1. Область изобретения
Настоящее изобретение относится к модифицированному полинуклеотиду, в котором инициирующий кодон заменен на ATG, к вектору, содержащему этот полинуклеотид, к микроорганизму, содержащему этот полинуклеотид и к способу получения L-лизина с использованием этого полинуклеотида.
2. Предшествующий уровень техники
Штаммы рода Corynebacterium, а именно Corynebacterium glutamicum, представляют собой грамположительные микроорганизмы, которые широко используются для получения L-лизина. L-лизин используется в кормах для животных, в лекарственных средствах и косметических средствах для людей, и его получают путем ферментации штамма Corynebacterium.
Традиционно известен штамм рода Corynebacterium, имеющий улучшенные гены биосинтеза лизина и способ получения L-лизина с использованием этого штамма. Например, в патенте США 6746855 раскрыт способ получения L-лизина путем культивирования Corynebacterium, который имеет повышенную экспрессию гена lysE (ген экспортирующего лизин носителя), и дополнительно введенные гены, выбранные из группы, состоящей из гена dapA, гена lysC, гена рус и гена dapB.
Другой способ заключается в амплификации генов пути биосинтеза лизина для модификации промотора. Например, в корейских выложенных патентных заявках 2009-0082702 и 2009-0084099 раскрыт способ получения L-лизина путем введения улучшенных промоторов ddh и lysC-asd оперона в Corynebacterium. В корейской выложенной патентной заявке 2008-0025355 раскрыт способ улучшения продуцирования лизина путем увеличения числа генов пути биосинтеза лизина, представляющих собой aspB, lysC, asd, dapA, dapB, lysA и рус, в хромосоме.
Между тем, инициирующий кодон, который распознается рибосомами для инициации трансляции на хромосоме, обычно представляет собой ATG. Трансляция может контролироваться, соответственно, инициирующим кодоном гена, и последовательность инициирующего кодона играет важную роль в регуляции активности белка. Однако, в то время как ATG является обычным инициирующим кодоном в генах биосинтеза лизина, происходящих из Corynebacterium glutamicum, инициирующим кодоном генов lysC и рус является GTG, и ген tkt на пентозофосфатном пути, содержит инициирующий кодон TTG (ссылка: J. Biotechnol., 104: 5-
Авторы настоящего изобретения предприняли многочисленные попытки для обнаружения способа улучшения продуцирования лизина и в результате обнаружили, что инициирующий кодон генов дикого типа lysC, tkt и рус может быть заменен на ATG для увеличения активностей аспартат-киназы, транскетолазы и пируваткарбоксилазы по сравнению с их эндогенной активностью, что составляет настоящее изобретение.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Задачей настоящего изобретения является создание модифицированного полинуклеотида, кодирующего аспартаткиназу (ЕС:2.7.2.4; ниже называемую LysC), транскетолазу (ЕС:2.2.1.1; ниже называемую Tkt) или пируваткарбоксилазу (ЕС:6.4.1.1; ниже называемую Рус), где инициирующий кодон полинуклеотида заменен на ATG.
Другая задача настоящего изобретения заключалась в предложении вектора, содержащего один или более модифицированных полинуклеотидов, кодирующих аспартаткиназу, транскетолазу или пируваткарбоксилазу, в которых инициирующий кодон полинуклеотида заменен на ATG
Еще одна задача настоящего изобретения заключается в предложении микроорганизма, в котором один или более ферментов имеют повышенную активность по сравнению с их эндогенной активностью.
Еще одна задача настоящего изобретения заключается в предложении способа получения L-лизина, включающего стадии культивирования микроорганизма и выделения L-лизина из этого культивируемого микроорганизма или культуральной жидкости.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВОПЛОЩЕНИЙ
В одном аспекте настоящего изобретения предложен модифицированный полинуклеотид, кодирующий аспартаткиназу (ЕС:2.7.2.4; ниже называемую LysC), транскетолазу (ЕС:2.2.1.1; ниже называемую Tkt) или пируваткарбоксилазу (ЕС:6.4.1.1; ниже называемую Рус), где инициирующий кодон полинуклеотида замещен на ATG.
Каждый полинуклеотид, кодирующий аспартаткиназу, транскетолазу или пируваткарбоксилазу, также может включать частичную замену, делецию, вставку или добавление в полинуклеотид, при условии, что каждый из них обладает ферментативной активностью и может иметь 70% или более гомологии, в частности 80% или более гомологии, более конкретно 90% или более гомологии, и еще более конкретно 95% или более гомологии, и наиболее конкретно 100% гомологии по отношению к известному полинуклеотиду.
При использовании здесь термин "гомология (гомологичный)" означает степень подобия нуклеотидных последовательностей гена lysC, гена tkt или гена рус дикого типа и нуклеотидных последовательностей соответствующих им модифицированных генов, то есть модифицированного гена lysC, гена tkt или гена рус, где часть полинуклеотидов заменена, делетирована, вставлена или добавлена.
При использовании здесь термин "инициирующий кодон" означает 3 нуклеотида, соответствующих точке начала трансляции, когда кодирующая последовательность мРНК (матричная РНК) транслируется в белок. В общем случае, инициирующие кодоны, обнаруженные в хромосоме микроорганизмов, представляют собой ATG (AUG в РНК), GTG (GUG в РНК) и TTG (UUG в РНК), и они находятся в соотношении 62,5%~66,5%, 23,1%~24,3% и 10,3%~13,2% в соответствии с результатами анализа полной последовательности генома Corynebacterium glutamicum (ссылка: Handbook of Corynebacterium glutamicum, 40p, Lothar Eggeling & Michael Bott, 2005).
Среди генов биосинтеза лизина, происходящих из Corynebacterium, о которых сообщалось до настоящего времени, lysC и рус имеют инициирующий кодон GTG, a tkt имеет инициирующий кодон TTG. Инициирующим кодоном этих генов не является ATG, который рассматривается, как уникальная характеристика Corynebacterium.
Модифицированный полинуклеотид в соответствии с настоящим изобретением характеризуется тем, что инициирующий кодон гена lysC, tkt или рус замещен на ATG, и эти модифицированные полинуклеотиды, имеющие такие замещенные инициирующие кодоны, впервые были модифицированы авторами настоящего изобретения. Более подробно, инициирующий кодон GTG полинуклеотида, кодирующего аспартаткиназу (LysC) или пируваткарбоксилазу (Рус), замещен на ATG, и инициирующий кодон TTG полинуклеотида, кодирующего транскетолазу (Tkt), замещен на ATG в настоящем изобретении. Более конкретно, последовательности генов lysC, tkt и рус представляют собой SEQ ID NO: 13, 14 и 15 соответственно; последовательности генов, имеющих замещенный инициирующий кодон, такой как ATG, представлены в SEQ ID NO: 16, 17 и 18 соответственно.
Замены оснований в кодонах инициации могут быть выполнены любым способом, известным в данной области техники, например посредством сайт-специфического мутагенеза, гомологичной рекомбинации, без ограничения ими.
В другом аспекте настоящего изобретения предлагается вектор, содержащий один или более модифицированных полинуклеотидов из модифицированных полинуклеотидов, кодирующих аспартаткиназу, транскетолазу или пируваткарбоксилазу, в которых инициирующий кодон полинуклеотида замещен на ATG.
Модифицированные полинуклеотиды, имеющие замещенный инициирующий кодон, такой как ATG, и кодирующие аспартаткиназу, транскетолазу или пируваткарбоксилазу, которые находятся в векторе по настоящему изобретению, могут включать те, в которых часть полинуклеотидов заменена, удалена, вставлена или добавлена, при условии, что они обладают ферментативной активностью и могут иметь 70%-ную или более гомологию, в частности 80%-ную или более гомологию, более конкретно 90%-ную или более гомологию, еще более конкретно 95%-ную или более гомологию, и наиболее конкретно 100%-ную гомологию.
Кроме того, модифицированные полинуклеотиды, имеющие замененный инициирующий кодон, такой как ATG, и кодирующие аспартаткиназу, транскетолазу или пируваткарбоксилазу, которые находятся в векторе по настоящему изобретению, также могут включать только часть гена, кодирующего аспартаткиназу, транскетолазу или пируваткарбоксилазу, при условии, что они имеют замещенный инициирующий кодон, такой как ATG.
При использовании здесь термин "вектор" относится к ДНК-конструкции, которая содержит нуклеотидную последовательность, функционально связанную с соответствующей контролирующей последовательностью, для экспрессии целевого гена в подходящем хозяине. Контрольные последовательности могут включать промотор для инициации транскрипции, определенную последовательность оператора для контроля над такой транскрипцией, последовательность, кодирующую подходящий участок связывания рибосомы на мРНК, и последовательность контроля окончания транскрипции и трансляции. Вектор, используемый в настоящем изобретении, не является конкретно ограниченным и может представлять собой любой вектор, известный в данной области, при условии, что он реплицируется в хозяине. Например, вектор может быть плазмидой, фаговой частицей или потенциальной геномной вставкой и конкретно представляет собой pDZ (корейский патент 10-0924065), но не ограничен им. При трансформации подходящего хозяина вектор может реплицироваться или функционировать независимо от генома хозяина, или может быть интегрирован в сам геном.
В частности, получена нуклеотидная последовательность (SEQ ID NO. 13, 14 или 15), содержащая инициирующий кодон гена lysC, tkt или рус, и на основе этой последовательности были синтезированы праймеры, имеющие замещенный инициирующий кодон, такой как ATG. ПЦР выполняли с использованием праймеров и хромосомной ДНК L-лизин-продуцирующего штамма в качестве матрицы, так чтобы получить ДНК, один конец которой замещен на ATG. Таким образом полученный фрагмент ДНК клонировали в вектор с получением конечного рекомбинантного вектора. Более конкретно, в настоящем изобретении были сконструированы векторы pDZ-lysC(ATG), pDZ-tkt(ATG) и pDZ-pyc(ATG) соответственно.
В еще одном аспекте настоящего изобретения предлагается микроорганизм, который включает один или более модифицированных полинуклеотидов, имеющих замененный инициирующий кодон, такой как ATG, и кодирующих ферменты, выбранные из группы, состоящей из аспартаткиназы, транскетолазы и пируваткарбоксилазы, с последующим улучшением уровня трансляции мРНК, транскрибируемой с полинуклеотидов в белки, что приводит к повышению активностей одного или более ферментов по сравнению с их эндогенной активностью. Микроорганизм по настоящему изобретению может иметь повышенные активности аспартаткиназы, транскетолазы или пируваткарбоксилазы при использовании модифицированных полинуклеотидов в комбинации из одного, двух или трех из них, где модифицированные полинуклеотиды кодируют соответствующие ферменты и имеют инициирующий кодон, замененный на ATG.
Для замены инициирующих кодонов целевых генов в хромосоме микроорганизма на ATG можно использовать различные методы, известные в данной области. Например, последовательности инициирующих кодонов эндогенных генов lysC, tkt и рус микроорганизма могут быть заменены в хромосоме. Альтернативно, соответствующие гены, имеющие замененные последовательности инициирующего кодона, могут быть введены в микроорганизм в виде плазмиды.
Микроорганизм может представлять собой любой штамм, при условии, что он продуцирует L-лизин. В частности, это могут быть микроорганизмы из Corynebacterium sp. или Brevibacterium sp. Примеры микроорганизмов Corynebacterium sp. или Brevibacterium sp. включают Corynebacterium glutamicum ATCC13032, Corynebacterium thermoaminogenes FERM BP-1539, Brevibacterium flavum ATCC 14067, Brevibacterium lactofermentum ATCC 13869. Кроме того, могут быть включены L-лизин-продуцирующие варианты или штаммы, происходящие от них, например Corynebacterium glutamicum KССМ11016Р (этот микроорганизм был раскрыт как KFCC10881 и редепонирован международным органом по депонированию в соответствии с Будапештским договором с учетным номером KССМ11016Р, корейский патент 10-0159812, корейский патент 10-0397322) и Corynebacterium glutamicum KFCC 11001. Кроме того, это может быть Corynebacterium glutamicum с учетным номером KССМ11016Р.
В частности, Corynebacterium glutamicum с учетным номером KССМ11016Р был трансформирован вектором, содержащим полинуклеотид, кодирующий аспартаткиназу, транскетолазу или пируваткарбоксилазу, где инициирующий кодон замещен на ATG, с получением рекомбинантного Corynebacterium glutamicum. Более конкретно, вектор pDZ-lysC(ATG), pDZ-tkt(ATG) или pDZ-pyc(ATG) вводили в Corynebacterium glutamicum с учетным номером KССМ11016Р, соответственно, так чтобы получить для каждого рекомбинантный Corynebacterium glutamicum.
Кроме того, микроорганизм может быть трансформирован с помощью вектора, содержащего два или более полинуклеотидов, кодирующих аспартаткиназу, транскетолазу или пируваткарбоксилазу и имеющих такой замененный инициирующий кодон, как ATG. В частности, вектор pDZ-tkt(ATG), содержащий ген, в котором инициирующий кодон заменен на ATG и который кодирует транскетолазу, используют для трансформации Corynebacterium glutamicum KCCM11016P-lysC, где инициирующий кодон GTG гена lysC заменен на ATG, и в результате второго кроссовера инициирующие кодоны lysC и tkt в хромосоме заменены на ATG, так чтобы получить Corynebacterium glutamicum KCCM11016P-lysC-tkt. Кроме того, вектор pDZ-pyc(ATG), содержащий полинуклеотид, у которого инициирующий кодон заменен на ATG и который кодирует пируваткарбоксилазу, использовали для трансформации KCCM11016P-lysC, где инициирующий кодон GTG в lysC замещен на ATG, и в результате второго кроссовера инициирующие кодоны lysC и рус в хромосоме заменены на ATG, так чтобы получить KCCM11016P-lysC-pyc. Кроме того, вектор, pDZ-tkt(ATG) используют для трансформации KCCM11016P-lysC-pyc, как описано выше, где инициирующий кодон GTG в lysC и рус заменен на ATG и, в результате второго кроссовера, инициирующие кодоны lysC, рус и tkt в хромосоме заменены на ATG, так чтобы получить KCCM11016P-lysC-pyc-tkt. Было подтверждено, что инициирующие кодоны lysC, рус и tkt также могут быть замещены на ATG в других лизин-продуцирующих штаммах, принадлежащих к Corynebacterium glutamicum, KFCC10750 (этот микроорганизм раскрыт, как KFCC10750 и редепонирован международным органом по депонированию в соответствии с Будапештским договором с учетным номером KССМ11347Р, корейский патент 10-0073610), KССМ10770Р (корейский патент 10-0924065), CJ3P (Genome Biology 2012, 13:R40) таким же образом. Эти результаты свидетельствуют, что модифицированные полинуклеотиды по настоящему изобретению могут быть стабильно введены в различные штаммы, принадлежащие к Corynebacterium sp., посредством чего увеличивая продуцирование L-лизина.
В частности, трансформант KCCM11016P-lysC-pyc-tkt, который был получен путем введения векторов pDZ-lysC(ATG), pDZ-tkt(ATG) и pDZ-pyc(ATG), включающих полинуклеотиды, кодирующие аспартаткиназу, транскетолазу и пируваткарбоксилазу, в Corynebacterium glutamicum KССМ11016Р, был обозначен как Corynebacterium glutamicum СА01-2059 и депонирован 2 мая, 2011 года, в Корейском центре культур микроорганизмов (ниже сокращенно "KССМ"), который является международным органом по депонированию в соответствии с Будапештским договором, под учетным номером KССМ11188Р. Он был депонирован международным органом по депонированию в соответствии с условиями Будапештского договора.
Микроорганизм по настоящему изобретению характеризуется тем, что инициирующие кодоны генов дикого типа, кодирующих указанные выше ферменты, заменены на ATG и, следовательно, микроорганизм имеет повышенные активности аспартаткиназы, транскетолазы и пируваткарбоксилазы по сравнению с их эндогенными активностями в результате значительного улучшения уровней трансляции мРНК, транскрибируемых с генов lysC, tkt и рус, в белки.
При использовании здесь термин "эндогенная активность" означает активность фермента в природном микроорганизме и, например, активность аспартаткиназы, транскетолазы или пируваткарбоксилазы в природном микроорганизме, принадлежащем к Corynebacterium sp. Термин "повышенная эндогенная активность" означает, что активность дополнительно повышена по сравнению с природным ферментом.
В частности, при сравнении аспартаткиназной активности штамма, в котором инициирующий кодон гена lysC был заменен на ATG, и родительского штамма KССМ11016Р, наблюдалось повышение аспартаткиназной активности 2,73 раза (Таблица 4). Кроме того, при сравнении транскетолазной активности штамма, в котором инициирующий кодон гена tkt был заменен на ATG, и родительского штамма KССМ11016Р, наблюдалась повышение транскетолазной активности в 3,5 раза (Таблица 5). Кроме того, при сравнении пируваткарбоксилазной активности штамма, в котором инициирующий кодон гена рус был заменен на ATG, и родительского штамма KССМ11016Р, наблюдалось повышение пируваткарбоксилазной активности в 1,89 раза (Таблица 6).
Было обнаружено, что продуцирование L-лизина микроорганизмом может быть увеличено путем повышения активности аспартаткиназы, транскетолазы, пируваткарбоксилазы или их комбинации.
В настоящем изобретении измеряли количества L-лизина, продуцируемые в L-лизин-продуцирующих штаммах, Corynebacterium glutamicum KCCM11016P-lysC, KCCM11016P-tkt, KCCM11016P-pyc, KCCM11016P-lysC-tkt, KCCM11016P-lysC-pyc и KCCM11016P-lysC-pyc-tkt, и в результате они показали значительное улучшение в продуцировании L-лизина по сравнению с родительским штаммом KССМ11016Р (Таблица 7). Также, другие лизин-продуцирующие штаммы, принадлежащие к Corynebacterium glutamicum, KFCC10750 (корейский патент 10-0073610), KССМ10770Р (корейский патент 10-0924065) и CJ3P (Genome Biology 2012, 13:R40), в которых инициирующие кодоны lysC, рус и tkt были заменены на ATG, продемонстрировали значительное улучшение продуцирования L-лизина по сравнению с родительским штаммом (Таблицы 8~10). Эти результаты свидетельствуют о том, что микроорганизмы, имеющие один тип модифицированных полинуклеотидов, кодирующих LysC, Tkt или Рус, или имеющие два типа модифицированных полинуклеотидов, кодирующих эти ферменты, или имеющие три типа модифицированных полинуклеотидов, кодирующих эти ферменты, и имеющие такой замененный инициирующий кодон, как ATG, в хромосоме, также демонстрировали значительно улучшенное продуцирование L-лизина по сравнению с микроорганизмом дикого типа, имеющим инициирующие кодоны GTG или TTG.
В еще одном аспекте настоящее изобретение предложен способ получения L-лизина, включающий стадии культивирования микроорганизма, как описано выше; и выделения L-лизина из культивируемых микроорганизмов или культуральной жидкости.
Для культивирования могут быть использованы различные методы получения L-лизина с использованием микроорганизмов, которые широко известны в этой области техники. Культивирование может быть осуществлено в соответствии с широко известным способом, и условия культивирования, включая температуру, время, рН среды и т.д. можно должным образом контролировать. Подробное описание культивирования приведено в следующих документах [Chmiel; Bioprozesstechnik 1. Einfuhrung in die Bioverfahrenstechnik (Gustav Fischer Verlag, Stuttgart, 1991), и Storhas; Bioreaktoren und periphere Einrichtungen (Vieweg Verlag, Braunschweig / Wiesbaden, 1994)]. Кроме того, культивирование может включать периодическую культуру, непрерывную культуру и культуру с подпиткой. В частности, процесс периодического, с подпиткой или с повторной подпиткой культивирования может выполняться в непрерывном режиме, но настоящее изобретение не ограничивается им.
Для использования в культивировании среда должна удовлетворять требованию конкретного использованного штамма. Культуральные среды для микроорганизмов, принадлежащих к Corynebacterium sp, хорошо известны (например Manual of Methods for General Bacteriology. American Society for Bacteriology. Washington D.C., USA, 1981). Используемые источники углерода могут включать сахариды и углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; масла и жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота, линолевая кислота; спирты, такие как глицерин и этанол; и органические кислоты, такие как уксусная кислота. Эти вещества можно использовать по отдельности или в комбинации. Используемые источники азота могут включать пептон, дрожжевой экстракт, мясной бульон, солодовый экстракт, жидкий кукурузный экстракт, соевую муку и мочевину, или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота можно использовать по отдельности или в комбинации. Используемые источники фосфора могут включать гидрофосфат калия, дигидрофосфат калия и соответствующие натриевые соли. Кроме того, культуральная среда может содержать соли металлов, такие как сульфат магния или сульфат железа, необходимые для роста. Наконец, в дополнение к вышеуказанным веществам можно использовать необходимые питательные вещества, такие как аминокислоты и витамины. Кроме того, в культуральную среду могут быть добавлены соответствующие предшественники. Эти вещества могут быть должным образом добавлены в культуру во время культивирования в периодическом или непрерывном режиме.
Значение рН культуральной среды может быть скорректировано с помощью щелочного соединения, такого как гидроксид натрия, гидроксид калия или аммония, или кислотного соединения, такого как фосфорная кислота или серная кислота. Образование пены можно сдерживать с помощью пеногасителя, такого как полигликолевый эфир жирной кислоты. Культуральная среда может храниться в аэробных условиях посредством введения в нее кислорода или кислородсодержащего газа (например воздуха). Температура культивирования обычно находится между 20 и 45°С и, в частности, между 25 и 40°С. Культивирование продолжают вплоть до получения максимального количества L-лизина. Для этого оно может быть закончено в пределах 10-160 часов. После продуцирования L-лизин может выделяться в культуральную среду или может оставаться в клетках.
Кроме того, способ получения L-лизина по настоящему изобретению включает стадию выделения L-лизина из культивируемого микроорганизма или культуральной жидкости. Способ выделения L-лизина из микроорганизмов или культуральной жидкости широко известен в данной области техники. Примеры способа выделения L-лизина могут включать фильтрацию, анионообменную хроматографию, кристаллизацию и ВЭЖХ, но не ограничены ими.
Примеры
Ниже настоящее изобретение будет описано более подробно со ссылкой на Примеры. Однако эти Примеры служат только для иллюстративных целей, и не предполагается ограничивать данное изобретение этими Примерами.
В Примерах были разработаны рекомбинантные векторы для замены инициирующим кодоном ATG инициирующих кодонов GTG или TTG гена lysC, кодирующего аспартаткиназу, гена tkt, кодирующего транскетолазу и гена рус, кодирующего пируваткарбоксилазу, происходящих из лизин-продуцирующего штамма, Corynebacterium glutamicum KССМ11016Р. Вектором трасформировали штамм Corynebacterium glutamicum KССМ11016Р с целью получения штамма, имеющего замененный инициирующий кодон в хромосоме, посредством чего получали штамм, обладающий улучшенным продуцированием лизина.
Штамм Corynebacterium glutamicum KССМ11016Р, используемый в настоящем изобретении, представляет собой штамм, который устойчив к S-(2-аминоэтил)цистеину (ниже называемому АЕС) и является гомосерин-ликовым, полученным посредством искусственной мутации с использованием Corynebacterium glutamicum (ATCC 13032) дикого типа в качестве родительского штамма (раскрытого как KFCC10881. См. корейский патент 10-0159812 и корейский патент 10-0397322). Кроме того, штамм KFCC 10750 представляет собой L-лизин-продуцирующий штамм Corynebacterium glutamicum, который является гомосериновым ауксотрофом и устойчив к аналогу L-лейцина, 4-азалейцину и к антибиотику рифампицину, полученный посредством искусственной мутации (корейский патент 10-0073610), штамм KССМ10770Р, представляет собой L-лизин-продуцирующий штамм, происходящий от KССМ11016Р, который сохраняет две копии 6 типов генов, составляющих путь биосинтеза лизина, в хромосоме (Корейский патент 10-0924065), и штамм CJ3P представляет собой штамм Corynebacterium glutamicum, который обладает L-лизин-продуцированием в результате введения каждой из P458S, V59A и T311I мутаций в три типа генов рус, hom и lysC дикого типа, на основании описания Binder et al. (Genome Biology 2012, 13:R40).
Пример 1: Конструирование рекомбинантного вектора (pDZ-lysC(ATG)). имеющего замененный на ATG инициирующий кодон в lysC, происходящем из Corynebacterium glutamicum и получение штамма, имеющего замененный инициирующий кодон
pDZ (см. корейский патент 10-0924065) использовали в качестве основного вектора для рекомбинантного вектора, который был сконструирован следующим образом.
(1) Конструирование рекомбинантного вектора pDZ-lysC(ATG)
Для того чтобы получить ген lysC, происходящий из Corynebacterium glutamicum, в качестве матрицы использовали хромосомную ДНК из лизин-продуцирующего штамма (Corynebacterium glutamicum KССМ11016Р), полученную с помощью искусственной мутации. На основе генбанка Национального института здоровья США (NIH GenBank) получали нуклеотидную последовательность (SEQ ID NO. 13), содержащую область инициирующего кодона гена lysC (NCBI (Национальный центр биотехнологической информации) учетный номер NC_003450, Ncgl0247) и на основе этой последовательности синтезировали две пары праймеров (Таблица 1, SEQ ID NO: 1~4) для замены инициирующего кодона GTG на ATG.
ПЦР выполняли, используя хромосомную ДНК из KССМ11016Р в качестве матрицы и праймеры из следующей ниже Таблицы 1. Высокоточную ДНК-полимеразу PfuUltra™ (Stratagene) использовали в качестве полимеразы, и при проведении ПЦР выполняли 30 циклов денатурации при 96°С в течение 30 секунд; отжига при 55°С в течение 30 секунд; и полимеризации при 72°С в течение 30 секунд. Два фрагмента ДНК, полученных таким образом, клонировали в вектор pDZ, обработанный рестриктазой Xbal, с использованием ПЦР-набора для клонирования In-Fusion PCR cloning kit (Clontech) и, наконец, конструировали рекомбинантный вектор pDZ-lysC(ATG).
(2) Получение штамма
Таким образом сконструированный вектор pDZ-lysC(ATG) вводили в KССМ11016Р посредством методом электрического импульса (с использованием метода трансформации согласно Appl. Microbiol.Biotechnol. (1999) 52: 541-545), и затем штаммы, у которых ген был встроен в хромосому посредством гомологичной рекомбинации, отбирали на селективной среде, содержащей 25 мг/л канамицина. Успешную хромосомную вставку вектора подтверждал синий цвет колоний на твердой среде, содержащей X-gal (5-бром-4-хлор-3-индолил-(β-D-галактозид). Штамм с первой хромосомной вставкой выращивали в питательной среде (30°С, 8 часов) при покачивании. Затем культивируемый штамм последовательно разводили от 10-4 до 10-10, и разбавленную культуру высевали на твердую среду, содержащую X-gal. Большинство колоний имели синий цвет, в то время как белые колонии также присутствовали на низком уровне. Путем отбора белых колоний, отбирали штаммы, у которых нуклеотидную последовательность в области инициирующего кодона lysC заменяли при помощи второго кроссовера. Нуклеотидная замена инициирующего кодона в выбранном штамме была окончательно подтверждена посредством ПЦР с использованием праймеров SEQ ID NO: 1 и 4 и затем посредством анализа нуклеотидной последовательности сайта-мишени.

Пример 2: Конструирование рекомбинантного вектора (pDZ-tkt(ATG)) с замененным инициирующим кодоном ATG в tkt полученного из Corynebacterium glutamicum и получение штамма, имеющего замененный инициирующий кодон
Имеется два кодона, которые, как ожидается, представляют собой инициирующие кодоны в последовательности гена tkt. Основываясь на расстоянии от RBS (сайт связывания рибосомы) и протеомике, расположенный ниже кодон определяли, как инициирующий кодон в настоящем изобретении.
(1) Конструирование рекомбинантного вектора pDZ-tkt(ATG)
Для получения гена tkt, происходящего из Corynebacterium glutamicum, в качестве матрицы использовали хромосомную ДНК KССМ11016Р. На основе генбанка Национального института здоровья США (NIH GenBank) получали нуклеотидную последовательность (SEQ ID NO. 14), содержащую область инициирующего кодона гена tkt (NCBI, учетный номер NC_003450, Ncgl1512), и на основании этой последовательности синтезировали две пары праймеров (Таблица 2, SEQ ID NO: 5~8) для замены инициирующего кодона TTG на ATG.
ПЦР выполняли, используя хромосомную ДНК из KССМ11016Р в качестве матрицы и праймеры из следующей ниже Таблицы 2 при условиях, используемых в Примере 1-1. Таким образом полученные два фрагмента ДНК клонировали в вектор pDZ, обработанный рестриктазой Xbal, используя ПЦР-набор для клонирования In-Fusion PCR cloning kit (Clontech), и в результате был сконструирован рекомбинантный вектор pDZ-tkt(ATG).
(2) Получение штамма
Таким образом сконструированным вектором pDZ-tkt(ATG) трансформировали лизин-продуцирующий штамм KССМ11016Р аналогично Примеру 1-2, и получали KCCM11016P-tkt, в котором инициирующий кодон tkt в хромосоме был заменен на ATG посредством второго кроссовера. Нуклеотидную замену инициирующего кодона гена окончательно подтверждали с помощью ПЦР, используя праймеры SEQ ID NO: 5 и 8, и затем анализируя нуклеотидную последовательность сайта-мишени.

Пример 3: Конструирование рекомбинантного вектора (pDZ-pyc(ATG)), имеющего замененный инициирующий кодон ATG в рус, происходящем из Corynebacterium glutamicum, и получение штамма, имеющего замененный инициирующий кодон
(1) Конструирование рекомбинантного вектора pDZ-pyc(ATG)
Для получения гена рус, происходящего из Corynebacterium glutamicum, в качестве матрицы использовали хромосомную ДНК KССМ11016Р. На основе генбанка Национального института здоровья США (NIH GenBank) получали нуклеотидную последовательность (SEQ ID NO. 15), содержащую область инициирующего кодона гена рус (NCBI учетный номер NC_003450, Ncgl0659) и на основании этой последовательности синтезировали две пары праймеров (Таблица 3, SEQ ID NO: 9~12) для замены инициирующего кодона GTG на ATG.
ПЦР выполняли, используя хромосомную ДНК KССМ11016Р в качестве матрицы и праймеры из следующей ниже Таблицы 3 в условиях, используемых в Примере 1-1. Два фрагмента ДНК, полученные таким образом, клонировали в вектор pDZ, обработанный рестриктазой Xbal с использованием набора для ПЦР-клонирования In-Fusion PCR cloning kit (Clontech) и в результате конструировали рекомбинантный вектор pDZ-pyc(ATG).
(2) Получение штамма
Сконструированным таким образом вектором pDZ-pyc(ATG) трансформировали лизин-продуцирующий штамм KССМ11016Р аналогично Примеру 1-2 и получали KССМ11016Р-рус, в котором инициирующий кодон рус в хромосоме был заменен на ATG посредством второго кроссовера. Нуклеотидную замену инициирующего кодона этого гена окончательно подтверждали с помощью ПЦР, используя праймеры SEQ ID NO: 9 и 12, и затем анализируя нуклеотидную последовательность сайта-мишени.

Пример 4: Измерение ферментативной активности аспартаткиназы в штамме с замененным инициирующим кодоном ATG в гене lysC
Клетки в экспоненциальной фазе собирали посредством центрифугирования (5,000 об/мин, 15 минут) и промывали три раза 0,1% Tris⋅HCl (рН 8,0) буфером и затем суспендировали в том же буфере до достижения значения мутности 160 при 610 нм. Клетки разрушали в течение 6 минут с использованием шариковой мельницы после добавления стеклянных шариков к суспензии в количестве 1,25 г/1,5 мл суспензии. Супернатант собирали центрифугированием (15000 об/мин, 20 минут) и содержание белка количественно измеряли с помощью метода Брэдфорда (Bradford, М.М 1976. Anal. Biochem. 72:248-254) и использовали в качестве неочищенного белкового раствора для измерения ферментативной активности аспартаткиназы (LysC). Чтобы количественно определить ферментативную активность LysC, добавляли примерно 0,05 мл раствора неочищенного белка к реакционному раствору, содержащему 0,1 М Tris⋅HCl (рН 8,0), 0,01 М хлорида магния (MgCl2), 0,6 М гидроксиламина HCl (рН 7,0), 4 мМ АТР и 0,2 М аспартата для инициации реакции. Смесь оставляли взаимодействовать при 30°С в течение 30 минут и для окончания реакции добавляли стоп-раствор (10% FeCl2, 3,3% ТСА, 0,7 н. HCl). Супернатант собирали посредством центрифугирования и измеряли поглощение при 540 нм. Единицу (U) ферментативной активности LysC определяли как количество нмоль аспартат-гидроксамата, продуцируемого 1 мг белка в течение 1 минуты.
Обнаружено, что штамм KCCM11016P-lysC имеет активность LysC в 2,73-раза выше активности родительского штамма KССМ11016Р (Таблица 4).

Пример 5: Измерение ферментативной активности транскетолазы в штамме с замененным инициирующим кодоном ATG в гене tkt
Клетки в экспоненциальной фазе собирали посредством центрифугирования (5,000 об/мин, 15 минут) и промывали три раза 0,1% Tris⋅HCl (рН 7,5) буфером и затем суспендировали в том же буфере до достижения значения мутности 160 при 610 нм. Клетки разрушали в течение 6 минут, используя шариковую мельницу, после добавления стеклянных шариков к суспензии в количестве 1,25 г/1,5 мл суспензии. Супернатант собирали посредством центрифугирования (15000 об/мин, 20 минут), количественно измеряли содержание белка с помощью метода Брэдфорда (Bradford, М.М 1976. Anal. Biochem. 72:248-254) и использовали в качестве неочищенного белкового раствора для измерения ферментативной активности транскетолазы (Tkt). Для того чтобы количественно определить ферментативную активность Tkt, неочищенный белковый раствор добавляли к реакционному раствору, содержащему 0,1 М Tris⋅HCl (рН7,5), 10 мМ D-R5P, 2 мМ D-Xu5P, 10 мкМ ThDP, 1,2 мМ MgCl2, 100 мкМ NADH, 1 единицу триозофосфатизомеразы, 1 единицу глицерин-3-фосфат-дегидрогеназы на 1 мл для инициирования реакции. Смесь оставляли взаимодействовать при 30°С в течение 20~30 минут, и измеряли поглощение при 340 нм. Единицу (U) Tkt-ферментативной активности определяли, как количество (мг) фермента, катализирующего получение 1 мкмоль глицеральдегид-3-фосфата в течение 1 минуты, и удельную активность определяли, как единицы/мг (Biochem.J. (2004)382, 759-767).
Обнаружено, что штамм KCCM11016P-tkt имел активность Tkt в 3,5-раза выше, чем активность Tkt родительского штамма KССМ11016Р (Таблица 5).

Пример 6: Измерение ферментативной активности пируваткарбоксилазы в штамме с замененным инициирующим кодоном ATG в гене рус
Клетки в экспоненциальной фазе собирали посредством центрифугирования (5000 об/мин, 15 минут) и промывали два раза 50 мМ Tris⋅HCl (рН 6,3) буфером, содержащим 50 мМ хлористого натрия (NaCl) и затем суспендировали в 100 мМ HEPES (рН 7,5) буфере, содержащем 20% глицерина. К суспензии добавляли СТАВ (цетилтриметиламмоний бромид-ЦТАБ) до концентрации 0,3% и оставляли на льду в течение 1 минуты. Клетки собирали центрифугированием (5000 об/мин, 10 минут) и затем суспендировали в 100 мМ Tris⋅HCl (рН 7,3) буфере. Содержание белка количественно измеряли с помощью метода Брэдфорда и использовали в качестве неочищенного белкового раствора для измерения ферментативной активности пируваткарбоксилазы (Рус). Для того чтобы количественно определить ферментативную активность Рус, неочищенный белковый раствор добавляли к реакционному раствору, содержащему 25 мМ NaHCO3, 5 мМ MgCl2, 3 мМ пирувата и 4 мМ АТР для инициирования реакции. Смесь оставляли взаимодействовать при 30°С в течение 1,5 минут, и для прекращения реакции добавляли 80 мкл стоп-раствора (30% орто-фосфорной кислоты). Супернатант собирали посредством центрифугирования (12000 об/мин, 15 мин, 4°С). На 1 мл добавляли 50 мкл супернатанта, 150 мМ Tris⋅HCl (рН 7,8), 150 мкМ NADH и 2,5 Ед лактатдегидрогеназы, и поглощение при 340 нм измеряли при 37°С. Единицу (U) ферментативной активности Рус определяли как количество нмоль лактата, продуцируемого 1 мг белка за 1 минуту.
Было замечено, что штамм KССМ11016Р имеет активность Рус в 1,89-раз выше, чем активность Рус родительского штамма KССМ11016Р (Таблица 6).

Пример 7: Создание происходящих от KССМ11016Р штаммов с замененными инициирующими кодонами ATG в двух или более генах lysC, tkt и рус
Так как использовались рекомбинантные векторы pDZ-lysC(ATG) pDZ-tkt(ATG) pDZ-pyc(ATG), полученные в Примерах 1, 2 и 3, процесс получения был следующим.
Вектор pDZ-tkt(ATG) использовали для трансформации KССМ11016Р-lysC из Примера 1, где инициирующий кодон GTG гена lysC заменяли на ATG, и в результате второго кроссовера инициирующие кодоны lysC и tkt в хромосоме были заменены на ATG так, чтобы получить KCCM11016P-lysC-tkt.
Нуклеотидная замена инициирующего кодона гена была окончательно подтверждена при помощи ПЦР с использованием праймеров SEQ ID NO: 5 и 8 и затем посредством анализа нуклеотидной последовательности сайта-мишени.
Так же как в Примере 1, вектором DZ-pyc(ATG) трансформировали KCCM11016P-lysC из Примера 1, где инициирующий кодон GTG гена lysC был заменен на ATG и в результате второго кроссовера инициирующие кодоны lysC и рус в хромосоме были заменены на ATG, так чтобы получить KССМ11016Р lysC-pyc. Нуклеотидная замена инициирующего кодона гена была окончательно подтверждена посредством ПЦР с использованием праймеров SEQ ID NO: 9 и 12 и затем путем анализа нуклеотидной последовательности сайта-мишени.
Вектор pDZ-tkt(ATG) был введен для трансформации в KССМ11016Р-lysC-pyc настоящего Примера, где инициирующий кодон GTG генов lysC и рус был заменен на ATG, и в результате второго кроссовера инициирующие кодоны lysC, рус и tkt в хромосоме были заменены на ATG так, чтобы получить KCCM11016P-lysC-pyc-tkt. Нуклеотидная замена инициирующего кодона гена была окончательно подтверждена при помощи ПЦР с использованием праймеров SEQ ID NO: 5 и 8 и затем путем анализа нуклеотидной последовательности сайта-мишени.
Вышеуказанные комбинации генов представлены только в иллюстративных целях, без ограничения ими объема комбинаций генов.
Пример 8: Продуцирование лизина в штамме с замененным инициирующим кодоном ATG
KCCM11016P-lysC, KCCM11intended016P-tkt, KССМ11016Р-рус, KCCM11016P-lysC-tkt, KCCM11016P-lysC-pyc, KCCM11016P-lysC-pyc-tkt, окончательно полученные в Примерах 1, 2, 3 и 7, культивировали с целью продуцирования L-лизина следующим способом.
Родительский штамм KССМ11016Р и KCCM11016P-lysC, KCCM11016P-tkt, KССМ11016Р-рус, KCCM11016P-lysC-tkt, KCCM11016P-lysC-pyc, KCCM11016P-lysC-pyc-tkt инокулировали в соответствующие 250 мл колбы с угловыми перегородками, содержащие 25 мл среды для посева, описанной ниже, и полученный продукт культивировали при 30°С при встряхивании (200 об/мин) в течение 20 часов. 1 мл полученной посевной культуральной жидкости инокулировали в 250 мл колбу с угловой перегородкой, содержащую 24 мл среды для продуцирования, описанной ниже, и культивировали при 30°С при встряхивании (200 об/мин) в течение 120 часов.
После завершения культивирования количество произведенного L-лизина измеряли с помощью ВЭЖХ. Результаты измерения L-лизина в культуральных жидкостях KССМ11016Р и KCCM11016P-lysC, KCCM11016P-tkt, KССМ11016Р-рус, KCCM11016P-lysC-tkt, KCCM11016P-lysC-pyc, KCCM11016P-lysC-pyc-tkt показаны в следующей Таблице 7.

Среда для посева (рН 7,0)
20 г сахара-сырца, 10 г пептона, 5 г дрожжевого экстракта, 1,5 г мочевины, 4 г KH2PO4, 8 г K2НРO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина-HCl, 2000 мкг пантотената кальция, 2000 мкг никотинамида (в 1 литре дистиллированной воды)
Среда для продуцирования (рН 7,0)
100 г глюкозы, 40 г (NH4)2SO4, 2,5 г соевого белка, 5 г твердой фазы кукурузного экстракта, 3 г мочевины, 1 г KH2PO4, 0,5 г MgSO4⋅7H2O, 100 мкг биотина, 1000 мкг тиамина-HCl, 2000 мкг пантотената кальция, 3000 мкг никотинамида, 30 г СаСО3 (в 1 л дистиллированной воды).
Как показано в Таблице 7, было обнаружено, что Corynebacterium glutamicum KCCM11016P-lysC, KCCM11016P-tkt, KCCM11016P-pyc, KCCM11016P-lysC-tkt и KCCM11016P-lysC-pyc с замененными инициирующими кодонами ATG демонстрировали 4~9% увеличение продуцирования L-лизина по сравнению с родительским штаммом KССМ11016Р. В частности, было установлено, что KCCM11016P-lysC-pyc-tkt со всеми тремя введенными генами демонстрировал увеличение продуцирования L-лизина на 12% по сравнению с родительским штаммом KССМ11016Р.
Пример 9: Создание KFСС10750-полученного штамма с замененными инициирующими кодонами ATG в генах lysC, tkt и рус и сравнение продуцирования лизина
Для того чтобы исследовать, влияет ли замена инициирующих кодонов на ATG в генах lysC, рус и tkt на продуцирование лизина также и в других лизин-продуцирующих штаммах, принадлежащих к Corynebacterium glutamicum, в L-лизин-продуцирующий штамм Corynebacterium glutamicum KFCC10750 (корейский патент 10-0073610) вводили все три гена, которые демонстрировали самое существенное влияние на увеличение продуцирования лизина в Примере 8, так чтобы получить рекомбинантный штамм, который был назван KFCC10750-lysC-pyc-tkt. Штамм культивировали так же, как в Примере 8, и определяли концентрацию L-лизина. (Таблица 8).
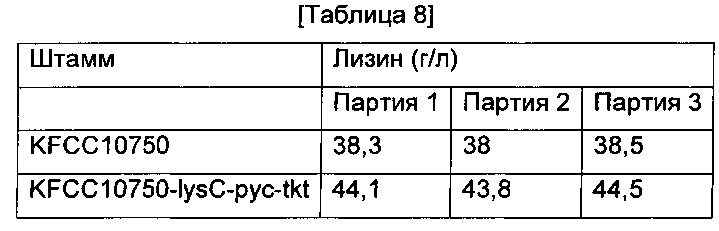
Как показано в Таблице 8, Corynebacterium glutamicum KFCC10750-lysC-pyc-tkt со всеми тремя введенными генами демонстрировал 15%-ное увеличение продуцирования лизина по сравнению с родительским штаммом KFCC10750.
Пример 10: Создание происходящего от KССМ10770Р штамма с замененными инициирующими кодонами ATG в генах lysC, tkt, рус и сравнение продуцирования лизина
В другой L-лизин-продуцирующий штамм Corynebacterium glutamicum KССМ10770Р (корейский патент 10-0924065) вводили все три гена, показавшие самое существенное влияние на увеличение продуцирования лизина в Примере 8, так чтобы получить рекомбинантный штамм, который был назван KCCM10770P-lysC-pyc-tkt. Штамм культивировали аналогично Примеру 8 и определяли концентрацию L-лизина (Таблица 9).
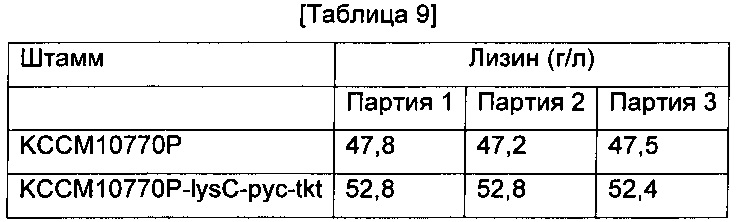
Как показано в Таблице 9, Corynebacterium glutamicum KCCM10770P-lysC-pyc-tkt со всеми тремя введенными генами демонстрирует 11%-ное увеличение продуцирования лизина по сравнению с родительским штаммом KССМ10770Р.
Пример 11: Создание происходящего от CJ3P штамма с замененными инициирующими кодонами ATG в генах lysC, tkt, рус и сравнение продуцирования лизина
В еще один L-лизин-продуцирующий штамм Corynebacterium glutamicum CJ3P (Binder et al. Genome Biology 2012, 13:R40) были введены все три гена, продемонстрировавшие наиболее существенное влияние на увеличение продуцирования лизина в Примере 8, так чтобы получить рекомбинантный штамм, который был назван CJ3P-lysC-pyc-tkt. Этот штамм культивировали так же, как в Примере 8, и определяли концентрацию L-лизина (Таблица 10).
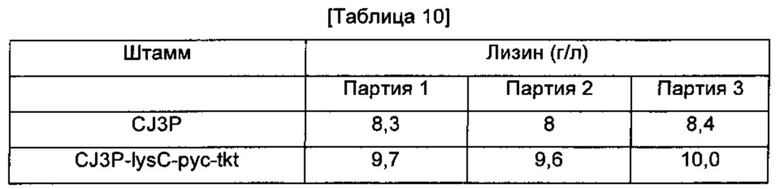
Как показано в Таблице 10, Corynebacterium glutamicum CJ3P-lysC-pyc-tkt со всеми тремя введенными генами демонстрировал 18%-ное увеличение продуцирования лизина по сравнению с родительским штаммом CJ3P.
Результат изобретения
В настоящем изобретении предложен микроорганизм Corynebacterium sp., обладающий улучшенным продуцированием L-лизина, где инициирующие кодоны одного или более генов, кодирующих аспартаткиназу, транскетолазу или пируваткарбоксилазу, заменены с целью увеличения активности соответствующих ферментов по сравнению с их эндогенной активностью в природном микроорганизме.














Группа изобретений относится к области биотехнологии. Представлен модифицированный полинуклеотид, кодирующий транскетолазу (Tkt), где инициирующий кодон полинуклеотида заменен на ATG. Представлен вектор экспрессии, содержащий указанный модифицированный полинуклеотид. Представлен модифицированный микроорганизм рода Corynebacterium для продукции L-лизина, который обладает повышенной активностью транскетолазы по сравнению с ее эндогенной активностью, где инициирующий кодон полинуклеотида, кодирующего транскетолазу, заменен на ATG. Группа изобретений позволяет получать L-лизин с высокой продуктивностью. 4 н. и 5 з.п. ф-лы, 10 табл., 11 пр.
1. Модифицированный полинуклеотид, кодирующий транскетолазу (Tkt), где инициирующий кодон полинуклеотида заменен на ATG.
2. Модифицированный полинуклеотид по п. 1, где инициирующий кодон полинуклеотида, кодирующего транскетолазу (Tkt), представляет собой TTG.
3. Модифицированный полинуклеотид по п. 1, который представлен нуклеотидной последовательностью SEQ ID NO: 17.
4. Вектор экспрессии, содержащий модифицированный полинуклеотид по п. 1.
5. Модифицированный микроорганизм рода Corynebacterium для продукции L-лизина, который обладает повышенной активностью транскетолазы по сравнению с ее эндогенной активностью, где инициирующий кодон полинуклеотида, кодирующего транскетолазу, заменен на ATG.
6. Модифицированный микроорганизм по п. 5, который дополнительно обладает повышенной активностью пируваткарбоксилазы по сравнению с ее эндогенной активностью, и инициирующий кодон полинуклеотида, кодирующего пируваткарбоксилазу, заменен на ATG.
7. Модифицированный микроорганизм по п. 5, трансформированный вектором экспрессии по п. 4.
8. Модифицированный микроорганизм по п. 5, где микроорганизм представляет собой Corynebacterium glutamicum.
9. Способ получения L-лизина, включающий стадии культивирования модифицированного микроорганизма по любому из пп. 5-8; и выделения L-лизина из культивируемого микроорганизма или культуральной жидкости.
| Теплообменник | 1984 |
|
SU1179084A2 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| SILBERBACH M., et al | |||
| Adaptation of Corynebacterium glutamicum to Ammonium Limitation: a Global Analysis Using Transcriptome and Proteome Techniques | |||
| APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Vol | |||
| Контрольный стрелочный замок | 1920 |
|
SU71A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| РЕОСТАТ НАКАЛА ДЛЯ КАТОДНЫХ ЛАМП | 1925 |
|
SU2391A1 |
| RU 2001108535 A, 20.06.2003. | |||

