Область техники
Настоящее изобретение имеет отношение к антителам и антигенсвязывающим фрагментам, связывающимся с высокой аффинностью с человеческим и мышиным рецептором фактора роста гепатоцитов (HGF), также известным как MET. Антитела и антигенсвязывающие фрагменты являются агонистами MET как у людей, так и у мышей, приводящими к молекулярным и клеточным эффектам, сходным с эффектами HGF-связывания. Изобретение также имеет отношение к терапевтическому применению антител и антигенсвязывающих фрагментов, являющихся агонистами MET.
Уровень техники
HGF представляет собой плейотропный цитокин мезенхимального происхождения, опосредующий характерный набор биологических функций, включая клеточную пролиферацию, подвижность, дифференцировку и выживаемость. HGF рецептор, также известный как MET, экспрессируется рядом тканей, включая весь эпителий, мышечные клетки, нервные клетки, остеобласты, гематопоэтические клетки и различные компоненты иммунной системы.
Cигнальный путь HGF и MET играет важную роль во время развития эмбриона, поскольку он направляет миграцию клеток-предшественников и обусловливает выживание или гибель клетки. У взрослых сигнальный путь HGF/MET обычно находится в состоянии покоя и возобновляется во время заживления ран и регенерации ткани. Некоторые виды рака и опухолей «захватывают» сигнальный путь HGF/MET, для того, чтобы способствовать выживаемости и пролиферации опухоли в организме-хозяине. Поэтому ингибирование HGF-MET оси становится популярной мишенью для противораковой терапии, хотя с ограниченным успехом.
Благодаря его роли в заживлении ткани и регенерации, рекомбинантный HGF также исследуется в качестве варианта лечения целого ряда состояний, включая дегенеративные болезни, воспалительные болезни, аутоиммунные заболевания, метаболические болезни и нарушения, связанные с трансплантацией. Однако, рекомбинантный HGF обладает плохими фармакологическими свойствами: ему необходима протеолитическая активация для того, чтобы стать биологически активным; после того, как он активируется, он имеет крайне короткое время полужизни in vivo; и его промышленное производство является сложным и дорогим.
Агонистические анти-MET-антитела, которые активируют MET, в некоторой степени подражая HGF, предлагаются в качестве альтернативы.
Описаны следующие антитела, имитирующие, по меньшей мере частично, HGF-активность: (i) 3D6 мышиное анти-человек MET антитело (патент США № 6,099,841); (ii) 5D5 мышиное анти-человек MET антитело (патент США № 5,686,292); (iii) NO-23 мышиное анти-человек MET антитело (патент США № 7,556,804 B2); (iv) B7 человеческое наивное анти-человек MET антитело (патентная заявка США № 2014/0193431 A1); (v) DO-24 мышиное анти-человек MET антитело (Prat et al., Mol Cell Biol. 11, 5954-5962, 1991; Prat et al., J Cell Sci. 111, 237-247, 1998); и (vi) DN-30 мышиное анти-человек MET антитело (Prat et al., Mol Cell Biol. 11, 5954-5962, 1991; Prat et al., J Cell Sci. 111, 237-247, 1998).
Краткое раскрытие изобретения
Агонистические анти-MET антитела, полученные к настоящему времени, например, описанные в разделе Уровень техники, часто получают в виде побочных продуктов процессов, направленных на идентификацию антагонистических молекул и явным образом не предназначенных для получения агонистических молекул в расчете на терапевтическое использование. Более того, самым очевидным ограничением анти-MET антител предшествующего уровня является то, что они вырабатываются в мышиной системе (за исключением B7, которое было идентифицировано с использованием человеческой наивной фаговой библиотеки); по этой причине маловероятно, что эти антитела будут проявлять перекрестную реактивность с мышиным MET. Даже в том случае, если минимальная перекрестная реактивность с собственными антигенами в принципе возможна, эти взаимодействия в большинстве случаев обладают очень низкой аффинностью.
При том, что отсутствие перекрестной реактивности не имеет отношения к мышиным моделям рака (поскольку они используют человеческие ксенотрансплантаты), перекрестная реактивность антител между человеческим и мышиным MET является важным требованием к доклиническим мышиным моделям регенеративной медицины или неонкологических болезней человека, которые нуждаются в функционировании антитела на мышиных тканях и клетках.
Для того, чтобы антитело было оценено в доклинических моделях, необходимо не только чтобы агонистическое анти-MET антитело перекрестно реагировало с мышиным MET, но также желательно, чтобы антитело связывалось с мышиным MET с той же самой или сходной аффинностью с его аффинностью в отношении человеческого MET, и также чтобы антитело вызывало эффекты в мышиных системах, такие же или сходные с эффектами, которые оно вызывает в человеческих системах - в ином случае эксперименты, проводимые на доклинических моделях, не будут прогностическими в ситуации с человеком. Как продемонстрировано в Примерах, ни одно из анти-MET агонистических антител предшествующего уровня техники не проявляет аффинность в отношении мышиного MET, и определенно ни одно из антител предшествующего уровня не демонстрирует такое же или сходное связывание и агонистические эффекты в обеих системах, и мышиной и человеческой.
Настоящая заявка предоставляет анти-MET агонистические антитела, созданные с целью связывания как с человечесим так и с мышиным MET с высокой аффинностью. Эти антитела: (i) демонстрируют агонистическую активность в обеих и в человеческой и в мышиной MET биологических системах - другими словами они индуцируют MET передачу сигнала - с эффективностью приблизительно сходной или превышающей таковую HGF; (ii) вызывают полный спектр HGF-индуцированной биологической активности, таким образом, представляя собой обоснованную замену рекомбинантного HGF; (iii) демонстрируют превосходное связывание с мышиным MET при прямом сравнении с антителами предшествующего уровня техники; (iv) проявляют биологически значимую агонистическую активность при концентрациях до 1 pM; (v) демонстрируют время полужизни в сыворотке у мышей несколько дней, достигая фармакологически насыщающих концентраций уже при дозе 1 мкг/кг, которая является очень низкой для терапевтического антитела; (vi) поддерживают почечную функцию и почечную целостность на мышиной модели острого повреждения почек; (vii) предотвращают повреждение печени и противодействуют повреждению гепатоцитов на мышиной модели острого поражения печени; (viii) демонстрируют анти-фиброзирующую, противовоспалительную и про-регенеративную активность на мышиной модели хронического поражения печени; (ix) предотвращают потерю веса, ослабляют кишечное кровотечение, сохраняют целостность кишечника, подавляют воспаление и способствуют регенерации эпителия на мышиной модели язвенного колита и мышиной модели воспалительного заболевания кишечника; (x) способствуют инсулин-независимому поглощению глюкозы на мышиной модели диабета I типа; (xi) преодолевают резистентность к инсулину на мышиной модели диабета II типа; (xii) улучшают состояние при жировой дистрофии печени, подавляют фиброз и восстанавливают функцию печени на мышиной модели неалкогольного стеатогепатита (NASH); (xiii) ускоряют заживление ран на мышиной модели диабетической язвы; (xiv) вступают в перекрестную реакцию с Rattus norvegicus MET и Macaca fascicularis MET, таким образом, обеспечивая возможность проведения токсикологических и фармакологических исследований на этих двух позвоночных животных, необходимых перед проведением исследований, впервые осуществляемых с участием людей; (xv) распознают эпитопы, консервативные среди человека, мыши, крысы и яванского макака, в силу этого обеспечивая более широкую применимость животных моделей.
Следовательно, в первом аспекте настоящее изобретение предоставляет антитело или его антигенсвязывающий фрагмент, который связывается с человеческим MET белком (hMET) с высокой аффинностью и связывается с мышиным MET белком (mMET) с высокой аффинностью, при этом антитело или его антигенсвязывающий фрагмент является hMET агонистом и mMET агонистом. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент содержит, по меньшей мере, один вариабельный домен тяжелой цепи (VH) и, по меньшей мере, один вариабельный домен легкой цепи (VL), при этом указанный VH и VL домен, в тех случаях, когда тестируется как Fab фрагмент, демонстрирует скорость диссоциации (koff измеряли с помощью Biacore) в отношении hMET в пределах от 1 x 10-3 s-1 до 1 x 10-2 s-1, необязательно от 1 x 10-3 s-1 до 6 x 10-3 s-1, и демонстрирует скорость диссоциации (koff измеряли с помощью Biacore) в отношении mMET в пределах от 1 x 10-3 s-1 до 1 x 10-2 s-1, необязательно от 1 x 10-3 s-1 до 6 x 10-3 s-1. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент обладает равной по величине аффинностью по отношению к hMET и mMET.
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент индуцирует фосфорилирование hMET и индуцирует фосфорилирование mMET. В некоторых вариантах осуществления антитело или антигенсвязывающий фрагмент индуцирует фосфорилирование hMET с EC50 (если оно измерено с помощью фосфо-MET ELISA) меньше чем 3,0 нM, необязательно меньше чем 2,0 нM и индуцирует фосфорилирование mMET с EC50 (если оно измерено с помощью фосфо-MET ELISA) меньше чем 3,0 нM, необязательно меньше чем 2,0 нM. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент индуцирует фосфорилирование hMET и mMET равноценно.
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент демонстрирует высокую эффективность фосфорилирования в отношении hMET и демонстрирует высокую эффективность фосфорилирования в отношении mMET. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент индуцирует фосфорилирование hMET с EC50 меньше чем 1нM и/или Emax (в виде процента от HGF-индуцированной активации в фосфо-MET ELISA), по меньшей мере, 80% и индуцирует фосфорилирование mMET с EC50 меньше чем 1нM и/или Emax (в виде процента от HGF-индуцированной активации в фосфо-MET ELISA) по меньшей мере, 80%. В некоторых альтернативных вариантах осуществления антитело или его антигенсвязывающий фрагмент демонстрирует низкую эффективность фосфорилирования в отношении hMET и демонстрирует низкую эффективность фосфорилирования в отношении mMET. В некоторых подобных вариантах осуществления антитело или его антигенсвязывающий фрагмент индуцирует фосфорилирование hMET при EC50 1нM-5нM и/или Emax (в виде процента от HGF-индуцированной активации в фосфо-MET ELISA) 60-80% и индуцирует фосфорилирование mMET при EC50 1нM-5нM и/или an Emax (в виде процента от HGF-индуцированной активации в фосфо-MET ELISA) 60-80%.
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент индуцирует HGF-подобный клеточный ответ при контактировании с человеческой клеткой и индуцирует HGF-подобный клеточный ответ при контактировании с мышиной клеткой. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент полностью вызывает HGF-подобный клеточный ответ при контактировании с человеческой клеткой и при контактировании с мышиной клеткой. В некоторых вариантах осуществления полная индукция HGF-подобного клеточного ответа поддается измерению при помощи одного, любых двух или всех способов из числа:
(i) при помощи анализа клеточного рассеивания, антитело или его антигенсвязывающий фрагмент вызывает клеточное рассеивание сравнимое с максимальным HGF-индуцированным рассеиванием, когда антитело или его антигенсвязывающий фрагмент находится в концентрации 0,1-1,0 нM;
(ii) при помощи анти-апоптотического клеточного анализа, антитело или его антигенсвязывающий фрагмент демонстрирует EC50 меньше, чем 1,1x значение для HGF, и/или при Emax (измеренном в виде % от общего ATP содержания в неапоптотических контрольных клетках) больше чем на 90% того, которое наблюдалось в отношении HGF; и/или
(iii) при помощи анализа морфогенеза ветвления, клетки, обработанные антителом, демонстрируют ответвлений на сфероид больше на 90%, чем число ответвлений, индуцированных такой же (ненулевой) концентрацией HGF.
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент частично индуцирует HGF-подобный клеточный ответ при контактировании с человеческой клеткой и при контактировании с мышиной клеткой. В некоторых вариантах осуществления частичная индукция HGF-подобного клеточного ответа измеряется как:
(i) анализ клеточного рассеивания, антитело или его антигенсвязывающий фрагмент вызывает клеточное рассеивание, равное, по меньшей мере, 25% от индуцированного 0,1 нM гомологичным HGF, когда концентрация антитела составляет 1 нM или ниже;
(ii) анти-апоптотический клеточный анализ, антитело или его антигенсвязывающий фрагмент демонстрирует EC50 не более чем 7,0x значение в отношении HGF и/или Emax жизнеcпособность клеток, по меньшей мере, 50% от наблюдаемой в отношении HGF; и/или
(ii) анализ морфогенеза ветвления, клетки, обработанные антителом, демонстрируют, по меньшей мере, 25% числа ответвлений на сфероид, индуцированных такой же (ненулевой) концентрацией HGF;
и антитело или антигенсвязывающий фрагмент не полностью индуцирует HGF-подобный клеточный ответ.
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент является HGF конкурентом. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент конкурирует с hHGF за связывание с hMET при значении IC50 не более чем 5нM и/или Imax по меньшей мере 50% и конкурирует с mHGF за связывание с mMET при значении IC50 не более чем 5нM и/или Imax по меньшей мере 50%. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент конкурирует с hHGF и mHGF эквивалентно. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент является полным HGF конкурентом. В некоторых подобных вариантах осуществления антитело или его антигенсвязывающий фрагмент конкурирует с hHGF со значением IC50 меньше чем 2 нM и/или Imax больше чем 90%, и конкурирует с mHGF с IC50 меньше чем 2 нM и/или Imax больше чем 90%. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент является частичным HGF-конкурентом. В некоторых подобных вариантах осуществления антитело или его антигенсвязывающий фрагмент конкурирует с hHGF с IC50 2-5нM и/или Imax 50%-90% и конкурирует с mHGF с IC50 2-5нM и/или Imax 50%-90%.
Антитела или их антигенсвязывающий фрагмент изобретения может демонстрировать перекрестную реактивность с MET обезьяньего происхождения, например, яванского макака (Macaca cynomolgus) MET, и могут демонстрировать перекрестную реактивность с MET крысиного происхождения (Rattus norvegicus).
Антитела или их антигенсвязывающий фрагмент изобретения могут связываться с эпитопом человеческого MET от аминокислотного остатка 123 до 223 человеческого MET (на всем протяжении документа нумерация человеческого MET имеет в виду GenBank последовательность # X54559). Также предоставляются антитела или их антигенсвязывающий фрагмент изобретения, который может связываться с эпитопом человеческого MET между аминокислотами 224-311 человеческого MET. Также предоставляются антитела или их антигенсвязывающий фрагмент изобретения, который может связываться с эпитопом человеческого MET между аминокислотами 314-372 человеческого MET. Также предоставляются антитела или их антигенсвязывающий фрагмент изобретения, который может связываться с эпитопом человеческого MET между аминокислотами 546-562 человеческого MET.
Также предоставляются антитела изобретения или их антигенсвязывающий фрагмент, который может связываться с эпитопом человеческого MET, содержащим аминокислотный остаток Ile367. Также предоставляются антитела изобретения или их антигенсвязывающий фрагмент, который может связываться с эпитопом человеческого MET, содержащим аминокислотный остаток Asp372 человеческого MET. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент связывается с эпитопом человеческого MET, содержащим аминокислотные остатки Ile367 и Asp372 человеческого MET.
Также предоставляются антитела изобретения или их антигенсвязывающий фрагмент, который может связываться с эпитопом человеческого MET, содержащим аминокислотный остаток Thr555 человеческого MET.
Изобретение дополнительно предоставляет антитело или его антигенсвязывающий фрагмент, содержащий вариабельный домен тяжелой цепи, включающий H-CDR1, H-CDR2 и H-CDR3, и вариабельный домен легкой цепи, включающий L-CDR1, L-CDR2 и L-CDR3, при этом:
H-CDR1 содержит аминокислотную последовательность, выбранную из SEQ ID NO:2, 9, 16, 23, 30, 37, 44, 51, 58, 65 и 72;
H-CDR2 содержит аминокислотную последовательность, выбранную из SEQ ID NO:4, 11, 18, 25, 32, 39, 46, 53, 60, 67 и 74;
H-CDR3 содержит аминокислотную последовательность, выбранную из SEQ ID NO:6, 13, 20, 27, 34, 41, 48, 55, 62, 69 и 76,
L-CDR1 содержит аминокислотную последовательность, выбранную из SEQ ID NO:79, 86, 93, 100, 107, 114, 121, 128, 135, 142, и 149;
L-CDR2 содержит аминокислотную последовательность, выбранную из SEQ ID NO:81, 88, 95, 102, 109, 116, 123, 130, 137, 144 и 151; и
L-CDR3 содержит аминокислотную последовательность, выбранную из SEQ ID NO:83, 90, 97, 104, 111, 118, 125, 132, 139, 146 и 153.
[71G2] В одном варианте осуществления изобретение предоставляет антитело или антигенсвязывающий фрагмент, содержащий вариабельный домен тяжелой цепи, включающий H-CDR1, H-CDR2 и H-CDR3, и вариабельный домен легкой цепи, включающий L-CDR1, L-CDR2 и L-CDR3, при этом:
H-CDR1 содержит аминокислотную последовательность, показанную как SEQ ID NO:44,
H-CDR2 содержит аминокислотную последовательность, показанную как SEQ ID NO:46,
H-CDR3 содержит аминокислотную последовательность, показанную как SEQ ID NO:48,
L-CDR1 содержит аминокислотную последовательность, показанную как SEQ ID NO:121,
L-CDR2 содержит аминокислотную последовательность, показанную как SEQ ID NO:123, и
L-CDR3 содержит аминокислотную последовательность, показанную как SEQ ID NO:125.
[71G2] В некоторых подобных вариантах осуществления вариабельный домен тяжелой цепи антитела или фрагмент содержит аминокислотную последовательность SEQ ID NO:167, или последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную этой, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO:168, или последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную этой.
[71D6] В другом варианте осуществления изобретение предоставляет антитело или антигенсвязывающий фрагмент, который содержит вариабельный домен тяжелой цепи, включающий H-CDR1, H-CDR2 и H-CDR3, и вариабельный домен легкой цепи, содержащий L-CDR1, L-CDR2 и L-CDR3, при этом:
H-CDR1 содержит аминокислотную последовательность, показанную как SEQ ID NO:30,
H-CDR2 содержит аминокислотную последовательность, показанную как SEQ ID NO:32,
H-CDR3 содержит аминокислотную последовательность, показанную как SEQ ID NO:34,
L-CDR1 содержит аминокислотную последовательность, показанную как SEQ ID NO:107,
L-CDR2 содержит аминокислотную последовательность, показанную как SEQ ID NO:109, и
L-CDR3 содержит аминокислотную последовательность, показанную как SEQ ID NO:111.
[71D6] В некоторых подобных вариантах осуществления вариабельный домен тяжелой цепи антитела или антигенсвязывающий фрагмент содержит аминокислотную последовательность SEQ ID NO:163, или последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную этой, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO:164, или последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную этой.
[71G3] В дополнительном варианте осуществления изобретение предоставляет антитело или антигенсвязывающий фрагмент, содержащий вариабельный домен тяжелой цепи, включающий H-CDR1, H-CDR2 и H-CDR3, и вариабельный домен легкой цепи, включающий L-CDR1, L-CDR2 и L-CDR3, при этом:
H-CDR1 содержит аминокислотную последовательность, показанную как SEQ ID NO:9,
H-CDR2 содержит аминокислотную последовательность, показанную как SEQ ID NO:11,
H-CDR3 содержит аминокислотную последовательность, показанную как SEQ ID NO:13,
L-CDR1 содержит аминокислотную последовательность, показанную как SEQ ID NO:86,
L-CDR2 содержит аминокислотную последовательность, показанную как SEQ ID NO:88, и
L-CDR3 содержит аминокислотную последовательность, показанную как SEQ ID NO:90.
[71G3] В некоторых вариантах осуществления вариабельный домен тяжелой цепи антитела или антигенсвязывающий фрагмент содержит аминокислотную последовательность SEQ ID NO:157, или последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную этой, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO:158, или последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную этой.
В дополнительных вариантах осуществления изобретение предоставляет антитело или антигенсвязывающий фрагмент, содержащий вариабельный домен тяжелой цепи, включающий H-CDR1, H-CDR2 и H-CDR3, и вариабельный домен легкой цепи, содержащий L-CDR1, L-CDR2 и L-CDR3, при этом H-CDR1, H-CDR2 и H-CDR3 выбирают из набора CDRs (CDR1, CDR2 и CDR3) для Fab, показанного в Таблице 3, и L-CDR1, L-CDR2 и L-CDR3 представляют собой соответствующие CDRs (CDR1, CDR2 и CDR3) для того же самого Fab, показанного в Таблице 4.
В некоторых вариантах осуществления вариабельный домен тяжелой цепи антитела или антигенсвязывающий фрагмент содержит VH аминокислотную последовательность из Таблицы 5 или последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную этой, и вариабельный домен легкой цепи содержит соответствующую VL аминокислотную последовательность из Таблицы 5 или последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную этой.
Варианты осуществления, в которых аминокислотная последовательность VH домена демонстрирует менее чем 100% идентичность последовательности с установленной аминокислотной последовательностю VH домена (например, SEQ ID NO: x) могут, тем не менее, включать CDR тяжелой цепи, которые являются идентичными с HCDR1, HCDR2 и HCDR3 VH последовательности SEQ ID NO: x, при этом демонстрируя изменение аминокислотной последовательности в пределах каркасных участков. Например, один или более аминокислотных остатков каркасного участка может замещаться аминокислотным остатком, который содержится в эквивалентном положении в человеческом VH домене, кодируемым человеческой зародышевой линией. Аналогично, варианты осуществления, в которых аминокислотная последовательность VL домена демонстрирует менее чем 100% идентичность последовательности с установленной аминокислотной последовательностью VL домена (например, SEQ ID NO:y) может несмотря на это содержать CDR легкой цепи, которые являются идентичными LCDR1, LCDR2 и LCDR3 VL последовательности SEQ ID NO:y, при этом демонстрируя изменение аминокислотной последовательности в пределах каркасных участков. Например, один или более аминокислотных остатков каркасного участка может замещаться аминокислотным остатком, который содержится в эквивалентном положении в человеческом VL домене, кодируемом человеческой зародышевой линией.
Изобретение также предоставляет антитела и антигенсвязывающие фрагменты, содержащие гуманизированные/модифицированные на уровне генов зародышевой линии варианты VH и VL доменов вышеупомянутых антител, плюс аффинные варианты и варианты, содержащие консервативные аминокислотные замены, как определено в данном документе. Конкретнее, предоставляются химерные антитела, содержащие VH и VL домены Fab, происходящих от ламы, описанные выше, или человеческие модифицированные на уровне генов зародышевой линии их варианты, слитые с константными доменами человеческих антител, в частности IgG1, IgG2, IgG3 или IgG4 человека. Вариабельные домены тяжелой и легкой цепи вышеупомянутых антител или модифицированные на уровне генов зародышевой линии варианты, аффинные варианты или их консервативные варианты могут быть включены в обычное четырех-цепочечное антитело или другие антигенсвязывающие белки, такие как, например Fab, Fab', F(ab')2, биспецифические Fab и Fv фрагменты, диатела, линейные антитела, молекулы одноцепочечных антител, одноцепочечный вариабельный фрагмент (scFv) и мультиспецифические антитела. Вариабельные домены тяжелой цепи или модифицированный на уровне генов зародышевой линии вариант, аффинный вариант или их консервативный вариант также могут использоваться в виде однодоменных антител.
В дополнительных аспектах изобретение также предоставляет изолированный полинуклеотид, который кодирует антитело или антигенсвязывающий фрагмент изобретения, вектор экспрессии, содержащий указанный полинуклеотид, функционально связанный с регуляторными последовательностями, которые обеспечивают экспрессию антитела или его антигенсвязывающего фрагмента в клетке-хозяине или бесклеточной экспрессирующей системе, и клетку-хозяина или бесклеточную экспрессирующую систему, содержащую указанный вектор экспрессии. Изобретение дополнительно предоставляет способ продуцирования рекомбинантного антитела или его антигенсвязывающего фрагмента, который включает культивирование указанной клетки-хозяина или бесклеточной экспрессирующей системы при условиях, которые дают возможность экспрессии антитела или антигенсвязывающего фрагмента, и извлечение экспрессированного антитела или антигенсвязывающего фрагмента.
В дополнительном аспекте изобретение предоставляет фармацевтическую композицию, содержащую антитело или антигенсвязывающий фрагмент изобретения и, по меньшей мере, один фармацевтически приемлемый носитель или эксципиент.
В дополнительном аспекте изобретение предоставляет антитело или антигенсвязывающий фрагмент изобретения или фармацевтическую композицию изобретения для использования в терапии.
В дополнительном аспекте изобретение предоставляет способ лечения или предотвращения поражения печени у пациента, необязательно острого поражения печени или хронического поражения печени, который включает введение нуждающемуся в этом пациенту терапевтически эффективного количества MET агонистического антитела. В некоторых вариантах осуществления MET агонистическое антитело является антителом или антигенсвязывающим фрагментом согласно изобретению.
В дополнительном аспекте изобретение предоставляет способ лечения или предотвращения поражения почки у пациента, необязательно острого поражения почки, который включает введение нуждающемуся в этом пациенту терапевтически эффективного количества MET агонистического антитела. В некоторых вариантах осуществления MET агонистическое антитело является антителом или антигенсвязывающим фрагментом согласно изобретению.
В дополнительном аспекте изобретение предоставляет способ лечения или предотвращения воспалительной болезни кишечника у человека-пациента, необязательно язвенного колита, который включает введение нуждающемуся в этом пациенту терапевтически эффективного количества MET агонистического антитела. В некоторых вариантах осуществления MET агонистическое антитело является антителом или антигенсвязывающим фрагментом согласно изобретению.
В дополнительном аспекте изобретение предоставляет способ лечения или предотвращения диабета у человека-пациента, необязательно диабета I типа или II типа, который включает введение нуждающемуся в этом пациенту терапевтически эффективного количества MET агонистического антитела.
В некоторых вариантах осуществления MET агонистическое антитело является антителом или антигенсвязывающим фрагментом согласно изобретению.
В дополнительном аспекте изобретение предоставляет способ лечения или предотвращения неалкогольного стеатогепатита у человека-пациента, который включает введение нуждающемуся в этом пациенту терапевтически эффективного количества MET агонистического антитела. В некоторых вариантах осуществления MET агонистическое антитело является антителом или антигенсвязывающим фрагментом согласно изобретению.
В дополнительном аспекте изобретение предоставляет способ лечения или содействия заживлению ран у человека-пациента, необязательно пациента, имеющего диабет, который включает введение нуждающемуся в этом пациенту терапевтически эффективного количества MET агонистического антитела. В некоторых вариантах осуществления MET агонистическое антитело является антителом или антигенсвязывающим фрагментом согласно изобретению.
Краткое описание чертежей
Фигура 1. Иммунный ответ у лам, иммунизированных человеческим MET-Fc, установленный методом ELISA. Рекомбинантный белок человеческий MET ECD (hMET) или мышиный MET ECD (mMET) был иммобилизован на твердой фазе и подвергнут воздействию серийных разведений сыворотки лам до (PRE) или после (POST) иммунизации. Связывание было установлено при помощи мышиного анти-лама IgG1 и HRP-конъюгированного ослиного анти-мышиного антитела. OD, оптическая плотность; AU, относительные единицы.
Фигура 2. Схематическое изображение человеческих делеционных мутантов MET , использованных для идентификации доменов MET, ответственных за связывание mAb. ECD, внеклеточный домен; aa, аминокислота; L. пептид, лидерный пептид; SEMA, домен гомологии семафоринов; PSI или P, домен гомологии плексин-семафорин-интегринов; IPT, домен гомологии иммуноглобулин-транскрипционный фактор-плексинов. Справа приведены соответствующие остатки человеческого MET согласно UniProtKB # P08581.
Фигура 3. Схематическое изображение лама-человеческих химерных MET белков, использованных для точного картирования эпитопов, распознаваемых анти-MET антителами. Внеклеточные участки MET ламы и человеческого MET состоят из 931 и 932 аминокислот (aa), соответственно (MET ламы имеет 2 aa более коротких лидерных пептида, но имеет вставку после aa 163). Оба эктодомена рецепторна содержат лидерный пептид, домен гомологии семафоринов (SEMA), домен гомологии плексин-семафорин-интегринов (PSI или P) и четыре домена гомологии иммуноглобулин-транскрипционный фактор-плексинов (IPT). Химеры CH1-5 имеют N-концевой участок ламы, за которым следует C-концевой человеческий участок. Химеры CH6-7 имеют N-концевой человеческий участок, за которым следует C-концевой участок ламы.
Фигура 4. Агонистическая активность человеческих/мышиных эквивалентных анти-MET антител в человеческих и мышиных клетках, измеренная при помощи вестерн-блоттинга. Клетки карциномы легких человека A549 и клетки-предшественники печени мыши MLP29 содержали в бессывороточной среде, а затем стимулировали увеличивающимися концентрациями mAb или рекомбинантного человеческого HGF (hHGF; A549) или мышиного HGF (mHGF; MLP29). Аутофосфорилирование MET определяли с помощью вестерн-блоттинга с использованием анти-фосфо-MET антител (тирозины 1234-1235). Те же самые клеточные лизаты также исследовали с помощью вестерн-блоттинга с использованием антител против тотального человеческого MET (A549) или антител против тотального мышиного MET (MLP29).
Фигура 5. Биологическая активность человеческих/мышиных эквивалентных анти-MET антител, определенная путем анализа морфогенеза ветвления с использованием эпителиальных клеток почки человека LOC и клеток-предшественников печени мыши MLP29. Клеточные сфероиды высевали внутрь коллагенового слоя и затем подвергали воздействию возрастающих концентраций mAb или рекомбинантного человеческого HGF (LOC) или мышиного HGF (MLP29). Морфогенез ветвления наблюдали в течение длительного периода времени с помощью микроскопии, колонии фотографировали через 5 дней.
Фигура 6. Сравнение с антителами предшествующего уровня техники: перекрестная реактивность человек-мышь. Человеческий или мышиный MET ECD иммобилизировали на твердой фазе и подвергали воздействию возрастающих концентраций антител (все в мышином IgG/λ формате) в растворе. Связывание выявляли с помощью ELISA с использованием HRP-конъюгированных анти-мышиных Fc антител.
Фигура 7. Сравнение с антителами предшествующего уровня техники: аутофосфорилирование MET. Клетки карциномы легких человека A549 и клетки-предшественники печени мыши MLP29 были лишены сывороточных факторов роста в течение 48 часов, а затем их стимулировали возрастающими концентрациями антител. После 15 минут стимулирования клетки лизировали и определяли уровни фосфо-MET с помощью ELISA с использованием анти-MET антител для захвата и анти-фосфо-тирозин антител для обнаружения.
Фигура 8. Сравнение с антителами предшествующего уровня техники: морфогенез ветвления. Сфероиды эпителиальных клеток почки человека LOC высевали в коллагеновый слой и затем инкубировали с возрастающими концентрациями mAb. Морфогенез ветвления наблюдали в течение длительного периода времени с помощью микроскопии, колонии фотографировали через 5 дней.
Фигура 9. Сравнение с антителами предшествующего уровня техники: морфогенез ветвления. Сфероиды клеток-предшественников печени мыши MLP29 высевали в коллагеновый слой и затем инкубировали с возрастающими концентрациями mAb. Морфогенез ветвления наблюдали в течение длительного периода времени с помощью микроскопии, колонии фотографировали через 5 дней.
Фигура 10. Стабильность в плазме человеческих/мышиных эквивалентных анти-MET антител. Однократную дозу 1 мг/кг или 10 мг/кг антител вводили i.p., образцы крови брали из хвостовой вены через 3, 6, 12 и 24 часов после инъекции. Образцы крови обрабатывали и определяли концентрацию антител в плазме с помощью ELISA. (A) Пиковый и остаточный уровни инъецированных антител. (B) Время полужизни антитела в плазме было вычислено с помощью линейной аппроксимации Ln преобразований концентрации антитела.
Фигура 11. Модель острой печеночной недостаточности: концентрация в плазме маркеров функции печени. Острую печеночную недостаточность индуцировали у мышей BALB/c с помощью подкожной инъекции раствора CCl4. Вскоре после первой инъекции CCl4 мышей случайным образом разделили на 4 группы, которые получали одноразовую дозу 71G3, 71D6, 71G2 или только разбавитель (PBS). Антитела вводили с помощью i.p. инъекции в дозе 5 мг/кг. Каждая группа содержала три подгруппы мышей, которых умерщвляли в разные моменты времени после интоксикации (12, 24 и 48 часов). Образцы крови собирали в разные временные точки после инъекции (0, 12, 24 и 48 часов). При аутопсии кровь и печень собирали для проведения анализов. Уровни в плазме печеночных маркеров аспартат трансаминазы (AST), аланин аминотрансферазы (ALT) и билирубина (BIL) определяли с помощью стандартных клинических биохимических методов.
Фигура 12. Модель острой печеночной недостаточности: гистологическое исследование срезов печени. Острую печеночную недостаточность индуцировали у мышей BALB/c, как описано в подписи к Фигуре 11. При аутопсии печень извлекали, экстрагировали и заливали в парафин для проведения гистологических анализов. Срезы окрашивали гематоксилином и эозином и исследовали под микроскопом. Показано характерное изображение для каждой лечебной группы. Увеличение: 100X.
Фигура 13. Модель хронического поражения печени: концентрация в плазме маркеров функции печени. Поражение печени и фиброз у мышей BALB/c индуцировали путем длительного воздействия CCl4 в теение нескольких недель. Вскоре после первой инъекции CCl4 мышей случайным образом разделили на 4 группы, которые получали лечение 71G3, 71D6, 71G2 или только разбавителем (PBS), соответственно. Антитела вводили три раза в неделю с помощью i.p. инъекции в дозе 1 мг/кг. Дополнительная пятая контрольная группа не получала CCl4 или антитело и служила в качестве здорового контроля. Мышей умерщвляли после 6 недель хронической CCl4 интоксикации. При аутопсии кровь и печень извлекали для проведения анализов. Уровни в плазме печеночных маркеров аспартат трансаминазы (AST) и аланин аминотрансферазы (ALT) определяли с помощью стандартных клинических биохимических методов.
Фигура 14. Модель хронического поражения печени: гистологическое исследование срезов печени, окрашенных пикросириусом красным. Поражение печени и фиброз у BALB/c мышей индуцировали путем длительного воздействия CCl4, как описано в подписи к Фигуре 13. При аутопсии печень извлекали, экстрагировали и заливали в парафин для проведения иммуногистохимических анализов. Срезы окрашивали пикросириусом красным. Показано характерное изображение для каждой лечебной группы. Увеличение: 100X.
Фигура 15. Модель хронического поражения печени: гистологическое исследование срезов печени, окрашенных антителами к альфа-актину гладких мышц (α-SMA). Поражение печени и фиброз у мышей BALB/c индуцировали путем длительного воздействия CCl4, как описано в подписи к Фигуре 13. При аутопсии печень извлекали, экстрагировали и заливали в парафин для проведения иммуногистохимических анализов. Срезы окрашивали антителами к альфа-актину гладких мышц (α-SMA). Показаны характерные изображения для каждой лечебной группы. Увеличение: 100X.
Фигура 16. Модель острой почечной недостаточности: уровни в плазме маркеров почечной функции. Острую почечную недостаточность индуцировали у мышей BALB/c с помощью i.p. инъекции однократной дозы HgCl2. Вскоре после HgCl2 интоксикации мышей случайным образом разделили на 4 группы, которые получали лечение 71G3, 71D6, 71G2 или только разбавителем (PBS). Антитела вводили с помощью i.p. инъекции каждые 24 часа в дозе 5 мг/кг. Мышей умерщвляли через 72 часа после инъекции HgCl2. При аутопсии кровь и почки отбирали для проведения анализов. Уровень азота мочевины в крови (BUN) и уровень креатинина в плазме (CRE) определяли с помощью стандартных клинических биохимических методов.
Фигура 17. Модель острой почечной недостаточности: гистологическое исследование срезов почки. Острую почечную недостаточность индуцировали у мышей BALB/c с помощью инъекции HgCl2, как описано в подписи к Фигуре 16. При аутопсии почки извлекали, экстрагировали и заливали в парафин для проведения гистологических анализов. Срезы почек окрашивали гематоксилином и эозином. Показано характерное изображение для каждого лечебной группы. Увеличение: 400X.
Фигура 18. Модель язвенного колита: вес тела, индекс активности болезни (DAI) и длина толстой кишки. Язвенный колит индуцировали у мышей BALB/c добавлением декстрансульфата натрия (DSS) к питьевой воде в течение 10 дней. На день 10 обработку DSS прерывали и мышей возвращали на потребление нормальной воды. Начиная с 1 дня, мышей случайным образом разделили на 7 групп, которые получали лечение 71G3, 71D6, 71G2 (в дозе 1 мг/кг или 5 мг/кг) или только разбавитель (PBS). Дополнительная восьмая контрольная нруппа не получала DSS или антитело и служила в качестве здорового контроля. Мышей умерщвляли на день 12, т.е. через 2 дня после того, как введение DSS было прекращено. При аутопсии извлекали толстый кишечник, промывали и определяли длину с помощью рулетки. После измерения кишку заливали в парафин и обрабатывали для проведения гистологических анализов. В течение всего эксперимента регулярно контролировали вес мышей и оценивали клинические симптомы язвенного колита путем обнаружения крови в фекалиях, ректального кровотечения и консистенции стула. Каждый параметр оценивался в баллах от 0 (отсутствие симптомов) до 3 (максимальное проявление симптома). Баллы, относящиеся к отдельным параметрам, суммировали вместе, чтобы выразить DAI в пределах от 0 до 9. (A) Вес тела в течение времени (% относительно времени 0). (B) DAI в течение времени. (C) Длина толстой кишки при аутопсии. Данные в группах 1 мг/кг и группах 5 мг/кг показаны на отдельных графиках для ясности.
Фигура 19. Модель язвенного колита: гистологическое исследование срезов кишки. Язвенный колит индуцировали у мышей BALB/c воздействием декстрансульфата натрия (DSS), как описано в подписи к Фигуре 18. При аутопсии толстую кишку извлекали, измеряли, заливали в парафин и обрабатывали для проведения гистологических анализов. Срезы кишки окрашивали гематоксилином и эозином, исследовали с помощью микроскопии и фотографировали. Экспериментальная группа, доза антитела и увеличение указаны на каждом изображении. В отношении анализа изображений просим обращаться к основному тексту.
Фигура 20. Модель воспалительной болезни кишечника: вес тела и длина кишки. Поражение толстой кишки и воспаление индуцировали у C57BLKS/J мышей с помощью внутриректальной инъекции 2,4,6-тринитробензолсульфоновой кислоты (TNBS), растворенной в этаноле. Вскоре после введения TNBS мышей случайным образом разделили на 4 группы, которые обрабатывали 71G3, 71D6, 71G2 или только растворителем (PBS). Дополнительная, пятая контрольная группа не получала TNBS или антитело и служила в качестве здорового контроля. Мышей умерщвляли через 5 дней после введения TNBS. При аутопсии кишку извлекали и измеряли. После измерения кишку заключали в парафин и обрабатывали для проведения гистологических исследований. В течение всего эксперимента каждый день определяли вес мышей. (A) Вес тела в течение времени (% относительно времени 0). (B) Длина кишки при аутопсии.
Фигура 21. Модель воспалительной болезни кишечника: гистологическое исследование срезов кишки. Поражение толстой кишки и воспаление индуцировали у мышей BALB/c с помощью внутриректальной инъекции 2,4,6- тринитробензолсульфоновой кислоты (TNBS), как описано в подписи к Фигуре 20. При аутопсии кишку извлекали и измеряли. После измерения кишку заключали в парафин и обрабатывали для проведения гистологических исследований. Срезы кишечника окрашивали гематоксилином и эозином, исследовали с помощью микроскопии и фотографировали. В отношении анализа изображений просим обращаться к основному тексту.
Фигура 22. Модель диабета I типа: стимулирование поглощения глюкозы и взаимодействие с инсулином у мышей с диабетом. Деградацию β-клеток поджелудочной железы индуцировали у мышей BALB/c с помощью i.p. инъекции стрептозотоцина (STZ). Мыши, обработанные STZ, демонстрировали в два раза более высокую среднюю базальную гликемию по сравнению с необработанными мышами. STZ-обработанных мышей разделили случайным образом на 4 группы, которые получали лечение 71G3, 71D6, 71G2 или только разбавителем (PBS), соответственно. Дополнительная, пятая контрольная группа не получала STZ или антитело и служила в качестве здорового контроля. Концентрацию глюкозы в крови натощак контролировали в течение 5 недель. В конце 5 недели были проведены тест на толерантность к глюкозе (GTT) и тест на толерантность к инсулину (ITT). (A) Анализ в течение времени базальных уровней глюкозы в крови натощак. (B) GTT: после перорального введения глюкозы голодному животному, уровни глюкозы в крови контролировали в течение времени. (C) ITT: после i.p. инъекции инсулина частично голодному животному, уровни глюкозы в крови контролировали в течение времени.
Фигура 23. Модель диабета I типа: стимулирование поглощения глюкозы и взаимодействие с инсулином в культивируемых клетках. Мышиные миобласты C2C12 стимулировали к дифференцировке в миоциты, а затем инкубировали с человеческими/мышиными эквивалентными агонистическими анти-MET антителами (71G3, 71D6, 71G2). Через 24 часа, клетки, обработанные антителами, разделили на три 3 группы, которые были подвергнуты острой стимуляции 0 нM, 100 нM или 1000 нM человеческого рекомбинантного инсулина в течение 1 часа в присутствии флуоресцентного аналога глюкозы 2-(N-(7-нитробенз-2-окса-1,3-диазол-4-ил)амино)-2-дезоксиглюкозы (2-NBDG). 2-NBDG поглощение определяли с помощью проточной цитометрии. (A) Индукция поглощения 2-NBDG человеческими/мышиными эквивалентными агонистическими анти-MET антителами или инсулином. (B) Индукция поглощения 2-NBDG 71G3 при отсутствии или в присутствии инсулина. (C) Индукция 2-NBDG поглощения 71D6 в отсутствие или в присутствии инсулина. (D) Индукция 2-NBDG поглощения 71G2 в отсутствие или в присутствии инсулина.
Фигура 24. Модель диабета II типа: нормализация уровня глюкозы в крови и преодоление резистентности к инсулину у мышей db/db. Самок db/db мышей 8-недельного возраста (C57BLKS/J вариант, несущий точечную мутацию в гене лептинового рецептора lepr) разделили случайным образом на 4 группы, которые обрабатывали 71G3, 71D6, 71G2 или только разбавителем (PBS), соответственно. Антитела вводили два раза в неделю с помощью i.p. инъекции в дозе 1 мг/кг. Концентрацию глюкозы в крови натощак контролировали каждые 10 дней в течение 7 недель. В конце обработки, т.е. когда мыши достигали 15-недельного возраста, были проведены тест на толерантность к глюкозе (GTT) и тест на толерантность к инсулину (ITT) с использованием C57BLKS/J мышей дикого типа в качестве контроля по возрастным группам. (A) Концентрация глюкозы в крови в течение времени. (B) GTT: после перорального введения глюкозы голодному животному уровни глюкозы в крови контролировали в течение некоторого времени. (C) ITT: после i.p. инъекции инсулина отчасти голодному животному уровни глюкозы в крови контролировали в течение некоторого времени. (D) ITT: концентрация глюкозы в крови через 15 минут после i.p. инъекции инсулина.
Фигура 25. Мышиная модель неалкогольного стеатогепатита (NASH): улучшение состояния при жировой дистрофии печени, определенное с помощью гистологического исследования. Мышей 8-недельного возраста db/db случайным образом разделили на 4 группы, которые получали лечение 71G3, 71D6, 71G2 или только разбавителем (PBS), соответственно. Антитела вводили два раза в неделю с помощью i.p. инъекции в дозе 1 мг/кг. После 8 недель обработки мышей умерщвляли и подвергали аутопсии. Кровь собирали для проведения исследования маркеров функции печени. Печень извлекали, заливали в парафин и обрабатывали для гистологического исследования. Срезы печени окрашивали гематоксилином и эозином. Цитоплазма жировых клеток выглядит пустой и белой, потому что липиды вымываются во время обработки образца спиртом. Показано характерное изображение для каждой лечебной группы. Увеличение: 200X.
Фигура 26. Мышиная модель неалкогольного стеатогепатита (NASH): подавление фиброза, установленное с помощью окрашивания пикросириусом красным. Мышей с диабетом 8-недельного возраста db/db случайным образом разделили на группы и обработали, как описано в подписи к Фигуре 25. При аутопсии печень собирали для проведения гистологического исследования. Срезы печени окрашивали пикросириусом красным, чтобы продемонстрировать фиброз. Показано характерное изображение для каждой лечебной группы. Увеличение: 200X.
Фигура 27. Мышиная модель неалкогольного стеатогепатита (NASH): нормализация маркеров печеночной функции. Мыши с диабетом db/db 8-недельного возраста были обработаны очищенными 71G3, 71D6, 71G2 или только разбавителем, как описано в подписи к Фигуре 25. Через 7 недель после обработки кровь собирали для анализа маркеров печеночной функции. (A) Уровни в плазме аспартат-трансаминазы (AST). (B) Уровни в плазме аланин-аминотрансферазы (ALT).
Фигура 28. Мышиная модель диабетических язв: ускоренное заживление ран. Мышей db/db 8-недельного возраста с диабетом обезболили, а затем с помощью инструмента с круговым лезвием шириной 0,8 см для взятия биопсийного образца кожи вырезали круглую рану на правом боку. Весь эпидермальный слой был удален. Через день после хирургической процедуры мышей случайным образом разделили на 4 группы, которые получали лечение очищенным 71G3, 71D6 и 71G2 или только разбавителем (PBS). Антитела вводили через день с помощью i.p инъекции в дозе 5 мг/кг. Диаметр раны измеряли каждый день, используя штангенциркуль. (A) Площадь раны в течение времени. (B) Средняя скорость реэпитализации, как определено усреднением ежедневного % закрытия раны.
Фигура 29. Rattus norvegicus и Macaca fascicularis перекрестная реактивность, определенная с помощью ELISA. Для того чтобы протестировать пан-видовую перекрестную реактивность, была выбрана ограниченная панель антител, характерных как для SEMA-связывающих антител (71D6, 71C3, 71D4, 71A3, 71G2), так и PSI-связывающих антител (76H10, 71G3). Антитело 5D5 предшествующего уровня техники использовали в качестве контроля. Человеческий, мышиный, крысиный или обезьяний MET ECD был иммобилизирован на твердой фазе и подвергнут воздействию возрастающих концентраций mAbs (в их человеческом IgG1/λ формате) в растворе. Связывание обнаруживали с помощью HRP-конъюгированных анти-человек Fc антител.
Фигура 30. Выравнивание аминокислотной последовательности между MET ECD доменами от H. sapiens, M. musculus, R. norvegicus, M. fascicularis и L. glama. (A) Выравнивание последовательности относительно участка, распознаваемого SEMA-связывающими антителами (71D6, 71C3, 71D4, 71A3 и 71G2) (человеческая MET последовательность SEQ ID NO: 239; мышиная MET последовательность SEQ ID NO: 240; крысиная MET последовательность SEQ ID NO: 241, яванского макака MET последовательность SEQ ID NO: 242, ламы MET последовательность SEQ ID NO: 243). Аминокислоты, идентифицированные при помощи подхода с использованием химеры человек-лама и показанные в Таблице 12, подчеркнуты. В пределах этого участка имеется пять остатков, которые являются консервативными в MET человека и мыши, но не в MET ламы (Ala 327, Ser 336, Phe 343, Ile 367, Asp 372). Эти аминокислоты указаны с помощью черных квадратов и порядковых номеров 1-5. Из них четыре остатка также являются консервативными у MET крысы и яванской макаки (Ala 327, Ser 336, Ile 367, Asp 372). Аминокислоты, ответственные за связывание с SEMA-связывающими антителами, указаны с помощью “S” (SEMA). Аминокислоты, ответственные за связывание с 5D5/онартузумаб указаны с помощью “O” (онартузумаб). (B) Выравнивание последовательностей относительно участка, распознаваемого PSI-связывающими антителами 76H10 и 71G3 (человеческая MET последовательность SEQ ID NO: 244; мышиная MET последовательность SEQ ID NO: 245; крысиная MET последовательность SEQ ID NO: 246, яванского макака MET последовательность SEQ ID NO: 247, MET последовательность ламы SEQ ID NO: 248). Аминокислоты, установленные с помощью подхода с использованием химеры человек-лама и показанные в Таблице 12, подчеркнуты. В пределах этого участка существует три остатка, которые являются консервативными у человека и мыши MET, но не у MET ламы (Arg 547, Ser 553, Thr 555). Эти аминокислоты указаны с помощью черных квадратов и порядковых номеров 6-8. Из них два остатка также являются консервативными в MET крысы и яванской макаки (Ser 553 и Thr 555). Аминокислота, отсветственная за связывание с PSI-связывающими антителами указана с помощью “P” (PSI).
Фигура 31. Схематическое изображение MET мутантов, использованных для точного картирования эпитопов. Используя человеческий MET ECD в качестве матрицы, ключевые остатки, обозначенные порядковыми номерами 1-8 на Фигуре 30, были подвергнуты мутагенезу при использовании разных перестановок, с получением мутантов A-L. Каждый из этих мутантов является полностью человеческим, за исключением указанных остатков, которые являются остатками от ламы.
Подробное описание изобретения
Использованный в описании термин "иммуноглобулин" включает полипептид, имеющий комбинацию двух тяжелых и двух легких цепей, обладает он или нет какой-либо релевантной специфической иммунореактивностью. Термин "антитела" относится к таким структурам, которые обладают значительной известной специфической иммунореактивной активностью в отношении представляющего интерес агнтигена (например, MET). Термин “MET антитела” или “анти-MET антитела” при использовании в описании относится к антителам, которые демонстрируют иммунологическую специфичность в отношении MET белка. Антитела и иммуноглобулины содержат легкую и тяжелую цепи, с или без межцепочечной ковалентной связи между ними. Структуры основных иммуноглобулинов у позвоночных относительно хорошо изучены.
Собирательный термин “иммуноглобулин” включает пять различных классов антител, которые можно отличить биохимически. Хотя все пять классов антител включаются в рамки настоящего изобретения, следующее обсуждение будет в основном относиться к IgG классу молекул иммуноглобулинов. Что касается IgG, иммуноглобулины содержат две идентичные легкие полипептидные цепи с молекулярной массой приблизительно 23000 дальтон, и две идентичные тяжелые цепи с молекулярной массой 53000-70000. Четыре цепи соединяются дисульфидными связями в "Y" конфигурации, при этом легкие цепи «скрепляют» тяжелые цепи, начиная с «горлышка» "Y" и проходя по всей длине вариабельного участка.
Легкие цепи антитела классифицируются как каппа или лямбда (κ, λ). Каждый класс тяжелых цепей может быть связан или с каппа или лямбда легкой цепью. В общем, легкая и тяжелая цепи являются ковалентно связанными друг с другом, и "хвостовые" участки двух тяжелых цепей соединяются друг с другом с помощью ковалентных дисульфидных связей или нековалентных связей, когда иммуноглобулины вырабатываются B-клетками или генетически сконструированными клетками-хозяевами. В тяжелой цепи аминокислотные последовательности проходят от N-конца на раздвоенных концах Y конфигурации и до C-конца у основания каждой цепи. Специалистам в данной области техники понятно, что тяжелые цепи классифицируются как гамма, мю, альфа, дельта или эпсилон (γ, μ, α, δ, ε) при наличии нескольких подклассов среди них (например, γ1-γ4). Природа этой цепи определяет "класс" антитела как IgG, IgM, IgA, IgD или IgE, соответственно. Подклассы иммуноглобулинов (изотип) например, IgG1, IgG2, IgG3, IgG4, IgA1 и т.д. хорошо охарактеризованы и, как известно, соответствуют функциональной специализации. Специалисту в данной области будут очевидны модифицированные версии каждого из этих классов и изотипов с точки зрения настоящего раскрытия, которые соответственно включаются в рамки настоящего изобретения.
Как указано выше, вариабельный участок антитела дает возможность антителу селективно распознавать и специфически связываться с эпитопами на антигенах. Иначе говоря, VL домен и VH домен антитела объединяются с образованием вариабельного участка, который определяет трехмерный антигенсвязывающий сайт. Эта четвертичная антительная структура образует антигенсвязывающий сайт, присутствующий на конце каждого «плеча» Y. Конкретнее, антигенсвязывающий сайт определяется тремя определяющими комплементарность участками (CDR) на каждой VH и VL цепи.
Использованные в описании термины "MET белок" или "MET антиген" или "MET" используются взаимозаменяемым образом и относятся к рецепторной тирозинкиназе, которая в форме белка дикого типа связывается с фактором роста гепатоцитов (HGF). Термины “человеческий MET белок” или “человеческий MET рецептор” или “человеческий MET” или “hMET” используются взаимозаменяемым образом и относятся к человеческому MET (учетный номер в GenBank X54559), включая нативный человеческий MET белок, в естественных условиях экспрессированный у человека-хозяина и/или на поверхности культивируемых клеточных линий человека, а также рекомбинантные формы и их фрагменты, а также мутантные формы природного происхождения. Термины “мышиный MET белок” или “мышиный MET рецептор” или “мышиный MET” или “mMET” используются взаимозаменяемым образом и относятся к мышиному MET (учетный номер в GenBank: NM_008591), включая нативный мышиный MET белок, в естественных условиях экспрессированный у мыши-хозяина и/или на поверхности культивируемых мышиных клеточных линий, а также рекомбинантные формы и их фрагменты, а также мутантные формы природного происхождения.
Использованный в описании термин “сайт (участок) связывания” включает участок полипептида, который является ответственным за селективное связывание с представляющим интерес антигеном-мишенью (например, hMET). Домены связывания содержат, по меньшей мере, один сайт связывания. Характерные домены связывания включают вариабельный домен антитела. Молекулы антител изобретения могут содержать один сайт связывания или несколько (например, два, три или четыре) сайтов связывания.
Использованный в описании термин "происходящий из" означенный белок (например, MET антитело или его антигенсвязывающий фрагмент) имеет отношение к происхождению полипептида. В одном варианте осуществления полипептид или аминокислотная последовательность, которая происходит из конкретного исходного полипептида, является CDR последовательностью или последовательностью, связанной с ней. В одном варианте осуществления аминокислотная последовательность, которая происходит из конкретного исходного полипептида, не является непрерывной. Например, в одном варианте осуществления одна, две, три, четыре, пять или шесть CDR происходят из исходного антитела. В одном варианте осуществления полипептид или аминокислотная последовательность, которая происходит из конкретного исходного полипептида или аминокислотной последовательности, имеет аминокислотную последовательность, которая в основном является идентичной исходной последовательности, или части ее, при этом часть состоит, по меньшей мере, из 3-5 аминокислот, по меньшей мере, из 5-10 аминокислот, по меньшей мере, 10-20 аминокислот, по меньшей мере, 20-30 аминокислот или, по меньшей мере, 30-50 аминокислот, или которая может быть иным способом идентифицирована средним специалистом в данной области, как последовательность, основанная на исходной последовательности. В одном варианте осуществления одна или более CDR последовательностей, происходящих от исходного антитела, подвергаются преобразованию для того, чтобы получить вариантные CDR последовательности, например, варианты с разной аффинностью (аффинные варианты), при этом вариант CDR последовательности сохраняет MET связывающую активность.
“Верблюжьего происхождения” --- В некоторых предпочтительных вариантах осуществления молекулы MET антитела изобретения содержат каркасные аминокислотные последовательности и/или CDR аминокислотные последовательности, происходящие из обычного верблюжьего антитела, индуцированного активной иммунизацией животного, относящегося к семейству верблюдовых, с использованием MET-происходящего антигена. Однако, MET-антитела, содержащие аминокислотные последовательности верблюжьего происхождения, могут быть сконструированы так, чтобы они содержали последовательности каркасных и/или константных участков, полученных из человеческой аминокислотной последовательности (т.е. антитела человека) или других видов млекопитающих, не относящихся к верблюдовым. Например, человеческий каркасный участок или каркасный участок отличного от человека примата, участок тяжелой цепи и/или шарнирный участок может быть включен в предметные MET антитела. В одном варианте осуществления одна или более неверблюжьих аминокислот может присутствовать в каркасном участке MET антитела “верблюжьего происхождения”, например, верблюжья каркасная аминокислотная последовательность может содержать одну или более аминокислотных мутаций, в которой присутствует соответствующий человеческий аминокислотный остаток или аминокислотный остаток от животного, не являющегося человеком. Более того, VH и VL домены верблюжьего происхождения или их гуманизированные варианты могут быть связаны с константными доменами человеческих антител, чтобы получить химерную молекулу, как подробно описывается в другом месте данного документа.
Использованный в описании термин "консервативная аминокислотная замена" представляет собой замену, при которой аминокислотный остаток заменяется аминокислотным остатком, имеющим сходную боковую цепь. В данной области техники установлены семейства аминокислотных остатков, имеющих сходные боковые цепи, включая основные боковые цепи (например, лизин, аргинин, гистидин), кислые боковые цепи (например, аспарагиновая кислота, глутаминовая кислота), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, остатки заменимых аминокислот в полипептиде иммуноглобулина могут быть заменены остатком другой аминокислоты из боковых цепей того же самого семейства. В другом варианте осуществления цепочка аминокислот может быть заменена структурно сходной цепочкой, которая отличается порядком и/или составом членов семейства боковой цепи.
Использованный в описании термин “участок тяжелой цепи” включает аминокислотные последовательности, происходящие из константных доменов тяжелой цепи иммуноглобулина. Полипептид, содержащий участок тяжелой цепи, включает, по меньшей мере, одно из следующего: CH1 домен, шарнирный (например, верхнюю, среднюю и/или нижнюю шарнирную область) домен, CH2 домен, CH3 домен или их вариант или фрагмент. В одном варианте осуществления антитело или антигенсвязывающий фрагмент изобретения может содержать Fc участок тяжелой цепи иммуноглобулина (например, шарнирный участок, CH2 домен и CH3 домен). В другом варианте осуществления в антителе или антигенсвязывающем фрагменте изобретения может отсутствовать, по меньшей мере, часть константного домена (например, весь или часть CH2 домена). В некоторых вариантах осуществления, по меньшей мере, один и предпочтительно все константные домены происходят из тяжелой цепи иммуноглобулина человека. Например, в одном предпочтительном варианте осуществления участок тяжелой цепи содержит полный шарнирный домен человека. В других предпочтительных вариантах осуществления участок тяжелой цепи содержит полностью человеческий Fc участок (например, последовательности шарнирного домена, CH2 и CH3 доменов из человеческого иммуноглобулина).
В некоторых вариантах осуществления составные константные домены участка тяжелой цепи происходят из разных молекул иммуноглобулина. Например, участок тяжелой цепи полипептида может содержать CH2 домен, происходящий из IgG1 молекулы, и шарнирный участок, происходящий из IgG3 или IgG4 молекулы. В других вариантах осуществления константные домены являются химерными доменами, содержащими участки разных молекул иммуноглобулинов. Например, шарнирный участок может содержать первый участок из IgG1 молекулы и второй участок из IgG3 или IgG4 молекулы. Как изложено выше, среднему специалисту в данной области техники понятно, что константные домены участка тяжелой цепи могут быть модифицированы так, что они отличаются в аминокислотной последовательности от молекулы иммуноглобулина природного происхождения (дикого типа). Другими словами, полипептиды изобретения, раскрытые в описании, могут содержать изменения или модификации в одном или более из константных доменов тяжелой цепи (CH1, шарнирный домен, CH2 или CH3) и/или константном домене легкой цепи (CL). Типичные модификации включают вставки, делеции или замены одной или более аминокислот в одном или более доменах.
Использованный в описании термин "химерный" белок содержит первую аминокислотную последовательность, соединенную со второй аминокислотной последовательностью, с которой она не соединяется в естественных условиях в природе. Аминокислотные последовательности могут при обычных условиях существовать в отдельных белках, которые соединяются вместе в гибридном полипептиде, или они могут при обычных условиях существовать в одном белке, но располагаются в новом порядке в гибридном полипептиде. Химерный белок может быть создан, например, с помощью химического синтеза, или путем создания и трансляции полинуклеотида, в котором пептидные участки закодированы в желательном взаиморасположении. Характерные химерные MET антитела включают гибридные белки, содержащие VH и VL домены верблюжьего происхождения, или их гуманизированные варианты, соединенные с константными доменами человеческого антитела, например, человеческого IgG1, IgG2, IgG3 или IgG4, или соединенные с константными доменами мышиного антитела, например, мышиного IgG1, IgG2a, IgG2b, IgG2c или IgG3.
Использованные в описании термины "вариабельный участок" и "вариабельный домен" используются взаимозаменяемым образом и имеют равноценное значение. Термин "вариабельный" имеет отношение к тому факту, что некоторые участки вариабельных доменов VH и VL в значительной степени отличаются по последовательности среди антител и используются для специфического связывания каждого конкретного антитела с его антигеном-мишенью. Однако вариабельность распределяется неравномерно на всем протяжении вариабельных доменов антител. Вариабельность сосредоточивается в трех сегментах, называемых "гипервариабельными петлями", в каждом VL домене и VH домене, которые образуют часть антигенсвязывающего сайта. Первая, вторая и третья гипервариабельные петли Vлямбда домена легкой цепи называются в настоящем документе L1(λ), L2(λ) и L3(λ) и могут быть охарактеризованы, как содержащие остатки 24-33 (L1(λ) состоит из 9, 10 или 11 остатков аминокислот), 49-53 (L2(λ) состоит из 3 остатков) и 90-96 (L3(λ) состоит из 5 остатков) в VL домене (Morea et al., Methods 20, 267-279, 2000). Первая, вторая и третья гипервариабельные петли Vкаппа домена легкой цепи называются в настоящем документе L1(κ), L2(κ) и L3(κ) и могут быть охарактеризованы, как содержащие остатки 25-33 (L1(κ) состоит из 6, 7, 8, 11, 12 или 13 остатков), 49-53 (L2(κ) состоит из 3 остатков) и 90-97 (L3(κ) состоит из 6 остатков) в VL домене (Morea et al., Methods 20, 267-279, 2000). Первая, вторая и третья гипервариабельные петли VH домена называются в настоящем документе H1, H2 и H3 и могут быть охарактеризованы, как содержащие остатки 25-33 (H1 состоит из 7, 8 или 9 остатков), 52-56 (H2 состоит из 3 или 4 остатков) и 91-105 (H3, высоко изменчивый по длине) в VH домене (Morea et al., Methods 20, 267-279, 2000).
Если не указано иное, термины L1, L2 и L3 соответственно относятся к первой, второй и третьей гипервариабельным петлям VL домена, и включают гипервариабельные петли, полученные от обоих изотипов Vкаппа и Vлямбда. Термины H1, H2 и H3 соответственно относятся к первой, второй и третьей гипервариабельным петлям VH домена, и включают гипервариабельные петли, полученные от любого известного изотипа тяжелой цепи, включая γ, ε, δ, α или μ.
Каждая из гипервариабельных петель L1, L2, L3, H1, H2 и H3 может содержать часть "определяющей комплементарность области" или "CDR", как определено ниже. Термины "гипервариабельная петля" и "определяющая комплементарность область" не являются строго равнозначными, поскольку гипервариабельные петли (HV) определяются на основе структуры, тогда как определяющие комплементарность области (CDR) определяются на основе изменчивости последовательности (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991), и длины (предельные величины) HVs и CDRs могут быть разными в некоторых VH и VL доменах.
CDR VL и VH доменов в большинстве случаев могут быть определены, как содержащие следующие аминокислоты: остатки 24-34 (CDRL1), 50-56 (CDRL2) и 89-97 (CDRL3) в вариабельном домене легкой цепи, и остатки 31-35 или 31-35b (CDRH1), 50-65 (CDRH2) и 95-102 (CDRH3) в вариабельном домене тяжелой цепи; (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991). Таким образом, HV могут содержаться в пределах соответствующих CDR, и ссылки в данном описании на "гипервариабельные петли" VH и VL доменов следует интерпретировать как также включающие соответствующие CDR, и наоборот, если не указано иное.
Более высококонсервативные участки вариабельных доменов называются каркасными участками (FR), как определено ниже. Каждый из вариабельных доменов нативных тяжелой и легкой цепей содержит четыре FR (FR1, FR2, FR3 и FR4, соответственно), в большинстве случаев принимая β-листовую конфигурацию, связанную тремя гипервариабельными петлями. Гипервариабельные петли в каждой цепи удерживаются вместе в непосредственной близости при помощи FR и вместе с гипервариабельными петлями из другой цепи способствуют формированию антигенссвязывающих сайтов антител. Структурный анализ антител обнаружил взаимосвязь между последовательностью и формой сайта связывания, образованного определяющими комплементарность участками (Chothia et al., J. Mol. Biol. 227, 799-817, 1992; Tramontano et al., J. Mol. Biol, 215, 175-182, 1990). Несмотря на высокую вариабельность последовательности, пять из шести петель принимают только лишь небольшой набор конформаций основной цепи, называемый “каноническими структурами”. Эти конформации определяются, прежде всего, длиной петель и, во-вторых, присутствием ключевых остатков в определенных положениях в петлях и в каркасных участках, которые определяют конформацию по их упаковке, водородному связыванию или способности принимать необычные конформации основной цепи.
Использованный в описании термин "CDR" или "определяющая комплементарность область" означает не непрерывные (состоящие из несмежных участков) антигенсязывающие центры антитела, присутствующие в пределах вариабельного участка полипептидов как тяжелой, так и легкой цепи. Эти отдельные участки описаны в Kabat et al., J. Biol. Chem. 252, 6609-6616, 1977; Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991; Chothia et al., J. Mol. Biol. 196, 901-917, 1987 и MacCallum et al., J. Mol. Biol. 262, 732-745, 1996, в которых определения включают перекрывание или наборы аминокислотных остатков при сравнении между собой. Аминокислотные остатки, входящие в CDR согласно определению каждой из цитированных выше ссылок, предлагаются для сравнения. Предпочтительно, термин “CDR” является CDR согласно определению по Kabat, исходя из сравнения последовательностей.
Таблица 1: Определения CDR.
1Нумерация остатков придерживается номенклатуры Kabat et al., выше
2Нумерация остатков придерживается номенклатуры Chothia et al., выше
3Нумерация остатков придерживается номенклатуры MacCallum et al., выше
Использованный в описании термин “каркасный участок” или “FR участок” включает аминокислотные остатки, которые являются частью вариабельного участка, но не являются частью CDR (например, используя Kabat определения CDR). В связи с этим, вариабельный каркасный участок содержит примерно 100-120 аминокислот в длину, но включает только аминокислоты, не входящие в CDR. В виде конкретного примера вариабельного домена тяжелой цепи и CDR по определению Kabat et al., каркасный участок 1 соответствует домену вариабельного участка, включающего аминокислоты 1-30; каркасный участок 2 соответствует домену вариабельного участка, включающего аминокислоты 36-49; каркасный участок 3 соответствует домену вариабельного участка, включающего аминокислоты 66-94, и каркасный участок 4 соответствует домену вариабельного участка от аминокислоты 103 до конца вариабельного участка. Каркасные участки легкой цепи подобным образом отделяются вариабельными участками легкой цепи CDR. Аналогично, используя определение CDR по Chothia et al. или McCallum et al., границы каркасного участка отделяются соответствующими CDR концами, как описано выше. В предпочтительных вариантах осуществления CDR соответствуют определению согласно Kabat.
В антителах природного происхождения шесть CDR, присутствующих на каждом мономерном антителе, являются короткими, не непрерывными последовательностями аминокислот, которые располагаются особым образом, чтобы сформировать антигенсвязывающий сайт, в связи с тем, что антитело принимает свою трехмерную конфигурацию в водной окружающей среде. Остальные части вариабельных доменов тяжелой и легкой цепи показывают меньшую межмолекулярную вариабельность в аминокислотной последовательности и называются каркасными участками. Каркасные участки в большинстве случаев принимают β-листовую конформацию, и CDR образуют петли, которые соединяют, а в некоторых случаях образуют часть β-листовой структуры. Таким образом, эти каркасные участки работают на формирование каркаса, который обеспечивает расположение шести CDR в правильной ориентации с помощью межцепочечных нековалентных взаимодействий. Антигенсвязывающий сайт, образованный позиционированными CDR, определяет поверхностную комплементарность к эпитопу на иммунореактивном антигене. Эта комплементарная поверхность способствует нековалентному связыванию антитела с эпитопом иммунореактивного антигена. Положение CDR может быть легко установлено средним специалистом в данной области техники.
Использованный в описании термин “шарнирная область” включает часть тяжелой цепи молекулы, которая соединяет CH1 домен с CH2 доменом. Эта шарнирная область содержит приблизительно 25 остатков и является подвижной, таким образом, давая возможность двум N-концевым антигенсвязывающим участкам двигаться независимо. Шарнирные области могут подразделяться на три разных домена: верхний, средний и нижний шарнирные домены (Roux et al., J. Immunol. 161, 4083-4090, 1998). MET антитела, содержащие “полностью человеческую” шарнирную область, могут содержать одну из последовательностей шарнирной области, показанных в Таблице 2 ниже.
Таблица 2: Человеческие последовательности шарнирных областей.
(SEQ ID NO:227)
(SEQ ID NO:228)
(SEQ ID NO:229)
(SEQ ID NO:230)
(SEQ ID NO:231)
(SEQ ID NO:232)
(SEQ ID NO:233)
(SEQ ID NO:234)
(SEQ ID NO:235)
(SEQ ID NO:236)
(SEQ ID NO:237)
(SEQ ID NO:238)
Использованный в описании термин “CH2 домен” включает часть тяжелой цепи молекулы, которая продолжается, например, примерно от остатка 244 до остатка 360 антитела, при использовании обычного порядка нумерации (остатки от 244 до 360, Kabat система нумерации; и остатки 231-340, EU система нумерации; Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991). CH2 домен является уникальным в том отношении, что он не образует близкую пару с другим доменом. Скорее, две N-связанных разветвленных углеводных цепи помещаются между двумя CH2 доменами интактной нативной IgG молекулы. Также хорошо известно, что CH3 домен продолжается от CH2 домена до C-конца IgG молекулы и содержит приблизительно 108 остатков.
Использованный в описании термин “фрагмент” имеет отношение к части или участку антитела или цепи антитела, содержащей меньшее количество аминокислотных остатков, чем интактное или полное антитело или цепь антитела. Термин “антигенсвязывающий фрагмент” относится к фрагменту полипептида иммуноглобулина или антитела, который связывает антиген или конкурирует с интактным антителом (т.е., с интактным антителом, из которого они были получены) за связывание с антигеном (т.е., специфическое связывание с hMET и mMET). Использованный в описании термин “фрагмент” молекулы антитела включает антигенсвязывающие фрагменты антител, например, вариабельный домен легкой цепи (VL) антитела, вариабельный домен (VH) тяжелой цепи антитела, одноцепочечное антитело (scFv), F(ab’)2 фрагмент, Fab фрагмент, Fd фрагмент, Fv фрагмент и фрагмент одноцепочечного антитела (DAb). Фрагменты могут быть получены, например, посредством химической или ферментативной обработки интактного или полного антитела или цепи антитела или с помощью рекомбинантных способов.
Использованный в описании термин “валентность” имеет отношение к количеству возможных целевых участков связывания в полипептиде. Каждый целевой участок связывания специфически связывается с одной целевой молекулой или специфическим сайтом на целевой молекуле. Когда полипептид содержит более чем один целевой участок связывания, каждый целевой участок связывания может специфически связываться с теми же самыми или разными молекулами (например, может связываться с разными лигандами или разными антигенами, или разными эпитопами на том же самом антигене). Предметные связывающие молекулы имеют, по меньшей мере, один сайт связывания, специфический для hMET.
Использованный в описании термин “специфичность” имеет отношение к способности связываться (например, вступать в иммунную реакцию с) с данной мишенью, например, hMET, mMET. Полипептид может быть моноспецифическим и содержать один или более участков связывания, которые специфически связываются с мишенью, или полипептид может быть мультиспецифическим и содержать два или более участков связывания, которые специфически связываются с одной и той же или разными мишенями. В одном варианте осуществления антитело изобретения является специфическим более чем для одной мишени. Например, в одном варианте осуществления мультиспецифическая связывающая молекула изобретения связывается с hMET и второй целевой молекулой. В этом контексте вторая целевая молекула является молекулой, отличной от hMET или mMET.
Термин "эпитоп" имеет отношение к участку(ам) антигена (например, человеческого MET), который контактирует с антителом. Эпитопы могут быть линейными, т.е., вовлекающими в связывание одну последовательность аминокислот, или конформационными, т.е. вовлекающими в связывание две или более последовательностей аминокислот в разных участках антигена, которые необязательно могут не быть обязательно непрерывными. Антитела, предоставленные в этом документе, могут связываться с разными (перекрывающимися или неперекрывающимися) эпитопами во внеклеточном домене человеческого MET белка.
Использованный в описании термин “синтетический” в отношении полипептидов включает полипептиды, содержащие аминокислотную последовательность, не существующую в природе. Например, неприродные полипептиды, которые являются модифицированными формами природных полипептидов (например, содержащими мутацию, такую как вставка, замена или делеция), или которые содержат первую аминокислотную последовательность (которая может быть природной или неприродной), которая скрепляется в линейную последовательность аминокислот со второй аминокислотной последовательностью (которая может быть природной или неприродной), с которой она не соединяется в естественных природных условиях.
Использованный в описании термин “инженерный (сконструированный)” включает манипуляцию с молекулами нуклеиновой кислоты или полипептида с использованием синтетических методов (например, при помощи рекомбинантных методов, пептидного синтеза in vitro, при помощи ферментативного или химического соединения пептидов или комбинации этих методов). Предпочтительно антитела изобретения создаются инженерным путем, включая, например, гуманизированные и/или химерные антитела, и антитела, которые были созданы с целью улучшения одного или более свойств, таких как, связывание антигена, стабильность/время полужизни или эффекторная функция.
Использованный в описании термин “модифицированное антитело” включает синтетические формы антител, которые изменяются так, что они являются неприродными, например, антитела, которые содержат, по меньшей мере, два участка тяжелой цепи, но не две полные тяжелые цепи (такие как, антитела с удаленным доменом или минитела); мультиспецифические формы антител (например, биспецифические, триспецифические и т.д.) измененные для того, чтобы связываться с двумя или более различными антигенами или с разными эпитопами на одном антигене; молекулы тяжелых цепей, соединенные с scFv молекулами и тому подобное. ScFv молекулы известны в данной области техники и описаны, например, в патенте США 5,892,019. В дополнение к этому термин “модифицированное антитело” включает мультивалентные формы антител (например, трехвалентные, четырехвалентные и т.д. антитела, которые связываются с тремя или более копиями того же самого антигена). В другом варианте осуществления модифицированное антитело изобретения является гибридным белком, содержащим, по меньшей мере, один участок тяжелой цепи, утративший CH2 домен и содержащий домен связывания полипептида, содержащего участок связывания одного члена пары рецептор-лиганд.
Термин “модифицированное антитело” также может использоваться в данном описании, для того, чтобы сослаться на варианты аминокислотной последовательности MET антитела изобретения. Среднему специалисту в данной области техники должно быть понятно, что MET антитело изобретения может быть модифицировано с целью получения варианта MET антитела, который различается по аминокислотной последовательности по сравнению с MET антителом, из которого оно было получено. Например, могут быть сделаны нуклеотидные или аминокислотные замены, приводящие к консервативным заменам или изменениям в остатках "не относящихся к незаменимым" аминокислот (например, в CDR и/или каркасных участках). Аминокислотные замены могут включать замены одной или более аминокислот на природную или неприродную аминокислоту.
Использованный в описании термин “гуманизирующие замены” относится к аминокислотным заменам, в которых аминокислотный остаток, присутствующий в конкретном положении в VH или VL домене MET антитела изобретения (например, происходящее от верблюдовых MET антитело) заменяется аминокислотным остатком, который находится в эквивалентном положении в исходном человеческом VH или VL домене. Исходный человеческий VH или VL домен может быть VH или VL доменом, кодированным человеческой зародышевой линией. Гуманизирующие замены могут быть сделаны в каркасных участках и/или CDR MET антитела, определенного в данном документе.
Использованный в описании термин “гуманизированный вариант” относится к варианту антитела, который содержит одну или более “гуманизирующих замен” по сравнению с исходным MET антителом, в котором участок исходного антитела (например, VH домен и/или VL домен или их части, содержащие, по меньшей мере, один CDR) имеет аминокислотную последовательность, происходящую от видов, не являющихся человеком, и “гуманизирующие замены” находятся в пределах аминокислотной последовательности, происходящей от видов, не являющихся человеком.
Термин “модифицированный на уровне зародышевой линии вариант” при использовании в описании относится конкретно к “гуманизированным вариантам”, в которых “гуманизирующие замены” приводят к замене одного или более остатков аминокислот, присутствующих в конкретном положении (ях) в VH или VL домене MET антитела изобретения (например, происходящего от верблюдовых MET антитела), аминокислотным остатком, который находится в эквивалентном положении в исходном человеческом VH или VL домене, кодированном человеческой зародышевой линией. Характерно, что в отношении того или иного “модифицированного на уровне зародышевой линии варианта” замещающие аминокислотные остатки, замененные в модифицированном на уровне зародышевой линии варианте, берутся исключительно, или преимущественно, из одного и того же человеческого, кодированного в зародышевой линии, VH или VL домена. Термины “гуманизированный вариант” и “модифицированный на уровне зародышевой линии вариант” часто используются в описании взаимозаменяемым образом. Введение одной или более “гуманизирующих замен” в происходящем от верблюдовых (например, происходящем от ламы) VH или VL домене приводит к получению “гуманизированного варианта” VH или VL домена, происходящего от верблюдовых (ламы). Если замещенные аминокислотные остатки происходят преимущественно или исключительно из одной и той же человеческой кодированной в зародышевой линии последовательности VH или VL домена, тогда результатом может быть “человеческий модифицированный на уровне зародышевой линии вариант” VH или VL домена, происходящего от верблюдовых(ламы).
Использованный в описании термин “аффинный вариант” имеет отношение к варианту антитела, который демонстрирует одно или более изменений в аминокислотной последовательности по сравнению с исходным MET антителом изобретения, при этом аффинный вариант демонстрирует измененную аффинность в отношении hMET и/или mMET по сравнению с исходным антителом. Предпочтительно аффинный вариант будет демонстрировать улучшенную аффинность в отношении hMET и/или mMET, по сравнению с исходным MET антителом. Улучшение может быть видно по более низкому значению KD в отношении hMET и/или в отношении mMET, или более медленной скорости диссоциации в отношении hMET и/или в отношении mMET. Как правило, аффинные варианты демонстрируют одно или более изменений в аминокислотной последовательности в CDR по сравнению с исходным MET антителом. Такие замены могут приводить к замене исходной аминокислоты, присутствующей в данном положении в CDR, другим остатком аминокислоты, который может быть природным аминокислотным остатком или несуществующим в природе остатком аминокислоты. Аминокислотные замены могут быть консервативными или неконсервативными.
Использованный в описании термин антитела, имеющие “высокую гомологию с человеческими” имеет отношение к антителам, содержащим вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), которые, взятые вместе, демонстрируют, по меньшей мере, 90% идентичность аминокислотной последовательности с ближайшими соответствующими человеческими VH и VL последовательностями зародышевой линии. Антитела, имеющие высокую гомологию с человеческими, могут включать антитела, содержащие VH и VL домены нативных нечеловеческих антител, которые демонстрируют достаточно высокий % идентичности последовательности с последовательностями зародышевой линии человека, включая, например, антитела, содержащие VH и VL домены верблюжьих обычных антител, а также инженерные, в частности, гуманизированные или модифицированные на уровне зародышевой линии, варианты таких антител, а также “полностью человеческие” антитела.
В одном варианте осуществления VH домен антитела с высокой гомологией с человеческим может демонстрировать идентичность аминокислотной последовательности или гомологию последовательности 80% или больше с одним или более человеческими VH доменами в пределах каркасных участков FR1, FR2, FR3 и FR4. В других вариантах осуществления идентичность аминокислотной последовательности или гомология последовательности между VH доменом полипептида изобретения и ближайшей соответствующей последовательностью VH домена зародышевой линии человека может быть 85% или больше, 90% или больше, 95% или больше, 97% или больше, или вплоть до 99% или даже 100%.
В одном варианте осуществления VH домен антитела с высокой гомологией с человеческим может содержать одно или более (например, от 1 до 10) несовпадений в аминокислотной последовательности в пределах каркасных участков FR1, FR2, FR3 и FR4 по сравнению с ближайшей соответствующей человеческой VH последовательностью.
В другом варианте осуществления VL домен антитела с высокой гомологией с человеческим может демонстрировать идентичность последовательности или гомологию последовательности 80% или больше с одним или более человеческими VL доменами в пределах каркасных участков FR1, FR2, FR3 и FR4. В других вариантах осуществления идентичность аминокислотной последовательности или гомология последовательности между VL доменом полипептида изобретения и ближайшей соответствующей последовательностью VL домена зародышевой линии человека может быть 85% или больше 90% или больше, 95% или больше, 97% или больше, или вплоть до 99% или даже 100%.
В одном варианте осуществления VL домен антитела с высокой гомологией с человеческими может содержать одно или более (например, от 1 до 10) несовпадений в аминокислотной последовательности в пределах каркасных участков FR1, FR2, FR3 и FR4 по сравнению с ближайшей соответствующей человеческой VL последовательностью.
Перед анализом процента идентичности последовательности между антителом с высокой гомологией с человеческим и человеческим VH и VL зародышевой линии, может быть определена каноническая укладка, которая дает возможность установления семейства сегментов зародышевой линии человека с идентичной комбинацией канонических укладок для H1 и H2 или L1 и L2 (и L3). Затем выбирают члена семейства человеческой зародышевой линии, который имеет самую высокую степень гомологии последовательности с вариабельным участком представляющего интерес антитела, для подсчета гомологии последовательности. Методы определения ближайшей соответствующей человеческой зародышевой линии и определения % идентичности/гомологии последовательности хорошо известны кавилифицированным специалистам.
Антитела с высокой гомологией с человеческими могут содержать гипервариабельные петли или CDR, имеющие каноническую укладку человека или подобную человеческой каноническую укладку. В одном варианте осуществления, по меньшей мере, одна гипервариабельная петля или CDR или в VH домене или VL домене антитела с высокой гомологией с человеческим может быть получена или может происходить от VH или VL домена нечеловеческого антитела, например, обычного антитела от видов семейства Верблюжьих, при этом демонстрирует предсказанную или действительную каноническую укладку, которая в основном является идентичной с канонической укладкой, которая наблюдается в человеческих антителах. В одном варианте осуществления оба и H1 и H2 в VH домене антитела с высокой гомологией с человеческими демонстрируют предсказанную или действительную каноническую укладку, которая в основном является идентичной канонической укладке, имеющейся в человеческих антителах.
Антитела с высокой гомологией с человеческими могут содержать VH домен, в котором гипервариабельные петли H1 и H2 образуют комбинацию канонических укладок, которая является идентичной комбинации канонических структур, которые, как известно, встречаются, по меньшей мере, в одном человеческом VH домене зародышевой линии. Наблюдались случаи, что только некоторые комбинации канонических укладок H1 и H2 действительно наблюдаются в VH доменах, кодированных человеческой зародышевой линией. В одном варианте осуществления H1 и H2 в VH домене антитела с высокой гомологией с человеческими могут быть получены из VH домена видов, не относящихся к человеку, например, видов семества Верблюдовых, при этом образуют комбинацию предсказанных или действительных канонических укладок, которая является идентичной с комбинацией канонических укладок, которая, как известно, наблюдается в человеческой зародышевой линии или соматически мутированном VH домене. В неограничивающих вариантах осуществления H1 и H2 в VH домене антитела с высокой гомологией с человеческими могут быть получены из VH домена видов, не являющихся человеком, например, видов семества Верблюдовых, и образуют одну из следующих комбинаций канонических укладок: 1-1, 1-2, 1-3, 1-6, 1-4, 2-1, 3-1 и 3-5. Антитело с высокой гомологией с человеческим может содержать VH домен, который демонстрирует и высокую идентичность последовательности и гомологию последовательности с человеческой VH, и при этом содержит гипервариабельные петли, демонстрирующие структурную гомологию с человеческой VH.
Для канонических укладок, присутствующих в H1 и H2 в VH домене антитела с высокой гомологией с человеческим, и их комбинации предпочтительно быть "правильными" в отношении VH последовательности зародышевой линии человека, которая представляет собой лучшее совпадение с VH доменом антитела с высокой гомологией с человеческим, имея в виду общую идентичность первичной аминокислотной последовательности. В качестве примера, если лучшим совпадением последовательности является совпадение с VH3 доменом зародышевой линии человека, тогда предпочтительно для H1 и H2 может быть образовывать комбинацию канонических укладок, которая также наблюдается в природе в человеческом VH3 домене. Это может быть особенно важно в случае антител с высокой гомологией с человеческими, которые происходят от видов, не относящихся к человеку, например, антител, содержащих VH и VL домены, которые происходят от обычных верблюжьих антител, особенно антител, содержащих гуманизированные верблюжьи VH и VL домены.
Таким образом, в одном варианте осуществления VH домен MET антитела с высокой гомологией с человеческим может демонстрировать идентичность последовательности или гомологию последовательности 80% или больше, 85% или больше, 90% или больше, 95% или больше, 97% или больше, или до 99% или даже 100% с человеческим VH доменом в пределах каркасных участков FR1, FR2 , FR3 и FR4, и вместе с тем, H1 и H2 в том же самом антителе являются полученными из нечеловеческого VH домена (например, происходящего от видов семейства Верблюдовых, предпочтительно ламы), но образуют комбинацию предсказанных или действительных канонических укладок, которая является такой же как известная комбинация канонических укладок, встречающаяся в природе в том же самом человеческом VH домене.
В других вариантах осуществления каждый из L1 и L2 в VL домене антитела с высокой гомологией с человеческим получен из VL домена от видов, не относящихся к человеку (например, полученный от верблюдовых VL домен), при этом каждый демонстрирует предсказанную или действительную каноническую укладку, которая в основном является идентичной канонической укладке, которая наблюдается в человеческих антителах.
L1 и L2 в VL домене антитела с высокой гомологией с человеческим могут формировать комбинацию предсказанных или действительных канонических укладок, которая является идентичной комбинации канонических укладок, которая, как известно, наблюдается в VL домене зародышевой линии человека. В неограничивающих вариантах осуществления L1 и L2 в VLambda домене антитела с высокой гомологией с человеческим (например, антитела, содержащего происходящий от верблюдовых VL домен или его гуманизированный вариант) могут формировать одну из следующих комбинаций канонических укладок: 11-7, 13-7(A,B,C), 14-7(A,B), 12-11, 14-11 и 12-12 (как определено в Williams et al., J. Mol. Biol. 264, 220-232, 1996, и как показано на сайте http://www.bioc.uzh.ch/antibody/Sequences/Germlines/VBase_hVL.html). В неограничивающих вариантах осуществления L1 и L2 в Vkappa домене могут формировать одну из следующих комбинаций канонических укладок: 2-1, 3-1, 4-1 и 6-1 (как определено в Tomlinson et al., EMBO J. 14, 4628-4638, 1995 и как показано на сайте http://www.bioc.uzh.ch/antibody/Sequences/Germlines/VBase_hVK.html). В следующем варианте осуществления все три L1, L2 и L3 в VL домене антитела с высокой гомологией с человеческим могут демонстрировать по существу человеческую структуру. Предпочтительно, что VL домен антитела с высокой гомологией с человеческим демонстрирует как высокую идентичность последовательности, так и гомологию последовательности с человеческим VL, а также, что гипервариабельные петли в VL домене демонстрируют структурную гомологию с человеческим VL.
В одном варианте осуществления VL домен MET антитела с высокой гомологией с человеческим может демонстрировать идентичность последовательности 80% или больше, 85% или больше, 90% или больше, 95% или больше, 97% или больше, или вплоть до 99% или даже 100% с человеческим VL доменом в пределах каркасных участков FR1, FR2, FR3 и FR4, и кроме того, гипервариабельная петля L1 и гипервариабельная петля L2 могут формировать комбинацию предсказанных или действительных канонических укладок, которая является такой же, как известная комбинация канонических укладок, встречающаяся в природе в том же самом человеческом VL домене.
Безусловно, предусматривается, что VH домены, демонстрирующие высокую идентичность последовательности /гомологию последовательности с человеческим VH, а также структурную гомологию с гипервариабельными петлями человеческого VH будут объединяться с VL доменами, демонстрирующими высокую идентичность последовательности /гомологию последовательности с человеческим VL, а также структурную гомологию с гипервариабельными петлями человеческого VL, чтобы предоставить антитела с высокой гомологией с человеческими, содержащие VH/VL спаривания (например, спаривания VH/VL, происходящих от верблюдовых) с максимальной гомологией последовательностей и структурной гомологией с человеческими VH/VL спариваниями.
Методы оценки происходящих от семейства верблюдовых (например, происходящих от ламы) CDR, VH доменов или VL доменов в отношении присутствия подобных человеческим канонических укладок, описаны в WO 2010/001251 и WO 2011/080350, содержание которых полностью включается в описание путем отсылки.
Использованный в описании термин “аффинность” или “аффинность связывания” следует понимать, исходя из обычного значения в данной области техники в контексте связывания с антителом, термин отражает силу и/или устойчивость связывания между антигеном и участком связывания на антителе или его антигенсвязывающем фрагменте.
Анти-MET антитела, предоставленные в описании, характеризуются высокой аффинностью связывания с человеческим MET (hMET) и также высокой аффинностью связывания с мышиным MET (mMET). Аффинность связывания в отношении hMET и mMET может быть оценена с помощью стандартных методов, известных специалистам в данной области техники.
В одном варианте осуществления аффинность связывания Fab клона, содержащего определенное VH/VL спаривание, может быть оценена с использованием поверхностного плазмонного резонанса, например, с использованием системы Biacore™. Fab клоны, содержащие VH/VL спаривания антител и антигенсвязывающих фрагментов изобретения, как правило, демонстрируют скорость диссоциации hMET, измеренную с помощью Biacore™, в пределах от 1 x 10-3 до 1 x 10-2 s-1, необязательно от 1 x 10-3 до 6 x 10-3 s-1. Скорость диссоциации в этих пределах может быть принята в качестве критерия того, что Fab, и соответствующее двухвалентное mAb, демонстрирует высокую аффинность связывания с hMET. Аналогично, Fab клоны, содержащие VH/VL спаривания антител, и антигенсвязывающие фрагменты изобретения, как правило, демонстрируют скорость диссоциации mMET, измеренную с помощью Biacore™, как описано в прилагаемых примерах, в пределах от 1 x 10-3 до 1 x 10-2 s-1, необязательно от 1 x 10-3 до 6 x 10-3 s-1. Скорость диссоциации в пределах этого диапазона может быть принята, как критерий того, что Fab, и соответствующее двухвалентное mAb, демонстрирует высокую аффинность связывания с mMET. Следовательно, Fab, которые демонстрируют скорость диссоциации в отношении и человеческого и мышиного MET, попадающую в установленный диапазон, показывают высокую аффинность связывания в отношении hMET и высокую аффинность связывания в отношении mMET - то есть, Fab являются перекрестно-реагирующими между hMET и mMET. Двухвалентные mAb, содержащие два Fab, которые (в отдельности) показывают скорости диссоциации в отношении человеческого и мышиного MET в установленных пределах, также принимаются как демонстрирующие высокую аффинность связывания с человеческим MET и высокую аффинность связывания с мышиным MET.
Аффинность связывания также может быть выражена как константа диссоциации для конкретного антитела или KD. Меньшее значение KD, более сильное связывающее взаимодействие между антителом и его целевым антигеном. KD можно определить, например, путем объединения Kon и Koff скорости, определенной при помощи SPR- измерения. В большинстве случаев антитела и антигенсвязывающие фрагменты изобретения, при измерении как mAbs, демонстрируют KD для mMET и для hMET менее чем 0,1 нмоль/л.
Аффинность связывания с человеческим и мышиным MET также может быть оценена при использовании клеточной системы, как описано в сопутствующих примерах, в которых mAb тестируются в отношении связывания с клеточными линиями млекопитающих, которые экспрессируют MET, например, с помощью ELISA или проточной цитометрии. На высокую аффинность в отношении hMET или mMET может указывать, например, значение EC50 не более чем 0,5 нM при анализе ELISA, например, как описано в Примере 3.
Как суммируется выше, изобретение имеет отношение, по меньшей мере, отчасти, к антителам и их антигенсвязывающим фрагментам, которые связываются с hMET и mMET с высокой аффинностью. Свойства и характеристики MET антител и фрагментов антител согласно изобретению далее будут описываться более подробно.
Высокоаффинные hMET и mMET перекрестные антитела и антигенсвязывающие фрагменты, описанные в этом документе, являются MET агонистами. В контексте данного документа MET агонисты индуцируют (частично или полностью) MET-сигнальный путь, при связывании с MET рецептором. MET агонистические антитела и антигенсвязывающие фрагменты согласно изобретению являются агонистами hMET и mMET. Агонистическая активность в отношении связывания hMET или mMET антителами, описанными в этом документе, может определяться по молекулярным и/или клеточным ответам, которые (по меньшей мере, частично) имитируют молекулярные и клеточные ответы, индуцированные после гомологичного HGF-MET связывания (т.е. связывание hMET с человеческим HGF, связывание mMET с мышиным HGF). Антитела, стимулирующие такой ответ, также упоминаются в описании как “анти-MET-агонисты”, “агонистические антитела” и их грамматические варианты. Подобным образом, антитела частично или полностью стимулирующие такие ответы называются в описании “частичными MET агонистами” или “частичными агонистами” или “полными MET агонистами” или “полными агонистами”, соответственно. Подчеркивается, что антитела и антигенсвязывающие фрагменты изобретения индуцируют MET-сигнальный путь в обеих системах, и в человеческой и в мышиной системах, то есть, они являются агонистами hMET и mMET. Таким образом, следующее обсуждение применяется и к ответу, индуцированному связыванием hMET антителами и антигенсвязывающими фрагментами изобретения, и к ответу, индуцированному связыванием mMET антителами и антигенсвязывающими фрагментами изобретения.
MET агонизм у антител и антигенсвязывающих фрагментов изобретения может быть выявлен по молекулярным ответам, таким как фосфорилирование MET рецептора и/или клеточные ответы, например, ответам, которые обнаруживаются в анализе клеточного рассеивания, анти-апоптотическом анализе и/или анализе морфогенеза ветвления. Эти молекулярные и клеточные ответы описаны далее:
(i) Фосфорилирование MET рецептора. В этом контексте MET агонистическое антитело или антигенсвязывающий фрагмент фосфорилирует MET, когда связывание антитела или антигенсвязывающего фрагмента вызывает аутофосфорилирование MET при отсутствии рецептор-лигандного связывания - то есть, связывания антитела или антигенсвязывающего фрагмена с человеческим hMET приводит к фосфорилированию hMET в отсутствие hHGF, и связывание антитела или антигенсвязывающего фрагмента с mMET приводит к фосфорилированию mMET в отсутствие mHGF. Фосфорилирование MET может быть определено с помощью известных в данной области техники методов, например, вестерн-блоттинга или фосфо-MET ELISA (как описано в Примере 6 и в работе Basilico et al., J Clin Invest. 124, 3172-3186, 2014). Антитела и антигенсвязывающие фрагменты, описанные в этом документе, могут демонстрировать “высокую активность фосфорилирования” или “низкую активность фосфорилирования” в отношении hMET и могут демонстрировать “высокую активность фосфорилирования” или “низкую активность фосфорилирования” в отношении mMET. В этом контексте, антитело или антигенсвязывающий фрагмент демонстрирует “высокую активность фосфорилирования”, когда антитело или фрагмент демонстрирует активность в отношении mMET при значении EC50, сходном с HGF (<1нM) и/или EMAX, по меньшей мере, 80% (в виде процента от максимальной HGF-индуцированной активации) и активность в отношении hMET при значении EC50, сходном с HGF (<1нM) и/или EMAX, по меньшей мере, 80% (в виде процента от максимальной HGF-индуцированной активации). Антитело или антигенсвязывающий фрагмент демонстрирует “низкую активность фосфорилирования”, когда антитело демонстрирует активность в отношении mMET при значении EC50 1н-5нM и/или EMAX 60-80% (в виде процента от максимальной HGF-индуцированной активации) и демонстрирует активность в отношении hMET при значении EC50 1нM-5нM и/или EMAX 60-80% (в виде процента от максимальной HGF-индуцированной активации).
(ii) Индуцирование HGF-подобных клеточных ответов. MET агонизм может быть измерен с помощью таких методов, как анализ клеточного рассеивания, анти-апоптотический анализ и/или анализ морфогенеза ветвления, описанных в предоставленных Примерах. В этом контексте MET агонистические антитела или антигенсвязывающие фрагменты согласно изобретению индуцируют ответ в таких как эти клеточных анализах, которые сходны (по меньшей мере, частично) с ответом, наблюдаемым после воздействия гомологичным HGF. Например, MET агонист может быть определен по увеличению клеточного рассеивания в ответ на антитело по сравнению с клетками, подвергнутыми воздействию контрольного антитела (например, IgG1); по защитной активности против апоптоза, индуцированного лекарственным средством, со значением EC50 менее чем 32 нM и/или Emax клеточной жизнеспособности больше чем 20% по сравнению с необработанными клетками; и/или по увеличению числа ветвлений на сфероид в препаратах клеточных сфероидов, подвергнутых воздействию антитела или антигенсвязывающего фрагмента.
Антитела и антигенсвязывающие фрагменты, описанные в этом документе, могут “полностью индуцировать” или “частично индуцировать” HGF-подобные клеточные ответы при контактировании с человеческой клеткой и могут “полностью индуцировать” или “частично индуцировать” HGF-подобные клеточные ответы при контактировании с мышиной клеткой, в зависимости от используемого метода анализа.
В этом контексте “полная индукция” HGF-подобных клеточных ответов антителом или фрагментом может быть измерена:
в анализе клеточного рассеивания, антитело или антигенсвязывающий фрагмент вызывает увеличение клеточного рассеивания, по меньшей мере, эквивалентное 0,1 нM гомологичного HGF, когда концентрация антитела составляет 0,1-1 нM;
в анти-апоптотическом анализе, антитело или антигенсвязывающий фрагмент демонстрирует EC50 не больше чем 1,1x в отношении HGF и/или Emax жизнеспособность клеток больше чем 90% жизнеспособности, которая наблюдалась в отношении HGF; и/или
в анализе морфогенеза ветвления, клетки, обработанные антителом или антигенсвязывающим фрагментом, демонстрируют больше чем 90% от числа ветвлений на сфероид, индуцированных той же самой (ненулевой) концентрацией HGF.
В этом контексте, если антитело или антигенсвязывающий фрагмент не “полностью индуцирует” HGF-подобные клеточные ответы, как определено выше, “частичная индукция” HGF-подобных клеточных ответов может быть измерена:
в анализе клеточного рассеивания, антитело или антигенсвязывающий фрагмент индуцирует уровень клеточного рассеивания, составляющий, по меньшей мере, 25% от индуцированного 0,1 нM гомологичного HGF, когда концентрация антитела составляет 1 нM или ниже;
в анти-апоптотическом анализе, антитело или антигенсвязывающий фрагмент демонстрирует EC50 не больше чем 7,0x значение для HGF и/или Emax жизнеспособность клеток, по меньшей мере, 50% от наблюдаемой для HGF;
в анализе морфогенеза ветвления, клетки, обработанные антителом или антигенсвязывающим фрагментом, демонстрируют, по меньшей мере, 25% от числа ветвлений на сфероид, индуцированных той же самой (ненулевой) концентрацией HGF.
Как уже было описано, антитела и антигенсвязывающие фрагменты согласно изобретению являются hMET агонистами и mMET агонистами. Таким образом, в вариантах осуществления, где антитела вызывают (частичные или полные) HGF- подобные клеточные ответы, HGF-подобные клеточные ответы индуцируются (частично или полностью), когда антитело или антигенсвязывающий фрагмент контактирует с человеческой клеткой, и индуцируются (частично или полностью), когда антитело или антигенсвязывающий фрагмент контактирует с мышиной клеткой.
Картирование участка связывания (Пример 4) демонстрирует, что анти-MET антитела изобретения распознают эпитопы MET или в PSI домене MET или в SEMA домене MET. Следовательно, в некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения распознают эпитоп в PSI домене MET, предпочтительно человеческого MET. В некоторых альтернативных вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения распознают эпитоп SEMA домена MET, предпочтительно человеческого MET.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, распознающие эпитоп в SEMA домене, распознают эпитоп, расположенный на «лопасти» SEMA β-пропеллера. В некоторых вариантах осуществления эпитоп располагается на «лопасти» 4 или 5 SEMA β-пропеллера. В некоторых таких вариантах осуществления эпитоп располагается между аминокислотами 314-372 человеческого MET. В некоторых вариантах осуществления эпитоп располагается на «лопастях» 1-4 или 1-3 SEMA β-пропеллера. В некоторых вариантах осуществления эпитоп располагается между аминокислотами 27-313 человеческого MET, или между аминокислотами 27-225 человеческого MET.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, распознающие эпитоп в PSI домене MET, распознают эпитоп, расположенный между аминокислотами 516-545 MET, предпочтительно человеческого MET. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, распознающие эпитоп в PSI домене MET, распознают эпитоп, расположенный между аминокислотами 546-562 MET, предпочтительно человеческого MET.
В некоторых аспектах антитела, описанные в этом документе, распознают эпитопы во внеклеточном домене MET, который содержит один или более аминокислотных остатков, консервативных для человеческого и мышиного MET. В предпочтительных вариантах осуществления антитела, описанные в этом документе, распознают эпитопы во внеклеточном домене MET, который содержит один или более аминокислотных остатков, консервативных для человеческого MET, мышиного MET, крысиного MET и обезьяньего (например, циномолгус) MET.
В некоторых вариантах осуществления антитела изобретения содержат эпитоп человеческого MET, расположенного на участке от аминокислотного остатка 123 до остатка 223 человеческого MET. В некоторых вариантах осуществления антитела изобретения распознают эпитоп человеческого MET, расположенного на участке от аминокислотного остатка 224 до остатка 311 человеческого MET. В некоторых вариантах осуществления антитела изобретения распознают эпитоп человеческого MET, расположенного на участке от аминокислотного остатка 314 до остатка 372 человеческого MET. В некоторых вариантах осуществления антитела изобретения распознают эпитоп человеческого MET, расположенного на участке от аминокислотного остатка 546 до остатка 562 человеческого MET.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения распознают эпитоп человеческого MET, содержащий аминокислотный остаток lle367. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения распознают эпитоп человеческого MET, содержащий аминокислотный остаток Asp372 человеческого MET. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения распознают эпитоп человеческого MET, содержащий аминокислотные остатки lle367 и Asp372.
В некоторых таких вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения распознают эпитоп человеческого MET, расположенного на участке от аминокислотного остатка 314 до остатка 372 человеческого MET, при этом данный эпитоп содержит аминокислотный остаток lle367. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения распознают эпитоп человеческого MET, расположенного на участке от аминокислотного остатка 314 до остатка 372 человеческого MET, при этом эпитоп содержит аминокислотный остаток Asp371. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения распознают эпитоп человеческого MET, расположенного на участке от аминокислотного остатка 314 до остатка 372 человеческого MET, при этом эпитоп содержит аминокислотные остатки lle367 и Asp372.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения связываются с эпитопом человеческого MET, содержащим аминокислотный остаток Thr555 человеческого MET.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения распознают эпитоп человеческого MET, расположенного на участке от аминокислотного остатка 546 до остатка 562 человеческого MET, при этом эпитоп содержит аминокислотный остаток Thr555.
Следует принимать во внимание, что антитело или его антигенсвязывающий фрагмент может распознавать эпитоп, состоящий из некоторого количества аминокислотных остатков. Эпитоп может быть линейным, конформационным или комбинацией. Когда эпитоп определяется, как находящийся в определенном участке аминокислот, эпитоп может быть образован одной или более аминокислотами на том участке, который входит в контакт с антителом или фрагментом. Поэтому, следует иметь в виду, что в некоторых вариантах осуществления изобретения антитела или их фрагменты могут распознавать эпитоп, состоящий из нескольких аминокислотных остатков (последовательных или непоследовательных) в пределах точно определенного участка (например, от аминокислоты 314-372, или от 546 до 562), при условии, что распознаваемый эпитоп включает точно установленный остаток аминокислоты (например, lle367, Asp372, Thr555). Способы определения остатков, распознаваемых как часть эпитопа антитела, известны специалисту и включают, например, описанные в Примерах 4 и 26.
Поскольку анти-MET антитела и антигенсвязывающие фрагменты изобретения связываются с эпитопами, перекрывающимися с или близко от домена связывания, распознаваемого HGF, антитела и антигенсвязывающие фрагменты способны (по меньшей мере, частично) конкурировать с HGF за связывание гомологичного MET (т.е. конкурируют с человеческим HGF за hMET связывание и конкурируют с мышиным HGF за mMET связывание). Другими словами, антитела или антигенсвязывающие фрагменты напрямую или опосредованно предохраняют HGF от связывания с гомологичным MET в анализе связывания, например, ELISA, например, как описано в Примере 5. Следовательно, в некоторых вариантах осуществления MET антитела и антигенсвязывающие фрагменты изобретения конкурируют с мышиным и человеческим HGF за связывание с гомологичным MET. Антитело или антигенсвязывающий фрагмент, конкурирующий с HGF таким образом, также называется в описании “HGF конкурент”. Методы анализа для установления, конкурирует ли антитело или антигенсвязывающий фрагмент с HGF за связывание с MET, хорошо известны специалистам, - например, в конкурентном анализе ELISA HGF-конкурент может показывать значение IC50 не более чем 5нM и/или Imax (максимальный процент конкуренции при насыщении) по меньшей мере, 50%. Антитела и антигенсвязывающие фрагменты изобретения конкурируют с мышиным HGF за связывание с mMET и с человеческим HGF за связывание с hMET.
Антитело или антигенсвязывающий фрагмент изобретения может “полностью конкурировать” или “частично конкурировать” с HGF за связывание с гомологичным MET. В этом контексте “полным конкурентом” может быть антитело или антигенсвязывающий фрагмент, который в конкурентном анализе, например, ELISA, демонстрирует IC50 меньше чем 2 нM и/или Imax по меньшей мере, 90%. В некоторых вариантах осуществления “полный конкурент” демонстрирует IC50 меньше чем 1 нM и/или Imax больше чем 90%. “Частичным конкурентом” может быть антитело или антигенсвязывающий фрагмент, который в конкурентном анализе, например, ELISA, демонстрирует IC50 2-5 нM и/или Imax 50-90%. Данные значения применяются к конкуренции с мышиным HGF и человеческим HGF за связывание с гомологичным MET.
Как было описано ранее, антитела и антигенсвязывающие фрагменты изобретения представляют интерес, благодаря их способности распознавать как человеческий, так и мышиный MET. Антитела или их антигенсвязывающие фрагменты, описанные в этом документе, представляют особенный интерес в том случае, когда они демонстрируют равноценные свойства при связывании с mMET и с hMET. Эта равноценность дает возможность анализировать антитела в доклинических мышиных моделях болезней с ожиданием того, что антитела будут проявлять такие же или сходные свойства в отношении человека.
Следовательно, в некоторых вариантах осуществления антитела и связывающие фрагменты изобретения демонстрируют эквивалентную аффинность связывания в отношении hMET и mMET. В этом контексте “эквивалентная аффинность связывания” принимается за значение аффинности антитела или антигенсвязывающего фрагмента в отношении hMET и составляет 0.5-1.5 кратную аффинность этого антитела в отношении mMET. В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты изобретения демонстрируют аффинность в отношении hMET, 0,8-1,2-кратную аффинности этого антитела или антигенсвязывающего фрагмента в отношении mMET.
В качестве разъяснения и примера, антитела или антигенсвязывающие фрагменты, имеющие равноценную аффинность в отношении mMET и hMET, могут, при оценивании в виде Fab фрагмента, демонстрировать скорость диссоциации в отношении hMET, которая составляет 0,5-1,5х скорость диссоциации, демонстрируемую в отношении mMET. Например, антитело, имеющее равноценную аффинность в отношении mMET и hMET, которое демонстрирует скорость диссоциации 2,6 x10-3 s-1 в отношении mMET, может показывать скорость диссоциации в отношении hMET 1,3-3,9 x10-3 s-1. В качестве следующего примера, антитела или антигенсвязывающие фрагменты, имеющие равноценную аффинность в отношении mMET и hMET, могут демонстрировать EC50 в отношении hMET (определенную, например, методом ELISA или с помощью проточной цитометрии) 0,5-1,5х EC50 антитела или фрагмента в отношении mMET. Например, антитело, имеющее равноценную аффинность в отношении mMET и hMET, которое демонстрирует EC50 в отношении mMET 0,1 нмоль/л, может показывать EC50 в отношении hMET 0,05-0,15 нмоль/л.
В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты изобретения являются равноценными агонистами mMET и hMET. В этом контексте “равноценность (эквивалентность)” принимается как значение уровня MET агонизма, индуцированного после связывания hMET и составляет 0,5-1,5х уровень сигнального пути, индуцированного после связывания mMET. В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты изобретения индуцируют MET сигнальный путь после связывания hMET, составляющий 0,8-1,2х уровень сигнального пути, индуцированного после связывания mMET.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты изобретения являются равноценными mMET и hMET агонистами, при измерении, по меньшей мере, при помощи одного метода анализа MET агонизма, описанного в этом документе. Например, антитела или антигенсвязывающие фрагменты изобретения могут индуцировать эквивалентное фосфорилирование MET, демонстрировать равноценную защитную эффективность против апоптоза, вызванного лекарственным средством, и/или вызывать эквивалентные уровни ветвления в анализе морфогенеза ветвления. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты демонстрируют эквивалентный MET агонизм при измерении всеми описанными методами анализа.
С целью разъяснения, эквивалентное фосфорилирование MET антителом изобретения может быть обнаружено как EC50 для этого антитела в отношении hMET, будучи 0,5-1,5x EC50 в отношении mMET. Например, если EC50 в отношении mMET составляет 2,9 нM, то антитело будет равноценно индуцировать hMET фосфорилирование, если EC50 в отношении hMET находится в диапазоне 1,45-4,35 нM. Аналогично, равноценный MET агонизм, показанный в анти-апоптотическом методе анализа, может быть обнаружен как Emax в клетках человека, будучи 0,5-1,5x Emax в мышиных клетках. Например, если Emax в мышиных клетках был 37,5%, то антитело может быть эквивалентным hMET агонистом, если Emax для человеческих клеток находится в пределах 18,75-56,25%. Равноценный MET агонизм, показанный в анализе морфогенеза ветвления, может обнаруживаться как число ветвлений, наблюдаемых после воздействия антитела на человеческие клеточные сфероиды антитело, будучи 0,5-1,5x число ветвлений, наблюдаемых после воздействия на мышиные клеточные сфероиды той же самой (ненулевой) концентрации антитела. Например, если число ветвлений, демонстрируемое мышиными клетками после воздействия 0,5 нM антитела, было 14, то антитело может быть эквивалентным hMET агонистом, если число ветвлений, демонстрируемое человеческими клетками после воздействия 0,5 нM антитела, находится в пределах 7-21.
Аналогично, эквивалентный агонизм hMET и mMET может быть выявлен по равноценному клеточному рассеиванию. Особенность получения результатов при таком анализе означает применение 0,5-1,5 фактора нецелесообразным. При использовании анализа клеточного рассеивания эквивалентный агонизм hMET и mMET может быть выявлен путем подсчета в баллах клеточного рассеивания в отношении человеческих клеток, подвергнутых воздействию антитела, составляя при этом +/- 1 балл при подсчете в баллах клеточного рассеивания в отношении мышиных клеток, подвергнутых воздействию того же самого антитела в той же (ненулевой) концентрации. Например, если мышиные клетки, подвергнутые воздействию 0,33 nM антитела, показали оценку в баллах клеточного рассеивания 2, тогда антитело может быть равноценным агонистом hHGF, если человеческие клетки, подвергнутые воздействию 0,33 nM того же самого антитела, показали оценку в баллах клеточного рассеивания 1-3.
В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты изобретения демонстрируют равноценную HGF конкуренцию между mMET и hMET. В этом контексте “равноценная HGF конкуренция” принимается как значение уровня конкуренции, демонстрируемой антителом или антигенсвязывающим фрагментом, с человеческим HGF в отношении hMET и составляет 0,5-1,5 раза от уровня конкуренции, демонстрируемой антителом или антигенсвязывающим фрагментом, с мышиным HGF в отношении mMET. В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты изобретения демонстрируют уровень конкуренции с человеческим HGF, составляющий 0,8-1,2 раз от уровня конкуренции, демонстрируемой этим антителом или антигенсвязывающим фрагментом, с мышиным HGF в отношении mMET.
В качестве примера, равноценная конкуренция антитела с человеческим HGF и мышиным HGF может быть установлена как IC50 для этого антитела, конкурирующего со связыванием человеческий HGF-hMET, будучи 0,5-1,5х IC50 для этого антитела, конкурирующего со связыванием мышиный HGF-mMET. Например, если IC50 в отношении mHGF-mMET связывания составляет 0,34 nM, антитело конкурирует с hHGF и mHGF равноценно, если IC50 в отношении hHGF-hMET связывания находится в диапазоне 0,17-0,51 nM.
В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты изобретения являются перекрестно-реактивными с крысиным MET и/или MET макаки. Перекрестная реактивность с одним или двумя и крысиным и MET макаки имеет преимущество, заключающееся в том, что токсикологические исследования могут проводиться на модельной системе крысы и/или макаки. В этом отношении, проявляет или нет антитело перекрестную реактивность с крысиным MET или MET яванской макаки, можно определить с помощью ELISA, такого как описанный в сопроводительном Примере 25.
Антитела или их антигенсвязывающие фрагменты, описанные в этом документе, могут содержать, по меньшей мере, одну гипервариабельную петлю или определяющую комплементарность область, полученную из VH домена или VL домена видов из семейства Верблюдовых. В частности, антитело или антигенсвязывающий фрагмент может содержать VH и/или VL домены, или их CDRs, полученные путем активной иммунизации аутбредных верблюдовых, например, лам, с помощью человеческого MET антигена.
Под "гипервариабельной петлей или определяющей комплементарность областью, полученной из VH домена или VL домена видов семейства Верблюдовых" подразумевается, что гипервариабельная петля (HV) или CDR имеет аминокислотную последовательность, которая является идентичной или в основном идентичной с аминокислотной последовательностью гипервариабельной петли или CDR, которая кодируется геном иммуноглобулина Верблюдовых. В этом контексте "ген иммуноглобулина" включает гены зародышевой линии, гены иммуноглобулина, которые подвергались реаранжировке, и также соматически мутированные гены. Таким образом, аминокислотная последовательность HV или CDR, полученная из VH или VL домена видов семейства Верблюдовых, может быть идентична аминокислотной последовательности HV или CDR, присутствующих в зрелом обычном антителе Верблюдовых. Термин "полученный из" в этом контексте подразумевает структурное взаимоотношение, в том смысле, что HVs или CDRs MET антитела содержат аминокислотную последовательность (или ее минорные варианты), которая первоначально кодировалась геном иммуноглобулина Верблюдовых. Однако, это не предполагает в обязательном порядке особого отношения применительно к способу получения, используемого для получения MET антитела.
Происходящие от верблюдовых MET антитела могут быть получены от любых видов семейства Верблюдовых, включая в числе прочих, ламу, одногорбого верблюда, альпаку, викунью, гуанако или верблюда.
MET антитела, содержащие происходящие от верблюдовых VH и VL домены или их CDR, являются в большинстве случаев рекомбинантно экспрессированными полипептидами и могут быть химерными полипептидами. Термин "химерный полипептид" относится к искусственному (неприродному) полипептиду, который создается путем соединения двух или более пептидных фрагментов, которые в другом случае не могут находиться рядом. В это определение включаются "виды" химерных полипептидов, созданные соединением пептидных фрагментов, кодированных двумя или более видами животных, например, верблюдовым и человеком.
Происходящие от верблюдовых CDR могут содержать одну из CDR последовательностей, показанных в Таблицах 3 и 4 ниже.
В одном варианте осуществления полный VH домен и/или полный VL домен может быть получен из видов семейства Верблюдовых. В специфических вариантах осуществления происходящий от верблюдовых VH домен может содержать аминокислотную последовательность, показанную в виде SEQ ID NOs:155, 157, 159, 161, 163, 165, 167, 169, 171, 173 или 175, тогда как происходящий от верблюдовых VL домен может содержать аминокислотную последовательность, показанную в виде SEQ ID NOs: 156, 158, 160, 162, 164, 166, 168, 170, 172, 174 или 176. Происходящий от верблюдовых VH домен и/или происходящий от верблюдовых VL домен затем может подвергаться белковой инженерии, при которой одна или более аминокислотных замен, вставок или делеций вводится в аминокислотную последовательность Верблюдовых. Эти инженерные изменения предпочтительно включают аминокислотные замены, относящиеся к последовательности Верблюдовых. Такие изменения включают "гуманизацию" или "модифицирование на уровне зародышевой линии", при этом один или более аминокислотных остатков в кодируемом верблюдовым VH или VL домене заменяются эквивалентными остатками из гомологичных кодируемых человеком VH или VL доменов.
Изолированные верблюдовые VH и VL домены, полученные с помощью активной иммунизации животного из семейства Верблюдовых (например, ламы) человеческим MET антигеном, могут использоваться в качестве основы для создания MET антител согласно изобретению. Начиная с интактных VH и VL доменов Верблюдовых, можно инженерным путем создать одну или более аминокислотных замен, вставок или делеций, которые отклоняются от исходной последовательности верблюда. В некоторых вариантах осуществления такие замены, вставки или делеции могут присутствовать в каркасных участках VH домена и/или VL домена. Целью таких изменений в первичной аминокислотной последовательности может быть уменьшение возможных нежелательных свойств (например, иммуногенности у человека-хозяина (так называемая гуманизация), участков, возможно продуцирующих гетерогенность и/или нестабильность (гликозилирования, дезаминирования, изомеризации и т.д.) или улучшение некоторых других желательных свойств молекулы (например, растворимости, стабильности, биодоступности и т.д.). В других вариантах осуществления изменения в первичной аминокислотной последовательности могут быть выполнены в одной или более из гипервариабельных петель (или CDR) Верблюдового VH и/или VL домена, полученного с помощью активной иммунизации. Такие изменения могут быть введены для того, чтобы увеличить антигенсвязывающую аффинность и/или специфичность или уменьшить возможные нежелательные свойства, например, иммуногенность у человека-хозяина (так называемая гуманизация), участки, возможно продуцирующие гетерогенность и/или нестабильность, гликозилирование, дезаминирование, изомеризацию и т.д., или усилить некоторое другое благоприятное свойство молекулы, например, растворимость, стабильность, биодоступность и т.д.
Таким образом, в одном варианте осуществления изобретение предоставляет вариант MET антитела, который содержит, по меньшей мере, одну аминокислотную замену, по меньшей мере, в одном каркасном или CDR участке или VH домена или VL домена по сравнению с происходящим от верблюдового VH или VL доменом, примеры которого включают, но не ограничиваются этим, VH домены Верблюдовых, содержащие аминокислотные последовательности, показанные как SEQ ID NOs:155, 157, 159, 161, 163, 165, 167, 169, 171, 173 или 175, и VL домены Верблюдовых, содержащие аминокислотные последовательности, показанные как SEQ ID NO: 156, 158, 160, 162, 164, 166, 168, 170, 172, 174 или 176.
В некоторых вариантах осуществления предоставляются молекулы "химерных" антител, содержащие происходящие от Верблюдовых VH и VL домены (или их варианты, созданные инженерным путем) и один или более константных доменов от антител не Верблюдовых, например кодированных человеком константных доменов (или их вариантов, созданных инженерным путем). В таких вариантах осуществления является предпочтительным, что оба и VH домен и VL домен получены от одного и того же вида семейства Верблюдовых, например и VH и VL могут происходить от ламы (до введения инженерного изменения аминокислотной последовательности). В таких вариантах осуществления оба домена и VH и VL домен могут происходить от одного животного, конкретнее от одного животного, которое было активно иммунизировано человеческим MET антигеном.
Изобретение может в некоторых вариантах осуществления включать химерные верблюжьи/человеческие антитела, и в частности, химерные анттела, в которых VH и VL домены являются полностью последовательностями Верблюдового (например, ламы или альпаки) и остальная часть антитела является полностью человеческой последовательностью. MET антитела могут включать антитела, содержащие "гуманизированные" или "модифицированные на уровне зародышевой линии" варианты происходящих от Верблюдовых VH и VL доменов, или их CDR, и верблюдовые/человеческие химерные антитела, в которых VH и VL домены содержат одну или более аминокислотных замен в каркасных участках по сравнению с VH и VL доменами Верблюдовых, полученными путем активной иммунизации верблюда с помощью человеческого MET антигена. Такая "гуманизация" увеличивает % идентичности последовательности с VH или VL доменами человеческой зародышевой линии путем замены несоответствующих аминокислотных остатков в исходном VH или VL домене Верблюдовых на эквивалентный остаток, обнаруженный в кодированном зародышевой линией человека VH или VL домене.
Изобретение может в некоторых вариантах осуществления рассматривать химерные верблюжьи/мышиные антитела, и в частности химерные антитела, в которых VH и VL домены являются полностью последовательностью Верблюдовых (например, ламы или альпаки) и остальная часть антитела является полностью мышиной последовательностью.
MET антитела и антигенсвязывающие фрагменты изобретения также могут быть CDR-«привитыми» антителами, в которых CDR (или гипервариабельные петли), происходящие от антитела Верблюдовых, например MET антитела Верблюдовых, полученные путем активной иммунизации с использованием человеческого MET белка, или иначе кодированные геном верблюдовых, «пересажены» в человеческий VH и VL каркас, при этом остальная часть антитела также является полностью человеческого происхождения. Такие CDR-«привитые» MET антитела могут содержать CDR, имеющие аминокислотные последовательности, показанные в Таблицах 3 и 4 ниже.
Полученные от верблюдовых MET антитела включают варианты, в которых гипервариабельная петля(и) или CDR(s) VH домена и/или VL домена получены из обычного верблюжьего антитела, индуцированного против человеческого MET, но в котором, по меньшей мере, одна из указанных (полученных от верблюдовых) гипервариабельных петель или CDRs может быть создана инженерным путем, чтобы включать одну или более аминокислотных замен, вставок или делеций относительно последовательности, кодированной верблюдовым. Такие изменения включают “гуманизацию” гипервариабельных петель/CDR. Полученные от верблюдовых HVs/CDRs, которые могут быть созданы таким способом, могут все еще демонстрировать аминокислотную последовательность, которая является "практически идентичной" с аминокислотной последовательностью, кодированной верблюдовыми HV/CDR. В этом контексте "значительная степень идентичности" может допустить не более чем одно или не более чем два несоответствия аминокислотной последовательности с кодированной верблюдовыми HV/CDR. Конкретные варианты осуществления MET антитела могут содержать гуманизированные варианты CDR последовательностей, показанных в Таблицах 3 и 4.
Обычные антитела Вербюдовых (например, ламы) обеспечивают благоприятную отправную точку для получения антител, полезных для человека в качестве терапевтических средств, благодаря следующим факторам, обсужденным в США 12/497,239, который включается в описание путем отсылки:
1) Высокий % гомологии последовательности между VH и VL доменами Верблюдовых и их человеческими эквивалентами;
2) Высокая степень структурной гомологии между CDR VH и VL доменов Верблюдовых и их человеческими эквивалентами (т.е. подобными человеческим каноническими укладками и комбинациями подобных человеческим канонических укладок).
Платформа на основе Верблюдовых (например, ламы) также предоставляет значительное преимущество в отношении функционального разнообразия MET антител, которые могут быть получены.
Функциональность MET антител, содержащих VH и/или VL домены Верблюдовых, для терапии человека может быть дополнительно усовершенствована путем “гуманизации” природных VH и VL доменов Верблюдовых, например, для придания им меньшей иммуногенности у человека-хозяина. Конечной целью гуманизации является получение молекулы, в которой VH и VL домены демонстрируют минимальную иммуногенность при введении человеку, при этом сохраняя специфичность и аффинность антигенсвязывающего участка, образованного исходными VH и VL доменами.
Один подход к гуманизации, так называемое “модифицирование на уровне зародышевой линии”, включает инженерные изменения в аминокислотной последовательности camelid VH или VL домена Верблюдовых, чтобы «привести» их ближе к последовательности зародышевой линии человеческого VH или VL домена.
Определение гомологии между VH (или VL) доменом Верблюдовых и человеческим VH (или VL) доменом является решающей стадией в процессе гуманизации, как для выбора аминокислотных остатков Верблюдовых, которые должны быть заменены (в данном VH или VL домене), так и для выбора соответствующего заменяющего аминокислотного остатка(ов).
Подход к модифицированию на уровне зародышевой линии обычных антител Верблюдовых был разработан на основе выравнивания большого числа новых последовательностей VH (и VL) домена Верблюдовых, в большинстве случаев соматически мутированных VH (или VL) доменов, которые как известно связываются с целевым антигеном, с человеческими VH (или VL) последовательностями зародышевой линии, человеческими VH (и VL) консенсусными последовательностями, а также на основе информации о последовательности зародышевой линии, доступной в отношении llama pacos (одомашненной ламы).
Этот метод, описанный в WO 2011/080350, содержание которого включается в описание путем отсылки, может применяться для (i) выбора “верблюжьих” аминокислотных остатков для замены в полученном от верблюдовых VH или VL домене или их CDR, и (ii) выбора заменяющего “человеческого” аминокислотного остатка, чтобы подставить его при гуманизации любого данного VH (или VL) домена Верблюдовых. Этот подход может использоваться для получения гуманизированных вариантов происходящих от верблюдовых CDR, имеющих аминокислотные последовательности, показанные в Таблицах 3 и 4, и также для модифицирования на уровне зародышевой линии происходящих от верблюдовых VH и VL доменов, имеющих последовательности, показанные в Таблице 5.
MET антитела могут иметь разнообразные варианты осуществления, в которых присутствуют и VH домен и VL домен. Термин "антитело" в этом документе используется в самом широком смысле и включает, но не ограничивается этим, моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела), при условии, что они демонстрируют подходящую иммунологическую специфичность в отношении человеческого MET белка и мышиного MET белка. Термин "моноклональное антитело", использованный в описании, имеет отношение к антителу, полученному из популяции практичнески однородных антител, т.е., отдельные антитела, составляющие популяцию, являются одинаковыми, за исключением возможных природных мутаций, которые могут присутствовать в минимальных количествах. Моноклональные антитела являются высокоспецифическими, будучи направленными против одной антигенной детерминанты. Более того, в противоположность обычным (поликлональным) препаратам антител, которые в большинстве случаев включают различные антитела, направленные против разных детерминант (эпитопов) на антигене, каждое моноклональное антитело направлено против одной детерминанты или эпитопа на антигене.
"Фрагменты антитела" содержат часть полноразмерного антитела, в большинстве случаев его антигенсвязывающий или вариабельный домен. Примеры фрагментов антитела включают Fab, Fab', F(ab')2, биспецифические Fab's и Fv фрагменты, диатела, линейные антитела, молекулы одноцепочечных антител, одноцепочечный вариабельный фрагмент (scFv) и мультиспецифические антитела, образованные из фрагментов антител (смотри Holliger и Hudson, Nature Biotechnol. 23:1126-1136, 2005, содержание которой включается в описание путем отсылки).
В неограничивающих вариантах осуществления MET антитела, предоставленные в данном документе, могут содержать CH1 домены и/или CL домены, аминокислотная последовательность которых является полностью или в основном человеческой. В том случае, если MET антитело предназначается для терапевтического применения у человека, тогда для полного константного участка антитела или, по меньшей мере, его части, характерно иметь полностью или в оновном человеческую аминокислотную последовательность. Следовательно, одна или более или любая комбинация CH1 домена, шарнирного участка, CH2 домена, CH3 домена и CL домена (и CH4 домена, если он присутствует) может быть полностью или в основном человеческой, что касается аминокислотной последовательности. Такие антитела могут быть антителами любого человеческого изотипа, например IgG1.
Предпочтительно, CH1 домен, шарнирный участок, CH2 домен, CH3 домен и CL домен (и CH4 домен, если он присутствует) могут иметь полностью или в основном человеческую аминокислотную последовательность. Что касается константного участка гуманизированного или химерного антитела, или фрагмента антитела, термин "в основном человеческая" относится к аминокислотной последовательности идентичной, по меньшей мере, на 90%, или по меньшей мере, на 92%, или по меньшей мере, на 95%, или по меньшей мере, на 97%, или по меньшей мере, на 99% человеческому константному участку. Термин “человеческая аминокислотная последовательность” в этом контексте относится к аминокислотной последовательности, которая кодируется геном иммуноглобулина человека, включая зародышевые, реаранжированные и соматически мутированные гены. Такие антитела могут быть любого человеческого изотипа, при этом человеческий IgG4 и IgG1 являются особенно предпочтительными.
Также предоставляются MET антитела, содержащие константные домены “человеческой” последовательности, которая изменена путем одной или более аминокислотных вставок, делеций или замен что касается человеческой последовательности, заисключением тех вариантов осуществления, где присутствие “полностью человеческого” шарнирного участка определенно является необходимым.
Присутствие “полностью человеческой” шарнирной области в MET антителах изобретения может быть полезно и для того, чтобы свести к минимуму иммуногенность и чтобы оптимизировать стабильность антитела.
MET антитела, предоставленные в данном описании, могут быть антителами любого изотипа. Антитела, предназначенные для применения в терапевтических целях у человека, в большинстве случаев будут антителами IgA, IgD, IgE IgG, IgM типа, часто IgG типа, в таком случае они могут принадлежать к любому из четырех подклассов IgG1, IgG2a и b, IgG3 или IgG4. В пределах каждого из этих подклассов допускается введение одной или более аминокислотных замен, вставок или делеций в пределах Fc участка, или введение других структурных модификаций, например чтобы увеличить или уменьшить Fc-зависимую функциональные возможности.
В неограничивающих вариантах осуществления следует иметь в виду, что одна или более аминокислотных замен, вставок или делеций могут быть сделаны в пределах константного участка тяжелой и/или легкой цепи, в частности, в пределах Fc участка. Результатом аминокислотных замен может быть замена одной аминокислоты другой аминокислотой, природной или неприродной или модифицированной аминокислотой. Также разрешаются другие структурные модификации, такие как, например, изменения в профиле гликозилирования (например, путем добавления или делеции N- или O-связанных сайтов гликозилирования). В зависимости от предполагаемого использования MET антитела, может быть желательно, модифицировать антитело изобретения в отношении его свойств связывания с Fc рецепторами, например, чтобы модулировать эффекторную функцию.
В некоторых вариантах осуществления MET антитела могут содержать Fc участок данного изотипа антитела, например, человеческого IgG1, который модифицируется для того, чтобы уменьшить или практически устранить одну или более эффекторных функций антитела, в естественных условиях связанных с этим изотипом антитела. В неограничивающих вариантах осуществления MET антитело может быть практически лишено любых эффекторных функций антитела. В этом контексте “эффекторные функции антитела” включают одну или более или все из числа антителозависимой клеточной цитотоксичности (ADCC), комплемензависимой цитотоксичности (CDC) и антителозависимого клеточного фагоцитоза (ADCP).
Аминокислотная последовательность Fc участка MET антитела может содержать одну или более мутаций, таких как аминокислотные замены, делеции или вставки, которые оказывают воздействие на снижение одной или более эффекторных функций антитела (по сравнению с эквивалентным антителом дикого типа, не имеющим указанной мутации). Некоторые такие мутации известны в области инженерии антител. Неограничивающие примеры, подходящие для включения в MET антитела, описанные в этом документе, включают следующие мутации в Fc домене человеческого IgG4 или человеческого IgG1: N297A, N297Q, LALA (L234A, L235A), AAA (L234A, L235A, G237A) или D265A (нумерация аминокислотных остатков согласно EU системе нумерации в человеческом IgG1).
Моноклональные антитела или их антигенсвязывающие фрагменты, которые "перекрестно-конкурируют" с MET антителами, раскрытыми в этом описании, - это такие антитела, которые связываются с человеческими MET в сайте(ах), который является идентичным или перекрывающимся с сайтом(ами), через который связываются предлагаемые MET антитела, и связываются с мышиным MET в сайте(ах), который является идентичным или перекрывающимся с сайтом(ами), через который связываются предлагаемые MET антитела. Конкурирующие моноклональные антитела или их антигенсвязывающие фрагменты можно идентифицировать, например, посредством конкурентного анализа антител. Например, образец очищенного или частично очищенного человеческого MET может быть связан с твердой подложкой. Затем, добавляют молекулу антитела или его антигенсвязывающий фрагмент настоящего изобретения и моноклональное антитело или его антигенсвязывающий фрагмент, предположительно способный конкурировать с такой молекулой антитела изобретения. Одна из двух молекул при этом является меченой. Если меченая молекула и немеченая молекула связывается с отдельными и разобщенными сайтами на MET, меченая молекула будет связываться на таком же уровне в любом случае, присутствует или нет предполагаемая конкурирующая молекула. Однако, если сайты, с которыми происходит взаимодействие, являются идентичными или перекрывающимися, немеченая молекула будет конкурировать, и количество меченой молекулы, связанной с антигеном, будет уменьшаться. Если немеченая молекула присутствует в избытке, очень небольшое количество, если вообще будет, меченой молекулы будет связываться. В контексте настоящего изобретения конкурирующие моноклональные антитела или их антигенсвязывающие фрагменты являются такими, которые уменьшают связывание предлагаемого антитела с MET примерно на 50%, примерно на 60%, примерно на 70%, примерно на 80%, примерно на 85%, примерно на 90%, примерно на 95% или примерно на 99%. Подробности методик проведения таких конкурентных анализов хорошо известны в данной области техники, их можно найти, например, в Harlow и Lane, Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1988, 567-569, 1988, ISBN 0-87969-314-2. При использовании очищенных антител такие анализы могут быть оценены количественно. Стандартную кривую получают путем титрования одного антитела в сравнении с самим собой, т.е., одно и то же антитело используется для мечения и в качестве конкурента. Способность немеченого конкурирующего моноклонального антитела или его антигенсвязывающего фрагмента ингибировать связывание меченой молекулы с чашкой определяется титрованием. Результаты наносятся на график, и сравниваются концентрации, необходимые для достижения желательной степени ингибирования связывания.
Изобретение также предоставляет молекулы полинуклеотида, кодирующие MET антитела изобретения, также векторы экспрессии, содержащие нуклеотидные последовательности, которые кодируют MET антитела изобретения, функционально связанные с регуляторными последовательностями, которые обеспечивают экспрессию антигенсвязывающего полипептида в клетке-хозяине или бесклеточной системе, и клетку-хозяина и бесклеточную систему, содержащую этот вектор экспресии.
Молекулы полинуклеотидов, кодирующие MET антитела изобретения включают, например, молекулы рекомбинантных ДНК. Термины "нуклеиновая кислота", “полинуклеотид” или "молекула полинуклеотида" используются в описании взаимозаменяемым образом и относятся к любой молекуле ДНК или РНК, или одноцепочечной или двухцепочечной и, в случае одноцепочечной молекулы к молекуле его комплементарной последовательности. При обсуждении молекул нуклеиновой кислоты последовательность или структура отдельной молекулы нуклеиновой кислоты может описываться в данном документе согласно обычному правилу предоставления последовательности в направлении от 5' к 3'. В некоторых вариантах осуществления изобретения нуклеиновые кислоты или полинуклеотиды являются "изолированными”. Этот термин при использовании применительно к молекуле нуклеиновой кислоты, относится к молекуле нуклеиновой кислоты, которая отделяется от последовательностей, с которыми она непосредственно соприкасается в природном геноме организма, из которого она происходит. Например, "изолированная нуклеиновая кислота" может включать молекулу ДНК, вставленную в вектор, такой как плазмида или вирусный вектор, или интегрированную в геномную ДНК прокариотической или эукариотической клетки или организма-хозяина, не являющегося человеком. При применении к РНК термин "изолированный полинуклеотид" относится в первую очередь к молекуле РНК, кодированной изолированной молекулой ДНК, как определено выше. Альтернативно, термин может иметь отношение к молекуле РНК, которая может быть очищена/отделена от других нуклеиновых кислот, с которыми она может быть соединена в своем естественном состоянии (т.е. в клетках или тканях). Изолированный полинуклеотид (или ДНК или РНК), кроме того, может представлять молекулу, полученную непосредственно при помощи биологических или синтетических способов и отделенную от других компонентов, присутствующих в процессе ее продуцирования.
Для рекомбинантного получения MET антитела согласно изобретению, может быть получен рекомбинантный полинуклеотид, кодирующий его, (с помощью стандартных методов молекулярной биологии) и вставлен в реплицируемый вектор для экспрессии в выбранной клетке-хозяине или бесклеточной системе экспрессии. Подходящие клетки-хозяева могут быть прокариотическими, дрожжевыми клетками или клетками высших эукариот, в частности, клетками млекопитающего. Примерами подходящих линий клеток-хозяев млекопитающих являются линия клеток почки обезьяны CV1, трансформированных SV40 (COS-7, ATCC CRL 1651); линия клеток почки эмбриона человека (293 или 293 клетки, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen. Virol. 36:59-74, 1977); клетки почки новорожденного хомяка (BHK, ATCC CCL 10); клетки яичника китайского хомяка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216, 1980); мышиные клетки Сертоли (TM4; Mather, Biol. Reprod. 23:243-252, 1980); мышиные миеломные клетки SP2/0-AG14 (ATCC CRL 1581; ATCC CRL 8287) или NS0 (HPA коллекция культур № 85110503); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы линии Buffalo (BRL 3A, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); TRI клетки (Mather et al., Annals N.Y. Acad. Sci. 383:44-68, 1982); MRC 5 клетки; FS4 клетки; и линия клеток гепатомы человека (Hep G2), а также DSM’s PERC-6 клеточная линия. Векторы экспресии, подходящие для использования в каждой из этих клеток-хозяев, также в основном известны в данной области техники.
Следует отметить, что термин "клетка-хозяин" в большинстве случаев относится к культивированным клеточным линиям. Человеческий организми «целиком», которому был введен вектор экспрессии, кодирующий антигенсвязывающий полипептид согласно изобретению, недвусмысленно исключается из определения "клетки-хозяина".
В одном важном аспекте изобретение также предоставляет способ получения MET антитела изобретения, который включает культивирование клетки-хозяина (или бесклеточной системы), содержащей полинуклеотид (например, вектор экспрессии), кодирующий MET антитело при условиях, обеспечивающих экспрессию MET антитела, и извлечение экспрессированного MET антитела. Процесс рекомбинантной экспрессии может использоваться для производства в больших масштабах MET антител согласно изобретению, включая моноклональные антитела, предназначенные для терапевтического применения при лечении человека. Подходящие векторы, клеточные линии и технологические процессы для производства в больших масштабах рекомбинантных антител, пригодных для терапевтического применения in vivo, являются общедоступными в данной области техники и хорошо известны специалистам.
MET антитела, предоставленные в этом документе, полезны при проведении терапии, в частности, терапевтическом лечении болезни, особенно состояний, при которых польза извлекается в результате стимулировании MET функции, включая, но не ограничиваясь этим, дегенеративные болезни, воспалительные болезни, аутоиммунные болезни, метаболические болезни, нарушения, связанные с трансплантацией, и заживление ран. В этом отношении MET антитела, предоставленные в этом документе, являются примерами обширного класса MET агонистов, например HGF, обладающих терапевтической пригодностью при лечении указанных состояний.
Гепатоциты экспрессируют MET и являются основной мишенью HGF, который способствует их пролиферации и защищает их от апоптоза. MET антитела, индуцирующие MET сигнальный путь и показанные в этом документе (Примеры 16 и 17), защищают гепатоциты на мышиных моделях поражения печени, как острого, так и хронического поражения печени. Как уже было сказано, антитела изобретения демонстрируют равноценные свойства, как в человеческой системе, так и в мышиной системе и таким образом, можно ожидать, что они обеспечат подобные защитные эффекты в отношении поражения печени человека. Следовательно, в одном аспекте изобретение предоставляет способ лечения или предотвращения поражения печени у пациента-человека, который включает введение пациенту, нуждающемуся в этом, терапевтически эффективного количества MET антитела, индуцирующего MET сигнальный путь. В некоторых вариантах осуществления способ является способом лечения или предотвращения острого поражения печени. В некоторых вариантах осуществления данный способ является способом лечения или предотвращения хронического поражения печени. В некоторых вариантах осуществления антитело является антителом, описанным в данном документе.
Эпителиальные клетки почки экспрессируют значительные уровни MET и являются чувствительными к HGF стимуляции. MET антитела, индуцирующие MET сигнальный путь и показанные в этом документе (Пример 18), обеспечивают защиту на мышиной модели от острого поражения почек. Как уже было сказано, антитела изобретения демонстрируют равноценные свойства, как в человеческой системе, так и в мышиной системе и таким образом, можно ожидать, что они обеспечат подобные защитные эффекты в отношении поражения почек человека. Следовательно, в одном аспекте изобретение предоставляет способ лечения или предотвращения поражения почек у пациента-человека, который включает введение пациенту, нуждающемуся в этом, терапевтически эффективного количества MET антитела, индуцирующего MET сигнальный путь. В некоторых вариантах осуществления данный способ является способом лечения или предотвращения острого поражения почек. В некоторых вариантах осуществления антитело представляет собой антитело, описанное в этом документе.
Кроме того, было показано (Примеры 19 и 20), что введение MET антител, индуцирующих MET сигнальный путь, обеспечивает эффективное лечение на мышиных моделях воспалительного заболевания кишечника (IBD), например, язвенного колита. Следовательно, в одном аспекте изобретение предоставляет способ лечения или предотвращения IBD у пациента-человека, который включает введение нуждающемуся в этом пациенту терапевтичеки эффективного количества MET антитела, индуцирующего MET сигнальный путь. В некоторых вариантах осуществления данный способ является способом лечения или предотвращения язвенного колита. В некоторых вариантах осуществления антитело представляет собой антитело, описанное в этом документе.
Кроме того, было показано, что введение MET антител, индуцирующих MET сигнальный путь, способно восстановить метаболическую функцию при диабете, включая оба типа диабета и I тип и II тип (Примеры 21 и 22). В частности, на модели диабета I типа (Пример 21) показано, что MET антитела способствуют усвоению глюкозы. Кроме того, введение MET антител вместе с инсулином приводит к синергетическому эффекту в отношении усвоения глюкозы. На модели диабета II типа (Пример 22) показано, что MET антитела нормализуют контроль над глюкозой и уменьшают резистентность к инсулину. Следовательно, в одном аспекте изобретение предоставляет способ лечения или предотвращения диабета у пациента-человека, который включает введение нуждающемуся в этом пациенту терапевтичеки эффективного количества MET антитела, индуцирующего MET сигнальный путь. В некоторых вариантах осуществления данный способ является способом лечения или предотвращения диабета I типа. В некоторых вариантах осуществления способ включает введение инсулина пациенту. В некоторых вариантах осуществления является способом лечения диабета II типа. В некоторых вариантах осуществления антитело представляет собой антитело, описанное в этом документе.
Кроме того, в данном описании было показано, что введение MET антител, индуцирующих MET сигнальный путь, способно уменьшать степень жировой дистрофии печени на мышиной модели неалкогольного стеатогепатита (NASH) (Пример 23). В частности, MET антитела были способны уменьшить количество жировых клеток и уровень фиброза. Следовательно, в одном аспекте изобретение предоставляет способ лечения или предотвращения NASH у пациента-человека, который включает введение нуждающемуся в этом пациенту терапевтичеки эффективного количества MET антитела, индуцирующего MET сигнальный путь. В некоторых вариантах осуществления антитело представляет собой антитело, описанное в этом документе.
Кроме того, было показано, что введение MET антител, индуцирующих MET сигнальный путь, может оказывать содействие заживлению раны (Пример 24). Более того, MET антитела были способны оказывать содействие заживлению раны у диабетических мышей, у которых отмечается ухудшенное заживление ран. Следовательно, в одном аспекте изобретение предоставляет способ содействия заживлению раны у пациента-человека, который включает введение пациенту, нуждающемуся в этом, терапевтически эффективного количества MET антитела, которое индуцирует MET сигнальный путь. В некоторых вариантах осуществления человек-пациент имеет диабет, необязательно диабет I типа. В некоторых вариантах осуществления антитело представляет собой антитело, описанное в этом документе.
Примеры
Далее изобретение будет трактоваться со ссылкой на следующие неограничивающие экспериментальные примеры.
Пример 1: Иммунизация лам
Иммунизацию лам, сбор лимфоцитов периферической крови (PBL), а также последующую экстракцию РНК и амплификацию фрагментов антител проводили, как описано (De Haard et al., J. Bact. 187:4531-4541, 2005). Двух взрослых лам (Lama glama) иммунизировали с помощью внутримышечной инъекции химерного белка, состоящего из внеклеточного домена (ECD) человеческого MET, слитого с Fc участком человеческого IgG1 (MET-Fc; R&D Systems). Каждая лама получала одну инъекцию в неделю в течение шести недель, всего шесть инъекций. Каждая инъекция, проводимая в шею, содержала 0,2 мг белка в неполном адьюванте Фрейнда.
Образцы крови по 10 мл собирали до и после иммунизации, чтобы изучить иммунный ответ. Приблизительно через одну неделю после последней иммунизации, 400 мл крови было собрано, и PBL были получены с использованием метода фиколл-пак. Общую РНК получали фенол-гуанидин-тиоцианатным методом (Chomczynski et al., Anal. Biochem. 162:156-159, 1987) и использовали в качесте матрицы для случайного кДНК синтеза с использованием набора SuperScriptTM III First-Strand Synthesis System kit (Life Technologies). Амплификацию кДНК, кодирующих VH-CH1 участки IgG1 ламы и VL-CL домены (κ и λ), и субклонирование в фагмидный вектор pCB3 проводили, как описано в (de Haard et al., J Biol Chem. 274:18218-18230, 1999). Штамм E. coli TG1 (Коллекция культур бактерий Нидерландов) был трансформирован с использованием рекомбинантных фагмид для получения 4 разных Fab-экспрессирующих фаговых библиотеки (одна λ и одна κ библиотека на иммунизированную ламу). Разнообразие было в пределах 108-109.
Иммунный ответ на антиген был изучен с помощью ELISA. С этой целью мы получили ECDs человеческого MET (UniProtKB # P08581; aa 1-932) и мышиного MET (UniProtKB # P16056.1; aa 1-931) с помощью стандартных методов белковой инженерии. Человеческий или мышиный MET ECD рекомбинантный белок был иммобилизован на твердой фазе (100 нг/лунку в 96-луночном планшете) и был подвергнут воздействию серийных разведений сыворотки от лам до (день 0) или после (день 45) иммунизации. Связывание обнаруживали с использованием мышиного анти-лама IgG1 (Daley et al., Clin. Vaccine Immunol. 12, 2005) и HRP-конъюгированного ослиного анти-мышь антитела (Jackson Laboratories). Как показано на Фигуре 1, обе ламы показали иммунный ответ против человеческого MET ECD. В соответствии с представлением, что внеклеточный участок человеческого MET демонстрирует 87% гомологию с его мышиным ортологом, двольно хорошая выраженность перекрестной реактивности также наблюдалась с мышиным MET ECD.
Пример 2: Выбор и скрининг Fab, связывающихся и с человеческим и с мышиным MET
Fab-экспрессирующие фаги из описанных выше библиотек были получены в соответствии со стандартными протоколами фагового дисплея. Для селекции фаги сначала адсорбировали, чтобы иммобилизировать рекомбинантный человеческий MET ECD, промыли, а затем элюировали с использованием трипсина. После двух циклов селекции с человеческим MET ECD, таким же образом проводили два других цикла с использованием мышиного MET ECD. Параллельно, мы также отобрали фаги, чередуя человеческий MET ECD цикл с мышиным MET ECD циклом, всего четыре цикла. Фаги, отобранные путем двух подходов, объединили вместе и затем их использовали для инфицирования TG1 E. coli. Изолировали отдельные колонии и секрецию Fabs индуцировали при помощи IPTG (Fermentas). Fab-содержащую периплазматическую фракцию бактерий собирали и тестировали в отношении их способности связываться с человеческим и мышиным MET ECD с помощью поверхностного плазмонного резонанса (SPR). Человеческий или мышиный MET ECD был иммобилизирован на CM-5 чипе при помощи соединения с аминной группой в натрий-ацетатном буфере (GE Healthcare). Fab-содержащие периплазматические экстракты «загружали» в прибор BIACORE 3000 (GE Healthcare) при скорости потока 30 мкл/мин. Скорость диссоциации Fab (koff) измеряли в течение двухминутного периода. Связывание Fab с человеческим и мышиным MET было, кроме того, охарактеризовано с помощью ELISA с использованием MET ECD на твердой фазе и периплазматическом сыром экстракте в растворе. Поскольку Fab были сконструированы с использованием MYC flag, связывание обнаруживали при помощи HRP-конъюгированных анти-MYC антител (ImTec Diagnostics).
Были отобраны Fab, которые связываются и с человеческим и с мышиным MET, в двух анализах SPR и ELISA, и были секвенированы их соответствующие фаги (LGC Genomics). Перекрестно-реактивные Fab последовательности были поделены на семейства, исходя из длины и состава последовательности VH CDR3. VH семействам были даны внутренние номера без учета IMTG (Международная информационная система по иммуногенетике) номенклатуры. В общем, мы смогли идентифицировать 11 разных еловеческих/мышиных перекрестно-реагирующих Fab, относящихся к 8 VH семействам. CDR и FR последовательности вариабельных участков тяжелой цепи показаны в Таблице 3. CDR и FR последовательности вариабельных участков легкой цепи показаны в Таблице 4. Полные аминокислотные последовательности вариабельных участков тяжелой цепи и легкой цепи показаны в Таблице 5. Полные последовательности ДНК вариабельных участков тяжелой цепи и легкой цепи показаны в Таблице 6.
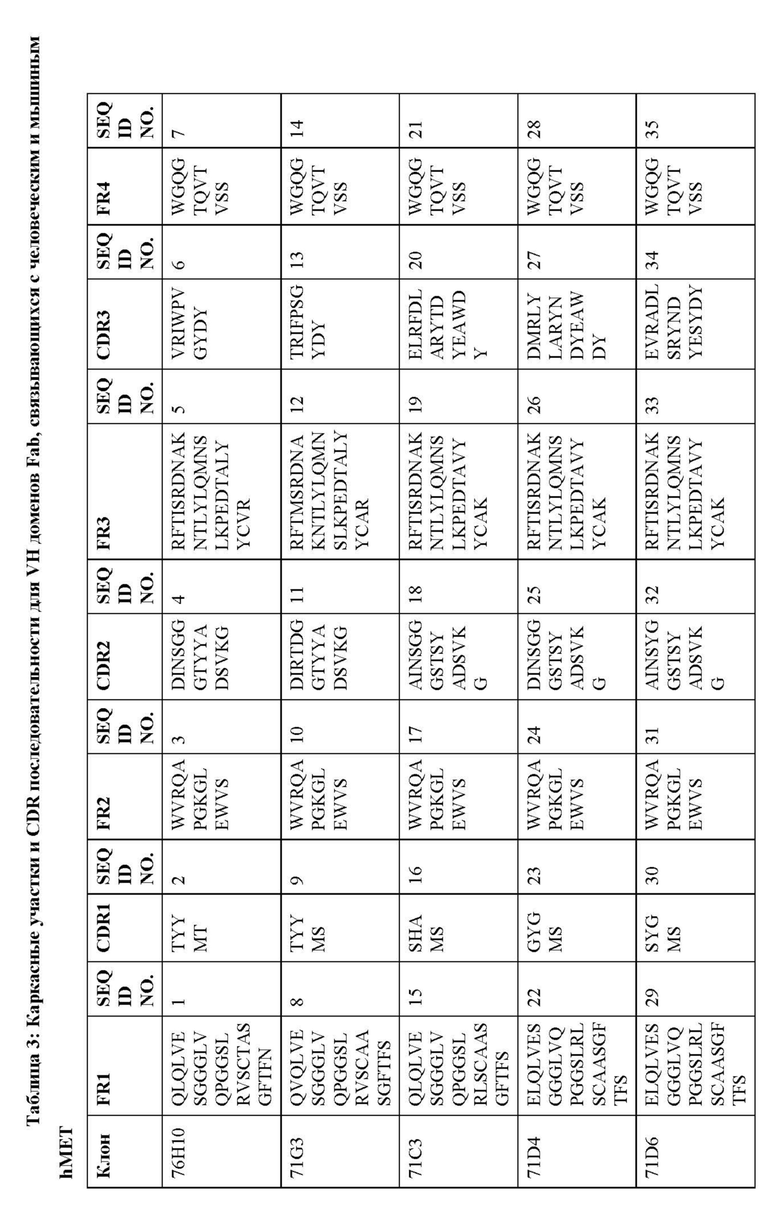
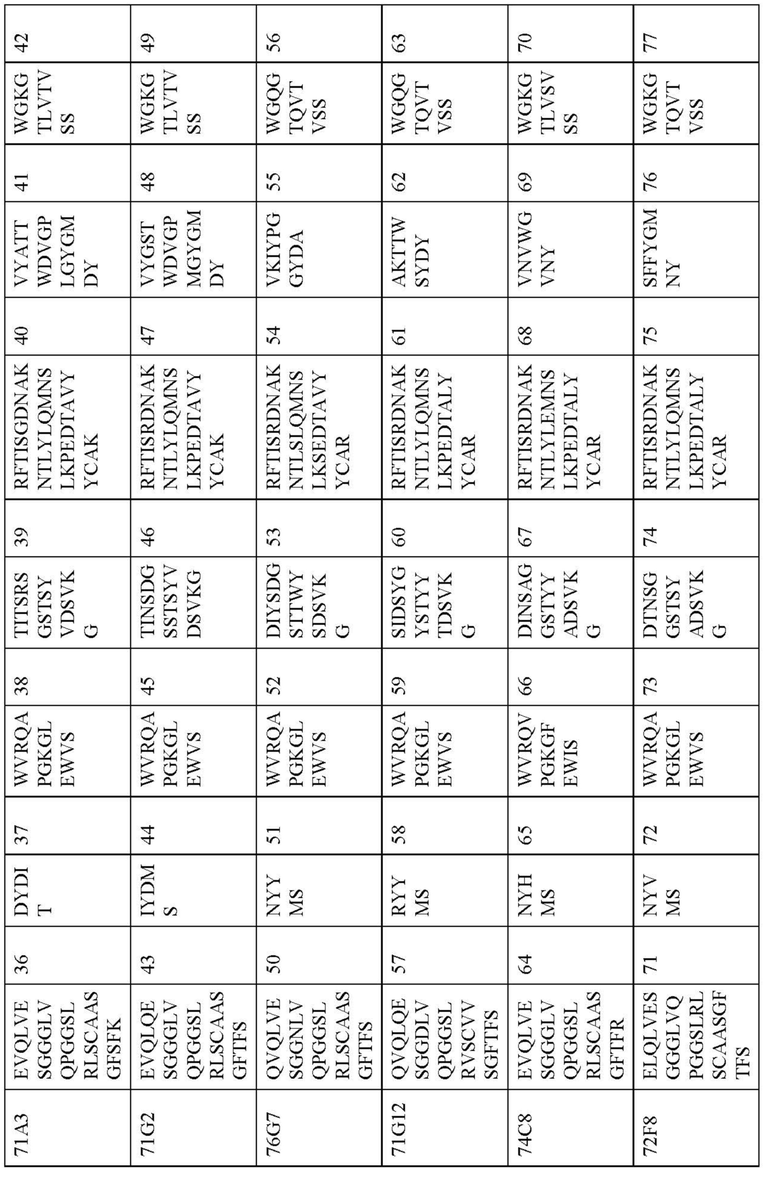
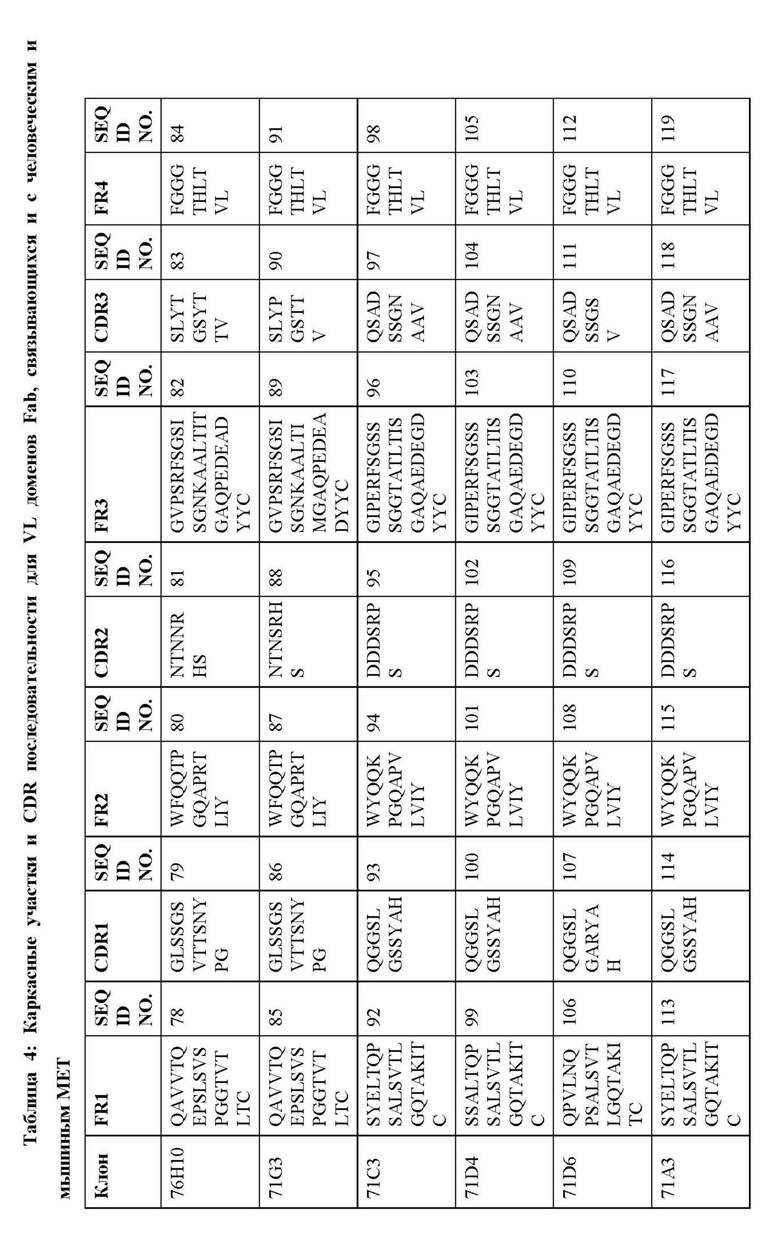







Различные семейства Fab и их способность связываться с человеческим и с мышиным MET показаны в Таблице 7.
Таблица 7: Связывание Fab и с человеческим MET (hMET) и с мышиным MET (mMET). Fab группируются в семейства, исходя из их VH CDR3 последовательности. Связывание Fab с человеческим и мышиным MET ECD было определено с помощью поверхностного плазмонного резонанса (SPR) и с помощью ELISA. SPR значения представлены в виде koff (s-1). ELISA значения представлены в виде оптической плотности (OD) при 450 нм (AU, относительные единицы). Оба анализа и SPR и ELISA проводились с использованием сырых периплазматических экстрактов. Концентрация Fab в экстракте не определялась. Значения представляют собой среднее из трех независимых измерений.
Пример 3: Химеризация Fab в mAb
кДНК, кодирующие VH и VL (κ или λ) домены отобранных Fab фрагментов, были сконструированы в два отдельных pUPE вектора экспрессии млекопитающего (U-protein Express), содержащих кДНК, кодирующие CH1, CH2 и CH3 человеческого IgG1 или человеческого CL (κ или λ), соответственно. Полные аминокислотные последовательности тяжелой цепи и легкой цепи химерного антитела лама-человек показаны в Таблице 8.






Получение (путем транзиентной трансфекции клеток млекопитающих) и очистка (с помощью аффинной хроматографии с белком A) полученных химерных лама-человек IgG1 молекул была осуществлена внешней компанией U-protein Express. Связывание химерных mAbs с MET определяли методом ELISA с использованием hMET или mMET ECD на твердой фазе и возрастающих концентраций антител (0-20 nM) в растворе. Связывание обнаруживали при помощи HRP-конъюгированных анти-человек Fc антител (Jackson Immuno Research Laboratories). Этот анализ показал, что все химерные лама-человек антитела связывались с человеческим и мышиным MET с пикомолярной аффинностью, демонстрируя EC50 в пределах 0,06 нM и 0,3 нM. Связывающая способность (EMAX) варьировала от антитела к антителу, возможно из-за частичной экспозиции эпитопа в иммобилизованном антигене, но не была сходной при человеческом и мышином «окружении». Значения EC50 и EMAX показаны в Таблице 9.
Таблица 9: Связывание химерных mAb с человеческим и мышиным MET, определенное методом ELISA с использованием иммобилизованного MET ECD на твердой фазе и возрастающих концентраций (0-20 нM) антител в растворе. EC50 значения выражаются в виде нмоль/л. EMAX значения выражаются в виде оптической плотности (OD) при 450 нм (AU, относительные единицы).
Мы также проанализировали, связываются ли химерные анти-MET антитела с нативным человеческим и мышиным MET в живых клетках. С этой целью возрастающие концентрации антител (0-100 нM) инкубировали с клетками карциномы легких человека A549 (Американская коллекция типовых культур) или мышиными клетками-предшественниками печени MLP29 (подарены профессором Enzo Medico, Университет Турина, Strada Provinciale 142 km 3.95, Candiolo, Torino, Италия; Medico et al., Mol Biol Cell 7, 495-504, 1996), и те и другие экспрессируют физиологические уровни MET. Связывание антитела с клетками было проанализировано с помощью проточной цитометрии с использованием конъюгированных с фикоэритрином анти-человек IgG1 антител (eBioscience) и CyAn ADP анализатора (Beckman Coulter). В качестве положительного контроля в отношении связывания человеческого MET мы использовали коммерческие мышиные анти-человек MET антитела (R&D Systems) и конъюгированные с фикоэритрином анти-мышиные IgG1 антитела (eBioscience). В качестве положительного контроля в отношении связывания мышиного MET мы использовали коммерческие козьи анти-мышиные MET антитела (R&D Systems) и конъюгированные с фикоэритрином анти-козьи IgG1 антитела (eBioscience). Все антитела демонстрировали дозозависимое связывание как с человеческими, так и с мышиными клетками, при этом EC50 варьировал в пределах 0,2 nM и 2,5 nM. В соответствии с результатами, полученными методом ELISA, максимальное связывание (EMAX) варьировало в зависимости от антитела, но было сходным в человеческих и мышиных клетках. Эти результаты показывают, что химерные лама-человеческие антитела распознают мембрано-связанный MET в его нативной конформации в обеих системах клеточных системах, и человеческой и мышиной. Значения EC50 и EMAX показаны в Таблице 10.
Таблица 10: Связывание химерных mAb с человеческими и мышиными клетками, определенное с помощью проточной цитометрии с использованием возрастающих концентраций (0-50 нM) антител. Значения EC50 выражаются как нмоль/л. EMAX значения выражаются в виде % относительно контроля.
Пример 4: Рецепторные области, отвечающие за связывание антитела
Для того чтобы картировать рецепторные области, распознаваемые антителами, связывающимися и с человеческим и с мышиным MET (в дальнейшем именуемые человек/мышь эквивалентные анти-MET антитела), мы измерили их способность связываться с панелью сконструированных белков, происходящих от человеческого MET, созданного как описано (Basilico et al, J Biol. Chem. 283, 21267-21227, 2008). Эта панель включала (Фигура 2): полный MET ECD (Decoy MET); MET ECD, утративший IPT домены 3 и 4 (SEMA-PSI-IPT 1-2); MET ECD, утративший IPT домены 1-4 (SEMA-PSI); изолированный SEMA домен (SEMA); фрагмент, содержащий IPT домены 3 и 4 (IPT 3-4). Созданные инженерным путем белки MET были иммобилизированы на твердой фазе и подвергнуты воздействию возрастающих концентраций химерных антител (0-50 nM) в растворе. Связывание обнаруживали при помощи HRP-конъюгированных анти-человек Fc антител (Jackson Immuno Research Laboratories). Как показано в Таблице 11, этот анализ показал, что 7 mAbs распознают эпитоп в пределах SEMA домена, тогда как другие 4 распознают эпитоп в пределах PSI домена.
Таблица 11: Связывание человек/мышь эквивалентных анти-MET антител с панелью мутантов с делецией MET, описанных на Фигуре 2. MET домен, ответственный за связывание с антителом, показан в последней колонке справа.
Чтобы более точно картировать участки MET, отвечающие за связывание антитела, мы использовали отсутствие перекрестной реактивности между нашими антителами и MET ламы (организм, использованный для получения этих иммуноглобулинов). С этой целью мы создали серию лама-человек и человек-лама химерных MET белков, охватывающих полный MET ECD, как описано (Basilico et al., J Clin Invest. 124, 3172-3186, 2014). Химеры (Фигура 3) были иммобилизированы на твердой фазе и затем подвергнуты воздействию возрастающих концентраций mAbs (0-20 nM) в растворе. Связывание обнаруживали при помощи HRP-конъюгированных анти-человек Fc антител (Jackson Immuno Research Laboratories). Этот анализ выявил, что 5 SEMA-связывающие mAb (71D6, 71C3, 71D4, 71A3, 71G2) распознают эпитоп, локализованный между aa 314-372 человеческого MET, участок, который соответствует «лопастям» 4-5 семилопастного SEMA β-пропеллера (Stamos et al., EMBO J. 23, 2325-2335, 2004). Другие 2 SEMA- связывающие mAb (74C8, 72F8) распознают эпитоп, локализованный между aa 123-223 и 224-311, в указанном порядке, которые соответствуют «лопастям» 1-3 и 1-4 SEMA β-пропеллера. PSI-связывающие mAb (76H10, 71G3, 76G7, 71G12), по-видимому, не проявили какого-либо значительного связывания с любой из двух PSI химер. Принимая во внимание результаты, представленные в Таблице 11, эти антитела вероятно распознают эпитоп, локализованный между aa 546 и 562 человеческого MET. Эти результаты суммированы в Таблице 12.
Таблица 12: Картирование эпитопов, распознаваемых человек/мышь эквивалентными анти-MET антителами, определенное методом ELISA. Человеческий MET ECD (hMET) или MET ECD ламы (lMET), а также лама-человек MET химерные белки, описанные на Фигуре 3 (CH1-7), были иммобилизированы на твердой фазе и затем подвергнуты воздействию возрастающих концентраций mAb.
Пример 5: HGF конкурентный анализ
Вышеизложенный анализ дает основание предполагать, что эпитопы, распознаваемые некоторыми человек/мышь эквивалентными анти-MET антителами, могут частично совпадать (перекрываться) с эпитопами, «занятыми» HGF, при связывании с MET (Stamos et al., EMBO J. 23, 2325-2335, 2004; Merchant et al., Proc Natl Acad Sci USA 110, E2987-2996, 2013; Basilico et al., J Clin Invest. 124, 3172-3186, 2014). Чтобы исследовать дальше эту связь, мы протестировали конкуренцию между mAb и HGF с помощью ELISA. Рекомбинантные человеческие и мышиные HGF (R&D Systems) были биотинилированы на N-концах с использованием NHS-LC-биотина (Thermo Scientific). Белок MET-Fc или человеческий или мышиный (R&D Systems), был иммобилизирован на твердой фазе, а затем подвергался воздействию 0,3 nM биотинилированного HGF, или человеческого или мышиного, в присутствии возрастающих концентраций антител (0-120 нM). HGF связывание с MET обнаруживали при помощи HRP-конъюгированного стрептавидина (Sigma-Aldrich). Как показано в Таблице 13, этот анализ позволил разделить человек/мышь эквивалентные анти-MET mAb на две группы: полные HGF конкуренты (71D6, 71C3, 71D4, 71A3, 71G2) и частичные HGF конкуренты (76H10, 71G3, 76G7, 71G12, 74C8, 72F8).
Таблица 13: Способность человек/мышь эквивалентных анти-MET антител конкурировать с HGF за связывание с MET, как определено методом ELISA. MET-Fc химерный белок (или человеческий или мышиный) иммобилизировали на твердой фазе и подвергали воздействию фиксированных концентраций биотинилированного HGF (или человеческого или мышиного), в присутствии возрастающих концентраций антител. HGF связывание с MET обнаруживали при помощи HRP-конъюгированного стрептавидина. Антитело-HGF конкуренция выражается как IC50 (концентрация, при которой достигается 50% конкуренция) и IMAX (максимальный % конкуренции, достигаемый при насыщении).
Как правило, SEMA-связывающие вытесняют HGF более эффективно, чем PSI-связывающие антитела. В частности, те антитела, которые распознают эпитоп в пределах «лопастей» 4 и 5 SEMA β-пропеллера, были наиболее сильными HGF конкурентами (71D6, 71C3, 71D4, 71A3, 71G2). Это наблюдение согласуется с мнением, что SEMA «лопасть» 5 содержит высокоаффинный связывающий участок для α-цепи HGF (Merchant et al., Proc Natl Acad Sci USA 110, E2987-2996, 2013). Показано, что PSI домен взаимодействует непосредственно с HGF, однако делается предположение, что он функционирует в качестве ‘шарнира’, регулирующего размещение HGF между SEMA доменом и IPT участком (Basilico et al., J Clin Invest. 124, 3172-3186, 2014). Вследствие этого вероятно, что mAbs связывание с PSI (76H10, 71G3, 76G7, 71G12) препятствует HGF связыванию с MET, вмешиваясь в этот процесс или в результате стерического несоответствия, а не в результате прямой конкуренции с лигандом. Наконец, показано, что «лопасти» 1-3 SEMA β-пропеллера являются ответственными за низкоаффинное связывание β-цепи HGF, которая играет основную роль в MET активации, но только частично способствует HGF-MET связывающей способности (Stamos et al., EMBO J. 23, 2325-2335, 2004). Это может объяснить, почему mAbs связывание с этим участком MET (74C8, 72F8) является частичным конкурентом HGF.
Пример 6: Анализ MET активации
Вследствие их двухвалентной природы, иммуноглобулины, направленные против рецепторных тирозинкиназ, могут проявлять рецепторную агонистическую активность, подражая эффекту природных лигандов. Для дальнейшего изучения в этом направлении мы протестировали способность человек/мышь эквивалентных анти-MET антител способствовать MET ауто-фосфорилированию в анализе активации рецептора. A549 клетки карциномы легких человека и MLP29 мышиные клетки-предшественники печени лишали сывороточных ростовых факторов в течение 48 часов, а затем стимулировали возрастающими концентрациями (0-5 нM) антител или рекомбинантным HGF (A549 клетки, рекомбинантный человеческий HGF, R&D Systems; MLP29 клетки, рекомбинантный мышиный HGF, R&D Systems). После 15-минутной стимуляции клетки два раза промывали охлажденным до температуры льда фосфатно-солевым буферным раствором (PBS) и затем лизировали, как описано (Longati et al., Oncogene 9, 49-57, 1994). Лизаты белков разгоняли с помощью электрофореза и затем анализировали с использованием вестерн-блоттинга и антител, специфических к фосфорилированной форме MET (тирозины 1234-1235), независимо от того человеческие или мышиные (Cell Signaling Technology). Те же самые лизаты анализировали с помощью вестерн-блоттинга с использованием антител против тотального человеческого MET (Invitrogen) или антител против тотального мышиного MET (R&D Systems). Этот анализ показал, что все человек/мышь эквивалентные антитела демонстрируют MET агонистическую активность. Как показано на Фигуре 4, некоторые антитела способствовали MET ауто-фосфорилированию в степени, сравнимой с ауто-фосфорилированием HGF (71G3, 71D6, 71C3, 71D4, 71A3, 71G2, 74C8). Некоторые другие (76H10, 76G7, 71G12, 72F8) были менее активными, причем это было особенно очевидно при более низких концентрациях антител. Очевидная корреляция между MET активирующей активностью и HGF-конкурентной активностью не неблюдалась.
Для получения более количесвенных показателе агонистическая активность антител была также охарактеризована с помощью фосфо-MET ELISA. С этой целью A549 и MLP29 клетки выдерживали без сыворотки, как описано выше, а затем стимулировали возрастающими концентрациями (0-25 нM) mAb. Рекомбинантный человеческий (A549) или мышиный (MLP29) HGF использовали в качестве контроля. Клетки лизировали и определяли уровни фосфо-MET с помощью ELISA, как описано (Basilico et al., J Clin Invest. 124, 3172-3186, 2014). Коротко, 96-луночные планшеты покрывали мышиными анти-человек MET антителами или крысиными анти-мышь MET антителами (и те и другие от компании R&D Systems) и затем инкубировали с клеточными лизатами. После промывки захваченные белки инкубировали с биотин-конъюгированными анти-фосфо-тирозин антителами (Thermo Fisher), а связывание обнаруживали при помощи HRP-конъюгированного стрептавидина (Sigma-Aldrich).
Результаты этого анализа соответствуют данным, полученным вестерн-блоттингом. Как показано в Таблице 14, 71G3, 71D6, 71C3, 71D4, 71A3, 71G2 и 74C8 сильно активировали MET, тогда как 76H10, 76G7, 71G12 и 72F8 вызывали менее выраженный эффект. В любом случае все антитела показали сравнимый эффект в человеческих и в мышиных клетках.
Таблица 14: Агонистическая активность человек/мышь эквивалентных анти-MET антител в человеческих и в мышиных клетках, измеренная методом ELISA. A549 клетки карциномы легких человека и MLP29 мышиные клетки-предшественники печени выдерживали без сыворотки, а затем стимулировали возрастающими концентрациями mAb. Рекомбинантный человеческий HGF (hHGF; A549) или мышиный HGF (mHGF; MLP29) использовали в качестве контроля. Клеточные лизаты анализировали методом ELISA с использованием антител к тотальному MET для захвата и анти-фосфотирозин антител для обнаружения. Агонистическая активность выражалась как EC50 (nM) и EMAX (% HGF активности).
Пример 7: Анализ рассеивания
Чтобы оценить, может ли агонистическая активность человек/мышь эквивалентных анти-MET антител трансформироваться в биологическую активность, мы провели анализ рассеивания и с человеческими и с мышиными эпителиальными клетками. С этой целью клетки аденокарциномы поджелудочной железы человека HPAF-II (Американская коллекция типовых культур) и мышиные клетки-предшественники печени MLP29 стимулировали возрастающими концентрациями рекомбинантного HGF (человеческого или мышиного; и то и другое от компании R&D Systems) и клеточное рассеивание определяли через 24 часа с помощью микроскопии, как описано ранее (Basilico et al., J Clin Invest. 124, 3172-3186, 2014). Этот предварительный анализ показал, что HGF-индуцированное клеточное рассеивание имеет линейный вид, до тех пор, пока не достигает насыщения приблизительно при 0,1 нM в обеих клеточных линиях. На основе этих HGF стандартных кривых мы разработали систему баллов в пределах от (полное отсутствие клеточного рассеивания при отсутствии HGF) до 4 (максимальное клеточное рассеивание в присутствии 0,1 нM HGF). HPAF-II и MLP29 клетки стимулировали возрастающими концентрациями человек/мышь эквивалентных анти-MET антител, и клеточное рассеивание определяли через 24 часа после использования системы подсчета очков, описанной выше. Как показано в Таблице 15, этот анализ показал, что все протестированные mAb способствовали клеточному рассеиванию в обеих и в человеческой и в мышиной клеточной системах, в основном с перекрывающимися результатами для обоих видов. 71D6 и 71G2 продемонстрировали ту же самую активность как HGF; 71G3 и 71A3 были только немного более сильными, чем HGF; 71C3 и 74C8 требовали существенно более высокой концентрации для того, чтобы соответствовать активности HGF; 71D4, 76G7, 71G12 и 72F8 не достигали насыщения в этом анализе.
Таблица 15: Биологическая активность человеческих/мышиных эквивалентных анти-MET антител, измеренная в анализе клеточного рассеивания. HPAF-II клетки аденокарциномы поджелудочной железы человека и MLP29 мышиные клетки-предшественники печени стимулировали возрастающими концентрациями человек/мышь эквивалентных анти-MET антител, и клеточное рассеивание определяли через 24 часа после использования системы подсчета баллов, описанной в тексте (0, отсутствие клеточного рассеивания; 4, максимальное клеточное рассеивание).
HPAF-II клетки аденокарциномы поджелудочной железы человека
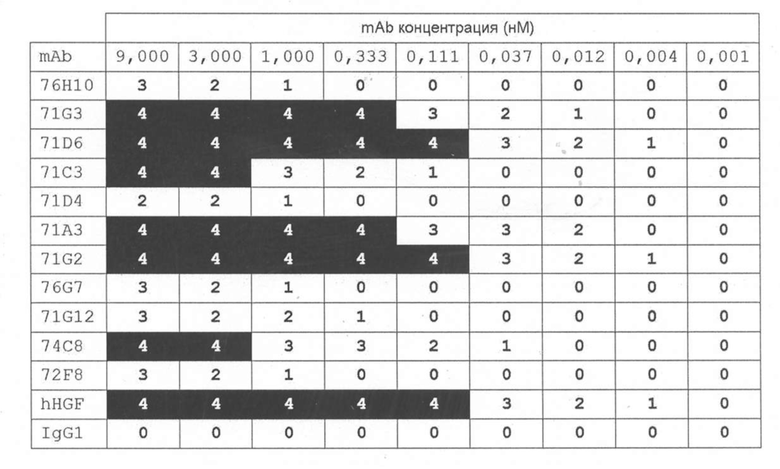

Пример 8: Защита от апоптоза, индуцированного лекарственным средством
Некоторое количество экспериментальных данных показывает, что HGF демонстрирует сильный анти-апоптотический эффект на MET-экспрессирующих клетках (Nakamura et al., J Gastroenterol Hepatol. 26 Suppl 1, 188-202, 2011). Чтобы проверить сильную анти-апоптотическую активность человек/мышь эквивалентных анти-MET антител, мы провели анализ выживаемости клеток при воздействии лекарственного средства. MCF10A эпителиальные клетки молочной железы человека (Американская коллекция типовых культур) и MLP29 мышиные клетки-предшественники печени инкубировали с возрастающими концентрациями стауроспорина (Sigma Aldrich). Через 48 часов жизнеспособность клеток определяли путем измерения общей концентрации ATP с использованием набора Cell Titer Glo kit (Promega) с помощью многоканального планшетного ридера Victor X4 (Perkin Elmer). Этот предварительный анализ показал, что концентрация лекарственного средства, которая индуцирует примерно 50% клеточную смерть, составляет 60 нM для клеток MCF10A и 100 нM для клеток MLP29. Затем, мы инкубировали MCF10A клетки и MLP29 клетки с установленными выше концентрациями лекарственного средства в присутствии возрастающих концентраций (0-32 nM) анти-MET mAb или рекомбинантного HGF (человеческого или мышиного; и тот и другой от компании R&D Systems). Жизнеспособность клеток определяли через 48 часов, как описано выше. Результаты этого анализа, представленные в Таблице 16, дают основание считать, что человек/мышь эквивалентные антитела защищали человеческие и мышиные клетки от индуцированной стауроспорином клеточной смерти в сопоставимой степени. В то время как некоторые mAb продемонстрировали защитную активность, сходную или превосходящую активность HGF (71G3, 71D6, 71G2), другие молекулы показали только частичную защиту (76H10, 71C3, 71D4, 71A3, 76G7, 71G12, 74C8, 72F8), или в человеческой или в мышиной клеточной системе.
Таблица 16: Биологическая активность человек/мышь эквивалентных анти-MET антител, измеренная с помощью клеточного анализа апоптоза, индуцированного лекарственным средством. MCF10A эпителиальные клетки молочной железы человека и MLP29 мышиные клетки-предшественники печени инкубировали с фиксированной концентрацией стауроспорина в присутствии возрастающих концентраций анти-MET mAb или рекомбинантного HGF (человеческого или мышиного), и общее содержание ATP определяли через 48 часов. Жизнеспособность клеток вычисляли как % от общего содержания ATP относительно клеток, не обработанных ни стауроспорином, ни антителами, и выражали в виде EC50 и EMAX.
Пример 9: Анализ морфогенеза ветвления
Как обсуждалось в разделе Уровень техники, HGF представляет собой плейотропный цитокин, способствующий гармоничной регуляции независимых видов биологической активности, включая клеточную пролиферацию, подвижность, инвазию, дифференцировку и выживаемость. Клеточным анализом, который в большей степени выявляет все эти виды активности, является анализ морфогенеза ветвления, воссоздающий формирование трубчатых органови и желез в ходе эмбриогенеза (см. обзор Rosário и Birchmeier, Trends Cell Biol. 13, 328-335, 2003). При проведении этого анализа сфероид из эпителиальных клеток вносят внутрь 3D коллагеновой матрицы и стимулируют HGF к развитию «трубочек», которые в итоге формируют разветвленные структуры. Эти разветвленные «трубочки» напоминают полые структуры эпителиальных желез, например, молочной железы, в том смысле, что они демонстрируют просвет, окруженный поляризованными клетками. Этот анализ является наиболее полным HGF анализом, который может выполняться in vitro.
Для того чтобы выяснить, проявляют ли человек/мышь эквивалентные анти-MET антитела агонистическую активность в этом анализе, мы высеяли LOC эпителиальные клетки почки человека (Michieli et al. Nat Biotechnol. 20, 488-495, 2002) и MLP29 мышиные клетки-предшественники печени в коллагеновый слой, как описано (Hultberg et al., Cancer Res. 75, 3373-3383, 2015), и затем подвергли их воздействию возрастающих концентраций mAbs или рекомбинантного HGF (человеческого или мышиного; и тот и другой от компании R&D Systems). Морфогенез ветвления в течение некоторого периода времени сопровождался микроскопией, при этом колонии фотографировали через 5 дней. Характерные изображения представлены на Фигуре 5. Количественная оценка активности морфогенеза ветвления была выполнена путем подсчета числа ветвлений на каждый сфероид. Как показано в Таблице 17, все протестированные антитела вызывали дозо-зависимое образование разветвленных «трубочек». Вместе с тем, в соответствии с результатами, полученными в анализе MET ауто-фосфорилирования и анализе клеточного рассеивания, 71D6, 71A3 и 71G2 продемонстрировали наиболее сильную агонистическую активность, сходную или превышающую активность рекомбинантного HGF.
Таблица 17: Анализ морфогенеза ветвления. Препараты клеточных сфероидов LOC эпителиальных клеток почки человека или MLP29 мышиных клеток-предшественников печени высевали в коллагеновый слой и затем инкубировали с возрастающими концентрациями (0; 0,5; 2,5 и 12,5 нM) mAb или рекомбинантного HGF (LOC, человеческий HGF; MLP29, мышиный HGF). Морфогенез ветвления в течение некоторого времени сопровождался микроскопией, и колонии фотографировали через 5 дней. Количественная оценка ветвления осуществлялась путем подсчета числа ветвлений для каждого сфероида (первичное ветвление плюс вторичное ветвление).
LOC клетки
MLP29 клетки
Пример 10: Человек-мышь эквивалентные агонистические анти-MET антитела обладают большими возможностями в отношении модулирования MET активности
Исходя из биохимического и биологического анализов, описанных до этого, мы провели комплексный анализ, нацеленный на сравнение функций антитела. Характеристики различных mAb, полученных в проведенных анализах, суммированы в Таблице 18. При анализе этой Таблицы выяснилось, что человек-мышь эквивалентные агонистические анти-MET антитела демонстрируют широкий спектр биохимической и биологической активности, предоставляя большие возможности для модулирования MET активности. В зависимости от выбора применения в трансляционном процессе или в терапевтических целях, из идентифицированных антител могут быть выбраны антитела, которые полностью или частично конкурируют с HGF, которые сильно или слабо вызывают MET активацию, которые сильно или слабо способствуют клеточной инвазивности или которые сильно или мягко противодействуют апоптозу. В этом контексте агонистические антитела являются намного более практичными и гибкими по сравнению с HGF, поскольку они дают возможность получения более дозированного ответа по сравнению с характером HGF «включен-или-выключен».
С точки зрения фармакологии возможность вызывать специфичную биологическую активность последующих звеньев пути MET может быть весьма полезной. Например, некоторые применения в области онкологии извлекают пользу от лигандов, которые отделяют трофические свойства HGF от его про-инвазивной активности (Michieli et al., Nat Biotechnol. 20, 488-495, 2002). Другие варианты использования в области гепатологии теоретически испытывают необходимость в факторах, которые защищают гепатоциты от апоптоза без активации клеточной инвазии (Takahara et al., Hepatology, 47, 2010-2025, 2008). При применении в области мышечной дистрофии дифференцировка миобластов в миоциты требует «выключения» HGF-индуцированной пролиферации, с одной стороны, и защиты от связанного с дифференцировкой апоптоза, с другой стороны (Cassano et al., PLoS One 3, e3223, 2008). Во всех этих вариантах применения и в других подобных случаях, можно представить себе применение частичных агонистических mAbs, которые заменяют эндогенный HGF, с одной стороны, и вызывают умеренную MET активацию, с другой, увеличивая, таким образом, некоторую биологическую активность HGF, в то же время, уменьшая другие.
Но, в то же время, разные применения в области регенеративной медицины требуют сильных сигналов, направляющих к выживаемости и быстрой репарации ткани, для того, чтобы предотвратить необратимое клеточное повреждение или дегенерацию. Например, эта ситуация обнаруживается в случае резкой печеночной недостаточности, острого поражения почки или тяжелого панкреатита (см. обзор Nakamura et al., J Gastroenterol Hepatol. 26 Suppl 1, 188-202, 2011). Во всех этих вариантах применения и в других подобных случаях, может быть предпочтительным использование полных агонистических mAbs, которые наиболее эффективно подталкивают заживление и регенерацию ткани. HGF конкуренция реально не играет роли в этом случае, потому что полные агонистические mAbs являются сильными, если не более сильными, чем HGF, и могут достигать фармакологических концентраций более высоких, чем физиологические уровни, характерные для эндогенного лиганда.
В другой патологической ситуации, которая касается неканонических, менее характерных функций HGF, например таких, которые вовлекают иммунную систему (воспалительные заболевания, аутоиммунные нарушения, осложнения, связанные с транплантацией), гематопоэтическую систему (мобилизацию стволовых клеток, гемопоэз) и нервную состему (рост нервов, дегенерацию нейронов), роль HGF/MET пути остается малоизученной. Наряду с тем, что некоторые экспериментальные данные показывает, что рекомбинантный HGF или HGF генная терапия улучшает эти нарушения в доклинических моделях (смотри Nakamura et al., J Gastroenterol Hepatol. 26 Suppl 1, 188-202, 2011), мы не имеем достаточно информации для определения того, являются ли все функции HGF полезными или только их часть. Для этих применений в лечебных целях тоно так же возможность «точной настройки» MET активности с весьма разнообразной панелью MET-агонистических антител является потенциально предпочтительной по сравнению с HGF (не говоря уже о многочисленных фармакологических проблемах, предполагаемых при использовании рекомбинантного HGF в качестве лекарственного средства, обсуждаемых в разделе Раскрытие изобретения).
В заключение, мы полагаем, что все идентифицированные человек-мышь эквивалентные анти-MET антитела могут быть потенциально полезными для применения в терапевтических целях, являются ли они полными или частичными конкурентами с HGF, или полными или частичными активаторами MET рецептора.
Таблица 18: Основные биохимические и биологические характеристики человек/мышь эквивалентных анти-MET антител. Эта Таблица суммирует способность каждого mAb связываться с очищенным MET ECD (ELISA), распознавать нативный MET на MET-экспрессирующих клетках (FACS), конкурировать с HGF за MET связывание (HGF конкуренция), активировать MET в анализе рецепторного ауто-фосфорилирования (MET активация), способствовать клеточному рассеиванию (анализ рассеивания), защищать клетки от апоптоза, индуцированного лекарственными средствами, (анализ выживаемости) и способствовать морфогенезу ветвления сфероидов эпителиальных клеток (морфогенез ветвления). Для каждого анализа подсчет баллов основывается на относительной активности любого данного mAb относительно других антител (+, ниже 50%; ++, выше 50%). Поскольку все антитела проявили сходную активность в человеческой и мышиной системах, показан только один подсчет баллов на анализ.
Пример 11: Обмен константных участков не изменяет биохимические и биологические свойства человек/мышь эквивалентных антител
Поскольку целью изобретения является создание и идентифицирование агонистических анти-MET антител, которые работают одинаково хорошо в человеческой и мышиной системах, мы попытались установить, влияет ли обмен человеческих константных участков тяжелой цепи и легкой цепи с соответствующими мышиными константными участками на основную биохимическую и биологическую активность репрезентативной панели антител. С этой целью мы выбрали 3 характерные молекулы из панели человек/мышь эквивалентных антител (71G3, частичный конкурент HGF и частичный агонист в биологических анализах; 71D6 и 71G2, полные конкуренты HGF и полные агонисты в биологических анализах). VH и VL участки 71G3, 71D6 и 71G2 были введены в каркасы мышиного IgG1/λ антитела. Последовательности всех мышиных вариантов иммуноглобулина доступны в общедоступных базах данных, таких как ImMunoGeneTics information system (www.imgt.org). Соединение с желательными вариабельными участками може достигаться с помощью стандартных методов генной инженерии. Полные аминокислотные последовательности тяжелых цепей и легких цепей созданных лама-мышь химерных антител показаны в Таблице 19.
Таблица 19: Полные аминокислотные последовательности тяжелой цепи и легкой цепи лама-мышь химерных mAb, связывающихся и с человеческим и с мышиным MET
Получение и очистка рекомбинантных иммуноглобулинов могут быть выполнены путем транзиентной трансфекции в клетках млекопитающих и аффинной хроматографии, соответственно, следуя надежно отработанным протоколам. В дальнейшем, мы сравнили биохимическую и биологическую активности 71G3, 71D6 и 71G2 в мышином формате с теми же самыми антителами в человеческом формате.
Мы оценили способность антитела связываться с очищенным человеческим или мышиным MET ECD при помощи ELISA, распознавать нативный MET на человеческих или мышиных клетках с помощью FACS, индуцировать рассеивание человеческих и мышиных эпителиальных клеток и способствовать морфогенезу ветвления в коллагене. Результаты этого анализа, суммированные в Таблице 20, показывают, что обмен человеческих и мышиных константных участков не влияет существенным образом на какие-либо из исследованных свойств.
Таблица 20: Обмен константных участков не изменяет биохимические и биологические свойства человек/мышь эквивалентных антител. Три характерных агонистических антитела (71G3, 71D6 и 71G2) или в мышином или в человеческом формате были подвергнуты нескольким in vitro анализам, имеющим целью охарактеризовать их основные биохимические и биологические свойства.
(ед. измер.)
(EC50, nM)
± 0.024
± 0.026
± 0.015
± 0.014
± 0.038
± 0.023
(EC50, nM)
± 0.035
± 0.028
± 0.022
± 0.025
± 0.029
± 0.021
(EMAX, % CTR)
± 15.3
± 17.2
± 9.7
± 11.4
± 19.1
± 12.5
(EMAX, % CTR)
± 11.3
± 13.2
± 14.1
± 15.8
± 18.3
± 12.1
(EMAX, % HGF)
± 9.8
± 8.9
± 7.9
± 9.5
± 9.6
± 8.2
(EMAX, % HGF)
± 8.8
± 8.4
± 8.5
± 7.9
± 9.8
± 4.7
(ветвлен n.)
± 2.1
± 3.5
± 1.5
± 3.7
± 3.5
± 4.1
(ветвлен n.)
± 8.0
± 7.3
± 8.0
± 6.1
± 7.0
± 6.3
Пример 12: Сравнение с антителами предшествующего уровня: перекрестная реактивность человек-мышь
Как подробно обсуждалось в разделе Уровень техники, некоторое количество других исследований уже описывали агонистические анти-MET антитела, которые имитируют HGF активность, по меньшей мере, частично. На момент написания данной работы, сюда можно было включить: (i) 3D6 мышиное анти-человек MET антитело (патент США № 6,099,841); (ii) 5D5 мышиное анти-человек MET антитело (патент США № 5,686,292); (iii) NO-23 мышиное анти-человек MET антитело (патент США № 7,556,804 B2); (iv) B7 человеческое нативное анти-человек MET антитело (патентная заявка США № 2014/0193431 A1); (v) DO-24 мышиное анти-человек MET антитело (Prat et al., Mol Cell Biol. 11, 5954-5962, 1991; Prat et al., J Cell Sci. 111, 237-247, 1998); и (vi) DN-30 мышиное анти-человек MET антитело (Prat et al., Mol Cell Biol. 11, 5954-5962, 1991; Prat et al., J Cell Sci. 111, 237-247, 1998).
Мы получили все агонистические анти-MET антитела предшествующего уровня, как описано далее. 3D6 гибридома была куплена в Американской коллекции типовых культур (кат. номер ATCC-HB-12093). 3D6 антитело было очишено из кондиционированной среды гибридомы с помощью стандартных протоколов аффинной хроматографии.
кДНК, кодирующая вариабельные участки 5D5 антитела, двухвалентный предшественник антагонистического анти-MET антитела онартузумаб (Merchant et al., Proc Natl Acad Sci USA 110, E2987-2996, 2013) были синтезированы на основе VH и VL последовательностей, опубликованных в патенте США № 7,476,724 B2. Полученные ДНК фрагменты были «слиты» с мышиными константными IgG1/λ доменами и получены в виде двухвалентных моноклональных антител с помощью стандартных протоколов инженерии белков.
NO-23 антитело было получено от профессора Maria Prat, Университет г. Новара, Италия (изобретатель NO-23; патент США № 7,556,804 B2). NO-23 антитело также можно получить, затребовав соответствующую гибридому в международном депозитарии Interlab Коллекции клеточных линий (ICLC) при Биотехнологическом центре (ABC) в г. Генуя, Италия (клон № ICLC 03001).
кДНК, кодирующие вариабельные участки B7 антитела, были синтезированы на основе VH и VL последовательностей, опубликованных в патентной заявке США № 2014/0193431 A1. Полученные ДНК фрагменты были «слиты» с мышиными константными IgG1/λ доменами и получены в виде двухвалентных моноклональных антител, как описано выше.
DO-24 и DN-30 антитела были получены от профессора Maria Prat, Университет г. Новара, Италия (она впервые идентифицировала и охарактеризовала DO-24 и DN-30; Prat et al., Mol Cell Biol. 11, 5954-5962, 1991; Prat et al., J Cell Sci. 111, 237-247, 1998). DO-24 антитело, теперь снятое с производства, было коммерчески доступно в течение нескольких лет от компании Upstate Biotechnology. DN-30 антитело антитело также можно получить, затребовав соответствующую гибридому в международном международном депозитарии Interlab Коллекции клеточных линий (ICLC) при Биотехнологическом центре (ABC) в г. Генуя, Италия (клон № ICLC PD 05006).
Поскольку подавляющая часть животных моделей болезней человека использует мышь в качестве хозяина, перекрестная реактивность с мышиным антигеном является необходимым условием для антитела, биологическая активность которого должна быть подтверждена в доклинических системах. Так как все антитела предшествующего уровня были получены в мыши (за исключением B7, которое было идентифицировано с использованием человеческой нативной фаговой библиотеки), маловероятно, что эти молекулы демонстрируют перекрестную реактивность с мышиным MET. Даже если минимальная перекрестная реактивность с собственным антигеном в принципе является возможной, эти взаимодействия в обычных условиях обладают очень низкой аффинностью.
Как подробно описано в патенте США № 6,099,841, 3D6 антитело не связывается с мышиным MET, и изобретатели были вынуждены использовать хорьков и норок, чтобы продемонстрировать, что их антитело обладает in vivo активностью. Совершенно понятно, было установлено, что эти животные модели не представляют собой ни идеальных систем моделирования болезней человека ни их использования в предклинической медицине. Более того, изобретатели не предоставляют каких-либо количественных данных относительно различия аффинности и активности антител между человеческими системами и системами хорьков или норок.
Было однозначно показано, что 5D5 антитело и его производные не связываются с мышиным MET (Merchant et al., Proc Natl Acad Sci USA 110, E2987-2996, 2013). Доступная информация о его перекрестной реактивности с другими доклиническими видами отсутствует.
Таким же образом, патентная заявка США № 2014/0193431 A1 не предоставляет информации относительно перекрестной реактивности B7 антитела с мышиным MET или MET других видов.
Патент США № 7,556,804 B2 заявляет, что NO-23 антитело вступает в перекрестную реакцию с мышиным, крысиным и собачьим MET, однако не предоставляется количественных экспериментальных данных для подтверждения этого положения. Изобретатели используют однократную насыщающую дозу NO-23, чтобы иммуно-преципитировать MET из лизатов мышиных, крысиных, человеческих или собачьих клеток, и затем инкубируют иммуно-преципитированные белки с радиоактивным 32P-ATP. После введения радиоактивной метки включенный 32P-ATP визуализируют с помощью авторадиографии. Этот метод является чрезвычайно чувствительным, но ни в коей мере не означает количественную оценку; не представляется возможным говорить, какому проценту перекрестной реактивности соответствуют полосы на геле.
Аналогично, предполагается, что DO-24 антитело перекрестно реагирует с мышиным MET, потому что содержащая DO-24 матригелевая гранула (Matrigel pellet) способствует восстановлению кровеносных сосудов при имплантации в брюшную полость мыши (Prat et al., J Cell Sci. 111, 237-247, 1998). Однако, это также может быть следствием повышенного воспаления, при этом не предоставляется прямого доказательства того, что DO-24 взаимодействует с мышиным MET. В разных исследованиях, показано, что однократная насыщающая доза DO-24 (20 nM) вызывает ауто-фосфорилирование MET в линии клеток сердечной мышцы крысы H9c2 и в линии клеток сердечной мышцы мыши HL-5 (Pietronave et al., Am J Physiol Heart Circ Physiol. 298, H1155-65, 2010; Fig. 1). В том же самом эксперименте показано, что более низкая доза рекомбинантного HGF (0,5 нM) вызывает MET фосфорилирование в сравнимой степени. Как подтверждают сами авторы в разделе Обсуждение, эти результаты дают основание предполагать, что DO-24 является существенно менее сильным, чем HGF, в этих клеточных линиях грызунов. Поскольку те же авторы заявляют, что DO-24 является полными агонистическим mAb, которое соответствует HGF активности в человеческих клеточных моделях (Prat et al., J Cell Sci. 111, 237-247, 1998), следует заключить, что DO-24 не добиваются такой же эффективности или активности в человеческих и в мышиных клетках. Кроме того, следует отметить, что эксперименты, продемонстрированные Pietronave et al., не являются количественными и не являются полезными для получения точных сведений о степени перекрестной реактивности, которая наблюдается между DO-24 и мышиным или крысиным MET и измерение которой требует обстоятельного исследования зависимости доза-эффект, подобного тому, который проведен нами (смотри ниже). В третьей работе смесь DO-24 и DN-30 антител используется для иммунопреципитации MET из клеточных лизатов мышиных мезенхимальных стволовых клеток (Forte et al., Stem Cells. 24, 23-33, 2006). И присутствие DN-30 и тип анализа (иммунопреципитация из лизатов клеток) препятствуют получению точной информации о способности DO-24 взаимодействовать с нативным мышиным MET. В заключение, вообще не существует экспериментального доказательства того, что DO-24 антитело вызывает сравнимые биологические ответы в человеческих и мышиных клетках.
Наконец, было однозначно показано, что DN-30 антитело не взаимодействует с мышиным MET (Prat et al., J Cell Sci. 111, 237-247, 1998; и дополн. материал Petrelli et al., Proc Natl Acad Sci USA 103, 5090-9095, 2006).
Для того, чтобы прямо установить реагируют ли перекрестно, и в какой степени, агонистические анти-MET антитела предшествующего уровня с мышиным MET и сравнить их с нашими человек/мышь эквивалентными анти-MET антителами, мы провели анализ ELISA. Поскольку все антитела предшествующего уровня были получены или созданы с использованием мышиного IgG/λ формата, мы использовали мышиную IgG/λ версию 71G3, 71D6 и 71G2. Человеческий или мышиный MET ECD был иммобилизирован на твердой фазе (100 нг/лунку в 96-луночном планшете) и подвергался воздействию возрастающих концентраций антител (0-40 нM) в растворе. Связывание обнаруживали при помощи HRP-конъюгированных анти-мышь Fc антител (Jackson Immuno Research Laboratories). Как видно из Таблицы 21, этот анализ показал, при том что антитела предшествующего уровня связывались с человеческим MET при значении KD в пределах от 0,059 нM (B7) до 4,935 нM (3D6), ни одно из них не проявило какой-либо аффинности в отношении мышиного MET, даже при концентрации вплоть до 40 нM. Среди протестированных антител только 71G3, 71D6 и 71G2 связывались и с человеческим и с мышиным MET, причем они делали это с неразличимой аффинностью и способностью. Полный профиль связывания всех антител показан на Фигуре 6.
Таблица 21: Аффинность связывания и способность к связыванию анти-MET антител с человеческоим и мышиным MET, определенная методом ELISA. Аффинность выражается в виде EC50 (нмоль/л). Способность к связыванию выражается в виде EMAX (оптическая плотность при 450 нм; n.c., не соед.). Полные профили связывания смотри на Фигуре 6.
Пример 13: Сравнение с антителами предшествующего уровня: MET ауто-фосфорилирование
Для того, чтобы сравнить агонистическую активность антител предшествующего уровня с активностью человек/мышь эквивалентных анти-MET антител, мы провели эксперимент по ауто-фосфорилированию MET с использованием человеческих и мышиных клеток. Клетки карциномы легких человека A549 и мышиные клетки-предшественники печени MLP29 были лишены сывороточных ростовых факторов в течение 48 часов, а затем их стимулировали возрастающими концентрациями антител (0-25 нM). После 15-минутного стимулирования клетки промывали два раза охлажденным до температуры льда фосфатно-буферным солевым раствором (PBS) и затем лизировали, как описано (Longati et al., Oncogene 9, 49-57, 1994). Фосфо-MET уровни определяли с помощью ELISA, как описано (Basilico et al., J Clin Invest. 124, 3172-3186, 2014) с использованием анти-MET антител для захвата (R&D Systems) и анти-фосфотирозинов для обнаружения (R&D Systems).
Этот анализ показал два основных различия между антителами предшествующего уровня и человек/мышь эквивалентными анти-MET антителами, описанными в настоящем документе. Во-первых, в соответствии с результатами, полученными в экспериментах по связыванию, только 71G3, 71D6 и 71G2 могут способствовать MET ауто-фосфорилированию и в человеческих и в мышиных клетках. Антитела предшествующего уровня, включая DO-24 и NO-23, вызывали MET активацию только в человеческих клетках; не было обнаружено активности в мышиных клетках в системе, которую мы исследовали. Во-вторых, все антитела предшествующего уровня неизменно демонстрировали более низкую агонистическую активность по сравнению с 71G3, 71D6 и 71G2. Наиболее агонистическими mAb предшествующего уровня были 5D5 и B7, которые показывали активность немного ниже, чем 71G3, 71D6 и 71G2. Наименее агонистическим mAb предшествующего уровня было 3D6. Другие молекулы проявляли промежуточную активность. Результаты этого анализа показаны на Фигуре 7.
Пример 14: Сравнение с антителами предшествующего уровня: морфогенез ветвления
Для того, чтобы сравнить биологическую активность антител предшествующего уровня с активностью человек/мышь эквивалентных анти-MET антител, мы провели анализ морфогенеза ветвления. Этот анализ воспроизводит все релевантные виды биологической активности HGF, включая пролиферацию клеток, рассеивание, дифференцировку и выживаемость. Эпителиальные клетки почки человека LOC и мышиные клетки-предшественники печени MLP29 высевали в коллагеновый слой, как описано выше, и затем инкубировали с возрастающими концентрациями mAb или рекомбинантного HGF (человеческого или мышиного, оба от компании R&D Systems). Морфогенез ветвления в течение некоторого времени сопровождался микроскопией, колонии фотографировали через 5 дней. Количественная оценка активности морфогенеза ветвления проводилась путем подсчета числа разветвленных «трубочек», прорастающих из каждого сфероида, и показана в Таблице 22. Характерные изображения сфероидов показаны на Фигуре 8 (LOC клетки) и на Фигуре 9 (MLP29 клетки).
Таблица 22: Анализ морфогенеза ветвления. Препараты клеточных сфероидов LOC эпителиальных клеток почки человека или MLP29 клеток-предшественников печени человека высевали в коллагеновый слой и затем инкубировали с возрастающими концентрациями (0; 0,04; 0,2; 1 и 5 нM) mAb или рекомбинантного человеческого HGF (LOC) или мышиного HGF (MLP29). Морфогенез ветвления в течение некоторого времени сопровождался микроскопией, колонии фотографировали через 5 дней. Ветвление количественно оценивали путем подсчета числа ветвлений для каждого сфероида (первичные ветвления плюс вторичные ветвления).
LOC клетки
MLP29 клетки
Представленные данные привели к следующим соображениям. В человеческих клетках 71D6, 71G2 и 5D5 продемонстрировали активность сравнимую с человеческим HGF; 71G3, 3D6, B7 и DO-24 вели себя как частичные агонисты; NO-23 и DN-30 проявили очень небольшую агонистическую активность. В мышиных клетках только 71G3, 71D6 и 71G2 эффективно индуцировали образование разветвленных «трубочек»; все другие антитела, в соответствии с их неспособностью связываться с мышиным MET в анализе ELISA, не индуцировали морфогенез ветвления вообще.
Мы пришли к выводу, что антитела предшествующего уровня, в противоположность человек/мышь эквивалентным анти-MET антителам, вызывают разную биологическую активность в человеческой и мышиной системах.
Пример 15: Период полужизни в плазме человек/мышь эквивалентных анти-MET антител
Затем, мы взяли отобранные человек/мышь эквивалентные анти-MET антитела для дальнейших исследований. В качестве предварительного анализа, мы определили их пиковые и текущие уровни у мышей. С этой целью мы инъецировали аффинно-очищенные 71G3, 71D6 и 71G2 (в их мышином IgG/λ формате) самкам мышей BALB/c 7 недельного возраста (Charles River) с помощью i.p. инъекции. Однократно вводили 1 мг/кг или 10 мг/кг, образцы крови брали из хвостовой вены через 3, 6, 12 и 24 часов после инъекции. Образцы крови подвергали обработке и определяли концентрацию антител в плазме с помощью ELISA. Стандартные 96-луночные планшеты покрывали человеческим MET ECD (100 нг/лунку), как описано в Примере 1, и затем подвергали воздействию увеличивающихся разведений мышиной плазмы для захвата анти-MET антител. После повторной промывки PBS присутствие анти-MET антител обнаруживали с помощью HRP-конъюгированного ослиного анти-мышь антитела (Jackson Laboratories). Чтобы определить количество связанного антитела, мы получили стандартную кривую очищенных 71G3, 71D6 и 71G2 в тех же самых условиях.
Результаты этого анализа показаны на Фигуре 10. Концентрации антител в плазме были сходны для всех протестированных антител и прямо пропорциональны количеству инъецированного белка. Через 24 часа концентрация антитела в плазме была приблизительно 15 нM в случае введения 1 мг/кг и 250 нM в случае введения 10 мг/кг. Принимая во внимание, что агонистическая активность этих антител в самых необходимых анализах (анализ морфогенеза ветвления) достигает насыщения при концентрации 5 нM или ниже, мы можем с уверенностью сделать вывод о том, что уровни в плазме антител, полученные путем i.p. инъекции являются релевантными с биологической точки зрения при введении только 1 мг/кг.
Кроме того, мы подсчитали время полужизни в плазме инъецированных антител. Это достигалось путем преобразования концентрации антител в натуральный логарифм (Ln), выстраивания данных в прямую линию и затем вычисления наклона кривой. Этот анализ привел к выводу, что время полужизни 71G3, 71D6 и 71G2 является очень сходным и соответствует приблизительно 3 дням для введения 1 мг/кг и 9 дням для введения 10 мг/кг. Эта стабильность является значительно более высокой по сравнению со стабильностью рекомбинантного HGF, который, как сообщалось, имел период полужизни 2,4 минут у грызунов (Ido et al., Hepatol Res. 30, 175-181, 2004). Стабильность в плазме данных в отношении всей панели суммируется в Таблице 23.
Эти данные подтвердили, что человек/мышь эквивалентные анти-MET антитела могут успешно заменять рекомбинантный HGF во всех случаях клинического применения, при которых требуется системное введение HGF.
Таблица 23. Стабильность в плазме человек/мышь эквивалентных антител. Однократное введение с помощью i.p. инъекции (1 мг/кг или 10 мг/кг) аффинно-очищенных 71G3, 71D6 и 71G2 самкам мышей BALB/c 7-недельного возраста. Образцы крови были взяты из хвостовой вены через 3, 6, 12 и 24 часов после инъекции, и концентрация антитела в плазме определяли методом ELISA. Период полужизни в плазме вычисляли путем преобразования концентраций антител в натуральный логарифм.
Пример 16: In vivo активность: Защита против острого поражения печени
Гепатоциты экспрессируют MET и являются основной мишенью HGF, который способствует их пролиферации и защищает их от апоптоза (см. обзор Nakamura et al., J Gastroenterol Hepatol. 1, 188-202, 2011). По этой причине мы протестировали, демонстрируют ли человек/мышь эквивалентные агонистические анти-MET антитела защитную активность на мышиных моделях острого поражения печени. С этой целью, мы инъецировали однократную дозу CCl4 (0.2 мл 10% раствора в оливковом масле; и то и другое от компании Sigma-Aldrich) в подкожный компартмент самок BALB/c мышей 7-недельного возраста (Charles River). Вскоре после CCl4 инъекции мышей случайным образом разбили на 4 группы по 6 мышей в каждой, которые получали однократную болюсную инъекцию очищенных 71G3, 71D6, 71G2 или только разбавитель (PBS). Антитела вводили с помощью i.p. инъекции в дозе 5 мг/кг. Образцы крови брали в разные моменты времени после инъекции (0, 12, 24 и 48 часов). Кроме того, пятая контрольная группа содержала 6 мышей, которые не получали ни CCl4 ни антитело, и которых умерщвляли в конце эксперимента. При аутопсии кровь и печень собирали для анализов. Уровни в плазме печеночных маркеров аспартат трансаминазы (AST), аланин аминотрансферазы (ALT) и билирубина (BIL) были определены с помощью стандартных клинических биохимических методов. Печень заливали в парафин и обрабатывали для гистологического исследования с использованием стандартных протоколов.
Как показано на Фигуре 11, CCl4 инъекция у контрольных мышей вызывала быстрое и значительное увеличение уровней всех трех проанализированных параметров крови, которые достигали пика через 12-24 часов после интоксикации. В контрольной группе инъекция CCl4 повышала уровни AST, ALT и билирубина в 286, 761 и 13 раз, соответственно. Во всех группах с введением антител, это увеличение было значительно меньше (71G3, 53%, 62%, и 46%; 71D6, 37%, 34% и 48%; 71G2, 50%, 39% и 54%, соответственно). В отношении защиты печени наиболее сильным было антитело 71D6.
Гистологическое исследование печени при аутопсии показало, что CCl4 индуцировал заметное поражение ткани центральной вены каждой доли печени, характеризующееся эозинофильным окрашиванием и большой цитоплазмой, типичной для пораженных гепатоцитов. Межклеточные взаимодействия выглядели неплотными, обеспечивая инфильтрацию красных клеток крови, пропускаемых из поврежденных сосудов. В группах, обработанных антителом, эти перицентральные поврежденные участки были меньше и показывали меньше признаков повреждения, что было видно по менее выраженному эозинофильному окрашиванию, нормальному размеру цитоплазмы и меньшей инфильтрации клеток крови. Характериные изображения срезов печени, окрашенных гематоксилином и эозином, показаны на Фигуре 12.
Эти результаты дают основание полагать, что человек/мышь эквивалентные агонистические анти-MET антитела могут использоваться в клинике для лечения острых поражений печени, характеризующихся быстрым развитием печеночной дисфункции, которая в большинство случаев приводит к ненормальным значениям биохимических показателей печени, разлитию желчи, коагулопатии, отеку головного мозга и энцефалопатии. Эти патологические состояния включают, но не ограничиваются этим, передозировку парацетамола, идиосинкратические реакции на лекарственные препараты (например, тетрациклин), употребление наркотиков (экстези, кокаин), вирусные инфекции (гепатит A, B, E).
Пример 17: In vivo активность: Защита от хронического поражения печени
Также мы проверили, оказывают ли человек/мышь эквивалентные агонистические MET антитела терапевтическое действие на мышиной модели хронического поражения печени. В действительности известно, что HGF обладает анти-фиброзной активностью в отношении печени (см. обзор Matsumoto и Nakamura, Ciba Found Symp. 212, 198-211; discussion 211-214, 1997). С этой целью, самок мышей BALB/c 7-недельного возраста (Charles River) подвергали хроническому воздействию CCl4 в течение нескольких недель. В течение первой недели мышам подкожно вводили два раза 0,1 мл 5% раствора CCl4 в оливковом масле (от компании Sigma-Aldrich). В течение последующих недель доза CCl4 была увеличена (0,1 мл 10% раствора в оливковом масле), при этом частота введения оставалась такой же (два раза в неделю). Вскоре после первой инъекции мышей случайным образом разбили на 4 группы по 7 мышей в каждой, которые получали лечение очищенными 71G3, 71D6, 71G2 или только разбавителем (PBS), соответственно. Антитела вводили три раза в неделю с помощью i.p. инъекции в дозе 1 мг/кг. В дополнение к этому, пятая контрольная группа содержала 7 мышей, которые не получали ни CCl4 ни антитело, и служила в качестве здорового контроля. Мышей забивали после 6-недельной хронической CCl4 интоксикации. При аутопсии кровь и печень собирали для анализов. Уровни в плазме гепатических маркеров аспартат трансаминазы (AST) и аланин аминотрансферазы (ALT) определяли с помощью стандартных методов клинической биохимии. Печень заливали в парафин и готовили для гистологических анализов, используя стандартные протоколы.
Как показано на Фигуре 13, хроническое воздействие CCl4 у контрольных мышей приводило к oслаблению функции печени, что было установлено по более высоким уровням в плазме AST и ALT. В противоположность острой модели, которая вызывает резкий, но временный всплеск уровней печеночных маркеров, хроническая CCl4- интоксикация вызывала более умеренное увеличение уровней AST и ALT, приблизительно в 5 раз по сравнению с необработанными мышами. Стоит отметить, что обработка антителом может полностью предотвратить увеличение концентрации AST, в действительности понижая его ниже базальных уровней. Антитела также могут в значительной степени предотвратить всплеск уровней ALT, хотя не так впечатляюще, как наблюдалось в отношении AST.
Срезы печени окрашивали с помощью различных методов, нацеленных на обнаружение фиброзной ткани, включая окраску трихром по Массону, окраску пикросириусом красным и антителами к альфа-актину гладких мышц (α-SMA). Также было выполнено окрашивание гематоксилином и эозином, для того, чтобы исследовать общее гистологическое строение. Этот анализ показал, что хроническая обработка CCl4 вызывала образование значительного количества фиброзных тканей в междольковом пространстве, которое характеризуется положительным окрашиванием пикросириусом красным и антителами к альфа-актину гладких мышц (α-SMA). Фиброзная ткань образовывала своего рода ‘ленту’, которая соединяла портальные тракты, подтверждая гексагональную форму частей печени. Следует отметить, что срезы печени, полученные от животных, получивших и CCl4 и агонистические анти-MET-антитела, продемонстрировали значительно более умеренный фиброз, что следует из интенсивности окрашивания, и участок фиброза ограничивался пери-портальным пространством. Характерные изображения срезов печени, окрашенных пикросириусом красным и анти-α-SMA антителами, показаны на Фигуре 14 и Фигуре 15, соответственно.
Эти результаты означают, что человек/мышь эквивалентные агонистические анти-MET антитела могут использоваться в клинике для лечения патологических состояний, связанных с хроническим поражением печени, которое характеризуется прогрессирующим разрушением и регенерацией паренхимальной ткани печени, приводя к циррозу и фиброзу. Агонистические анти-MET антитела могут использоваться для уменьшения или предотвращения фиброза, приводя к восстановлению структуры и функции печени. Кроме того, они могут использоваться для подавления воспаления и иммунной реакции, которые зачастую усугубляют хронические заболевания печени.
Пример 18: In vivo активность: Защита от острого поражения почек
Эпителиальные клетки почки экспрессируют значительные уровни MET и являются очень чувствительными к HGF стимулированию (см. обзор Mizuno et al. Front Biosci. 13, 7072-7086, 2008). Поэтому, мы проверили, демонстрируют ли человек/мышь эквивалентные агонистические анти-MET-антитела защитные свойства на мышиной модели острого поражения почек. С этой целью мы индуцировали тубулярное повреждение у самок мышей BALB/c 7-недельного возраста (Charles River) с помощью однократной инъекции HgCl2 (3 мг/кг). Вскоре после HgCl2 интоксикации мышей случайным образом разбили на 4 группы, которые подвергались обработке 71G3, 71D6, 71G2 или только разбавителем (PBS). Антитела вводили с помощью i.p. инъекции каждые 24 часа в дозе 10 мг/кг. Каждая группа содержала 6 мышей, которых забивали через 72 часа после HgCl2 инъекции. При аутопсии собирали кровь и почки для исследования. Уровни азота мочевины в крови (BUN) и креатинина (CRE) в плазме определяли с помощью стандартных клинических биохимических методов. Почки обрабатывали для проведения гистологического исследования с помощью стандартных протоколов.
Как видно на Фигуре 16, у контрольных мышей инъекция HgCl2 вызывала быстрое повышение уровней BUN и CRE. В контрольной группе BUN и CRE повышались в 6 и 12 раз, соответственно. Во всех группах с введением антител эти повышения были значительно меньше (71G3, 52%, и 54%; 71D6, 39% и 30%; 71G2, 45% и 44%, соответственно). Самую сильную защиту почки обеспечивало антитело 71D6.
Гистологическое исследование почек показало, что распространенное тубулярное HgCl2 повреждение характеризуется расширением проксимальных канальцев, атрофией и некрозом. Клубочковые структуры сжимаются и отсоединяются от окружающей стромы, существенно увеличивая перигломерулярное пространство. В группах, обработанных антителами, проксимальные тубулярные клетки были менее некротическими, при этом гистологическая структура клубочков выглядела неповрежденной. Характерные изображения срезов почки, окрашенных гематоксилином и эозином, показаны на Фигуре 17.
Мы полагаем, что человек/мышь эквивалентные агонистические анти-MET антитела могут использоваться в клинике для лечения патологических состояний, связанных с острым поражением почек, которые могут быть вызваны, например ишемическим или нефротоксическим повреждением, гиповолемическим шоком, нарушением проходимости системы сбора мочи, атеросклерозом, сипсисом, сахарным диабетом, аутоиммунными заболеваниями или рабдомиолизом. Агонистические анти-MET антитела могут использоваться для предотвращения или восстановления острого поражения почек, защиты тубулярных эпителиальных клеток от апоптоза, ускорения регенерации эпителиальных клеток и восстановления функции почек.
Пример 19: In vivo активность: Защита от острого поражения кишечника, уменьшение воспаления и оказание содействия регенерации на мышиной модели язвенного колита
Хорошо известно, что клетки кишечного эпителия экспрессируют MET, и что HGF играет центральную роль в гомеостазе и регенерации желудочно-кишечного тракта (см. озор Nakamura et al., J Gastroenterol Hepatol. 1, 188-202, 2011). Поэтому мы проверили, могут ли человек/мышь эквивалентные агонистические анти-MET антитела способствовать защите и регенерации кишечника на мышиной модели язвенного колита. С этой целью мы подвергли самок мышей BALB/c 7 недельного возраста (Charles River) обработке декстрансульфатом натрия (DSS) в питьевой воде в течение 10 дней. На день 10 DSS-обработку прекратили и мышей перевели обратно на потребление нормальной воды. Начиная с 1 дня, мышей случайным образом разбили на 7 групп по 7 мышей в каждой, которых обрабатывали 71G3, 71D6, 71G2 (в дозе 1 мг/кг или 5 мг/кг) или только разбавителем (PBS). Антитела вводили три раза в неделю с помощью i.p. инъекции. В дополнение к этому, восьмая контрольная группа содержала 7 мышей, которые не получали DSS или антитела, и служила в качестве здорового контроля. Мышей умерщвляли на день 12, т.е. через 2 дня после того, как введение DSS было прекращено. При аутопсии извлекали кишку, тщательно промывали и измеряли ее длину с помощью рулетки. После измерения, толстую кишку заливали в парафин и подготавливали для проведения гистологического исследования.
В течение всего периода проведения эксперимента регулярно контролировали вес мышей и оценивали клинические симптомы язвенного колита по наличию крови в фекалиях, ректальному кровотечению и консистенции стула. Количественную оценку проводили с помощью стандартной системы подсчета баллов, используемой в доклинических моделях (Kim et al., J Vis Exp. 60, pii: 3678, 2012): каждый параметр оценивали в баллах от 0 (отсутствие симптома) до 3 (максимальное проявление симптома). Баллы в отношении отдельных параметров суммировали вместе, чтобы получить Индекс активности болезни (DAI) в диапазоне от 0 до 9.
Как видно на Фигуре 18, воздействие DSS в группе с введением PBS вызывало потерю веса до 25%; DAI увеличивался до 4 баллов или выше; а длина кишки уменьшалась на 40%. Следует отметить, что при использовании всех исследованных антител эффекты изменялись дозозависимым образом, демонстрируя значительную активность уже при более низкой проверяемой дозе. 71D6 было самым «сильным» антителом: после временного снижения наблюдался возврат веса тела к нормальным значениям, сравнимым с теми, которые наблюдались в группе с PBS; антитело сдерживало увеличение DAI, значительно ингибируя все клинические симптомы; и предотвращало укорочение толстой кишки, ограничивая его незначительными изменениями.
Срезы кишки окрашивали гематоксилином и эозином и исследовали с помощью микроскопа. Как показано на Фигуре 19, введение DSS вызывало выраженное поражение слизистой толстой кишки. Эпителиальный слой выглядел разрушенным и был инфильтрован лимфоцитами. По слизистой оболочке толстой кишки были распространены места скрытых абсцессов, а слизистая оболочка была сильно колонизирована пенистыми макрофагами, ответственными за разрушение ткани. Перивисцеральные лмфатические узлы были увеличены. Слизистые железы характеризовались атрофией и демонстрировали заметное истощение слизистой, вместо которой появлялся воспалительный инфильтрат, содержащий пенистые макрофаги, лимфоциты и нейтрофилы. Некоторые язвы были заполнены гранулоцитарным или макрофагальным экссудатом, что приводило к полному исчезновению железистого компонента. Следует отметить, что мыши, обработанные и DSS и агонистическими анти-MET-антителами, продемонстрировали более умеренные признаки дегенерации и воспаления. В частности, элементы острого воспаления отсутствовали, включая макрофаги и гранулоциты; слизистая была повреждена только в небольшой степени, демонстрируя редкую деформацию и разрежение железы; секреция муцина была восстановлена, и эрозии и язвы полностью отсутствовали. Несмотря на то, что эти защитные эффекты были дозо-зависимыми во всех группах с введением антитела, они были очевидны уже при дозе 1 мг/кг, показывая, что концентрации антител, обеспеченные этой дозой, являются очень близкими к насыщению (смотри устойчивость в плазме в Примере 15). В этой модели также наиболее эффективным антителом было 71D6.
Пример 20: In vivo активность: защита от острого поражения кишечника, уменьшение воспаления и иммуносупрессии на мышиной модели воспалительного заболевания кишечника
Приведенные выше результаты подтолкнули нас к исследованию, проявляют ли агонистические анти-MET антитела терапевтический эффект на более специфической модели воспалительного заболевания кишечника. С этой целью, мы индуцировали острое поражение толстой кишки у самок мышей C57BL/6 7-недельного возраста (Charles River) при помощи внутри-ректальной инъекции 2,4,6-тринитробензолсульфоновой кислоты (TNBS), растворенной в этаноле. Известно, что комбинация TNBS/этанол индуцирует колоректальное воспаление в результате как иммунологического, так и эрозийного процессов (см. обзор Jones-Hall и Grisham, Pathophysiology 21, 267-288, 2014). TNBS, растворенную в 50% этаноле, вводили с помощью клизмы в дозе 5 мг/кг. Вскоре после введения TNBS, мышей рандомизировали на 4 группы по 6 мышей, каждая из которых получала лечение очищенными 71G3, 71D6, 71G2 или только разбавитель (PBS). Антитела вводили через день с помощью i.p. инъекции в дозе 1 мг/кг. Кроме того, пятая контрольная группа содержала 6 мышей, которые не получали TNBS или антитело, и служила в качестве здорового контроля. Вес мышей контролировали ежедневно. Мышей умерщвляли через 5 дней после введения TNBS. При аутопсии извлекали толстую кишку и измеряли, как описано выше. После измерения кишку заливали в парафин и подготавливали для гистологического исследования.
Как показано на Фигуре 20, воздействие TNBS вызывало потерю веса приблизительно на 15% и уменьшало длину кишки более чем на 20%. Эти эффекты, хоть и более умеренные по сравнению с эффектами, вызванными DSS, значительно отличались от тех, которые наблюдались во всех группах с антителами. Фактически, лечение 71G3, 71D6 и 71G2 ингибировало TNBS-индуцированную потерю веса и укорочение кишки почти полностью, что делает животных, обработанных антителами, едва различимыми от здоровых контрольных мышей.
Срезы кишки окрашивали гематоксилином и эозином и исследовали под микроскопом. Как показано на Фигуре 21, введение TNBS вызывало появление типичных признаков лимфоцитарного колита, отличающегося увеличенными перивисцеральными лимфатическими узлами, появлением агрегации лимфоцитов в подслизистой и слизистой оболочках и повышенной инфильтрацией лимфоцитов. Были видны несколько язв на полную глубину, которые связаны со стромальной гиперпролиферацией и инфильтрацией лимфоцитами и нейтрофилами. Все эти патологические процессы были сильно ингибированы в группах с введением агонистических анти-MET-антител, в которых наблюдалась уменьшенная инфильтрация лимфоцитов и уменьшенное поражение слизистой. Даже в том случае, где лимфоциты присутствовали, это не было связано с истощением слизи или повреждением эпителия.
Эти результаты и результаты, приведенные в предыдущем примере показывают, что человек/мышь эквивалентные агонистические анти-MET-антитела могут использоваться в клинике для лечения патологических состояний, связанных с язвенным колитом или вообще с воспалительными заболеваниями кишечника. Лечение агонистическими анти-MET-антителами может уменьшать интестинальные повреждения, способствовать пролиферации эпителиальных клеток и уменьшать инфильтрацию воспалительных клеток, улучшая таким образом клиническое течение болезни.
Пример 21: In vivo активность: Содействие поглощению глюкозы и взаимодействию с инсулином на мышиной модели диабета I типа
Сообщалось, что HGF способствует инсулин-зависимому поглощению глюкозы в культивируемых мышиных клетках скелетных мышц (Perdomo et al., J Biol Chem. 283, 13700-13706, 2008). Поэтому мы исследовали, могут ли наши агонистические анти-MET антитела уменьшать высокие уровни глюкозы в крови на мышиной модели диабета I типа. С этой целью мы индуцировали разрушение β-клеток поджелудочной железы у самок мышей BALB/c 7-недельного возраста (Charles River) с помощью i.p. инъекции стрептозотоцина (STZ; Sigma Aldrich). STZ вводили в дозе 40 мг/кг каждый день в течение 5 последовательных дней. Через неделю после последней инъекции, уровни глюкозы в крови в состоянии натощак определяли, используя стандартные полоски для определения глюкозы (GIMA). В это же время у мышей, обработанных STZ, наблюдалась умеренная базальная гликемия в два раза выше по сравнению с необработанными мышами (240 мг/дл vs. 120 мг/дл). Мышей случайным образом разбили на 4 группы по 7 мышей в каждой, исходя из базальной гликемии, которые получали лечение очищенными 71G3, 71D6, 71G2 или только разбавителем (PBS), соответственно. Антитела вводили два раза в неделю с помощью i.p. инъекции в дозе 1 мг/кг. В дополнение к этому пятая контрольная группа содержала 7 мышей, которые не получали STZ или антитело, и служила в качестве здорового контроля. Концентрацию глюкозы в крови в состоянии натощак контролировали в течение 5 недель. В конце пятой недели мы провели тест на устойчивость к глюкозе (GTT) и тест на устойчивость к инсулину (ITT). Тест GTT заключается во введении глюкозы голодным животным через желудочный зонд и затем измерении уровней глюкозы в крови в разные моменты времени. Тест ITT заключается во введении инсулина голодным животным с помощью i.p или i.v. инъекции и затем измерении уровней глюкозы в крови в разные моменты времени.
Как показано на Фигуре 22A, базальные уровни глюкозы в крови у мышей, обработанных STZ, продолжали увеличиваться на всем протяжении эксперимента. Это происходит в результате хронического воспаления поджелудочной железы, которое постепенно усугубляет поражение органа. В противоположность этому, леченные антителами животные демонстрировали неуклонно снижающиеся уровни гликемии, которые в конечном итоге достигали плато после второй недели лечения. Введение антитела не полностью нормализовало гликемию, но снижало ее на 25%, таким образом, доведя ее уровень примерно до половины между уровнями, наблюдаемыми у STZ-обработанных мышей и у контрольных мышей. Принимая во внимание, что на этой модели гипергликемия является следствием отсутствия инсулина, вырабатываемого β-клетками, мы задались вопросом, были ли более низкие уровни глюкозы в группах с введением антитела следствием повышенных уровней инсулина. Однако, анализ ELISA на образцах крови показал, что это не так (данные не показаны). В тесте GTT, мыши, получавшие лечение антителом, несмотря на начало с более низких уровней глюкозы в крови, были неспособны показать нормальную кривую поглощения глюкозы (Фигура 22B). Напротив, леченные антителом мыши не продемонстрировали более быстрого ответа на инсулин в тесте ITT (Фигура 22C). Через пятнадцать минут после инъекции инсулина уровни глюкозы в крови у мышей, подвергнутых хроническому лечению антителами, понизились приблизительно на 30-40% относительно момента времени ноль, что значительно меньше, чем уровни, наблюдаемые как у STZ-обработанных мышей, так и у контрольных животных (Фигура 22D). Эти результаты говорят о том, что агонистические анти-MET-антитела способствуют поглощению глюкозы при отсутствии инсулина. Они также означают, что агонистические анти-MET-антитела и инсулин, в том случае, когда оба они присутствуют, взаимодействуют при содействии поглощению глюкозы.
Эта гипотеза была проверена в клеточных анализах с использованием мышиных клеток скелетных мышц. C2C12 мышиные миобласты (полученные из Американской коллекции типовых культур) индуцировали к дифференцировке в миоциты в соответствии с рекомендациями поставщика, а затем c инкубировали с человек/мышь эквивалентными агонистическими анти-MET-антителами (71G3, 71D6, 71G2). Через 24 часа клетки, обработанные антителами, разбили на три 3 группы, которые подвергались острой стимуляции с использованием 0 нM, 100 нM или 1000 нM человеческого рекомбинантного инсулина (Sigma Aldrich) в течение 1 часа и в присутствии флуоресцентного аналога глюкозы 2-(N-(7-нитробенз-2-окса-1,3-диазол-4-ил)амино)-2-дезоксиглюкозы (2-NBDG; Life Technologies). Поглощение 2-NBDG определяли с помощью проточной цитометрии.
Как видно на Фигуре 23, 71G3, 71D6 и 71G2 способствовали поглощению глюкозы дозо-зависимым образом. Комбинация инсулина и агонистических анти-MET-антител приводила к суммарному эффекту и способствовала более высокому поглощению глюкозы по сравнению как с инсулином в отдельности, так и с антителами в отдельности. Эти результаты согласуются с установлением того, что HGF и инсулин взаимодействуют при регулировании метаболизма глюкозы в культивируемых клетках (Fafalios et al. Nat Med. 17, 1577-1584, 2011), и подтверждают нашу гипотезу о том, что агонистические анти-MET антитела способны увеличивать как инсулин-независимое, так и инсулин-зависимое поглощение глюкозы.
Пример 22: In vivo активность: нормализация уровня глюкозы в крови и преодоление резистентности к инсулину на мышиной модели диабета II типа
Под впечателением наблюдения, что человек/мышь эквивалентные агонистические анти-MET антитела могут взаимодействовать с инсулином при оказании содействия поглощению глюкозы, мы проверили их терапевтический потенциал на мышиной модели диабета II типа. Диабет II типа харектеризуется высокими уровнями глюкозы в крови, гиперинсулинемией и резистентностью к инсулину. Одной из наиболее охарактеризованных мышиных моделей диабета II типа является модель на мышах db/db, штамм C57BLKS/J, несущий точечную мутацию в гене рецептора лептина lepr. Эта мутация вызывает потерю чувства насыщения и вследствие этого неограниченное потребление пищи, что приводит к ожирению и упомянутым выше клиническим признакам диабета II типа (см. обзор Wang et al. Curr Diabetes Rev. 10, 131-145, 2014).
Самки мышей db/db были получены от компании Charles River (JAX™ Mice Strain BKS.Cg-Dock7m+/+LeprdbJ) в возрасте 7 недель. Через неделю мышей случайным образом разделили на 4 группы по 5 мышей в каждой, котрые получали лечение очищенными 71G3, 71D6, 71G2 или только разбавителем (PBS), соответственно. Антитела вводили два раза в неделю с помощью i.p. инъекции в дозе 1 мг/кг. Концентрацию глюкозы в крови в состоянии натощак проверяли каждые 14 дней в течение 8 недель. Через 7 недель лечения, т.е. когда мыши достигали 15-недельного возраста, проводили тест на устойчивость к глюкозе (GTT) и тест на устойчивость к инсулину.
Как видно на Фигуре 24, средняя базальная концентрация глюкозы в крови в PBS-группе на момент рандомизации составляла приблизительно 230 мг/дл, что полностью соответствует диабетическим уровням. Эти уровни имели тенденцию к увеличению в течение времени и в конце эксперимента, т.е. на 8 недель позже, средняя базальная концентрация глюкозы в крови в PBS-группе была приблизительно 330 мг/дл. В противоположность этому, в группах, получавших лечение антителами, базальная гликемия в условиях натощак неизменно понижалась в течение времени. В конце эксперимента средняя концентрация глюкозы в крови в группах с введением 71G3, 71D6 и 71G2 составляла 173 мг/дл, 138 мг/дл и 165 мг/дл, соответственно.
Через 7 недель лечения, т.е. когда мыши достигали 15-недельного возраста, мы тестировали их острый ответ на провокацию глюкозой и инсулином. Следует иметь в виду, что эти мыши, в отличие от мышей, обработанных STZ (смотри пример 21), являются гиперинсулинемичными и демонстрируют высокие уровни глюкозы в крови, потому что они являются инсулин-резистентными. Фактически, при стимулировании глюкозой в GTT, мыши из группы с PBS неспособны показывать нормальный профиль поглощения глюкозы. Все мыши показали резкое увеличение гликемии, которая оставалась повышенной в течение всего периода теста. Симметрично, при проведении теста ITT те же самые мыши показали парадоксальную реакцию на инсулин, демонстрируя слабое и временное увеличение уровней глюкозы. Этот парадоксальный ответ является признаком резистентности к инсулину, по меньшей мере, в доклинических моделях.
Мыши из группы с введением антител, при том что начинали с более низких базальных уровней, также не продемонстрировали нормальный профиль поглощения глюкозы в тесте GTT, таким образом, свидетельствуя, что агонистические анти-MET антитела не являются способными нейтрализовать резкое повышение уровней глюкозы в крови. Несмотря на это, стоит отметить, что лечение антителом значительно улучшало ответ на инсулин в тесте ITT, полностью изменяя парадоксальный эффект, наблюдаемый в группе с PBS, и делая ITT профиль более похожим на профиль, демонстрируемый недиабетическими мышами (C57BLKS/J; Charles River). Мы сделали заключение, что длительное лечение агонистическими анти-MET антителами улучшает состояние при диабете II типа у db/db мышей и частично преодолевает резистентность к инсулину.
На основе этих результатов и результатов, представленных в предыдущем примере, мы пришли к заключению, что человек/мышь эквивалентные агонистические анти-MET антитела могут использоваться в клинике для лечения патологических состояний, связанных с высокими уровнями глюкозы в крови. К ним можно отнести диабет I типа, диабет II типа или другие диабето-подобные патологии, характеризующиеся высоким уровнем глюкозы и/или резистентностью к инсулину (например, метаболический синдром).
Пример 23: In vivo активность: улучшение состояния при жировой дистрофии печени на мышиной модели неалкогольного стеатогепетита
Показано, что целевая генетическая мутация MET в печени приводит к развитию тяжелого неалкогольного стеатогепатита (NASH) у мышей (Kroy et al. J Hepatol. 61, 883-890, 2014). В независимом исследовании (Kosone et al., Am J Physiol Gastrointest Liver Physiol. 293, G204-210, 2007) было показано, что HGF улучшал состояние при вызванной рационом с высоким содержанием жира жировой дистрофии печени путем активации микросомальных белков-переносчиков триглицеридов (MTP) и аполипопротеина B (ApoB), таким образом, сводя к минимуму депонирование жирных кислот.
Гиперсулинемические db/db мыши также широко используются в качестве модели NASH, и в более общих чертах, болезни жировой дистрофии печени. На нормальной диете эти мыши накапливают значительное количество жира в гепатоцитах, что приводит к стеатозу (жировой дистрофии) печени, фиброзу и хронической печеночной недостаточности. Это состояние может быть еще больше ухудшено установлением для мышей рациона с высоким содержанием жира (см. обзор Anstee и Goldin, Int J Exp Pathol. 87, 1-16, 2006).
Под впечателением приведенных выше наблюдений и выводов, мы протестировали, могут ли человек/мышь эквивалентные агонистические анти-MET антитела улучшать умеренный стеатоз печени у db/db мышей при содержании на нормальной диете. С этой целью, мы получили самок мышей db/db, как описано выше. Когда животные достигали 8-недельного возраста, их случайным образом делили на 4 группы по 6 мышей в каждой, которые получали лечение очищенными 71G3, 71D6, 71G2 или только разбавителем (PBS), соответственно. Антитела вводили два раза в неделю с помощью i.p. в дозе 1 мг/кг. Через 8 недель лечения, мышей забивали и проводили аутопсию. Печени извлекали, заливали в парафин и обрабатывали для гистологического исследования. Образцы крови собирали для исследования маркеров функции печени.
Срезы печени окрашивали гематоксилином и эозином или пикросириусом красным с целью выявления фиброза. Как показано на Фигуре 25, на печени из группы с PBS было видно заметное жировое перерождение (стеатоз), как правило, сконцентрированное вокруг центральных вен. Гепатоциты были значительно увеличены и наполнены жиром. Гепатоциты, содержащие жир, были перемешаны с нормальными гепатоцитами, при этом стеатоз занимал до 60% перицентрального пространства. В противоположность этому, печени мышей, обработанных антителами, содержали значительно меньше жирных кислот и выглядели вполне нормально. Как видно на Фигуре 26, окраска пикросириусом красным выявила умеренный пери-портальный фиброз в группе с PBS, характеризующийся истончением стромального слоя вокруг печеночной триады (воротной вены, печеночной артерии и желчного протока), иногда распространением в междольковое пространство. Следует ометить, что срезы печени из всех групп с антителами продемонстрировали более низкую выраженность фиброза, если он вообще наблюдался. Анализ маркеров функции печени AST и ALT в плазме подтвердил эти наблюдения (смотри Фигуру 27). Фактически, у животных, которых лечили агонистическими анти-MET-антителами, отмечались крайне низкие концентрации в плазме AST и ALT, примерно в 2,5 раза ниже, чем в группе с PBS, и в 2 раза ниже, чем средние уровни AST и ALT у нормальных мышей.
Эти данные говорят о том, что человек/мышь эквивалентные агонистические анти-MET антитела могут использоваться в клинике для лечения NASH и других патологических состояний, связанных с жировой дистрофией печени. Агонистические анти-MET антитела могут использоваться для ингибирования накопления жира в гепатоцитах, предотвращения или инвертирования гепатостеатоза и подавления порочного цикла, который происходит между накоплением жирных кислот и инфильтрацией макрофагов. Хроническое воспаление неизбежно приводит к образованию внеклеточного матрикса. Следовательно, агонистические анти-MET антитела могут использоваться для уменьшения фиброза, связанного со стеатозом (жировой дистрофией).
Пример 24: In vivo активность: заживление ран у мышей с диабетом
Важным с клинической точки зрения осложнением диабета является повышенное образование язв и плохое заживление ран. Поскольку HGF был вовлечен в заживление ран (Nakamura et al., J Gastroenterol Hepatol. 1, 188-202, 2011), мы попытались установить, способны ли человек/мышь эквивалентные анти-MET-антитела способствовать заживлению ран, связанных с диабетом. С этой целью, мы получили db/db диабетических мышей, как описано выше. В возрасте 8 недель животных обезболивали и затем вырезали круглую рану 0,8 см в ширину на правом боку, используя инструмент с круговым лезвием для взятия биопсийного образца кожи (GIMA). Весь эпидермальный слой был удален. Через день после хирургической процедуры мышей случайным образом разделили на 4 группы, которые получали лечение очищенным 71G3, 71D6 и 71G2 или только разбавителем (PBS). Антитела вводили через день с помощью i.p инъекции в дозе 5 мг/кг. Диаметр раны измеряли каждый день, используя штангенциркуль.
Как показано на Фигуре 28, лечение антителом значительно ускоряло закрытие раны и реэпитализацию. В то время как в контрольной группе репарация экспериментальной раны происходила со средней скоростью 5% в день, это значение увеличивалось до 8% в группе 71G3, до 12% в группе 71D6 и до 11% в группе 71G2.
Мы полагаем, что человек/мышь эквивалентные агонистические анти-MET антитела могут использоваться в клинике для лечения язв и ран, связанных с диабетом, которые, как правило, демонстрируют плохое заживление. Связанные с диабетом язвы представляют собой неудовлетворенную медицинскую потребность. В Соединенных Штатах диабет является основной причиной нетравматических ампутаций нижних конечностей. Агонистические анти-MET-антитела могут использоваться для ускорения заживления, улучшения реэпитализации и оказания содействия васкулярицации язв, вызванных высоким уровнем глюкозы в крови.
Приимер 25: Перекрестная реактивность с Rattus norvegicus и Macaca fascicularis MET
Поскольку подавляющее болшинство животных моделей болезней человека используют мышь в качестве хозяина, перекрестная реактивность с мышиным антигеном является необходимым условием для антитела, которое должно подвергнуться проверке в доклинических системах. Это было основной причиной, подтолкнувшей нас к идентификации человеческих-мышиных эквивалентных анти-MET антител. Однако, некоторые доклинические исследования предпочтительно проводятся на более крупных грызунах или приматах (например, трансплантация органов и другие исследуемые применения на практике, требующие сложных хирургических вмешательств). Кроме того, фармакодинамические и фармакокинетические исследования предпочтительно проводятся на высших позвоночных, как правило, крысах и обезьянах. В заключение и самое главное, токсикологическая оценка терапевтических антител проводится на обезьянах или альтернативно - в том случае, если это невозможно - на двух разных видах грызунов. Следовательно, в идеале также желательно исследование перекрестной реактивности с крысиным и обезьяним MET.
С этой целью мы исследовали, реагируют ли перекрестно наши человек/мышь эквивалентные анти-MET-антитела с MET от других видов, включая крысиный (Rattus norvegicus) и яванского макака (Macaca fascicularis). Крысиный MET ECD (NCBI # NP_113705.1; aa 1-931) и обезьяний MET ECD (NCBI # XP_005550635.2; aa 1-948) были получены с помощью обычных методов инженерии белков. Человеческий и мышиный MET ECD использовали в качестве контролей. Была выбрана ограниченная панель антител и SEMA-связывающих (71D6, 71C3, 71D4, 71A3, 71G2) и PSI-связывающих (76H10, 71G3) антител. Антитело 5D5 предшествующего уровня использовали в качестве контроля. MET ECD белки иммобилизировали на твердой фазе (100 нг/лунку в 96-луночном планшете) и подвергли воздействию возрастающими концентрациями (0-40 нM) антител (с человеческми константными участками) в растворе. Связывание устанавливали при помощи HRP-конъюгированных анти-человек Fc антител (Jackson Immuno Research Laboratories). Как видно на Фигуре 29, все протестированные человек/мышь эквивалентные антитела связывались с человеческим, мышиным, крысиным и обезьяним MET со сходной аффинностью и способностью, тогда как 5D5 связывалось только с человеческим и обезьяним MET. Мы пришли к заключению, что 71D6, 71C3, 71D4, 71A3, 71G2, 76H10 и 71G3 антитела связываются со сходной аффинностью и способностью с человеческим MET, мышиным MET, крысиным MET и обезьяним MET, по меньшей мере, как определено с помощью ELISA.
Пример 26: Точное картирование эпитопа
Для того, чтобы точно картировать эпитопы MET, распознаваемые человек/мышь эквивалентными анти-MET-антителами, мы придерживались следующей стратегии. Мы полагали, что если антитело, выработанное в ламе и направленное против человеческого MET, перекрестно реагирует с мышиным MET, тогда это антитело вероятно распознает остаток (или несколько остатков), который является (или являются) консервативным между H. sapiens и M. Musculus, но не среди H. sapiens, M. musculus и L. glama. Точно такое распознавание может быть распространено на R. norvegicus и M. fascicularis.
Для проведения дальнейших исседований в этом направлении, мы выровняли и сравнили аминокислотные последовательности человека (UniProtKB # P08581; aa 1-932), мыши (UniProtKB # P16056.1; aa 1-931), крысы (NCBI # NP_113705.1; aa 1-931), яванского макака (NCBI # XP_005550635.2; aa 1-948) и ламы MET (GenBank # KF042853.1; aa 1-931) между друг другом (Фигура 30). Со ссылкой на Таблицу 12, мы сосредоточили наше внимание в пределах участков MET, ответственных за связывание с 71D6, 71C3, 71D4, 71A3 и 71G2 антителами (aa 314-372 человеческого MET) и с 76H10 и 71G3 антителами (aa 546-562 человеческого MET). В пределах первого участка человеческого MET (aa 314-372) имеется пять остатков, которые являются консервативными в человеческом и мышином MET, но не в MET ламы (Ala 327, Ser 336, Phe 343, Ile 367, Asp 372). Эти аминокислоты выделены черными прямоугольниками и порядковыми номерами 1-5 на Фигуре 30. Из них четыре остатка также являются консервативными у крысы и яванского макака MET (Ala 327, Ser 336, Ile 367, Asp 372). В пределах последнего участка человеческого MET (aa 546-562) имеется три остатка, которые являются консервативными в человеческом и мышином MET, но не у MET ламы (Arg 547, Ser 553, Thr 555). Эти аминокислоты выделены черными прямоугольниками и порядковыми номерами 6-8 на Фигуре 30. Из них два остатка также являются консервативными у крысиного и MET яванского макака (Ser 553 и Thr 555).
Используя человеческий MET в качестве матрицы, мы подвергли мутагенезу каждый из этих остатков в разных перестановках, получив серию MET мутантов, которые были полностью человеческими, за исключением отдельных остатков, которые были остатками ламы. Схематическое изображение мутантов показано на фигуре Фигура 31. Затем, мы протестировали аффинность отобранных SEMA-связывающих mAb (71D6, 71C3, 71D4, 71A3, 71G2) и PSI-связывающих mAb (76H10 и 71G3) для этих MET мутантов с помощью ELISA. С этой целью различные MET белки были иммобилизированы натвердой фазе (100 нг/лунку в 96-луночном планшете) и затем подвергнуты воздействию возрастающих концентраций антител (0-50 nM) в растворе. Поскольку использованные антитела были человеческими в их константном участке, связывание выявляли, используя HRP-конъюгированное анти-человек Fc вторичное антитело (Jackson Immuno Research Laboratories). Человеческий MET дикого типа использовали в качестве положительного контроля. Результаты этого анализа представлены в Таблице 24.
Таблица 24. Эпитопы MET, отвечающие за связывание агонистического антитела, имеющего консервативные остатки среди H. sapiens, M. musculus, R. norvegicus, M. fascicularis но не среди тех же видов и L. glama. Применимость остатков, консервативных среди человека, мыши, крысы, яванского макака, но не MET ламы, для связывания с агонистическими mAb, было исследовано методом ELISA. Дикого типа (WT) или мутантный (MT) человеческий MET ECD был иммобилизирован на твердой фазе и подвергался воздействию возрастающих концентраций mAb в растворе. Связывание выявляли с помощью анти-человек Fc вторичных антител. Все значения связывания были нормализованы относительно WT белка и выражались в виде % связывания (EMAX) по сравнению с WT MET. Каждый мутант (A-L), содержащий, по меньшей мере, 2 из мутаций (1-8) показан на Фигуре 31.
Представленные выше результаты предоставляют законченную и ясную картину остатков, релевантных в отношении связывания с нашими агонистическими антителами.
Все протестированные SEMA-связывающие антитела (71D6, 71C3, 71D4, 71A3, 71G2), по-видимому, связываются с тем же самым эпитопом, который содержит две ключевые аминокислоты, консервативные у человеческого, мышиного, яванского макака и крысиного MET, но не у MET ламы, расположенные в пределах «лопасти» 5 SEMA β-пропеллера: Ile 367 и Asp 372. Фактически мутация Ala 327, Ser 336 или Phe 343 совсем не влияет на связывание; мутация Ile 367 частично ухудшает связывание; мутация Ile 367 и Asp 372 полностью отменяет связывание. Мы сделали вывод, что и Ile 367 и Asp 372 человеческого MET являются решающими для связывания с проверенными SEMA-направленными антителами. Эти два остатка указаны с помощью “S” (для SEMA) на Фигуре 30.
Также протестированные PSI связывающие антитела (76H10, 71G3), по-видимому, связываются с тем же самым эпитопом. В противоположность SEMA эпитопу, однако, PSI эпитоп содержит только одну ключевую аминокислоту, также консервативную у человеческого, мышиного, яванского макака и крысиного MET, но не у MET ламы: Thr 555. Фактически, мутация Arg 547 или Ser 553 совсем не влияет на связывание, тогда как мутация Thr 555 полностью отменяет его. Мы сделали заключение, что Thr 555 представляет собой критический решающий фактор в отношении связывания с протестированными PSI-антителами. Этот остаток показан с помощью “P” (для PSI) на Фигуре 30.
Пример 27: Уникальность человек/мышь эквивалентных агонистических антител
Результаты точного картирования эпитопов, представленные в Примере 26, предоставляют подробную иллюстрацию на молекулярном уровне о том, что агонистические антитела, предоставленные этим изобретением, обладают уникальными свойствами, которые не имеют чего-либо общего с молекулами предшествующего уровня. Эта уникальность станет более понятна после следующего анализа.
В отношении большинства антител предшествующего уровня, обсужденных в Примерах 12-14, отсутствует доступная информация в отношении точных эпитопов, которые они распознают на MET. Однако, нам известно, что эти эпитопы должны быть отличными от эпитопов, распознаваемых нашими антителами, потому что ни одна из молекул предшествующего уровня не реагировала перекрестно с мышиным MET. Хорошая иллюстрация этого отличия предоставляется 5D5/онартузумабом, единственным анти-MET антителом предшествующего уровня, снабженным подробной молекулярной информацией по поводу его взаимодействия с MET. 5D5/онартузумаб распознает 4 разных остатка, которые располагаются в пределах «лопасти» 5 SEMA β-пропеллера, очень близко к аминокислотным остаткам, ответственным за взаимодействие с нашими SEMA-связывающими антителами (Merchant et al., Proc Natl Acad Sci USA 110, E2987-2996, 2013). Эти результаты, показанные с помощью “O” (для онартузумаба) на Фигуре 30, соответствуют Gln 328, Arg 331, Leu 337 и Asn 338.
Интересно отметить, что ни один из этих остатков не является консервативным между H. sapiens и M. musculus. Это полностью согласуется с идеей о том, что D5/онартузумаб был получен с использованием мыши в качестве хозяина, и объясняет, почему он не реагирует перекрестно с мышиным MET (Merchant et al., см. выше, и наши данные представленные на Фигуре 6 и Фигуре 29). Более того, ни один из этих остатков не является консервативным среди H. sapiens и R. norvegicus, однако все они являются консервативными между H. sapiens и M. fascicularis. Это объясняет, почему 5D5/онартузумаб не связывается с крысиным MET, но связывается с MET яванского макака (Фигура 29).
В противоположность 5D5/онартузумабу и всем другим предшествующим молекулам, обсуждаемым в данном документе, наши человек/мышь эквивалентные антитела были получены с использованием ламы в качестве хозяина и были непосредственно скринированы в отношении их способности перекрестно реагировать и с человеческим и с мышиным MET. Если учесть, что большинство из немногих аминокислот, которые являются консервативными между H. sapiens и M. musculus, но не среди H. sapiens, M. musculus и L. glama (показанных в черных квадратах на Фигура 30), также являются консервативными в R. norvegicus и M. fascicularis, случай определил, что выбранные антитела также являются перекрестно-реагирующими с крысиным и обезьяним.
В заключение, и стратегия иммунизации и дизайн скрининга делают антитела этого изобретения уникальными. С одной стороны, виды, использованные для иммунизации (L. glama), являются достаточно удаленными от H. sapiens, M. musculus, R. norvegicus и M. Fascicularis, чтобы гарантировать наличие достаточного количества ошибочных спариваний среди аминокислотных последовательностей MET ламы по сравнению с MET других видов (смотри Фигура 30). Эти ошибочные спаривания являются определяющими, потому что иммунизированный хозяин не может индуцировать антитела против эпитопа, который он распознает как ‘свой’. С другой стороны, протокол двойного человек/мышь скринирования ускоряет селекцию тех антител, которые распознают эпитопы, консервативные между этими двумя видами. Эта стадия является также необходимой, потому что без двух-видового пэннинга можно просто отобрать антитела, которые являются наиболее представленными или демонстрируют более высокую аффинность, но не обязательно являются перекрестно-реагирующими. Введение обоих этих критериев (пятого вида и протокола ‘двойного получения’) дают нам возможность идентифицировать антитела с новыми, уникальными свойствами.
Ссылки
Anstee QM и Goldin RD, Int J Exp Pathol. 87, 1-16, 2006
Basilico C et al, J Biol. Chem. 283:21267-21227, 2008
Basilico C et al., J Clin Invest. 124, 3172-3186, 2014
Cassano M et al., PLoS One 3, e3223, 2008
Chomczynski P et al., Anal. Biochem. 162:156-159, 1987
Daley LP et al., Clin. Vaccine Immunol. 12, 2005
de Haard H et al., J Biol Chem. 274:18218-18230, 1999
De Haard H et al., J. Bact. 187:4531-4541, 2005
Fafalios A et al. Nat Med. 17, 1577-1584, 2011
Forte G et al., Stem Cells. 24, 23-33, 2006
Hultberg A et al., Cancer Res. 75, 3373-3383, 2015
Ido A et al., Hepatol Res. 30, 175-181, 2004
Jones-Hall YL и Grisham MB, Pathophysiology 21, 267-288, 2014
Kim JJ et al., J Vis Exp. 60, pii: 3678, 2012
Kosone T et al., Am J Physiol Gastrointest Liver Physiol. 293, G204-210, 2007
Kroy DC et al. J Hepatol. 61, 883-890, 2014
Longati P et al., Oncogene 9, 49-57, 1994
Matsumoto K и Nakamura T, Ciba Found Symp. 212, 198-214, 1997
Medico E et al., Mol Biol Cell 7, 495-504, 1996
Merchant M et al., Proc Natl Acad Sci USA 110, E2987-2996, 2013
Michieli P et al. Nat Biotechnol. 20, 488-495, 2002
Mizuno S et al. Front Biosci. 13, 7072-7086, 2008
Nakamura T et al., J Gastroenterol Hepatol. 26 Suppl 1, 188-202, 2011
Perdomo G et al., J Biol Chem. 283, 13700-13706, 2008
Petrelli A et al., Proc Natl Acad Sci USA 103, 5090-9095, 2006
Pietronave S et al., Am J Physiol Heart Circ Physiol. 298, H1155-65, 2010
Prat M et al., Mol Cell Biol. 11, 5954-5962, 1991
Prat M et al., J Cell Sci. 111, 237-247, 1998
Rosário M и Birchmeier W, Trends Cell Biol. 13, 328-335, 2003
Stamos J et al., EMBO J. 23, 2325-2335, 2004
Takahara et al., Hepatology, 47, 2010-2025, 2008
Wang B et al. Curr Diabetes Rev. 10, 131-145, 2014
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> AGOMAB THERAPEUTICS BVBA
<120> ANTI-MET ANTIBODIES AND USES THEREOF
<130> BET18P4323
<140> PCT/EP2017/065599
<141> 2017-06-23
<150> GB1611123.9
<151> 2016-06-27
<160> 248
<170> PatentIn version 3.5
<210> 1
<211> 30
<212> PRT
<213> Lama glama
<400> 1
Gln Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Val Ser Cys Thr Ala Ser Gly Phe Thr Phe Asn
20 25 30
<210> 2
<211> 5
<212> PRT
<213> Lama glama
<400> 2
Thr Tyr Tyr Met Thr
1 5
<210> 3
<211> 14
<212> PRT
<213> Lama glama
<400> 3
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 4
<211> 16
<212> PRT
<213> Lama glama
<400> 4
Asp Ile Asn Ser Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 5
<211> 32
<212> PRT
<213> Lama glama
<400> 5
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Val Arg
20 25 30
<210> 6
<211> 10
<212> PRT
<213> Lama glama
<400> 6
Val Arg Ile Trp Pro Val Gly Tyr Asp Tyr
1 5 10
<210> 7
<211> 11
<212> PRT
<213> Lama glama
<400> 7
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
1 5 10
<210> 8
<211> 30
<212> PRT
<213> Lama glama
<400> 8
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Val Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 9
<211> 5
<212> PRT
<213> Lama glama
<400> 9
Thr Tyr Tyr Met Ser
1 5
<210> 10
<211> 14
<212> PRT
<213> Lama glama
<400> 10
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 11
<211> 16
<212> PRT
<213> Lama glama
<400> 11
Asp Ile Arg Thr Asp Gly Gly Thr Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 12
<211> 32
<212> PRT
<213> Lama glama
<400> 12
Arg Phe Thr Met Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Ala Arg
20 25 30
<210> 13
<211> 10
<212> PRT
<213> Lama glama
<400> 13
Thr Arg Ile Phe Pro Ser Gly Tyr Asp Tyr
1 5 10
<210> 14
<211> 11
<212> PRT
<213> Lama glama
<400> 14
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
1 5 10
<210> 15
<211> 30
<212> PRT
<213> Lama glama
<400> 15
Gln Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 16
<211> 5
<212> PRT
<213> Lama glama
<400> 16
Ser His Ala Met Ser
1 5
<210> 17
<211> 14
<212> PRT
<213> Lama glama
<400> 17
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 18
<211> 17
<212> PRT
<213> Lama glama
<400> 18
Ala Ile Asn Ser Gly Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 19
<211> 32
<212> PRT
<213> Lama glama
<400> 19
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
20 25 30
<210> 20
<211> 17
<212> PRT
<213> Lama glama
<400> 20
Glu Leu Arg Phe Asp Leu Ala Arg Tyr Thr Asp Tyr Glu Ala Trp Asp
1 5 10 15
Tyr
<210> 21
<211> 11
<212> PRT
<213> Lama glama
<400> 21
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
1 5 10
<210> 22
<211> 30
<212> PRT
<213> Lama glama
<400> 22
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 23
<211> 5
<212> PRT
<213> Lama glama
<400> 23
Gly Tyr Gly Met Ser
1 5
<210> 24
<211> 14
<212> PRT
<213> Lama glama
<400> 24
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 25
<211> 17
<212> PRT
<213> Lama glama
<400> 25
Asp Ile Asn Ser Gly Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 26
<211> 32
<212> PRT
<213> Lama glama
<400> 26
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
20 25 30
<210> 27
<211> 17
<212> PRT
<213> Lama glama
<400> 27
Asp Met Arg Leu Tyr Leu Ala Arg Tyr Asn Asp Tyr Glu Ala Trp Asp
1 5 10 15
Tyr
<210> 28
<211> 11
<212> PRT
<213> Lama glama
<400> 28
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
1 5 10
<210> 29
<211> 30
<212> PRT
<213> Lama glama
<400> 29
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 30
<211> 5
<212> PRT
<213> Lama glama
<400> 30
Ser Tyr Gly Met Ser
1 5
<210> 31
<211> 14
<212> PRT
<213> Lama glama
<400> 31
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 32
<211> 17
<212> PRT
<213> Lama glama
<400> 32
Ala Ile Asn Ser Tyr Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 33
<211> 32
<212> PRT
<213> Lama glama
<400> 33
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
20 25 30
<210> 34
<211> 17
<212> PRT
<213> Lama glama
<400> 34
Glu Val Arg Ala Asp Leu Ser Arg Tyr Asn Asp Tyr Glu Ser Tyr Asp
1 5 10 15
Tyr
<210> 35
<211> 11
<212> PRT
<213> Lama glama
<400> 35
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
1 5 10
<210> 36
<211> 30
<212> PRT
<213> Lama glama
<400> 36
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Lys
20 25 30
<210> 37
<211> 5
<212> PRT
<213> Lama glama
<400> 37
Asp Tyr Asp Ile Thr
1 5
<210> 38
<211> 14
<212> PRT
<213> Lama glama
<400> 38
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 39
<211> 17
<212> PRT
<213> Lama glama
<400> 39
Thr Ile Thr Ser Arg Ser Gly Ser Thr Ser Tyr Val Asp Ser Val Lys
1 5 10 15
Gly
<210> 40
<211> 32
<212> PRT
<213> Lama glama
<400> 40
Arg Phe Thr Ile Ser Gly Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
20 25 30
<210> 41
<211> 17
<212> PRT
<213> Lama glama
<400> 41
Val Tyr Ala Thr Thr Trp Asp Val Gly Pro Leu Gly Tyr Gly Met Asp
1 5 10 15
Tyr
<210> 42
<211> 11
<212> PRT
<213> Lama glama
<400> 42
Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 43
<211> 30
<212> PRT
<213> Lama glama
<400> 43
Glu Val Gln Leu Gln Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 44
<211> 5
<212> PRT
<213> Lama glama
<400> 44
Ile Tyr Asp Met Ser
1 5
<210> 45
<211> 14
<212> PRT
<213> Lama glama
<400> 45
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 46
<211> 17
<212> PRT
<213> Lama glama
<400> 46
Thr Ile Asn Ser Asp Gly Ser Ser Thr Ser Tyr Val Asp Ser Val Lys
1 5 10 15
Gly
<210> 47
<211> 32
<212> PRT
<213> Lama glama
<400> 47
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys
20 25 30
<210> 48
<211> 17
<212> PRT
<213> Lama glama
<400> 48
Val Tyr Gly Ser Thr Trp Asp Val Gly Pro Met Gly Tyr Gly Met Asp
1 5 10 15
Tyr
<210> 49
<211> 11
<212> PRT
<213> Lama glama
<400> 49
Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 50
<211> 30
<212> PRT
<213> Lama glama
<400> 50
Gln Val Gln Leu Val Glu Ser Gly Gly Asn Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 51
<211> 5
<212> PRT
<213> Lama glama
<400> 51
Asn Tyr Tyr Met Ser
1 5
<210> 52
<211> 14
<212> PRT
<213> Lama glama
<400> 52
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 53
<211> 17
<212> PRT
<213> Lama glama
<400> 53
Asp Ile Tyr Ser Asp Gly Ser Thr Thr Trp Tyr Ser Asp Ser Val Lys
1 5 10 15
Gly
<210> 54
<211> 32
<212> PRT
<213> Lama glama
<400> 54
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 55
<211> 10
<212> PRT
<213> Lama glama
<400> 55
Val Lys Ile Tyr Pro Gly Gly Tyr Asp Ala
1 5 10
<210> 56
<211> 11
<212> PRT
<213> Lama glama
<400> 56
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
1 5 10
<210> 57
<211> 30
<212> PRT
<213> Lama glama
<400> 57
Gln Val Gln Leu Gln Glu Ser Gly Gly Asp Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Val Ser Cys Val Val Ser Gly Phe Thr Phe Ser
20 25 30
<210> 58
<211> 5
<212> PRT
<213> Lama glama
<400> 58
Arg Tyr Tyr Met Ser
1 5
<210> 59
<211> 14
<212> PRT
<213> Lama glama
<400> 59
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 60
<211> 17
<212> PRT
<213> Lama glama
<400> 60
Ser Ile Asp Ser Tyr Gly Tyr Ser Thr Tyr Tyr Thr Asp Ser Val Lys
1 5 10 15
Gly
<210> 61
<211> 32
<212> PRT
<213> Lama glama
<400> 61
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Ala Arg
20 25 30
<210> 62
<211> 9
<212> PRT
<213> Lama glama
<400> 62
Ala Lys Thr Thr Trp Ser Tyr Asp Tyr
1 5
<210> 63
<211> 11
<212> PRT
<213> Lama glama
<400> 63
Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
1 5 10
<210> 64
<211> 30
<212> PRT
<213> Lama glama
<400> 64
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg
20 25 30
<210> 65
<211> 5
<212> PRT
<213> Lama glama
<400> 65
Asn Tyr His Met Ser
1 5
<210> 66
<211> 14
<212> PRT
<213> Lama glama
<400> 66
Trp Val Arg Gln Val Pro Gly Lys Gly Phe Glu Trp Ile Ser
1 5 10
<210> 67
<211> 17
<212> PRT
<213> Lama glama
<400> 67
Asp Ile Asn Ser Ala Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 68
<211> 32
<212> PRT
<213> Lama glama
<400> 68
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Glu
1 5 10 15
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Ala Arg
20 25 30
<210> 69
<211> 8
<212> PRT
<213> Lama glama
<400> 69
Val Asn Val Trp Gly Val Asn Tyr
1 5
<210> 70
<211> 11
<212> PRT
<213> Lama glama
<400> 70
Trp Gly Lys Gly Thr Leu Val Ser Val Ser Ser
1 5 10
<210> 71
<211> 30
<212> PRT
<213> Lama glama
<400> 71
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
20 25 30
<210> 72
<211> 5
<212> PRT
<213> Lama glama
<400> 72
Asn Tyr Val Met Ser
1 5
<210> 73
<211> 14
<212> PRT
<213> Lama glama
<400> 73
Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10
<210> 74
<211> 16
<212> PRT
<213> Lama glama
<400> 74
Asp Thr Asn Ser Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 75
<211> 32
<212> PRT
<213> Lama glama
<400> 75
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu Gln
1 5 10 15
Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Ala Arg
20 25 30
<210> 76
<211> 8
<212> PRT
<213> Lama glama
<400> 76
Ser Phe Phe Tyr Gly Met Asn Tyr
1 5
<210> 77
<211> 11
<212> PRT
<213> Lama glama
<400> 77
Trp Gly Lys Gly Thr Gln Val Thr Val Ser Ser
1 5 10
<210> 78
<211> 22
<212> PRT
<213> Lama glama
<400> 78
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys
20
<210> 79
<211> 14
<212> PRT
<213> Lama glama
<400> 79
Gly Leu Ser Ser Gly Ser Val Thr Thr Ser Asn Tyr Pro Gly
1 5 10
<210> 80
<211> 15
<212> PRT
<213> Lama glama
<400> 80
Trp Phe Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr Leu Ile Tyr
1 5 10 15
<210> 81
<211> 7
<212> PRT
<213> Lama glama
<400> 81
Asn Thr Asn Asn Arg His Ser
1 5
<210> 82
<211> 32
<212> PRT
<213> Lama glama
<400> 82
Gly Val Pro Ser Arg Phe Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala
1 5 10 15
Leu Thr Ile Thr Gly Ala Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys
20 25 30
<210> 83
<211> 10
<212> PRT
<213> Lama glama
<400> 83
Ser Leu Tyr Thr Gly Ser Tyr Thr Thr Val
1 5 10
<210> 84
<211> 10
<212> PRT
<213> Lama glama
<400> 84
Phe Gly Gly Gly Thr His Leu Thr Val Leu
1 5 10
<210> 85
<211> 22
<212> PRT
<213> Lama glama
<400> 85
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys
20
<210> 86
<211> 14
<212> PRT
<213> Lama glama
<400> 86
Gly Leu Ser Ser Gly Ser Val Thr Thr Ser Asn Tyr Pro Gly
1 5 10
<210> 87
<211> 15
<212> PRT
<213> Lama glama
<400> 87
Trp Phe Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr Leu Ile Tyr
1 5 10 15
<210> 88
<211> 7
<212> PRT
<213> Lama glama
<400> 88
Asn Thr Asn Ser Arg His Ser
1 5
<210> 89
<211> 32
<212> PRT
<213> Lama glama
<400> 89
Gly Val Pro Ser Arg Phe Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala
1 5 10 15
Leu Thr Ile Met Gly Ala Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys
20 25 30
<210> 90
<211> 9
<212> PRT
<213> Lama glama
<400> 90
Ser Leu Tyr Pro Gly Ser Thr Thr Val
1 5
<210> 91
<211> 10
<212> PRT
<213> Lama glama
<400> 91
Phe Gly Gly Gly Thr His Leu Thr Val Leu
1 5 10
<210> 92
<211> 22
<212> PRT
<213> Lama glama
<400> 92
Ser Tyr Glu Leu Thr Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys
20
<210> 93
<211> 11
<212> PRT
<213> Lama glama
<400> 93
Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala His
1 5 10
<210> 94
<211> 15
<212> PRT
<213> Lama glama
<400> 94
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
1 5 10 15
<210> 95
<211> 7
<212> PRT
<213> Lama glama
<400> 95
Asp Asp Asp Ser Arg Pro Ser
1 5
<210> 96
<211> 32
<212> PRT
<213> Lama glama
<400> 96
Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly Gly Thr Ala Thr
1 5 10 15
Leu Thr Ile Ser Gly Ala Gln Ala Glu Asp Glu Gly Asp Tyr Tyr Cys
20 25 30
<210> 97
<211> 11
<212> PRT
<213> Lama glama
<400> 97
Gln Ser Ala Asp Ser Ser Gly Asn Ala Ala Val
1 5 10
<210> 98
<211> 10
<212> PRT
<213> Lama glama
<400> 98
Phe Gly Gly Gly Thr His Leu Thr Val Leu
1 5 10
<210> 99
<211> 22
<212> PRT
<213> Lama glama
<400> 99
Ser Ser Ala Leu Thr Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys
20
<210> 100
<211> 11
<212> PRT
<213> Lama glama
<400> 100
Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala His
1 5 10
<210> 101
<211> 15
<212> PRT
<213> Lama glama
<400> 101
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
1 5 10 15
<210> 102
<211> 7
<212> PRT
<213> Lama glama
<400> 102
Asp Asp Asp Ser Arg Pro Ser
1 5
<210> 103
<211> 32
<212> PRT
<213> Lama glama
<400> 103
Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly Gly Thr Ala Thr
1 5 10 15
Leu Thr Ile Ser Gly Ala Gln Ala Glu Asp Glu Gly Asp Tyr Tyr Cys
20 25 30
<210> 104
<211> 11
<212> PRT
<213> Lama glama
<400> 104
Gln Ser Ala Asp Ser Ser Gly Asn Ala Ala Val
1 5 10
<210> 105
<211> 10
<212> PRT
<213> Lama glama
<400> 105
Phe Gly Gly Gly Thr His Leu Thr Val Leu
1 5 10
<210> 106
<211> 22
<212> PRT
<213> Lama glama
<400> 106
Gln Pro Val Leu Asn Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys
20
<210> 107
<211> 11
<212> PRT
<213> Lama glama
<400> 107
Gln Gly Gly Ser Leu Gly Ala Arg Tyr Ala His
1 5 10
<210> 108
<211> 15
<212> PRT
<213> Lama glama
<400> 108
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
1 5 10 15
<210> 109
<211> 7
<212> PRT
<213> Lama glama
<400> 109
Asp Asp Asp Ser Arg Pro Ser
1 5
<210> 110
<211> 32
<212> PRT
<213> Lama glama
<400> 110
Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly Gly Thr Ala Thr
1 5 10 15
Leu Thr Ile Ser Gly Ala Gln Ala Glu Asp Glu Gly Asp Tyr Tyr Cys
20 25 30
<210> 111
<211> 9
<212> PRT
<213> Lama glama
<400> 111
Gln Ser Ala Asp Ser Ser Gly Ser Val
1 5
<210> 112
<211> 10
<212> PRT
<213> Lama glama
<400> 112
Phe Gly Gly Gly Thr His Leu Thr Val Leu
1 5 10
<210> 113
<211> 22
<212> PRT
<213> Lama glama
<400> 113
Ser Tyr Glu Leu Thr Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys
20
<210> 114
<211> 11
<212> PRT
<213> Lama glama
<400> 114
Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala His
1 5 10
<210> 115
<211> 15
<212> PRT
<213> Lama glama
<400> 115
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
1 5 10 15
<210> 116
<211> 7
<212> PRT
<213> Lama glama
<400> 116
Asp Asp Asp Ser Arg Pro Ser
1 5
<210> 117
<211> 32
<212> PRT
<213> Lama glama
<400> 117
Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly Gly Thr Ala Thr
1 5 10 15
Leu Thr Ile Ser Gly Ala Gln Ala Glu Asp Glu Gly Asp Tyr Tyr Cys
20 25 30
<210> 118
<211> 11
<212> PRT
<213> Lama glama
<400> 118
Gln Ser Ala Asp Ser Ser Gly Asn Ala Ala Val
1 5 10
<210> 119
<211> 10
<212> PRT
<213> Lama glama
<400> 119
Phe Gly Gly Gly Thr His Leu Thr Val Leu
1 5 10
<210> 120
<211> 22
<212> PRT
<213> Lama glama
<400> 120
Ser Ser Ala Leu Thr Gln Pro Ser Ala Leu Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys
20
<210> 121
<211> 11
<212> PRT
<213> Lama glama
<400> 121
Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala His
1 5 10
<210> 122
<211> 15
<212> PRT
<213> Lama glama
<400> 122
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
1 5 10 15
<210> 123
<211> 7
<212> PRT
<213> Lama glama
<400> 123
Gly Asp Asp Ser Arg Pro Ser
1 5
<210> 124
<211> 32
<212> PRT
<213> Lama glama
<400> 124
Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly Gly Thr Ala Thr
1 5 10 15
Leu Thr Ile Ser Gly Ala Gln Ala Glu Asp Glu Asp Asp Tyr Tyr Cys
20 25 30
<210> 125
<211> 10
<212> PRT
<213> Lama glama
<400> 125
Gln Ser Thr Asp Ser Ser Gly Asn Thr Val
1 5 10
<210> 126
<211> 10
<212> PRT
<213> Lama glama
<400> 126
Phe Gly Gly Gly Thr Arg Leu Thr Val Leu
1 5 10
<210> 127
<211> 22
<212> PRT
<213> Lama glama
<400> 127
Gln Ala Gly Leu Thr Gln Pro Pro Ser Val Ser Gly Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys
20
<210> 128
<211> 14
<212> PRT
<213> Lama glama
<400> 128
Ala Gly Asn Ser Ser Asp Val Gly Tyr Gly Asn Tyr Val Ser
1 5 10
<210> 129
<211> 15
<212> PRT
<213> Lama glama
<400> 129
Trp Tyr Gln Gln Phe Pro Gly Met Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 130
<211> 7
<212> PRT
<213> Lama glama
<400> 130
Leu Val Asn Lys Arg Ala Ser
1 5
<210> 131
<211> 32
<212> PRT
<213> Lama glama
<400> 131
Gly Ile Thr Asp Arg Phe Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser
1 5 10 15
Leu Thr Ile Ser Gly Leu Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys
20 25 30
<210> 132
<211> 10
<212> PRT
<213> Lama glama
<400> 132
Ala Ser Tyr Thr Gly Ser Asn Asn Ile Val
1 5 10
<210> 133
<211> 10
<212> PRT
<213> Lama glama
<400> 133
Phe Gly Gly Gly Thr His Leu Thr Val Leu
1 5 10
<210> 134
<211> 23
<212> PRT
<213> Lama glama
<400> 134
Glu Ile Val Leu Thr Gln Ser Pro Ser Ser Val Thr Ala Ser Val Gly
1 5 10 15
Gly Lys Val Thr Ile Asn Cys
20
<210> 135
<211> 17
<212> PRT
<213> Lama glama
<400> 135
Lys Ser Ser Gln Ser Val Phe Ile Ala Ser Asn Gln Lys Thr Tyr Leu
1 5 10 15
Asn
<210> 136
<211> 15
<212> PRT
<213> Lama glama
<400> 136
Trp Tyr Gln Gln Arg Pro Gly Gln Ser Pro Arg Leu Val Ile Ser
1 5 10 15
<210> 137
<211> 7
<212> PRT
<213> Lama glama
<400> 137
Tyr Ala Ser Thr Arg Glu Ser
1 5
<210> 138
<211> 32
<212> PRT
<213> Lama glama
<400> 138
Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Thr Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Val Gln Pro Glu Asp Ala Ala Val Tyr Tyr Cys
20 25 30
<210> 139
<211> 8
<212> PRT
<213> Lama glama
<400> 139
Gln Gln Ala Tyr Ser His Pro Thr
1 5
<210> 140
<211> 10
<212> PRT
<213> Lama glama
<400> 140
Phe Gly Gln Gly Thr Lys Val Glu Leu Lys
1 5 10
<210> 141
<211> 22
<212> PRT
<213> Lama glama
<400> 141
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys
20
<210> 142
<211> 14
<212> PRT
<213> Lama glama
<400> 142
Gly Leu Ser Ser Gly Ser Val Thr Thr Ser Asn Tyr Pro Gly
1 5 10
<210> 143
<211> 15
<212> PRT
<213> Lama glama
<400> 143
Trp Phe Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr Leu Ile Tyr
1 5 10 15
<210> 144
<211> 7
<212> PRT
<213> Lama glama
<400> 144
Asn Thr Asn Ser Arg His Ser
1 5
<210> 145
<211> 32
<212> PRT
<213> Lama glama
<400> 145
Gly Val Pro Ser Arg Phe Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala
1 5 10 15
Leu Thr Ile Thr Gly Ala Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys
20 25 30
<210> 146
<211> 10
<212> PRT
<213> Lama glama
<400> 146
Ser Leu Tyr Pro Gly Ser Tyr Thr Asn Val
1 5 10
<210> 147
<211> 10
<212> PRT
<213> Lama glama
<400> 147
Phe Gly Gly Gly Thr His Leu Thr Val Leu
1 5 10
<210> 148
<211> 22
<212> PRT
<213> Lama glama
<400> 148
Gln Ser Ala Leu Thr Gln Pro Pro Ser Leu Ser Ala Ser Pro Gly Ser
1 5 10 15
Ser Val Arg Leu Thr Cys
20
<210> 149
<211> 14
<212> PRT
<213> Lama glama
<400> 149
Thr Leu Ser Ser Gly Asn Asn Ile Gly Ser Tyr Asp Ile Ser
1 5 10
<210> 150
<211> 15
<212> PRT
<213> Lama glama
<400> 150
Trp Tyr Gln Gln Lys Ala Gly Ser Pro Pro Arg Tyr Leu Leu Asn
1 5 10 15
<210> 151
<211> 11
<212> PRT
<213> Lama glama
<400> 151
Tyr Tyr Thr Asp Ser Arg Lys His Gln Asp Ser
1 5 10
<210> 152
<211> 34
<212> PRT
<213> Lama glama
<400> 152
Gly Val Pro Ser Arg Phe Ser Gly Ser Lys Asp Ala Ser Ala Asn Ala
1 5 10 15
Gly Leu Leu Leu Ile Ser Gly Leu Gln Pro Glu Asp Glu Ala Asp Tyr
20 25 30
Tyr Cys
<210> 153
<211> 11
<212> PRT
<213> Lama glama
<400> 153
Ser Ala Tyr Lys Ser Gly Ser Tyr Arg Trp Val
1 5 10
<210> 154
<211> 10
<212> PRT
<213> Lama glama
<400> 154
Phe Gly Gly Gly Thr His Val Thr Val Leu
1 5 10
<210> 155
<211> 118
<212> PRT
<213> Lama glama
<400> 155
Gln Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Val Ser Cys Thr Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asn Ser Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Val
85 90 95
Arg Val Arg Ile Trp Pro Val Gly Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Gln Val Thr Val Ser Ser
115
<210> 156
<211> 110
<212> PRT
<213> Lama glama
<400> 156
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Thr Thr Ser
20 25 30
Asn Tyr Pro Gly Trp Phe Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr
35 40 45
Leu Ile Tyr Asn Thr Asn Asn Arg His Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Leu Tyr Thr Gly Ser
85 90 95
Tyr Thr Thr Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 157
<211> 118
<212> PRT
<213> Lama glama
<400> 157
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Val Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Arg Thr Asp Gly Gly Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Met Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Ala
85 90 95
Arg Thr Arg Ile Phe Pro Ser Gly Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Gln Val Thr Val Ser Ser
115
<210> 158
<211> 109
<212> PRT
<213> Lama glama
<400> 158
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Thr Thr Ser
20 25 30
Asn Tyr Pro Gly Trp Phe Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr
35 40 45
Leu Ile Tyr Asn Thr Asn Ser Arg His Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala Leu Thr Ile Met Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Leu Tyr Pro Gly Ser
85 90 95
Thr Thr Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105
<210> 159
<211> 126
<212> PRT
<213> Lama glama
<400> 159
Gln Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asn Ser Gly Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Glu Leu Arg Phe Asp Leu Ala Arg Tyr Thr Asp Tyr Glu Ala
100 105 110
Trp Asp Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 160
<211> 108
<212> PRT
<213> Lama glama
<400> 160
Ser Tyr Glu Leu Thr Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Asn Ala
85 90 95
Ala Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105
<210> 161
<211> 126
<212> PRT
<213> Lama glama
<400> 161
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asn Ser Gly Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Met Arg Leu Tyr Leu Ala Arg Tyr Asn Asp Tyr Glu Ala
100 105 110
Trp Asp Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 162
<211> 108
<212> PRT
<213> Lama glama
<400> 162
Ser Ser Ala Leu Thr Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Asn Ala
85 90 95
Ala Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105
<210> 163
<211> 126
<212> PRT
<213> Lama glama
<400> 163
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asn Ser Tyr Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Glu Val Arg Ala Asp Leu Ser Arg Tyr Asn Asp Tyr Glu Ser
100 105 110
Tyr Asp Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 164
<211> 106
<212> PRT
<213> Lama glama
<400> 164
Gln Pro Val Leu Asn Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys Gln Gly Gly Ser Leu Gly Ala Arg Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Ser Val
85 90 95
Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105
<210> 165
<211> 126
<212> PRT
<213> Lama glama
<400> 165
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Lys Asp Tyr
20 25 30
Asp Ile Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Thr Ser Arg Ser Gly Ser Thr Ser Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gly Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Tyr Ala Thr Thr Trp Asp Val Gly Pro Leu Gly Tyr Gly
100 105 110
Met Asp Tyr Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 166
<211> 108
<212> PRT
<213> Lama glama
<400> 166
Ser Tyr Glu Leu Thr Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Asn Ala
85 90 95
Ala Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105
<210> 167
<211> 126
<212> PRT
<213> Lama glama
<400> 167
Glu Val Gln Leu Gln Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ile Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Asn Ser Asp Gly Ser Ser Thr Ser Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Tyr Gly Ser Thr Trp Asp Val Gly Pro Met Gly Tyr Gly
100 105 110
Met Asp Tyr Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 168
<211> 107
<212> PRT
<213> Lama glama
<400> 168
Ser Ser Ala Leu Thr Gln Pro Ser Ala Leu Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Asp Asp Tyr Tyr Cys Gln Ser Thr Asp Ser Ser Gly Asn Thr
85 90 95
Val Phe Gly Gly Gly Thr Arg Leu Thr Val Leu
100 105
<210> 169
<211> 119
<212> PRT
<213> Lama glama
<400> 169
Gln Val Gln Leu Val Glu Ser Gly Gly Asn Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Tyr Ser Asp Gly Ser Thr Thr Trp Tyr Ser Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Lys Ile Tyr Pro Gly Gly Tyr Asp Ala Trp Gly Gln Gly
100 105 110
Thr Gln Val Thr Val Ser Ser
115
<210> 170
<211> 110
<212> PRT
<213> Lama glama
<400> 170
Gln Ala Gly Leu Thr Gln Pro Pro Ser Val Ser Gly Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Ala Gly Asn Ser Ser Asp Val Gly Tyr Gly
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln Phe Pro Gly Met Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Leu Val Asn Lys Arg Ala Ser Gly Ile Thr Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Tyr Thr Gly Ser
85 90 95
Asn Asn Ile Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 171
<211> 118
<212> PRT
<213> Lama glama
<400> 171
Gln Val Gln Leu Gln Glu Ser Gly Gly Asp Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Val Ser Cys Val Val Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asp Ser Tyr Gly Tyr Ser Thr Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Ala Lys Thr Thr Trp Ser Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Gln Val Thr Val Ser Ser
115
<210> 172
<211> 112
<212> PRT
<213> Lama glama
<400> 172
Glu Ile Val Leu Thr Gln Ser Pro Ser Ser Val Thr Ala Ser Val Gly
1 5 10 15
Gly Lys Val Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Phe Ile Ala
20 25 30
Ser Asn Gln Lys Thr Tyr Leu Asn Trp Tyr Gln Gln Arg Pro Gly Gln
35 40 45
Ser Pro Arg Leu Val Ile Ser Tyr Ala Ser Thr Arg Glu Ser Gly Ile
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Thr Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Pro Glu Asp Ala Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Ala Tyr Ser His Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Leu Lys
100 105 110
<210> 173
<211> 117
<212> PRT
<213> Lama glama
<400> 173
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Asn Tyr
20 25 30
His Met Ser Trp Val Arg Gln Val Pro Gly Lys Gly Phe Glu Trp Ile
35 40 45
Ser Asp Ile Asn Ser Ala Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Val Asn Val Trp Gly Val Asn Tyr Trp Gly Lys Gly Thr Leu
100 105 110
Val Ser Val Ser Ser
115
<210> 174
<211> 110
<212> PRT
<213> Lama glama
<400> 174
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Thr Thr Ser
20 25 30
Asn Tyr Pro Gly Trp Phe Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr
35 40 45
Leu Ile Tyr Asn Thr Asn Ser Arg His Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Leu Tyr Pro Gly Ser
85 90 95
Tyr Thr Asn Val Phe Gly Gly Gly Thr His Leu Thr Val Leu
100 105 110
<210> 175
<211> 116
<212> PRT
<213> Lama glama
<400> 175
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Val Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Thr Asn Ser Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Ala
85 90 95
Arg Ser Phe Phe Tyr Gly Met Asn Tyr Trp Gly Lys Gly Thr Gln Val
100 105 110
Thr Val Ser Ser
115
<210> 176
<211> 117
<212> PRT
<213> Lama glama
<400> 176
Gln Ser Ala Leu Thr Gln Pro Pro Ser Leu Ser Ala Ser Pro Gly Ser
1 5 10 15
Ser Val Arg Leu Thr Cys Thr Leu Ser Ser Gly Asn Asn Ile Gly Ser
20 25 30
Tyr Asp Ile Ser Trp Tyr Gln Gln Lys Ala Gly Ser Pro Pro Arg Tyr
35 40 45
Leu Leu Asn Tyr Tyr Thr Asp Ser Arg Lys His Gln Asp Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Lys Asp Ala Ser Ala Asn Ala Gly Leu
65 70 75 80
Leu Leu Ile Ser Gly Leu Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys
85 90 95
Ser Ala Tyr Lys Ser Gly Ser Tyr Arg Trp Val Phe Gly Gly Gly Thr
100 105 110
His Val Thr Val Leu
115
<210> 177
<211> 354
<212> DNA
<213> Lama glama
<400> 177
cagttgcagc tggtggagtc tgggggaggc ttggtgcagc ctggggggtc tctgagagtt 60
tcctgtacag cctctggatt caccttcaat acctactaca tgacctgggt ccgccaggct 120
ccagggaagg ggctcgagtg ggtctcagat attaatagtg gtggtggtac atactatgca 180
gactccgtga agggccgatt caccatctcc agagacaacg ccaagaacac gctatatctg 240
caaatgaaca gcctgaaacc tgaggacacg gccctgtatt actgtgtaag agttcgtatt 300
tggccagtgg gatatgacta ctggggccag gggacccagg tcaccgtttc ctca 354
<210> 178
<211> 330
<212> DNA
<213> Lama glama
<400> 178
caggctgtgg tgacccagga gccgtccctg tcagtgtctc caggagggac ggtcacactc 60
acctgcggcc tcagctctgg gtctgtcact accagtaact accctggttg gttccagcag 120
acaccgggcc aggctccacg cactcttatc tacaacacaa acaaccgcca ctctggggtc 180
cccagtcgct tctccggatc catctctggg aacaaagccg ccctcaccat cacgggggcc 240
cagcccgagg acgaggccga ctattactgt tctctatata ctggcagtta cactactgtg 300
ttcggcggag ggacccatct gaccgtcctg 330
<210> 179
<211> 354
<212> DNA
<213> Lama glama
<400> 179
caggtgcagc tcgtggagtc tgggggaggc ttggtgcagc ctggggggtc tctgagagtc 60
tcctgtgcag cctctggatt caccttcagt acctactaca tgagctgggt ccgccaggct 120
ccagggaagg ggctcgagtg ggtctcagat attcgtactg atggtggcac atactatgca 180
gactccgtga agggccgatt caccatgtcc agagacaacg ccaagaacac gctgtatcta 240
caaatgaaca gcctgaaacc tgaggacacg gccctgtatt actgtgcaag aactcgaatt 300
ttcccctcgg ggtatgacta ctggggccag gggacccagg tcaccgtctc ctca 354
<210> 180
<211> 327
<212> DNA
<213> Lama glama
<400> 180
caggctgtgg tgacccagga gccgtccctg tcagtgtctc caggagggac ggtcacactc 60
acctgcggcc tcagctctgg gtctgtcact accagtaact accctggttg gttccagcag 120
acaccaggcc aggctccgcg cactcttatc tacaacacaa acagccgcca ctctggggtc 180
cccagtcgct tctccggatc catctctggg aacaaagccg ccctcaccat catgggggcc 240
cagcccgagg acgaggccga ctattactgt tctctgtacc ctggtagtac cactgtgttc 300
ggcggaggga cccatctgac cgtcctg 327
<210> 181
<211> 378
<212> DNA
<213> Lama glama
<400> 181
cagttgcagc tggtggagtc tgggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt agccatgcca tgagctgggt ccgccaggct 120
ccaggaaagg ggctcgagtg ggtctcagct attaatagtg gtggtggtag cacaagctat 180
gcagactccg tgaagggccg attcaccatc tccagagaca acgccaagaa cacgctgtac 240
ctgcaaatga acagcctgaa acctgaggac acggccgtgt attactgtgc aaaagagctg 300
agattcgacc tagcaaggta taccgactat gaggcctggg actactgggg ccaggggacc 360
caggtcaccg tctcctca 378
<210> 182
<211> 324
<212> DNA
<213> Lama glama
<400> 182
tcctatgagc tgactcagcc ctccgcgctg tccgtaacct tgggacagac ggccaagatc 60
acctgccaag gtggcagctt aggtagcagt tatgctcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatgatgat gacagcaggc cctcagggat ccctgagcgg 180
ttctctggct ccagctctgg gggcacagcc accctgacca tcagcggggc ccaggccgag 240
gacgagggtg actattactg tcagtcagca gacagcagtg gtaatgctgc tgtgttcggc 300
ggagggaccc atctgaccgt cctg 324
<210> 183
<211> 378
<212> DNA
<213> Lama glama
<400> 183
gagttgcagc tggtggagtc tgggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt ggctatggca tgagctgggt ccgccaggct 120
ccaggaaagg ggctcgagtg ggtctcagat attaatagtg gtggtggtag cacaagctat 180
gcagactccg tgaagggccg attcaccatc tccagagaca acgccaagaa cacgctgtat 240
ctgcaaatga acagcctgaa acctgaggac acggccgtgt attactgtgc aaaagatatg 300
agattatacc tagcaaggta taacgactat gaggcctggg actactgggg ccaggggacc 360
caggtcaccg tctcctca 378
<210> 184
<211> 324
<212> DNA
<213> Lama glama
<400> 184
tcctctgcac tgactcagcc ctccgcgctg tccgtaacct tgggacagac ggccaagatc 60
acctgccaag gtggcagctt aggtagcagt tatgctcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatgatgat gacagcaggc cctcagggat ccctgagcgg 180
ttctctggct ccagctctgg gggcacagcc accctgacca tcagcggggc ccaggccgag 240
gacgagggtg actattactg tcagtcagca gacagcagtg gtaatgctgc tgtgttcggc 300
ggagggaccc atctgaccgt cctg 324
<210> 185
<211> 378
<212> DNA
<213> Lama glama
<400> 185
gagttgcagc tggtggagtc tgggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatggca tgagctgggt ccgccaggct 120
ccaggaaagg ggctcgagtg ggtctcagct attaatagtt atggtggtag cacaagctat 180
gcagactccg tgaagggccg attcaccatc tccagagaca acgccaagaa cacgctgtat 240
ctgcaaatga acagcctgaa acctgaggac acggccgtgt attactgtgc aaaagaagtg 300
cgggccgacc taagccgcta taacgactat gagtcgtatg actactgggg ccaggggacc 360
caggtcaccg tctcctca 378
<210> 186
<211> 318
<212> DNA
<213> Lama glama
<400> 186
cagccggtgc tgaatcagcc ctccgcgctg tccgtaacct tgggacagac ggccaagatc 60
acctgccaag gtggcagctt aggtgcgcgt tatgctcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatgatgat gacagcaggc cctcagggat ccctgagcgg 180
ttctctggct ccagctctgg gggcacagcc accctgacca tcagcggggc ccaggccgag 240
gacgagggtg actattactg tcagtcagca gacagcagtg gttctgtgtt cggcggaggg 300
acccatctga ccgtcctg 318
<210> 187
<211> 378
<212> DNA
<213> Lama glama
<400> 187
gaggtgcagc tcgtggagtc tgggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt cagcttcaag gactatgaca taacctgggt ccgccaggct 120
ccgggaaagg ggctcgagtg ggtctcaact attactagtc gtagtggtag cacaagctat 180
gtagactccg taaagggccg attcaccatc tccggagaca acgccaagaa cacgctgtat 240
ctgcaaatga acagcctgaa acctgaggac acggccgtgt attactgtgc aaaagtttac 300
gcgactacct gggacgtcgg ccctctgggc tacggcatgg actactgggg caaggggacc 360
ctggtcaccg tctcctca 378
<210> 188
<211> 324
<212> DNA
<213> Lama glama
<400> 188
tcctatgagc tgactcagcc ctccgcgctg tccgtaacct tgggacagac ggccaagatc 60
acctgccaag gtggcagctt aggtagcagt tatgctcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatgatgat gacagcaggc cctcagggat ccctgagcgg 180
ttctctggct ccagctctgg gggcacagcc accctgacca tcagcggggc ccaggccgag 240
gacgagggtg actattactg tcagtcagca gacagcagtg gtaatgctgc tgtgttcggc 300
ggagggaccc atctgaccgt cctg 324
<210> 189
<211> 378
<212> DNA
<213> Lama glama
<400> 189
gaggtgcagc tgcaggagtc ggggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt atatatgaca tgagctgggt ccgccaggct 120
ccaggaaagg ggctcgagtg ggtctcaact attaatagtg atggtagtag cacaagctat 180
gtagactccg tgaagggccg attcaccatc tccagagaca acgccaagaa cacgctgtat 240
ctgcaaatga acagcctgaa acctgaggac acggccgtgt attactgtgc gaaagtttac 300
ggtagtacct gggacgtcgg ccctatgggc tacggcatgg actactgggg caaagggacc 360
ctggtcactg tctcctca 378
<210> 190
<211> 321
<212> DNA
<213> Lama glama
<400> 190
tcctctgcac tgactcagcc ctccgcgctg tccgtgtcct tgggacagac ggccaggatc 60
acctgccaag gtggcagctt aggtagcagt tatgctcact ggtaccagca gaagccaggc 120
caggcccctg tgctggtcat ctatggtgat gacagcaggc cctcagggat ccctgagcgg 180
ttctctggct ccagctctgg gggcacagcc accctgacca tcagcggggc ccaggccgag 240
gacgaggatg actattactg tcagtcaaca gacagcagtg gtaatactgt gttcggcgga 300
gggacccgac tgaccgtcct g 321
<210> 191
<211> 357
<212> DNA
<213> Lama glama
<400> 191
caggtgcagc tggtggagtc tgggggaaac ttggtgcagc ctgggggttc tctgagactc 60
tcctgtgcag cctctggatt caccttcagt aactactaca tgagctgggt ccgccaggct 120
ccagggaagg ggctggaatg ggtgtccgat atttatagtg acggtagtac cacatggtat 180
tcagactccg tcaagggccg attcaccatc tccagagaca acgccaagaa cacgctgtct 240
ctgcaaatga acagtctgaa atctgaggac acggccgtct attactgtgc gcgcgtgaag 300
atctatccgg gggggtatga cgcctggggc caggggaccc aggtcaccgt ctcctca 357
<210> 192
<211> 330
<212> DNA
<213> Lama glama
<400> 192
caggcagggc tgactcagcc tccctccgtg tctgggtctc caggaaagac ggtcaccatc 60
tcctgtgcag gaaacagcag tgatgttggg tatggaaact atgtctcctg gtaccagcag 120
ttcccaggaa tggcccccaa actcctgata tatctcgtca ataaacgggc ctcagggatc 180
actgatcgct tctctggctc caagtcaggc aacacggcct ccctgaccat ctctgggctc 240
cagtctgagg acgaggctga ttattactgt gcctcatata caggtagcaa caatatcgtg 300
ttcggcggag ggacccatct aaccgtcctc 330
<210> 193
<211> 354
<212> DNA
<213> Lama glama
<400> 193
caggtgcagc tgcaggagtc ggggggagac ttggtgcagc ctggggggtc tctgagagtc 60
tcctgtgtag tctctggatt caccttcagt cgctactaca tgagctgggt ccgccaggct 120
ccagggaagg ggctcgagtg ggtctcatct attgatagtt atggttacag cacatactat 180
acagactccg tgaagggccg attcaccatc tccagagaca acgccaagaa cacgctgtat 240
ctgcaaatga acagcctgaa acctgaggac acggccctgt attactgtgc aagagcgaaa 300
acgacttgga gttatgacta ctggggccag gggacccagg tcaccgtctc ctca 354
<210> 194
<211> 336
<212> DNA
<213> Lama glama
<400> 194
gaaattgtgt tgacgcagtc tcccagctcc gtgactgcat ctgtaggagg gaaggtcact 60
atcaactgta agtccagcca gagcgtcttc atagcttcta atcagaaaac ctacttaaac 120
tggtaccagc agagacctgg acagtctccg aggttggtca tcagctatgc gtccacccgt 180
gaatcgggga tccctgatcg attcagcggc agtgggtcca caacagattt cactctcacg 240
atcagcagtg tccagcctga agatgcggcc gtgtattact gtcagcaggc ttatagccat 300
ccaacgttcg gccaggggac caaggtggaa ctcaaa 336
<210> 195
<211> 351
<212> DNA
<213> Lama glama
<400> 195
gaggtgcagc tcgtggagtc tgggggaggc ttggtgcaac ctgggggttc tctgagactc 60
tcctgtgcag cctctggatt caccttcagg aattaccaca tgagttgggt ccgccaggtt 120
ccagggaagg ggttcgagtg gatctcagat attaatagtg caggtggtag cacatactat 180
gcagactccg tgaagggccg attcaccatc tccagagaca acgccaagaa cacgctgtat 240
ctggaaatga acagcctgaa acctgaggac acggccctgt attactgtgc aagagtcaac 300
gtctgggggg tgaactactg gggcaaaggg accctggtca gcgtctcctc a 351
<210> 196
<211> 330
<212> DNA
<213> Lama glama
<400> 196
cagactgtgg tgactcagga gccgtccctg tcagtgtctc caggagggac ggtcacactc 60
acctgcggcc tcagctctgg gtctgtcact accagtaact accctggttg gttccagcag 120
acaccaggcc aggctccacg cactcttatc tacaacacaa acagccgcca ctctggggtc 180
cccagtcgct tctccggatc catctctggg aacaaagccg ccctcaccat cacgggggcc 240
cagcccgagg acgaggccga ctattactgt tctctgtacc ctggtagtta cactaatgtg 300
ttcggcggag ggacccatct gaccgtcctg 330
<210> 197
<211> 348
<212> DNA
<213> Lama glama
<400> 197
gagttgcagc tggtggagtc tgggggaggc ttggtgcagc ctggggggtc tctgagactc 60
tcctgtgcag cctctggatt caccttcagc aactatgtca tgagctgggt ccgccaggct 120
ccaggaaagg ggctcgagtg ggtctcagat actaatagtg gtggtagcac aagctatgca 180
gactccgtga agggccgatt caccatctct agagacaacg ccaagaacac gctgtatttg 240
caaatgaaca gcctgaaacc tgaggacacg gcattgtatt actgtgcgag atcatttttc 300
tacggcatga actactgggg caaagggacc caggtcaccg tgtcctca 348
<210> 198
<211> 351
<212> DNA
<213> Lama glama
<400> 198
cagtctgccc tgactcagcc gccctccctc tctgcatctc cgggatcatc tgtcagactc 60
acctgcaccc tgagcagtgg aaacaatatt ggcagctatg acataagttg gtaccagcag 120
aaggcaggga gccctccccg gtacctcctg aactactaca ccgactcacg caagcaccag 180
gactccgggg tcccgagccg cttctctggg tccaaagatg cctcggccaa cgcagggctt 240
ctgctcatct ctgggcttca gcccgaggac gaggctgact attactgttc tgcatacaag 300
agtggttctt accgttgggt gttcggcgga gggacgcacg tgaccgtcct g 351
<210> 199
<211> 450
<212> PRT
<213> Lama glama
<400> 199
Gln Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Val Ser Cys Thr Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Tyr Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asn Ser Gly Gly Gly Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Val
85 90 95
Arg Val Arg Ile Trp Pro Val Gly Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Gln Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Ser Glu Val Phe Val Pro Gln Ser Arg Lys
435 440 445
Val Ile
450
<210> 200
<211> 215
<212> PRT
<213> Lama glama
<400> 200
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Thr Thr Ser
20 25 30
Asn Tyr Pro Gly Trp Phe Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr
35 40 45
Leu Ile Tyr Asn Thr Asn Asn Arg His Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Leu Tyr Thr Gly Ser
85 90 95
Tyr Thr Thr Val Phe Gly Gly Gly Thr His Leu Thr Val Leu Gln Pro
100 105 110
Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys Ser
210 215
<210> 201
<211> 450
<212> PRT
<213> Lama glama
<400> 201
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Val Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Arg Thr Asp Gly Gly Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Met Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Ala
85 90 95
Arg Thr Arg Ile Phe Pro Ser Gly Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Gln Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Ser Glu Val Phe Val Pro Gln Ser Arg Lys
435 440 445
Val Ile
450
<210> 202
<211> 214
<212> PRT
<213> Lama glama
<400> 202
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Thr Thr Ser
20 25 30
Asn Tyr Pro Gly Trp Phe Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr
35 40 45
Leu Ile Tyr Asn Thr Asn Ser Arg His Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala Leu Thr Ile Met Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Leu Tyr Pro Gly Ser
85 90 95
Thr Thr Val Phe Gly Gly Gly Thr His Leu Thr Val Leu Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 203
<211> 458
<212> PRT
<213> Lama glama
<400> 203
Gln Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asn Ser Gly Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Glu Leu Arg Phe Asp Leu Ala Arg Tyr Thr Asp Tyr Glu Ala
100 105 110
Trp Asp Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Ser Glu
435 440 445
Val Phe Val Pro Gln Ser Arg Lys Val Ile
450 455
<210> 204
<211> 213
<212> PRT
<213> Lama glama
<400> 204
Ser Tyr Glu Leu Thr Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Asn Ala
85 90 95
Ala Val Phe Gly Gly Gly Thr His Leu Thr Val Leu Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 205
<211> 458
<212> PRT
<213> Lama glama
<400> 205
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Gly Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asn Ser Gly Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Met Arg Leu Tyr Leu Ala Arg Tyr Asn Asp Tyr Glu Ala
100 105 110
Trp Asp Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Ser Glu
435 440 445
Val Phe Val Pro Gln Ser Arg Lys Val Ile
450 455
<210> 206
<211> 213
<212> PRT
<213> Lama glama
<400> 206
Ser Ser Ala Leu Thr Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Asn Ala
85 90 95
Ala Val Phe Gly Gly Gly Thr His Leu Thr Val Leu Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 207
<211> 458
<212> PRT
<213> Lama glama
<400> 207
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asn Ser Tyr Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Glu Val Arg Ala Asp Leu Ser Arg Tyr Asn Asp Tyr Glu Ser
100 105 110
Tyr Asp Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Ser Glu
435 440 445
Val Phe Val Pro Gln Ser Arg Lys Val Ile
450 455
<210> 208
<211> 211
<212> PRT
<213> Lama glama
<400> 208
Gln Pro Val Leu Asn Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys Gln Gly Gly Ser Leu Gly Ala Arg Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Ser Val
85 90 95
Phe Gly Gly Gly Thr His Leu Thr Val Leu Gln Pro Lys Ala Ala Pro
100 105 110
Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn Lys
115 120 125
Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val Thr
130 135 140
Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu Thr
145 150 155 160
Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser Tyr
165 170 175
Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser Cys
180 185 190
Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro Thr
195 200 205
Glu Cys Ser
210
<210> 209
<211> 458
<212> PRT
<213> Lama glama
<400> 209
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Lys Asp Tyr
20 25 30
Asp Ile Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Thr Ser Arg Ser Gly Ser Thr Ser Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Gly Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Tyr Ala Thr Thr Trp Asp Val Gly Pro Leu Gly Tyr Gly
100 105 110
Met Asp Tyr Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Ser Glu
435 440 445
Val Phe Val Pro Gln Ser Arg Lys Val Ile
450 455
<210> 210
<211> 213
<212> PRT
<213> Lama glama
<400> 210
Ser Tyr Glu Leu Thr Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Asn Ala
85 90 95
Ala Val Phe Gly Gly Gly Thr His Leu Thr Val Leu Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 211
<211> 458
<212> PRT
<213> Lama glama
<400> 211
Glu Val Gln Leu Gln Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ile Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Asn Ser Asp Gly Ser Ser Thr Ser Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Tyr Gly Ser Thr Trp Asp Val Gly Pro Met Gly Tyr Gly
100 105 110
Met Asp Tyr Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
130 135 140
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
195 200 205
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
210 215 220
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
225 230 235 240
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
245 250 255
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
260 265 270
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
275 280 285
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
290 295 300
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
305 310 315 320
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
325 330 335
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
340 345 350
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
355 360 365
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
370 375 380
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
385 390 395 400
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
405 410 415
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
420 425 430
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Ser Glu
435 440 445
Val Phe Val Pro Gln Ser Arg Lys Val Ile
450 455
<210> 212
<211> 212
<212> PRT
<213> Lama glama
<400> 212
Ser Ser Ala Leu Thr Gln Pro Ser Ala Leu Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Asp Asp Tyr Tyr Cys Gln Ser Thr Asp Ser Ser Gly Asn Thr
85 90 95
Val Phe Gly Gly Gly Thr Arg Leu Thr Val Leu Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 213
<211> 451
<212> PRT
<213> Lama glama
<400> 213
Gln Val Gln Leu Val Glu Ser Gly Gly Asn Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Tyr Ser Asp Gly Ser Thr Thr Trp Tyr Ser Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Lys Ile Tyr Pro Gly Gly Tyr Asp Ala Trp Gly Gln Gly
100 105 110
Thr Gln Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Ser Glu Val Phe Val Pro Gln Ser Arg
435 440 445
Lys Val Ile
450
<210> 214
<211> 215
<212> PRT
<213> Lama glama
<400> 214
Gln Ala Gly Leu Thr Gln Pro Pro Ser Val Ser Gly Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Ala Gly Asn Ser Ser Asp Val Gly Tyr Gly
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln Phe Pro Gly Met Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Leu Val Asn Lys Arg Ala Ser Gly Ile Thr Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ser Tyr Thr Gly Ser
85 90 95
Asn Asn Ile Val Phe Gly Gly Gly Thr His Leu Thr Val Leu Gln Pro
100 105 110
Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys Ser
210 215
<210> 215
<211> 450
<212> PRT
<213> Lama glama
<400> 215
Gln Val Gln Leu Gln Glu Ser Gly Gly Asp Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Val Ser Cys Val Val Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asp Ser Tyr Gly Tyr Ser Thr Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Ala Lys Thr Thr Trp Ser Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Gln Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Ser Glu Val Phe Val Pro Gln Ser Arg Lys
435 440 445
Val Ile
450
<210> 216
<211> 219
<212> PRT
<213> Lama glama
<400> 216
Glu Ile Val Leu Thr Gln Ser Pro Ser Ser Val Thr Ala Ser Val Gly
1 5 10 15
Gly Lys Val Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Phe Ile Ala
20 25 30
Ser Asn Gln Lys Thr Tyr Leu Asn Trp Tyr Gln Gln Arg Pro Gly Gln
35 40 45
Ser Pro Arg Leu Val Ile Ser Tyr Ala Ser Thr Arg Glu Ser Gly Ile
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Thr Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Pro Glu Asp Ala Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Ala Tyr Ser His Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Leu Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 217
<211> 449
<212> PRT
<213> Lama glama
<400> 217
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Asn Tyr
20 25 30
His Met Ser Trp Val Arg Gln Val Pro Gly Lys Gly Phe Glu Trp Ile
35 40 45
Ser Asp Ile Asn Ser Ala Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Glu Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Arg Val Asn Val Trp Gly Val Asn Tyr Trp Gly Lys Gly Thr Leu
100 105 110
Val Ser Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Ser Glu Val Phe Val Pro Gln Ser Arg Lys Val
435 440 445
Ile
<210> 218
<211> 215
<212> PRT
<213> Lama glama
<400> 218
Gln Thr Val Val Thr Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Thr Thr Ser
20 25 30
Asn Tyr Pro Gly Trp Phe Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr
35 40 45
Leu Ile Tyr Asn Thr Asn Ser Arg His Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala Leu Thr Ile Thr Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Leu Tyr Pro Gly Ser
85 90 95
Tyr Thr Asn Val Phe Gly Gly Gly Thr His Leu Thr Val Leu Gln Pro
100 105 110
Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro
130 135 140
Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala
145 150 155 160
Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala
165 170 175
Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg
180 185 190
Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr
195 200 205
Val Ala Pro Thr Glu Cys Ser
210 215
<210> 219
<211> 448
<212> PRT
<213> Lama glama
<400> 219
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Val Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Thr Asn Ser Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Ala
85 90 95
Arg Ser Phe Phe Tyr Gly Met Asn Tyr Trp Gly Lys Gly Thr Gln Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Ser Glu Val Phe Val Pro Gln Ser Arg Lys Val Ile
435 440 445
<210> 220
<211> 222
<212> PRT
<213> Lama glama
<400> 220
Gln Ser Ala Leu Thr Gln Pro Pro Ser Leu Ser Ala Ser Pro Gly Ser
1 5 10 15
Ser Val Arg Leu Thr Cys Thr Leu Ser Ser Gly Asn Asn Ile Gly Ser
20 25 30
Tyr Asp Ile Ser Trp Tyr Gln Gln Lys Ala Gly Ser Pro Pro Arg Tyr
35 40 45
Leu Leu Asn Tyr Tyr Thr Asp Ser Arg Lys His Gln Asp Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Lys Asp Ala Ser Ala Asn Ala Gly Leu
65 70 75 80
Leu Leu Ile Ser Gly Leu Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys
85 90 95
Ser Ala Tyr Lys Ser Gly Ser Tyr Arg Trp Val Phe Gly Gly Gly Thr
100 105 110
His Val Thr Val Leu Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe
115 120 125
Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys
130 135 140
Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala
145 150 155 160
Asp Ser Ser Pro Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys
165 170 175
Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro
180 185 190
Glu Gln Trp Lys Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu
195 200 205
Gly Ser Thr Val Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215 220
<210> 221
<211> 442
<212> PRT
<213> Lama glama
<400> 221
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Val Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Arg Thr Asp Gly Gly Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Met Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Ala
85 90 95
Arg Thr Arg Ile Phe Pro Ser Gly Tyr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Gln Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser Val Tyr Pro
115 120 125
Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val Thr Leu Gly
130 135 140
Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val Thr Trp Asn
145 150 155 160
Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro Ser Ser Pro
180 185 190
Arg Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro Ala Ser Ser
195 200 205
Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly Cys Lys Pro
210 215 220
Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys Val Thr Cys
245 250 255
Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln Phe Ser Trp
260 265 270
Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln Pro Arg Glu
275 280 285
Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu Pro Ile Met
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg Val Asn Ser
305 310 315 320
Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
325 330 335
Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro Lys Glu Gln
340 345 350
Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr Asp Phe Phe
355 360 365
Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln Pro Ala Glu
370 375 380
Asn Tyr Lys Asn Thr Gln Pro Ile Met Asn Thr Asn Gly Ser Tyr Phe
385 390 395 400
Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu Ala Gly Asn
405 410 415
Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn His His Thr
420 425 430
Glu Lys Ser Leu Ser His Ser Pro Gly Lys
435 440
<210> 222
<211> 219
<212> PRT
<213> Lama glama
<400> 222
Gln Ala Val Val Thr Gln Glu Pro Ser Leu Ser Val Ser Pro Gly Gly
1 5 10 15
Thr Val Thr Leu Thr Cys Gly Leu Ser Ser Gly Ser Val Thr Thr Ser
20 25 30
Asn Tyr Pro Gly Trp Phe Gln Gln Thr Pro Gly Gln Ala Pro Arg Thr
35 40 45
Leu Ile Tyr Asn Thr Asn Ser Arg His Ser Gly Val Pro Ser Arg Phe
50 55 60
Ser Gly Ser Ile Ser Gly Asn Lys Ala Ala Leu Thr Ile Met Gly Ala
65 70 75 80
Gln Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Leu Tyr Pro Gly Ser
85 90 95
Thr Thr Val Phe Gly Gly Gly Thr His Leu Thr Val Leu Gly Gln Pro
100 105 110
Lys Ser Ser Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu
115 120 125
Glu Thr Asn Lys Ala Thr Leu Val Cys Thr Ile Thr Asp Phe Tyr Pro
130 135 140
Gly Val Val Thr Val Asp Trp Lys Val Asp Gly Thr Pro Val Thr Gln
145 150 155 160
Gly Met Glu Thr Glu Thr Thr Gln Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Met Glu Thr Ala Ser Ser Tyr Leu Thr Leu Thr Ala Arg Ala Trp
180 185 190
Glu Arg His Ser Ser Tyr Ser Cys Gln Val Thr His Glu Gly His Thr
195 200 205
Val Glu Lys Ser Leu Ser Arg Ala Asp Cys Ser
210 215
<210> 223
<211> 450
<212> PRT
<213> Lama glama
<400> 223
Glu Leu Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Asn Ser Tyr Gly Gly Ser Thr Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Glu Val Arg Ala Asp Leu Ser Arg Tyr Asn Asp Tyr Glu Ser
100 105 110
Tyr Asp Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser Ala Lys
115 120 125
Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln
130 135 140
Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser
180 185 190
Ser Val Thr Val Pro Ser Ser Pro Arg Pro Ser Glu Thr Val Thr Cys
195 200 205
Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val
210 215 220
Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val
225 230 235 240
Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile
245 250 255
Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp
260 265 270
Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His
275 280 285
Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg
290 295 300
Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr
340 345 350
Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu
355 360 365
Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp
370 375 380
Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile
385 390 395 400
Met Asn Thr Asn Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln
405 410 415
Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His
420 425 430
Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro
435 440 445
Gly Lys
450
<210> 224
<211> 216
<212> PRT
<213> Lama glama
<400> 224
Gln Pro Val Leu Asn Gln Pro Ser Ala Leu Ser Val Thr Leu Gly Gln
1 5 10 15
Thr Ala Lys Ile Thr Cys Gln Gly Gly Ser Leu Gly Ala Arg Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Asp Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Ala Asp Ser Ser Gly Ser Val
85 90 95
Phe Gly Gly Gly Thr His Leu Thr Val Leu Gly Gln Pro Lys Ser Ser
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Glu Thr Asn
115 120 125
Lys Ala Thr Leu Val Cys Thr Ile Thr Asp Phe Tyr Pro Gly Val Val
130 135 140
Thr Val Asp Trp Lys Val Asp Gly Thr Pro Val Thr Gln Gly Met Glu
145 150 155 160
Thr Glu Thr Thr Gln Pro Ser Lys Gln Ser Asn Asn Lys Tyr Met Glu
165 170 175
Thr Ala Ser Ser Tyr Leu Thr Leu Thr Ala Arg Ala Trp Glu Arg His
180 185 190
Ser Ser Tyr Ser Cys Gln Val Thr His Glu Gly His Thr Val Glu Lys
195 200 205
Ser Leu Ser Arg Ala Asp Cys Ser
210 215
<210> 225
<211> 450
<212> PRT
<213> Lama glama
<400> 225
Glu Val Gln Leu Gln Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ile Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Asn Ser Asp Gly Ser Ser Thr Ser Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Tyr Gly Ser Thr Trp Asp Val Gly Pro Met Gly Tyr Gly
100 105 110
Met Asp Tyr Trp Gly Lys Gly Thr Leu Val Thr Val Ser Ser Ala Lys
115 120 125
Thr Thr Pro Pro Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln
130 135 140
Thr Asn Ser Met Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser
180 185 190
Ser Val Thr Val Pro Ser Ser Pro Arg Pro Ser Glu Thr Val Thr Cys
195 200 205
Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val
210 215 220
Pro Arg Asp Cys Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val
225 230 235 240
Ser Ser Val Phe Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile
245 250 255
Thr Leu Thr Pro Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp
260 265 270
Asp Pro Glu Val Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His
275 280 285
Thr Ala Gln Thr Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg
290 295 300
Ser Val Ser Glu Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Phe Lys Cys Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr
340 345 350
Thr Ile Pro Pro Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu
355 360 365
Thr Cys Met Ile Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp
370 375 380
Gln Trp Asn Gly Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile
385 390 395 400
Met Asn Thr Asn Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln
405 410 415
Lys Ser Asn Trp Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His
420 425 430
Glu Gly Leu His Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro
435 440 445
Gly Lys
450
<210> 226
<211> 217
<212> PRT
<213> Lama glama
<400> 226
Ser Ser Ala Leu Thr Gln Pro Ser Ala Leu Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gln Gly Gly Ser Leu Gly Ser Ser Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Asp Asp Ser Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Gly Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Glu
65 70 75 80
Asp Glu Asp Asp Tyr Tyr Cys Gln Ser Thr Asp Ser Ser Gly Asn Thr
85 90 95
Val Phe Gly Gly Gly Thr Arg Leu Thr Val Leu Gly Gln Pro Lys Ser
100 105 110
Ser Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Glu Thr
115 120 125
Asn Lys Ala Thr Leu Val Cys Thr Ile Thr Asp Phe Tyr Pro Gly Val
130 135 140
Val Thr Val Asp Trp Lys Val Asp Gly Thr Pro Val Thr Gln Gly Met
145 150 155 160
Glu Thr Glu Thr Thr Gln Pro Ser Lys Gln Ser Asn Asn Lys Tyr Met
165 170 175
Glu Thr Ala Ser Ser Tyr Leu Thr Leu Thr Ala Arg Ala Trp Glu Arg
180 185 190
His Ser Ser Tyr Ser Cys Gln Val Thr His Glu Gly His Thr Val Glu
195 200 205
Lys Ser Leu Ser Arg Ala Asp Cys Ser
210 215
<210> 227
<211> 10
<212> PRT
<213> Homo sapiens
<400> 227
Glu Pro Lys Ser Cys Asp Lys Thr His Thr
1 5 10
<210> 228
<211> 5
<212> PRT
<213> Homo sapiens
<400> 228
Cys Pro Pro Cys Pro
1 5
<210> 229
<211> 8
<212> PRT
<213> Homo sapiens
<400> 229
Ala Pro Glu Leu Leu Gly Gly Pro
1 5
<210> 230
<211> 12
<212> PRT
<213> Homo sapiens
<400> 230
Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr
1 5 10
<210> 231
<211> 5
<212> PRT
<213> Homo sapiens
<400> 231
Cys Pro Arg Cys Pro
1 5
<210> 232
<211> 8
<212> PRT
<213> Homo sapiens
<400> 232
Ala Pro Glu Leu Leu Gly Gly Pro
1 5
<210> 233
<211> 7
<212> PRT
<213> Homo sapiens
<400> 233
Glu Ser Lys Tyr Gly Pro Pro
1 5
<210> 234
<211> 5
<212> PRT
<213> Homo sapiens
<400> 234
Cys Pro Ser Cys Pro
1 5
<210> 235
<211> 8
<212> PRT
<213> Homo sapiens
<400> 235
Ala Pro Glu Phe Leu Gly Gly Pro
1 5
<210> 236
<211> 3
<212> PRT
<213> Homo sapiens
<400> 236
Glu Arg Lys
1
<210> 237
<211> 10
<212> PRT
<213> Homo sapiens
<400> 237
Cys Cys Val Glu Cys Pro Pro Pro Cys Pro
1 5 10
<210> 238
<211> 7
<212> PRT
<213> Homo sapiens
<400> 238
Ala Pro Pro Val Ala Gly Pro
1 5
<210> 239
<211> 100
<212> PRT
<213> Homo sapiens
<400> 239
Arg Phe Cys Ser Ile Asn Ser Gly Leu His Ser Tyr Met Glu Met Pro
1 5 10 15
Leu Glu Cys Ile Leu Thr Glu Lys Arg Lys Lys Arg Ser Thr Lys Lys
20 25 30
Glu Val Phe Asn Ile Leu Gln Ala Ala Tyr Val Ser Lys Pro Gly Ala
35 40 45
Gln Leu Ala Arg Gln Ile Gly Ala Ser Leu Asn Asp Asp Ile Leu Phe
50 55 60
Gly Val Phe Ala Gln Ser Lys Pro Asp Ser Ala Glu Pro Met Asp Arg
65 70 75 80
Ser Ala Met Cys Ala Phe Pro Ile Lys Tyr Val Asn Asp Phe Phe Asn
85 90 95
Lys Ile Val Asn
100
<210> 240
<211> 100
<212> PRT
<213> Mus musculus
<400> 240
Arg Phe Cys Ser Val Asp Ser Gly Leu His Ser Tyr Met Glu Met Pro
1 5 10 15
Leu Glu Cys Ile Leu Thr Glu Lys Arg Arg Lys Arg Ser Thr Arg Glu
20 25 30
Glu Val Phe Asn Ile Leu Gln Ala Ala Tyr Val Ser Lys Pro Gly Ala
35 40 45
Asn Leu Ala Lys Gln Ile Gly Ala Ser Pro Ser Asp Asp Ile Leu Phe
50 55 60
Gly Val Phe Ala Gln Ser Lys Pro Asp Ser Ala Glu Pro Val Asn Arg
65 70 75 80
Ser Ala Val Cys Ala Phe Pro Ile Lys Tyr Val Asn Asp Phe Phe Asn
85 90 95
Lys Ile Val Asn
100
<210> 241
<211> 100
<212> PRT
<213> Rattus norvegicus
<400> 241
Arg Phe Cys Ser Val Asp Ser Gly Leu His Ser Tyr Met Glu Met Pro
1 5 10 15
Leu Glu Cys Ile Leu Thr Glu Lys Arg Arg Lys Arg Ser Thr Arg Glu
20 25 30
Glu Val Phe Asn Ile Leu Gln Ala Ala Tyr Val Ser Lys Pro Gly Ala
35 40 45
Asn Leu Ala Lys Gln Ile Gly Ala Ser Pro Tyr Asp Asp Ile Leu Tyr
50 55 60
Gly Val Phe Ala Gln Ser Lys Pro Asp Ser Ala Glu Pro Met Asn Arg
65 70 75 80
Ser Ala Val Cys Ala Phe Pro Ile Lys Tyr Val Asn Asp Phe Phe Asn
85 90 95
Lys Ile Val Asn
100
<210> 242
<211> 100
<212> PRT
<213> Macaca fascicularis
<400> 242
Arg Phe Cys Ser Leu Asn Ser Gly Leu His Ser Tyr Met Glu Met Pro
1 5 10 15
Leu Glu Cys Ile Leu Thr Glu Lys Arg Lys Lys Arg Ser Thr Lys Lys
20 25 30
Glu Val Phe Asn Ile Leu Gln Ala Ala Tyr Val Ser Lys Pro Gly Ala
35 40 45
Gln Leu Ala Arg Gln Ile Gly Ala Ser Leu Asn Asp Asp Ile Leu Phe
50 55 60
Gly Val Phe Ala Gln Ser Lys Pro Asp Ser Ala Glu Pro Met Asp Arg
65 70 75 80
Ser Ala Met Cys Ala Phe Pro Ile Lys Tyr Val Asn Asp Phe Phe Asn
85 90 95
Lys Ile Val Asn
100
<210> 243
<211> 100
<212> PRT
<213> Lama glama
<400> 243
Arg Phe Cys Ser Val Asp Ser Gly Leu His Ser Tyr Met Glu Met Pro
1 5 10 15
Leu Glu Cys Ile Leu Thr Glu Lys Arg Arg Arg Arg Ser Thr Lys Glu
20 25 30
Glu Val Phe Asn Ile Leu Gln Ala Ala Tyr Val Ser Lys Pro Gly Ser
35 40 45
Gln Leu Ala Lys Gln Ile Gly Ala Asn Leu Asn Asp Asp Ile Leu Tyr
50 55 60
Gly Val Phe Ala Gln Ser Lys Pro Asp Ser Ala Glu Pro Met Asn Arg
65 70 75 80
Ser Ala Val Cys Ala Phe Pro Val Lys Tyr Val Asn Glu Phe Phe Asn
85 90 95
Lys Ile Val Asn
100
<210> 244
<211> 50
<212> PRT
<213> Homo sapiens
<400> 244
Leu Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys Cys
1 5 10 15
Val Arg Ser Glu Glu Cys Leu Ser Gly Thr Trp Thr Gln Gln Ile Cys
20 25 30
Leu Pro Ala Ile Tyr Lys Val Phe Pro Asn Ser Ala Pro Leu Glu Gly
35 40 45
Gly Thr
50
<210> 245
<211> 50
<212> PRT
<213> Mus musculus
<400> 245
Leu Ser Ala Pro Tyr Phe Ile Gln Cys Gly Trp Cys His Asn Gln Cys
1 5 10 15
Val Arg Phe Asp Glu Cys Pro Ser Gly Thr Trp Thr Gln Glu Ile Cys
20 25 30
Leu Pro Ala Val Tyr Lys Val Phe Pro Thr Ser Ala Pro Leu Glu Gly
35 40 45
<210> 246
<211> 50
<212> PRT
<213> Rattus norvegicus
<400> 246
Leu Ser Ala Pro Tyr Phe Ile Gln Cys Gly Trp Cys His Asn Arg Cys
1 5 10 15
Val His Ser Asn Glu Cys Pro Ser Gly Thr Trp Thr Gln Glu Ile Cys
20 25 30
Leu Pro Ala Val Tyr Lys Val Phe Pro Thr Ser Ala Pro Leu Glu Gly
35 40 45
Gly Thr
50
<210> 247
<211> 50
<212> PRT
<213> Macaca fascicularis
<400> 247
Leu Ser Ala Pro Pro Phe Val Gln Cys Gly Trp Cys His Asp Lys Cys
1 5 10 15
Val Arg Ser Glu Glu Cys Pro Ser Gly Thr Trp Thr Gln Gln Ile Cys
20 25 30
Leu Pro Ala Ile Tyr Lys Val Phe Pro Thr Ser Ala Pro Leu Glu Gly
35 40 45
Gly Thr
50
<210> 248
<211> 50
<212> PRT
<213> Lama glama
<400> 248
Leu Ser Ala Pro Ser Phe Val Gln Cys Gly Trp Cys His Asp Lys Cys
1 5 10 15
Val Gln Leu Glu Glu Cys Ser Gly Gly Ile Trp Thr Gln Glu Ile Cys
20 25 30
Leu Pro Thr Ile Tyr Lys Val Leu Pro Thr Ser Ala Pro Leu Glu Gly
35 40 45
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-PVRIG АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2732042C2 |
| АНТИ-PVRIG АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2016 |
|
RU2827926C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2015 |
|
RU2743464C2 |
| Антитела против CXCR2 и их применение | 2019 |
|
RU2807067C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ ИНТЕРЛЕЙКИНА-33 (IL-33) | 2015 |
|
RU2829846C2 |
| ЛИГАНДСВЯЗЫВАЮЩАЯ МОЛЕКУЛА С РЕГУЛИРУЕМОЙ ЛИГАНДСВЯЗЫВАЮЩЕЙ АКТИВНОСТЬЮ | 2017 |
|
RU2803067C2 |
| ИММУНОАКТИВИРУЮЩАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2722788C2 |
| АНТИТЕЛА К МИОСТАТИНУ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2789884C2 |
| Новые варианты интерлейкина-2 для лечения рака | 2020 |
|
RU2833537C2 |
| КОМБИНАЦИЯ АНТИТЕЛА ПРОТИВ HGFR И HEGFR ДЛЯ ЛЕЧЕНИЯ ОПУХОЛИ И/ИЛИ МЕТАСТАЗА | 2019 |
|
RU2788606C2 |
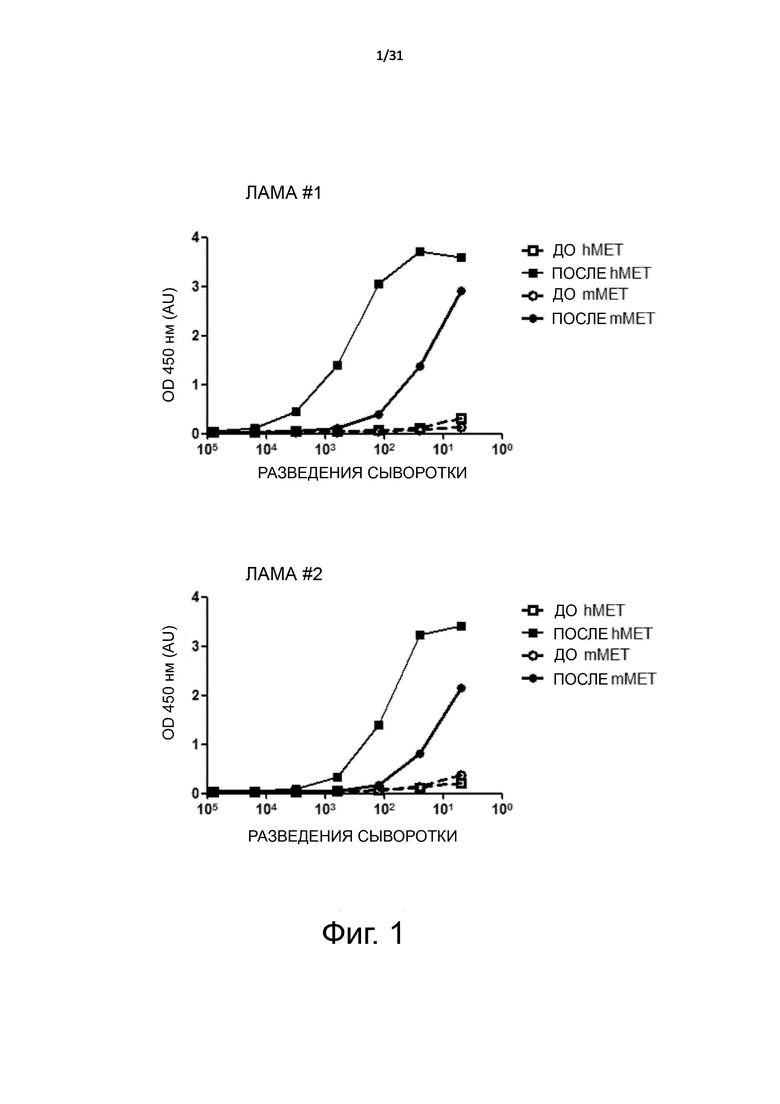




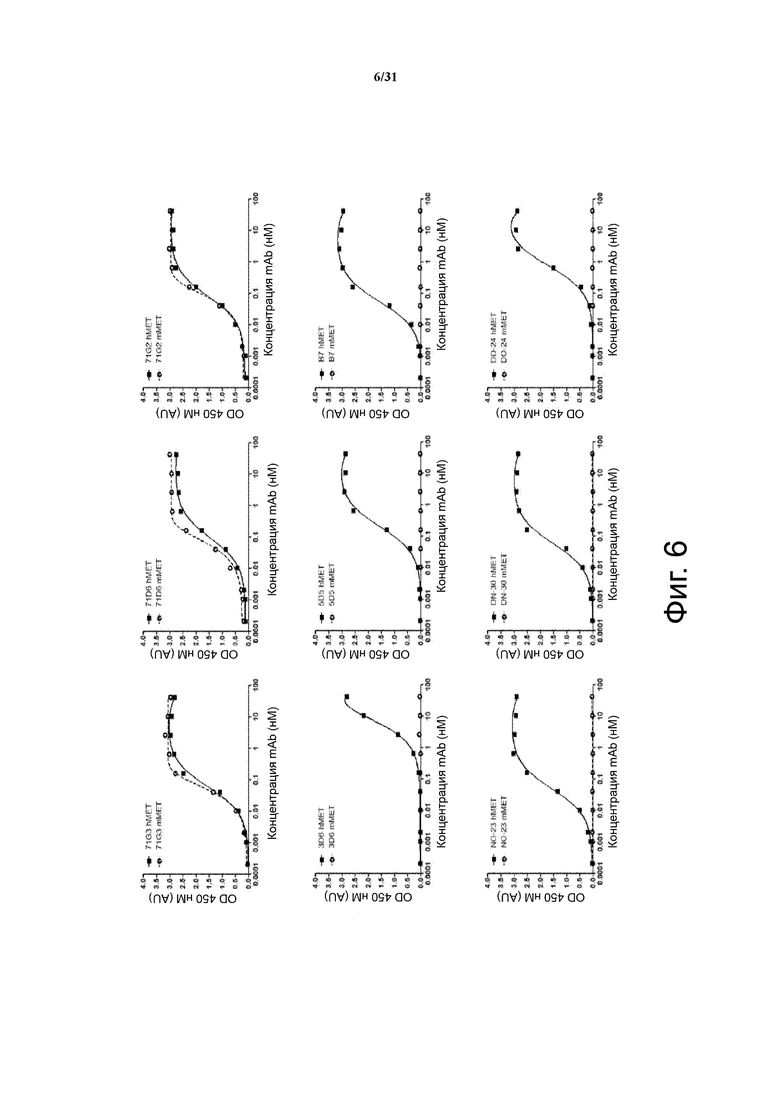

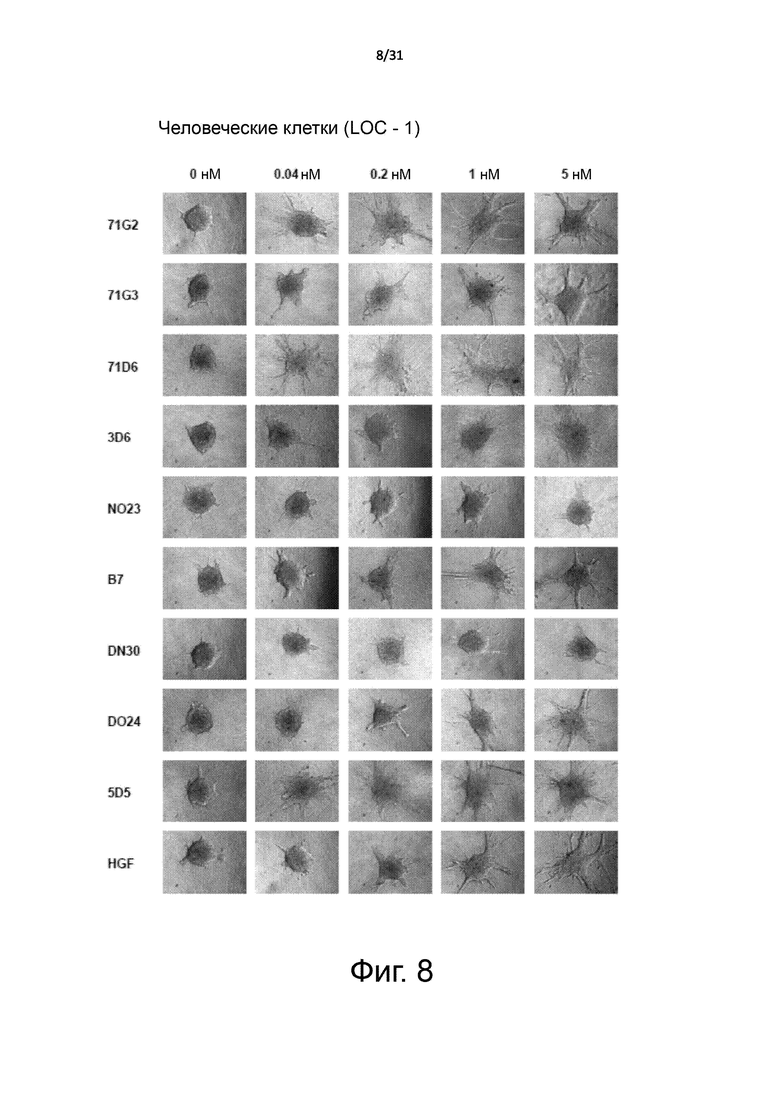




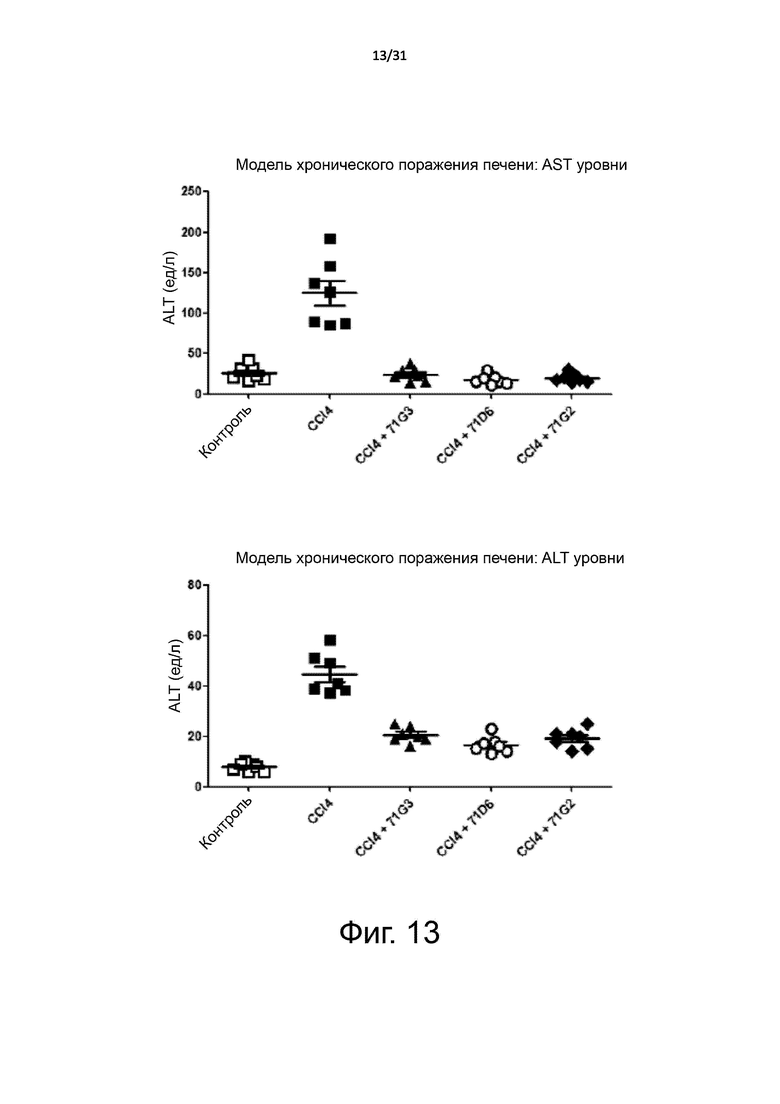
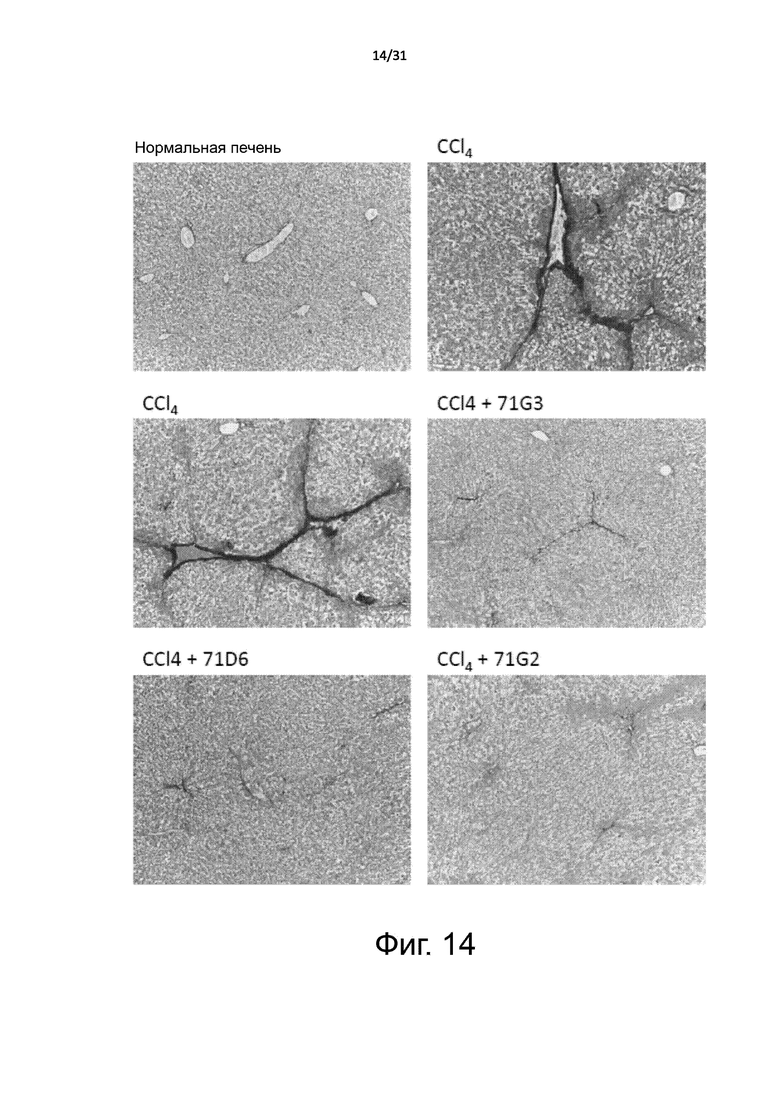

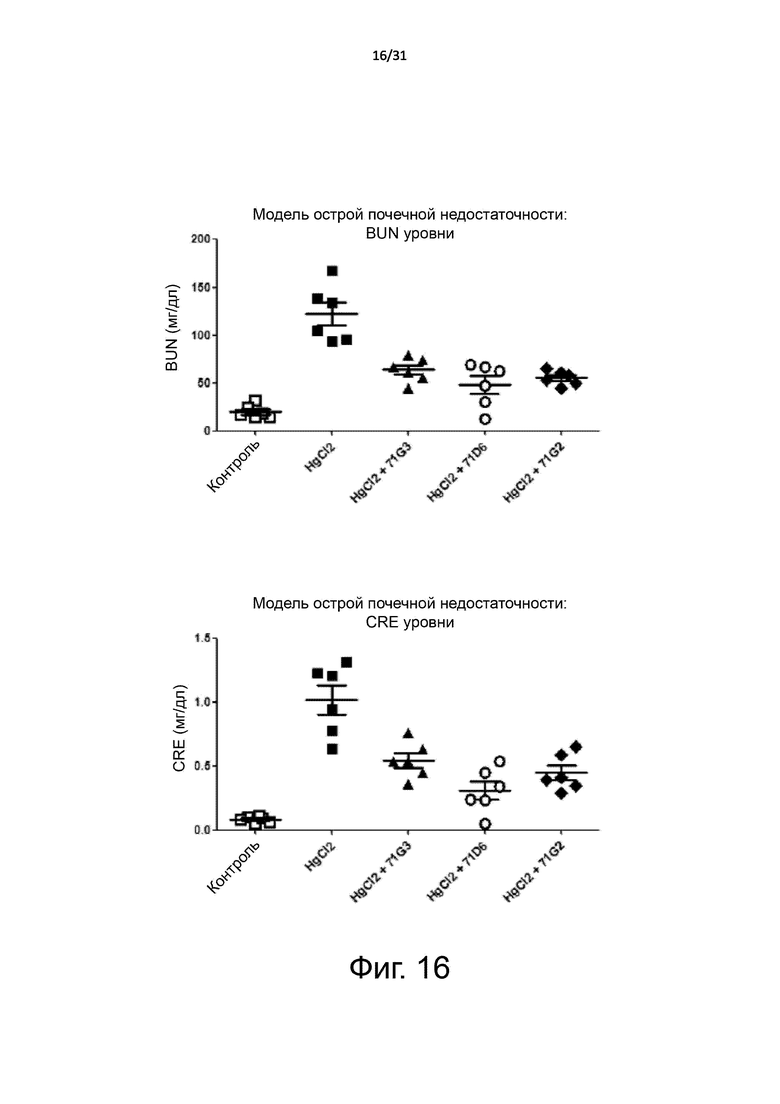
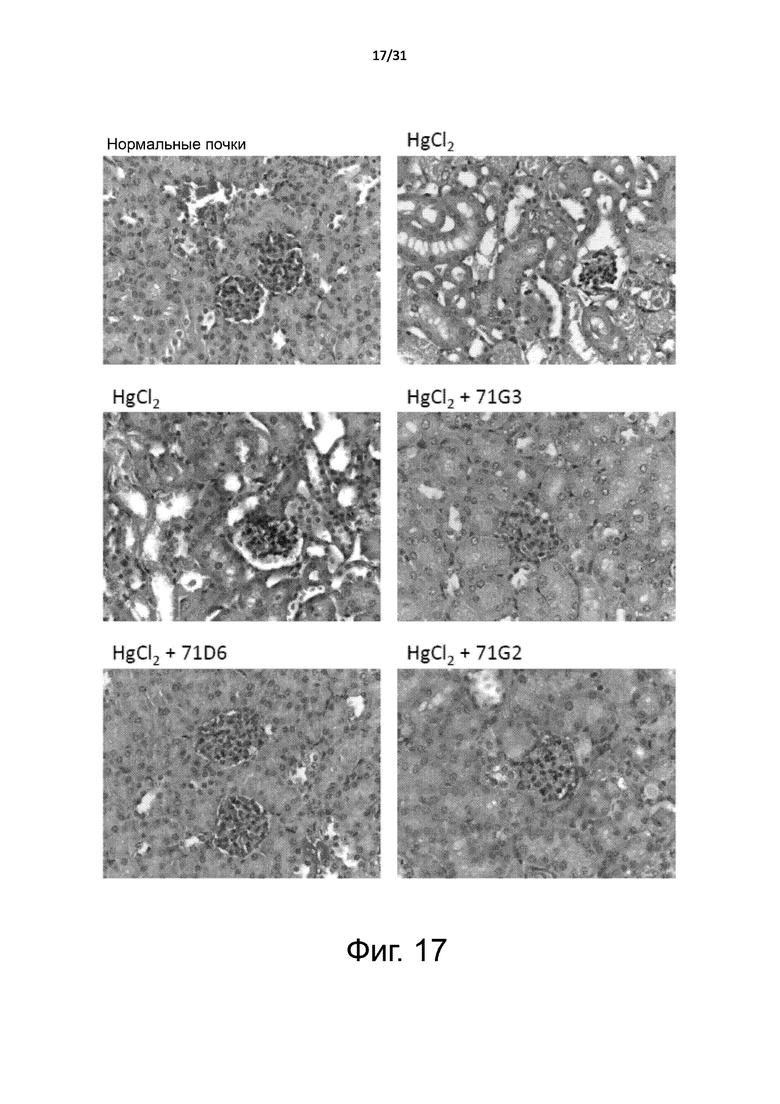







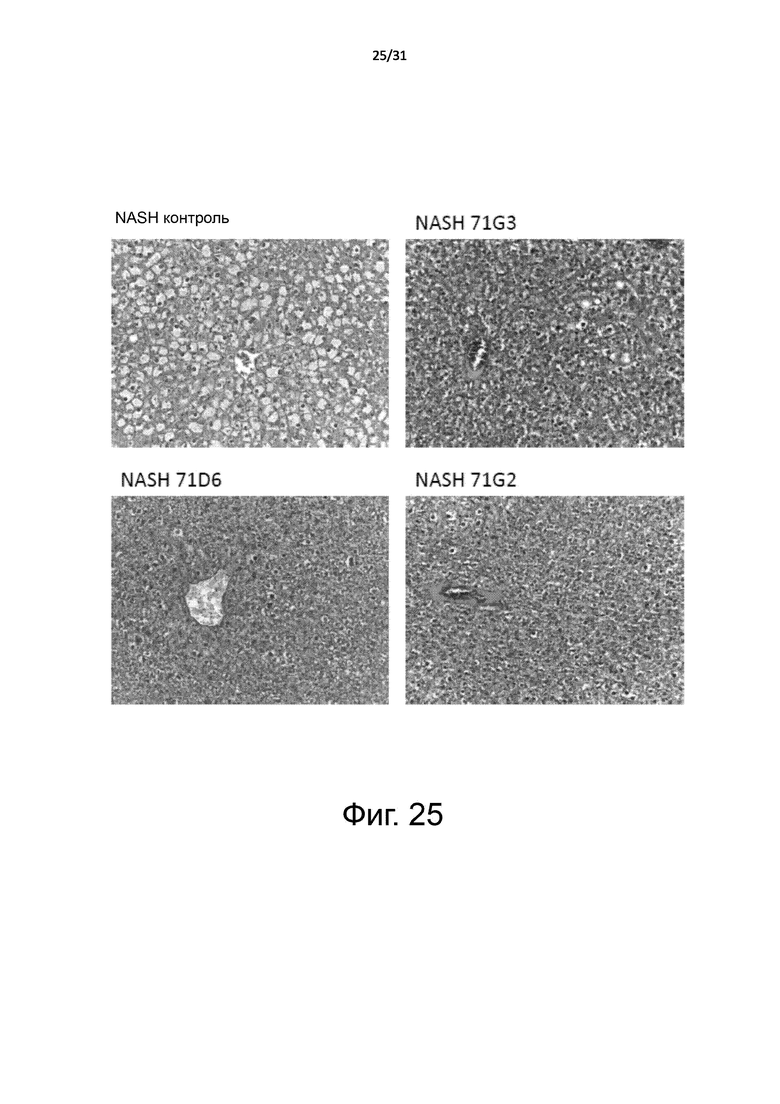

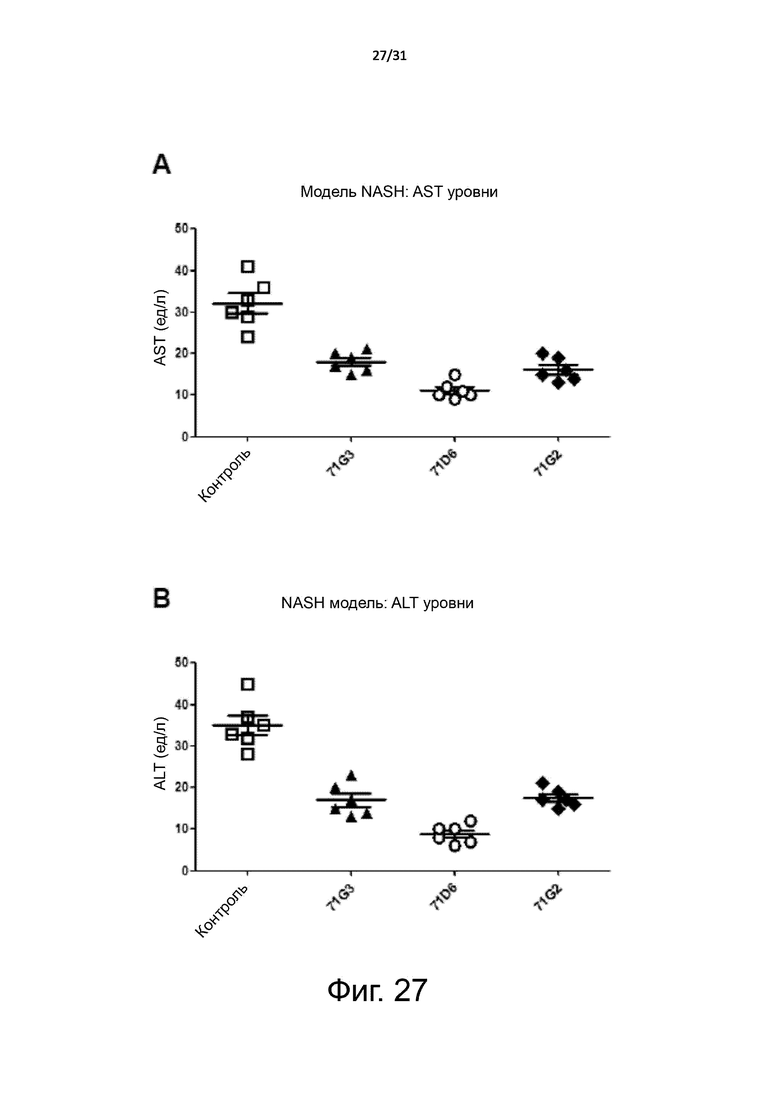
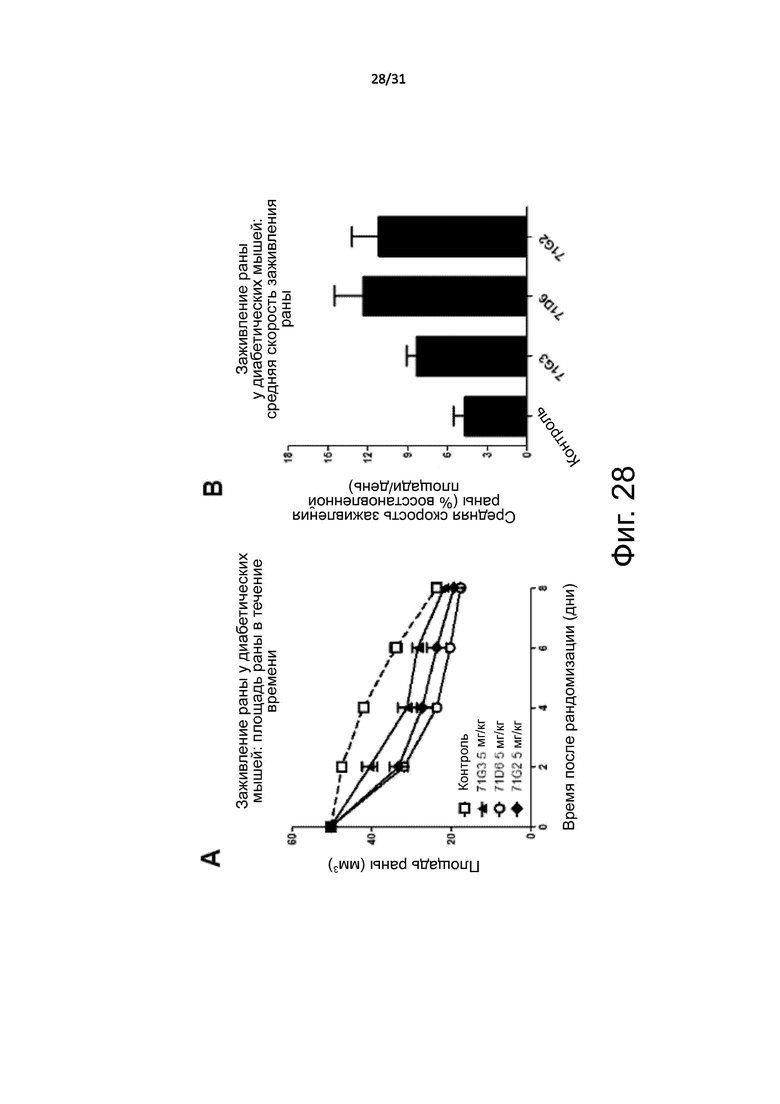



Данное изобретение имеет отношение к агонистическим анти-MET антителам и их применению при лечении болезней. Эти антитела связываются с высокой аффинностью с человеческим и мышиным рецептором фактора роста гепатоцитов (HGF), также известным как MET, и являются агонистами MET как у людей, так и у мышей, приводящими к молекулярным и клеточным эффектам, сходным с эффектами HGF-связывания, что позволяет эксперименты, проводимые на доклинических моделях, сделать прогностическими в ситуации с человеком. 7 н. и 7 з.п. ф-лы, 31 ил., 24 табл., 27 пр.
1. Антитело или его антигенсвязывающий фрагмент, которые являются агонистом MET человека и агонистом MET мыши и содержат вариабельный домен тяжелой цепи, включающий H-CDR1, H-CDR2 и H-CDR3, и вариабельный домен легкой цепи, включающий L-CDR1, L-CDR2 и L-CDR3, при этом:
H-CDR1 содержит аминокислотную последовательность, показанную как SEQ ID NO:30,
H-CDR2 содержит аминокислотную последовательность, показанную как SEQ ID NO:32,
H-CDR3 содержит аминокислотную последовательность, показанную как SEQ ID NO:34,
L-CDR1 содержит аминокислотную последовательность, показанную как SEQ ID NO:107,
L-CDR2 содержит аминокислотную последовательность, показанную как SEQ ID NO:109, и
L-CDR3 содержит аминокислотную последовательность, показанную как SEQ ID NO:111.
2. Антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельный домен тяжелой цепи содержит аминокислотную последовательность SEQ ID NO:163 или последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную этой, и вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO:164 или последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную этой.
3. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1, 2, являющиеся полноразмерным моноклональным антителом или фрагментом F(ab’)2.
4. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3, являющиеся иммуноглобулином изотипа IgG.
5. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-4, выполняющие одно или более из следующих условий:
- содержат шарнирный участок, CH2 домен и/или CH3 домен человеческого IgG;
- содержат шарнирный участок, CH2 домен и/или CH3 домен человеческого IgG1;
- имеют последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную человеческому IgG,
- имеют последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную человеческому IgG1,
- имеют последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную человеческому IgG, в котором CH2 и/или CH3 домен модифицирован для того, чтобы уменьшить или практически устранить одну или более эффекторных функций антитела;
- имеют последовательность, по меньшей мере, на 90%, 95%, 97% или 99% идентичную человеческому IgG1, в котором CH2 и/или CH3 домен модифицирован для того, чтобы уменьшить или практически устранить одну или более эффекторных функций антитела.
6. Антитело или его антигенсвязывающий фрагмент по любому из пп. 1-5, содержащие H-CDR1 последовательности SEQ ID NO:30, H-CDR2 последовательности SEQ ID NO:32, H-CDR3 последовательности SEQ ID NO:34, L-CDR1 последовательности SEQ ID NO:107, L-CDR2 последовательности SEQ ID NO:109 и L-CDR3 последовательности SEQ ID NO:111, привитые в антитело или антигенсвязывающий фрагмент полностью человеческого происхождения.
7. Изолированный полинуклеотид, кодирующий антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6.
8. Вектор экспрессии, содержащий полинуклеотид по п. 7, функционально связанный с регуляторными последовательностями, которые обеспечивают экспрессию антитела или его антигенсвязывающего фрагмента в организме-хозяине или в бесклеточной системе экспрессии.
9. Клетка-хозяин, содержащая экспрессирующий вектор по п. 8, для экспрессии антитела или его антигенсвязывающего фрагмента по любому из пп. 1-6.
10. Бесклеточная система экспрессии, содержащая экспрессирующий вектор по п. 8, для экспрессии антитела или его антигенсвязывающего фрагмента по любому из пп. 1-6.
11. Способ получения рекомбинантного антитела или его антигенсвязывающего фрагмента, включающий культивирование клетки-хозяина по п. 9 или бесклеточной системы экспрессии по п. 10 при условиях, которые обеспечивают экспрессию антитела или его антигенсвязывающего фрагмента и извлечение экспрессированного антитела или его антигенсвязывающего фрагмента.
12. Фармацевтическая композиция, имеющая агонистическую активность в отношении MET человека и/или MET мыши, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-6 и, по меньшей мере, один фармацевтически приемлемый носитель или эксципиент.
13. Антитело или антигенсвязывающий фрагмент по любому из пп. 1-6 или фармацевтическая композиция по п. 12, предназначенная для использования в терапии.
14. Антитело или антигенсвязывающий фрагмент или фармацевтическая композиция, предназначенная для использования по п. 13, при этом указанная терапия предназначается для:
- лечения или предотвращения поражения печени у пациента-человека, необязательно острого поражения печени или хронического поражения печени,
- лечения или предотвращения поражения почки у пациента-человека, необязательно острого поражения почки,
- лечения или предотвращения воспалительного заболевания кишечника у пациента-человека, необязательно язвенного колита,
- лечения или предотвращения диабета у пациента-человека, необязательно диабета I типа или II типа,
- лечения или предотвращения неалкогольного стеатогепатита у пациента-человека или
- лечения или содействия заживлению ран у пациента-человека, необязательно пациента, имеющего диабет.
| LI K, TAVARE R, ZETTLITZ KA et al | |||
| "Anti-MET immunoPET for non-small cell lung cancer using novel fully human antibody fragments", Mol Cancer Ther., 2014, Vol | |||
| Насос | 1917 |
|
SU13A1 |
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Сотенные или десятичные весы | 1925 |
|
SU2607A1 |
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |

