ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Областью изобретения являются генетически модифицированные, не являющиеся человеком, животные, которые производят антитела, имеющие только тяжелую цепь, в частности, генетически модифицированные животные, которые содержат делецию нуклеотидной последовательности в последовательности гена иммуноглобулина гамма (IgG), кодирующей домен CH1 (или домен CH1 и шарнирную область), но которые способны к экспрессии IgM, у которого не отсутствует функциональный домен CH1, и, в частности, мыши, которые способны к выработке молекулы IgM дикого типа (т.е. с доменами CH1), но которые производят тяжелую цепь антител IgG, лишенную функционального домена CH1 (или домена CH1 и шарнирной области).
УРОВЕНЬ ТЕХНИКИ
У большинства животных нормальные тяжелые цепи иммуноглобулинов хорошо экспрессируются только тогда, когда они спарены с их родственными легкими цепями. У людей одиночные тяжелые цепи обнаружены при болезни тяжелых цепей, которая проявляется наличием дисфункциональных тяжелых цепей, у которых потеряны последовательности вариабельного домена, домена CH1, или вариабельного и CH1 доменов. Только тяжелые цепи, без легких цепей, встречаются у определенных видов рыб и у верблюдов. У таких тяжелых цепей отсутствует функциональный домен CH1 и имеются не относящиеся к человеку особенности в вариабельных доменах тяжелых цепей. Были предприняты попытки сделать камелизированные антитела посредством модификации мышей для экспрессии камелизированных генов, которые имитируют домены VHH, обнаруженные у верблюдов или определенных видов рыб, частично за счет удаления доменов CH1 IgM и IgG и сборки вариабельных областей тяжелых цепей для того, чтобы они походили на таковые у верблюдов и/или определенных видов рыб. Однако можно ожидать, что камелизированные антитела будут индуцировать иммунные ответы у животных, не являющихся верблюдами.
Существует необходимость в данной области в генетически модифицированных, не являющихся человеком, животных, которые производят антитела, имеющие только тяжелую цепь с не-верблюжьими доменами VH.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Фигура 1 изображает локус IgG1 дикого типа у мыши (IgG1, сверху), демонстрируя генный сегмент области JH, слитый с генным сегментом CH1, с последующей шарнирной областью, генным сегментом CH2 и генным сегментом CH3; локус IgG1, являющийся мишенью для конструкции, которая делетирует домен CH1 (IgG1∆CH1, середина); и локус IgG1, являющийся мишенью для конструкции, которая делетирует как домен CH1, так и шарнирную область (IgG∆CH1-∆шарнир, внизу).
Фигура 2 изображает мышиный ген-мишень IgG1 для создания генетически модифицированного локуса, который экспрессирует IgG1 без домена CH1.
Фигура 3 изображает мышиный ген-мишень IgG1 для создания генетически модифицированного локуса, который экспрессирует IgG1 без домена CH1 и шарнирной области.
Фигура 4 изображает локус-мишень константной области тяжелой цепи мыши для создания генетически модифицированного локуса, который экспрессирует IgG1 без домена CH1 и не экспрессирует IgG2b или IgG2a.
Фигура 5 изображает константную область тяжелой цепи мыши, являющейся мишенью для конструкции, которая делетирует как домен CH1, так и шарнирную область и которая делетирует гены IgG2b и IgG2.
Фигура 6 изображает константную область тяжелой цепи генетически модифицированной мыши с IgG1, у которого отсутствует домен CH1 или отсутствуют домен CH1 и шарнирная область (вверху), и константную область тяжелой цепи генетически модифицированной мыши с IgG1, у которого отсутствует домен CH1 или отсутствуют домен CH1 и шарнирная область, и у мыши отсутствует ген IgG2a и ген IgG2b (внизу).
На фигуре 7 представлены вестерн-блоттинги супернатантов из клеток CHO, сконструированных для того, чтобы независимо экспрессировать контроль (цитокиновый эктодомен, слитый с мышиным Fc), химерное антитело (человеческий VR)/(мышиный Fc), имеющее только тяжелую цепь, в которой отсутствует домен CH1 (hVR-mFc∆CH1), камелизированное химерное антитело (человеческий VR)/(мышиный Fc), имеющее только тяжелую цепь, в которой отсутствует домен CH1 (hVR*-mFc∆CH1), химерное антитело (человеческий VR)/(мышиный Fc), имеющее только тяжелую цепь (hVR-mFc), камелизированное химерное антитело (человеческий VR)/(мышиный Fc), имеющее только тяжелую цепь (hVR*-mFc), mFc с доменом CH1 (mFc) или без домена CH1 (mFc∆CH1).
На фигуре 8 представлены изображения вестерн-блоттингов с восстанавливающего SDS-PAGE мышиной сыворотки от мыши дикого типа (слева) и от генетически модифицированной мыши, у IgG1 которой отсутствуют домен CH1 и шарнирная область (гетерозиготная) (справа), блоты делали с анти-мышиным IgG; предоставлены схемы тяжелых цепей, поскольку они являются маркерами положения молекулярных масс.
На фигуре 9 представлены изображения Вестерн-блоттингов с невосстанавливающего SDS-PAGE мышиной сыворотки от мыши дикого типа (WT) и четырех генетически модифицированных мышей, у IgG1 которых отсутствует домен CH1 и шарнирная область (гомозиготные; обозначены как HO 1, HO 2, HO 3, HO 4, соответственно), блоты делали с анти-мышиным IgG; каждая мышь (WT или HO) представлена двумя дорожками, обозначенными квадратными скобками над линиями, соответствующими разведениям сыворотки 1:5 и 1:10 от каждого животного (последовательные полосы слева направо для каждого).
На фигуре 10 представлена схематическая диаграмма нормального антитела IgG1 (слева) и антитела, имеющего только тяжелую цепь, у которой отсутствуют домен CH1 и шарнирная область.
Фигура 11 показывает раздельные анализы сывороточных иммуноглобулинов IgG1 и IgG2b мышей дикого типа (WT) и генетически модифицированных мышей, у которых IgG1 не содержит домена CH1 и шарнирной области (HO; гомозиготная мышь, которая экспрессирует тяжелую цепь антитела, в которой отсутствуют домен CH1 и шарнирная область). Контроль представляет собой смешанную сыворотку человека.
На фигуре 12 представлены белковые последовательности одиннадцати независимых RT-ПЦР клонов, амплифицированных из РНК спленоцитов мышей, несущих последовательности генов тяжелой цепи мыши в модифицированном эндогенном локусе мышиной тяжелой цепи, лишенной последовательностей областей CH1 IgG1 и шарнира. B1=SEQ ID NO:19; B2=SEQ ID NO:21; B3=SEQ ID NO:23; B5=SEQ ID NO:25; D2=SEQ ID NO:27; D5=SEQ ID NO:29; D6=SEQ ID NO:31; E2=SEQ ID NO:33; E8=SEQ ID NO:35; E10=SEQ ID NO:37; F6=SEQ ID NO:39. Строчные буквы оснований указывают на основания, не относящиеся к зародышевой линии, которые являются результатом или мутации и/или добавления некодирующих нуклеотидов во время рекомбинации. Точки представляют искусственные пропуски в последовательности для точного выравнивания каркаса (FR) и определяющих комплементарность областей (CDR), которые обозначены над последовательностями. Для каждого клона показаны первые девять аминокислот из области CH2 эндогенной константной области IgG1 (CH2).
На фигуре 13 представлены белковые последовательности семи независимых RT-ПЦР клонов, амплифицированных из РНК спленоцитов мышей, несущих последовательности генов тяжелой цепи человека в модифицированном локусе мышиной эндогенной тяжелой цепи, лишенной последовательности области CH1 IgG1. A8=SEQ ID NO:51; C2=SEQ ID NO:53; D9=SEQ ID NO:55; C4=SEQ ID NO:57; H8=SEQ ID NO:59; A5=SEQ ID NO:61; A2=SEQ ID NO:63. Строчные буквы оснований указывают на основания, не относящиеся к зародышевой линии, которые являются результатом или мутации и/или добавления некодирующих нуклеотидов во время рекомбинации. Точки представляют искусственные пропуски в последовательности для точного выравнивания каркаса (FR) и определяющих комплементарность областей (CDR), которые обозначены над последовательностями. Для каждого клона показаны первые семь аминокислот из тринадцати аминокислот шарнирной области эндогенной константной области IgG1 (HINGE).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Предлагаются генетически модифицированные клетки, не принадлежащие человеку эмбрионы, животные, не являющиеся человеком, и способы и композиции для их создания и применения, где животные генетически модифицированы для потери последовательности функционального CH1 в иммуноглобулине G (IgG), необязательно модифицированы для потери функциональной шарнирной области IgG в модифицированном IgG, и где клетки, эмбрионы и животные содержат последовательность функционального CH1 IgM. В некоторых аспектах мыши содержат замену одного или нескольких, или всех, эндогенных генных сегментов вариабельной области тяжелой цепи иммуноглобулина мыши одним или несколькими генными сегментами вариабельной области тяжелой цепи иммуноглобулина человека. В некоторых аспектах все эндогенные генные сегменты V, D и J мыши замещены одним или несколькими генными сегментами V человека, одним или несколькими генными сегментами D человека, и одним или несколькими генными сегментами J человека.
В одном из аспектов предлагается генетически модифицированная мышь, где генетическая модификация включает модификацию нуклеотидной последовательности, кодирующей константную область IgG, где модификация приводит к потере функции домена CH1 константной области IgG. В одном из вариантов осуществления модификация, связанная с потерей функции, представляет собой делецию нуклеотидной последовательности, кодирующей домен CH1, или делецию внутри нуклеотидной последовательности, кодирующей домен CH1.
В одном из вариантов осуществления IgG выбран из IgG1, IgG2a, IgG2b и их сочетания. В одном из вариантов осуществления IgG представляет собой IgG1. В одном из вариантов осуществления IgG представляет собой IgG1, IgG2a и IgG2b.
В одном из вариантов осуществления модификация дополнительно включает делецию нуклеотидной последовательности шарнирной области IgG, который содержит модификацию CH1.
В одном из вариантов осуществления генетически модифицированная мышь выбрана из линии 129, линии C57BL/6 и помеси 129×C57BL/6. В конкретном варианте осуществления мышь является на 50% 129 и 50% C57BL/6.
В одном из вариантов осуществления генетически модифицированная мышь представляет собой линию 129, выбранную из группы, состоящей из 129P1, 129P2, 129P3, 129X1, 129S1 (например, 129S1/SV, 129S1/Svlm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129T1, 129T2 (см., например, Festing et al. (1999) Revised nomenclature for strain 129 mice, Mammalian Genome 10:836). В одном из вариантов осуществления генетически модифицированная мышь представляет собой линию C57BL, в конкретном варианте осуществления выбранную из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr, C57BL/Ola. В конкретном варианте осуществления генетически модифицированная мышь представляет собой помесь вышеуказанной линии 129 и вышеуказанной линии C57BL/6. В другом конкретном варианте осуществления мышь представляет собой помесь вышеуказанных линий 129, или помесь вышеуказанных линий BL/6. В конкретном варианте осуществления линия 129 помеси представляет собой линию 129S6 (129/SvEvTac).
В одном из вариантов осуществления мышь содержит один или несколько нереаранжированных эндогенных генных сегментов вариабельной области тяжелой цепи иммуноглобулина мыши (mVR), функционально связанных с модифицированной последовательностью константной области IgG. В одном из вариантов осуществления один или несколько генных сегментов mVR представляют собой семейство генов VH мыши, выбранное из VH1, VH3, VH5, VH7, VH14 и их сочетания. В одном из вариантов осуществления один или несколько генных сегментов mVR выбраны из mVH 1-26, 1-42, 1-50, 1-58, 1-72, 3-6, 5-6, 7-1, 14-2 и их сочетания.
В одном из вариантов осуществления мышь содержит реаранжированный ген, который кодирует FR1, FR2 и FR3 в тяжелой цепи IgG, в которой отсутствует функциональная область CH1, где каждый из FR1, FR2 и FR3 независимо идентичны, по меньшей мере, на 90%, 95%, 96%, 97%, 98% или 99% FR1, FR2 и FR3, полученным из последовательности зародышевой линии mVH, выбранной из семейства генов VH1, VH3, VH5, VH7 и VH14. В одном из вариантов осуществления последовательность зародышевой линии mVH выбрана из 1-26, 1-42, 1-50, 1-58, 1-72, 3-6, 5-6, 7-1 и 14-2 последовательностей.
В одном из вариантов осуществления мышь содержит CDR3, полученную из генного сегмента DH, выбранного из DH 1-1, 2-14, 3-1, 3-2, 3-3, 4-1 и их сочетания. В одном из вариантов осуществления мышиная CDR3 содержит последовательность, кодируемую JH, который представляет собой JH1, JH2, JH3 или JH4.
В одном из вариантов осуществления мышь содержит реаранжированную последовательность антитела, кодирующую CDR3, которая получена реаранжировкой DH 1-1, 2-14, 3-1, 3-2, 3-3, 4-1 и JH1, JH2, JH3 или JH4.
В одном из вариантов осуществления мышь содержит реаранжированный ген, который кодирует FR4 в тяжелой цепи IgG, в которой отсутствует функциональная область CH1, где FR4 является идентичным, по меньшей мере, на 90%, 95%, 96%, 97%, 98% или 99% FR4, который кодируется реаранжировкой DH 1-1, 2-14, 3-1, 3-2, 3-3 или 4-1 с JH1, JH2, JH3 или JH4.
В одном из вариантов осуществления мышь содержит нереаранжированный генный сегмент вариабельной области тяжелой цепи иммуноглобулина человека (hVR) в эндогенном локусе вариабельной области тяжелой цепи мыши. В одном из вариантов осуществления мышь содержит нереаранжированный генный сегмент hVR, функционально связанный с модифицированной последовательностью константной области IgG в эндогенном локусе вариабельной области тяжелой цепи мыши. В одном из вариантов осуществления генные сегменты hVR принадлежат к семейству генов человека VH, выбранному из VH1, VH3, VH4 и их сочетания. В одном из вариантов осуществления один или несколько генных сегментов hVR выбраны из 1-2, 1-8, 1-18, 1-46, 1-69, 3-21, 3-72 и 4-59. В конкретном варианте осуществления один или несколько генных сегментов hVR выбраны из 1-8, 1-18 и 1-69.
В одном из вариантов осуществления все или по существу все генные сегменты V тяжелой цепи мыши замещены одним или несколькими генными сегментами V тяжелой цепи человека. В одном из вариантов осуществления все генные сегменты V и D тяжелой цепи мыши замещены одним или несколькими генными сегментами V и D тяжелой цепи человека. В одном из вариантов осуществления генные сегменты V, D и J тяжелой цепи мыши замещены одним или несколькими генными сегментами V тяжелой цепи человека, одним или несколькими генными сегментами D тяжелой цепи человека и одним или несколькими генными сегментами J тяжелой цепи человека. В этих вариантах осуществления генные сегменты V и/или D и/или J тяжелой цепи человека находятся в эндогенном локусе тяжелой цепи мыши и функционально связаны с геном(ами) константной области мыши или модифицированным геном(ами) константной области мыши.
В одном из вариантов осуществления мышь содержит нуклеотидную последовательность, которая кодирует последовательности FR1, FR2 и FR3 тяжелой цепи IgG, в которой отсутствует функциональная область CH1, которые, по меньшей мере, на 80% идентичны FR1, FR2 и FR3 из нуклеотидной последовательности зародышевой линии человека из генного сегмента 1-8, 1-18 или 1-69 вариабельной области тяжелой цепи иммуноглобулина человека; где FR1+FR2+FR3 последовательность модифицированной мыши оптимально выравнена с упомянутой последовательностью зародышевой линии человека безотносительно к последовательностям CDR мыши и человека (т.е., оптимальное выравнивание FR, без обсуждения идентичности аминокислот любых сравниваемых CDR в сравнении). В конкретных вариантах осуществления FR1, FR2 и FR3 идентичны приблизительно на 85%, 90%, 95%, 96%, 97%, 98% или 99% с последовательностью FR1+FR2+FR3 генного сегмента вариабельной области тяжелой цепи зародышевой линии человека, который представляет собой генный сегмент 1-8, 1-18 или 1-69.
В одном из вариантов осуществления мышь дополнительно содержит FR4, который является идентичным, по меньшей мере, на 80% FR4, образованному реаранжировкой D6-19/J6 человека, реаранжировкой D6-7/J4, реаранжировкой D4-4/J4, реаранжировкой D6-6/J2, реаранжировкой D3-16/J6, реаранжировкой D6-6/J4 и реаранжировкой D1-7/J4. В конкретных вариантах осуществления FR4 является идентичным приблизительно на 85%, 90%, 95%, 96%, 97%, 98% или 99% FR4, образованному вышеуказанной реаранжировкой D/J.
В одном из вариантов осуществления мышь содержит нуклеотидную последовательность, кодирующую FR1, чья аминокислотная последовательность отличается не более чем на 1, не более чем на 2, не более чем на 3, не более чем на 4 или не более чем на 5 аминокислот от FR1, который кодируется врожденной последовательностью генного сегмента вариабельной области тяжелой цепи человека, выбранной из V1-8, V1-18 и V1-69. В конкретном варианте осуществления нуклеотидная последовательность, кодирующая FR1, представляет собой реаранжированную последовательность, функционально связанную с последовательностью, кодирующей константную область IgG, в которой отсутствует последовательность функционального CH1.
В одном из вариантов осуществления мышь содержит нуклеотидную последовательность, кодирующую FR2, чья аминокислотная последовательность отличается не более чем на 1, не более чем на 2, не более чем на 3, не более чем на 4 или не более чем на 5 аминокислот от FR2, который кодируется врожденной последовательностью генного сегмента вариабельной области тяжелой цепи человека, выбранной из V1-8, V1-18 и V1-69. В конкретном варианте осуществления нуклеотидная последовательность, кодирующая FR2, представляет собой реаранжированную последовательность, функционально связанную с последовательностью, кодирующей константную область IgG, в которой отсутствует последовательность функционального CH1.
В одном из вариантов осуществления мышь содержит нуклеотидную последовательность, кодирующую FR3, чья аминокислотная последовательность отличается не более чем на 1, не более чем на 2, не более чем на 3, не более чем на 4, не более чем на 5, не более чем на 6, не более чем на 7, не более чем на 8, не более чем на 9, не более чем на 10 или не более чем на 11 аминокислот от FR3, который кодируется врожденной последовательностью генного сегмента вариабельной области тяжелой цепи человека, выбранной из V1-8, V1-18 и V1-69. В конкретном варианте осуществления нуклеотидная последовательность, кодирующая FR3, представляет собой реаранжированную последовательность, функционально связанную с последовательностью, кодирующей константную область IgG, в которой отсутствует последовательность функционального CH1.
В одном из вариантов осуществления мышь содержит нуклеотидную последовательность, кодирующую FR4, чья аминокислотная последовательность отличается не более чем на 1, не более чем на 2, не более чем на 3 аминокислоты от FR4, который кодируется реаранжировкой D6-19/J6, D6-7/J4, D4-4/J4, D6-6/J2, D3-16/J6, D6-6/J4 и D1-7/J4 человека. В конкретном варианте осуществления нуклеотидная последовательность, кодирующая FR4, представляет собой реаранжированную последовательность, функционально связанную с последовательностью, кодирующей константную область IgG, в которой отсутствует последовательность функционального CH1.
В одном из вариантов осуществления мышь содержит нуклеотидную последовательность, кодирующую CDR3 тяжелой цепи, полученную из генного сегмента области D тяжелой цепи человека (hDH). В одном из вариантов осуществления hDH выбрана из D1-7, D3-16, D4-4, D6-6, D6-7 и D6-19.
В одном из вариантов осуществления мышь содержит нуклеотидную последовательность, кодирующую CDR3 тяжелой цепи, полученную из генного сегмента соединительного пептида тяжелой цепи (JH) человека. В конкретном варианте осуществления JH выбран из J2, J4 и J6.
В одном из вариантов осуществления мышь содержит CDR3 тяжелой цепи, которая кодируется нуклеотидной последовательностью, полученной из реаранжировки DH и JH человека. В конкретном варианте осуществления CDR3 получена из реаранжировки D1-7/J4, D3-16/J6, D4-4/J4, D6-6/J2, D6-6/J4, D6-7/J4 или D6-19/J6.
В одном из вариантов осуществления мышь содержит замену эндогенного генного сегмента mVR генным сегментом hVR. В конкретном варианте осуществления замена эндогенного генного сегмента mVR генным сегментом hVR представляет собой тот же аллель, что и модифицированная константная область тяжелой цепи. В другом конкретном варианте осуществления замена генного сегмента mVR генным сегментом hVR представляет собой аллель, отличный от аллеля модифицированной константной области тяжелой цепи.
В одном из вариантов осуществления 90-100% генных сегментов mVR замещены, по меньшей мере, одним генным сегментом hVR. В конкретном варианте осуществления все или по существу все эндогенные генные сегменты mVR замещены, по меньшей мере, одним генным сегментом hVR. В одном из вариантов осуществления замена происходит при участии, по меньшей мере, 18, по меньшей мере, 39 или, по меньшей мере, 80 или 81 генного сегмента hVR. В одном из вариантов осуществления замена происходит при участии, по меньшей мере, 12 генных сегментов функциональной hVR, по меньшей мере, 25 генных сегментов функциональной hVR или, по меньшей мере, 43 генных сегментов функциональной hVR.
В одном из вариантов осуществления генетически модифицированная мышь содержит трансген, который состоит, по меньшей мере, из одного нереаранжированного генного сегмента hVR, по меньшей мере, из одного нереаранжированного сегмента D человека, по меньшей мере, из одного нереаранжированного сегмента J человека и, по меньшей мере, из одной константной последовательности тяжелой цепи человека. В одном из вариантов осуществления эндогенные локусы вариабельной области тяжелой цепи мыши и вариабельной области легкой цепи каппа функционально выключены. В конкретном варианте осуществления мышь способна к транс-переключению для выработки химерного антитела человек/мышь, содержащего вариабельный домен тяжелой цепи человека, примыкающий к последовательности IgG мыши, в которой отсутствует функциональный домен CH1 и, необязательно, отсутствует шарнирная область IgG, в котором нет функционального домена CH1. В конкретном варианте осуществления трансген дополнительно содержит последовательность IgG, в которой отсутствует домен CH1, и необязательно содержит IgM с функциональным доменом CH1. В дополнительном конкретном варианте осуществления в последовательности IgG отсутствует шарнирная область.
В одном из вариантов осуществления мышь содержит первый аллель вариабельной области тяжелой цепи и второй аллель вариабельной области тяжелой цепи, где первый аллель и второй аллель принадлежат одной и той же линии мыши. В одном из вариантов осуществления первый аллель относится к первой линии мыши и второй аллель относится ко второй линии мыши. В одном из вариантов осуществления один аллель из первого и второго аллелей содержит замену mVR, по меньшей мере, одной hVR. В другом варианте осуществления оба аллеля содержат замену mVR, по меньшей мере, одной hVR.
В одном из аспектов предлагается генетически модифицированная мышь, где мышь экспрессирует IgM, содержащий домен CH1, и мышь экспрессирует IgG, у которого отсутствует функциональный домен CH1, или которая экспрессирует IgG, у которого отсутствуют как функциональный домен CH1, так и функциональная шарнирная область.
В одном из вариантов осуществления IgG представляет собой IgG1.
В одном из вариантов осуществления мышь экспрессирует четыре IgG, а именно: модифицированный IgG1 и IgG3, IgG2a и IgG2b дикого типа. В другом варианте осуществления мышь экспрессирует не более чем два IgG, а именно: модифицированный IgG1 и IgG3 дикого типа. В конкретном варианте осуществления мышь экспрессирует изотипы тяжелой цепи, а именно: IgM дикого типа, IgD дикого типа, IgG3 дикого типа, модифицированный IgG1, IgG2a дикого типа, IgG2b дикого типа, IgA дикого типа и IgE дикого типа. В другом конкретном варианте осуществления мышь экспрессирует изотипы тяжелой цепи, а именно: IgM дикого типа, IgD дикого типа, IgG3 дикого типа, модифицированный IgG1, IgA дикого типа и IgE дикого типа. В различных вариантах осуществления модификация IgG1 включает делецию домена CH1 и, необязательно, делецию шарнирной области.
В одном из вариантов осуществления мышь относится к линии, выбранной из 129, C56BL/6 и смешанной 129×C57BL/6.
В одном из аспектов предлагается мышь, которая экспрессирует тяжелую цепь антитела, где антитело, имеющее только тяжелую цепь, состоит по существу из димерной тяжелой цепи, где в тяжелой цепи отсутствует функциональный домен CH1 или отсутствуют функциональный домен CH1 и функциональная шарнирная область; тяжелая цепь содержит вариабельный домен тяжелой цепи млекопитающего, который включает последовательность, не идентичную вариабельному домену тяжелой цепи млекопитающего, который кодируется геном вариабельной области зародышевой линии, и тяжелая цепь содержит домен CH2 человека или мыши и домен CH3 человека или мыши; где мышь экспрессирует IgM дикого типа человека или мыши.
В одном из вариантов осуществления мышь содержит функциональный генетический локус легкой цепи иммуноглобулина.
В одном из вариантов осуществления вариабельный домен тяжелой цепи млекопитающего представляет собой вариабельный домен тяжелой цепи человека или мыши.
В одном из вариантов осуществления антитело, имеющее только тяжелую цепь, состоит, в основном, из димерной тяжелой цепи, в которой отсутствуют функциональный домен CH1 и функциональная шарнирная область, где тяжелая цепь включает человеческий вариабельный домен, который содержит, по меньшей мере, одну соматическую мутацию, и включает домен CH2 и домен CH3. В конкретном варианте осуществления домен CH2 и домен CH3 независимо выбраны из доменов мыши и человека. В конкретном варианте осуществления домены CH2 и CH3 являются человеческими; в другом варианте осуществления домены CH2 и CH3 являются мышиными.
В одном из аспектов предлагается антитело, имеющее только тяжелую цепь, где антитело, имеющее только тяжелую цепь, содержит тяжелую цепь, включающую не-верблюжий вариабельный домен и константную область тяжелой цепи, в которой отсутствует домен CH1.
В одном из вариантов осуществления у антитела, имеющего только тяжелую цепь, дополнительно отсутствует шарнирная область.
В одном из вариантов осуществления антитело, имеющее только тяжелую цепь, содержит константную область, которая состоит по существу из шарнирной области, домена CH2 и домена CH3. В другом варианте осуществления антитело, имеющее только тяжелую цепь, содержит константную область, которая состоит по существу из домена CH2 и домена CH3.
В одном из вариантов осуществления не-верблюжий вариабельный домен представляет собой соматически мутированный вариабельный домен тяжелой цепи человека или мыши, полученный из нуклеотидной последовательности, кодирующей IgM или IgG, в B-клетке мыши или генетически модифицированной мыши, несущей генный сегмент вариабельной области тяжелой цепи человека. В конкретном варианте осуществления мышь содержит гуманизированный генный сегмент вариабельной области тяжелой цепи. В другом варианте осуществления мышь содержит замену эндогенного локуса генного сегмента вариабельной области тяжелой цепи мыши, по меньшей мере, одним генным сегментом вариабельной области человека. В другом варианте осуществления мышь содержит замену эндогенного локуса тяжелой цепи мыши, по меньшей мере, одним генным сегментом вариабельной области человека, по меньшей мере, одним генным сегментом D человека и, по меньшей мере, одним генным сегментом J человека. В конкретном варианте осуществления эндогенный локус вариабельной области иммуноглобулина мыши полностью или по существу полностью замещен локусом вариабельной области иммуноглобулина человека, содержащим множество генных сегментов V, D и J человека.
В одном из вариантов осуществления не-верблюжий вариабельный домен представляет собой вариабельный домен человека или мыши. В другом варианте осуществления не-верблюжий вариабельный домен представляет собой вариабельный домен человека или мыши, который содержит одну или несколько камелизирующих модификаций. В конкретном варианте осуществления камелизирующая модификация выбрана из L11S, V37F, G44E, L45C, L45R и W47G (нумерация по Kabat). В конкретном варианте осуществления камелизирующая модификация выбрана из V37F, G44E и L45C. В конкретном варианте осуществления вариабельный домен тяжелой цепи включает определяющую комплементарность область 3 (CDR3), которая содержит два цистеина.
В одном из вариантов осуществления антитело, имеющее только тяжелую цепь, содержит димер первой тяжелой цепи, включающей первый вариабельный домен тяжелой цепи, и второй тяжелой цепи, включающей второй вариабельный домен тяжелой цепи, где в каждой из первой и второй тяжелых цепей отсутствует домен CH1 (или отсутствует домен CH1 и шарнирная область). В одном из вариантов осуществления человеческий вариабельный домен первой тяжелой цепи димера связывает первый эпитоп, и человеческий вариабельный домен второй тяжелой цепи димера связывает второй эпитоп, где первый и второй эпитопы не являются идентичными. В конкретном варианте осуществления вариабельные домены тяжелых цепей первой и второй тяжелых цепей содержат вариабельные домены тяжелых цепей человека и/или области FR тяжелой цепи человека, как описано в настоящем документе.
В одном из аспектов предлагается генетически модифицированная не принадлежащая человеку клетка, где генетическая модификация включает делецию домена CH1 IgG, и клетка экспрессирует функциональный IgM. В конкретном варианте осуществления клетка содержит ген IgM, включающий последовательность, кодирующую домен CH1.
В одном из вариантов осуществления клетка выбрана из не принадлежащих человеку эмбриональной стволовой (ES) клетки, плюрипотентной клетки и тотипотентной клетки. В конкретном варианте осуществления не принадлежащая человеку ES клетка выбрана из ES клетки мыши и ES клетки крысы.
В одном из аспектов предлагается генетически модифицированный, не принадлежащий человеку эмбрион, где генетическая модификация включает модификацию, описанную в настоящем документе. В одном из вариантов осуществления генетическая модификация включает делецию домена CH1 IgG, и не принадлежащий человеку эмбрион экспрессирует функциональный IgM. В конкретном варианте осуществления не принадлежащий человеку эмбрион содержит ген IgM, включающий домен CH1.
В одном из вариантов осуществления не принадлежащий человеку эмбрион представляет собой эмбрион мыши или эмбрион крысы.
В одном из аспектов предлагается не принадлежащий человеку эмбрион, содержащий донорскую клетку, где донорская клетка генетически модифицирована, и где генетическая модификация представляет собой модификацию, описанную в настоящем документе. В одном из вариантов осуществления генетическая модификация включает делецию домена CH1 IgG, и клетка содержит ген IgM, включающий домен CH1.
В одном из вариантов осуществления не принадлежащий человеку эмбрион представляет собой эмбрион мыши или эмбрион крысы, и донорская клетка представляет собой ES клетку мыши или ES клетку крысы, соответственно.
В одном из аспектов предлагается конструкция ДНК, где конструкция ДНК содержит (a) мышиное гомологичное плечо, гомологичное первой последовательности 5' и непосредственно примыкающее к началу области CH1 IgG; (b) маркер или кассету для отбора по устойчивости к лекарственному средству; и (c) гомологичное плечо, гомологичное второй последовательности 3' и непосредственно примыкающее к концу области CH1 IgG, или, альтернативно, гомологичное плечо, гомологичное второй последовательности 3' и непосредственно примыкающее к концу шарнирной области IgG.
В одном из аспектов предлагается способ получения антитела, в котором отсутствует домен CH1, включающий: (a) иммунизацию не являющегося человеком животного, как описано в настоящем документе, у которого отсутствует функциональный домен CH1 в IgG или отсутствует функциональный домен CH1 и отсутствует функциональная шарнирная область в IgG, где мышь экспрессирует IgM, который содержит функциональный домен CH1; (b) содержание не являющегося человеком животного в условиях, достаточных для выработки антитела; (c) идентификацию антитела, выработанного мышью, в котором отсутствует функциональный домен CH1 или в котором отсутствует функциональная шарнирная область; и (d) выделение из мыши антитела, клетки, которая вырабатывает антитело, или нуклеотидной последовательности, которая кодирует последовательность антитела.
В одном из вариантов осуществления не являющееся человеком животное содержит генетический локус функциональной легкой цепи иммуноглобулина.
В одном из аспектов предлагается способ для гуманизации мышиного антитела, имеющего только тяжелую цепь, включающий иммунизацию посредством интересующего антигена генетически модифицированной мыши, вырабатывающей антитела, имеющие только тяжелую цепь; возможность для мыши нарастить иммунный ответ; идентификацию у мыши мышиной области VH, которая кодируется в B-клетке мыши, где B-клетка специфически связывает интересующий антиген, и гуманизацию области VH.
В одном из вариантов осуществления генетически модифицированная мышь, которая вырабатывает антитела, имеющие только тяжелую цепь, представляет собой мышь, описанную в настоящем документе. В одном из вариантов осуществления мышь содержит, по меньшей мере, один генный сегмент mVR, функционально связанный с константным локусом тяжелой цепи, мышь содержит интактный ген IgM и ген IgG, в котором отсутствует домен CH1 или отсутствуют домен CH1 и шарнирный домен. В одном из вариантов осуществления ген IgG представляет собой ген IgG1. В одном из вариантов осуществления ген IgG выбран из IgG1, IgG2A, IgG2B, IgG3 и их сочетания.
В одном из вариантов осуществления способ дополнительно содержит клонирование нуклеотидной последовательности, кодирующей гуманизированную область VH к нуклеотидной последовательности константной области иммуноглобулина человека.
В одном из вариантов осуществления генный сегмент мыши mVR относится к генному семейству VH мыши, выбранному из VH1 и VH14, и гуманизация включает замену каркасной области мыши из VH1 или VH14 каркасной областью из гена человека VH1. В одном из вариантов осуществления ген человека VH1 выбран из 1-2, 1-3, 1-8, 1-17, 1-18, 1-24, 1-45, 1-46, 1-58 и 1-69. В конкретных вариантах осуществления ген mVR представляет собой ген 1-58, и ген человека представляет собой ген 1-18; ген mVR представляет собой ген 1-26, и ген человека представляет собой ген 1-2; ген mVR представляет собой ген 1-50, и ген человека представляет собой ген 1-46; ген mVR представляет собой ген 1-17, и ген человека представляет собой ген 1-2; ген mVR представляет собой ген 1-42, и ген человека представляет собой ген 1-2; ген mVR представляет собой ген 14-1, и ген человека представляет собой ген 1-2; или mVR представляет собой ген 14-2, и ген человека представляет собой ген 1-2.
В одном из вариантов осуществления генный сегмент mVR представляет собой ген мыши VH, выбранный из генов VH4, VH5, VH6, VH7, VH10, VH11 и VH13, и гуманизация включает замену каркасной области мыши каркасной областью из гена человека VH3. В одном из вариантов осуществления ген человека VH3 выбран из 3-7, 3-9, 3-11, 3-13, 3-15, 3-16, 3-20, 3-21, 3-23, 3-30, 3-33, 3-35, 3-38, 3-43, 3-48, 3-49, 3-53, 3-64, 3-66, 3-72, 3-73 и 3-74. В конкретном варианте осуществления ген mVR представляет собой ген 7-1, и ген человека представляет собой ген 3-72; ген mVR представляет собой ген 3-6, и ген человека представляет собой ген 4-59; ген mVR представляет собой ген 5-6, и ген человека представляет собой ген 3-21.
В одном из вариантов осуществления генный сегмент mVR относится к генному семейству VH мыши, выбранному из VH3 и VH12, и гуманизация включает замену каркасной области мыши каркасной областью из гена человека VH4. В одном из вариантов осуществления ген человека VH4 выбран из 4-4, 4-28, 4-31, 4-34, 4-39, 4-59 и 4-61.
В одном из вариантов осуществления генный сегмент mVR относится к генному семейству VH4 мыши, и гуманизация включает замену каркасной области мыши каркасной областью из гена человека VH6. В одном из вариантов осуществления ген человека VH6 представляет собой 6-1.
В одном из вариантов осуществления генный сегмент mVR относится к генному семейству VH9 мыши, и гуманизация включает замену каркасной области мыши VH9 каркасной областью из гена VH человека из семейства человека VH7. В одном из вариантов осуществления ген VH человека выбран из 7-4-1 и 7-81.
В одном из вариантов осуществления гуманизация дополнительно содержит создание одной или нескольких консервативных или неконсервативных замен, одной или нескольких делеций и/или одной или нескольких вставок в CDR мыши таким образом, что CDR мыши более соответствует CDR человека.
В одном из вариантов осуществления гуманизация дополнительно включает создание одной или нескольких консервативных или неконсервативных замен, одной или нескольких делеций и/или одной или нескольких вставок в каркасную область человека таким образом, что каркасная область человека более соответствует каркасной области мыши.
В одном из аспектов предлагается генетически модифицированная мышь, которая содержит функциональный ген легкой цепи иммуноглобулина, где мышь экспрессирует антитело, имеющее только тяжелую цепь, в котором отсутствует легкая цепь и в котором отсутствует область CH1 или отсутствуют область CH1 и шарнирная область.
В одном из вариантов осуществления мышь содержит ген иммуноглобулина, в котором отсутствует последовательность, кодирующая область CH1, или отсутствует последовательность, кодирующая шарнир и область CH1. В одном из вариантов осуществления ген иммуноглобулина, в котором отсутствует последовательность, представляет собой один или несколько генов константной области тяжелой цепи. В конкретном варианте осуществления ген иммуноглобулина, в котором отсутствует последовательность, выбран из генов IgG1, IgG2a, IgG2b и IgG3. В конкретном варианте осуществления мышь содержит ген IgM, в котором присутствует область CH1 и/или шарнирная область и/или область CH1 и шарнирная область.
В одном из вариантов осуществления антитело экспрессируется в ответ на антиген, и антитело специфически связывается с антигеном.
В одном из вариантов осуществления антитело содержит мышиный домен VH. В конкретном варианте осуществления мышиный домен VH включает генный сегмент VH мыши, выбранный из 1-26, 1-42, 1-50, 1-58, 1-72, 3-6, 5-6, 7-1, 14-1 и 14-2.
В одном из вариантов осуществления антитело содержит человеческий домен VH. В конкретном варианте осуществления человеческий домен VH включает последовательность, полученную из генного сегмента VH человека, выбранного из 1-2, 1-18, 1-46, 3-21, 3-72 и 4-59.
В одном из аспектов предлагается генетически модифицированная мышь, которая экспрессирует связывающий белок, состоящий по существу из двух тяжелых цепей IgG1, в каждой из которых отсутствует домен CH1, где мышь экспрессирует IgM, который содержит область CH1, и где мышь не способна экспрессировать из своего генома мРНК, которая содержит нуклеотидную последовательность, кодирующую домен CH1 IgGl.
В одном из вариантов осуществления тяжелые цепи иммуноглобулинов, в каждой из которых отсутствует домен CH1, состоят по существу, от N-конца к C-концу, из вариабельной области тяжелой цепи иммуноглобулина человека или мыши, необязательно, шарнирной области, области CH2 мыши и области CH3 мыши. В конкретном варианте осуществления вариабельная область тяжелой цепи иммуноглобулина представляет собой вариабельную область человека, присутствует шарнирная область, и мышь содержит функциональный генетический локус легкой цепи иммуноглобулина.
В одном из аспектов предлагается мышь, которая экспрессирует антитело, имеющее только тяжелую цепь, в котором отсутствует легкая цепь, и в котором отсутствует область CH1 полностью или частично, где мышь экспрессирует B-клеточный рецептор на B-клетке, где на поверхности B-клеточного рецептора расположена связывающая молекула, которая включает вариабельный домен тяжелой цепи иммуноглобулина, слитый непосредственно с шарнирной областью иммуноглобулина или слитый непосредственно с областью CH2, где у связывающей молекулы отсутствует область CH1. В одном варианте осуществления связывающая молекула содержит области CH2 и CH3 IgG1.
В одном из аспектов предлагается способ для получения антитела, имеющего только тяжелую цепь, включающий иммунизацию мыши интересующим антигеном, где мышь содержит ген IgG, в котором отсутствует последовательность, кодирующая область CH1, где мышь содержит интактный ген константной области IgM, что позволяет мыши наращивать иммунный ответ против интересующего антигена; и выделение из мыши клетки или белка, которые специфически распознают интересующий антиген, где клетка или белок содержат антитело, имеющее только тяжелую цепь, в котором отсутствует домен CH1, и в котором отсутствует родственная легкая цепь и которое специфически связывается с интересующим антигеном.
В одном из вариантов осуществления мышь содержит функциональный ген легкой цепи. В одном из вариантов осуществления мышь содержит функциональный ген легкой цепи, выбранный из лямбда, каппа и их сочетания.
В одном из вариантов осуществления мышь содержит замену всех или по существу всех генных сегментов V, D, J тяжелой цепи мыши одним или несколькими генными сегментами V, D, J человека.
В одном из вариантов осуществления ген IgG, в котором отсутствует последовательность, кодирующая CH1, выбран из IgG1, IgG2a, IgG2b, IgG3 и их сочетания.
В одном из вариантов осуществления ген IgG, в котором отсутствует последовательность CH1, представляет собой IgG1, и у мыши отсутствует ген, кодирующий IgG2a, IgG2b, IgG3 или их сочетание. В одном из вариантов осуществления ген IgG, в котором отсутствует последовательность CH1, представляет собой IgG2a, и у мыши отсутствует ген, кодирующий IgG1, IgG2b, IgG3 или их сочетание. В одном из вариантов осуществления ген IgG, в котором отсутствует последовательность CH1, представляет собой IgG2b, и у мыши отсутствует ген, кодирующий IgG1, IgG2a, IgG3 или их сочетание. В одном из вариантов осуществления ген IgG, в котором отсутствует последовательность CH1, представляет собой IgG3, и у мыши отсутствует ген, кодирующий IgG1, IgG2a, IgG2b или их сочетание.
В одном из вариантов осуществления мышь содержит B-клетку, несущую на своей поверхности B-клеточный рецептор, где B-клеточный рецептор содержит VDJ-реаранжированную тяжелую цепь, которая связывает интересующий антиген, и где B-клеточный рецептор содержит IgM с областью CH1, и где IgM содержит легкую цепь. В одном из вариантов осуществления легкая цепь является VJ-реаранжированной. В конкретном варианте осуществления легкая цепь представляет собой легкую цепь каппа или лямбда, которая является родственной с VDJ-реаранжированной тяжелой цепью, связывающей интересующий антиген.
В одном из аспектов предлагаются мышиное антитело, имеющее только тяжелую цепь, человеческое антитело, имеющее только тяжелую цепь, или химерное антитело человек/мышь, имеющее только тяжелую цепь, произведенные в мыши по изобретению.
В одном из аспектов предлагаются мышиное антитело, имеющее только тяжелую цепь, человеческое антитело, имеющее только тяжелую цепь, химерное антитело человек/мышь, имеющее только тяжелую цепь, или гуманизированное антитело, имеющее только тяжелую цепь, созданные с использованием нуклеотидной последовательности вариабельной области тяжелой цепи или ее фрагмента и произведенные в мыши по изобретению.
Другие варианты осуществления описаны и будут ясны специалистам в данной области из обзора последующего подробного описания.
ПОДРОБНОЕ ОПИСАНИЕ
Изобретение не ограничено конкретными способами и описанными экспериментальными условиями, поскольку такие способы и условия могут варьировать. Терминология, применяемая в настоящем документе, используется только с целью описания конкретных вариантов осуществления, и не является ограничивающей, поскольку объем настоящего изобретения ограничен только формулой изобретения.
Если не определено иное, все технические и научные термины, применяемые в настоящем документе, имеют то же значение, в каком их обычно понимают специалисты в той области, к которой относится изобретение. Хотя любые способы и материалы, аналогичные или эквивалентные способам и материалам, описанным в настоящем документе, можно использовать в практическом осуществлении или тестировании по настоящему изобретению, конкретные способы и материалы описаны здесь. Все публикации, упомянутые в настоящем документе, включены в настоящий документ в качестве ссылки в полном объеме.
Домены CH1 и выработка антител
Предлагаются генетически модифицированные, не являющиеся человеком, животные, которые вырабатывают антитела без домена CH1, в том числе антитела, имеющие только тяжелую цепь, т.е. антитела без легких цепей. Генетически модифицированные, не являющиеся человеком, животные содержат генетическую модификацию, которая включает отсутствие функционального домена тяжелой цепи иммуноглобулина (домена CH1), например, домена CH1 IgG1, и в некоторых вариантах осуществления дополнительную модификацию, включающую делецию шарнирной области в тяжелой цепи иммуноглобулина, в которой отсутствует функциональный домен CH1, где не являющееся человеком животное экспрессирует функциональный IgM. Другие модификации включают превращение изотипов, иных чем IgG1 и IgM, в нефункциональные, например, создание делеций в генах, или делеции генов для IgD, IgG3, IgG2a, IgG2b, IgA и IgE. Также предлагаются генетически модифицированные не принадлежащие человеку эмбрионы, клетки, и нацеленные конструкции для получения не являющихся человеком животных, не принадлежащих человеку эмбрионов и клеток.
Усилия, направленные на создание генетически модифицированных клеток, которые могут вырабатывать антитела, имеющие только тяжелую цепь (т.е. антитела, у которых отсутствует легкая цепь), сфокусированы на имитации антител, имеющих только тяжелую цепь, у других видов, например, у верблюдовых и некоторых рыб. Этот подход был использован для генетической модификации мышиной ES клетки для делеции доменов CH1 в генах константной области иммуноглобулинов IgM и IgG, и также для введения вариабельных областей тяжелой цепи, которые принадлежали верблюдовым или были камелизированы (т.е. VHH или VHH-подобные), в ES клетку. Делецию доменов CH1 IgM и IgG осуществляют, по-видимому, для того чтобы предотвратить образование эндогенных природных антител, которое конкурирует с образованием камелизированного антитела из генетически модифицированного локуса. Добавление генных сегментов VHH осуществляют, вероятно, для того чтобы имитировать образование антитела, имеющего только тяжелую цепь, в комбинации с делецией CH1. Антитела, имеющие только тяжелую цепь, от таких животных будут содержать генный сегмент VHH. Генные сегменты VHH, предположительно, считаются необходимыми для надлежащей экспрессии антитела, имеющего только тяжелую цепь, поскольку исследования in vitro указывают, что не-верблюжьи домены VH не могут удовлетворительно формировать способные к экспрессии антитела, имеющие только тяжелую цепь, когда они присутствуют в тяжелых цепях с отсутствием домена CH1.
У верблюдовых, однако, (и у некоторых хрящевых рыб) есть гены, которые включают домены CH1 или CH1-подобные домены. Считают, что антитела, содержащие VHH без доменов CH1, являются результатом сплайсинга РНК или перестройки последовательностей ДНК, которые могут кодировать область CH1. Таким образом, даже верблюдовые сохранили последовательности ДНК, кодирующие области CH1. Поскольку люди (при определенных обстоятельствах) могут вырабатывать антитела, имеющие только тяжелую цепь, в которой область CH1 отсутствует полностью или частично (например, болезни тяжелых цепей человека), можно было бы заставить не-верблюдовых, таких как мыши, при данных обстоятельствах формировать тяжелые цепи, в которых отсутствует область CH1. Этот подход основан на том, чтобы, не нарушая структуру CH зародышевой линии, вместо этого превратить локус легкой цепи животного в нефункциональный. Этот подход предполагает, что при наличии нефункционального локуса легкой цепи, те тяжелые цепи, которым для экспрессии необходима родственная легкая цепь, (например, полноразмерные тяжелые цепи с областями CH1) не будут вырабатываться из-за отсутствия легкой цепи каппа или лямбда, таким образом, будут экспрессироваться и секретироваться только тяжелые цепи, которые могут экспрессироваться и секретироваться без легкой цепи (т.е. тяжелые цепи, в которых отсутствует область CH1). Подход основан на отсутствии функциональных генных сегментов каппа или лямбда, которые могут перестраиваться для образования функционального гена легкой цепи, и на отсутствии любого функционального перестроенного гена легкой цепи, и, таким образом, требует генетических манипуляций (например, нокаута) для нарушения функциональности обоих локусов легкой цепи зародышевой линии. Подход основан на "природных" процессах, приводящих к неиспользованию эндогенной нуклеотидной последовательности CH1, и на том, что "природный" процесс сайленсинга CH1 происходит при переключении класса. Кажется, нет никакой возможности использовать такой способ у любого животного, которое содержит функциональный ген легкой цепи. Кроме того, судя по всему, "природный" процесс включает синтез больших количеств нормальной РНК, т.е. РНК, которая содержит область, кодирующую CH1.
Изобретение относится к композициям и способам для получения мыши, которая продуцирует антитело, в котором отсутствует иммуноглобулиновый домен CH1 (и, необязательно, шарнирная область), в том числе антитела, имеющие только тяжелую цепь, и в том числе антитела, которые содержат домены VH (например, домены VH мыши или человека). Способы предусматривают выборочное превращение эндогенной, не относящейся к IgM, области CH1 в нефункциональную (например, посредством делеции последовательности домена CH1), и использование или нереаранжированных эндогенных генных сегментов вариабельной области (mVR) мыши или нереаранжированных генных сегментов вариабельной области (hVR) человека в эндогенном локусе вариабельной области мыши для получения химерного антитела человек/мышь у мыши. Делецию домена CH1 производят в одном или нескольких генах IgG, но не в гене IgM. Такой подход выборочно превращает один или несколько доменов CH1 IgG в нефункциональные, в то же время сохраняя функциональный IgM. В дополнение к делеции одного или нескольких доменов CH1 IgG, дополнительный вариант осуществления предлагает удаление или превращение в нефункциональную шарнирной области иммуноглобулина(ов) IG, в которых домен CH1 удален или сделан нефункциональным.
Подход с делецией CH1 IgG использует относительно консервативное нарушение в развитии природной B-клетки у животного, поскольку не все изотипы Ig генетически модифицированного не являющегося человеком животного будут обладать нефункциональным CH1 или делецией домена CH1 (и, необязательно, шарнира). Таким образом, модификация CH1 не происходит в молекулах IgM и, таким образом, не нарушает те шаги в раннем развитии B-клетки, которые зависят от IgM с функциональным CH1. Поскольку IgM не модифицирован, животные, несущие одну или несколько делеций домена CH1 IgG (и, необязательно, шарнирной области IgG), но не домена CH1 IgM, должны быть способны производить удовлетворительно большой репертуар вариабельных областей на этапах клональной селекции перед презентацией вариабельного домена в случае IgG. Таким образом, в различных вариантах осуществления любое вредное воздействие генетической модификации(ий) на разнообразие вариабельных областей, доступных для применения в антителе, имеющем только тяжелую цепь, не должно негативно влиять на совокупность вариабельных областей, доступных для селекции в случае IgG. Дополнительно, когда последовательность CH1, ставшая нефункциональной в зародышевой линии (например, делетированная), находится в IgG1, мышь теряет способность производить любую РНК, которая кодирует домен CH1.
Генетическое изменение не являющегося человеком животного для превращения домена CH1 или домена CH1 и шарнирной области одного или нескольких изотипов IgG в нефункциональные может привести к мыши, которая способна к селекции из полного или по существу полного репертуара областей VH подходящей области VH для экспрессии в антителе, имеющем только тяжелую цепь. Выборочное изменение изотипов IgG (но не IgM) позволяет избежать потенциального снижения числа областей VH, которые проходят селекцию за счет отсутствия домена CH1 или отсутствия домена CH1 в IgM. Таким образом, доступен более полный репертуар областей VH для селекции в случае IgG (в котором отсутствует домен CH1 или отсутствуют домен CH1 и шарнирная область). Таким образом, селекция домена VH у генетически модифицированной мыши по изобретению не зависит, например, от того, какой домен VH может помочь преодолеть трудности на ранних этапах IgM-зависимого развития B-клетки, которые связаны с изменением структуры IgM. Наоборот, ранние этапы, зависимые от IgM, должны происходить в обычном режиме, приводя к большому репертуару тяжелых цепей, доступных для селекции в отношении их пригодности для экспрессии в случае IgG, в котором отсутствует домен CH1 или отсутствуют домен CH1 и шарнирная область.
Таким образом, в различных вариантах осуществления генетически модифицированная мышь по изобретению должна поддерживать экспрессию функционального IgM, которая должна обеспечивать возможность для более естественного процесса клональной селекции. Например, с функциональным IgM (например, IgM, у которого не потерян домен CH1), как суррогатная легкая цепь, так и родственная легкая цепь будут способны к ассоциации посредством домена CH1 IgM и участию в процессах селекции на ранних этапах развития B-клетки. Считают, что у генетически модифицированной мыши по изобретению переключение классов на изотип IgG представляет собой первый шаг селекции, где встречается любой выбор вариабельных доменов тяжелых цепей, которые могут экспрессироваться в случае константного домена, в котором отсутствует функциональный домен CH1 или в котором отсутствует функциональный домен CH1 и функциональный шарнир.
IgM в развитии B-клетки
Хотя наблюдения у верблюдовых, некоторых рыб и при патологических состояниях выявили, что при некоторых обстоятельствах антитело, в котором отсутствует домен CH1 в константной области тяжелой цепи, может экспрессироваться в отсутствие родственной легкой цепи, нормальное развитие антитело-продуцирующих B-клеток, как правило, требует наличия домена CH1. Все изотипы тяжелой цепи, включая IgM, содержат домен CH1. Полагают, что как суррогатная легкая цепь, так и родственная легкая цепь взаимодействуют с данной тяжелой цепью посредством домена CH1 тяжелой цепи в случае IgM. Поскольку формирование антител, имеющих только тяжелую цепь, зависит от структурной целостности или функциональности изотипа тяжелой цепи IgM, нарушение структурной целостности или функции IgM было бы нежелательно.
Для нормального формирования антител необходимо, чтобы антитела выживали на протяжении множества комплексных схем селекции, результатом которой является выживание и итоговая экспрессия функционального и пригодного антитела. Нарушения в структуре антитела могут оказаться вредными для выживания и итоговой экспрессии антитела, поскольку структурное нарушение приводит к неспособности антитела эффективно конкурировать и развиваться в соответствии с требованиями одной или нескольких природных схем селекции антител.
На раннем этапе формирования антител, тяжелые цепи антител проходят процесс селекции, где природа выбирает посредством ряда селекционных схем подходящие тяжелые цепи для прохождения дальнейшего отбора и образования в конечном итоге функциональных антител с созревшей аффинностью. Тяжелые цепи антител, которые экспрессировались с рекомбинировавших генных сегментов тяжелой цепи в предшественнике B-клеток (или про-B-клетках) в норме спарены с суррогатной легкой цепью для презентации на поверхности про-B-клетки в изотипе IgM для формирования структуры (которая включает другие сорецепторы), обозначаемой пре-B-клеточный рецептор, или пре-BCR. Полагают, что как только пре-BCR презентирован на поверхности клетки, он подает сигнал клетке путем соответствующего формирования комплекса, эффективно информируя клетку, что тяжелая цепь прошла этот ранний этап селекции. Таким образом, клетка информирована, что тяжелая цепь может проходить дальнейшую селекцию. Если тяжелая цепь содержит дефект, который вредит формированию пре-BCR при презентации в случае IgM и суррогатной легкой цепи, клетка подвергается апоптозу. Если клетка подвергается апоптозу, пригодность или вклад в разнообразие тяжелых цепей вариабельной области тяжелой цепи будет утерян. Таким образом, на самом раннем этапе селекции антитела необходима презентация тяжелой цепи вместе с суррогатной легкой цепью в случае изотипа IgM. Полагают, что суррогатная легкая цепь взаимодействует с IgM, по меньшей мере частично, посредством домена CH1 IgM. Недостаток или нарушение в структуре антитела на этом раннем этапе (например, нефункциональный домен CH1) может привести к неудаче клональной селекции, потере про-B-клетки, которая экспрессирует тяжелую цепь, и потере возможности использовать конкретный вариабельный домен тяжелой цепи в пригодном антителе.
Как только клетка, несущая пре-BCR, проходит данный этап селекции, для следующего этапа селекции необходимо, чтобы тяжелая цепь была спарена с родственной легкой цепью (например, каппа или лямбда у мышей и людей). Спаренная структура тяжелая цепь/родственная легкая цепь снова презентируется на поверхности клетки, теперь уже наивной пре-B-клетки, в случае изотипа IgM посредством домена CH1 IgM. Результатом этого комплекса на поверхности является функциональный мембраносвязанный B-клеточный рецептор (BCR). Полагают, что этот BCR сигнализирует клетке, что данная тяжелая цепь подходит для дальнейшей селекции, и что клетка может теперь совершать экспрессию этой конкретной легкой цепи и приступать к дальнейшим этапам созревания B-клетки, включая созревание аффинности и переключение классов. Если тяжелая цепь содержит дефект, который вредит формированию BCR при презентации в случае IgM и родственной легкой цепи, клетка подвергается апоптозу. Если клетка подвергается апоптозу, пригодность или вклад в разнообразие тяжелых цепей вариабельной области тяжелой цепи будет утерян. Таким образом, на самом раннем этапе селекции антитела требуется презентация тяжелой цепи вместе с суррогатной легкой цепью применительно к изотипу IgM. Снова, недостаток или нарушение в структуре антитела на этом раннем этапе (например, нефункциональный домен CH1) может привести к неудаче клональной селекции и сопутствующей потере про-B-клетки, которая экспрессирует тяжелую цепь.
Пережившая селекцию до настоящего времени, пре-B-клетка, которая презентирует тяжелую цепь, спаренную с родственной ей легкой цепью применительно к IgM, затем проходит процесс созревания, который, в конечном счете, приводит к переключению классов и дальнейшим механизмам селекции, когда тяжелая цепь и родственная легкая цепь презентируются на поверхности В-клетки применительно к изотипу IgG. На этом этапе будет происходить любая селекция тяжелых цепей IgG, у которых отсутствует домен CH1 или у которых отсутствуют домен CH1 и шарнирная область. Полагают, что у животных по изобретению обычный репертуар вариабельных областей тяжелых цепей будет доступен для селекции в зависимости от того, выживет ли вариабельный домен для того, чтобы экспрессироваться в тяжелой цепи IgG, в которой отсутствует домен CH1 или в которой отсутствуют домен CH1 и шарнирная область. В отличие от этого, мыши с нарушенным IgM, вероятно, не смогут презентировать полный репертуар вариабельных областей тяжелой цепи, поскольку для переключения классов будут доступны только те вариабельные области, которые способны пережить селекцию применительно к нарушенному IgM.
Таким образом, животное, у которого отсутствует функциональный IgM, может проявлять значительное снижение способности создавать популяцию B-клеток после реаранжировки иных подходящих генных сегментов вариабельной области тяжелой цепи. В таком случае даже там, где имеется достаточный запас вариабельных областей тяжелой цепи (т.е. у животного есть подходящее количество генных сегментов вариабельной области тяжелой цепи, способных к реаранжировке), может не образоваться удовлетворительная популяция B-клеток, демонстрирующая желаемую степень разнообразия, из-за нарушения IgM, которое затрудняет выживание тяжелых цепей во время процесса селекции.
Выработка антитела, имеющего только тяжелую цепь, при функциональном гене IgM
Желательно поддерживать подходящее количество реаранжированных вариабельных областей тяжелой цепи, которые могут эффективно перенести селекцию, когда презентируются в процессе развития B-клетки применительно к IgM, для того чтобы создать достаточное разнообразие при выработке антител при иммунизации интересующим антигеном животного, которое не является человеком. Таким образом, генетически модифицированное, не являющееся человеком животное, которое содержит нефункциональный домен CH1 или нефункциональный домен CH1 и нефункциональную шарнирную область в тяжелой цепи иммуноглобулина, не должно содержать делецию CH1 в обоих аллелях IgM.
В некоторых вариантах осуществления нежелательно удалять домены CH1 во всех изотипах Ig для получения антитела, имеющего только тяжелую цепь, у генетически модифицированного животного. Таким образом, предлагаются способы и композиции для получения антитела, имеющего только тяжелую цепь, у генетически модифицированного, не являющегося человеком животного посредством повреждения, удаления или каким-либо другим образом превращения в нефункциональную нуклеотидной последовательности, кодирующей домен CH1 или его фрагмент в IgG (и в некоторых вариантах осуществления также повреждение, удаление или каким-либо другим образом превращение в нефункциональную нуклеотидной последовательности, кодирующей шарнирную область в IgG), в то время как для других изотипов (например, IgM) допускается сохранение функциональных доменов CH1. Полагают, что функциональность доменов CH1 другого изотипа (иного, чем один или несколько выбранных доменов CH1 IgG) приводит к процессу развития B-клетки, который не нарушает или по существу не нарушает этапы развития, на которых вариабельный домен тяжелой цепи презентируется применительно к не-IgG изотипу, например, в изотипе IgM. Таким образом, нарушение, например, IgM-зависимых этапов во время развития B-клетки относительно минимизировано. Не ограничиваясь изобретением (которое описывается формулой изобретения), авторы изобретения предполагают, что минимизированное нарушение ранних этапов селекции, связанных с презентацией вариабельного домена тяжелой цепи в случае IgM, приводит к большему числу клеток, несущих вариабельные области тяжелой цепи, выживших, чтобы пройти переключение классов на изотип IgG и селекцию применительно к IgG, в котором отсутствует функциональный домен CH1 или в котором отсутствуют функциональный домен CH1 и функциональная шарнирная область.
Таким образом, предлагается генетически модифицированное, не являющееся человеком животное, а также способы и композиции для создания животного, где генетическая модификация приводит к потере функционального домена CH1 (в дополнительном варианте осуществления - к потере функциональной шарнирной области) в домене Ig, который не является доменом IgM. В различных вариантах осуществления последовательность, кодирующая CH1 или CH1 и шарнирную область (или по существу их функциональную часть), удалена из генома генетически модифицированного животного. Генетически модифицированное, не являющееся человеком животное подходит для выработки антител, имеющих только тяжелую цепь (т.е. антител, у которых отсутствует легкая цепь), включая полные антитела человека (у мыши, генетически модифицированной для включения генов иммуноглобулина человека) и химерные антитела человек/мышь [например, у мыши, генетически модифицированной для включения генных сегментов вариабельной области человека, областей D и областей J, или у мыши, имеющей трансген человека, способный к транс-переключению на генетически модифицированный изотип IgG, в котором отсутствует функциональный домен CH1 или в котором отсутствуют функциональный домен CH1 и функциональная шарнирная область).
Антитела, имеющие только тяжелую цепь
Антитела пригодны в качестве терапевтического средства для человека. Антитела, имеющие только тяжелую цепь, т.е. антитела, в которых отсутствует легкая цепь, также пригодны в качестве терапевтического средства для человека. Поскольку в антителах, имеющих только тяжелую цепь, отсутствует легкая цепь, они меньше, и, таким образом, ожидается, что они будут лучше проникать в ткани, чем антитела, которые содержат легкие цепи, при этом обладая сходным или более благоприятным фармакокинетическим профилем и сохраняя сходную эффекторную функцию по сравнению с обычным антителом. Поскольку они меньше, антитела, имеющие только тяжелую цепь, также можно вводить в более высокой дозе в определенном объеме. Обычным способом введения антител является подкожная инъекция, и уменьшение вводимого объема при определенной дозировке антитела может обеспечить благоприятные эффекты для пациентов и избавить от осложнений и боли, вызванных большими объемами подкожной инъекции.
Другим преимуществом антител, имеющих только тяжелую цепь, является возможность получать биспецифические антитела за счет гетеродимеризации тяжелых цепей со специфичностью к двум различным эпитопам в одном терапевтическом средстве. Поскольку у антител, имеющих только тяжелую цепь, отсутствует легкая цепь, они особенно подходят для изготовления биспецифических антител, поскольку не требуется создавать легкую общую цепь, которая бы не мешала аффинности связывания или специфичности любой из тяжелых цепей и позволяла удобно экспрессировать биспецифическое антитело.
Генетически модифицированных животных по изобретению можно использовать для получения широкого спектра антител, имеющих только тяжелую цепь. Генетические модификации, описываемые в настоящем документе, можно производить, например, в любой подходящей линии мыши. Мышиная линия может иметь любую генетическую основу, подходящую для выработки антитела, имеющего только выбранную тяжелую цепь. Некоторые генетические основы, которые охватывают конкретные варианты осуществления, предлагаются ниже.
Генетически модифицированное животное может быть мышью, которая содержит генетическую модификацию в соответствии с изобретением и один или несколько нереаранжированных генных сегментов вариабельной области человека, один или несколько нереаранжированных генных сегментов области D и один или несколько нереаранжированных генных сегментов области J, замещающих эндогенный локус вариабельной области тяжелой цепи мыши. У такой мыши гуманизированный локус вариабельной области способен к рекомбинации с образованием реаранжированного гена вариабельной области перед эндогенными последовательностями константного домена мыши (где один или несколько генов константной области иммуноглобулина модифицированы, как описано в настоящем документе). Мышь, таким образом, будет способна вырабатывать химерное антитело, имеющее только тяжелую цепь с вариабельной областью человека/константной областью мыши. Под воздействием интересующего иммуногена мышь будет способна производить антитело, имеющее только тяжелую цепь, в соответствии с изобретением; антитело является аффинно зрелым и способно к специфическому связыванию эпитопа интересующего иммуногена.
Генетически модифицированное животное может быть мышью, содержащей эндогенную вариабельную область мыши, которая включает нереаранжированные эндогенные генные сегменты вариабельной области мыши, нереаранжированные эндогенные генные сегменты области D мыши и нереаранжированные эндогенные генные сегменты области J мыши, где мышь содержит генетическую модификацию константной области тяжелой цепи мыши, описанную в настоящем документе. Мышь, таким образом, будет способна вырабатывать мышиное антитело, имеющее только тяжелую цепь. Под воздействием интересующего иммуногена мышь будет способна производить антитело, имеющее только тяжелую цепь, в соответствии с изобретением; антитело является аффинно зрелым и способно к специфическому связыванию эпитопа интересующего иммуногена.
Генетически модифицированное животное может быть мышью, содержащей трансген человека, который включает нереаранжированные генные сегменты вариабельной области человека, нереаранжированные генные сегменты D человека и нереаранжированные генные сегменты J человека, ген мю и последовательность, которая позволяет транс-переключение. Мышь будет дополнительно содержать модификацию константной области тяжелой цепи мыши, описанную в настоящем документе. Мышь, таким образом, будет способна вырабатывать полное антитело IgM человека, и посредством транс-переключения химерное антитело с вариабельной областью человека/константной областью мыши, где константный домен содержит генетическую модификацию, описанную в настоящем документе. Под воздействием интересующего иммуногена мышь будет способна производить антитело, имеющее только тяжелую цепь, в соответствии с изобретением; антитело является аффинно зрелым и способно к специфическому связыванию эпитопа интересующего иммуногена.
Экспрессия антител, имеющих только тяжелую цепь, in vitro
Авторы изобретения установили, что нормальную вариабельную область тяжелой цепи человека или мыши (hVR или mVR) можно экспрессировать в системе in vitro применительно к IgG, в котором отсутствует функциональный домен CH1. Авторы изобретения экспрессировали hVR из нереаранжированного минилокуса hVR в мыши с мышиным IgM дикого типа. Экспрессированный hVR клонировали в IgG2b, в котором отсутствует домен CH1, и полученный hVR-IgG2b∆CH1 экспрессировался и секретировался клеткой CHO, которая была временно трансфицирована конструкцией hVR-IgG2b∆CH1, эффективно доказывая, что hVR, выбранный у мыши с IgM дикого типа, может экспрессироваться и секретироваться клеткой при переключении на IgG, в котором отсутствует функциональный домен CH1, т.е. в качестве антитела, имеющего только тяжелую цепь.
Авторы изобретения сконструировали систему in vitro в клетках CHO для экспрессии тяжелых цепей, в которых отсутствуют домены CH1 и в которых присутствуют hVR или камелизированные VR (hVR*) человека. VR получали от мыши RAG, у которой эндогенный локус тяжелой цепи мыши замещен минилокусом вариабельной области тяжелой цепи человека (содержит три генных сегмента области V человека, 6-1, 1-2 и 1-3, все генные сегменты DH человека и все генные сегменты JH человека). Эндогенные локусы легких цепей каппа и лямбда иммуноглобулина мыши были интактными и функциональными.
Были созданы конструкции химерной тяжелой цепи (hVR-mFc) и камелизированной тяжелой цепи (hVR*-mFc) для экспрессии в клетках CHO, с использованием последовательностей VR, полученных от мыши, несущей вышеописанный минилокус. Химерные тяжелые цепи были продуктом обычной рекомбинации V-D-J во время развития B-клетки у мыши для создания функционального антитела, содержащего химерную тяжелую цепь (hVR-mFc) и легкую цепь мыши. Конструкции hVR-mFc и hVR*-mFc делали как с наличием домена CH1, так и с отсутствием домена CH1.
Временная трансфекция конструкциями hVR-mFc и hcVR-mFc клеток CHO показала, что в отсутствие домена CH1, тяжелые цепи с hVR и hVR* экспрессировались и оставались растворенными в супернатанте. В присутствии домена CH1 тяжелые цепи, содержащие или hVR или hVR*, не экспрессировались в супернатант. Это наблюдение указывает на то, что такие антитела, имеющие только тяжелую цепь, могут быть созданы без использования верблюжьих доменов VHH, например, с доменами VH человека или мыши в антителах, имеющих только тяжелую цепь, в которой отсутствует домен CH1.
Гуманизированные антитела, имеющие только тяжелую цепь
Для выработки гуманизированной версии антитела, имеющего только тяжелую цепь, по настоящему изобретению, животное, гомозиготное по модификации, иммунизируют антигеном, и после получения специфического иммунного ответа животного клетки из селезенки иммунизированного животного сливают с подходящей иммортализованной клеткой (например, миеломной клеткой) для получения гибридомных клеток. Альтернативно, антитела можно получать непосредственно из B-клеток иммунизированного животного. Супернатанты из гибридомных клеток (или, например, из выделенных B-клеток) проверяют на наличие антитела посредством твердофазного иммуноферментного анализа (ELISA) и можно выбрать антитела, специфичные к антигену, на основании желаемых характеристик.
Нуклеиновые кислоты вариабельной области тяжелой цепи (VH) можно выделять из гибридомы и/или B-клеток с использованием стандартных молекулярно-биологических способов, известных в данной области (Sambrook et al. 1989. Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y.; Ausubel et al. 1995. Short Protocols in Molecular Biology, 3rd ed., Wiley & Sons). После того как определена последовательность нуклеиновой кислоты VH, можно получить предсказанную аминокислотную последовательность и сравнить с другими последовательностями VH человека, для того чтобы выявить группу родственных последовательностей VH, которые имеют сходную последовательность. Родственные последовательности VH можно получать с использованием баз данных по антителам, которые доступны специалистам в данной области, например, The International ImMunoGeneTics Information System® (IMGT®). Это сравнение можно проводить путем выравнивания последовательностей, которое осуществляется либо на глаз или, альтернативно, в электронном виде с использованием программ выравнивания (например, CLUSTAL). В этом сравнении идентифицируются определяющие комплементарность области (CDR) и каркасные области (FR). Остатки CDR и FR определяют согласно определению стандартной последовательности (например, Kabat et al. 1987, Sequences of Proteins of Immunological Interest, National Institutes of Health, Bethesda Md.; Chothia и Lesk, 1987. J. Mol Biol. 196:901-917). Специалистам в данной области понятно, что иногда могут существовать расхождения в способах нумерации и определения областей CDR и FR в последовательности тяжелой цепи иммуноглобулина. В таких случаях предпочтительным является структурное определение, однако, остатки, идентифицированные способом определения последовательности, считаются важными остатками FR для определения того, какие каркасные остатки заменить на основании сравнения последовательностей тяжелой цепи.
После выравнивания идентифицируют взаимозаменяемые положения в последовательностях VH. Если идентичность аминокислоты в положении в изолированной последовательности VH варьирует при сравнении с другими последовательностями VH человека, такое положение оценивают на пригодность для замещения в этом положении в изолированной последовательности VH. Таким образом, любое положение в выделенной последовательности VH, которое отличается от другой родственной последовательности(ей) VH человека, с которыми сравнивают, может потенциально являться положением, которое можно заместить аминокислотой в соответствующем положении, находящейся в одной или нескольких родственных последовательностях VH человека. Положения, которые идентичны другим родственным последовательностям VH человека, т.е. те, которые не демонстрируют вариабельности, определяют как невзаимозаменяемые положения. В различных вариантах осуществления вышеописанные способы используют для создания консенсусной последовательности человеческого антитела, имеющего только тяжелую цепь.
Гуманизированное антитело, имеющее только тяжелую цепь, для целей, описываемых в настоящем документе, представляет собой вариант аминокислотной последовательности тяжелой цепи иммуноглобулина или ее фрагмент, который способен связывать заданный антиген и который содержит область FR, имеющую аминокислотную последовательность, по существу сходную или идентичную по сравнению с аминокислотной последовательностью FR человека, и CDR, имеющую аминокислотную последовательность, по существу сходную или идентичную с не принадлежащей человеку аминокислотной последовательностью CDR. Как правило, гуманизированное антитело, имеющее только тяжелую цепь, имеет один или несколько аминокислотных остатков, которые получены из источника, не принадлежащего человеку. Такие остатки, как правило, получены из вариабельного домена тяжелой цепи. Дополнительно, эти остатки могут иметь сопутствующие характеристики, такие как, например, аффинность и/или специфичность, а также другую желаемую биологическую активность, связанную с функцией антитела.
В различных вариантах осуществления гуманизированное антитело, имеющее только тяжелую цепь, содержит по существу все из, по меньшей мере, одного, и в других вариантах осуществления, по меньшей мере, двух доменов VH, в которых все или по существу все области CDR соответствуют таковым в домене VH, не принадлежащем человеку, и все или по существу все области FR - из последовательности домена VH человека. Гуманизированное антитело, имеющее только тяжелую цепь, будет содержать уникальную константную область иммуноглобулина (Fc), в которой в одном из вариантов осуществления отсутствует, по меньшей мере, домен CH1, и в одном из вариантов осуществления также отсутствует шарнирная область Fc человека. В одном из вариантов осуществления антитело, имеющее только тяжелую цепь, не содержит легкую цепь и содержит области CH2 и CH3 константной области тяжелой цепи иммуноглобулина G (IgG). В одном из вариантов осуществления константная область антитела, имеющего только тяжелую цепь, включает шарнир и области CH2 и CH3 Fc тяжелой цепи IgG. В одном из вариантов осуществления константная область антитела, имеющего только тяжелую цепь, будет включать область CH1 IgM.
Гуманизированное антитело, имеющее только тяжелую цепь, выбирают из любого класса IgG, включая IgG1, IgG2, IgG3 и IgG4. В различных вариантах осуществления константная область может содержать последовательности более чем от одного класса IgG, и выбор конкретных константных области для оптимизации желаемых эффекторных функций находится в пределах компетенции специалиста в данной области.
Как правило, области тяжелой цепи FR и CDR гуманизированного антитела, имеющего только тяжелую цепь, не должны точно соответствовать родительским последовательностям; например, не принадлежащая человеку CDR тяжелой цепи или FR тяжелой цепи человека могут быть изменены путем замены, вставки или делеции, по меньшей мере, одного остатка таким образом, что остаток CDR тяжелой цепи или остаток FR тяжелой цепи в указанном сайте не соответствуют или последовательности FR тяжелой цепи человека или не принадлежащей человеку последовательности CDR тяжелой цепи. Такие мутации, однако, не должны быть протяженными. В одном из вариантов осуществления, по меньшей мере, 75% остатков гуманизированного антитела, имеющего только тяжелую цепь, будут соответствовать таковым в родительских последовательностях FR и CDR тяжелой цепи, в другом варианте осуществления 90%, и в другом варианте осуществления более чем 95%.
Гуманизированные антитела, имеющие только тяжелую цепь, описанную в настоящем документе, в одном из вариантов осуществления получены посредством процесса анализа родительских последовательностей и различных концептуальных гуманизированных составных последовательностей in silico, с использованием компьютерных программ, доступных и известных специалистам в данной области. Модификации последовательности для создания гуманизированных версий и/или для изменения характеристик, таких как иммуногенность, аффинность и т.д., производят с использованием известных в данной области способов (например, US 5565332 Hoogenboom et al.; US 5639641 Pedersen et al.; US 5766886 Studnicka et al.; US 5859205 Adair et al.; US 6054297 Carter et al.; US 6407213 Carter et al.; US 6639055 Carter et al.; US 6849425 Huse et al.; US 6881557 Foote; US 7098006 Gorman et al.; US 7175996 Watkins et al.; US 7235643 Nicolaides et al.; US 7393648 Rother et al.; US 7462697 Couto et al.).
В различных вариантах осуществления желаемые замены в последовательности родительского антитела, имеющего только тяжелую цепь, для получения варианта родительского антитела, имеющего только тяжелую цепь, представляют собой те, которые в одном из вариантов осуществления сохраняют, или в другом варианте осуществления повышают, активность связывания антигена родительским антителом, имеющим только тяжелую цепь. Как правило, вариант родительского антитела, имеющего только тяжелую цепь, также имеющий только тяжелую цепь, обладает аффинностью связывания антигена, которая составляет по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 100% (например, по меньшей мере 150%, по меньшей мере 200%, по меньшей мере 500%, по меньшей мере 1000% или по меньшей мере вплоть до 10000%) от аффинности связывания конкретного антигена родительским антителом, имеющим только тяжелую цепь. В некоторых вариантах осуществления вариантное антитело, имеющее только тяжелую цепь, содержит одну замену по сравнению с родительским антителом, имеющим только тяжелую цепь. Однако, в других вариантах осуществления несколько аминокислот, например, вплоть до приблизительно 5 или 10 или более, замещены по сравнению с последовательностью родительского антитела, имеющего только тяжелую цепь; аминокислоты получены из других последовательностей тяжелой цепи человека, которые идентичны по данному положению. Замены в одном из вариантов осуществления являются консервативными (т.е. аминокислота имеет сходные свойства с замещаемым остатком), и в другом варианте осуществления являются неконсервативными (т.е. аминокислота имеет свойства, отличающиеся от свойств замещаемого остатка). В различных вариантах осуществления полученное вариантное антитело, имеющее только тяжелую цепь, тестируют для подтверждения того, что из-за замены остатков не произошло значительного снижения желаемой аффинности связывания и/или специфичности. В некоторых вариантах осуществления улучшенное вариантное антитело, имеющее только тяжелую цепь, получают путем замены аминокислот из отличающейся последовательности тяжелой цепи человека.
Показано, что природные антитела, имеющие только тяжелую цепь, (например, обнаруженные у верблюдовых) содержат уникальные аминокислотные остатки в положениях, соответствующих границе между вариабельными областями тяжелой и легкой цепей в обычных молекулах антитела (т.е. две тяжелые цепи и две легкие цепи). Известно, что эти остатки на границе влияют на близость или ориентацию двух родственных друг другу цепей в обычных антителах. Хотя известно, что эти природные антитела, имеющие только тяжелую цепь, содержат замены остатков, которые коррелируют с отсутствием вариабельных областей легкой цепи, они сохраняют остатки в других положениях в последовательности по сравнению с обычными антителами для сохранения характерного изгиба иммуноглобулина. Замены, обнаруженные в природных антителах, имеющих только тяжелую цепь, представляют собой L11S, V37F, G44E, L45R или L45C, W47G и дополнительные остатки цистеина, которые участвуют в образовании дисульфидной связи между CDR1 и CDR3 вариабельной области тяжелой цепи. В некоторых вариантах осуществления антитела по настоящему изобретению, имеющие только тяжелую цепь, могут сохранять остаток родительского антитела в этих положениях. В других вариантах осуществления родительское антитело может проявлять мутации в этих положениях, которые ассоциированы с остатками в природных антителах, имеющих только тяжелую цепь. В некоторых вариантах осуществления может быть желательным сохранять такой же остаток, который находится в родительском антителе, имеющем только тяжелую цепь по меньшей мере в одном из этих положений или, в одном из вариантов осуществления, во всех этих положениях, при создании гуманизированного антитела, имеющего только тяжелую цепь, полученную из изолированной последовательности VH от генетически модифицированной мыши, описанной в настоящем документе. В различных вариантах осуществления специалисту в данной области ясно, что нелогично ожидать, что эти остатки на границе будут вовлечены в межцепочечные взаимодействия в антителах, имеющих только тяжелую цепь, которые производятся генетически модифицированной мышью, описанной в настоящем документе.
Создание генетически модифицированных животных
Генетические модификации для создания животного, которое экспрессирует антитело, имеющее только тяжелую цепь, удобно описывать с использованием мыши в качестве иллюстрации. Генетически модифицированную мышь по изобретению можно создать рядом способов, конкретные варианты осуществления которых описаны ниже.
Схематическая иллюстрация (не в масштабе) локуса IgG1 приведена на фигуре 1 (вверху), для того чтобы показать расположение домена CH в локусе IgG1. Как показано на фигуре, домены CH1, CH2 и CH3 и шарнирная область находятся в легко узнаваемых интервалах нуклеотидов ниже области переключения.
Генетически модифицированная мышь, у которой отсутствует нуклеотидная последовательность, кодирующая домен CH1 IgG1, но которая содержит шарнирную область, может быть получена любым известным в данной области способом. Например, можно создать нацеленный вектор, который замещает ген IgG1 геном усеченного IgG1, в котором отсутствует домен CH1, но присутствует шарнир. Фигура 2 изображает геном мыши (вверху), который является мишенью для нацеленной конструкции с 5' (по отношению к направлению транскрипции геномного гена IgG1) гомологичным плечом, содержащим последовательность выше эндогенного домена CH1, с последующими нуклеотидными последовательностями, которые кодируют шарнир IgG1, домен CH2 IgG1, домен CH3 IgG1, кассету для селекции по устойчивости к лекарственному средству (например, ген устойчивости, фланкированный последовательностями lox), и трансмембранный домен IgG1, и 3' гомологичным плечом, включающим последовательность 3' по отношению к трансмембранному домену. При гомологичной рекомбинации в локусе и удалении кассеты для селекции по устойчивости к лекарственному средству (например, посредством обработки Cre), эндогенный IgG1 замещается IgG1, в котором отсутствует домен CH1 (внизу на фигуре 2; сайт lox не показан). Фигура 1 (IgG1∆CH1, в центре) показывает структуру полученного локуса, который будет экспрессировать IgG1, в котором последовательность области J слита с последовательностью шарнира.
Генетически модифицированную мышь, у которой отсутствует нуклеотидная последовательность, кодирующая домен CH1 IgG1 и у которой отсутствует нуклеотидная последовательность, кодирующая шарнирную область, можно создавать любым известным в данной области способом. Например, можно создать направленный вектор, который замещает ген IgG1 усеченным IgG1, в котором отсутствует последовательность, кодирующая домен CH1, и в котором отсутствует нуклеотидная последовательность, кодирующая шарнирную область. Фигура 3 изображает геном мыши (вверху), который является мишенью для нацеленной конструкции с 5' (по отношению к направлению транскрипции геномного гена IgG1) гомологичным плечом, содержащим последовательность выше эндогенного домена CH1, с последующими нуклеотидными последовательностями, которые кодируют домен CH2 IgG1, домен CH3 IgG1, кассету для селекции по устойчивости к лекарственному средству (например, ген устойчивости, фланкированный последовательностями lox), и трансмембранный домен IgG1, и 3' гомологичным плечом, включающим последовательность 3' по отношению к трансмембранному домену. При гомологичной рекомбинации в локусе и удалении кассеты для селекции по устойчивости к лекарственному средству (например, посредством обработки Cre), эндогенный IgG1 замещается IgG1, в котором отсутствует домен CH1 (внизу на фигуре 3; сайт lox не показан). Фигура 1 (IgG1∆CH1-∆шарнир, внизу) показывает структуру полученного локуса, который будет экспрессировать IgG1, в котором последовательность области J слита с доменом CH2.
Генетически модифицированная мышь, у которой отсутствует последовательность CH1 IgG1 (IgG1∆CH1) или у которой отсутствует последовательность CH1 IgG1 и у которой отсутствует шарнир (IgG1∆CH1-∆шарнир), может быть дополнительно модифицирована для удобного использования модифицированного изотипа IgG1 посредством удаления одного или нескольких других изотипов IgG, например, посредством делеции или функционального выключения последовательностей, кодирующих IgG2b и IgG2a. Например, делают нацеленную конструкцию с 5' гомологичным плечом, содержащим последовательность выше эндогенной последовательности шарнирной области (или выше эндогенной последовательности домена CH1), последовательностями, которые кодируют домены CH2 и CH3 IgG1, кассетой для селекции по устойчивости к лекарственному средству, с последующей последовательностью, кодирующей трансмембранный домен IgG1, с последующей кассетой для селекции по устойчивости к другому лекарственному средству, при желании. При гомологичной рекомбинации в локусе и удалении кассеты для селекции по устойчивости к лекарственному средству(ам) (например, посредством обработки Cre), эндогенный константный локус тяжелой цепи содержит только два гена IgG: эндогенный IgG3 и IgG1∆CH1 (см. фигуру 4, внизу; сайт(ы) рекомбиназы не показаны; см. фигуру 6, внизу) или IgG1∆CH1-∆шарнир (см. фигуру 5, внизу; сайт(ы) рекомбиназы не показаны; см. фигуру 6, внизу).
IgG1, который экспрессируется у генетически модифицированной мыши с аллелем IgG1∆CH1-∆шарнир или IgG1∆CH1∆IgG2a∆IgG2b, будет иметь структуру, показанную на правой панели фигуры 10, т.е. домен VH будет слит с доменом CH2. На левой панели на фигуре 10 представлено, для сравнения, антитело IgG1 дикого типа, в котором домен CH1 связан посредством шарнирной области с доменом CH2 и связан посредством дисульфидной связи с константным доменом CL легкой цепи. Напротив, у антитела, которое производит генетически модифицированная мышь, отсутствует шарнир и домены CH1 и, таким образом, отсутствует любой домен CL.
Генетически модифицированных мышей, описанных выше, и других, получают путем введения подходящей нацеленной конструкции в подходящую ES клетку мыши (посредством одного или нескольких независимых нацеливаний), и выявляют и выращивают позитивные клоны, содержащие маркер или кассету для селекции нацеленной конструкции. Клоны затем используют в качестве донорских ES клеток для эмбриона-хозяина в условиях, подходящих для создания химерной мыши или мыши, полностью полученной из ES клетки. Маркер или кассету для селекции можно по желанию удалить, или на стадии ES клетки или из химерной или полученной из ES клетки мыши, например, с использованием кассеты с последовательностями lox и скрещиванием с линией, несущей Cre, или путем электропорации ES клетки с Cre-экспрессирующим вектором.
Генетически модифицированную мышь с аллелем IgG1∆CH1-∆шарнир (гетерозиготным) получали в соответствии с вариантом осуществления изобретения. У мыши выделяли сыворотку и проводили вестерн-блоттинг (восстановительные условия) с использованием антитела к IgG1 мыши для выявления тяжелой цепи. В отличие от мыши дикого типа, которая демонстрирует полосу, соответствующую размеру тяжелой цепи IgG1 дикого типа, мышь, генетически модифицированная для создания аллеля IgG1∆CH1-∆шарнир, также экспрессирует тяжелую цепь, которая реагирует с антителом против мышиного IgG1, которая имеет ожидаемые размеры антитела, имеющего только тяжелую цепь, которая включает домены VH, CH2 и CH3 (см. фигуру 8).
ПРИМЕРЫ
Пример 1: Экспрессия in vitro антител, имеющих только тяжелую цепь
Конструкции химерных тяжелых цепей получали с использованием молекулярно-биологических способов (например, см. Maniatis et al. 1982. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory) для слияния вариабельных областей человека с константной областью IgG2b мыши (mIgG2b). Генный сегмент вариабельной области человека для каждой конструкции представлял собой полноразмерный генный сегмент вариабельной области, содержащий оба экзона (т.е. лидерная последовательность плюс зрелая последовательность), идентифицированные из hVR IgM, выделенного из наивной мыши RAG, которая содержит замену эндогенного мышиного локуса тяжелой цепи иммуноглобулина тремя генными сегментами hVR, всеми генными сегментами hDH и всеми генными сегментами hJH. Легкая цепь антитела IgM представляла собой легкую цепь мыши.
Использовали две версии последовательности mIgG2b, одну с доменом CH1 и одну без домена CH1. Были также созданы несколько других конструкций, которые служили в качестве контролей трансфекции и экспрессии. Первая контрольная конструкция была сделана с использованием рецептора цитокина, слитого с доменами CH2 и CH3 константной области IgG2a мыши (mIgG2a) (Контроль I). Два других контроля конструировали путем слияния сигнальной последовательности ROR мышей с последовательностью IgG2a мышей с наличием и отсутствием доменов CH1 (Контроль II и III, соответственно).
Были также сделаны камелизированные версии каждой вариабельной области человека с использованием способов сайт-специфического мутагенеза на основе ПЦР (например, см. Hutchinson et al. 1978. Mutagenesis at a specific position in DNA sequence. J. Biol. Chem. 253(18):6551-60). Для каждой вариабельной области использовали два специфических набора праймеров для создания конкретных мутаций внутри последовательности вариабельной области человека, приводящих к последовательности вариабельной области человека, содержащей особенности, присущие верблюдам. Праймеры L1 (SEQ ID NO:1) и HH1.2 mut BOT (SEQ ID NO:2) использовали для амплификации одного продукта, содержащего 5' половину вариабельной области, в то время как праймеры HH1.2 mut TOP (SEQ ID NO:3) и m18.3.1 (SEQ ID NO:4) для амплификации 3' половины вариабельной области. Эти продукты очищали и смешивали вместе, чтобы они служили матрицей для третьей реакции ПЦР с использованием праймеров L1 и m18.3.1. Полученный ПЦР-продукт камелизированной вариабельной области человека клонировали, очищали и подтверждали секвенированием.
Конструкции полноразмерной тяжелой цепи (вариабельная и константная области) получали путем амплификации вариабельных областей человека (камелизированных и некамелизированных) и константных областей при помощи праймеров, содержащих участки распознавания рестрикционных ферментов, для того чтобы обеспечить последующее лигирование друг с другом посредством липких концов. Все конструкции полноразмерных тяжелых цепей клонировали в экспрессирующие векторы, очищали и еще раз подтверждали секвенированием. Таблица 1 излагает каждую конструкцию тяжелой цепи, их SEQ ID NO и краткое описание для каждой конструкции.
Конструкции химерных тяжелых цепей были временно трансфицированы в клетки китайского хомячка (CHO-K1) для исследования экспрессии в отсутствие легкой цепи иммуноглобулина. Супернатанты и клеточные лизаты исследовали при помощи вестерн-блоттинга для выявления присутствия тяжелой цепи посредством хемилюминесценции с использованием пероксидазы хрена (HRP), конъюгированной с антителом против IgG мыши (Promega). Все конструкции химерных тяжелых цепей участвовали во временной трансфекции независимо по шесть (6) раз. Типичный вестерн-блоттинг трансфекций представлен на фигуре 7.
Все конструкции химерных тяжелых цепей, с наличием и отсутствием домена CH1, а также контрольные конструкции, были обнаружены в клеточном лизате. В супернатантах наблюдали только конструкции, в которых отсутствует домен CH1 (фигура 7, слева). Также были выявлены Контроль I и Контроль III (белок Fc мыши, в котором отсутствует домен CH1) (фигура 7), но белок Fc мыши, содержащий домен CH1, выявлен не был. Как некамелизированные, так и камелизированные конструкции тяжелой цепи, содержащие домен CH1, не были выявлены в супернатанте ни в одной из трансфекций (фигура 7, справа). Однако, как некамелизированные, так и камелизированные конструкции тяжелой цепи, в которых отсутствует домен CH1, были обнаружены в супернатанте во всех трансфекциях. В совокупности, результаты доказывают, что hVR (нормальные или камелизированные), в которых отсутствует домен CH1, могут экспрессироваться и секретироваться временно трансфицированными клетками CHO, в отсутствие легкой цепи иммуноглобулина, в то время как hVR (нормальные или камелизированные), которые содержат домен CH1, не могут секретироваться в отсутствие легкой цепи.
Пример 2: Модификация константной области тяжелой цепи IgG1 мыши
A. Получение вектора, нацеленного на IgG1-CH1-шарнир мыши (фигура 3)
Была создана нацеленная конструкция для введения делеции в CH1 и шарнирные области константного домена IgG1 мыши для аллеля C57BL/6 из ES клетки мыши VELOCIMMUNE® (описано ниже).
Нацеленная конструкция была создана с использованием технологии VELOCIGENE® (см., например, патент США № 6586251 и Valenzuela et al. (2003) High throughput engineering of mouse genome coupled with high-resolution expression analysis, Nature Biotech. 21 (6):652-659) для модификации Бактериальной Искусственной Хромосомы (BAC) BMQ 70p08. ДНК BAC BMQ 70p08 модифицировали для удаления CH1 и шарнирных областей константного домена IgG1, одновременно оставляя интактным остаток гена IgG1 (например, CH2, CH3 и трансмембранные экзоны).
В кратком изложении, верхнее и нижнее гомологичные плечи были получены с использованием праймеров m102 (SEQ ID NO:13) и m104 (SEQ ID NO:14), и m100 (SEQ ID NO:15) и m99 (SEQ ID NO:16), соответственно. Эти гомологичные плечи применяли для создания кассеты, которая удаляет CH1 и шарнирные области константного домена IgG1, одновременно сохраняя CH2, CH3 и трансмембранные области константного домена IgG1 (см., например, фигуру 3). Нацеленная конструкция содержала фланкированный последовательностями lox ген устойчивости к гигромицину, расположенный между CH3 и экзонами трансмембранного домена гена IgG1. Гены, расположенные выше экзонов CH1 и шарнира (например, IgG3, IgD, IgM) и ниже трансмембранного экзона IgG1 (например, IgG2b, IgG21, IgE, IgA и т.д.), не были модифицированы нацеленной конструкцией. Области переключения для всех константных доменов не были модифицированы нацеленной конструкцией. Нуклеотидная последовательность между концами делеции включала следующую последовательность, которая указывает на акцепторный сайт сплайсинга, находящийся в точке делеции: TGACAGTGTA ATCACATATA CTTTTTCTTG T(AG)TCCCAGA AGTATCATC (SEQ ID NO:17). Последовательность делеции содержит акцепторный сайт сплайсинга (AG, в скобках выше) с пре-CH1 последовательностями с 5' от акцепторного сайта сплайсинга и последовательности экзона CH2 с 3' от акцепторного сайта сплайсинга.
B. Получение вектора, нацеленного на IgG1-CH1 мыши (фигура 2)
Вторая нацеленная конструкция для введения делеции в CH1 константного домена IgG1 мыши для аллеля 129/SvEvTac из ES клетки мыши (описано ниже) была создана сходным образом, как описано в разделе A данного Примера.
Нацеленная конструкция была создана с использованием технологии VELOCIGENE® (см., например, патент США № 6586251 и Valenzuela et al. (2003) High throughput engineering of mouse genom coupled with high-resolution expression analysis, Nature Biotech. 21 (6):652-659) для модификации Бактериальной Искусственной Хромосомы (BAC) BMQ 70p08. ДНК BAC BMQ 70p08 модифицировали для удаления области CH1 константного домена IgG1, одновременно оставляя интактным остаток гена IgG1 (например, шарнир, CH2, CH3 и трансмембранные экзоны; см. фигуру 2).
Гомологичные плечи для второй нацеленной конструкции были получены таким же образом, что и для нацеленного вектора CH1-шарнир (как описано выше в разделе A данного Примера). Эти гомологичные плечи применяли для создания кассеты, которая удаляет область CH1 константного домена IgG1, одновременно сохраняя шарнир, CH2, CH3 и трансмембранные области константного домена IgG1 (см., например, фигуру 2). Нацеленная конструкция содержала фланкированный последовательностями lox ген устойчивости к гигромицину, расположенный между экзонами CH3 и трансмембранного домена гена IgG1. Гены, расположенные выше экзона CH1 (например, IgG3, IgD, IgM) и ниже трансмембранного экзона IgG1 (например, IgG2b, IgG21, IgE, IgA и т.д.), не были модифицированы нацеленной конструкцией. Области переключения для всех константных доменов не были модифицированы нацеленной конструкцией. Нуклеотидная последовательность между концами делеции включала следующую последовательность, которая указывает на акцепторный сайт сплайсинга, находящийся в точке делеции: TGACAGTGTA ATCACATATA CTTTTTCTTG T(AG)TGCCCAG GGATTGTGGT TGTAAGCCTT GCATATGTAC AGGTAAGTCA GTAGGCCTTT CACCCTGACC C (SEQ ID NO:64). Последовательность делеции содержит акцепторный сайт сплайсинга (AG, в скобках выше) с пре-CH1 последовательностями с 5' от акцепторного сайта сплайсинга и последовательностями экзонов шарнира с 3' от акцепторного сайта сплайсинга.
Пример 3: Модификация константной области тяжелой цепи мыши в ES клетках
A. Направленное воздействие на ES клетки мыши с использованием нацеленного вектора IgG1-CH1-шарнир
ES клетка мыши являлась мишенью для нацеленной конструкции, описанной выше (т.е. нацеленной конструкции, вносящей делецию областей CH1 и шарнира гена IgG1). ES клетка была получена от VELOCIMMUNE® мыши, которая являлась 50/50 помесью линии 129 и линии C57BL/6 и несла генетические модификации, которые включали замену генных сегментов вариабельных областей тяжелой цепи и легкой цепи мыши неаранжированными генными сегментами вариабельных областей тяжелой цепи и легкой цепи человека. Линия 129, использованная для скрещивания с C57BL/6, представляет собой линию, которая содержит замену генных сегментов вариабельных областей тяжелой и легкой цепи мыши генными сегментами вариабельных областей тяжелой и легкой цепи человека.
Гетерозиготные мыши VELOCIMMUNE® несут единичный набор эндогенных генов константной области тяжелой цепи мыши от линии 129 по одному аллелю и единичный набор эндогенных генов константной области тяжелой цепи мыши от линии C57BL/6 по другому аллелю. Аллель тяжелой цепи 129 граничит с локусом генных сегментов вариабельной области тяжелой цепи, которые представляют собой генные сегменты вариабельной области тяжелой цепи человека и которые заместили эндогенные генные сегменты вариабельной области тяжелой цепи мыши (т.е. в эндогенном локусе мыши). Аллель тяжелой цепи BL/6 граничит с генными сегментами вариабельной области тяжелой цепи мыши дикого типа. Мыши VELOCIMMUNE® также несут эндогенные гены константной области легкой цепи мыши дикого типа. Таким образом, за счет нацеливания на аллель 129 конструкции, содержащей делецию CH1 IgG, D, E или A, можно получать химерное антитело человек/мышь, имеющее только тяжелую цепь, в то время как за счет нацеливания на аллель C57BL/6 аналогичной конструкции можно получать полностью мышиное антитело, имеющее только тяжелую цепь, в которой отсутствует домен CH1 и в которой отсутствует шарнир.
Вышеописанные ES клетки из мышей VELOCIMMUNE® подвергали электропорации с линейным нацеленным вектором из раздела A в Примере 2 и отбирали по наличию гена устойчивости к гигромицину.
B. Направленное воздействие на ES клетки мыши с использованием нацеленного вектора IgG1-CH1
Аналогично, ES клетка мыши являлась мишенью для нацеленной конструкции CH1, описанной в разделе B в Примере 2 (см. также фигуру 2). ES клетка была получена от мыши VELOCIMMUNE®, которая являлась 50/50 помесью линии 129/SvEvTac и линии C57BL/6 и несла генетические модификации, которые включали замену генных сегментов вариабельных областей тяжелой цепи и легкой цепи мыши неаранжированными генными сегментами вариабельных областей тяжелой цепи и легкой цепи человека. Линия 129/SvEvTac, использованная для скрещивания с C57BL/6, представляет собой линию, которая содержит замену генных сегментов вариабельных областей тяжелой и легкой цепи мыши генными сегментами вариабельных областей тяжелой и легкой цепи человека.
Гетерозиготные VELOCIMMUNE® мыши несут единичный набор эндогенных генов константной области тяжелой цепи мыши от линии 129/SvEvTac по одному аллелю и единичный набор эндогенных генов константной области тяжелой цепи мыши от линии C57BL/6 по другому аллелю. Аллель тяжелой цепи 129/SvEvTac граничит с локусом генных сегментов вариабельной области тяжелой цепи, которые представляют собой генные сегменты вариабельной области тяжелой цепи человека и которые заместили эндогенные генные сегменты вариабельной области тяжелой цепи мыши (т.е. в эндогенном локусе мыши). Аллель тяжелой цепи BL/6 граничит с генными сегментами вариабельной области тяжелой цепи мыши дикого типа. Мыши VELOCIMMUNE® также несут эндогенные гены константной области легкой цепи мыши дикого типа. Таким образом, за счет нацеливания на аллель 129/SvEvTac конструкции, содержащей делецию CH1 IgG, D, E или A, можно получать химерное антитело человек/мышь, имеющее только тяжелую цепь, в то время как за счет нацеливания на аллель C57BL/6 аналогичной конструкции можно получать полностью мышиное антитело, имеющее только тяжелую цепь, в которой отсутствует домен CH1 и в которой отсутствует шарнир.
Вышеописанные ES клетки из мышей VELOCIMMUNE® подвергали электропорации с линейным нацеленным вектором, описанным в разделе В в Примере 2, и отбирали по наличию гена устойчивости к гигромицину.
Пример 4: Создание мышей, несущих модифицированную константную область IgG1
A. Мыши, несущие делецию IgG1-CH1-шарнир
Вышеописанные ES клетки-мишени применяли в качестве донора ES клеток и вносили в эмбрион мыши на стадии 8 клеток посредством способа VELOCIMOUSE® (см., например, патент США № 7294754 и Poueymirou et al. (2007) F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses. Nature Biotech. 25(1):91-99). VELOCIMICE® (F0 мыши, полностью происходящие из донорской ES клетки), несущие C57BL/6 IgG1 аллели после нацеленного воздействия, идентифицировали при помощи генотипирования с использованием модификации аллельного анализа (Valenzuela et al., выше), которая выявляет наличие последовательностей, расположенных выше и ниже делетированных областей шарнира и CH1.
Мышей, прогенотипированных на наличие делеции CH1 и шарнира IgG1 (в аллеле C57BL/6, т.е. мышином аллеле), скрестили с линией мышей Cre deleter (см., например, публикацию международной заявки на патент № WO2009/114400), для того чтобы удалить кассету устойчивости к гигромицину с сайтами lox, расположенную ниже экзона CH3 IgG1 и выше трансмембранного экзона IgG1, которая была вставлена посредством нацеленной конструкции (см., например, фигуру 3). Детенышей генотипировали и выбирали детеныша, гетерозиготного по делеции CH1 и шарнира IgG1, для анализа тяжелой цепи IgG1, экспрессирующейся с аллеля C57BL/6, в сыворотке детеныша.
B. Мыши, несущие делецию IgG1-CH1
Аналогично, ES клетки-мишени, несущие делецию области CH1 IgG1, применяли в качестве донора ES клеток и вносили в эмбрион мыши на стадии 8 клеток посредством способа VELOCIMOUSE® (см., например, патент США № 7294754 и Poueymirou et al. (2007) F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses. Nature Biotech. 25(1):91-99). VELOCIMICE® (F0 мыши, полностью происходящие из донорской ES клетки), несущие 129SvEv/Tac аллели после нацеленного воздействия, идентифицировали при помощи генотипирования с использованием модификации аллельного анализа (Valenzuela et al., выше), которая выявляет наличие последовательностей, расположенных выше и ниже делетированных области CH1.
Мышей, прогенотипированных на наличие делеции CH1 IgG1 (в аллеле 129/SvEvTac, т.е. человеческом аллеле), скрестили с линией мышей Cre deleter (см., например, публикацию международной заявки на патент № WO2009/114400), для того чтобы удалить кассету устойчивости к гигромицину с сайтами lox, расположенную ниже экзона CH3 IgG1 и выше трансмембранного экзона IgG1, которая была вставлена посредством нацеленной конструкции (см., например, фигуру 2). Детенышей генотипировали и выбирали детеныша, гомозиготного по делеции CH1 IgG1, для анализа экспрессии модифицированной тяжелой цепи IgG1.
Пример 5: Антитела, имеющие только тяжелую цепь от мышей, несущих модифицированный ген IgG1
A. Мыши IgG1-∆CH1-∆шарнир
Брали кровь у детеныша мыши, идентифицированного выше, как носитель делеции CH1 и шарнира, и детеныша дикого типа, и получали сыворотки из крови мышей для вестерн-блоттинга для определения любого экспрессировавшегося IgG в сыворотке с использованием антитела к mIgG1. В кратком изложении, 10 мкл мышиной сыворотки, разведенной 1:100, использовали для восстанавливающего SDS-PAGE, и гель переносили на поливинилиденфторидную (PVDF) мембрану. Блот блокировали в течение ночи 5% обезжиренным молоком в TBS с 0,05% Tween-20 (TBST; Sigma), отмывали 4 раза TBST по 5 минут на отмывку, а затем обрабатывали первичным антителом (козье анти-mIgG1, конъюгированное с HRP, Southern Biotech), разведенным 1:1000 в 1% обезжиренном молоке в TBST, в течение двух часов при комнатной температуре. Блот отмывали 6 раз по 5 минут на отмывку. Блот проявляли в течение 5 минут при помощи SUPERSIGNAL™ West Pico Chemiluminescent Substrate (Thermo Scientific), а затем экспонировали на пленку в течение 1 минуты.
Сыворотка от VELOCIMOUSE® (50% дикого типа BL/6; 50% ∆CH1-∆шарнир BL/6), полученная из донорских ES клеток-мишеней, показала смесь полос: одна полоса приблизительно 57,5 кДа, ожидаемый размер для IgG дикого типа, и одна полоса приблизительно 45 кДа, ожидаемый размер для IgG, в котором отсутствует домен CH1 и шарнир (фигура 8). Результаты соответствуют VELOCIMOUSE®, экспрессирующей нормальную тяжелую цепь мыши с аллеля дикого типа BL/6 и ∆CH1-∆шарнир тяжелую цепь мыши с аллеля ∆CH1-∆шарнир BL/6. Этот результат подтверждает, что генетически модифицированные мыши, несущие функциональный ген IgM и ген IgG, в котором отсутствуют домен CH1 и шарнирный домен, способны к экспрессии антител, имеющих только тяжелую цепь, в сыворотку.
B. Мыши IgG1-∆CH1
Аналогично, брали кровь у детенышей мыши, гомозиготных по делеции CH1, и детенышей дикого типа. Плазму и сыворотку (для пяти гомозигот; двух дикого типа) из крови мышей для Вестерн-блоттинга получали для определения любого экспрессировавшегося IgG в сыворотке с использованием антитела к mIgG1 (описано выше). Вестерн-блоттинги сыворотки и плазмы от мышей, гомозиготных по делеции IgG1-∆CH1, выявили смесь полос: одна полоса приблизительно 45 кДа, ожидаемый размер для IgG, в котором отсутствует домен CH1, и одна полоса приблизительно 75 кДа, ожидаемый размер для димера IgG, в котором отсутствует домен CH1 (данные не показаны). Результаты соответствуют гомозиготной VELOCIMICE®, экспрессирующей тяжелую цепь IgG1-∆CH1 с одного или обоих локусов тяжелой цепи. Этот результат подтверждает, что генетически модифицированные мыши, несущие функциональный ген IgM и ген IgG, в котором отсутствует домен CH1, способны к экспрессии антител, имеющих только тяжелую цепь, в периферическом компартменте лимфоцитов иммунной системы животных.
Пример 6: Характеристика мышей, гомозиготных по делеции IgG1-CH1-шарнир
Скрещивали VELOCIMICE®, гетерозиготных по делеции CH1-шарнир, для получения мышей, гомозиготных по делеции. Четыре детеныша мыши были определены, как гомозиготные по IgG1-∆CH1-∆шарнир. Брали кровь у этих четырех мышей и мыши дикого типа и получали сыворотку из крови мышей для вестерн-блоттинга для определения любого экспрессировавшегося IgG в сыворотке с использованием антитела к mIgG1 (как описано выше). На фигуре 9 представлена пленка, проявленная с PVDF-мембраны, использованной в этом эксперименте. Сыворотки разбавляли 1:5 и 1:10 и 10 мкл каждого разведения наносили на гель последовательно для каждой мыши. В верхней части изображений гелей помечены линии для каждой мыши, а также контроли IgG1 (1) и IgG2a (2a).
Сыворотка от мыши дикого типа показала ожидаемый паттерн для мыши дикого типа, которая экспрессирует нормальные антитела, содержащие две тяжелые цепи и две легкие цепи (приблизительно 150 кДа). Каждая из четырех мышей (гомозиготных по IgG1-∆CH1-∆шарнир) показала смесь полос: одну полосу приблизительно 150 кДа, ожидаемый размер для IgG дикого типа, иного чем IgG1 (например, IgG2a, IgG2b или IgG3), и одну полосу приблизительно 45 кДа, ожидаемый размер для IgG, в котором отсутствует домен CH1 и шарнир (фигура 9). Эти результаты соответствуют мыши, экспрессирующей антитело IgG1, имеющее только тяжелую цепь, в которой отсутствует домен CH1 и шарнирная область, и в котором отсутствует легкая цепь. Этот результат дополнительно подтверждает, что генетически модифицированные мыши, несущие функциональный ген IgM и ген IgG, в котором отсутствует домен CH1 и шарнирная область, способны к экспрессии антител, имеющих только тяжелую цепь, в сыворотку.
В другом эксперименте сывороточную экспрессию IgG определяли у мышей, гомозиготных по IgG1-∆CH1-∆шарнир, с использованием анализа ELISA. В кратком изложении, антитела, специфичные или для mIgG1 или для mIgG2b (Pharmingen), разводили раздельно и покрывали планшеты 100 мкл/лунку при концентрации 2 мкг/мл в 1×PBS (Irvine Scientific), и инкубировали при 4°C в течение ночи. На следующий день планшеты отмывали четыре раза PBS с 0,05% Tween-20 (PBST; Sigma). После четвертой отмывки планшеты блокировали PBST с 5% BSA (Sigma) в количестве 250 мкл/лунку и инкубировали при комнатной температуре в течение одного часа. Делали серийное разведение сыворотки и стандартов (фактор разведения 0,316) в PBST в 0,5% BSA вдоль планшета (сверху вниз) в начальной концентрации 400 нг/мл (mIgG1) или 600 нг/мл (mIgG2b). После блокирования планшеты снова отмывали четыре раза PBST. После четвертой отмывки в планшеты добавляли 100 мкл сыворотки или стандарта и инкубировали в течение одного часа при комнатной температуре. Планшеты снова отмывали четыре раза PBST. После отмывки в планшеты добавляли 100 мкл биотинилированного детектирующего антитела (10 нг/мл крысиного анти-mIgG1 или 250 нг/мл анти-mIgG2b; Pharmingen) и инкубировали в течение одного часа при комнатной температуре. Планшеты снова отмывали, как описано выше. После отмывки в планшеты добавляли 100 мкл/лунку пероксидазы хрена, конъюгированной со стрептавидином (HRP-SA), в разведении 1:20000 в PBST и планшеты инкубировали в течение 30 минут при комнатной температуре. Планшеты затем отмывали шесть раз PBST, после чего добавляли 100 мкл/лунку Субстрата A и B (BD OPTEIA™; BD Biosciences) в разведении 1:1 и оставляли планшеты в темноте. Реакция происходит в темноте и останавливается по желанию (примерно 15 мин) при помощи 1н фосфорной кислоты. Остановленные реакции считывали на спектрофотометре для чтения планшетов Wallac 1420 Work Station VICTOR™ при длине волны поглощения 450 нм (1,0 с/чтение) и результаты отображали на графике (фигура 11).
Сыворотка от мышей дикого типа продемонстрировала нормальные уровни IgG1 и IgG2b. Мыши, гомозиготные по IgG1-∆CH1-∆шарнир, были способны экспрессировать IgG1, в котором отсутствуют домен CH1 и шарнирная область, на периферию (сыворотка; левая сторона фигуры 11). Далее, сывороточные уровни других изотипов IgG (например, IgG2b) не были заметно снижены по сравнению с уровнями дикого типа (правая сторона фигуры 11). Этот результат дополнительно подтверждает, что генетически модифицированные мыши, несущие функциональный ген IgM и ген IgG, в котором отсутствуют домен CH1 и шарнирная область, способны к экспрессии модифицированного изотипа IgG1 (т.е. в котором отсутствуют домен CH1 и шарнир), который можно обнаружить в сыворотке.
Пример 7: Анализ V-D-J реаранжировок у мышей с модифицированным IgG1
A. Мыши, гомозиготные по делеции IgG1-CH1-шарнир
У мышей, гомозиготных по модификации IgG1-∆CH1-∆шарнир анализировали рекомбинацию V-D-J и использование генов тяжелой цепи при помощи полимеразной цепной реакции с обратной транскриптазой (RT-ПЦР) с использованием РНК, выделенной из спленоцитов.
В кратком изложении, собирали селезенки и перфузировали 10 мл RPMI-1640 (Sigma) с 5% HI-FBS в стерильных одноразовых пакетах. Каждый пакет, содержащий одну селезенку, затем помещали в STOMACHER™ (Seward) и гомогенизировали при средних параметрах в течение 30 секунд. Гомогенизированные селезенки фильтровали с использованием 0,7-мкм клеточного сита, а затем осаждали центрифугированием (1000 об/мин в течение 10 мин) и эритроциты (RBC) лизировали в BD PHARM LYSE™ (BD Biosciences) в течение трех минут. Спленоциты разбавляли RPMI-1640 и снова центрифугировали с последующим ресуспендированием в 1 мл PBS (Irvine Scientific). РНК выделяли из осажденных спленоцитов с использованием стандартных способов, известных в данной области.
RT-ПЦР проводили на РНК спленоцитов с использованием набора вырожденных праймеров, специфичных для генных сегментов вариабельной области тяжелой цепи мыши (VH) (Novagen) и праймера CH2 IgG1 мыши (CGATGGGGGC AGGGAAAGCT GCAC; SEQ ID NO:40). ПЦР продукты выделяли из геля и клонировали в pCR2.1-TOPO TA (Invitrogen) и секвенировали с праймерами M13 Forward (GTAAAACGAC GGCCAG; SEQ ID NO:41) и M13 Reverse (CAGGAAACAG CTATGAC; SEQ ID NO:42), локализованными внутри последовательности вектора в положениях, фланкирующих клонированный участок. Были секвенированы девятнадцать клонов для определения использования генов тяжелой цепи и последовательности соединения реаранжированных VH и CH2 константной области IgG1 (Таблица 2).
На фигуре 12 представлено выравнивание последовательностей доменов VH, реаранжированных с CH2 константной области IgG1, для одиннадцати из девятнадцати RT-ПЦР клонов. Последовательности, представленные на фигуре 12, показывают уникальные реаранжировки, включающие различные генные сегменты V, D и J тяжелой цепи мыши и IgG1 мыши, лишенного областей CH1 и шарнира. Мыши, гомозиготные по делеции CH1 и шарнирных областей эндогенного гена константной области IgG1, были способны производить тяжелые цепи, содержащие домены VH мыши, функционально связанные с областью CH2-CH3 из константной области мышиного IgG1, лишенного CH1 и шарнирных областей, и были способны производить B-клетки, которые экспрессировали тяжелые цепи мышиного IgG1, лишенного CH1 и шарнирных областей и в котором отсутствует легкая цепь (фигуры 8 и 9). Эти реаранжировки демонстрируют, что модифицированные локусы были способны независимо реаранжировать генные сегменты тяжелой цепи мыши во множестве независимых B-клеток у этих мышей для выработки антител, имеющих только тяжелую цепь, которые похожи на те, что обычно находят у верблюдов. Дополнительно, этот Пример демонстрирует, что делеция эндогенных областей CH1 и шарнира IgG1 не приводит к бездействию локуса и не предотвращает рекомбинацию, в которую вовлечена модифицированная константная область IgG1. Эти мыши производят функциональные антитела, имеющие только тяжелую цепь, включая IgG1, лишенный CH1 и шарнирных областей, как часть эндогенного репертуара без каких-либо выявляемых дефектов развития B-клетки.
B. Мыши, гомозиготные по делеции IgG1-CH1
Аналогично, у мышей, гомозиготных по модификации IgG1-∆CH1, анализировали рекомбинацию V-D-J и использование генов тяжелой цепи при помощи полимеразной цепной реакции с обратной транскриптазой (RT-ПЦР) с использованием РНК, выделенной из спленоцитов.
В кратком изложении, выделяли селезенки у двух гомозиготных по IgG1-∆CH1 мышей, как описано выше в разделе A данного Примера. CD19+ B-клетки выделяли с использованием магнитной сортировки клеток (MACS, Miltenyi Biotec) из общего пула спленоцитов. РНК экстрагировали из отобранных CD19+ B-клеток с использованием Qiagen ALLPREP™ ДНК/РНК mini kit (Qiagen). Первую цепь кДНК синтезировали при помощи обратной транскриптазы SUPERSCRIPT™ III и праймеров Oligo (dT)20 (Invitrogen). кДНК затем использовали в качестве матрицы для ПЦР, которую проводили с праймером, специфичным для 3' шарнира IgG1 мыши, и 5' вырожденными праймерами, предназначенными для связывания с лидерными последовательностями вариабельной области тяжелой цепи человека (Таблица 3). ПЦР продукты клонировали в pCR2.1 TOPO™ TA вектор (Invitrogen) и секвенировали с праймерами M13 Forward и M13 Reverse (как описано выше в разделе A данного Примера).
Для определения использования генов тяжелой цепи у мышей, гомозиготных по IgG1-∆CH1, были секвенированы двадцать восемь RT-ПЦР клонов. Среди этих клонов наблюдали семь уникальных реаранжировок генных сегментов V, D и J человека (Таблица 4).
На фигуре 13 представлено выравнивание последовательностей доменов VH, реаранжированных с шарниром-CH2-CH3 константной области IgG1, для семи реаранжировок, показанных в таблице 4. Последовательности, представленные на фигуре 13, показывают уникальные реаранжировки, включающие различные генные сегменты V, D и J тяжелой цепи человека и IgG1 мыши, лишенного области CH1. Мыши, гомозиготные по делеции области CH1 эндогенного гена константной области IgG1, были способны производить тяжелые цепи, содержащие домены VH человека, функционально связанные с областью шарнир-CH2-CH3 из константной области мышиного IgG1, лишенного CH1, и были способны производить B-клетки, которые экспрессировали тяжелые цепи мышиного IgG1, лишенного областей CH1 и в котором отсутствует легкая цепь (данные не показаны). Эти реаранжировки демонстрируют, что один или оба модифицированных локуса (IgG1-∆CH1-∆шарнир и IgG1-∆CH1) были способны независимо реаранжировать генные сегменты тяжелой цепи (мыши и человека) во множестве независимых B-клеток у этих мышей для выработки антител, имеющих только тяжелую цепь, которые похожи на те, что обычно находят у верблюдов. Дополнительно, этот Пример демонстрирует, что делеция эндогенного CH1 IgG1 не приводит к бездействию локуса и не предотвращает рекомбинацию, в которую вовлечены генные сегменты V, D и J тяжелой цепи человека и модифицированная константная область IgG1 мыши. Эти мыши производят функциональные антитела, имеющие только тяжелую цепь, содержащие V-домены тяжелой цепи человека и мышиный IgG1, лишенный CH1, как часть эндогенного репертуара без каких-либо выявляемых дефектов развития B-клетки.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ ДЛЯ ТЕСТИРОВАНИЯ ТЕРАПЕВТИЧЕСКИХ АГЕНТОВ | 2019 |
|
RU2805212C2 |
| МЫШИ, У КОТОРЫХ ВЫРАБАТЫВАЮТСЯ СВЯЗЫВАЮЩИЕ БЕЛКИ, СОДЕРЖАЩИЕ VL-ДОМЕНЫ | 2011 |
|
RU2612903C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664181C2 |
| ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА ЖИВОТНЫЕ, ЭКСПРЕССИРУЮЩИЕ ЧУВСТВИТЕЛЬНЫЕ К pН ПОСЛЕДОВАТЕЛЬНОСТИ ИММУНОГЛОБУЛИНОВ | 2013 |
|
RU2664473C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ, КОТОРЫЕ ЭКСПРЕССИРУЮТ ТЯЖЕЛЫЕ ЦЕПИ, СОДЕРЖАЩИЕ ДОМЕНЫ VL | 2013 |
|
RU2770016C2 |
| МЫШИ, У КОТОРЫХ ВЫРАБАТЫВАЮТСЯ СВЯЗЫВАЮЩИЕ БЕЛКИ, СОДЕРЖАЩИЕ V-ДОМЕНЫ | 2011 |
|
RU2750219C2 |
| ГУМАНИЗИРОВАННЫЕ НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕКУ ЖИВОТНЫЕ С ОГРАНИЧЕННЫМИ ЛОКУСАМИ ТЯЖЕЛОЙ ЦЕПИ ИММУНОГЛОБУЛИНА | 2013 |
|
RU2656155C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664232C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2787781C2 |
| МЫШЬ С ОБЩЕЙ ЛЕГКОЙ ЦЕПЬЮ | 2013 |
|
RU2683514C2 |

























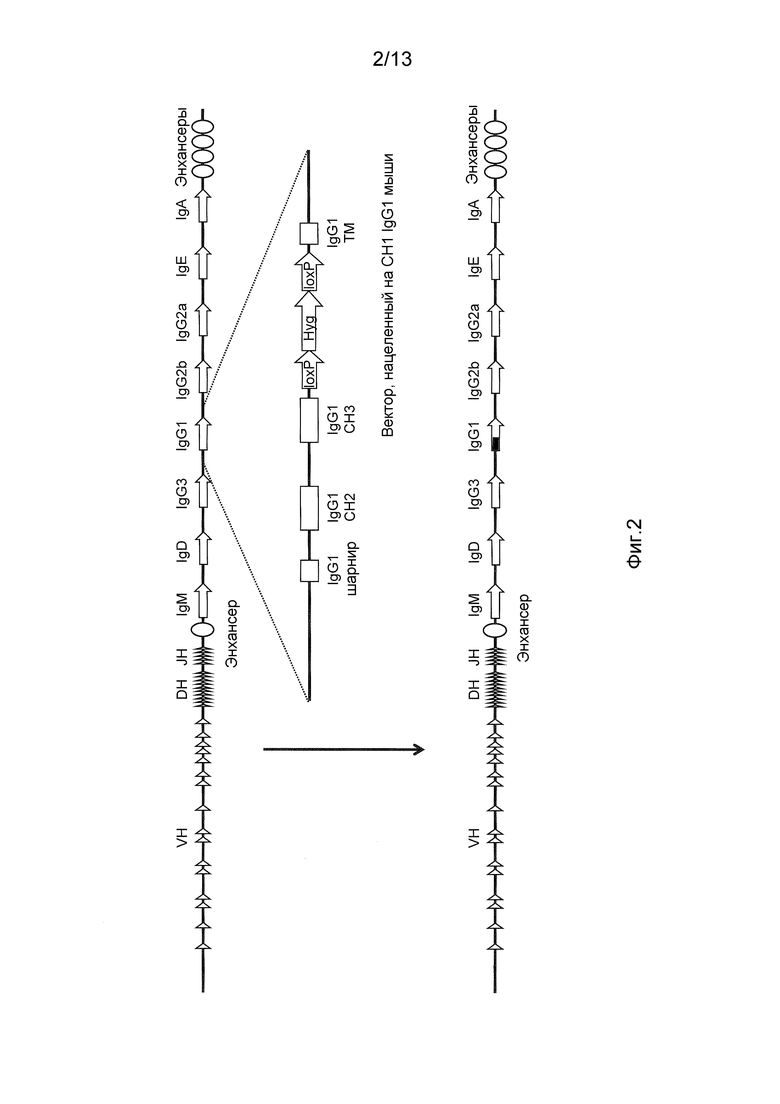




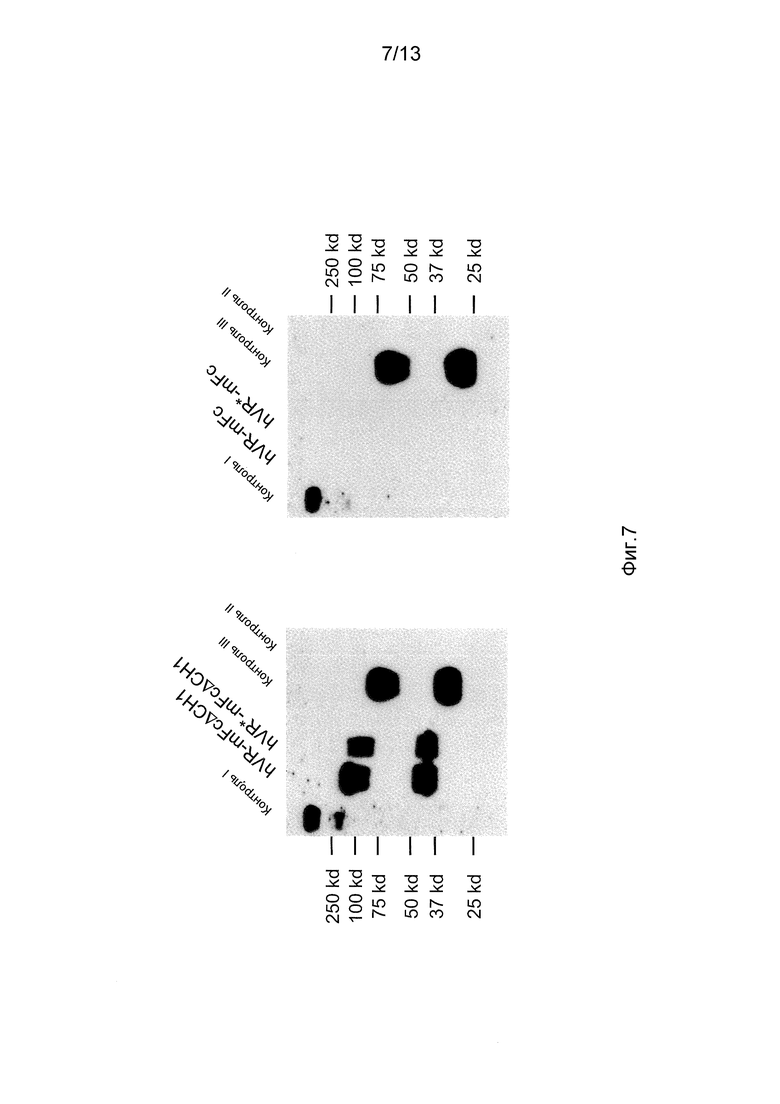

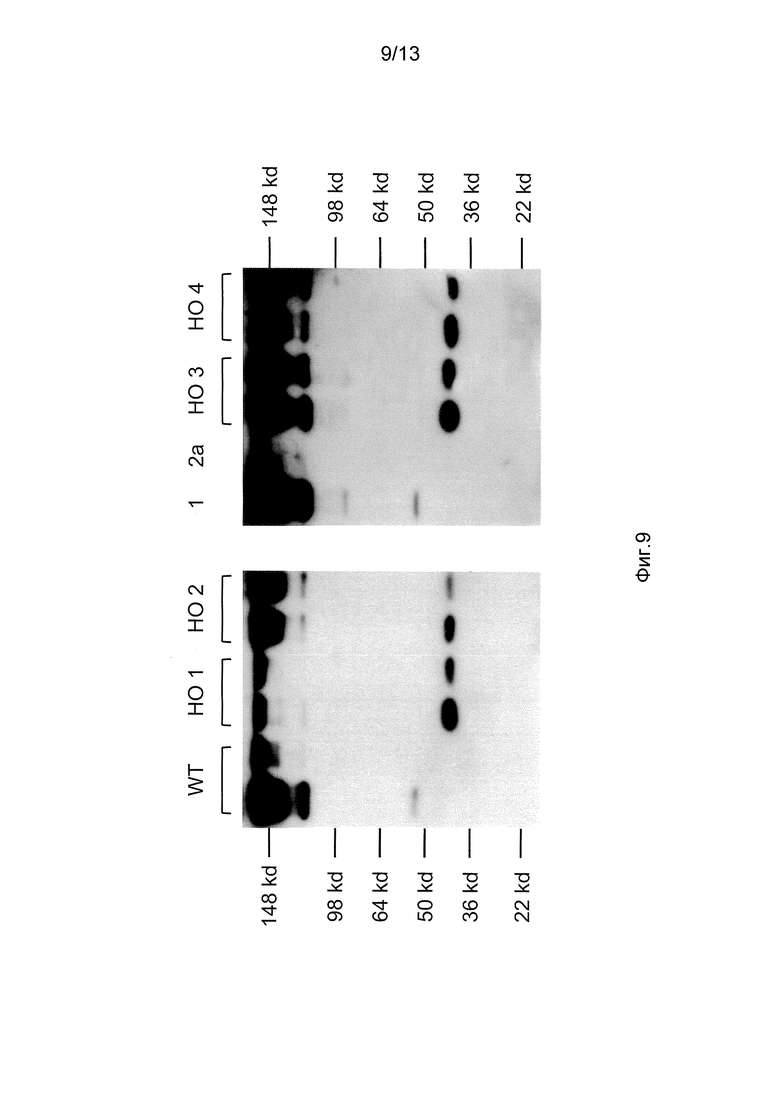

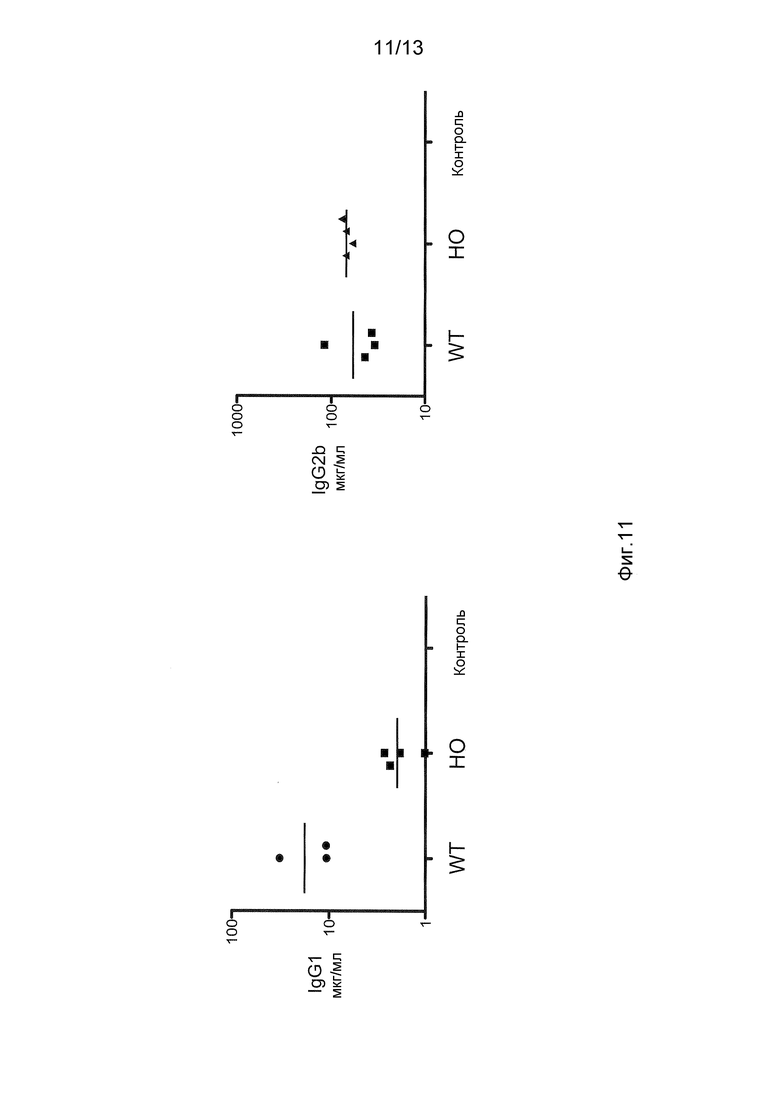

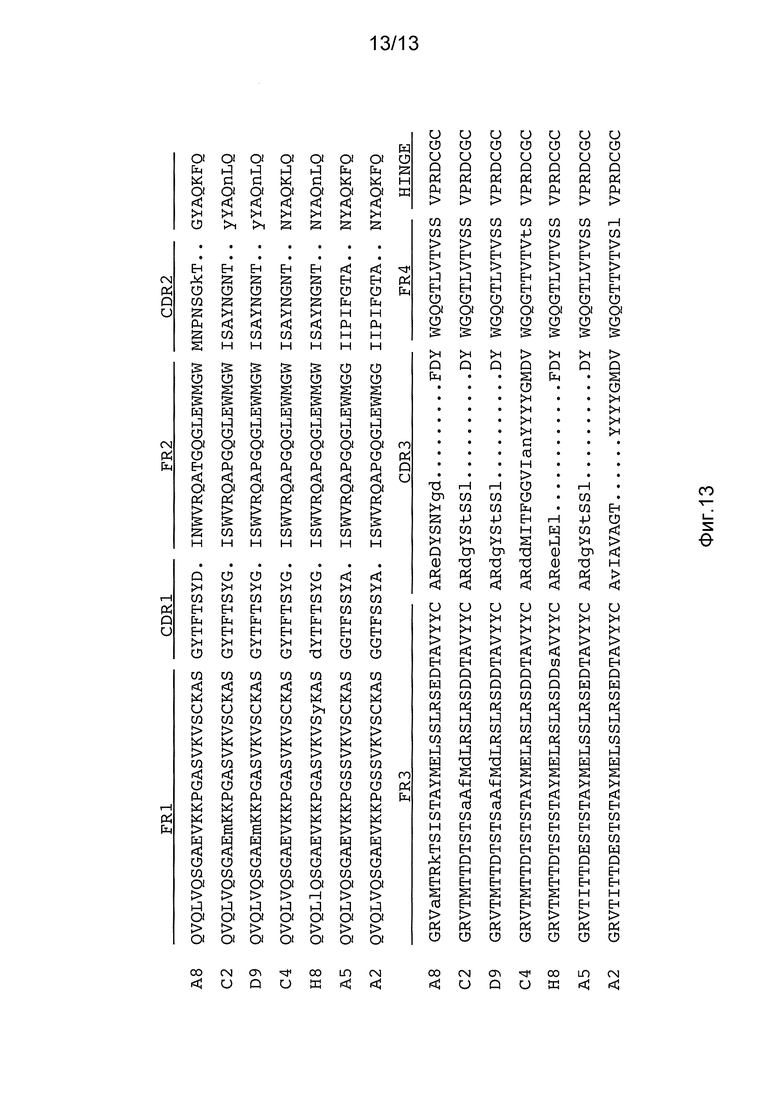
Изобретение относится к области биотехнологии. Описаны генетически модифицированные мыши, включая мышей с функциональным геном IgM и модифицированных для получения делеции домена CH1 и шарнирной области в константном домене тяжелой цепи, которая не является цепью IgM, например, в константном домене тяжелой цепи IgG. Предлагаются генетически модифицированные мыши, которые производят химерные антитела с вариабельной областью человека/константной областью мыши, имеющие только тяжелые цепи (антитела, в которых отсутствует легкая цепь), полностью мышиные антитела, имеющие только тяжелую цепь, или полностью человеческие антитела, имеющие только тяжелую цепь. Предложенная группа изобретений может быть использована в медицине. 15 н. и 48 з.п. ф-лы, 13 ил., 4 табл., 7 пр.
1. Мышь для продукции IgG антител, имеющих только тяжелые цепи, содержащая генетическую модификацию зародышевой линии, которая включает:
делецию нуклеотидной последовательности, кодирующей домен СН1, эндогенного гена константной области IgG;
где мышь экспрессирует ген константной области IgM, которая кодирует функциональный домен СН1, и мышь экспрессирует в сыворотку антитело IgG, в котором отсутствует домен СН1 полностью или частично и в котором отсутствует родственная легкая цепь.
2. Мышь по п. 1, где ген константной области IgG выбран из группы, состоящей из гена константной области IgG1, гена константной области IgG2b, гена константной области IgG2a и их сочетания.
3. Мышь по п. 1, где ген константной области IgG представляет собой ген константной области IgG1.
4. Мышь по п. 1, которая содержит функциональный генетический локус легкой цепи иммуноглобулина.
5. Мышь по п. 4, где генетический локус легкой цепи иммуноглобулина представляет собой генетический локус κ легкой цепи.
6. Мышь по п. 4, где генетический локус легкой цепи иммуноглобулина представляет собой генетический локус λ легкой цепи.
7. Мышь по п. 1, где антитело IgG, в котором отсутствует родственная легкая цепь, содержит:
(a) вариабельный домен человека и константный домен мыши, в котором отсутствует домен СН1, или
(b) вариабельный домен мыши и константный домен мыши, в котором отсутствует домен СН1.
8. Мышь по п. 1, которая дополнительно содержит:
(a) делецию гена константной области IgG2a;
(b) делецию гена константной области IgG2b;
(c) делецию гена константной области IgG2a и делецию гена константной области IgG2b; или
(d) делецию шарнирной области гена константной области IgG, который содержит делецию по меньшей мере части нуклеотидной последовательности, кодирующей домен СН1.
9. Мышь по п. 1, характеризующаяся тем, что в тяжелой цепи антитела IgG полностью отсутствует домен СН1.
10. Мышь по любому из пп. 1-9, которая дополнительно содержит замену одного или нескольких эндогенных генных сегментов вариабельной области тяжелой цепи мыши одним или несколькими генными сегментами вариабельной области тяжелой цепи человека, при этом один или несколько генных сегментов вариабельной области тяжелой цепи человека способны рекомбинировать для образования реаранжированного гена вариабельной области человека, функционально связанного с эндогенным геном константной области мыши.
11. Мышь по п. 10, где один или несколько генных сегментов вариабельной области тяжелой цепи человека представляют собой генные сегменты семейства VH1, VH3 или VH4 человека.
12. Мышь по п. 11, где генные сегменты семейства вариабельной области тяжелой цепи человека представляют собой генные сегменты, выбранные из группы генных сегментов VH человека 1-2, 1-8, 1-18, 1-46, 1-69, 3-21, 3-72 и 4-59.
13. Мышь по п. 11, где все эндогенные генные сегменты вариабельной области тяжелой цепи мыши замещены 18, 39 или 81 генными сегментами вариабельной области человека.
14. Мышь по п. 11, где генные сегменты вариабельной области тяжелой цепи человека выбраны из 1-8, 1-18 и 1-69.
15. Мышь по п. 10, где IgG антитело, имеющее только тяжелые цепи, содержит вариабельную область человека, полученную из гена вариабельной области человека, и константную область, полученную из гена константной области, при этом константная область содержит шарнир IgG1, домен СН2 и домен СН3.
16. Мышь по п. 1 или 10, характеризующаяся тем, что мышь дополнительно эксперссирует:
a) белок IgG3 дикого типа,
b) белок IgG2a дикого типа и
c) белок IgG2b дикого типа.
17. Мышь по п. 16, дополнительно характеризующаяся тем, что мышь эксперссирует:
e) белок IgM дикого типа,
f) белок IgD дикого типа,
g) белок IgA дикого типа и
h) белок IgE дикого типа.
18. Мышь по п. 1 или 10, где указанная мышь получена из линии выбранной из группы, состоящей из линии 129, линии C57BL/6 и помеси 129 x C57BL/6.
19. Мышь по п. 18, где мышь является на 50% 129 и 50% C57BL/6.
20. Мышь по п. 1, которая содержит замену одного или нескольких эндогенных генных сегментов вариабельной области тяжелой цепи мыши одним или несколькими камелизированными генными сегментами вариабельной области тяжелой цепи человека.
21. Генетически модифицированная мышь для продукции IgG антител, имеющих только тяжелые цепи, которая содержит замену всех или, по существу, всех генных сегментов V, D и J одним или несколькими генными сегментами V человека, D человека и J человека, где один или несколько генных сегментов V, D и J человека функционально связаны в эндогенном локусе мыши с генетическим локусом константной области тяжелой цепи мыши, где генетический локус константной области тяжелой цепи мыши содержит полноразмерный ген константной области IgM, ген константной области IgG, который содержит делецию нуклеотидной последовательности, кодирующей домен СН1, в гене константной области IgG, выбранном из IgG1, IgG2a, IgG2b, IgG3 и их сочетания; где мышь экспрессирует B-клеточный рецептор, который содержит IgM с функциональной областью СН1, и где IgM содержит тяжелую цепь, ассоциированную с родственной легкой цепью лямбда или каппа.
22. Генетически модифицированная мышь по п. 21, где один или несколько генных сегментов V, D и J человека содержат генный сегмент человека из семейства вариабельной области, выбранной из VH1, VH3, VH4 и их сочетания.
23. Генетически модифицированная мышь по п. 22, где один или несколько генных сегментов человека выбраны из 1-2, 1-8, 1-18, 1-46, 1-69, 3-21, 3-72, 4-59 и их сочетания.
24. Генетически модифицированная мышь по п. 22, где один или несколько генных сегментов V, D и J человека содержат D1-7, D3-16, D4-4, D6-6, D6-7, D6-19, J2, J3, J4 и их сочетание.
25. Изолированная клетка мыши по любому из пп. 1-24 для продукции IgG антител, имеющих только тяжелые цепи, при этом клетка содержит нуклеотидную последовательность, кодирующую функциональный домен СН1 эндогенной константной области IgM и делецию нуклеотидной последовательности, кодирующей домен СН1, эндогенного гена константной области IgG.
26. Изолированная клетка мыши по п. 25, в которой клетка представляет собой В-клетку.
27. Изолированная эмбриональная стволовая клетка (ES) мыши по любому из пп. 1-24 для получения мыши, способной к продукции IgG антител, имеющих только тяжелую цепь, при этом клетка содержит нуклеотидную последовательность, кодирующую функциональный домен СН1 эндогенного гена константной области IgM, и делецию нуклеотидной последовательности, кодирующей домен СН1 эндогенного гена константной области IgG.
28. Способ получения мыши, которая экспрессирует IgG антитела, имеющие только тяжелые цепи, в которых отсутствует домен СН1 полностью или частично, а также экспрессирует антитела IgM, состоящие из двух легких цепей и двух тяжелых цепей, в которых каждая из тяжелых цепей IgM включает функциональный домен СН1, при этом способ включает этап удаления из генома мыши нуклеотидной последовательности, кодирующей домен СН1 эндогенного гена IgG.
29. Способ получения IgG антител, имеющих только тяжелые цепи, в которых отсутствует домен СН1, при этом способ включает следующие этапы:
a) получения мыши, зародышевая линия которой содержит
i - ген константной области IgG, в которой отсутствует функциональная СН1 последовательность IgG;
ii - константную область IgM, включающую функциональную СН1 последовательность IgM; и
b) содержания мыши таким образом, что указанная мышь экспрессирует:
i - антитела IgM, состоящие из двух тяжелых цепей IgM, в которых каждая из тяжелых цепей IgM включает функциональный домен СН1, и двух родственных легких цепей; и
ii - IgG антитела, имеющие только тяжелые цепи, состоящие из тяжелых цепей IgG, в которых отсутствует домен СН1, и которые не включают родственные легкие цепи.
30. Способ получения соматически мутированного антитела, имеющего только тяжелые цепи, включающий этапы:
a) иммунизации мыши антигеном, где мышь содержит в зародышевой линии делецию нуклеотидной последовательности по меньшей мере одного аллеля, который кодирует домен СН1, и где мышь экспрессирует ген константной области IgM, который содержит функциональный домен СН1;
b) содержания мыши в условиях, достаточных для инициации иммунного ответа на антиген; и
c) выделения из мыши клетки или белка, которые специфически связываются с антителом, где клетка или белок включают соматически мутированное антитело, имеющее только тяжелые цепи, которое специфически связывается с антигеном.
31. Способ по п. 30, в котором по меньшей мере одна константная область IgG выбрана из группы, состоящей из IgG1, IgG2b, IgG2a и их сочетания.
32. Способ по п. 31, в котором ген константной области IgG представляет собой ген константной области IgG1.
33. Способ по п. 32, в котором мышь дополнительно экспрессирует а) белок IgG3 дикого типа, b) белок IgG2a дикого типа и с) белок IgG2b дикого типа.
34. Способ по п. 33, в котором мышь дополнительно экспрессирует d) белок IgM дикого типа, е) белок IgD дикого типа, f) белок IgA дикого типа и h) белок IgE дикого типа.
35. Способ по п. 30, в котором у антитела, имеющего только тяжелые цепи, полностью отсутствует домен СН1.
36. Способ по п. 35, в котором антитело, имеющее только тяжелые цепи, содержит шарнир IgG1, домен СН2 и домен СН3.
37. Способ по п. 30, в котором мышь содержит функциональный локус легкой цепи иммуноглобулина.
38. Способ по п. 37, в котором локус легкой цепи иммуноглобулина представляет собой генетический локус κ легкой цепи.
39. Способ по п. 37, в котором локус легкой цепи иммуноглобулина представляет собой генетический локус λ легкой цепи.
40. Способ по п. 30, в котором мышь получена из линии, выбранной из группы, состоящей из линии 129, линии C57BL/6 и помеси 129 x C57BL/6.
41. Способ по п. 40, в котором мышь является на 50% 129 и 50% C57BL/6.
42. Способ по п. 30, в котором мышь дополнительно содержит замену всех или, по существу, всех эндогенных генных сегментов VH, DH и JH мыши одним или несколькими генными сегментами VH человека, DH человека и JH человека, где один или несколько генных сегментов VH, DH и JH человека функционально связаны в эндогенном локусе мыши с генетическим локусом константной области тяжелой цепи мыши, и где генетический локус константной области тяжелой цепи мыши содержит ген константной области IgM, имеющей функциональный домен СН1, и ген IgG, который содержит делецию нуклеотидной последовательности, кодирующей домен СН1, в гене IgG, выбранном из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3 и их сочетания.
43. Способ по п. 42, в котором антитело, имеющее только тяжелые цепи, содержит вариабельную область тяжелой цепи.
44. Способ по п. 43, дополнительно включающий в качестве этапа:
d) выделение нуклеиновых кислот вариабельной области тяжелой цепи из клеток, выделенных на этапе с).
45. Способ по п. 44, дополнительно включающий в качестве последних этапов:
e) трансфекцию клетки вектором, содержащим вторую нуклеиновую кислоту, функционально связанную с геном константной области тяжелой цепи человека, в котором нуклеотидная последовательность, кодирующая домен СН1, удалена, при этом указанная нуклеиновая кислота идентична или, по существу, идентична нуклеиновым кислотам тяжелой цепи, выделенным на этапе (с), и
f) культивирование клетки в условиях, достаточных для экспрессии вектора.
46. Способ по п. 45, в котором ген константной области тяжелой цепи представляет собой IgG человека, выбранный из группы, состоящей из IgG1, IgG2a, IgG2b и IgG3.
47. Способ по п. 30, дополнительно включающий в качестве этапа:
d) выделение нуклеиновых кислот вариабельной области тяжелой цепи из клеток, выделенных на этапе (с).
48. Способ по п. 47, дополнительно включающий в качестве последних этапов:
e) трансфекцию клетки вектором, содержащим вторую нуклеиновую кислоту, функционально связанную с геном константной области тяжелой цепи человека, в которой нуклеотидная последовательность, кодирующая домен СН1, удалена, при этом нуклеиновая кислота идентична или, по существу, идентична нуклеиновым кислотам тяжелой цепи, выделенным на этапе (с), и
f) культивирование клетки в условиях, достаточных для экспрессии вектора.
49. Способ по п. 48, в котором ген константной области тяжелой цепи представляет собой IgG человека, выбранный из группы, состоящей из IgG1, IgG2a, IgG2b и IgG3.
50. Способ по п. 30, дополнительно включающий этапы:
d) получения гибридомы из клетки, выделенной на этапе (с),
e) культивирования гибридомы и
f) отбора антител, имеющих только тяжелые цепи, специфичных к антигену, из культурального супернатанта, полученного на этапе (е).
51. Способ по п. 50, дополнительно включающий в качестве этапа:
g) выделение нуклеиновых кислот вариабельной области тяжелой цепи из гибридомы, полученной на этапе (d).
52. Способ по п. 51, дополнительно включающий в качестве последних этапов:
h) трансфекцию клетки вектором, содержащим вторую нуклеиновую кислоту, функционально связанную с геном константной области тяжелой цепи, в котором нуклеотидная последовательность, кодирующая домен СН1, удалена, при этом нуклеиновая кислота, по существу, идентична нуклеиновым кислотам тяжелой цепи, выделенным на этапе (g), и
i) культивирование клетки в условиях, достаточных для экспрессии вектора, и
j) выделение белка, кодируемого вектором.
53. Способ по п. 52, в котором ген константной области тяжелой цепи представляет собой IgG человека, выбранный из группы, состоящей из IgG1, IgG2a, IgG2 и IgG3.
54. Гибридома для получения соматически мутированного IgG антитела, имеющего только тяжелые цепи, при этом гибридома получена способом, включающим следующие этапы:
а) иммунизации мыши антигеном, где мышь содержит в зародышевой линии делецию нуклеотидной последовательности по меньшей мере одного аллеля, который кодирует домен СН1, и где мышь экспрессирует ген константной области IgM, который содержит функциональный домен СН1;
b) содержания мыши в условиях, достаточных для инициации иммунного ответа на антиген;
c) выделения из мыши клетки, которая специфически связывается с антителом, где клетка включает соматически мутированное IgG антитело, имеющее только тяжелые цепи, которое специфически связывается с антигеном;
d) получения гибридомы из клетки, выделенной на этапе (с).
55. Нуклеиновая кислота, кодирующая вариабельную область тяжелой цепи человека, выделенная способом, включающим следующие этапы:
a) иммунизации мыши антигеном, где мышь содержит в зародышевой линии делецию нуклеотидной последовательности по меньшей мере одного аллеля, который кодирует домен СН1 по меньшей мере одной констатной области IgG,
при этом мышь дополнительно содержит замену всех или, по существу, всех эндогенных генных сегментов VH, DH и JH мыши одним или несколькими генными сегментами VH человека, DH человека и JH человека, где один или несколько генных сегментов VH, DH и JH человека функционально связаны в эндогенном локусе мыши с генетическим локусом константной области тяжелой цепи мыши, и где генетический локус константной области тяжелой цепи мыши содержит ген константной области IgM, имеющей функциональный домен СН1, и ген IgG, который содержит делецию по меньшей мере в части нуклеотидной последовательности, кодирующей домен СН1, в гене IgG, выбранном из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3 и их сочетания,
и при этом мышь экспрессирует ген константой области IgM, имеющей функциональный домен СН1;
b) содержания мыши в условиях, достаточных для инициации иммунного ответа на антиген; и
c) выделения из мыши клетки или белка, которые специфически связываются с антителом, где клетка или белок включают соматически мутированное IgG антитело, имеющее только тяжелые цепи, которое специфически связывается с антигеном;
d) выделение нуклеиновых кислот вариабельной области тяжелой цепи из клеток, выделенных на этапе (с).
56. Нуклеиновая кислота, кодирующая вариабельную область тяжелой цепи, при этом нуклеиновая кислота выделена способом, включающим следующие этапы:
a) иммунизации мыши антигеном, где мышь содержит в зародышевой линии делецию нуклеотидной последовательности по меньшей мере одного аллеля, который кодирует домен СН1, и где мышь экспрессирует ген константной области IgM, который содержит функциональный домен СН1;
b) содержания мыши в условиях, достаточных для инициации иммунного ответа на антиген;
c) выделения из мыши клетки или белка, которые специфически связываются с антителом, где клетка или белок включают соматически мутированное IgG антитело, имеющее только тяжелые цепи, которое специфически связывается с антигеном; и
d) выделения нуклеиновых кислот вариабельной области тяжелой цепи из клеток, выделенных на этапе (с).
57. Нуклеиновая кислота, кодирующая вариабельную область тяжелой цепи, выделенная из гибридомы, при этом нуклеиновая кислота выделена способом, включающим следующие этапы:
a) иммунизации мыши антигеном, где мышь содержит в зародышевой линии делецию нуклеотидной последовательности по меньшей мере одного аллеля, который кодирует домен СН1, и где мышь экспрессирует ген константной области IgM, который содержит функциональный домен СН1;
b) содержания мыши в условиях, достаточных для инициации иммунного ответа на антиген;
c) выделения из мыши клетки или белка, которые специфически связываются с антителом, где клетка или белок включают соматически мутированное IgG антитело, имеющее только тяжелые цепи, которое специфически связывается с антигеном;
d) получения гибридомы из клетки, выделенной на этапе (с);
e) выделения нуклеиновых кислот вариабельной области тяжелой цепи из гибридомы, полученной на этапе (d).
58. Клетка не являющегося человеком животного для продукции вариабельного домена тяжелой цепи IgG, содержащая нуклеиновую кислоту по п. 55.
59. Клетка не являющегося человеком животного для продукции вариабельного домена тяжелой цепи IgG, содержащая нуклеиновую кислоту по п. 56.
60. Клетка не являющегося человеком животного для продукции вариабельного домена тяжелой цепи IgG, содержащая нуклеиновую кислоту по п. 57.
61. Антитело IgG, имеющее только тяжелые цепи, полученное способом, включающим этапы:
a) иммунизации мыши антигеном, где мышь содержит в зародышевой линии делецию нуклеотидной последовательности по меньшей мере одного аллеля, который кодирует домен СН1 по меньшей мере одной эндогенной константной области IgG, при этом мышь дополнительно содержит замену всех или, по существу, всех эндогенных генных сегментов VH, DH и JH мыши одним или несколькими генными сегментами VH человека, DH человека и JH человека, где один или несколько генных сегментов VH, DH и JH человека функционально связаны в эндогенном локусе мыши с генетическим локусом константной области тяжелой цепи мыши, и где генетический локус константной области тяжелой цепи мыши содержит ген константной области IgM, имеющей функциональный домен СН1, и ген IgG, который содержит делецию нуклеотидной последовательности, кодирующей домен СН1, в гене IgG, выбранном из группы, состоящей из IgG1, IgG2a, IgG2b, IgG3 и их сочетания, и где мышь экспрессирует ген константной области IgM, которая содержит функциональный домен СН1;
b) содержания мыши в условиях, достаточных для инициации иммунного ответа на антиген; и
c) выделения из мыши соматически мутированного антитела, имеющего только тяжелые цепи, содержащего вариабельную область тяжелой цепи человека, которое специфически связывается с антигеном.
62. Антитело IgG, имеющее только тяжелые цепи, по п. 61, где способ дополнительно включает в качестве этапов:
d) выделение нуклеиновых кислот вариабельной области тяжелой цепи из клеток, выделенных на этапе (с);
e) трансфекцию клетки вектором, содержащим вторую нуклеиновую кислоту, функционально связанную с геном константной области тяжелой цепи человека, в котором по меньшей мере часть домена СН1 удалена, при этом указанная нуклеиновая кислота идентична или, по существу, идентична нуклеиновым кислотам тяжелой цепи, выделенным на этапе (d);
f) культивирование клетки в условиях, достаточных для экспрессии вектора; и
g) выделение белка, кодируемого вектором.
63. Антитело IgG, имеющее только тяжелые цепи, по п. 62, где ген константной области тяжелой цепи человека представляет собой IgG человека, выбранный из группы, состоящей из IgG1, IgG2a, IgG2b и IgG3.
| JANSSENS R | |||
| et al., "Generation of heavy-chain-only antibodies in mice", Proc Natl Acad Sci U S A | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| RIECHMANN L | |||
| et al., "Single domain antibodies: comparison of camel VH and camelised human VH domains", J Immunol Methods | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |
| BRUGGEMANN M | |||
| et al., "Heavy-chain-only antibody expression and B-cell | |||

