ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Люди выращивают кукурузу ради применений в качестве пищи и энергии. Насекомые поедают и повреждают растения кукурузы и, таким образом, подрывают эти затраты человеческого труда. Сельскохозяйственными насекомыми-вредителями, приводящими к крупному уничтожению кукурузы, являются совка травяная (FAW, Spodoptera frugiperda) и кукурузный мотылек (ECB, Ostrinia nubilalis).
В настоящее время борьбу с этими вредителями с использованием вводимых в растения трансгенов успешно проводят благодаря экспрессии в растениях гена кристаллического (Cry) дельта-эндотоксина, кодирующего белок Cry1Fa из Bacillus thuringiensis. Cry1Fa является белком-токсином, в настоящее время присутствующим в сорте HerculexTM семян трансгенной кукурузы Dow AgroSciences (Herculex, Herculex-Extra, и Herculex-RW), которые являются устойчивыми к насекомым-вредителям FAW и ECB. Это белок функционирует посредством связывания со специфическим рецептором(ами), располагающимся в средней кишке насекомых, и образует поры в клетках кишки. Образование этих пор не позволяет насекомым регулировать баланс осмотически активных веществ, что приводит к их гибели.
Однако некоторые опасаются, что у насекомых, возможно, сможет развиться устойчивость к действию Cry1Fa за счет генетических изменений рецепторов в их кишке, которые связывают Cry1Fa. Насекомые, продуцирующие рецепторы с уменьшенной способностью к связыванию Cry1Fa, могут быть устойчивыми к активности Cry1Fa и, следовательно, продолжать существовать на растениях, которые экспрессируют этот белок.
В случае единственного токсина Cry, постоянно присутствующего в растении в условиях роста, существует опасение, что у насекомых могла бы развиться устойчивость к активности этого белка за счет генетических изменений рецептора, который связывает токсин Cry1Fa, в кишке насекомого. Уменьшение связывания токсина вследствие этих изменений в рецепторе будет приводить к уменьшенной токсичности Cry1Fa, которая, возможно, будет приводить к конечной уменьшенной эффективности белка после экспрессии в сельскохозяйственной культуре.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится частично к сложению определенных генов Cry наряду с Cry1Fa, чтобы иметь результатом продукты, которые являются более стойкими и менее подвержены насекомым, у которых развивается устойчивость к активности любого из токсинов (таких как Cry1Fa) самого по себе. Варианты партнеров по сложению с Cry1F включают Cry2Aa и/или Cry1I - для воздействия на совку травяную (FAW; Spodoptera frugiperda), и Cry1E - для воздействия на кукурузного мотылька (ECB; Ostrinia nubilalis).
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Настоящее изобретение включает использование по крайней мере одного из являющихся партнерами по сложению токсинов вместе с токсинами Cry1Fa в качестве пары. Некоторыми предпочтительными парами для воздействия на FAW (совку травяную; Spodoptera frugiperda) являются белок Cry1Fa плюс белок Cry1Ea. Некоторые предпочтительные пары для воздействия на ECB (кукурузного мотылька; Ostrinia nubilalis) включают белок Cry1Fa плюс белок Cry1I и/или Cry2Aa.
Настоящее изобретение также относится частично к сложениям трех токсинов или «пирамидам» из трех (или более) токсинов, при этом Cry1Fa и являющийся партнером по сложению токсин являются основной парой. Одна предпочтительная пирамида обеспечивает два механизма действия против двух вредителей - FAW и ECB. Этот тип пирамиды «2 MOA» включает Cry1Fa плюс Cry2Aa плюс Cry1Ab (предпочтительный вариант для борьбы с ECB) и Cry1Fa плюс Cry1Ea (предпочтительный вариант для борьбы с FAW). Под «независимыми механизмами действия» подразумевают, что белки не приводят к перекрестной устойчивости друг к другу.
В некоторых предпочтительных вариантах пирамид выбранные токсины обеспечивают три независимых механизма действия (активных ингредиента, которые не приводят к перекрестной устойчивости) против ECB. Предпочтительными комбинациями в пирамидах являются Cry1Fa плюс второй токсин для IRM (управления устойчивостью насекомых) плюс третий токсин для IRM. Их примерами является следующее.
Пирамиды против ECB настоящего изобретения включают токсин Cry1Fa плюс токсин Cry2Aa в качестве второго токсина для IRM и третий токсин для IRM, выбираемый из группы, состоящей из токсинов Cry1Be, Cry1Ab, DIG-3 и Cry1I.
Пирамиды против ECB настоящего изобретения включают токсин Cry1Fa плюс токсин Cry1I в качестве второго токсина для IRM и третий токсин для IRM, выбираемый из группы, состоящей из токсинов Cry1Ab, Cry1Be, DIG-3 и Cry2Aa.
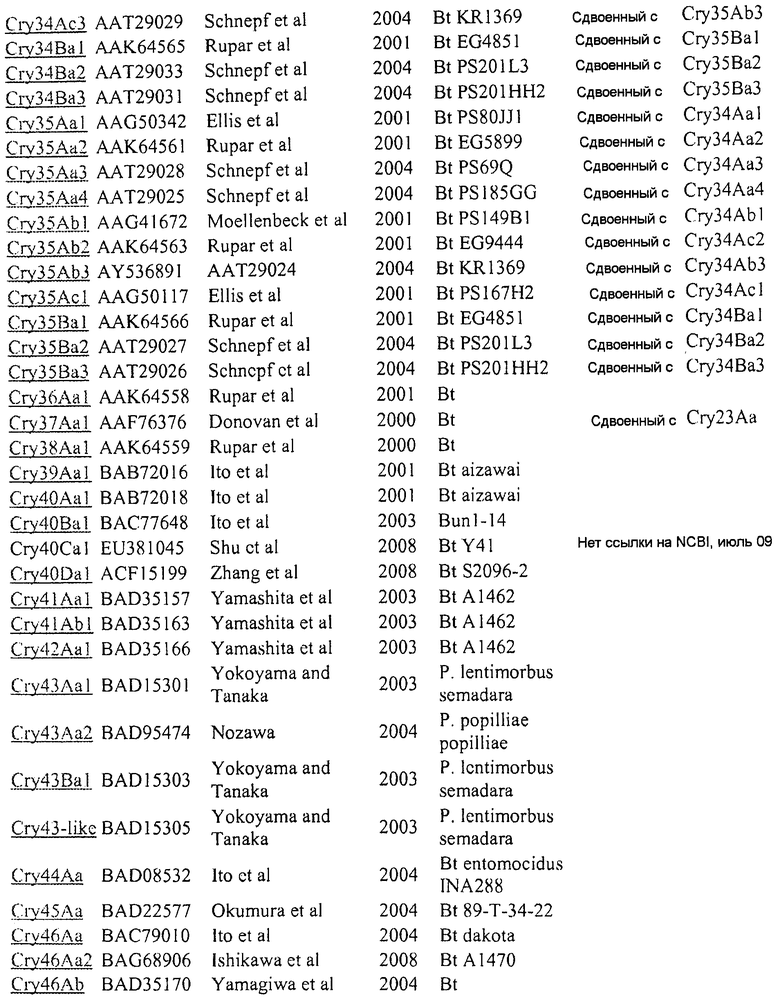
Эти различные токсины (и другие) перечислены в присоединенном приложении A. Эти номера GENBANK могут также использоваться для получения последовательностей любого из генов и белков, описываемых или упоминаемых в настоящем описании.
Для получения соответствующих последовательностей можно также использовать патенты. Например, в патенте США № 5188960 и в патенте США № 5827514 описываются содержащие основную часть токсина Cry1Fa белки, подходящие для применения при осуществлении настоящего изобретения. В патенте США № 6218188 описываются оптимизированные для растений последовательности ДНК, кодирующие содержащие основную часть токсина Cry1Fa белки, которые подходят для применения в настоящем изобретении. Заявка на патент США № 2010-00269223 относится к белкам DIG-3.
Настоящее изобретение также относится в целом к применению трех инсектицидных белков (белков Cry в некоторых предпочтительных вариантах осуществления), которые не конкурируют друг с другом/не приводят к перекрестной устойчивости, хотя являются эффективными против одного целевого вредителя.
В объем настоящего изобретения включены растения (и площадь земли (в акрах), засеянная такими растениями), которые продуцируют любую из рассматриваемых пар, или три (или более) токсина. Могут быть также добавлены дополнительные токсины/гены, но эти сложения трех конкретных токсинов будут, в соответствии с настоящим изобретением, преимущественно и как ни удивительно обеспечивать три механизма действия (не приводящие к перекрестной устойчивости активности) против FAW и/или ECB. Это может содействовать сокращению или устранению требования в отношении площади земли (в акрах) резервата (например, менее чем 40%, менее чем 20%, менее чем 10%, менее чем 5% или даже 0% резервата). Засеянное таким образом поле площадью свыше 10 акров включено, таким образом, в настоящее изобретение.
Полинуклеотид(ы) по изобретению находится предпочтительно в генетической конструкции под контролем (функционально связан/включает) промотора не из Bacillus-thuringiensis. Полинуклеотиды по изобретению могут включать частоту использования кодонов, характерную для растения, для увеличения экспрессии в растении.
Для нейтрализации способности насекомых к развитию устойчивости к Cry1Fa авторы настоящего изобретения идентифицировали токсины Cry, которые неконкурентно (с Cry1Fa) связываются с препаратами клеток кишки FAW и/или ECB. Cry1Fa не вытесняет связывание в кишке личинок насекомых FAW и ECB идентифицированных здесь белков Cry - белков Cry, таких как Cry2Aa. Способность этих токсинов Cry быть токсичными по отношению к личинкам FAW и/или ECB, несмотря на взаимодействие с абсолютно с теми же сайтами, что и Cry1Fa, указывает на то, что их токсичность не будет нарушать насекомых, у которых произошли генетические изменения их рецептора Cry1Fa в качестве механизма для становления устойчивыми к токсичности Cry1Fa.
Таким образом, насекомые, у которых развилась устойчивость к Cry1Fa за счет уменьшения способности их рецепторов кишки к связыванию Cry1Fa, будут все еще чувствительными к токсичности белков Cry2Aa, например, которые связываются с альтернативными сайтами. Авторы настоящего изобретения получили биохимические данные, подтверждающие это. Наличие комбинаций этих белков, экспрессируемых в трансгенных растениях, обеспечивает, таким образом, полезный и ценный механизм для уменьшения вероятности развития устойчивости у насекомых в поле и, следовательно, вызова снижения требования в отношении резерватов. Была исследована эффективность других белков Cry против других основных насекомых-вредителей, как чувствительных, так и устойчивых к Cry1Fa (rFAW и rECB). Как показано в таблице 1, Cry1I и Cry2Aa являются эффективными против как устойчивых, так и чувствительных личинок ECB. Cry1Ea является эффективным против как устойчивой, так чувствительной FAW. Можно получить относящиеся к связыванию данные для этих токсинов Cry против этих насекомых-вредителей. В любом случае представленные ниже данные указывают на то, что токсины взаимодействуют в независимых по сравнению с Cry1Fa сайтах мишени в кишке насекомого и будут поэтому отличными партнерами по сложению.
Сложение экспрессирующих Cry1Fa сельскохозяйственных культур с одним или более дополнительных генов Cry, таких как те, которые описаны здесь, приводит к эффективной стратегии управления для предотвращения возможности развития у насекомых устойчивости к активности трансгенных растений, экспрессирующих эти белки-токсины. Поскольку авторы настоящего изобретения демонстрируют, что эти белки Cry взаимодействуют в отличных и/или перекрывающихся сайтах по сравнению с Cry1Fa, в случае возникновения устойчивости за счет изменений сродства рецепторов кишки насекомых, которые связываются с токсинами Cry, изменения должны возникать одновременно в по крайней мере двух различных рецепторах, чтобы позволить насекомым продолжать существовать на растениях, экспрессирующих множество белков. Вероятность этого случая является чрезвычайно далекой, что тем самым увеличивает длительность отражения насекомых, которые способны к развитию устойчивости к белкам, с помощью трансгенного продукта.
Авторы настоящего изобретения подвергли йодированию с использованием радиоактивного йода усеченные с помощью трипсина формы белков-токсинов Cry и использовали методы радиорецепторного анализа для определения их взаимодействия в виде связывания с предполагаемыми рецепторными белками, расположенными в оболочках кишки насекомых. Эти оболочки кишки были приготовлены в виде мембранных везикул из щеточной каемки (BBMV) с помощью способа Вольферсбергера. Йодирование токсинов проводили, используя либо Iodo-Beads, либо обработанные пробирки Iodogen от Pierce Chemicals. Удельная активность меченного радиоактивным изотопом токсина составляла приблизительно 1-4 мкКи/мкг белка. Анализы связывания проводили по существу согласно процедурам Лианга.
Представленные в настоящем описании данные указывают на то, что токсины, взаимодействуют в независимых по сравнению с Cry1Fa сайтах мишени в кишке насекомого, и будут поэтому отличными партерами по сложению.
Настоящее изобретение можно использовать с множеством растений. Примеры включают кукурузу, сою и хлопчатник.
Гены и токсины, применимые в соответствии с настоящим изобретением, включают не только полноразмерные последовательности, которые описаны, но также фрагменты этих последовательностей, варианты, мутанты и слитые белки, которые сохраняют характеристическую пестицидную активность токсинов, конкретно приведенных здесь в качестве примера. Используемые здесь термины «варианты» или «вариации» генов относятся к нуклеотидным последовательностям, которые кодируют одинаковые токсины или которые кодируют эквивалентные токсины, обладающие пестицидной активностью. Используемый здесь термин «эквивалентные токсины» относится к токсинам, обладающим биологической активностью против целевых вредителей, одинаковой или по существу одинаковой с таковой заявленных токсинов.
Как используются в настоящем описании, пределы представляют собой составляющую приблизительно 95% (например, с Cry1Fa), 78% (например, с Cry1F) и 45% (Cry1) идентичность последовательностей согласно "Revision of the Nomenclature for the Bacillus thuringiensis Pesticidal Crystal Proteins," N. Crickmore, D.R. Zeigler, J. Feitelson, E. Schnepf, J. Van Rie, D. Lereclus, J. Baum, and D.H. Dean. Microbiology and Molecular Biology Reviews (1998) Vol. 62: 807-813. Эти отсечки могут также распространяться лишь на основные части белков (в случае основных частей белков Cry1F и Cry1Fa, например). Смотри имеющий отношение к этому созданный Crickmore et al. веб-сайт lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/.
Фрагменты и эквиваленты, которые сохраняют пестицидную активность приводимых в качестве примера токсинов, будут находиться в объеме настоящего изобретения. Также из-за избыточности генетического кода ряд отличных последовательностей ДНК может кодировать аминокислотные последовательности, описанные в настоящем описании. Создание этих альтернативных последовательностей ДНК, кодирующих одинаковые или по существу одинаковые токсины, находится полностью в пределах квалификации специалиста в данной области техники. Эти варианты последовательностей ДНК находятся в объеме настоящего изобретения. Как здесь используется, ссылка на «по существу одинаковую» последовательность относится к последовательностям, которые содержат замены, делеции или вставки аминокислот, которые фактически не влияют на пестицидную активность. Фрагменты генов, кодирующие белки, сохраняющие пестицидную активность, также включены в это определение.
Дополнительный способ идентификации кодирующих токсины генов и частей генов, применимых в соответствии с настоящим изобретением, происходит посредством использования олигонуклеотидных зондов. Эти зонды являются детектируемыми нуклеотидными последовательностями. Эти последовательности могут быть детектируемыми благодаря подходящей метке или можно сделать в основе флуоресцентными, как описано в международной заявке № W093/16094. Как хорошо известно в данной области техники, если молекула зонда и образец нуклеиновых кислот гибридизуются посредством образования прочной связи между двумя молекулами, можно обосновано предположить, что зонд и образец обладают значительной гомологией последовательностей. Предпочтительно, когда гибридизацию проводят в жестких условиях с использованием методов, хорошо известных в данной области техники, как описано, например, в Keller, G. H., M. M. Manak (1987) DNA Probes, Stockton Press, New York, N.Y., pp. 169-170. Некоторые примеры комбинаций концентраций солей и температур являются следующими (в порядке увеличения жесткости): 2X SSPE или SSC при комнатной температуре; 1X SSPE или SSC при 42ºC; 0,1X SSPE или SSC при 42ºC; 0,1X SSPE или SSC при 65ºC. Детектирование зонда обеспечивает способ определения известным образом того, произошла ли гибридизация. Такое исследование с использованием зондов обеспечивает быстрый способ идентификации кодирующих токсины генов настоящего изобретения. Нуклеотидные сегменты, используемые в качестве зондов в соответствии с настоящим изобретением, можно синтезировать, используя ДНК-синтезатор и стандартные процедуры. Эти нуклеотидные последовательности могут также использоваться в качестве праймеров для ПЦР для амплификации генов настоящего изобретения.
Некоторые белки настоящего изобретения были конкретно приведены здесь в качестве примера. Поскольку эти белки являются только примерами белков настоящего изобретения, со всей очевидностью будет вытекать, что настоящее изобретение включает являющиеся вариантами или эквивалентные белки (и нуклеотидные последовательности, кодирующие эквивалентные белки), обладающие пестицидной активностью, одинаковой или схожей с таковой приводимого в качестве примера белка. Эквивалентные белки будут иметь аминокислотную последовательность, гомологичную таковой приводимого в качестве примера белка. Эта гомология аминокислотных последовательностей будет обычно превышать 75%, предпочтительно 90% и наиболее предпочтительно 95%. Гомология аминокислотных последовательностей будет наибольшей в важных районах белка, которые отвечают за биологическую активность или вовлечены в определение трехмерной конфигурации, которая в конечном счете ответственна за биологическую активность. В связи с этим определенные аминокислотные замены являются допустимыми и могут быть ожидаемыми, если эти замены находятся в районах, которые не важны для активности или являются консервативными аминокислотными заменами, которые не оказывает влияние на трехмерную конфигурацию молекулы. Например, можно заменить аминокислоту в следующих классах: неполярные, незаряженные полярные, основные и кислотные. Консервативные замены, при которых аминокислоту одного класса заменяют на другую аминокислоту того же типа, находятся в объеме настоящего изобретения при условии, что замена не изменяет фактически биологическую активность соединения. Перечень примеров аминокислот, относящихся к каждому классу, является следующим. Важным фактором является то, что эти замены не должны значительно уменьшать биологическую активность белка.
Стратегии управления устойчивостью насекомых (IRM). Roush и др., например, кратно излагают стратегии с использованием двух токсинов, также называемые «возведением пирамиды» или «сложением», для управления инсектицидными трансгенными сельскохозяйственными культурами (The Royal Society. Phil. Trans. R. Soc. Lond. B. (1998) 353, 1777-1786).
На своем веб-сайте Управление охраны окружающей среды Соединенных Штатов (epa.gov/oppbppdl/biopesticides/pips/bt_corn_refuge_2006.htm) публикует следующие требования в отношении обеспечения засеянных нетрансгенными (т.е. без гена B.t.) культурами резерватов (участка культур/кукурузы без B.t.) для использования вместе с трансгенными сельскохозяйственными культурами, продуцирующими один белок B.t., эффективный против целевых вредителей.
«Конкретными структурными требованиями в отношении защищенных от кукурузного мотылька продуктов кукурузы с B.t. (Cry1Ab или Cry1IF) являются следующее:
Структурированные резерваты:
20% резерват кукурузы без чешуекрылых насекомых, B.t. в кукурузном поясе;
50% резерват без чешуекрылых насекомых, B.t. в поясе хлопчатника
Блоки
Внутренние (т.е. внутри поля с культурой с B.t.)
Внешние (т.е. изолированные поля в пределах ½ мили (1/4 мили, если возможно) относительно поля с культурой с B.t. для максимизации панмиксии).
Полосы на поле
Ширина полос должна составлять по крайней мере 4 ряда (предпочтительно 6 рядов) для уменьшения эффектов перемещения личинок».
Кроме того, Национальная ассоциация кукурузоводов, на своем веб-сайте (ncga.com/insect-resistance-management-fact-sheet-bt-corn) также дает схожие руководства, что касается требований в отношении резервата. Например:
«Требования IRM кукурузного мотылька:
- Засейте по крайней мере 20% Ваших акров для кукурузы для обеспечения резервата для гибридов;
- В районах выращивания хлопчатника резерват должен равняться 50%;
- Засев должен осуществляться в пределах ½ мили от гибридов в резервате;
- Резерват можно засеять в виде полос в поле с культурой с B.t.; ширина полос резервата должна составлять по крайней мере 4 ряда;
- Резерват можно обрабатывать обычными пестицидами только в случае достижения практических пределов для целевого насекомого;
- Разбрызгиваемые инсектициды на основе B.t. нельзя использовать для кукурузы в резервате;
- Соответствующий резерват должен засеиваться в каждом хозяйстве кукурузой с B.t.».
Как изложено Roush и др. (на стр. 1780 и 1784 в правом столбце, например), сложение двух различных белков, каждый из которых является эффективным против целевых вредителей и приводящим к незначительной перекрестной устойчивости или не приводящим к ней, или возведение пирамиды из них, может позволить использовать резерват меньшего размера. Roush предполагает, что в случае успешного сложения размер резервата, составляющий менее чем 10%, может обеспечить управление устойчивостью, сопоставимое с таковым в случае приблизительно 50% резервата для одного (не в виде пирамиды) признака. Для имеющихся в настоящее время продуктов кукурузы с B.t. в виде пирамиды Управление охраны окружающей среды Соединенных Штатов требует засева значительно меньшего (как правило, 5%) структурированного резервата кукурузы без B.t., чем для продуктов с одним признаком (как правило, 20%).
Существуют различные способы обеспечения эффектов резервата на IRM, включающие различные геометрические структуры засева полей (как отмечено выше) и перемешивания семян в мешке, обсуждаемые, кроме того, Roush и др. (выше) и в патенте США № 6551962.
Вышеуказанные проценты или схожие показатели для резервата, могут использоваться для рассматриваемых сложений двух или трех токсинов или пирамид из двух или трех токсинов. В случае сложений трех токсинов с тремя сайтами действия против одного целевого вредителя, целью будет равный нулю резерват (или менее чем 5% резерват, например). Это особенно верно в отношении коммерческой площади земли в более 10 акров, например.
Все патенты, заявки на патенты, предварительные заявки и публикации, упомянутые или приведенные здесь, полностью включены посредством ссылки в пределах, в которых они не противоречат точно сформированным идеям этого описания изобретения.
За исключением случаев, когда это специально оговорено или следует из контекста, грамматические формы единственного числа следует понимать как означающие “по крайней мере один”.
Следующими являются примеры, которые поясняют методы осуществления на практике настоящего изобретения. Эти примеры не должны рассматриваться как ограничение. Все проценты являются весовыми, а все доли растворителя в смесях являются объемными, кроме особо оговоренных случаев. Все температуры представлены в градусах по Цельсию.
СПИСОК ЛИТЕРАТУРЫ

Пример 1 - Биоанализ
Была исследована эффективность рассматриваемых белков Cry против других основных насекомых-вредителей как чувствительных, так и устойчивых к Cry1Fa (rFAW и rECB). Как показано в таблице 1, Cry1I и Cry2Aa являются эффективными против как устойчивых, так и чувствительных личинок ECB. Cry1Ea является эффективным против как устойчивой, так чувствительной FAW. (Ради общего рассмотрения этого вредителя смотри, например, Tabashnik, PNAS (2008), vol. 105 no. 49, 19029-19030).
| название | год | авторы | номер документа |
|---|---|---|---|
| ИНСЕКТИЦИДНЫЕ БЕЛКОВЫЕ КОМБИНАЦИИ, СОДЕРЖАЩИЕ CRY1AB И CRY2AA, ДЛЯ РЕГУЛИРОВАНИЯ КУКУРУЗНОГО МОТЫЛЬКА И СПОСОБЫ БОРЬБЫ С УСТОЙЧИВОСТЬЮ НАСЕКОМЫХ | 2010 |
|
RU2569460C2 |
| КОНТРОЛЬ УСТОЙЧИВОСТИ НАСЕКОМЫХ С ПОМОЩЬЮ КОМБИНАЦИИ БЕЛКОВ Cry1Be И Cry1F | 2010 |
|
RU2575611C2 |
| ПРИМЕНЕНИЕ CRY1AB В КОМБИНАЦИИ С CRY1BE ДЛЯ УПРАВЛЕНИЯ РЕЗИСТЕНТНОСТЬЮ НАСЕКОМЫХ | 2010 |
|
RU2583288C2 |
| ПРИМЕНЕНИЕ Cry1Da В СОЧЕТАНИИ С Cry1Be ДЛЯ УПРАВЛЕНИЯ УСТОЙЧИВЫМИ НАСЕКОМЫМИ | 2010 |
|
RU2590592C2 |
| ПРИМЕНЕНИЕ Cry1Da В СОЧЕТАНИИ С Cry1Ca ДЛЯ УПРАВЛЕНИЯ УСТОЙЧИВЫМИ НАСЕКОМЫМИ | 2010 |
|
RU2569108C2 |
| ПРИМЕНЕНИЕ Vip3Ab В СОЧЕТАНИИ С Cry1Ca ДЛЯ УПРАВЛЕНИЯ УСТОЙЧИВЫМИ НАСЕКОМЫМИ | 2010 |
|
RU2575084C2 |
| ПРИМЕНЕНИЕ ИНСЕКТИЦИДНОГО КРИСТАЛЛИЧЕСКОГО БЕЛКА DIG3 В КОМБИНАЦИИ С CRY1AB ДЛЯ РЕГУЛИРОВАНИЯ УСТОЙЧИВОСТИ К КУКУРУЗНОМУ МОТЫЛЬКУ | 2012 |
|
RU2624031C2 |
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ БЕЛКОВ Cry1Da И Cry1Fa ДЛЯ ВЫРАБАТЫВАНИЯ РЕЗИСТЕНТНОСТИ К НАСЕКОМЫМ | 2010 |
|
RU2603257C2 |
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ БЕЛКОВ Vip3Ab И Cry1Fa ДЛЯ ВЫРАБАТЫВАНИЯ РЕЗИСТЕНТНОСТИ К НАСЕКОМЫМ | 2010 |
|
RU2607666C2 |
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ VIP3AB И CRY1AB ДЛЯ РЕГУЛИРОВАНИЯ УСТОЙЧИВЫХ НАСЕКОМЫХ | 2011 |
|
RU2608500C2 |



















Изобретение относится к области биохимии, в частности к трансгенному растению, которое обладает устойчивостью к насекомым-вредителям кукурузным мотылькам, содержащее ДНК, кодирующую белок Cry1Fa, ДНК, кодирующую второй белок, выбираемый из группы, состоящей из Cry2Aa и Cry1I, а также к его семени. Раскрыто множество трансгенных растений на поле растений, включающее растения без гена Bacillus thuringiensis и множество вышеуказанных трансгенных растений, а также раскрыта смесь семян, включающая нетрансгенные семена и множество вышеуказанных трансгенных семян. Изобретение также относится к способу борьбы с насекомым кукурузным мотыльком посредством приведения указанного насекомого в контакт с белком Cry1F и вторым инсектицидным белком, выбираемым из группы, состоящей из Cry2Aa с SEQ ID NO:2 и белка Cry1I. Изобретение позволяет эффективно бороться с насекомым кукурузным мотыльком. 4 н. и 15 з.п. ф-лы, 1 табл., 1 пр.
1. Трансгенное растение, которое обладает устойчивостью к насекомым-вредителям кукурузным мотылькам (ECB, Ostrinia nubilalis), содержащее ДНК, кодирующую инсектицидный белок Cry1Fa с SEQ ID NO:1, и ДНК, кодирующую второй инсектицидный белок, выбираемый из группы, состоящей из инсектицидного белка Cry2Aa с SEQ ID NO:2 и инсектицидного белка Cry1I с SEQ ID NO:3.
2. Трансгенное растение по п. 1, дополнительно содержащее ДНК, кодирующую третий инсектицидный белок, при этом указанный третий белок выбирают из группы, состоящей из Cry1Be, CryAb и DIG-3.
3. Трансгенное растение по п. 2, где указанным третьим белком является Cry1Be, и указанное растение дополнительно содержит ДНК, кодирующую четвертый инсектицидный белок, выбираемый из группы, состоящей из Cry1Ca, Cry1Da и Vip3Ab.
4. Семя растения по п. 1, содержащее ДНК, кодирующую инсектицидный белок Cry1Fa с SEQ ID NO:1, и ДНК, кодирующую второй инсектицидный белок, выбираемый из группы, состоящей из инсектицидного белка Cry2Aa с SEQ ID NO:2 и инсектицидного белка Cry1I с SEQ ID NO:3, где указанное семя засеивают для контроля развития устойчивости кукурузного мотылька к указанным инсектицидным белкам Cry1Fa, Cry2Aa и Cry1I.
5. Множество трансгенных растений на поле растений, включающее растения без гена Bacillus thuringiensis (без B.t.), которые не экспрессируют трансгенные инсектицидные белки, и множество трансгенных растений, которые обладают устойчивостью к насекомым кукурузным мотылькам, где указанное множество трансгенных растений содержит ДНК, кодирующую второй инсектицидный белок, выбираемый из группы, состоящей из инсектицидного белка Cry2Aa с SEQ ID NO:2 и инсектицидного белка Cry1I с SEQ ID NO:3, где указанные растения без B.t. охватывают менее 40% от всех сельскохозяйственных растений на указанном множестве растений, где указанное множество растений замедляет развитие устойчивости к белкам Cry1Fa, Cry2Aa и Cry1I насекомыми кукурузными мотыльками, где указанное множество растений включает по меньшей мене одно растение без B.t.
6. Множество трансгенных растений на поле по п. 5, где указанные растения без B.t. охватывают менее 30% от всех сельскохозяйственных растений на указанном множестве растений.
7. Множество трансгенных растений на поле по п. 5, где указанные растения без B.t. охватывают менее 20% от всех сельскохозяйственных растений на указанном множестве растений.
8. Множество трансгенных растений на поле по п. 5, где указанные растения без B.t. охватывают менее 10% от всех сельскохозяйственных растений на указанном множестве растений.
9. Множество трансгенных растений на поле по п. 5, где указанные растения без B.t. охватывают менее 5% от всех сельскохозяйственных растений на указанном множестве растений.
10. Множество трансгенных растений на поле по п. 5, где указанные растения без B.t. собраны в блоках или на полосах.
11. Смесь семян, включающая нетрансгенные семена от растений без B.t., которые не экспрессируют трансгенные инсектицидные белки, и множество трансгенных семян по п. 4, где указанные нетрансгенные семена охватывают менее 40% от всех семян в смеси, где указанную смесь семян засеивают для контроля развития устойчивости кукурузного мотылька к инсектицидным белкам Cry1Fa, Cry2Aa и Cry1I, где указанная смесь семян включает по меньшей мене одно нетрансгенное семя.
12. Смесь семян по п. 11, где указанные нетрансгенные семена охватывают менее 30% от всех семян в смеси.
13. Смесь семян по п. 11, где указанные нетрансгенные семена охватывают менее 20% от всех семян в смеси.
14. Смесь семян по п. 11, где указанные нетрансгенные семена охватывают менее 10% от всех семян в смеси.
15. Смесь семян по п. 11, где указанные нетрансгенные семена охватывают менее 5% от всех семян в смеси.
16. Множество трансгенных растений по любому из пп. 5-10, где указанные растения занимают более 10 акров.
17. Растение по любому из пп. 1-4, которое выбирают из группы, состоящей из кукурузы, сои и хлопчатника.
18. Растение по п. 17, которое является кукурузой.
19. Способ борьбы с насекомым кукурузным мотыльком посредством приведения указанного насекомого в контакт с инсектицидным белком Cry1F с SEQ ID NO:1 и вторым инсектицидным белком, выбираемым из группы, состоящей из инсектицидного белка Cry2Aa с SEQ ID NO:2 и инсектицидного белка Cry1I с SEQ ID NO:3.
| BRAVO A.et al., How to cope with insect resistance to Bt toxins?, 2008, Vol | |||
| Прибор для получения стереоскопических впечатлений от двух изображений различного масштаба | 1917 |
|
SU26A1 |
| Котел для водяного отопления с внутренним перегревателем воды для побуждения циркуляции в сети и с регулятором наружной температуры котла | 1924 |
|
SU573A1 |
| US2007006340 A1, 04.01.2007 | |||
| JURAT-FUENTES JL et al., Dual resistance to Bacillus thuringiensis Cry1Ac and Cry2Aa toxins in Heliothis virescens suggests multiple mechanisms of resistance, Appl Environ Microbiol, 2003, Vol.69, N.10, pp.5898-5906. | |||

