Изобретение относится к медицине, в частности к психофармакологии, и касается известного средства (композиции), представляющего собой глицин, иммобилизованный на частицах детонационного наноалмаза размером 2-10 нм [1] (далее нанолицин), применяемого для лечения и профилактики расстройств аутистического спектра, независимо от причин, вызывающих различные виды аутизма.
Под расстройствами аутистического спектра понимают группу комплексных нарушений психического развития с отсутствием способности к социальному взаимодействию, общению, стереотипностью поведения. Для больных аутизмом характерны фобии, возбуждение, нарушения пищевого поведения и другие неспецифические симптомы [2].
Впервые об аутизме как заболевании заговорили всего около 60 лет назад, но тогда контроль подобных расстройств был крайне редким. В настоящее время в мире количество детей больных аутизмом уже достигает одного на 200 (например, в Великобритании болеет один ребенок из 60), 75% из них отстают в умственном развитии [3]. Аутизм встречается среди мальчиков в 3-4 раза чаще, чем среди девочек. Этой болезнью страдают представители всех социальных слоев населения; она также обнаружена во всех странах мира, в которых проводились соответствующие исследования [2, 3].
До сих пор эффективного лекарственного средства или метода лечения расстройств аутистического спектра не найдено [2-4]. Однако существуют методы, которые способны помочь аутистам в той или иной степени. Причем добиться наибольших результатов можно, только применяя одновременно несколько из них. Одним из этих методов является медикаментозный метод лечения аутизма. Однако стандартных лекарств для лечения аутизма в настоящее время не существует. На данном этапе развития психофармакологии медикаментозное лечение аутизма является достаточно условным понятием, более точно говорить о «коррекции» расстройств аутистического спектра или лечении отдельной симптоматики аутизма. Медикаментозная терапия аутизма выступает в роли важного, но вспомогательного элемента общего терапевтического плана. И, несмотря на то что современный уровень психофармакологии делает невозможной каузальную медикаментозную терапию раннего детского аутизма, при помощи лекарственных средств можно воздействовать на определенные проявления, симптомы аутистического расстройства или группы симптомов. Например, медикаментозное лечение аутизма может положительно повлиять на двигательное возбуждение, саморазрушающее поведение аутиста, нейтрализовать или снизить психомоторные, депрессивные расстройства, воздействовать на развитие речевых функций и т.д. Помимо этого медикаментозный метод лечения аутизма незаменим при необходимости кризисных вмешательств [3]. Максимально раннее начало медикаментозной терапии имеет существенное значение как прогностический благоприятный фактор. Это обусловлено закономерностями развития головного мозга, положительными тенденциями онтогенеза при остановке активного течения болезни. Лекарственные средства выбирают с учетом психопатологической структуры расстройства, наличия или отсутствия сопутствующих психологических, неврологических и соматических нарушений [3, 4].
Если во взрослой практике лечение направлено не только на ликвидацию болезненных явлений, но и на оптимальное приспособление к среде, то у аутичных детей улучшение их адаптации в каждый момент обеспечивает в дальнейшем возможность более адекватного развития психики [3].
Дополнительные сложности проведения психофармакотерапии расстройств аутистического спектра заключаются, прежде всего, в том, что лекарственные средства нового поколения (атипичные нейролепики, антидепрессанты), как правило, не рекомендованы к применению в детском возрасте. Первым препаратом, одобренным в США для лечения аутоагрессии и приступов гнева при детском аутизме, в 2006 году стал атипичный антипсихотик рисперидон [4], который при умеренной эффективности переносится лучше, чем такой эталонный препарат как галоперидол. Поэтому арсенал лекарственных средств для лечения расстройств аутистического спектра достаточно ограничен [3, с. 127; 4]. Это также связано с тем, что после наиболее часто применяемой антипсихотической фармакотерапии проявляются тяжелые побочные эффекты, в том числе экстрапирамидные расстройства. При применении антидепрессантов (кломипрамин, амитриптилин, сертралин, флуоксетин) возможны обострения различных продуктивных расстройств. При наличии в структуре аутистических расстройств тревоги, нарушений сна, обсессивно-компульсивных проявлений, выраженного возбуждения с симптомами интоксикации применяют анксиолитики (транквилизаторы) и гипнотики, которые также имеют широкий ряд токсических и побочных проявлений. При наличии в клинической картине выраженных аффективных расстройств часто применяют антиконвульсанты (вальпроат натрия, карбамазепин, ламиктал, конвулекс). При патогенетически обоснованной каузальной терапии аутистических расстройств в последнее время стал применяться глуматергический препарат актинол-мемантин. Эти препараты, как правило, также проявляют различные побочные эффекты и влияют на кроветворение. При всех видах аутистических расстройств широко используются ноотропы и вещества с ноотропным действием. Для преодоления когнитивного дефицита нейролептики сочетают с иммунными препаратами (кагоцел, тенатен и др.). Широко используются и гомеопатические средства (церебрум-композитум, коэнзим-композитум и др.), как правило, с парентеральным введением [3, с. 129-131]. Однако эффективность этих средств невысока.
Многочисленные попытки найти эффективные препараты и успешный способ терапии аутистических расстройств пока не привели к успеху [2-4]. Поэтому разработка и безопасное применение эффективных фармакологических средств для лечения и профилактики аутизма является чрезвычайно важной и актуальной социально-медицинской задачей.
Расстройства аутистического спектра диагностируются наличием нескольких критериев, основными из которых являются:
- стереотипное, повторяющееся поведение, настойчивое требование постоянных, одинаковых условий;
- нарушение социального взаимодействия;
- нарушение процессов обучения и, особенно, затруднение процессов переобучения;
- дефицит общения;
- ограниченность интересов;
- повышенная тревожность и др.
На основе этих представлений в последние годы разработаны специальные психофармакологические экспериментальные модели аутизма у животных, позволяющие воспроизвести нарушения центральной нервной системы (ЦНС) в эксперименте на мышах [5-13].
Наиболее известными и часто используемыми экспериментальными тестами на животных являются:
1) тест оценки повторяющегося, настойчивого (упорного) моторного поведения;
2) тест оценки коммуникационных отношений по обонятельному взаимодействию.
Известно, что заменимая аминокислота глицин (NH2CH2COOH) помимо участия в образовании важнейших биологически активных соединений - пуриновых нуклеотидов, гема, креатина и др., являясь центральным нейромедиатором тормозного типа действия, проявляет седативное действие и улучшает метаболические процессы в тканях мозга, контролирует процессы формирования тонкой моторики пластических процессов и тонусных реакций поперечнополосатой мускулатуры [14, 15]. Основная масса глицина сосредоточена в спинном мозге, где аминокислота, высвобождаясь из окончаний клеток Реншоу, опосредует постсинаптическое высвобождение (торможение) мотонейронов. Поэтому глицин широко используется в неврологической практике для устранения повышенного мышечного тонуса.
Регуляция активности NMDA-глутаматных рецепторов также осуществляется глицином. Он имеет собственный сайт в составе большинства глутаматных возбуждающих рецепторов. При взаимодействии с магнием глицин оказывает тормозящее воздействие, в свободном виде - стимулирующее воздействие.
В современной психофармакотерапевтической практике глицин применяют как средство, ослабляющее депрессивные нарушения, повышенную раздражительность, влечение к алкоголю, уменьшающее явления абстиненции, нормализующее сон, усиливающее антипсихотическую терапию, а также в комплексном лечении нарушений мозгового кровообращения [14]. В основе фармакологического действия глицина лежит эффект амплификации метаболических и нейротрансмиттерных процессов, возникающих за счет усиления его эндогенного синтеза. Увеличить внутриклеточный синтез глицина можно, только используя пути передачи сигнала, обусловленные взаимодействием с рецепторными системами. Его взаимодействие с глициновыми рецепторами приводит к открытию хлорных каналов, гиперполяризации мембраны и распространению торможения. Наряду с этим глицин способен выступать в роли аллостерического коагониста глутаматных рецепторов. Связываясь в специфическом сайте, он усиливает способность глутамата и N-метил-D-аспарата (NMDA) открывать катионный канал [15, 16]. Применяемый фармакопейный глицин назначают в виде таблеток по 0,1 г сублингвально (под язык) 3-4 раза в день [14].
Известен глицин, иммобилизованный на частицах детонационного наноалмаза размером 2-10 нм [1] (далее, нанолицин), обладающий при разных дозах седативным, антидепрессивным, антипсихотическим и анксиолитическим действиями, существенно превосходящими соответствующие виды специфической активности фармакопейного глицина (в субстанции), и способ его получения [17-20]. По антидепрессивному действию алмацин не уступает по глубине эффекта эталонным антидепрессантам амитриптилину и флуоксетину. При этом в дозе, превышающей более чем в 20 раз терапевтическую дозу глицина, у него отсутствуют побочные эффекты и токсическое действие.
Применение глицина, иммобилизованного на частицах детонационного наноалмаза (нанолицина), как средства для лечения и профилактики расстройств аутистического спектра, в научной и патентной литературе не описано.
Целью изобретения является применение глицина, иммобилизованного на частицах детонационного наноалмаза размером 2-10 нм (нанолицина) для лечения и профилактики расстройств аутистического спектра без проявления каких-либо побочных эффектов и токсических действий, повышение психофармакологической активности глицина и расширение номенклатуры лекарственных средств, применяемых при лечении и профилактики аутизма у детей и взрослых.
В соответствии с изобретением описывается средство для лечения и профилактики расстройств аутистического спектра, представляющее собой глицин, иммобилизованный на частицах детонационного наноалмаза размером 2-10 нм с содержанием глицина до 21±3% масс.
Для определения перспективности применения глицина, иммобилизованного на частицах детонационного наноалмаза размером 2-10 нм с содержанием глицина до 21±3% масс, (нанолицина), в качестве средства для лечения и профилактики расстройств аутистического спектра было проведено изучение его специфического психофармакологического действия в сравнении с фармакопейным глицином и эталонным атипичным нейролептиком трифтазином (препарат сравнения).
Исследование специфического психофармакологического действия нанолицина и сравниваемых с ним лекарственных средств было проведено по тесту обонятельной габитуации/дисгабитуации, моделирующему нарушение адекватного реагирования на обонятельные стимулы, показателям процесса обучения пространственному навыку и способности осуществления нового пространственного движения при переобучении в водном лабиринте на 330 мышах-самцах линии Balb/C возраста 5-7 недель, 54 аутбредных мышах-самцах возрастом 2 месяца и 18 мышах-самцах линии C57BL/6 возрастом 2-3 месяца. Работа выполнялась согласно методикам [20, 7], соответственно.
Статистическую обработку экспериментальных данных проводили с помощью программы «Statistica v6.0», используя однофакторный дисперсионный анализ; непараметрический анализ для независимых переменных (U-тест Манна-Уитни) и парный критерий Стьюдента для внутригрупповых сравнений.
В результате проведенного фармакологического исследования неожиданно было обнаружено, что:
1) нанолицин в дозе 1 мг/кг при однократном введении вызывал более быструю адаптацию (привыкание) мышей к обонятельным стимулам при их повторных предъявлениях и улучшал распознавание нового социального запаха. Нанолицин в дозе 10 мг/кг также вызывал более быстрое привыкание животных к обонятельным стимулам, однако при распознавании новых запахов поведение животных на его фоне не отличалось от контрольных значений. Препарат сравнения трифтазин (0,5 мг/кг) тоже обладал способностью улучшать распознавание нового запаха и ускорять процессы адаптации к нему. По эффективности нанолицин в обеих дозах не уступал препарату сравнения трифтазину. Фармакопейный глицин (субстанция) в дозе 10 мг/кг обладал слабой эффективностью в данном тесте и существенно уступал нанолицину в дозах 1 и 10 мг/кг. Детонационные наноалмазы не проявляли способность улучшать распознавание новых запахов;
2) нанолицин в дозе 10 мг/кг (введение в течение 6-ти дней ежедневно) достоверно улучшал процессы обучения мышей в условиях водного лабиринта как до, так и после пространственной сшибки, а в дозе 1 мг/кг - улучшал процессы обучения мышей только на второй день обучения. Препараты сравнения (введение в течение 6-ти дней ежедневно): трифтазин (0,5 мг/кг) и фармакопейный глицин (10 мг/кг) не обнаруживали влияния на процессы обучения животных, аналогично детонационным наноалмазам.
Изобретение иллюстрируется следующими примерами.
Пример 1. Изучение влияния нанолицина в сравнении с глицином, трифтазином и наноалмазами на проявления аутизма в тесте обонятельной габитуации/дисгабитуации.
При аутизме нарушается способность адекватного реагирования на социальные и несоциальные обонятельные стимулы. Экспериментальные модели этих нарушений учитывают установленный факт, что взаимоотношения у мышей, в первую очередь, происходят через обонятельные сигналы. Поэтому обонятельное привыкание/непривыкание в эксперименте изучается с помощью повторных предъявлений мышам несоциальных и социальных запахов в тесте обонятельной габитуации/дисгабитуации [5, 12, 21].
Эксперимент проводили на мышах-самцах линии Balb/C 5-7 недельного возраста, массой 13-15 г. В качестве социального стимула использовали мышей линии С57В1/6 того же пола, что и тестируемые мыши. В качестве дополнительного контроля использовали аутбредных белых мышей-самцов возрастом 2-3 месяца, массой 24-30 г.
Животные были получены из питомника РАМН «Столбовая» (Московская область). Содержание животных в виварии соответствовало Приказу Минздравсоцразвития РФ №708н от 23 августа 2010 г. «Об утверждении правил лабораторной практики». Животные содержались на постоянном доступе к корму и воде - предоставлялся полный рацион экструдированного брикетированного корма (ГОСТ на корм Р50258-92) и питьевая вода; при температурном режиме 20-22°С, при световом режиме - 12 часов свет/12 часов темнота; в полипропиленовых клетках с решеткой из цинкохромовой стали, с обеспыленной подстилкой из деревянной стружки по 10 мышей в каждой клетке (Т/3С). Содержание животных осуществлялось в соответствии с нормативным документом №1045-73 от 06.04.1973 г. «Санитарные правила по устройству, оборудованию и содержанию вивариев», утвержденным Главным государственным санитарным врачом.
При проведении Примера 1 использовали:
- глицин, иммобилизованный на частицах детонационного наноалмаза размером 2-10 нм с содержанием глицина до 21±3% масс, (нанолицин) в дозах 1 и 10 мг/кг;
- детонационные наноалмазы в соответствующей концентрации для дозы глицина 10 мг/кг;
- субстанцию фармакопейного глицина (препарат сравнения) в дозе 10 мг/кг;
- трифтазин (препарат сравнения) в дозе 0,5 мг/кг.
Вещества вводили однократно внутрибрюшинно в объеме 0,1 мл на 10 г массы мыши за 40 мин до эксперимента.
Контрольным животным линии Balb/C и аутбредным мышам вводили однократно внутрибрюшинно физиологический раствор в объеме 0,1 мл на 10 г массы мыши за 40 мин до эксперимента.
Тест обонятельной габитуации/дисгабитуации используется для выявления способности животных отвечать на социальные и несоциальные обонятельные стимулы. Исследование проводили согласно методике [21], предложенной в 2010 году.
Процедура эксперимента. Мыши перед началом тестирования, по отдельности, помещают в клетку с чистыми опилками на 30 мин. Ватный тампон длиной 15 см смачивают в воде, пропускают через крышку клетки так, чтобы он располагался на высоте 5 см от выстилающих дно клетки опилок. Выполняют по 3 предъявления каждого запаха:
3 предъявления - вода;
3 предъявления - несоциальный стимул: разбавленный цветочный запах (экстракт лимона, разбавленный в соотношении 1:100 (вода);
3 предъявления - социальный стимул (запах из мышиной «незнакомой» клетки с грязными опилками).
Для социального стимула используют ватный тампон, который предварительно смочили водой и протерли им дно грязной «чужой» мышиной клетки зигзагообразными движениями так, чтобы захватить все углы и середину клетки. В «чужой» клетке должно сидеть не менее 3-х мышей линии С57В1/6 того же пола, что и тестируемые мыши. Опилки не должны меняться не менее 3 дней.
Наблюдение проводят в течение 2 мин. Регистрируют следующие показатели:
- число поднятий и поворотов головы (носа) в сторону тампона с запахом, находящегося на расстоянии не более 2 см;
- число подходов к тампону;
- число обнюхивания тампона;
- число залезаний на тампон;
- число жеваний тампона.
Через 2 мин ватный тампон меняют на такой же, который предъявляют также 2 мин.
Результаты эксперимента
Поведение аутбредных мышей и мышей линии Balb/C при предъявлении обонятельных стимулов по показателю реакций на обонятельные стимулы.
Характерной чертой поведения интактных аутбредных мышей в течение всего эксперимента было распознавание нового запаха, что выражалось в повышенной реакции мышей на тампон с нанесенным на него запахом при первом предъявлении каждого обонятельного стимула и быстрое привыкание к запаху при его повторном предъявлении.
При первом тестировании запаха «Лимон» у аутбредных мышей отмечено увеличение на 23,4% реакции мышей на обонятельный стимул по сравнению с предшествующим этому предъявлению нейтральным запахом «Вода» (3-е предъявление), ставшим уже привычным. А в случае первого предъявления обонятельного социального стимула «Запах мышей С57В1/6» относительно третьего предъявления запаха «Лимон», ставшего на тот момент привычным, увеличение реакции мышей на тампон с запахом составило 5,5 раза (Таблица 1).
Динамика реагирования мышей на запах при его повторных предъявлениях представлена в Таблице 1. Как следует из данных Таблицы 1, аутбредные мыши при втором предъявлении запаха «Вода» относительно первого тестирования этого запаха реагировали на обонятельный стимул на 24,2% меньше. При тестировании запахов «Лимон» и «Запах мышей линии С57В1/6» снижение реакции животных было более значительно и соответствовало 74,1% и 51,6%, соответственно.
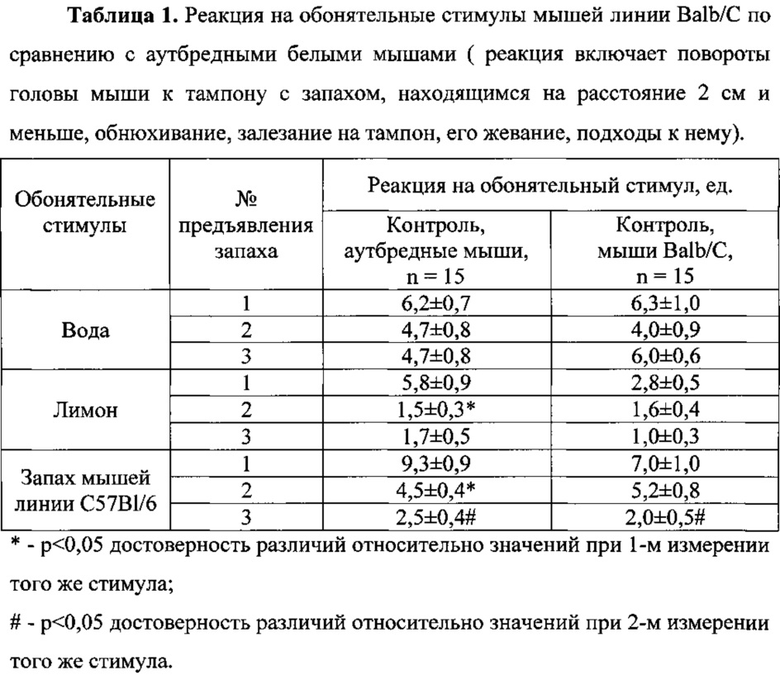
У мышей линии Balb/C при повторном предъявлении запахов «Лимон» и «Запах мышей линии С57В1/6» выраженность адаптации к обонятельным стимулам была ниже, чем в группе аутбредных мышей. Так, при повторном предъявлении запахов «Лимон» и «Запах мышей линии С57В1/6» снижение реакции мышей линии Balb/C на стимул относительно первого предъявления подобного запаха составило 42,9% и 25,7%, соответственно, что было в 1,7 и 2,0 раза меньше, соответственно, чем в группе аутбредных мышей (Таблица 1).
Способность распознавать обонятельные стимулы «Вода» и «Запах мышей линии С57В1/6» у мышей линии Balb/C не отличалась от аналогичных способностей аутбредных мышей, о чем свидетельствует отсутствие различий между реакциями на новый запах в обеих контрольных группах. Однако реакция на запах «Лимон» у мышей линии Balb/C была в 2,1 раза ниже по сравнению с группой аутбредных животных (Таблица 1).
Влияние алмацина на поведение мышей линии Balb/C при предъявлении обонятельных стимулов по показателю реакций на обонятельные стимулы.
Мыши линии Balb/C, получавшие алмацин в дозах 1 и 10 мг/кг, быстрее привыкали к обонятельным стимулам «Вода» и «Запах мышей линии С57В1/6» при их повторных предъявлениях по сравнению с контрольными животными линии Balb/C (Таблица 2). Так, в группе «Алмацин, 1 мг/кг» при повторном предъявлении обонятельных стимулов «Вода» и «Запах мышей линии С57В1/6» снижение реакции мышей было достоверно относительно первого предъявления запаха и соответствовало 32,5% и 61,2%. В группе «Алмацин, 10 мг/кг» снижение реакции мышей на запахи «Вода» и «Запах мышей линии С57В1/6» соответствовало 43,4% и 65,4% (р<0,05). Снижение реакции на третье предъявление обонятельных стимулов «Вода» и «Запах мышей линии С57В1/6» относительно в группе животных «Алмацин, 1 мг/кг» было на уровне тенденции. Кроме того, реакция животных, получавших алмацин в дозе 10 мг/кг, на третье предъявление запаха «Лимон» была достоверно на 68% ниже, чем реакция на его второе предъявление.
При этом у животных, получавших алмацин в дозе 1 мг/кг, способность к распознаванию впервые предъявленного стимула «Запах мышей линии С57В1/6» была в 2,1 выше, чем в группе контрольных мышей линии Balb/C (р<0,05).
Наноалмаз не изменял способность к распознаванию обонятельных стимулов по сравнению с контролем. Однако на его фоне животные быстрее привыкали к стимулам «Вода» и «Запах мышей линии С57В1/6» по сравнению с контрольными животными линии Balb/C (Таблица 2).
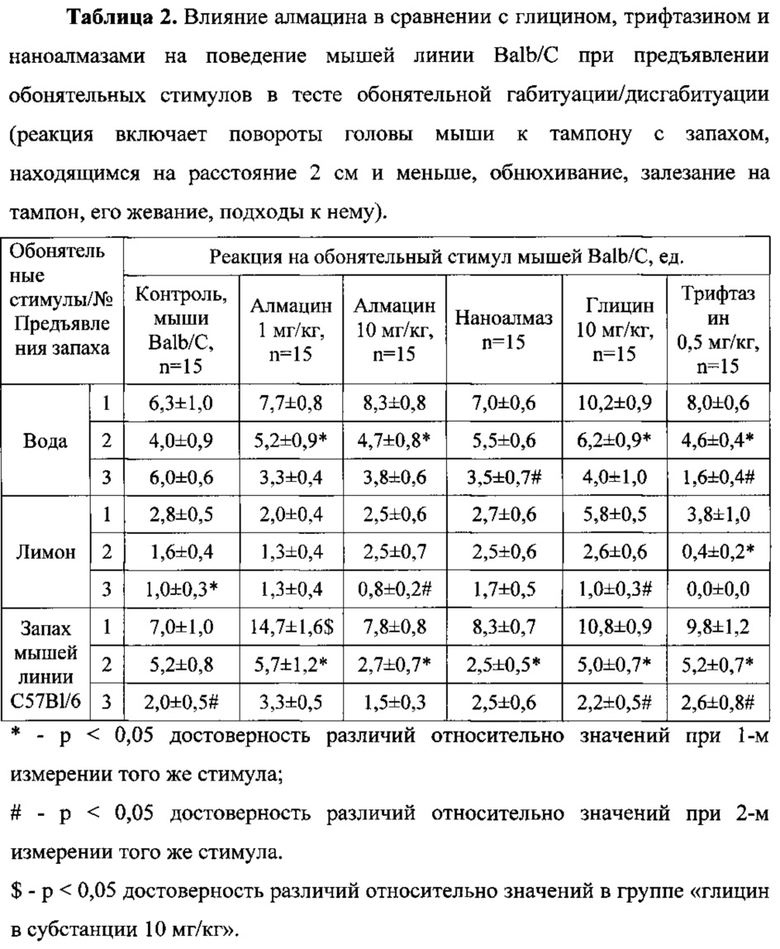
На фоне препаратов сравнения - глицина в дозе 10 мг/кг и трифтазина в дозе 0,5 мг/кг - привыкание к обонятельным стимулам также происходило быстрее по сравнению с контрольными животными линии Balb/C.
У животных, получавших глицин в дозе 10 мг/кг, снижение реакции на второе предъявление нейтрального обонятельного стимула «Вода» было достоверным относительно первого предъявления и составляло 39,2%. Глицин вызывал достоверно более быстрое привыкание к каждому последующему предъявлению обонятельного стимула «Запах мышей линии С57В1/6». В этой группе реакция на третье предъявление запаха «Лимон» также достоверно снижалась в сравнении с его вторым предъявлением. В целом, для этой группы животных характерна достоверна (р<0,01) более выраженная реакция к распознаванию всех обонятельных стимулов по сравнению с группой контрольных животных линии Balb/C.
Второй препарат сравнения трифтазин вызывал достоверно более быстрое привыкание к каждому последующему предъявлению обонятельных стимулов «Вода» и «Запах мышей линии С57В1/6». Кроме того, на его фоне реакция животных на третье предъявление запаха «Лимон» была достоверно в 9,5 раза ниже, чем реакция на его второе предъявление. Распознаванию всех обонятельных стимулов при первом предъявлении животными этой группы хотя и было более выражено по сравнению с контрольной группой, не было достоверно (Таблица 2).
Таким образом, алмацин в дозах 1 и 10 мг/кг улучшает распознавание нового социального запаха и адаптацию к обонятельным стимулам в тесте обонятельной габитуации/дисгабитуации. По эффективности алмацин в обеих дозах не уступает препарату сравнения трифтазину (0,5 мг/кг) и превосходит глицин (10 мг/кг). Детонационные наноалмазы не улучшают распознавание новых запахов у мышей по сравнению с контролем.
Пример 2. Изучение влияния алмацина в сравнении с глицином, трифтазином и наноалмазами на проявления аутизма в тестах обучения и переобучения после пространственной «сшибки» в водном лабиринте Морриса.
Зацикленное, повторяющееся, стереотипное поведение, ограниченный круг интересов и активности являются характерными проявлениями аутизма [22, 5]. Повторяющееся, стереотипное поведение у мышей включает вращательные движения, повороты назад, повторяющееся обнюхивание одних и тех же мест в пространстве или объектов, а также чрезмерный грумминг и повышенную нецеленаправленную двигательную активность [23, 8].
Повторяющееся зацикленное поведение мышей линии Balb/C выявляется в неспособности животных принять решение и осуществить правильное пространственное движение при смене локализации платформы при переобучении в водном лабиринте [8].
Эксперимент проводили на мышах-самцах линии Balb/C возрастом 5-7 недель массой 13-15 г. Источник получения и условия содержания идентичны Примеру 1.
При проведении Примера 2 исследуемые вещества использовали аналогично Примеру 1.
Вещества вводили в объеме 0,1 мл на 10 г массы мыши внутрибрюшинно, в течение 6-ти дней. Контрольным животным линии Balb/C и аутбредным мышам вводили внутрибрюшинно физиологический раствор в объеме 0,1 мл на 10 г массы мыши за 40 мин до эксперимента.
Для оценки влияния алмацина на симптомы расстройств аутистического спектра применялась методика, предложенная в 2007 году в работе [7]. Изучалось влияние испытуемых веществ на процесс обучения пространственному навыку и на способность осуществить новое пространственное движение при переобучении в водном лабиринте.
Исследование проводили в водном лабиринте Морриса. Сначала у мышей проводили предварительное обучение навыку находить платформу в водном лабиринте и воспроизводить этот пространственный навык. А затем животным предъявляли новое местоположение платформы и, таким образом, осуществляли пространственную «сшибку». При переобучении - смене локализации платформы в водном лабиринте - животные должны принять новое решение и осуществить правильное пространственное движение.
Лабиринт Морриса представляет собой круглый бассейн диаметром 122 см, глубиной 25 см, заполненный водой температурой 25-28°С. В бассейн помещали круглую платформу диаметром 12 см. Центр платформы располагали на расстоянии 30 см от краев бассейна.
Процедура эксперимента
1 этап. Ознакомление с условиями опыта. Платформу располагают на 0,5 см выше уровня воды. Мышь помещают на платформу на 20 секунд. Затем мышь опускают в воду на противоположной стороне бассейна и позволяют в течение 60 сек найти платформу и взобраться на нее, где оставляют животное на 20 секунд. Процесс повторяют, опуская мышь в воду с другой, отличной от первой попытки стороны бассейна. Каждому животному предлагается 4 попытки найти платформу.
Если животное не в состоянии самостоятельно найти платформу в течение 60 сек, экспериментатор помогает ему переместиться к платформе и взобраться на нее.
2 этап. Обучение животных пространственному навыку. В течение 2-х последующих дней платформа располагается на 0,5 см ниже уровня воды. Ежедневно животным предоставляют по 4 попытки найти платформу в течение 60 сек. Интервал между попытками составляет 20 сек, в течение которого они находятся на платформе. Каждый день перед первой попыткой животное на 20 сек помещают на платформу. Регистрируют время, прошедшее от момента спуска животного в воду до влезания на платформу, и количество результативных попыток поиска платформы. Животных опускают в воду в 3-х различных точках на половине бассейна, противоположной по отношению к платформе.
3 этап. Воспроизведение пространственного навыка. Через 24 часа после второго дня обучения оценивают воспроизведение пространственного навыка: платформу убирают, и животных однократно помещают в бассейн на 60 сек. Регистрируют время, в течение которого животное находится в квадранте, где в дни обучения располагалась платформа, что служит показателем эффективности обучения и воспроизведения пространственного навыка.
4 этап. Сшибка пространственного навыка - переобучение и новое воспроизведение. На следующий день после воспроизведения пространственного навыка платформу перемещают в область бассейна, диагонально противоположную прежнему ее местоположению. Платформа должна быть погружена в воду, как и предыдущие дни обучения на 0,5 см ниже уровня воды. Повторяются процедуры обучения (2 дня) и воспроизведения по описанной выше схеме.
Результаты эксперимента
Влияние алмацина на обучение и воспроизведение пространственного навыка мышами линии Balb/C в водном лабиринте Морриса
В первый день обучения животных в водном лабиринте Морриса для мышей линии Balb/C было зарегистрировано достоверное увеличение времени поиска платформы (на 24,0%) и уменьшение количества результативных попыток (в 3,6 раза) по сравнению с аутбредными животными (Таблица 3).
Для групп мышей линии Balb/C, получавших алмацин в дозах 1 и 10 мг/кг, наноалмаз, глицина и трифтазин, достоверных различий с группой линейных мышей не обнаружено. При этом отмечена тенденция (р<0,1, критерий Стьюдента) к снижению времени поиска платформы мышами, получавшими алмацин в дозе 1 мг/кг, относительно контрольных мышей линии Balb/C (Таблица 3).
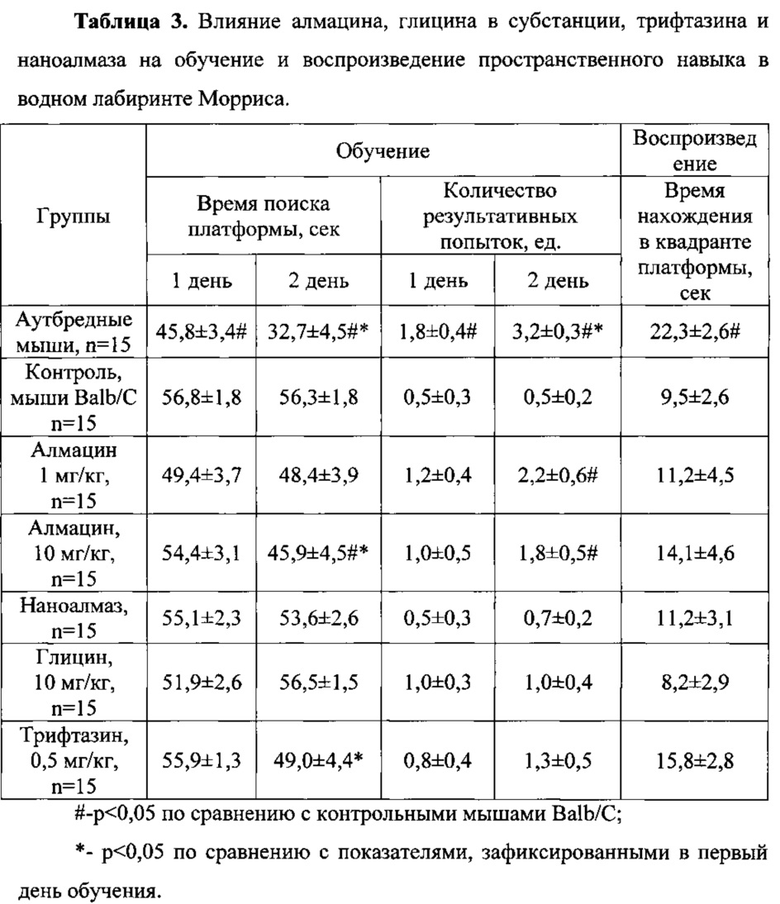
Во второй день обучения поведение линейных мышей также достоверно отличалось от поведения аутбредных животных по времени поиска платформы (увеличение на 72,2%) и по количеству результативных попыток (уменьшение в 6,4 раза). При этом внутригрупповые показатели первого и второго дня обучения мышей линии Balb/C не отличались между собой, тогда как аутбредные животные во второй день обучения достоверно быстрее находили платформу (на 28,6%) и совершали большее количество результативных попыток (в 1,8 раза) по сравнению с первым днем обучения (Таблица 3).
Животные, получавшие алмацин в дозе 10 мг/кг, затрачивали на 18,5% достоверно меньше времени на поиск платформы и совершали значимо (в 3,6 раза) большее количество результативных попыток по сравнению с контрольной группой. При этом алмацин в дозе 10 мг/кг положительно влиял на способность животных к обучению, о чем свидетельствует достоверное снижение времени поиска платформы (на 15,6%) и увеличение количества результативных попыток (на 80%) во второй день по сравнению с первым днем обучения.
В дозе 1 мг/кг алмацин достоверно увеличивал (в 4,4 раза) количество результативных попыток поиска платформы по сравнению с контрольной группой линейных мышей. При этом снижение времени поиска платформы на фоне введения препарата было на уровне тенденции (р<0,1, критерий Стьюдента) и составляло 14%.
Препарат сравнения трифтазин не оказывал влияния на обучение животных в водном лабиринте по сравнению с контрольной группой животных. Однако при внутригрупповом сравнении показано, что трифтазин во второй день обучения вызывал достоверное уменьшение времени поиска платформы на 12,3% и увеличение количества результативных попыток на 62,5% по сравнению с первым днем (Таблица 3).
При воспроизведении пространственного навыка продолжительность нахождения линейных мышей в квадранте, где при обучении находилась платформа, была достоверно в 2,3 раза меньше по сравнению с показателем аутбредных животных (Таблица 3). Продолжительность нахождения в данном квадранте животных, получавших исследуемые вещества, не достигала значимых различий от показателя в группе контрольных линейных мышей.
Влияние алмацина на поведение мышей линии Balb/C после пространственной сшибки (переобучения) в водном лабиринте Морриса.
Обнаружено, что непосредственно после «сшибки» (процесс переобучения мышей) время поиска платформы и количество результативных попыток у контрольных мышей линии Balb/C достоверно отличалось от группы аутбредных животных как в первый, так и во второй день переобучения (Таблица 4).
Для групп мышей линии Balb/C, получавших алмацин в дозе 1 мг/кг, наноалмаз, глицин и трифтазин, достоверных различий с контрольной группой не обнаружено по обоим регистрируемым параметрам как в первый, так и во второй дни переобучения (Таблица 4).
На фоне алмацина в дозе 1 мг/кг отмечено достоверное улучшение переобучения. Так, в первый день переобучения животные совершили в 2,4 раза больше результативных попыток нахождения платформы по сравнению с контрольными животными; во второй день количество результативных попыток превышало данный показатель контрольной группы на 80% (Таблица 4).
Мыши, получавшие алмацин в дозе 10 мг/кг, во второй день переобучения затрачивали достоверно меньше времени на поиск платформы и совершали достоверно большее количество результативных попыток по сравнению с первым днем переобучения, что свидетельствует о позитивном влиянии препарата при переобучении («сшибке») (Таблица 4).
При воспроизведении навыка, полученного после пространственной «сшибки», обнаружено, что время нахождения контрольных животных в квадранте, где находилась платформа при переобучении, достоверно меньше показателя у аутбредных мышей (Таблица 4). Алмацин в дозе 1 мг/кг достоверно в 3,4 раза увеличивал время пребывания животных в «правильном» квадранте. Достоверное увеличение этого параметра было также и в группе животных, получавших наноалмаз, которое составляло 90,1%. Препараты сравнения трифтазин и глицин, также как алмацин в дозе 10 мг/кг, не оказывали влияния на воспроизведение вновь приобретенного навыка.
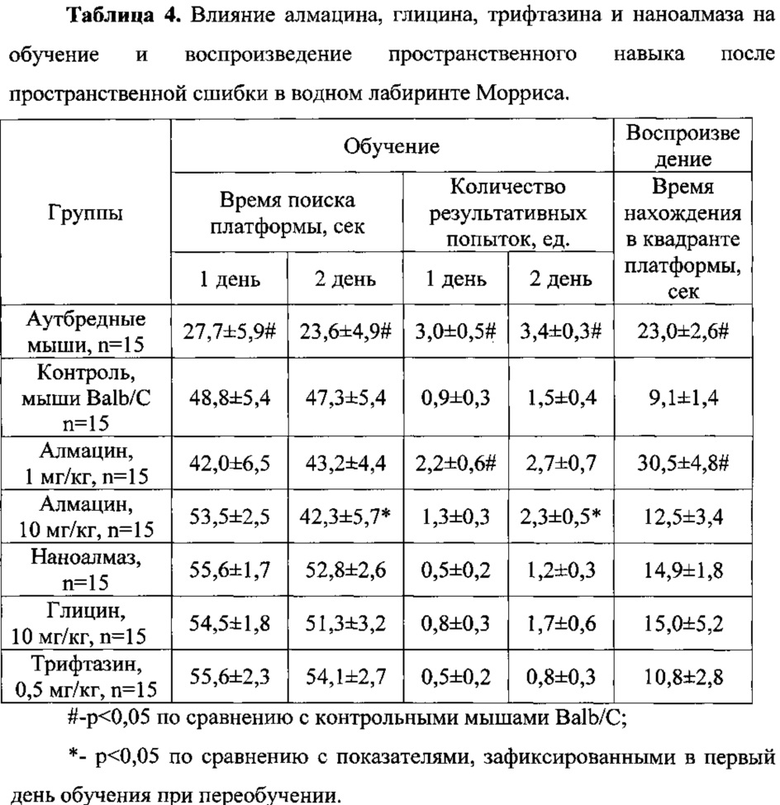
Таким образом, алмацин в дозе 10 мг/кг достоверно улучшает процессы обучения мышей линии Balb/C в условиях водного лабиринта Морриса как до, так и после пространственной сшибки. Алмацин в дозе 1 мг/кг улучшает процессы обучения мышей линии Balb/C только на второй день обучения. Препараты сравнения трифтазин и глицин не проявили влияние на процессы обучения животных. Все испытуемые вещества не оказывали влияние на воспроизведение полученного пространственного навыка. Однако при воспроизведении пространственного навыка в условиях «сшибки» пространственной памяти алмацин в дозе 1 мг/кг достоверно увеличивал время нахождения в квадранте платформы у мышей линии Balb/C.
Полученные в Примерах 1 и 2 результаты эксперимента свидетельствуют о наличии у заявляемого средства в дозах 1 и 10 мг/кг (при однократном введении) выраженной способности улучшать распознавание нового социального запаха и ускорять адаптацию к обонятельным стимулам в тесте обонятельной габитуации/дисгабитуации (Olfactory habituation/ dishabituation). По эффективности заявляемое средство в обеих дозах не уступает препарату сравнения трифтазину (0,5 мг/кг) и превосходит глицин (10 мг/кг). Детонационные наноалмазы не обладают способностью улучшать распознавание новых запахов. Заявляемое средство в дозе 10 мг/кг (введение в течение 6-ти дней ежедневно) достоверно улучшает процесс обучения мышей линии Balb/C в условиях водного лабиринта Морриса как до, так и после пространственной сшибки. Препараты сравнения трифтазин (0,5 мг/кг) и глицин (10 мг/кг), а также наноалмазы (введение в течение 6-ти дней ежедневно) не оказывают влияния на процессы обучения животных. В условиях сшибки пространственной памяти заявляемое средствов дозе 1 мг/кг (введение в течение 6-ти дней ежедневно) улучшает воспроизведение пространственного навыка, достоверно увеличивает время нахождения животных в квадранте платформы. Препараты сравнения трифтазин (0,5 мг/кг) и глицин (10 мг/кг), а также наноалмазы (введение в течение 6-ти дней ежедневно) не оказывают влияния на воспроизведение пространственного навыка в условиях сшибки пространственной памяти.
Список литературы
1. Яковлев Р.Ю. Конъюгат наноалмаза с глицином и способ его получения. Пат. РФ 2560700, 2015.
2. Рациональная фармакотерапия в психиатрической практике: руководство для практикующих врачей. Под общ. ред. Ю.А. Александровского, Н.Г. Незнанова. - М.: Литтера, 2014, с. 714-732.
3. Расстройства аутистического спектра у детей. Научно-практическое руководство. Под ред. Н.В. Симашковой. - М.: Авторская академия, 2013. 264 с.
4. Руководство по клинической психофармакологии / А.Ф. Шацберг, Дж.О. Коул, Ч. ДеБатиста. Пер. с англ.; под общ. ред. А.Б. Смулевича, C.B. Иванова. 2-е изд. – М.: МЕДпресс-информ, 2014. С. 175-220.
5. Roullet F.I., Crawley J.N. Mouse Models of Autism: Testing Hypotheses About Molecular Mechanisms / J.J. Hagan (ed.) Molecular and Functional Models in Neuropsychiatry. Curr. Topics in Behav. Neurosci. 2010. V.7. P. 187-212.
6. Moy S.S., Nadler J.J., Perez A. et al. Sociability and preference for social novelty in five inbred strains: an approach to assess autistic-like behavior in mice // Genes Brain Behav. 2004; 3(5): 287-302.
7. Moy S.S., Nadler J.J., Young N.B. et al. Mouse behavioral tasks relevant to autism: phenotypes of 10 inbred strains // Behav Brain Res. 2007. V. 176 №1. P. 4-20.
8. Moy S.S., Nadler J.J., Poe M.D. et al. Development of a mouse test for repetitive, restricted behaviors: relevance to autism // Behav. Brain Res. 2008. V. 188. №1. P. 178-194.
9. Nadler J.J., Moy S.S., Dold G. et al. Automated apparatus for quantitation of social approach behaviors in mice // Genes Brain Behav. 2004. V. 3. P. 303-314.
10. Kwon CH., Luikart B.W., Powell C.M. et al. Pten regulates neuronal arborization and social interaction in mice // Neuron. 2006 V. 50. P. 377-388.
11. Crawley J.N., Chen T., Puri A. et al. Social approach behaviors in oxytocin knockout mice: comparison of two independent lines tested in different laboratory environments // Neuropeptides. 2007 V. 41. №3. P. 145-163.
12. Chadman K.K., Gong S., Scattoni M.L.S. et al. Minimal aberrant behavioral phenotypes of neuroligin-3 R451C knockin mice // Autism Res. 2008. V. 1. P. 147-158.
13. Moon J., Beaudin AE., Verosky S. et al. Attentional dysfunction, impulsivity, and resistance to change in a mouse model of fragile X syndrome // Behav. Neurosci. 2006 V. 120. P. 1367-1379.
14. Машковский M.Д. Лекарственные средства. 16-е изд., перераб., испр. и доп. - М.: Новая волна: Издатель Умеренков, 2012. С. 661.
15. Комисарова И.Α., Нарциссов Я.Р. Молекулярные механизмы действия лекарственного препарата «Глицин» // Terra medica. 2001. №1. С 23-25.
16. Беспалов А.Ю., Звартау Э.Э. Нейропсихофармакология антагонистов NMDA-рецепторов. СПб.: Невский Диалект, 2000. 297 с.
17. Леонидов Н.Б., Яковлев Р.Ю., Лисичкин Г.В. Седативное средство и способ его получения. Пат. РФ 2506075, 2013.
18. Леонидов Н.Б., Яковлев Р.Ю., Соломатин А.С, Лисичкин Г.В. Антидепрессант и способ его получения. Пат. РФ 2519759, 2013.
19. Леонидов Н.Б., Яковлев Р.Ю., Лисичкин Г.В. Антипсихотическое средство и способ его получения. Пат. РФ 2519761, 2013.
20. Леонидов Н.Б., Яковлев Р.Ю., Кулакова И.И., Лисичкин Г.В. Анксиолитик и способ его получения. Пат. РФ 2519755, 2013.
21. Ryan B.C., Young N.B., Crawley J.N., et al. Social deficits, stereotypy and early emergence of repetitive behavior in the C58/J inbred mouse strain // Behav. Brain Res. 2010 V. 208. №1. P. 178-188.
22. South M., Ozonoff S., McMahon W.M. Repetitive behavior profiles in Asperger syndrome and highfunctioning autism // J. Autism Dev Disord. 2005 V. 35. P. 145-158.
23. Crawley J.N. Mouse behavioral assays relevant to the symptoms of autism // Brain Pathol. 2007 V. 17. P. 448-459.
| название | год | авторы | номер документа |
|---|---|---|---|
| Средства, улучшающие процесс обучения, памяти и когнитивные функции, а также для симптоматической терапии при аутистических расстройствах | 2015 |
|
RU2612791C1 |
| СРЕДСТВО ДЛЯ КОРРЕКЦИИ РАССТРОЙСТВ АУТИСТИЧЕСКОГО СПЕКТРА | 2017 |
|
RU2666598C1 |
| АНКСИОЛИТИК И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2013 |
|
RU2519755C1 |
| СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ И ПРОФИЛАКТИКИ АЛКОГОЛИЗМА | 2014 |
|
RU2574001C1 |
| СПОСОБ ПОЛУЧЕНИЯ СОПОЛИМЕРА НАТРИЙКАРБОКСИМЕТИЛЦЕЛЛЮЛОЗЫ И ГОССИПОЛА И ЕГО ПРИМЕНЕНИЕ В КОМПЛЕКСНОЙ ТЕРАПИИ ПАЦИЕНТОВ С АУТИСТИЧЕСКИМИ РАССТРОЙСТВАМИ И КОГНИТИВНЫМИ НАРУШЕНИЯМИ | 2010 |
|
RU2453559C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ ПРОТИВОИНСУЛИТНЫМ ДЕЙСТВИЕМ, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2013 |
|
RU2521404C1 |
| АНТИДЕПРЕССАНТ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2013 |
|
RU2519759C1 |
| АНКСИОЛИТИЧЕСКОЕ СРЕДСТВО И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ АНКСИОЛИТИЧЕСКОГО ДЕЙСТВИЯ | 1999 |
|
RU2155065C1 |
| Способ определения приоритета периферического зрения, характерного для людей с расстройством аутистического спектра | 2019 |
|
RU2724018C1 |
| ПРИМЕНЕНИЕ СЕЛЕКТИВНОГО АНТАГОНИСТА ГЛУТАМАТНЫХ РЕЦЕПТОРОВ Ro-256981 ДЛЯ НАПРАВЛЕННОГО ВЛИЯНИЯ НА КОГНИТИВНЫЕ ФУНКЦИИ | 2011 |
|
RU2496495C2 |
Изобретение относится к области медицины, в частности к психофармакологии, и касается средства для лечения и профилактики расстройств аутистического спектра, представляющего собой глицин, иммобилизованный на частицах детонационного наноалмаза размером 2-10 нм, с содержанием глицина от 1 до 21±3 мас.% Описываемое средство позволяет повысить результативность медикаментозного лечения и профилактики аутизма и расширить ассортимент эффективных и безопасных психотропных препаратов. 5 табл., 2 пр.
Средство для лечения и профилактики расстройств аутистического спектра, представляющее собой глицин, иммобилизованный на частицах детонационного наноалмаза размером 2-10 нм, с содержанием глицина от 1 до 21±3% масс.
| Н | |||
| В | |||
| СИМАШКОВА | |||
| Эффективная фармакотерапия и реабилитация больных с расстройствами аутистического спектра//Ж: Эффективная фармакотерапия | |||
| Неврология и психиатрия | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| АНТИДЕПРЕССАНТ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2013 |
|
RU2519759C1 |
| АНТИПСИХОТИЧЕСКОЕ СРЕДСТВО И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2013 |
|
RU2519761C1 |
| АНКСИОЛИТИК И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2013 |
|
RU2519755C1 |
| БОРОДИНА Л | |||
| Г | |||
| Лекарственная терапия расстройств аутичтисекого спектра у детей: опыт зарубежных психофармакологов//Ж: Аутизм и нарушения развития, N4 (39), 2012, с.1-18. | |||

