ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По данной заявке испрашивается приоритет предварительной заявки на патент США номер 61/349012, поданной 27 мая 2010 г., содержание которой во всей своей полноте включено в настоящий документ посредством ссылки.
УРОВЕНЬ ТЕХНИКИ
Наноразмерные компьютеры потенциально могут быть реализованы с использованием молекул. Такие компьютеры могут подходить для решения определенных вычислительных задач. В частности, компьютеры, в которых используются биологические молекулы, могут быть совместимыми с биологическими средами, и потенциально могут быть применены для диагностики или даже лечения сложных заболеваний.
Способность преобразования одной последовательности нуклеиновых кислот в другую может быть использована для построения логических элементов и сетей с нуклеиновыми кислотами. Такие элементы и схемы управляются двумя событиями: гибридизацией и замещением цепи. Оба события обычно являются термодинамически предпочтительными; то есть, они предполагают переход от высокоэнергетического состояния к низкоэнергетическому. Таким образом, оба события могут самопроизвольно происходить в системе.
Гибридизация включает в себя свободные однонитевые удлинения нуклеиновых кислот. Соответственно, сеть нуклеиновых кислот может регулироваться доступностью таких свободных нитей.
"Событие блокирования" позволяет сделать определенные последовательности условно доступными для остальной части сети. Такие события дают возможность построения преобразователей, которые преобразуют одну однонитевую последовательность нуклеиновых кислот в отличающуюся однонитевую последовательность нуклеиновых кислот. Такие преобразователи являются основой, на которой с использованием нуклеиновых кислот могут быть построены базовые логические операторы, такие как AND, NOT, OR, NAND, NOR, XOR и XNOR. Из этих и других логических компонентов могут быть построены сети большего размера, которые включают в себя такие компоненты, как усилители. В результате данные события преобразования являются важными для информационной обработки с использованием нуклеиновых кислот и для молекулярных вычислений.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В соответствии с одним из аспектов настоящего изобретения представлен способ для улучшения системы молекулярных вычислений, основанной на нуклеиновых кислотах. Способ включает в себя: (A) идентификацию вычислительной системы, состоящей из (i) структуры из нуклеиновой кислоты, которая включает в себя дуплексный домен с неполным спариванием оснований, (ii) по меньшей мере, одной полинуклеотидной молекулы перестановки, которая связывается со структурой из нуклеиновой кислоты в условиях гибридизации таким образом, что структура из нуклеиновой кислоты подвергается переходу в другое энергетическое состояние из-за реакции миграции ветви, в которую вовлечен дуплексный домен, и (iii) полинуклеотидной молекулы столкновения, которая конкурирует с полинуклеотидной молекулой перестановки за связывание со структурой из нуклеиновой кислоты в условиях гибридизации, но которая не может привести к реакции миграции ветви, в которую вовлечен дуплексный домен; затем (B) изменение конфигурации, по меньшей мере, одной молекулы из молекулы перестановки и структуры из нуклеиновой кислоты, соответственно, с целью внедрения химической модификации по отношению к первой референсной молекуле, которая содержит природные нуклеозиды и имеет тот же состав последовательности, что и молекула перестановки или структура из нуклеиновой кислоты, в зависимости от конкретной ситуации. Указанная ранее модификация приводит к тому, что связь молекулы перестановки и структуры из нуклеиновой кислоты имеет свободную энергию гибридизации, отличающуюся от энергии первого референсного связывания между молекулой перестановки или структурой из нуклеиновой кислоты и первой референсной молекулой, что больше способствует реакции миграции ветви по сравнению с первым референсным связыванием. После или вместо этапа (B) следует этап (C) изменения конфигурации, по меньшей мере, одной молекулы из полинуклеотидной молекулы столкновения и структуры из нуклеиновой кислоты, соответственно, с целью внедрения химической модификации по отношению ко второй референсной молекуле, которая содержит природные нуклеозиды и имеет тот же состав последовательности, что и молекула столкновения или структура из нуклеиновой кислоты, в зависимости от конкретной ситуации. Модификация приводит к тому, что связь молекулы столкновения и структуры из нуклеиновой кислоты имеет свободную энергию гибридизации, отличающуюся от энергии второго референсного связывания между молекулой столкновения или структурой из нуклеиновой кислоты и второй референсной молекулой, в результате чего связывание молекулы столкновения затрудняется по сравнению со вторым референсным связыванием.
Изобретение также предоставляет в другом своем аспекте систему, которая включает в себя (A) структуру из нуклеиновой кислоты, которая включает в себя дуплексный домен с неполным спариванием оснований; (B) по меньшей мере, одну полинуклеотидную молекулу перестановки для осуществления связывания со структурой из нуклеиновой кислоты в условиях гибридизации таким образом, что структура из нуклеиновой кислоты подвергается переходу в другое энергетическое состояние из-за реакции миграции ветви, в которую вовлечен дуплексный домен; и (C) по меньшей мере, одну полинуклеотидную молекулу столкновения, способную связываться со структурой из нуклеиновой кислоты в условиях гибридизации таким образом, что такая структура из нуклеиновой кислоты и молекула столкновения связываются и предотвращается связывание полинуклеотидной молекулы перестановки. По меньшей мере, одна молекула из молекулы перестановки и структуры из нуклеиновой кислоты, соответственно, имеет химическую модификацию по отношению к референсной молекуле, которая содержит природные нуклеозиды и имеет тот же состав последовательности, что и молекула перестановки или структура из нуклеиновой кислоты, в зависимости от конкретной ситуации. Модификация приводит к тому, что связь имеет свободную энергию гибридизации, отличающуюся от энергии референсного связывания между молекулой перестановки или структурой из нуклеиновой кислоты и референсной молекулой, что больше способствует реакции миграции ветви по сравнению с референсным связыванием. В дополнение или в качестве альтернативы, по меньшей мере, одна молекула из молекулы столкновения и структуры из нуклеиновой кислоты, соответственно, имеет химическую модификацию по отношению к референсной молекуле, которая содержит природные нуклеозиды и имеет тот же состав последовательности, что и молекула перестановки или структура из нуклеиновой кислоты, в зависимости от конкретной ситуации. Модификация приводит к тому, что связь имеет свободную энергию гибридизации, отличающуюся от энергии референсного связывания между молекулой перестановки или структурой из нуклеиновой кислоты и референсной молекулой, в результате чего связывание молекулы столкновения затрудняется по сравнению референсным связыванием.
В еще одном аспекте представлена система распространения информации. Система включает в себя первую молекулу и вторую молекулу, сконфигурированную для связывания с первой молекулой. По меньшей мере, одна молекула из первой молекулы и второй молекулы, соответственно, имеет химическую модификацию относительно референсной молекулы, которая имеет тот же состав последовательности, что и первая молекула или вторая молекула, в зависимости от конкретной ситуации. Модификация приводит к тому, что связь имеет свободную энергию гибридизации, отличающуюся от энергии между референсной молекулой и первой или второй молекулой, в результате чего в системе реализуется, по меньшей мере, одно из следующего: вероятность продуктивного связывания повышается или вероятность непродуктивного связывания снижается.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
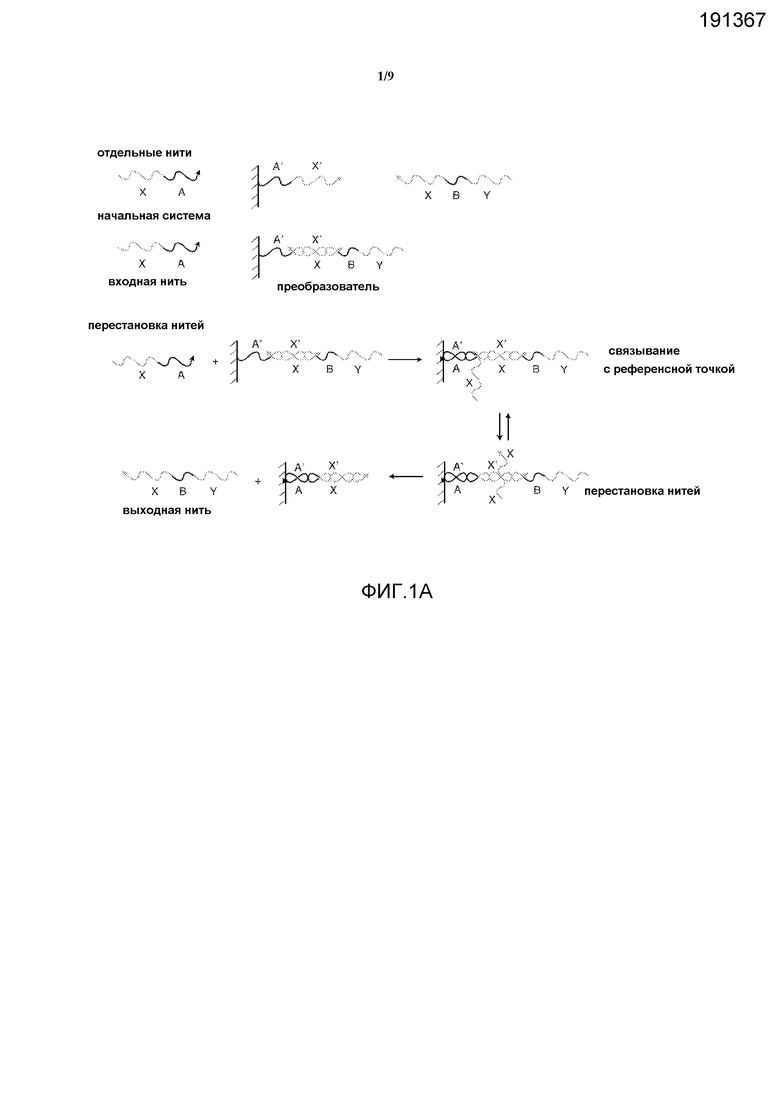
Фигура 1A иллюстрирует твердофазную изолированную реализацию преобразователя нуклеиновых кислот, при этом отдельные секции (A, B и т.д.) представляют собой фрагменты олигонуклеотидов произвольной длины и последовательности.
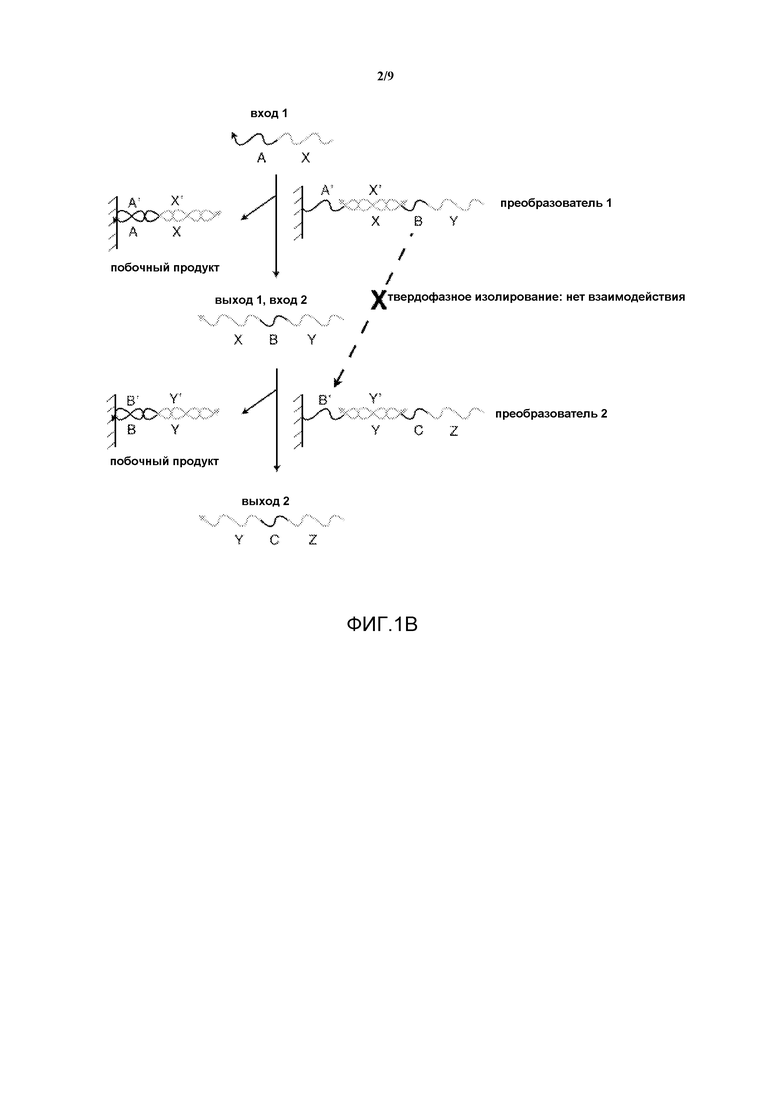
Фигура 1B иллюстрирует, каким образом твердофазная реализация преобразователя нуклеиновых кислот изолирует фрагменты олигонуклеотидов.
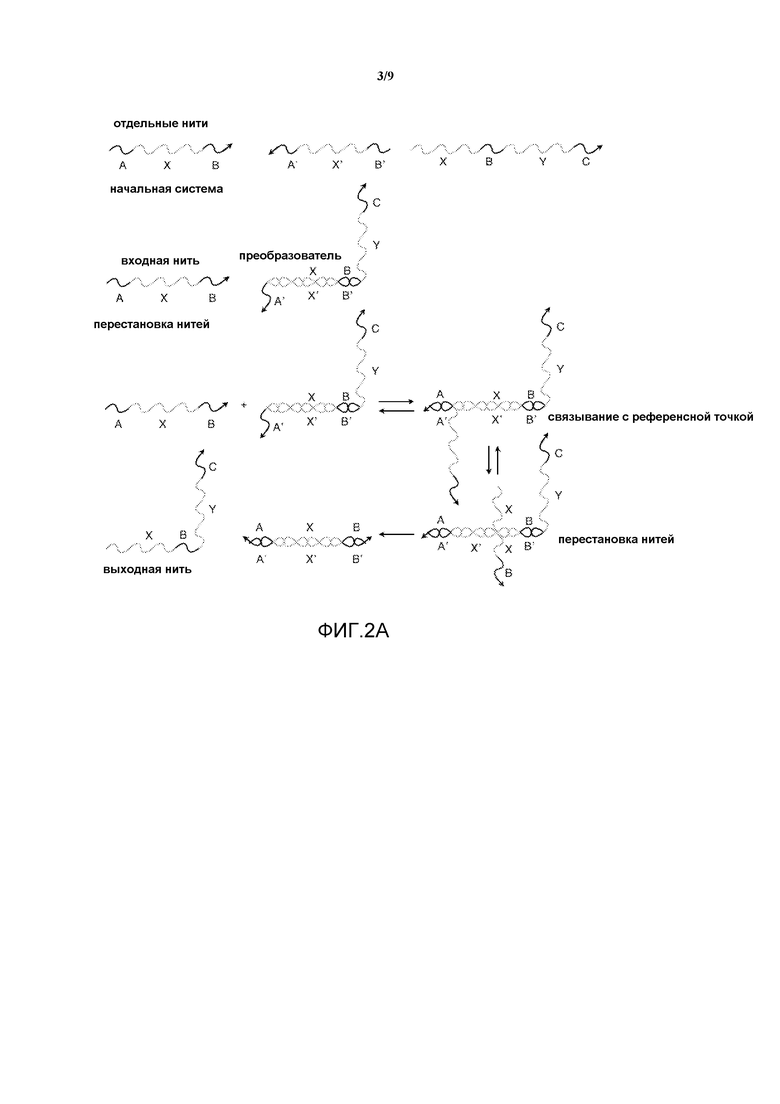
Фигура 2A иллюстрирует реализацию преобразователя нуклеиновых кислот с "точкой опоры". Снова отдельные секции представляют собой фрагменты олигонуклеотидов произвольной длины и последовательности.
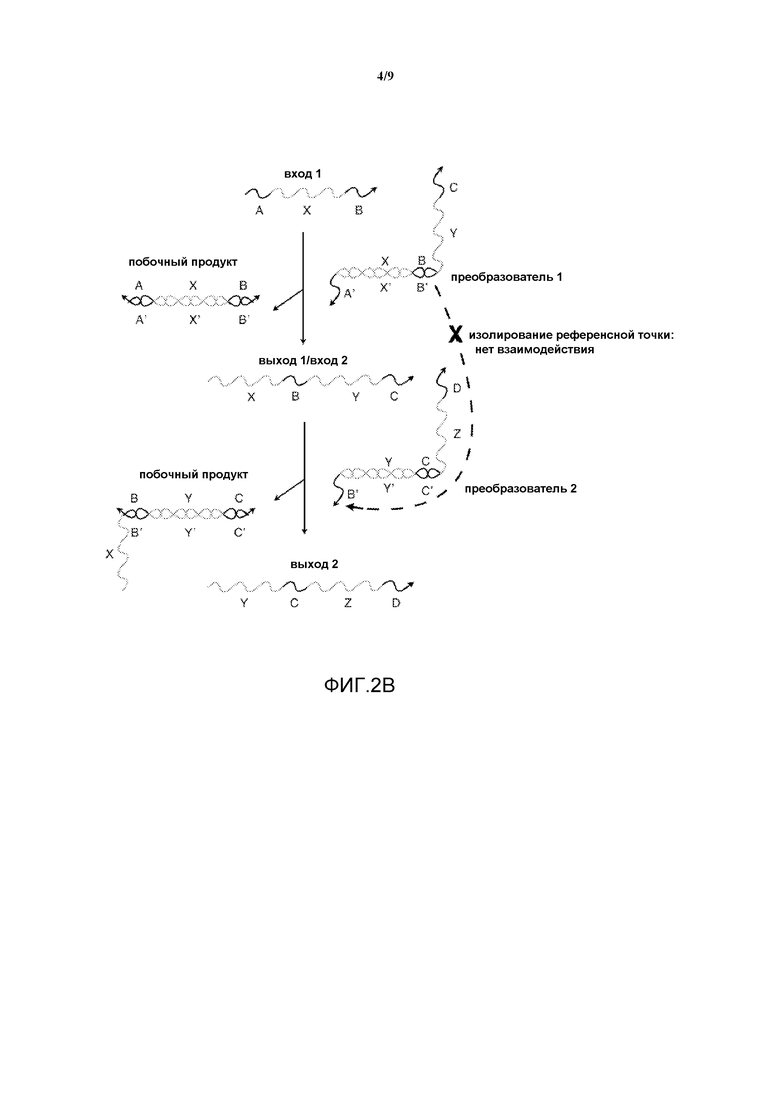
Фигура 2B иллюстрирует, каким образом реализация преобразователя нуклеиновых кислот с "точкой опоры" изолирует фрагменты олигонуклеотидов.
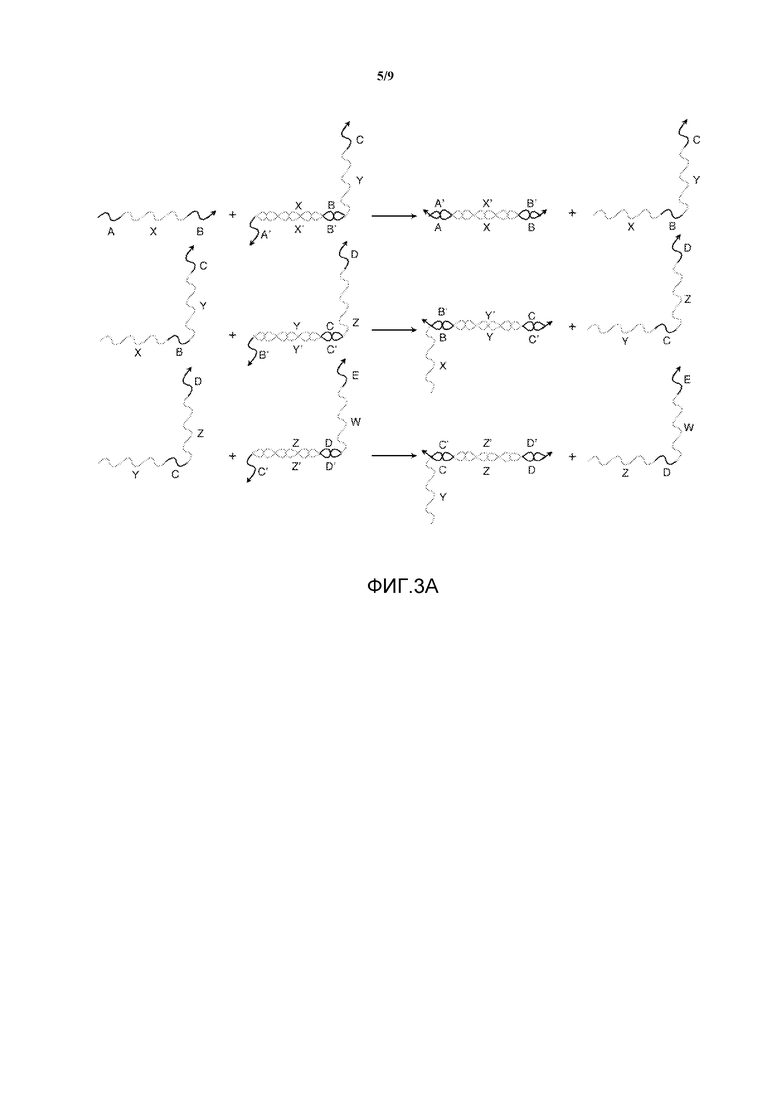
На фигуре 3A показана система с тремя преобразователями нуклеиновых кислот с точкой опоры. Все показанные реакции представляют собой реакции перестановки нити, которые проходят посредством такого же механизма миграции ветви, как на фигуре 2A.
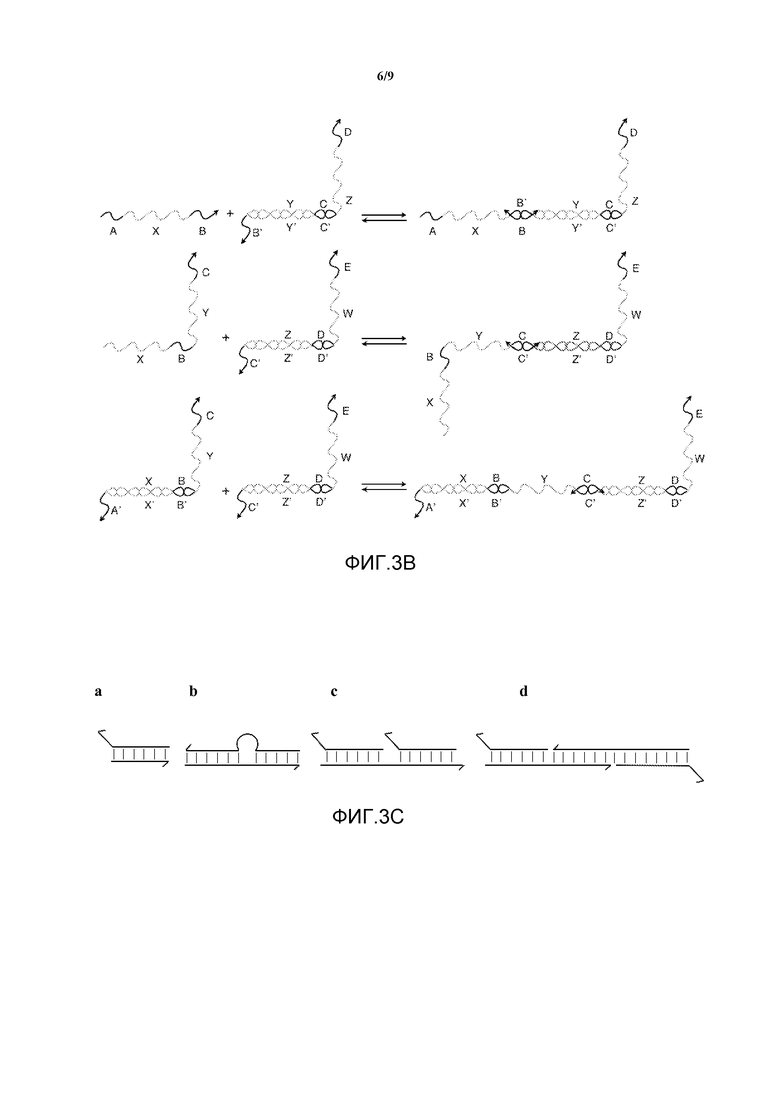
На фигуре 3B показана система с теми же олигонуклеотидами, что показаны на фигуре 3A, но вместо реакций перестановки нити показаны "столкновения" точек опоры, при этом референсная точка связана последовательностью, которая не может вызвать реакцию перестановки нити. Данное событие связывания занимает точку опоры, в результате чего желаемая нить не может связаться.
Фигура 3C иллюстрирует различные архитектуры для структур из нуклеиновой кислоты с не полностью спаренными основаниями, включающие в себя: (a) конец; (b) внутреннюю петлю; и (c) и (d) комплекс из множества частей.
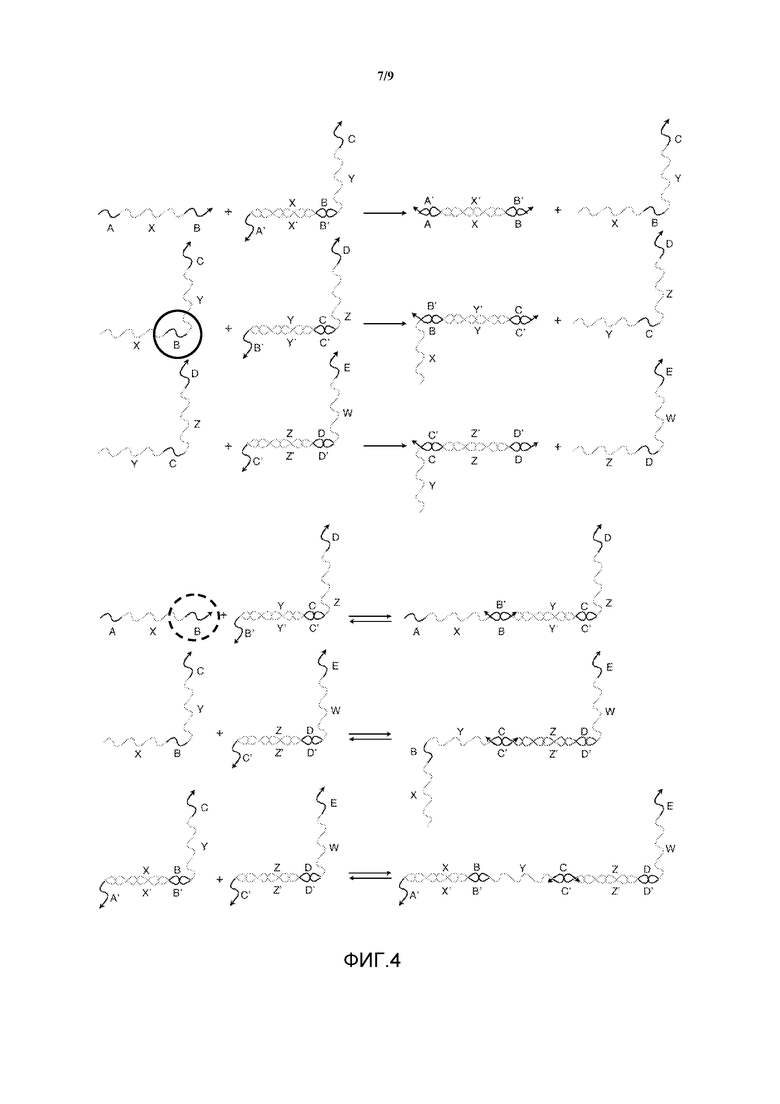
Фигура 4 представляет собой схематическое изображение того, каким образом в системе преобразователей нуклеиновых кислот, показанной на фигуре 3A, равновесие может быть сдвинуто в пользу реакций перестановки нити относительно столкновения точек опоры посредством замены фрагментов олигонуклеотидов химическими модификациями, которые повышают связывающую способность, и/или замены фрагментов олигонуклеотидов модификациями, которые понижают связывающую способность.
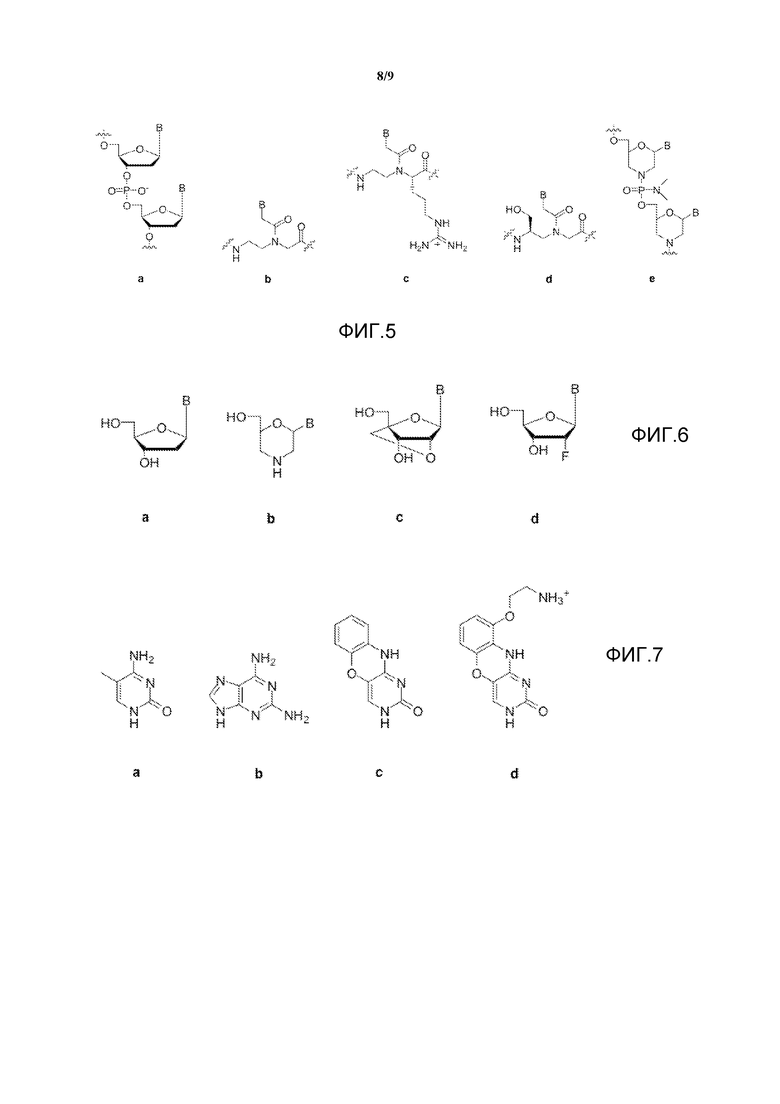
На фигуре 5 показаны модифицированные структуры остова для аналогов нуклеиновых кислот, при этом "B" представляет собой произвольное нуклеиновое основание и (a) показывает природный фосфодиэфирный остов ДНК, (b) показывает пептидные нуклеиновые кислоты, (c) показывает гуанидиновые пептидные нуклеиновые кислоты, (d) показывает L-сериновые производные гамма-ПНК, и (e) показывает фосфородиамидаты (здесь - с морфолиновым сахаром).
Фигура 6 иллюстрирует модифицированные структуры сахаров для аналогов нуклеиновых кислот, при этом "B" представляет собой произвольное нуклеиновое основание, и (a) показывает природный дезоксирибозный сахар РНК, (b) показывает морфолины, (c) показывает запертые нуклеиновые кислоты и (d) показывает модифицированную фтором производную РНК.
Фигура 7 иллюстрирует модифицированные структуры для аналогов нуклеиновых кислот метилцитозина (a), диаминопурина (b), феноксазина (c) и G-clamp (d).
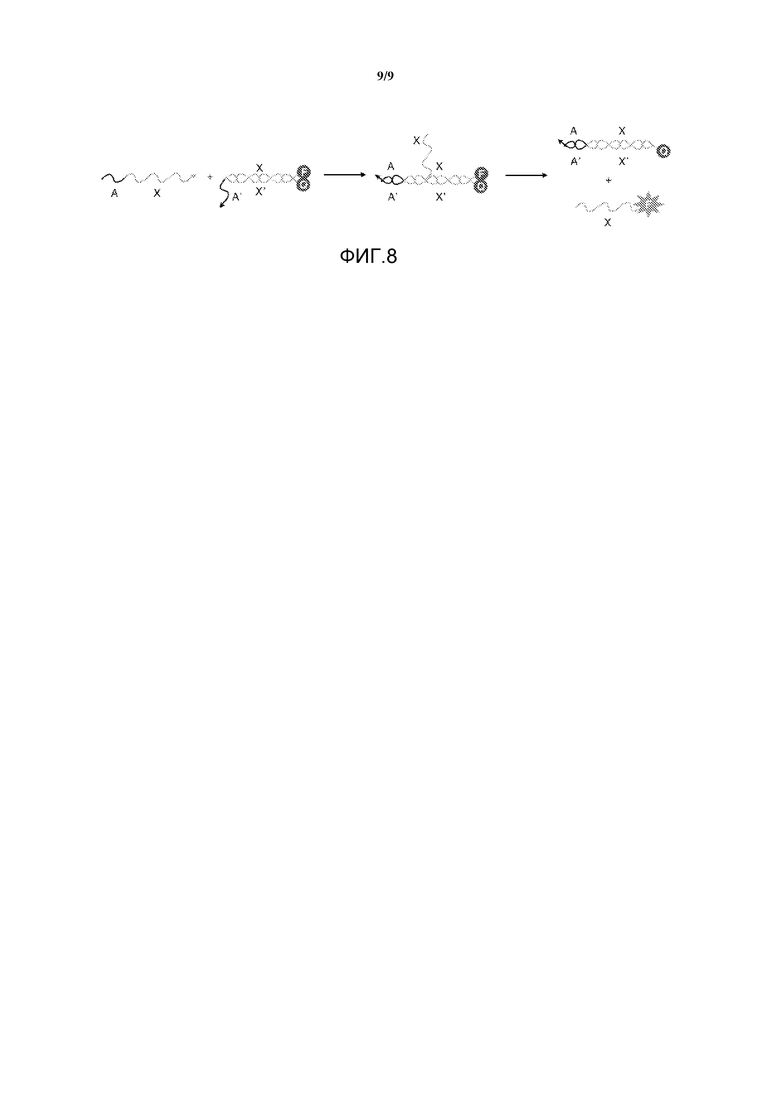
Фигура 8 иллюстрирует флуоресцентный кинетический анализ для демонстрации изменения скорости реакции вследствие химической модификации по изобретению.
ПОДРОБНОЕ ОПИСАНИЕ
Различные подходы, такие как твердофазное изолирование, изолирование референсной точки и обмен референсными точками, могут быть использованы в преобразовании последовательностей нуклеиновых кислот с целью построения логических операторов и сетей. Данные три подхода, подробнее описанные ниже, поясняются через геометрические элементы, в которых используются опосредованные референсными точками реакции миграции ветви в трех направлениях. Однако для реакций миграции ветви также возможны дополнительные механизмы, включая миграцию ветви в четырех направлениях, ускоренную миграцию в четырех направлениях и комплексную миграцию множества нитей, но не ограничиваясь перечисленным.
Таким образом, несмотря на то, что приведенные ниже варианты осуществления описываются, в целях иллюстрации, с использованием миграции ветви в трех направлениях, настоящим изобретением также предусматриваются логические элементы и сети ДНК, построенные с использованием других способов миграции ветви. И наоборот, варианты осуществления изобретения могут быть применены к произвольной реакции миграции ветви.
В твердофазном изолировании производится физическое разделение соответствующих последовательностей/нитей в пространстве с использованием для этой цели гранул, наночастиц или поверхностей. В данном подходе используются принципы местной изоляции, которые широко использовались в области органической химии. В твердофазных изолированных геометрических элементах появление событий перестановки может контролироваться путем регулирования того, находятся ли необходимые нити в растворе или твердой фазе системы.
На фигуре 1A показана базовая для твердофазного изолирования настройка преобразователя - компонента, который позволяет системе заменять одну нуклеиновую кислоту на другую. Здесь нить A'-X' (где A и X представляют собой фрагменты олигонуклеотидов произвольной длины и последовательности, и X' и A' представляют собой соответствующие им обратно комплементарные фрагменты) связана с твердой подложкой и первоначально гибридизуется в Y-B-X, образуя структуру из нуклеиновой кислоты в форме дуплекса с неполным спариванием, который может функционировать в качестве преобразователя. В данной конфигурации нить Y-B-X подвергается твердофазному изолированию и не может взаимодействовать с остальными элементами системы. Однако в присутствии нити X-A, называемой "полинуклеотидная молекула перестановки", нить Y-B-X может быть перемещена из твердой подложки и помещена в фазу раствора системы, при этом нить X-A связана с подложкой. Данная операция включает в себя два этапа, первый из которых представляет собой гибридизацию комплементарных последовательностей A и A' (часто называемую связыванием в референсной точке). На втором этапе область X нити X-A связывается с областью X' нити A'-X', переставляя область X нити Y-B-X и высвобождая данную нить в раствор, при этом оставляя X-A связанной с твердой подложкой (данный этап часто называется реакцией миграции ветви). Данный двухэтапный процесс позволяет осуществлять эффективное преобразование свободной нити X-A в свободную нить Y-B-X.
На фигуре 1B показана система, имеющая входную нить X-A, взаимодействующую с дуплексом с неполным спариванием, "преобразователем 1". Выход включает в себя дуплекс с полным спариванием, A'-X'/X-A, который считается побочным продуктом, и выходную нить Y-B-X, которая обозначается как "выход 1" и может быть использована в качестве "входа 2" при продолжении реакции. "Вход 2" взаимодействует с "преобразователем 2", в результате чего образуется "выход 2" и другой побочный продукт. На данной фигуре область B нити Y-B-X иллюстрирует изолирование последовательностей в данной сети. В начале Y-B-X не может гибридизироваться с областью B' преобразователя 2, поскольку они изолированы на отдельных твердых подложках. После того, как вход 1 связывается с преобразователем 1 и высвобождает Y-B-X в раствор, Y-B-X может взаимодействовать с преобразователем 2. Таким образом, способность Y-B-X и преобразователя 2 к взаимодействию зависит от присутствия входа 1.
Нити, связанные с одной твердой поверхностью, очень медленно взаимодействуют с нитями на другой твердой поверхности из-за стерических эффектов. Следовательно, нити в фазе раствора являются единственными компонентами, которые могут взаимодействовать с операторами твердой фазы.
Изолирование референсной точки и обмен референсными точками представляют собой разные подходы, в которых используются сходные взаимодействия образования пар, но с различной геометрией. В обоих подходах могут выполняться одни и те же операции твердофазных преобразователей, но они функционируют путем поддержания связи фрагментов последовательности в дуплексе. Так же, как и в твердофазном преобразователе, событие перестановки может высвобождать представляющую интерес последовательность. Для обеих геометрий референсной точки все нити могут находиться в растворе вместе, вследствие чего события перестановки регулируются доступностью референсных точек, то есть, коротких фрагментов однонитевых последовательностей нуклеиновых кислот, которые предоставляют начальную точку для события перестановки.
На фигуре 2A показан преобразователь с изолированием референсной точки, аналогичный преобразователю с фигуры 1A, но основанный на изолировании референсной точки, а не на твердофазном изолировании. В данном примере область A' преобразователя представляет собой референсную точку, которая связывает входную нить и позволяет выполнить реакцию перестановки нити.
На фигуре 2B показана основанная на референсной точке система, имеющая входную нить A-X-B, взаимодействующую с дуплексом с неполным спариванием оснований, "преобразователем 1". Выход включает в себя побочный продукт, то есть, дуплекс с полным спариванием оснований, B'-X'-A'/A-X-B, и "выход 1" - нить X-B-Y-C, которая может быть использована как "вход 2" при продолжении реакции. "Вход 2" взаимодействует с "преобразователем 2", в результате чего образуется "выход 2" и другой побочный продукт. На данной фигуре область B нити X-B-Y-C изолируется в преобразователе 1 посредством гибридизации с комплементарной областью B', и, следовательно, не может взаимодействовать с областью B' преобразователя 2. Способность X-B-Y-C взаимодействовать с преобразователем 2 зависит от присутствия входа 1 (A-X-B) в системе.
Геометрические элементы референсной точки потенциально являются очень полезными, но их использование до настоящего времени было ограничено скоростью, с которой системы, содержащая такие геометрические элементы референсной точки, могут распространять информацию. Это происходит из-за неотъемлемых ограничений, присутствующих в имеющихся подходах с изолированием референсной точки, которые замедляют их до биологически нецелесообразных сроков. Данные кинетические "узкие места" являются результатом непродуктивных реакций, называемых здесь "столкновения референсных точек", которые происходят, когда референсная точка связывается молекулой, имеющей комплементарную последовательность или "нить столкновения", которая не может произвести реакцию перестановки.
На фигуре 3A показана система из трех преобразователей нуклеиновых кислот с изолированием референсной точки, аналогичных преобразователю на фигуре 2A. Однако если все три нити находятся в растворе вместе, то других событий связывания происходить не может. Фигура 3B иллюстрирует некоторые непродуктивные события связывания или столкновения, которые могут иметь место. За счет использования "полинуклеотидной молекулы столкновения" данные события не вызывают реакцию перестановки, но могут замедлить работу системы, поскольку воздействие нити столкновения блокирует связывание нитей, которое может привести к реакции перестановки.
Референсные точки могут быть короткими в целях снижения влияния данных столкновений на систему: чем короче референсная точка, тем больше может быть скорость включения/выключения комплементарной последовательности. Таким образом, обычно референсные точки имеют длину пять или шесть нуклеотидов, поскольку если при таких длинах происходит событие непродуктивного связывания, то время, проведенное в двухнитевом состоянии ("столкновении"), небольшое. Однако в данном подходе имеется упомянутое выше "узкое место", поскольку событие продуктивного связывания ограничивается теми же термодинамическими параметрами; следовательно, вероятно, что входящая нить не будет сильно связана с этими референсными точками. В результате, желаемая перестановка не всегда будет происходить при связывании правильной входящей нити, поскольку она должна находиться в связанном состоянии достаточно долго для инициирования реакции перестановки. Таким образом, использование коротких референсных точек увеличивает временной интервал, требующийся для проведения заданной операции и выдачи выхода. Другими словами, реакция перестановки не может начаться до того, как произойдет множество событий связывания, как с нитями столкновения, так и с желаемыми нитями. Данная неэффективность ограничивает полезность системы путем замедления распространения информации до временных интервалов, которые являются нецелесообразно длинными.
В соответствии с одним из аспектов изобретения, химическая модификация структуры из нуклеиновой кислоты и/или полинуклеотидной молекулы перестановки способствует продуктивности взаимодействий. Вследствие этого она является целесообразной при построении цепи из отдельных логических элементов с формированием сетей произвольного размера для биологических приложений. Конкретнее, представлен подход для определения того, каким образом не способствовать взаимодействиям столкновения и способствовать продуктивным взаимодействиям без изменения информационного содержания последовательностей.
В соответствии с изобретением, данные подходы улучшают термодинамику связывания для желаемой нити и/или ухудшают термодинамику для нитей столкновений, при этом не изменяя информационное содержание последовательности. В частности, при использовании химически модифицированных структур свободная энергия Гиббса (ΔG) для желаемой реакции гибридизации между заданным олигонуклеотидом и комплементарной ему ДНК или РНК снижается и/или ΔG взаимодействия столкновения повышается. На молекулярном уровне это соответствует более сильному связыванию (более высокой связывающей способности) для желаемой реакции гибридизации и менее сильному связыванию (менее высокой связывающей способности) для нитей столкновения, соответственно, что является результатом модификации структур.
В первом случае происходит сдвиг равновесия для желаемых событий связывания от референсной точки в направлении дуплексного (связанного) состояния, что увеличивает вероятность возникновения реакции перестановки, при этом не влияя на равновесие связывания между референсной точкой и нитями столкновения. В последнем случае ухудшаются условия для связывания нити(-ей) столкновения, делая предпочтительным для данных взаимодействий несвязанное состояние. Эти два изменения могут быть использованы совместно или по отдельности в целях способствования связыванию желаемой нити или противодействия связыванию нити(-ей) столкновения.
В вариантах осуществления в соответствии с настоящим изобретением используется преимущество того факта, что скорость реакции для любой опосредованной референсной точкой перестановки нити связана с термодинамическим предпочтением двух связываемых нитей или комплексов нуклеиновых кислот. За счет того, что данные термодинамические условия делаются более предпочтительными для событий связывания без столкновения и/или менее предпочтительными для событий связывания со столкновением, можно направить систему в сторону продуктивных реакций перестановки. Это увеличит скорость, с которой распространяется информация, до такого значения, которое сделает временные интервалы биологически целесообразными. В подходе по настоящему изобретению, описанному здесь, это достигается через химические модификации фрагментов олигонуклеотидов в системе.
Посредством такой химической модификации структуры из нуклеиновой кислоты и/или полинуклеотидной молекулы, в соответствии с изобретением, расширяется диапазон эффективной концентрации, в котором реакция является оптимальной. Это происходит потому, что процент нуклеиновой кислоты в дуплексе при заданной температуре является функцией концентрации и ΔG реакции гибридизации. Таким образом, чем более концентрированным является множество комплементарных нуклеотидов и ниже (предпочтительнее) ΔG, тем выше процент образования дуплексов. Следовательно, изменение ΔG для заданного взаимодействия изменяет процент дуплексов при заданной концентрации. Снижение ΔG (то есть, взаимодействие становится более благоприятным) означает, что процент дуплексов будет более высоким при заданной концентрации, тогда как повышение ΔG (взаимодействие становится менее благоприятным) означает, что процент дуплексов будет менее высоким при заданной концентрации. Данный эффект расширяет диапазон оптимальных концентраций, при которых данные реакции могут быть использованы на практике, поскольку желаемые дуплексы могут быть сформированы при более низких концентрациях, и нежелательные дуплексы не будут сформированы при более высоких концентрациях относительно референсного множества немодифицированных олигонуклеотидов.
Данные подходы применимы к любым геометриям реакции, опосредованной референсной точкой, включая реакции миграции ветви в трех направлениях, обсуждаемые выше, но не ограничиваясь ими. Могут быть использованы различные архитектуры для дуплекса с неполным спариванием оснований, в которых разветвление или петли размещаются в различных точках структуры (см., например, фигуру 3C). Ориентация ветвей или петель не определяется направленностью нити. Таким образом, данные структуры могут включать в себя (a) концевую структуру, (b) внутреннюю петлю или (c) и (d) комплекс из множества частей, или любые другие возможные архитектуры для дуплекса с неполным спариванием оснований.
Данные структуры из нуклеиновой кислоты могут преобразовывать активную "входную" последовательность (полинуклеотид перестановки) в активную "выходную" последовательность (высвобождаемый полинуклеотид).
Структуры из нуклеиновой кислоты, называемые выше "преобразователями", могут находиться в своих соответствующих термодинамических минимумах, то есть, они представляют собой наиболее стабильные структуры, которые могут быть сформированы конкретным множеством последовательностей нуклеиновых кислот. Данные структуры могут быть сформированы путем отжига двух или более отдельных нитей нуклеиновых кислот. Например, все нити могут быть смешаны вместе, нагреты до температуры, значительно превышающей точку плавления для любой структуры, которая может быть сформирована, и затем медленно охлаждены. Это позволяет осуществить гибридизацию нитей в энергетическом состоянии с наименьшей возможной энергией (термодинамическом минимуме).
Данная процедура может быть одинаковой для природных нуклеиновых кислот, таких как ДНК и РНК, нуклеиновых кислот с модифицированными остовами, сахарами или основаниями, для химер, сделанных из природных и модифицированных нуклеиновых кислот.
Избирательное введение специфических химических модификаций может способствовать связыванию нитей, способных к продуктивным реакциям перестановки, относительно событий связывания со столкновениями референсных точек. Например, на фигуре 4 показаны те же нити, что и на фигурах 3A и 3B. Однако если определенные фрагменты нуклеиновых кислот замещаются химически модифицированными аналогами, то в результате улучшается термодинамика связывания, и кинетика для столкновений сдвигается в сторону реакций перестановки. Например, если фрагмент "B" олигонуклеотидов в нити X-B-Y-C (обведены окружностью из сплошной линии) имеет более высокую связывающую способность для фрагмента B', чем для фрагмента "B" олигонуклеотидов в нити A-X-B, то равновесие смещается в пользу продуктивной реакции перестановки нити, что повышает вероятность или частоту, с которой возникает данная реакция. Оба фрагмента B' имеют одну и ту же последовательность спаривания оснований по Уотсону-Крику, но взаимодействие между X-B-Y-C и референсной точкой B' является более предпочтительным из-за химических модификаций.
Аналогично, если определенные фрагменты нуклеиновых кислот заменяются аналогами, которые не благоприятствуют связыванию, то равновесие также сместится от столкновений. Например, если "B" в нити A-X-B (обведен окружностью из пунктирной линии) имеет более низкое связывание для B', чем "B" в X-B-Y-C, то равновесие сместится от столкновения референсных точек, улучшая кинетику для реакции перестановки нитей.
Оба подхода увеличивают скорость, с которой сеть нуклеиновых кислот может обрабатывать сигналы, поскольку в обоих подходах возрастает время нахождения желаемой нити на референсной точке относительно времени нахождения нити или нитей столкновения. Это увеличивает вероятность того, что реакция перестановки нити произойдет в заданный период времени и, таким образом, повысится скорость, с которой сеть оценивает заданный вход или множество входов. Данные модификации ранее не использовались для способствования или противодействия реакциям перестановки, для построения значимых логических элементов или сетей ДНК, или для распространения информации.
Имеется множество хорошо охарактеризованных модификаций нуклеиновых кислот, которые могут быть использованы в соответствии с вариантами осуществления изобретения для улучшения или ухудшения термодинамических свойств связывания с природной ДНК или РНК. Они включают в себя изменения в остове, сахаре или нуклеиновом основании олигонуклеотида. Данные модификации также могут быть использованы по отдельности или совместно друг с другом; то есть, использование модифицированного остова не исключает использования модифицированного нуклеинового основания на той же нити.
Аналоги остовов нуклеиновых кислот могут быть использованы для улучшения связывания нитей, способных к выполнению реакции перестановки. Имеется ряд различных аналогов, которые могут быть использованы, при этом все они обеспечивают большую связывающую способность с ДНК и РНК, чем природные нуклеиновые кислоты. Данные аналоги включают в себя, но не ограничиваются перечисленным, незаряженные остовы (пептидные нуклеиновые кислоты или фосфородиамидаты), положительно заряженные остовы (гуанидиниевые пептидные нуклеиновые кислоты) и группы образования водородной связи, которые предусматривают предварительную организацию (гамма-пептидные нуклеиновые кислоты).
Общие структуры для определенных аналогов показаны на фигуре 5. Все эти аналоги улучшают термодинамику реакций гибридизации нуклеиновых кислот, что повышает степень связывания с референсной точкой и, следовательно, повышает скорость перестановки. Посредством использования этих аналогов в конкретных местах логической сети желаемые реакции перестановки могут получить большое преимущество по сравнению с взаимодействиями столкновения, в результате чего увеличивается скорость обработки информации.
Использование модифицированных сахарных колец также может изменить термодинамику связывания с ДНК или РНК для олигонуклеотида. Чаще всего используемыми аналогами являются морфолино, запертые нуклеиновые кислоты (LNA) и производные LNA. В литературе также документированы другие модифицированные сахара, которые могут привести к аналогичному результату относительно изменения термодинамики связывания. Иллюстрацией таких сахаров являются модификации в 2', 3' или 4'-положении и сахара с различными атомами, замененными на кислород в циклопентановом кольце рибозы. Данные аналоги проиллюстрированы на фигуре 6.
Модификации нуклеиновых оснований также могут быть использованы для достижения того же результата, что и при использовании аналогов остова и сахаров; а именно для изменения термодинамики конкретных реакций гибридизации. Такие основания включают в себя метилцитозин, диаминопурин, G-clamp и феноксазин (фигура 7), при этом все они повышают связывающую способность нити для обратно комплементарной ей нити. Другая возможность модификации нуклеиновых оснований относится к псевдокомплементарным основаниям. Данный класс аналогов оснований формирует слабые пары оснований друг с другом, но формирует сильные пары оснований со стандартными основаниями. Одной из таких пар оснований является 2-аминоаденин (nA) и 2-тиотимин (sT). Данные основания могут быть использованы для способствования связыванию одной нити и противодействия связыванию другой, например, повышения вероятности события продуктивного связывания при снижении в то же время вероятности столкновения.
Другая химическая модификация, которая может быть использована для изменения термодинамики связывающих взаимодействий, состоит во включении заряженных полимеров, таких как хитозан, который, как было показано в литературе, увеличивает скорость реакций перестановки. Однако поскольку данные полимеры ускоряют реакции неспецифично, то они должны были бы использоваться совместно с одной из других модификаций, упомянутых выше, для того, чтобы можно было различать желательные и нежелательные события связывания.
Различные подходы могут быть использованы для синтеза молекул с химическими модификациями, обсуждаемыми выше. Например, химические свойства остова могут учитываться при построении модифицированных молекул. Химические свойства остова - это то, что используется для соединения отдельных мономеров в более длинную нить. Модификации, затрагивающие нуклеиновое основание или сахар, но которые сохраняют природный диэфирный остов ДНК/РНК, могут быть синтезированы через стандартные фосфорамидатные химические реакции, которые используются для природных мономеров. Иллюстрации данных методов можно найти, например, в работах Beaucage, S., and R. Iyer, Tetrahedron 48: 2223 (1992); Brown, D. M. A, "Brief history of oligonucleotide synthesis", 20 METHODS IN MOLECULAR BIOLOGY (Protocols for Oligonucleotides and Analogs) 1-17 (1993); Reese, Colin B., Organic & Biomolecular Chemistry 3: 3851 (2005) и Iyer, R. P. and S.L. Beaucage, "7.05. Oligonucleotide synthesis," 7 COMPREHENSIVE NATURAL PRODUCTS CHEMISTRY (DNA and Aspects of Molecular Biology) 105-52 (1999), соответствующее содержание которых настоящим включено в данный документ посредством ссылки во всей их полноте.
Если остов изменяется в некоторой модификации, то могут быть использованы другие химические реакции. Такие химические реакции модификации общеизвестны в научной литературе. Таким образом, пептидные нуклеиновые кислоты (ПНК) и их производные опираются на амидные связи для соединения отдельных мономеров. Следовательно, вместо использования фосфорамидатных химических реакций, нити данных мономеров создаются с помощью условий формирования амидных связей и связующих реагентов, таких как HBTU. Исследование способов, использованных для создания ПНК или ПНК-подобных олигонуклеотидов можно найти, например, в работе F. Beck, "Solid Phase Synthesis of RNA Oligomers", METHODS IN MOLECULAR BIOLOGY SERIES (Peptide Nucleic Acids), Humana Press, http://www.springerlink.com/content/mr571738x7t65067/.
Другой подход к модификации остова включает в себя использование химерных олигонуклеотидов. Они представляют собой нити олигонуклеотидов, которые содержат различные химические вещества в одной и той же молекуле. Например, если требуется получить нить, которая наполовину состоит из остова ПНК и наполовину из остова ДНК, то нужно будет некоторым образом соединить эти два химических вещества. Создание таких химерных нитей также общеизвестно в технике. В приведенном выше примере химеры ПНК/ДНК различие в химических свойствах может быть преодолено путем использования модифицированных мономеров ДНК или ПНК. В ДНК 5'-диметокситритил (DMT)-защищенная гидроксильная группа заменяется на монометокситритил (MMT)-защищенный амин, который может реагировать с карбоновой кислотой ПНК после снятия защиты. В ПНК защищенный N-концевой азот заменяется на DMT-защищенную гидроксильную группу, которая может реагировать с фосфорамидатной группой на ДНК после снятия защиты. Данные подходы подробнее описаны, например, в работе E. Uhlmann, et al., Angew. Chem. (Int'l ed.) 37: 2796-823 (1998).
Все эти модификации, применяемые по отдельности или совместно друг с другом, могут повлиять на термодинамические условия конкретных взаимодействий в произвольной сети нуклеиновых кислот таким образом, что связывание желаемых нитей или комплексов получает предпочтение над взаимодействиями столкновения без изменения содержания последовательности. Все эти взаимодействия могут применяться к произвольным опосредованным ветвью реакциям миграции, независимо от того, являются ли они миграциями ветви в трех направлениях, таким как твердофазное изолирование, изолирование референсной точки или обмен референсными точками, или миграции ветви происходят посредством других механизмов, например, миграции ветви в четырех направлениях, ускоренной миграции ветви в четырех направлениях или комплексной многонитевой миграцией.
В соответствии с некоторыми вариантами осуществления, кинетика химически модифицированных молекул для улучшенной реакции миграции ветви может быть протестирована. Один из подходов состоит в измерении улучшения, которое обеспечивают различные модификации относительно ускорения или замедления реакции миграции ветви по сравнению с природным полинуклеотидом, с применением флуоресцентного анализа кинетики (см., например, фигуру 8). Здесь структура преобразователя имеет гаситель на одной нити и флуорофор на другой. При отсутствии нити перестановки флуоресценция флуорофора гасится, что дает фоновые характеристики реакции. В случае, когда молекула перестановки, которая могла быть изменена в области референсной точки и/или не в области референсной точки, добавляется к преобразователю, она заменит нить с гасителем и в процессе этого выдаст флуоресцентный сигнал. Таким образом, скорость "включения" флуоресценции в системе является показателем кинетики реакции перестановки. Данный анализ может быть усложнен посредством введения требования по увеличению цикла оценки до окончательной выдающей флуоресценцию реакции перестановки или до того, как нити столкновения смогут быть добавлены для конкуренции с нитями перестановки за референсную точку.
Подходя в соответствии с изобретением также могут применяться к произвольным молекулам, природным или искусственным, которые подходят для распространения информации, при условии, что имеется первая молекула и вторая молекула, при этом последняя сконфигурирована для связи с первой. По меньшей мере, одна молекула из первой молекулы и второй молекулы, соответственно, имеет химическую модификацию относительно референсной молекулы, которая имеет тот же состав последовательности, что и первая молекула или вторая молекула, в зависимости от конкретной ситуации. Модификация приводит к тому, что связь имеет свободную энергию гибридизации, отличающуюся от энергии референсного связывания между референсной молекулой и первой или второй молекулой, в результате чего в системе реализуется, по меньшей мере, одно из следующего: вероятность продуктивного связывания повышается; или вероятность непродуктивного связывания снижается.
Как указано выше, в соответствии с изобретением многоэтапный процесс может быть применен для получения улучшенной сети нуклеиновых кислот. Это повлечет за собой исследование полной сети рассматриваемых преобразователей нуклеиновых кислот, включая любые взаимодействия столкновения, и последующую идентификацию фрагментов последовательности, которые должны быть модифицированы в целях наилучшего способствования событиям перестановки и минимизации столкновений.
В отношении произвольной реакции перестановки нитей, данные модификации могут быть выполнены для желаемой молекулы перестановки, для молекулы столкновения, или для обеих молекул. Это возможно потому, что для любого преобразователя или логического элемента с доступной референсной точкой будет иметь место конкуренция между желаемой молекулой перестановки и произвольной молекулой столкновения за связывание с референсной точкой. Кроме того, улучшение способности молекулы перестановки связываться с референсной точкой относительно молекулы столкновения повысит скорость, с которой выполняется реакция перестановки. Таким образом, улучшение способности молекулы перестановки связываться (снижение ΔG связывания) или ухудшение способности молекул(-ы) столкновения связываться (увеличение ΔG связывания) приведет к повышению скорости реакции. Поскольку оба этих изменения ΔG могут быть достигнуты через химические модификации, в соответствии с изобретением, то при рассмотрении того, какие фрагменты последовательности будут модифицированы, важно исследовать возможность изменения только молекулы перестановки, только молекулы столкновения, или обеих молекул вместе.
В соответствии с вариантами осуществления изобретения два параметра, которые могут изменяться в данном анализе, это (i) константы скорости для взаимодействия между любыми двумя олигонуклеотидами и (ii) длина референсных точек для всех олигонуклеотидов в сети. Первый параметр будет определяться химическим составом олигонуклеотида и ΔG его взаимодействий с другими олигонуклеотидами. Последний параметр будет представлять собой количество нуклеотидов в области референсной точки рассматриваемой последовательности.
Константы кинетической скорости, требующиеся для моделирования сети нуклеиновых кислот, могут быть вычислены по ΔG гибридизации между любыми двумя олигонуклеотидами. Для природных оснований и многих их модификаций данные значения, которые получаются на основании параметров ближайшего соседа, были описаны в литературе. Если они еще не известны, то они могут быть определены экспериментально через анализ Вант-Гоффа для данных температуры плавления, дифференциальную сканирующую калориметрию или изотермическую титрационную калориметрию. Каждый из указанных способов позволит определить термодинамические параметры для исследуемого взаимодействия нуклеиновых кислот, которые затем могут быть использованы для определения ΔG реакции при произвольной температуре. Затем значение ΔG может быть использовано для определения константы равновесия (Keq) взаимодействия. Константа равновесия будет пропорциональна отношению констант скорости; следовательно, можно использовать известные константы для нахождения нужной константы кинетической скорости для заданной реакции.
Таким образом, полное множество констант скорости может быть получено для рассматриваемых модификаций для использования в логической сети нуклеиновых кислот, совместно с длинами всех референсных точек. При наличии данной информации систему можно смоделировать для определения поведения сети с течением времени, и также для определения того, где должны быть сделаны конкретные модификации для оптимизации продуктивных реакций перестановки относительно взаимодействий столкновения. Дополнительно в модели может изменяться длина конкретных областей референсных точек. Сеть может быть оптимизирована путем изменения модификаций в конкретных положениях и путем моделирования реакций в сети с различными длинами референсных точек. Данные вычисления могут быть использованы для определения оптимальной модификации и длины референсной точки для каждого компонента, которые обеспечивают наиболее предпочтительное поведение сети с течением времени.
Сеть также может быть оптимизирована посредством экспериментального изменения модификаций и длин референсных точек и последующего исследования поведения с течением времени в лаборатории, в отличие от моделирования поведения с помощью математической модели.
Если имеется полностью оптимизированная сеть, то необходимые олигонуклеотиды могут быть синтезированы с помощью стандартной фосфорамидитной химии или с помощью подхода(-ов), описанных в литературе для конкретных модификаций. Любая структура преобразователей или элементов, которая состоит из более чем одного олигонуклеотида, может быть сконструирована путем смешивания отдельных олигонуклеотидов и последующего отжига, путем первоначального нагревания смеси до температуры выше точки плавления всех дуплексов и последующего медленного ее охлаждения. Это приведет к гибридизации олигонуклеотидов с образованием наиболее стабильной структуры, которую может принять конкретное множество последовательностей нуклеиновых кислот. Данные структуры могут быть очищены до того, как они будут использоваться в сети. После того, как будут спроектированы, синтезированы и подвергнуты отжигу все преобразователи, элементы и другие компоненты, сеть может быть применена в целевой аналитической, диагностической или биологической системе.
Несмотря на то, что были обсуждены конкретные варианты осуществления рассматриваемого изобретения, они являются только иллюстративными и не ограничивают изобретение. В результате изучения данной спецификации специалистам в данной области техники станет понятно множество вариаций изобретения. Полный объем изобретения должен определяться со ссылкой на приведенные ниже пункты формулы изобретения, а также на полный диапазон их эквивалентов, и на спецификацию, с учетом таких вариаций.
ПРОЦИТИРОВАННЫЕ ПУБЛИКАЦИИ
Приведенные ниже публикации могут помочь в понимании или практическом применении вариантов осуществления настоящего изобретения. Каждая процитированная публикация включена в настоящий документ посредством ссылки во всей своей полноте.
(1) Picuri, J. M.; Frezza, B. M.; Ghadiri, M. R. J Am Chem Soc 2009, 131, 9368-77.
(2) Li, Q.; Luan, G.; Guo, Q.; Liang, J. Nucleic Acids Res 2002, 30, E5.
(3) Zhang, D. Y.; Turberfield, A. J.; Yurke, B.; Winfree, E. Science 2007, 318, 1 121-5.
(4) Frezza, B. M.; Cockroft, S. L.; Ghadiri, M. R. J Am Chem Soc 2007, 129, 14875-9.
(5) Voelcker, N. Н; Guckian, K. M.; Saghatelian, A.; Ghadiri, M. R. Small 2008, 4, 427-31.
(6) Yashin, R.; Rudchenko, S.; Stojanovic, M. N. J Am Chem Soc 2007, 129, 15581-4.
(7) Seelig, G.; Soloveichik, D.; Zhang, D. Y.; Winfree, E. Science 2006, 314, 1585-8.
(8) Zhang, D. Y.; Winfree, E. J Am Chem Soc 2009, 131, 17303-14.
(9) Biswas, I.; Yamamoto, A.; Hsieh, P. J Mol Biol 1998, 279, 795-806.
(10) Panyutin, I. G.; Hsieh, P. J Mol Biol 1993, 230, 413-24.
(11) Panyutin, I. G.; Hsieh, P. Proc Natl Acad Sci USA 1994, 91, 2021-5.
(12) Maugh, T. H., 2nd Science 1982, 217, 719-720.
(13) Tajima, T.; Nakajima, A. Journal of the American Chemical Society 2008, 130, 10496-7.
(14) Voit, B. Angew Chem Int Ed Engl 2006, 45, 4238-40.
(15) Kahan, M.; Gil, B.; Adar, R.; Shapiro, E. Physica D-Nonlinear Phenomena 2008, 237, 1165-1172.
(16) Nielsen, P. E.; Egholm, M. Curr Issues Mol Biol 1999, 1, 89-104.
(17) Dragulescu-Andrasi, A.; Zhou, P.; He, G.; Ly, D. H. Chem Commun (Camb) 2005, 244-6.
(18) Sahu, B.; Chenna, V.; Lathrop, K. L.; Thomas, S. M.; Zon, G.; Livak, K. J.; Ly, D. H. J Org Chem 2009, 74, 1509-16.
(19) Zhou, P.; Wang, M.; Du, L.; Fisher, G. W.; Waggoner, A.; Ly, D. H. J Am Chem Soc 2003, 125, 6878-9.
(20) Dragulescu-Andrasi, A.; Rapireddy, S.; Frezza, B. M.; Gayathri, C; Gil, R. R.; Ly, D. H. J Am Chem Soc 2006, 128, 10258-67.
(21) Freier, S. M.; Altmann, K. H. Nucleic Acids Research 1997, 25, 4429-4443.
(22) Braasch, D. A.; Corey, D. R. Chemistry & Biology 2001, 8, 1-7.
(23) Summerton, J.; Weller, D. Antisense & Nucleic Acid Drug Development 1997, 7, 187-195.
(24) Ortega, J. A.; Blas, J. R.; Orozco, M.; Grandas, A.; Pedroso, E.; Robles, J. Org Lett 2007, 9, 4503-6.
(25) Kutyavin, I. V.; Rhinehart, R. L.; Lukhtanov, E. A.; Gorn, V. V.; Meyer, R. B., Jr.; Gamper, H. B., Jr. Biochemistry 1996, 35, 11170-6.
(26) Lee, D.; Singha, K.; Jang, M. K.; Nah, J. W.; Park, I. K.; Kim, W. J. Mol Biosyst 2009, 5, 391-6.
(27) Патент США № 7538202.
(28) Публикация заявки на патент США № 2005/0112614.
(29) Публикация заявки на патент США № 2007/0072215.
(30) Публикация заявки на патент США № 2009/0191546.
Настоящее изобретение относится к области биоинформатики. Предложен способ для приготовления улучшенной вычислительной системы, основанной на нуклеиновых кислотах, включающий синтезирование в водном растворе варианта системы молекулярных вычислений, отличающегося включением химической модификации, изменяющей энергию гибридизации молекул нуклеиновых кислот в системе. Также рассмотрена вычислительная система, основанная на нуклеиновых кислотах и содержащая химические модификации в соответствии со способом по настоящему изобретению. Предложенное изобретение позволяет увеличивать вероятность желаемых событий связывания молекул нуклеиновых кислот в составе системы. 2 н. и 14 з.п. ф-лы, 8 ил.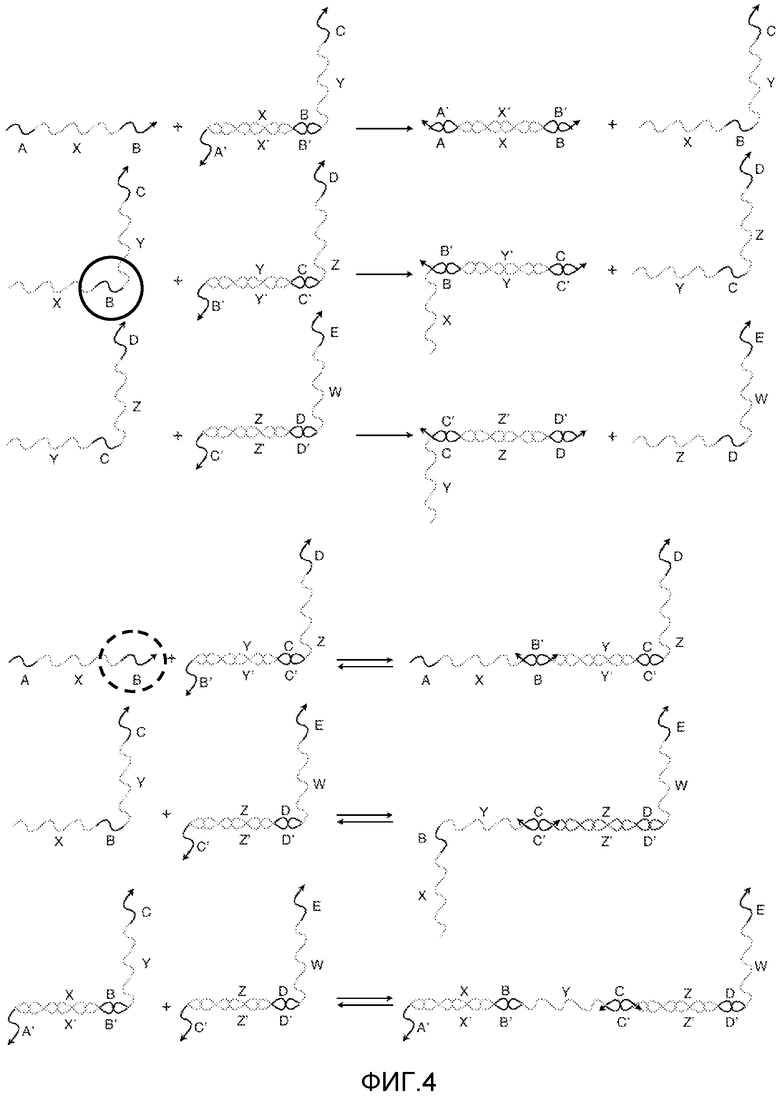
1. Способ приготовления улучшенной вычислительной системы, основанной на нуклеиновых кислотах, включающий
(1) идентификацию вычислительной системы, основанной на нуклеиновых кислотах, которая включает:
(i) входную нить нуклеиновой кислоты, включающую первый фрагмент (А) и второй фрагмент (В),
(ii) мишень, представляющую собой дуплекс нуклеиновой кислоты с неполным спариванием оснований, включающий первую и вторую нити,
где указанная первая нить включает первый фрагмент (В′) и второй фрагмент (А′), указанная вторая нить включает первый фрагмент (В) и второй фрагмент (С), а указанный дуплекс-мишень включает дуплексную область, образованную спариванием оснований между В и В′ и двумя однонитиевыми областями А′ и С, и
где указанная входная нить может связываться с первой нитью дуплекса-мишени при гибридизационных условиях, чтобы вызвать реакцию перестановки нити с получением выходного дуплекса, образованного (А-В) входной нити и первой нитью дуплекса-мишени (В′-А′), и
(iii) столкновение, представляющее собой дуплекс нуклеиновой кислоты с неполным спариванием оснований, включающий первую и вторую нити,
где указанная первая нить включает первый фрагмент (С') и второй фрагмент (В′), указанная вторая нить включает первый фрагмент (С) и второй фрагмент (D), а указанный дуплекс столкновения включает дуплексную область, образованную спариванием оснований между С и С' и двумя однонитиевыми областями В′ и D,
где вторая нить дуплекса-мишени может связываться с первой нитью дуплекса столкновения при гибридизационных условиях, чтобы вызвать реакцию перестановки нити с получением выходного дуплекса, образованного второй нитью дуплекса-мишени (В-С) и первой нитью дуплекса столкновения (С'-В′), и
где второй фрагмент (В) входной нити может связываться с однонитиевой областью В′ дуплекса столкновения при гибридизационных условиях; после чего
(2) синтезирование в водном растворе варианта системы, идентифицированной в соответствии с (1), который отличается от последней
(i) включением в первый фрагмент (В) второй нити дуплекса-мишени первой химической модификации, которая приводит к тому, что связывание со вторым фрагментом (В′) первой нити дуплекса столкновения имеет пониженную свободную энергию гибридизации; и/или
(ii) включением во второй фрагмент (В) входной нити второй химической модификации, которая приводит к тому, что связывание со вторым фрагментом (В′) первой нити дуплекса столкновения имеет повышенную свободную энергию гибридизации,
так что первый фрагмент (В) второй нити дуплекса-мишени по сравнению со вторым фрагментом (В) входной нити связывается со вторым фрагментом (В′) первой нити дуплекса столкновения с образованием более стабильной области дуплекса,
в силу чего указанный вариант является улучшенным по сравнению с указанной системой, идентифицированной в соответствии с (1), в отношении пониженной частоты случаев гибридизации между входной нитью и дуплексом столкновения.
2. Способ по п.1, в котором указанный вариант содержит первую химическую модификацию.
3. Способ по п.1, в котором указанный вариант содержит вторую химическую модификацию.
4. Способ по п.1, в котором указанные входная нить, дуплекс-мишень и дуплекс столкновения образуют часть не содержащего ферменты логического элемента на нуклеиновых кислотах, выбираемого из группы, состоящей из элемента AND, элемента NOT, элемента OR, элемента NAND, элемента NOR, элемента XOR и элемента XNOR.
5. Способ по п.1, в котором указанные первая и вторая химические модификации выбираются из группы, состоящей из (i) замены сахарофосфодиэфирного остова указанных нуклеотидов на псевдопептидный остов, (ii) модификации сахарной составляющей указанных нуклеотидов и (iii) замены азотистого основания его аналогом по меньшей мере в одном из указанных нуклеотидов.
6. Способ по п.5, в котором указанные первая и вторая химические модификации включают замену сахарофосфодиэфирного остова указанных нуклеотидов на псевдопептидный остов, в который встроена функциональная группа гуанидиния.
7. Способ по п.5, в котором указанные первая и вторая химические модификации включают замену азотистого основания на трициклический аналог цитозина по меньшей мере в одном из указанных нуклеотидов.
8. Способ по п.5, в котором указанные первая и вторая химические модификации включают введение гетероатома в 2′-позицию указанной сахарной составляющей.
9. Способ по п.1, в котором указанная реакция перестановки нити включает опосредованную референсной точкой перестановку нити.
10. Вычислительная система, основанная на нуклеиновых кислотах, включающая:
(i) входную нить нуклеиновой кислоты, включающую первый фрагмент (А) и второй фрагмент (В),
(ii) мишень, представляющую собой дуплекс нуклеиновой кислоты с неполным спариванием оснований, включающий первую и вторую нити,
где указанная первая нить включает первый фрагмент (В′) и второй фрагмент (А′),
где указанная вторая нить включает первый фрагмент (В) и второй фрагмент (С), а указанный дуплекс-мишень включает дуплексную область, образованную спариванием оснований между В и В′ и двумя однонитиевыми областями А′ и С, и
где указанная входная нить может связываться с первой нитью дуплекса-мишени при гибридизационных условиях, чтобы вызвать реакцию перестановки нити с получением выходного дуплекса, образованного (А-В) входной нити и первой нитью дуплекса-мишени (В′-А′), и
(iii) столкновение, представляющее собой дуплекс нуклеиновой кислоты с неполным спариванием оснований, включающий первую и вторую нити,
где указанная первая нить включает первый фрагмент (С′) и второй фрагмент (В′),
где указанная вторая нить включает первый фрагмент (С) и второй фрагмент (D), а указанный дуплекс столкновения включает дуплексную область, образованную спариванием оснований между С и С' и двумя однонитиевыми областями В′ и D,
где вторая нить дуплекса-мишени может связываться с первой нитью дуплекса столкновения при гибридизационных условиях, чтобы вызвать реакцию перестановки нити с получением выходного дуплекса, образованного второй нитью дуплекса-мишени (В-С) и первой нитью дуплекса столкновения (С′-В′), и
где второй фрагмент (В) входной нити может связываться с однонитиевой областью В′ дуплекса столкновения при гибридизационных условиях;
где
первый фрагмент (В) второй нити дуплекса-мишени содержит первую химическую модификацию по отношению к природной нуклеиновой кислоте, которая приводит к тому, что связывание со вторым фрагментом (В′) первой нити дуплекса столкновения имеет пониженную свободную энергию гибридизации; и/или
второй фрагмент (В) входной нити содержит вторую химическую модификацию по отношению к природной нуклеиновой кислоте, которая приводит к тому, что связывание со вторым фрагментом (В′) первой нити дуплекса столкновения имеет повышенную свободную энергию гибридизации,
так что первый фрагмент (В) второй нити дуплекса-мишени по сравнению со вторым фрагментом (В) входной нити связывается со вторым фрагментом (В′) первой нити дуплекса столкновения с образованием более стабильной области дуплекса.
11. Система по п.10, в которой указанные входная нить, дуплекс-мишень и дуплекс столкновения образуют часть не содержащего ферменты логического элемента на нуклеиновых кислотах, выбираемого из группы, состоящей из элемента AND, элемента NOT, элемента OR, элемента NAND, элемента NOR, элемента XOR и элемента XNOR.
12. Система по п.10, в которой указанные первая и вторая химические модификации выбираются из группы, состоящей из (i) замены сахарофосфодиэфирного остова указанных нуклеотидов на псевдопептидный остов, (ii) модификации сахарной составляющей указанных нуклеотидов и (iii) замены азотистого основания его аналогом по меньшей мере в одном из указанных нуклеотидов.
13. Система по п.12, в которой указанные первая и вторая химические модификации включают замену сахарофосфодиэфирного остова указанных нуклеотидов на псевдопептидный остов, в который встроена функциональная группа гуанидиния.
14. Система по п.13, в которой указанные первая и вторая химические модификации включают замену азотистого основания на трициклический аналог цитозина по меньшей мере в одном из указанных нуклеотидов.
15. Система по п.13, в которой указанные первая и вторая химические модификации включают введение гетероатома в 2′-позицию указанной сахарной составляющей.
16. Система по п.10, в которой указанная реакция перестановки нити включает опосредованную референсной точкой перестановку нити.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| TANAKA FUMIAKI et al | |||
| "Design of nucleic acid sequences for DNA computing based on a thermodynamic approach." Nucleic Acids Research, 2005, 33(3): 903-911 | |||
| SHORTREED M.R | |||
| et al | |||
| "A thermodynamic approach to designing structure-free combinatorial DNA word | |||

