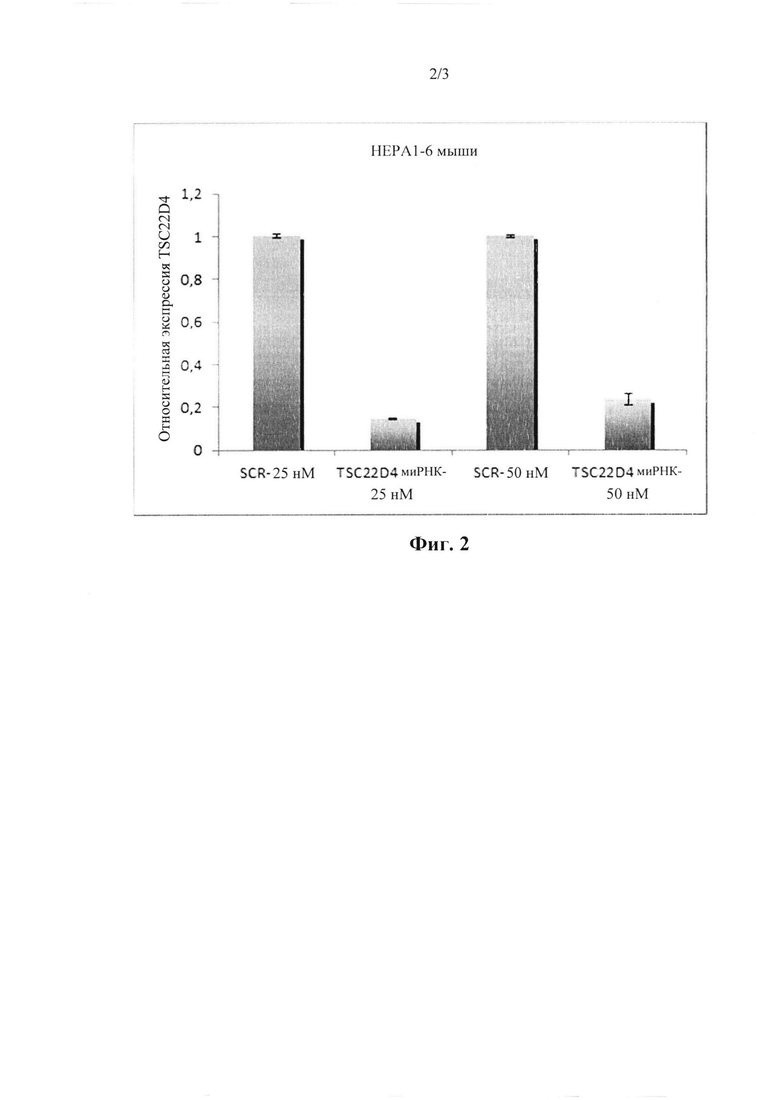
На фиг. 2 показана эффективность нокдауна mhD4-siRNA1 при трансфекции в мышиные клетки гепатомы Нера1-6 в отношении мышиного TSC22D4. Показано относительное содержание мРНК.
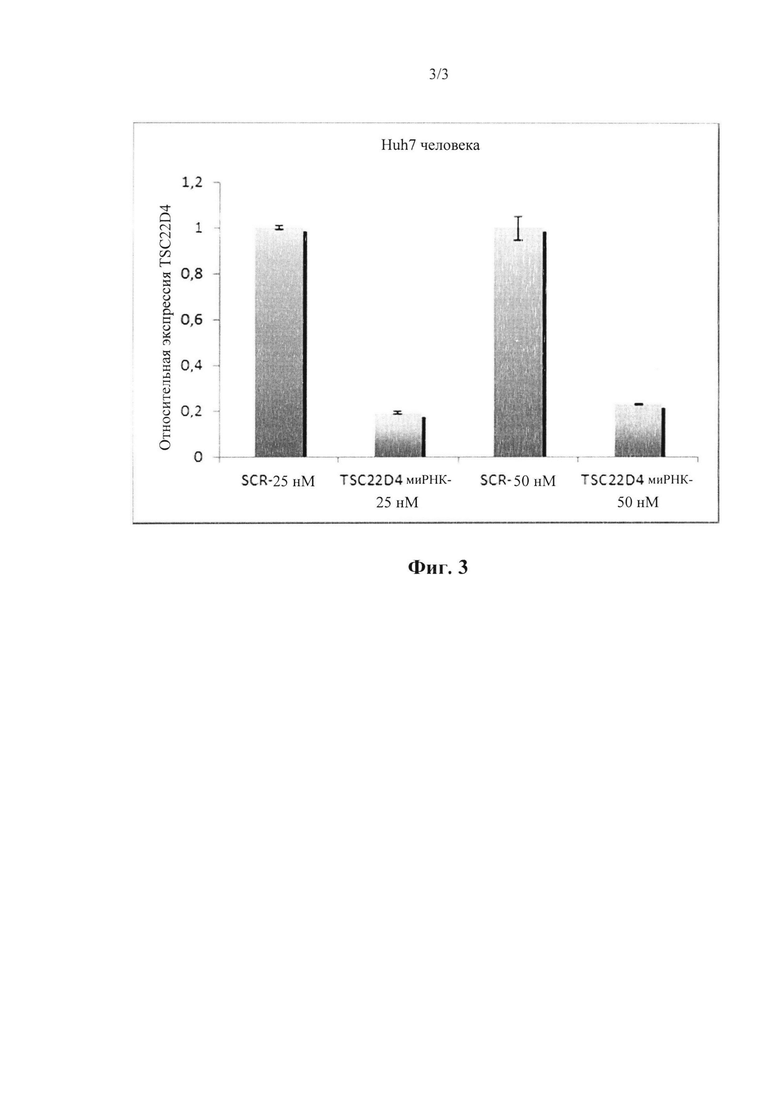
На фиг. 3 показана эффективность нокдауна mhD4-siRNA1 при трансфекции в клетки гепатомы человека Huh7 по отношению к человеческому TSC22D4. Показано относительное содержание мРНК.
Последовательности с ID NO: 1-6 показывают олигонуклеотидные последовательности в соответствии с настоящим изобретением.
ПРИМЕРЫ
Рекомбинантные вирусы
Аденовирусы, экспрессирующие TSC22D4 или неспецифическую кшРНК под контролем промотора U6, или кДНК TSC22D4 под контролем промотора CMV клонировали с использованием экспрессионной системы RNAi Adenoviral BLOCK-iT (Invitrogen, Karlsruhe, Германия). Вирусы очищали способом с помощью хлорида цезия и диализовали против забуференного фосфатом солевого буфера, содержащего 10% глицерин перед инъекцией животным, как описано ранее (Herzig S, Hedrick S, Morantte I, Koo SH, Galimi F, Montminy M. CREB controls hepatic lipid metabolism through nuclear hormone receptor PPAR-gamma. Nature. 2003; 426: 190-193. Herzig S, Long F, Jhala US, Hedrick S, Quinn R, Bauer A, Rudolph D, Yoon C, Puigserver P, Spiegelman B, et al. CREB regulates hepatic gluconeogenesis through the coactivator PGC-1. Nature. 2001; 413: 179-183). Кодирующий AAV контроль или специфические к TSC22D4 миРНК под контролем специфического для гепатоцитов промотора устанавливали, как описано ранее (Rose AJ, Frosig С, Kiens В, Wojtaszewski JF, Richter EA. Effect of endurance exercise training on Ca2+ calmodulin-dependent protein kinase II expression and signaling in skeletal muscle of humans. J Physiol. 2007; 583: 785-795).
Эксперименты на животных
Самцов мышей С57В1/6 в возрасте 8-12 недель и мышей db/db в возрасте 10 недель получали из лабораторий Charles River (Брюссель, Бельгия) и содержали в условиях с 12 часовым циклом день-ночь с регулярным неограниченным питанием. До проведения исследований на толерантность к инсулину и глюкозе животных не кормили в течение 4 часов. В противном случае животных кормили ad libitum и они имели свободный доступ к воде. Для инъекций аденовируса 1-2×109 бляшкообразующих единиц (pfu) на рекомбинантный вирус вводили путем инъекции в хвостовую вену. Для экспериментов AAV через хвостовую вену вводили 5×1011 вирусов. В каждом эксперименте 6-12 животных получали идентичное воздействие и их анализировали в условиях голодания (голодание в течение 18 ч), случайного питания или условиях сытости (голодание в течение 18 ч, затем повторное питание в течение 6 ч), как указано. Органы, включая в себя печень, эпидидимальные и брюшные жировые подушечки и икроножные мышцы, собирали через определенные периоды времени, взвешивали, замораживали и использовали для дальнейшего анализа. Общее содержание жира в организме определяли с помощью анализатора состава тела Echo MRI (Echo Medical Systems, Хьюстон, США). Содержание животных и эксперименты проводились в соответствии с руководящими принципами NIH и утверждались местными властями.
Для исследований на толерантность к инсулину готовили исходный раствор 1 Ед инсулина/мл с использованием 0,9% хлорида натрия. Мыши голодали в течение 4 часов до эксперимента. Определяли массу тела всех животных и измеряли содержание глюкозы в крови путем отрезания кончика хвоста лезвием бритвы. Каплю крови помещали на полоску глюкометра и измеряли. 1 Ед инсулина/кг массы тела инъецировали мышам С57В1/6, а 1,5 Ед инсулина/кг массы тела инъецировали мышам db/db внутрибрюшинно. Содержание глюкозы в крови контролировали через 20, 40, 60, 80 и 120 мин.
Для исследования толерантности к глюкозе готовили раствор 20% глюкозы с использованием 0,9% хлорида натрия. Мыши голодали в течение 4 часов до эксперимента. Определяли массу тела всех животных и измеряли содержание глюкозы в крови путем отрезания кончика хвоста лезвием бритвы. Каплю крови помещали на полоску глюкометра и измеряли. 5 мкл на грамм 20% раствора глюкозы вводили мышам С57В1/6 и db/db внутрибрюшинно. Содержание глюкозы в крови контролировали через 20, 40, 60, 80 и 120 мин.
Количественная ОТ-ПЦР Taqman
Общую РНК экстрагировали из гомогенизированной мышиной печени или клеточных лизатов с использованием реагента Qiazol (Qiagen, Хильден, Германия). кДНК получали путем обратной транскрипции с использованием фермента M-MuLV и праймера dT Oligo (Fermentas, Санкт-Леон-Рот, Германия). кДНК амплифицировали с использованием наборов для запрашиваемого анализа и детектора последовательности ABIPRISM 7700 (Applied Biosystems, Дармштадт, Германия). Данные экспрессии РНК нормировали до уровней РНК связывающего ТАТА-box белка (ТВР).
Экспрессию мРНК TSC22D4 человека измеряли с помощью количественной ОТ-ПЦР в реальном времени в флуоресцентном температурном циклере с использованием анализа TaqMan, а флуоресценцию обнаруживали на детекторе последовательностей ABI PRISM 7000 (Applied Biosystems, Дармштадт, Германия). Общую РНК выделяли с использованием TRIzol (Life technologies, Гранд-Айленд, Нью-Йорк), и 1 мкг РНК обратно транскрибировали посредством стандартных реагентов (Life Technologies, Гранд-Айленд, Нью-Йорк). Из каждой ОТ-ПЦР 2 мкл амплифицировали в 26 мкл ПЦР-реакции с использованием набора реагентов Brilliant SYBR green QPCR Core из Stratagene (La Jolla, CA) в соответствии с инструкциями производителя. Образцы инкубировали в детекторе последовательностей ABI PRISM 7000 для начальной денатурации при 95°С в течение 10 минут с последующими 40 циклами ПЦР, каждый цикл предусматривал температуру 95°С в течение 15 с, 60°С в течение 1 мин и 72°С для 1 мин. Экспрессию мРНК человеческих TSC22D4 и Obp2a (LCN13) (определяемые по Hs00229526_m1 и Hs01062934_g1, соответственно) (Applied Biosystems, Дармштадт, Германия) рассчитывали относительно экспрессии мРНК гипоксантинфосфорибозилтрансферазы 1 (HPRT1), определяемой посредством предварительным запрашиваемого анализа для HPRT1 (Hs01003267_m1) (Applied Biosystems, Дармштадт, Германия). Амплификацию специфических транскриптов подтверждали профилями кривой плавления (охлаждение образца до 68°С и медленное нагревание до 95°С с измерением флуоресценции) в конце каждой ПЦР. Специфичность ПЦР дополнительно подтверждали, подвергая продукты амплификации электрофорезу в агарозном геле.
Анализ белка
Белок экстрагировали из замороженных образцов органов или культивируемых гепатоцитов в буфере для лизиса клеток (Rose AJ, Frosig С, Kiens В, Wojtaszewski JF, Richter EA. Effect of endurance exercise training on Ca2+ calmodulin-dependent protein kinase II expression and signaling in skeletal muscle of humans. J Physiol. 2007; 583: 785-795), и 20 мкг белка загружали на 4-12% ДСН-полиакриламидные гели и промокали на нитроцеллюлозные мембраны. Вестерн-блот-анализ проводили, как описано (Herzig et al., 2001) с использованием антител, специфичных к TSC22D4 (Abeam, Кембридж, Великобритания или Sigma, Мюнхен, Германия), АКТ, р-АКТ, GSK, p-GSK (Cell signaling, Данверс, США) или VCP (Abcam).
Плазмиды и РНК-интерференция
Для экспериментов с кшРНК олигонуклеотиды, нацеленные на мышь и TSC22D4 (SEQ ID No. 1-3), клонировали в вектор кшРНК pENTR/U6 (Invitrogen).
Клеточная культура и анализы временной трансфекции
Первичные мышиные гепатоциты выделяли и культивировали, как описано (Klingmuller U, Bauer A, Bohl S, Nickel PJ, Breitkopf K, Dooley S, Zellmer S, Kern C, Merfort I, Sparna T, et al. Primary mouse hepatocytes for systems biology approaches: a standardized in vitro system for modelling of signal transduction pathways. IEE Proc Syst Biol. 2006; 153: 433-447). Вкратце, 8-12-недельным самцам мышей С57 В1/6 для анестезии вводили внутрибрюшинно инъекцию 100 мг/кг массы тела кетамина гидрохлорида и 5 мг/кг массы тела ксилазина гидрохлорида. После вскрытия брюшной полости печень перфузировали при 37°С с помощью HANKS I (8 г NaCl, 0,4 г KCl, 3,57 г Hepes, 0,06 г Na2HPO4 × 2 H2O, 0,06 г KH2PO4 в 1 л дистиллированной H2O, 2,5 мМ EGTA, 0,1% глюкозы, доводили до рН 7,4) через портальную вену в течение 5 мин, а затем с помощью HANKS II (8 г NaCl, 0,4 г KCl, 3,57 г Hepes, 0,06 г Na2HPO4 × 2 H2O, 0,06 г КН2РО4 в 1 л дистиллированной H2O, 0,1% глюкозы, 3 мг/мл коллагеназы CLSII, 5 мМ CaCl2, доводили до рН 7,4) в течение 5-7 минут до тех пор, пока не наблюдалась дезинтеграция структуры печени. Капсулу печени удаляли и клеточную суспензию фильтровали через сетку размером 100 мкм. Клетки промывали и затем жизнеспособность клеток определяли окрашиванием трипановым синим. 1000000 живых клеток/лунку высевали на 6-луночные планшеты с покрытием коллагеном I. Через 24 ч клетки инфицировали рекомбинантными аденовирусами с множественностью заражения 100. Для экспериментов на стимуляцию первичные гепатоциты обрабатывали PBS (контрольная среда) или инсулином в концентрации 100 нМ/6 лунок в течение 10 минут. Клетки собирали через 48 часов после инфицирования.
Цистроновый анализ TSC22D4 печени
Анализ KEGG-Pathway результатов чип-секвенирования сортировали по значимости. Установлено, что сигнальный путь инсулина значительно регулируется (р=0,00005). Чип-секвенирование проводили в экстрактах печени от инъецированных аденовирусной кДНК мышей Flag-TSC22D4 самцов мышей С57В1/6 через 7 дней после инъекции.
Результаты
Последовательности работали очень эффективно как для мышиных, так и для человеческих TSC, как показано в 4 независимых экспериментах (смотрите фигуры). Существует неспецифический липкий конец dTdT, прикрепленный к каждой последовательности. Последовательности соответствовали как мышиным, так и человеческим TSC до 100%.
Исходя из этих результатов, последовательности в соответствии с настоящим изобретением (SEQ ID №1-3) выбирали в качестве первичных кандидатов для применения в терапевтических целях, поскольку они демонстрируют превосходную эффективность нокдауна в отношении TSC22D4 и нацеливании на различные виды, включая в себя мышь, нечеловекообразных приматов и людей. Последовательности идентифицировали, функционально исследовали и проверяли различные миРНК, направленные против последовательности мРНК TSC22D4, в исследованиях нокдауна in vitro с использованием мышиного Нера1.6, а также клеток гепатомы Huh7 человека. В частности, mhD4-siRNA1 показала превосходную эффективность нокдауна в отношении TSC22D4 и нацеленно воздействует на множество видов, включая в себя мышь, нечеловекообразных приматов и людей.

--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДОЙЧЕС КРЕБСФОРШУНГСЦЕНТРУМ ШТИФТУНГ ДЕС ЭФФЕНТЛИХЕН РЕХТС
<120> ОЛИГОНУКЛЕОТИДНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ, НАЦЕЛЕННЫЕ НА ТРАНСКРИПЦИОННЫЙ ФАКТОР TSC22D4, ДЛЯ ЛЕЧЕНИЯ РЕЗИСТЕНТНОСТИ К ИНСУЛИНУ
<130> D31385WO
<150> 15160259.6
<151> 03-23-2015
<160> 6
<170> PatentIn version 3.5
<210> 1
<211> 19
<212> RNA
<213> Mus musculus
<400> 1
ggacgugugu ggauguuua 19
<210> 2
<211> 19
<212> RNA
<213> mus musculus
<400> 2
uaaacaucca cacacgucc 19
<210> 3
<211> 19
<212> RNA
<213> mus musculus
<400> 3
ggauguuuac gagagagau 19
<210> 4
<211> 19
<212> RNA
<213> mus musculus
<400> 4
aucucucucg uaaacaucc 19
<210> 5
<211> 19
<212> RNA
<213> homo sapiens
<400> 5
agucccaccu cauguuugc 19
<210> 6
<211> 19
<212> RNA
<213> homo sapiens
<400> 6
gcaaacauga ggugggacu 19
<---
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая ингибитор экспрессии TSC22D4, рекомбинантный вектор экспрессии, рекомбинантную клетку, фармацевтическую композицию, применение вышеуказанного ингибитора, вектора, клетки или композиции для профилактики, регуляции и/или лечения заболевания, выбранного из резистентности к инсулину, гипертонии, дислипидемии, заболевания коронарной артерии, метаболического синдрома и/или сахарного диабета 1 или 2 типа и/или для улучшения чувствительности к инсулину и терапевтический набор для профилактики, регуляции и/или лечения вышеуказанных заболеваний. В одном из вариантов ингибитор экспрессии TSC22D4 представляет собой интерферирующую рибонуклеиновую кислоту, белковую нуклеиновую кислоту или закрытую нуклеиновую кислоту и содержит по меньшей мере одну из следующих последовательностей:
5'-GGACGUGUGUGGAUGUUUAdTdT-3';
5'-GGAUGUUUACGAGAGAGAUdTdT-3';
5'-AGUCCCACCUCAUGUUUGCdTdT-3'.
Изобретение расширяет арсенал средств для ингибирования экспрессии TSC22D4. 6 н. и 7 з.п. ф-лы, 3 ил., 6 пр.
1. Ингибитор экспрессии TSC22D4, выбранный из олигонуклеотида, который представляет собой интерферирующую рибонуклеиновую кислоту, PNA (белковую нуклеиновую кислоту) или LNA (закрытую нуклеиновую кислоту), содержащий по меньшей мере одну из следующих последовательностей:
5'-GGACGUGUGUGGAUGUUUAdTdT-3';
5'-GGAUGUUUACGAGAGAGAUdTdT-3';
5'-AGUCCCACCUCAUGUUUGCdTdT-3';
их антисмысловой последовательности.
2. Ингибитор по п. 1, в котором интерферирующая рибонуклеиновая кислота представляет собой малую интерферирующую рибонуклеиновую кислоту (миРНК) или короткую шпилечную рибонуклеиновую кислоту (кшРНК) или микрорибонуклеиновую кислоту (микроРНК) или их комбинации.
3. Ингибитор по п. 1 или 2, в котором миРНК составляет в длину от 19 до 30 нуклеотидов.
4. Ингибитор по любому из пп. 1-3, в котором миРНК состоит из последовательности: в соответствии с SEQ ID No. 1-3.
5. Рекомбинантный вектор экспрессии, содержащий олигонуклеотидный ингибитор по любому из пп. 1-4.
6. Рекомбинантная клетка, предпочтительно рекомбинантная гепатоцитарная клетка, для экспрессии олигонуклеотидного ингибитора экспрессии TSC22D4, содержащая олигонуклеотидный ингибитор по любому из пп. 1-4 или рекомбинантный вектор по п. 5.
7. Фармацевтическая композиция для профилактики, регуляции и/или лечения заболевания, которое выбрано из резистентности к инсулину, гипертонии, дислипидемии, заболевания коронарной артерии, метаболического синдрома и/или сахарного диабета 1 или 2 типа и/или для улучшения чувствительности к инсулину, содержащая эффективное количество по меньшей мере одного из ингибитора по любому из пп. 1-4, рекомбинантного вектора экспрессии по п. 5 и рекомбинантной клетки по п. 6, вместе с фармацевтически приемлемым носителем.
8. Фармацевтическая композиция по п. 7, причем указанная фармацевтическая композиция предназначена для введения пероральным, ректальным, трансмукозальным, трансдермальным, внутрикишечным, парентеральным, внутримышечным, интратекальным, прямо интравентрикулярным, внутривенным, внутрибрюшинным, интраназальным, внутриглазным или подкожным путем.
9. Применение ингибитора по любому из пп. 1-4, рекомбинантного вектора по п. 5, рекомбинантной клетки по п. 6 или фармацевтической композиции по п. 7 или 8, для профилактики, регуляции и/или лечения заболевания, выбранного из резистентности к инсулину, гипертонии, дислипидемии, заболевания коронарной артерии, метаболического синдрома и/или сахарного диабета 1 или 2 типа и/или для улучшения чувствительности к инсулину.
10. Применение по п. 9, причем резистентность к инсулину вызвана синдромом инсулинорезистентности, который представляет собой индуцированную диетой инсулинорезистентность и/или индуцированную ожирением инсулинорезистентность.
11. Применение по п. 9, где чувствительность к инсулину представляет собой чувствительность к инсулину в контексте опухолевого заболевания.
12. Терапевтический набор для профилактики, регуляции и/или лечения заболевания, которое выбрано из резистентности к инсулину, гипертонии, дислипидемии, заболевания коронарной артерии, метаболического синдрома и/или сахарного диабета 1 или 2 типа и/или для улучшения чувствительности к инсулину, содержащий ингибитор по любому из пп. 1-4, рекомбинантный вектор по п. 5, рекомбинантную клетку по п. 6 или фармацевтическую композицию по п. 7 или 8, необязательно вместе с подходящими буферами и вспомогательными веществами, и инструкции по использованию.
13. Терапевтический набор по п. 12, где чувствительность к инсулину представляет собой чувствительность к инсулину в контексте опухолевого заболевания.
| WO 2014202602 A1, 24.12.2014 | |||
| US 7943306 B2, 17.05.2011 | |||
| WO 2000074634 A2, 14.12.2000 | |||
| СПОСОБ ВЫЯВЛЕНИЯ ГЕНОВ-МИШЕНЕЙ ДЛЯ ДИАГНОСТИКИ И ТЕРАПИИ ЛЕЙКОЗОВ ЧЕЛОВЕКА | 2014 |
|
RU2573450C1 |
| Allan Jones et al | |||
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |

