ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке испрашивается приоритет в соответствии с 35 U.S.C. § 119(e) предварительной патентной заявки США с серийным № 61/754339, зарегистрированной 18 января 2013 года, и в соответствии с 35 U.S.C. § 120 предварительной патентной заявки США с серийным № 13/801762, зарегистрированной 13 марта 2013 года, содержание каждой из которых полностью включено в настоящий документ в качестве ссылки.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
[0002] Получение молекулярных компьютеров с наноразмерами обещает значительный потенциал, отчасти потому, что такие компьютеры могут очень хорошо подходить для решения определенных задач по обработке информации. В частности, компьютеры с использованием биомолекул могут быть совместимыми с биологическими средами, что делает их пригодными для применения в диагностике комплексных заболеваний или даже в лекарственных средствах.
[0003] Способность транслировать одну последовательность нуклеиновой кислоты в другую теоретически позволяет построение логических вентилей и схем с использованием нуклеиновых кислот. Эти вентили и схемы находятся под управлением двух процессов, гибридизации и замещения цепей, которые, как правило, оба являются термодинамически выгодными, например, они включают переход из более высокого в более низкое энергетическое состояние. Таким образом, оба процесса в системе могут происходить спонтанно.
[0004] В гибридизации участвуют свободные одноцепочечные участки нуклеиновых кислот. Таким образом, систему нуклеиновых кислот можно регулировать доступностью этих свободных цепей.
[0005] "Процесс изоляции" позволяет определенным последовательностям быть доступными для остальной системы в зависимости от определенных условий. Такие процессы дают возможность конструировать трансляторы, которые преобразуют одну одноцепочечную последовательность нуклеиновой кислоты в другую одноцепочечную последовательность нуклеиновой кислоты. Эти трансляторы являются основанием, на котором с использованием нуклеиновых кислот можно строить основные логические операторы, такие как AND, NOT, OR, NAND, NOR, XOR и XNOR. На основе этих и других логических компонентов можно конструировать более крупные схемы, которые включают такие компоненты, как амплификаторы. Таким образом, эти процессы трансляции важны для обработки информации с использованием нуклеиновых кислот и молекулярных вычислений.
СУЩНОСТЬ
[0006] По одному из аспектов настоящего изобретения предоставлена композиция, содержащая первый и второй комплексы нуклеиновых кислот, каждый из которых содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая из цепей содержит последовательно первый, второй и третий фрагменты, где цепи нуклеиновых кислот определены как B-X-D, 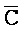 -
-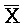 -
-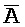 ,
, 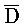 -
-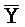 -
-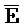 и F-Y-C для первой, второй, третьей и четвертой цепей первого комплекса, соответственно, и
и F-Y-C для первой, второй, третьей и четвертой цепей первого комплекса, соответственно, и 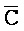 -
-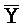 -
-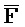 , E-Y-D, F-Z-G,
, E-Y-D, F-Z-G, 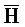 -
-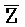 -
-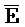 для первой, второй, третьей и четвертой цепей второго комплекса, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где: каждый из первого и второго комплексов содержит первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (X::
для первой, второй, третьей и четвертой цепей второго комплекса, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где: каждый из первого и второго комплексов содержит первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (X::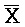 и
и 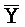 ::Y, в первом и втором комплексах, соответственно), вторую область дуплекса, сформированную между вторыми фрагментами третьей и четвертой цепей (Y::
::Y, в первом и втором комплексах, соответственно), вторую область дуплекса, сформированную между вторыми фрагментами третьей и четвертой цепей (Y::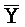 и Z::
и Z::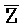 ), третью область дуплекса, сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи (D::
), третью область дуплекса, сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи (D::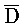 и
и 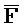 ::F), и четвертую область дуплекса, сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи (
::F), и четвертую область дуплекса, сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи (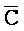 ::C и E::
::C и E::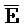 ); в каждом из первого и второго комплексов первый фрагмент (B и
); в каждом из первого и второго комплексов первый фрагмент (B и 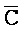 в первом и втором комплексах, соответственно) первой цепи, третий фрагмент (
в первом и втором комплексах, соответственно) первой цепи, третий фрагмент (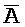 и D) второй цепи, третий фрагмент (
и D) второй цепи, третий фрагмент (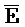 и G) третьей цепи и первый фрагмент (F и
и G) третьей цепи и первый фрагмент (F и 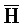 ) четвертой цепи являются одноцепочечными; третья цепь первого комплекса (
) четвертой цепи являются одноцепочечными; третья цепь первого комплекса (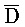 -
-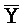 -
-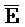 ) обладает подходящей комплементарностью последовательности со второй цепью второго комплекса (E-Y-D) для обеспечения связывания между ними в условиях гибридизации; и четвертая цепь первого комплекса (F-Y-C) обладает подходящей комплементарностью последовательности с первой цепью второго комплекса (
) обладает подходящей комплементарностью последовательности со второй цепью второго комплекса (E-Y-D) для обеспечения связывания между ними в условиях гибридизации; и четвертая цепь первого комплекса (F-Y-C) обладает подходящей комплементарностью последовательности с первой цепью второго комплекса (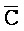 -
-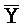 -
-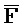 ) для обеспечения связывания между ними в условиях гибридизации.
) для обеспечения связывания между ними в условиях гибридизации.
[0007] В одном из аспектов композиция дополнительно содержит третий комплекс нуклеиновых кислот, содержащий первую и вторую цепи нуклеиновых кислот, где каждая из цепей содержит последовательно первый, второй и третий фрагменты, где первая и вторая цепи определены как E-Z-H и 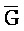 -
-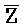 -
-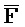 , соответственно, где третий комплекс нуклеиновых кислот содержит область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (Z::
, соответственно, где третий комплекс нуклеиновых кислот содержит область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (Z::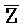 ), а первый фрагмент (E) первой цепи и третий фрагмент второй цепи (
), а первый фрагмент (E) первой цепи и третий фрагмент второй цепи (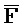 ) являются одноцепочечными, и где:
) являются одноцепочечными, и где:
третья цепь второго комплекса (F-Z-G) обладает подходящей комплементарностью последовательности со второй цепью третьего комплекса (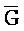 -
-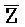 -
-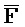 ) для обеспечения связывания между ними в условиях гибридизации; а четвертая цепь второго комплекса (
) для обеспечения связывания между ними в условиях гибридизации; а четвертая цепь второго комплекса (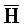 -
-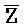 -
-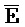 ) обладает подходящей комплементарностью последовательности с первой цепью третьего комплекса (E-Z-H) для обеспечения связывания между ними в условиях гибридизации.
) обладает подходящей комплементарностью последовательности с первой цепью третьего комплекса (E-Z-H) для обеспечения связывания между ними в условиях гибридизации.
[0008] В определенных аспектах третий фрагмент (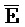 ) третьей цепи первого комплекса и первый фрагмент (E) первой цепи третьего комплекса обладают комплементарностью последовательностей, но в нормальных условиях гибридизации стабильно друг с другом не связываются вследствие химической модификации любого из двух или обоих фрагментов; и первый фрагмент (F) четвертой цепи первого комплекса и третий фрагмент (
) третьей цепи первого комплекса и первый фрагмент (E) первой цепи третьего комплекса обладают комплементарностью последовательностей, но в нормальных условиях гибридизации стабильно друг с другом не связываются вследствие химической модификации любого из двух или обоих фрагментов; и первый фрагмент (F) четвертой цепи первого комплекса и третий фрагмент (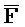 ) первой цепи третьего комплекса обладают комплементарностью последовательностей, но в нормальных условиях гибридизации стабильно друг с другом не связываются вследствие химической модификации любого из двух или обоих фрагментов.
) первой цепи третьего комплекса обладают комплементарностью последовательностей, но в нормальных условиях гибридизации стабильно друг с другом не связываются вследствие химической модификации любого из двух или обоих фрагментов.
[0009] Химические модификации, подходящие для практического осуществления технологии по настоящему изобретению, в качестве неограничивающих примеров включают замену сахарофосфатного каркаса фрагмента нуклеиновой кислоты конъюгированной с мини-ПЭГ получаемой из серина гамма-ПНК. В определенных аспектах химическая модификация включает замену азотистого основания по меньшей мере одного нуклеозида во фрагменте нуклеиновой кислоты на трициклический аналог цитозина. В одном из аспектов химическая модификация включает введение гетероатома в 2'-положение сахарной группы в нуклеотиде.
[0010] Иллюстрации комплексов и их взаимосвязи можно найти на ФИГ. 6A, где транслятор 102 соответствует первому комплексу, транслятор 103 соответствует второму комплексу, а транслятор 104 соответствует третьему комплексу. Подобным образом, на ФИГ. 10-13, 14A-D и 15-16 также проиллюстрированы трансляторы (комплексы нуклеиновых кислот) в объеме настоящего изобретения, включающие каждый отдельный транслятор и их комбинации в виде наборов трансляторов (или в форме композиции).
[0011] В одном из аспектов длина каждого фрагмента составляет от приблизительно 5 оснований до приблизительно 50 оснований. В другом аспекте длина каждого фрагмента, который является одноцепочечным, составляет от приблизительно 5 оснований до приблизительно 30 оснований.
[0012] Также в объем изобретения входит то, что каждая композиция может дополнительно необязательно содержать фармацевтически приемлемый носитель. В определенных вариантах осуществления настоящего изобретения дополнительно предоставлена клетка, содержащая композицию или комплекс по настоящему изобретению.
[0013] Также в настоящем документе предоставлены компьютерно-реализованные способы и энергонезависимые считываемые компьютером носители, подходящие для моделирования, проектирования, регистрации, представления или анализа определенных аспектов описываемой технологии.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0014] В качестве вариантов осуществления настоящего изобретения предоставлены чертежи, которые являются только иллюстративным примером и не являются ограничением, где:
[0015] на ФИГ. 1A проиллюстрировано осуществление транслятора на основе нуклеиновых кислот посредством изоляции на твердой фазе, где отдельные отрезки (A, B и т.д.) представляют собой участки олигонуклеотидов произвольной длины и последовательности;
[0016] на ФИГ. 1B проиллюстрировано, как осуществление транслятора на основе нуклеиновых кислот посредством изоляции на твердой фазе изолирует участки олигонуклеотидов;
[0017] на ФИГ. 2A проиллюстрировано осуществление транслятора на основе нуклеиновых кислот посредством изоляции посредством "фиксаторов". Снова, отдельные отрезки представляют собой участки олигонуклеотидов произвольной длины и последовательности;
[0018] на ФИГ. 2B проиллюстрировано, как посредством осуществления на основе изолирования фиксаторами, представленного на ФИГ. 2A, изолировать участки олигонуклеотидов;
[0019] на ФИГ. 3A представлена система из трех изолированных фиксаторами трансляторов на основе нуклеиновых кислот. Все приведенные реакции представляют собой реакции замещения цепей, проходящие по тому же механизму, что миграции цепи, что и на ФИГ. 2A;
[0020] на ФИГ. 3B представлена система с теми же олигонуклеотидами, которые представлены на ФИГ. 3A, но где вместо реакций замещения цепей представлено образование "замыканий" фиксаторов, где фиксатор связывается с последовательностью, которая не может участвовать в реакции замещения цепей. Это событие связывания оккупирует фиксатор так, что желаемая цепь не может связаться;
[0021] на ФИГ. 4a-b продемонстрированы реакции замещения с использованием (a) или без использования (b) фиксаторов в буфере PBS (137 мМ NaCl, 2,7 мМ KCl, 10 мМ Na2HPO4, 1,76 мМ KH2PO4, pH 7,4) при 37°C. Флуоресцентные сигналы преобразовывали в концентрации посредством использования стандартной кривой. a) 50 нМ [F-(A24)::(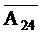 )-Q]0 дуплекс инкубировали с титрованием 10-200 нМ [(A24)]0 с шагом 10 нМ (от синего к красному) и концентрации свободного [F-(A24)]t контролировали посредством резонансного переноса энергии флуоресценции (FRET). b) 50 нМ [F-(A24)::()-Q]0 дуплекс смешивали с титрованием 10-200 нМ [(A24)::()]0 с шагом 10 нМ (от меньшего к большему) и концентрации [F-(A24)::()]t контролировали посредством FRET;
)-Q]0 дуплекс инкубировали с титрованием 10-200 нМ [(A24)]0 с шагом 10 нМ (от синего к красному) и концентрации свободного [F-(A24)]t контролировали посредством резонансного переноса энергии флуоресценции (FRET). b) 50 нМ [F-(A24)::()-Q]0 дуплекс смешивали с титрованием 10-200 нМ [(A24)::()]0 с шагом 10 нМ (от меньшего к большему) и концентрации [F-(A24)::()]t контролировали посредством FRET;
[0022] на ФИГ. 5 проиллюстрирован подход замены фиксаторов при проектировании трансляторов;
[0023] на ФИГ. 6A-6B проиллюстрированы комплексы нуклеиновых кислот (трансляторы), применяемые в изолирующей посредством чередования системе;
[0024] на ФИГ. 7 представлены модифицированные каркасные структуры аналогов нуклеиновых кислот, где "B" представляет произвольное нуклеиновое основание, и в (a) представлен природный фосфодиэфирный каркас, присутствующий в ДНК, в (b) представлены пептидные нуклеиновые кислоты, в (c) представлены гуанидиниевые пептидные нуклеиновые кислоты, в (d) представлены получаемые из L-серина гамма-ПНК, в (e) представлены фосфородиамидаты (на этом рисунке с морфолиновым сахаром), и в (f) и (g) представлены конъюгированные с мини-ПЭГ, получаемые из серина гамма-ПНК;
[0025] на ФИГ. 8 проиллюстрированы структуры модифицированных сахаров аналогов нуклеиновых кислот, где "B" представляет произвольное нуклеиновое основание, и в (a) представлен природный дезоксирибозный сахар, присутствующий в ДНК, в (b) представлены морфолино, в (c) представлены закрытые нуклеиновые кислоты и в (d) представлены модифицированное фтором производное РНК;
[0026] на ФИГ. 9 проиллюстрированы структуры модифицированных нуклеиновых оснований аналогов нуклеиновых кислот метилцитозина (a), диаминопурина (b), феноксазина (c) и G-зажим (d);
[0027] на ФИГ. 10 представлен адаптер иРНК;
[0028] на ФИГ. 11 представлен адаптер РНКи;
[0029] на ФИГ. 12 представлен антисмысловой адаптер;
[0030] на ФИГ. 13 представлен набор трансляторов с объединением на входе;
[0031] на ФИГ. 14A-14D представлены четыре набора трансляторов с разделением на выходе. Два из этих наборов продуцируют два или более идентичных транслятора, таким образом, работая в качестве амплификаторов (A и B), тогда как другие два набора порождают два или более различных последующих трансляторов (C и D);
[0032] на ФИГ. 15 проиллюстрирован зависимый от соединения набор трансляторов;
[0033] на ФИГ. 16 представлен инвертер;
[0034] на ФИГ. 17A-M представлены результаты имитации систем с изолирующими посредством чередования трансляторами (RST) и систем с трансляторами с изолированными фиксаторами (THST), в отношении основного сигнала и фонового сигнала для различных размеров фиксаторов в диапазоне от 3 нуклеотидов (н.) до 15 н. (как указано). Все имитации проводили со 100 нМ трансляторами в 3 этапа;
[0035] на ФИГ. 18A-D представлены результаты имитации для систем с RST и THST с различными глубинами (т.е. количеством участвующих трансляторов/стадий, как указано) систем. Все имитации проводили с фиксатором длиной 8 н. с концентрациями трансляторов 100 нМ; и
[0036] на ФИГ. 19A-N представлены результаты имитации с системами средних размеров (5 этапов) для длинных (13 н.) и коротких (8 н.) фиксаторов и показано, как изменяется поведение системы при варьировании указанных концентраций трансляторов.
[0037] Некоторые или все фигуры представляют собой схематические представления для иллюстрации; таким образом, на них не обязательно изображены действительные относительные размеры или положения представленных элементов. Фигуры представлены с целью иллюстрации одного или нескольких вариантов осуществления с четким пониманием того, что их не следует использовать для ограничения объема или смысла приведенной ниже формулы изобретения.
ПОДРОБНОЕ ОПИСАНИЕ
[0038] Среди различных подходов, которые можно использовать при трансляции последовательностей нуклеиновых кислот для построения логических операторов и схем, находятся изоляция на твердой фазе, изоляция фиксаторов и замена фиксаторов. Более подробно описанные ниже, эти три конкретных подхода проиллюстрированы конфигурациями, в которых использованы реакции трехсторонней, опосредованной фиксаторами миграции цепей. Для реакций миграции цепей возможны дополнительные механизмы, включая в качестве неограничивающих примеров четырехстороннюю миграцию цепей, четырехстороннюю ускоренную миграцию и миграцию комплекса из множества цепей.
[0039] Таким образом, хотя ниже описаны варианты осуществления, где в иллюстративных целях используют трехстороннюю миграцию цепей, настоящее изобретение предусматривает логические вентили и схемы с использованием ДНК, построенные с использованием других способов миграции цепей. И наоборот, варианты осуществления изобретения можно использовать для любой реакции миграции цепи.
[0040] На всем протяжении настоящего описания и на сопровождающих фигурах прописные буквы, например, A, B, C, X, Y, Z, необязательно с подстрочными индексами или надстрочными индексами, используют для обозначения участков, также обозначаемых как "фрагменты", олигонуклеотидов произвольной длины. Соответствующие A', B', C', X', Y', Z' или взаимозаменяемо 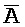 ,
, 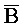 ,
, 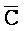 ,
, 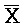 ,
, 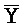 ,
, 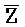 представляют соответствующие обратно комплементарные участки.
представляют соответствующие обратно комплементарные участки.
[0041] Термины "олигонуклеотиды", "полинуклеотиды" и "нуклеиновые кислоты" используют в настоящем документе для обозначения всех форм молекул нуклеиновых кислот. Без ограничения эта категория включает рибонуклеиновые кислоты (РНК), дезоксирибонуклеиновые кислоты (ДНК), пептидные нуклеиновые кислоты (ПНК) и их производные с модификациями и без, соответственно.
Изоляция на твердой фазе
[0042] Изоляция на твердой фазе включает физическое разделение соответствующих последовательностей/цепей в пространстве посредством гранул, наночастиц или поверхностей. В этом подходе используют принципы изоляции участка, который находит широкое применение в органической химии. Временные рамки этих событий замещения в твердофазных изолирующих конфигурациях можно контролировать посредством регуляции нахождения необходимых цепей в жидкой фазе или в твердой фазе системы.
[0043] На ФИГ. 1A представлена основная для транслятора схема изоляции на твердой фазе, компонент, который позволяет системе замещать одну последовательность нуклеиновой кислоты другой. Для цепи A'-X' каждый из A и X представляет собой участок олигонуклеотида произвольной длины и последовательности, а X' и A' представляют собой соответствующие им участки обратной комплементарности. Являясь связанной с твердой подложкой, цепь A'-X' сначала гибридизуется с Y-B-X, формируя структуру нуклеиновой кислоты в форме дуплекса с неполным спариванием оснований, который может функционировать в качестве транслятора. В этой конфигурации цепь Y-B-X изолирована на твердой фазе и, таким образом, не может взаимодействовать с остальной системой. Однако в присутствии цепи X-A, обозначаемой как "полинуклеотидная замещающая молекула", цепь Y-B-X может быть вытеснена с твердой подложки и экспонирована в жидкую фазу системы, в то время как цепь X-A будет связана с подложкой. Эта операция включает два этапа; первый представляют собой гибридизацию комплементарных последовательностей A и A' (часто обозначаемую как "связывание фиксаторов"). На втором этапе область X цепи X-A связывается с областью X' A'-X', замещая область X Y-B-X и высвобождая эту цепь в раствор, оставляя при этом X-A связанной с твердой подложкой (этот этап часто обозначают как "реакция миграции цепи"). Этот двухэтапный процесс эффективно осуществляет трансляцию свободной цепи X-A в свободную цепь Y-B-X.
[0044] На ФИГ. 1B представлена система с входной цепью X-A, взаимодействующей с дуплексом с неполным спариванием оснований, "транслятором 1". Выход включает дуплекс с полностью спаренными основаниями, A'-X'/X-A, который рассматривают в качестве "отработанного" продукта, и выходную цепь Y-B-X, которую обозначают как "выход 1", и ее можно использовать в качестве "входа 2" в дальнейшей реакции. "Вход 2" взаимодействует с "транслятором 2" и продуцирует "выход 2" и другой отработанный продукт. На этой фигуре область B цепи Y-B-X иллюстрирует изоляцию последовательностей в этой системе. В начале Y-B-X не может гибридизоваться с областью B' транслятора 2, так как оба изолированы на разных твердых подложках. Когда вход 1 связывается с транслятором 1 и высвобождает Y-B-X в раствор, тогда Y-B-X может взаимодействовать с транслятором 2. Таким образом, способность Y-B-X и транслятора 2 взаимодействовать обусловлена присутствием входа 1.
[0045] Цепи, связанные на одной твердой поверхности, крайне медленно взаимодействуют с цепями на другой твердой поверхности вследствие пространственных эффектов. Таким образом, компонентами, которые могут взаимодействовать с операторами на твердой фазе, являются только цепи в жидкой фазе.
Изоляция фиксаторов
[0046] Изоляция фиксаторов осуществляет те же операции что и трансляторы на твердой фазе, но функционирует, оставляя участки последовательности связанными в дуплексе. Также как и в трансляторе на твердой фазе, событие замещения могут высвобождать представляющую интерес последовательность. Все цепи могут находиться в растворе вместе, где последовательность их событий замещения регулирует доступность фиксаторов. Здесь термин "фиксатор" относится к коротким участкам одноцепочечных последовательностей нуклеиновых кислот, которые обеспечивают точку старта для события замещения.
[0047] На ФИГ. 2A представлен транслятор с изолированными фиксаторами, сходный с транслятором на ФИГ. 1A, но на основе фиксатора, а не на основе изоляции на твердой фазе. В этом примере область A' транслятора представляет собой фиксатор, который связывается с входной цепью и обеспечивает прохождение реакции замещения цепей.
[0048] На ФИГ. 2B представлена система на основе фиксаторов со входной цепью A-X-B, взаимодействующей с дуплексом с неполным спариванием оснований, "транслятором 1". Выход включает отработанный продукт, например, дуплекс с полностью спаренными основаниями, B'-X'-A'/A-X-B, и "выход 1", цепь X-B-Y-C, которую можно использовать в качестве "входа 2" в дальнейшей реакции. "Вход 2" взаимодействует с "транслятором 2" и продуцирует "выход 2" и другой отработанный продукт. На этой фигуре область B X-B-Y-C изолирована в трансляторе 1 посредством гибридизации с комплементарной областью B' и, таким образом, не может взаимодействовать с областью B' транслятора 2. Способность X-B-Y-C взаимодействовать с транслятором 2 обусловлена присутствием в системе входа 1 (A-X-B).
[0049] Несмотря на очень высокую потенциальную пользу, применения конфигураций фиксаторов до настоящего времени были ограничены скоростью, с которой система, содержащая такие конфигурации фиксаторов, могла передавать информацию. Неотъемлемые ограничения при обычных подходах с изоляцией фиксаторов часто могут замедлять конечную передачу ниже биологически пригодных сроков. Более конкретно, трансляторы с изолированными фиксаторами функционируют с приемлемыми скоростями только в очень узком динамическом диапазоне концентраций, как следствие замыкающих взаимодействий. Регулируя длину фиксаторов, замыкающие взаимодействия при низких концентрациях можно делать обратимыми. Однако по мере роста концентрации образование замыканий будет доминировать, и система прекратит работу. С другой стороны, если концентрацию снизить, опосредованное фиксаторами замещение также прекратится. В связи с этим, следует отметить, что в биологическом окружении концентрации могут широко варьировать; таким образом, такие трансляторы плохо подходят для применения в настоящем биологическом окружении. Полагают, что кинетические "узкие места" приводят к непродуктивным реакциям, обозначаемым в настоящем документе как "замыкание фиксаторов", которое происходит, когда фиксатор связан с молекулой с комплементарной последовательностью или с "замыкающей цепью", которая не может приводить к реакции замещения.
[0050] Например, на ФИГ. 3A представлена система из трех трансляторов на основе нуклеиновых кислот с изолированными фиксаторами, подобных транслятору на ФИГ. 2A. Однако если все три цепи находятся в растворе вместе, существуют другие события связывания, которые могут происходить. На ФИГ. 3B проиллюстрированы некоторые из событий непродуктивного связывания или замыканий, которые могут происходить. Вовлекая "полинуклеотидную замыкающую молекулу", эти события не приводят к реакции замещения, а могут замедлять систему, поскольку образование замыкающей цепи блокирует связывание цепей, которое может приводить к реакции замещения. Такие события замыкания могут становиться все более преобладающими по мере роста концентраций трансляторов на основе нуклеиновых кислот в системе.
[0051] Другим ограничением этого подхода является потенциальная токсичность, являющаяся результатом доступности длинных одноцепочечных областей трансляторов. Например, когда одноцепочечная область связывается с эндогенной ДНК или РНК в клетке, она может изменять транскрипцию, трансляцию или другую функцию молекулы ДНК или РНК. Такое "побочное" событие может приводить к нежелательным последствиям в клетке.
[0052] Другим ограничением является потенциальная постоянная утечка сигнала вследствие фоновых реакций трансляторов в отсутствие открытых фиксаторов, что также может приводить к нежелательным последствиям. Например, на ФИГ. 4a представлено, что химическая активность между транслятором и фиксатором с увеличением концентрации фиксатора возрастает. Однако в отсутствие фиксатора фоновую химическую активность также наблюдают (ФИГ. 4b).
[0053] Одним из предлагаемых решений является получение коротких фиксаторов для ослабления действия замыканий на систему: чем более коротким является фиксатор, тем быстрее может быть скорость прямой/обратной реакций с комплементарной последовательностью. Таким образом, обычными являются фиксаторы длиной пять или шесть нуклеотидов, так как при этих длинах, если происходит событие непродуктивного связывания, время, проведенное в двухцепочечном "замкнутом" состоянии, является коротким.
[0054] Однако этот подход создает указанное выше кинетическое "узкое место", так как событие продуктивного связывания ограничено теми же термодинамическими параметрами; таким образом, входящая цепь также с этими фиксаторами прочно не связывается. Таким образом, при корректном связывании входящей цепи желаемое замещение происходит не всегда, так как для инициации реакции замещения необходимо, чтобы она была в связанном состоянии достаточно долго. Таким образом, использование коротких фиксаторов увеличивает период, необходимый для осуществления данной операции и получения выходящей цепи. Другими словами, реакция замещения не может происходить без наступления множества событий связывания с замыкающими цепями и желаемыми цепями. Эта неэффективность ограничивает пользу системы, замедляя передачу информации до сроков, которые являются слишком длительными, чтобы являться пригодными.
[0055] Более короткие фиксаторы, наряду с более короткими одноцепочечными областями на трансляторах, также могут помочь в уменьшении токсичности. Однако более короткие фиксаторы не решают проблемы с утечкой сигнала.
Замена фиксаторов
[0056] При "замене фиксаторов" используют сходные взаимодействия спаривания, как и при изоляции фиксаторов, но с другими конфигурациями. При замене фиксаторов делают попытку разрешить проблему замыкания фиксаторов, возникающую при подходе изоляции фиксаторов. Со ссылкой на ФИГ. 5, исходный фиксатор, A-B, только частично комплементарен одноцепочечной области в дуплексе справа от фиксатора (например, A::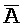 ). Подобным образом, когда получают фиксатор B-C-D посредством реакции замещения фиксатором A-B, фиксатор B-C-D также только частично комплементарен одноцепочечной области дуплекса за ним. По существу, замыкания фиксаторов происходить не может.
). Подобным образом, когда получают фиксатор B-C-D посредством реакции замещения фиксатором A-B, фиксатор B-C-D также только частично комплементарен одноцепочечной области дуплекса за ним. По существу, замыкания фиксаторов происходить не может.
[0057] Однако у подхода замены фиксаторов также существуют свойственные ему ограничения. Во-первых, также как и при изоляции фиксаторов, длинные одноцепочечные области трансляторов могут вызывать нежелательное токсическое действие вследствие побочного связывания с эндогенными нуклеиновыми кислотами в клетке.
[0058] Другая проблема относится к динамическому диапазону трансляции, что сходно, хотя и не одинаково с проблемами, с которыми сталкиваются при замыкании фиксаторов. Однако эта проблема при замене фиксаторов является менее тяжелой, так как она больше влияет на скорость трансляции, чем на выход. В этом случае длины фиксаторов определяют скорость спонтанной диссоциации, которая является более высокой при более коротких фиксаторах. С другой стороны, они также определяют скорость прямого замещения, но прямое замещение более быстрым делают более длинные фиксаторы. Таким образом, между этими ограничениями существует зависимый от концентрации баланс.
[0059] Важным ограничением подхода замены фиксаторов является низкий выход, так как все реакции являются двунаправленными, что приводит к отсутствию предпочтительного конечного продукта за исключением случаев, когда, например, вся трансляция связана с необратимой реакцией в конце.
Изолирующие посредством чередования трансляторы
[0060] В настоящем изобретении предоставлен ряд трансляторов, которые решают проблемы, с которыми сталкиваются в подходах, подробно описанных выше. С использованием этих трансляторов минимизировано токсическое побочное действие и фоновая реактивность, можно избежать замыканий фиксаторов, общий выход близок к полному и скорость трансляции все еще значительно повышена.
[0061] На ФИГ. 6A проиллюстрированы некоторые основные компоненты системы "изолирующего посредством чередования транслятора". Наверху представлен комплекс нуклеиновых кислот (101), формируемый между двумя цепями, A-X-C и 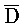 -
-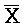 -
-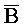 . Вследствие комплементарности их последовательностей, формируется область дуплекса X::
. Вследствие комплементарности их последовательностей, формируется область дуплекса X::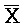 , тогда как A, C,
, тогда как A, C, 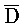 и
и 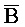 остаются одноцепочечными. Такой частичный дуплекс далее обозначают как "двойной фиксатор".
остаются одноцепочечными. Такой частичный дуплекс далее обозначают как "двойной фиксатор".
[0062] Второй комплекс нуклеиновых кислот (102), представленный на ФИГ. 6A, состоит из четырех цепей нуклеиновых кислот, B-X-D, 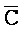 -
-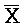 -
-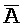 ,
, 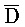 -
-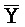 -
-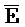 и F-Y-C. Как показано, эти цепи формируют между собой четыре области дуплексов, а на концах остаются четыре одноцепочечных области. Одна из этих четырех цепей,
и F-Y-C. Как показано, эти цепи формируют между собой четыре области дуплексов, а на концах остаются четыре одноцепочечных области. Одна из этих четырех цепей, 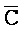 -
-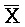 -
-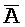 , комплементарна одной из цепей, A-X-C, в первом комплексе, тогда как другая цепь, B-X-D, комплементарна
, комплементарна одной из цепей, A-X-C, в первом комплексе, тогда как другая цепь, B-X-D, комплементарна 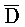 -
-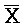 -
-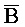 , другой цепи первого комплекса.
, другой цепи первого комплекса.
[0063] Вследствие комплементарности последовательностей в свисающих одноцепочечных областях, первый комплекс (101) обладает подходящей комплементарностью последовательности так, чтобы быть способным к связыванию со вторым комплексом (102), что инициирует реакцию замещения цепей, в которую вовлечены шесть цепей. Продукт реакции замещения цепей на выходе включает два полностью отожженных дуплекса (105 и 106) и новый двойной фиксатор (107), содержащий в середине область дуплекса и четыре разделенных одноцепочечных области (ФИГ. 6B).
[0064] На ФИГ. 6B представлено, что полученные комплексы нуклеиновых кислот (105, 106 и 107) находятся в более низком энергетическом состоянии вследствие двух полных дуплексов, A-X-C::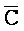 -
-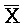 -
-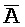 (105) и B-X-D::
(105) и B-X-D::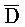 -
-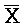 -
-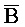 (106). Другими словами, такое замещение цепей с большим предпочтением проходит в прямом направлении и, таким образом, не так легко обратимо.
(106). Другими словами, такое замещение цепей с большим предпочтением проходит в прямом направлении и, таким образом, не так легко обратимо.
[0065] Затем вновь сформированный двойной фиксатор, состоящий из 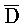 -
-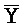 -
-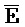 и F-Y-C (107), может инициировать другую реакцию замещения цепей с третьим комплексом нуклеиновых кислот (103) на ФИГ. 6A (см. стрелки), продолжая процесс трансляции. Подобным образом, новый двойной фиксатор, получаемый после этой реакции замещения, может взаимодействовать с четвертым комплексом (104) и так далее.
и F-Y-C (107), может инициировать другую реакцию замещения цепей с третьим комплексом нуклеиновых кислот (103) на ФИГ. 6A (см. стрелки), продолжая процесс трансляции. Подобным образом, новый двойной фиксатор, получаемый после этой реакции замещения, может взаимодействовать с четвертым комплексом (104) и так далее.
[0066] Как объяснено выше, реакции замещения цепей в этом подходе достигают высокой скорости выхода, так как в каждой из этих реакций продукты на выходе образуются в намного меньшем энергетическом состоянии, чем молекулы нуклеиновых кислот на входе. Кроме того, как проиллюстрировано в таблице 1, экспериментальные данные демонстрируют, что такие опосредованные двойными фиксаторами реакции также могут быть очень быстрыми. Следует сравнить столбец "двойные фиксаторы" со столбцами под заголовками "3'-фиксатор" или "5'-фиксатор".
Сравнение скорости реакции у опосредованных одиночными фиксаторами и двойными фиксаторами реакций замещения цепей
[0067] В соответствии с этими данными в настоящем изобретении предоставлен ряд трансляторов и наборов трансляторов, иногда обозначаемых как "адаптеры". Транслятор или группа трансляторов принимает в качестве входа молекулу нуклеиновой кислоты (например, одноцепочечную нуклеиновую кислоту, полностью или частично двухцепочечные нуклеиновые кислоты или более сложную структуру нуклеиновой кислоты или трансляторы) и осуществляет одну или несколько реакций замещения цепей с получением одной или нескольких молекул нуклеиновой кислоты в качестве выхода. Каждый из этих входов и выходов может служить в качестве выходов или входов для других трансляторов или групп трансляторов в системе. Таким образом, каждый из этих трансляторов и наборов трансляторов функционирует в качестве логического оператора, и в комбинации они реализуют желательные вычисления и передачу информации.
[0068] Например, на ФИГ. 6B транслятор 102 в качестве входа принимает частичный дуплекс, фиксатор 101, и подвергается ряду реакций замещения цепей, что приводит к высвобождению трех различных молекул нуклеиновых кислот, дуплексов 105 и 106 и нового фиксатора 107. Теперь новый фиксатор 107 (например, дистальная/правая половина транслятора 102) может служить в качестве входа для других трансляторов. Образование дуплексов 105 и 106 из комплекса вследствие их высокой стабильности помогает проводить реакции замещения в желательном направлении до завершения.
[0069] Таким образом, в одном из вариантов осуществления в настоящем изобретении предоставлена композиция, содержащая первый и второй комплексы нуклеиновых кислот, где каждый содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот. Каждая из цепей содержит последовательно (например, от 5'-конца до 3'-конца, подряд или разделенные промежуточными нуклеотидами) первый фрагмент, второй фрагмент и третий фрагмент, где цепи нуклеиновых кислот определены как:
B-X-D, 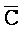 -
-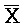 -
-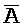 ,
, 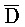 -
-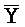 -
-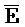 и F-Y-C для первой, второй, третьей и четвертой цепей первого комплекса, соответственно, и
и F-Y-C для первой, второй, третьей и четвертой цепей первого комплекса, соответственно, и
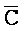 -
-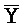 -
-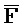 , E-Y-D, F-Z-G,
, E-Y-D, F-Z-G, 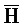 -
-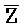 -
-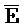 для первой, второй, третьей и четвертой цепей второго комплекса, соответственно.
для первой, второй, третьей и четвертой цепей второго комплекса, соответственно.
Каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь.
[0070] В одном из аспектов каждый из первого и второго комплексов содержит первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (X::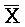 и
и 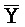 ::Y в первом и втором комплексах, соответственно), вторую область дуплекса, сформированную между вторыми фрагментами третьей и четвертой цепей (Y::
::Y в первом и втором комплексах, соответственно), вторую область дуплекса, сформированную между вторыми фрагментами третьей и четвертой цепей (Y::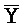 и Z::
и Z::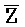 ), третью область дуплекса, сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи (D::
), третью область дуплекса, сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи (D::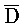 и
и 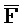 ::F), и четвертую область дуплекса, сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи (
::F), и четвертую область дуплекса, сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи (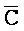 ::C и E::
::C и E::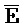 ).
).
[0071] В каждом из первого и второго комплексов первый фрагмент (B и 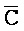 в первом и втором комплексах, соответственно) первой цепи, третий фрагмент (
в первом и втором комплексах, соответственно) первой цепи, третий фрагмент (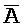 и D) второй цепи, третий фрагмент (
и D) второй цепи, третий фрагмент (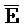 и G) третьей цепи и первый фрагмент (F и
и G) третьей цепи и первый фрагмент (F и 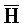 ) четвертой цепи являются одноцепочечными.
) четвертой цепи являются одноцепочечными.
[0072] Кроме того, третья цепь первого комплекса (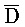 -
-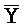 -
-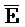 ) обладает подходящей комплементарностью последовательности со второй цепью второго комплекса (E-Y-D) для обеспечения связывания между ними в условиях гибридизации; а четвертая цепь первого комплекса (F-Y-C) обладает подходящей комплементарностью последовательности с первой цепью второго комплекса (
) обладает подходящей комплементарностью последовательности со второй цепью второго комплекса (E-Y-D) для обеспечения связывания между ними в условиях гибридизации; а четвертая цепь первого комплекса (F-Y-C) обладает подходящей комплементарностью последовательности с первой цепью второго комплекса (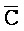 -
-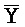 -
-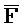 ) для обеспечения связывания между ними в условиях гибридизации.
) для обеспечения связывания между ними в условиях гибридизации.
[0073] В одном из аспектов композиция дополнительно содержит третий комплекс нуклеиновых кислот, содержащий первую и вторую цепи нуклеиновых кислот, где каждая из цепей содержит последовательно первый, второй и третий фрагменты, где первая и вторая цепи определены как E-Z-H и 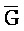 -
-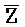 -
-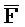 , соответственно, где третий комплекс нуклеиновых кислот содержит область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (Z::
, соответственно, где третий комплекс нуклеиновых кислот содержит область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (Z::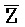 ), и первый фрагмент (E) первой цепи и третий фрагмент второй цепи (
), и первый фрагмент (E) первой цепи и третий фрагмент второй цепи (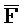 ) являются одноцепочечными, и где: третья цепь второго комплекса (F-Z-G) обладает подходящей комплементарностью последовательности со второй цепью третьего комплекса (
) являются одноцепочечными, и где: третья цепь второго комплекса (F-Z-G) обладает подходящей комплементарностью последовательности со второй цепью третьего комплекса (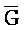 -
-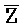 -
-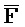 ) для обеспечения связывания между ними в условиях гибридизации; а четвертая цепь второго комплекса (
) для обеспечения связывания между ними в условиях гибридизации; а четвертая цепь второго комплекса (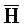 -
-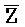 -
-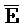 ) обладает подходящей комплементарностью последовательности с первой цепью третьего комплекса (E-Z-H) для обеспечения связывания между ними в условиях гибридизации.
) обладает подходящей комплементарностью последовательности с первой цепью третьего комплекса (E-Z-H) для обеспечения связывания между ними в условиях гибридизации.
Химические модификации для предотвращения образования замыканий фиксаторов
[0074] В настоящем изобретении также предоставлена методология предотвращения образования замыканий фиксаторов. Как показано на ФИГ. 6A, образование замыканий фиксаторов все еще может происходить, например, между фиксатором (101) и комплексом (103) вследствие доступных одноцепочечных областей C/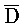 и
и  /D.
/D.
[0075] В одном из таких способов каждый фрагмент нуклеиновой кислоты, который в ходе процесса трансляции потенциально может быть доступен в виде одноцепочечного фрагмента, является объектом потенциальной химической модификации. В дополнение к его природному состоянию (состояние N, указываемое как подстрочный индекс, например, в CN), каждый такой фрагмент можно модифицировать с формированием двух состояний, состояния R (например, с правой стороны) и L (с левой стороны), соответственно, при условии, что такая модификация допускает (или способствует) и препятствует (или подавляет) связывание между комплементарными цепями в определенных состояниях, как показано в таблице 2 ниже.
Требования для связывания для химической модификации
[0076] С такими модификациями в одном из аспектов концевой фрагмент ( ) третьей цепи первого комплекса (102) обладает обратной комплементарностью последовательностей с головным фрагментом (ER) первого фрагмента первой цепи третьего комплекса (104), но стабильно с ним не связывается. Подобным образом, в другом аспекте головной фрагмент (FL) четвертой цепи первого комплекса (102) обладает обратной комплементарностью последовательностей с концевым фрагментом (
) третьей цепи первого комплекса (102) обладает обратной комплементарностью последовательностей с головным фрагментом (ER) первого фрагмента первой цепи третьего комплекса (104), но стабильно с ним не связывается. Подобным образом, в другом аспекте головной фрагмент (FL) четвертой цепи первого комплекса (102) обладает обратной комплементарностью последовательностей с концевым фрагментом ( ) первого фрагмента первой цепи третьего комплекса (104), но стабильно с ним не связывается. Таким образом, образование замыканий фиксаторов происходить не может, и такие модификации обеспечивают прохождение реакций замещения контролируемым образом. Таким образом, в одном из аспектов термин "чередуемые" используют для обозначения модификаций (L и R), которые модулируют изоляцию, вместо используемых реакций обмена.
) первого фрагмента первой цепи третьего комплекса (104), но стабильно с ним не связывается. Таким образом, образование замыканий фиксаторов происходить не может, и такие модификации обеспечивают прохождение реакций замещения контролируемым образом. Таким образом, в одном из аспектов термин "чередуемые" используют для обозначения модификаций (L и R), которые модулируют изоляцию, вместо используемых реакций обмена.
[0077] Как используют в настоящем документе, термин "стабильно не связывается" означает, что две цепи нуклеиновых кислот не формируют стабильных дуплексных структур, даже если их последовательности комплементарны. "Стабильная" дуплексная структура в определенных аспектах относится к дуплексу, который в реакционном растворе может существовать в устойчивой форме в концентрации более 0,1%, или 1%, или 5% или 10% от общей концентрации каждой отдельной цепи.
[0078] Как дополнительно описано ниже, химические модификации, посредством которых можно достигать требований для связывания, определенных в таблице 2, известны. В этом контексте "химическая модификация" не ограничена химическими модификациями молекул нуклеиновых кислот после их получения; предпочтительнее фраза также включает встраивание при получении молекулы нуклеиновой кислоты химически модифицированных нуклеотидов.
[0079] Существует множество хорошо охарактеризованных модификаций нуклеиновых кислот, которые можно использовать для модуляции термодинамических свойств связывания с природной ДНК или РНК. Они включают изменения каркаса, сахара или нуклеинового основания олигонуклеотида. Модификации также можно использовать раздельно или в сочетании друг с другом; т.е. использование модифицированного каркаса не исключает использование модифицированного нуклеинового основания в той же цепи.
[0080] Для улучшения связывания цепей, способных к участию в реакции замещения, можно использовать аналоги каркаса нуклеиновой кислоты. Существует ряд различных аналогов, которые можно использовать, которые все обеспечивают более высокие аффинности связывания с ДНК и РНК, чем природные нуклеиновые кислоты. Эти аналоги в качестве неограничивающих примеров включают аналоги с незаряженными каркасами (пептидные нуклеиновые кислоты или фосфородиамидаты), положительно заряженными каркасами (гуанидиниевые пептидные нуклеиновые кислоты) и с водородсвязывающими группами, которые допускают предорганизацию (гамма-пептидные нуклеиновые кислоты). В одном из аспектов каркасы включают конъюгированные с мини-ПЭГ, получаемые из серина гамма-ПНК, как проиллюстрировано на ФИГ. 7f-g.
[0081] На ФИГ. 7 представлены общие структуры определенных аналогов. Все эти аналоги улучшают термодинамику реакций гибридизации нуклеиновых кислот, обеспечивая более крепкое связывание фиксаторов и таким образом более быстрое замещение. Используя эти аналоги в конкретных местах логической схемы, желаемые реакции замещения можно сделать намного более предпочтительными по сравнению с замыкающими взаимодействиями, таким образом, увеличивая скорость желаемых реакций замещения цепей.
[0082] Использование модифицированных сахарных колец также может изменять термодинамику связывания олигонуклеотида с ДНК или РНК. Наиболее широко используемыми аналогами являются морфолино, закрытые нуклеиновые кислоты (ЗНК) и производные ЗНК. В литературе описаны другие модифицированные сахара, которые также могут приводить к сходному результату в отношении изменения термодинамики связывания. Их иллюстрацией являются сахара с модификациями в 1', 2', 3' или 4' положении и сахара с различными атомами, замещающими кислород в циклопентановом цикле рибозы. Эти аналоги проиллюстрированы на ФИГ. 8.
[0083] Также для достижения такого же эффекта, как и у аналогов каркаса и сахаров, а именно изменения термодинамики конкретных реакций гибридизации, можно использовать модификации нуклеиновых оснований. Эти основания включают метилцитозин, диаминопурин, G-зажим и феноксазин (ФИГ. 9), которые все улучшают аффинность связывания цепи с обратно комплементарной ей цепью. Другая существующая возможность с использованием модификаций нуклеиновых оснований включает псевдокомплементарность оснований. Этот класс аналогов оснований формирует слабые пары оснований друг с другом, но формирует сильные пары оснований со стандартными основаниями. Одной такой парой оснований является 2-аминоаденин (nA) и 2-тиотимин (sT). Эти основания можно использовать для предпочтительного связывания одной цепи, при пренебрежении другой, что является примером увеличения вероятности события продуктивного связывания при одновременном снижении вероятности замыкания.
[0084] Другой химической модификацией, которую можно использовать для изменения термодинамики взаимодействий связывания, является встраивание заряженных полимеров, таких как хитозан, для которых в литературе описано, что они ускоряют скорость реакций замещения. Однако так как эти полимеры ускоряют реакции неспецифически, то для обеспечения различия между желательными и нежелательными событиями связывания их необходимо использовать в сочетании с одной из других модификаций, указанных выше.
[0085] Для синтеза молекул с описанными выше химическими модификациями можно использовать различные подходы. Например, при конструировании модифицированных молекул можно рассматривать каркасные химические структуры. Каркасные химические структуры представляет собой то, что используют для связывания отдельных мономеров в более длинную цепь. Модификации, которые затрагивают нуклеиновое основание или сахар, но оставляют природный фосфодиэфирный каркас ДНК/РНК, можно получать посредством стандартных химических структур фосфорамидитов, как используют для природных мономеров. Иллюстрации этих способов находятся, например, в Beaucage, S., and R. Iyer, Tetrahedron 48: 2223 (1992), в Brown, D. M. A, "Brief history of oligonucleotide synthesis", 20 Methods in Molecular Biology (Protocols for Oligonucleotides and Analogs) 1-17 (1993), в Reese, Colin B., Organic & Biomolecular Chemistry 3: 3851 (2005) и в Iyer, R. P.; and S.L. Beaucage, "7.05. Oligonucleotide synthesis", 7 Comprehensive Natural Products Chemistry (DNA and Aspects of Molecular Biology) 105-52 (1999). Соответствующие содержания указанных выше публикаций таким образом полностью включены в настоящий документ в качестве ссылки.
[0086] Если каркас изменяют посредством конкретной модификации, можно использовать другие химические структуры. Такие химические структуры для получения модификаций описаны в научной литературе. Таким образом, пептидные нуклеиновые кислоты (ПНК) и их производные основаны на амидных связях, связывающих отдельные мономеры. Таким образом, вместо использования химических структур фосфорамидитов, цепи этих мономеров получают в условиях формирования амидных связей и с использованием связывающих реагентов, подобных HBTU. Исследование способов, используемых для получения олигонуклеотидов ПНК или ПНК-подобных олигонуклеотидов, можно найти, например, в F. Beck, "Solid Phase Synthesis of PNA Oligomers", Methods in Molecular Biology Series (Peptide Nucleic Acids), Humana Press, http://www.springerlink.com/content/mr571738x7t65067/.
[0087] Эти химические модификации включают введение хиральности во фрагмент ДНК или ПНК. Хиральность ДНК, в частности ПНК, является хорошо охарактеризованной характеристикой, и способы получения хиральности также известны. См., например, Corradini et al. "Control of helical handedness in DNA and PNA nanostructures", Methods Mol Biol. 749:79-92 (2011). Левовращающие ДНК или ПНК не связываются с правовращающими ДНК или ПНК, при этом оба варианта могут связываться с их природными партнерами.
[0088] Другой подход к модификации каркаса включает химерные олигонуклеотиды. Они представляют собой олигонуклеотидные цепи, которые содержат различные каркасные химические структуры в одной и той же молекуле. Например, если необходима цепь, которая наполовину представляет собой каркас ПНК и наполовину представляет собой каркас ДНК, необходим способ для связывания этих двух различных каркасных химических структур. Получение этих химерных цепей также широко известно в данной области. В приведенном выше примере химерных ПНК/ДНК различие в химических структурах можно преодолеть, используя модифицированные мономеры ДНК или ПНК. Для ДНК защищенный 5'-диметокситритилом (DMT) гидроксил замещают защищенным монометокситритилом (MMT) амином, который после снятия защиты может реагировать с карбоновой кислотой ПНК. Для ПНК защищенный N-концевой азот замещают защищенным DMT гидроксилом, который после снятия защиты может реагировать с фосфорамидитной группой на ДНК. Эти подходы дополнительно описаны, например, в E. Uhlmann et al., Angew. Chem. (Int'l ed.) 37: 2796-823 (1998).
[0089] Все эти модификации, используемые отдельно или в сочетании друг с другом, могут влиять на термодинамические условия конкретных взаимодействий в произвольной системе нуклеиновых кислот так, что связывание желательных цепей или комплексов будет предпочтительным по сравнению с замыкающими взаимодействиями без изменения состава последовательностей. Все эти взаимодействия можно использовать в любых опосредованных миграциями цепей реакциях, являются ли они трехсторонними миграциями цепей, такими как изоляция на твердой фазе, изоляция фиксаторов или замена фиксаторов, или они представляют собой миграции цепей, которые происходят по другому механизму, например, четырехсторонней миграции цепей, четырехсторонней ускоренной миграции цепей или миграции комплекса из множества цепей.
Адаптеры и наборы трансляторов
[0090] Трансляторы, предоставляемые по настоящему изобретению, можно использовать для построения компьютеров на основе нуклеиновых кислот, которые могут быть полезны в медицине. Например, трансляторы могут "распознавать" присутствие вирусной нуклеиновой кислоты, а затем осуществлять ряд актов трансляции, что приведет к "высвобождению" регуляторной РНК (например, миРНК, антисмысловой РНК), которую можно использовать для инициации противовирусного ответа или апоптоза клетки-хозяина для элиминации вируса.
I. Адаптер иРНК
[0091] "Распознание" можно реализовывать как инициацию одной или нескольких реакций замещения цепей, принимая нуклеиновую кислоту (например, вирусную ДНК/РНК, опухолевую ДНК/РНК) в качестве входа и высвобождая один или несколько фиксаторов в качестве выхода. Трансляторы, которые осуществляют одну или несколько таких реакций замещения цепей, называют "адаптерами". Например, в случае распознания опухолевой иРНК, такой адаптер можно называть "адаптер иРНК".
[0092] Адаптер иРНК проиллюстрирован на ФИГ. 10. В иРНК (201) определена уникальная последовательность, содержащая участок из фрагментов  РНК-
РНК-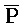 РНК или
РНК или 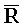 РНК-
РНК-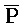 РНК-
РНК-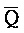 РНК. Первый комплекс нуклеиновых кислот (202) адаптера иРНК сконструирован так, чтобы содержать две цепи, одна из которых содержит комплементарную иРНК последовательность (S-Q-P). Другая цепь, также представленная на ФИГ. 10, формирует область дуплекса с первой цепью (Q-P::
РНК. Первый комплекс нуклеиновых кислот (202) адаптера иРНК сконструирован так, чтобы содержать две цепи, одна из которых содержит комплементарную иРНК последовательность (S-Q-P). Другая цепь, также представленная на ФИГ. 10, формирует область дуплекса с первой цепью (Q-P::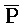 -
-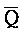 ) и содержит одноцепочечную область, A-
) и содержит одноцепочечную область, A-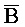 .
.
[0093] Второй комплекс в адаптере иРНК (203) содержит по меньшей мере три цепи. Первая цепь (P-B-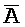 ) частично комплементарна второй цепи комплекса 202, формируя две отдельные области дуплексов (
) частично комплементарна второй цепи комплекса 202, формируя две отдельные области дуплексов (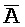 ::A и B::
::A и B::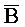 ) со второй и третьей цепями. В отсутствие первой цепи, вторая и третья цепи принимают форму двойного фиксатора, как описано выше.
) со второй и третьей цепями. В отсутствие первой цепи, вторая и третья цепи принимают форму двойного фиксатора, как описано выше.
[0094] Таким образом, в клетке, содержащей адаптер иРНК, включающий комплексы 202 и 203, при появлении молекулы иРНК 201 иРНК инициирует реакцию замещения цепей с комплексом 202, таким образом, высвобождая вторую цепь (A-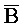 -
-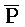 -
-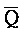 ). Затем высвободившаяся цепь взаимодействует с комплексом 203, что приводит к реакции замещения цепей с высвобождением комплекса (204), который содержит вторую и третью цепи, в форме двойного фиксатора. Затем такой двойной фиксатор (204) может инициировать дополнительные реакции замещения цепей, как описано в настоящем документе. См., например, ФИГ. 6A.
). Затем высвободившаяся цепь взаимодействует с комплексом 203, что приводит к реакции замещения цепей с высвобождением комплекса (204), который содержит вторую и третью цепи, в форме двойного фиксатора. Затем такой двойной фиксатор (204) может инициировать дополнительные реакции замещения цепей, как описано в настоящем документе. См., например, ФИГ. 6A.
[0095] Такой "адаптер иРНК" не только распознает молекулу иРНК. Он может распознавать любую одноцепочечную нуклеиновую кислоту, включая в качестве неограничивающих примеров вирусную ДНК и мутантную опухолевую ДНК.
[0096] Таким образом, настоящее изобретение относится к композиции, содержащей первый комплекс нуклеиновых кислот (например, 202 на ФИГ. 10) и второй комплекс нуклеиновых кислот (203). Первый комплекс (202) содержит (a) первую цепь, последовательно содержащую первый фрагмент (S), второй фрагмент (Q), третий фрагмент (P) и четвертый фрагмент (Q), и (b) вторую цепь, последовательно содержащую первый фрагмент (A), второй фрагмент (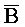 ), третий фрагмент (
), третий фрагмент (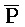 ) и четвертый фрагмент (
) и четвертый фрагмент (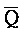 ). Первый комплекс содержит область дуплекса, сформированную между вторым и третьим фрагментами (Q-P) первой цепи и третьим и четвертым фрагментами (
). Первый комплекс содержит область дуплекса, сформированную между вторым и третьим фрагментами (Q-P) первой цепи и третьим и четвертым фрагментами (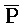 -
-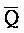 ) второй цепи.
) второй цепи.
[0097] Дополнительно второй комплекс (203) содержит (a) первую цепь, последовательно содержащую первый фрагмент (P), второй фрагмент (B) и третий фрагмент (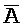 ), обладающие подходящей комплементарностью последовательностей с первым (A), вторым (
), обладающие подходящей комплементарностью последовательностей с первым (A), вторым (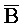 ) и третьим (
) и третьим (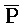 ) фрагментами второй цепи первого комплекса, соответственно, для обеспечения связывания между ними в условиях гибридизации, (b) вторую цепь, последовательно содержащую первый фрагмент (A), второй фрагмент (X) и третий фрагмент (C), и (c) третью цепь, последовательно содержащую первый фрагмент (
) фрагментами второй цепи первого комплекса, соответственно, для обеспечения связывания между ними в условиях гибридизации, (b) вторую цепь, последовательно содержащую первый фрагмент (A), второй фрагмент (X) и третий фрагмент (C), и (c) третью цепь, последовательно содержащую первый фрагмент (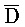 ), второй фрагмент (
), второй фрагмент (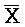 ) и третий фрагмент (
) и третий фрагмент (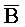 ). Второй комплекс (203) содержит первую область дуплекса, сформированную между третьим фрагментом (
). Второй комплекс (203) содержит первую область дуплекса, сформированную между третьим фрагментом (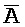 ) первой цепи и первым фрагментом (A) второй цепи, вторую область дуплекса, сформированную между вторым фрагментом (B) первой цепи и третьим фрагментом (
) первой цепи и первым фрагментом (A) второй цепи, вторую область дуплекса, сформированную между вторым фрагментом (B) первой цепи и третьим фрагментом (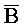 ) третьей цепи, и третью область дуплекса, сформированную между вторым фрагментом (X) второй цепи и вторым фрагментом (
) третьей цепи, и третью область дуплекса, сформированную между вторым фрагментом (X) второй цепи и вторым фрагментом (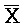 ) третьей цепи.
) третьей цепи.
[0098] Как проиллюстрировано на ФИГ. 10, предусмотрены химические модификации фрагментов нуклеиновых кислот, которые могут быть экспонированы в виде одиночных цепей, для уменьшения или устранения образования замыканий фиксаторов.
[0099] По одному из аспектов изобретения один из вариантов осуществления изобретения относится к композиции, содержащей: (a) первый комплекс нуклеиновых кислот, содержащий (i) первую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как S-Q-P, где каждая буква означает фрагмент, а последовательность букв, соединенных "-", означает цепь, и (ii) вторую цепь, последовательно содержащую первый, второй, третий и четвертый фрагменты и определенную как A-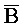 -
-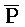 -
-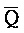 , где первый комплекс содержит область дуплекса (Q-P::
, где первый комплекс содержит область дуплекса (Q-P::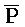 -
-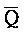 ), сформированную между вторым и третьим фрагментами первой цепи и третьим и четвертым фрагментами второй цепи; и (b) второй комплекс нуклеиновых кислот, содержащий (i) первую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как P-B-
), сформированную между вторым и третьим фрагментами первой цепи и третьим и четвертым фрагментами второй цепи; и (b) второй комплекс нуклеиновых кислот, содержащий (i) первую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как P-B-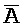 , (ii) вторую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как A-X-C, и (iii) третью цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как
, (ii) вторую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как A-X-C, и (iii) третью цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как 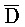 -
-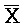 -
-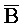 , где второй комплекс содержит первую область дуплекса (
, где второй комплекс содержит первую область дуплекса (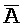 ::A), сформированную между третьим фрагментом первой цепи и первым фрагментом второй цепи, вторую область дуплекса (B::
::A), сформированную между третьим фрагментом первой цепи и первым фрагментом второй цепи, вторую область дуплекса (B::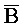 ), сформированную между первым фрагментом первой цепи и третьим фрагментом третьей цепи, и третью область дуплекса (X::
), сформированную между первым фрагментом первой цепи и третьим фрагментом третьей цепи, и третью область дуплекса (X::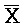 ), сформированную между вторым фрагментом второй цепи и вторым фрагментом третьей цепи, где первый (S) фрагмент первой цепи первого комплекса, первый фрагмент (A) и второй фрагмент (
), сформированную между вторым фрагментом второй цепи и вторым фрагментом третьей цепи, где первый (S) фрагмент первой цепи первого комплекса, первый фрагмент (A) и второй фрагмент (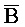 ) второй цепи первого комплекса, первый фрагмент (P) третьей цепи второго комплекса, третий фрагмент (C) второй цепи второго комплекса и первый фрагмент (
) второй цепи первого комплекса, первый фрагмент (P) третьей цепи второго комплекса, третий фрагмент (C) второй цепи второго комплекса и первый фрагмент (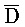 ) третьей цепи второго комплекса являются одноцепочечными; и где первый, второй и третий фрагменты (A,
) третьей цепи второго комплекса являются одноцепочечными; и где первый, второй и третий фрагменты (A, 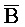 ,
, 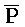 ) второй цепи первого комплекса обладают подходящей комплементарностью последовательностей с третьим, вторым и первым фрагментами (
) второй цепи первого комплекса обладают подходящей комплементарностью последовательностей с третьим, вторым и первым фрагментами (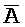 , B, P) первой цепи второго комплекса для обеспечения связывания между ними в условиях гибридизации, соответственно.
, B, P) первой цепи второго комплекса для обеспечения связывания между ними в условиях гибридизации, соответственно.
[0100] В одном из аспектов первая цепь (S-Q-P-R) первого комплекса обладает подходящей комплементарностью последовательности с фрагментом в нуклеиновой кислоте патогенной нуклеиновой кислоты для обеспечения связывания между ними в условиях гибридизации.
[0101] В одном из аспектов патогенная нуклеиновая кислота представляет собой вирусную ДНК, вирусную РНК, бактериальную ДНК, бактериальную РНК, мутантную опухолевую ДНК или опухолевую РНК.
II. Адаптер РНКи
[0102] Дополнительно предоставлен адаптер, который принимает нуклеиновую кислоту в качестве входа, и высвобождает молекулу РНКи (например, миРНК) в качестве выхода так, что миРНК после высвобождения может осуществлять определенные биологические функции. Такой адаптер проиллюстрирован на ФИГ. 11 и включает комплексы 303 и 304.
[0103] На ФИГ. 11 комплекс 301 представляет двойной фиксатор, который можно получать посредством предшествующих трансляторов и который может взаимодействовать с комплексом 302 с реакцией замещения цепей, приводящей к высвобождению нового двойного фиксатора, который содержит две цепи нуклеиновых кислот дистальной части комплекса.
[0104] Комплекс 303 сходен с трансляторами 102-104 на ФИГ. 6A, но отличается от них содержанием двух молекул РНКи ( РНК и IРНК), которые на дистальных концах связываются с одноцепочечными цепями (например, с правой стороны, как представлено на фигуре). С другой стороны, другой комплекс (304) похож на двойной фиксатор (например, фиксатор 101 на ФИГ. 6A), но отличается содержанием на дистальных концах двух молекул РНКи (
РНК и IРНК), которые на дистальных концах связываются с одноцепочечными цепями (например, с правой стороны, как представлено на фигуре). С другой стороны, другой комплекс (304) похож на двойной фиксатор (например, фиксатор 101 на ФИГ. 6A), но отличается содержанием на дистальных концах двух молекул РНКи (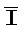 РНК и J).
РНК и J).
[0105] Таким образом, после высвобождения дистальной части (например, с правой стороны) комплекса 302 в качестве нового двойного фиксатора, новый фиксатор инициирует реакцию замещения цепей с комплексом 303, что приводит к высвобождению комплекса, содержащего дистальную часть комплекса вместе с цепями РНКи. Затем такой новый комплекс может инициировать реакцию замещения цепей с комплексом 304, что приводит к высвобождению всех молекул РНКи ( РНК, IРНК,
РНК, IРНК, 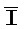 РНК и J).
РНК и J).
[0106] Таким образом, в одном из вариантов осуществления предоставлена композиция, содержащая первый комплекс нуклеиновых кислот, содержащий (a) первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты, и (b) первую и вторую цепи РНК, где первый комплекс содержит: (i) первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей; (ii) вторую область дуплекса, сформированную между вторыми фрагментами третьей и четвертой цепей; (iii) третью область дуплекса, сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи; (iv) четвертую область дуплекса, сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; (v) пятую область дуплекса, сформированную между третьим фрагментом третьей цепи и первой РНК; и (vi) шестую область дуплекса, сформированную между первым фрагментом четвертой цепи и второй РНК, и первый фрагмент (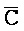 ) первой цепи и третий фрагмент (D) второй цепи являются одноцепочечными.
) первой цепи и третий фрагмент (D) второй цепи являются одноцепочечными.
[0107] В одном из аспектов композиция дополнительно содержит второй комплекс нуклеиновых кислот, содержащий первую и вторую цепи нуклеиновых кислот и первую и вторую цепи РНК, где: второй комплекс содержит первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса, сформированную между третьим фрагментом первой цепи и первой цепью РНК, и третью область дуплекса, сформированную между первым фрагментом второй цепи и второй цепью РНК, где первый фрагмент (E) первой цепи и третий фрагмент (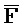 ) второй цепи являются одноцепочечными; третья цепь первого комплекса и вторая цепь второго комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации; и четвертая цепь первого комплекса и первая цепь второго комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации.
) второй цепи являются одноцепочечными; третья цепь первого комплекса и вторая цепь второго комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации; и четвертая цепь первого комплекса и первая цепь второго комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации.
III. Антисмысловой адаптер
[0108] Также предоставлен адаптер для получения сигналов от предшествующих трансляторов и высвобождения антисмысловой РНК. Со ссылкой на ФИГ. 12 комплекс 401 представляет собой типичный двойной фиксатор, как описано выше, а комплекс 402 сходен с типичным изолирующим посредством чередования транслятором (например, 102-104 на ФИГ. 6A), за исключением того, что одна из цепей необязательно содержит более длинную одноцепочечную область (F). Такая более длинная одноцепочечная область (F) способствует поддержанию стабильности транслятора 403, обеспечивая существование более короткой другой одноцепочечной области (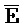 ). Таким образом, это позволяет одноцепочечной области (
). Таким образом, это позволяет одноцепочечной области (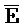 ) транслятора 402 быть достаточно короткой для минимизации нежелательной реакции между транслятором 402 и мишенью, 404.
) транслятора 402 быть достаточно короткой для минимизации нежелательной реакции между транслятором 402 и мишенью, 404.
[0109] Таким образом, после высвобождения из комплекса 402 вследствие реакции замещения цепей, инициируемой фиксатором 401, дистальная часть комплекса (402), которая представляет собой новый двойной фиксатор, взаимодействует с комплексом 403 и высвобождает частичный дуплекс, который содержит F-Z и 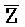 -
-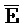 .
.
[0110] Таким образом, после контакта с клеточной молекулой РНК, содержащей фрагмент E-Z, цепь 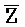 -
-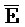 затем свободно связывается с клеточной РНК, претерпевая замещение цепей и высвобождая цепь F-Z в раствор. Связывание
затем свободно связывается с клеточной РНК, претерпевая замещение цепей и высвобождая цепь F-Z в раствор. Связывание 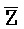 -
-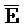 и клеточной РНК может приводить к ингибированию активности (например, трансляции и сплайсинга) клеточной РНК, где фрагмент - служит в качестве антисмысловой нуклеиновой кислоты.
и клеточной РНК может приводить к ингибированию активности (например, трансляции и сплайсинга) клеточной РНК, где фрагмент - служит в качестве антисмысловой нуклеиновой кислоты.
[0111] Уникальным преимуществом такой схемы является то, что цепь - может быть достаточно длинной. В области применения антисмысловых последовательностей известно, что чем длиннее является антисмысловая нуклеинов кислота, тем более эффективной является блокировка трансляции. Однако, к сожалению, в общепринятых технологиях более длинные антисмысловые нуклеиновые кислоты приводят к сильному побочному действию (например, токсичности). Это происходит потому, что такие антисмысловые нуклеиновые кислоты являются одноцепочечными. В настоящем изобретении предоставлена антисмысловая нуклеиновая кислота (например, цепь 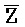 -
-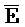 ) в форме частичного дуплекса (например, часть одной из или обе цепи являются одноцепочечными). Таким образом, пока антисмысловая нуклеиновая кислота не связана клеточной РНК-мишенью, антисмысловая нуклеиновая кислота, даже очень большой длины, содержит только очень небольшую одноцепочечную часть, выставленную в раствор. Таким образом, такая уменьшенная экспозиция снижает потенциальное побочное действие.
) в форме частичного дуплекса (например, часть одной из или обе цепи являются одноцепочечными). Таким образом, пока антисмысловая нуклеиновая кислота не связана клеточной РНК-мишенью, антисмысловая нуклеиновая кислота, даже очень большой длины, содержит только очень небольшую одноцепочечную часть, выставленную в раствор. Таким образом, такая уменьшенная экспозиция снижает потенциальное побочное действие.
[0112] Таким образом, в способе по настоящему изобретению в биологическую систему допустимо вводить антисмысловую нуклеиновую кислоту с длиной более чем, например, 30 н., 35, н., 40 н., 45 н., 50 н., 55 н., 60 н., 65 н., 70 н., 80 н., 90 или 100 н.
[0113] Таким образом, один из вариантов осуществления относится к композиции, содержащей: (a) первый комплекс нуклеиновых кислот, содержащий (i) первую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как B-X-D, (ii) вторую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как 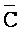 -
-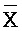 -
-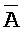 , (iii) третью цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как
, (iii) третью цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как 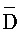 -
-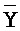 -
-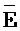 , и (iv) четвертую цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как F-Y-C, где первый комплекс содержит первую область дуплекса (X::
, и (iv) четвертую цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как F-Y-C, где первый комплекс содержит первую область дуплекса (X::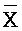 ), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (
), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (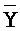 ::Y), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D::
::Y), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D::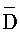 ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (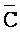 ::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; и (b) второй комплекс нуклеиновых кислот, содержащий (i) первую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как
::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; и (b) второй комплекс нуклеиновых кислот, содержащий (i) первую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как 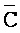 -
-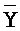 -
-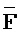 , (ii) вторую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как E-Y-D, (iii) третью цепь, последовательно содержащую первый и второй фрагменты и определенную как F-Z, и (iv) четвертую цепь, последовательно содержащую первый и второй фрагменты и определенную как
, (ii) вторую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как E-Y-D, (iii) третью цепь, последовательно содержащую первый и второй фрагменты и определенную как F-Z, и (iv) четвертую цепь, последовательно содержащую первый и второй фрагменты и определенную как 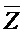 -
-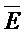 , где второй комплекс содержит первую область дуплекса (
, где второй комплекс содержит первую область дуплекса (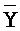 ::Y), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (Z::
::Y), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (Z::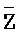 ), сформированную между вторым фрагментом третьей цепи и первым фрагментом четвертой цепи, третью область дуплекса (
), сформированную между вторым фрагментом третьей цепи и первым фрагментом четвертой цепи, третью область дуплекса (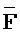 ::F), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (E::
::F), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (E::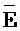 ), сформированную между первым фрагментом второй цепи и вторым фрагментом четвертой цепи, где первый (B) фрагмент первой цепи, третий фрагмент (
), сформированную между первым фрагментом второй цепи и вторым фрагментом четвертой цепи, где первый (B) фрагмент первой цепи, третий фрагмент (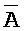 ) второй цепи, третий фрагмент (
) второй цепи, третий фрагмент (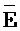 ) третьей цепи и первый фрагмент (F) четвертой цепи первого комплекса и первый (
) третьей цепи и первый фрагмент (F) четвертой цепи первого комплекса и первый (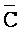 ) фрагмент первой цепи и третий фрагмент второй цепи (D) второго комплекса являются одноцепочечными; где первый, второй и третий фрагменты (
) фрагмент первой цепи и третий фрагмент второй цепи (D) второго комплекса являются одноцепочечными; где первый, второй и третий фрагменты (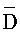 ,
, 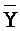 и
и 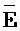 ) третьей цепи первого комплекса обладают подходящей комплементарностью последовательностей с третьим, вторым и первым фрагментами (D, Y, и E) второй цепи второго комплекса для обеспечения связывания между ними в условиях гибридизации, соответственно; и где третий, второй и первый фрагменты (C, Y и F) четвертой цепи первого комплекса обладают подходящей комплементарностью последовательностей с первым, вторым и третьим фрагментами (
) третьей цепи первого комплекса обладают подходящей комплементарностью последовательностей с третьим, вторым и первым фрагментами (D, Y, и E) второй цепи второго комплекса для обеспечения связывания между ними в условиях гибридизации, соответственно; и где третий, второй и первый фрагменты (C, Y и F) четвертой цепи первого комплекса обладают подходящей комплементарностью последовательностей с первым, вторым и третьим фрагментами (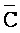 ,
, 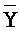 и
и 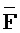 ) первой цепи второго комплекса для обеспечения связывания между ними в условиях гибридизации, соответственно.
) первой цепи второго комплекса для обеспечения связывания между ними в условиях гибридизации, соответственно.
[0114] В определенных аспектах первый фрагмент (F) четвертой цепи первого комплекса по меньшей мере на 5 нуклеотидов (н.) или 6 н., 7 н., 8 н., 9 н., 10 н., 15 н., 20 н. длиннее, чем третий фрагмент (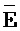 ) третьей цепи первого комплекса. В определенных аспектах длина первого фрагмента (
) третьей цепи первого комплекса. В определенных аспектах длина первого фрагмента (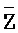 ) четвертой цепи второго комплекса составляет по меньшей мере 30 нуклеотидов (н.) или 35 н., 40 н., 45 н., 50 н., 55 н., 60 н., 65 н., 70 н. или 80 н.
) четвертой цепи второго комплекса составляет по меньшей мере 30 нуклеотидов (н.) или 35 н., 40 н., 45 н., 50 н., 55 н., 60 н., 65 н., 70 н. или 80 н.
IV. Объединение на входе
[0115] Также предоставлен набор трансляторов с "объединением на входе". Набор трансляторов с объединением на входе может принимать на входе две или более различных нуклеиновых кислот в качестве входного сигнала и продуцировать один и тот же выход. Другими словами, набор трансляторов с объединением на входе может осуществлять операцию OR. Набор с объединением на входе проиллюстрирован на ФИГ. 13.
[0116] В иллюстрируемом наборе трансляторов с объединением на входе каждый из трансляторов 502-503 и 505-506 сходен с 102-103 на ФИГ. 6A и сходен друг с другом. Вследствие различия их последовательностей, интерференция между ними отсутствует. Однако у трансляторов 503 и 506 присутствует одинаковая дистальная часть (F-Z-G/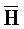 -
-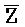 -
-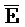 ). Это означает, что после ряда реакций замещения цепей, проходящих в предшествующих трансляторах, трансляторы 503 и 506 продуцируют одинаковый выходной фиксатор, который взаимодействует с комплексом 507.
). Это означает, что после ряда реакций замещения цепей, проходящих в предшествующих трансляторах, трансляторы 503 и 506 продуцируют одинаковый выходной фиксатор, который взаимодействует с комплексом 507.
[0117] Таким образом, каким бы ни был фиксатор, 501 или 504, будет инициирована реакция замещения цепей в трансляторе 507.
[0118] Таким образом, в одном из вариантов осуществления предоставлена композиция, содержащая первый и второй комплексы нуклеиновых кислот, где каждый содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая из цепей содержит последовательно первый, второй и третий фрагменты, где цепи нуклеиновых кислот определены как: B-X-D, 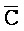 -
-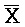 -
-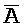 ,
, 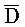 -
-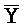 -
-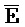 и F-Y-C для первой, второй, третьей и четвертой цепей первого комплекса, соответственно, и
и F-Y-C для первой, второй, третьей и четвертой цепей первого комплекса, соответственно, и 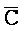 -
-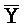 -
-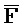 , E-Y-D, F-Z-G,
, E-Y-D, F-Z-G, 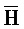 -
-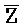 -
-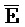 для первой, второй, третьей и четвертой цепей второго комплекса, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где: каждый из первого и второго комплексов содержит первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (X::
для первой, второй, третьей и четвертой цепей второго комплекса, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где: каждый из первого и второго комплексов содержит первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (X::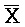 и
и 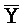 ::Y в первом и втором комплексах, соответственно), вторую область дуплекса, сформированную между вторыми фрагментами третьей и четвертой цепей (Y::
::Y в первом и втором комплексах, соответственно), вторую область дуплекса, сформированную между вторыми фрагментами третьей и четвертой цепей (Y::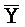 и Z::
и Z::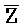 ), третью область дуплекса, сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи (D::
), третью область дуплекса, сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи (D::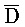 и
и 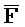 ::F), и четвертую область дуплекса, сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи (
::F), и четвертую область дуплекса, сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи (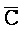 ::C и E::
::C и E::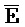 ); в каждом из первого и второго комплексов первый фрагмент (B и
); в каждом из первого и второго комплексов первый фрагмент (B и 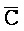 в первом и втором комплексах, соответственно) первой цепи, третий фрагмент (
в первом и втором комплексах, соответственно) первой цепи, третий фрагмент (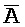 и D) второй цепи, третий фрагмент (
и D) второй цепи, третий фрагмент (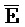 и G) третьей цепи и первый фрагмент (F и
и G) третьей цепи и первый фрагмент (F и 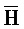 ) четвертой цепи являются одноцепочечными; третья цепь первого комплекса (
) четвертой цепи являются одноцепочечными; третья цепь первого комплекса (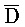 -
-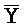 -
-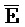 ) обладает подходящей комплементарностью последовательности со второй цепью второго комплекса (E-Y-D) для обеспечения связывания между ними в условиях гибридизации; и четвертая цепь первого комплекса (F-Y-C) обладает подходящей комплементарностью последовательности с первой цепью второго комплекса (
) обладает подходящей комплементарностью последовательности со второй цепью второго комплекса (E-Y-D) для обеспечения связывания между ними в условиях гибридизации; и четвертая цепь первого комплекса (F-Y-C) обладает подходящей комплементарностью последовательности с первой цепью второго комплекса (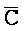 -
-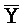 -
-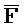 ) для обеспечения связывания между ними в условиях гибридизации; дополнительно содержит третий комплекс нуклеиновых кислот, содержащий первую и вторую цепи нуклеиновых кислот, где каждая из цепей содержит последовательно первый, второй и третий фрагменты, где первая и вторая цепи определены как E-Z-H и
) для обеспечения связывания между ними в условиях гибридизации; дополнительно содержит третий комплекс нуклеиновых кислот, содержащий первую и вторую цепи нуклеиновых кислот, где каждая из цепей содержит последовательно первый, второй и третий фрагменты, где первая и вторая цепи определены как E-Z-H и 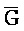 -
-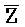 -
-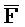 , соответственно, где третий комплекс нуклеиновых кислот содержит область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (Z::
, соответственно, где третий комплекс нуклеиновых кислот содержит область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (Z::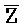 ), и первый фрагмент (E) первой цепи и третий фрагмент второй цепи (
), и первый фрагмент (E) первой цепи и третий фрагмент второй цепи (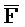 ) являются одноцепочечными, и где: третья цепь второго комплекса (F-Z-G) обладает подходящей комплементарностью последовательности со второй цепью третьего комплекса (
) являются одноцепочечными, и где: третья цепь второго комплекса (F-Z-G) обладает подходящей комплементарностью последовательности со второй цепью третьего комплекса (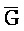 -
-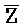 -
-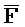 ) для обеспечения связывания между ними в условиях гибридизации; и четвертая цепь второго комплекса (
) для обеспечения связывания между ними в условиях гибридизации; и четвертая цепь второго комплекса (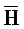 -
-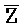 -
-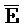 ) обладает подходящей комплементарностью последовательности с первой цепью третьего комплекса (E-Z-H) для обеспечения связывания между ними в условиях гибридизации; и еще дополнительно содержит четвертый и пятый комплексы нуклеиновых кислот, где каждый комплекс содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая из этих цепей последовательно содержит первый, второй и третий фрагменты, где цепи определены как: K-R-M,
) обладает подходящей комплементарностью последовательности с первой цепью третьего комплекса (E-Z-H) для обеспечения связывания между ними в условиях гибридизации; и еще дополнительно содержит четвертый и пятый комплексы нуклеиновых кислот, где каждый комплекс содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая из этих цепей последовательно содержит первый, второй и третий фрагменты, где цепи определены как: K-R-M, 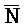 -
-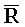 -
- ,
, 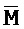 -
-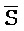 -
-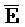 и F-S-N для первой, второй, третьей и четвертой цепей четвертого комплекса, соответственно; и
и F-S-N для первой, второй, третьей и четвертой цепей четвертого комплекса, соответственно; и 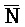 -
-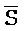 -
-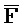 , E-S-M, F-Z-G и
, E-S-M, F-Z-G и 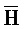 -
-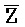 -
-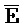 для первой, второй, третьей и четвертой цепей пятого комплекса, соответственно, где: каждый из четвертого и пятого комплексов содержит первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (R::
для первой, второй, третьей и четвертой цепей пятого комплекса, соответственно, где: каждый из четвертого и пятого комплексов содержит первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей (R::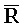 и
и 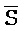 ::S, в четвертом и пятом комплексах, соответственно), вторую область дуплекса, сформированную между вторыми фрагментами третьей и четвертой цепей (
::S, в четвертом и пятом комплексах, соответственно), вторую область дуплекса, сформированную между вторыми фрагментами третьей и четвертой цепей (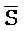 ::S и Z::
::S и Z::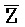 ), третью область дуплекса, сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи (M::
), третью область дуплекса, сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи (M::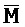 и
и 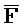 ::F), и четвертую область дуплекса, сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи (
::F), и четвертую область дуплекса, сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи (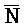 ::N и E::
::N и E::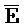 ); в каждом из четвертого и пятого комплексов первый фрагмент (K и
); в каждом из четвертого и пятого комплексов первый фрагмент (K и 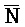 в первом и втором комплексах, соответственно) первой цепи, третий фрагмент (
в первом и втором комплексах, соответственно) первой цепи, третий фрагмент ( и M) второй цепи, третий фрагмент (
и M) второй цепи, третий фрагмент (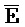 и G) третьей цепи и первый фрагмент (F и
и G) третьей цепи и первый фрагмент (F и 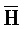 ) четвертой цепи являются одноцепочечными; третья цепь четвертого комплекса (
) четвертой цепи являются одноцепочечными; третья цепь четвертого комплекса (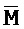 -
-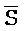 -
-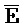 ) обладает подходящей комплементарностью последовательности со второй цепью пятого комплекса (E-S-M) для обеспечения связывания между ними в условиях гибридизации; четвертая цепь четвертого комплекса (F-S-N) обладает подходящей комплементарностью последовательности с первой цепью пятого комплекса (
) обладает подходящей комплементарностью последовательности со второй цепью пятого комплекса (E-S-M) для обеспечения связывания между ними в условиях гибридизации; четвертая цепь четвертого комплекса (F-S-N) обладает подходящей комплементарностью последовательности с первой цепью пятого комплекса (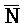 -
-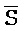 -
-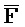 ) для обеспечения связывания между ними в условиях гибридизации; третья цепь второго комплекса (F-Z-G) и третья цепь пятого комплекса (F-Z-G) обладают подходящей идентичностью последовательности для обеспечения их связывания с одной и той же нуклеиновой кислотой-мишенью в условиях гибридизации; и четвертая цепь второго комплекса (
) для обеспечения связывания между ними в условиях гибридизации; третья цепь второго комплекса (F-Z-G) и третья цепь пятого комплекса (F-Z-G) обладают подходящей идентичностью последовательности для обеспечения их связывания с одной и той же нуклеиновой кислотой-мишенью в условиях гибридизации; и четвертая цепь второго комплекса (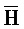 -
-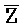 -
-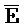 ) и четвертая цепь пятого комплекса (
) и четвертая цепь пятого комплекса (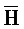 -
-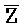 -
-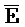 ) обладают подходящей идентичностью последовательности для обеспечения их связывания с одной и той же нуклеиновой кислотой-мишенью в условиях гибридизации.
) обладают подходящей идентичностью последовательности для обеспечения их связывания с одной и той же нуклеиновой кислотой-мишенью в условиях гибридизации.
V. Разделение на выходе
[0119] В противоположность набору трансляторов с объединением на входе, набор трансляторов с "разделением на выходе" принимает один вход и продуцирует несколько выходящих нуклеиновых кислот. В одном из аспектов несколько выходящих нуклеиновых кислот обладают различными последовательностями так, что они могут осуществлять различные функции или последующие реакции. В другом аспекте несколько выходящих нуклеиновых кислот являются идентичными, и таким образом набор трансляторов с разделением на выходе служит в качестве амплификатора.
[0120] На ФИГ. 14A проиллюстрирован набор трансляторов с разделением на выходе, который принимает один фиксатор в качестве входа и продуцирует два новых идентичных фиксатора. Подобным образом, набор с разделением на выходе на ФИГ. 14B продуцирует четыре новых идентичных фиксатора. С другой стороны, на ФИГ. 14C-D представлены наборы трансляторов с разделением на выходе, которые продуцируют два или более новых различных фиксаторов.
[0121] На ФИГ. 14A нуклеиновая кислота 601 представляет собой типичный двойной фиксатор, как описано в настоящем документе. Комплекс 602 отличается от трансляторов 102 и 103 на ФИГ. 6A тем, что одноцепочечные области на дистальном конце являются более длинными и содержат два, а не один, последовательных фрагмента. Как видно в комплексе 603, каждый набор последовательных фрагментов (A-B) способен высвобождать новый фиксатор, который после такого высвобождения содержит две отдельные одноцепочечные области, A и B, на двух отдельных цепях. Таким образом, любой из этих новых фиксаторов может инициировать новую реакцию замещения цепей с транслятором 604.
[0122] Трансляторы на ФИГ. 14B являются сходными с трансляторами на ФИГ. 14A, за исключением того, что каждая из одноцепочечных областей в трансляторе 606 содержит четыре последовательных фрагмента, способных к высвобождению двух новых фиксаторов.
[0123] Набор трансляторов на ФИГ. 14C может на входе принимать тот же фиксатор (609), что и на ФИГ. 14A-B (601 и 605), но транслятор 610 после контакта с входящей последовательностью высвобождает фиксатор, отличающийся от фиксатора на ФИГ. 14A-B. Такое различие является более очевидным, как отражено в соответствующих цепях в трансляторе 611. Т.е., после контакта с высвобожденным из транслятора 610 фиксатором и реакций замещения цепей, транслятор 611 высвобождает два различных фиксатора. Один из этих двух фиксаторов способен инициировать новую реакцию замещения цепей с транслятором 612, а другой - с транслятором 613. Таким образом, набор трансляторов на ФИГ. 14B способен порождать из одного входящего фиксатора (609) два новых различных выходящих фиксатора, с трансляторов 612 и 613, соответственно.
[0124] Подобно набору трансляторов на ФИГ. 14C, набор трансляторов на ФИГ. 14D также может принимать один фиксатор (614) в качестве входа и продуцировать четыре новых различных фиксатора в качестве выходов, где каждый высвобождается с трансляторов 617-620, соответственно.
[0125] Таким образом, в одном из вариантов осуществления изобретения предоставлен комплекс нуклеиновых кислот, содержащий первую, вторую, третью, четвертую, пятую и шестую цепи нуклеиновых кислот, где: первая цепь последовательно содержит первый, второй, третий и четвертый фрагменты и определена как 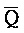 -
-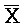 -
-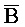 -
-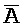 , где каждая буква означает фрагмент, а последовательность букв, соединенных "-", означает цепь; вторая цепь последовательно содержит первый, второй, третий и четвертый фрагменты и определена как
, где каждая буква означает фрагмент, а последовательность букв, соединенных "-", означает цепь; вторая цепь последовательно содержит первый, второй, третий и четвертый фрагменты и определена как 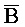 -
-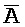 -
-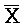 -P; каждая из третьей и пятой цепей последовательно содержит первый, второй и третий фрагменты и определена как A-Y-C; и каждая из четвертой и шестой цепей последовательно содержит первый, второй и третий фрагменты и определена как
-P; каждая из третьей и пятой цепей последовательно содержит первый, второй и третий фрагменты и определена как A-Y-C; и каждая из четвертой и шестой цепей последовательно содержит первый, второй и третий фрагменты и определена как 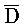 -
-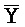 -B; и где комплекс содержит: первую область дуплекса (
-B; и где комплекс содержит: первую область дуплекса (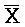 :X), сформированную между вторым фрагментом первой цепи и третьей цепью второй цепи; вторую область дуплекса (
:X), сформированную между вторым фрагментом первой цепи и третьей цепью второй цепи; вторую область дуплекса (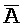 ::A), сформированную между четвертым фрагментом первой цепи и первой цепью третьей цепи; третью область дуплекса (
::A), сформированную между четвертым фрагментом первой цепи и первой цепью третьей цепи; третью область дуплекса (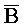 ::B), сформированную между третьим фрагментом первой цепи и третьим фрагментом четвертой цепи; четвертую область дуплекса (
::B), сформированную между третьим фрагментом первой цепи и третьим фрагментом четвертой цепи; четвертую область дуплекса (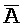 ::A), сформированную между вторым фрагментом второй цепи и первым фрагментом пятой цепи; пятую область дуплекса (
::A), сформированную между вторым фрагментом второй цепи и первым фрагментом пятой цепи; пятую область дуплекса (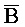 ::B), сформированную между первым фрагментом второй цепи и третьим фрагментом шестой цепи; шестую область дуплекса (Y::
::B), сформированную между первым фрагментом второй цепи и третьим фрагментом шестой цепи; шестую область дуплекса (Y::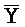 ), сформированную между вторыми фрагментами третьей и четвертой цепей; и седьмую область дуплекса (Y::
), сформированную между вторыми фрагментами третьей и четвертой цепей; и седьмую область дуплекса (Y::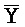 ), сформированную между вторыми фрагментами шестой и седьмой цепей, и где первый фрагмент (
), сформированную между вторыми фрагментами шестой и седьмой цепей, и где первый фрагмент (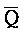 ) первой цепи, четвертый фрагмент (P) второй цепи, третий фрагменты (C) третьей и пятой цепей и первые фрагменты (
) первой цепи, четвертый фрагмент (P) второй цепи, третий фрагменты (C) третьей и пятой цепей и первые фрагменты (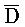 ) четвертой и шестой цепей являются одноцепочечными.
) четвертой и шестой цепей являются одноцепочечными.
[0126] Подобным образом, также предоставлен комплекс нуклеиновых кислот, который разветвляет два или более различных фиксаторов.
VI. Зависимый от соединения
[0127] "Зависимый от соединения" набор трансляторов осуществляет операцию AND. Зависимому от соединения набору для активации и высвобождения выходящей нуклеиновой кислоты необходимо два отдельных входных сигнала.
[0128] На ФИГ. 15 проиллюстрирован зависимый от соединения набор трансляторов, который содержит по меньшей мере три трансляторных комплекса нуклеиновых кислот 703, 704 и 705. Транслятор 703 в качестве входа принимает фиксатор 701 и продуцирует первый новый фиксатор, содержащий дистальную часть 703. Подобным образом, транслятор 704 принимает в качестве входа фиксатор 702 и продуцирует второй новый фиксатор, содержащий дистальную часть 704.
[0129] Транслятор 705 содержит по меньшей мере шесть цепей, составляющих слева направо три дуплексных пары. Только после контакта с первым новым фиксатором, высвобождаемым из транслятора 703, транслятор 705 может претерпевать реакцию замещения цепей, приводящую к экспозиции двух одноцепочечных областей (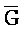 и H) из середины дуплекса.
и H) из середины дуплекса.
[0130] Затем, после контакта со вторым новым фиксатором, продуцируемым транслятором 704, происходит другая реакция замещения цепей и высвобождает третью дуплексную пару 705 в качестве третьего нового фиксатора. Затем этот третий новый фиксатор необязательно может инициировать дальнейшие реакции, например, посредством транслятора 706.
[0131] Таким образом, в одном из вариантов осуществления предоставлена композиция, содержащая первый, второй и третий комплексы нуклеиновых кислот, где: первый комплекс содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как B-W-D, 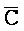 -
-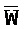 -
-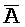 ,
, 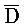 -
-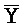 -H и
-H и 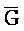 -Y-C, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где первый комплекс содержит первую область дуплекса (W::
-Y-C, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где первый комплекс содержит первую область дуплекса (W::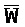 ), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (
), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (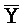 ::Y), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D::
::Y), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D::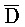 ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи; и четвертую область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи; и четвертую область дуплекса (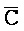 ::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; второй комплекс содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как F-X-H,
::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; второй комплекс содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как F-X-H, 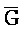 -
-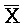 -
-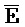 ,
, 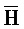 -
-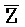 -
-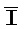 и J-Z-G, соответственно, где второй комплекс содержит первую область дуплекса (X::
и J-Z-G, соответственно, где второй комплекс содержит первую область дуплекса (X::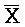 ), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (
), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (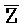 ::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (H::
::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (H::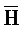 ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи; и четвертую область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи; и четвертую область дуплекса (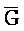 ::G), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; третий комплекс содержит первую, вторую, третью, четвертую, пятую и шестую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как
::G), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; третий комплекс содержит первую, вторую, третью, четвертую, пятую и шестую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как 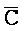 -
-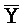 -G,
-G, 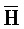 -Y-D,
-Y-D, 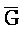 -
-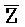 -
- , I-Z-H, J-Q-M и
, I-Z-H, J-Q-M и 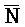 -
-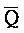 -
-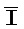 , соответственно, где третий комплекс содержит первую область дуплекса (
, соответственно, где третий комплекс содержит первую область дуплекса (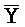 ::Y), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (
::Y), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (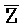 ::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (Q::
::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (Q::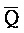 ), сформированную между вторыми фрагментами пятой и шестой цепей, четвертую область дуплекса (G::
), сформированную между вторыми фрагментами пятой и шестой цепей, четвертую область дуплекса (G::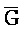 ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи; пятую область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи; пятую область дуплекса (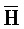 ::H), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи, шестую область дуплекса (
::H), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи, шестую область дуплекса ( ::J), сформированную между третьим фрагментом третьей цепи и первым фрагментом пятой цепи, и седьмую область дуплекса (I::
::J), сформированную между третьим фрагментом третьей цепи и первым фрагментом пятой цепи, и седьмую область дуплекса (I::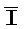 ), сформированную между первым фрагментом четвертой цепи и третьим фрагментом шестой цепи; где в первом комплексе первый фрагмент (B) первой цепи, третий фрагмент (
), сформированную между первым фрагментом четвертой цепи и третьим фрагментом шестой цепи; где в первом комплексе первый фрагмент (B) первой цепи, третий фрагмент (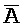 ) второго фрагмента, третий фрагмент (H) третьей цепи и первый фрагмент (
) второго фрагмента, третий фрагмент (H) третьей цепи и первый фрагмент (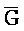 ) четвертой цепи являются одноцепочечными; во втором комплексе первый фрагмент (F) первой цепи, третий фрагмент (
) четвертой цепи являются одноцепочечными; во втором комплексе первый фрагмент (F) первой цепи, третий фрагмент (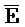 ) второго фрагмента, третий фрагмент (
) второго фрагмента, третий фрагмент (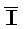 ) третьей цепи и первый фрагмент (J) четвертой цепи являются одноцепочечными; и в третьем комплексе первый фрагмент (
) третьей цепи и первый фрагмент (J) четвертой цепи являются одноцепочечными; и в третьем комплексе первый фрагмент (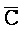 ) первой цепи, третий (D) фрагмент второй цепи, третий фрагмент (M) пятого фрагмента и первый фрагмент (
) первой цепи, третий (D) фрагмент второй цепи, третий фрагмент (M) пятого фрагмента и первый фрагмент (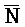 ) шестой цепи являются одноцепочечными, и где: третья цепь (
) шестой цепи являются одноцепочечными, и где: третья цепь (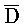 -
-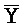 -H) первого комплекса и вторая цепь (
-H) первого комплекса и вторая цепь (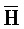 -Y-D) третьего комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации; четвертая цепь (
-Y-D) третьего комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации; четвертая цепь (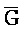 -Y-C) первого комплекса и первая цепь (
-Y-C) первого комплекса и первая цепь (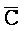 -
-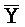 -G) третьего комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации; третья цепь (
-G) третьего комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации; третья цепь (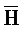 -
-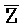 -
-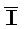 ) второго комплекса и четвертая цепь (I-Z-H) третьего комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации; и четвертая цепь (J-Z-G) второго комплекса и третья цепь (
) второго комплекса и четвертая цепь (I-Z-H) третьего комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации; и четвертая цепь (J-Z-G) второго комплекса и третья цепь (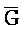 -
-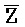 -
- ) третьего комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации.
) третьего комплекса обладают подходящей комплементарностью последовательностей для обеспечения связывания между ними в условиях гибридизации.
VII. Инвертер
[0132] "Инвертер" состоит из одного или нескольких трансляторов, которые после контакта с входящим сигналом продуцируют нуклеиновую кислоту ("затвор"), которая блокирует реакцию замещения цепей, которая в ином случае происходит в системе при инициации другим входящим сигналом.
[0133] Как проиллюстрировано на ФИГ. 16, инвертерный комплекс нуклеиновых кислот 804 состоит из трех цепей, две из которых, F-Y-D и 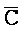 -
-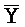 -
-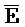 , формируют область дуплекса между средними фрагментами (Y::
, формируют область дуплекса между средними фрагментами (Y::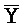 ). Третья цепь содержит два фрагмента, C и
). Третья цепь содержит два фрагмента, C и 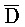 , каждый из которых комплементарен одному фрагменту двух других цепей. После контакта с фиксатором 802 происходит реакция замещения цепей, которая приводит к высвобождению третьей цепи, C-
, каждый из которых комплементарен одному фрагменту двух других цепей. После контакта с фиксатором 802 происходит реакция замещения цепей, которая приводит к высвобождению третьей цепи, C-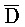 .
.
[0134] Комплекс нуклеиновых кислот 803 представляет собой транслятор, который в качестве входа принимает фиксатор 801 и в качестве выхода продуцирует дистальную часть комплекса, новый фиксатор. Если этот новый фиксатор контактирует с транслятором 805, можно осуществлять последующую реакцию замещения цепей с потенциальной последующей реакцией с транслятором 806.
[0135] Однако этот путь обработки сигнала 801-803-805-806 не может проходить, если перед тем, как произойдет замещение цепей между фиксатором 801 и транслятором 803, фиксатор 802 придет в контакт с инвертером 804 и высвободит цепь-затвор C-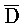 . Это происходит потому, что цепь-затвор C-
. Это происходит потому, что цепь-затвор C-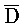 связывается с доступными одноцепочечными областями,
связывается с доступными одноцепочечными областями, 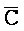 и D, таким образом блокируя реакцию замещения цепей между 805 и фиксатором, продуцируемым с транслятора 803.
и D, таким образом блокируя реакцию замещения цепей между 805 и фиксатором, продуцируемым с транслятора 803.
[0136] Таким образом, в одном из вариантов осуществления предоставлена композиция, содержащая первый и второй комплексы нуклеиновых кислот, где: первый комплекс содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как B-X-D, 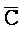 -
-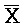 -
-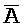 ,
, 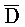 -
-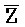 -
-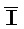 и J-Z-C, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где первый комплекс содержит первую область дуплекса (X::
и J-Z-C, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где первый комплекс содержит первую область дуплекса (X::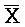 ), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (
), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (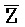 ::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D::
::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D::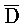 ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (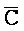 ::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; и второй комплекс содержит первую и вторую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как F-Y-D и
::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; и второй комплекс содержит первую и вторую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как F-Y-D и 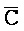 -
-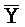 -
-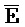 , соответственно, и третья нуклеиновая кислота последовательно содержит первый и второй фрагменты и определена как
, соответственно, и третья нуклеиновая кислота последовательно содержит первый и второй фрагменты и определена как 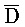 -C, где второй комплекс содержит первую область дуплекса (Y::
-C, где второй комплекс содержит первую область дуплекса (Y:: ), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (D::
), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (D::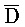 ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и третью область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и третью область дуплекса (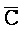 ::C), сформированную между первым фрагментом второй цепи и вторым фрагментом третьей цепи; где первый фрагмент (B) первой цепи первого комплекса, третий фрагмент (
::C), сформированную между первым фрагментом второй цепи и вторым фрагментом третьей цепи; где первый фрагмент (B) первой цепи первого комплекса, третий фрагмент (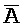 ) второй цепи первого комплекса, третий фрагмент (
) второй цепи первого комплекса, третий фрагмент (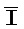 ) третьей цепи первого комплекса, первый фрагмент (J) четвертой цепи первого комплекса, первый фрагмент (F) первой цепи второго комплекса и третий фрагмент (
) третьей цепи первого комплекса, первый фрагмент (J) четвертой цепи первого комплекса, первый фрагмент (F) первой цепи второго комплекса и третий фрагмент ( ) второй цепи второго комплекса являются одноцепочечными; и где: третья цепь (
) второй цепи второго комплекса являются одноцепочечными; и где: третья цепь ( -C) второго комплекса и комбинация первого фрагмента (
-C) второго комплекса и комбинация первого фрагмента ( ) третьей цепи первого комплекса и третьего фрагмента (C) четвертой цепи первого комплекса, последовательно, обладают подходящей идентичностью последовательности для осуществления их связывания с одной и той же последовательностью нуклеиновой кислоты-мишени.
) третьей цепи первого комплекса и третьего фрагмента (C) четвертой цепи первого комплекса, последовательно, обладают подходящей идентичностью последовательности для осуществления их связывания с одной и той же последовательностью нуклеиновой кислоты-мишени.
[0137] Для любого из указанных выше трансляторов и наборов трансляторов предусмотрено, что химические модификации можно использовать для уменьшения или устранения образования замыканий фиксаторов, т.е. непредусмотренного связывания фиксаторов и одноцепочечных областей любых трансляторов в системе, таких как клетка. Такие модификации можно легко спроектировать с использованием предоставляемой в этом описании информации. Кроме того, для каждого набора трансляторов, на соответствующей иллюстративной фигуре предоставлены иллюстративные модификации, как отмечено подстрочными индексами N, R и L. Желаемые свойства таких модификаций приведены в таблице 2.
[0138] В определенных аспектах следует отметить, что если фрагменты нуклеиновых кислот конкретно не указаны в качестве части области дуплекса, их следует считать одноцепочечными. Признаки одноцепочечности также видны на сопровождающих фигурах.
[0139] Длину каждой из цепей нуклеиновых кислот или фрагментов можно определять посредством вычислений или экспериментально. В таблице 1 проиллюстрировано влияние такой длины, в частности на фиксаторы. В одном из аспектов длина каждого из фрагментов приблизительно составляет от 3 оснований до приблизительно 50 оснований. Альтернативно, длина фрагментов составляет по меньшей мере приблизительно 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 оснований. В другом аспекте длина фрагментов составляет не более чем приблизительно 50, 45, 40, 35, 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 28, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6 или 5 оснований. Также фрагменты, которые существуют в одноцепочечных формах, такие как фиксаторы, могут быть относительно короче, например, от приблизительно 3, 4, 5, 6, 7, 8, 9, 10 оснований до приблизительно 30, 29, 28, 27, 26, 25, 24, 23, 22, 21, 20, 19, 28, 17, 16 или 15 оснований. В одном из аспектов длина фрагмента фиксатора составляет по меньшей мере приблизительно 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 оснований. В другом аспекте длина фрагмента фиксатора составляет не более чем приблизительно 20, 19, 28, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6 или 5 оснований. В отличие от этого, фрагменты, которые формируют области дуплексов в трансляторе, могут быть относительно длиннее.
Способы и компьютерное моделирование
[0140] В настоящем изобретении предоставлены способы использования трансляторов по настоящему изобретению для осуществления обработки информации и, в частности, передачи информации в биологическом окружении. Например, адаптер иРНК можно использовать для детекции присутствия патогенной или неопластической нуклеиновой кислоты и продуцировать выходящий сигнал, например, в форме фиксатора. Если необходимо, для обеспечения надежной детекции можно использовать несколько адаптеров иРНК со специфичностью к нескольким нуклеиновым кислотам.
[0141] Затем можно использовать другие трансляторы в системе, такие как трансляторы с объединением на входе, с разделением на выходе и инвертеры для обработки сигналов, генерируемых адаптерами иРНК, что, в конечном счете, приводит к продукции регуляторных нуклеиновых кислот, например, РНКи и антисмысловых РНК, с соответствующих адаптеров РНКи и антисмысловых адаптеров. Посредством тщательно подобранной обработки можно точно контролировать точные последовательности и количества этих регуляторных РНК. Таким образом, этими регуляторными РНК можно инициировать биологические реакции, такие как апоптоз и иммунный ответ, обеспечивая терапевтическую или диагностическую пользу.
[0142] Можно легко понять, что трансляторы, композиции и системы, описанные в настоящем документе, могут включать биологические материалы, но их также можно моделировать математически на компьютере. Таким образом, в настоящем изобретении также предоставлены компьютерные способы, системы и содержащие программный код носители для осуществления такого моделирования.
[0143] Таким образом, предоставлен реализованный на компьютере способ, соответствующий каждому из трансляторов, композиций или систем в настоящем описании, в котором компьютер сконфигурирован для представления их последовательности, структуры, синтеза и реакции замещения цепей.
[0144] Описанную в настоящем документе методологию можно реализовывать в компьютерной системе или сети. Подходящая компьютерная система может включать по меньшей мере процессор и память; необязательно, машиночитаемый носитель, который хранит компьютерный код для выполнения процессором. После выполнения кода компьютерная система выполняет описанные способы.
[0145] В этом отношении "процессор" представляет собой электронную схему, которая может выполнять компьютерные программы. Подходящие процессоры иллюстрируются, но не ограничены центральными процессорами, микропроцессорами, графическими процессорами, физическими процессорами, процессорами цифровых сигналов, сетевыми процессорами, препроцессорами, сопроцессорами, процессорами для обработки данных и аудиопроцессорами. Термин "память" включает электрическое устройство, хранящее данные для извлечения. Таким образом, в одном из аспектов подходящая память представляет собой блок компьютера, который хранит данные и поддерживает вычислительный процесс. В более общем смысле известны подходящие способы и устройства для обеспечения необходимой сетевой передачи данных.
[0146] Также предусмотрен энергонезависимый машиночитаемый носитель, который содержит исполняемый код для выполнения описанных способов. В определенных вариантах осуществления носитель дополнительно содержит данные или базы данных, необходимые для таких способов.
Пример
[0147] Настоящее изобретение дополнительно проиллюстрировано посредством ссылки на этот пример, который представляет собой результаты кинетической имитации динамики систем изолирующих посредством чередования трансляторов (RST) по настоящему изобретению, с одновременным сравнением с кинетическими имитациями систем трансляторов с изолированными фиксаторами (THST), указанными выше в подразделе, озаглавленном "изоляция фиксаторов", со ссылкой на ФИГ. 2 и ФИГ. 3.
[0148] Получали схемы, которые включали последовательности из N трансляций, обозначаемые как "количество этапов", у которых их константы скоростей прямых реакций получали из Frezza, B.M., Orchestration of Molecular Information Through Higher Order Chemical Recognition: A Thesis Presented, The Scripps Research Institute, La Jolla, California (2010), at Chapter 3. Схемы также включали фоновую "утечку" трансляторов, также с константами скоростей реакций из Frezza (2010) и взаимодействиями с "образованием замыканий". У этих последних их константы скоростей реакций получали на основе свободной энергии связывания ближайшего соседа, см. Allawi and SantaLucia, Biochemistry 36: 10581-94 (1997), с использованием стандартной константы скорости прямой реакции для гибридизации (105 моль в секунду) и полного равновесия для получения константы скорости обратной реакции.
[0149] Для каждой имитации проводили имитации основных сигналов и фоновых сигналов для обеих систем RST и THST. "Основной сигнал" относится к концентрации получаемой конечной выходной последовательности, с введением исходной входной последовательности в концентрации, равной с другими трансляторами в имитации. "Фоновый сигнал" означает концентрацию получаемой конечной выходной последовательности, без введения исходной входной последовательности.
[0150] Эти имитации демонстрируют идеализированный ответ, который каждая конструкция системы должна обеспечивать в условиях варьирования длины фиксаторов, концентраций трансляторов и размера системы (количества этапов).
[0151] На ФИГ. 17A-M представлены результаты имитации для RST и THST в отношении основного сигнала и фонового сигнала для различных размеров фиксаторов, в диапазоне от 3 до 15 нуклеотидов (н.). Все имитации проводили со 100 нМ трансляторами в три этапа. На ФИГ. 17 продемонстрировано, что при экстремально коротких фиксаторах RST фактически продуцируют очень четко выраженные фоновые сигналы. Однако по мере увеличения длины фиксаторов этот фонд падает почти до нуля (8 н. и более; например, см. ФИГ. 17F), при этом THST во всех случаях обладают достаточно однородной и относительно высокой скоростью фоновой трансляции, также обозначаемой как "утечка". Кроме того, после увеличения длины фиксаторов более 11 н., замыкающие взаимодействия в THST начинали доминировать в системе и значительно замедлять основную активность, тогда как RST с более длинными фиксаторами функционировали номинально (см., например, ФИГ. 17I-M).
[0152] В другой серии имитаций фиксатор длиной 8 н. при концентрациях трансляторов 100 нМ продуцировал для RST и THST сходные основные сигналы. Системы исследовали с увеличивающейся глубиной системы в отношении количества этапов, например, количества трансляторов. Как представлено на ФИГ. 18A-D, с увеличением количества этапов фонов сигналы RST и THST значительно увеличивались. Однако в случае THST степень этого увеличения была значительно больше.
[0153] В еще одной серии имитаций исследовали системы умеренного размера (5 этапов) с длинными (13 н.) и короткими (8 н.) фиксаторами и продемонстрировали, как их поведение изменяется при изменениях концентраций трансляторов. Результаты представлены на ФИГ. 19A-N. Более высокие концентрации трансляторов (10 мкМ) действовали очень быстро, а очень низкие концентрации трансляторов (100 пМ) действовали очень медленно. Это поведение соответствует тому факту, что системы RST и THST подчиняются кинетике реакций второго порядка, таким образом, время до достижения равновесия зависит от общей концентрации системы.
[0154] В случае длинных фиксаторов для RST сроки оставались биологически значимыми до очень низких концентраций, когда небольшую активность наблюдали вплоть до 100 пМ, тогда как THST для получения любой значимой активности требовали намного более высоких концентраций (10 мкМ) (ФИГ. 19A-G). В случае коротких фиксаторов RST функционировали подобным образом, хотя и со значительно более высокими фоновыми сигналами (ФИГ. 19H-N). THST с этими более короткими фиксаторами регулировали с получением значимой активности при низких концентрациях, но при высоких концентрациях утечка, вносящая вклад в фоновые сигналы THST, была катастрофически высокой.
[0155] Взятые вместе, эти данные демонстрируют, что RST обеспечивают возможности конструирования, в результате чего фоновую активность (утечку) можно снижать почти до невыявляемых уровней при использовании более длинных фиксаторов без потери активности посредством экстремального ингибирования, с которым сталкиваются THST. Кроме того, показано, что RST могут функционировать с идеальным соотношением основной и фоновой реактивности в широчайшем динамическом диапазоне концентраций, что делает их идеально подходящими для использования в биологических условиях.
[0156] Несмотря на то, что описаны конкретные варианты осуществления настоящего изобретения, они являются только иллюстративными и не ограничивают изобретения. Обзор настоящего описания для специалистов в области техники, к которой относится изобретение, делает очевидными множество вариаций по изобретению. Полный объем изобретения необходимо определять на основе приведенной ниже формулы изобретения, наряду с их полным диапазоном эквивалентов, и на основе описания с такими вариантами.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРАНСЛЯТОРЫ, ИЗОЛИРУЮЩИЕ ПОСРЕДСТВОМ ЧЕРЕДОВАНИЯ | 2014 |
|
RU2599444C1 |
| СИСТЕМА И СПОСОБ ДЛЯ РАСПРОСТРАНЕНИЯ ИНФОРМАЦИИ С ИСПОЛЬЗОВАНИЕМ МОДИФИЦИРОВАННЫХ НУКЛЕИНОВЫХ КИСЛОТ | 2011 |
|
RU2603746C2 |
| ОПОСРЕДОВАННАЯ НУКЛЕАЗОЙ СБОРКА ДНК | 2015 |
|
RU2707911C2 |
| НАБОР ЗОНДОВ ДЛЯ АНАЛИЗА ОБРАЗЦОВ ДНК И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2016 |
|
RU2753883C2 |
| СПОСОБ АМПЛИФИКАЦИИ ДНК НА ОСНОВЕ ВНЕДРЕНИЯ В ЦЕПЬ | 2015 |
|
RU2729983C2 |
| СПОСОБ ОБНАРУЖЕНИЯ ВИРУСА ГЕПАТИТА С (ВАРИАНТЫ), НАБОР, СПОСОБ ОПРЕДЕЛЕНИЯ ОБРАЗЦА КРОВИ, РЕАГЕНТ | 1991 |
|
RU2180354C2 |
| СПОСОБ СОЗДАНИЯ КВАДРУПЛЕКСА И СПОСОБ ОБНАРУЖЕНИЯ НУКЛЕИНОВОЙ КИСЛОТЫ | 2002 |
|
RU2304168C2 |
| СПОСОБ ВЫЯВЛЕНИЯ ПАРВОВИРУСА B19 ЧЕЛОВЕКА В БИОЛОГИЧЕСКОМ ОБРАЗЦЕ (ВАРИАНТЫ) И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2002 |
|
RU2301263C2 |
| ОЛИГОМЕР (ВАРИАНТЫ), СПОСОБ ОБНАРУЖЕНИЯ ПОСЛЕДОВАТЕЛЬНОСТИ HCV (ВАРИАНТЫ), НАБОР ДЛЯ ОБНАРУЖЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ КРОВИ | 1990 |
|
RU2145635C1 |
| МОЛЕКУЛЯРНЫЙ ЗОНД | 1991 |
|
RU2107730C1 |

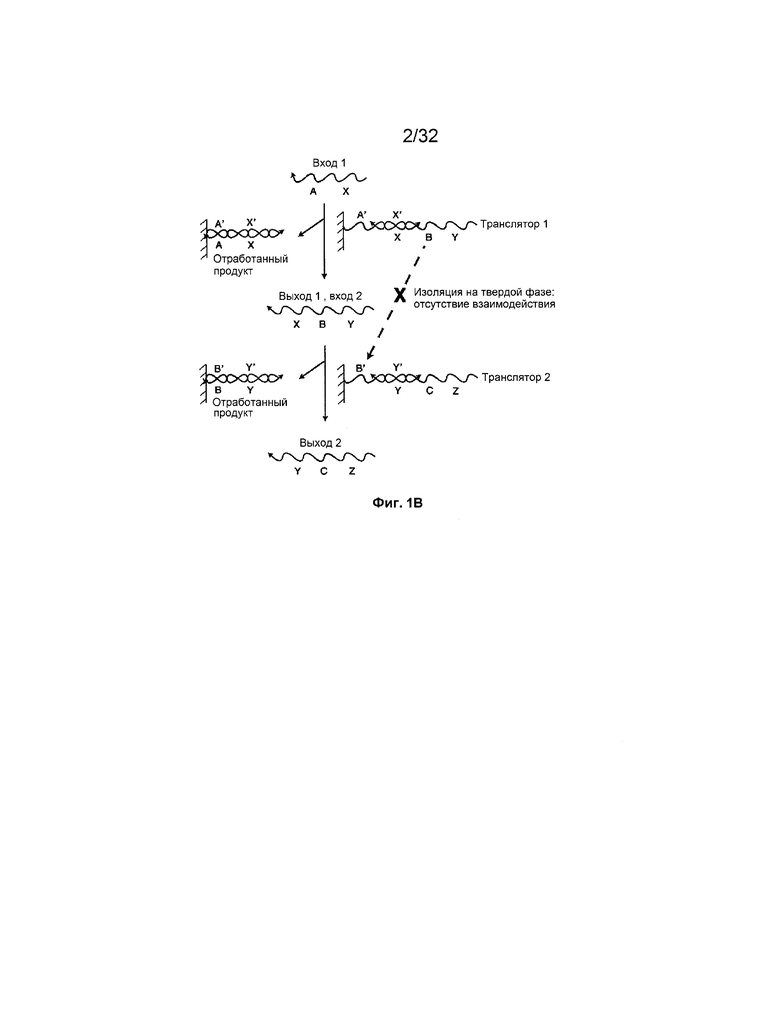
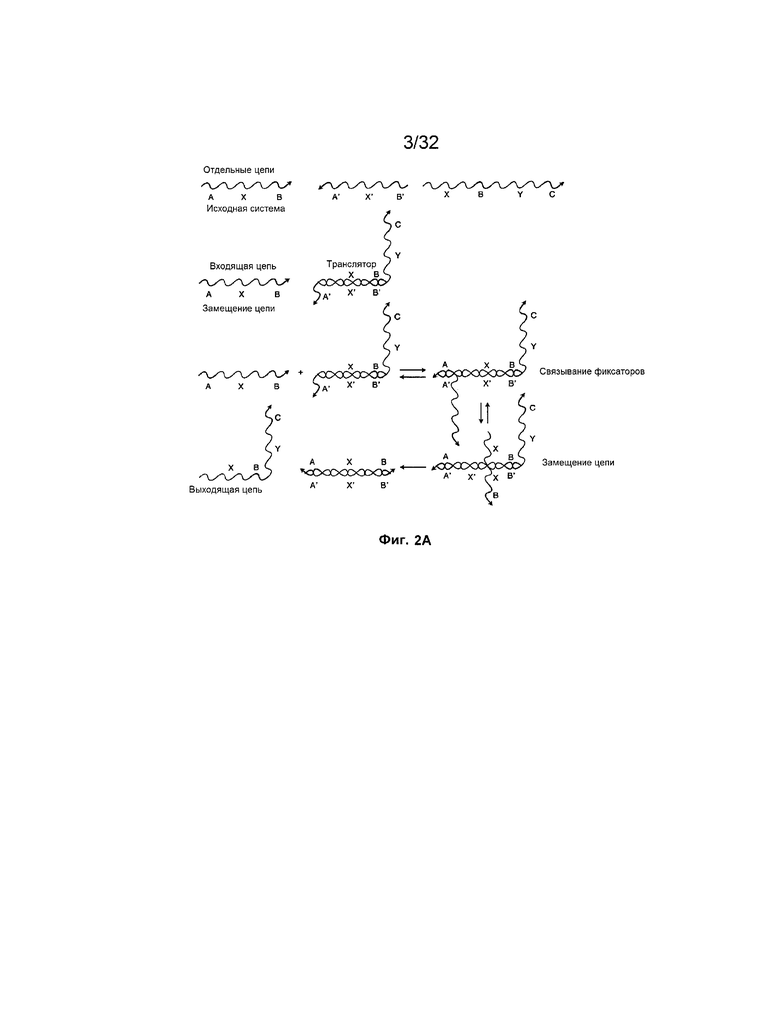
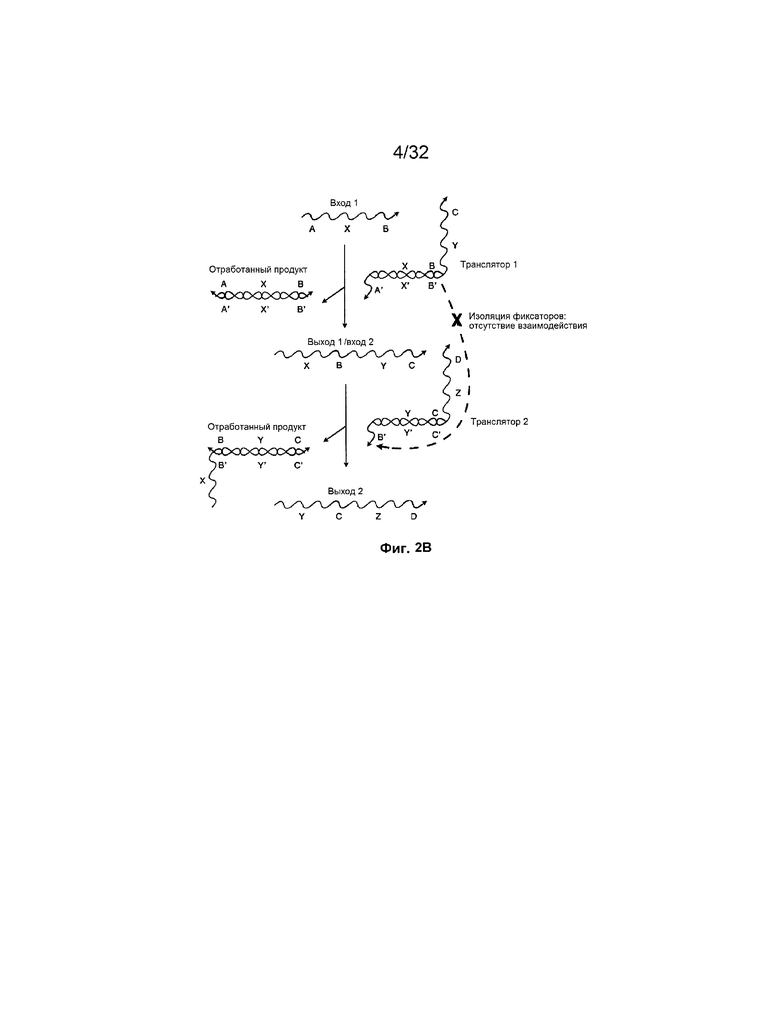
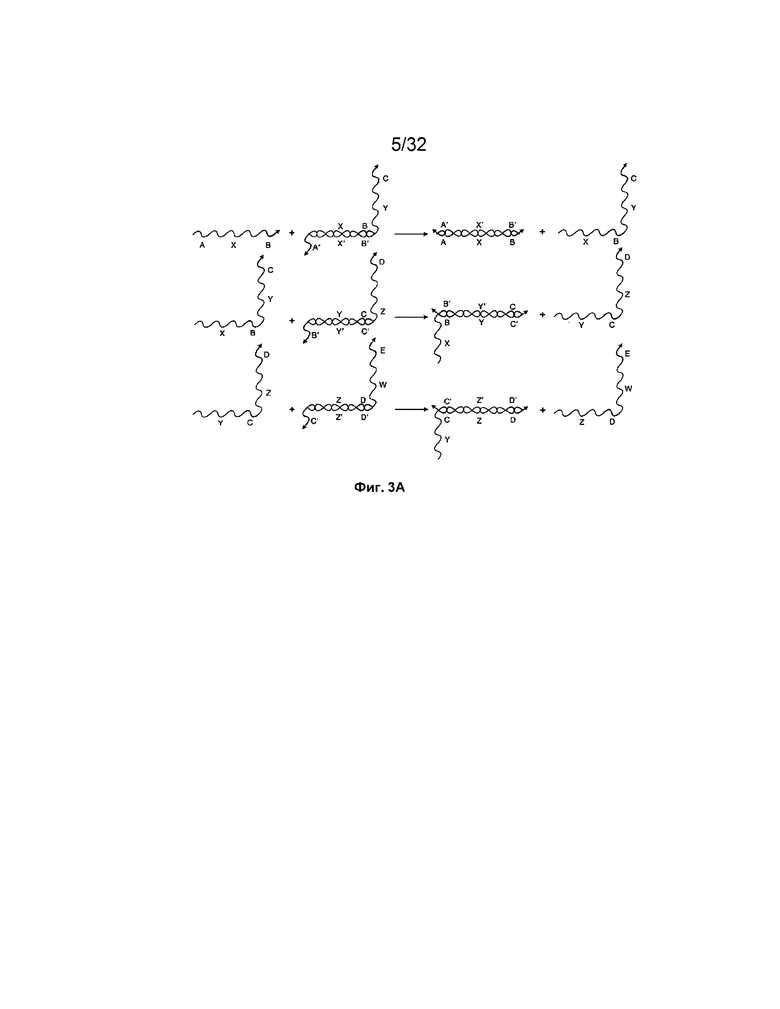
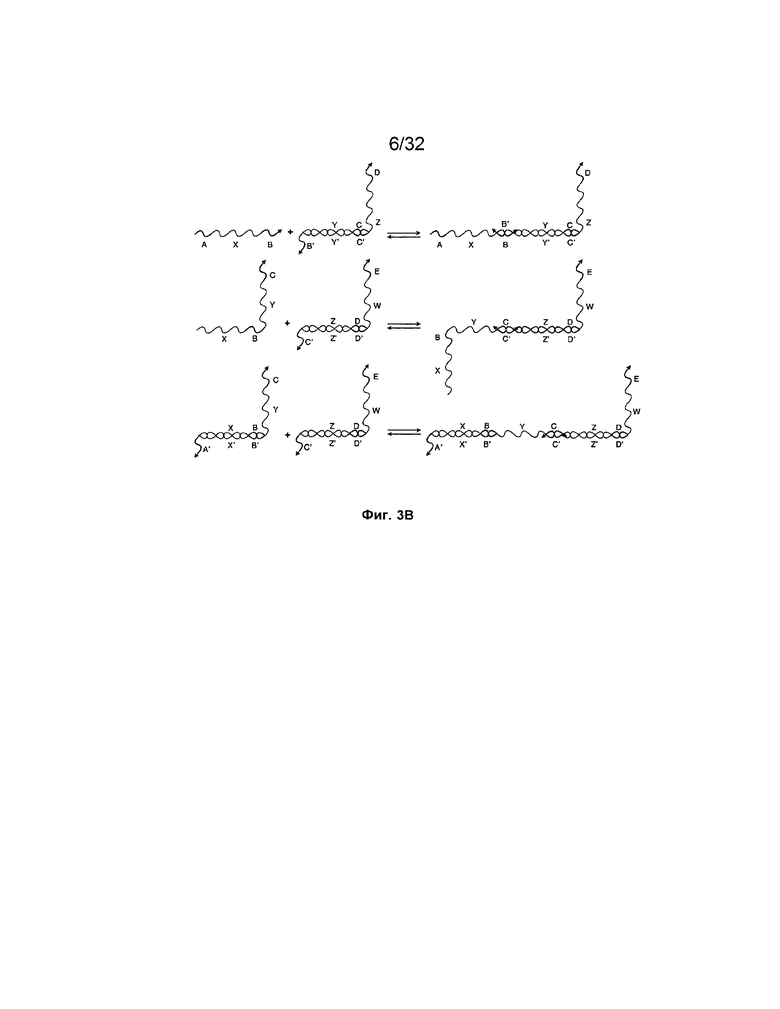
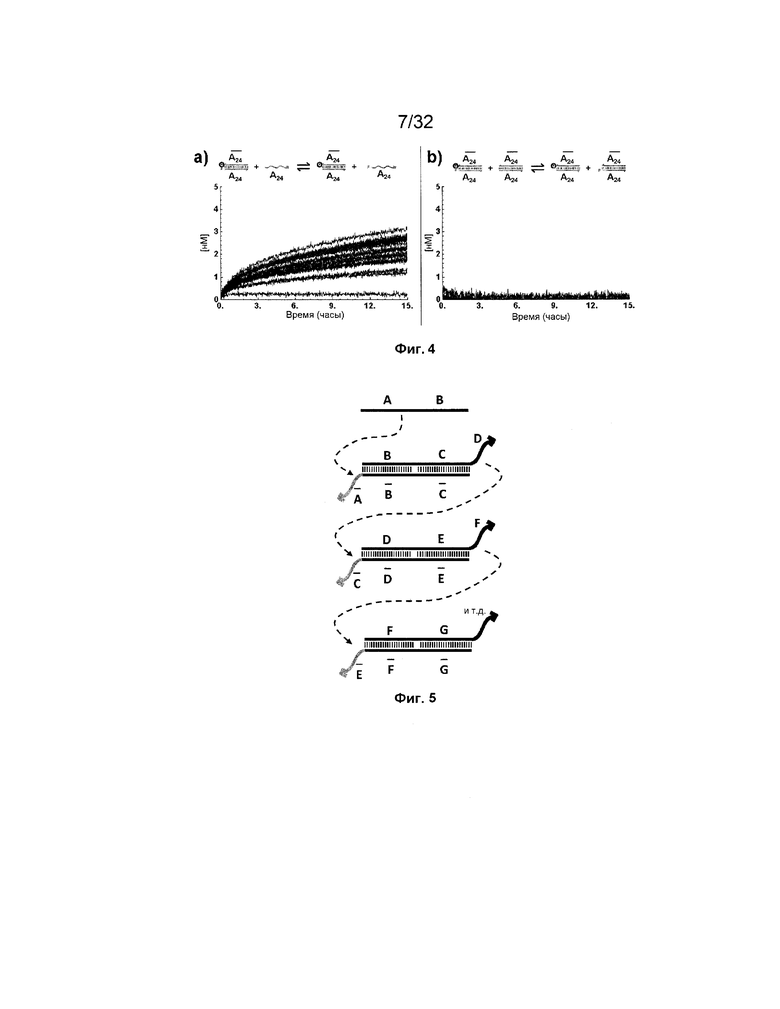
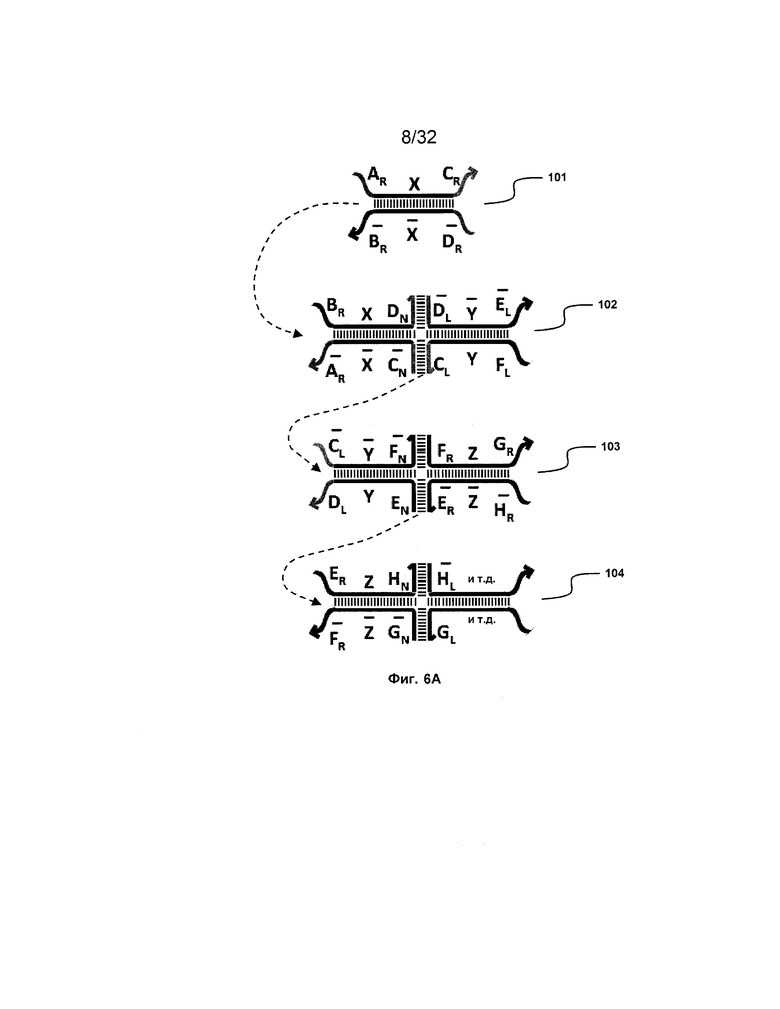
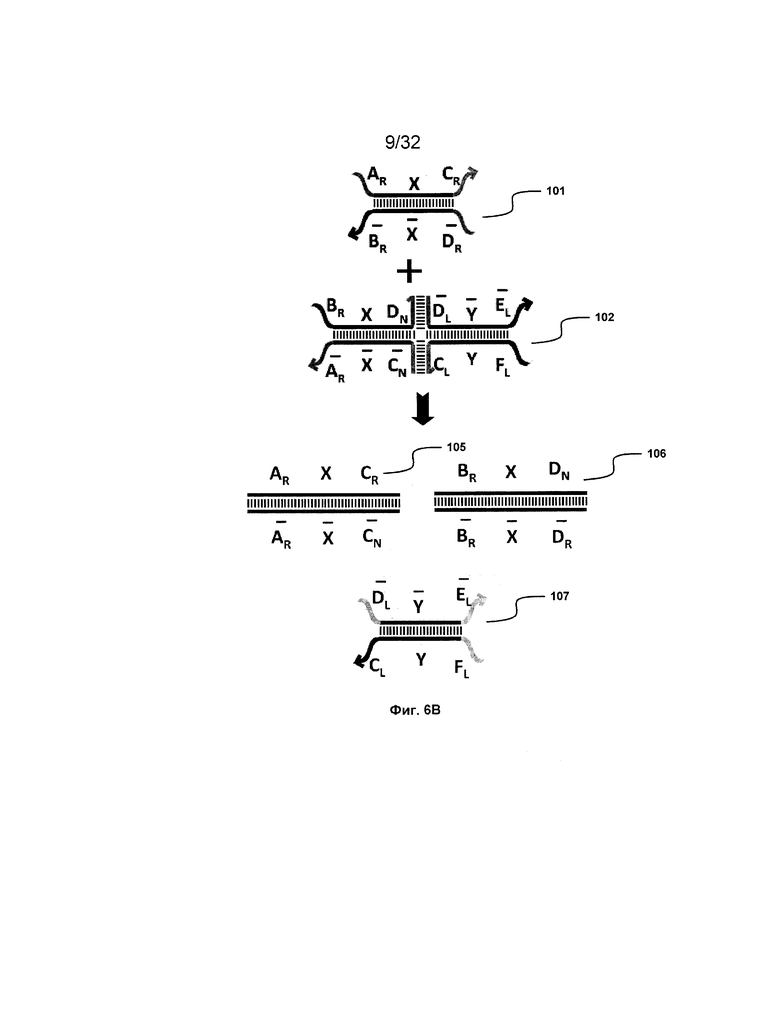
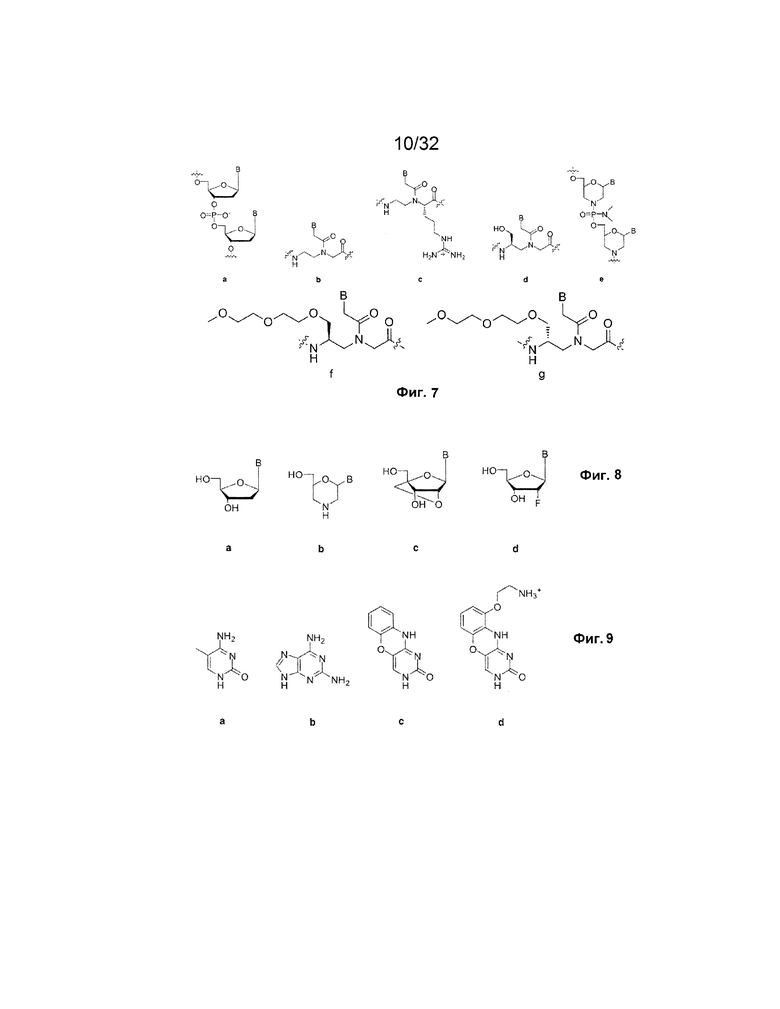
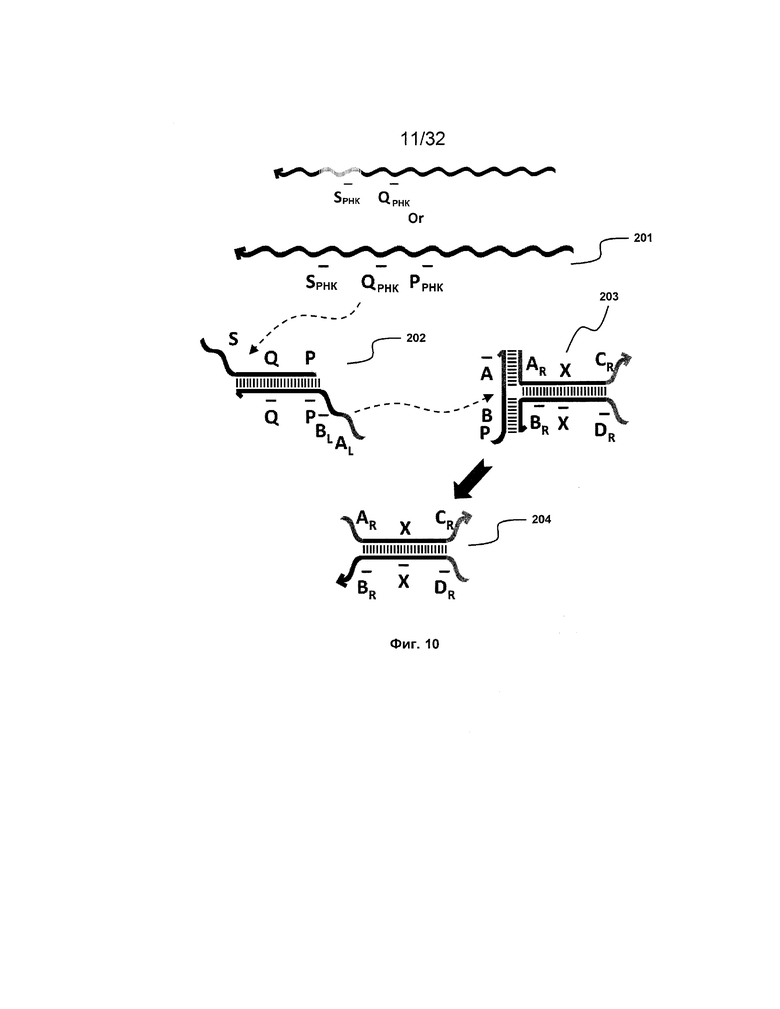
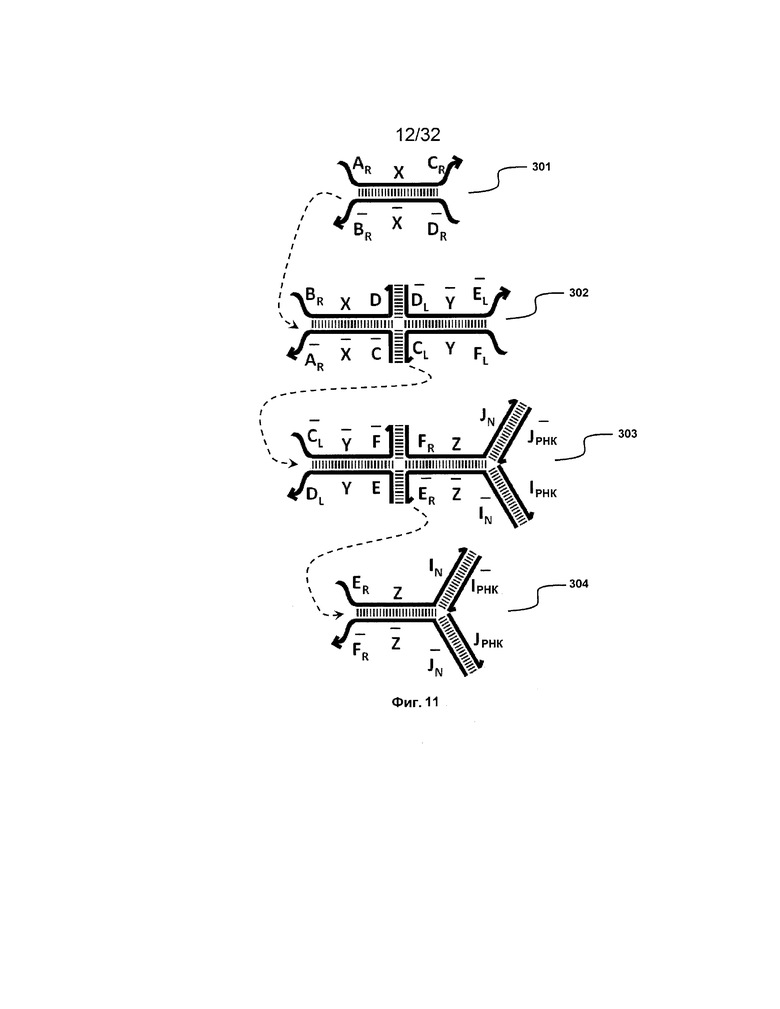
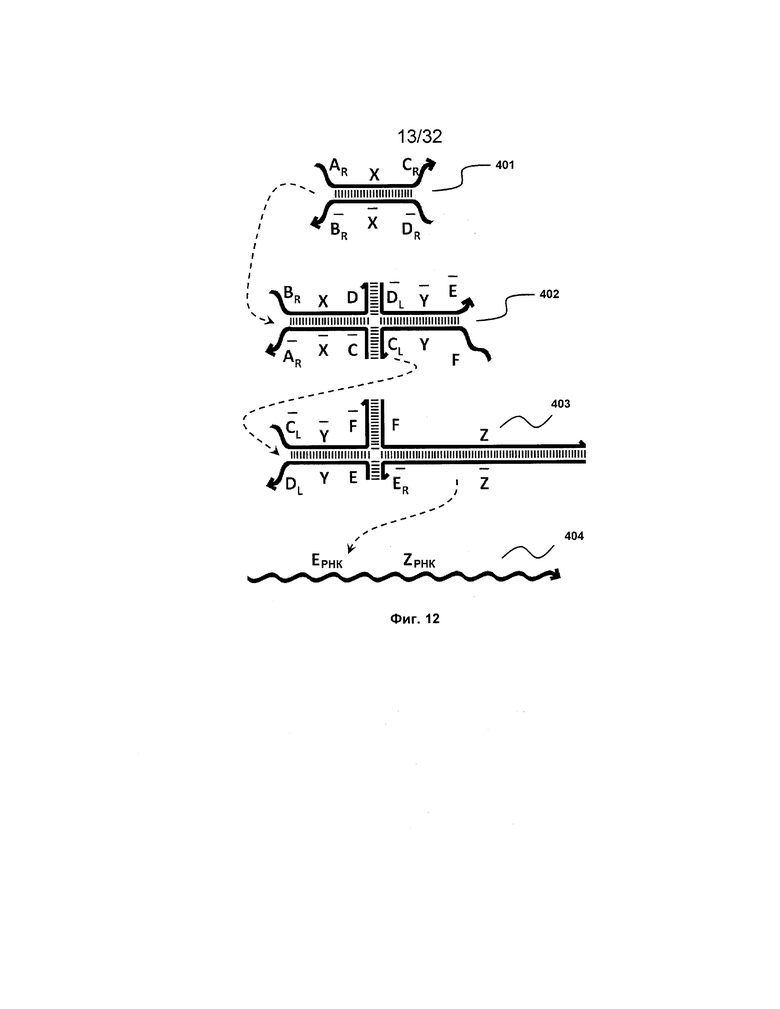
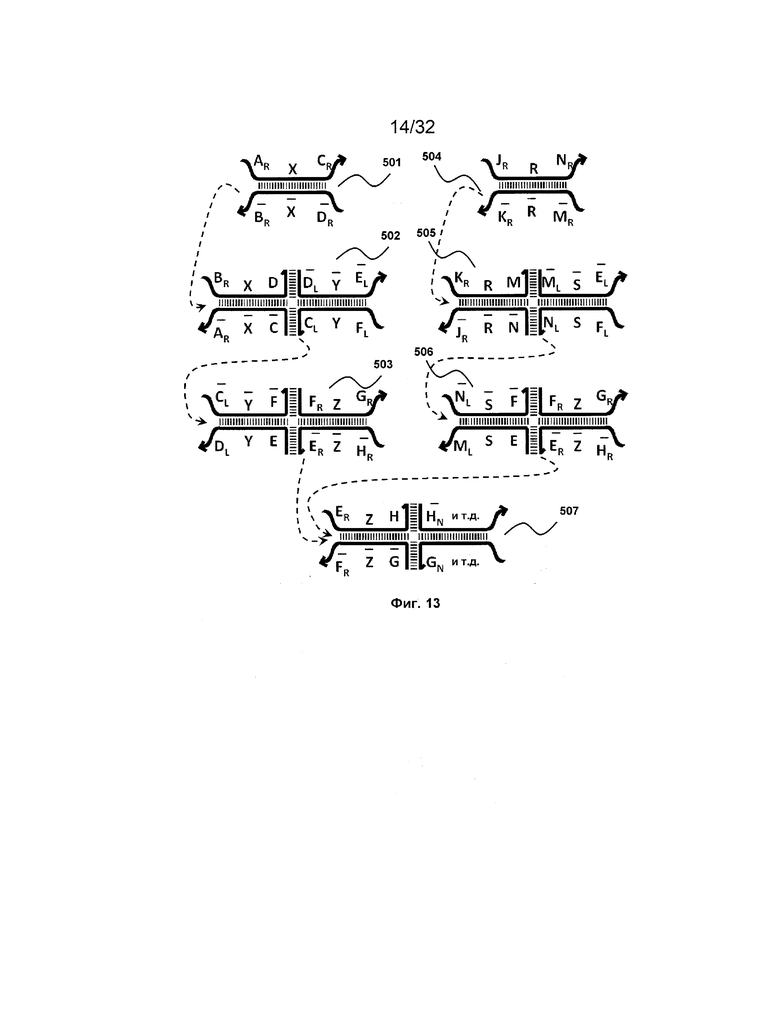
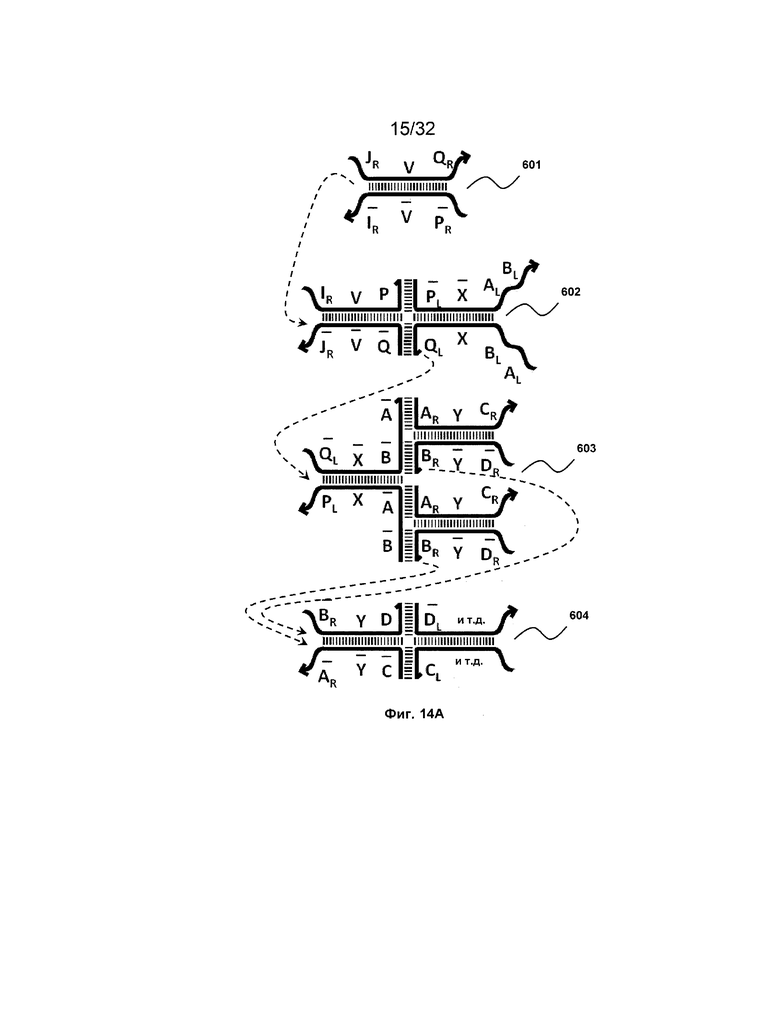
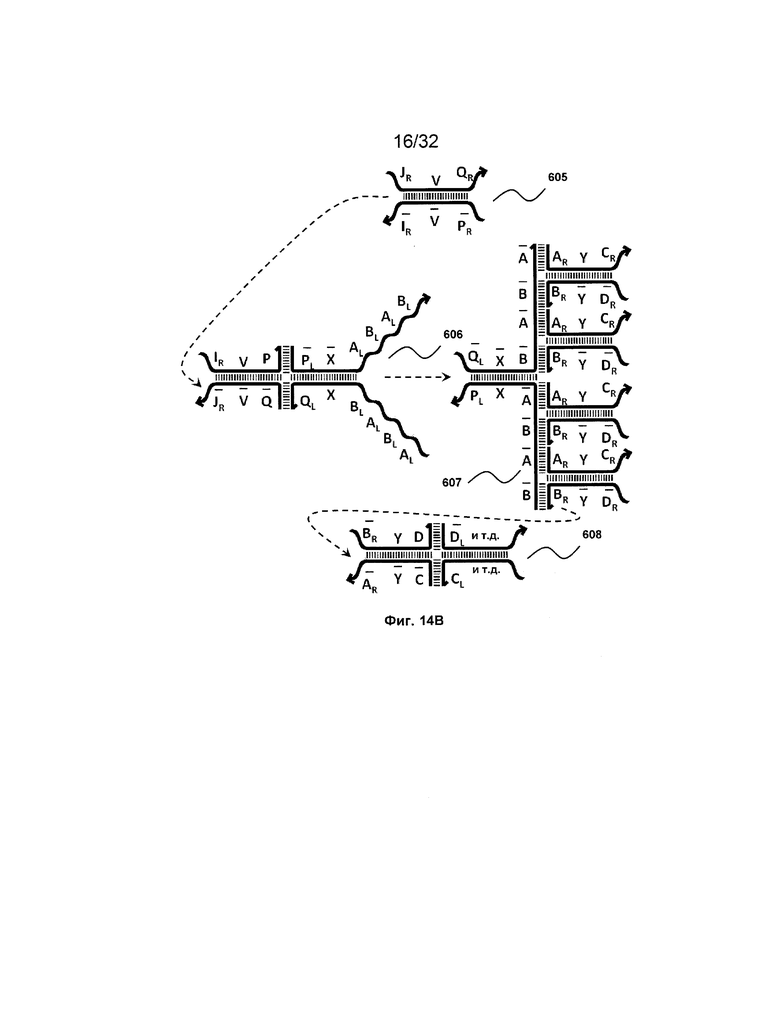
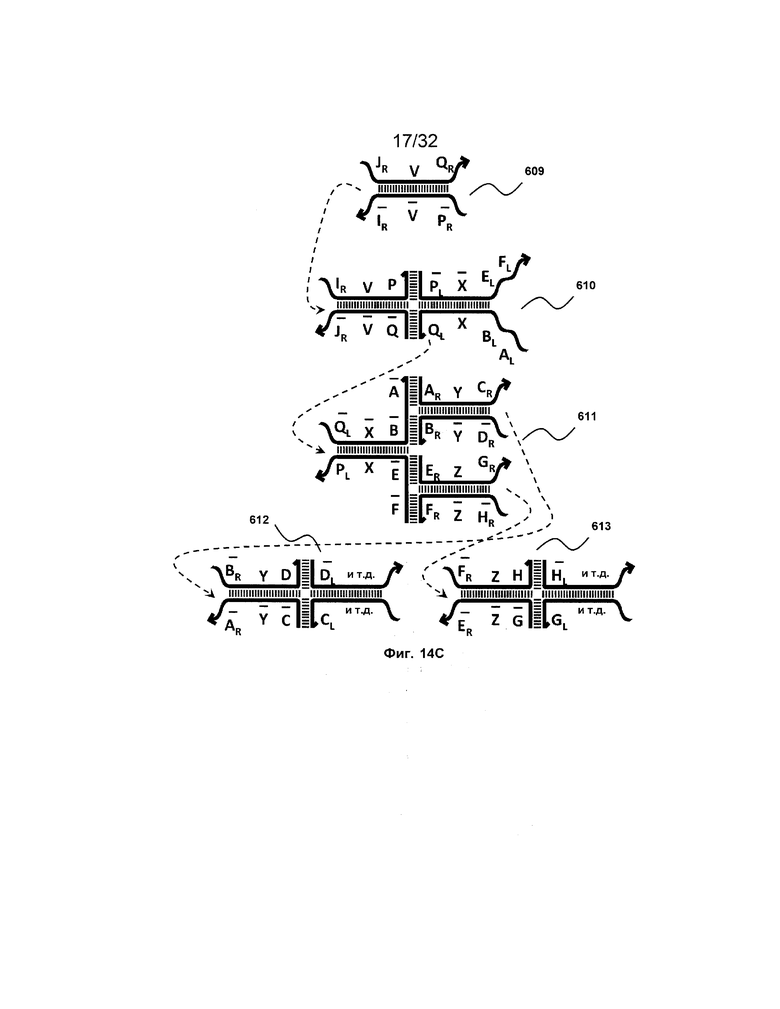
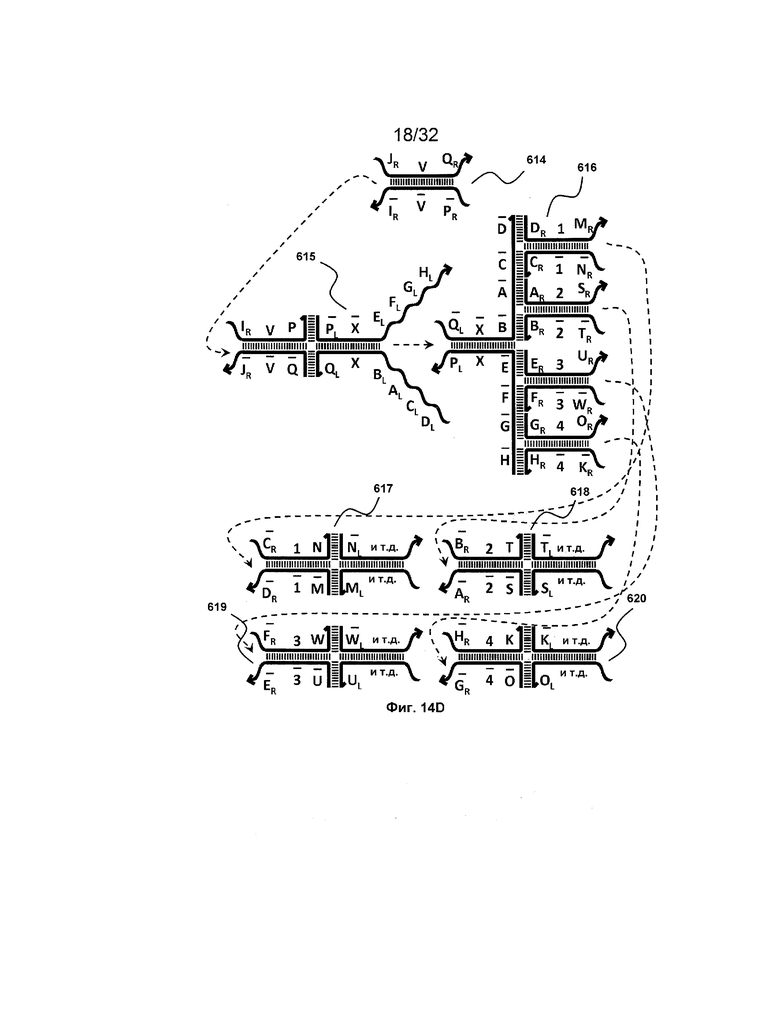
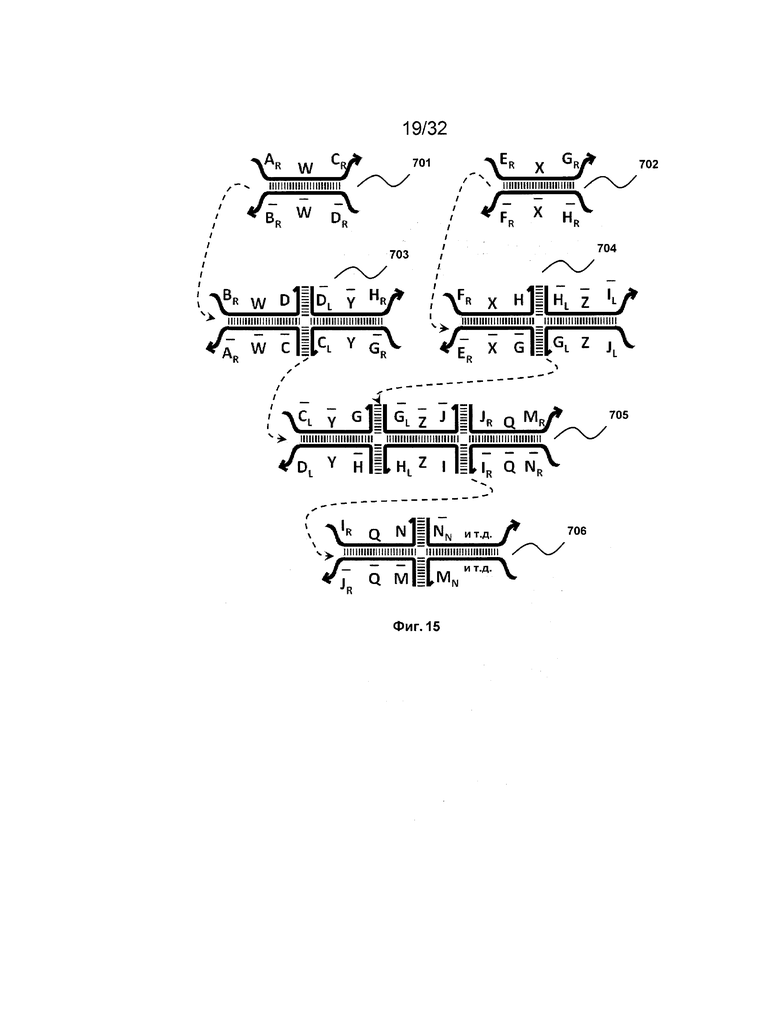
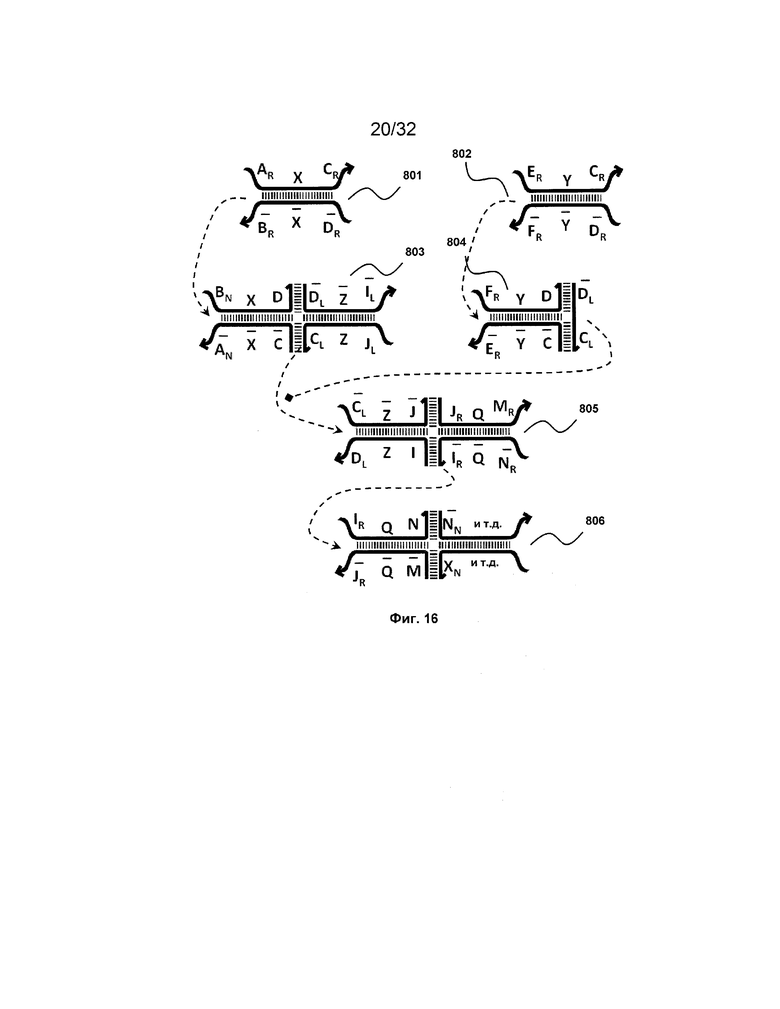
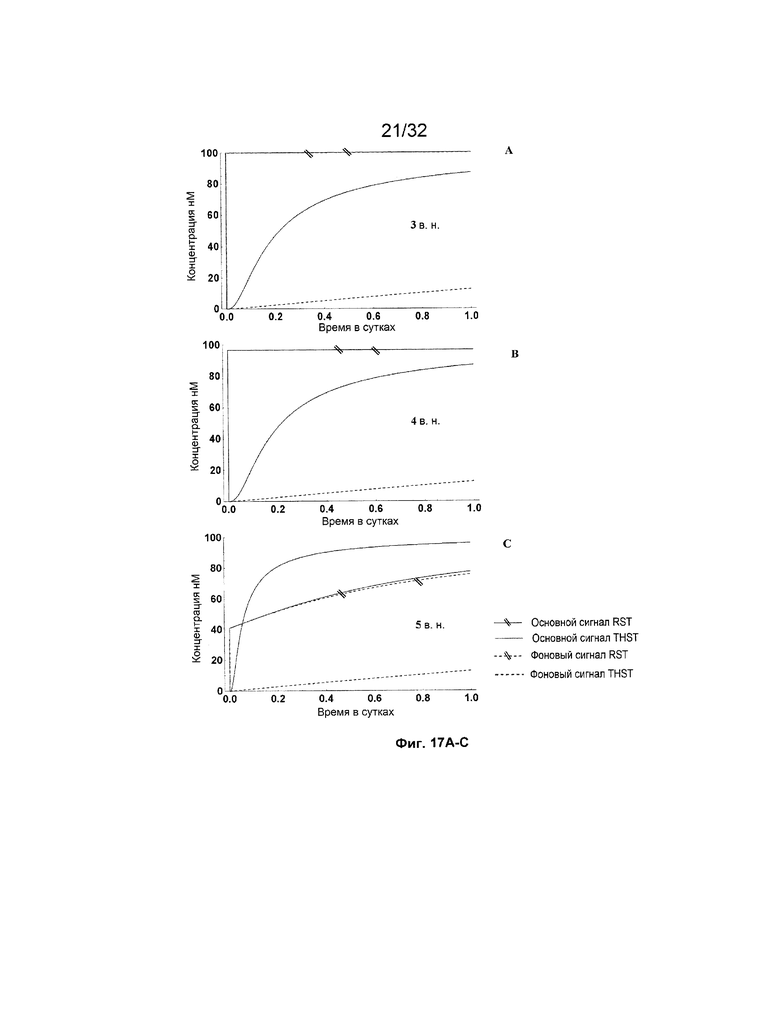
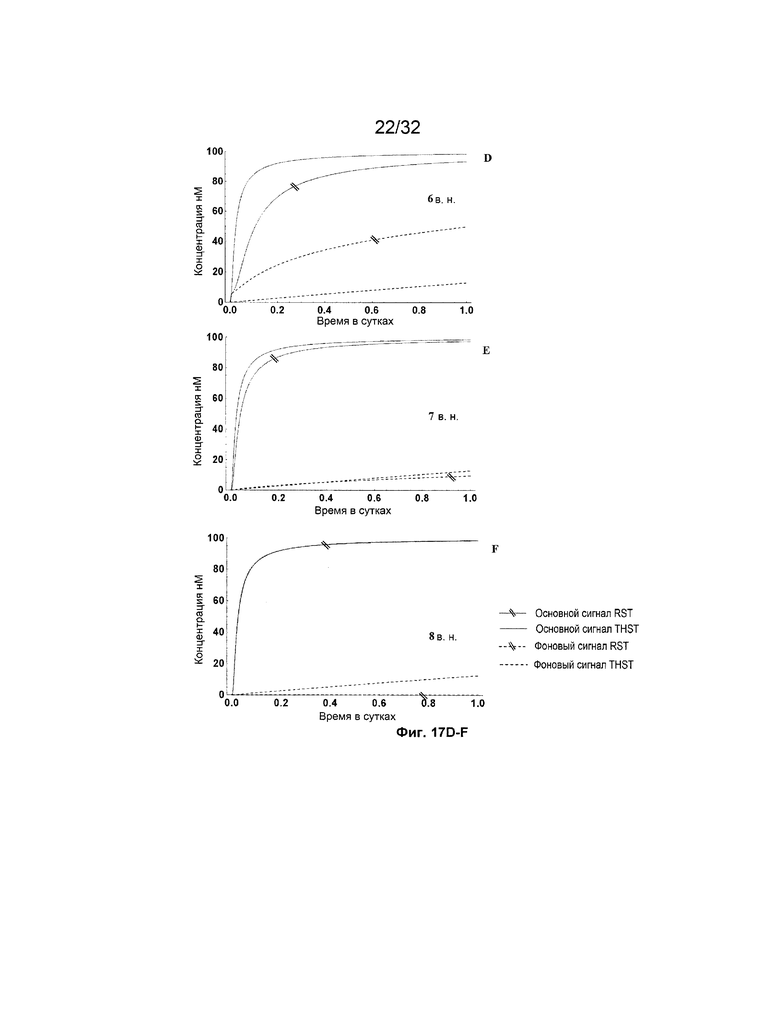
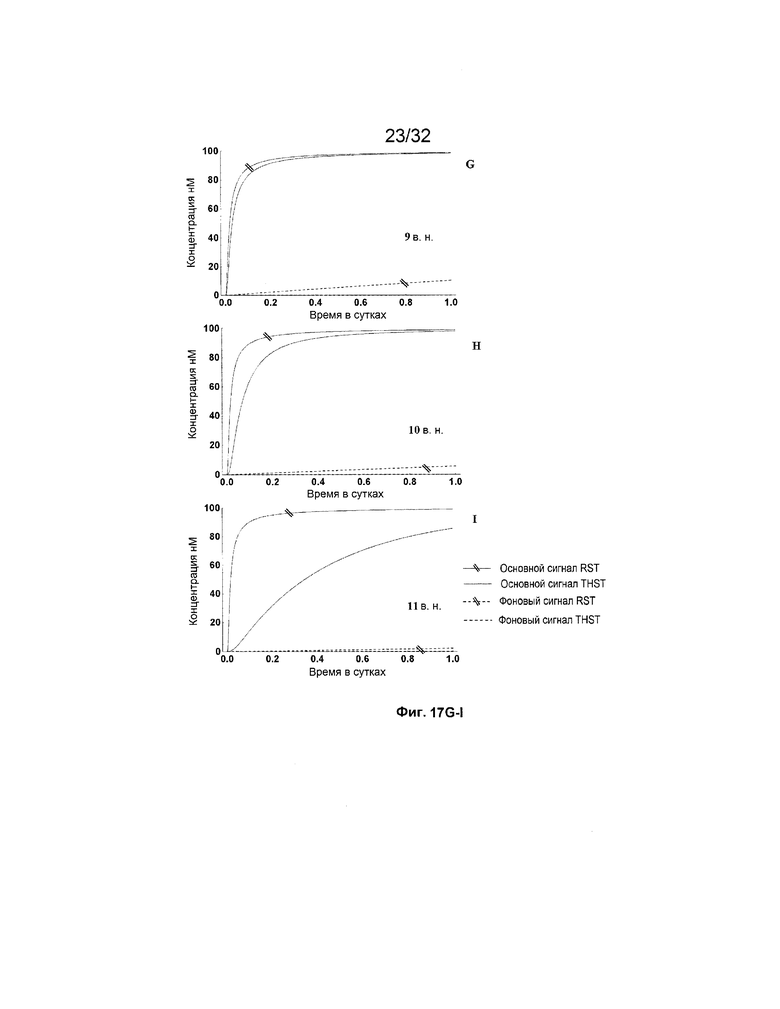
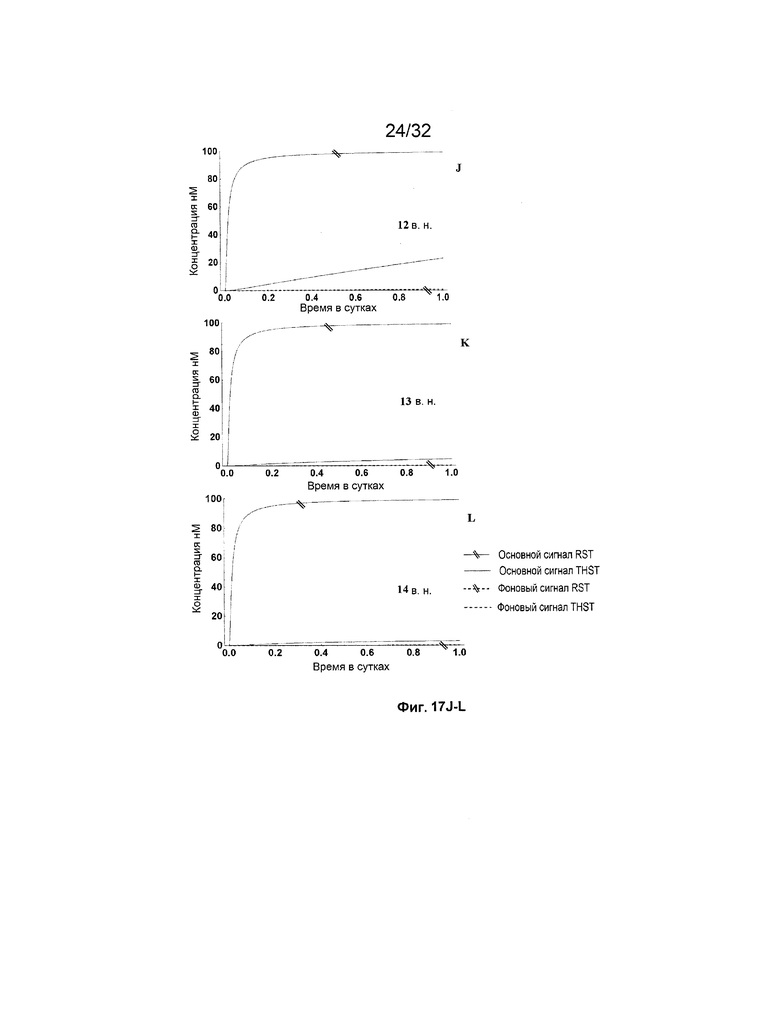
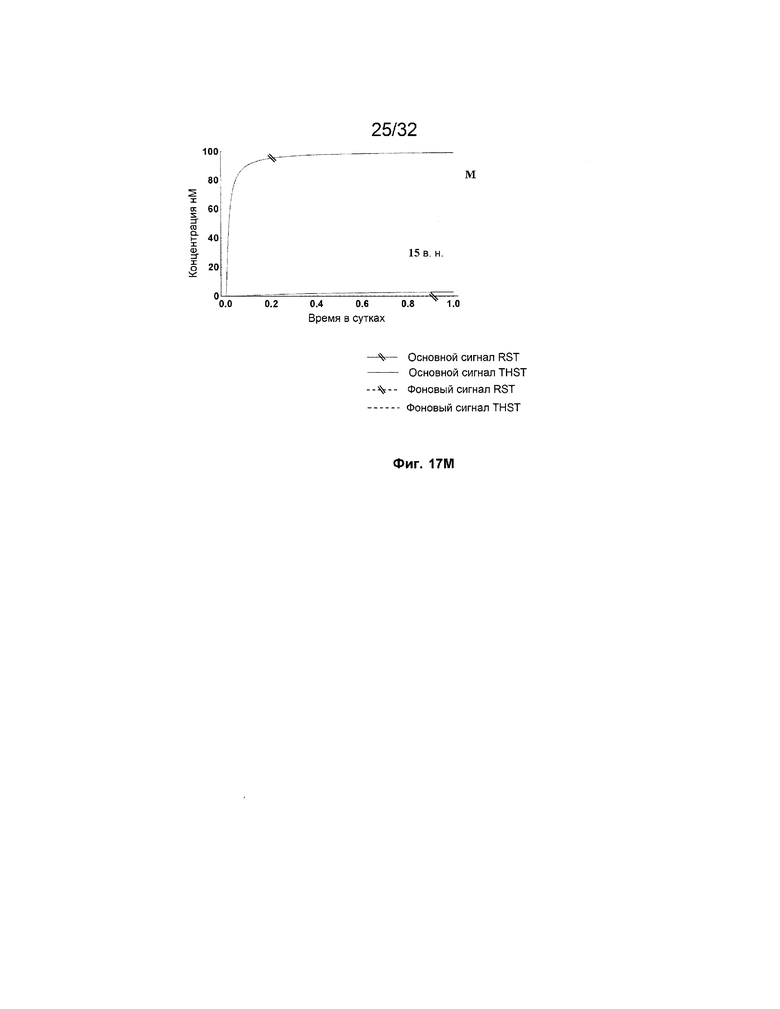
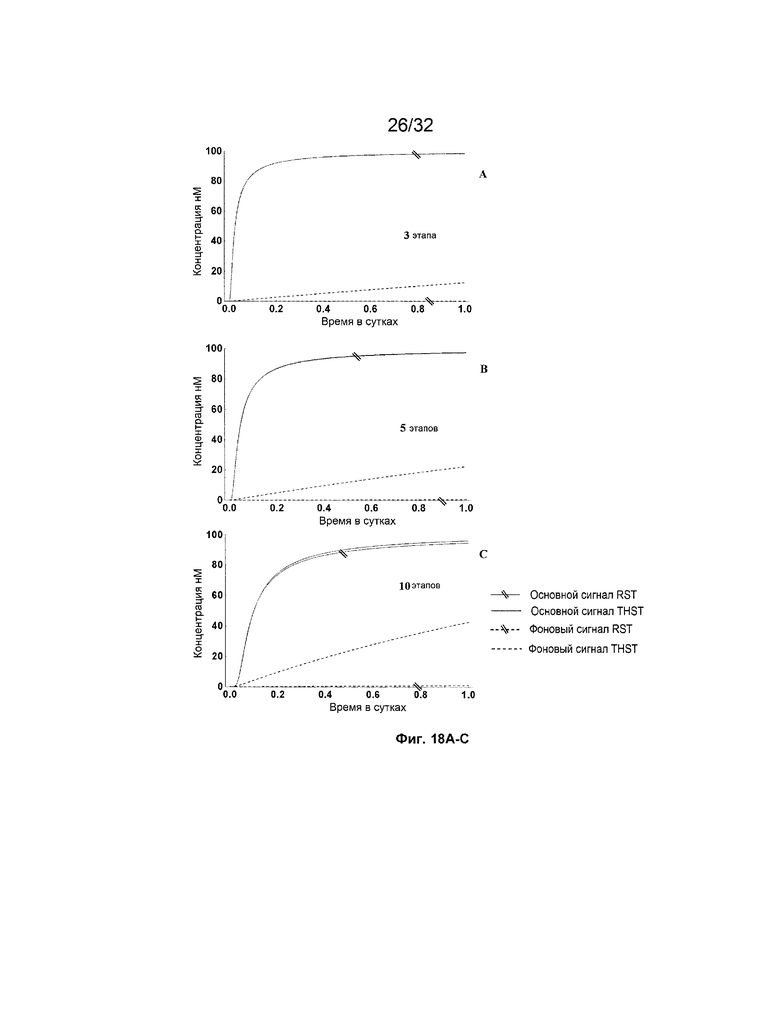
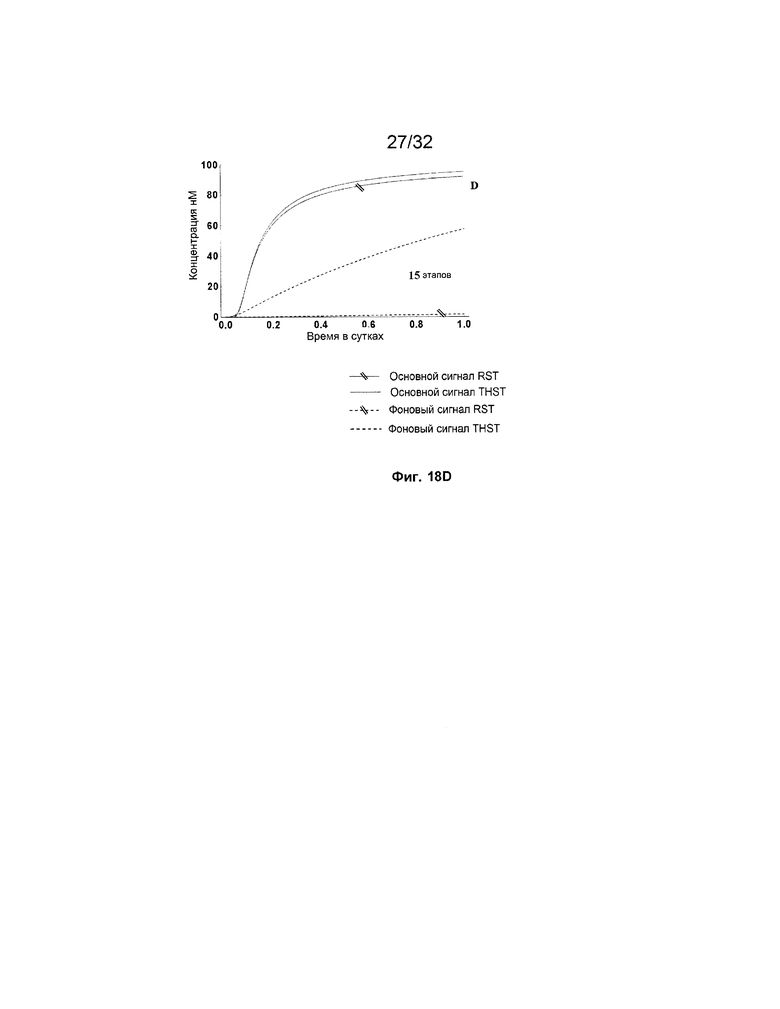
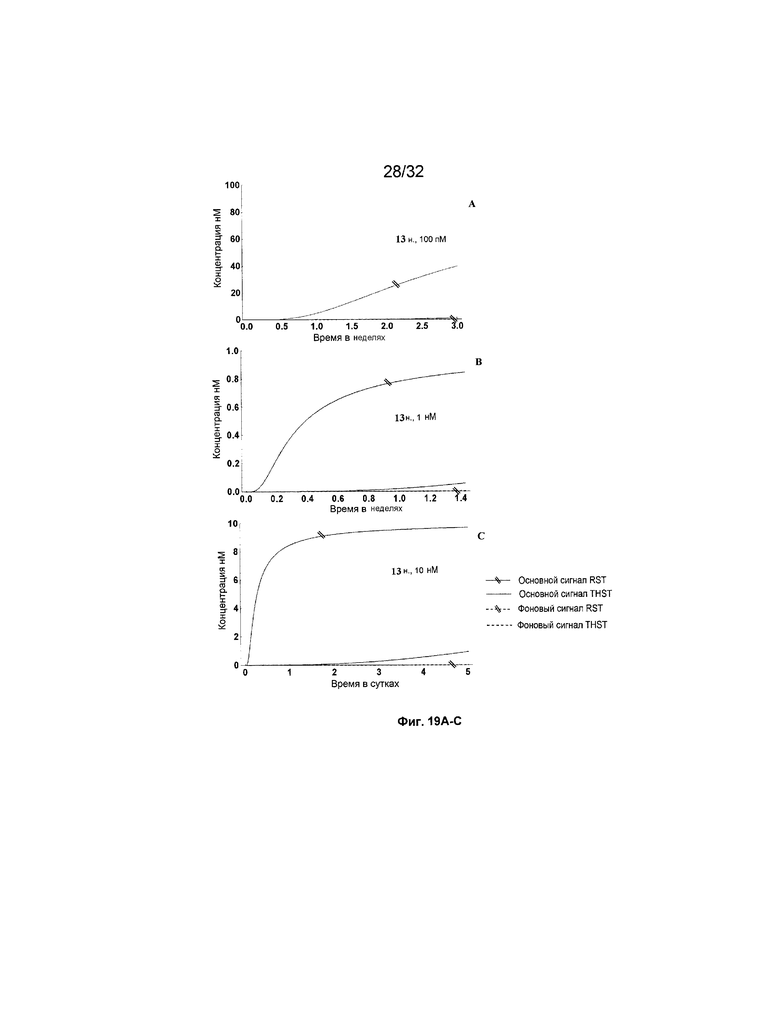
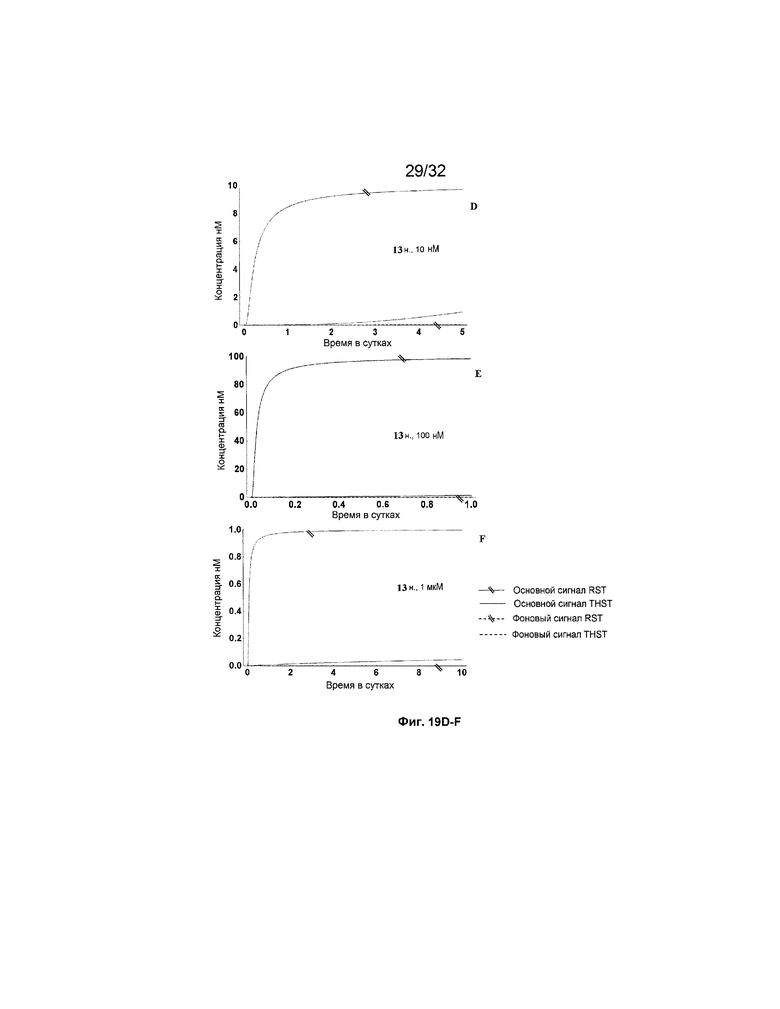
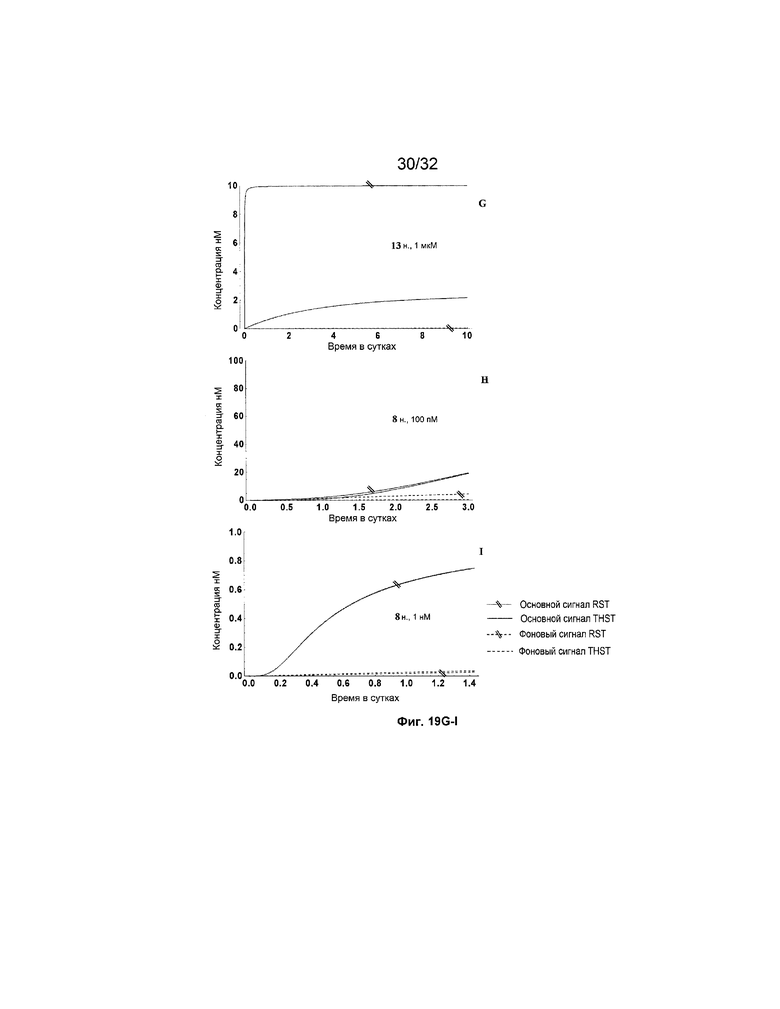
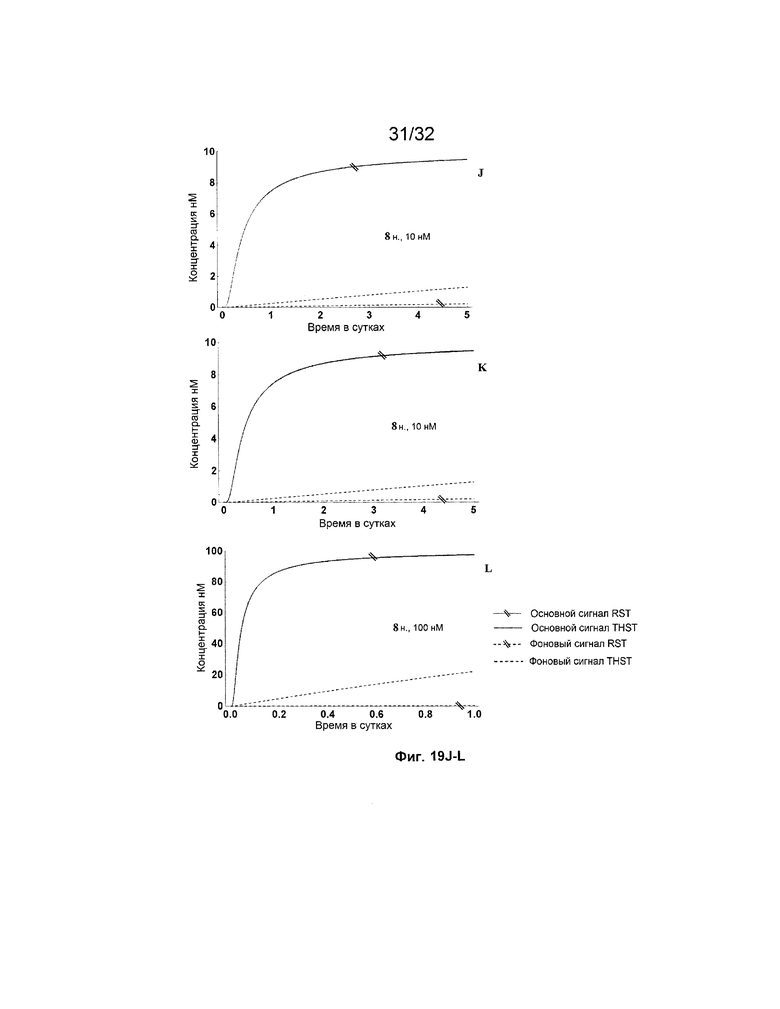
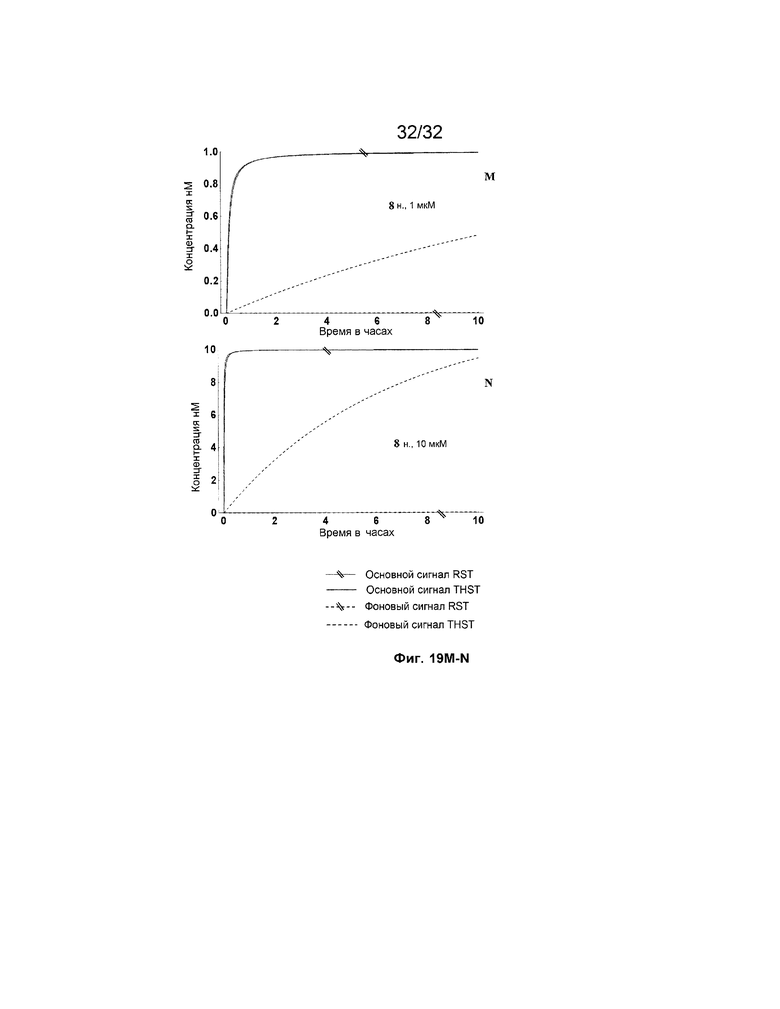
Изобретение относится к биотехнологии. Описаны трансляторы на основе нуклеиновых кислот, способные осуществлять логические операции с улучшенной эффективностью, максимизированным выходом и сниженным побочным действием, в частности в биологической системе. Также предоставлены способы использования этих трансляторов для передачи сигнала. 6 н. и 12 з.п. ф-лы, 19 ил., 2 табл., 1 пр.
1. Композиция для осуществления реакции замещения цепей нуклеиновых кислот, содержащая:
(a) первый комплекс нуклеиновых кислот, содержащий (i) первую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как S-Q-P, где каждая буква означает фрагмент, а последовательность букв, соединенных "-", означает цепь, и (ii) вторую цепь, последовательно содержащую первый, второй, третий и четвертый фрагменты и определенную как A- -
- -
-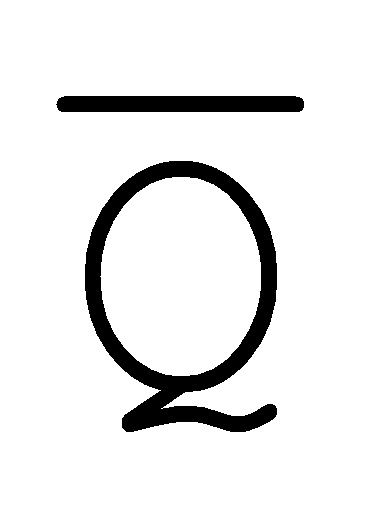 , где первый комплекс содержит область дуплекса (Q-P::
, где первый комплекс содержит область дуплекса (Q-P:: -
-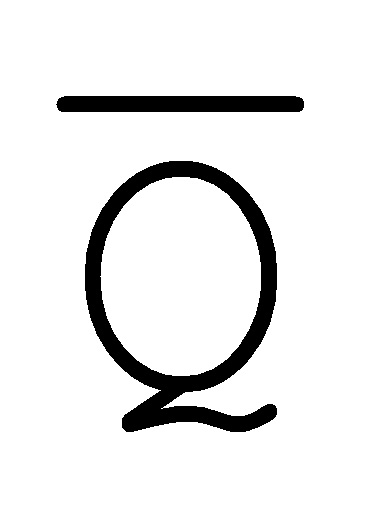 ), сформированную между вторым и третьим фрагментами первой цепи и третьим и четвертым фрагментами второй цепи; и
), сформированную между вторым и третьим фрагментами первой цепи и третьим и четвертым фрагментами второй цепи; и
(b) второй комплекс нуклеиновых кислот, содержащий (i) первую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как P-B- , (ii) вторую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как A-X-C, и (iii) третью цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как
, (ii) вторую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как A-X-C, и (iii) третью цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как  -
- -
- , где второй комплекс содержит первую область дуплекса (
, где второй комплекс содержит первую область дуплекса ( ::A), сформированную между третьим фрагментом первой цепи и первым фрагментом второй цепи, вторую область дуплекса (B::
::A), сформированную между третьим фрагментом первой цепи и первым фрагментом второй цепи, вторую область дуплекса (B:: ), сформированную между первым фрагментом первой цепи и третьим фрагментом третьей цепи, и третью область дуплекса (X::
), сформированную между первым фрагментом первой цепи и третьим фрагментом третьей цепи, и третью область дуплекса (X:: ), сформированную между вторым фрагментом второй цепи и вторым фрагментом третьей цепи,
), сформированную между вторым фрагментом второй цепи и вторым фрагментом третьей цепи,
где первый (S) фрагмент первой цепи первого комплекса, первый (A) и второй ( ) второй цепи первого комплекса, первый фрагмент (P) третьей цепи второго комплекса, третий фрагмент (C) второй цепи второго комплекса и первый фрагмент (
) второй цепи первого комплекса, первый фрагмент (P) третьей цепи второго комплекса, третий фрагмент (C) второй цепи второго комплекса и первый фрагмент ( ) третьей цепи второго комплекса являются одноцепочечными; и
) третьей цепи второго комплекса являются одноцепочечными; и
где первый, второй и третий фрагменты (A,  ,
,  ) второй цепи первого комплекса обладают комплементарностью последовательностей с третьим, вторым и первым фрагментами (
) второй цепи первого комплекса обладают комплементарностью последовательностей с третьим, вторым и первым фрагментами ( , B, P) первой цепи второго комплекса для обеспечения связывания между ними, соответственно.
, B, P) первой цепи второго комплекса для обеспечения связывания между ними, соответственно.
2. Композиция по п. 1, где первая цепь (S-Q-P) первого комплекса обладает комплементарностью последовательности с фрагментом в нуклеиновой кислоте патогенной нуклеиновой кислоты для обеспечения связывания между ними.
3. Композиция по п. 2, где патогенная нуклеиновая кислота представляет собой вирусную ДНК, вирусную РНК, бактериальную ДНК, бактериальную РНК, мутантную опухолевую ДНК или опухолевую РНК.
4. Композиция для осуществления реакции замещения цепей нуклеиновых кислот, содержащая первый комплекс нуклеиновых кислот, содержащий (a) первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты, и (b) первую и вторую цепи РНК, где первый комплекс содержит:
(i) первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей;
(ii) вторую область дуплекса, сформированную между вторыми фрагментами третьей и четвертой цепей;
(iii) третью область дуплекса, сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи;
(iv) четвертую область дуплекса, сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи;
(v) пятую область дуплекса, сформированную между третьим фрагментом третьей цепи и первой РНК; и
(vi) шестую область дуплекса, сформированную между первым фрагментом четвертой цепи и второй РНК,
и первый фрагмент ( ) первой цепи и третий фрагмент (D) второй цепи являются одноцепочечными.
) первой цепи и третий фрагмент (D) второй цепи являются одноцепочечными.
5. Композиция по п. 4, дополнительно содержащая второй комплекс нуклеиновых кислот, содержащий первую и вторую цепи нуклеиновых кислот и первую и вторую цепи РНК, где:
второй комплекс содержит первую область дуплекса, сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса, сформированную между третьим фрагментом первой цепи и первой цепью РНК, и третью область дуплекса, сформированную между первым фрагментом второй цепи и второй цепью РНК, где первый фрагмент (E) первой цепи и третий фрагмент ( ) второй цепи являются одноцепочечными;
) второй цепи являются одноцепочечными;
третья цепь первого комплекса и вторая цепь второго комплекса обладают комплементарностью последовательностей для обеспечения связывания между ними; и
четвертая цепь первого комплекса и первая цепь второго комплекса обладают комплементарностью последовательностей для обеспечения связывания между ними.
6. Композиция для осуществления реакции замещения цепей нуклеиновых кислот, содержащая:
(a) первый комплекс нуклеиновых кислот, содержащий (i) первую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как B-X-D, (ii) вторую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как  -
- -
-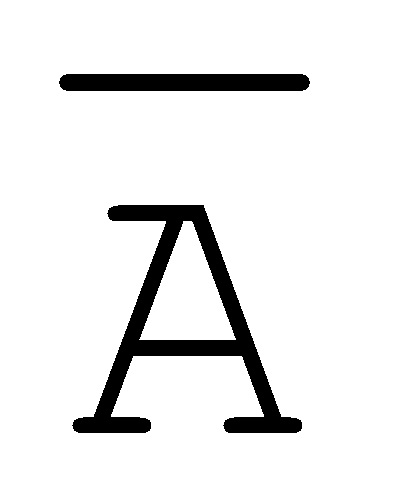 , (iii) третью цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как
, (iii) третью цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как  -
-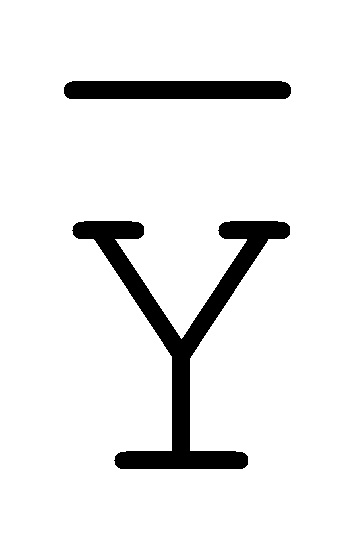 -
-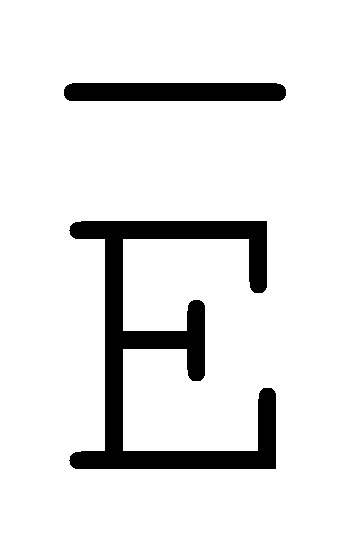 , и (iv) четвертую цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как F-Y-C, где первый комплекс содержит первую область дуплекса (X::
, и (iv) четвертую цепь, последовательно содержащую первый, второй и третий фрагменты и определенную как F-Y-C, где первый комплекс содержит первую область дуплекса (X::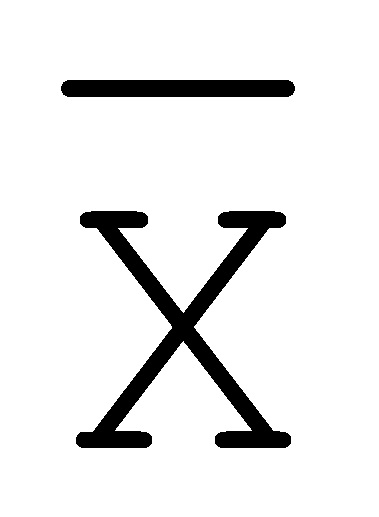 ), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (
), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (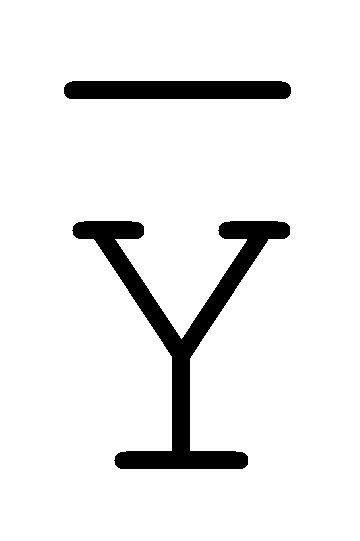 ::Y), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D::
::Y), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D:: ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (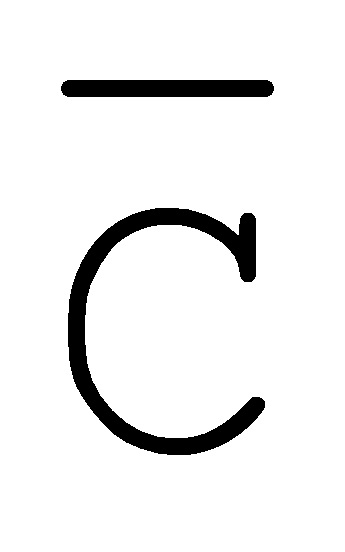 ::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; и
::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; и
(b) второй комплекс нуклеиновых кислот, содержащий (i) первую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как 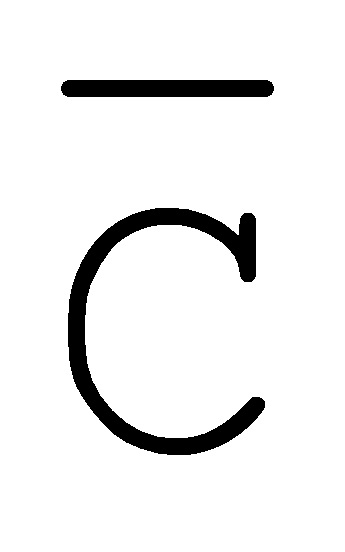 -
-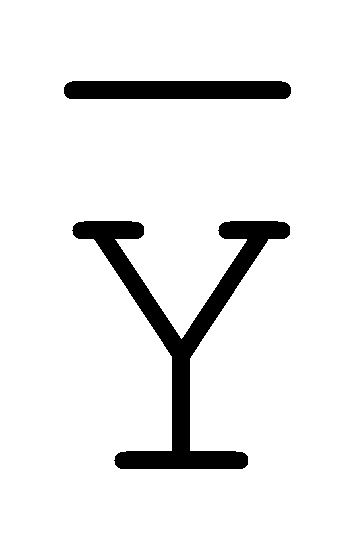 -
-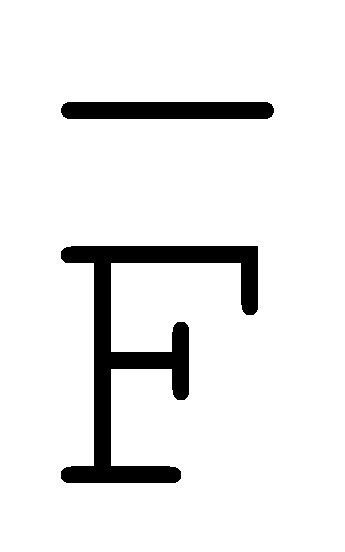 , (ii) вторую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как E-Y-D, (iii) третью цепь, последовательно содержащую первый и второй фрагменты и определенную как F-Z, и (iv) четвертую цепь, последовательно содержащую первый и второй фрагменты и определенную как
, (ii) вторую цепь нуклеиновой кислоты, последовательно содержащую первый, второй и третий фрагменты и определенную как E-Y-D, (iii) третью цепь, последовательно содержащую первый и второй фрагменты и определенную как F-Z, и (iv) четвертую цепь, последовательно содержащую первый и второй фрагменты и определенную как 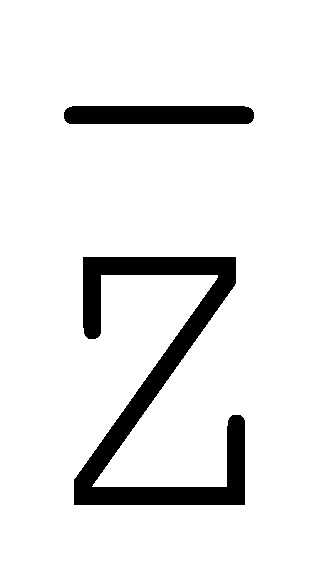 -
-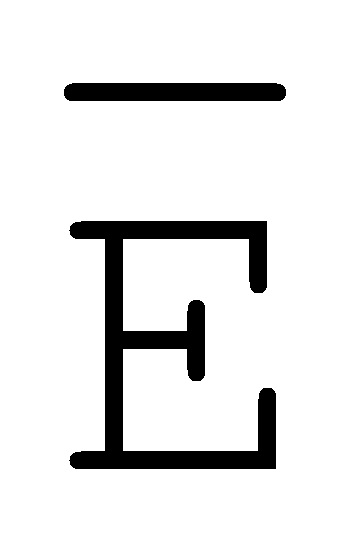 , где второй комплекс содержит первую область дуплекса (
, где второй комплекс содержит первую область дуплекса (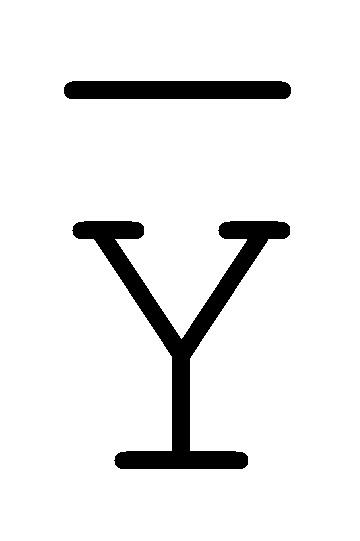 ::Y), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (Z::
::Y), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (Z:: ), сформированную между вторым фрагментом третьей цепи и первым фрагментом четвертой цепи, третью область дуплекса (
), сформированную между вторым фрагментом третьей цепи и первым фрагментом четвертой цепи, третью область дуплекса ( ::F), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (E::
::F), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (E:: ), сформированную между первым фрагментом второй цепи и вторым фрагментом четвертой цепи,
), сформированную между первым фрагментом второй цепи и вторым фрагментом четвертой цепи,
где первый (B) фрагмент первой цепи, третий фрагмент ( ) второй цепи, третий фрагмент (
) второй цепи, третий фрагмент (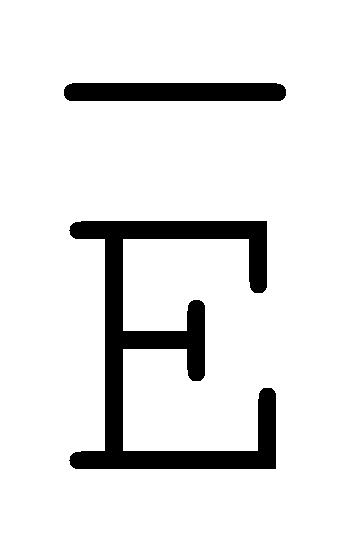 ) третьей цепи и первый фрагмент (F) четвертой цепи первого комплекса и первый (
) третьей цепи и первый фрагмент (F) четвертой цепи первого комплекса и первый (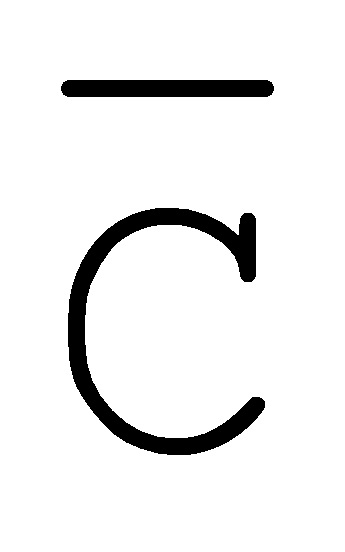 ) фрагмент первой цепи и третий фрагмент второй цепи (D) второго комплекса являются одноцепочечными;
) фрагмент первой цепи и третий фрагмент второй цепи (D) второго комплекса являются одноцепочечными;
где первый, второй и третий фрагменты ( ,
, 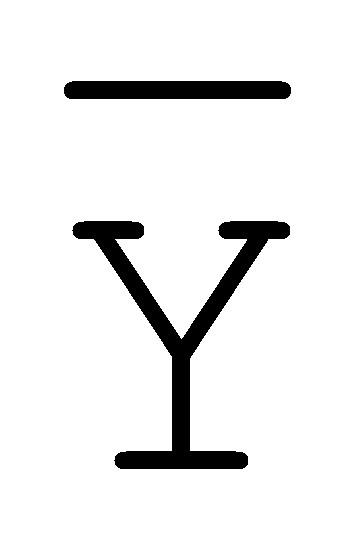 и
и 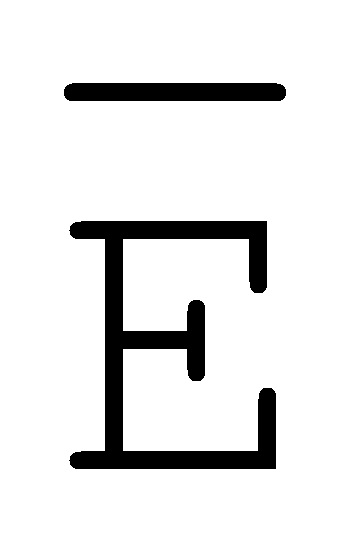 ) третьей цепи первого комплекса обладают комплементарностью последовательностей с третьим, вторым и первым фрагментами (D, Y и E) второй цепи второго комплекса для обеспечения связывания между ними, соответственно; и
) третьей цепи первого комплекса обладают комплементарностью последовательностей с третьим, вторым и первым фрагментами (D, Y и E) второй цепи второго комплекса для обеспечения связывания между ними, соответственно; и
где третий, второй и первый фрагменты (C, Y и F) четвертой цепи первого комплекса обладают комплементарностью последовательностей с первым, вторым и третьим фрагментами (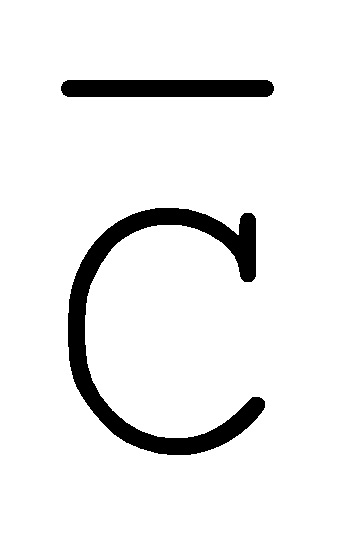 ,
, 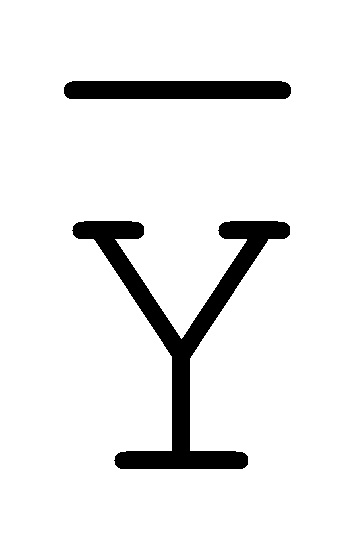 и
и 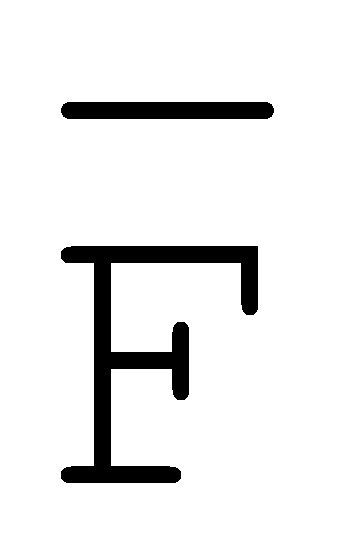 ) первой цепи второго комплекса для обеспечения связывания между ними, соответственно.
) первой цепи второго комплекса для обеспечения связывания между ними, соответственно.
7. Композиция по п. 6, где первый фрагмент (F) четвертой цепи первого комплекса по меньшей мере на 5 нуклеотидов (н.) длиннее, чем третий фрагмент (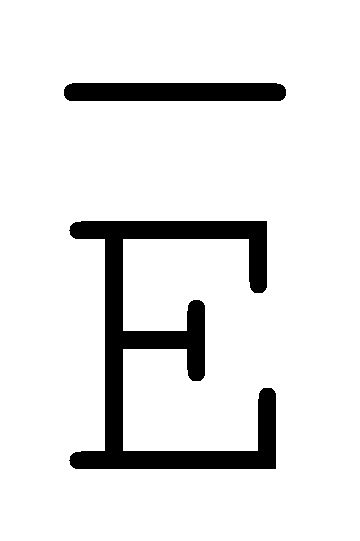 ) третьей цепи первого комплекса.
) третьей цепи первого комплекса.
8. Композиция по п. 6, где первый фрагмент (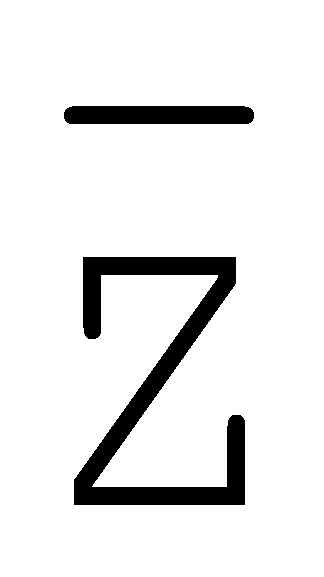 ) четвертой цепи второго комплекса длиннее по меньшей мере на 30 нуклеотидов (н.).
) четвертой цепи второго комплекса длиннее по меньшей мере на 30 нуклеотидов (н.).
9. Композиция по п. 6, где третий фрагмент (C) первой цепи первого комплекса и первый фрагмент () первой цепи третьего комплекса обладают комплементарностью последовательностей, но в виду химической модификации одного или двух из фрагментов не связываются друг с другом стабильно по сравнению со связыванием между третьим фрагментом (C) первой цепи первого комплекса и первым фрагментом () второй цепи второго комплекса; и первый фрагмент () второй цепи первого комплекса и третий фрагмент (D) первой цепи третьего комплекса обладают комплементарностью последовательностей, но в виду химической модификации одного или двух из фрагментов, друг с другом не связываются стабильно по сравнению со связыванием между первым фрагментом () второй цепи первого комплекса и третьим фрагментом (D) первой цепи второго комплекса.
10. Композиция по п. 9, где химическая модификация включает замену сахарофосфатного каркаса фрагмента нуклеиновой кислоты конъюгированными с мини-ПЭГ, получаемыми из серина гамма-ПНК.
11. Композиция по п. 10, где химическая модификация включает замену азотистого основания по меньшей мере одного нуклеозида во фрагменте нуклеиновой кислоты на трициклический аналог цитозина.
12. Композиция по п. 10, где химическая модификация включает введение гетероатома в 2'-положение молекулы сахара в нуклеотиде.
13. Комплекс нуклеиновых кислот для осуществления реакции замещения цепей нуклеиновых кислот, содержащий первую, вторую, третью, четвертую, пятую и шестую цепи нуклеиновых кислот, где:
первая цепь последовательно содержит первый, второй, третий и четвертый фрагменты и определена как 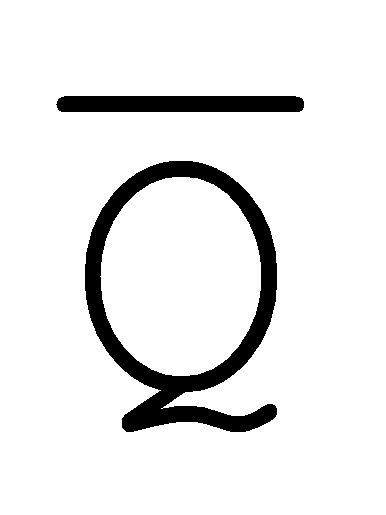 -
-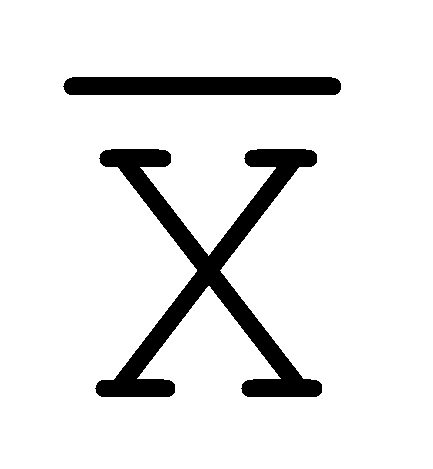 -
- -
- , где каждая буква означает фрагмент, а последовательность букв, соединенных "-", означает цепь;
, где каждая буква означает фрагмент, а последовательность букв, соединенных "-", означает цепь;
вторая цепь последовательно содержит первый, второй, третий и четвертый фрагменты и определена как  -
- -
-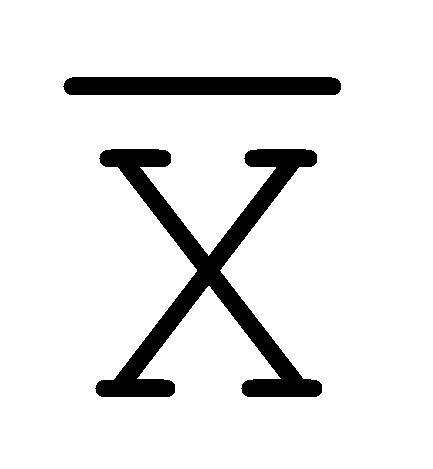 -P;
-P;
каждая из третьей и пятой цепей последовательно содержит первый, второй и третий фрагменты и определена как A-Y-C; и
каждая из четвертой и шестой цепей последовательно содержит первый, второй и третий фрагменты и определена как  -
- -B;
-B;
и где комплекс содержит:
первую область дуплекса ( :X), сформированную между вторым фрагментом первой цепи и третьей цепью второй цепи;
:X), сформированную между вторым фрагментом первой цепи и третьей цепью второй цепи;
вторую область дуплекса ( ::A), сформированную между четвертым фрагментом первой цепи и первой цепью третьей цепи;
::A), сформированную между четвертым фрагментом первой цепи и первой цепью третьей цепи;
третью область дуплекса ( ::B), сформированную между третьим фрагментом первой цепи и третьим фрагментом четвертой цепи;
::B), сформированную между третьим фрагментом первой цепи и третьим фрагментом четвертой цепи;
четвертую область дуплекса ( ::A), сформированную между вторым фрагментом второй цепи и первым фрагментом пятой цепи;
::A), сформированную между вторым фрагментом второй цепи и первым фрагментом пятой цепи;
пятую область дуплекса ( ::B), сформированную между первым фрагментом второй цепи и третьим фрагментом шестой цепи;
::B), сформированную между первым фрагментом второй цепи и третьим фрагментом шестой цепи;
шестую область дуплекса (Y:: ), сформированную между вторыми фрагментами третьей и четвертой цепей; и
), сформированную между вторыми фрагментами третьей и четвертой цепей; и
седьмую область дуплекса (Y:: ), сформированную между вторыми фрагментами пятой и шестой цепей,
), сформированную между вторыми фрагментами пятой и шестой цепей,
и где первый фрагмент ( ) первой цепи, четвертый фрагмент (P) второй цепи, третьи фрагменты (C) третьей и пятой цепей и первые фрагменты (
) первой цепи, четвертый фрагмент (P) второй цепи, третьи фрагменты (C) третьей и пятой цепей и первые фрагменты ( ) четвертой и шестой цепей являются одноцепочечными.
) четвертой и шестой цепей являются одноцепочечными.
14. Композиция для осуществления реакции замещения цепей нуклеиновых кислот, содержащая первый, второй и третий комплексы нуклеиновых кислот, где:
первый комплекс содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как B-W-D,  -
- -
- ,
,  -
- -H и
-H и  -Y-C, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где первый комплекс содержит первую область дуплекса (W::
-Y-C, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где первый комплекс содержит первую область дуплекса (W:: ), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (
), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса ( ::Y), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D::
::Y), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D:: ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса ( ::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи;
::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи;
второй комплекс содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как F-X-H,  -
- -
- ,
,  -
- -
- и J-Z-G, соответственно, где второй комплекс содержит первую область дуплекса (X::
и J-Z-G, соответственно, где второй комплекс содержит первую область дуплекса (X::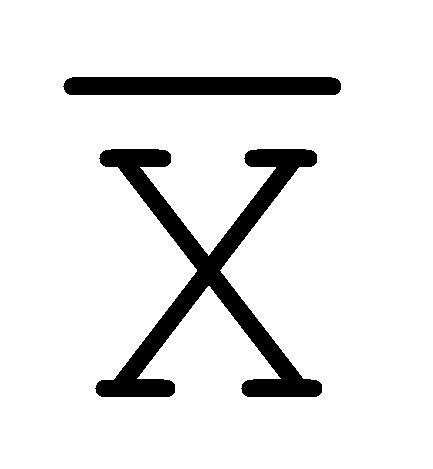 ), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (
), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса ( ::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (H::
::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (H::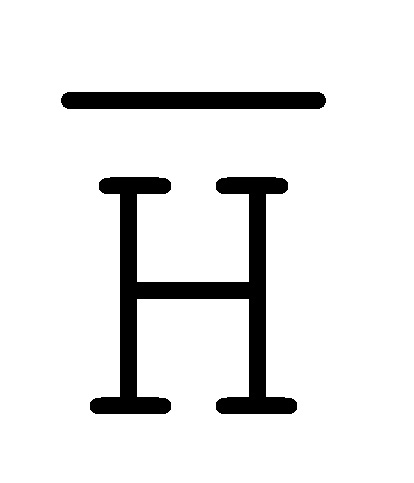 ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (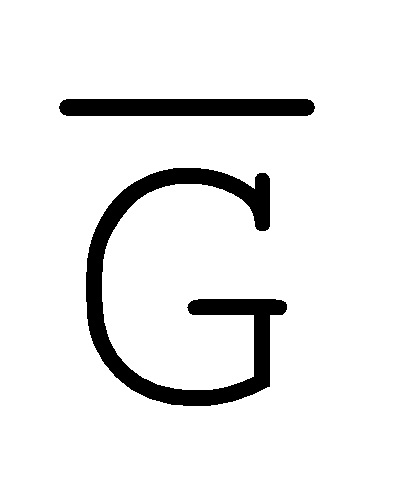 ::G), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи;
::G), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи;
третий комплекс содержит первую, вторую, третью, четвертую, пятую и шестую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как  -
-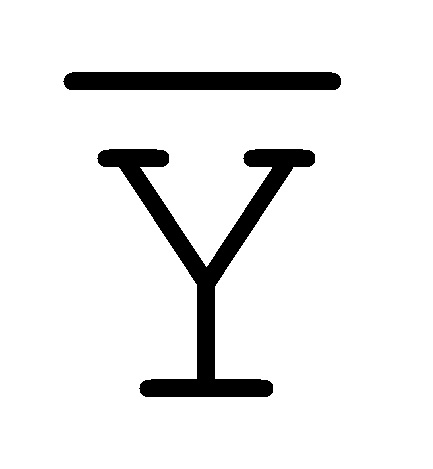 -G,
-G, 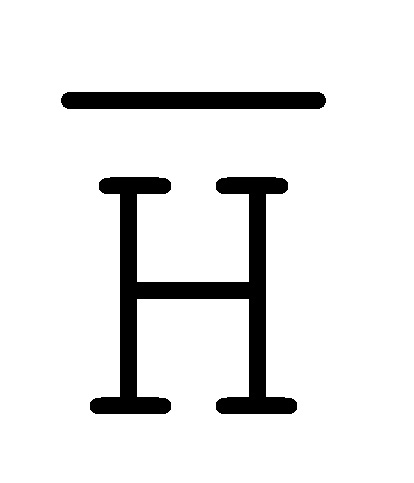 -Y-D,
-Y-D, 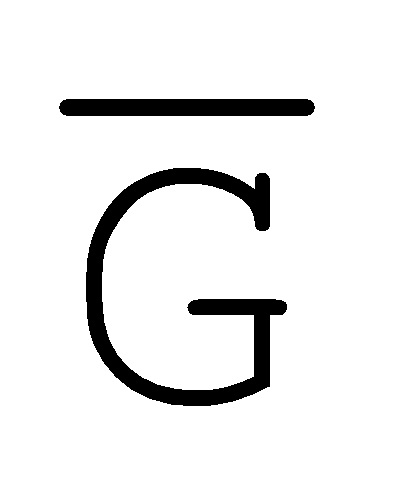 -
- -
- , I-Z-H, J-Q-M и
, I-Z-H, J-Q-M и  -
-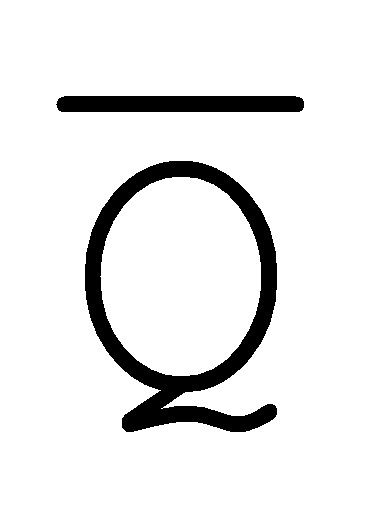 -
-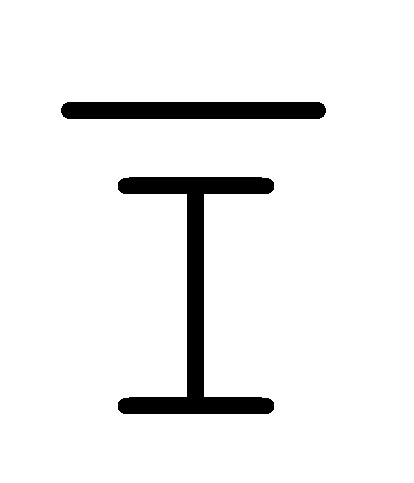 , соответственно, где третий комплекс содержит первую область дуплекса (
, соответственно, где третий комплекс содержит первую область дуплекса (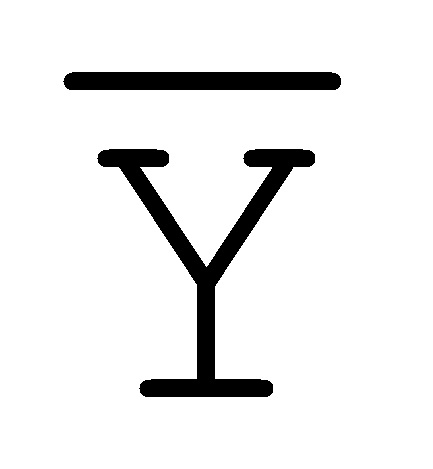 ::Y), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (
::Y), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса ( ::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (Q::
::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (Q::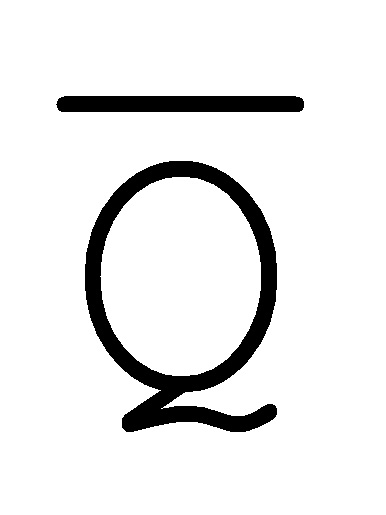 ), сформированную между вторыми фрагментами пятой и шестой цепей, четвертую область дуплекса (G::
), сформированную между вторыми фрагментами пятой и шестой цепей, четвертую область дуплекса (G::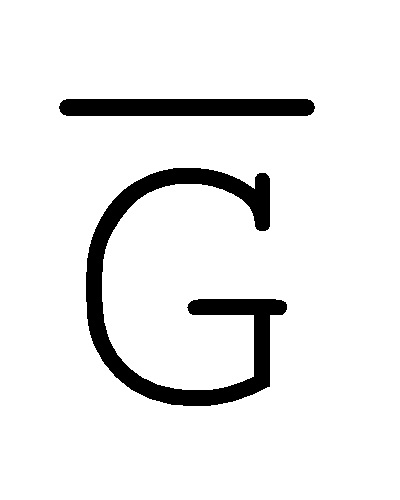 ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, пятую область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, пятую область дуплекса (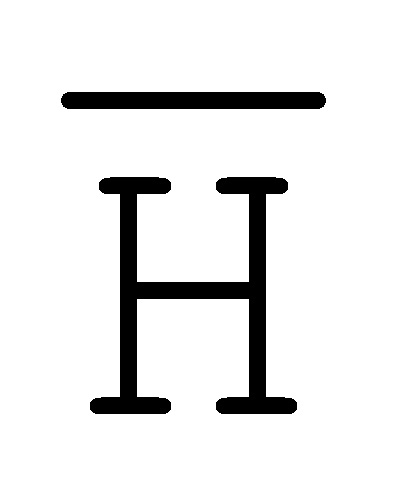 ::H), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи, шестую область дуплекса (
::H), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи, шестую область дуплекса ( ::J), сформированную между третьим фрагментом третьей цепи и первым фрагментом пятой цепи, и седьмую область дуплекса (I::
::J), сформированную между третьим фрагментом третьей цепи и первым фрагментом пятой цепи, и седьмую область дуплекса (I::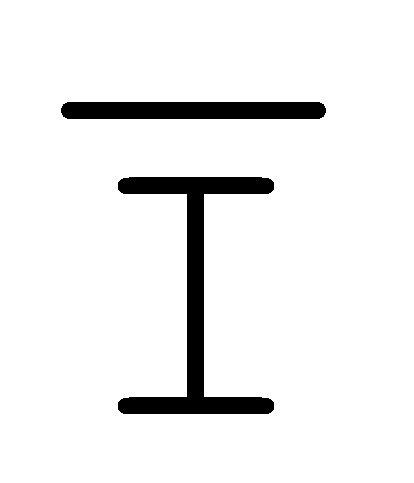 ), сформированную между первым фрагментом четвертой цепи и третьим фрагментом шестой цепи;
), сформированную между первым фрагментом четвертой цепи и третьим фрагментом шестой цепи;
где в первом комплексе первый фрагмент (B) первой цепи, третий фрагмент ( ) второго фрагмента, третий фрагмент (H) третьей цепи и первый фрагмент (
) второго фрагмента, третий фрагмент (H) третьей цепи и первый фрагмент (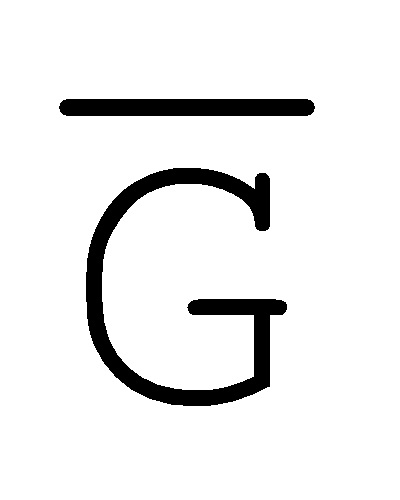 ) четвертой цепи являются одноцепочечными; во втором комплексе первый фрагмент (F) первой цепи, третий фрагмент (
) четвертой цепи являются одноцепочечными; во втором комплексе первый фрагмент (F) первой цепи, третий фрагмент ( ) второго фрагмента, третий фрагмент (
) второго фрагмента, третий фрагмент (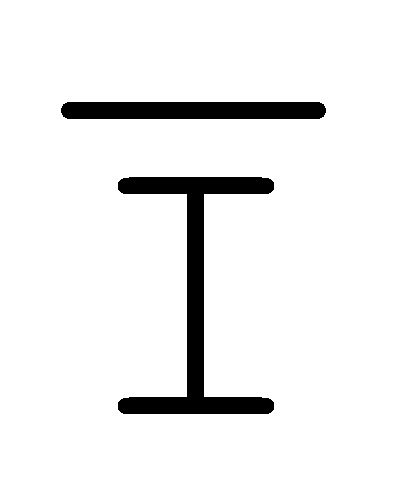 ) третьей цепи и первый фрагмент (J) четвертой цепи являются одноцепочечными; и в третьем комплексе первый фрагмент (
) третьей цепи и первый фрагмент (J) четвертой цепи являются одноцепочечными; и в третьем комплексе первый фрагмент ( ) первой цепи, третий (D) фрагмент второй цепи, третий фрагмент (M) пятого фрагмента и первый фрагмент (
) первой цепи, третий (D) фрагмент второй цепи, третий фрагмент (M) пятого фрагмента и первый фрагмент ( ) шестой цепи являются одноцепочечными,
) шестой цепи являются одноцепочечными,
и где:
третья цепь ( -
-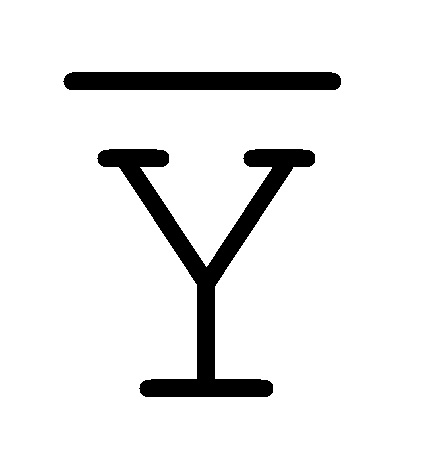 -H) первого комплекса и вторая цепь (
-H) первого комплекса и вторая цепь (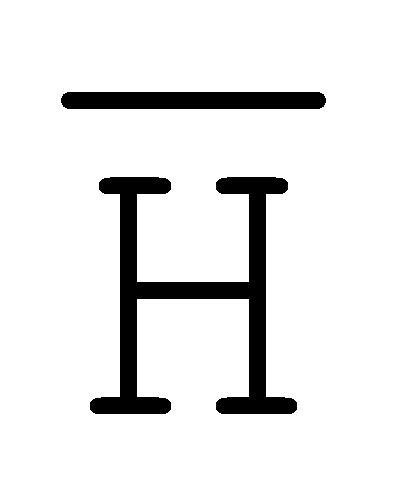 -Y-D) третьего комплекса обладают комплементарностью последовательностей для обеспечения связывания между ними;
-Y-D) третьего комплекса обладают комплементарностью последовательностей для обеспечения связывания между ними;
четвертая цепь (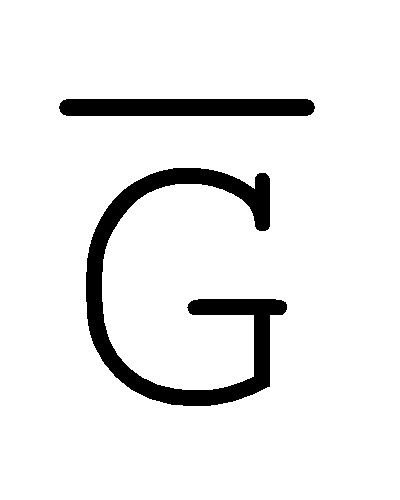 -Y-C) первого комплекса и первая цепь (
-Y-C) первого комплекса и первая цепь ( -
-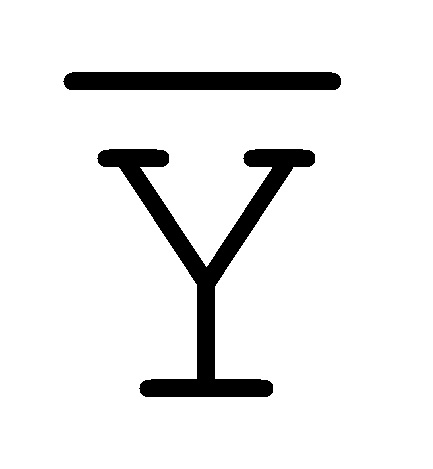 -G) третьего комплекса обладают комплементарностью последовательностей для обеспечения связывания между ними;
-G) третьего комплекса обладают комплементарностью последовательностей для обеспечения связывания между ними;
третья цепь (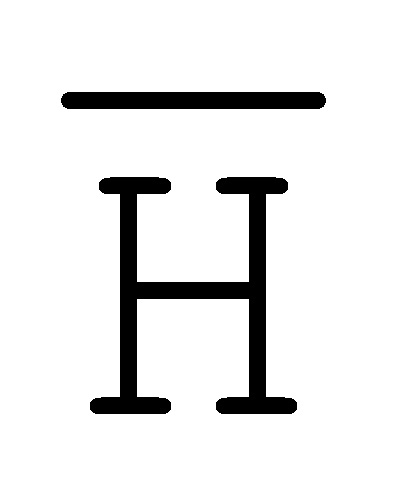 -
- -
- ) второго комплекса и четвертая цепь (I-Z-H) третьего комплекса обладают комплементарностью последовательностей для обеспечения связывания между ними; и
) второго комплекса и четвертая цепь (I-Z-H) третьего комплекса обладают комплементарностью последовательностей для обеспечения связывания между ними; и
четвертая цепь (J-Z-G) второго комплекса и третья цепь ( -
- -
- ) третьего комплекса обладают комплементарностью последовательностей для обеспечения связывания между ними.
) третьего комплекса обладают комплементарностью последовательностей для обеспечения связывания между ними.
15. Композиция для осуществления реакции замещения цепей нуклеиновых кислот, содержащая первый и второй комплексы нуклеиновых кислот, где:
первый комплекс содержит первую, вторую, третью и четвертую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как B-X-D, - -
- ,
,  -
- -
- и J-Z-C, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где первый комплекс содержит первую область дуплекса (X::
и J-Z-C, соответственно, где каждая буква означает фрагмент, а каждая последовательность букв, соединенных "-", означает цепь, и где первый комплекс содержит первую область дуплекса (X::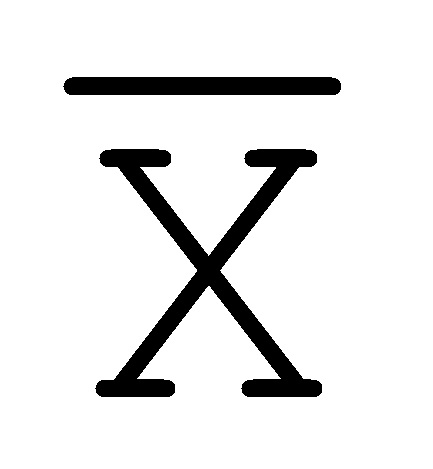 ), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (
), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса ( ::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D::
::Z), сформированную между вторыми фрагментами третьей и четвертой цепей, третью область дуплекса (D:: ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и четвертую область дуплекса ( ::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; и
::C), сформированную между первым фрагментом второй цепи и третьим фрагментом четвертой цепи; и
второй комплекс содержит первую и вторую цепи нуклеиновых кислот, где каждая последовательно содержит первый, второй и третий фрагменты и определена как F-Y-D и  -
-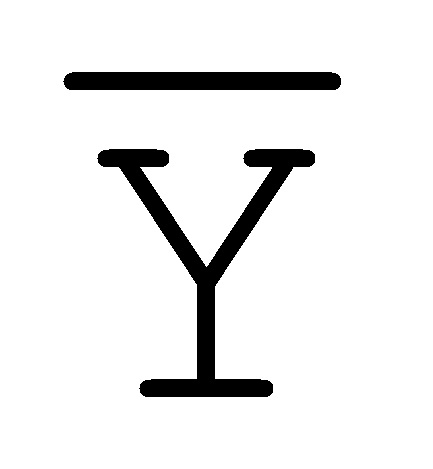 -
- , соответственно, и третью нуклеиновую кислоту, последовательно содержащую первый и второй фрагменты и определенную как
, соответственно, и третью нуклеиновую кислоту, последовательно содержащую первый и второй фрагменты и определенную как  -C, где второй комплекс содержит первую область дуплекса (Y::
-C, где второй комплекс содержит первую область дуплекса (Y::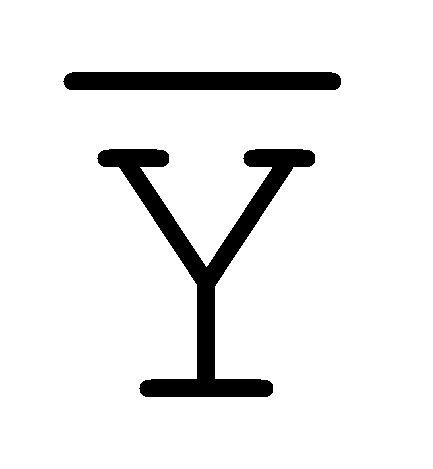 ), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (D::
), сформированную между вторыми фрагментами первой и второй цепей, вторую область дуплекса (D:: ), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и третью область дуплекса (
), сформированную между третьим фрагментом первой цепи и первым фрагментом третьей цепи, и третью область дуплекса ( ::C), сформированную между первым фрагментом второй цепи и вторым фрагментом третьей цепи;
::C), сформированную между первым фрагментом второй цепи и вторым фрагментом третьей цепи;
где первый фрагмент (B) первой цепи первого комплекса, третий фрагмент ( ) второй цепи первого комплекса, третий фрагмент (
) второй цепи первого комплекса, третий фрагмент (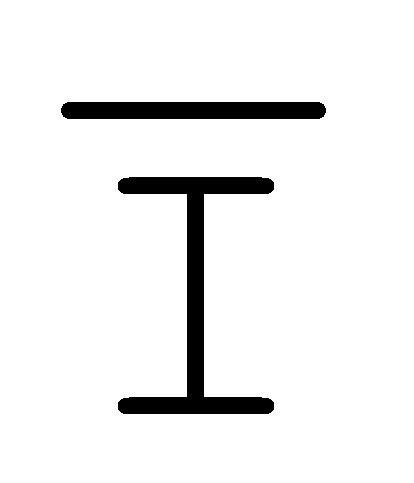 ) третьей цепи первого комплекса, первый фрагмент (J) четвертой цепи первого комплекса, первый фрагмент (F) первой цепи второго комплекса и третий фрагмент (
) третьей цепи первого комплекса, первый фрагмент (J) четвертой цепи первого комплекса, первый фрагмент (F) первой цепи второго комплекса и третий фрагмент ( ) второй цепи второго комплекса являются одноцепочечными;
) второй цепи второго комплекса являются одноцепочечными;
и где:
третья цепь ( -C) второго комплекса и комбинация первого фрагмента (
-C) второго комплекса и комбинация первого фрагмента ( ) третьей цепи первого комплекса и третьего фрагмента (C) четвертой цепи первого комплекса последовательно обладают подходящей идентичностью последовательности для осуществления их связывания с одной и той же последовательностью нуклеиновой кислоты-мишени.
) третьей цепи первого комплекса и третьего фрагмента (C) четвертой цепи первого комплекса последовательно обладают подходящей идентичностью последовательности для осуществления их связывания с одной и той же последовательностью нуклеиновой кислоты-мишени.
16. Композиция по п. 1, где длина каждого фрагмента составляет от приблизительно 5 оснований до приблизительно 50 оснований.
17. Композиция по п. 1, где длина каждого фрагмента, который является одноцепочечным, составляет от приблизительно 5 оснований до приблизительно 30 оснований.
18. Композиция по п. 1, дополнительно содержащая фармацевтически приемлемый носитель.
| WO2012151328 A2, 08.11.2012 | |||
| СПОСОБ АМПЛИФИКАЦИИ МАТРИЧНОЙ НУКЛЕИНОВОЙ КИСЛОТЫ | 2001 |
|
RU2284357C2 |

