Изобретение относится к медицине, а именно к разработке и применению нового анксиолитического средства. Может быть использовано для лечения направленного на повышение мотивационной устойчивости и адекватного адаптивного поведения при психиатрических и неврологических заболеваниях для коррекции страха, тревоги, составляющих патогенетическую основу многих психоэмоциональных расстройств. Для лечения и реабилитации больных при нейрохирургических операциях и черепномозговых травмах, а также для повышения адаптационно-приспособительских возможностей здорового человека в экстремальных ситуациях.
Актуальность разработки препарата определяется возрастанием стрессовых ситуаций в современной жизни, связанных с техногенными и природными катастрофами, ухудшающейся экологической обстановкой, урбанизацией и нестабильностью политической и экономической обстановки в современном обществе, общей тенденцией к постарению общества и недостаточностью в современной медицине анксиолитиков, не обладающих побочными эффектами.
Известны в качестве анксиолитиков различные транквилизаторы, преимущественно бенздиазепинового ряда, нашедшие широкое применение в клинической практике. Однако они характеризуются большим числом побочных эффектов: седация, миорелаксация, гипо-седативное, снотворное действие, нарушение обучения, феномена отмены и другие.
Новым направлением в области создания эффективных и безопасных лекарственных средств для адекватного купирования страха, тревоги является создание анксиолитиков на основе эндогенных регуляторных пептидов, высокоэффективных и безвредных в силу их принадлежности к родственным организму биологическим структурам.
Известен гептапептид общей формулы (Thr-Lys-Pro-Arg-Pro-Gly-Pro) в качестве стимулятора памяти (Патент СССР N 1124544, приоритет от 30 июня 1983 г.). Однако возможность использовать его в качестве лекарственного препарата анксиолитического действия неизвестно. Техническим результатом, достигаемым при реализации изобретения, является обнаружение широкого спектра лечебного действия известного стимулятора памяти, что определяет возможность его использования в низких дозах в качестве анксиолитического средства, в виде удобной для применения лекарственной формы без нежелательных побочных эффектов, характерных для транквилизаторов, с хорошей переносимостью.
Выраженность анксиолитического действия этого нового лекарственного препарата в дозах 200-300 мкг/кг более сопоставимо с действием гидазепама (7 мг/кг), диазепама (0.3 мг/кг), феназепама (0.1 мг/кг), альпразолама (0.2 мг/кг) - любой из которых можно рассматривать в качестве прототипа предлагаемого нового лекарственного препарата. Однако в отличие от вышеперечисленных транквилизаторов, которые рассматриваются в качестве прототипа, увеличение дозы нового лекарственного препарата в 30-50 раз и до 200-500 раз в различных тестах не сопровождается развитием нежелательных побочных эффектов.
Вышеописанный технический результат достигается тем, что известный гептапептид общей формулы: Thr-Lys-Pro-Arg-Pro-Gly-Pro применяется в качестве анксиолитического средства.
В лекарственном составе анксиолитического средства, содержащего активное вещество и дистиллированную воду, в качестве активного вещества содержится гептапептид Thr-Lys-Pro-Arg-Pro-Gly-Pro и дополнительно консервирующее вещество нипагин при следующем соотношении компонентов, г/л:
Гептапептид Thr-Lys-Pro-Arg-Pro-Gly-Pro - 1,45-1,55
Нипагин - 0,95-1,05
Вода дистиллированная - Остальное
Лекарственной форме присвоено название Селанк.
Далее в тексте лекарственная форма будет называться Селанк, а активное вещество гептапептид Thr-Lys-Pro-Arg-Pro-Gly-Pro - просто гептапептид.
Установлено, что гептапептид Thr-Lys-Pro-Arg-Pro-Gly-Pro, известный как стимулятор памяти, обладает широким спектром действия. Наряду с анксиолитическим действием гептапептид оказывает оптимизирующее и активирующее действие на мнестические и когнитивные функции мозга, что составляет важную положительную часть его специфической фармакологической активности. Гептапептид активирует процессы обучения и памяти (фиксацию, консолидацию, воспроизведение у нормальных животных и при экспериментальной патологии этих функций мозга). Эти свойства отличают фармакологическое действие гептапептида от фармакологического действия транквилизаторов.
Список сокращений:
ВМГ - вентромедиальное ядро гипоталамуса
ОП - открытое поле
ОР - ориентировочные реакции
ПКЛ - приподнятый крестообразный лабиринт
ТСК - темно-светлая камера
УРПИ - условная реакция пассивного избегания
УРАИ - условная реакция активного избегания
ЭСР - эмоционально-стрессовая реакция
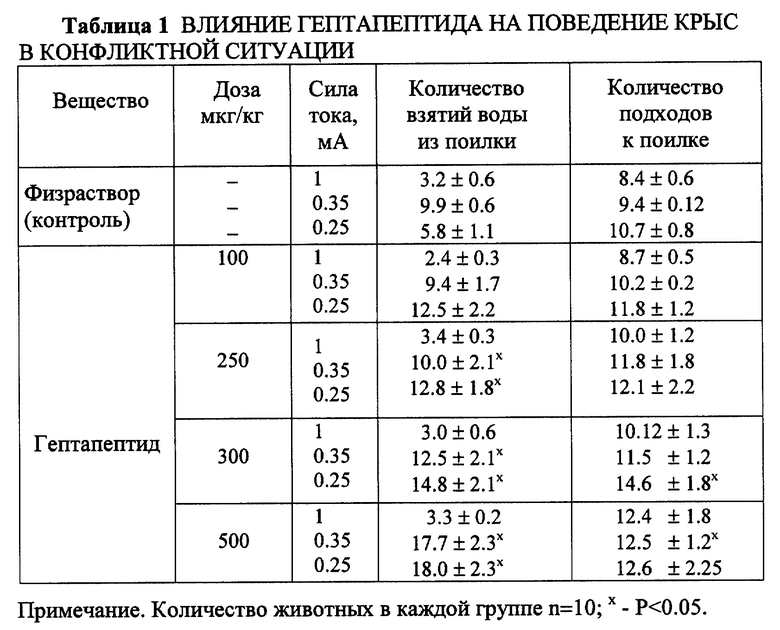
Пример 1. Влияние на поведение крыс в тесте конфликтной ситуации, вызванной столкновением оборонительного и пищевого рефлексов.
Метод конфликтной ситуации, основанной на столкновении питьевого и оборонительного рефлексов при действии внезапного болевого или аверсивного раздражителя, является общепринятым для выявления транквилизующего и анксиолитического действия соединений (Т.А. Воронина и соавторы, 1982). В зависимости от силы тока животное может получить интенсивное болевое ("повреждающее") раздражение или аверсивное ("наказующее", отвергаемое), эмоционально негативное.
Эксперимент начинали с выработки у крыс навыка взятия воды из поилки, установленной в экспериментальной камере. За 48 часов до опыта животных лишали воды и кормили только сухим кормом (питьевая депривация). Затем ежедневно (в течение 3 дней) в одно и то же время суток крыс помещали в экспериментальную камеру (на 10 минут), где они могли брать воду из поилки. На 4-й день обучения (после стабилизации питьевого рефлекса) крыс вновь помещали в экспериментальную камеру, но через пять секунд после начала питья наносили электрическое раздражение пропусканием тока через поилку. Использовались сила тока двух интенсивностей: 1 мА - надпороговое болевое раздражение; 0.25 и 0.35 мА - пороговое болевое или аверсивное, эмоционально-негативное. Столкновением двух рефлексов, питьевого и оборонительного, создавалась экстремальная конфликтная ситуация, при которой страх получения болевого (повреждающего) или аверсивного (негативного, отвергаемого) раздражения удерживало животных от взятия воды, несмотря на питьевую депривацию. В течение 20 минут регистрировали: количество взятий воды из поилки, несмотря на получение электрического раздражения; число подходов к поилке; общую двигательную активность и общее эмоциональное состояние по таким признакам, как: голосовые реакции, неадекватность реагирования на предъявляемые тест-стимулы, резкие колебания дыхания, видимая тахикардия. Гептапептид вводили интраназально в дозах от 100 до 500 мкг/кг. Для сравнения эффективности гептапептида с препаратом, обладающим анксиолитическим действием в этих условиях эксперимента, был использован транквилизатор бенздиазепинового ряда диазепам (в дозе 1 мг/кг) внутрибрюшинно.
Установлено, что гептапептид в дозах от 100 до 500 мкг/кг достоверно увеличивает количество взятий воды из поилки при использовании аверсивного раздражения (ток силой I = 0.25 мА и 0.35 мА). Наиболее эффективными оказались дозы 250 мкг/кг и 300 мкг/кг, при которых количество взятий воды из поилки возрастало в среднем в 2.3 раза, количество подходов без взятия воды - в 1.5 раза ( P < 0.05) (табл. 1).
Многие животные не отходили от поилки и часто совершали движения, напоминающие незавершенные попытки взятия воды. Внешне видимые проявления эмоционального напряжения (страха), наблюдаемые в контроле, сменялись исследовательской активностью с повторным возвращением к поилке. При использовании тока силой 1 мА (интенсивное болевое раздражение) количество взятия воды из поилки также возрастало, однако, не достигало критерия достоверности, тем не менее, животные чаще подходили к полке, визуально регистрируемые признаки страха-тревоги (пилоэрекция, голосовые реакции, мышечное напряжение) устранялись. У животных, получавших диазепам (1 мг/кг), наблюдалось высокодостоверное (P < 0.01) увеличение числа попыток взятия воды (фиг. 1) как при использовании аверсивного, так и интенсивного болевого раздражения. Несмотря на повторные удары током, животные не только подходили, но и брали воду из поилки.
Таким образом, результаты свидетельствуют, что на модели конфликтной ситуации гептапептид оказывает действие, аналогичное классическому транквилизатору диазепаму и ассоциируемое с устранением страха получения аверсивного воздействия. Однако, при 4-кратном увеличении интенсивности силы тока эффект гептапептида ослабевает, тогда как у диазепама сохраняется. Данное наблюдение отражает, по-видимому, факт сохранности адекватности оценки ситуации на фоне действия гептапептида, несмотря на уменьшение негативности ее восприятия.
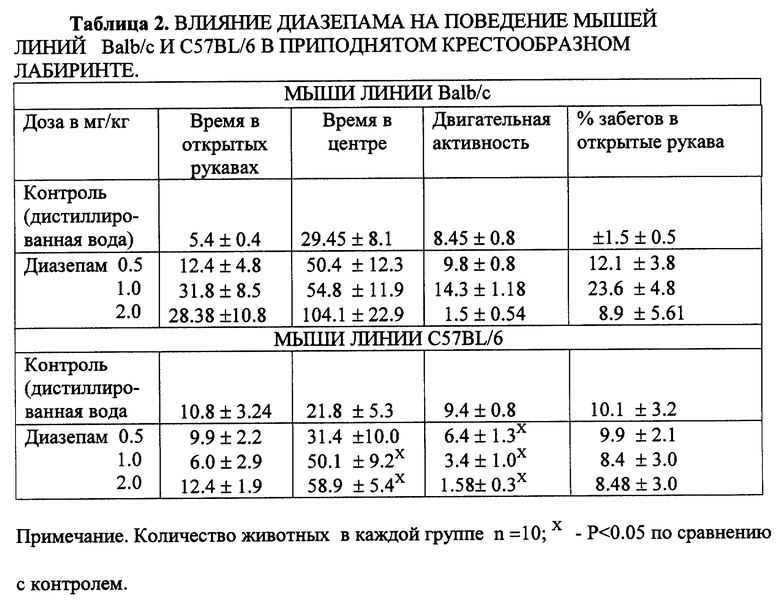
Пример 2. Влияние на поведение мышей с наследственно контролируемым фенотипом эмоционально-стрессовой реакции в тестах приподнятый крестообразный лабиринт (ПКЛ).
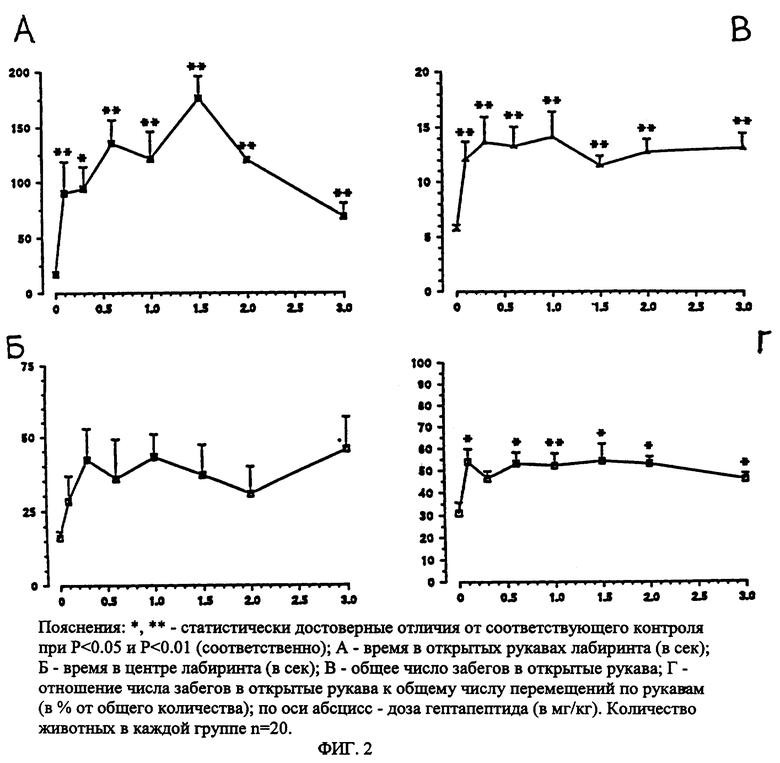
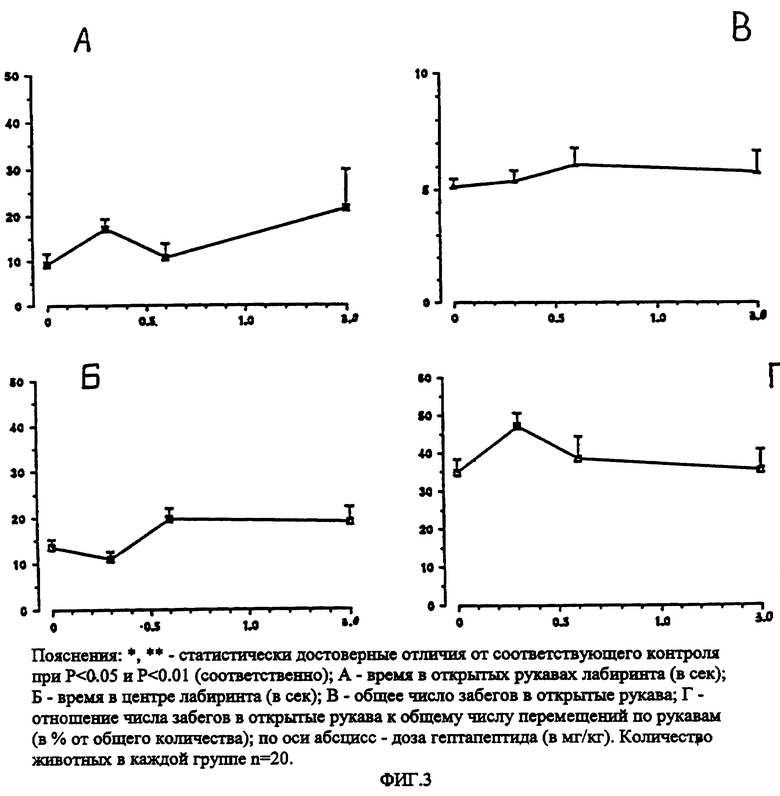
Метод "приподнятый крестообразный лабиринт" (ПКЛ) используется в настоящее время в мировой и отечественной экспериментальной психофармакологии в качестве адекватного теста для выявления анксиолитических свойств известных препаратов и новых соединений (Pellow Sh. et. al.,1985; Ю.А. Бледнов и соавторы, 1994; S. B. Seredenin et. al, 1995). Негативная ситуация создается столкновением двух противоположных естественных для грызунов форм поведения разной модальности: тревога и поисковая активность. Важно, что эксперимент идет без применения вредных (noxions) или повреждающих стимулов, таких как электрошок, болевое раздражение, манипуляции с пищевой депривацией и др. Эксперименты выполнены на мышах с "пассивным" фенотипом ЭСР и высоким уровнем тревоги (линия Balb/c) и "активным" фенотипом ЭСР (линия C57BL/6) (С.Б. Середенин, Ведерников, 1979; С.Б. Середенин и соавторы, 1989; Ю.А. Бледнов и соавторы, 1994). Согласно стандартной программе в опытах регистрировалось: время пребывания в "открытых" (светлых) и "закрытых" (темных) рукавах ПКЛ, общее время пребывания в светлых отсеках, в темных рукавах, общее число перемещений и соотношение числа забегов в светлые рукава к общему числу забегов, длительность наблюдения - 5 минут. Гептапептид вводили в дозах от 100 до 6000 мкг/кг внутрибрюшинно за 30 минут до тестирования. Статистическая обработка приведена с использованием метода однофакторного дисперсивного анализа с последующим межгрупповым сравнением по методу Ньюмена-Кьюпса (фиг. 2 и 3).
Установлено, что гептапептид во всем диапазоне испытанных доз от 100 до 3000 мкг/кг дозозависимо увеличивает время пребывания мышей линии Balb/c в светлых рукавах и центре ПКЛ (фиг. 2 А, Б). Число забегов в светлые рукава ПКЛ возрастает почти в 3 раза.
Можно видеть, что диазепам в дозах 0.5-1.0 мг/кг оказывал на поведение мышей линии Balb/c в тесте ПКЛ сходное с гептапептидом действие, т.е. увеличивая время пребывания в открытых рукавах лабиринта, повышая (хотя и менее значительно) двигательную активность и процент забегов в открытые рукава (табл. 2); однако, в дозе 2 мг/кг оказывал, в отличие от гептапептида, достовернозначимое седативное действие: общая двигательная активность резко снижалась, увеличивалось время задержки в центре лабиринта. На поведение мышей линии C57BL/6 диазепам оказывал седативное действие в меньших дозах 0.5-1.0 мг/кг (табл. 2).
Таким образом в тесте ПКЛ гептапептид подобно диазепаму оказывал анксиолитическое действие на поведение мышей линии Balb/c, гептапептид практически не влиял на поведение мышей линии C57BL/6 в тесте ПКЛ, тогда как диазепам оказывал седативное действие на мышей данной линии уже в дозе 0.5 мг/кг. Анксиолитическое действие гептапептида на поведение мышей линии Balb/c не сопровождалось снижением двигательной активности даже при 30-кратном увеличении дозы, тогда как 2-кратное увеличение дозы диазепама вызывало снижение двигательной активности. Куполообразная зависимость "доза-эффект", наблюдаемая при введении диазепама, отражает, по мнению ряда авторов (Ю.А. Бледнов и соавт. , 1995), развитие общеседативного и миорелаксантного действия, типичного для бенздиазепинов. Отсутствие такого рода зависимости в действии гептапептида характеризует его как анксиолитик, лишенный побочных (седативного, миорелаксантного) эффектов бенздиазепиновых транквилизаторов.
Таким образом, использование фармакогенетического методологического подхода и высокоспецифического для выявления анксиолитического действия препаратов теста ПКЛ позволило установить наличие и селективность (в отношении побочных эффектов бенздиазепинов) анксиолитического действия изучаемого препарата. В широком диапазоне доз (от 100 до 3000 мкг/кг) без каких-либо побочных эффектов анксиолитическое действие гептапептида в тесте ПКЛ формируется у мышей с исходно пассивной формой ЭСР и высоким уровнем тревожности. В том же диапазоне доз гептапептид практически не меняет поведение мышей с активной формой ЭСР и исходно низкой тревожностью.
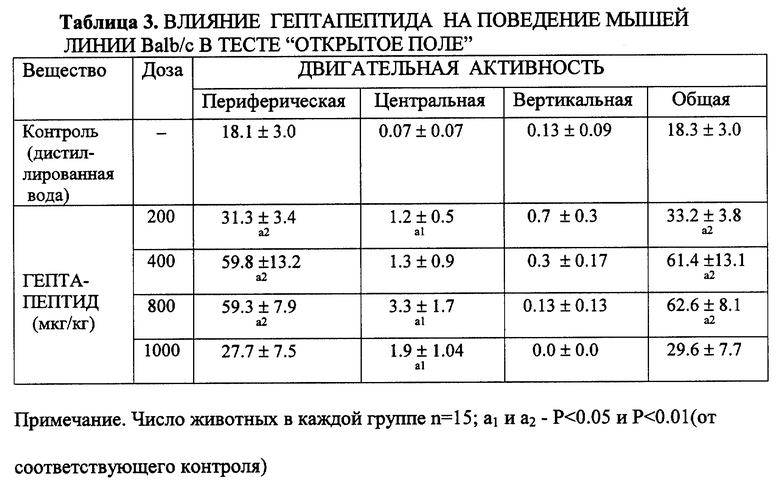
Пример 3. Влияние на реакцию затаивания в тестах открытое поле (ОП) и темно-светлая камера (ТСК) (реакция предпочтения).
Известно, что в основе психоэмоциональной реакции на стресс лежит эмоционально-негативное состояние, страх и тревога. Характер и сила эмоционально- стрессовой реакции является наследственно контролируемой. Эксперименты выполнены на мышах линии Balb/c и C57BL/6 с противоположным наследственноконтролируемым "пассивным" и "активным" фенотипом ЭСР. Использовались две стандартные модели воспроизведения стресс-реакции у мышей: неизбегаемый стресс по методу Grawby I.N. с использованием установки "темно-светлая камера" (ТСК) и тест открытое поле (ОП).
В опытах регистрировалось: число пересеченных квадратов на периферии и в центральных областях, число вертикальных стоек, суммарно-общее число перемещений (общая двигательная активность). Настоящие и ранее выполненные исследования показали, что поведение мышей линий Balb/c (с) и C57BL/6 (B6) в стресс-ситуации ОП принципиально отличается. У мышей линии с возникает фризинг-реакция. Затаивание у стенки лабиринта, очень низкая двигательная активность в пределах 1-3 квадратов в периферической части ОП, стойки практически отсутствуют, выходов в центральные части ОП не бывает. У мышей линии B6, напротив, активность была более чем в 6 раз выше, наблюдались частые стойки и выходы в центральные отделы ОП.
Гептапептид вводили внутрибрюшинно в дозах от 200 до 1000 мкг/кг за 30 минут до тестирования. Установлено, что во всем диапазоне испытанных доз гептапептид оказывает выраженное антистрессорное действие только у мышей линии с, с "пассивным" фенотипом ЭСР (табл. 3, 4). В условиях ОП пептид устраняет экологически-значимые проявления страха-тревоги. Фризинг-реакция не возникает, двигательная активность возрастает в 2 раза, горизонтальные перемещения происходят как в периферической, так и центральной зоне, причем, животное повторно пересекает центр поля. Возрастает количество вертикальных стоек (табл. 3).
В том же диапазоне доз гептапептид не оказывает достоверного влияния на поведение мышей с исходно "активным" фенотипом ЭСР (табл. 4).
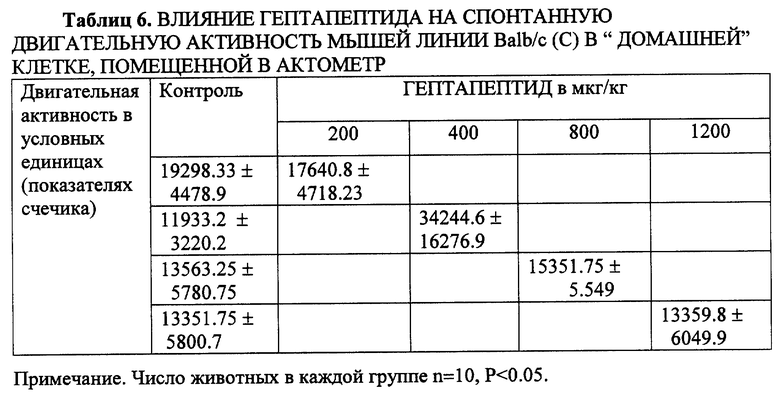
Гептапептид не оказывал также достоверного влияния на спонтанную двигательную активность мышей линии с, находящихся в "домашней клетке", вне конфликтной стрессогенной ситуации (табл. 6). И не вызывал седации, угнетения, миорелаксации, сопровождающих, как известно, анксиолитическое действие известных транквилизаторов на поведение мышей и крыс в тесте ОП.
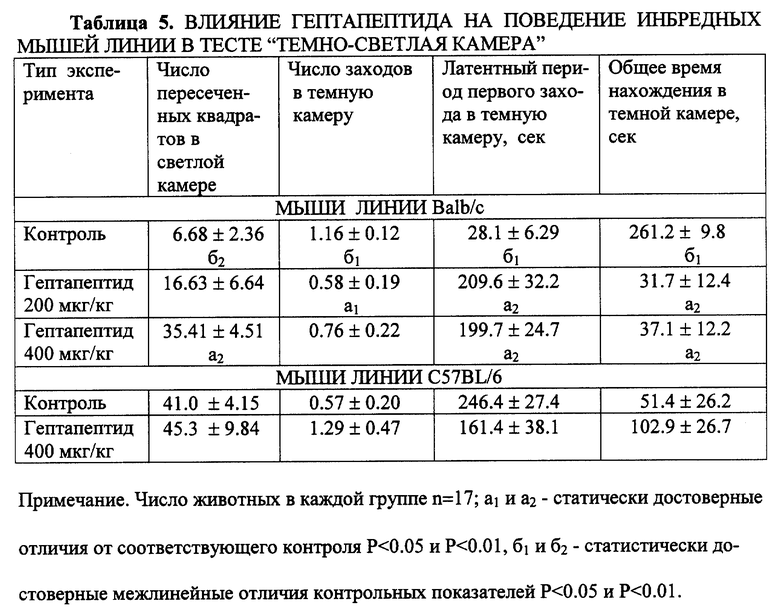
Было изучено влияние гептапептида на поведение мышей обеих линий в тесте темно-светлая камера (реакция "предпочтения" или избегаемый стресс). Реакцию моделировали по методу Grawby I.N. с использованием установки "темно-светлая камера" (ТСК). Гептапептид вводился по той же схеме в дозах 200 и 400 мкг/кг. Регистрировалось: общая двигательная активность по числу пересеченных квадратов, число заходов в темную камеру, латентный период первого захода и общее время нахождения в светлой или темной частях камеры. Длительность наблюдения - 5 минут. Исходное поведение мышей линии с и В6 в данной стресс-ситуации принципиально различалось. У мышей, исходно тревожных (линия с), число пересеченных квадратов было в 5 раз меньше, латентный период захода в темный отсек - в 8-9 раз короче, чем у мышей линии В6. Обнаружив возможность избегания стресс-ситуации путем захода в темную камеру, мыши уже не выходили из нее. Время пребывания в темной камере достигало 87% от общего времени наблюдения. Мыши линии В6, напротив, долго обследовали территорию, не заходя в темную часть установки ТСК, время нахождения в темной камере составляло 17% от общего времени.
Установлено, что гептапептид принципиально изменял поведение высокотревожных животных линии с в данной стрессогенной ситуации. Время пребывания в темной камере сокращалось в 7 раз, причем большая часть животных вообще не покидала светлую камеру, продолжая обследовать ярко освещенную территорию. Латентный период первого захода возрастал почти в 30 раз. У мышей линии В6 достоверного активирующего влияния не обнаруживалось (табл. 5).
Таким образом, на мышах с высокотревожным генетически детерминированным "пассивным" типом эмоционально-стрессовой реакции (линия Balb/c) выявлено анксиолитическое действие гептапептида в ситуациях открытого поля (ОП) и темно-светлой камеры (ТСК). У мышей с "активной" формой ЭСР (линия C57BL/6) анксиолитическое действие гептапептида на тех же моделях и в тех же диапазонах доз не выявляется. В отличие от действия диазепама, анксиолитический эффект гептапептида развивается без побочных эффектов.
С целью дополнительного анализа избирательности анксиолитического действия гептапептида, который у мышей линии Balb/c сопровождается "активацией" двигательной активности в открытых рукавах ПКЛ, устранением фризинг-реакции и повышением двигательной активности в тестах ОП и ТСК, была проведена специальная серия экспериментов по выявлению действия препарата на спонтанную двигательную активность. Эксперименты выполнялись без перемещения мышей в незнакомую ситуацию, т.е. группа из 10 животных, находящихся в знакомой ("домашней") клетке, устанавливалась на специальную платформу (с опорой на пьезодатчики), снабженную также фотоэлементами, регистрирующими перемещения мышей с фиксацией на самописце. Как видно из табл. 6, гептапептид в широком диапазоне доз (от 200 до 1200 мкг/кг) не оказывал достоверного влияния на спонтанную двигательную активность животных, при их спокойном состоянии в "домашней" клетке.
Таким образом, действие гептапептида не является результатом прямого активирующего влияния на двигательную активность, поскольку вне стресс-ситуации спонтанная двигательная активность на фоне тех же доз достоверно не изменяется. Данные отражают избирательность анксиолитического действия гептапептида, поскольку устранение тревоги-страха и "активация поведения" в стресс-ситуации выявляется только на мышах линии Balb/c (высокотревожных, с "пассивным" фенотипом ЭСР). У мышей обеих линий не наблюдается также и достоверного снижения двигательной активности даже на фоне очень больших доз препарата.
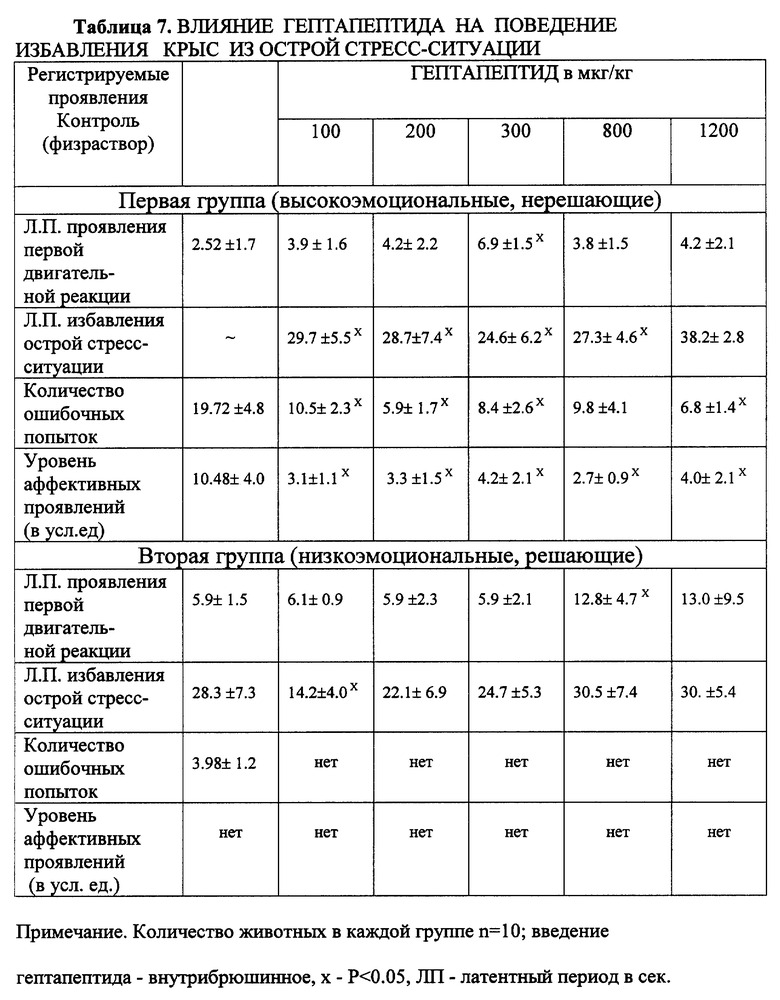
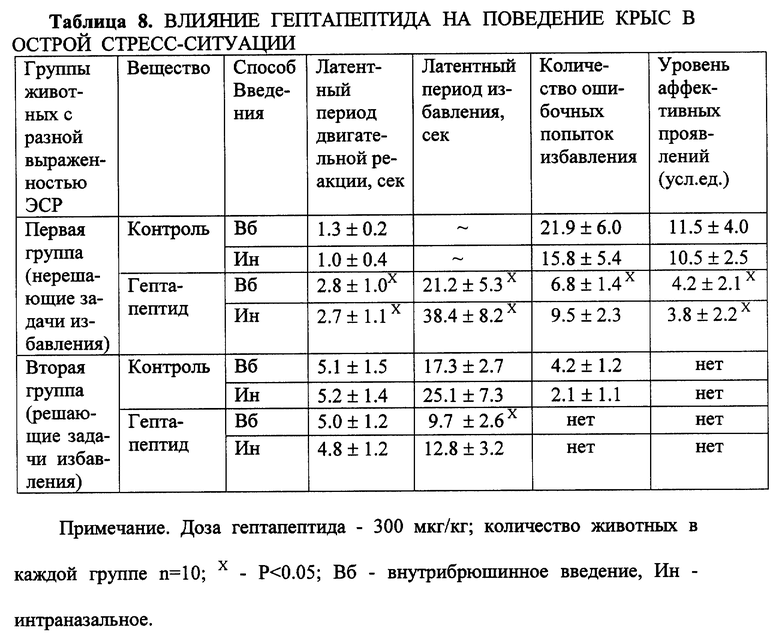
Пример 4. Влияние на реакцию избавления крыс разной топологической принадлежности в тесте подныривания по Хендерсону.
Для изучения влияния гептапептида на эмоционально-стрессовую реакцию крыс и способность к реализации избавления из стресс-ситуации использовали метод, предложенный Hendtrson (1970) в модификации Н.А. Бондаренко (1980). Животных помещали в стеклянный цилиндр диаметром 12 см, высотой 22 см. Нижний конец цилиндра погружен в "бассейн" с водой (глубина бассейна - 50 см) на глубину 2.5 см. На внутренней стороне бассейна помещена лесенка. Животное, погруженное в цилиндр, заполненный водой (хвостом вниз), могло, преодолев страх, погрузиться с головой в воду и, поднырнув под стенки цилиндра, найти лесенку и залезть на край бассейна. Такой сложный путь избавления требует от животного решения, которое модулируется его эмоциональным состоянием. Животное помещалось в цилиндр на 2 минуты. Регистрировали: латентный период первой попытки к избавлению, количество безуспешных попыток избавления (ошибки), количество ориентировочных движений в цилиндре, уровень аффективных проявлений (вокализация). Гептапептид вводили в дозах от 70 до 1200 мкг/кг внутрибрюшинно и, в серии экспериментов, интраназально и внутрибрюшинно в наиболее эффективной дозе - 300 мкг/кг.
Установлено, что гептапептид в диапазоне доз от 100 до 1200 мкг/кг оказывал "успокаивающее" (транквилизующее) действие на поведение высокоэмоциональных животных первой группы: выраженность аффективных реакций уменьшалась, количество ошибочных действий достоверно снижалось, практически все животные быстро реализовали реакцию избегания, поднырнув под стенки цилиндра (табл. 7).
Отмечается неярковыраженная дозозависимость эффекта с оптимальным выражением в дозе 300 мкг/кг. На фоне большой дозы (1200 мкг/кг) общее количество движений животного в цилиндре уменьшается, латентный период реализации реакции избавления возрастает, однако сама реакция осуществляется четко. У животных второй группы, с низким уровнем ЭСР, в данной ситуации несколько увеличивается латентный период избавления, при этом увеличение дозы до 1200 мкг/кг не оказывает дополнительного влияния на поведение. В отдельной серии экспериментов было сопоставлено действие гептапептида в наиболее эффективной дозе (300 мкг/кг) при внутрибрюшинном и интраназальном способах введения (табл. 8).
Установлено, что различия в действии гептапептида при разных способах его введения крысам не выявляется, что подтверждает аналогичные результаты, полученные на других экспериментальных моделях.
Таким образом, гептапептид оказывал выраженное антистрессорное действие, способствовал реализации поведения избавления крыс из острой стресс-ситуации. При этом динамика компонентов стресс-обусловленного поведения на фоне гептапептида зависела от особенностей индивидуально- типологического реагирования животных на стресс-ситуацию.
Данные свидетельствуют о модулирующем действии гептапептида на эмоциональную реактивность и его способности оптимизировать поведение избавления из стресс-ситуации.
Пример 5. Влияние на реакцию тревоги-страха в тесте реакция избегания (escape reaction) при электрической стимуляции "триггерных зон страха" вентромедиального ядра гипоталамуса (ВМГ) кроликов.
Метод прямой электрической стимуляции генетически детерминированных мотивационных центров гипоталамуса традиционно используется в экспериментальной психофармакологии (А. В. Вальдман, М.М. Козловская, 1969; А.В. Вальдман и соавторы, 1976; 1979; К.В. Судаков, 1971, 1996). При электрическом раздражении "центров страха" вентромедиального гипоталамуса (К.В. Судаков, 1996) у кроликов возникает оборонительное поведение, замирание, прыжок, побег и длительно сохраняющееся состояние тревоги.
В наших экспериментах изучение анксиолитического, противотревожного действия гептапептида проводилось на кроликах-самцах породы Шиншилла (массой 3000 - 3200 г) с биполярными электродами, стереотоксически вживленными в вентромедиальное ядро гипоталамуса по ранее опубликованному методу (В.И. Бадиков и соавторы, 1985). Параметры тестирующих стимулов: частота и длительность прямоугольных импульсов - 100 Гц и 0.3 Мсек, длительность повторных (через каждые 10 минут) раздражений 1-4 секунды (в зависимости от реакции животного), при общей длительности опыта 3 часа. Тестирующая амплитуда раздражения (сила тока) подбиралась индивидуально до получения реакции "прыжок-побег" и сохранялась постоянной в течение всего эксперимента. В 1-4-секундный интервал прямой стимуляции ВМГ у кролика возникало моментальное настораживание типа "alarm", прыжок и побег (escape reaction), ограниченный размером экспериментальной камеры. В 10-минутные интервалы между раздражениями сохранялась реакция эмоционального напряжения, тревоги и трансформация общей типичной для животного структуры поведения.
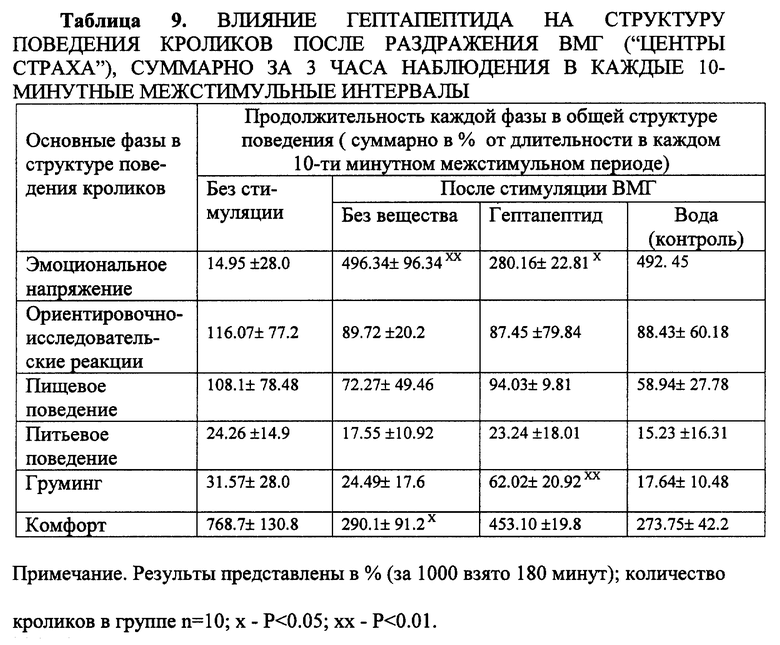
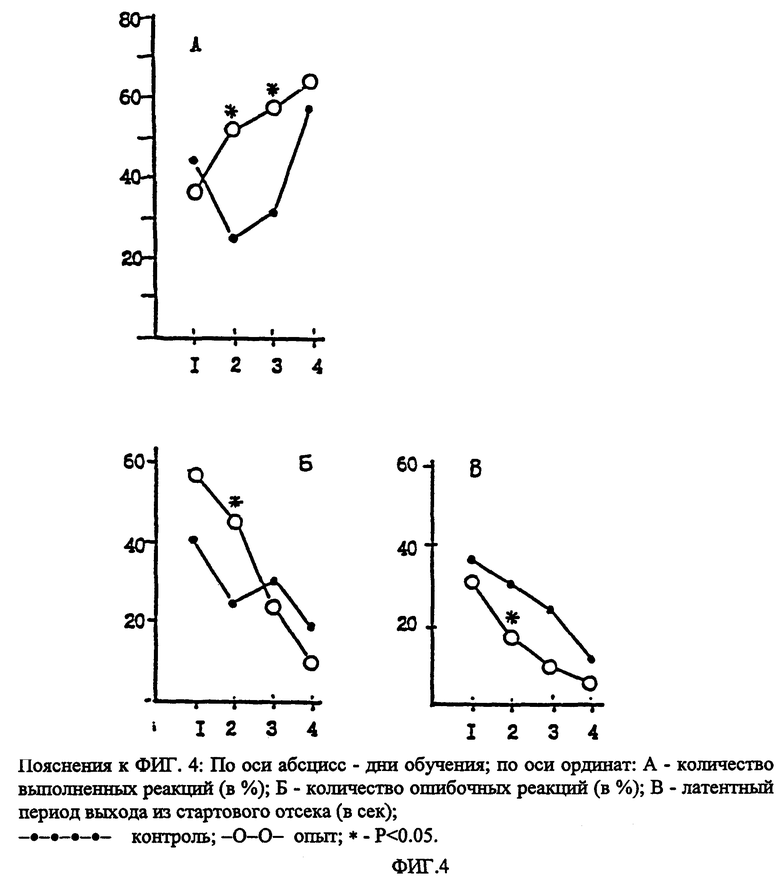
В опытах регистрировалось: латентный период реакции побега и изменения в структуре поведения, отражающие состояние эмоционального напряжения и тревоги. Каждые 10 минут межстимульного периода хронометрировалась продолжительность (в секундах) разных фаз (паттернов) поведения, входящих в общую структуру спонтанного поведения кроликов. Учитывались следующие фазы: эмоциональное напряжение (характерная неподвижная поза на выпрямленных лапах с высоко поднятыми ушами, мышечным напряжением, резким вздрагиванием и побегом на афферентные стимулы), ориентировочная реакция (ОР), агрессивность (спонтанное или спровоцированное стучание лапами, кусание предметов), груминг, пищевая (подход к кормушке, еда), питьевая (подход к поилке, питье) реакции, состояние "покоя" или "комфорта" (спокойное бездействие на грани засыпания, мышечное расслабление, прикрывание глаз). У каждого животного в 1-й день наблюдения фиксировали типичную структуру, сменяемость и продолжительность деленных шести фаз спонтанного поведения, во второй день - динамику изменения структуры поведения на фоне тревоги, вызванной стимуляцией, в третий день - то же после предварительного введения гептапептида или дистиллированной воды (контроль).
Гептапептид вводился в дозе 200 мкг/кг интраназально за 15 минут до тестирования. Математическая обработка результатов проводилась индивидуально по каждому животному и с оценкой действия препарата в группе животных (10 кроликов). Учитывалось абсолютное (в сек) и относительное (в %) содержание каждой фазы поведения в составе каждого 10-минутного отрезка времени, а также суммарно в пределах первого, второго и третьего часа наблюдения. Установлено, что электрическая стимуляция ВМГ ("центров страха"), вызывающая реакцию побега (escape reaction) с латентным периодом 1-2 секунды, достоверно изменяет структуру поведения кроликов (табл. 9).
Доминирующей (P < 0.05) становится фаза эмоционального напряжения, которая является компонентом эмоционального стресса и тревоги. Общая длительность фазы эмоционального напряжения возрастает почти в 40 раз, тогда как паттерны поведения, ассоциируемые с состоянием покоя, комфорта или удовлетворением собственных мотиваций (еда, питье), подавляются, продолжительность их сокращается более чем в 5 раз (суммарно). На фоне гептапептида длительность фазы эмоционального напряжения сокращается в 2.5 раза (P < 0.05), тогда как длительность фазы комфорта, напротив, возрастает в 2.3 раза (P < 0.05). Увеличивается длительность фаз пищевого и питьевого поведения. Данные, представленные в табл. 9, в общем виде иллюстрируют положительную динамику восстановления структуры спонтанного поведения кроликов (n = 10) на фоне гептапептида и достоверное снижение уровня эмоционального напряжения в 3-часовой межстимульный период. При прямой стимуляции ВМГ ("центры страха") на фоне гептапептида не подавляется полностью, однако, латентный период ее реализации возрастает (фиг. 4). Таким образом анксиолитическое действие гептапептида было выявлено при разных способах введения (внутривенно, внутрибрюшинно, интраназально) при использовании общепринятых тестов. Анксиолитический эффект сопоставим с действием известных транквилизаторов (феназепама, альпразолама, гидазепама). Данные выявили селективность анксиолитического действия и отсутствие побочных эффектов (миорелаксации, седации, сонливости), свойственные бенздизепинам. Таким образом гептапептид в широком диапазоне доз (от 100 до 6000 мкг/кг) оказывает избирательное (селективное) анксиолитическое действие.
Проведенные исследования показали, что гептапептид не только не вызывает побочных эффектов, свойственных транквилизаторам (миорелаксацию, седацию, сонливость), но и обладает выраженным положительным влиянием на мнестические функции, что подтверждают следующие примеры.
Пример 6. Влияние на условный рефлекс пассивного избегания.
Выработку условного рефлекса пассивного избегания (УРПИ) проводили по модифицированной методике Буреша. Экспериментальная установка состояла из 2 камер равной величины (25х25) с электродным полом разделенным перегородкой с небольшим отверстием для перехода крысы из одной камеры в другую.
Мышь помещали в светлую камеру хвостом к переходному отверстию и в течение 3 минут регистрировали время пребывания в светлой и темной камере и латентный период первого перехода в темную камеру. Затем в темной камере крыса получала через электродный пол однократный удар током силой 1.5 мА от стимулятора, имеющего выход по току с цифровой индексацией в миллиамперах. Раздражение продолжали до момента выхода крысы в светлую камеру через переходное отверстие (обучение). Тест на воспроизведение УРПИ проводили через 24 часа после обучения. Установлено, что гептапептид в дозе 300 мкг/кг облегчал выработку УРПИ. Достоверно (P < 0.05) увеличивал латентный период входа животных в темную камеру и суммарное время пребывания в ней в течение всего периода тестирования (3 минуты). При проверке сохранности рефлекса через 24 часа после обучения число животных, достигших критерия обученности (> 300 с) в опытной группе, составляло 151% от контроля (P < 0.05), что свидетельствовало о лучшем запоминании экспериментальной ситуации (табл. 10).
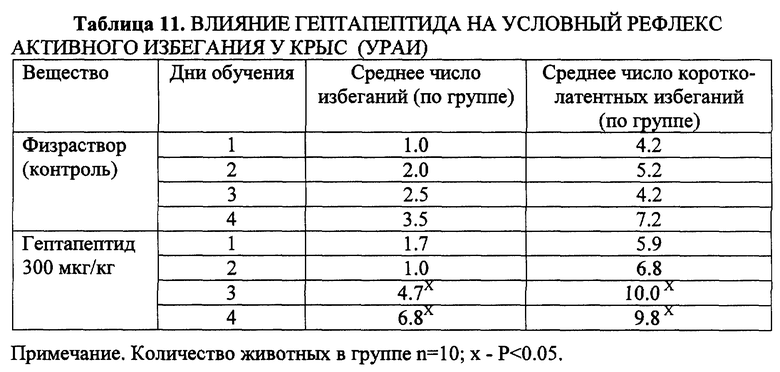
Пример 7. Влияние на условный рефлекс активного избегания (УРАИ).
Выработку условного рефлекса активного избегания (УРАИ) проводили с использованием челночной камеры производства фирмы "Ugo-Basile" (Италия). Камера состояла из двух равных отсеков, разделенных перегородкой и отверстием диаметром 9 см. Безусловное электрокожное (болевое) раздражение подавалось через пластинки электродного пола на лапы животного. Продолжительность каждого раздражения - 4 секунды. Изолированное действие условных сигналов (свет и звук) составляло 4 секунды, интервал между предъявлениями условных сигналов составлял 2 секунды. Для избегания болевого воздействия крыса должна обучиться переходить смежный отсек камеры в течение 4 секунд после включения условных сигналов. Параметры тока: частота 50 Гц, сила тока 0.5-0.8 мА, в зависимости от индивидуальной выраженности ответа.
УРАИ вырабатывали в течение 4 дней по 25 сочетаний раздражителей ежедневно. Регистрировали: количество условных реакций избегания и количество коротколатентных реакций избавления (< 1с). Гептапептид вводили внутрибрюшинно за 15 минут перед каждым сеансом обучения в дозе 300 мкг/кг.
Установлено, что гептапептид в дозе 300 мкг/кг облегчал выработку УРАИ: на третий и четвертый день обучения отмечалось достоверное (P < 0.05) увеличение числа избеганий в сравнении с контрольными животными. Увеличивалось также число коротколатентных реакций избавления, что свидетельствовало о готовности животных к выполнению данной (целенаправленной) деятельности (табл. 11).
Пример 8. Влияние на условный пищевой лабиринтный рефлекс.
Изучение влияния гептапептида на условный рефлекс с положительным пищевым подкреплением проводилось с использованием сложного многоходового лабиринта. Предварительно обучали крыс находить кормушку с пищей. Обучение проводили на животных с относительной пищевой депривацией в течение 4 дней. Ежедневно задача поиска пищи предъявлялась 5 раз, длительность каждой попытки не превышала 5 минут (в противном случае задача считалась невыполненной). В качестве подкрепления использованы хлебные шарики диаметром 10 мм. Регистрировали: количество выполненных реакций в каждый день опыта и латентный период их выполнения; число ошибок, т.е. число заходов в тупиковые коридоры лабиринта; количество невыполненных реакций (ограничение - 5 минут нахождения в лабиринте). Латентный период выхода из стартового отсека. Гептапептид вводили внутрибрюшинно повторно за 30 минут до каждого сеанса обучения в дозе 300 мкг/кг.
Установлено, что гептапептид оказывает значительное и достоверное облегчающее действие на формирование сложного условно-пищевого рефлекса, обеспечивая ускорение его образования. Со 2-го дня сеанса обучения у опытных животных наблюдается достоверное (P < 0.05) увеличение числа выполненных пищедобывательных реакций; число обучившихся животных возрастает в 1.5-2 раза (по сравнению с контрольной группой). Латентный период выхода из стартового отсека лабиринта прогрессивно сокращается при повторных введениях гептапептида.
Расширенное экспериментальное изучение действия гептапептида на обучение и память позволяет сделать заключение о выраженном положительном влиянии на мнестические процессы, связанные с фиксацией консолидацией и воспроизведением.
В представленных выше примерах гептапептид вводили преимущественно в виде водного раствора. Как указывалось выше, лекарственная форма под названием Селанк представляет собой 0.15% водный раствор гептапептида с добавлением 0.1% нипагина во флаконах-капельницах по 3 мл. Результаты, полученные в процессе изучения лекарственной формы Селанк, позволили определить оптимальный диапазон доз для проявления специфического фармакологического действия Селанка, которое полностью совпадает с вышеописанным действием гептапептида.
Установлено, что в диапазоне доз 150-400 мкг/кг наблюдается характерная зависимость "доза-эффект" с нарастанием активности к дозам 250-300-400 мкг/кг и установлением "плато-активности" при дальнейшем увеличении дозы. Установлено, что Селанк не обнаруживает явлений токсичности при введении в дозах, в 200 и более раз превышающих терапевтическую.
Следующие примеры демонстрируют идентичность фармакологического действия гептапептида и Селанка, которые были получены при проведении экспериментов, выполненных в один и тот же день, на одной и той же партии экспериментальных животных при соблюдении стандартного режима проведения опытов.
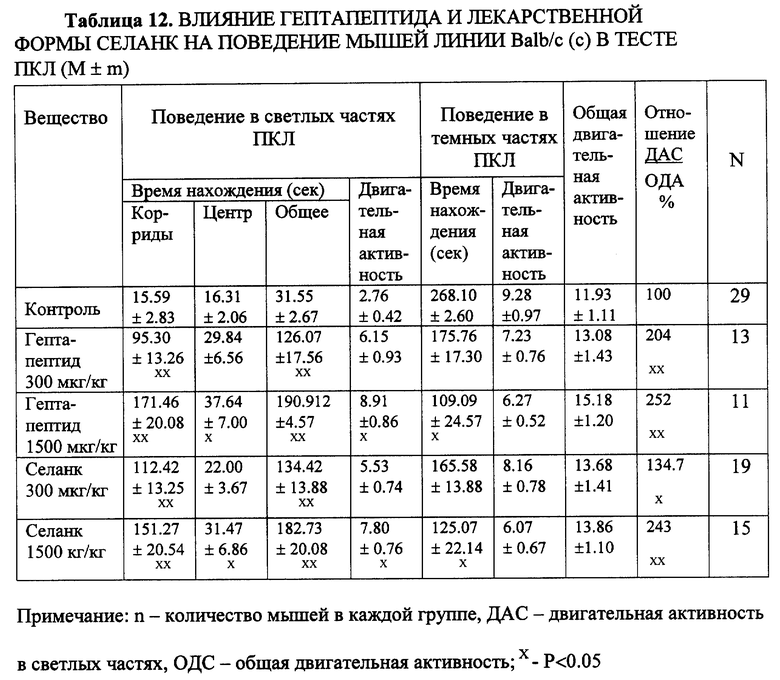
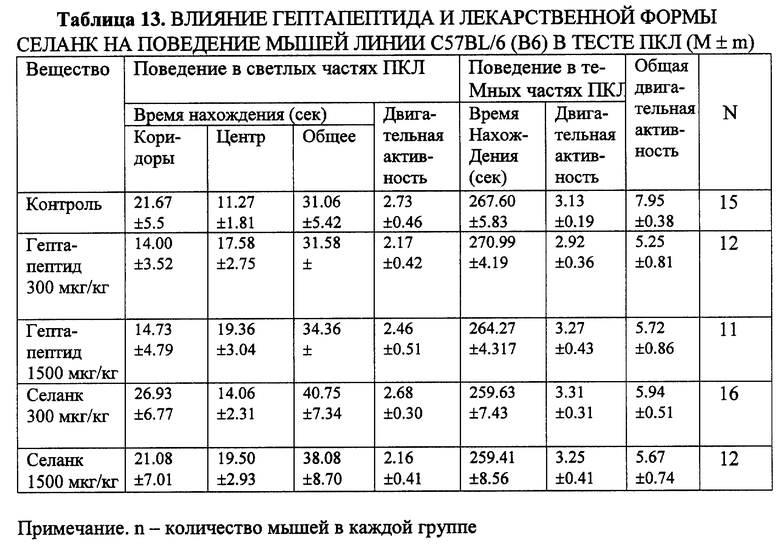
Пример 9. Сопоставление действия гептапептида и Селанка по анксиолитическому действию в тестах ПКЛ на мышах с пассивным и активным фенотипом ЭСР (линия Balb/c и C57В1/6).
Как уже отмечалось выше, селективное анксиолитическое действие достоверно выявляется у мышей линии Balb/c, поведение которых при эмоциональном воздействии в конфликтной стрессогенной ситуации (ОП, ПКЛ) характеризуется freezing-реакцией.
Установлено, что как водный раствор гептапептида, так и лекарственная форма Селанк, обладают выраженным селективным анксиолитическим действием в тесте ПКЛ. У мышей с выраженной freezing-реакцией, высоким уровнем тревоги достоверно увеличивается число забегов в светлые рукава лабиринта, общее время пребывания в светлых частях лабиринта и соотношение числа забегов в светлые рукава к общему числу перемещений по рукавам лабиринта. Последний показатель является наиболее важным и общепринятым критерием, характеризующим наличие анксиолитических свойств соединений (File, 1995). Десятикратное увеличение дозы незначительно повышает время пребывания мышей в центре лабиринта и не вызывает снижения двигательной активности или каких-либо других побочных явлений (седации, миорелаксации, атаксии) (табл. 12). Поведение мышей с "активным" фенотипом ЭСР (линия C57BL/6) существенно не менялось на фоне изучаемых соединений (табл. 13).
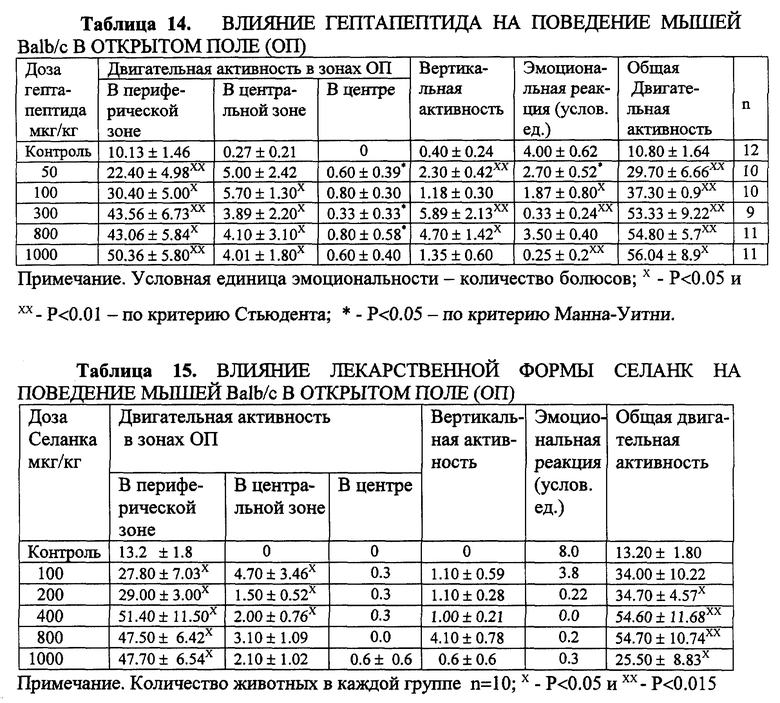
Пример 10. Сопоставление действия гептапептида и Селанка по анксиолитическому действию в тестах ОП на мышах линии Balb/c.
Поведение мышей линии Balb/c (с) в открытом поле исходно характеризовалось развитием freezing-реакции и крайне низким уровнем двигательной активности, реализующейся в виде горизонтальных перемещений по периферии арены открытого поля, без выходов в центральные зоны. В данной серии экспериментов установлено, что как гептапептид (табл. 14), так и Селанк (табл. 15) достоверно повышают общую двигательную активность, появляются выходы в средние зоны и даже пересечение центра арены открытого поля, freezing-реакции не возникает. Высокий уровень активности сохраняется и при увеличении дозы.
Таким образом, в специфических ситуациях ОП и ПКЛ, используемых для выявления анксиолитического действия известных и новых соединений, как гептапептид, так и лекарственная форма Селанк активировали поведение мышей с "пассивным типом" ЭСР. Те же дозы гептапептида и Селанка не оказывали выраженного влияния на поведение мышей с "активным" типом ЭСР в тех же условиях. Фармакологический эффект обеих изучаемых форм (гептапептида и лекарственной формы Селанк) может быть по аналогии с бенздиазепиновыми транквилизаторами представлен как анксиолитический, транквило-активирующий, не переходящий, однако, транквило-седативный.
Пример 11. Сопоставление действия на процессы обучения и памяти гептапептида и Селанка.
Для сравнительной оценки действия гептапептида и лекарственной формы Селанк на процессы обучения и памяти использовалась методика выработки условнорефлекторной пищедобывательной реакции у крыс. Выработку реакции проводили в течение одного сеанса на протяжении 30-ти тренировок в камере размером 150х16х23 см, имеющей стартовый, центральный и целевой отсеки. В целевом отсеке находилось и пищевое подкрепление. Передвижение животных и время выполнения реакции регистрировали с помощью фотоэлементов. Тестирование сохранения навыка проводили через 24 часа, 7 суток и 1 месяц. Вещества вводили однократно в дозе 300 мкг/кг интраназально во время сеанса обучения после первых 10-ти тренировок (табл. 16).
Установлено, что как введение гептапептида, так и Селанка облегчает формирование условнорефлекторной двигательной пищедобывательной реакции и ее воспроизведение даже через 30 дней после обучения. Оба соединения вызывают сокращение времени выполнения реакции, наиболее выраженное на заключительном этапе обучения: 40 секунд - контроль, 25 секунд - на фоне гептапептида, 15 секунд - на фоне лекарственной формы Селанк. Положительный эффект на условнорефлекторную пищедобывательную реакцию сохранялся через 24 часа, 7 суток и месяц после обучения. Животные демонстрируют достоверно более высокий уровень сохранения навыка, что выражается более низкими значениями времени (скорости) выполнения реакции по сравнению с контролем.
Таким образом, как гептапептид, так и лекарственная форма Селанк, оказывали активирующее влияние на обучение и память крыс. При этом не было выявлено каких-либо различий в характере их активности.
Экспериментальные исследования Селанка позволяют предложить его в качестве селективного анксиолитика, активирующего мнестические функции.
Селанк является нетоксичным, удобным для применения в клинической практике (капли в нос), с хорошей переносимостью лекарственным препаратом.
Сравнение Селанка с современными транквилизаторами выявили его существенные преимущества.
Преимущества Селанка заключаются в том, что он состоит из природных, встречающихся в организме аминокислот и быстро метаболизирует. Исходя из метаболизма пептидов и аминокислот в организме не образуются токсические продукты. Для Селанка не удалось обнаружить каких-либо нарушений в состоянии животных при увеличении действующей дозы в 200 и более раз.
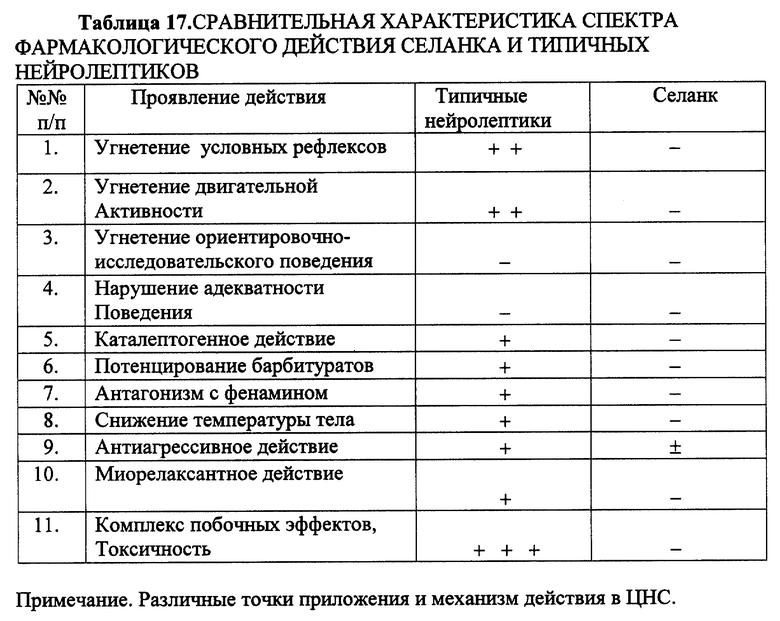
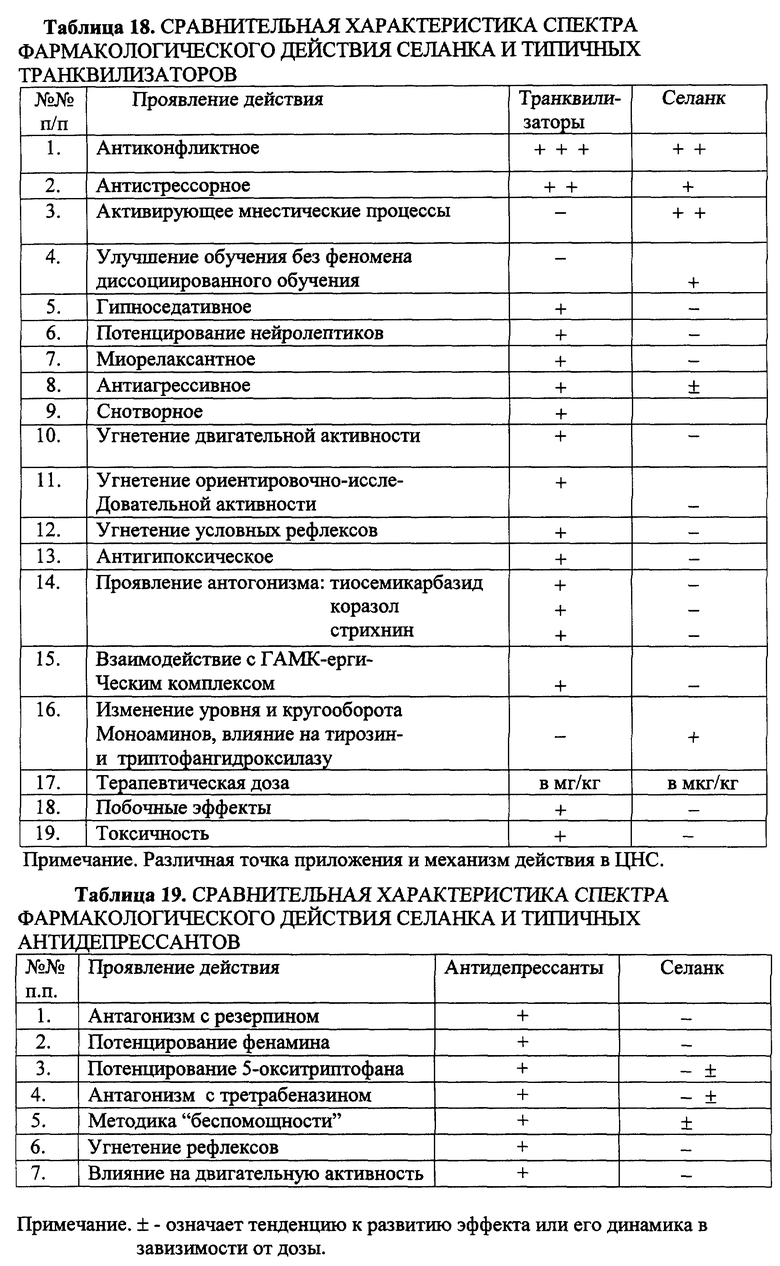
Сравнение Селанка с нейролептиками, транквилизаторами, антидепрессантами показало отсутствие у Селанка побочных эффектов, свойственных последним (табл. 17-19).
| название | год | авторы | номер документа |
|---|---|---|---|
| АНКСИОЛИТИЧЕСКОЕ СРЕДСТВО | 1999 |
|
RU2175229C2 |
| АНКСИОЛИТИЧЕСКОЕ СРЕДСТВО И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ АНКСИОЛИТИЧЕСКОГО ДЕЙСТВИЯ | 2014 |
|
RU2580311C1 |
| ПРОИЗВОДНЫЕ 1,4-БЕНЗОДИАЗЕПИНА, ОБЛАДАЮЩИЕ СЕЛЕКТИВНОЙ АНКСИОЛИТИЧЕСКОЙ АКТИВНОСТЬЮ | 1995 |
|
RU2133248C1 |
| ПРОИЗВОДНЫЕ 2-МЕРКАПТОБЕНЗИМИДАЗОЛА, ОБЛАДАЮЩИЕ СЕЛЕКТИВНОЙ АНКСИОЛИТИЧЕСКОЙ АКТИВНОСТЬЮ | 1994 |
|
RU2061686C1 |
| ГЕПТАПЕПТИД С АДАПТОГЕННОЙ И АНКСИОЛИТИЧЕСКОЙ АКТИВНОСТЬЮ | 1999 |
|
RU2161500C1 |
| ЗАМЕЩЕННЫЕ 2-[2-(3-ОКСОМОРФОЛИН-4-ИЛ)ЭТИЛТИО]БЕНЗИМИДАЗОЛЫ, ОБЛАДАЮЩИЕ АНКСИОЛИТИЧЕСКОЙ АКТИВНОСТЬЮ | 2007 |
|
RU2373202C2 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ АНКСИОЛИТИЧЕСКИМ ДЕЙСТВИЕМ | 2008 |
|
RU2394588C1 |
| СРЕДСТВО ДЛЯ КУПИРОВАНИЯ АЛКОГОЛЬНОГО АБСТИНЕНТНОГО СИНДРОМА | 2012 |
|
RU2521226C2 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ АНКСИОЛИТИЧЕСКОЙ, НООТРОПНОЙ, ПРОТИВОСУДОРОЖНОЙ, АНТИДЕПРЕССИВНОЙ, ЦЕРЕБРОПРОТЕКТОРНОЙ АКТИВНОСТЬЮ И СПОСОБНОСТЬЮ НОРМАЛИЗОВАТЬ ПРОЦЕССЫ ПЕРЕДАЧИ В СИНАПСАХ МОЗГА | 2007 |
|
RU2335293C1 |
| АНКСИОЛИТИЧЕСКОЕ, ПРОТИВОАЛКОГОЛЬНОЕ, ЦЕРЕБРОПРОТЕКТОРНОЕ ЛЕКАРСТВЕННОЕ СРЕДСТВО | 1999 |
|
RU2145855C1 |



Изобретение относится к медицине, в частности к разработке и применению нового анксиолитического средства. Предложено применение гептапептида Тhr-Lys-Рrо-Аrg-Рrо-Gly-Рrо в качестве анксиолитического средства. Предложен новый состав анксиолитического средства, содержащий активное вещество в виде указанного гептапептида (1,45-1,55 г/л), консервант в виде нипагина (0,95-1,05 г/л) и вода дистиллированная - остальное. Состав высокоэффективен в низких дозах и прост в применении: используется в виде капель в нос. 2 с.п. ф-лы, 4 ил., 19 табл.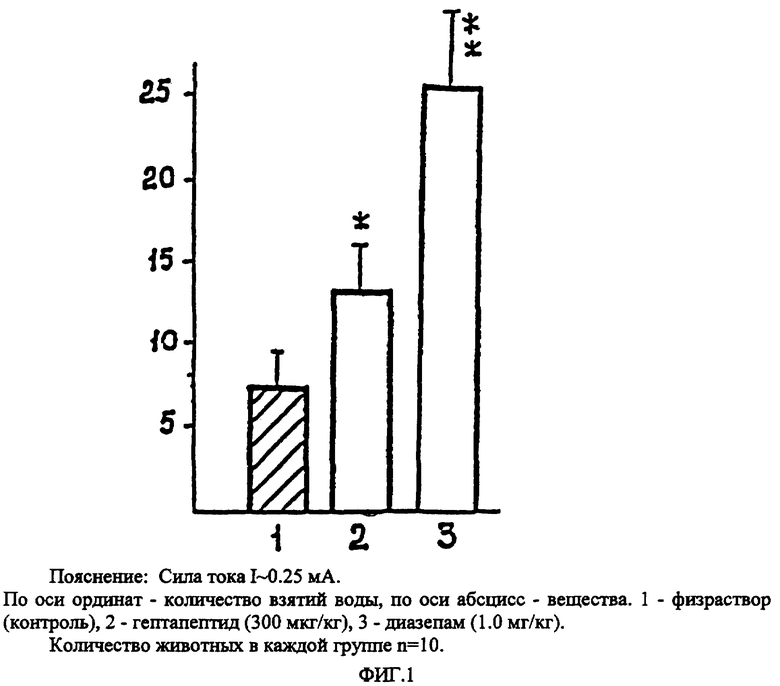
Гептапептид указанной формулы - 1,45 - 1,55
Нипагин - 0,95 - 1,05
Дистиллированная вода - Остальное
| МАШКОВСКИЙ М.Д | |||
| Лекарственные средства | |||
| - Вильнюс, 1994, ч | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Нефтяной конвертер | 1922 |
|
SU64A1 |
| НООТРОПНОЕ СРЕДСТВО И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НООТРОПНОГО ДЕЙСТВИЯ | 1994 |
|
RU2045958C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПЕПТИДОВ, ВОССТАНАВЛИВАЮЩИХ ФУНКЦИЮ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ | 1994 |
|
RU2089204C1 |

