Область техники, к которой относится изобретение
Данное изобретение относится к новым гуманизированным антителам против человеческого CD52 и их использованию для лечения или профилактики заболеваний человека.
Уровень техники
CD52 является гликозилированным белком клеточной поверхности, заякоренным гликозилфосфатидилинозитолом (ГФИ), который в больших количествах обнаружен на различных нормальных и злокачественных лимфоидных клетках, в частности, на В- и Т-клетках (Gilleece et al, Blood 82 807-812 (1993); Hale et al, J Biol Regul Homeost Agents, 15 стр. 386-391 (2001); Rodig et al, Clin Cancer Res 12, стр. 7174-7179 (2006)). CD52 экспрессируется на более низких уровнях на миелоидных клетках, например, моноцитах, макрофагах и дендритных клетках (ДК) с небольшой экспрессией, что можно видеть на зрелых естественных киллерах (ЕК), нейтрофилах и стволовых клетках крови. CD52 также продуцируется эпителиальными клетками в придатках яичка и семявыносящих протоках и захватывается спермой при прохождении через половой тракт (Haie et al, там же; Domagala et al, Med Sci Monit 7, стр. 325-331 (2001)). Точная биологическая функция CD52 остается неясной, но некоторые данные свидетельствуют о том, что он может участвовать в миграции и костимуляции Т-клеток (Masuyama et al., J Exp Med 189, 979-989 (1999); Watanabe et al, Clin Immunol 120 247-259 (2006)).
Campath-1H (алемтузумаб, Campath®, MabCampath®) представляет собой гуманизированное моноклональное антитело, действующее против человеческого CD52, которое демонстрирует сильную антителозависимую клеточно-опосредованную цитотоксичность (АЗКЦ) и комплемент-зависимую цитотоксичность (КЗЦ) in vitro. CD52 присутствует в по крайней мере 95% всех человеческих лимфоцитов периферической крови и моноцитов/макрофагов (Haie G. et al., CAMP ΑΤΗ-1 antigen (CD52). Tissue Antigens 1990,35:118-127). Campath-1H распознает эпитоп, который состоит из карбоксиконцевых четырех аминокислот зрелого белка CD52 и части отрицательно заряженного ГФИ-якоря. Благодаря своим значительным цитотоксическим эффектам Campath-1H может разрушать положительные клетки CD52 в живом организме, и одобрен для терапии первой линии и терапии третьей линии хронического лимфолейкоза (ХЛЛ). Campath-1H оценивали с точки зрения полезности в терапии некоторых аутоиммунных заболеваний, включая ревматоидный артрит, васкулит, миозит, болезнь Вегенера и диабет. Самые передовые исследования Campath-1H проводят в отношении терапии рецидивирующего рассеянного склероза (PC). Эти исследования показали значительное увеличение времени до развития рецидива по сравнению с человеческим интерфероном бета-1a (Rebif® (то есть, интерфероном бета-1а)).
Главное ограничение Campath-1H состоит в иммуногенности, в связи с чем антитела индуцируются у до 70% пациентов (Therapeutic Monoclonal Antibodies: From Bench to Clinic, ed. Zhiqiang An (2009) ISBN: 978-0-470-11791-0). Для улучшения клинического применения анти-СD52 антител крайне важно предложить улучшенные анти-СD52 антитела, которые не связаны с существенной иммуногенностью у пациентов.
Раскрытие изобретения
Изобретение относится к гуманизированным иммуноглобулинам, которые имеют специфическое связывание с человеческим CD52 (huCD52). В данном изобретении также предложены гуманизированные антитела, которые связываются с человеческим CD52 с равновесной константой диссоциации (Kd) не менее 10-8 М. В данном изобретении также предложены гуманизированные антитела, которые специфически связываются с человеческим CD52, либо имеющие тяжелую цепь антител IgG1, IgG2, IgG3 или IgG4, либо использующие мутированную константную область IgG, в частности, константную область, которая повышает АЗКЦ (антителозависимую клеточную цитотоксичность) или КЗЦ (комплемент-зависимую цитотоксичность). В данном изобретении также предложены гуманизированные антитела, у которых легкая цепь представляет собой каппа легкую цепь. Гуманизированное антитело может быть закодировано нуклеиновыми кислотами человеческой тяжелой цепи IgG и человеческой каппа легкой цепи, которые кодируют белковые последовательности в своих вариабельных областях, как указано в SEQ ID NO: 20 - SEQ ID №: 28.
В настоящем изобретении также предложены гуманизированные антитела, которые специфически связываются с человеческим CD52, за счет чего вариабельные области антител были селектированы или модифицированы таким образом, чтобы исключить один или несколько эпитопов для человеческого CD4+Τ клетки. В настоящем изобретении также предложены гуманизированные антитела, которые специфически связываются с человеческим CD52, в результате чего образуются вариабельные области антител, главным образом, посредством слияния сегментов последовательностей, полностью полученных из существующих последовательностей вариабельных областей человеческих антител.
В настоящем изобретении также предложены гуманизированные анти-СD52 антитела по изобретению, содержащие аминокислотные последовательности гипервариабельных участков CDR1, CDR2 и CDR3 тяжелой цепи, RYGMS (SEQ ID №5), MMKTKGGRTYYPDSVKG (SEQ ID №6) и DGYY (SEQ ID №7), соответственно, и аминокислотные последовательности CDR1, CDR2, CDR3 легкой цепи KSSQSLLHSDGKTYLN (SEQ ID №8), LVSKLDS (SEQ ID №9) и WQGTHLWT (SEQ ID №10), соответственно. В настоящем изобретении также предложены гуманизированные анти-СD52 антитела по изобретению, содержащие аминокислотные последовательности вариабельной области тяжелой цепи, соответствующие SEQ ID №№20-24 для тяжелой цепи и SEQ ID №№25-28 для легкой цепи. В предпочтительном варианте изобретения предложено гуманизированное анти-СD52 антитело по настоящему изобретению, содержащее аминокислотные последовательности вариабельной области тяжелой цепи, соответствующее SEQ ID №22 для тяжелой цепи и SEQ ID №28 для легкой цепи.
Гуманизированные антитела по настоящему изобретению могут состоять из любой из вышеуказанных последовательностей гипервариабельных участков SEQ ID №5 - SEQ ID №10 и вариантов этих последовательностей гипервариабельных участков с незначительными отклонениями, в которых изменения одной или нескольких аминокислот не снижают существенно связывание с человеческим CD52. Гуманизированные антитела могут быть созданы путем соединения последовательностей гипервариабельных участков с каркасами вариабельных областей человека, если такие каркасные последовательности получены из одной или нескольких иных каркасных последовательностей вариабельной области человеческих антител. Обычно такие каркасные последовательности вариабельных областей человеческих антител содержат одну или несколько мутаций, которые способствуют оптимальному или улучшенному связыванию гуманизированных антител с CD52. В предпочтительном варианте настоящего изобретения такие каркасные последовательности вариабельных областей человека в гуманизированных антителах получены полностью из последовательностей в других вариабельных областях человеческих антител, как описано в ЕР 1844074. Эти последовательности содержат соединенные сегменты последовательностей из других вариабельных областей человеческих антител вместе с константными областями антител человека. В частности, такие гуманизированные антитела также содержат последовательности гипервариабельных участков, полученные полностью из последовательностей гипервариабельных участков в других вариабельных областях человеческих антител, в том числе соединенные сегменты последовательностей других человеческих гипервариабельных участков вместе с человеческими константными областями, вследствие чего образуются гуманизированные антитела, в которых последовательности вариабельных областей полностью получены из последовательностей в других вариабельных областях человеческих антител вместе с человеческими константными областями, что создает «полностью человеческие» антитела.
В настоящем изобретении также предложены гуманизированные антитела, которые специфически связываются с человеческим CD52, при этом указанное гуманизированное антитело получают с помощью прокариотической или эукариотической клетки, в частности, из линии клеток млекопитающих, в частности, клеток яичников китайского хомячка (СНО) или миеломы мыши (NS0). В изобретении также предложено гуманизированное антитело, которое специфически связывается с человеческим CD52, который представляет собой фрагмент участка связывания антигена Fab или одноцепочечный вариабельный фрагмент (scFv). В изобретении также предложены мультиспецифические белки, содержащие по крайней мере одно гуманизированное антитело из последовательностей SEQ ID №№: 20-24 для тяжелой цепи и SEQ ID №№: 25-28 для легкой цепи, в котором мультиспецифический белок специфически связывается с человеческим CD52 и, дополнительно, связывается или взаимодействует с одной или несколькими другими молекулами. Различные антитела или белки, которые могут быть включены в каждое мультиспецифическое антитело, могут быть связаны друг с другом либо ковалентно, либо нековалентно.
В изобретении также предложен лекарственный препарат, содержащий гуманизированное антитело (в виде либо белковоподобного антитела, либо гена, кодирующего антитело), который специфически связывается с человеческим CD52 и фармацевтически приемлемый носитель. Лекарственный препарат может дополнительно содержать одно или несколько химиотерапевтических средств, либо связанных, либо не связанных с гуманизированным антителом.
В изобретении предложен способ лечения ХЛЛ и других лейкозов; нескольких аутоиммунных заболеваний, включая рассеянный склероз, ревматоидный артрит, васкулит, миозит, болезнь Вегенера и диабет; и отторжения трансплантатов органов и реакции «трансплантат против хозяина», в каждом случае включающий в себя введение пациенту эффективной дозы гуманизированного антитела (в виде либо белковоподобного антитела, либо гена, кодирующего антитело), который специфически связывается с человеческим CD52, при этом антитело вызывает разрушение или апоптоз клеток-мишеней CD52+, например, В- и Т-клеток. Кроме того, в изобретении также предложен способ диагностики вышеуказанных заболеваний, например, посредством введения гуманизированного антитела, прикрепленного к детектируемой метке, и определения связывания гуманизированного антитела in vivo для обеспечения основы для выявления клеток CD52+, например, в локализованных опухолевых образованиях или в воспалительных поражениях. Кроме того, гуманизированные антитела по настоящему изобретению могут быть использованы для испытаний in vitro для клеток CD52+ в качестве средства для выявления заболеваний, а также для испытаний in vitro антител, которые могут связываться с гуманизированными антителами, используемыми в терапевтических целях. Соответственно, такие гуманизированные антитела по изобретению могут быть использованы в качестве диагностических или терапевтических агентов в живом организме in vivo и in vitro.
Гуманизированные антитела по изобретению могут включать в себя различные изотипы антител или их смеси, например, IgG1, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgAsec, IgD, IgE, или мутированные формы этих IgG, например мутации, которые увеличивают связывание с Fc-рецепторами (например, Horton et al., Blood 116 (2010), стр. 3004-3012) или с комплементом (например, Natsume et al., Cancer Res 68 (2008), стр. 3863-3872). Обычно гуманизированные антитела включают в себя константные области тяжелой цепи IgG1 и константные области κ-легкой цепи. Гуманизированные антитела могут иметь полную длину (например IgG1/κ антитело) или могут содержать только антигенсвязывающую часть (например, фрагмент Fab, F(ab′)2, Fv или scFv).
Некоторые гуманизированные анти-СD52 антитела по настоящему изобретению можно охарактеризовать одним или несколькими из следующих свойств: а) специфичность к человеческому CD52 (специфическое связывание с человеческим CD52); b) способность связывания с человеческим CD52 с равновесной константой диссоциации (Kd) не менее 10-8 М.
В другом аспекте изобретения предложены молекулы нуклеиновых кислот, кодирующие гуманизированные антитела или антигенсвязывающие части по изобретению. Соответственно, рекомбинантные экспрессирующие векторы, которые содержат нуклеиновые кислоты, кодирующие антитела по изобретению, а также клетки-хозяева, трансфицированные такими векторами, также включены в настоящее изобретение, также как и способы получения антител по изобретению путем культивирования таких клеток-хозяев.
Гуманизированные моноклональные антитела против человеческого CD52 по изобретению или их антигенсвязывающие участки (например, Fab), могут быть получены или связаны с другой функциональной молекулой, например, другим пептидом или белком (например, фрагментом Fab′). Например, антитело или антигенсвязывающая часть гуманизированных антител по изобретению могут быть функционально связаны (например, путем химического связывания, слияния генов, нековалентной связи или иным образом) с одной или несколькими другими молекулярными структурами. Например, гуманизированное анти-СD52 антитело или его антигенсвязывающий фрагмент могут быть конъюгированы с терапевтической функциональной группой, например, цитотоксическим лекарственным средством, ферментативно активным токсином или его фрагментом, радиоизотопом, терапевтической нуклеиновой кислотой или низкомолекулярным противоопухолевым препаратом. Антитела по изобретению также могут быть конъюгированы с цитотоксическими препаратами, например, мечеными радиоактивными изотопами с цитотоксическими средствами, например, I, или могут быть связаны с белком, инактивирующим рибосому, например, экзотоксином синегнойной палочки (фрагмент РЕ38, растительные или бактериальные токсины, например, рицин, α-цепь рицина, сапорин, антивирусный белок фитолакки, дифтерийный токсин или экзотоксин A Pseudomonas (Kreitman and Pastan (1998) Adv. Drug Delivery Rev. 31:53.).
В другом аспекте в настоящем изобретении предложены композиции, например, фармацевтические и диагностические композиции, содержащие фармацевтически приемлемый носитель и по крайней мере одно гуманизированное моноклональное антитело по изобретению, или его антигенсвязывающую часть, которые специфически связываются с человеческим CD52. Некоторые композиции также могут содержать комбинацию гуманизированных антител или их антигенсвязывающих участков по изобретению. Такие композиции могут также представлять собой комбинации одной или нескольких других биологически активных молекул в виде отдельных молекул, например, комбинацию по меньшей мере одного гуманизированного моноклонального антитела по изобретению и иной биологически активной молекулы, или могут содержать комбинации одной или нескольких других биологически активных молекул в одной и той же молекуле, например, в виде биспецифической или мультиспецифической молекулы, либо в виде комбинации двух или нескольких гуманизированных антител по изобретению, или в виде комбинации одной или нескольких иных биологически активных молекул.
Для методов in vivo гуманизированные антитела или их антигенсвязывающие участки (либо биспецифическая, либо мультиспецифическая молекула по изобретению) могут быть введены человеку, страдающему заболеванием, связанным с клетками CD52+, или заболеванием, которое можно облегчить или предотвратить путем терапии гуманизированными антителами по изобретению.
Композиции гуманизированных моноклональных антител также можно вводить в сочетании с другими известными терапиями, например, противораковой терапией, терапией аутоиммунных заболеваний, таких как ревматоидный артрит, или терапией рассеянного склероза. Соответственно, в настоящем изобретении предложен способ лечения рака или воспалительных заболеваний у пациента, включающий в себя введение пациенту терапевтически эффективного количества фармацевтической композиции гуманизированного антитела вместе с фармацевтическим носителем.
В еще одном аспект настоящего изобретения предложен способ, в котором антитела по изобретению используют для обнаружения in vitro или in vivo наличия антигена человеческого CD52 в образце, например, для диагностики заболевания, связанного с человеческим CD52. В некоторых методах это обеспечивается путем контакта тестируемого образца, вместе с контрольным образцом, с гуманизированным моноклональным антителом по изобретению или его антигенсвязывающим участком (или биспецифической или мультиспецифической молекулой), в условиях, обеспечивающих возможность образования комплекса между антителом и человеческим CD52. Затем выполняют обнаружение комплексообразования (например, с помощью иммуносорбентного ферментного анализа) в тестовых образцах, и любое статистически значимое увеличение образования комплексов между тестовым и контрольным образцами указывает на наличие антигена человеческого CD52 в тестовом образце.
Специалистам в соответствующей области техники очевидно, что гуманизированные антитела по настоящему изобретению будут иметь дополнительное использование для иных целей или в других составах, помимо описанных в настоящем документе, во всех случаях, когда гуманизированное антитело связывается с антигеном к человеческому CD52, и такие способы использования и составы включены в объем изобретения. Специалистам в данной области техники очевидно, что последовательности вариабельных областей гуманизированных антител по настоящему изобретению (SEQ ID №: 20 - SEQ ID №: 28) или гипервариабельные участки гуманизированных антител по настоящему изобретению (SEQ ID №: 5 - SEQ ID №: 10) могут быть подвержены вариациям, которые не меняют в значительной степени свойства гуманизированных антител по настоящему изобретению, вследствие чего такие варианты также входят в объем настоящего изобретения. Кроме того, такие изменения либо в вариабельной области, либо в последовательностях гипервариабельного участка гуманизированных антител входят в объем настоящего изобретения, если они в значительной степени гомологичны последовательностям гуманизированных антител по настоящему изобретению. Например, может быть определено, что вариантная нуклеиновая кислота входит в объем настоящего изобретения, если она содержит последовательности, содержащие или по существу идентичные SEQ ID №: 11 - SEQ ID №: 19, как определено на основании ее способности к гибридизации в строгих условиях с нуклеиновой кислотой по настоящему изобретению. В одном варианте может быть определено, что последовательность нуклеиновой кислоты входит в объем настоящего изобретения (например, по существу идентичная SEQ ID №: 11 - SEQ ID №: 19), на основании ее способности к гибридизации в строгих условиях с нуклеиновой кислотой, входящей в объем изобретения (например, SEQ ID №: 11 - SEQ ID №: 19). Термин «гибридизация» относится к связыванию, дупликации или гибридизации молекулы к определенной нуклеотидной последовательности при жестких условиях гибридизации, когда такая последовательность присутствует в сложной смеси (например, общая ДНК или РНК клетки или библиотеки), при этом определенная нуклеотидная последовательность обнаружена в объеме, по крайней мере, приблизительно в 10 раз больше фонового значения. Строгими условиями гибридизации могут быть выбраны, например, температура на 5-10°С ниже, чем точка плавления (Tm) для конкретной последовательности при определенной ионной силе рН. Также следует понимать, что гуманизированные антитела по настоящему изобретению могут быть модифицированы в константных областях тяжелой цепи для улучшения АЗКЦ и КЗЦ. Для увеличения АЗКЦ формы гуманизированных антител, лишенные фукозы, могут быть получены путем экспрессии антител в определенных клетках млекопитающих, в том числе вариантной линии СНО, Lec13 (Shields et al, J Biol Chem 277 (2002) стр. 26733-26740), линии клеток крысиной гибридомы YB2/0 (Shinkawa et al., J Biol Chem 278 (2003) стр. 3466-3473) и FUT8 (a-1,6-фукозилтрансфераза) линии клеток нокаутных СНО (Yamane-Ohnuki et al., Biotechnol Bioeng 87 (2004) стр. 614-622). Кроме того, мутации в константных областях тяжелой цепи могут быть использованы для увеличения АЗКЦ, как описано у Shields et al., J Biol Chem 276 (2001) стр. 6591-6604 и Lazar et al,. Proc Natl Acad Sci USA 2006; 103 (2006) p 4005-4010. В ином случае, мутации в константных областях тяжелой цепи могут быть использованы для повышения КЗЦ, например, посредством использования антител смешанного человеческого изотипа IgG1/IgG3 (Natsume et al., там же).
Специалистам в данной области техники понятно, на основании других известных источников, в частности, клинических исследований с Campath-1H (An Zhiqiang, там же), что антитела, которые связываются с антигеном к человеческому CD52, в основном иммуногены у пациентов, вероятно, из-за свойственной цитотоксичности анти-СD52 антител, которая действует как со-стимулирующий сигнал на эпитопы к CD4+Т-клеткам антител, в результате чего возникают ответы CD4+Т-хелперных клеток и иммуногенность. Таким образом, специалистам в данной области техники очевидно, что антитела по настоящему изобретению удивительно лишены таких ответов СD4+Т-хелперных клеток, которые были выявлены в исследованиях in vitro с человеческой кровью (см. пример 9) и что такие анти-СD52 антитела с низкими ответами CD4+T-клеток (<=4% ответов Т-клеток в исследованиях человеческих Т-клеток) являются новыми.
Краткое описание чертежей
В условных обозначениях, прилагаемых к фигурам, обозначения 2Е8 или ANT01 использованы взаимозаменяемо для обозначения гуманизированных антител мышей химерных или гуманизированных антител, полученных из моноклональных антител мышей 2Е8.
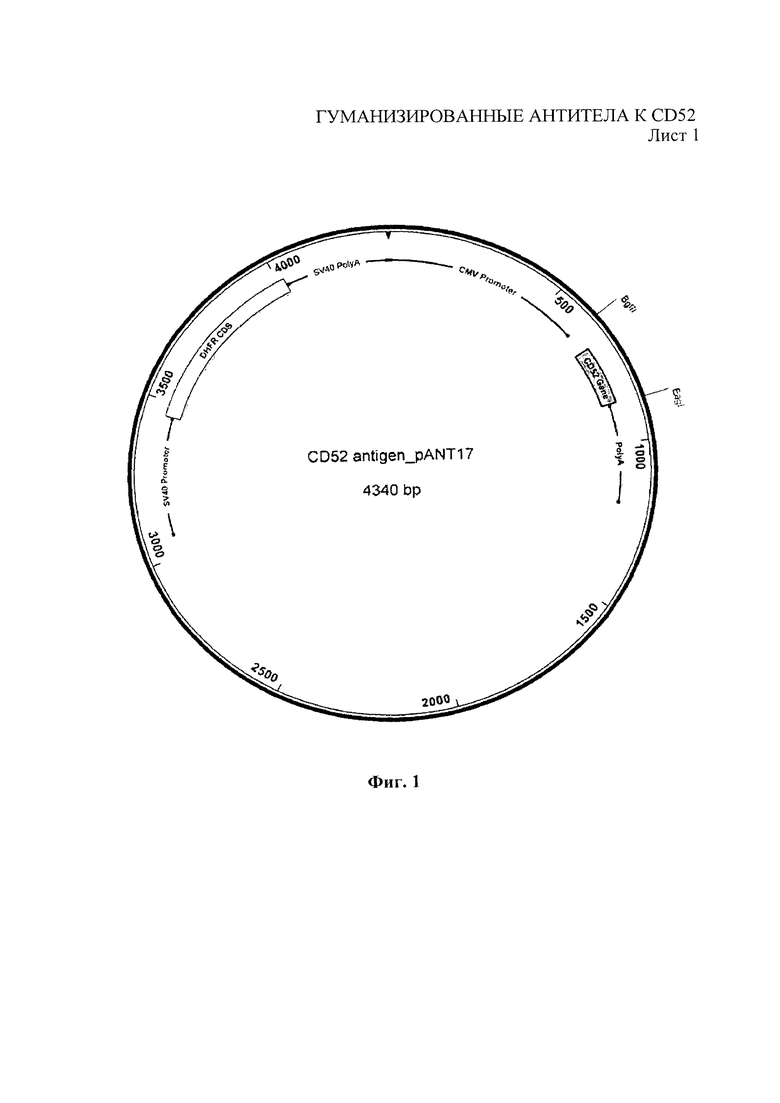
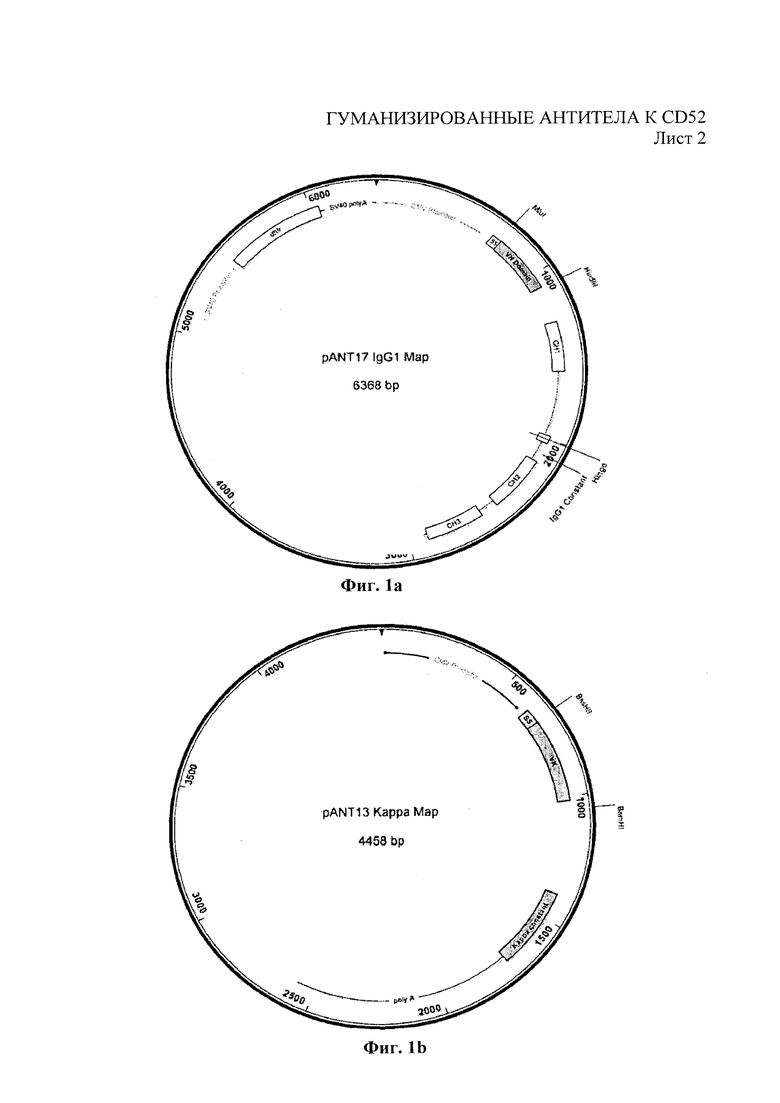
На Фиг. 1 показаны плазмидные векторы, используемые для экспрессии химерных и гуманизированных антител в клетках млекопитающих, содержащие pANT17 для тяжелых цепей и pANT13 для легких цепей.
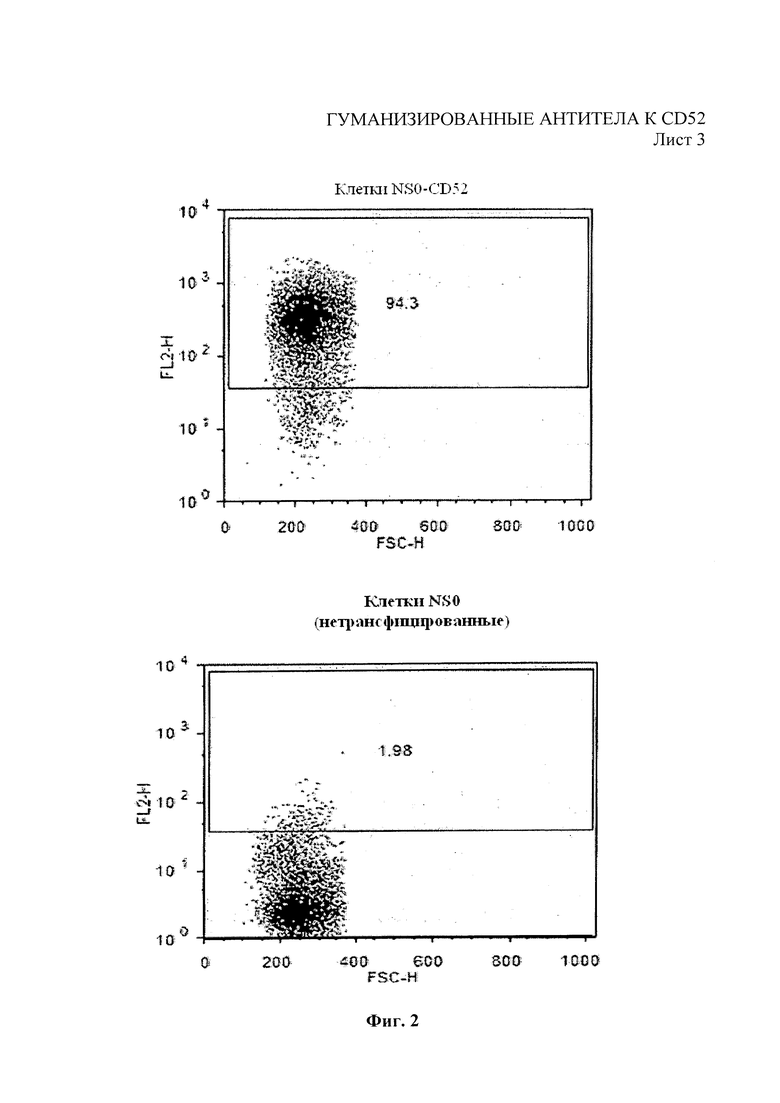
На Фиг. 2 показан анализ связывания моноклонального антитела мышей 2Е8 с NS0 клетками, трансфицированными человеческим CD52, с помощью проточной цитометрии, в сравнении со связыванием с NS0 CD52-. Окрашивание выполнено с конъюгированным антителом антимышиным IgG-PE, с сигналом, полученным из РЕ по оси Y.
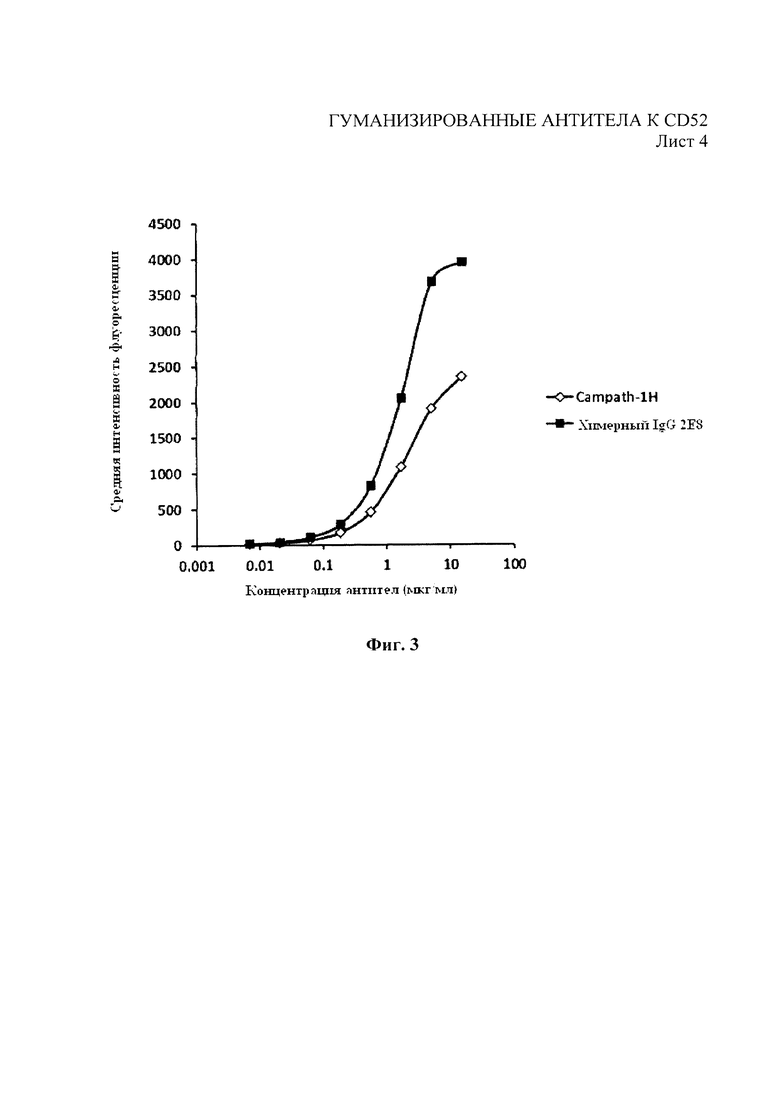
На Фиг. 3 показан анализ связывания разведений химерного 2Е8 в клетках Hut78 с помощью проточной цитометрии в сравнении с Campath-1Н. Окрашивание выполнено с конъюгированным антителом против IgG-PE человека.
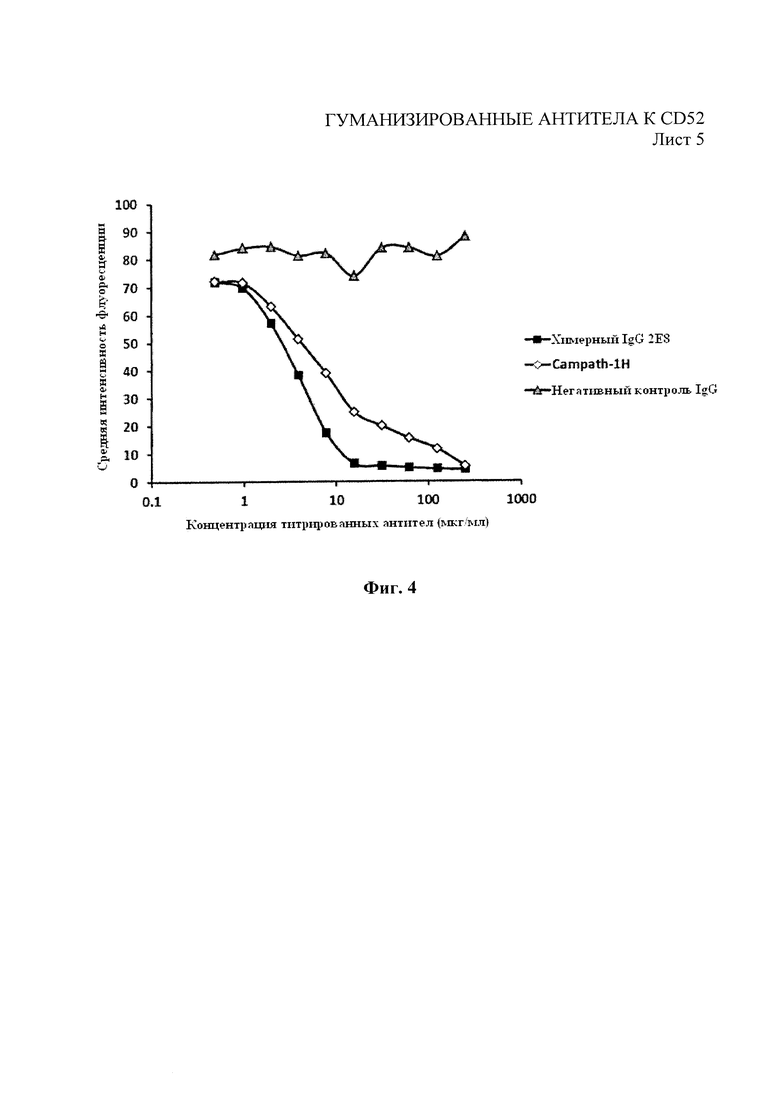
На Фиг. 4 показан анализ конкурентного связывания с помощью проточной цитометрии с использованием Campath-1H-PE в конкуренции с химерным 2Е8 и Campath-1H за связывание с клетками Hut78.
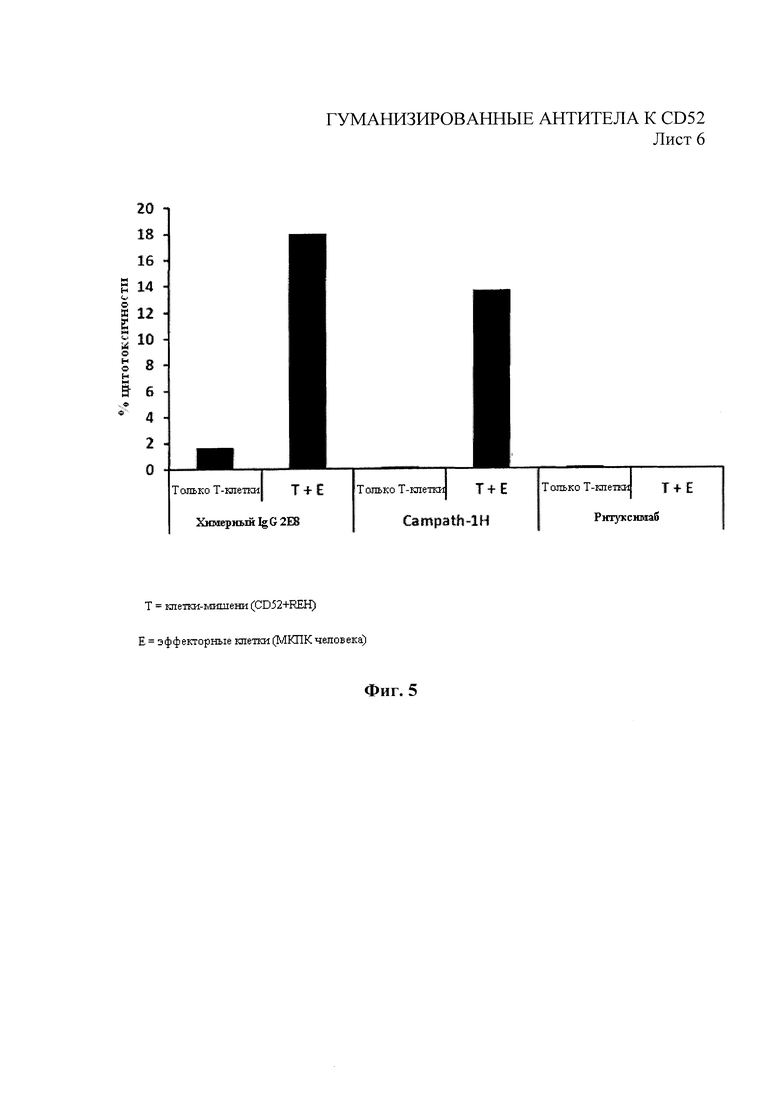
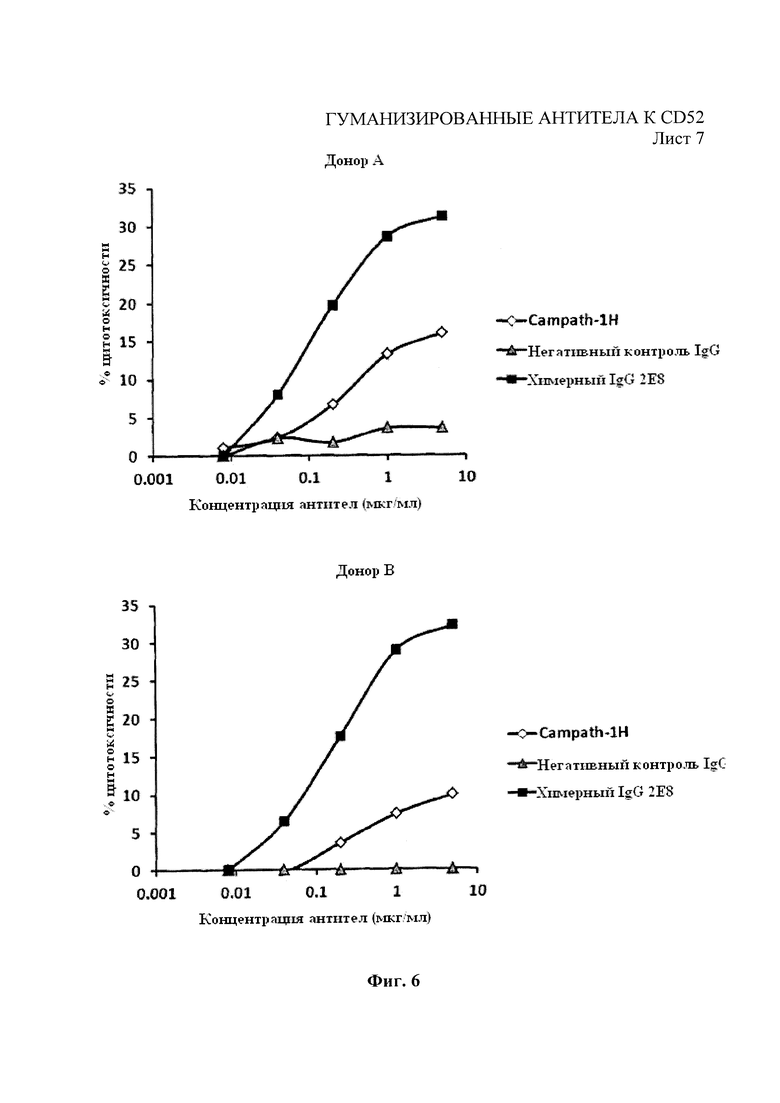
На Фиг. 5 показана средняя цитотоксичность 5 образцов МКПК человека, использованных в качестве эффекторных клеток в анализе АЗКЦ для химерных 2Е8 и Campath-1H с клетками-мишенями REH.
Фиг. 6 аналогична Фиг. 5, за исключением 2 отдельных МКПК с разведениями химерных 2Е8 и Campath-1H с клетками-мишенями REH с высокой экспрессией.
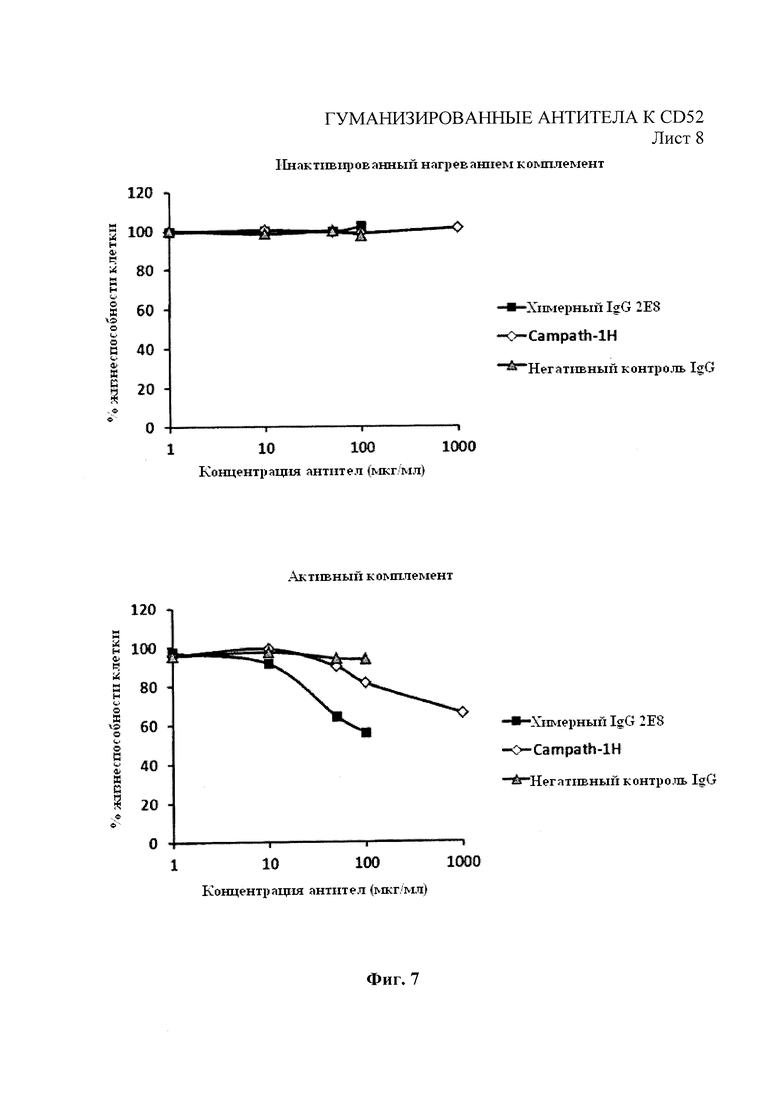
Фиг. 7 аналогична Фиг. 6, за исключением использования человеческого комплемента для анализов КЗЦ с разведениями химерных 2Е8 и Campath-1H и клеток-мишеней REH с высокой экспрессией.
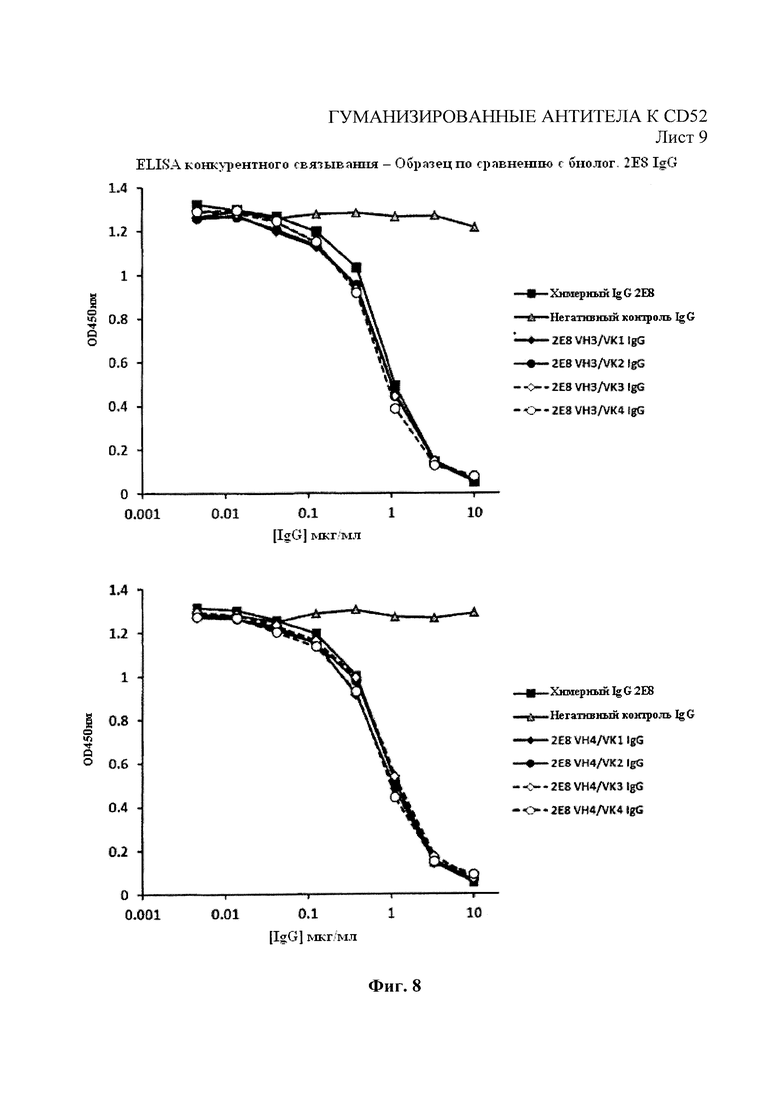
На Фиг. 8 показан иммуносорбентный ферментный анализ конкурентного связывания пептидов CD52 в отношении связывания гуманизированных вариантов 2Е8 в конкуренции с биотинилированными химерными 2Е8.
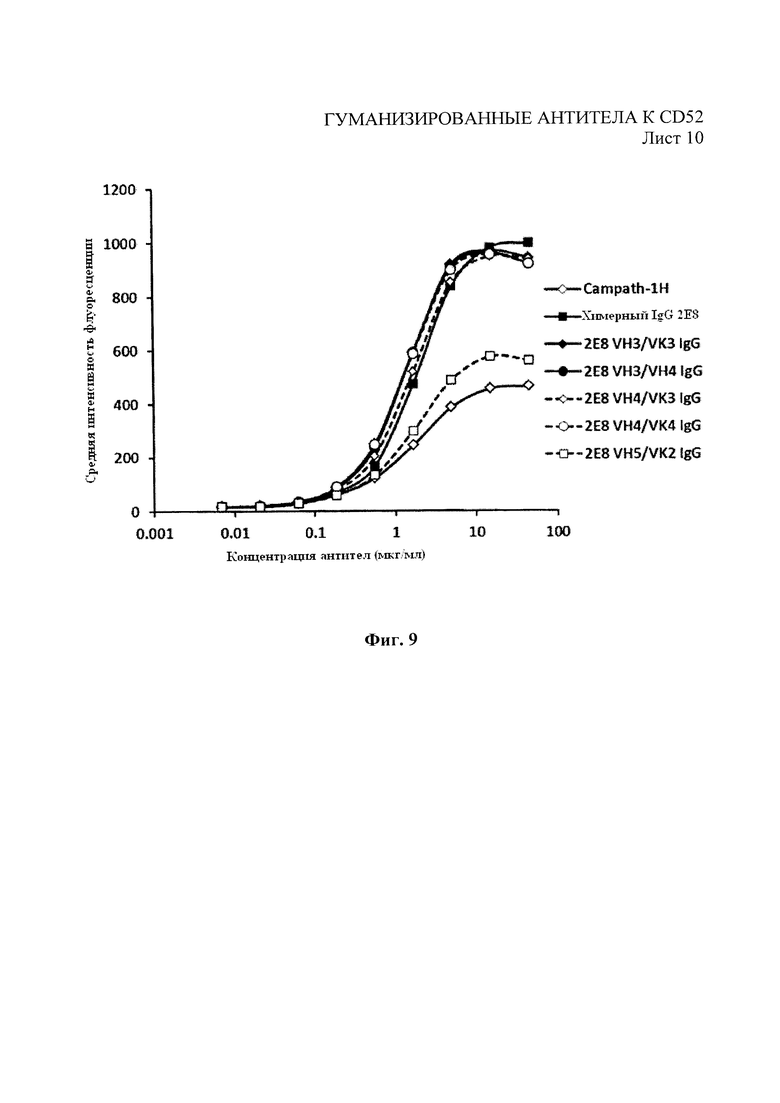
На Фиг. 9 показан анализ с помощью проточной цитометрии связывания разведений гуманизированных вариантов и Campath-H с клетками REH.
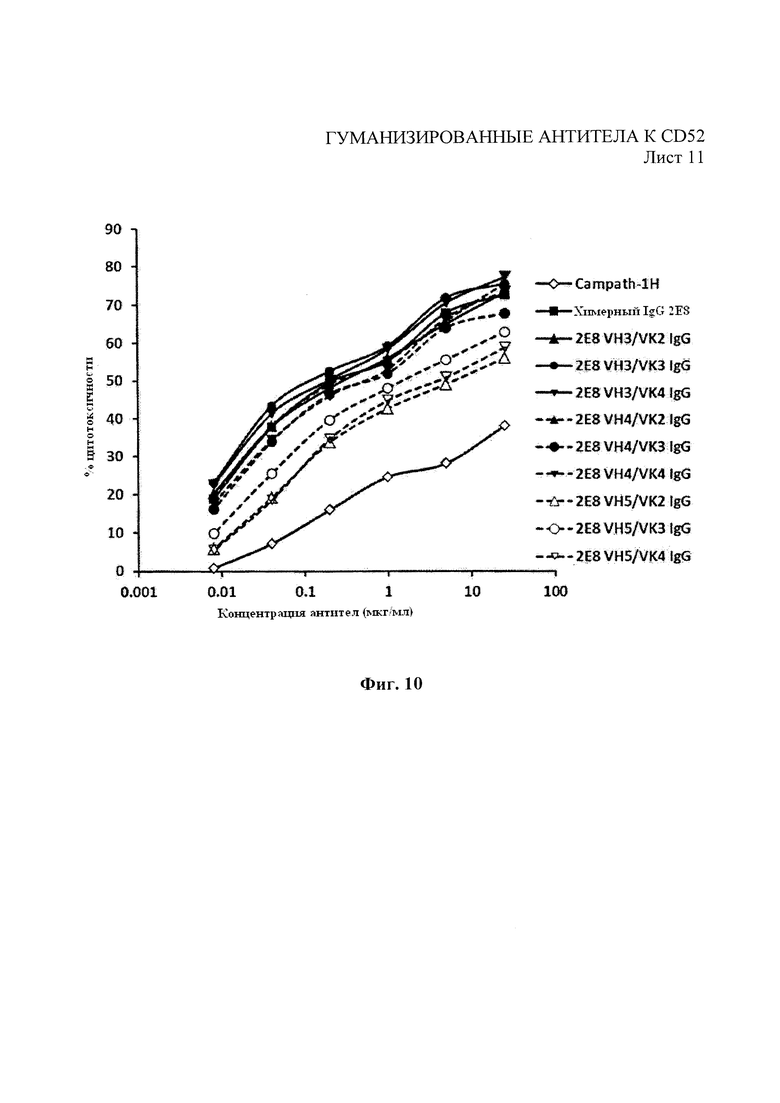
На Фиг. 10 показана средняя цитотоксичность 4 образцов МКПК человека, использованных в качестве эффекторных клеток, в анализе АЗКЦ для гуманизированных вариантов 2Е8 и Campath-1H с клетками-мишенями REH.
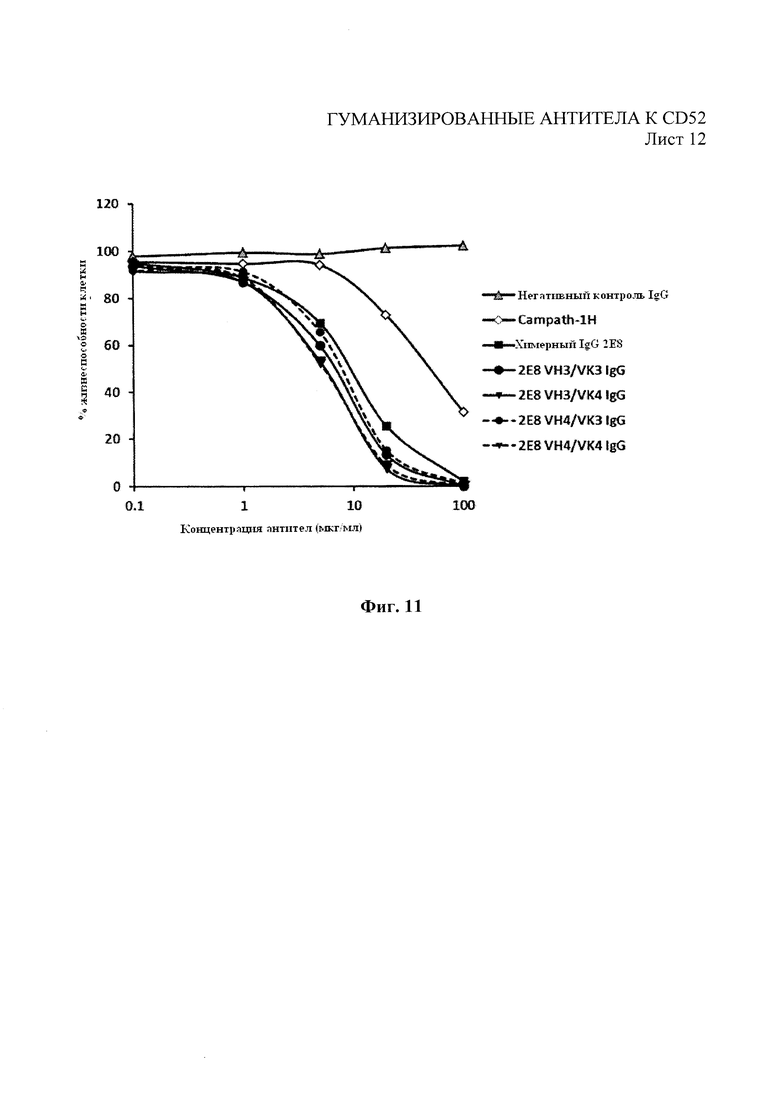
На Фиг. 11 показана КЗЦ для гуманизированных вариантов 2Е8 и Campath-1H с клетками-мишенями REH с высокой экспрессией CD52 и человеческим комплементом.
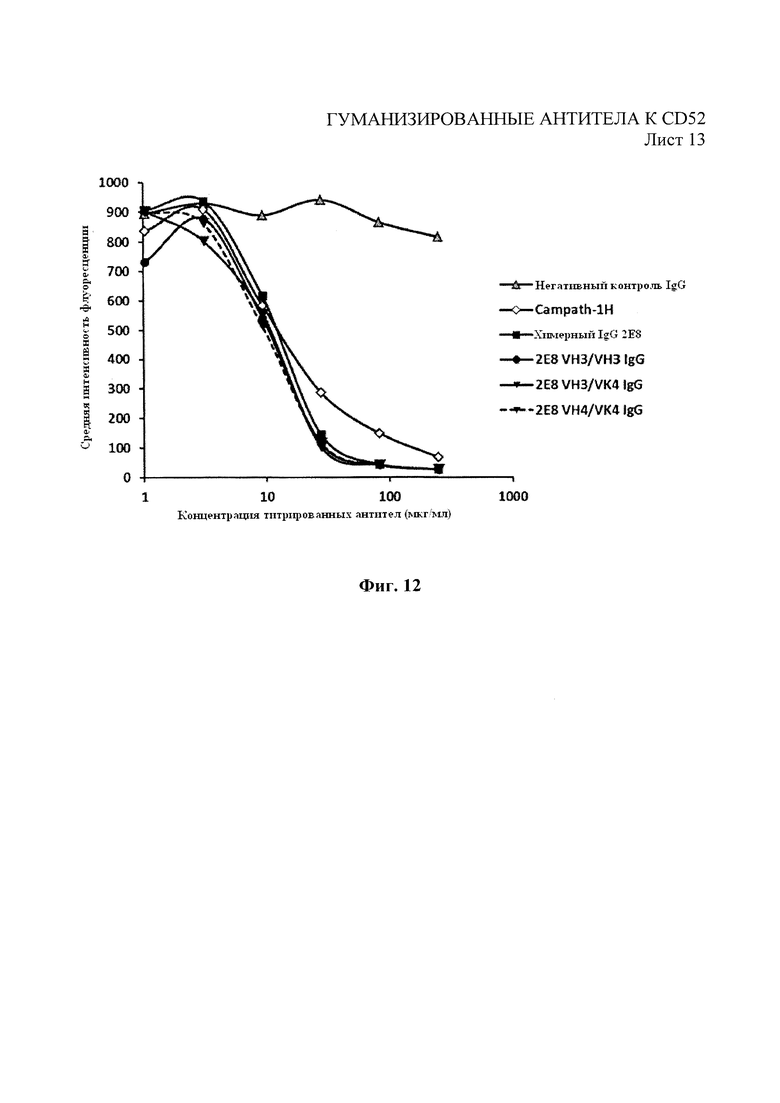
На Фиг. 12 показан анализ конкурентного связывания с помощью проточной цитометрии с использованием Campath-1H-PE в конкуренции с химерным 2Е8, Campath-1H и выбранными вариантами за связывание с клетками REH.
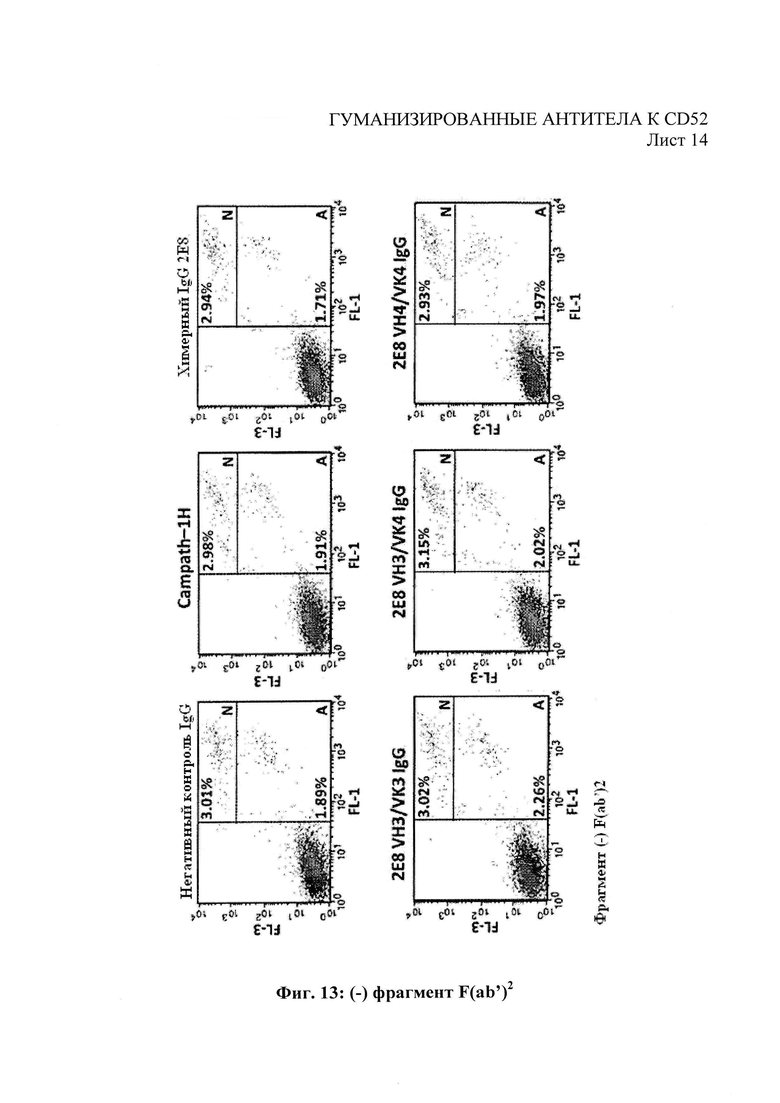
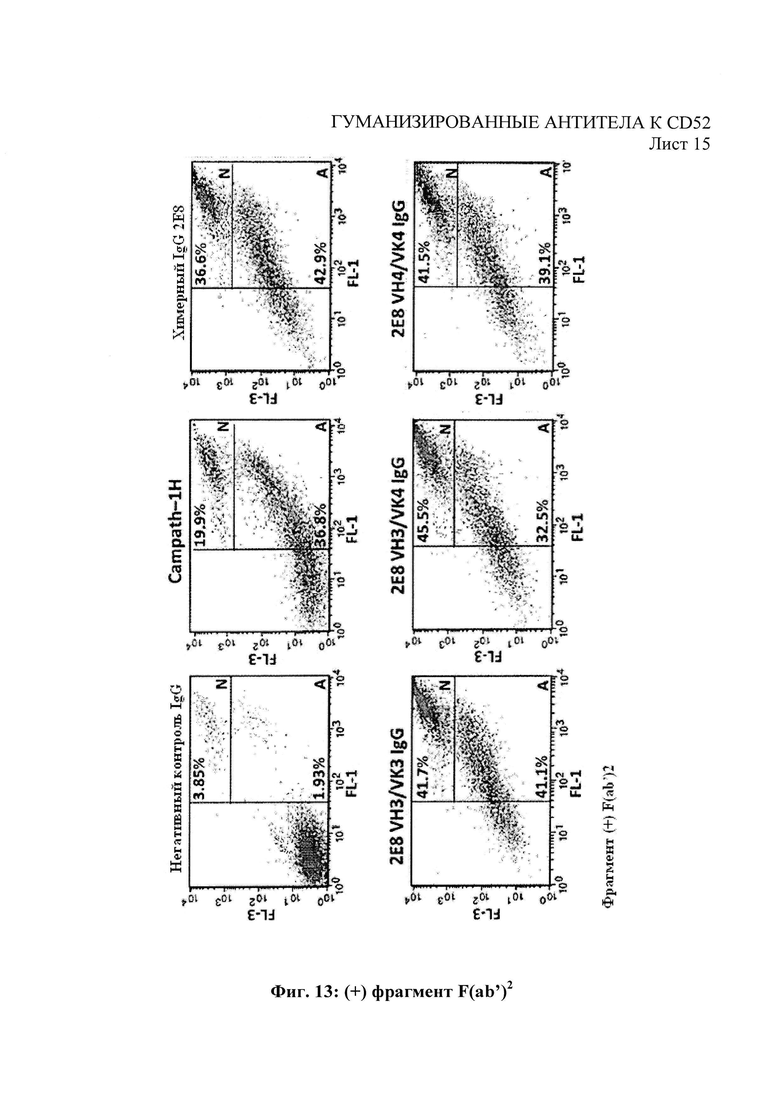
На Фиг. 13 показаны прямые цитотоксические эффекты воздействия антител к человеческому CD52 на клетки REH, измеряемые в плане апоптоза и некроза.
На Фиг. 14 показан участок Каплана-Мейера для мышей с ТКИН, которым были пересажены клетки Раджи человеческой лимфомы Беркитта после обработки Campath-1H и лидерным вариантом VH3/VK4 гуманизированного 2Е8 (SEQ ID №№22 и 28 области V).
Осуществление изобретения
Далее в примерах указаны коммерчески доступные реагенты, которые были использованы в соответствии с инструкциями производителя, если только не указано иное. Источником клеток, идентифицированных в примерах и по всему описанию с помощью учетных номеров ЕСАСС, является Европейское собрание клеточных культур (European Collection of Cell Cultures (ЕСАСС)), Солсбери, Англия. Если не указано иное, все используемые технические и научные термины имеют значение, понимаемое средним специалистом в области техники, к которой принадлежит данное изобретение. Методики и материалы, приводимые в качестве примеров, описаны ниже, однако в процессе реализации или испытания данного изобретения также могут быть использованы методики и материалы, похожие или равнозначные описанным. Материалы, методики и примеры приведены только для пояснения и не могут считаться ограничивающими объем изобретения.
Пример 1 - Генерация мышиных моноклональных антител
Пептид CD52 (GQNDTSQTSSPSC) был индивидуально синтезирован и конъюгирован либо с гемоцианином лимфы улитки, либо с БСА посредством линкера малеимидокапроил-N-гидроксисукцинимид («Мимотопс» (Mimotopes), Виррал, Чешир, Великобритания), при этом N-конец пептида остается свободным. Клетки Раджи и НuТ78 были получены из Европейской коллекции культивируемых клеток животных. Линии клеток NSO, экспрессирующие CD52, были получены следующим образом: человеческий CD52, кодирующий ДНК (эталонная последовательность НЦБИ: NM_001803.2) (полноразмерная последовательность, включающая сигнальный пептид N-конца, сигнальный пептид смещенного ГФИ-якоря С-конца и зрелый поверхностный пептид с ГФИ-якорем), был амплифицирован с помощью ПЦР и субклонирован в экспрессирующие векторы антител pANT (Фиг. 1а) через сайты BglII и Eagl. Транскрипцию гена CD52 контролировали с помощью немедленно-раннего промотора ЦМВ (US5168062 и US5385839, университет штата Айова). Плазмида с экспрессией pANT содержала мутантный миниген dhfr (Simonsen & Levinson 1983, PNAS 80:2495-2499) под контролем промотора SV40 и последовательности polyA для селекции в эукариотических клетках, а также ген бета-лактамазы (ApR) для прокариотической селекции и точку рМВ1 начала репликации для размножения в прокариотических клетках. Плазмида экспрессии была репродуцирована в XL 1-Blue кишечной палочки Е. coli (Stratagene Cat. No. 200130). Стабильные линии клеток с экспрессией CD52 были получены путем трансфекции клеток NS0 путем электропорации и размещения в селектируемые клетки с 200 нМ метотрексата. Клетки выращивали и размножали, затем тестировали с помощью проточной цитометрии на экспрессию CD52. Линии клеток с высокой экспрессией CD52 заморозили и использовали для иммунизации мышей, как описано ниже.
Самок мышей линии Balb/c либо первично иммунизировали путем введения внутрь малоберцовой кости (i.р.) 50 мкг конъюгата пептида CD52-гемоцианина лимфы улитки в полном адъюванте Фрейнда (ПАФ), либо первично иммунизировали 1×106 клетками РАДЖИ, экспрессирующими CD52, в фосфатно-солевом буфере (ФСБ). После четырех недель всех мышей повторно иммунизировали путем введения внутрь малоберцовой кости 106 клеток HUT-78 в ФСБ с дополнительной повторной инъекцией через две недели. Через четыре недели всем мышам была сделана третья инъекция 3×106 клеток NS0, экспрессирующих CD52, в ФСБ введением внутрь малоберцовой кости. Две последующих повторных инъекции 107 клеток NS0, экспрессирующих CD52, в ФСБ были введены внутрь малоберцовой кости с двухнедельными интервалами, и некоторым мышам дополнительно ввели дополнительную дозу 5 мкг пептида CD52 - гемоцианина лимфы улитки (KLH).
За три дня до слияния миеломы двум мышам, показывающим самый высокий титр антител, повторно внутрь малоберцовой кости ввели 107 клеток NS0, экспрессирующих CD52, в ФСБ. В день слияния обеих мышей умертвили, селезенки удалили и клетки из каждой полной селезенки объединили, промыли в бессывороточной культуральной среде и разделили на два равных по размеру образца. Провели слияние половины клеток селезенок с клетками миеломы F0 и ПЭГ- опосредованное слияние другой половины с клетками миеломы P3X63Ag8U.1. В планшетах 1-4 находились слившиеся клетки F0, в планшетах 5-8 находились клетки P3X63Ag8U.1. Полная среда слияния состояла из модифицированной по способу Дульбекко среды Игла, 2% L-глутамина, 1% пенициллина-стрептомицина, 10% фетальной бычьей сыворотки, 5% среды клонирования гибридом BriClone (Национальный институт клеточной биотехнологии, Дублин, Ирландия) и гипоксантина-аминоптерина-тимидина (ГАТ). Полученные слияния высевали в 96-луночные планшеты объемом 200 мкл в каждую лунку. Остальные не помещенные на планшеты слившиеся клетки стабилизировали в культуре в течение срока до трех дней, затем заморозили и хранили в жидком азоте. Помещенные на планшеты слившиеся клетки культивировали при 37°С в 5% СО2 в течение двух недель, перенесли на 96-луночные планшеты и тестировали на наличие выделенных анти-СD52 антител с помощью иммуносорбентного ферментного анализа пептида CD52-гемоцианина лимфы улитки, как описано ниже. Клетки из 24 иммунопозитивных лунок размножили в культуре и тестировали на наличие CD52-специфических антител с помощью иммуносорбентного ферментного анализа пептида CD52, иммуносорбентного ферментного анализа на основе клеток NS0-CD52 и проточной цитометрии.
Для иммуносорбентного ферментного анализа CD52 пептида, планшеты иммуносорбентного ферментного анализа («Ви-Даблью-Ар» (VWR), Латтерворт, Великобритания) оставили на ночь при 4°С покрытыми 100 мкм на лунку либо пептида CD52-гемоцианина лимфы улитки, пептида CD52-БСА, только гемоцианина лимфы улитки или только БСА в 0,5 мкг/мл в ФСБ. Планшеты промыли и фиксировали 150 мкл на лунку ФСБ, содержащего 2% БСА. Супернатанты культуры клеток или очищенные антитела разводили в ФСБ/2% БСА и добавляли 100 мкл к каждому планшету с последующей инкубацией в течение 1 часа при комнатной температуре. Планшеты промыли три раза в ФСБ-полисорбате Твин (0,05%) и инкубировали в течение 1 часа с 100 мкл на лунку козьего антимышиного Ig (Fab-специфического), конъюгированного с пероксидазой хрена («Сигма-Олдрич» (Sigma-Aldrich)). Планшеты трехкратно промыли ФСБ-полисорбатом, после чего добавили субстрат о-фенилендиамина дигидрохлорида SigmaFast («Сигма-Олдрич») и инкубировали при комнатной температуре в темноте для обеспечения проявления цвета. Реакцию остановили путем добавления 50 мкл 3М НСl. Результаты реакций на планшетах считывали при 490 нм с помощью планшет-ридера Dynex («Дайнекс» (Dynex), Уортинг, Великобритания). Гибридомами, специфичными к пептиду CD52, были гибридомы, которые были связаны с пептидом CD52-гемоцианином лимфы улитки и пептидом CD52-БСА, но не те, которые были связаны только с гемоцианином лимфы улитки или только с БСА.
Для иммуносорбентного ферментного анализа на основе клеток NS0-CD52 3×105 клеток на лунку (клетки дикого типа NS0 или NS0, экспрессирующие CD52) высеяли в 96-луночный планшет с V-образным дном. Планшет центрифугировали, супернатанты удалили, а планшет промокнули фильтровальной бумагой. Образцы гибридом разводили в пропорции 1 в 2 в буфере для флуоресцентной сортировки клеток (фосфатно-солевой буфер Дульбекко, содержащий 1% БСА и 0,05% азида натрия) и 100 мкл перенесли в каждый из двух планшетов, содержащих либо NS0 клетки (планшет 1), либо клетки NS0-CD52 (планшет 2). После инкубации при комнатной температуре в течение 1 часа планшеты промывали дважды центрифугированием планшетов и ресуспендированием клеток в 200 мкл буфера для флуоресцентной сортировки клеток между центрифугированиями. После центрифугирования клетки ресуспендировали в 100 мкл буфера для флуоресцентной сортировки клеток, содержащего антимышиный IgG (специфический к Fab) («Сигма»), разведенного в пропорции 1:500. После инкубации в течение 1 часа при комнатной температуре планшеты промывали дважды центрифугированием и ресуспендированием клеток в ФСБ. После центрифугирования клетки ресуспендировали в 50 мкл ФСБ и перенесли на планшет для иммуносорбентного ферментного анализа. Добавили 100 мкл субстрата ТМВ («Инвитроджен» (Invitrogen)) и инкубировали при комнатной температуре в темноте для обеспечения проявления цвета. Реакцию остановили добавлением 50 мкл 3М НСl. Результаты реакций на планшетах считывали при 450 нм с использованием планшет-ридера Dynex. CD52-специфическими клонами были клоны, которые специфически связывались с клетками NS0-CD52 по сравнению с клетками NS0 дикого типа.
Для проточной цитометрии окрасили 3×105 клеток NS0-CD52 или NS0 дикого типа с использованием разведения 1 в 2 антител к гибридомам против CD52 с вместе разведением 1 в 100 конъюгированного антитела антимышиного IgG-PE («Сигма»). Мышный IgG («Сигма») также включили в качестве отдельного контрольного образца для различных мышиных изотипов, присутствующих в гибридомах. Клетки окрашивали в течение 1 часа при 4°С. Также был включен контрольный образец, содержащий только конъюгированное антитело антимышиного IgG-PE. Клетки дважды промывали буфером для флуоресцентной сортировки клеток, в конце ресуспендировали в буфере для флуоресцентной сортировки клеток и выполнили проточную цитометрию с использованием Beckton Dickinson FACSCalibur («Бектон Диккинсон» (Becton Dickinson), Оксфорд, Великобритания). Настройки прибора были определены путем анализа соответствующих антител изотипического контроля.
На основании результатов иммуносорбентного ферментного анализа CD52 пептидов, иммуносорбентного ферментного анализа на основе клеток NS0-CD52 и проточной цитометрии клонировали гибридомы, специфические к huCD52, размножили в культуре, заморозили как родительские запасы и хранили в жидком азоте. Каждую из отобранных гибридом развели в среде клонирования и высеяли в 96-луночные планшеты с плотностью одна клетка на три лунки. Среда клонирования состояла из модифицированной по Дульбекко среды Игла, 2% L-глутамина, 1% пенициллина-стрептомицина, 10% фетальной бычьей сыворотки, 5% среды клонирования гибридом BriClone и гипоксантина-тимидина (ГТ). Культуры выдержали при 37°С в 5% СО2 в течение 2 недель с клонированными клетками, в которые добавили свежую среду через одну неделю содержания в культуре. Через две недели после клонирования супернатанты из всех лунок с засеянным материалом перенесли на новые 96-луночные планшеты и испытали на наличие анти-CD52 антител с помощью иммуносорбентного ферментного анализа пептида CD52 и проточной цитометрии, как описано выше. Материал из показавших положительный результат лунок размножили в культуре и испытали повторно. Положительные клетки дополнительно размножили и испытали на изотип антител. Анти-CD52 положительные субклоны заморозили, сохранили в жидком азоте и использовали для получения моноклональных антител для дальнейших исследований.
Моноклональные антитела изотипировали с помощью Набора для изотипирования с мышиными антителами для быстрого ИФА («Пербио» (Perbio), Крамлингтон, Великобритания). Антитела очистили на колонке с 1 мл белка А-сефарозы («Джи-И Хелфкер» (GE Healthcare), Литтл Чалфонт, Великобритания). До очистки трубки и колонку белка А депирогенизировали обработкой 0,4 M NaOH. Колонку повторно кондиционировали 20 колоночными объемами ФСБ с рН 7,4. Супернатанты культуры клеток гибридом собрали, довели до объема 1 × ФСБ с рН 7,4 с помощью 10 объемов ФСБ и стерилизовали фильтрованием. Отфильтрованный супернатант прокачали через колонку с сефарозой белка А со скоростью 0,5 мл/мин. Колонку промыли 1 × ФСБ с рН 7,4 и элюировали IgG с помощью 0,1 M стерильного цитрата натрия с рН 3 с 0,9 мл фракций, собранных и нейтрализованных 0,1 мл стерильного 1М Tris-HCl с рН 9. В стерильных условиях произвели замену буфера ФСБ с рН 7,4 в продукте с удалением элюирующего буфера для концентрации образца. После концентрирования произвели количественный подсчет антител методом OD280hm с коэффициентом затухания ЕС (0,1%) 1,4. Очищенные антитела анализировали методом SDS-PAGE с использованием системы электрофореза Novex NuPAGE с 4-12% геля NuPage («Инвитроджен» (Invitrogen), Пейсли, Великобритания) и подвижного буфера МЭС кислоты. 1 мкг антитела приготовили с добавлением 4 объемов буфера для образцов NuPAGE с добавлением бета-меркаптоэтанола и нагрели. Гель окрасили окрашивающим раствором InstantBlue («Экспедион» (Expedeon), Кембридж, Великобритания) и оценили молекулярный размер путем сравнения окрашенных полос в лэддере предварительно окрашенных белков PageRuler™ Plus Prestained Protein Ladder («Ферментас» (Fermentas), Йорк, Великобритания). Было определено, что две полосы для каждого антитела не имеют поддающегося обнаружению загрязнения. Очищенные антитела тестировали методом проточной цитометрии CD52 пептида, как описано выше. По данным анализа поточной цитометрии (фигура 2) было показано, что ведущее моноклональное антитело, обозначенное 2Е8, связывается избирательно с клетками NS0-CD52.
Пример 2 - Секвенсирование вариабельных областей генов
Всю РНК экстрагировали из клеток гибридом 2Е8 с использованием набора RNAqueous-4PCR («Амбион» (Ambion), Уоррингтон, Великобритания) и использовали для синтеза кДНК. Фрагменты вариабельной области (V) тяжелой и каппа легкой цепи мышиного иммуноглобулина амплифицировали посредством полимеразной цепной реакции с использованием вырожденных праймеров мышиных лидерных последовательностей («Сигма») и уникальных праймеров константных областей («Сигма»), как показано в таблице 1. Полученные фрагменты ПЦР субклонировали в векторной системе pGEM-Т Easy I («Промега» (Promega), Саутгемптон, Великобритания) и секвенсировали вставки с использованием вектор-специфического праймера M13Forward ((«Сигма»). Все секвенсирование ДНК было выполнено компанией «Джинсервис Лтд.» (Geneservice Ltd), Кембридж, Великобритания). Полученные нуклеотидные последовательности вариабельной области показаны в виде SEG ID №1 и SEG ID №2, и соответствующие аминокислотные последовательности показаны в виде SEG ID №3 и SEG ID №4 для вариабельных областей тяжелой и легкой цепей, соответственно.


Последовательности гипервариабельных областей 2Е8 (CDR) были определены следующим образом:
SEG ID №5 CDRH1 RYGMS
SEG ID №6 CDRH2 MMKTKGGRTYYPDSVKG
SEQ ID №7 CDRH3 DGYY
SEQ ID №8 CDRL1 KSSQSLLHSDGKTYLN
SEQ ID №9 CDRL2 LVSKLDS
SEQ ID №10 CDRL3 WQGTHLWT
Пример 3 - Генерация химерных антител
Последовательности вариабельных областей тяжелых и легких цепей моноклональных антител 2Е8 амплифицировали посредством ПЦР и субклонировали в экспрессирующие векторы антител pANT (Фиг. 1b), при этом вариабельные области тяжелых и легких цепей клонировали в pANT17 и pANT13, соответственно. Гены вариабельной области тяжелой цепи клонировали в pANT17 на сайтах MluI и HindIII внутри рамки считывания с геном тяжелой цепи гамма-1 человека (аллотип Glm3 (Glm(f))) и гены вариабельной области легкой цепи клонировали в pANT13 на сайтах BssHII и BamHI внутри рамки считывания с геном константной области каппа легкой цепи человека (аллотип Кm3). Транскрипцию генов как тяжелых, так и легких цепей выполняли под контролем немедленно-раннего промотора ЦМВ (US 5168062 и US 5385839, университет штата Айова), а плазмиду pANT17, содержащую мутантный миниген dhfr (Simonsen & Levinson 1983, PNAS 80: 2495-2499), - под контролем промотора SV40 и последовательности полиА для селекции в эукариотических клетках. Как pANT17, так и pANT13 содержали ген бета-лактамазы (ApR) для прокариотной селекции и точку начала репликации рМВ1 для размножения в прокариотических клетках. Все плазмиды были размножены в XL1-Blue кишечной палочки Е. coli (Stratagene Cat. No. 200130). Праймеры, использованные для амплификации генов вариабельной области для клонирования в экспрессирующих векторах pANT, приведены в таблице 2.

Затем экспрессирующие конструкции тяжелых и легких цепей трансфицировали вместе либо временно в клетки НЕК293 посредством трансфекции на основе фосфата кальция, либо постоянно трансфицировали в клетки NS0 путем электропорации. Выделенные антитела очистили от супернатантов клеточной культуры посредством хроматографии с белком А. Как показано на фиг. 3, при использовании проточной цитометрии, подробно описанной в примере 1, разведения антитела 2Е8 показали улучшение профиля связывания с клетками СD52+НuТ78 по сравнению с Campath-1H. Как показано на фиг. 4, антитело 2Е8 показало улучшение профиля конкурентного связывания, полученного посредством анализа методом проточной цитометрии, когда оно конкурировало с Campath-1H за связывание с клетками СD52+НuТ78.
Пример 4 - Антителозависимая клеточная цитотоксичность (АЗКЦ)
Анализы АЗКЦ проводили с помощью моноклональных антител 2Е8 следующим образом. Клетки-мишени (клетки REH или Раджи) собрали и предварительно внесли в них 25 мкМ (конечного) Кальцеина-АМ («Сигма»). После инкубации с кальцеином в течение 1 часа при 37°С клетки промыли в средах для удаления неинкорпорировавшегося кальцеина. Добавили 1×104 клеток-мишеней на прозрачный планшет с V-образным дном, содержащий антитела 2Е8 или контрольные антитела в 50 мкг/мл конечного объема, как показано на фиг.5 или на фиг.6, и инкубировали в течение 1 часа для предварительной опсонизации клеток-мишеней. МНПК (эффекторные клетки) выделили из лейкоцитарных пленок здоровых доноров (из крови, взятой не позднее, чем 24 часа назад), полученных от Национальной службы переливания крови Великобритании (больница Адденбрука, Кембридж, Великобритания) и в соответствии с разрешением, предоставленным местным комитетом по этике в научных исследованиях больницы Адденбрука. МНПК выделили из лейкоцитарных пленок посредством центрифугирования в градиенте плотности на Lymphoprep («Аксис-Шилд» (Axis-Shield), Данди, Великобритания). 5×105 эффекторных клеток добавили в каждую лунку планшета, содержащего клетки-мишени и антитела в конечном объеме 250 мкл (соотношение эффекторных клеток к клеткам-мишеням: 50:1). Образцы инкубировали в течение 4 часов при температуре 37°С в 5% СO2. Через 4 часа добавили Triton Х-100 в контрольные лунки, содержащие клетки (эффекторные и (или) клетки-мишени) для установления максимального контроля лизиса. После центрифугирования 150 мкл среды перенесли из каждой лунки на 96-луночный планшет с прозрачным дном и черными стенками, измерили флуоресценцию планшета при 520 нм. Результаты для % цитотоксичности были выражены следующим образом:
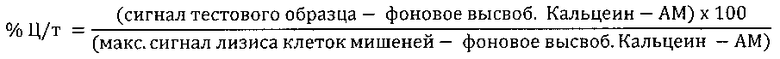
На Фиг. 5 показана средняя АЗКЦ для МКПК от 5 людей-доноров для химерных 2Е8 (′ANT01′) и Campath-1H для клеток-мишеней СD52+REH, при этом АЗКЦ для химерных 2Е8 существенно улучшена. Впоследствии выделили линию вариантных клеток REH с высокой экспрессией СD52 путем флуоресцентной сортировки клеток, которая показала связывание Campath-1H приблизительно в два раза выше по сравнению с родительскими клетками REH. На Фиг. 6 показана серия разведений химерных 2Е8 и Campath-1H для АЗКЦ на клетках с высоким СD52+REH при использовании МКПК двух отдельных доноров. Здесь также показано значительное улучшение профиля АЗКЦ для химерного 2Е8 по сравнению с Campath-1H.
Пример 5 - Комплементзависимая цитотоксичность (КЗЦ)
Анализы КЗЦ проводили на моноклональных антителах 2Е8 следующим образом. Клетки-мишени (клетки REH или Раджи) собрали и добавили к ним 5×104 клеток на лунку в 96-луночный планшет с черной стенками и прозрачным плоским дном. 2Е8 или контрольные антитела для конечных концентраций, как показано на Фиг. 7, добавили вместе с активной или инактивированной нагреванием (при 60°С в течение 30 мин) человеческой сывороткой («Пафуэй Дайагностикс Лтд.» (Pathway Diagnostics Ltd), Доркинг, Великобритания) на лунку (конечная концентрация сыворотки - 25%). Образцы инкубировали в течение 3 часов при температуре 37°С в 5% СO2. Через 3 часа добавили Triton X-100 в контрольные лунки, содержащие клетки, для установления максимального контроля лизиса. Реагент для определения жизнеспособности клеток Prestoblue (10×) («Инвироджен») разбавили питательной средой, использованной в анализе, и добавили в каждую лунку для получения конечного разведения PrestoBlue 1 в 10. После инкубации в течение 1 часа при 37°С в 5% СО2 измерили флуоресценцию планшета при 590 нм. Результаты для % жизнеспособности были выражены следующим образом:
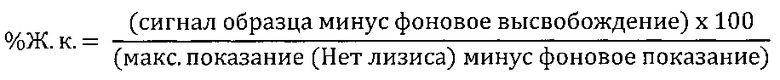
На Фиг. 7 показан значительно улучшенный профиль КЗЦ для химерного 2Е8 по сравнению с Campath-1H.
Пример 6 - Генерация гуманизированных антител
Гуманизированные антитела были получены методами, описанными в ЕР 1844074 («Антитоуп Лтд.» (Antitope Ltd)). Структурные модели вариабельных областей мышей 2Е8 получили посредством использования швейцарской базы данных белковых структур и проанализировали с целью выявления важных аминокислот, которые, с наибольшей вероятностью, были важны для свойств антитела связываться с CD52 («сдерживающие остатки»). База данных последовательностей вариабельных областей человека была использована для выявления сегментов последовательностей вариабельных областей человека, каждый из которых содержал сдерживающие остатки, которые использованы в проектировании гуманизированных антител. Обычно использовали два или более вариантных сегмента последовательности вариабельной области для получения каждого сдерживающего остатка, что приводило к образованию большого диапазона возможных последовательностей вариабельных областей гуманизированных анти-CD52 антител для 2Е8. Затем эти последовательности анализировали для прогнозирования связывания пептида не-зародышевой линии КТС класса II с помощью анализа путем компьютерного моделирования, как описано у Fothergill et al. (WO 9859244, на имя Eclagen Ltd), а также для известных эпитопов к CD4+T клетки с использованием баз данных, в том числе проекта «Ресурс баз данных и анализа эпитопов, распознаваемых иммунной системой», http://www.immuneepitope.org/. Не учитывались последовательности вариабельных областей с прогнозируемыми связывающими пептидами незародышевой линии КТС класса II или с существенными попаданиями из баз данных эпитопов Т-клеток. Вследствие этого был получен уменьшенный набор последовательностей вариабельной области. Отдельные комбинации сегментов последовательностей вариабельной области затем объединили для получения аминокислотных последовательностей вариабельной области тяжелой и легкой цепей гуманизированных антител. Для 2Е8 отобрали последовательности пяти тяжелых цепей и четырех легких цепей (обозначенные VH1-VH5 и Vκ1-Vκ4, соответственно) (SEQ ID №20-24 и 25-28, соответственно).
Вариантные вариабельные области гуманизированных антител, кодирующие ДНК, синтезировали и субклонировали в экспрессирующие векторы pANT17 и pANT13 (Фиг. 1), как описано в примере 3. Все комбинации гуманизированных цепей VH и Vκ (т.е., в общей сложности 20 пар для 2Е8) временно трансфицировали в НЕК293, а также трансфицировали в клетки NS0, и антитела очистили с помощью хроматографии белка А из культуральных супернатантов, как описано в примере 3.
Пример 7 - Анализ гуманизированных антител
Связывание гуманизированных вариантов 2Е8, произведенных на основе НЕК и NS0, с пептидом CD52 оценивали посредством иммуносорбентного ферментного анализа конкурентного связывания по сравнению с родительскими химерными антителами. Родительское химерное антитело 2Е8 биотинилировали с помощью набора Biotin Tag™ Micro Biotinylation («Сигма-Олдрич»). На 96-лучночные планшеты MaxiSorp («Нанк» (Nunc)) нанесли 0,025 мкг/мл пептида CD52-гемоцианина лимфы улитки в ФСБ Дюльбекко («Пи-Эй-Эй Лабораториз» (РАА Laboratories), Йовил, Великобритания) (конечный объем 100 мкл) при 4°С и выдержали в течение ночи. Планшеты фиксировали в ФСБ Дюльбекко - 2% БСА в течение 1 часа при комнатной температуре. Планшеты промыли 3 раза промывочным буфером (0,05% Полисорбата 20 в ФСБ Дюльбекко). Тестовые гуманизированные антитела с различными концентрациями предварительно смешали с биотинилированным родительским химерным антителом (0,035 мкг/мл конечной концентрации) и затем добавили к планшету с пептидом CD52-гемоцианином лимфы улитки (100 мкл конечного объема). Все образцы испытали в двух экземплярах. Планшеты инкубировали в течение 1 часа при комнатной температуре и промыли 3 раза промывочным буфером. Добавили 100 мкл комплекса стрептавидин-пероксидаза хрена с разведением 1 в 1000 («Сигма-Олдрич») и инкубировали в течение 1 часа при комнатной температуре. Планшеты промыли 3 раза промывочным буфером и добавили 100 мкл субстрата ТМВ («Инвитроджен») и инкубировали при комнатной температуре в темноте для обеспечения проявления цвета. Реакцию остановили посредством добавления 50 мкл 3М НСl. Результаты реакций на планшетах были считаны при 450 нм посредством использования планшет-ридера Dynex.
Как показано на Фиг. 8, все лидерные гуманизированные варианты 2Е8 показали профили конкурентного связывания, аналогичные родительским химерным антителам. Гуманизированные варианты впоследствии испытывали на связывание с помощью проточной цитометрии, как описано в примере 1, на АЗКЦ, как описано в примере 4, и КЗЦ, как описано в примере 5. Как показано на Фиг. 9, гуманизированные варианты показали улучшенный профиль связывания, полученный с помощью анализа методом проточной цитометрии, в сравнении с Campath-1Н за связывание с клетками REH. Как показано на Фиг. 10 и 11, гуманизированные варианты также показали улучшение профилей АЗКЦ и КЗЦ при использовании клеток-мишеней REH для АЗКЦ при соотношении клеток-мишеней к эффекторным клеткам 50:1 или высокую линию клеток CD52+REH для КЗЦ (как в примере 4) в сравнении с Campath-1Н.
Пример 8 - Получение scFv и Fab
Гуманизированные варианты 2Е8 из примера 6 преобразовали в scFv и клонировали в векторы отображения фагов M13, как описано в Benhar I. and Reiter Y., Current Protocols in Immunology, Unit 10.19B, Wiley Online Library, May 2002 (http://www.currentprotocols.com/protocol/im1019b) с помощью модуля экспрессии анализа зашиты РНК-азы векторов pCANTAB5E («Амершам Фармасия Байотек» (Amersham Pharmacia Biotech), Литтл Чалфонт, Великобритания). Гены гуманизированных VH и VK амплифицировали при помощи праймеров, вследствие чего образовались концевые сайты рестрикции Sfil и Notl, внутренний линкер Gly4Ser и С-концевая Гистидинб метка. Конструкции ScFv были вставлены в вектор pCANTAB5E в виде фрагментов Sfil-Notl и трансформированы в E. coli НВ2151, в результате чего scFv был экспортирован в периплазму и частично в среду роста. scFv были очищены от среды роста посредством аффинной хроматографии хелата никеля при помощи использования картриджей HIS-Select HF («Сигма-Олдрич»). Очищенные scFv 2Е8 были испытаны посредством описанного в примере 7 анализа конкуренции за связывание с пептидом CD52, и все гуманизированные scFv показали конкурентное связывание с пептидом CD52. Гуманизированные варианты 2Е8 из примера 6 также были преобразованы в Fab посредством метода, использованного для scFv, за исключением того, что амплифицированные гуманизированные гены VH и VK были дополнительно амплифицированы генами константных областей СН1 и Сκ для образования фрагментов VH-CH1 и VK-Сκ, которые были дополнительно амплифицированы праймерами, для присоединения этих фрагментов к лидерной последовательности pelB 22 аминокислот (Lei S.P. et al., J Bacteriol. 169 (1987) стр. 4379-4383) между расположенным выше фрагментом гена VH-СН1 и расположенным далее фрагментом гена VK-Сκ, что привело к образованию бицистронного гена Fab. Fab из гуманизированных вариантов 2Е8 получили и очистили в порядке, описанном выше для scFv, и испытали посредством анализа конкуренции пептида CD52, как описано в примере 7. Все гуманизированные Fab показали конкурентное связывание с пептидом CD52.
Пример 9 - Анализ ответов CD4+ Т-клетки
МНГЖ выделили из лейкоцитарных пленок здоровых общественных доноров (из крови, взятой не позднее, чем за 24 часа), полученных от Национальной службы переливания крови Великобритании (больница Адденбрука, Кембридж, Великобритания) в соответствии с разрешением, предоставленным местным комитетом по этике в научных исследованиях больницы Адденбрука. МНПК выделили из лейкоцитарных пленок посредством центрифугирования в градиенте плотности на Lymphoprep («Аксис-Шилд» (Axis-Shield), Данди, Великобритания), и CD8+ Т-клетки были разрушены с помощью CD8+ RosetteSep™ («СтемСелл Текнолоджиз Инк.» (StemCell Technologies Inc), Лондон, Великобритания). При помощи комплекта («Байотест» (Biotest), Солихалл, Великобритания) для тканевого типирования на основе метода ПЦР АЛЧ с сиквенсспецифическими праймерами было установлено, что доноры имели гаплотипы HLA-DR. Также были определены ответы Т-клеток на контрольные антигены, включая тетанотоксин «воскресшего» антигена (гемоцианин лимфы улитки «Пирс» (Pierce), Крамлингтон, Великобритания, и пептиды, полученные из вирусов гриппа А и болезни Эпштейна-Барра). Затем МКПК заморозили и хранили в жидком азоте до тех пор, пока они не потребовались.
Для приготовления дендритных клеток (ДК), полученных из моноцитов, были отобраны 50 различных донорских МНПК для обеспечения распределения с частотой аллотипов HLA-DR и HLA- DQ, аналогичных частоте их обнаружения у всего населения мира. МНПК оживили в культуральной среде AIM-V®, и выделяли клетки CD14+ с использованием колонок Miltenyi CD14 Microbead и LS («Милтений Байотек» (Miltenyi Biotech), Оксфорд, Великобритания). Моноциты ресуспендировали в AIM-V®, добавили 1000 ед/мл Интерлейкина-4 и 1000 ед/мл ГМ-КСФ («культуральная среда ДК») к 4-6×106 МНПК/мл, и затем разливали в 24-луночные планшеты (2 мл конечного объема культуры). На второй день произвели замену среды клеток на половину объема культуральной среды ДК. К третьему дню моноциты дифференцировались в полузрелые ДК, которые были предварительно инкубированы с использованием либо 40 мкг/мл Campath-1Н, химерных антител 2Е8, гуманизированных антител 2Е8, 100 мкг/мл гемоцианина лимфы улитки, либо только среды. Полузрелые ДК инкубировали с антигеном в течение 24 часов, после чего избыток тестового антитела удаляли путем промывки клеток два раза и ресуспендирования в культуральной среде ДК с добавлением 50 нг/мл ФНО-α («Пепротек» (Peprotech), Лондон, Великобритания). На седьмой день в ДК произвели замену половины объема культуральной среды ДК (с добавлением 50 нг/мл ФНОα) перед сбором зрелых ДК на восьмой день. Произвели подсчет собранных зрелых ДК и оценку жизнеспособности путем исключения при помощи трипанового синего. Затем ДК подвергли облучению гамма-лучами (4000 рад) и ресуспендировали в 2×105 клеток на мл в среде AIM-V перед использованием в анализе ELISpot и анализе пролиферации. Кроме того, на 8-й день также приготовили свежие CD4+ Т-клетки. Для очистки CD4+ Т-клетки МНПК оживили в культуральной среде AIM-V®, и клетки CD14+ выделяли с использованием колонок Miltenyi CD 14 Microbead и LS («Милтений Байотек», Оксфорд, Великобритания) и ресуспендировали в среде AIM-V® с уровнем 2×106 клеток/мл.
На 8-й день провели анализы пролиферации Т-клеток, в которых 1×105 аутологичных клеток CD4+Τ добавили к 1×104 ДК, в которые были внесены гуманизированные антитела 2Е8 или химерные антитела 2Е8 (в соотношении 10:1) на 96-луночных планшетах с U-образным дном, с добавлением среды AIM-V® до конечного объема 200 мкл/лунку). На 14-й день на планшеты, на которых выполняли анализы, произвели воздействие импульсами luCi [3Η] («Перкин Элмер» (Perkin Elmer), Биконсфилд, Великобритания) на лунку в 25 мкл AIMV в течение 6 часов перед сбором на фильтровальных подложках («Перкин Элмер») с помощью коллектора клеток Tomtec Mach III («Хамден Си-Ти» (Hamden СТ), США). Количество импульсов в минуту для каждой лунки определяли при помощи счетчика сцинтилляций Meltilex™ («Перкин Элмер») на основании счетчика-сцинтиллятора 1450 Microbeta Wallac Trilux Liquid («Перкин Элмер») с методом подсчета с низким фоном Paralux. Количество импульсов в минуту для каждого образца антитела были нормализованы до контрольного образца, содержащего только среду.
Для анализов ELISpot планшеты ELISpot («Миллипор» (Millipore), Уотфорд, Великобритания) покрывали 100 мкл/лунку иммобилизованных антител интерлейкина-2 («Ар энд Ди системз» (R&D Systems), Абингдон, Великобритания) в ФСБ. Затем планшеты промыли дважды в ФСБ, инкубировали в течение ночи в фиксирующем буфере (1% БСА («Сигма») в ФСБ) и промыли в среде AIM V®. На 8-й день добавили 1×105 аутологичных CD4+ Т-клетки в 1×104 ДК с добавленным антигеном (в соотношении 10:1) в 96-луночных планшетах для ELISPOT. Все препараты испытали в шести культурах. Для каждого образца донорских МКПК также были выполнены отрицательный контроль (только среда AIM V®), отсутствие контроля клеток и положительный контроль фитогемагглютинином (10 мкг/мл).
После дополнительного 7-дневного инкубационного периода планшеты для ELISpot обработали, используя три последовательных промывки в dH2O и ФСБ перед добавлением 100 мкл отфильтрованного биотинилированного детекторного антитела («Ар энд Ди Системз», Абингдон, Великобритания) в ФСБ/1% БСА. После инкубации при 37°С в течение 1,5 часов планшеты дополнительно промыли три раза в ФСБ; и добавили 100 мкл отфильтрованного комплекса стрептавидин - щелочная фосфатаза («Ар энд Ди Системз») в ФСБ/1% БСА на 1 час (инкубация при комнатной температуре). Комплекс стрептавидин-щелочная фосфатаза слили, а планшеты промыли четыре раза в ФСБ. Добавили 5-бромо-4-хлоро-3-индолил-фосфат/нитросиний тетразолий («Ар энд Ди Системз») в каждую лунку и инкубировали в течение 30 минут при комнатной температуре. Образование пятен остановили посредством трехкратной промывки лунок и задних сторон лунок посредством dH2O. Высушенные планшеты отсканировали на анализаторе Immunoscan™ и определили количество пятен на лунку при помощи Версии 4 ПО Immunoscan™.
Для анализа пролиферации и анализа ELISpot Интерлейкина-2 результаты были выражены в форме индекса стимуляции (ИС), определенного как соотношение импульсов в минуту (анализ пролиферации) или количества пятен (анализ ELISpot) для тестируемого антитела к контрольному варианту, содержащему только среду, при этом пороговое значение ИС было равно или больше 2 (ИС≥2,0) для положительных ответов Т-клеток. Результаты показали, что и Campath-1Н, и химерное антитело 2Е8 индуцировали ответы Т-клеток в 10 или более из 50 испытанных донорских МКПК (>=20%), в то время как ни одно из гуманизированных антител 2Е8 не индуцировало ответа Т-клеток у более чем 2 из 50 доноров (<=4%), что демонстрирует эффективность процесса гуманизации в удалении ответов Т-клеток из вариабельных областей.
Пример 10 - Анализ прямой цитотоксичности
Прямые цитотоксические эффекты антител против человеческого CD52 были оценены с помощью совместного окрашивания аннексином V/иодидом пропидия в качестве маркеров апоптоза и некроза, соответственно. 1×105 клеток REH высеяли в присутствии 100 мкг/мл тестовых антител против человеческого CD52 или контрольного антитела соответствующего изотипа +/- 100 мкг/мл сшивающего антитела F(ab′) (Jackson ImmunoResearch, Cat no. 109-006-008) (600 мкл конечного объема). Клетки инкубировали в течение 72 часа до промывания в ФСБ/ 2% БСА с последующим совместным окрашиванием аннексином V / иодидом пропидия в соответствии с протоколом, рекомендованным производителями (Invitrogen, Cat no. VI3245). Построили диаграммы рассеяния посредством анализа флуоресцентной сортировки клеток и разделили на три области для количественного анализа живых клеток (неокрашенных), апоптозных клеток (FL1, положительная реакция на аннексии V) и некротических клеток (FL1, положительная реакция на аннексии V и FL3, положительная реакция на иодид пропидия). Как показано на Фиг. 13, гуманизированные антитела 2Е8 показали увеличение апоптоза и некроза по сравнению с Campath-1H клеток-мишеней REH, при этом процент некротических клеток > 40% с гуманизированными антителам по сравнению с 19,9% для Campath-1H. На Фиг. 13 «N» обозначает некроз: клетки, значительно окрашенные как аннексином V, так и иодидом пропидия (каналы FL1 и FL3 соответственно). «А» обозначает апоптоз: клетки, положительно окрашенные аннексином V с меньшей окраской иодидом пропидия.
Пример 11 - Экспериментальная модель опухоли на животном
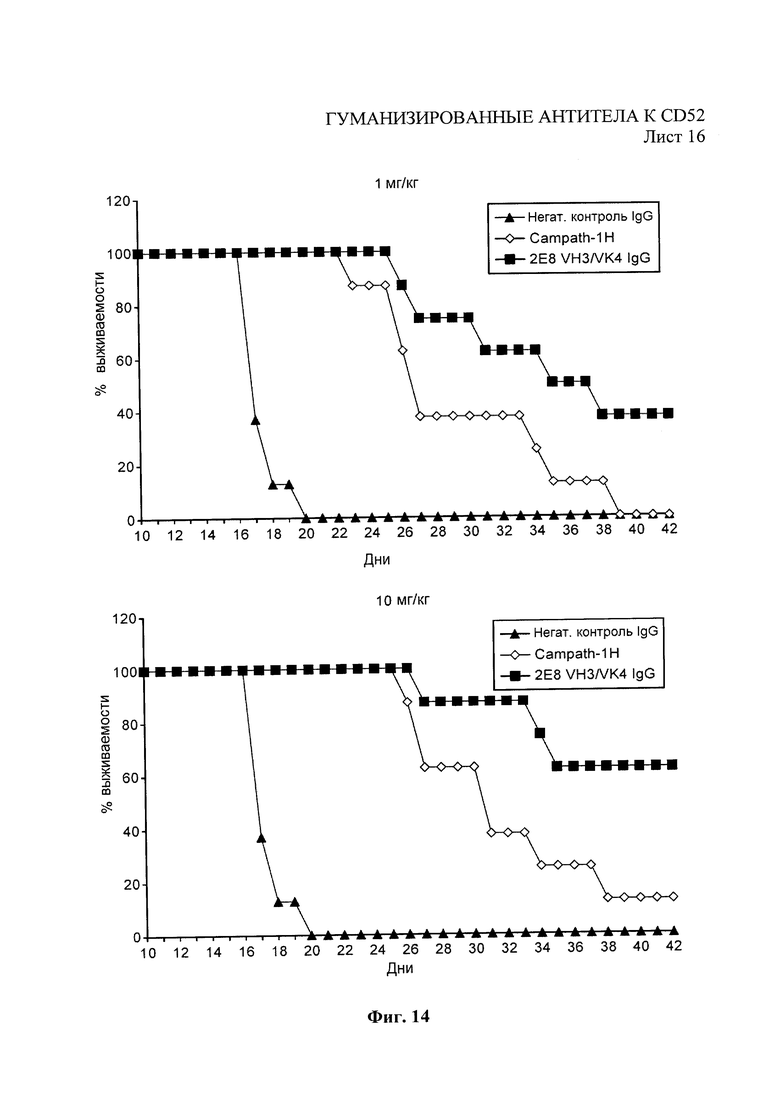
Экспериментальную модель опухоли на животном использовали для анализа in vivo антител против человеческого CD52 в ингибировании роста опухоли. В модели клетки Раджи человеческой лимфомы Беркитта пересадили мышам с ТКИН и животным, обработанным антителами против человеческого CD52. 7-недельных самкам мышей с ТКИН Fox Chase SCID («Чарльз Ривер» Charles River), Моррисвилль, Северная Каролина, США) ввели 1×106 клеток Раджи (Американская коллекция типовых культур, 0,2 мл клеточной суспензии) с помощью болюсной инъекции в хвостовую вену (в/в). Тестовые антитела против человеческого CD52 или контрольное антитело соответствующего изотипа вводили внутрибрюшинно (в/б) один раз в день через день, пока не было введено семь доз, введение было начато через три дня после инъекции опухолевых клеток. Объем дозирования 10 мл/кг (0,20 мл/20 г мышь) отмеряли в соответствии с массой тела каждого животного, определяемой дважды в неделю. Результаты, показанные на Фиг. 14, продемонстрировали улучшение уровня выживаемости при дозах 1 и 10 мг/кг ведущим анти-СD52 антителом VH3/VK4 (SEQ ID вариабельных областей: 22 и 28) по сравнению с Campath 1Н.







| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОГЛОБУЛИНЫ K CD52 ЧЕЛОВЕКА | 2010 |
|
RU2603743C2 |
| АНТИТЕЛО ПРОТИВ ЛИГАНДА 1 ЗАПРОГРАММИРОВАННОЙ ГИБЕЛИ КЛЕТОК (PD-L1), ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2727914C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CTLA4 | 2012 |
|
RU2629768C2 |
| АНТИТЕЛО IGG С АФФИННОСТЬЮ СВЯЗЫВАНИЯ В ОТНОШЕНИИ АНТИГЕННОГО КОМПЛЕКСА CD3, РЕКОМБИНАНТНЫЕ НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ ЛЕГКУЮ И ТЯЖЕЛУЮ ЦЕПИ АНТИТЕЛА, СПОСОБ ПОЛУЧЕНИЯ СИСТЕМЫ, СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛА, СПОСОБ ЛЕЧЕНИЯ ПАЦИЕНТА | 1999 |
|
RU2244720C2 |
| КОМПОЗИЦИИ АНТИТЕЛА К ИНТЕГРИНУ β1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2681994C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ ДЛЯ ПЕПТИДА-6, ПОЛУЧЕННОГО ИЗ HSP65, СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2010 |
|
RU2596925C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ПРОТИВ TAU PS422 ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ ГОЛОВНОГО МОЗГА | 2010 |
|
RU2536247C2 |
| АНТИТЕЛО ПРОТИВ c-Met | 2009 |
|
RU2560257C2 |
| АНТИТЕЛО ПРОТИВ КЛАУДИНА 18А2 И ЕГО ПРИМЕНЕНИЕ | 2017 |
|
RU2793445C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО К PD-L1 | 2017 |
|
RU2665790C1 |












Изобретение относится к области биохимии. Предложено антитело к человеческому СD52. Антитело применяется для лечения или диагностики заболеваний, связанных с клетками СD52+, таких как ХЛЛ и другие лейкозы, аутоиммунные заболевания, отторжение трансплантата органа, реакция «трансплантат-против-хозяина». Изобретение позволяет эффективно связывать CD52. 7 н. и 13 з.п. ф-лы, 14 ил., 2 табл., 11 пр.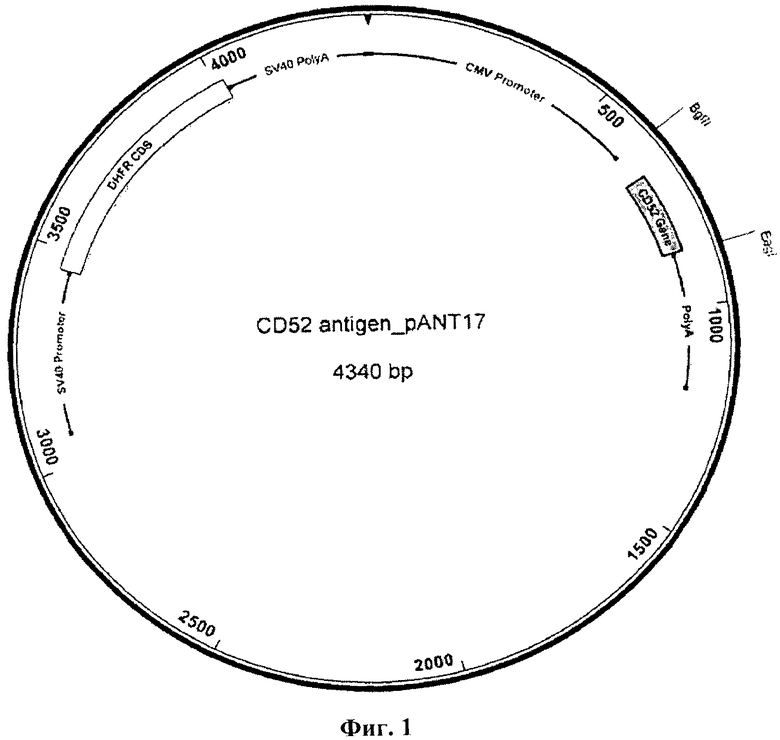
1. Антитело к CD52, содержащее последовательности гипервариабельных участков:
(i) CDRH1, содержащий последовательность RYGMS (SEQ ID №: 5);
(ii) CDRH2, содержащий последовательность MMKTKGGRTYYPDSVKG (SEQ ID №: 6);
(iii) CDRH3, содержащий последовательность DGYY (SEQ ID №: 7);
(iv) CDRL1, содержащий последовательность KSSQSLLHSDGKTYLN (SEQ ID №: 8);
(v) CDRL2, содержащий последовательность LVSKLDS (SEQ ID №: 9);
(vi) CDRL3, содержащий последовательность WQGTHLWT (SEQ ID №: 10).
2. Антитело по п.1, которое содержит одну или несколько последовательностей вариабельных участков, выбранных из группы, состоящей из SEQ ID №№: 20-24 для вариабельной области тяжелой цепи в комбинации с одной или несколькими последовательностями, выбранными из группы, состоящей из SEQ ID №№: 25-28 для вариабельной области легкой цепи.
3. Антитело по п.1, содержащее SEQ ID №: 22 для вариабельной области тяжелой цепи в сочетании с SEQ ID №: 28 для вариабельной области легкой цепи.
4. Антитело по п.1, которое при проверке in vitro на индукцию ответов Т-клеток хелперов CD4+ по меньшей мере в 50 образцах человеческой крови с распределением аллотипов HLA-DR у человеческой популяции характеризуется увеличением количества ответов Т-клеток на <=4%.
5. Антитело по п.1, в котором последовательности вариабельных областей полностью получены из последовательностей вариабельных областей человеческих антител.
6. Антитело по п.5, которое состоит из вариабельных областей вместе с человеческими константными областями.
7. Антитело по п.6, в котором человеческая константная область тяжелой цепи представляет собой либо изотип IgG1, IgG2, IgG3 или IgG4, либо мутированную константную область IgG, а человеческая константная область легкой цепи человека представляет собой изотип каппа.
8. Антитело по п.6, в котором человеческие константные области представляют собой IgG1 и изотип каппа.
9. Антитело по п.6, в котором человеческие константные области представляют собой IgG4 и изотип каппа.
10. Антитело по п.1, в котором антитело представляет собой scFv или Fab.
11. Антитело по п.1, которое является компонентом мультиспецифического белка, который специфически связывается с человеческим CD52 и, кроме того, связывается или взаимодействует с одной или несколькими другими молекулами.
12. Полинуклеотид, кодирующий антитело по любому из пп.1-10.
13. Вектор для экспрессии антитела к CD52, содержащий полинуклеотид по п.12.
14. Клетка-хозяин для экспрессии антитела к CD52, содержащая вектор по п.13.
15. Клетка-хозяин по п.14, которая представляет собой прокариотическую клетку, эукариотическую клетку или клетку млекопитающего.
16. Композиция для лечения или диагностики заболеваний, связанных с CD52, которая содержит антитело к CD52 по любому из пп.1-11.
17. Способ лечения заболевания, связанного с клетками CD52+, в том числе ХЛЛ и других лейкозов; аутоиммунных заболеваний, включая рассеянный склероз, ревматоидный артрит, васкулит, миозит и диабет; и отторжения трансплантата органа и реакции «трансплантат-против-хозяина», в котором пациенту, нуждающемуся в таком лечении, вводят эффективную дозу антитела по любому из пп.1-11 или композиции по п.16.
18. Способ по п.17, в котором параллельно вводят эффективное количество химиотерапевтического вещества.
19. Способ по п.17, в котором параллельно вводят фармацевтический носитель.
20. Способ обнаружения присутствия антигена человеческого CD52 в образце, который содержит следующие этапы:
(a) контактирование тестируемого образца с антителом по любому из пп.1-11 при условиях, обеспечивающих возможность образования комплекса между антителом и антигеном CD52 человека; и
(b) обнаружение присутствия полученного комплекса.
| WO 2010132659 A2,18.11.2010 | |||
| WO 2005042581 A2, 12.05.2005 | |||
| Способ определения сорбционной способности гемосорбентов | 1987 |
|
SU1508155A1 |
| ENBLAD G | |||
| et al., A pilot study of alemtuzumab (anti-CD52 monoclonal antibody) therapy for patients with relapsed or chemotherapy-refractory peripheral T-cell lymphomas, BLOOD, 2004, vol.103, no.8, pp.2920-2924 | |||
| RU 2007147414 A, 10.07.2009. | |||

