Предшествующий уровень техники
Кукурузу выращивают для применения в пищу и получения энергии. Выращивают также многие другие сельскохозяйственные культуры, включая сою и хлопок. Насекомые поедают и повреждают растения и, таким образом, подрывают эти усилия человека. Миллиарды долларов расходуются каждый год для борьбы с насекомыми-вредителями и еще миллиарды теряются с ущербом, который они наносят. Основными средствами, применяемыми для борьбы с насекомыми-вредителями, являются синтетические органические химические инсектициды, но в некоторых регионах важную роль играют биологические инсектициды, такие как инсектицидные белки, получаемые из Bacillus thuringiensis (Bt). Способность получать устойчивые к насекомым растения путем транформации генами из Bt, экспрессирующими инсектицидные белки, в корне изменила современное сельское хозяйство и повысила важность и значение инсектицидных белков и экспрессирующих их генов.
Несколько белков из Bt применяли для создания устойчивых к насекомым трансгенных растений, которые на настоящий момент успешно зарегистрированы и коммерциализированы. Эти белки включают Cry1Ab, Cry1Ac, Cry1F и Cry3Bb в кукурузе, Cry1Ac и Cry2Ab в хлопке и Cry3A в картофеле.
Коммерческие продукты, вырабатывающие эти белки, вырабатывают единственный белок кроме случаев, когда желателен объединенный инсектицидный спектр из 2 белков (например, Cry1Ab и Cry3Bb объединены в кукурузе для придания устойчивости к чешуекрылым вредителям и повреждающим корни личинкам соответственно) или когда независимое действие белков делает их полезными в качестве средства для задержки развития устойчивости у восприимчивых популяций насекомых (например, Cry1Ac и Cry2Ab в хлопке, объединенные для обеспечения регулирования развития устойчивости у табачного почкоеда). См. также патент США 2009 0313717, который относится к белку Cry2 плюс Vip3Aa, Cry1F или Cry1A для борьбы с Helicoverpa zea или armigera. WO 2009 132850 относится к Cry1F или Cry1A и Vip3Aa для борьбы с Spodoptera frugiperda. Патент США 2008 0311096 относится частично к Cry1Ab для борьбы с Cry1F-устойчивым ECB.
Таким образом, некоторые качества трансгенных растений, обладающих устойчивостью к насекомым, которые привели к быстрому и широко распространенному внедрению этой технологии, также создали проблему того, что популяции вредителей развивают устойчивость к инсектицидным белкам, вырабатываемым этими растениями. Было предложено несколько стратегий для сохранения полезности признаков устойчивости к насекомым, основанных на Bt, которые включают применение белков в большой дозе в комбинации с резерватами и изменение при помощи или совместное применение различных токсинов (McGaughey et al. (1998), «B.t. Resistance Management» Nature Biotechnol. 16:144-146).
Необходимо, чтобы белки, выбираемые для применения в IRM стеке, проявляли свои инсектицидные действия независимо так, чтобы устойчивость, выработанная к одному белку, не придавала устойчивость ко второму белку (т.е. отсутствие перекрестной устойчивости к белкам). Если, например, популяция вредителя, выбранная по устойчивости к «белку А», была бы чувствительной к «белку Б», можно было бы заключить, что перекрестной устойчивости нет и что комбинация белка A и белка Б будет эффективной для задержки развития устойчивости к белку А отдельно.
В отсутствие устойчивых популяций насекомых можно делать оценки на основе других характеристик, которые, как предполагается, связаны с механизмом действия и потенциалом перекрестной устойчивости. Предложена возможность идентификации инсектицидных белков, скорее всего не проявляющих перекрестной устойчивости, путем рецептор-опосредованного связывания (van Mellaert et al., 1999). Ключевым показателем недостатка перекрестной устойчивости, присущим этому подходу, является то, что инсектицидные белки не конкурируют за рецепторы у чувствительных видов насекомых.
В случае, когда два Bt токсина конкурируют за один и тот же рецептор, то если этот рецептор мутирует у этого насекомого так, что один из токсинов больше не связывается с этим рецептором и, таким образом, больше не является инсектицидным в отношении насекомого, может случиться так, что насекомое также будет устойчивым ко второму токсину (который избирательно связывается с тем же самым рецептором). Таким образом, говорят, что насекомое обладает перекрестной устойчивостью к обоим Bt токсинам. Однако, если два токсина связываются с двумя различными рецепторами, это может указывать на то, что насекомое не является устойчивым одновременно к этим двум токсинам.
Cry1Fa полезен в борьбе со многими видами чешуекрылых вредителей, включая мотылька кукурузного (ECB; Ostrinia nubilalis (Hubner)) и совку травяную (FAW; Spodoptera frugiperda), и активен против точильщика стеблей сахарного тростника (SCB; Diatraea saccharalis). Белок Cry1Fa, образующийся в растениях кукурузы, содержащий фрагмент TC1507, отвечает за ведущий для промышленности признак устойчивости к насекомым для борьбы с FAW. Cry1Fa дополнительно применяется в продуктах Herculex®, SmartStax™ и WideStrike™.
Дополнительные Cry токсины перечислены на веб-сайте официального комитета по спецификации B.t. (Crickmore et al.; lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/). В настоящее время существует почти 60 главных групп токсинов «Cry» (Cry1-Cry59) с дополнительными Cyt-токсинами и VIP-токсинами и т.п. Многие из каждой числовой группы содержат подгруппы с заглавной буквой, и подгруппы с заглавной буквой содержат подподгруппы со строчными буквами (Cry1 содержит A-L, и Cry1A содержит a-i, например).
Сущность изобретения
Изобретение частично относится к открытию того, что Vip3Ab и Cry1Ab не конкурируют друг с другом за связывание с рецепторами кишечника у Heticoverpa zea (совка хлопковая; CEW). Изобретение также относится частично к удивительному открытию того, что Vip3Ab является активным против капустных молей (DBM), которые устойчивы к Cry1Ab.
Как будет понятно специалисту в области техники, благоприятный эффект этого раскрытия заключается в том, что растения, экспрессирующие Vip3Ab и Cry1Ab, или их инсектицидные части будут полезными для задержки или предотвращения развития устойчивости к каждому из этих инсектицидных белков в отдельности.
Таким образом, изобретение имеет отношение частично к применению белка Vip3Ab в комбинации с белком Cry1Ab. Растения (и площадь земли в акрах, засаженная такими растениями), которые вырабатывают Vip3Ab плюс Cry1Ab, включены в объем изобретения.
Изобретение также имеет отношение частично к тройным стекам или «пирамидам» из трех (или более) токсинов с Vip3Ab и Cry1Ab в качестве основной пары. Такие тройные стеки могут обеспечивать три белка, оказывая неконкурентное действие против CEW. Это может помочь дополнительно снизить или избавить от необходимости создания площади насаждений резерватных растений.
Краткое описание чертежей
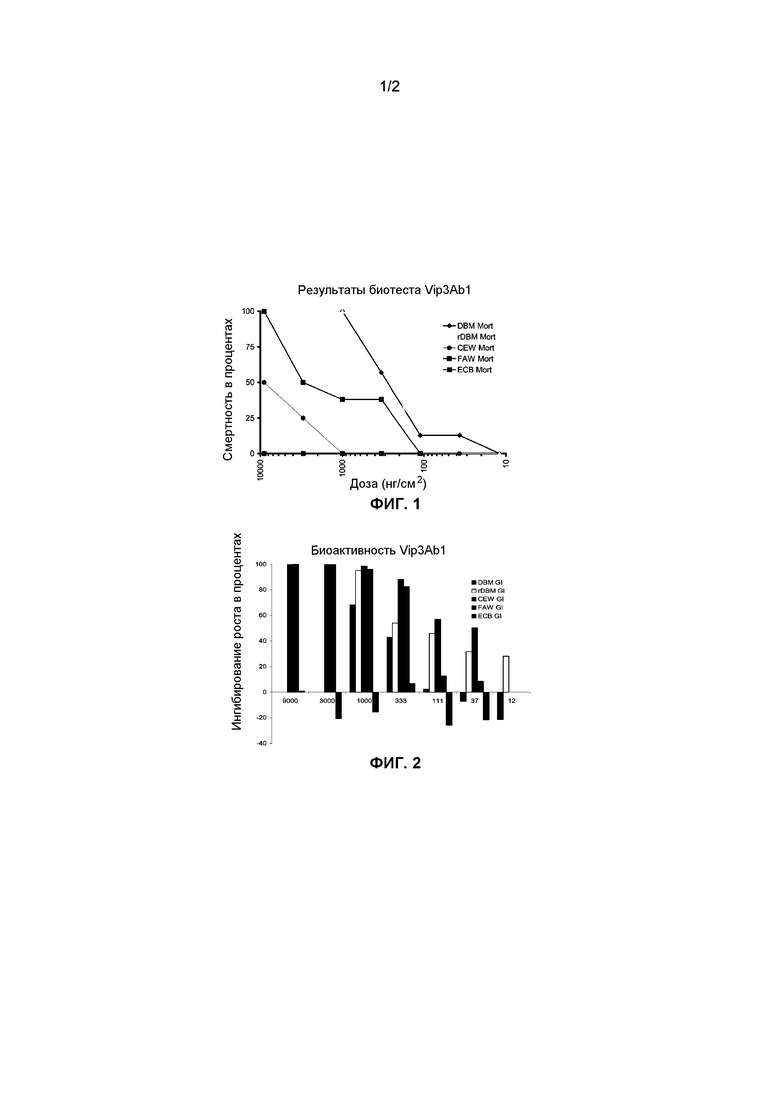
Фиг.1. Реакция на смертельную дозу полноразмерного Vip3Ab1 у личинок Plutella xylostella (Linnaeus) (DBM) и устойчивого к Cry1A Plutella xylostella (rDBM), Heliothis zea (CEW), Spodoptera frugiperda (J.E. Smith), (FAW) и Ostrinia nubilalis (Hubner), (ECB), когда очищенный токсин применяли местно к искусственной пище насекомого. Летальность в процентах основана на наблюдении за 8 насекомыми для каждой дозы через 5 дней после экспонирования токсину.
Фиг.2. Реакция на дозы ингибирования роста полноразмерного Vip3Ab1 у личинок Plutella xylostella (Linnaeus) (DBM) и устойчивого к Cry1A Plutella xylostella (rDBM), Heliothis zea (CEW), Spodoptera frugiperda (J.E. Smith), (FAW) и Ostrinia nubilalis (Hubner), (ECB), когда очищенный токсин применяли местно к искусственной пище насекомого. Ингибирование роста в процентах основано на сравнении среднего веса 8 личинок, обработанных только буфером, к весу личинок, экспонированных к действию токсина в течение 5 дней.
Фиг.3. Кривые конкурентного замещения при связывании 125I Cry1Ab с белком из BBMV, полученным из H. zea. Концентрация белка из BBMV составляла 0,10 мг белка/мл и концентрация 125I Cry1Ab составляла 0,25 нМ. 100% специфическое связывание 125I Cry1Ab измеряли как общее связывание в отсутствие немаркированного Cry1Ab минус неспецифическое связывание, измеренное в присутствии 500 нМ Cry1Ab. Vip3Ab1 тестировали в таких высоких концентрациях как 1000 нМ (в 4000 раз выше, чем меченный радиоактивным изотопом замещающий лиганд), и он не вытеснял связанный 125I Cry1Ab. Немеченый радиоактивным изотопом лиганд замещал 125I Cry1Ab на 50% при концентрации приблизительно 2 нМ.
Краткое описание последовательностей
SEQ ID NO:1 представляет собой N-концевой фрагмент инсектицидного белка Cry1Ab.
SEQ ID NO:2 представляет собой полноразмерный белок Vip3Ab, который, как использовалось в исследованиях связывания, обрабатывали трипсином до корового фрагмента (остатки 200-788).
SEQ ID NO:3 представляет собой полноразмерный белок Cry1Ab, который, как использовалось в исследованиях связывания, обрабатывали трипсином до корового фрагмента (остатки 29-612).
Подробное описание изобретения
Изобретение частично поддерживается открытием того, что Vip3Ab и Cry1Ab не конкурируют друг с другом за связывание с рецепторами в кишечнике у Helicoverpa zea (совка хлопковая; CEW).
Изобретение включает применение Vip3Ab и Cry1Ab для защиты кукурузы и других экономически важных видов растений от повреждения и потери урожая, вызываемых поеданием CEW, и предотвращения развития у популяций CEW устойчивости к любому из этих белков.
Изобретение описывает композиции для борьбы с чешуекрылыми вредителями, включающие клетки, вырабатывающие коровый токсинсодержащий белок Cry1Ab и коровый токсинсодержащий белок Vip3Ab.
Изобретение дополнительно включает хозяина, трансформированного для выработки как инсектицидного белка Cry1Ab, так и инсектицидного белка Vip3Ab, где указанный хозяин является микроорганизмом или растительной клеткой. В некоторых вариантах осуществления изобретения растительные клетки являются неразмножающимися/нетотипотентными клетками. Рассматриваемый полинуклеотид(ы) в генетической конструкции предпочтительно находится под контролем (функционально связанный с/включающий) промотора не из Bacillus thuringiensis. Рассматриваемые полинуклеотиды могут включать кодон растений для усиленной экспрессии в растении.
Дополнительно предполагается, что изобретение обеспечивает способ борьбы с чешуекрылыми вредителями, включающий приведение в контакт указанных вредителей или среды указанных вредителей с эффективным количеством композиции, содержащей коровый токсинсодержащий белок Cry1Ab и дополнительно содержащей коровый токсинсодержащий белок Vip3Ab.
Вариант осуществления изобретения включает растение кукурузы, содержащее экспрессирующий в растении ген, кодирующий коровый токсинсодержащий белок Vip3Ab, и экспрессирующий в растении ген, кодирующий коровый токсинсодержащий белок Cry1Ab, и семя такого растения.
Дополнительный вариант осуществления изобретения включает растение кукурузы, где в указанное растение кукурузы и семя такого растения введен экспрессирующий в растении ген, кодирующий коровый токсинсодержащий белок Vip3Ab, и экспрессирующий в растении ген, кодирующий коровый токсинсодержащий белок Cry1Ab.
Как описано в примерах, исследования конкурентного связывания с рецептором с применением меченного радиоактивным изотопом белка Cry1Ab показывают, что коровый токсинсодержащий белок Cry1Ab не конкурирует за связывание в тканях насекомого CEW с рецептором, с которыми связывается Vip3Ab. Эти результаты также указывают на то, что комбинация белков Cry1Ab и Vip3Ab представляет собой эффективное средство для снижения развития устойчивости к Cry1Ab у популяций CEW (и аналогично, развития устойчивости к Vip3Ab) и, вероятно, увеличивает уровень устойчивости к этому вредителю у растений кукурузы, вырабатывающих оба белка. Таким образом, частично на основании данных, приведенных в настоящем описании, считается, что совместная выработка (стекирование) белков Vip3Ab и Cry1Ab может быть применена для получения большой дозы стека IRM для CEW.
К этой паре могут быть добавлены другие белки для расширения спектра борьбы с насекомыми. Другим вариантом применения может быть применение белков Cry1Ab и Vip3Ab в комбинации с другим, третьим токсином/геном, и применение этого тройного стека для снижения развития устойчивости к любому из этих токсинов у CEW. Таким образом, другой вариант применения изобретения состоит в том, чтобы применять два, три или более белков в регионах, где выращивают сельскохозяйственные культуры, где у CEW могут развиваться устойчивые популяции.
Соответственно изобретение также частично имеет отношение к тройным стекам или «пирамидам» из трех (или более) токсинов с токсинами Cry1Ab и Vip3Ab, представляющими собой основную пару. В некоторых предпочтительных вариантах осуществления пирамид выбранные белки обеспечивают неконкурентное действие против CEW.
Растения (и территория, засаженная такими растениями), которые вырабатывают любую из рассматриваемых комбинаций белков, включены в объем изобретения. Также могут быть добавлены дополнительные токсины/гены, но конкретные стеки, рассмотренные выше с преимуществом и к удивлению обеспечивают много способов действия против CEW. Это может способствовать снижению или отказу от необходимости выделения площади сельскохозяйственной земли под резерватные растения. Поле, засаженное таким образом на площади более 10 акров, таким образом, включено в объем рассматриваемого изобретения.
GENBANK также может быть применен для получения последовательностей для любого из генов и белков, раскрытых или указанных в настоящем описании.
Комбинации белков, описанных в настоящем описании, могут быть применены для борьбы с чешуекрылыми вредителями. Взрослые чешуекрылые, например бабочки и мотыльки, главным образом питаются нектаром цветков и являются важным фактором опыления. Почти все личинки чешуекрылых, т.е. гусеницы, питаются растениями, и многие из них являются серьезными вредителями. Гусеницы питаются на или внутри листвы или на корнях или стебле растения, лишая растение питательных веществ и часто разрушая структуру физической опоры растения. Дополнительно гусеницы питаются плодами, тканями и запасами зерна и мукой, делая невозможным продажу этих продуктов или сильно снижая их стоимость. Как применено в настоящем описании, ссылка на чешуекрылых вредителей относится к различным стадиям жизненного цикла вредителя, включая личиночные стадии.
Некоторые химерные токсины по изобретению включают целиком N-концевую часть корового Bt токсина и, в некоторой точке после конца коровой части токсина белок имеет переход к гетерологической последовательности протоксина. N-концевая, инсектицидно активная токсиновая часть Bt токсина указана в качестве «корового» токсина. Переход от сегмента корового токсина к сегменту гетерологического протоксина может встретиться приблизительно в области соединения токсина/протоксина или, альтернативно, часть нативного протоксина (простирающийся через часть корового токсина) может быть сохранена с переходом к гетерологической части протоксина, имеющим место ниже.
В качестве примера один химерный токсин изобретения представляет собой целую часть корового токсина Cry1Ab (приблизительно первые 600 аминокислот) и гетерологический протоксин (остаток от молекулы до C-окончания). В одном предпочтительном варианте осуществления часть химерного токсина, включающую протоксин, получают из другого Cry1Ab белкового токсина.
Специалисту в области техники будет понятно, что Bt токсины даже в пределах определенного класса, такого как Cry1A, изменяются до некоторой степени в длине и точном расположении перехода от части корового токсина до части протоксина. Как правило, токсины Cry1Ab содержат от приблизительно 1150 до приблизительно 1200 аминокислот в длину. Переход от части корового токсина до части протоксина, как правило, встречается между приблизительно 50% и приблизительно 60% общей длины токсина. Химерный токсин изобретения включает всю протяженность этой N-концевой части корового токсина. Таким образом, химерный токсин включает, по меньшей мере, приблизительно 50% общей длины Cry1Ab Bt белкового токсина. Это, как правило, представляет собой, по меньшей мере, приблизительно 590 аминокислот. В отношении части протоксина общая протяженность части протоксина Cry1Ab составляет от конца части корового токсина до C-конца молекулы.
Гены и токсины. Гены и токсины, полезные согласно изобретению, включают не только полноразмерные раскрытые последовательности, но также и фрагменты этих последовательностей, варианты, мутантные и сшитые белки, сохраняющие характерное пестицидное действие токсинов, конкретно проиллюстрированных в настоящем описании. Как применено в настоящем описании, термины «варианты» или «вариации» генов относятся к нуклеотидным последовательностям, кодирующим те же самые токсины или кодирующим эквивалентные токсины, обладающие пестицидным действием. Как применено в настоящем описании, термин «эквивалентные токсины» относится к токсинам, обладающим той же самой или по существу той же самой биологической активностью против целевых вредителей как патентуемые токсины.
Как применено в настоящем описании, граничные значения составляют приблизительно 95% (Cry1Ab и Vip3Ab’s), 78% (Cry1F и Vip3A’s) и 45% (Cry1’s и Vip3’s) идентичности последовательности по «Revision of the Nomenclature for the Bacillus thuringiensis Pesticidal Crystal Proteins» N. Crickmore, D.R. Zeigler, J. Feitelson, E. Schnepf, J. Van Rie, D. Lereclus, J. Baum и D.H. Dean. Microbiology and Molecular Biology Reviews (1998), том 62: 807-813. Эти фрагменты также могут быть применены только к коровым токсинам (для Cry1Ab, например). Белки для применения согласно изобретению могут быть, например, по меньшей мере, на 75%, 85%, 90%, 95% или 99% (и любое целочисленное приращение в пределах этого диапазона) идентичны (идентичность по аминокислотам) белку, приведенному в качестве примера или специфически предложенному в настоящем описании. Они включают белки, кодируемые полинуклеотидами/ДНК, применяемыми согласно изобретению.
Специалисту в области техники будет понятно, что гены, кодирующие активные токсины, могут быть идентифицированы и получены несколькими путями. Конкретные гены или части генов, приведенные в настоящем описании в качестве примера, могут быть получены из изолятов, депонированных в депозитарии культур. Эти гены или их части или варианты также могут быть получены синтетическим путем, например, при помощи генного синтезатора. Вариации генов могут быть легко получены с применением стандартных способов точковой мутации. Кроме того, фрагменты этих генов могут быть получены с применением коммерчески доступных экзонуклеаз или эндонуклеаз стандартными способами. Например, для систематического отрезания нуклеотидов на концах этих генов могут быть применены ферменты, такие как Bal31, или сайт-направленный мутагенез. Гены, кодирующие активные фрагменты, также могут быть получены с применением множества рестрикционных ферментов. Для прямого получения активных фрагментов этих белковых токсинов могут быть применены протеазы.
Фрагменты и эквиваленты, которые сохраняют пестицидное действие токсинов, приведенных в качестве примера, входят в объем изобретения. Кроме того, благодаря избыточности генетического кода многие различных последовательностей ДНК могут кодировать последовательности аминокислот, раскрытых в настоящем описании. Специалист в области техники с легкостью может получить эти альтернативные последовательности ДНК, кодирующие такие же или по существу такие же токсины. Эти различные последовательности ДНК входят в объем изобретения. Как применено в настоящем описании, ссылка на «по существу такую же» последовательность относится к последовательностям, содержащим замены, удаления, добавления или вставки аминокислот, которые фактически не влияют на пестицидное действие. Фрагменты генов, кодирующие белки, которые сохраняют пестицидное действие, также включены в это определение.
Дополнительным способом идентификации генов, кодирующих токсины, и части генов, полезные согласно изобретению, является применение олигонуклеотидных зондов. Эти зонды представляют собой обнаруживаемые нуклеотидные последовательности. Эти последовательности могут быть обнаружены по соответствующей метке или могут быть созданы флуоресцентными по своей природе, как описано в международной заявке № WO93/16094. Как известно в области техники, если тест-молекула и контрольная нуклеиновая кислота гибридизируют путем образования прочной связи между этими двумя молекулами, можно обоснованно предполагать, что тест-молекула и контрольная нуклеиновая кислота существенно гомологичны. Предпочтительно гибридизация проводится в жестких условиях способами, известными в области техники, как описано, например, в Keller, G.H., М.М. Manak (1987) LNA Probes, Stockton Press, Нью Йорк, N.Y., с. 169-170. Некоторые примеры комбинаций концентрации соли и температуры приведены далее (в порядке увеличения жесткости условий): 2X SSPE или SSC при комнатной температуре; 1X SSPE или SSC при 42°С; 0,1Х SSPE или SSC при 42°C; 0,1X SSPE или SSC при 65°С. Обнаружение тест-молекулы обеспечивает средство для определения того, произошла ли гибридизация. Такой зондовый анализ обеспечивает быстрый способ идентификации генов, кодирующих токсины по изобретению. Нуклеотидные сегменты, применяемые в качестве тест-молекул согласно изобретению, могут быть синтезированы при помощи ДНК синтезатора и стандартных способов. Эти нуклеотидные последовательности также могут быть применены в качестве праймеров ПЦР для амплификации генов по изобретению.
Варианты токсинов. В частности, для примера в настоящем описании приведены определенные токсины по изобретению. Поскольку эти токсины являются типовыми для токсинов по изобретению, сразу должно быть очевидно, что изобретение включает варианты токсинов или эквивалентные токсины (и нуклеотидные последовательности, кодирующие эквивалентные токсины), обладающие таким же или схожим пестицидным действием с действием токсина, приведенного для примера. Эквивалентные токсины будут гомологичны по аминокислотам токсину, приведенному для примера. Эта аминокислотная гомологичность, как правило, составляет боле 75%, предпочтительно, более 90% и наиболее предпочтительно более 95%. Аминокислотная гомологичность является наиболее высокой в критических частях токсина, которые отвечают за биологическую активность или вовлечены в создание трехмерной конфигурации, которая, в конечном счете, отвечает за биологическую активность. В этом отношении определенные замены аминокислот приемлемы и могут быть ожидаемы, если эти замены находятся в частях, которые не являются критически важными в отношении активности или являются консервативными заменами аминокислот, не влияющими на трехмерную конфигурацию молекулы. Например, аминокислоты могут быть классифицированы следующим образом: неполярные, незаряженные полярные, основные и кислотные. Консервативные замены, при которых аминокислота одного класса заменяется другой аминокислотой того же типа, входят в объем изобретения, если при этом замена фактически не изменяет биологическую активность соединения. Ниже приведен список примеров аминокислот, принадлежащих каждому классу.
В некоторых случаях также могут быть сделаны неконсервативные замены. Критическим фактором является то, что эти замены не должны значительно нарушать биологическую активность токсина.
Рекомбинантные хозяева. Гены, кодирующие токсины по изобретению, могут быть введены большому разнообразию растений-хозяев или микроорганизмов-хозяев. Экспрессия токсинового гена приводит напрямую или косвенно к внутриклеточному образованию и сохранению пестицида. Конъюгационный перенос и рекомбинантный перенос могут быть применены для получения штамма Bt, который вырабатывает оба токсина по изобретению. Другие организмы-хозяева также могут быть модифицированы одним или обоими токсиновыми генами и затем применены для осуществления синергического действия. С подходящими микроорганизмами-хозяевами, например, Pseudomonas, микроорганизмы могут быть применены к месту нахождения вредителя, где они будут распространяться и попадать внутрь вредителя. Результатом является борьба с вредителем. Альтернативно микроорганизм-хозяин токсинового гена можно обрабатывать в условиях, продлевающих действие токсина и стабилизирующих клетку. Затем обработанная клетка, сохраняющая токсическое действие, может быть применена к среде целевого вредителя.
В случаях, когда в микроорганизм-хозяин вводят Bt токсиновый ген при помощи подходящего вектора, и указанный микроорганизм-хозяин применяют к среде в живом состоянии, необходимо применение определенных микроорганизмов-хозяинов. Выбирают такие микроорганизмы-хозяева, о которых известно, что они занимают «фитосферу» (филлоплану, филлосферу, прикорневую зону и/или ризоплану) одной или более представляющих интерес сельскохозяйственных культур. Эти микроорганизмы выбирают так, чтобы они были способны успешно конкурировать в конкретной окружающей среде (сельскохозяйственная культура и другие среды обитания насекомого) с микроорганизмами дикого типа, обеспечивать устойчивое сохранение и экспрессию гена, экспрессирующего полипептидный пестицид, и, по желанию, обеспечивать улучшенную защиту пестицида от разложения и инактивации в окружающей среде.
Известно, что большое количество микроорганизмов населяет филлоплану (поверхность листьев растения) и/или прикорневую зону (почву, окружающую корни растения) большого разнообразия важных сельскохозяйственных культур. Эти микроорганизмы включают бактерии, водоросли и грибы. Особенный интерес представляют микроорганизмы, такие как бактерии, например, рода Pseudomonas, Erwinia, Serratia, Klebsiella, Xanthomonas, Streptomyces, Rhizobium, Rhodopseudomonas, Methylophilius, Agrobactenum, Acetobacter, Lactobacillus, Arthrobacter, Azotobacter, Leuconostoc и Alcaligenes; грибы, в частности, дрожжи, например, рода Saccharomyces, Cryptococcus, Kluyveromyces, Sporobolomyces, Rhodotorula и Aureobasidium. Особенный интерес представляют такие виды фитосферных бактерий, как Pseudomonas syringae, Pseudomonas fluorescens, Serratia marcescens, Acetobacter xylinum, Agrobactenium tumefaciens, Rhodopseudomonas spheroides, Xanthomonas campestris, Rhizobium melioti, Alcaligenes entrophus и Azotobacter vinlandii; и виды фитосферных дрожжей, такие как Rhodotorula rubra, R. glutinis, R. marina, R. aurantiaca, Cryptococcus albidus, C. diffluens, C. laurentii, Saccharomyces rosei, S. pretoriensis, S. cerevisiae, Sporobolomyces roseus, S. odorus, Kluyveromyces veronae и Aureobasidium pollulans. В частности, интересны пигментированные микроорганизмы.
Существует большое разнообразие способов для введения Bt гена, кодирующего токсин в микроорганизме-хозяине при условиях, позволяющих устойчивое сохранение и экспрессию гена. Эти способы известны специалистам в области техники и описаны, например, в патенте США № 5135867, включенном в настоящее описание посредством ссылки.
Обработка клеток. Bacillus thuringiensis или рекомбинантные клетки, образующие Bt токсины, можно обрабатывать так, чтобы продлить действие токсина и стабилизировать клетку. Образуемая пестицидная микрокапсула включает Bt токсин или токсины внутри стабилизированной клеточной структуры, которая защищает токсин, когда микрокапсулу применяют к среде обитания целевого вредителя. Подходящие клетки-хозяева могут включать как прокариотов, так и эукариотов, обычно ограничиваясь теми клетками, которые не образуют токсичные вещества для высших организмов, таких как млекопитающие. Однако организмы, образующие токсичные вещества для высших организмов, могут быть применены в случае, когда токсичные вещества неустойчивы или норма внесения является достаточно низкой, такой, чтобы исключить любую возможность токсичности для млекопитающего-хозяина. В качестве хозяев особенно интересны прокариоты и низшие эукариоты, такие как грибы.
Клетка, как правило, является интактной и во время обработки находится по существу в пролиферативной фазе, а не в фазе споры, хотя в некоторых случаях споры могут быть применены.
Обработка микробной клетки, например, микроорганизма, содержащего ген или гены B.t. токсина, может быть осуществлена химическими или физическими средствами или комбинацией химических и/или физических средств в случае, если этот способ не имеет вредного воздействия на свойства токсина и не снижает способность клетки защищать токсин. Примерами химических реагентов являются галоидирующие агенты, в частности галогены с атомным номером 17-80. Более конкретно, для достижения желаемых результатов может быть применен иод в мягких условиях и в течение достаточного количества времени. Другие подходящие способы включают обработку альдегидами, такими как глютаральдегид; противоинфекционными средствами, такими как хлорид зефирана и хлорид цетилпиридиния; спиртами, такими как изопропиловый и этиловый спирты; различными гистологическими фиксаторами, такими как иодный раствор Люголя, фиксатор Буэна, различные кислоты и фиксатор Хелли (см. Humason, Gretchen L., Animal tissue techniques, W.H. Freeman and Company, 1967); или комбинацией физических (высокая температура) и химических агентов, сохраняющих и продлевающих действие токсина, образующегося в клетке, когда клетку вносят в среду обитания хозяина. Примерами физических агентов являются коротковолновая радиация, такая как гамма-излучение и рентгеновское излучение, замораживание, УФ излучение, лиофилизация и т.п. Способы обработки микробных клеток раскрыты в патентах США №№ 4695455 и 4695462, включенных в настоящее описание посредством ссылки.
В целом, клетки будут обладать повышенной структурной устойчивостью, которая увеличит устойчивость к факторам окружающей среды. Когда пестицид находится в виде проформы, способ обработки клетки необходимо выбирать так, чтобы не ингибировать перевод проформы в итоговую форму пестицида целевым патогенным организмом вредителя. Например, формальдегид сшивает белки и может ингибировать трансформацию проформы полипептидного пестицида. Способ обработки должен сохранять, по меньшей мере, существенную долю биодоступности или биоактивность токсина.
Характеристики, представляющие особенный интерес при отборе клетки-хозяина с целью получения включают легкость введения B.t. гена или генов хозяину, пригодность систем экспрессии, эффективность экспрессии, устойчивость пестицида в хозяине и наличие вспомогательных наследственных характеристик. Представляющие интерес характеристики при применении в качестве пестицидной микрокапсулы включают защитные свойства для пестицида, такие как толстостенность клеточных оболочек, пигментация и внутриклеточная упаковка или образование телец-включений; выживаемость в водных средах; низкая токсичность для млекопитающих; привлекательность для заглатывания вредителями; легкость уничтожения и фиксации без повреждения токсина; и т.п. Другие принимаемые в рассмотрение факторы включают легкость при составлении в композицию и обращении, экономичность, устойчивость при хранении и т.п.
Рост клеток. Хозяин клетки, содержащий B.t. инсектицидный ген или гены, может быть выращен в любой удобной питательной среде, в которой конструкция ДНК обеспечивает селективное преимущество, обеспечивая селективную питательную среду таким образом, чтобы существенно все или все клетки сохраняли B.t. ген. Затем эти клетки можно собирать в соответствии с традиционными способами. Альтернативно клетки можно обрабатывать до их сбора.
B.t. клетки, вырабатывающие токсины по изобретению, могут быть культивированы с применением стандартных в области техники среды и способов ферментации. По завершению цикла ферментации бактерии могут быть собраны путем первоначального отделения B.t. спор и кристаллов от ферментативного бульона средствами, известными в области техники. Выделенные B.t. споры и кристаллы могут быть составлены в композицию в виде смачиваемого порошка, жидкого концентрата, гранул или в виде других композиций путем добавления поверхностно-активных веществ, диспергирующих агентов, инертных носителей и других компонент для облегчения обращения с ними и применения в отношении конкретных целевых вредителей. Все эти композиции и способы применения известны в области техники.
Композиции. Составленные в композицию приманочные гранулы, содержащие аттрактант и споры, кристаллы и токсины изолятов B.t., или рекомбинантные микроорганизмы, включающие гены, получаемые из изолятов B.t., раскрытых в настоящем описании, могут быть применены к почве. Составленный в композицию продукт также может быть применен в качестве покрытия для семян или обработки корня или обработки всего растения на более поздних стадиях цикла развития сельскохозяйственной культуры. Обработка растения и почвы клетками B.t. может быть осуществлена в виде смачиваемых порошков, гранул или пудр путем смешивания с различными инертными материалами, такими как неорганические минералы (филлосиликаты, карбонаты, сульфаты, фосфаты и т.п.) или растительные материалы (истертые кукурузные початки, рисовая шелуха, скорлупа грецкого ореха и т.п.). Композиции могут включать вспомогательные агенты, улучшающие растекание-связывание, стабилизирующие агенты, другие пестицидные добавки или поверхностно-активные вещества. Жидкие композиции могут быть водными или неводными и могут быть применены в виде пен, гелей, суспензий, эмульгируемых концентратов и т.п. Ингредиенты могут включать реологические агенты, поверхностно-активные вещества, эмульгаторы, диспергаторы или полимеры.
Специалисту в области техники понятно, что концентрация пестицида будет значительно различаться в зависимости от природы конкретной композиции, в частности, того, является ли она концентратом или будет применена непосредственно. Пестицид может составлять, по меньшей мере, 1% по массе и 100% по массе. В сухих композициях количество пестицида составляет приблизительно 1-95% по массе, в то время как в жидких композициях твердая фаза в жидкой фазе составляет приблизительно 1-60% по массе. Композиции, в целом, содержат приблизительно от 102 до приблизительно 104 клеток/мг. Эти композиции применяют в количестве от приблизительно 50 мг (жидкой или сухой) до 1 кг или более на гектар.
Композиции могут быть применены к среде обитания чешуекрылого вредителя, например листве или почве, путем разбрызгивания, распыливания, опрыскивания и т.п.
Модифицирование растений. Предпочтительным рекомбинантным хозяином для получения инсектицидных белков по изобретению является модифицированное растение. Гены, кодирующие токсиновые B.t. белки, как раскрыто в настоящем описании, могут быть введены в растительные клетки с применением множества способов, известных в области техники. Например, для подготовки к введению чужеродных генов в высшие растения доступно большое количество клонирующих векторов, включающих систему репликации в Escherichia Coli и маркерный ген, который позволяет выбирать модифицированные клетки. Векторы включают среди прочего, например, pBR322, ряд pUC, ряд M13mp, pACYC184. Соответственно фрагмент ДНК, содержащий последовательность, кодирующую Bt токсиновый белок, может быть введен в вектор на подходящем сайте рестрикции. Получаемую плазмиду применяют для введения в E. coli. Клетки E. coli клетки культивируют в подходящей питательной среде, затем собирают и подвергают лизису. Плазмиду регенерируют. В качестве методов анализа, как правило, применяют секвенирование, рестрикционный анализ, электрофорез и другие биохимические методы молекулярной биологии. После каждого этапа примененная последовательность ДНК может быть расщеплена и присоединена к следующей последовательности ДНК. Каждая последовательность плазмиды может быть клонирована в той же самой или других плазмидах. В зависимости от способа введения желаемых генов в растение могут потребоваться другие последовательности ДНК. Если, например, Ti- или Ri-плазмиду применяют для модифицирования растительной клетки, то, по меньшей мере, правый конец, но часто правый и левый концы T-ДНК Ti- или Ri-плазмиды, должны быть соединены в качестве фланкирующей области вставляемых генов. Применение T-ДНК для модифицирования растительных клеток интенсивно исследовано и достаточно описано в EP 120516, Lee и Gelvin (2008), Hoekema (1985), Fraley et al., (1986) и An et al., (1985), и широко известно в области техники.
Как только введенная ДНК интегрируется в геном растения, она становится относительно устойчивой. Модифицирующий вектор, как правило, содержит селектируемый маркер, который придает модифицированным растительным клеткам устойчивость к биоциду или антибиотику, такому как среди прочих Биафос, Канамицин, G418, Блеомицин или Гигромицин. Индивидуально применяемый маркерный ген должен соответственно позволять выбор модифицированных клеток, а не клеток, которые не содержат введенную ДНК.
Существует большое количество способов для введения ДНК в клетку растения-хозяина. Эти способы включают модифицирование посредством T-ДНК с применением Agrobacterium tumefaciens или Agrobacterium rhizogenes в качестве модифицирующего агента, слияние, инъекция, баллистическая трансфекция (бомбардировка микрочастицами) или электропорация, так же, как и другие возможные способы. Если для модифицирования применяют Agrobacteria, то вводимую ДНК необходимо клонировать в специальных плазмидах, а именно и во вспомогательном векторе или в бинарном векторе. Вспомогательные векторы могут быть интегрированы в Ti- или Ri-плазмиду путем гомологической рекомбинации в силу последовательностей, которые являются гомологическими последовательностям в T-ДНК. Ti- или Ri-плазмида также включает vir-область, необходимую для переноса T-ДНК. Вспомогательные векторы в Agrobacteria не могут реплицироваться. Вспомогательный вектор может быть перенесен в Agrobacterium tumefaciens посредством плазмиды-помощника (конъюгация). Бинарные векторы могут реплицироваться как в E. coli, так и в Agrobacteria. Они включают маркерный ген отбора и линкер или полилинкер, которые ограничены областями правой и левой границы T-ДНК. Они могут быть перенесены непосредственно в Agrobacteria (Holsters et al., 1978). Agrobacterium, применяемая в качестве клетки-хозяина, должна включать плазмиду, несущую vir-область. Vir-область необходима для переноса T-ДНК в растительную клетку. В состав может входить дополнительная T-ДНК. Модифицированную таким образом бактерию применяют для модифицирования растительных клеток. Эксплантаты растения с преимуществом могут быть культивированы с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в растительную клетку. Затем могут быть воссозданы целые растения из инфицированной растительной ткани (например, части листа, сегментов стебля, корней, а также протопластов или культивированных в суспензии клеток) в подходящей питательной среде, которая может содержать антибиотики или биоциды для отбора. Затем растения, получаемые таким образом, могут быть протестированы на наличие введенной ДНК. Не предъявляется никаких специальных требований к плазмидам в случае инъекции и электропорации. Допускается применение обычных плазмид, таких как, например, производные pUC.
Модифицированные клетки растут внутри растений обычным образом. Они могут образовывать зародышевые клетки и передавать модифицированный признак(и) растениям потомства. Такие растения могут быть выращены обычным образом и скрещены с растениями, содержащими такие же модифицированные наследственные признаки или другие наследственные признаки. Получаемые в результате гибридные растения обладают соответствующими фенотипическими признаками.
В предпочтительном варианте осуществления изобретения растения модифицируют генами, в которых частота использования кодона оптимизирована для растений (см., например, патент США № 5380831, который таким образом включен посредством ссылки). Одновременно с тем, что в настоящем описании для примера приведены некоторые укороченные токсины, в области исследования B.t. известно, что токсины с молекулярной массой 130 кДа (общая длина) содержат половину с N-концом, которая является коровым токсином и половину с C-концом, которая является протоксином «хвостом». Таким образом, подходящие «хвосты» могут быть применены с укороченными/коровыми токсинами по изобретению (см., например, патент США № 6218188 и патент США № 6673990). Кроме того, в области техники известны способы создания синтетических Bt генов для применения в растениях (Stewart and Burgin, 2007). Одним неограничивающим примером предпочтительного модифицированного растения является фертильное растение кукурузы, включающее экспрессируемый растением ген, кодирующий белок Cry1Ab и дополнительно включающее второй экспрессируемый растением ген, кодирующий белок Vip3Ab.
Перенос (или интрогрессия) Cry1Ab- и Vip3Ab-определяемого признака(ов) в инбредную линию кукурузы может быть осуществлено размножением с рекуррентной селекцией, например обратным скрещиванием. В этом случае желаемый рекуррентный родитель сначала скрещивают с инбредным растением-донором (нерекуррентный родитель), которое несет подходящий ген(ы) для Cry1F- и Vip3Ab-определяемых признаков. Потомство от этого скрещивания затем снова скрещивают с рекуррентным родителем с последующей селекцией в получаемом потомстве в отношении желаемого признака(ов), чтобы передать его от нерекуррентного родителя. После трех, предпочтительно четырех, более предпочтительно пяти или более поколений обратного скрещивания с рекуррентным родителем и с селекцией в отношении желаемого признака(ов), потомство будет гетерозиготным по локусам, отвечающим за передаваемый признак(и), но будет похожим на рекуррентного родителя по большинству или почти всем остальным генам (см., например, Poehlman & Sleper (1995) Breeding Field Crops, 4-е издание, 172-175; Fehr (1987) Principles of Cultivar Development, Том 1: Theory and Technique, 360-376).
Стратегии борьбы с устойчивостью у насекомых (IRM). Roush et al., например, выделяет стратегии с двумя токсинами, также называемые «пирамидирование» или «стекирование», для выращивания инсектицидных трансгенных сельскохозяйственных культур (The Royal Society. Phil. Транс. R. Soc. Lond. B. (1998) 353, 1777-1786).
На своем веб-сайте Управление по охране окружающей среды Соединенных Штатов Америки (epa.gov/oppbppdl/biopesticides/pips/bt_corn_refuge_2006.htm) публикует следующие требования для создания резерватов с резерватными растениями (т.е. не B.t.) (секция не Bt сельскохозяйственных культур/кукурузы) для применения вместе с трансгенными сельскохозяйственными культурами, вырабатывающими один Bt белок, активный против целевых вредителей.
«Конкретные структурные требования для продуктов Bt кукурузы (Cry1Ab или CryIF), защищенной от кукурузного мотылька, являются следующими:
Структурированные резерваты:
20% резерват Bt кукурузы для нечешуекрылых в кукурузном поясе;
50% резерват Bt кукурузы для нечешуекрылых в хлопковом поясе.
Блоки
Внутренний (т.е., в пределах B.t. поля)
Внешний (т.е., отдельные поля в пределах Ѕ мили (по возможности ј мили) от Bt поля для максимизирования случайного скрещивания)
Полоски внутри поля
Полоски должны иметь, по меньшей мере, 4 ряда в ширину (предпочтительно, 6 рядов) для уменьшения эффектов от передвижения личинок»
Кроме того, Национальная ассоциация производителей кукурузы на своем веб-сайте: (ncga.com/insect-resistance-management-fact-sheet-bt-corn) также приводит схожее руководство относительно требований к резерватам. Например:
«Требования для IRM кукурузного мотылька:
- по меньшей мере, 20% вашей территории посадок кукурузы должны быть заняты резерватными гибридами
- в областях выращивания хлопка резерваты должны составлять 50%
- необходимо делать посадки в пределах 1/2 мили от резерватных гибридов
- резерват может быть засажен полосами в пределах B.t. поля; резерватные полосы должны иметь, по меньшей мере, 4 ряда в ширину
- резерват можно обрабатывать одними только обычными пестицидами, если при этом достигается экономически выгодный эффект в отношении целевого насекомого
- распыляемые инсектициды на основе B.t. нельзя применять к резерватной кукурузе
- подходящий резерват должен быть посажен на каждой ферме с Bt кукурузой»
Как заявлено авторами Roush et al. (в правой колонке на страницах 1780 и 1784, например), стекирование или пирамидирование двух различных белков, каждый из которых эффективен против целевых вредителей и с небольшой или отсутствием перекрестной устойчивости, может позволить применение меньшего резервата. Roush предполагает, что в случае образования успешного стека, менее чем 10% резервата может обеспечить борьбу с устойчивостью, сопоставимую с таковой приблизительно 50% резервата в случае отдельного (непирамидированного) признака. Для доступных в настоящее время пирамидированных Bt продуктов кукурузы, Управление по охране окружающей среды США требует создания значительно менее (в целом, 5%) структурированного резервата не Bt кукурузы, чем для продуктов с одним признаком (в целом, 20%).
Существуют различные способы получения IRM эффектов от резервата, включая различные геометрические структуры посадки на полях (как указано выше) и пакетированные смеси семян, как рассмотрено дополнительно авторами Roush et al. (выше) и в патенте США № 6551962.
Вышеуказанные величины в процентах или подобные соотношения в резерватах могут быть применены с целью создания двойных или тройных стеков или пирамид. Для тройных стеков с тремя режимами действия против единственного целевого вредителя целью будет нулевой резерват (или, например, менее 5% под резерват). Это, в частности, верно для коммерческих площадей, например, размером более 10 акров.
Все патенты, патентные заявки, предварительные заявки и публикации, на которые сделаны ссылки или процитированные в настоящем описании, включены полностью посредством ссылки в той степени, в которой они не являются несогласующимися с явными указаниями в настоящем описании.
Если в частности не указано или подразумевается другое, неопределенная и определенная формы единственного числа, как применено в настоящем описании, обозначают «по меньшей мере» один.
Следующие примеры иллюстрируют способы осуществления изобретения. Эти примеры не должны быть рассмотрены как ограничивающие. Все проценты приведены по массе, и все соотношения растворимые компоненты в смеси приведены по объему, если не указано иное. Все температуры приведены в градусах Цельсия.
ПРИМЕРЫ
Пример 1 - Краткое описание примеров
Следующие примеры демонстрируют, что Vip3Ab1 вместе с Cry1Ab придает неперекрестную устойчивость к гусеницам хлопковой американской совки (CEW), таким образом, показывая, что эти два белка могут противодействовать развитию устойчивости у CEW к каждому из этих белков по отдельности.
Дополнительно было показано, что Vip3Ab1 действует против личинок Plutella xylostella (Linnaeus) (капустная моль) и против Cry1Ab-устойчивых личинок Plutella xylostella (Linnaeus). В биотесте включения в питание Vip3Ab1 оказался токсичным для обоих штаммов этого насекомого.
Дополнительное подтверждение этому борьбе с устойчивостью дают исследования конкурентного связывания с радиоактивной меткой с применением 125I Cry1Ab. Представленные данные показывают, что Cry1Ab меченый радиоактивным изотопом 125I прочно и специфично связывается со специфическим набором рецепторных белков, расположенных в препаратах мембранных везикулах щеточной каймы (BBMV) из средней кишки личинок Heliothis zea. Связывание радиоактивного Cry1Ab с его рецепторами может быть конкурентным образом заменено путем применения нерадиоактивного Cry1Ab в BBMV из этого насекомого. Однако Vip3Ab1 как в форме своей полной длины в 85 кДа, так и в ферментативно преобразованной трипсином форме до более низкомолекулярных преобразованных белков не заменяет связывание 125I Cry1Ab с его рецепторами у этого насекомого. Эти результаты показывают, что Vip3Ab проявляет свое биологическое действие на участке, отличном от того участка, на котором связывается Cry1Ab.
Пример 2 - Очистка и обработка трипсином белков Cry1Ab и Vip3Ab1
Гены, кодирующие протоксины Cry1Ab и Vip3Ab1 экспрессировались в экспрессирующих штаммах Pseudomonas fluorescens, и полноразмерные белки отделялись в виде нерастворимых внутриклеточных телец. Промытые внутриклеточные тельца растворяли путем перемешивания при 37°C в буфере, содержащем 20 мМ буфер CAPS с pH 11, + 10 мМ DDT, + 0,1% 2-меркаптоэтанол, в течение 2 ч. Раствор центрифугировался при 27000×g в течение 10 мин при 37°C, и надосадочный раствор обрабатывали 0,5% (мас./об.) трипсином, обработанным TCPK (Sigma). Этот раствор инкубировали при перемешивании в течение еще 1 ч при комнатной температуре, фильтровали, затем загружали в колонку Pharmacia Mono Q 1010, уравновешенную 20 мМ CAPS с М 10,5. После промывания заправленной колонки 2 колоночными объемами буфера укороченный токсин элюировали при помощи линейного градиента от 0 до 0,5М NaCl в 20 мМ CAPS в 15 колоночных объемах при объемной скорости потока 1,0 мл/мин. Очищенные укороченные трипсином Cry белки элюировались при концентрации NaCl приблизительно 0,2-0,3М. Чистоту белков проверяли при помощи SDS PAGE и с визуализацией с применением красителя Кумасси бриллиантовый голубой. В некоторых случаях объединенные фракции очищенного токсина концентрировали и загружали в колонку Superpose 6 (диаметром 1,6 см, длиной 60 см) и дополнительно очищали гель-хроматографией. Фракции, составляющие один пик с молекулярной массой мономера, объединяли и концентрировали, получая гомогенный на более чем 95% препарат белка с молекулярной массой приблизительно 60000 кДа.
Обработку Vip3Ab1 достигали схожим образом, начиная с очищенного полноразмерного белка в 85 кДа (DIG 307). Белок (12 мг) подвергали диализу в 50 мМ буфере фосфата натрия с pH 8,4, затем обрабатывали путем добавления 1 мг твердого трипсина и инкубирования в течение 1 ч при комнатной температуре. Раствор загружали в анионнообменную колонку MonoQ (диаметром 1 см, длиной 10 см) и элюировали при помощи линейного градиента NaCl от 0 до 500 мМ в 20 мМ буфере фосфата натрия с pH 8,4 более чем 7 объемами колонки. Элюирование белка наблюдали при помощи SDS-PAGE. Основная полоса обработки содержала соединения с молекулярной массой 65 кДа, что определяли при помощи SDS-PAGE с применением стандартов сравнения молекулярной массы.
Пример 3 - Биотест насекомых
Очищенные белки тестировали на инсектицидное действие в биотесте, проводимом с новорожденными личинками Plutella xylostella (Linnaeus) и Heliothis zea, находящимися на искусственном питании для насекомых. Cry1A-устойчивые P. xylostella развивались в режимах смены питания с применением коммерческого Bt продукта (DiPel®), получаемого из штамма NO-QA (Tabashnik et al., 1996; Tabashnik et al., 1997).
Биотест насекомых проводили в пластмассовых лотках для биотеста с 128 лунками (C-D International, Питман, Нью-Джерси). Каждая лунка содержала 0,5 мл корма для разных видов чешуекрылых (Southland Products, Лейк-Вилледж, Арканзас). Аликвоту объемом 40 мкл очищенного белка Cry или Vip3Ab1 разбавляли до различных концентраций в 10 мМ CAPS с pH 10,5, или в каждую лунку на поверхность корма площадью 1,5 см2 пипеткой наносили контрольный раствор (26,7 мкл/см2). Тестировали по 16 лунок на каждый образец. Отрицательным контролем служил буферный раствор, не содержащий белок. Положительные контроли включали препараты Cry1Ac или Cry1F. Обработанные лотки держали в вытяжном шкафу до тех пор, пока жидкость на поверхности корма не испарялась или не впитывалась в корм.
В течение нескольких часов после вылупления индивидуальные личинки собирали увлажненной щеткой из верблюжьей шерсти и располагали на обработанном корме по одной личинке в лунке. Затем зараженные лунки запечатывали липкими листами чистой пластмассы, через которые создавали вентиляцию для возможности газообмена (C-D International, Питман, Нью-Джерси). Лотки для биотеста держали в климатической камере (28°C, относительная влажность ~40%, фотопериод 16:8 [Свет:Темнота]). Через 5 дней всех насекомых подвергали воздействию каждого из белковых образцов, регистрировали количество мертвых насекомых и массу выживших насекомых.
Пример 4 - Иодирование Cry1Ab токсинов
Очищенный коровый токсин Cry1Ab, укороченный трипсином, иодировали с применением иодных шариков (Pierce). Вкратце, два иодных шарика промывали дважды при помощи 500 мкл PBS (20 мМ фосфат натрия, 0,15М NaCl, pH 7,5) и помещали в центрифужную пробирку на 1,5 мл за свинцовый экран. К этим иодным шарикам добавляли 100 мкл PBS. Под вытяжкой и соответствующими способами обращения с радиоактивными веществами к раствору PBS с иодными шариками добавляли 0,5 мКи Na125I (17,4 Ки/мг, Perkin Elmer). Компонентам позволили реагировать в течение 5 мин при комнатной температуре, затем 5 мкг укороченного Cry1Ab высокой степени очистки добавляли к раствору и позволяли реагировать в течение еще 3-5 мин. Реакцию завершали путем отделения раствора от иодных шариков и нанесения его на деминерализующую спин-колонку для очистки (G-20, GE biosciences), уравновешенную 10 мМ CAPS с pH 10,5. Иодные шарики промывали дважды 50 мкл PBS, и промывочный раствор также вносили в деминерализующую колонку. Радиоактивный раствор элюировали из деминерализующей колонки путем центрифугирования при 1000×g в течение 2 мин. Иодо-Cry1Ab, меченый радиоактивным изотопом 125I, измеряли в гамма-счетчике на количество радиоактивности и удельную активность, определяли на основе предположения о 80% эффективности регенерации исходного количества токсина.
Пример 5 - Получение и фракционирование растворенного препарата из BBMV
Стандартные способы количественного анализа белка и электрофорез на СДС-полиакриламидном геле применяли так, как описано, например, в публикации авторов Sambrook et al. (Sambrook и Russell, 2001) и ее обновлениях. Личинки H. zea в последней возрастной стадии не кормили в течение ночи и затем препарировали после охлаждения на льду в течение 15 мин. Ткань средней кишки удаляли из полости тела, оставляя присоединенной к наружной оболочке за задней кишкой. Среднюю кишку помещали в 9-кратный объем охлажденного льдом гомогенизирующего буфера (300 мМ маннитол, 5 мМ ЭГТА, 17 мМ Трис основание, pH 7,5) с добавлением смеси ингибиторов протеаз (Sigma-Aldrich P-2714), разбавленной согласно рекомендации производителя. Ткань гомогенизировали 15 тактами стеклянного гомогенизатора ткани. Препараты из BBMV получали методом осаждения Вульферсбергера при помощи MgCl2 (Wolfersberger, 1993). Вкратце, в равном объеме смешивали 24 мМ раствор MgCl2 в 300 мМ маннитоле с гомогенатом средней кишки, перемешивали в течение 5 мин и оставляли на льду в течение 15 мин. Раствор центрифугировали при 2500×g в течение 15 мин при 4°C. Надосадочный раствор сохраняли, а таблетку-осадок суспендировали в первоначальном объеме в 0,5х разбавленном гомогенизирующем буфере и снова центрифугировали. Эти два надосадочных раствора объединяли и центрифугировали при 27000×g в течение 30 мин при 4°C для получения фракции BBMV. Таблетку-осадок суспендировали в буфере для хранения BBMV (10 мМ HEPES, 130 мМ KCl, 10% глицерин, pH 7,4) до концентрации белка приблизительно 3 мг/мл. Концентрацию белка определяли с применением BSA в качестве стандарта.
Активность L-лейцин-п-нитроанилидаминопептидазы (маркерный фермент для фракции BBMV) определяли до замораживания образцов. Вкратце, 50 мкл L-лейцин-п-нитроанилида (1 мг/мл в PBS) добавляли к 940 мл 50 мМ Трис-HCl в стандартной кювете. Кювету помещали в спектрофотометр Cary 50 Bio, обнуляли измерение поглощения при длине волны 405 нм и инициировали реакцию путем добавления 10 мкл либо гомогената средней кишки насекомого или препарат из BBMV насекомого. Увеличение поглощения при 405 нм наблюдали в течение 5 мин при комнатной температуре. Удельную активность гомогената и препаратов BBMV определяли на основе кинетика увеличения поглощения со временем на участке линейного увеличения поглощения на единицу общего количества белка, добавленного для анализа, по следующему уравнению:
ΔOD/(мин*мг) =
Скорость аминопептидазы (ΔOD/мл*мин)/[белок](мг/мл)
Удельная активность этого фермента, как правило, увеличивалась в 7 раз по сравнению с таковой, обнаруживаемой в исходной фракции гомогената средней кишки. Аликвоты препаратов из BBMV добавляли в образцы объемом в 250 мкл, резко замораживали в жидком N2 и сохраняли при -80°C.
Пример 6 - Электрофорез
Анализ белков при помощи SDS-PAGE проводили в восстановительных условиях (т.е. в 5% β-меркаптоэтанол, BME) и денатурирующих (т.е. нагревание в течение 5 мин при 90°C в присутствии 4% СДС) условиях. Затем белки загружали в лунки с 4-20% трис-глицин полиакриломидным гелем (BioRad; Hercules, Калифорния) и разделяли при 200 В в течение 60 мин. Полоски белка обнаруживали путем окрашивания красителем Кумасси бриллиантовый синий R-250 (BioRad) в течение 1 ч и обесцвечивали раствором 5% метанола в 7% уксусной кислоте. Снимки гелей и анализ делали при помощи прибора Fluro-S Multi Imager™ компании BioRad. Относительные молекулярные массы белков в полосках определяли путем сравнения с подвижностью белков с известной молекулярной массой, наблюдаемых в образце BenchMark™ Protein Ladder (Invitrogen, Карлсбад, Калифорния), загруженном в одну лунку геля.
Пример 7 - Связывание 125I-маркированного Cry1Ab с препаратом BBMV из личинки H. zea
Строили кривую насыщения для определения оптимального количества белка в препарате BBMV для применения в тестах на связывание с коровым токсиновым Cry-белком. 0,5 нМ 125I-меченого радиоактивным изотопом корового токсинового белка Cry1Ab инкубировали в течение 1 ч при 28°C в буферном растворе (8 мМ NaHPO4, 2 мМ KH2PO4, 150 мМ NaCl, 0,1% BSA, pH 7,4) с количеством белка из BBMV в пределах от 0 мкг/мл до 500 мкг/мл (общий объем 0,5 мл). 125I-маркированный коровый токсиновый Cry-белок, связанный с белками из BBMV, отделяли от несвязанной фракции путем отбора 150 мкл реакционной смеси в трех параллелях в отдельные центрифужные пробирки объемом 1,5 мл и центрифугирования образцов при 14000×g в течение 8 мин при комнатной температуре. Надосадочный раствор аккуратно удаляли и таблетку-осадок промывали три раза ледяным буферным раствором. Основание центрифужной пробирки, содержащее таблетку-осадок, отрезали, помещали в стеклянную культуральную пробирку размером 13×75 мм, и каждый из образцов измеряли в течение 5 мин в гамма-счетчике. Строили график зависимости получаемого CPM (число импульсов в минуту) минус фоновое CPM (реакция в отсутствие белка из BBMV) от концентрации белка из BBMV. Оптимальную концентрацию белка из BBMV для применения в тесте на связывание определяли как равную 150 мкг/мл.
Тесты на гомологическое и гетерологическое конкурентное связывание проводили с применением 150 мкг/мл белка из BBMV и 0,5 нМ меченого радиоактивным изотопом 125I корового токсинового Cry-белка. Концентрации конкурентного немеченого радиоактивным изотопом корового токсинового Cry-белка, добавленного к реакционной смеси, изменялись от 0,045 нМ до 1000 нМ и ее добавляли одновременно с радиоактивного корового токсинового Cry-белка, чтобы гарантировать действительно конкурентное связывание. Инкубации проводили в течение 1 ч при 28°C, и количество 125I-маркированного корового токсинового Cry-белка, связанного с BBMV (суммарное связывание) измеряли как описано выше. Неспецифическое связывание представляли в виде счета, полученного в присутствии 1000 нМ гомологического корового токсинового Cry-белка, немеченого радиоактивым изотопом. Специфическое связывание измеряли путем вычитания полученного уровня неспецифического связывания из суммарного связывания. Стопроцентным специфическим связыванием считали количество связывания, полученное в отсутствие любого конкурентного лиганда, минус количество связывания, полученное в присутствии 1000 нМ гомологического корового токсинового Сry-белка, немеченого радиоактивным изотопом. Количество замещений гетерологическими лигандами сравнивали со 100% специфическим связыванием 125I Cry1Ab с его рецептором.
Пример 8 - Сущность результатов
Результаты смертности в биотестах с полноразмерным белком Vip3Ab1, тестированном в различных дозах против дикого типа и Cry1A-устойчивых личинок P. xylostella и личинок H. Zea, показаны на фиг.1. Результаты биотестов ингибирования роста, выраженные в процентах, представлены на фиг.2. Протестированные концентрации Vip3Ab1 составляли 9000, 3000, 1000, 333, 111, 37 и 12 нг/см2. Токсин был наиболее активен против личинок P. xylostella, показывая эквивалентный ответ на летальную дозу как для дикого типа, так и для Cry1Ab-устойчивых личинок. Половинная летальная доза LC-50 для P. xylostella составляла приблизительно 200 нг/см2. Ингибирование роста наблюдали у личинок H. zea, хотя для летального исхода во время теста требовались более высокие концентрации. Высокий уровень ингибирования роста, наблюдавшийся у личинок H. zea, указывает на то, что эти насекомые, наиболее вероятно, отмирали бы, если бы их оставляли на более длительное время.
Тесты на конкурентное связывание с радиоактивной меткой проводили для определения того, конкурирует ли укороченный трипсином Vip3Ab1 со связыванием меченными радиоактивным изотопом 125I Cry1Ab рецепторными белками, содержащимися в препаратах BBMV из H. zea.
Эксперименты проводили путем сравнения способности Vip3Ab1 конкурировать со связыванием 125I Cry1Ab из препаратов BBMV, полученных из личинок H. zea (фиг.3). В препаратах BBMV было показано, что немеченый радиоактивным изотопом Cry1Ab эффективно конкурировал со связыванием 125I Cry1Ab из белков BBMV H. zea, но Vip3Ab1 не замещал меченый радиоактивным изотопом лиганд Cry1Ab из этих препаратов BBMV. Эти три исследования демонстрируют, что Vip3Ab1 не конкурирует со связыванием Cry1Ab в личинках H. zea.
Насекомые могут развивать устойчивость к токсичности Cry-белков посредством нескольких различных биохимических механизмов, но наиболее распространенный механизм происходит из-за снижения способности токсинового Cry-белка связываться со своим специфическим рецептором в кишечнике насекомого (Heckel et al., 2007; Tabashnik et al., 2000; Xu et al., 2005). Это может быть вызвано небольшими точковыми мутациями, большими делециями гена или другими генетическими или биохимическими механизмами.
Vip3Ab1 дополняет действие Cry1Ab тем, что имеет биологическое действие против схожих насекомых, но при этом не связывается с теми же сайтами рецептора, как эти Cry-белки, и, таким образом, не зависит от механизмов устойчивости, которые вызывали бы снижение связывания Cry-токсина. Из этих исследований был сделан вывод, что Vip3Ab1 является превосходным токсином для насекомых и при объединении с Cry1Ab в качестве подхода к борьбе с устойчивостью у насекомых обеспечит биологическое действие против насекомых, которые могут развивать устойчивость к любому из этих белков, а также предотвращать выживание Cry1A-устойчивых насекомых. Так как Vip3Ab1 является токсичным для Cry1A-устойчивых насекомых, этот токсин будет превосходным партнером для стекирования при объединении с Cry1Ab для проявления активности как против дикого типа, так и против устойчивых насекомых.
Источники информации

| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ БЕЛКОВ Vip3Ab И Cry1Fa ДЛЯ ВЫРАБАТЫВАНИЯ РЕЗИСТЕНТНОСТИ К НАСЕКОМЫМ | 2010 |
|
RU2607666C2 |
| ПРИМЕНЕНИЕ Vip3Ab В СОЧЕТАНИИ С Cry1Ca ДЛЯ УПРАВЛЕНИЯ УСТОЙЧИВЫМИ НАСЕКОМЫМИ | 2010 |
|
RU2575084C2 |
| КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ БЕЛКИ Cry34Ab/35Ab И Cry3Aa, ДЛЯ ПРЕДОТВРАЩЕНИЯ РАЗВИТИЯ УСТОЙЧИВОСТИ У КУКУРУЗНЫХ КОРНЕВЫХ ЖУКОВ (Diabrotica spp.) | 2011 |
|
RU2582249C2 |
| ИНСЕКТИЦИДНЫЕ КОМБИНАЦИИ БЕЛКОВ ДЛЯ БОРЬБЫ С СОВКОЙ ТРАВЯНОЙ И КУКУРУЗНЫМ МОТЫЛЬКОМ И СПОСОБЫ УПРАВЛЕНИЯ УСТОЙЧИВОСТЬЮ НАСЕКОМЫХ | 2010 |
|
RU2577141C2 |
| КОМБИНИРОВАННОЕ ИСПОЛЬЗОВАНИЕ БЕЛКОВ CRY1Ca И CRY1Ab ДЛЯ КОНТРОЛЯ УСТОЙЧИВОСТИ НАСЕКОМЫХ | 2010 |
|
RU2596406C2 |
| КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ БЕЛКИ Cry3Aa И Cry6Aa, ДЛЯ ПРЕДОТВРАЩЕНИЯ РАЗВИТИЯ УСТОЙЧИВОСТИ У КУКУРУЗНЫХ КОРНЕВЫХ ЖУКОВ (Diabrotica spp.) | 2011 |
|
RU2576005C2 |
| КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ БЕЛКИ Cry34Ab/35Ab И Сry3Ba, ДЛЯ ПРЕДОТВРАЩЕНИЯ РАЗВИТИЯ УСТОЙЧИВОСТИ У КУКУРУЗНЫХ КОРНЕВЫХ ЖУКОВ (Diabrotica spp.) | 2011 |
|
RU2591519C2 |
| КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ БЕЛКИ Cry34Ab/35Ab И Cry6Aa, ДЛЯ ПРЕДОТВРАЩЕНИЯ РАЗВИТИЯ УСТОЙЧИВОСТИ У КУКУРУЗНЫХ КОРНЕВЫХ ЖУКОВ (Diabrotica spp. ) | 2011 |
|
RU2582262C2 |
| ПРИМЕНЕНИЕ CRY1AB В КОМБИНАЦИИ С CRY1BE ДЛЯ УПРАВЛЕНИЯ РЕЗИСТЕНТНОСТЬЮ НАСЕКОМЫХ | 2010 |
|
RU2583288C2 |
| ПРИМЕНЕНИЕ Cry1Da В СОЧЕТАНИИ С Cry1Be ДЛЯ УПРАВЛЕНИЯ УСТОЙЧИВЫМИ НАСЕКОМЫМИ | 2010 |
|
RU2590592C2 |











Изобретение относится к области биохимии, в частности к трансгенному растению, которое имеет устойчивость к насекомым Helicoverpa zea, включающему ДНК, кодирующую Vip3Ab1, и ДНК, кодирующую Cry1Ab, его семени и клетке, а также к способу задержки или предотвращения развития устойчивости у насекомых Helicoverpa zea к белкам Cry1Ab и Vip3Ab1 с его использованием. Также раскрыта совокупность растений, содержащая множество вышеуказанных трансгенных растений и не-Bacillus thuringiensis (не-Bt) растения, которые не экспрессируют трансгенные инсектицидные белки, и смесь семян, содержащая семена от не-Bt растений и множество вышеуказанных семян. Изобретение позволяет эффективно бороться с насекомым Helicoverpa zea. 7 н. и 4 з.п. ф-лы, 3 ил., 1 табл., 8 пр.
1. Трансгенное растение, которое имеет устойчивость к насекомым Helicoverpa zea, включающее ДНК, кодирующую инсектицидный белок Vip3Ab1 с SEQ ID NO: 2, и ДНК, кодирующую инсектицидный белок Cry1Ab с SEQ ID NO: 3.
2. Семя растения по п. 1, которое имеет устойчивость к насекомым Helicoverpa zea, включающее ДНК, кодирующую инсектицидный белок Vip3Ab1 с SEQ ID NO: 2, и ДНК, кодирующую инсектицидный белок Cry1Ab с SEQ ID NO: 3.
3. Совокупность растений для задержки или предотвращения развития устойчивости к белкам Vip3Ab1 и Cry1Ab у насекомых Helicoverpa zea, включающая не-Bacillus thuringiensis (не-Bt) растения, которые не экспрессируют трансгенные инсектицидные белки, и совокупность трансгенных растений по п. 1, где указанные не-Bt растения составляют 5-50% от всех растений сельскохозяйственных культур в указанной совокупности растений.
4. Совокупность растений по п. 3, в которой не-Bt растения расположены в блоках или полосах.
5. Смесь семян для задержки или предотвращения развития устойчивости к белкам Vip3Ab1 и Cry1Ab у насекомых Helicoverpa zea, включающая семена от не-Bacillus thuringiensis (не-Bt) растений, которые не экспрессируют трансгенные инсектицидные белки, и совокупность семян по п. 2, где указанные не-Bt семена составляют 5-50% от всех семян в смеси.
6. Способ задержки или предотвращения развития устойчивости у насекомых Helicoverpa zea к инсектицидному белку Cry1Ab и инсектицидному белку Vip3Ab1, получаемым из Bacillus thuringiensis, причем указанный способ включает пирамидирование инсектицидного белка Cry1Ab и инсектицидного белка Vip3Ab1 в семени растения и посев одного или более указанных семян для получения совокупности растений по п. 3.
7. Способ задержки или предотвращения развития устойчивости у насекомых Helicoverpa zea, причем указанный способ включает предоставление указанным насекомым эффективного количества композиции, где композиция включает клетки, которые экспрессируют инсектицидно эффективное количество, как инсектицидного белка Cry1Ab с SEQ ID NO: 1 или SEQ ID NO: 3, так и инсектицидного белка Vip3Ab1 с SEQ ID NO: 2.
8. Совокупность растений по п. 3, где указанная совокупность растений занимает более 10 акров.
9. Трансгенное растение по п. 1, где указанное растение выбрано из группы, состоящей из кукурузы, сои и хлопка.
10. Трансгеное растение по п. 1, в котором указанное растение является растением кукурузы.
11. Растительная клетка трансгенного растения по любому из пп. 1, 9 или 10, которая имеет устойчивость к насекомым Helicoverpa zea, где растительная клетка включает ДНК, кодирующую инсектицидный белок Vip3Ab1 с SEQ ID NO: 2, и ДНК, кодирующую инсектицидный белок Cry1Ab с SEQ ID NO: 3.
| BURKNESS E.C | |||
| et al., Novel Vip3A Bacillus thuringiensis (Bt) maize approaches high-dose efficacy against Helicoverpa zea (Lepidoptera: Noctuidae) under field conditions: Implications for resistance management, GM Crops, 2010, Vol.1, N.5, pp.337-343 | |||
| BRAVO A.et al., How to cope with insect resistance to Bt toxins?, 2008, Vol | |||
| Прибор для получения стереоскопических впечатлений от двух изображений различного масштаба | 1917 |
|
SU26A1 |
| Котел для водяного отопления с внутренним перегревателем воды для побуждения циркуляции в сети и с регулятором наружной температуры котла | 1924 |
|
SU573A1 |
| PARDO-LOPEZ L., et al., Strategies to improve the insecticidal activity of Cry toxins from Bacillus thuringiensis, Peptides, 2009, Vol.30, N.3, pp | |||
| Прибор для переработки спирта в газовую смесь для двигателей внутреннего сгорания | 1920 |
|
SU589A1 |
| ФРАГМЕНТ ГИБРИДНОГО ТОКСИНА Bacillus thuringiensis, ОБЛАДАЮЩИЙ ИНСЕКТИЦИДНОЙ АКТИВНОСТЬЮ, КОДИРУЮЩАЯ ЕГО РЕКОМБИНАНТНАЯ ДНК, СПОСОБ БОРЬБЫ С НАСЕКОМЫМИ | 1994 |
|
RU2210593C2 |

