Область техники, к которой относится изобретение
[0001] Настоящее изобретение относится к изопренсинтазе и кодирующему ее гену, и т.п.
Предшествующий уровень техники
[0002] Природный каучук представляет собой очень важное сырье для шинной и каучуковой промышленности. Несмотря на то что в будущем его потребности увеличатся вследствие моторизации в основном в странах с переходной экономикой, нелегкой задачей является увеличение сельскохозяйственных ферм, принимая во внимание обезлесивание и конкуренцию с пальмами, и едва ли ожидается повышение выхода природного каучука. Таким образом, прогнозируют, что баланс спроса и предложения станет напряженным. Синтезированный полиизопрен является доступным в качестве материала вместо природного каучука. Его исходный мономер (изопрен (2-метил-1,3-бутадиен)) в основном получают экстракцией из C5-фракции, получаемой крекингом нефти. Однако в последние годы с использованием установок для крекинга легкой фракции количество получаемого изопрена, как правило, уменьшается, и его поставка вызывает беспокойство. Также в последние годы вследствие того, что изменения цен на нефть оказывает значительное влияние, необходимо разработать систему, в которой изопрен, получаемый из ненефтяных источников, получают при низких затратах для стабильного обеспечения поставки изопренового мономера.
[0003] Касательно такой потребности, был описан способ, в котором мономер изопрена получают с использованием трансформанта, получаемого введением гена изопренсинтазы и соответствующего мутанта, получаемого из выделенных пуэрарии волосистой или тополя, в микроорганизм для ферментативной продукции (см. патентную литературу 1 и 2).
Ссылки на известный уровень техники
Патентная литература
[0004]
Патентная литература 1: JP публикация №2011-505841
Патентная литература 2: JP публикация №2011-518564
Непатентная литература
[0005]
Непатентная литература 1: Kesselmeier J. et al., Journal of Atmospheric Chemistry, vol.33, pp.23-88, 1999.
Непатентная литература 2: Monson R.K. et al., Plant Physiol., vol. 98, pp.117 5-1180, 1992.
Непатентная литература 3: Kuzma J. et al., Plant Physiol., vol. 101, pp.435-440, 1993.
Описание изобретения
Задача, подлежащая решению посредством изобретения
[0006] Однако ферментативная активность изопренсинтазы, описанной в патентной литературе 1 и 2, являлась низкой, и все еще существовали возможности для улучшения продукции изопрена с высокой продуктивностью, когда использовали такие гены изопренсинтазы.
[0007] Настоящее изобретение было создано исходя из указанных выше обстоятельств, и задачей настоящего изобретения является предоставление средств, пригодных для разработки превосходной системы, продуцирующей мономер изопрена.
Средства решения проблемы
[0008] В результате тщательного исследования для решения указанной выше проблемы авторы настоящего изобретения обнаружили, что изопренсинтаза, получаемая из мукуны (Mucuna pruriens), обладает превосходной способностью продуцировать мономер изопрена, и осуществили настоящее изобретение.
[0009] Таким образом, настоящее изобретение составляет следующее.
[1] Полинуклеотид из указанных ниже (a), (b) или (c):
(a) полинуклеотида, содержащего (i) нуклеотидную последовательность, представленную SEQ ID NO: 1, или (ii) нуклеотидную последовательность, состоящую из нуклеотидных остатков в положениях 133-1785 в нуклеотидной последовательности, представленной SEQ ID NO: 1;
(b) полинуклеотида, который содержит нуклеотидную последовательность, обладающую 90% или более идентичностью нуклеотидной последовательности (i) или (ii), и кодирует белок, обладающей активностью изопренсинтазы, или
(c) полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом, состоящим из нуклеотидной последовательности комплементарной нуклеотидной последовательности (i) или (ii), и кодирует белок, обладающий активностью изопренсинтазы.
[2] Полинуклеотид по пункту [1], где полинуклеотид получают из мукуны.
[3] Белок из указанных ниже (A), (B) или (C):
(A) белка, содержащего (i') полноразмерную аминокислотную последовательность, представленную SEQ ID NO: 2, или (ii') аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 45-594 в аминокислотной последовательности, представленной SEQ ID NO: 2;
(B) белка, который содержит аминокислотную последовательность, обладающую 90% или более идентичностью аминокислотной последовательности (i') или (ii'), и обладает активностью изопренсинтазы, или
(C) белка, который содержит аминокислотную последовательность, содержащую делецию, замену, добавление или вставку одной или более аминокислот в аминокислотной последовательности (i') или (ii'), и обладает активностью изопренсинтазы.
[4] Экспрессирующий вектор, содержащий полинуклеотид по пункту [1] или [2], или полинуклеотид, кодирующий белок по пункту [3].
[5] Трансформант, полученный введением экспрессирующего вектора по пункту [4] в хозяина.
[6] Трансформант по пункту [5], где хозяин обладает способностью синтезировать диметилаллилдифосфат метилэритриолфосфатным путем.
[7] Трансформант по пункту [6], где хозяин представляет собой Escherichia coli.
[8] Трансформант по любому из пунктов [5]-[7], где трансформант обладает способностью синтезировать диметилаллилдифосфат как мевалонатному пути, так и по метилэритриолфосфатному пути.
[9] Трансформант по пункту [5], где хозяином является микроорганизм, принадлежащий роду Corynebacterium, роду Pantoea, роду Enterobacter или роду Saccharomyces.
[10] Способ получения белка по пункту [3], включающий получение белка по пункту [3] с использованием трансформанта по любому из пунктов [5]-[9].
[11] Способ получения изопренового мономера, включающий получение изопренового мономера из диметилаллилдифосфата в присутствии белка по пункту [3].
[12] Способ по пункту [11], где мономер изопрена получают культивированием трансформанта по любому из пунктов [5]-[9].
[13] Способ по пункту [12], где обеспечивают поступление диметилаллилдифосфата из источника углерода в среде посредством культивирования трансформанта.
[14] Способ получения полимера изопрена, включающий (I) и (II):
(I) получение изопренового мономера способом по любому из пунктов [11]-[13], и
(II) полимеризацию изопренового мономера с образованием изопренового полимера.
Эффект изобретения
[0010] В соответствии с настоящим изобретением возможна разработка превосходной продуцирующей мономер изопрена системы.
Краткое описание фигур
[0011] На фиг. 1 представлен график, демонстрирующий количество изопрена, продуцируемого на единицу массы сухих листьев различных растений;
на фиг. 2 представлен график, демонстрирующий количество изопрена, продуцируемого относительно количества общего белка, экстрагируемого из листьев различных растений;
на фиг. 3 представлено изображение, на котором представлена схема правого элемента оперона и его проксимальной области мевалонатного пути на фиксированной хромосоме, и
на фиг. 4 представлено изображение, на котором представлены правый элемент оперона и его проксимальная область мевалонатного пути, регулируемая промотором tac, на хромосоме.
Варианты осуществления для реализации изобретения
[0012] Полинуклеотид, кодирующий изопренсинтазы
Настоящее изобретение относится к полинуклеотиду, кодирующему изопренсинтазу.
Изопренсинтаза представляет собой фермент, который преобразует диметилаллилдифосфат в изопрен. Авторы настоящего изобретения, во-первых, оценили способность продукции изопрена в различных растениях при одинаковых условиях при высокой температуре. В результате чего, было обнаружено, что способность мукуны (Mucuna bracteata) продуцировать изопрен являлась в несколько раз, в несколько десятков раз больше по сравнению со способностью продуцировать изопрен тополем (Populus nigra var. italic) и пуэрарии волосистой (Pueraria lobata). Однако не ясно, относится ли такая высокая способность продуцировать изопрен к высокой способности растения продуцировать диметилаллилдифосфат, высокой продукции изопренсинтазы растением или к высокой специфической ферментативной активности изопренсинтазы, которой обладает растение. В частности, исследовали зависимость от pH и т.п. активности изопренсинтазы в мукуне с использованием фермента, частично очищенного фракционированием сульфатом аммония согласно Monson and Kuzma et al., в непатентной литературе 2 и 3, но оставалось неясным, являлась ли специфическая активность в мукуне выше, чем в пуэрарии волосистой и тополе или не являлась. Таким образом, авторы настоящего изобретения частично очищали изопренсинтазу из листьев мукуны и пуэрарии волосистой фракционированием сульфатом аммония и исследовали специфическую активность раствора неочищенного экстракта. В результате было обнаружено, что специфическая активность изопренсинтазы, полученной из мукуны, являлась в несколько раз, несколько десятков раз выше, чем у изопренсинтазы, полученной из пуэрарии волосистой. Также было продемонстрировано, что измеряемая специфическая активность изопренсинтазы из мукуны являлась в несколько раз, несколько десятков раз выше, чем специфическая активность изопренсинтазы в неочищенном ферменте из тополя, опубликованная в патентной литературе 1 и 2. На основании указанного выше было сделано предположение, что мукуна потенциально содержит высокофункциональную изопренсинтазу.
[0013] Затем анализировали нуклеотидную последовательность гена изопренсинтазы из мукуны. Один из примеров способов анализа будет описан ниже.
Известно, что изопренсинтаза экспрессируется только в листьях растения, и уровень ее экспрессии повышается при высоком освещении и высокой температуре. Таким образом, сначала экстрагировали общую РНК из листьев мукуны, где индуцировали транскрипцию мРНК изопренсинтазы в условиях освещения при температуре 40°C. Подтверждали, что экстрагированная общая РНК не разрушалась и не являлась загрязненной геномной ДНК. Затем общую РНК преобразовывали в двойную цепь и фрагментировали, и анализировали только нуклеотидные последовательности, содержащие последовательность поли-A на 3'-конце с использованием высокопроизводительного секвенатора. Перекрывающиеся последовательности объединяли с получением совокупности последовательностей контига. Для таких последовательностей контига проводили поиск в программе BLAST, и детектировали последовательность контига с гомологией (идентичностью нуклеотидных последовательностей) с регистрируемыми последовательностями известных генов изопренсинтазы пуэрарии волосистой и тополя, и получали неполную последовательность гена изопренсинтазы мукуны. На основании такой частичной последовательности проводили 5' RACE (быструю амплификацию концов кДНК) стандартными способами анализа полноразмерной нуклеотидной последовательности гена изопренсинтазы мукуны, которая представлена SEQ ID NO: 1.
кДНК изопренсинтазы из мукуны можно получать, например, RT-ПЦР общей РНК, полученной, как указано выше, в качестве матрицы с использованием праймеров, сконструированных на основании информации об анализируемой нуклеотидной последовательности гена изопренсинтазы мукуны.
[0014] В одном из вариантов осуществления полинуклеотид по настоящему изобретению представляет собой полинуклеотид, содержащий (i) нуклеотидную последовательность, представленную SEQ ID NO: 1, или (ii) нуклеотидную последовательность, состоящую из нуклеотидных остатков в положениях 133-1785 в нуклеотидной последовательности, представленной SEQ ID NO: 1. Нуклеотидная последовательность, представленная SEQ ID NO: 1, может кодировать аминокислотную последовательность, представленную SEQ ID NO: 2, нуклеотидная последовательность, состоящая из нуклеотидных остатков в положениях 1-132, может кодировать предполагаемый сигнал локализации хлоропласта, и нуклеотидная последовательность, состоящая из нуклеотидных остатков в положениях 133-1785, может кодировать аминокислотную последовательность зрелой изопренсинтазы.
[0015] В другом варианте осуществления полинуклеотид по настоящему изобретению представляет собой полинуклеотид, который содержит нуклеотидную последовательность, обладающую 90% или более идентичностью с нуклеотидной последовательностью (i) или (ii), как указано выше, и кодирует белок, обладающий активностью изопренсинтазы. Процентная идентичность с нуклеотидной последовательностью может составлять 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, или более. Активность изопренсинтазы относится к активности в отношении образования изопрена из диметилаллилдифосфата (DMAPP) (это же обозначение будет применяться далее в настоящем описании).
[0016] Процентная идентичность нуклеотидных последовательностей и процентная идентичность аминокислотных последовательностей, как описано ниже, можно определять с использованием алгоритма BLAST (Pro. Natl. Acad. Sci. USA, 90, 5873 (1993)) согласно Karlin and Altschul и FASTA (Methods Enzymol., 183, 63 (1990)) согласно Pearson. Программы, обозначаемые как BLASTP и BLASTN, были разработаны на основе такого алгоритма BLAST (см. http://www.ncbi.nlm.nih.gov). Таким образом, процентную идентичность нуклеотидных последовательностей и аминокислотных последовательностей можно рассчитывать с использованием этих программ с настройками по умолчанию. Также в отношении гомологии аминокислотных последовательностей можно использовать, например, числовое значение, полученное вычислением сходства в виде процента при настройках "размер единицы для сравнения=2" с использованием полной длины участка полипептида, кодируемого в ORF, с использованием программного обеспечения GENETYX Ver. 7.0.9 от Genetyx Corporation, в котором применяют способ Липмана-Пирсона. Наименьшее значение среди значений, полученных в таких вычислениях, можно использовать качестве процентной идентичности нуклеотидных последовательностей и аминокислотных последовательностей.
[0017] В еще одном другом варианте осуществления полинуклеотид по настоящему изобретению представляет собой полинуклеотид, который гибридизуется в жестких условиях с полинуклеотидом, состоящим из нуклеотидной последовательности комплементарной нуклеотидной последовательности (i) или (ii), как указано выше, и кодирует белок, обладающий активностью изопренсинтазы.
[0018] "Жесткие условия" относятся к условиям, когда образуется так называемый специфический гибрид, при этом неспецифический гибрид не образуется. Четкое количественное определение таких условий вызывает затруднение. Тем не менее, ссылаясь на конкретный случай, такие условия представляют собой условия, когда по существу одинаковые полинуклеотиды с высокой идентичностью, например, полинуклеотиды с описанной выше процентной идентичностью гибридизуются друг с другом, тогда как полинуклеотиды с более низкой идентичностью по сравнению с указанными выше не гибридизуются друг с другом. В частности, такие условия могут включать гибридизацию в 6×SCC (хлорид натрия/цитрат натрия) приблизительно при 45°C с последующим одним или двумя, или более промываниями в 0,2×SCC и 0,1% SDS при 50-65°C.
[0019] Полинуклеотид по настоящему изобретению может представлять собой ДНК или РНК и предпочтительно представляет собой ДНК. Полинуклеотид по настоящему изобретению можно получать из мукуны. Полинуклеотид по настоящему изобретению также может представлять собой полинуклеотид, кодирующий белок по настоящему изобретению, описанный ниже.
[0020] Изопренсинтаза
Настоящее изобретение также относится к белку, обладающему активностью изопренсинтазы. Активность изопренсинтазы является такой, как описано выше.
[0021] В одном из вариантов осуществления белок по настоящему изобретению представляет собой белок, содержащий (i') полноразмерную аминокислотную последовательность, представленную SEQ ID NO: 2, или (ii') аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 45-594 в аминокислотной последовательности, представленной SEQ ID NO: 2. Аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 1-44 в аминокислотной последовательности, представленной SEQ ID NO: 2, может кодировать предполагаемый сигнал локализации хлоропласта, и аминокислотная последовательность, состоящая из аминокислотных остатков в положениях 45-594 в аминокислотной последовательности, представленной SEQ ID NO: 2, может кодировать зрелую изопренсинтазу.
[0022] В другом варианте осуществления белок по настоящему изобретению представляет собой белок, который содержит аминокислотную последовательность, обладающую 90% или более идентичностью с аминокислотной последовательностью (i') или (ii'), как указано выше, и обладает активностью изопренсинтазы. Процент идентичности с аминокислотной последовательностью может составлять 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, или более.
[0023] В еще одном другом варианте осуществления белок по настоящему изобретению представляет собой белок, который содержит аминокислотную последовательность, содержащую мутацию одной или более аминокислот в аминокислотной последовательности (i') или (ii'), как указано выше, и обладает активностью изопренсинтазы. Примеры мутации аминокислотных остатков может включать делецию, замену, добавление и вставку аминокислотных остатков. Мутацию одной или более аминокислот можно вводить в одну область или многие различные области в аминокислотной последовательности. Термин "один или более" указывает на диапазон, в котором трехмерная структура и активность белка существенно не ухудшается. В случае белка, число, представленное фразой "один или более" равно, например, 1-100, предпочтительно 1-80, более предпочтительно 1-50, 1-30, 1-20, 1-10 или 1-5. Белок по настоящему изобретению может содержать метку для очистки, такую как гистидиновая метка.
[0024] Белок по настоящему изобретению предпочтительно обладает активностью изопренсинтазы, которая составляет 50% или более, 60% или более, 70% или более, 80% или более, 90% или более или 95% или более активности изопренсинтазы белка, содержащего аминокислотную последовательность любой из (i') или (ii'), как указано выше, при измерении в одинаковых условиях. Касательно стабильности также предпочтительно, чтобы белок по настоящему изобретению обладал остаточной активностью, которая составляет 30% или более, 40% или более, 50% или более, 60% или более, или 65% или более исходной активности при хранении белка в определенном буфере [например, растворе 50 мМ Tris-HCl (pH 8,0) и 0,15 мМ MgCl2] при 4°C в течение 48 часов.
[0025] В белке по настоящему изобретению мутацию можно вводить в участки в каталитическом домене и в участки, отличные от каталитического домена, при условии, что сохраняется целевая активность. Положения аминокислотных остатков, которые необходимо подвергать мутации, которые способны сохранять целевую активность, известны специалисту в данной области. В частности, специалист в данной области может проводить корреляцию между структурой и функцией, т.к. специалист в данной области может 1) сравнивать аминокислотные последовательности многих белков, обладающих одним и тем же типом активности (например, аминокислотную последовательность, представленную SEQ ID NO: 2, и аминокислотные последовательности отличные от изопренсинтаз), 2) выяснять области, которые являются относительно консервативными, и области, которые не являются относительно консервативными, и затем 3) прогнозировать области, которые могут играть функционально важную роль, и области, которые не могут играть функционально важную роль, из областей, которые являются относительно консервативными, и областей, которые не являются относительно консервативными, соответственно. Таким образом, специалист в данной области может идентифицировать положения аминокислотных остатков, которые необходимо подвергнуть мутации, в аминокислотной последовательности изопренсинтазы.
[0026] Когда аминокислотный остаток подвергают мутации посредством замены, замена аминокислотного остатка может представлять собой консервативную замену. Как используют в настоящем описании, термин "консервативная замена" относится к замене определенного аминокислотного остатка на аминокислотный остаток со сходной боковой цепью. Семейства аминокислотных остатков со сходной боковой цепью хорошо известны в данной области. Примеры таких семейств могут включать аминокислоты, содержащие основную боковую цепь (например, лизин, аргинин, гистидин), аминокислоты, содержащие кислотную боковую цепь (например, аспарагиновая кислота, глутаминовая кислота), аминокислоты, содержащие с незаряженную полярную боковую цепь (например, аспарагин, глутамин, серин, треонин, тирозин, цистеин), аминокислоты, содержащие неполярную боковую цепь (например, глицин, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), аминокислоты, содержащие разветвленную боковую цепь в положении β (например, треонин, валин, изолейцин), аминокислоты, содержащие ароматическую боковую цепь (например, тирозин, фенилаланин, триптофан, гистидин), аминокислоты, содержащие боковую цепь, содержащую гидроксильную группу (например, спиртовую, фенольную) (например, серин, треонин, тирозин), и аминокислоты, содержащие боковую цепь, содержащую серу (например, цистеин, метионин). Предпочтительно консервативная замена аминокислот может представлять собой замену между аспарагиновой кислотой и глутаминовой кислотой, замену среди аргинина, лизина и гистидина, замену между триптофаном и фенилаланином, замену между фенилаланином и валином, замену между лейцином, изолейцином и аланином и замену между глицином и аланином.
[0027] Экспрессирующий вектор
Настоящее изобретение относится к экспрессирующему вектору. Экспрессирующий вектор по настоящему изобретению содержит полинуклеотид по настоящему изобретению или полинуклеотид, кодирующий белок по настоящему изобретению.
[0028] Примеры экспрессирующего вектора по настоящему изобретению могут включать вектор на основе клеточных систем, которые экспрессирует белок в клетке-хозяине, и вектор на основе бесклеточных систем, в котором используют систему трансляции белка. Экспрессирующий вектор также может представлять собой плазмиду или интегрирующий вектор. Экспрессирующий вектор может дополнительно представлять собой вектор на основе ДНК или вектор на основе РНК.
Известный вектор, подходящий для клетки-хозяина, используют в качестве вектора на основе клеточной системы. Примеры экспрессирующего вектора могут включать плазмиды на основе ColE, типичными представителями которых являются производные pBR322, плазмиды на основе pACYC с происхождением p15A, плазмиды на основе pSC и миниплазмиды F, полученные из фактора F Вас и т.п. в Escherichia coli. Кроме того, также можно использовать экспрессирующие векторы, содержащие триптофановый промотор, такой как trc и tac, промотор lac, промотор T7, промотор T5, промотор T3, промотор SP6, индуцируемый арабинозой промотор, промотор холодового шока, индуцируемый тетрациклином промотор и т.п.
Примеры вектора на основе бесклеточной системы могут включать экспрессирующие векторы, содержащие промотор T7, или содержащие промотор T3 в качестве примера вектора на основе клеточных систем, и векторы для синтеза белков в бесклеточной системе из зародышей пшеницы, такой как плазмиды на основе pEU, содержащие промотор SP6 или промотор T7.
[0029] При синтезе белка с использованием вектора на основе бесклеточных систем кДНК для целевого белка сначала транскрибируют для синтеза мРНК с использованием системы транскрипции. Такая система транскрипции может содержать системы, общеизвестные и опубликованные, в которых кДНК транскрибируется РНК-полимеразой. Примеры РНК-полимеразы могут включать РНК-полимеразу T7.
Затем, мРНК подвергается трансляции с синтезом белок с использованием бесклеточной системы синтеза белка, которая представляет собой систему трансляции. В эту систему входят факторы, такие как рибосома, фактор инициации трансляции, фактор элонгации трансляции, фактор диссоциации и аминоацил-тРНК-синтетаза, которые необходимы для трансляции. Примеры такой системы трансляции белка могут включать экстракт E. coli, экстракт ретикулоцитов кролика и экстракт зародышей пшеницы.
Кроме того, также может быть включена восстановленная бесклеточная система синтеза белка, состоящая из факторов, необходимых для указанной выше трансляции, которые независимо очищают.
Синтез белка с использованием вектора на основе клеточной системы описан ниже в разделе "Трансформанты".
Белок, синтезируемый с использованием вектора на основе клеточной системы или вектора на основе бесклеточной системы, можно подвергать очистке. Примеры способов очистки могут включать способы с использованием метода высаливания и различных хроматографических методов. Когда экспрессирующий вектор конструируют для экспрессии последовательности метки, такой как гистидиновая метка на N-конце или С-конце целевого белка, можно применять способ очистки с использованием аффинной хроматографии с использованием вещества, такого как никель или кобальт с аффинностью в отношении такой метки. Кроме того, чистоту белка по настоящему изобретению можно увеличивать подходящим методом очистки в комбинации с ионообменной хроматографией, гель-фильтрационной хроматографией или т.п.
[0030] Трансформанты
Настоящее изобретение относится к трансформанту, содержащему экспрессирующий вектор по настоящему изобретению. Трансформант по настоящему изобретению представляет собой трансформант, полученный введением экспрессирующего вектора по настоящему изобретению в хозяина. Хозяин, используемый для настоящего изобретения, предпочтительно представляет собой бактерию или гриб. Бактерия может представлять собой грамположительную бактерию или грамотрицательную бактерию.
[0031] Примеры грамположительных бактерий могут включать бактерии, принадлежащие роду Bacillus, Listeria, Staphylococcus, Streptococcus, Enterococcus, Clostridium, Corynebacterium и Streptomyces. Предпочтительными являются бактерии, принадлежащие роду Bacillus и Corynebacterium.
Примеры бактерий, принадлежащих роду Bacillus, могут включать Bacillus subtilis, Bacillus anthracis и Bacillus cereus. Bacillus subtilis является наиболее предпочтительной.
Примеры бактерий, принадлежащие роду Corynebacterium, могут включать Corynebacterium glutamicum, Corynebacterium efficiens и Corynebacterium callunae. Corynebacterium glutamicum является наиболее предпочтительной.
[0032] Примеры грамотрицательных бактерий могут включать бактерии, принадлежащие роду Escherichia, Pantoea, Salmonella, Vivrio, Serratia и Enterobacter. Предпочтительными являются бактерии, принадлежащие роду Escherichia, Pantoea и Enterobacter.
Escherichia coli является предпочтительной в качестве бактерий, принадлежащих роду Escherichia.
Примеры бактерий, принадлежащих роду Pantoea, могут включать Pantoea ananatis, Pantoea stewartii, Pantoea agglomerans и Pantoea citrea. Предпочтительными являются Pantoea ananatis и Pantoea citrea. В качестве бактерий, принадлежащих роду Pantoea, можно использовать штаммы, проиллюстрированные в публикации европейской патентной заявки 0952221. Примеры характерных штаммов бактерий, принадлежащих роду Pantoea, могут включать штамм AJ13355 Pantoea ananatis (FERM BP-6614) и штамм AJ13356 Pantoea ananatis (FERM BP-6615), описанные в публикации европейской патентной заявки 0952221.
Примеры бактерий, принадлежащих роду Enterobacter, могут включать Enterobacter agglomerans и Enterobacter aerogenes. Предпочтительным является Enterobacter aerogenes. Бактериальные штаммы, проиллюстрированные в публикации европейской патентной заявки 0952221, можно использовать в качестве бактерий, принадлежащих роду Enterobacter. Примеры характерных штаммов бактерий, принадлежащих роду Enterobacter, могут включать штамм ATCC12287 Enterobacter agglomerans, штамм TACC13048 Enterobacter aerogenes, штамм NBRC12010 Enterobacter aerogenes (Biotechnol. Bioeng., 2007, Mar. 27; 98 (2):340-348) и AJ110637 Enterobacter aerogenes (FERM BP-10955). Штамм AJ110637 Enterobacter aerogenes был депонирован в Международном депозитарии патентуемых микроорганизмов (IPOD), Национального института новых промышленных исследований и технологий (AIST) (Chuo № 6, Higashi 1-1-1, Tsukuba City, Ibaraki Pref., JP, Postal code 305-8566) 22 августа 2007 года, и перенесен в международный депозитарий на основании Будапештского договора 13 марта 2008 года, и ему был присвоен номер доступа FERM BP-10955.
[0033] Примеры гриба могут включать микроорганизмы, принадлежащие родам Saccharomyces, Schizosaccharomyces, Yarrowia, Trichoderma, Aspergillus, Fusarium и Mucor. Предпочтительными являются микроорганизмы, принадлежащие родам Saccharomyces, Schizosaccharomyces, Yarrowia или Trichoderma.
Примеры микроорганизмов, принадлежащих роду Saccharomyces, могут включать Saccharomyces carlsbergensis, Saccharomyces cerevisiae, Saccharomyces diastaticus, Saccharomyces douglasii, Saccharomyces kluyveri, Saccharomyces norbensis и Saccharomyces oviformis. Предпочтительным является Saccharomyces cerevisiae.
Schizosaccharomyces pombe является предпочтительным в качестве микроорганизмов, принадлежащих роду Schizosaccharomyces.
Yarrowia lypolytica является предпочтительной в качестве микроорганизмов, принадлежащих роду Yarrowia.
Примеры микроорганизмов, принадлежащих роду Trichoderma, могут включать Trichoderma harzianum, Trichoderma koningii, Trichoderma longibrachiatum, Trichoderma reesei и Trichoderma viride. Trichoderma reesei является предпочтительным.
Кроме того, хозяин, используемый для настоящего изобретения, конкретно не ограничен, при условии, что хозяин обладает способностью синтезировать диметилаллилдифосфат (DMAPP) через путь мевалоновой кислоты (MVA) и/или метилэритриолфосфатный (МЕР) путь, которые вовлечены в синтез диметилаллилдифосфата, который представляет собой субстрат изопренсинтазы и может являться клеткой насекомого, клеткой животного, растительной клеткой и т.п.
[0034] В трансформанте по настоящему изобретению можно активировать путь синтеза диметилаллилдифосфата (DMAPP), который является субстратом изопренсинтазы. Для такой активации в трансформант по настоящему изобретению можно вводить экспрессирующий вектор, который экспрессирует изопентенилдифосфат-дельта-изомеразу, обладающую способностью преобразовывать изопентенилдифосфат (IPP) в диметилаллилдифосфат (DMAPP). В трансформант по настоящему изобретению также можно вводить экспрессирующий вектор, который экспрессирует один или более ферментов, участвующих в мевалонатном пути и/или метилэритриолфосфатном пути, связанных с образованием IPP и/или DMAPP. Экспрессирующий вектор для такого фермента может представлять собой плазмиду или интегрирующий вектор. Экспрессирующий вектор для такого фермента также может представлять собой вектор на основе ДНК или вектор на основе РНК. Экспрессирующий вектор для такого фермента может дополнительно экспрессировать ряд ферментов (например, один, два, три или четыре, или более), участвующих в мевалонатном пути и/или метилэритриолфосфатном пути, и может представлять собой, например, экспрессирующий вектор для полицистронной мРНК. Происхождение одного или более ферментов, участвующих в мевалонатном путем и/или метилэритриолфосфатном пути, может быть гомологичное или гетерологичное по отношению к хозяину. Когда происхождение фермента, участвующего в мевалонатном пути и/или метилэритриолфосфатном пути, является гомологичным по отношению к хозяину, например, может представлять собой бактерию, как описано выше (например, Escherichia coli), и фермент, участвующий в мевалонатном пути, можно получать из гриба (например, Saccharomyces cerevisiae). Кроме того, когда хозяин по своей природе продуцирует фермент, участвующий в метилэритриолфосфатном пути, экспрессирующий вектор, который следует вводить хозяину, может экспрессировать фермент, участвующий в мевалонатном пути.
[0035] Примеры изопентенилдифосфат-дельта-изомеразы (EC:5.3.3.2) могут включать Idi1p (номер доступа NP_015208), AT3G02780 (номер доступа NP_186927), AT5G16440 (номер доступа NP_19714 8) и Idi (номер доступа NP_417365).
[0036] Примеры ферментов, участвующих в мевалонатном (MVA) пути, могут включать мевалонаткиназу (EC: 2.7.1.36; пример 1, Erg12p номер доступа NP_013935; пример 2, AT5G27450 номер доступа NP_001190411), фосфомевалонаткиназу (EC: 2.7.4.2; пример 1, Erg8p номер доступа NP_013947; пример 2, AT1G31910 номер доступа NP_001185124), дифосфомевалонатдекарбоксилазу (EC: 4.1.1.33; пример 1, Mvd1p номер доступа NP_014441; пример 2, AT2G38700 номер доступа NP_181404; пример 3, AT3G54250 номер доступа NP_566995), ацетил-CoA-C-ацетилтрансферазу (EC: 2.3.1.9; пример 1, Erg10p номер доступа NP_015297; пример 2, AT5G47720 номер доступа NP_001032028; пример 3, AT5G48230 номер доступа ID NP_568694), гидроксиметилглутарил-CoA-синтазу (EC: 2.3.3.10; пример 1, Erg13p номер доступа NP_013580; пример 2, AT4G11820 номер доступа NP_192919; пример 3, MvaS номер доступа AAG02438), гидроксиметилглутарил-CoA-редуктазу (EC: 1.1.1.34; пример 1, Hmg2p номер доступа NP_013555; пример 2, Hmg1p номер доступа NP_013636; пример 3, AT1G76490 номер доступа NP_177775; пример 4, AT2G17370 номер доступа NP_179329, EC: 1.1.1.88, пример, MvaA номер доступа P13702) и ацетил-CoA-C-ацетилтрансферазу/гидроксиметилглутарил-CoA-редуктазу (EC: 2.3.1.9/1.1.1.34, пример, MvaE номер доступа AAG02439).
[0037] Примеры ферментов, участвующих в метилэритриолфосфатном (МЕР) пути, могут включать 1-дезокси-D-ксилулоза-5-фосфатсинтетазу (EC: 2.2.1.7, пример 1, Dxs номер доступа NP_414954; пример 2, AT3G21500 номер доступа NP_5 66686; пример 3, AT4G15560 номер доступа NP_193291; пример 4, AT5G11380 номер доступа NP_001078570), 1-дезокси-D-ксилулоза-5-фосфатредкутоизомеразу (EC: 1.1.1.267; пример 1, Dxr номер доступа NP_414715; пример 2, AT5G62790 номер доступа NP_001190600), 4-дифосфоуитидил-2-C-метил-D-эритритолсинтазу (EC: 2.7.7.60; пример 1, IspD номер доступа NP_417227; пример 2, AT2G02500 номер доступа NP_565286), 4-дифосфоцитидил-2-C-метил-D-эритритолкиназу (EC: 2.7.1.148; пример 1, IspE номер доступа NP_415726; пример 2, AT2G26930 номер доступа NP_1802 61), 2-C-метил-D-эритритол-2,4-cуциклодифосфатсинтетазу (EC: 4.6.1,12; пример 1, IspF номер доступа NP_41722 6; пример 2, AT1G63970 номер доступа NP_564819), 1-гидрокси-2-метил-2-(E)-бутенил-4-дифосфатсинтетазу (EC: 1,17.7.1; пример 1, IspG номер доступа NP_417010; пример 2, AT5G60600 номер доступа NP_001119467) и 4-гидрокси-3-метил-2-бутенилдифосфатредуктазу (EC: 1.17.1.2; пример 1, IspH номер доступа NP_414570; пример 2, AT4G34350 номер доступа NP_567965).
[0038] Введение содержащего ген экспрессирующего вектора хозяину (трансформацию) можно проводить с применением известных способов. Примеры такого способа могут включать способ компетентных клеток с использованием микробной клетки, обработанной кальцием, и способ электропорации. Ген можно вводить путем инфекции микробной клетки с использованием фагового вектора, а не плазмидного вектора.
[0039] Кроме того, в трансформант по настоящему изобретению также можно вводить ген, кодирующий фермент, участвующий в мевалонатном пути или метилэритриолфосфатном пути, который синтезирует диметилаллилдифосфат, который является субстратом изопренсинтазы.
Такой фермент может включать 1-дезокси-D-ксилоза-5-фосфатсинтетазу, которая преобразует пируват и D-глицеральдегид-3-фосфат в 1-дезокси-D-ксилоза-5-фосфат, и изопентилдифосфатизомеразу, которая преобразует изопентенилдифосфат в диметилаллилдифосфат.
[0040] Белок по настоящему изобретению можно экстрагировать или выделять из трансформанта по настоящему изобретению, и изопрен можно получать культивированием трансформанта, который экспрессирует белок по настоящему изобретению.
[0041] Способы получения мономера изопрена и изопренового полимера
Настоящее изобретение относится к способу получения мономера изопрена. Способ получения мономера изопрена по настоящему изобретению включает получение мономера изопрена из диметилаллилдифосфата в присутствии белка по настоящему изобретению.
[0042] Способ получения мономера изопрена по настоящему изобретению конкретно не ограничен, при условии, что способ проводят в присутствии белка по настоящему изобретению, и его можно проводить с применением системы ферментативных реакций с самим белком (например, очищенным белком) по настоящему изобретению или культивированием трансформанта по настоящему изобретению, который продуцирует белок по настоящему изобретению. Предпочтительно его проводят культивированием трансформанта по настоящему изобретению. Когда трансформант по настоящему изобретению используют в способе получения мономера изопрена по настоящему изобретению, диметилаллилдифосфат, который является сырьем мономера изопрена, эффективно поставляется из источника углерода в среду для культивирования трансформантом по настоящему изобретению. Трансформант по настоящему изобретению продуцирует мономер изопрена в основном в результате дегазации источника углерода в среде для культивирования. Таким образом, мономер изопрена собирают путем сбора газа, продуцируемого трансформантом. Диметилаллилдифосфат, который является субстратом изопренсинтазы, синтезируется из источника углерода в среду для культивирования мевалонатным путем или метилэритриолфосфатным путем в хозяине.
Культуральная среда для культивирования трансформанта по настоящему изобретению предпочтительно содержит источник углерода, который преобразуется в изопрен. Источник углерода может содержать углеводы, такие как моносахариды, дисахариды, олигосахариды и полисахариды; инвертированные сахара, полученные гидролизом сахарозы; глицерин; соединения, содержащие один атом углерода (далее в настоящем описании, обозначаемые как C1-соединение), такие как метанол, формальдегид, формиат, моноксид углерода и диоксид углерода; масла, такие как кукурузное масло, пальмовое масло и соевое масло; ацетат; животные жиры; животные масла; жирные кислоты, такие как насыщенные жирные кислоты и ненасыщенные жирные кислоты; липиды; фосфолипиды; глицеролипиды; сложные эфиры глицерина и жирных кислот, такие как моноглицерид, диглицерид и триглицерид; полипептиды, такие как белки микроорганизмов и белки растений; возобновляемые источники углерода, такие как источники углерода на основе гидролизованной биомассы; дрожжевые экстракты или их комбинации. Для источника азота можно использовать неорганические аммонийные соли, такие как сульфат аммония, хлорид аммония и фосфат аммония, органический азот, такой как гидролизованные соевые бобы, аммиачный газ, аммиачная вода и т.п. Желательно, чтобы содержались необходимые вещества, такие как витамин B1 и L-гомосерин или дрожжевой экстракт и т.п., в соответствующем количестве в качестве источника органических микроэлементов. В дополнение к перечисленному, при необходимости, в небольшом количестве добавляют фосфат калия, сульфат магния, ионы железа, ионы марганца и т.п. Среда для культивирования, используемая в настоящем изобретении, может представлять собой природную среду или синтетическую среду, при условии, что она содержит источник углерода, источник азота, неорганические ионы и необязательно другие органические микроингредиенты.
[0043] Примеры моносахаридов могут включать триозу, такую как кетотриоза (дигидроксиацетон) и альдотриозу (глицеральдегид); тетрозу, такую как кетотетроза (эритрулоза) и альдотетроза (эритроза, треоза); пентозу, такую как кетопентоза (рибулоза, ксилулоза), альдопентоза (рибоза, арабиноза, ксилоза, ликсоза) и дезоксисахарид (дезоксирибоза); гексозу, такую как кетогексоза (фикоза, фруктоза, сорбоза, тагатоза), альдогексоза (аллоза, альтроза, глюкоза, манноза, гулоза, идоза, галактоза, таллоза) и дезоксисахарид (фукоза, фукроза, рамноза), и гептозу, такую как седогептулоза. Предпочтительными являются C6-сахара, такие как фруктоза, манноза, галактоза и глюкоза, и C5-сахара, такие как ксилоза и арабиноза.
Примеры дисахаридов могут включать сахарозу, лактозу, мальтозу, трегалозу, туранозу и целлобиозу. Предпочтительными являются сахароза и лактоза.
Примеры олигосахаридов могут включать трисахариды, такие как рафиноза, мелицитоза и мальтотриоза, тетрасахариды, такие как акарбоза и стахиоза, и другие олигосахариды, такие как фруктоолигосахарид (FOS), галактоолигосахарид (GOS) и маннанолигосахарид (MOS).
Примеры полисахаридов могут включать гликоген, крахмал (амилозу, амилопектин), целлюлозу, декстрин и глюкан (β1,3-глюкан), и предпочтительными являются крахмал и целлюлоза.
[0044] Примеры белка микроорганизмов могут включать полипептиды, полученные из дрожжей или бактерий.
Примеры белка растений могут включать полипептиды, полученные из соевых бобов, кукурузы, канолы, ятрофы, пальмы, арахиса, подсолнечника, кокосового ореха, горчицы, семян хлопчатника, косточкового пальмового масла, маслины, сафлора, кунжута и семян льна.
[0045] Примеры липидов могут включать вещества, содержащие одну или более насыщенных или ненасыщенных жирных кислот C4 или более.
[0046] Предпочтительно масло представляет собой липид, который содержит одну или более насыщенных или ненасыщенных жирных кислот C4 или более, и является жидким при комнатной температуре, и примеры масла могут включать липиды, полученные из соевых бобов, кукурузы, канолы, ятрофы, пальмы, арахиса, подсолнечника, кокосового ореха, горчицы, семян хлопчатника, косточкового пальмового масла, маслины, сафлора, кунжута, семян льна, маслосодержащих микроорганизмов, китайского сального дерева и комбинации двух или более перечисленных источников.
[0047] Примеры жирной кислоты могут включать соединения, представленные формулой RCOOH ("R" представляет собой углеводородную группу).
Ненасыщенная жирная кислота представляет собой соединение, содержащее по меньшей мере одну двойную связь между двумя атомами углерода в "R", и примеры ненасыщенной жирной кислоты могут включать олеиновую кислоту, вакценовую кислоту, линолевую кислоту, пальмитэлаидиновую кислоту и арахидоновую кислоту.
Насыщенная жирная кислота представляет собой соединение, где "R" представляет собой насыщенную алифатическую группу, и примеры насыщенной жирной кислоты могут включать докозановую кислоту, эйкозановую кислоту, октадекановую кислоту, гексадекановую кислоту, тетрадекановую кислоту и додекановую кислоту.
Среди них в качестве жирной кислоты предпочтительными являются кислоты, содержащие одну или более C2-C22 жирных кислот, и наиболее предпочтительными являются кислоты, содержащие C12-жирную кислоту, C14-жирную кислоту, C16-жирную кислоту, C18-жирную кислоту, C20-жирную кислоту и C22-жирную кислоту.
Источник углерода могут включать соли и производные этих жирных кислот и соли таких производных. Примеры соли могут включать литиевые соли, калиевые соли и натриевые соли.
[0048] Примеры источника углерода также могут включать комбинации углевода, такого как глюкоза, с липидом, маслом, жирами, жирной кислотой и сложным эфиром жирной кислоты и глицерина.
[0049] Примеры возобновляемого источника углерода могут включать источники углерода на основе гидролизованной биомассы.
Примеры источника углерода на основе биомассы могут включать субстраты на основе целлюлозы, такие как отходы производства лесоматериалов, сорта бумаги и целлюлозы, лиственные растения и мякоть плода, и части растений, такие как стебли, частицы зерна, корни и клубни.
Примеры растений, которые следует использовать в качестве источника углерода на основе биомассы, могут включать кукурузу, пшеницу, рожь, сорго, тритикале, рис, просо, ячмень, маниок, бобовые, такие как горох, картофель, сладкий картофель, бананы, сахарный тростник и тапиока.
[0050] Когда к среде для культивирования добавляют возобновляемый источник углерода, такой как биомасса, источник углерода предпочтительно подвергают предварительной обработке. Примеры предварительной обработки могут включать ферментативную предварительную обработку, химическую предварительную обработку и сочетание ферментативной предварительной обработки и химической предварительной обработки.
Предпочтительно возобновляемый источник углерода подвергают полному или частичному гидролизу перед добавлением в среду для культивирования.
[0051] Примеры источника углерода также могут включать дрожжевой экстракт и комбинацию дрожжевого экстракта с другим источником углерода, таким как глюкоза. Комбинация дрожжевого экстракта с C1-соединением, таким как диоксид углерода и метанол, является предпочтительным.
[0052] В способе культивирования трансформанта по настоящему изобретению предпочтительно культивировать клетку в стандартной среде, содержащей физиологический раствор и питательные вещества.
Среда для культивирования не является конкретно ограниченной, и примеры среды для культивирования могут включать готовые обычные среды, которые являются коммерчески доступными, такие как бульон Луриа-Бертани (LB), декстрозный бульон Сабуро (SD) и бульон на основе дрожжевой среды (YM). Для использования можно выбирать соответствующим образом среду, подходящую для культивирования конкретного хозяина.
[0053] Желательно, чтобы она содержала подходящие минералы, соли, вспомогательные элементы, буферы и ингредиенты, известные специалистам в данной области, чтобы являться подходящей для культивирования и чтобы облегчать продукцию изопрена в дополнение к подходящему источнику углерода в среде для клеток.
Для сохранения экспрессии белка по настоящему изобретению в трансформанте по настоящему изобретению предпочтительно к среде добавлять сахар, соль металла, противомикробное вещество и т.п.
[0054] Условия культивирования трансформанта по настоящему изобретению не являются конкретно ограниченными, при условии, что может экспрессироваться белок по настоящему изобретению, и можно использовать стандартные условия культивирования клеток.
Температура культуры предпочтительно составляет 20-37°C, композиция газа предпочтительно приблизительно составляет 6-84% концентрации CO2, и pH предпочтительно составляет приблизительно от 5 до приблизительно 9.
Трансформант предпочтительно культивируют в аэробных, не содержащих кислород или анаэробных условиях в зависимости от природы хозяина.
[0055] Примеры способа культивирования трансформанта могут включать способ с применением известного способа ферментации, такого как способ периодического культивирования, способ культивирования с подпиткой или способ непрерывного культивирования.
В способе периодического культивирования композицию среды добавляют при начале ферментации и инокулируют трансформант в композицию среды, и культивируют, регулируя pH и концентрацию кислорода.
При культивировании трансформанта способом периодического культивирования рост трансформанта начинается с умеренной индукционной фазы, проходит через логарифмическую фазу роста и, наконец, переходит в стационарную фазу, при которой скорость роста снижается или останавливается. Трансформант продуцирует изопрен в логарифмической фазе роста и стационарной фазе.
[0056] В способе культивирования с подпиткой в дополнение к указанному выше периодическому способу постадийно добавляют источник углерода в зависимости от протекания процесса ферментации. Способ культивирования с подпиткой является эффективным, когда количество источника углерода должно быть ограничено в среде, т.к. метаболизм трансформанта, как правило, снижается вследствие подавления продуктами катаболизма. Культивирование с подпиткой можно проводить с использованием ограниченного количества или избыточного количества источника углерода, такого как глюкоза.
[0057] В способе непрерывного культивирования постоянно привносят определенное количество среды в биореактор при постоянной скорости, при этом удаляют такое же количество среды. В способе непрерывного культивирования культуру можно постоянно поддерживать в высокой концентрации, и трансформант в среде для культивирования, как правило, находится в фазе логарифмического роста.
Подпитку можно дополнять полной или частичной заменой среды соответствующим образом, и можно предотвращать накопление побочных продуктов метаболизма, которые потенциально оказывают неблагоприятное действие на рост трансформанта и накопление погибших клеток.
[0058] Промотор, содержащийся в экспрессирующем векторе по настоящему изобретению, может включать конститутивные промоторы и индуцибельные промоторы. Когда экспрессирующий вектор по настоящему изобретению содержит индуцибельный промотор, такой как промотор lac, экспрессию белка по настоящему изобретению можно индуцировать добавлением IPTG (изопропил-β-тиогалактопиранозида) в среду для культивирования.
[0059] Примеры способа оценки количества изопрена, полученного культивированием трансформанта по настоящему изобретению, могут включать способ, в котором газовую фазу собирают парофазным способом и анализируют такую газовую фазу газовой хроматографией.
Более подробно мономер изопрена в свободном пространстве, который получают культивированием трансформанта в герметично запечатанном флаконе при перемешивании среды для культивирования, анализируют стандартной газовой хроматографией. Затем площадь, которую вычисляют по кривой, измеряемой посредством газовой хроматографии, преобразовывают в количество мономера изопрена, продуцируемого трансформантом, с использованием стандартной кривой.
[0060] Примеры способа сбора мономера изопрена, полученного культивированием трансформанта по настоящему изобретению, могут включать очистку газа, фракционную перегонку или диссоциацию мономер изопрена, адсорбированного на твердой фазе нагреванием или вакуумом, или экстрагирование растворителем.
При очистке газа газообразный изопрен непрерывно удаляют из выделяющегося газа. Такое удаление газообразного изопрена можно проводить различными способами. Примеры удаления могут включать адсорбцию на твердой фазе, разделение на жидкую фазу и способ, в котором непосредственно конденсируют газообразный изопрен.
[0061] Мономер изопрена можно собирать в одну стадию или в ходе многих стадий. Когда мономер изопрена собирают в одну стадию, мономер изопрена переводят в жидкую фазу одновременно с выделением мономера изопрена из выходящего газа. Мономер изопрена также можно непосредственно конденсировать из газа с получением жидкой фазы. Когда мономер изопрена собирают в ходе многих стадий, мономер изопрена выделяют из выделяющегося газа, и затем переводят в жидкую фазу. Например, мономер изопрена адсорбируют на твердой фазе и экстрагируют из твердой фазы растворителем.
[0062] Способ сбора мономера изопрена может дополнительно включать очистку мономера изопрена. Примеры очистки могут включать выделение из экстракта жидкой фазы перегонкой и различными хроматографическими методами.
[0063] Белок по настоящему изобретению обладает более высокой способностью продуцировать изопрен по сравнению с общепринятой изопренсинтазой. Таким образом, мономер изопрена можно эффективно получать с использованием трансформанта, который экспрессирует белок по настоящему изобретению.
[0064] Настоящее изобретение также относится к способу получения изопренового полимера. Способ получения изопренового полимера по настоящему изобретению включает следующие стадии (I) и (II):
(I) получение мономера изопрена способом по настоящему изобретению и
(II) полимеризацию мономера изопрена с образованием изопренового полимера.
[0065] Стадию (I) можно проводить таким же образом, как в способе получения мономера изопрена по настоящему изобретению, описанному выше. Полимеризацию мономер изопрена на стадии (II) можно проводить любым известным в данной области способом.
ПРИМЕРЫ
[0066] Далее в настоящем описании настоящее изобретение более подробно описано со ссылкой на примеры, но настоящее изобретение не ограничено следующими ниже примерами.
[0067] Пример 1: Оценка способности продукции изопрена в растениях
1-1) Измерение количества изопрена, образовавшегося на единицу массы сухих листьев
Сначала измеряли количество изопрена, образовавшегося на 1 г сухих листьев в растении для оценки способности продукции изопрена в растениях. В качестве растений использовали мукуну (Mucuna bracteata), плакучую иву (Salix babylonica), ликвидамбар смолоносный (Liquidambar styraciflua), мирт (Myrtus communis) и пуэрарию волосистую (Pueraria lobata).
При измерении количества образовавшегося изопрена эксикатор с возможностью замены газа (товарное наименование: Vacuum Desiccator, выпускаемый AS ONE Corporation) помещали в инкубатор (товарное наименование: Growth Chamber MLR-351H, выпускаемый SANYO), и инкубатор устанавливали в режиме условий индукции высокой температуры (освещенность 100 пмоль E/м2/с при 40°C), при этом вентилятор для перемешивания газа, который помещался в эксикаторе с возможностью замены газа, приводили в движение для перемешивания атмосферы в пространстве эксикатора с возможностью замены газа. После того, как температура атмосферы в эксикаторе с возможностью замены газа достигала 40°C, растительный организм мукуну, посаженную в ящик, содержали в нем и поддерживали в течение 3 часов в состоянии, при котором эксикатор с возможностью замены газа являлся герметически закрытым. Затем газовый компонент, выделяемый мукуной, отсасывали из пространства в эксикаторе с возможностью замены газа насосом для отсасывания через силиконовую трубку, адсорбционную пробирку и пробирку для сбора газа. Таким образом, водяные пары (влажность), содержащиеся в газовом компоненте, выделяемом мукуной, адсорбировали и выделяли в адсорбционной пробирке, газовый компонент, из которого выделяли водяные пары, подавали в пробирку для сбора газа и собирали газовый компонент в пробирке для сбора газа. Затем изопрен, содержащийся в газовом компоненте, собранный в пробирку для сбора газа, подвергали количественному анализу с использованием газового хроматографа (товарное наименование: GC-FID6890, выпускаемого Agilent).
Для определения массы сухих листьев для площади листа свежего индивидуального листа и сухой массы, когда свежий индивидуальный лист сушат сушилкой при 80°C в течение 8 часов, устанавливали очень хорошую положительную корреляцию. Таким образом, получали формулу для перехода от площади листа к сухой массе и определяли сухую массу из площади целого листа от растительного организма мукуны, используемой для определения количества образовавшегося изопрена.
Количество образовавшегося изопрена на 1 г сухой листвы получали делением количества образовавшегося изопрена от целого растительного организма мукуны на установленную массу целого растительного организма.
В результате было продемонстрировано, что мукуна являлась превосходной в отношении количества образовавшегося изопрена на единицу масса сухой листвы (фиг. 1).
1-2) Измерение количества образовавшегося изопрена на количество общего белка
Затем измеряли количество образовавшегося изопрена на количество общего белка, экстрагированного из листьев различных растений. В качестве растений использовали мукуну (образцы 1 и 2), плакучую иву, ликвидамбар смолоносный, мирт и пуэрарию волосистую.
Для экстракции белка получали буферный раствор (50 мМ Tris-HCl, 20 мМ MgCl, 5% глицерин, 0,02% Triton-X100, pH 8,0) и непосредственно перед использованием добавляли 10% Polyclar AT, 20 мМ DTT, полную таблетку протеазы (одна таблетка/50 мл) и 1 мМ бензамидина HCl (конечные концентрации для каждого), и использовали в качестве буфера для экстракции белка. 50 мл буфера для экстракции белка добавляли к 5 г образца, затем смесь хорошо измельчали в холодной ступке на льду и фильтровали через сложенный вдвое внахлест материал Miracloth. Фильтрат центрифугировали при 12000×g в течение 20 минут и 40000×g в течение 40 минут, с получением супернатанта и использовали супернатант в виде неочищенного экстракта.
Затем этот неочищенный экстракт фракционировали сульфатом аммония. Белки, выпадавшие в осадок в диапазоне от 40% до 55% конечных концентраций сульфата аммония, центрифугировали при 40000×g в течение 40 минут, и полученный осадок повторно растворяли в буфере для экстракции белка, с получением фракции сульфата аммония.
Массу общего (фракция сульфата аммония) белка рассчитывали посредством измерения фракции сульфата аммония с применением анализа по Брэдфорду. Реагент Брэдфорда подвергали взаимодействию со стандартным белком, бычьим сывороточным альбумином, и измеряли оптическая плотность при длине волны 595 нм с использованием спектрофотометра. Стандартную кривую для белка строили с использованием полученных значений оптической плотности. Также измеряли оптическую плотность при длине волны 595 нм во фракции сульфата аммония, разбавленной до 50 раз, и определяли количество общего (фракция сульфата аммония) белка из стандартной кривой для стандартного белка.
При измерении количества образовавшегося изопрена 100 мкл неочищенного экстракта или 100 мкл неочищенного раствора фермента, кипящего при 100°C, помещали в 4 мл стеклянный флакон, и затем добавляли в него 2 мкл 0,5M раствора MgCl2 и 5 мкл 0,2M раствора DMAPP. Флакон плотно закрывали завинчивающейся крышкой с диафрагмой, и затем флакон аккуратно перемешали вихревым способом и помешали в инкубатор при 40°C. Через 0,5, 1 и 2 часа отбирали образец от 0,5 до 2 мл газового слоя в свободном пространстве над продуктом посредством газонепроницаемого шприца и измеряли количество образовавшегося изопрена с использованием газового хроматографа (товарное наименование: GC-FID6890, выпускаемого Agilent). Количество образовавшегося изопрена рассчитывали с использованием неочищенного фермента через 0,5, 1 и 2 часа путем вычитания измеряемого значения в случае использования раствора неочищенного фермента, кипящего при 100°C, из измеряемого значения в случае использования неочищенного фермента. Ферментативная активность на 1 мг общего белка (специфическая активность) рассчитывали из количества образовавшегося изопрена за один час. Количество образовавшегося изопрена измеряли, сохраняя постоянным количество DMAPP, которое являлось субстратом изопренсинтазы.
В результате было продемонстрировано, что мукуна являлась превосходной в отношении количества образовавшегося изопрена на количество общего белка (фиг.2, таблица 1). Как описано выше, показано, что мукуна являлась превосходной в отношении способности продуцировать изопрен.
[0069]
Количество образовавшегося изопрена на количество общего белка (числовые показатели относительно случая пуэрарии волосистой)
[0070] Пример 2: Клонирование гена изопренсинтазы, полученного из мукуны
2-1) Оценка времени отбора образца
Отбирали образец газообразного изопрена, выделяемого из листьев мукуны, облучаемой светом в течение 1, 2, 3 и 5 часов при температуре 40°C, и определяли количество продуцируемого изопрена газовой хроматографией, описанной ниже, и подтверждали продукцию 4, 8, 12 и 10 мкг изопрена/г листвы DW. Таким образом, подтверждали, что оптимальное облучение светом составляло 3 часа.
[0071] 2-2) Экстракция лизирующим раствором для общей РНК
Общую РНК экстрагировали из листьев мукуны лизирующим раствором для общей РНК следующими ниже способами.
(1) Отбирали образец листьев мукуны, облучаемой светом в течение 3 часов при температуре 40°C.
(2) 100 мг ткани листа растирали в ступке, быстро замораживая ткань листа жидким азотом, затем ткань листа с жидким азотом диспергировали в не содержащей РНК 2 мл пробирке Eppendorf и переводили в газообразное состояние жидкий азот.
(3) К пробирке Eppendorf добавляли 450 мкл растворяющего буфера RLT (содержащего 2-меркаптоэтанола), прилагаемого к набору RNeasy Plant (выпускаемого Qiagen) и интенсивно перемешивали с использованием мешалки Vortex с получением лизата ткани листа.
(4) Лизат ткани листа наносили на центрифужную колонку QIAshredder, прилагаемую к набору RNeasy Plant, и центрифугировали при 15000 об/мин в течение 2 минут.
(5) Только супернатант элюата колонки переносили в новую не содержащую РНК 2 мл пробирку Eppendorf, затем к супернатанту добавляли половину объема супернатанта высокой степени очистки этанола и перемешивали полученный раствор пипетированием с получением приблизительно 650 мкл раствора.
(6) Раствор наносили на центрифужную колонку RNeasy, прилагаемую к набору RNeasy Plant, центрифугировали при 10000 об/мин в течение 15 секунд и удаляли фильтрат.
(7) В центрифужную колонку RNeasy добавляли 700 мкл буфера RW1, прилагаемого к набору RNeasy Plant, центрифугировали при 10000 об/мин в течение 15 секунд и удаляли фильтрат.
(8) В центрифужную колонку RNeasy добавляли 500 мкл буфера BPE, прилагаемого к набору RNeasy Plant, центрифугировали при 10000 об/мин в течение 15 секунд и удаляли фильтрат.
(9) В центрифужную колонку RNeasy снова добавляли 500 мкл буфера BPE, центрифугировали при 10000 об/мин в течение 2 минут и удаляли фильтрат.
(10) Центрифужную колонку RNeasy помещали в 2 мл пробирку для сбора, прилагаемую к набору RNeasy Plant, центрифугировали при 15000 об/мин в течение одной минуты и удаляли фильтрат.
(11) Центрифужную колонку RNeasy помещали в 1,5 мл пробирку для сбора, прилагаемую к набору RNeasy Plant.
(12) Не содержащую РНК дистиллированную воду, прилагаемую к набору RNeasy Plant, непосредственно добавляли на мембрану центрифужной колонки RNeasy с использованием Pipetman, центрифугировали при 10000 об/мин в течение одной минуты и собирали общую РНК. Этот этап повторяли дважды с получением приблизительно 100 мкг общей РНК.
[0072] 2-3) Анализ нуклеотидной последовательности гена изопренсинтазы, полученного из мукуны
Качество РНК в растворе экстрагируемой общей РНК проверяли с использованием наночипов для РНК, предоставляемых для BioAnalyzer (Agilent Technologies, Inc.), и подтверждали, что раствор не загрязнен геномной ДНК, и РНК не разрушалась в растворе.
Общую РНК преобразовывали в двухцепочечную с использованием обратной транскриптазы, и затем фрагментировали с использованием распылителя. Нуклеотидные последовательности 198179 фрагментов, содержащих последовательность поли-A на 3'-конце анализировали с использованием высокопроизводительного секвенатора 454 titanium FLX (выпускаемого Roche Applied Science). Выравнивали перекрывающиеся последовательности в полученных последовательностях фрагментов с получением 13485 последовательностей контига. Проводили поиск BLAST для этих последовательностей контига, и экстрагировали 6 последовательностей контига, обладающих гомологией (идентичностью нуклеотидных последовательностей) с регистрируемыми и известными последовательностями гена изопренсинтазы из пуэрарии волосистой и тополя. Последовательности дополнительно более подробно анализировали и было выявлено, что 3 последовательности из 6 последовательностей контига получают из одного и того же гена. Таким образом, получали частичную последовательность гена изопренсинтазы, полученного из мукуны. Проводили 5'-RACE на основании этой частичной последовательности с получением полноразмерной нуклеотидной последовательности гена изопренсинтазы, полученного из мукуны, который был представлен SEQ ID NO: 1.
[0073] Пример 3: Получение плазмиды для экспрессии изопренсинтазы, полученной из различных растений
3-1) Химический синтез изопренсинтазы, полученной из Pueraria montana var. lobata (пуэрарии волосистой)
Нуклеотидная последовательность и аминокислотная последовательность изопренсинтазы, полученной из Pueraria montana var. lobata, уже известны (доступ: AAQ84170: изопренсинтаза (IspS) P. montana var. lobata). Аминокислотная последовательность белка IspS, полученного из P. montana, и нуклеотидная последовательность его гена представлены SEQ ID NO: 3 и SEQ ID NO: 4, соответственно. Ген IspS оптимизировали по частоте использования кодона в E. coli для эффективной экспрессии гена IspS в E. coli, и в дальнейшем конструировали для отсечения сигнала локализации хлоропласта. Сконструированный ген обозначали как IspSK. Нуклеотидная последовательность IspSK представлена SEQ ID NO: 5. Ген IspSK химически синтезировали, затем клонировали в pUC57 (выпускаемой GenScript), и полученную плазмиду обозначали как pUC5-IspSK.
[0074] 3-2) Химический синтез изопренсинтазы, полученной из Populus alba × Populus tremula (тополя)
Нуклеотидная последовательность и аминокислотная последовательность изопренсинтазы, полученной из P. alba × P. tremula, уже известны (доступ: CAC35696: изопренсинтаза P. alba × P. tremula (тополя)). Аминокислотная последовательность белка IspS, полученного из P. alba × P. tremula, и нуклеотидная последовательность его гена представлены SEQ ID NO: 6 и SEQ ID NO: 7, соответственно. Конструировали ген IspS, который оптимизировали по частоте использования кодона в E. coli аналогичным способом, как указано выше, и в котором отсекали сигнал локализации хлоропласта и обозначали как IspSP. Нуклеотидная последовательность IspSP представлена SEQ ID NO:8. Ген IspSP химически синтезировали, затем клонировали в pUC57 (выпускаемую GenScript), и полученную плазмиду обозначали как pUC57-IspSP.
[0075] 3-3) Химический синтез изопренсинтазы, полученной из Mucuna pruriens (мукуны)
На основе нуклеотидной последовательности, полученной из M. pruriens, конструировали ген IspS, в котором оптимизировали частоту использования кодона в E. coli аналогичным способом, как указано выше. Изопренсинтазу, в которой обеспечивали сигнал локализации хлоропласта, обозначали как IspSM (L), и изопренсинтазу, в которой отсекали сигнал локализации хлоропласта, обозначали как IspSM. Нуклеотидные последовательности для IspSM (L) и IspSM представлены SEQ ID NO: 9 и SEQ ID NO: 10, соответственно. Ген IspSM и ген IspSM (L) химически синтезировали, и затем клонировали в pUC57 (выпускаемую GenScript), и полученные плазмиды обозначали как pUC57-IspSM и pUC57-IspSM (L).
[0076] 3-4) Конструкция плазмиды для экспрессии pSTV28-Ptac-Ttrp
Конструировали плазмиду для экспрессии pSTV28-Ptac-Ttrp для экспрессии IspS, полученной из различных растений, в E. coli. Сначала химически синтезировали фрагмент ДНК, содержащий область промотора tac (синоним: Ptac) (deBoer et al., (1983) Proc. Natl. Acad. Sci. USA, 80, 21-25) и концевую область оперона триптофана (синоним: Ttrp), полученную из E. coli (Wu et al., (1978) Proc. Natl. Acad. Sci. USA, 75, 442-5446) и содержащую участок KpnI на 5'-конце и участок BamHI на 3'-конце (нуклеотидная последовательность Ptac-Ttrp представлена SEQ ID NO:11). Полученный фрагмент ДНК Ptac-Ttrp расщепляли KpnI и BamHI и лигировали к pSTV28 (выпускаемой Takara Bio Inc.) аналогично расщепляемыми KpnI и BamHI реакцией лигирования с ДНК-лигазой. Полученную плазмиду обозначали как pSTV28-Ptac-Ttrp (ее нуклеотидная последовательность представлена SEQ ID NO: 12). Такая плазмида может увеличивать экспрессию гена IspS в результате клонирования гена IspS после Ptac.
[0077] 3-5) Конструкция плазмиды для экспрессии гена IspS, полученного из различных растений
Плазмиды для экспрессии гена IspSK, гена IspSP, гена IspSM и гена IspSM (L) in E. coli конструировали следующим способом. ПЦР проводили с полимеразой Prime Star (выпускаемой Takara Bio Inc.) с использованием синтезированных олигонуклеотидов, состоящих из нуклеотидных последовательностей, представленных SEQ ID NO: 13 и 14, в качестве праймеров с pUC57-IspSK в качестве матрицы, синтезированных олигонуклеотидов, состоящих из нуклеотидных последовательностей, представленных SEQ ID NO: 15 и 16, в качестве праймеров с pUC57-IspSP в качестве матрицы, синтезированных олигонуклеотидов, состоящих из нуклеотидных последовательностей, представленных SEQ ID NO: 17 и 18, в качестве праймеров с pUC57-IspSM в качестве матрицы или дополнительно синтезированных олигонуклеотидов, состоящих из нуклеотидных последовательностей, представленных SEQ ID NO: 19 и 20, в качестве праймеров с pUC57-IspSM (L) в качестве матрицы. Реакционный раствор получали в соответствии с рецептурой, прилагаемой к набору, и проводили реакцию при 98°C в течение 10 секунд, при 54°C в течение 20 секунд и при 68°C в течение 120 секунд в течение 40 циклов. В результате получали продукт ПЦР, содержащий ген IspSK, ген IspSP, ген IspSM или ген IspSM(L). Аналогично, проводили ПЦР с полимеразой Prime Star (выпускаемой Takara Bio Inc.) с использованием синтезированных олигонуклеотидов, состоящих из нуклеотидных последовательностей, представленных SEQ ID NO: 21 и 22, в качестве праймеров с pSTV28-Ptac-Ttrp в качестве матрицы. Реакционный раствор получали в соответствии с рецептурой, прилагаемой к набору, и проводили реакцию при 98°C в течение 10 секунд, при 54°C в течение 20 секунд и при 68°C в течение 210 секунд в течение 40 циклов. В результате получали продукт ПЦР, содержащий pSTV28-Ptac-Ttrp. Затем очищенные фрагменты гена IspSK, гена IspSP, гена IspSM и гена IspSM(L) лигировали с продуктом ПЦР для pSTV28-Ptac-Ttrp с использованием набора для клонирования In-Fusion HD (выпускаемого Clontech). Полученные плазмиды для экспрессии гена IspSK, гена IspSP, гена IspSM и гена IspSM(L) обозначали как pSTV28-Ptac-IspSK, pSTV28-Ptac-IspSP, pSTV28-Ptac-IspSM и pSTV28-Ptac-IspSM (L), соответственно.
[0078]
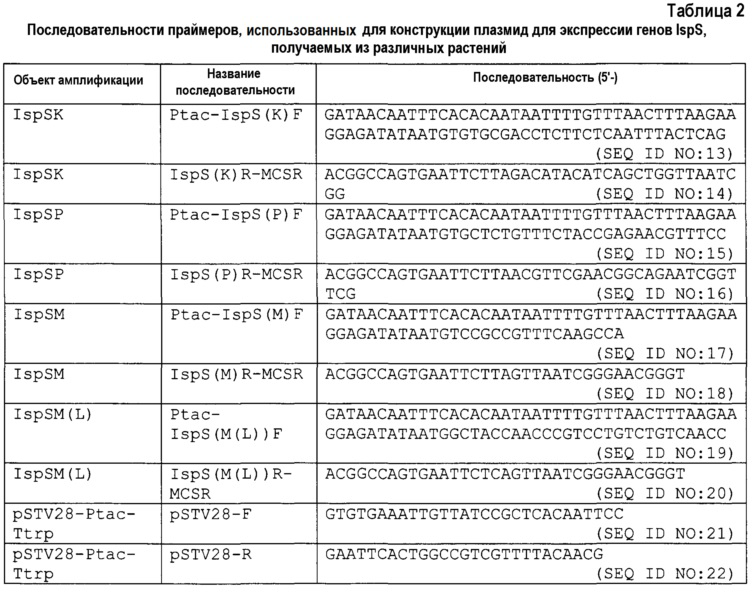
[0079] Пример 4: Измерение ферментативной активности изопренсинтазы, полученной из различных растений, с использованием неочищенного фермента, полученного из E. coli
4-1) Конструкция штамма MG1655 E. coli, обладающего способностью продуцировать изопрен
Получали компетентные клетки штамма MG1655 E. coli (ATCC 700926), и затем вводили в них pSTV28-Ptac-Ttrp, pSTV28-Ptac-IspSK, pSTV28-Ptac-IspSP, pSTV28-Ptac-IspSM или дополнительно pSTV28-Ptac-IspSM (L). Суспензию клеток равномерно наносили на чашку Петки с LB, содержащий 60 мг/л хлорамфеникола, и культивировали при 37°C в течение 18 часов. Затем из формированной чашки Петри получали трансформанты, которые являлись устойчивыми к хлорамфениколу. Штамм, в котором pSTV28-Ptac-Ttrp, pSTV28-Ptac-IspSK, pSTV28-Ptac-IspSP, pSTV28-Ptac-IspSM или дополнительно pSTV28-Ptac-IspSM (L) вводили в штамм MG1655 E. coli, обозначали как штамм MG1655/pSTV28-Ptac-Ttrp, MG1655/pSTV28-Ptac-IspSK, MG1655/pSTV28-Ptac-IspSP, MG1655/pSTV28-Ptac-IspSM или дополнительно MG1655/pSTV28-Ptac-IspSM (L), соответственно.
[0080] 4-2) Способ получения экстракта неочищенного фермента
Микробные клетки штамма MG1655/pSTV28-Ptac-Ttrp, MG1655/pSTV28-Ptac-IspSK, MG1655/pSTV2 8-Ptac-IspSP, MG1655/pSTV28-Ptac-IspSM или MG1655/pSTV28-Ptac-IspSM (L) равномерно наносили на чашку Петри с LB, содержащий 60 мг/л хлорамфеникола, и культивировали при 37°C в течение 18 часов. Микробные клетки, соответствующие 1/6 формированной чашки Петри, вносили в колбу Сакагути, в которую добавляли 20 мл LB, содержащего 60 мг/л хлорамфеникола, и культивировали при 37°C в течение 6 часов. Микробные клетки из среды для культивирования центрифугировали при 5000 об/мин при 4°C в течение 5 минут и дважды промывали ледяным буфером для изопренсинтазы (50 мМ Tris-HCl, pH 8,0, 20 мМ MgCl2, 5% глицерин). Промытые микробные клетки суспендировали в 1,8 мл того же буфера. Приблизительно 0,9 мл гранул для дезинтеграции (YBG01, диаметр 0,1 мм) и 0,9 мл суспензии микробных клеток помещали в 2 мл пробирку, специальную для проведения шока с использованием множества гранул, и подвергали микробные клетки дезинтеграции с использованием устройства для проведения шока с использованием множества гранул, выпускаемого Yasui Kikai Corporation, при 2500 об/мин при 4°C в течение 3 циклов "Включено" в течение 30 секунд/"Выключено" в течение 30 секунд. После дезинтеграции пробирку центрифугировали при 20000×g при 4°C в течение 20 минут и использовали супернатант в качестве экстракта неочищенного фермента.
4-3) Измерение активности изопренсинтазы
Экстракт неочищенного фермента от штамма MG1655/pSTV28-Ptac-Ttrp, MG1655/pSTV28-Ptac-IspSK, MG1655/pSTV2 8-Ptac-IspSP, MG1655/pSTV28-Ptac-IspSM, или MG1655/pSTV28-Ptac-IspSM (L) (содержащий 2 мг как количество общего белка) совместно с буфером на основе изопрена в общем объеме 0,5 мл помещали во флакон со свободным объемом (22 мл CLEAR CRIMP TOP VIAL (каталожный № B0104236), выпускаемый Perkin Elmer), затем к нему добавляли 0,025 мл 0,5М раствора MgCl2 и 0,01 мл 0,2М раствора DMAPP (выпускаемого Cayman, каталожный № 63180), и смесь аккуратно перемешивали смесь вихревым способом. Затем сразу же флакон плотно закрывали крышкой с бутилкаучуковой мембраной для флакона со свободным объемом (CRIMPS (каталожный № B0104240), выпускаемой Perkin Elmer) и выдерживали при 37°C в течение 2 часов.
После окончания реакции в свободном объеме флакона измеряли концентрацию изопрена газовой хроматографией. Параметры анализа газовой хроматографии будут описаны ниже.
[0082] Устройство для забора образцов из свободного объема (выпускаемое Perkin Elmer, Turbo Matrix 40)
Температура для поддержания флакона нагретым: 40°C
Период времени поддержания флакона нагретым: 30 минут
Время воздействия давления: 3,0 минуты
Время впрыска: 0,02 минуты
Температура иглы: 70°C
Температура переноса: 80°C
Давление газа носителя (гелий высокой чистоты): 124 кПа
[0083] Газовый хроматограф (выпускаемый Shimadzu Corporation, GC-2010 Plus AF)
Колонка (Rxi (зарегистрированный товарный знак)-1 мс: длина 30 м, внутренний диаметр 0,53 мм, толщина пленки жидкой фазы 1,5 мкм, каталожный №13370)
Температура колонки: давление 37°C: 24,8 кПа
Скорость потока колонки: 5 мл/мин
Способ поступления: разделение 1:0 (фактически измеряемое 1:18)
Несущий поток: 90 мл
Объем впрыска GC: 1,8 мл (несущий поток × время впрыска)
Объем впрыска образца в колонку: 0,1 мл
Температура на входе: 250°C
Детектор: FID (водород 40 мл/мин, воздух 400 мл/мин, вспомогательный газ гелий 30 мл/мин)
Температура детектора: 250°C
[0084] Получение стандартного образца изопрена
Реагент изопрен (удельная плотность 0,681) разбавляли в 10, 100, 1000, 10000 и 100000 раз холодным метанолом с получением стандартных растворов для добавления. Затем добавляли 1 мкл каждого стандартного раствора для добавления во флакон со свободным объемом, в который добавляли 1 мл воды, и использовали в качестве стандартного образца.
[0085] Количество образовавшегося изопрена после реакции каждого микробного штамма в течение 2 часов, описано в таблице 3.
[0086]
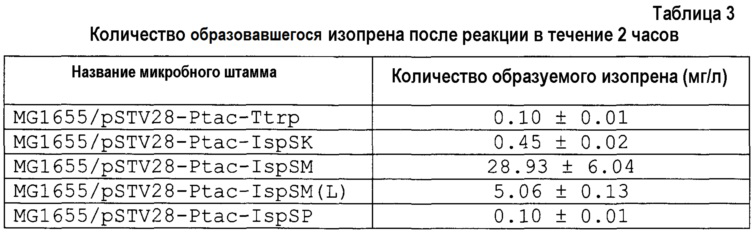
[0087] На основании результата в таблице 3 количество образовавшегося изопрена увеличивалось в порядке штаммов MG1655/pSTV28-Ptac-IspSM, MG1655/pSTV28-Ptac-IspSM (L) и MG1655/pSTV28-Ptac-IspSK и являлась практически одинаковым в штаммах MG1655/pSTV28-Ptac-IspSP и MG1655/pSTV28-Ptac-Ttrp. На основании указанного выше результата экстракт неочищенного фермента из штамма, в который вводили изопренсинтазу, полученную из мукуны, обладал наиболее высокой активностью образования изопрена.
[0088] Пример 5: Эффекты введения изопренсинтазы, полученной из различных растений, на штамм MG1655 E. coli
На основании результата активности неочищенного фермента в примере 4 наиболее высокую активность подтверждали для изопренсинтазы, полученной из мукуны, в которой удаляли сигнал локализации хлоропласта. Таким образом, способность продуцировать изопрен из глюкозы сравнивали во всех штаммах с введенной изопренсинтазов, в которых удаляли сигнал локализации хлоропласта. Микробные клетки штаммов MG1655/pSTV28-Ptac-Ttrp, MG1655/pSTV28-Ptac-IspSK, MG1655/pSTV28-Ptac-IspSP или MG1655/pSTV28-Ptac-IspSM равномерно наносили на чашку Петри с LB, покрытый 60 мг/л хлорамфеникола, и культивировали при 37°C в течение 18 часов. Одну петлю для посева микробных клеток из формированной чашки Петри инокулировали 1 мл среды M9 с глюкозой во флаконе со свободным объемом. Флакон плотно закрывали крышкой с бутилкаучуковой мембраной для флакона со свободным объемом (CRIMPS (каталожный № B0104240), выпускаемой Perkin Elmer), и культивировали микробные клетки при 30°C в течение 24 часов с использованием соответствующего аппарата для культивирования с возможностью встряхивания (120 об/мин). Композиция среды M9 с глюкозой является такой, как описано в таблице 4.
[0089]
Композиция среды M9 с глюкозой
Кроме того, хлорамфеникол добавляли в конечной концентрации 60 мг/л. Объем доводили до 1 л, и затем стерилизовали среду посредством фильтрации.
[0090] После окончания культивирования концентрацию изопрена в свободном объеме во флаконе измеряли газовой хроматографией. Также измеряли значение OD при 600 нм с использованием спектрофотометра (HITACHI U-2900). Концентрация изопрена и значение OD в каждом микробном штамме в момент времени окончания культивирования описаны в таблице 5.
[0091]
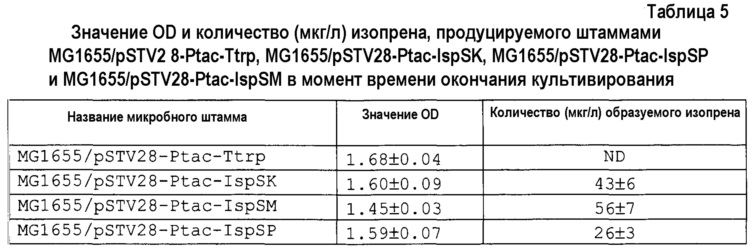
[0092] Из результатов в таблице 5 выявлено, что количество продуцируемого изопрена увеличивалось в порядке штаммов MG1655/pSTV28-Ptac-IspSM, MG1655/pSTV2 8-Ptac-IspSK, MG1655/pSTV28-Ptac-IspSP и MG1655/pSTV28-Ptac-Ttrp. На основании указанных выше результатов в штаммах дикого типа для штамма, в который вводили изопренсинтазу, полученную из мукуны, демонстрировали наиболее высокую активность продукции изопрена.
[0093] Пример 6: Эффекты введения изопренсинтазы, полученной из различных растений, на штамм MG1655 E. coli, в котором усиливали путь МЕР (метилэритриола).
6-1) Конструкция плазмиды для экспрессии гена dxs (pMW219-dxs
Уже было опубликовано, что количество образовавшегося изопрена увеличивалось (Appl. Microbiol. Biotechnol., (2011) 90, 1915-1922), когда усиливали экспрессию гена dxs (1-дезокси-D-ксилулоза-5-фосфатсинтетазы), который составляет путь МЕР, в штамме E. coli, в который вводили изопренсинтазу. Таким образом, подтверждали, отличалась ли также способность продуцировать изопрен вследствие природы изопренсинтазы в штамме, в котором усиливали экспрессию гена dxs. Полная геномная нуклеотидная последовательность штамма K-12 E. coli уже продемонстрирована (номер доступа GeneBank U00096) (Science, (1997) 277, 1453-1474). Для амплификации гена использовали pMW219 (выпускаемую Nippon Gene Co., Ltd.). Эта плазмида может повышать уровень экспрессии целевого гена, когда добавляют изопропил-β-тиогалактопиранозид (IPTG) посредством введения целевого гена в участок множественного клонирования. Синтезированные олигонуклеотиды синтезировали из нуклеотидных последовательностей, представленных SEQ ID NO: 23 и 24 на основе нуклеотидной последовательности гена dxs в геномной нуклеотидной последовательности E. coli. Затем проводили ПЦР с полимеразой Prime Star (выпускаемой Takara Bio Inc.) с использованием синтезированных олигонуклеотидов, состоящих из нуклеотидных последовательностей, представленных SEQ ID NO: 23 и 24, в качестве праймеров с геномной ДНК штамма MR1655 в качестве матрицы. Реакционный раствор получали в соответствии с рецептурой, прилагаемой к набору, и проводили реакцию при 98°C в течение 10 секунд, при 54°C в течение 20 секунд и при 68°C в течение 120 секунд в течение 40 циклов. В результате получали продукт ПЦР, содержащий ген dxs. Аналогично, проводили ПЦР с полимеразой Prime Star (выпускаемой Takara Bio Inc.) с использованием синтезированных олигонуклеотидов, состоящих из нуклеотидных последовательностей, представленных SEQ ID NO: 25 и 26, в качестве праймеров с pMW219 в качестве матрицы. Реакционный раствор получали в соответствии с рецептурой, прилагаемой к набору, и проводили реакцию при 98°C в течение 10 секунд, при 54°C в течение 20 секунд и при 68°C в течение 240 секунд в течение 40 циклов. В результате получали продукт ПЦР, содержащий pMW219. Затем, очищенный фрагмент гена dxs лигировали с продуктом ПЦР pMW219 с использованием набора для клонирования In-Fusion HD (выпускаемого Clontech). Полученную плазмиду для экспрессии гена dxs обозначали как pMW219-dxs.
[0094]

[0095] 6-2) Введение pMW219-dxs в штамм MG1655 E. coli, обладающий способностью продуцировать изопрен
Получали компетентные клетки штамма MG1655/pSTV28-Ptac-Ttrp, MG1655/pSTV2 8-Ptac-IspSK, MG1655/pSTV28-Ptac-IspSM или дополнительно MG1655/pSTV28-Ptac-IspSP и вводили в них pMW219-dxs способом электропорации. Клетки равномерно наносили на среду LB, содержащую 60 мг/л хлорамфеникола и 50 мг/л канамицина гидрохлорида, и культивировали клетки при 37°C в течение 18 часов. Из формированных чашек Петри с LB получали трансформанты, которые являлись устойчивыми к хлорамфениколу и канамицину. Штаммы, в которых pMW219-dxs вводили в штамм MG1655/pSTV28-Ptac-Ttrp, MG1655/pSTV28-Ptac-IspSK, MG1655/pSTV28-Ptac-IspSM или дополнительно MG1655/pSTV28-Ptac-IspSP, обозначали как штамм MG1655/pSTV28-Ptac-Ttrp/pMW219-dxs, MG1655/pSTV28-Ptac-IspSK/pMW219-dxs, MG1655/pSTV28-Ptac-IspSM/pMW219-dxs или дополнительно MG1655/pSTV28-Ptac-IspSP/pMW219-dxs, соответственно.
[0096] 6-3) Эффекты введения изопренсинтазы, полученной из различных растений, на штамм MG1655E. coli, в котором усилена экспрессия DXS
Штамм MG1655/pSTV28-Ptac-Ttrp/pMW219-dxs, MG1655/pSTV28-Ptac-IspSK/pMW219-dxs, MG1655/pSTV2 8-Ptac-IspSM/pMW219-dxs или дополнительно MG1655/pSTV28-Ptac-IspSP/pMW219-dxs раномерно наносили на среду LB, содержащую 60 мг/л хлорамфеникола и 50 мг/л канамицина гидрохлорида, и культивировали при 37°C в течение 18 часов. Затем культивирование во флаконе со свободным объемом оценивали, как описано в примере 5. Количество (мкг/л) продуцируемого изопрена и значение OD после окончания культивирования описаны в таблице 7.
[0097]

[0098] На основании результатов в таблице 7 количество продуцируемого изопрена увеличивалось в порядке штаммов MG1655/pSTV28-Ptac-IspSM/pMW219-dxs, MG1655/pSTV28-Ptac-IspSK/pMW219-dxs, MG1655/pSTV28-Ptac-IspSP/pMW219-dxs и MG1655/pSTV28-Ptac-Ttrp/pMW219-dxs. На основании указанных выше результатов в штаммах с усиленным путем передачи сигнала МЕР штамм, в который вводили изопренсинтазу, полученную из мукуны, также обладал наиболее выраженной способностью продуцировать изопрен.
[0099] Пример 7: Эффекты введения изопренсинтазы, полученной из различных растений на штамм MG1655 E. coli, в котором вводят путь MVA (мевалоновой кислоты)
7-1) Клонирование гена, регулирующего последующие звенья мевалонатного пути, который получают из дрожжей
Область в 5'-3' направлении мевалонатного пути, получали из Saccharomyces cerevisiae (WO 2009076676, Saccharomyces база данных геномов http://www.yeastgenome.org/# Nucleic Acids Res., Jan 2012; 40:D700-D705). Ген ERG12, кодирующий меалонаткиназу, ген ERG8, кодирующий фосфомеалонаткиназу, ген ERG19, кодирующий дифосфомевалонатдекарбоксилазу, и ген IDI1, кодирующий изопентенилдифосфат-дельта-изомеразу, амплифицировали посредством ПЦР с геномной ДНК S. cerevisiae в качестве матрицы с использованием праймера, указанного ниже (таблица 8). Для фермента ПЦР использовали Prime Star Max Premix, поставляемую Takara Bio Inc., и проводили реакцию при 98°C в течение 2 минут и в течение 30 циклов 98°C в течение 10 секунд, 55°C в течение 5 секунд и 72°C в течение 5 секунд/т.п.н. Клонирование и конструирование экспрессирующего вектора проводили введением фрагмента ПЦР в вектор pSTV28-Ptac-Ttrp (SEQ ID NO: 12), обработанный ферментом рестрикции SmaI, способом клонирования "in-fusion". DH5α E. coli трансформировали экспрессирующим вектором, выбирали клоны, содержащие допустимую длину последовательности каждого гена, плазмиду экстрагировали стандартными способами и подтверждали ее последовательность. Нуклеотидные последовательности таких амплифицированных генов и аминокислотные последовательности ферментов, кодируемых этими генами, являются доступными в базе данных геномов Saccharomyces http://www.yeastgenome.org/#.
[0100]

[0101] 7-2) Конструкция искусственного оперона в 5'-3' направлении мевалонатного пути
Последовательность, в которой ген, кодирующий мевалонаткиназу, и ген, кодирующий фосфомевалонаткиназу, располагали в прямом направлении, конструировали способом клонирования "in-fusion". Ген ERG12, кодирующий мевалонаткиназу, и ген ERG8, кодирующий фосфомевалонаткиназу, амплифицировали ПЦР с геномной ДНК от Saccharomyces cerevisiae в качестве матрицы с использованием праймеров, представленных в таблице 9. Для фермента ПЦР использовали KOD plus, поставляемый Toyobo, и проводили реакцию при 94°C в течение 2 минут и в течение 30 циклов 94°C в течение 15 секунд, 45°C в течение 30 секунд и 68°C в течение 1 минуты/т.п.н. Клонирование и конструирование экспрессирующего вектора проводили введением фрагмента ПЦР в вектор pUC118, обработанный фермент рестрикции SmaI способом клонирования "in-fusion". JM109 E. coli трансформировали экспрессирующим вектором, выбирали клоны, содержащие допустимую длину последовательности каждого гена, плазмиду экстрагировали стандартными способами и подтверждали ее последовательность. Полученную плазмиду обозначали как pUC-mvk-pmk. Нуклеотидная последовательность pUC-mvk-pmk представлена SEQ ID NO: 35.
[0102]

[0103] Последовательность, в которой ген, кодирующий дифосфомевалонатдекарбоксилaзу, и ген, кодирующий изопентенилдифосфат-дельта-изомеразу, располагали в прямом направлении, конструировали способом клонирования "in-fusion". Ген ERG19 ген, кодирующий дифосфомевалонатдекарбоксилaзу, и ген IDI1, кодирующий изопентенилдифосфат-дельта-изомеразу, амплифицировали ПЦР с геномной ДНК Saccharomyces cerevisiae в качестве матрицы с использованием праймеров, представленных в таблице 10. Для фермента ПЦР использовали KOD plus, поставляемый Toyobo, и проводили реакцию при 94°C в течение 2 минут и в течение 30 циклов 94°C в течение 15 секунд, 45°C в течение 30 секунд и 68°C в течение 1 минуты/т.п.н., и затем при 68°C в течение 10 минут. Клонирование и конструирование экспрессирующего вектора проводили введением фрагмента ПЦР в вектор TWV228, обработанный ферментом рестрикции SmaI, способом клонирования "in-fusion". DH5α E. coli трансформировали экспрессирующим вектором, выбирали клоны, содержащие допустимую длину последовательности каждого гена, экстрагировали плазмиду стандартными способами и подтверждали ее последовательность. Полученную плазмиду обозначали как pTWV-dmd-yidi. Нуклеотидная последовательность pTWV-dmd-yidi представлена SEQ ID NO: 40.
[0104]

[0105] Последовательность, в которой ген, кодирующий мевалонаткиназу, ген, кодирующий дифосфомевалонаткиназу, ген, кодирующий дифосфомевалонатдекарбоксилaзу, и ген, кодирующий изопентенилдифосфат-дельта-изомеразу, располагали в прямом направлении, конструировали способом клонирования "in-fusion". Экспрессирующий вектор, в которых гены этих четырех ферментов располагали в прямом направлении, конструировали посредством амплификации гена, кодирующего мевалонаткиназу, и гена, кодирующего дифосфомевалонаткиназу, ПЦР с pUC-mvk-pmk в качестве матрицы, с использованием праймеров, представленных в таблице 11, и амплификации гена, кодирующего дифосфомевалонатдекарбоксилaзу, и гена, кодирующего изопентенилдифосфат-дельта-изомеразу, ПЦР с pTWV-dmd-yidi в качестве матрицы с использованием праймеров shown в таблице 11, с последующим клонированием продуктов амплификации в pTrcHis2B вектор способом клонирования "in-fusion". Для фермента ПЦР использовали ДНК-полимераза Prime Star HS, поставляемую Takara Bio Inc., и проводили реакцию при 98°C в течение 2 минут с последующими 30 циклами 98°C в течение 10 секунд, 52°C в течение 5 секунд и 72°C в течение 1 минуты/т.п.н., и затем при 72°C в течение 10 минут. Для конструкции экспрессирующего вектора ПЦР фрагмент встраивали в вектор pTrcHis2B, обработанный ферментами рестрикции NcoI и PstI. JM109E. coli трансформировали экспрессирующим вектором, выбирали клоны с целевой длиной последовательности, экстрагировали плазмиду стандартными способами и подтверждали ее последовательность. Конструируемый экспрессирующий вектор обозначали как pTrc-KKDyI (β). Нуклеотидная последовательность pTrc-KKDyI (β) представлена SEQ ID NO: 45.
[0106]

[0107] 7-3) Фиксация области в направлении 5'-3' мевалонатного пути на хромосоме
Последовательность, в которой ген, кодирующий мевалонаткиназу, ген, кодирующий дифосфомевалонаткиназу, ген, кодирующий дифосфомевалонатдекарбоксилaзу, и ген, кодирующий изопентенилдифосфат-дельта-изомеразу, располагали в прямом направлении, экспрессировалась на хромосоме. Для экспрессии гена использовали промотор глюкоизомеразы и для терминации транскрипции использовали область терминации транскрипции гена aspA в E. coli (WO 2010031062). Участок транслокации Tn7 использовали в качестве хромосомного участка, который необходимо фиксировать (Mol. Gen. Genet., 1981, 183 (2):380-7). В качестве маркера лекарственной устойчивости использовали ген cat после фиксации на хромосоме. Область в 5'-3' направлении T7 в области хромосомы, которую необходимо фиксировать, амплифицировали ПЦР с геномной ДНК E. coli в качестве матрицы с использованием праймеров, представленных в таблице 12. Для фермента использовали ПЦР ДНК-полимераза Prime Star HS, поставляемую Takara Bio Inc., и проводили реакцию при 98°C в течение 2 минут с последующими 30 циклами 98°C в течение 10 секунд, 52°C в течение 5 секунд и 72°C в течение 1 минуты/т.п.н., и затем при 72°C в течение 10 минут. Область гена cat, содержащую участок прикрепления фага λ, амплифицировали ПЦР с плазмидой pMW118-attL-Cm-attR в качестве матрицы с использованием праймеров, представленных в таблице 12 (WO 2010-027022). Для фермента ПЦР использовали ДНК-полимеразу Prime Star HS, поставляемую Takara Bio Inc., и проводили реакцию при 95°C в течение 3 минут с последующими 2 циклами 95°C в течение 1 минуты, 34°C в течение 30 секунд и 72°C в течение 40 секунд, 2 циклами 95°C в течение 30 секунд, 50°C в течение 30 секунд и 72°C в течение 40 секунд, и затем при 72°C в течение 5 минут. Последовательность в направлении в 5'-3' направлении мевалонатного пути, к которой добавляли промотор и область терминации транскрипции (далее в настоящем описании сокращенно обозначаемой KKDyI) амплифицировали c pTrc-KKDyI (β) в качестве матрицы с использованием праймеров, представленных в таблице 12. Для фермента ПЦР использовали ДНК-полимеразу Prime Star HS, поставляемую Takara Bio Inc., и проводили реакцию при 98°C в течение 2 минут с последующими 30 циклами 98°C в течение 10 секунд, 52°C в течение 5 секунд и 72°C в течение 1 минуты/т.п.н., и затем при 72°C в течение 10 минут. Вектор конструировали с использованием продуктов ПЦР и pMW219, обработанной ферментом рестрикции SmaI, способом клонирования "in-fusion". JM109 E. coli трансформировали экспрессирующим вектором, выбирали клоны с целевой длиной последовательности, экстрагировали плазмиду стандартными способами и подтверждали ее последовательность. Полученную плазмиду обозначали как pMW219-KKDyI-TaspA. Нуклеотидная последовательность pMW219-KKDyI-TaspA представлена SEQ ID NO: 50.
[0108] Затем область в 3'-5' направлении Tn7 в области хромосомы, которую необходимо фиксировать, амплифицировали ПЦР с геномной ДНК E. coli в качестве матрицы с использованием праймеров, представленных в таблице 13. Для фермента ПЦР использовали ДНК-полимеразу Prime Star HS, поставляемую Takara Bio Inc., и проводили реакцию при 98°C в течение 2 минут с последующими 30 циклами 98°C в течение 10 секунд, 52°C в течение 5 секунд и 72°C в течение 1 минуты/т.п.н., и затем при 72°C в течение 10 минут. Вектор конструировали с использованием продукта ПЦР и pMW219-KKDyI-TaspA, обработанной ферментом рестрикции SalI, способом клонирования "in-fusion". JM109 E. coli трансформировали экспрессирующим вектором, выбирали клоны с целевой длиной последовательности, экстрагировали плазмиду стандартными способами и подтверждали ее последовательность. Полученную плазмиду обозначали как pMW-Tn7-Pgi-KKDyI-TaspA-Tn7. Последовательность конструируемой плазмиды представлена SEQ ID NO: 51.
[0109] Затем хромосому, содержащую область, содержащую ген устойчивости к хлорамфениколу, промотор глюкоизомеразы, оперон в 5'-3' направлении мевалонатного пути и область терминации транскрипции гена aspA, фиксировали с применением способа λ-Red. Фрагмент для фиксации хромосомы получали экстракцией плазмиды pMW-Tn7-Pgi-KKDyI-TaspA-Tn7, и затем ее обработкой ферментом рестрикции PvuI и SaiI, с последующей ее очисткой. Для электропорации использовали MG1655 E. coli, содержащий плазмиду pKD46, обладающую способностью температурно-чувствительной репликации (далее в настоящем описании, обозначаемый как MG1655/pKD46). Плазмида pKD46 [Proc. Natl. Acad. Sci. USA, 2000, vol.97, № 12, pр.6640-6645] содержит фрагмент ДНК общей длиной 2154 нуклеотидов (номер доступа GenBank/EMBL J02459, 31088-33241) фага λ, содержащего гены системы λ Red (λ, β, экзогены), контролируемые индуцируемым арабинозой промотором ParaB. После электропорации получали колонию, которая приобретала устойчивость к хлорамфениколу, затем экстрагировали геномную ДНК и подтверждали ее ПЦР с использованием праймеров, представленных в таблице 14, целевую область которой фиксировали на хромосоме. В дальнейшем последовательность целевой области подтверждали путем подтверждения последовательности фрагмента ПЦР. Нуклеотидная последовательность мевалонатного пути в 5'-3' направлении и ее проксимальная область, фиксированные на хромосоме, представлены SEQ ID NO: 52, и ее общая схема конструкции представлена на фиг. 3. Полученный мутант обозначали как MG1655 cat-Pgi-KKDyI.
[0110] Маркер лекарственной устойчивости в MG1655 cat-Pgi-KKDyI удаляли следующим ниже способом. Получали компетентные клетки MG1655 cat-Pgi-KKDyI, и затем вводили в них pMW-int-xis. pMW-int-xis представляет собой плазмиду, содержащую ген, кодирующий интегразу (Int) фага λ, и ген, кодирующий эксцизионную гидролазу (Xis) фага λ, и обладающую способностью температурно-чувствительной репликации (WO 2007/037460, JP 2005-058827).
[0111] Ген устойчивости к хлорамфениколу, располагаемый в области, лежащей между attL и attR, которые являются участком прикрепления фага λ, удаляли из хромосомы введением pMW-int-xis. В результате, известно, что хозяин теряет устойчивость к хлорамфениколу. Таким образом, из полученной колонии получали чувствительный к хлорамфениколу штамм, и затем культивировали в среде LB при 42°C в течение 6 часов. Культивируемые микробные клетки наносили на среду LB для чашек Петри, обеспечивая возможность прорастания колоний. Из этих колоний выбирали колонию, которая утрачивала устойчивость к ампициллину, для устранения лекарственной устойчивости. Полученный мутант, как указано выше, обозначали как MG1655 Pgi-KKDyI.
[0112]
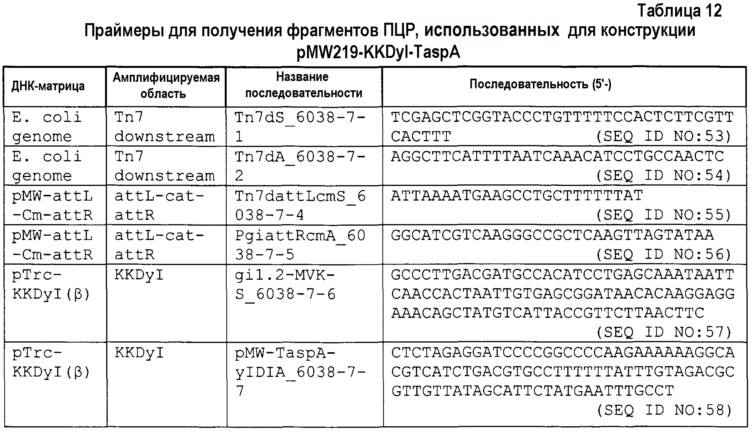
[0113]

[0114]
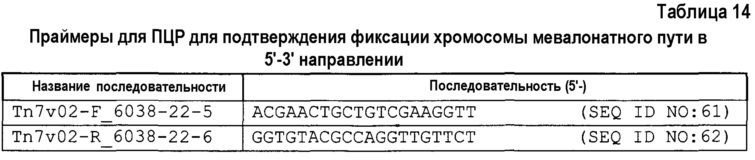
[0115] 7-4) Замена промотора в 5'-3' направлении мевалонатного пути на хромосоме
Промотор оперона в 5'-3' направлении мевалонатного пути на хромосоме заменяли способом λ-red. В качестве матрицы для ПЦР использовали геномный фрагмент, содержащий attL-Tet-attR-Ptac. Он представляет собой фрагмент, в котором промотор tac и attL и attR, которые представляют собой участки прикрепления маркера лекарственной устойчивости к тетрациклину и фага λ, являются выровненными. Такая последовательность представлена SEQ ID NO: 63. Фрагмент ПЦР получали с использованием промотора, представленного в таблице 15. Для фермента ПЦР использовали полимеразу LA-Taq, поставляемую Takara Bio Inc., и проводили реакцию при 92°C в течение 1 минуты, затем в течение 40 циклов 92°C в течение 10 секунд, 50°C в течение 20 секунд и 72°C в течение 1 минуты/т.п.н., и дополнительно при 72°C в течение 7 минут. Очищали продукт ПЦР. Для электропорации использовали MG1655 Pgi-KKDyI, содержащий плазмиду pKD46 (далее в настоящем описании, обозначаемый как MG1655 Pgi-KKDyI/pKD46), обладающий способностью температурно-чувствительной репликации. Плазмида pKD46 [Proc. Natl. Acad. Sci. USA, 2000, vol.97, № 12, pp.6640-6645] содержит фрагмент ДНК общей длиной 2154 нуклеотидов (номер доступа GenBank/EMBL J02459, 31088-33241) фага λ, содержащего гены системы Red (λ, β, экзогены), контролируемые индуцируемым арабинозой промотором ParaB. Плазмида pKD46 необходима для встраивания продукта ПЦР в MG1655 Pgi-KKDyI.
[0116] Компетентные клетки для электропорации получали, как указано ниже. MG1655 Pgi-KKDyI/pKD46 культивировали в среде LB, содержащей 100 мг/л ампициллина при 30°C в течение ночи, разбавляли в 100 раз 5 мл среды LB, содержащей ампициллин и L-арабинозу (1 мМ). Для использования для электропорации полученные клетки в разбавленной суспензии выращивали до получения OD600 приблизительно 0,6 в условиях вентиляции при 30°C, и затем промывали три раза ледяным 10% раствором глицерина. Электропорацию проводили с использованием 50 мкл компетентных клеток и приблизительно 100 нг продукта ПЦР. После электропорации клетки культивировали в 1 мл среды SOC [Molecular Cloning: Laboratory Manuals, 2nd Edition, Sambrook J. et al., Cold Spring Harbor Laboratory Press (1989)] при 37°C в течение одного часа и подвергали культурованию на твердой среде на агаровой среде LB при 37°C для отбора устойчивого к хлорамфениколу трансформанта. Затем для удаления плазмиды KD46 трансформант пассировали на агаровой среде LB, содержащей тетрациклин при 37°C. В полученных колониях исследовали устойчивость к ампициллину и получали устойчивый к ампициллину штамм, не содержащий pKD46. Получали мутант, содержащий замену промотора tac, который можно отличать по гену устойчивости к тетрациклину. Полученный мутант обозначали как MG1655 tet-Ptac-KKDyI.
[0117] Маркер устойчивости к антибиотику удаляли следующим ниже способом. Получали компетентные клетки MG1655 tet-Ptac-KKDyI, и затем вводили в них pMW-int-xis. pMW-int-xis представляет собой плазмиду, содержащую гены, кодирующие интегразу (Int) и эксцизионную гидролазу (Xis) фага λ, и обладающую способностью температурно-чувствительной репликации (WO 2007/037460, публикация № JP 2005-058827). Ген устойчивости к тетрациклину, располагаемый в области, находящейся между attL и attR, которые представляют собой участок прикрепления фага λ, удаляли из хромосомы введением pMW-int-xis. В результате, известно, что хозяин теряет устойчивость к тетрациклину. Таким образом, из полученных колоний получали чувствительный к тетрациклину штамм. Клетки такого штамма культивировали в среде LB при 42°C в течение 6 часов и наносили культивируемые клетки в среду LB для чашек Петри для обеспечения роста колоний. Для удаления лекарственной устойчивости выбирали клон, который терял устойчивость к ампициллину. Полученный мутант обозначали как MG1655 Ptac-KKDyI. Нуклеотидная последовательность мевалонатного пути в 5'-3' направлении и ее проксимальная область, контролируемая промотором tac на хромосоме, представлены SEQ ID NO: 64, и ее общая схема представлена на фиг. 4.
[0118]

[0119] 7-5) Введение изопренсинтазы, полученной из различных растений, в штамм MG1655 Ptac-KKDyI
Получали компетентные клетки штамма MG1655 Ptac-KKDyI, и затем водили в них pSTV28-Ptac-Ttrp, pSTV28-Ptac-IspSK, pSTV28-Ptac-IspSM или дополнительно pSTV28-Ptac-SP. Клетки равномерно наносили на чашку Петри с LB, содержащую 60 мг/л хлорамфеникола, и культивировали клетки при 37°C в течение 18 часов. Из формированной чашки Петри получали трансформанты, которые обладали устойчивостью к хлорамфениколу. Штамм, в котором pSTV28-Ptac-Ttrp, pSTV28-Ptac-IspSK, pSTV28-Ptac-IspSM или pSTV28-Ptac-IspSP вводили в штамм MG1655 Ptac-KKDyI, обозначали как MG1655 Ptac-KKDyI/pSTV28-Ptac-Ttrp, MG1655 Ptac-KKDyI/pSTV28-Ptac-IspSK, MG1655 Ptac-KKDyI/pSTV28-Ptac-IspSM или MG1655 Ptac-KKDyI/pSTV28-Ptac-IspSP, соответственно.
[0120] 7-6) Эффекты введения изопренсинтазы, полученной из различных растений, на штамм MG1655, в котором усилен путь MVA
Микробные клетки MG1655 Ptac-KKDyI/pSTV28-Ptac-Ttrp, MG1655 Ptac-KKDyI/pSTV28-Ptac-IspSK, MG1655 Ptac-KKDyI/pSTV28-Ptac-IspSM или дополнительно MG1655 Ptac-KKDyI/pSTV28-Ptac-IspSP равномерно наносили на чашку Петри с LB, содержащую 60 мг/л хлорамфеникола, и культивировали клетки при 37°C в течение 18 часов. Одну петлю для посева микробных клеток из формированной чашки Петри LB инокулировали в 1 мл среды M9 с глюкозой (содержащей мевалоновую кислоту) во флакон со свободным объемом (22 мл флакон CLEAR CRIMP TOP (каталожный № B0104236), выпускаемый Perkin Elmer), и затем оценивали культивирование способом, описанным в примере 2. Композиция среды M9 с глюкозой (содержащей мевалоновую кислоту) описана в таблице 16. Количество продуцируемого изопрена и значение OD после окончания культивирования описаны в таблице 17.
[0121]
Композиция среды M9 с глюкозой (содержащей мевалоновую кислоту)
Хлорамфеникол добавляли в конечной концентрации 60 мг/л.
Общий объем доводили до 1 л и стерилизовали среду посредством фильтрации.
[0122]
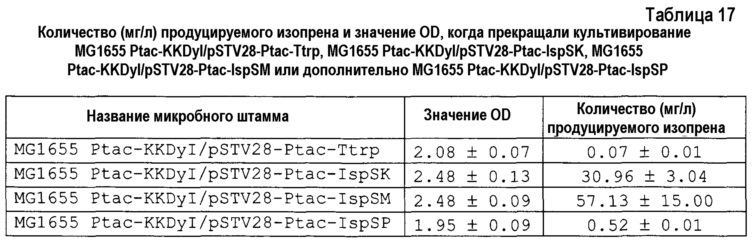
[0123] На основании результатов в таблице 17 количество продуцируемого изопрена увеличивалось в порядке штаммов MG1655 Ptac-KKDyI/pSTV28-Ptac-IspSM, MG1655 Ptac-KKDyI/pSTV28-Ptac-IspSK, MG1655 Ptac-KKDyI/pSTV2 8-Ptac-IspSP и MG1655 Ptac-KKDyI/pSTV28-Ptac-Ttrp. На основании указанных выше результатов в штаммах, в которые вводили путь MVA, штамм, в который вводили изопренсинтазу, полученную из мукуны, также обладал наиболее высокой способностью продуцировать изопрен.
[0124] Пример 8: Измерение ферментативной активности изопренсинтазы, полученной из различных растений, с использованием экстракта неочищенного фермента, полученного из Corynebacterium glutamicum
1) Конструкция плазмиды для экспрессии гена IspS, полученного из различных растений
В качестве коринеформной бактерии использовали штамм 2256 C. glutamicum (ATCC13869) (Okumura et al., 1962, Santamaria et al., 1984, Tsuchida et al., 1986). Плазмиды для экспрессии гена IspSK, гена IspSP и гена IspSM в C. glutamicum конструировали следующим ниже способом. Проводили ПЦР с полимеразой Prime Star (выпускаемой Takara Bio Inc.) с использованием синтезированных олигонуклеотидов, представленных SEQ ID NO: 67 и 68 в качестве праймеров с pUC57-IspSK в качестве матрицы, синтезированных олигонуклеотидов, представленных SEQ ID NO: 69 и 70 в качестве праймеров с pUC57-IspSP в качестве матрицы или синтезированных олигонуклеотидов, представленных SEQ ID NO: 71 и 72 в качестве праймеров с pUC57-IspSM в качестве матрицы. Реакционный раствор получали в соответствии с рецептурой, прилагаемой к набору, и проводили реакцию в течение 30 циклов 98°C в течение 10 секунд, 55°C в течение 5 секунд и 72°C в течение 120 секунд. В результате получали продукт ПЦР, содержащий ген IspSK, ген IspSP или ген IspSM. Затем с целью получения промоторной последовательности фактора элонгации Tu (далее в настоящем описании Р0480) проводили ПЦР с использованием хромосомной ДНК штамма 2256 C. glutamicum в качестве матрицы. Для объединения гена IspSK с Р0480 в качестве праймеров для ПЦР использовали синтезированные нуклеотиды, представленные SEQ ID NO: 73 и 74. Также для объединения гена IspSP с Р0480 в качестве праймеров для ПЦР использовали синтезированные нуклеотиды, представленные SEQ ID NO: 73 и 75. Также для объединения гена IspSM с Р0480 в качестве праймеров для ПЦР использовали синтезированные нуклеотиды, представленные SEQ ID NO: 73 и 76. Для ПЦР использовали полимеразу Prime Star (поставляемую Takara Bio Inc.). Реакционный раствор получали в соответствии с рецептурой, прилагаемой к набору, и проводили реакцию в течение 30 циклов 98°C в течение 10 секунд, 55°C в течение 5 секунд и 72°C в течение 30 секунд. В результате получали продукты ПЦР, содержащие Р0480. Сведения о последовательности Р0480 представлены SEQ ID NO: 77. Челночный вектор pVK9 для C. glutamicum и E. coli расщепляли ферментом рестрикции XbaI (поставляемым Takara Bio Inc.) (Miwa et al., 1985). Сведения о последовательности pVK9 представлены SEQ ID NO: 78. Очищенный фрагмент гена IspSK, гена IspSP или гена IspSM, продукта ПЦР Р0480 и очищенный pVK9, обработанный XbaI, лигировали с использованием набора для клонирования In-fusion HD (поставляемого Clontech). Полученные плазмиды для экспрессии гена IspSK, гена IspSP и гена IspSM обозначали как pVK9-Р0480-IspSK, pVK9-Р0480-IspSP и pVK9-Р0480-IspSM, соответственно, и сведения об их последовательности представлены SEQ ID NO: 79, 80 и 81, соответственно.
[0125] 2) Конструкция штамма 2256 C. glutamicum, обладающего способностью продуцировать изопрен
Микробные клетки штамма 2256 C. glutamicumstrain инокулировали в тестовую пробирку, содержащую 4 мл среды CM-Dex, и культивировали до логарифмической фазы роста при 30°C при 120 об/мин. Эти микробные клетки собирали, промывали два-три раза ледяным 10% глицерином, концентрировали приблизительно от 100 до 200 раз и использовали в качестве компетентных клеток. Компетентные клетки и каждую плазмиду из pVK9, pVK9-Р0480-IspSK, pVK9-Р0480-IspSP или pVK9-Р0480-IspSM тщательно перемешивали и применяли к ним импульс. Применение импульса проводили в условиях 1,8 кВ, 25 мкФ и 200 Ом с использованием GenePulserXCell, выпускаемого BIORAD. После применения импульса клетки восстанавливали культивированием в среде CM-Dex в течение приблизительно от 1-2 часов, затем помещали в среду CM-Dex в чашку Петри, содержащую 50 мкг/мл Km, и культивировали при 30°C в течение 18-24 часов. После культивирования из чашки Петри получали трансформанты, обладающие устойчивостью к канамицину. Композиция среды CM-Dex представлена в таблице 18. Штамм, в котором pVK9, pVK9-Р0480-IspSK, pVK9-Р0480-IspSP или pVK9-Р0480-IspSM вводили в штамм 2256 C. glutamicum, обозначали как 2256/pVK9, 2256/pVK9-Р0480-IspSK, 2256/pVK9-Р0480-IspSP или 2256/pVK9-Р0480-IspSM, соответственно.
[0126]
Композиция среды CM-Dex
Среду стерилизовали при 120°C в течение 20 минут. При необходимости добавляли канамицин в конечной концентрации 50 мг/л.
В случае чашки Петри, добавляли 20 г/л агара.
[0127] 3) Способ получения экстракта неочищенного фермента
Микробные клетки 2256/pVK9, 2256/pVK9-Р0480-IspSK, 2256/pVK9-Р0480-IspSP или 2256/pVK9-Р0480-IspSM равномерно наносили на среду CM-Dex в чашке Петри, содержащую 50 мг/л канамицина, и культивировали при 30°C в течение 18 часов. Микробные клетки соответствующие 1/6 от культивируемой чашки Петри инокулировали в колбу Сакагути, в которой содержалось 20 мл среды CM-Dex, содержащей 50 мг/л канамицина, и культивировали при 30°C в течение 6 часов. Микробные клетки выделяли из среды для культивирования центрифугированием при 5000 об./мин в течение 5 минут и дважды промывали буфером для изопренсинтазы (50 мМ Tris-HCl (pH 8,0), 20 мМ MgCl2, 5% глицерин). После промывания микробные клетки ресуспендировали в 1,8 мл того же буфера. Затем приблизительно 0,9 мл гранул для дезинтеграции (YBG01, диаметр 0,1 мм) и 0,9 мл суспензии микробных клеток помещали в 2 мл пробирку, специальную для устройства для проведения шока с использованием множества гранул. Микробные клетки подвергали дезинтеграции с использованием устройства для проведения шока с использованием множества гранул (модель MB701 (S)), выпускаемого Yasui Kikai Corporation, при 2500 об./мин в течение 6 циклов в положение "Вкл" в течение 60 секунд/в положение "Выкл" в течение 60 секунд.
После дезинтеграции пробирку центрифугировали при 20000×g в течение 20 минут и использовали полученный супернатант в качестве экстракта неочищенного фермента.
[0128] 4) Измерение активности изопренсинтазы
Концентрация белка экстрактов неочищенного фермента из штаммов 2256/pVK9, 2256/pVK9-Р0480-IspSK, 2256/pVK9-Р0480-IspSP и 2256/pVK9-Р0480-IspSM измеряли способом с бицинхониновой кислотой (способ BCA). В качестве реагентов для измерения использовали набор BCA protein assay reagent (Thermo Scientific Japan, каталожный № 23227). Объединяли и подготавливали 2 мг экстракта неочищенного фермента в качестве количества общего белка и изопреновый буфер с получением всего 0,5 мл. Указанное соединение помещали во флакон со свободным объемом, затем добавляли в него 0,025 мл 0,5M раствора MgCl2 и 0,01 мл 0,2M раствора DMAPP (выпускаемого Cayman, каталожный номер 63180) и слегка перемешивали смесь вихревым способом. Затем сразу же плотно закрывали флакон и проводили реакцию при 37°C в течение 2 часов. После проведения реакции измеряли концентрацию изопрена в свободном объеме во флаконе газовой хроматографией. В качестве флакона со свободным объемом использовали 22 мл флакон CLEAR CRIMP TOP (каталожный № B0104236), выпускаемый Perkin Elmer и в качестве крышки для герметичного закупоривания использовали крышку с бутилкаучуковой мембраной для флакона со свободным объемом (каталожный № B0104240), выпускаемую Perkin Elmer.
Количества изопрена, образовавшегося после реакции в течение 2 часов в различных микробных штаммах, описаны в таблице 19.
[0129]

[0130] На основании результатов в таблице 19 количество образовавшегося изопрена было больше в штамме 2256/pVK9-Р0480-IspSM по сравнению со штаммом 2256/pVK9-Р0480-IspSK. Количество образовавшегося изопрена в штамме 2256/pVK9-Р0480-IspSP было таким же низким, как в штамме 2256/pVK9. На основании указанных выше результатов экстракт неочищенного фермента, полученный из штамма, в который вводили изопренсинтазу, полученную из мукуны, обладал наиболее высокой активностью образования изопрена.
[0131] Пример 9: Эффекты введения изопренсинтазы, полученной из различных растений, на штамм 2256 Corynebacterium glutamicum
1) Эффекты введения изопренсинтазы, полученной из различных растений, на штамм 2256 C. glutamicum
Микробные клетки штамма 2256/pVK9, 2256/pVK9-P0480-IspSK, 2256/pVK9-Р0480-IspSP или 2256/pVK9-Р0480-IspSM равномерно наносили на чашку Петри с CM-Dex, содержащую 50 мг/л канамицина, и культивировали при 30°C в течение 18-24 часов. Из формированной чашки Петри инокулировали одну петлю для посева микробных клеток в 1 мл среды MM с глюкозой во флакон со свободным объемом. Затем флакон со свободным объемом плотно закрывали, и затем культивировали клетки в соответствующем аппарате с возможностью встряхивания при 30°C при 120 об/мин в течение 24 часов. В качестве флакона со свободным объемом использовали 22 мл флакон CLEAR CRIMP TOP (каталожный № B0104236), выпускаемый Perkin Elmer, и в качестве крышки для герметичного закупоривания использовали крышку с бутилкаучуковой мембраной для флакона со свободным объемом (каталожный № B0104240), выпускаемую Perkin Elmer.
Композиция среды MM с глюкозой является такой, как описано в таблице 20.
[0132]
Композиция среды MM с глюкозой
При необходимости добавляли канамицин в конечной концентрации 50 мг/л.
Общий объем доводили до 1 мл и стерилизовали среду посредством фильтрации.
[0133] После окончания культивирования измеряли концентрацию изопрена в свободном объеме во флаконе газовой хроматографией. Значение OD измеряли при 660 нм с использованием спектрофотометра (HITACHI U-2900).
Концентрации изопрена, образовавшегося из различных микробных штаммов после окончания культивирования, описаны в таблице 21.
[0134]
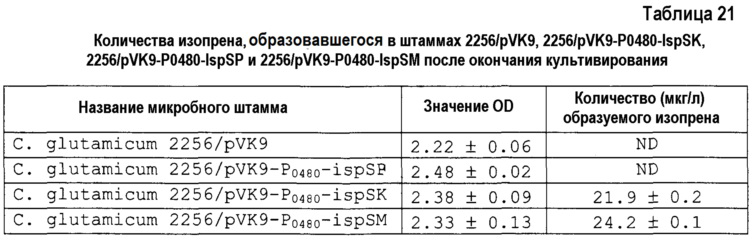
[0135] На основании результатов в таблице 21 количество продуцированного изопрена увеличивалось в порядке 2256/pVK9-Р0480-IspSM и 2256/pVK9-Р0480-IspSK. Изопрен не детектировали в 2256/pVK9 и 2256/pVK9-P0480-IspSP. На основании указанных выше результатов в штаммах 2256 C. glutamicum дикого типа штамм, в который вводили изопренсинтазу, полученную из мукуны, обладал наиболее высокой активностью продукции изопрен.
[0136] Пример 10: Измерение ферментативной активности изопренсинтазы, полученной из различных растений, с использованием экстракта неочищенного фермента, полученного из P. ananatis
1) Конструкция штамма AJ13355 P. ananatis, обладающего способностью продуцировать изопрен
Получали компетентные клетки AJ13355 P. ananatis, затем вводили в них pSTV28-Ptac-Ttrp, pSTV28-Ptac-IspSK, pSTV28-Ptac-IspSP или дополнительно pSTV28-Ptac-IspSM способом электропорации, и равномерно наносили клетки на чашку Петри с LB, содержащей 60 мг/л хлорамфеникола, и культивировали при 30°C в течение 18 часов. Затем из формированных чашек Петри получали трансформанты, обладающие устойчивостью к хлорамфениколу. Штамм, в котором pSTV28-Ptac-Ttrp, pSTV28-Ptac-IspSK, pSTV28-Ptac-IspSP или pSTV28-Ptac-IspSM вводили в штамм AJ13355 P. ananatis, обозначали как штамм AJ13355/pSTV28-Ptac-Ttrp, AJ13355/pSTV28-Ptac-IspSK, AJ13355/pSTV28-Ptac-IspSP или AJ13355/pSTV28-Ptac-IspSM, соответственно.
[0137] 2) Способ получения экстракта неочищенного фермента
Активность неочищенного фермента измеряли способом, описанным в примере 4. Микробные клетки штамма AJ13355/pSTV28-Ptac-Ttrp, AJ13355/pSTV28-Ptac-IspSK, AJ13355/pSTV28-Ptac-IspSP или AJ13355/pSTV28-Ptac-IspSM равномерно наносили на чашку Петри с LB, содержащей 60 мг/л хлорамфеникола, и культивировали при 30°C в течение 18 часов. Микробные клетки соответствующие 1/6 формированной чашки Петри инокулировали в колбу Сакагути, в которую добавляли 20 мл LB, содержащей 60 мг/л хлорамфеникола, и культивировали при 34°C в течение 6 часов. Микробные клетки из среда для культивирования центрифугировали при 6000 об/мин при 4°C в течение 5 минут и дважды промывали ледяным буфером для изопренсинтазы (50 мМ Tris-HCl, pH 8,0, 20 мМ MgCl2, 5% глицерин). Промытые микробные клетки суспендировали в 1,8 мл того же буфер. Приблизительно 0,9 мл гранул для дезинтеграции (YBG01, диаметр 0,1 мм) и 0,9 мл суспензии микробных клеток помещали в 2 мл пробирку, специальную для устройства для проведения шока с использованием множества гранул, и подвергали микробные клетки диезинтеграции с использованием устройства для проведения шока с использованием множества гранул, выпускаемого Yasui Kikai Corporation, при 2500 об/мин при 4°C в течение 3 циклов в положении "Включено" в течение 30 секунд/в положении "Выключено" в течение 30 секунд. После дезинтеграции пробирку центрифугировали при 20000×g при 4°C в течение 20 минут и использовали супернатант в качестве экстракта неочищенного фермента.
[0138] 3) Измерение активности изопренсинтазы
Общий объем 0,5 мл экстракта неочищенного фермента из штамма AJ13355/pSTV28-Ptac-Ttrp, AJ13355/pSTV28-Ptac-IspSK, AJ13355/pSTV28-Ptac-IspSP или AJ13355/pSTV28-Ptac-IspSM (для которого способом Бредфорда определяли, что он количественно составляет 2 мг в виде количества общего белка) и изопреновый буфер помещали во флакон со свободным объемом (22 мл флакон CLEAR CRIMP TOP (каталожный № B0104236), выпускаемый Perkin Elmer), затем добавляли в него 0,025 мл 0,5M раствора MgCl2 и 0,01 мл 0,2M раствора DMAPP (выпускаемого Cayman, каталожный № 63180) и аккуратно перемешивали смесь вихревым способом. Затем сразу же плотно закрывали флакон крышкой с бутилкаучуковой мембраной для флакона со свободным объемом (CRIMPS (каталожный № B0104240), выпускаемой Perkin Elmer) и хранили при 37°C в течение 2 часов.
После окончания реакции в свободном объеме флакона измеряли концентрацию изопрена газовой хроматографией.
[0139]

[0140] На основании результатов в таблице 22 количество образовавшегося изопрена увеличивалось в порядке AJ13355/pSTV28-Ptac-IspSM, AJ13355/pSTV28-Ptac-IspSK, AJ13355/pSTV28-Ptac-IspSP и AJ13355/pSTV28-Ptac-Ttrp. На основании указанных выше результатов экстракт неочищенного фермента из штамма, в который вводили изопренсинтазу, полученную из мукуны, обладал наиболее высокой активностью образования изопрена.
[0141] Пример 11: Эффекты введения изопренсинтазы, полученной из различных растений, на штамм AJ13355 P. ananatis
В различных штаммах AJ13355 P. ananatis, в которых удаляли сигнал локализации хлоропласта и вводили изопренсинтазу, сравнивали способность продуцировать изопрен из глюкозы. Микробные клетки штамма AJ13355/pSTV28-Ptac-Ttrp, AJ13355/pSTV28-Ptac-IspSK, AJ13355/pSTV28-Ptac-IspSP или AJ13355/pSTV28-Ptac-IspSM равномерно наносили на чашку Петри с LB, содержащей 60 мг/л хлорамфеникола, и культивировали при 30°C в течение 18 часов. Из формированной чашки Петри инокулировали одну петлю для посева микробных клеток в 1 мл среды M9 с глюкозой во флакон со свободным объемом. Затем флакон плотно закрывали крышкой с бутилкаучуковой мембраной для флакона со свободным объемом (CRIMPS (каталожный № B0104240), выпускаемой Perkin Elmer) и культивировали в соответствующем аппарате для культивирования с возможностью встряхивания (120 об/мин) при 30°C в течение 24 часов. После окончания культивирования в свободном объеме во флаконе измеряли концентрацию изопрена газовой хроматографией. Значение OD измеряли при 600 нм с использованием спектрофотометра (HITACHI U-2900). Концентрация изопрена и значение OD в различных микробных штаммах после окончания культивирования описаны в таблице 23.
[0142]

[0143] На основании результатов в таблице 23 количество продуцируемого изопрена увеличивалось в порядке штаммов AJ13355/pSTV28-Ptac-IspSM, AJ13355/pSTV28-Ptac-IspSK, AJ13355/pSTV28-Ptac-IspSP и AJ13355/pSTV28-Ptac-Ttrp. На основании указанных выше результатов штамм, в который вводили изопренсинтазу, полученную из мукуны, также обладал наиболее высокой способностью продуцировать изопрен в штаммах AJ13355 P. ananatis.
[0144] Пример 12: Измерение ферментативной активности изопренсинтазы, полученной из различных растений, с использованием экстракта неочищенного фермента, полученного из AJ110637 E. aerogenes
1) Конструкция штамма AJ110637 E. aerogenes, обладающего способностью продуцировать изопрен
Получали компетентные клетки AJ110637 E. aerogenes, затем в них вводили pSTV28-Ptac-Ttrp, pSTV28-Ptac-IspSK, pSTV28-Ptac-IspSP или дополнительно pSTV28-Ptac-IspSM способом электропорации, и равномерно наносили клетки на чашку Петри с LB, содержащую 60 мг/л хлорамфеникола и культивировали при 37°C в течение 18 часов. Затем из формированных чашек Петри получали трансформанты, обладающие устойчивостью к хлорамфениколу. Штамм, в котором pSTV28-Ptac-Ttrp, pSTV28-Ptac-IspSK, pSTV28-Ptac-IspSP или pSTV28-Ptac-IspSM вводили в штамм AJ110637 E. aerogenes, обозначали как штамм AJ110637/pSTV28-Ptac-Ttrp, AJ110637/pSTV28-Ptac-IspSK, AJ110637/pSTV28-Ptac-IspSP или AJ110637/pSTV28-Ptac-IspSM, соответственно.
[0145] 2) Способ получения экстракта неочищенного фермента
Активность неочищенного фермента измеряли способом, описанным в примере 4. Микробные клетки штамма AJ110637/pSTV28-Ptac-Ttrp, AJ110637/pSTV28-Ptac-IspSK, AJ110637/pSTV28-Ptac-IspSP или AJ110637/pSTV28-Ptac-IspSM равномерно наносили на чашку Петри с LB, содержащую 60 мг/л хлорамфеникола, и культивировали при 37°C в течение 18 часов. Микробные клетки соответствующие 1/6 формированной чашки Петри инокулировали в колбу Сакагути, в которую добавляли 20 мл LB, содержащей 60 мг/л хлорамфеникола, и культивировали при 37°C в течение 4 часов. Микробные клетки из среды для культивирования центрифугировали при 8000 об/мин при 4°C в течение 10 минут и дважды промывали ледяным буфером для изопренсинтазы (50 мМ Tris-HCl, pH 8,0, 20 мМ MgCl2, 5% глицерин). Промытые микробные клетки суспендировали в 1,8 мл того же буфера. Приблизительно 0,9 мл гранул для дезинтеграции (YBG01, диаметр 0,1 мм) и 0,9 мл суспензии микробных клеток помещали в 2 мл пробирку, специальную для устройства для проведения шока с использованием множества гранул, и подвергали микробные клетки дезинтеграции с использованием устройства для проведения шока с использованием множества гранул (модель MB701 (S)), выпускаемого Yasui Kikai Corporation, при 2500 об/мин при 4°C в течение 3 циклов в положении "Включено" в течение 30 секунд/в положении "Выключено" в течение 30 секунд. После дезинтеграции пробирку центрифугировали при 20000×g при 4°C в течение 20 минут и использовали супернатант в качестве экстракта неочищенного фермента.
[0146] 3) Измерение активности изопренсинтазы
Общий объем 0,5 мл экстракта неочищенного фермента из штамма AJ110637/pSTV28-Ptac-Ttrp, AJ110637/pSTV28-Ptac-IspSK, AJ110637/pSTV28-Ptac-IspSP или AJ110637/pSTV28-Ptac-IspSM (для которого способом Бредфорда определяли, что он количественно составляет 2 мг в виде количества общего белка) и изопреновый буфер помещали во флакон со свободным объемом (22 мл флакон CLEAR CRIMP TOP (каталожный № B0104236), выпускаемый Perkin Elmer), затем добавляли в него 0,025 мл 0,5M раствора MgCl2 и 0,01 мл 0,2M раствора DMAPP (выпускаемого Cayman, каталожный № 63180), и аккуратно перемешивали смесь вихревым способом. Затем сразу же флакон плотно закрывали крышкой с бутилкаучуковой мембраной для флакона со свободным объемом (CRIMPS (каталожный № B0104240), выпускаемой Perkin Elmer) и хранили при 37°C в течение 2 часов.
После окончания реакции в свободном объеме флакона измеряли концентрацию изопрена газовой хроматографией.
Количество образовавшегося изопрена в каждом микробном штамме после реакции в течение 2 часов описано в таблице 24.
[0147]
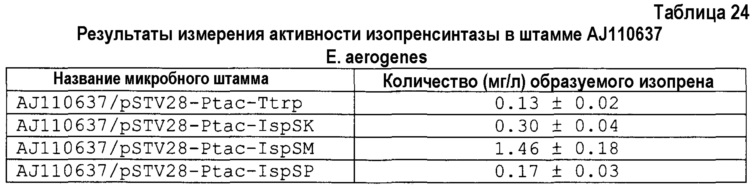
[0148] На основании результатов в таблице 24 количество образовавшегося изопрена увеличивалось в порядке штаммов AJ110637/pSTV28-Ptac-IspSM, AJ110637/pSTV28-Ptac-IspSK, AJ110637/pSTV28-Ptac-IspSP и AJ110637/pSTV28-Ptac-Ttrp. На основании указанных выше результатов экстракт неочищенного фермента из штамма, в которой вводили изопренсинтазу, полученную из мукуны, обладал наиболее высокой активностью образования изопрена.
[0149] Пример 13: Эффекты введения изопренсинтазы, полученной из различных растений, на штамм AJ110637 E. aerogenes
В различных штаммах AJ110637 E. aerogenes, в которых удаляли сигнал локализации хлоропласта и вводили изопренсинтазу, сравнивали способность продуцировать изопрен из глюкозы. Микробные клетки штамма AJ110637/pSTV28-Ptac-Ttrp, AJ110637/pSTV28-Ptac-IspSK, AJ110637/pSTV28-Ptac-IspSP или AJ110637/pSTV28-Ptac-IspSM равномерно наносили на чашку Петри с LB, содержащей 60 мг/л хлорамфеникола, и культивировали при 30°C в течение 18 часов. Из формированной чашки Петри инокулировали одну петлю для посева микробных клеток в 1 мл среды M9 с глюкозой во флакон со свободным объемом. Затем флакон плотно закрывали крышкой с бутилкаучуковой мембраной для флакона со свободным объемом (CRIMPS (каталожный № B0104240), выпускаемой Perkin Elmer) и культивировали в соответствующем аппарате для культивирования с возможностью встряхивания (120 об/мин) при 30°C в течение 24 часов. После окончания культивирования в свободном объеме во флаконе измеряли концентрацию изопрена газовой хроматографией. Значение OD измеряли при 600 нм с использованием спектрофотометра (HITACHI U-2900). Концентрация изопрена и значение OD в различных микробных штаммах после окончания культивирования описаны в таблице 25.
[0150]
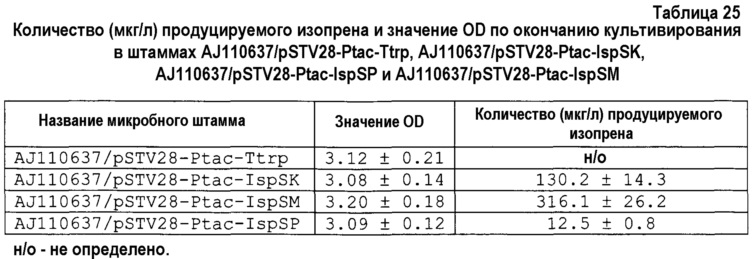
[0151] На основании результатов в таблице 25 количество продуцируемого изопрена увеличивалось в порядке AJ110637/pSTV28-Ptac-IspSM, AJ110637/pSTV28-Ptac-IspSK, AJ110637/pSTV28-Ptac-IspSP и AJ110637/pSTV28-Ptac-Ttrp. На основании указанных выше результатов в штаммах AJ110637 E. aerogenes штамм, в который вводили изопренсинтазу, полученную из мукуны, также обладал наиболее высокой способностью продуцировать изопрен.
[0152] Пример 14: Измерение ферментативной активности изопренсинтазы, полученной из различных растений, с использованием экстракта неочищенного фермента, полученного из Saccharomyces cerevisiae
1) Конструкция плазмиды для экспрессии гена IspS, полученного из различных растений
Плазмиды для экспрессии гена IspSK, гена IspSP и гена IspSM в S. cerevisiae конструировали следующим ниже способом. Проводили ПЦР с полимеразой Prime Star (выпускаемой Takara Bio Inc.) с использованием синтезированных олигонуклеотидов, представленных SEQ ID NO: 82 и 83, в качестве праймеров с pUC57-IspSK в качестве матрицы, синтезированных олигонуклеотидов, представленных SEQ ID NO: 84 и 85, в качестве праймеров с pUC57-IspSP в качестве матрицы или синтезированных олигонуклеотидов, представленных SEQ ID NO: 86 и 87, в качестве праймеров с pUC57-IspSM в качестве матрицы. Реакционный раствор получали в соответствии с рецептурой, прилагаемой к набору, и проводили реакцию в течение 30 циклов 98°C в течение 10 секунд, 55°C в течение 5 секунд и 72°C в течение 120 секунд. В результате получали продукты ПЦР, содержащие ген IspSK, ген IspSP и ген IspSM. В тоже время расщепляли челночный вектор pYES2 для S. cerevisiae и E. coli (Invitrogen, каталожный номер V825-20) ферментом рестрикции KpnI (выпускаемым Takara Bio Inc.). Затем очищенный фрагмент ПЦР, содержащий ген IspSK, лигировали с очищенным pYES2, обработанным KpnI, с использованием набора для клонирования In-fusion HD (выпускаемого Clontech). Полученную плазмиду для экспрессии гена IspSK обозначали как pYES2-IspSK. Аналогично, очищенный фрагмент ПЦР, содержащий ген IspSP, лигировали с очищенным pYES2, обработанным KpnI, с использованием набора для клонирования In-fusion HD (выпускаемого Clontech). Полученную плазмиду для экспрессии гена IspSP обозначали как pYES2-IspSP. Аналогично, очищенный фрагмент ПЦР, содержащий ген IspSM, лигировали с очищенным pYES2, обработанным KpnI, с использованием набора для клонирования In-fusion HD (выпускаемого Clontech). Полученную плазмиду для экспрессии гена IspSM обозначали как pYES2-IspSM.
[0153] 2) Конструкция штамма S288C S. cerevisiae, обладающего способностью продуцировать изопрен
Компетентные клетки S288C S. cerevisiae получали с использованием набора Frozen-EZ Yeast Transformation II (ZYMO RESEARCH, каталожный номер T2001). pYES2, pYES2-IspSK, pYES2-IspSP или pYES2-IspSM вводили в компетентные клетки штамма S288C S. cerevisiae согласно протоколу того же набора. Клетки равномерно наносили на чашку Петри с SD-Ura и культивировали при 30°C в течение 2 суток. Затем из формированных чашек Петри получали трансформанты, которые утрачивали потребность в Ura. Композиция среды SD-Ura является такой, как описано в таблице 26. Штамм, в который вводили pYES2, pYES2-IspSK, pYES2-IspSP или pYES2-IspSM в штамм S288C S. cerevisiae, обозначали как штамм S288C/pYES2, S288C/pYES2-IspSK, S288C/pYES2-IspSP или S288C/pYES2-IspSM, соответственно.
[0154]
Среда SD-Ura
[0155] 3) Способ получения экстракта неочищенного фермента
Микробные клетки штамма S288C/pYES2, S288C/pYES2-IspSK, S288C/pYES2-IspSP или S288C/pYES2-IspSM равномерно наносили на чашку Петри с SD-Ura и культивировали при 30°C в течение 24 часов. Микробные клетки, соответствующие 1/4 формированной чашки Петри, инокулировали в колбу Сакагути, в которую добавляли 50 мл SD-Ura с галактозой (среда SD-Ura, в которой глюкозу заменяли галактозой), и культивировали при 30°C в течение 16 часов. Композиция среды с SD-Ura галактозой представлена в таблице 27. Микробные клетки отделяли от среды для культивирования центрифугированием при 3000 об/мин в течение 5 минут и дважды промывали буфером для изопренсинтазы (50 мМ Tris-HCl, pH 8,0, 20 мМ MgCl2, 5% глицерин). Промытые микробные клетки суспендировали в 1,8 мл того же буфера. Приблизительно 0,9 мл гранул для дезинтеграции (YBG05, диаметр 0,5 мм) и 0,9 мл суспензии микробных клеток помещали в 2 мл пробирку, специальную для устройства для проведения шока с использованием множества гранул, и подвергали микробные клетки дезинтеграции с использованием устройства для проведения шока с использованием множества гранул (модель MB701 (S)), выпускаемого Yasui Kikai Corporation, при 2500 об./мин в течение 12 циклов в положении "Вкл" в течение 60 секунд/в положении "Выкл" в течение 60 секунд.
После дезинтеграции пробирку центрифугировали при 20000×g в течение 20 минут и использовали супернатант в качестве экстракта неочищенного фермента.
[0156]
Среда SD-Ura с галактозой
[0157] 4) Измерение активности изопренсинтазы
Концентрацию белка экстрактов неочищенного фермента из штаммов S288C/pYES2, S288C/pYES2-IspSK, S288C/pYES2-IspSP и S288C/pYES2-IspSM измеряли способом с бицинхониновой кислотой (способ BCA). В качестве реагентов для измерения использовали набор BCA protein assay reagent (Thermo Scientific Japan, каталожный №23227). 0,4 мг экстракта неочищенного фермента в качестве количества общего белка и изопреновый буфер объединяли и получали всего 0,25 мл. Указанное соединение помещали во флакон со свободным объемом, затем в него добавляли 0,0125 мл 0,5M раствора MgCl2 и 0,005 мл 0,2M раствора DMAPP (выпускаемого Cayman, каталожный номер 63180) и аккуратно перемешивали смесь вихревым способом. Затем сразу же флакон плотно закрывали крышкой с бутилкаучуковой мембраной для флакона со свободным объемом (каталожный № B0104240), выпускаемой Perkin Elmer, и проводили реакцию при 37°C в течение 2 часов. После реакции в свободном объеме во флаконе измеряли концентрацию изопрена газовой хроматографией. В качестве флакона со свободным объемом использовали 22 мл флакон CLEAR CRIMP TOP (каталожный № B0104236), выпускаемый Perkin Elmer использовали, и в качестве крышки для герметичного закупоривания использовали крышку с бутилкаучуковой мембраной для флакона со свободным объемом (каталожный № B0104240), выпускаемую Perkin Elmer.
Количества изопрена, образовавшегося в различных микробных штаммах, после реакции в течение 2 часов описаны в таблице 28.
[0158]

[0159] На основании результатов в таблице 28 количество образовавшегося изопрена являлось больше в штамме S288C/pYES2-IspSM по сравнению со штаммом S288C/pYES2-IspSK и меньше в штамме S288C/pYES2-IspSP и штамме S288C/pYES2. На основании указанных выше результатов экстракт неочищенного фермента из штамма, в который вводили изопренсинтазу, полученную из мукуны, обладал наиболее высокой активностью образования изопрена.
[0160] Пример 15: Эффекты введения изопренсинтазы, полученной из различных растений, на штамм S288C Saccharomyces cerevisiae
1) Эффекты введения изопренсинтазы, полученной из различных растений, на штамм S288C S. cerevisiae
Микробные клетки S288C/pYES2, S288C/pYES2-IspSK, S288C/pYES2-IspSP или S288C/pYES2-IspSM равномерно наносили на чашку Петри с YDP и культивировали при 30°C в течение 18-24 часов. Из формированной чашки Петри инокулировали одну петлю для посева микробных клеток в 1 мл среды SD-Ura2 (содержащей по 1 г/л глюкозы и галактозы) во флакон со свободным объемом (22 мл флакон CLEAR CRIMP TOP (каталожный № B0104236), выпускаемый Perkin Elmer), плотно закрывали флакон крышкой с бутилкаучуковой мембраной для флакона со свободным объемом (CRIMPS (каталожный № B0104240), впускаемой Perkin Elmer) и культивировали микробные клетки при 30°C при 120 об/мин в течение 24 часов с использованием подходящего аппарата для культивирования с возможностью встряхивания. Композиции среды YPD и среды SD-Ura2 описаны в таблицах 29 и 30, соответственно.
Композиция среды YPD
[0161]
Композиция среды YPD
Стерилизованная при 120°C в течение 15 минут
[0162]
Среда SD-Ura2
Стерилизованная при 120°C в течение 15 минут
[0163] После окончания культивирования в свободном объеме во флаконе измеряли концентрацию изопрена газовой хроматографией.
Значение OD измеряли при 600 нм с использованием спектрофотометра (HITACHI U-2900). Концентрация в каждом микробном штамме после окончания культивирования описана в таблице 31.
[0164]
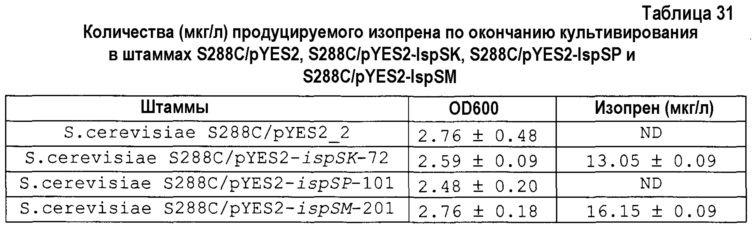
[0165] На основании результатов в таблице 31 количество продуцируемого изопрена являлось больше в S288C/pYES2-IspSM чем в S288C/pYES2-IspSK. Изопрен не детектировали в S288C/pYES2 и S288C/pYES2-IspSP. На основании указанных выше результатов штамм, в который вводили изопренсинтазу, полученную из мукуны, обладал наиболее высокой способностью продуцировать изопрен в штаммах дикого типа.
[0166] Пример 16: Сравнение стабильности изопренсинтазы
1) Конструкция плазмиды для экспрессии
Плазмиду для избыточной экспрессии изопренсинтазы, полученной из мукуны, конструировали следующим ниже способом. Вектор получали ПЦР с использованием синтезированных олигонуклеотидов, представленных SEQ ID NO: 88 и 89, с pCold-TF (выпускаемой Takara, №3365, данные об его последовательности получали из GenBank/EMBL/DDBJ ID доступа AB213654) в качестве матрицы. Вставку получали ПЦР с использованием синтезированных олигонуклеотидов, представленных SEQ ID NO: 90 и 91, с pUC57-IspSM в качестве матрицы. В качестве полимеразы для ПЦР использовали PrimeStar HS (выпускаемую Takara Bio Inc.). Реакционный раствор получали в соответствии с рецептурой, прилагаемой к набору. Реакцию проводили путем повторения цикла 95°C в течение 10 секунд, 55°C в течение 5 секунд и 72°C в течение 6 минут 30 раз. Эти полученные фрагменты ДНК лигировали с использованием набора для клонирования In-Fusion HD (выпускаемого Clontech). Конструируемую плазмиду обозначали как pCold-TF-IspSM. Нуклеотидная последовательность pCold-TF-IspSM представлена SEQ ID NO: 92.
Плазмиду для избыточной экспрессии изопренсинтазы, полученной из тополя, конструировали следующим ниже способом. Вектор получали ПЦР с использованием синтезированных олигонуклеотидов, представленных SEQ ID NO: 88 и 89, с pCold-TF в качестве матрицы. Вставку получали ПЦР с использованием синтезированных олигонуклеотидов, представленных SEQ ID NO: 93 и 94, с pUC57-ispSP в качестве матрицы. В качестве полимеразы для ПЦР использовали PrimeStar HS (выпускаемую Takara Bio Inc.). Реакционный раствор получали в соответствии с рецептурой, прилагаемой к набору. Реакцию проводили путем повторения цикла 95°C в течение 10 секунд, 55°C в течение 5 секунд и 72°C в течение 6 минут 30 раз. Эти полученные фрагменты ДНК лигировали с использованием набора для клонирования In-Fusion HD (выпускаемого Clontech). Конструируемую плазмиду обозначали как pCold-TF-IspSP. Нуклеотидная последовательность pCold-TF-IspSP представлена SEQ ID NO: 95.
Плазмиду для избыточной экспрессии изопренсинтазы, полученной из пуэрарии волосистой, конструировали следующим ниже способом. Вектор получали ПЦР с использованием синтезированных олигонуклеотидов, представленных SEQ ID NO: 88 и 89, с pCold-TF в качестве матрицы. Вставку получали ПЦР с использованием синтезированных олигонуклеотидов, представленных SEQ ID NO: 96 и 97, с pUC57-ispSK в качестве матрицы. В качестве полимеразы для ПЦР использовали PrimeStar HS (выпускаемую Takara Bio Inc.). Реакционный раствор получали в соответствии с рецептурой, прилагаемой к набору. Реакцию проводили путем повторения цикла 95°C в течение 10 секунд, 55°C в течение 5 секунд и 72°C в течение 6 минут 30 раз. Полученные фрагменты ДНК лигировали с использованием набора для клонирования In-Fusion HD (выпускаемого Clontech). Конструируемую плазмиду обозначали как pCold-TF-IspSK. Нуклеотидная последовательность pCold-TF-IspSK представлена SEQ ID NO: 98.
При конструкции экспрессирующих векторов изопренсинтазу конструировали так, чтобы она экспрессировалась в виде белка, слитого с инициирующим фактором (TF) на его N-конце. Слитый белок изопренсинтазы и TF обозначали как TF-IspS. Слитые белки, полученные из мукуны, тополя и пуэрарии волосистой, обозначали как TF-IspSM, TF-IspSP и TF-IspSK, соответственно.
Компетентные клетки BL21 (DE3) E. coli (one shot BL21 (DE3), выпускаемые Life Technologies) трансформировали pCold-TF-IspSM, pCold-TF-IspSP или pCold-TF-IspSK способом теплового шока. После теплового шока при 42°C в течение 30 секунд клетки восстанавливали культивированием в среде SOC при 37°C при 120 об/мин в течение одного часа. Затем все клетки высевали на чашку Петри с LB, содержащей 100 мг/л ампициллина, и культивировали в статических условиях при 37°C в течение 14 часов.
[0167] 2) Культивирование препарата изопренсинтазы в большом количестве
Образовавшуюся колонию накалывали, инокулировали в 5 мл среды LB, содержащей 100 мг/л ампициллина, и культивировали при 37°C при 200 об/мин до получения значения OD600, равного 1,0. После подтверждения, что значение OD равно 1,0, все клетки were инокулировали в 500 мл колбу Сакагути, в которую добавляли 100 мл среды LB, содержащей 100 мг/л ампициллина, и культивировали при 37°C при 200 об/мин до получения значения OD600 равного 1,0. Затем добавляли 1M изопропил-β-тиогалактопиранозид (IPTG) в конечной концентрации 1 мМ и культивировали клетки при 15°C при 100 об/мин в течение ночи. После окончания культивирования микробные клетки собирали центрифугированием при 8000×g в течение 10 минут и хранили при -20°C до проведения очистки.
[0168] 3) Очистка изопренсинтазы
Изопренсинтазу очищали с использованием колонки с His-меткой. Микробные клетки, полученные из 100 мл бульона после окончания культивирования суспендировали в 240 мл дезинтегрирующего буфера, состоящего из 50 мМ фосфатного буфера (pH 8,0) и 500 мМ NaCl, и дезинтегрировали с использованием ультразвуковой машины (Sonifier 250, выпускаемой Baransan) в условиях 50% рабочего цикла и управлением выхода 6 на льду в течение 8 минут. После обработки ультразвуком подвергнутый дезинтеграции супернатант получали центрифугированием при 14000×g в течение 20 минут. Все последующие стадии очистки проводили при 4°C. Гель для никель-аффинной хроматографии His-select (выпускаемый Sigma) соответствующий 2 мл объема слоя вносили в колонку "polyprep" (выпускаемую BioRad) и заполняли гравитационным способом. Затем в колонку вносили 10 мл дезинтегрирующего буфера и уравновешивали гравитационным способом. Общий объем подвергнутого дезинтеграции супернатанта после обработки ультразвуком вносили в колонку и адсорбировали TF-IspS на колонке.
Подтверждали, что общий объем вносимого подвергнутого дезинтеграции супернатанта, проходил через колонку, и затем колонку промывали 10 мл дезинтегрирующего буфера. Затем колонку дополнительно промывали 10 мл буфера для измерения активности, состоящего из 50 мМ Tris-HCl (pH 8,0) и 15 мМ MgCl2. Затем в колонку вносили 4 мл раствора для промывания, состоящего из 50 мМ Tris-HCl (pH 8,0), 15 мМ MgCl2 и 10 мМ имидазола, и удаляли выходящий раствор. В заключении в колонку вносили 4 мл элюента, состоящего из 50 мМ Tris-HCl (pH 8,0), 15 мМ MgCl2 и 200 мМ имидазола, для элюирования TF-IspS. Концентрировали TF-IspS после элюирования на колонке для гель-фильтрации (Amicon Ultra MWCO100k, выпускаемой Millipore).
Затем расщепляли слитый участок слитого белка TF-IspS. Для расщепления использовали фактор Xa (выпускаемый Novagen). Реакционный буфер состоял из 50 мМ Tris-HCl (pH 8,0), 100 мМ NaCl и 5 мМ СаСl2, добавляли 26 ед. фактора Xa и проводили реакцию при 4°C в течение 14 часов. Удаляли TF из реакционного раствора с использования того факта, что один TF адсорбируется на геле для никель-аффинной хроматографии His-select, путем пропускания реакционного раствора после реакции через гель для никель-аффинной хроматографии His-select. Затем концентрировали IspS на колонке для гель-фильтрации (Amicon Ultra MWCO50k, выпускаемой Millipore). Для тестирования чистоты, полученный концентрированный раствор проявляли на NuPAGE от 4 до 12% (выпускаемом Life Technologies). В результате окрашивания проявленного геля CBB не наблюдали полос полученных в результате примесей, отличных от IspS. Таким образом, определяли, что чистота составляет приблизительно 99%.
Полученные таким образом раствор IspSM и раствор IspSK хранили в буфере для измерения активности, в который добавляли глицерин в конечной концентрации 5%, в морозильнике при -80°C до проведения измерения активности. Раствор IspSP хранили в том же буфере, в который добавляли PEG600 в конечной концентрации 5%, в морозильнике при -80°C до проведения измерения активности.
[0169] 4) Измерение активности изопренсинтазы
IspSM, IspSK и IspSP размораживали на люду и удаляли глицерин и PEG600 заменой буфера. Концентрации белков выравнивали колориметрическим количественным определением на SDS-PAGE. 50 мкл изопренового реакционного раствора состояло из 50 мМ Tris-HCl, pH 8,0, 15 мМ MgCl2, 4 мМ DMAPP и 1 мкг IspS. 50 мкл изопренового реакционного раствора помещали в 0,2 мл пробирку для ПЦР (выпускаемую Nippon Genetics Co., Ltd.) и проделывали отверстие в ее крышке. Затем эту пробирку помещали в 22 мл флакон (выпускаемый Perkin Elmer), в который добавляли 450 мкл очищенной воды, и затем немедленно плотно закрывали флакон крышкой с бутилкаучуковой мембраной для флакона со свободным объемом (выпускаемой Perkin Elmer). Реакцию образования изопрена проводили при 40°C в течение 238 часов. Образуемый изопрен количественно определяли способом, описанным в 4-2) и 4-3). Результаты представлены в таблице 32. Как представлено в результатах, демонстрировано, что изопренсинтаза, полученная из мукуны, обладала приблизительно в 10 раз более высокой способностью продуцировать изопрен, чем изопренсинтаза, получаемая из пуэрарии волосистой.
[0170]
Сравнение накопленных количеств изопрена IspS (238 часов, реакция при 40°C)
**Чистота IspS.
[0171] 5) Сравнение стабильности изопренсинтазы
Каждую IspS, очищенную указанным выше способом, хранили в растворе 50 мМ Tris-HCl (pH 8,0) и 15 мМ MgCl2 при 4°C в течение 48 часов. Количество изопрена, продуцируемого ферментативной реакцией каждой из изопренсинтаз до хранения и после него анализировали газовой хроматографией указанным выше способом. Виды остаточной активности сравнивали путем деления количество изопрена, продуцируемого после хранения, на количество изопрена, продуцируемого до хранения. В результате было продемонстрировано, что изопренсинтаза, полученная из мукуны, обладала наиболее превосходной стабильностью, по сравнению с изопренсинтазой, полученной из пуэрарии волосистой, и изопренсинтазой, полученной из тополя.
[0172]
Сравнение остаточной активности каждой IspS (после хранения при 4°C в течение 48 часов)
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТЫ ИЗОПРЕНСИНТАЗЫ, ПРИМЕНЯЕМЫЕ ДЛЯ УЛУЧШЕНИЯ ПРОДУЦИРОВАНИЯ ИЗОПРЕНА МИКРООРГАНИЗМАМИ | 2009 |
|
RU2516343C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПОЛУЧЕНИЯ ИЗОПРЕНА | 2008 |
|
RU2545699C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПОЛУЧЕНИЯ ИЗОПРЕНА, НЕ СОДЕРЖАЩЕГО С5-УГЛЕВОДОРОДОВ В УСЛОВИЯХ НАРУШЕНИЯ ВЗАИМОСВЯЗИ МЕЖДУ ПРОДУЦИРОВАНИЕМ ИЗОПРЕНА И РОСТОМ КЛЕТОК И/ИЛИ В УСЛОВИЯХ ПРОДУЦИРОВАНИЯ ИЗОПРЕНА НА БЕЗОПАСНЫХ РАБОЧИХ УРОВНЯХ | 2009 |
|
RU2563513C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ ПРОДУКТА РЕАКЦИИ, КАТАЛИЗИРУЕМОЙ БЕЛКОМ, ОБЛАДАЮЩИМ АКТИВНОСТЬЮ 2-ОКСОГЛУТАРАТ-ЗАВИСИМОГО ФЕРМЕНТА, И СПОСОБ ПРОДУКЦИИ УКАЗАННОГО ПРОДУКТА | 2008 |
|
RU2444568C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |
| ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ФУМАРОВОЙ КИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ ФУМАРОВОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЭТОГО ШТАММА | 2014 |
|
RU2573936C1 |
| ТОПЛИВНЫЕ КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ПРОИЗВОДНЫЕ ИЗОПРЕНА | 2010 |
|
RU2531623C2 |
| ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ЯНТАРНОЙ КИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ЯНТАРНОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЭТОГО ШТАММА | 2015 |
|
RU2603004C1 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ 1-БУТАНОЛА | 2006 |
|
RU2429295C2 |
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКОЙ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ydiB, СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА НИЗШИХ АЛКИЛОВ АЛЬФА-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИНА | 2007 |
|
RU2355759C1 |
































































































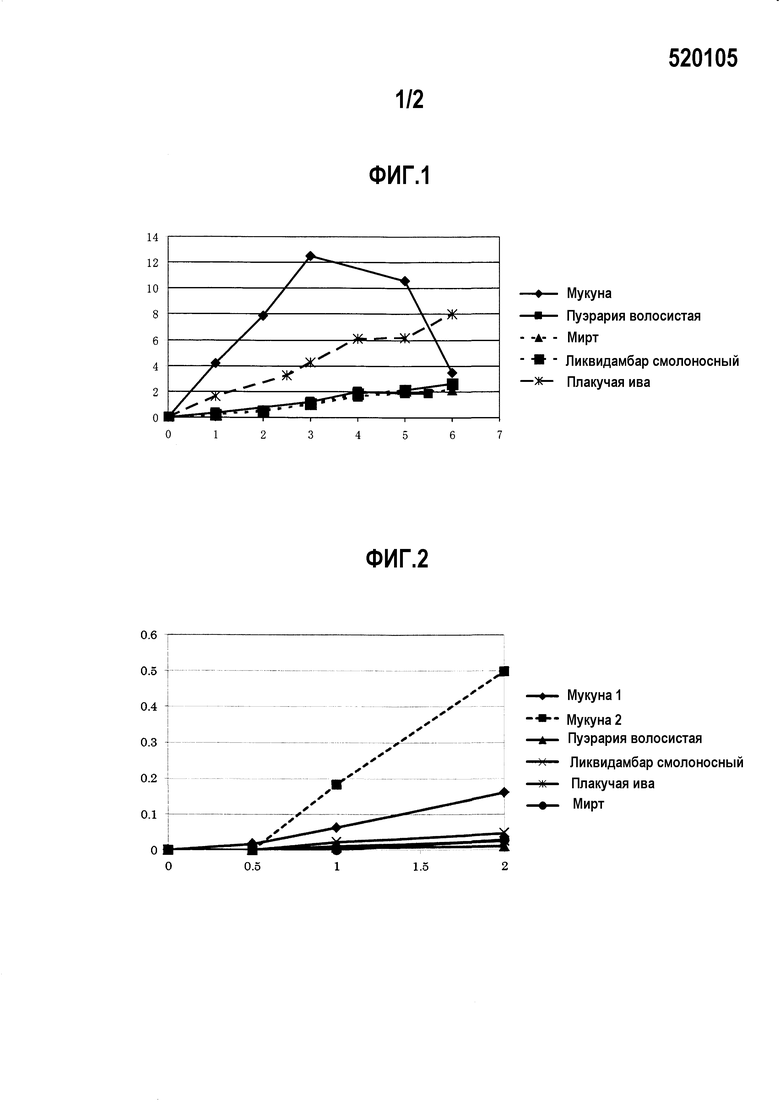
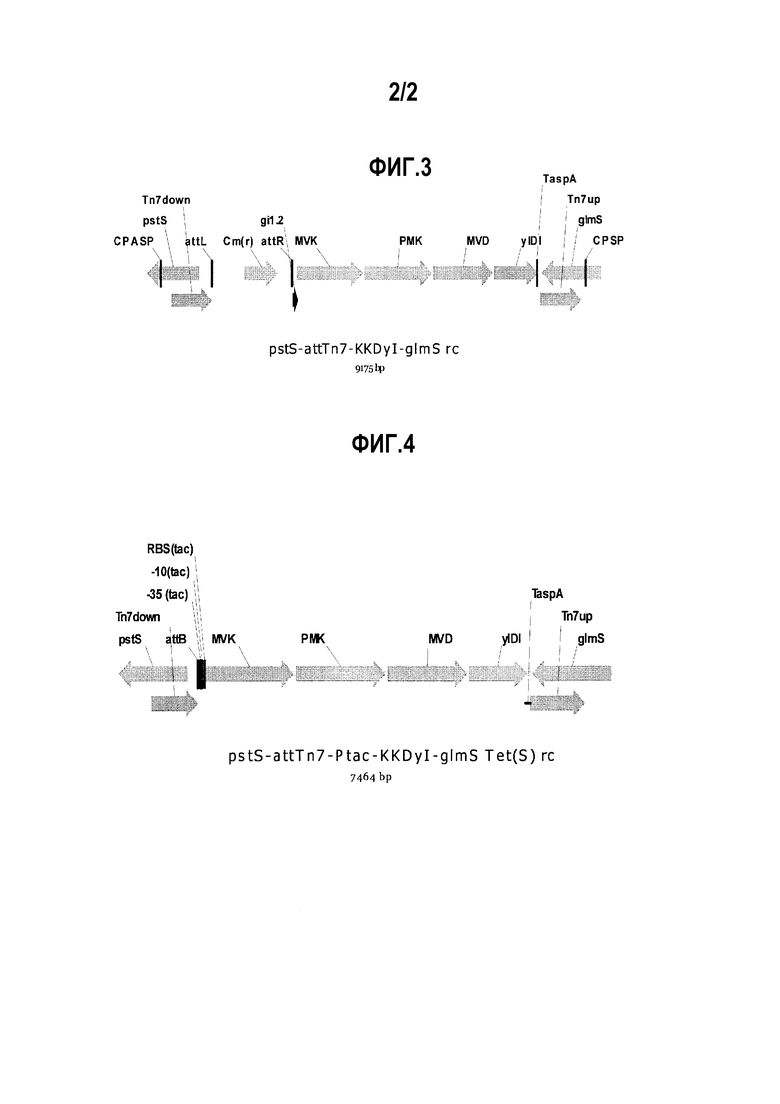
Группа изобретений относится к области биохимии. Предложен полинуклеотид, кодирующий белок, обладающий активностью изопренсинтазы. Предложен белок, обладающий активностью изопренсинтазы. Также предложен экспрессирующий вектор, содержащий указанный полинуклеотид или полинуклеотид, кодирующий указанный белок. Предложена клетка-хозяин для получения изопренсинтазы, содержащая указанный вектор. Предложен способ получения белка, обладающего активностью изопренсинтазы, с использованием указанной клетки-хозяина. Предложен способ получения мономера изопрена из диметилаллилдифосфата в присутствии указанного белка или с использованием указанной клетки-хозяина в среде с диметилаллилдифосфатом. Предложен способ получения изопренового полимера, включающий стадию получения мономера изопрена указанным способом. Группа изобретений позволяет повысить продукцию изопрена в несколько раз по сравнению с продукцией изопрена с использованием изопренсинтазы, полученной из пуэрарии волосистой или из тополя. 8 н. и 6 з.п. ф-лы, 4 ил., 33 табл., 16 пр.
1. Полинуклеотид из следующих (а), (b) или (с):
(a) полинуклеотида, содержащего (i) нуклеотидную последовательность, представленную SEQ ID NO: 1, и кодирующего белок, обладающий активностью изопренсинтазы, или (ii) нуклеотидную последовательность, состоящую из нуклеотидных остатков в положениях 133-1785 в нуклеотидной последовательности, представленной SEQ ID NO: 1, и кодирует белок, обладающий активностью изопренсинтазы;
(b) полинуклеотида, который содержит нуклеотидную последовательность, обладающую 90% или более идентичностью нуклеотидной последовательности (i) или (ii), и кодирует белок, обладающий активностью изопренсинтазы, или
(c) полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности (i) или (ii), и кодирует белок, обладающий активностью изопренсинтазы.
2. Полинуклеотид по п.1, где полинуклеотид получают из мукуны.
3. Белок из следующих (А), (В) или (С):
(A) белка, содержащего (i') полноразмерную аминокислотную последовательность, представленную SEQ ID NO: 2, или (ii') аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 45-594 в аминокислотной последовательности, представленной SEQ ID NO: 2, где указанный белок обладает активностью изопренсинтазы;
(B) белка, который содержит аминокислотную последовательность, обладающую 90% или более идентичностью аминокислотной последовательности (i') или (ii'), и обладает активностью изопренсинтазы, или
(C) белка, который содержит аминокислотную последовательность, содержащую делецию, замену, добавление или вставку от одной до 50 аминокислот в аминокислотной последовательности (i') или (ii'), и обладает активностью изопренсинтазы.
4. Экспрессирующий вектор, содержащий полинуклеотид по п.1 или 2, или полинуклеотид, кодирующий белок по п.3.
5. Клетка-хозяин для получения изопренсинтазы, включающая экспрессирующий вектор по п.4.
6. Клетка-хозяин по п.5, где клетка-хозяин обладает способностью синтезировать диметилаллилдифосфат по метилэритриолфосфатному пути.
7. Клетка-хозяин по п.6, где клетка-хозяин представляет собой Escherichia coli.
8. Клетка-хозяин по любому из пп.5-7, где клетка-хозяин обладает способностью синтезировать диметилаллилдифосфат по мевалонатному пути и метилэритриолфосфатному пути.
9. Клетка-хозяин по п.5, где клетка-хозяин представляет собой микроорганизм, принадлежащий к роду Corynebacterium, роду Pantoea, роду Enterobacter или роду Saccharomyces.
10. Способ получения белка, обладающего активностью изопренсинтазы, включающий культивирование клетки-хозяина по любому из пп.5-9 в среде с получением белка по п. 3.
11. Способ получения мономера изопрена, включающий получение мономера изопрена из диметилаллилдифосфата в присутствии белка по п.3.
12. Способ получения мономера изопрена, включающий культивирование клетки-хозяина по любому из пп.5-9 в среде с получением белка мономера изопрена, где в среде присутствует диметилаллилдифосфат.
13. Способ по п.12, где поступление диметилаллилдифосфата из источника углерода в среду обеспечивают культивированием клетки-хозяина.
14. Способ получения изопренового полимера, включающий (I) и (II):
(I) получение мономера изопрена способом по любому из пп.11-13 и
(II) полимеризацию мономера изопрена с образованием изопренового полимера.
| WO2009076676 A2, 18.06.2009 | |||
| RU2010147659 A, 27.05.2012 | |||
| R | |||
| K | |||
| MONSON et al., Relationships among Isoprene Emission Rate, Photosynthesis, and Isoprene Synthase Activity as Influenced by Temperature, Plant Physiology Vol | |||
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| Музыкальный инструмент | 1924 |
|
SU1175A1 |
| J | |||
| KUZMA et al., Leaf lsoprene Emission Rate 1s Dependent on Leaf Development and the Leve1 of lsoprene Synthase, Plant Physiology Vol | |||
| Приспособление для записи звуковых явлений на светочувствительной поверхности | 1919 |
|
SU101A1 |
| Способ получения твердых неплавких и нерастворимых продуктов уплотнения формальдегида с фонолами | 1925 |
|
SU435A1 |

