Область техники изобретения
Настоящее изобретение относится к области модификации гликозилирования белков. Более конкретно, настоящее изобретение относится к молекулам нуклеиновой кислоты, включая конструкции слияния, обладающие каталитической активностью, и их применение для модификации гликозилирования в клетке-хозяине с целью получения полипептидов с улучшенными терапевтическими свойствами, включая антитела с повышенной аффинностью связывания Fc-рецептора и повышенной эффекторной функцией.
Уровень техники
Гликопротеины опосредуют многие важнейшие функции в организме человека, других эукариотических организмах и у некоторых прокариот, включая каталитиз и передачу сигналов, межклеточные взаимодействия, распознавание молекул и их ассоциацию. Они составляют большинство нецитозольных белков в эукариотических организмах (Lis et al, Eur. J Biochem. 218:1-27 (1993)). Многие гликопротеины были использованы в терапевтических целях, и в течение двух последних десятилетий основным продуктом биотехнологического производства были рекомбинантные варианты встречающихся в природе, секретируемых, гликопротеинов. Примеры включают: эритропоэтин (ЭП), терапевтические моноклональные антитела (терапевтические МКАТ, mAb), тканевой активатор плазминогена (TAII), интерферон-бета (ИФН-β), фактор, стимулирующий колонии гранулоцитов/макрофагов (GM-CSF), хорионический гонадотропин человека (ХГч) (Cumming et al., Glycobiology 1:115-130 (1991)).
Олигосахаридный компонент может существенно влиять на свойства, имеющие отношение к эффективности терапевтического гликопротеина, включая физическую стабильность, устойчивость к действию протеаз, взаимодействие с иммунной системой, фармакокинетику и специфическую биологическую активность. Такие свойства могут зависеть не только от присутствия или отсутствия олигосахаридов, но и от их специфической структуры. Можно провести параллели между структурой олигосахаридов и функцией гликопротеина. Например, определенные олигосахаридные структуры
опосредуют, путем взаимодействия со специфическими углевод-связывающими белками, быстрое выведение (клиренс) гликопротеина из кровяного русла, в то время как другие могут быть связаны антителами и запускать нежелательные иммунные реакции (Jenkins et al., NatureBiotechnol 14:975-81 (1996)).
Клетки млекопитающих являются предпочтительными хозяевами для получения терапевтических гликопротеинов благодаря своей способности гликозилировать белки наиболее подходящим для последующего введения в организм человека образом (Cumming et al., Glycobiology 1:115-30 (1991); Jenkins et al, Nature Biotechnol. 14:975-81 (1996)). Бактерии чрезвычайно редко гликозилируют белки и, подобно другим типам обычных клеток-хозяев, таких как дрожжи, мицелиальные грибы, клетки насекомых и растений, создают паттерны гликозилирования, ассоциированные с быстрым клиренсом из кровяного русла, нежелательными иммунными взаимодействиями и, в некоторых специфических случаях, со снижением биологической активности. В течение двух последних десятилетий наиболее часто используемыми клетками млекопитающих были клетки яичников китайского хомячка (СНО). В дополнение к подходящим паттернам гликозилирования, использование этих клеток позволяет неизменно получать генетически стабильные, высокопродуктивные линии клеточных клонов. Их культуры могут быть доведены до высоких плотностей в простых биореакторах (культиваторах) с использованием бессывороточной среды, при этом возможна разработка безопасного воспроизводимого биотехнологического процесса. Другие обычно используемые клетки включают клетки почек хомячка (ВНК), клетки миеломы мыши NSO и SP2/0. Более поздние исследования относятся к получению гликопротеинов с использованием трансгенных животных (Jenkins et al, NatureBiotechnol. 14:975-81 (1996)).
Все антитела содержат углеводные структуры в консервативных положениях константных областей тяжелой цепи, причем каждый изотип (класс) обладает особым набором N-связанных (присоединенных к остатку аспарагина) углеводных структур, который вариабельно влияет на сборку, секрецию или функциональную активность белка (Wright A., Morrison, S.L., Trends Biotech. 15:26-32 (1997)). Структура N-связанного углевода, связанного с антителом, может значительно варьироваться в зависимости от степени процессинга и может включать как сильноразветвленные олигосахариды с высоким содержанием маннозы, так и «двухантенные» сложные олигосахариды (Wright, А., Morrison, S.L., Trends Biotech. 15:26-32 (1997). Обычно имеет место гетерогенный процессинг коровых (каркасных) олигосахаридных структур, прикрепленных в определенном сайте гликозилирования, что приводит к тому, что даже моноклональные антитела существуют в виде множественных гликоформ. Также было показано, что имеют место существенные отличия в гликозилировании между различными клеточными линиями, и незначительные - между клетками одной линии, культивируемыми в разных условиях (Lifely, M.R. et al., Glycobiology 5 (8): 813-22 (1995)).
Неконъюгированные моноклональные антитела (mAbs, МКАТ) могут быть полезны в качестве лекарств для лечения рака, что подтверждается одобрением Администрацией по контролю за продуктами питания и лекарствами США (FDA) применения Rituximab'a (Rituxan™; IDEC Pharmaceuticals, Сан-Диего, Калифорния, и GenentechInc, Сан-Франциско, Калифорния), для лечения CD20-положительной В-клеточной низкозлокачественной или фолликулярной лимфомы, отличной от лимфомы Ходжкина, Trastuzumab'a (Herceptin; GenentechInc,) для лечения запущенного рака груди (Grillo-Lopez, A.-J., et al., Semis. Oncol. 26:66-73 (1999); Goldenberg, M.M., Clin. Ther. 21:309-18 (1999)), Gemtuzumab'a (Mylotarg™, Celltech/Wyeth - Ayerst) для лечения рецидивирующей острой миелоидной лейкемии, и Alemtuzumab'a (САМРАТН™, MilleniumPharmaceuticals/Schering AG) для лечения В-клеточной хронической лимфоцитарной лейкемии. Успех этих продуктов является результатом не только их эффективности, но и их выдающейся безопасностью (Grillo-Lopez, A.-J., et al, Semin. Oncol. 26: 66-73 (1999); Goldenberg, M.M., Clin. Ther. 21:309-18 (1999)). Несмотря на достижения этих лекарственных средств, в настоящее время существует заинтересованность в получении более высокой специфической активности антител, чем та, которую обычно можно достичь при помощи терапии с использованием неконъюгированных МКАТ.
Один из способов достигнуть повышения эффективности, сохранив при этом простоту процесса получения и, если возможно, избежав значительных нежелательных побочных эффектов, состоит в том, чтобы усилить естественные опосредуемые клетками эффекторные функции МКАТ путем модификации их олигосахаридного компонента (Umana, P. et al, Nature Biotechnol 17:176-180 (1999)). Антитела типа IgG1, наиболее часто применяемые в иммунотерапии рака, представляют собой гликопротеины, которые содержат консервативный сайт N-гликозилирования в положении Asn297 каждого из доменов СН2. Сложные «двуантенные» олигосахариды, присоединенные к Asn297, скрыты между двумя доменами СН2, образуя множественные контакты с белковым каркасом. Их присутствие необходимо для осуществления эффекторной функции антитела, такой как антителозависимая клеточная цитотоксичность (АЗКЦ) (Lifely, М.R., et al., number 4 Glycobiology 5:813-822 (1995); Jefferis, R., et al, Immunol Rev. 163:59-76 (1998); Wright, A., Morrison, S.L., Trends Biotechnol 15:26-32 (1997)).
Авторами настоящего изобретения было ранее показано, что повышенная экспрессия бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII), гликозилтрансферазы, катализирующей образование разветвленных (bisected) олигосахаридов, in vitro значительно увеличивает активность АЗКЦ антинейробластомного химерного моноклонального антитела (chCE7), продуцируемого модифицированными клетками СНО (См. Umana, P. et al, Nature Biotechnol. 17:176-180 (1999), Международная публикация No. WO 99/54342, полное содержание каждого из источников полностью включено в описание изобретения путем ссылки). Антитело chCE7 принадлежит к обширному классу неконъюгированных МКАТ, которые обладают высокой аффинностью и специфичностью к опухолям; однако эффективность этих антител при продуцировании их стандартными промышленными клеточными линиями, дефицитными по ферменту GnTIII, слишком низка для клинического применения (Umana, Р., et al., Nature Biotechnol. 17:176-180 (1999)). Это исследование впервые показало, что сильное повышение активности АЗКЦ может быть достигнуто путем модификации продуцирующих антитела клеток для экспрессии GnTIII, которая также приводит к увеличению доли связанных с константной (Fc) областью разветвленных (bisected) олигосахаридов (включая разветвленные нефукозилированные олигосахариды) по сравнению с уровнем, обнаруженным у встречающихся в природе антител.
Результаты ряда исследований позволяют предположить, что Fc-рецептор-зависимые механизмы вносят существенный вклад в действие противоопухолевых цитотоксических антител, а также показывают, что оптимальное противоопухолевое антитело должно предпочтительно связываться с активирующим Fc-рецептором и как минимум - с ингибирующим партнером FcγRIIB (Clynes, R.A., et al., Nature Medicine 6(4):443-446 (2000)); Kalergis, A.M., Ravetch, J.V., J. Exp. Med. 195(12):1653-1659 (Июнь 2002)). Например, результаты по меньшей мере одного исследования позволяют предположить, что, в частности, рецептор FcγRIIIa имеет сильное влияние на эффективность лечения антителами (Cartron, G., et al., Blood 99(3): 754-757 (Февраль 2002)). Это исследование показало, что пациенты, гомозиготные по (гену) FcγRIIIa, лучше реагируют на Rituximab, чем гетерозиготные пациенты. Авторы пришли к заключению, что повышенный ответ был результатом лучшего связывания антитела с FcγRIIIa in vivo, которое привело к лучшей активности АЗКЦ против клеток лимфомы (Cartron, G., et al., Blood 99(3):754-757 (Февраль 2002)).
Кроме АЗКЦ, успешные противораковые антитела часто индуцируют Fc-независимый прямой сигнальный механизм, который определяет выживание, пролиферацию или смерть клетки-мишени через активацию сигнальных каскадов клетки или блокирования доступа к факторам роста (Selenko, N., et al, J Clin. Immunol. 22(3): 124-130 (2002)). Например, было показано, что лечение CD20+ В-клеток Rituximab'ом индуцирует, помимо АЗКЦ, комплемент опосредованный лизис и Mab-индуцируемую индукцию апоптоза (Selenko, N., et al., J. Clin. Immunol. 22(3): 124-130 (2002)). Более того, индуцированный Rituximab’ом апоптоз клеток лимфомы не только убивает клетки, но и способствует захвату и кросс-презентации белков - производных клеток лимфомы антиген-презентирующими дендритными клетками, индуцирует созревание дендритных клеток и делает возможным появление специфических цитотоксических Т-лимфоцитов (ЦТЛ).
Краткое описание изобретения
Принимая во внимание огромный потенциал антител с повышенной аффинностью связывания Fc-рецептора и повышенной эффекторной функцией, авторы настоящего изобретения разработали способ получения таких антител, который включает модификацию профиля гликозилирования Fc-области антитела.
Настоящее изобретение предусматривает, в общих чертах, на создание способа гликомодификации клетки-хозяина с целью изменения профиля гликозилирования одного или более полипептидов, продуцируемых этой клеткой-хозяином. Способы изобретения возможно применять для получения терапевтических антител с измененным гликозилированием в Fc-области, включая сниженное фукозилирование (перенос на олигосахарид остатков фукозы), где, в результате модификации гликозилирования, антитела обладают повышенной эффекторной функцией и/или повышенным связыванием Fc-рецептора. Гликомодифицированные антитела настоящего изобретения особенно полезны в терапевтическом лечении рака у пациентов. В одном воплощении клетку-хозяин данного изобретения подвергают модификации гликозилирования посредством экспрессии молекулы нуклеиновой кислоты, кодирующей полипептид с каталитической активностью GnTIII либо GalT. В предпочтительном воплощении конструкты слияния коэкспрессируются совместно с молекулой нуклеиновой кислоты, кодирующей полипептид, обладающий каталитической активностью ManII и/или с молекулой нуклеиновой кислоты, кодирующей полипептид с каталитической активностью GnTII. В другом воплощении гликомодифицированные полипептиды настоящего изобретения продуцируются клеткой-хозяином, которая была модифицирована для повышения уровня экспрессии молекулы нуклеиновой кислоты, кодирующей полипептид с каталитической активностью ManII.
В другом аспекте настоящее изобретение предусматривает клетку-хозяина, содержащую описанный выше вектор экспрессии, содержащий изолированную нуклеиновую кислоту, содержащую последовательность, кодирующую рекомбинантный полипептид слияния, где указанный полипептид слияния обладает активностью бета-1,4-галактозилрансферазы и содержит отвечающий за локализацию в комплексе Гольджи домен какого-либо полипептида-резидента комплекса Гольджи. В одном воплощении вектор экспрессии кодирует полипептид слияния, содержащий каталитический домен бета-1,4-галактозилрансферазы III, а домен, ответственный за локализацию в комплексе Гольджи выбирают из группы, состоящей из домена локализации маннозидазы II, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы I, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы II, домена локализации маннозидазы I и домена локализации альфа 1-6 коровой фукозилтрансферазы или нуклеиновой кислотой, кодирующей полипептид слияния, содержащей каталитический домен бета-1,4-N-ацетилглюкозаминилтрансферазы III или бета-1,4-галактозилтрансферазы и отвечающий за локализацию в комплексе Гольджи домен гетерологического полипептида-резидента комплекса Гольджи.
В другом аспекте настоящее изобретение предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей рекомбинантный белок слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III («GnTIII»), в количестве, достаточном для модификации олигосахаридов в Fc-полипептида полипептида, продуцируемого клеткой-хозяином, где указанный полипептид, продуцируемый указанной клеткой-хозяином, выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела, содержащего Fc-область и белка слияния, содержащего область, эквивалентную Fc-области иммуноглобулина. В одном воплощении полипептид, обладающий активностью GnTIII, содержит каталитический домен бета-1,4-N-ацетилглюкозаминилтрансферазы III и отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, который выбирают из группы, состоящей из домена локализации маннозидазы II, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы I, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы II, домена локализации маннозидазы I и домена локализации альфа1-6 кор-фукозилтрансферазы.
В другом аспекте настоящее изобретение предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей рекомбинантный белок слияния, обладающий активностью бета-1,4-галактозилтрансферазы («GalT»), в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, где указанный полипептид, продуцируемый указанной клеткой-хозяином, выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела, содержащего Fc-область, и белка слияния, содержащего область, эквивалентную Fc-области иммуноглобулина. В одном воплощении рекомбинантный полипептид, обладающий активностью GalT, содержит каталитический домен бета-1,4-галактозилтрансферазы и отвечающий за локализацию в комплексе Гольджи, домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, который выбирают из группы, состоящей из домена локализации маннозидазы II, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы I, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы II, домена локализации маннозидазы I и домена локализации альфа1-6 кор-фукозилтрансферазы.
Предпочтительно, отвечающий за локализацию в комплексе Гольджи домен представляет собой домен маннозидазы II или бета-1,2-N-ацетилглюкозаминилтрансферазы I, или, в качестве альтернативы, галактозилтрансферазы.
В другом аспекте настоящее изобретение предусматривает полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи. В одном воплощении полипептиды слияния согласно данному изобретению содержат каталитический домен бета-1,4-N-ацетилглюкозаминилтрансферазы III. В другом воплощении отвечающий за локализацию в комплексе Гольджи домен выбирают из группы, состоящей из домена локализации маннозидазы II, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы I, домена локализации маннозидазы I, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы II и домена локализации альфа 1-6 кор-фукозилтрансферазы.
В другом аспекте настоящее изобретение предусматривает полипептид слияния, обладающий активностью бета-1,4-галактозалтрансферазы и содержащий домен, отвечающий за локализацию в комплексе Гольджи, какого-либо гетерологичного полипептида-резидента комплекса Гольджи. В одном воплощении полипептиды слияния согласно данному изобретению содержат каталитический домен бета-1,4 галактозилтрансферазы. В другом воплощении отвечающий за локализацию в комплексе Гольджи домен выбирают из группы, состоящей из домена локализации маннозидазы II, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы I, домена локализации
маннозидазы I, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы II и домена локализации альфа 1-6 кор-фукозилтрансферазы.
Предпочтительно, отвечающий за локализацию в комплексе Гольджи домен представляет собой домен маннозидазы II или бета-1,2-N-ацетилглюкозаминилтрансферазы I («GnTI»), или, в качестве альтернативы, галактозилтрансферазы («GalT»).
В другом аспекте настоящее изобретение предусматривает способ получения рекомбинантного полипептида слияния, обладающего активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III, включающий культивирование клетки-хозяина согласно данному изобретению в культуральной среде в условиях, делающих возможной экспрессию нуклеиновой кислоты, кодирующей указанный полипептид слияния, и получение (выделение) полипептида из полученной в результате культуры клеток. В одном воплощении полипептид слияния содержит каталитический домен бета-1,4-N-ацетилглюкозаминилтрансферазы III. Предпочтительно, полипептид слияния содержит отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного белка-резидента комплекса Гольджи, который выбирают из группы, состоящей из домена локализации маннозидазы И, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы I, домена локализации маннозидазы I, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы II и домена локализации альфа1-6 кор-фукозилтрансферазы.
В другом аспекте настоящее изобретение предусматривает способ получения рекомбинантного полипептида слияния, обладающего активностью бета-1,4-N-галактозилтрансферазы, включающий культивирование в среде клетки-хозяина согласно настоящему изобретению в условиях, делающих возможной экспрессию нуклеиновой кислоты, кодирующей полипептид слияния, и получение (выделение) полипептида слияния из полученной в результате культуры. В одном воплощении полипептид слияния содержит каталитический домен бета-1,4-N-галактозилтрансферазы. Предпочтительно, полипептид слияния содержит отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного белка-резидента комплекса Гольджи, который выбирают из группы, состоящей из домена локализации маннозидазы II, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы I, домена локализации маннозидазы Гдомена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы II и домена локализации альфа1-6 кор-фукозилтрансферазы.
Предпочтительно, отвечающий за локализацию в комплексе Гольджи домен представляет собой домен маннозидазы II, или бета-1,2-N-ацетилглюкозаминилтрансферазы I («GnTI»), илигалактозилтрансферазы («GalT»).
В дальнейшем аспекте изобретение предусматривает способ модификации профиля гликозилирования полипептида, продуцируемого клеткой-хозяином, содержащей введенную в нее по меньшей мере одну нуклеиновую кислоту или вектор экспрессии данного изобретения. Предпочтительно, полипептид представляет собой IgG или его фрагмент, содержащий Fc-область этого полипептида(IgG). В особенно предпочтительном воплощении полипептид представляет собой IgG или его фрагмент, содержащий Fc-область этого полипептида. В качестве альтернативы полипептид представляет собой белок слияния, который включает область, эквивалентную Fc-области IgG человека.
В другом аспекте настоящее изобретение предусматривает способ получения рекомбинантного полипептида слияния в клетке-хозяине, включающий: (а) культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III(«GnTIII») или, в качестве альтернативы, активностью бета-1,4-галактозилтрансферазы («GalT»), в культуральной среде в условиях, которые допускают продукцию полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела, содержащего Fc-фрагмент и слитого белка, содержащего область, эквивалентный Fc-области иммуноглобулина, где указанный полипептид слияния экспрессируется в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином; и (b) выделение указанного полипептида. Предпочтительно, полипептид слияния содержит каталитический домен бета-1,4-N-ацетилглюкозаминилтрансферазы III или бета-1,4-галактозилтрансферазы («GalT») и, дополнительно содержит отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, который выбирают из группы, состоящей из домена локализации маннозидазы II, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы I, домена локализации маннозидазы I, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы II и домена локализации альфа 1-6 коровой фукозилтрансферазы. В предпочтительных воплощениях полипептид, продуцируемый клеткой-хозяином, обладает, в результате модификации, повышенной эффекторной функцией и/или повышенным связыванием Fc-рецептора. В особенно предпочтительных воплощениях повышенная эффекторная функция представляет собой повышенную Fc-опосредованную клеточную цитотоксичность, повышенное связывание NK-клеток, повышенное связывание макрофагов, повышенное связывание моноцитов, повышенное связывание полиморфноядерных клеток, повышенную прямую передачу индуцирующих апоптоз сигналов, повышенное созревание дендритных клеток и/или повышенную активность начально активизации Т-лимфоцитов (priming), а повышенное связывание Fc-рецептора представляет собой повышенное связывание активирующего Fc-рецептора, такого как рецептор FcγRIIIA. Предпочтительно полипептид, демонстрирующий повышенную эффекторную функцию и/или повышенное связывание Fc-рецептора, представляет собой антитело, фрагмент антитела или белок слияния, содержащий область, эквивалентную Fc-области иммуноглобулина, и обладает увеличенной долей нефукозилированных олигосахаридов в Fc-области.
В дальнейшем аспекте изобретение предусматривает фармацевтические составы, содержащие антитело, фрагмент антитела, содержащий Fc-область, или рекомбинантный полипептид слияния, содержащий Fc-область иммуноглобулина данного изобретения, и применение таких составов в лечении опухолей, таких как рак, или других расстройств. В одном воплощении лечение представляет собой снижение количества В-лимфоцитов путем назначения терапевтически эффективного количества такого фармацевтического состава пациенту, например, человеку, при необходимости. В дальнейшем аспекте изобретение предусматривает клетку-хозяина, содержащую вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид слияния, где указанный полипептид слияния обладает активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержит отвечающий за локализацию в комплексе Гольджи домен какого-либо полипептида-резидента комплекса Гольджи, а также вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид, где указанный полипептид слияния обладает активностью маннозидазы II. В предпочтительных воплощениях полипептид слияния содержит каталитический домен GnTIII, а отвечающий за локализацию в комплексе Гольджи домен выбирают из группы, состоящей из домена локализации ManII, домена локализации GnTI, домена локализации GnTII, домена локализации ManI и домена локализации альфа1-6 кор-фукозилтрансферазы. В одном воплощении клетка-хозяин дополнительно содержит вектор экспрессии, кодирующий полипептид, обладающий активностью GnTII. Молекулы нуклеиновой кислоты, кодирующие полипептид слияния, обладающий активностью ManII полипептид и обладающий активностью GnTII полипептид, могут содержаться либо в разных векторах экспрессии, либо в одном векторе экспрессии. В дополнительном аспекте изобретение предусматривает клетку-хозяина, содержащую вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую рекомбинантный полипептид слияния, где указанный полипептид слияния обладает активностью бета-1,4-галактозилтрансферазы (GalT) и содержит отвечающий за локализацию в комплексе Гольджи домен полипептида-резидента комплекса Гольджи, а также вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид, где указанный полипептид обладает активностью маннозидазы II (ManII). В предпочтительных воплощениях полипептид слияния содержит каталитический домен GnTIII, а отвечающий за локализацию в комплексе Гольджи домен выбирают из группы, состоящей из домена локализации ManII, домена локализации GnT I, домена локализации GnTII, домена локализации ManI и домена локализации альфа1-6 кор-фукозилтрансферазы. В одном воплощении клетка-хозяин дополнительно содержит вектор экспрессии, кодирующий полипептид, обладающий активностью GnTII. Молекулы нуклеиновых кислот, кодирующие полипептид слияния, обладающий активностью ManII полипептид и обладающий активностью GnTII полипептид, могут содержаться либо в разных векторах экспрессии, либо в одном векторе экспрессии.
В дальнейшем аспекте изобретение предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью GnT III и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью Man II, в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, где указанный полипептид, продуцируемый указанной клеткой-хозяином, выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела, содержащего Fc-фрагмент и слитого белка, который содержит область, эквивалентную Fc-области иммуноглобулина.
В дополнительном способе воплощения изобретение предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей слитый полипептид, обладающий активностью GnT III, по крайней мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью Man II, и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью GnT II, в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого указанной клеткой-хозяином, где указанный полипептид, продуцируемый указанной клеткой-хозяином, выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела и слитого белка, который содержит область, эквивалентную Fc-области иммуноглобулина.
В дальнейшем аспекте изобретение предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью GalT, и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью Man II, в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, где указанный полипептид, продуцируемый указанной клеткой-хозяином, выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела и слитого белка, который содержит область, эквивалентную Fc-области иммуноглобулина. В дополнительном аспекте изобретение предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей рекомбинантный полипептид слияния, обладающий активностью GalT, по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью Man II, и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью GnT II, в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого указанной клеткой-хозяином, где указанный полипептид, продуцируемый указанной клеткой-хозяином, выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела и слитого белка, который содержит область, эквивалентную Fc-области иммуноглобулина.
В дальнейшем аспекте изобретение предусматривает способ продуцирования полипептида клеткой-хозяином, включающий культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью GnT III, и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью Man II, в условиях, которые допускают продуцирование полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела и слитого белка, который содержит область, эквивалентную Fc-области иммуноглобулина, где указанный слитый полипептид экспрессируется в количестве, достаточном для модификации олигосахаридов в Fc-области указанного полипептида, продуцируемого клеткой-хозяином, а также выделение указанного полипептида.
В другом аспекте изобретение предусматривает способ продуцирования полипептида клеткой-хозяином, включающий культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью GalT, и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью Man II, в условиях, которые допускают продуцирование полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела и слитого белка, который содержит область, эквивалентную Fc-области иммуноглобулина, где указанный полипептид слияния экспрессируется в количестве, достаточном для модификации олигосахаридов в Fc-области указанного полипептида, продуцируемого указанной клеткой-хозяином, а также выделение указанного полипептида.
В дополнительном аспекте изобретение предусматривает способ продуцирования полипептида, обладающего повышенной Fc-опосредованной клеточной цитотоксичностью, клеткой-хозяином, включающий культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей GalT, и по меньшей мере одной нуклеиновой кислоты, кодирующей Man II, в условиях, которые допускают продуцирование полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела, который содержит Fc-область иммуноглобулина, где уровень экспрессии одного из ферментов GalT и Man II или их обоих достаточен для модификации олигосахаридов в Fc-области указанного полипептида, продуцируемого указанной клеткой-хозяином, и где указанный полипептид обладает, в результате указанной модификации, повышенной Fc-опосредованной клеточной цитотоксичностью.
В другом аспекте настоящее изобретение предусматривает способ продуцирования полипептида клеткой-хозяином, включающий: (а) культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью альфа-маннозидазы И, в условиях, которые допускают продуцирование полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела и белка слияния, который содержит область, эквивалентную Fc-области иммуноглобулина, где указанный полипептид, обладающий активностью альфа-маннозидазы II, экспрессируется в количестве, достаточном для модификации олигосахаридов в Fc-области указанного полипептида, продуцируемого указанной клеткой-хозяином; и (b) выделение указанного полипептида продуцируемого указанной клеткой-хозяином.
В другом аспекте изобретение предусматривает клетку-хозяина, модифицированную для экспрессии по крайней мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью альфа-маннозидазы II в условиях, которые допускают продуцирование полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела и белка слияния, который содержит фрагмент, эквивалентный Fc-фрагменту иммуноглобулина, где указанный полипептид, обладающий активностью альфа-маннозидазы II, экспрессируется в количестве, достаточном для модификации олигосахаридов в Fc-фрагменте полипептида, продуцируемого клеткой-хозяином.
В еще одном аспекте настоящее изобретение предусматривает полипептиды, продуцируемые такой клеткой-хозяином, в частности - антитела, обладающие, в результате указанной модификации олигосахаридов, повышенной эффекторной функцией и/или повышенным связыванием Fc-рецептора.
Краткое описание рисунков
Фиг. 1: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов рекомбинантного неизмененного (негликомодифицированного) anti-CD20 антитела класса IgG1, продуцируемого клетками ВНК. Клетки были трансфецированы вектором экспрессии антитела pETR1502. Очистку антитела от культуральной среды, а также приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» Примера 1.
Фиг. 2: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов рекомбинантного гликомодифицированного анти-CD20 антитела класса IgG1, продуцируемого клетками ВНК, модифицированных нуклеиновой кислотой, кодирующей GnTIII дикого типа («wt»). Клетки были подвергнуты ко-трансфекции вектором экспрессии антитела pETR1502 и вектором экспрессии GnTIII pETR1166. Очистку антитела от культуральной среды, а также приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» примера 1.
Фиг. 3: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов рекомбинантного гликомодифицированного анти-CD20 антитела класса IgG1, продуцируемого клетками ВНК, модифицированными нуклеиновой кислотой, кодирующей полипептид слияния («Gl-GnTIII») с активностью GnTIII, содержащий функциональный отвечающий за локализацию в комплексе Гольджи домен GnTI. Клетки были подвергнуты ко-трансфекции вектором экспрессии антитела pETR1502 и вектором экспрессии GnTIII pETR1425. Очистку антитела от культуральной среды, а также приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» примера 1.
Фиг. 4: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов рекомбинантного гликомодифицированного анти-CD20 антитела класса IgG1, продуцируемого клетками ВНК, модифицированными нуклеиновой кислотой, кодирующей полипептид слияния ("M2-GnTIII") с активностью GnTIII и лсодержащий функциональный обеспечивающий локализацию в комплексе Гольджи доменмманнозидазы И. Клетки были подвергнуты ко-трансфекции вектором экспрессии антитела pETR1502 и вектором экспрессии GnTIII pETR1506. Очистку антитела от культуральной среды, а также приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» примера 1.
Фиг. 5: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов рекомбинантного неизмененного (негликомодифицированного) анти-CD20 антитела класса IgG1, продуцируемого клетками HEK293-EBNA. Клетки были трансфецированы вектором экспрессии антитела pETR1502. Очистку антитела культуральной среды, а также приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» Примера 1.
Фиг. 6: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов рекомбинантного гликомодифицированного анти-CD20 антитела класса IgG1, продуцируемого клетками HEK293-EBNA, модифицированными нуклеиновой кислотой, кодирующей полипептид слияния ("M2-GnTIII") с активностью GnTIII, содержащий функциональный отвечающий за локализацию в комплексе Гольджи домен альфа-маннозидазы II. Клетки были подвергнуты ко-трансфекции вектором экспрессии антитела pETR1520 и вектором экспрессии GnTIII pETR1519. Очистку антитела от культуральной среды, а также приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» Примера 1.
Фиг. 7: Полученные на масс-спектрометре MALDI/TOF-MS спектры смеси нейтральных олигосахаридов рекомбинантного гликомодифицированного анти-CD20 антитела класса IgG1, продуцируемого клетками HEK293-EBNA, модифицированными нуклеиновой кислотой, кодирующей полипептид слияния ("M2-GnTIII") с активностью GnTIII, содержащий функциональный отвечающий за локализацию в комплексе Гольджи домен альфа-маннозидазы II. Клетки были подвергнуты ко-трансфекции вектором экспрессии антитела pETR1520 и вектором экспрессии GnTIII pETR1519. Очистку антитела от культуральной среды, а также приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» Примера 1. (а) Состав олигосахаридов, образующихся при обработке гликозидазой PNGaseF без дальнейшей ферментативной обработки. (b) Состав олигосахаридов, образующихся при обработке гликозидазой PNGaseF после дополнительного расщепления эндогликозидазой EndoH.
Фиг. 8: (а) Схематическое описание катализируемого эндогликозидазой EndoH расщепления олигосахаридов. Эндогликозидаза EndoH способна расщеплять гибридные (и разветвленные гибридные), но не сложные или сложные разветвленные олигосахариды. (b) Благодаря тому, что эндогликозидаза EndoH способна «различать» сложные и гибридные олигосахариды, обработка EndoH позволяет определить, каким структурам соответствуют пики в спектре олигосахаридов с одинаковыми отношениями m/z в спектрах MALDI/TOF-MS, первоначально полученных в результате обработки эндогликозидазой PNGase.
Фиг. 9: Антителозависимая клеточная цитотоксичность (АЗКЦ), гликомодифицированного при помощи «G1-GnTIII» рекомбинантного анти-CD20 химерного антитела класса IgG1 в сравнении с АЗКЦ неизмененного рекомбинантного анти-CD20 химерного антитела класса IgG1. Оба антитела были продуцированы клетками ВНК. Получение и профиль гликозилирования гликомодифицированного антитела изображены на Фиг. 3, а получение и профиль гликозилирования неизмененного антитела - на Фиг. 1. Клетки-мишени (Т) представляли собой лимфобластоидные клетки человека SKW6.4. Эффекторные клетки (Е) представляли собой свежевыделенные МНПК человека. Клетки Е и Т использовали (в отношении Е:Т=25:1) в анализе АЗКЦ с 4-часовой инкубацией, определяющем цитотоксичность по отношению высвобождения лактат дегидрогеназы (ЛДГ) к контролям максимального высвобождения (при использовании детергента вместо антитела) и спонтанного высвобождения (вместо антитела - культуральная среда). Детали анализа описаны в разделе «Материалы и методы» Примера 1.
Фиг. 10: Антителозависимая клеточная цитотоксичность (АЗКЦ) гликомодифицированного при помощи «M2-GnTIII» рекомбинантного анти-CD20 химерного антитела класса IgG1 в сравнении с АЗКЦ неизмененного рекомбинантного анти-CD20 химерного антитела класса IgG1. Оба антитела были продуцированы клетками HEK293-ENBA. Получение и профиль гликозилирования гликомодифицированного антитела изображены на Фиг. 6, а неизмененного антитела - на Фиг. 5. Клетки-мишени (Т) представляли собой лимфобластоидные клетки человека SKW6.4. Эффекторные клетки (Е) представляли собой свежевыделенные МНПК человека. Клетки Е и Т использовали (в отношении Е:Т=25:1) в анализе АЗКЦ с 4-часовой инкубацией, определяющем АЗКЦ по отношению высвобождения лактат дегидрогеназы (ЛДГ) к контролям максимального высвобождения (при использовании детергента вместо антитела) и спонтанного высвобождения (вместо антитела - культуральная среда). Детали анализа описаны в разделе «Материалы и методы Примера 1».
Фиг. 11: Антителозависимая клеточная цитотоксичность (АЗКЦ) гликомодифицированного при помощи «M2-GnTIII» рекомбинантных анти-CD20 химерных антитела класса IgG1 в сравнении с АЗКЦ гликомодифицированного при помощи «wt-GnTIII» рекомбинантного анти-CD20 химерного антитела класса IgG1. Оба антитела были продуцированы клетками HEK293-ENBA. Получение и профиль гликозилирования гликомодифицированного антитела изображены на Фиг. 4, а гликомодифицированного при помощи «wt-GnTIII» антитела - на Фиг. 2. Клетки-мишени (Т) представляли собой лимфобластоидные клетки человека SKW6.4. Эффекторные клетки (Е) представляли собой свежевыделенные МНПК человека. Клетки Е и Т использовали (в отношении Е:Т=25:1) в анализе АЗКЦ с 4-часовой инкубацией, определяющем АЗКЦ по отношению высвобождения лактат дегидрогеназы (ЛДГ) к контролям максимального высвобождения (при использовании детергента вместо антитела) и спонтанного высвобождения (вместо антитела - культуральная среда). Детали анализа описаны в разделе «Материалы и методы» Примера 1.
Фиг. 12: Связывание рецептора FcgammaRIIIa на NK-клетках гликомодифицированным при помощи «M2-GnTIII» рекомбинантным химерным анти-CD20 антителом класса IgG1, в сравнении со связыванием того же рецептора неизмененным рекомбинантным химерным анти-CD20 антителом класса IgG1. Оба антитела были продуцированы клетками HEK293-ENBA. Получение и профиль гликозилирования гликомодифицированного антитела изображены на Фиг. 6, а неизмененного антитела - на Фиг. 5. Анализ связывания проводили, как описано в разделе «Материалы и методы» Примера 1. NK-клетки человека, экспрессирующие на своей поверхности рецептор FcgammaRIIIa выделяли из организма донора с генотипом, исключающим продукцию рецептора FcgammaRIIc (т.е., гомозиготного по варианту гена, который содержит внутрирамочный стоп-кодон внутри последовательности, кодирующей FcgammaRIIc). Среднее геометрическое интенсивности флуоресценции, измеренное методом «сортировки клеток с активацией флуоресценцией» (FACS) с использованием фрагментов анти-человеческого антитела класса IgG, меченого ФИТЦ (флуоресцин изотиоцианат), возрастет при увеличении количества связанного с NK-клетками рекомбинантного антитела. Как показывает использование конкурирующего фрагмента FcgammaRIIa-специфичного антитела, определяемое в этом анализе связывание является FcgammaRIIIa-специфичным (См. Фиг. 13).
Фиг. 13: Связывание рецептора FcgammaRIIIa на NK-клетках гликомодифицированным при помощи «M2-GnTIII» рекомбинантным химерным анти-CD20 антителом класса IgG1 в сравнении со связыванием того же рецептора неизмененным рекомбинантным химерным анти-CD20 антителом класса IgG1 в присутствии увеличивающихся концентраций конкурирующего фрагмента FcgammaRIIa-специфичного антитела. Оба рекомбинантных антитела были продуцированы клетками HEK293-ENBA. Получение и профиль гликозилирования гликомодифицированного антитела изображены на Фиг. 6, а неизмененного антитела - на Фиг. 5. Анализ связывания проводили, как описано в разделе «Материалы и методы» Примера 1, но инкубируя очищенные NK-клетки одновременно с рекомбинантным антителом (всегда в конечной концентрации 3 мкг/мл) и с фрагментом конкурирующего анти- FcgammaRIII антитела 3G8-Fab2 в увеличивающихся и меняющихся концентрациях (см.график). NK-клетки человека, экспрессирующие на своей поверхности рецептор FcgammaRIIIa выделяли из организма донора с генотипом, исключающим продукцию рецептора FcgammaRIIc (т.е., гомозиготного по варианту гена, который содержит внутрирамочный стоп-кодон внутри последовательности, кодирующей FcgammaRIIc). Среднее геометрическое интенсивности флуоресценции, измеренное методом «сортировки клеток с активацией флуоресценцией» (FACS) с использованием фрагментов анти-человеческого антитела IgG, меченого ФИТЦ (флуоресцин изотиоцианат), возрастет при увеличении количества связанного с NK-клетками рекомбинантного антитела.
Фиг. 14: Полученные на масс-спектрометре MALDI/TOF-MS спектры смесей нейтральных олигосахаридов рекомбинантных антител класса IgG1 «L19», распознающих изоформу ED-B+ фибронектина и продуцируемых клетками HEK293-ENBA. (а) Неизмененное антитело, продуцируемое клетками HEK293-ENBA, трансфецированными вектором экспрессии антитела pETR1546. (b) Антитело, гликомодифицированное при помощи М2-GnTIII, продуцируемое клетками HEK293-ENBA, подвергнутыми котрансфекции вектором экспрессии антитела pETR1546 и вектором экспрессии GnTIII pETR1519. Оба антитела очищали от культуральной среды методом аффинной хроматографии с применением белка А с последующей стадией вытесняющей хроматографии на матриксе Superdex200 (Amersham), при замене буфер афосфатным буферным раствором (ФБР, PBS). Приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» Примера 1.
Фиг. 15: Связывание рецептора FcgammaRIIb на лимфоидных клетках Raji (Лимфома Беркитта человека) гликомодифицированным при помощи «M2-GnTIII» рекомбинантным анти-ED-В+-фибронектин антителом класса IgG1 в сравнении со связыванием того же рецептора неизмененным рекомбинантным анти-ED-В+-фибронектин антителом класса IgG1. Оба антитела были продуцированы клетками HEK293-ENBA. Получение и профиль гликозилирования гликомодифицированного антитела изображен на Фиг. 14b, а неизмененного антитела - на Фиг. 14а. Анализ связывания проводили, как описано в разделе «Материалы и методы» Примера 1. Среднее геометрическое интенсивности флюоресценции, измеренное методом «сортировки клеток с активацией флуоресценцией» (FACS) с использованием фрагментов анти-человеческого антитела IgG, меченого ФИТЦ (флуоресцин изотиоцианат), возрастет при увеличении количества связанного с В-клетками лимфомы линии Raji.
Фиг. 16: Связывание рецептора FcgammaRIIIa на NK-клетках разных доноров гликомодифицированным при помощи «M2-GnTIII» рекомбинантным анти-CD20 антителом класса IgG1 в сравнении со связыванием того же рецептора неизмененным рекомбинантным анти-CD20 антителом класса IgG1. Оба рекомбинантных антитела были продуцированы клетками HEK293-ENBA. Получение и профиль гликозилирования гликомодифицированного антитела изображены на Фиг. 6, а неизмененного антитела - на Фиг. 5. Анализ связывания проводили, как описано в разделе «Материалы и методы» Примера 1. NK-клетки человека, экспрессирующие на свою поверхность рецептор FcgammaRIIIa выделяли из организма донора с генотипом, исключающим продукцию рецептора FcgammaRIIc (т.е., гомозиготного по варианту гена, который содержит внутрирамочный стоп-кодон внутри последовательности, кодирующей FcgammaRIIc). Генотипирование двух доноров показало, что они гомозиготны по «высокоаффинному» варианту гена FcgammaRIIIa (158V). Генотипирование двух других доноров показало, они гетерозиготны (т.е. их генотип- 158V/F, где 158V обозначает «высокоаффинный», a 158F-«низкоаффинный» варианты рецептора FcgammaRIIIa). Среднее геометрическое интенсивности флуоресценции, измеренное методом «сортировки клеток с активацией флуоресценцией» (FACS) с использованием фрагмента анти-человеческого антитела IgG, меченого ФИТЦ (флуоресцин изотиоцианат), возрастет при увеличении количества, связанного с NK-клетками рекомбинантного антитела. Как показывает использование конкурирующего фрагмента FcgammaRIIIa-специфичного антитела, определяемое в этом анализе связывание является FcgammaRIIIa-специфичным (См. Фиг. 13).
Фиг. 17: Анализ методом FACS экспрессии усеченного CD4(tCD4) (а) клетками ВНК-1502-28 (дикий тип) и (b) клоном ВКН-1502-28-11 (гликомодифицированными «М2-GnTIII») - стабильной клеточной линией, продуцирующей химерное анти-CD20 антитело класса IgG1. Экспрессия tCD4 функционально связана с экспрессией M2-GnTIII через элемент IRES вектора экспрессии GnTIII pETR1537, и таким образом может быть использована в качестве непрямого маркера экспрессии GnTIII. Среднее и геометрическое среднее интенсивностей флуоресценции составили, соответственно, 27.6 и 19.9 для гликомодифицированной клеточной линии и 4.7 и 4.1 - для клеточной линии дикого типа.
Фиг. 18: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов гликомодифицированного при помощи M2-GnTII рекомбинантного анти-CD20 химерного антитела класса IgG1, продуцируемого клеточной линией ВПК-1502-28-11. Клеточная линия, очистка антитела, а также приготовление и анализ олигосахаридов описаны в разделе «Материалы и методы» Примера 1.
Фиг. 19: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов полученной, путем высвобождения олигосахаридов при обработке гликозидазой PNGase с последующей обработкой эндогликозидоазой EndoH, из гликомодифицированного при помощи M2-GnTIII рекомбинантного анти-CD20 химерного антитела класса IgG1, продуцируемого клеточной линией ВНК-1502-28-11. Клеточная линия, очистка антитела, а также приготовление и анализ олигосахаридов описаны в разделе «Материалы и методы» Примера 1.
Фиг. 20: Связывание рецептора FcgammaRIIIa на NK-клетках гликомодифицированным при помощи «M2-GnTIII» рекомбинантным химерным анти-CD20 антителом класса IgG1 в сравнении со связыванием того же рецептора неизмененным рекомбинантным химерным анти-CD20 антителом класса IgG1. (антитела продуцируются стабильными клеточными линиями). Профиль гликозилирования гликомодифицированного антитела показан на Фиг. 18 и Фиг. 19. Анализ связывания проводили, как описано в разделе «Материалы и методы» Примера 1. NK-клетки человека, экспрессирующие на своей поверхности рецептор FcgammaRIIa выделяли из организма донора с генотипом, исключающим продукцию рецептор FcgammaRIIc (т.е., гомозиготного по варианту гена, который содержит внутрирамочный стоп-кодон внутри последовательности, кодирующей FcgammaRIIc). Среднее геометрическое интенсивности флуоресценции, измеренное методом «сортировки клеток с активацией флуоресценцией» (FACS) с использованием фрагмента анти-человеческого антитела класса IgG, меченого ФИТЦ (флуоресцин изотиоцианат), возрастет при увеличении количества связанного с NK-клетками рекомбинантного антитела. Как показывает использование конкурирующего фрагмента FcgammaRIIa-специфичного антитела, определяемое в этом анализе связывание является FcgammaRIIIa-специфичным (См. Фиг. 13).
Фиг. 21. Комплемент-опосредованный лизис (здесь КОЛ) гликомодифицированного при помощи «M2-GnTIII» рекомбинантного химерного анти-CD20 антитела класса IgG1 в сравнении с КОЛ неизмененного рекомбинантного химерного анти-CD20 антитела класса IgG1. Оба антитела были продуцированы клетками HEK293-ENBA. Получение и профиль гликозилирования гликомодифицированного антитела изображены на Фиг. 6, а неизмененного антитела - на Фиг. 5. Клетки-мишени (Т) представляли собой лимфобластоидные клетки человека SKW6.4. В анализе использовали комплемент человека. Лизис измеряли по высвобождению ЛДГ. Детали анализа описаны в разделе «Материалы и методы» Примера 1.
Фиг. 22: Полученные на масс-спектрометре MALDI/TOF-MS спектры смесей нейтральных олигосахаридов рекомбинантных химерных антител класса IgG1 «С225», распознающих рецептор к эпидермальному фактору роста (РЭФР) и продуцируемых клетками НЕК293-ENBA. (а) Неизмененное антитело, продуцируемое клетками HEK293-ENBA, трансфецированными вектором экспрессии антитела pETRURSI28 (b) гликомодифицированное при помощи M2-GnTIII антитело, продуцируемое клетками HEK293-ENBA, подвергнутыми котрансфекции вектором экспрессии антитела pETRURSI28 и вектором экспрессии GnTIII pETR1519. Оба антитела очищали от культуральной среды методом аффинной хроматографии с применением белка А с последующей стадией вытесняющей хроматографии на матриксе Superdex200(Amersham), заменяющим буфер фосфатным буферным раствором (ФБР, PBS). Приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» Примера 1.
Фиг. 23: Антителозависимая клеточная цитотоксичность (АЗКЦ) гликомодифицированного при помощи M2-GnTIII рекомбинантного анти-РЭФР химерного антитела класса IgG1 «С225» в сравнении с АЗКЦ неизмененного рекомбинантного анти-РЭФР химерного антитела класса IgG1 «С225». Оба антитела были продуцированы клетками HEK293-ENBA. Получение и профиль гликозилирования гликомодифицированного антитела изображены на Фиг. 22а, а неизмененного антитела - на Фиг. 22b. Клетки-мишени (Т) представляли собой клетки плоскоклеточной карциномы человека А431 (ЕСАСС No. 85090402). Эффекторные клетки (Е) представляли собой свежевыделенные МНПК человека. Клетки Е и Т использовали (в отношении Е:Т=25:1) в анализе АЗКЦ с 4-часовой инкубацией, определяющем АЗКЦ по отношению высвобождения лактат дегидрогеназы (ЛДГ) к контролям максимального высвобождения (при использовании детергента вместо антитела) и спонтанного высвобождения (вместо антитела - культуральная среда). Детали анализа описаны в разделе «Материалы и методы» Примера 1.
Фиг. 24: Последовательность нуклеиновой кислоты и последовательность аминокислот, соответственно, полипептида слияния ManII-GnTIII данного изобретения.
Фиг. 25: Последовательность нуклеиновой кислоты и последовательность аминокислот, соответственно, полипептида слияния GnTI-GnTIII данного изобретения.
Фиг. 26: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов неизмененного рекомбинантного анти-CD20 химерного антитела С2С8 класса IgG1 («Cwt»), продуцируемого клетками HEK293-EBNA, трансфецированными вектором экспрессии антитела pETR1520. Очистка антитела от культуральной среды, а также приготовление и анализ олигосахаридов описаны в разделе «Материалы и методы» Примера 5.
Фиг. 27: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов гликомодифицированного рекомбинантного анти-CD20 химерного антитела С2С8 класса IgG1 («Cbrt»), продуцируемого клетками HEK293-EBNA, подвергнутыми котрансфекции вектором экспрессии антитела pETR1520 и вектором экспрессии полипептида слияния GnTIII pETR1519. Очистку антитела от культуральной среды, а также приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» примера 5. (А) Состав олигосахаридов, высвобожденных при обработке гликозидазой PNGaseF без дальнейшей ферментативной обработки. (В) Состав олигосахаридов, высвобожденных при обработке гликозидазой PNGaseF после дополнительного расщепления эндогликозидазой EndoH.
Фиг. 28: Полученный на масс-спектрометре MALDI/TOF-MS спектр смеси нейтральных олигосахаридов гликомодифицированного рекомбинантного анти-CD20 химерного антитела С2С8 класса IgG1 («Cm»), продуцируемого клетками HEK293-EBNA, подвергнутого котрансфекции вектором экспрессии антитела pETR1520, вектором экспрессии полипептида слияния GnTIII (pETR1519) и вектором экспрессии полипептида маннозидазы II (pCLF9). Очистку антитела от культуральной среды, а также приготовление и анализ олигосахаридов проводили, как описано в разделе «Материалы и методы» примера 5. (А) Состав олигосахаридов, высвобожденных при обработке гликозидазой PNGaseF без дальнейшей ферментативной обработки. (В) Состав олигосахаридов, высвобожденных при обработке гликозидазой PNGaseF после дополнительного расщепления эндогликозидазой EndoH.
Фиг. 29: Антителозависимая клеточная цитотоксичность (АЗКЦ), опосредуемая химерными анти-CD20 антителами гликомодифицированными путем экспрессии в клетках HEK293-EBNA нуклеиновой кислоты, кодирующей белок слияния ManII-GnTIII, где каталитический домен GnTIII локализуется через обеспечивающий локализацию в комплексе Гольджи домен ManII, где кодирующая ManII-GnTIII нуклеиновая кислота экспрессируется либо сама по себе (Cbrt), либо совместно с нуклеиновой кислотой, кодирующей ManII (Cm). Cwt представляет собой неизмененное рекомбинантное анти-CD20 химерное антитело С2С8 класса IgG1 («Cwt»), продуцируемое клетками НЕК293-EBNA, трансфецированными вектором экспрессии антитела pETR1520. Детали анализа описаны в разделе «Материалы и методы» Примера 1.
Фиг. 30: Связывание рецептора FcgammaRIII химерными анти-CD20 антителами, гликомодифицированными путем экспрессии в клетках HEK293-EBNA нуклеиновой
кислоты, кодирующей полипептид слияния ManII-GnTIII, где каталитический домен, GnTIII локализуется через обеспечивающий локализацию в комплексе Гольджи домен ManII, где кодирующая ManII-GnTIII нуклеиновая кислота экспрессируется либо сама по себе (антитело Cbrt), либо совместно с кодирующей ManII нуклеиновой кислотой (Cm). Cwt представляет собой неизмененное рекомбинантное анти-CD20 химерное антитело С2С8 класса IgG1 («Cwt»), продуцируемое клетками HEK293-EBNA, трансфецированными вектором экспрессии антитела pETRT520. Анализ связывания проводили, как описано в разделе «Материалы и методы» Примера 1. NK-клетки человека, экспрессирующие на своей поверхности рецептор FcgammaRIIIa, выделяли из организма донора с генотипом, исключающем продукцию рецептора FcgammaRIIc (т.е., гомозиготных по варианту гена, который содержит внутрирамочный стоп-кодон внутри последовательности, кодирующей FcgammaRIIc). Среднее геометрическое интенсивности флуоресценции, измеренное методом «сортировки клеток с активацией флуоресценцией» (FACS) с использованием фрагментов анти-человеческого антитела IgG, меченого ФИТЦ (флуоресцин изотиоцианат), возрастает при увеличении количества связанного с NK-клетками рекомбинантного антитела. Как показывает использование конкурирующего фрагмента FcgammaRIIIa-специфичного антитела, определяемое в этом анализе связывание является FcgammaRIIIa-специфичным (См. Фиг. 13)
Фиг. 31: Комплемент-опосредованная цитотоксичность химерного анти-CD20 антитела, гликомодифицированного путем экспрессии в клетках HEK293-EBNA нуклеиновой кислоты, кодирующей полипептид слияния ManII-GnTIII, где каталитический домен, GnTIII локализуется через обеспечивающий локализацию в комплексе Гольджи домен ManII, где кодирующая ManII-GnTIII нуклеиновая кислота экспрессируется либо сама по себе (Cbrt), либо совместно с кодирующей ManII нуклеиновой кислотой(Cm). Cwt представляет собой неизмененное рекомбинантное анти-CD20 химерное антитело С2С8 класса IgG1 («Cwt»), продуцируемое клетками HEK293-EBNA, трансфецированными вектором экспрессии антитела pETR1520. Детали анализа описаны в разделе «Материалы и методы» Примера 1.
Фиг. 32 (А-С): Векторы экспрессии pCLF9(A), pETR1842(B) и pETR1843(С)
Фиг. 33 (А и В): Векторы экспрессии для белков слияния ManII-GalT(A) и GalT(B).
Фиг. 34: Состав олигосахаридов анти-CD20 моноклонального антитела, продуцируемого в присутствии альфа-маннозидазы II и относительное процентное содержание структур, связанных с Fc-частью антитела.
Фиг. 35 (А и В): Состав олигосахаридов анти-CD2 моноклонального антитела, продуцируемого в присутствии белка слияния ManII-GalT и относительное процентное содержание структур, связанных с Fc-частью антитела. Состав олигосахаридов после расщепления гликозидазами PNGaseF(А) и EndoH(В).
Фиг. 36: Антитело, продуцируемое в присутствии альфа-маннозидазы II (ManII) связывает рецепрор FcγRIIIA с более высокой, чем у антитела дикого типа, аффинностью.
Фиг. 37: Антителозависимая клеточная цитотоксичность, опосредуемая гликомодифицированным химерным анти-CD20 антителом.
Подробное описание изобретения
Научные и технические термины употребляются здесь в значениях, обычно употребляемых в данной области, если они дополнительно не определены следующим образом:
Термин «антитело» здесь включает целые молекулы антител, в т.ч. моноклональных, поликлональных и мультиспецифических (например, биспецифических), а также фрагменты антител, обладающие Fc-областью, и белки слияния, которые содержат области, эквивалентные Fc-области иммуноглобулина. Термин также охватывает гуманизированные и химерные антитела.
Термин «Fc-область» здесь относится С-концевой области тяжелой цепи IgG. Хотя границы Fc-области тяжелой цепи IgG могут слегка варьироваться, Fc-область IgG человека обычно определяют, как участок от аминокислотного остатка в положении Cys 226 до карбоксильного конца.
Термин «область, эквивалентная Fc-области иммуноглобулина» включает здесь встречающиеся в природе аллельные варианты Fc-области какого-либо иммуноглобулина, а также варианты, имеющие отличия, которые порождают замены, вставки или делеции, но которые не приводят к значительному снижению способности иммуноглобулина опосредовать эффекторные функции (такие как антителозависимая клеточная цитотоксичность). Например, одна или более аминокислот могут быть делетированы из N-конца или С-конца Fc-области иммуноглобулина без существенной потери биологической функции. Такие варианты могут быть выбраны, согласно общим правилам, известным в данной области, таким образом, чтобы влияние на активность было минимальным. См., например, (См. например, Bowie, J.U. et al., Science 247:1306-10 (1990).
Термин (рекомбинантный) полипептид слияния, «обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III» здесь относится к полипептидам слияния, которые способны катализировать присоединение при помощи β-1,-4 связи остатка N-ацетилглюкозамина (GlcNAc) к содержащей три остатка маннозы коровой части олигосахаридов. Это включает полипептиды слияния, демонстрирующие ферментативную активность сходную, но необязательно идентичную, с активностью бета-1,4-N-глюкозаминилтрансферазы III, также известной как бета-1,4-маннозил-гликопротеин4-бета-N-ацетилглюкозаминилтрансфераза (КФ 2.4.1.144 согласно номенклатуре NC-IUBMB - Комитета по Номенклатуре Международного Союза по Биохимии и Молекулярной Биологии), что определяется по результатам измерения в специальном биологическом анализе (с или без зависимости от дозы). В случае, если существует зависимость от дозы, она не обязательно должна быть идентичной зависимости, присущей бета-1,4-N-глюкозаминилтрансферазе III (т.е. исследуемый полипептид может проявлять активность большую, либо меньшую не более, чем в 25 раз, предпочтительно - меньшую не более, чем в 10 раз, наиболее предпочтительно - меньшую не более, чем в 3 раза, по отношению к активности бета-1,4-N-ацетилглюкозаминилтрансферазы III).
Термин (рекомбинантный) полипептид слияния, «обладающий активностью бета-1,4-галактозилтрансферазы» или «обладающий активностью GalT» здесь относится к полипептидам слияния, которые способны катализировать перенос остатка галактозы с УДФ-галактозы на невосстанавливающий концевой GlcNAc, обнаруженный в N-связанных олигосахаридах. Это включает полипептиды слияния, демонстрирующие ферментативную активность сходную, но необязательно идентичную, с активностью бета-1,4-N-галактозилтрансферазы, также известной как УДФ-Гал:N-ацетилглюкозамин бета-1,4-N-галактозилтрансфераза (КФ 2.4.1.38 согласно номенклатуре NC-IUBMB - Комитета по Номенклатуре Международного Союза по Биохимии и Молекулярной Биологии), что определяется по результатам измерения в специальном биологическом анализе (с или без зависимости от дозы). В случае, если существует зависимость от дозы, она не обязательно должна быть идентичной присущей бета-1,4-N-галактозилтрансферазе (т.е. исследуемый полипептид может проявлять активность большую, либо меньшую не более, чем в 25 раз, предпочтительно - меньшую не более, чем в 10 раз, наиболее предпочтительно - меньшую не более, чем в 3 раза, по отношению к активности бета-1,4-N-галактозилтрансферазы).
Утверждение, что нуклеиновая кислота или полинуклеотид имеет последовательность нуклеотидов, «идентичную» по меньшей мере, например, на 95% опорной (эталонной, послед-ти сравнения - «reference») последовательности нуклеотидов согласно настоящему изобретению, означает здесь, что последовательность нуклеотидов или полинуклеотид идентичен опорной последовательности за тем исключением, что эта последовательность полинуклеотида может включать до пяти точечных мутаций на каждые 100 нуклеотидов опорной последовательности нуклеотидов. Другими словами, чтобы получить полинуклеотид, обладающий последовательностью нуклеотидов, идентичной опорной последовательности нуклеотидов по меньшей мере на 95%, до 5% нуклеотидов опорной последовательности может быть подвергнуто делеции или замене на другой нуклеотид, либо может быть произведена инсерция нуклеотидов в количестве, не превышающем 5% общего числа нуклеотидов в опорной последовательности. Рассматриваемая последовательность может представлять собой целую последовательность, показанную на Фиг. 5 либо на Фиг. 6.
На практике, является ли некоторая молекула нуклеиновой кислоты или полипептида по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичной последовательности нуклеотидов или последовательности полипептида согласно настоящему изобретению, можно определить обычным способом с применением известных в данной области компьютерных программ. Предпочтительный способ определения «лучшего общего совпадения» между последовательностью сравнения (последовательностью согласно настоящему изобретению) и рассматриваемой последовательностью, также известный как «полное выравнивание последовательностей», может быть осуществлен при помощи компьютерной программы FASTDB, основанной на алгоритме, описанном в Bratlag et al., Comp. Арр. Biosci. 6: 237-245 (1990). При выравнивании и последовательность сравнения, и рассматриваемая последовательность представляют собой последовательности ДНК. Последовательность РНК можно сравнивать, заменив нуклеотиды У(U) на Т(Т). Результат выравнивания выражен в процентах идентичности. Следующие значения параметров предпочтительны для выравнивания последовательностей ДНК при помощи программы FASTDB: Matrix (матрица) = Unitary, k-tuple (размер участка максимального совпадения) = 4, MismatchPenalty (штраф за несовпадение) = 1, Joining Penalty (штраф за соединение) = 30, Randomization Group Length (длина группы рандомизации) = 0, Cutoff Score = 1 (пороговое значение), Gap Penalty (штраф на введение гэпов (делеций)) = 5, Gap Size Penalty (штраф на размер гэпа) = 0.05, Window Size (размер сегмента) = 500 или длине рассматриваемой последовательности, если она короче 500 нуклеотидов.
Если рассматриваемая последовательность короче последовательности сравнения вследствие делеций на концах 3' или 5', а не вследствие внутренних делеций, должна быть проведена коррекция результатов «вручную». Это необходимо, т.к. программа FASTDB не учитывает усечения концов 3' или 5' при расчете значения идентичности (%). Для рассматриваемых последовательностей, усеченных по сравнению с последовательностью сравнения на 3'- или 5'-конце, значение идентичности в процентах
корректируют путем расчета процентного отношения числа оснований последовательности сравнения, которые представляют собой 3' или 5' рассматриваемой последовательности и которые не совпадают/не выровнены, к общему числу оснований последовательности сравнения. Совпадение/ выравнивание нуклеотида определяют по результатам выравнивания программой FASTDB. Полученное процентное отношение затем вычитают из значения идентичности (%), вычисленного ранее при помощи программы FASTDB, с использованием указанных параметров, и получают конечное значение идентичности. Откорректированное значение идентичности и применяют для целей данного изобретения. Только основания за пределами 3'- и 5'- концов рассматриваемой последовательности (что показывает выравнивание программой FASTDB), которые не совпадают/не выровнены с последовательностью сравнения, берут в расчет с целью корректировки значения идентичности.
Например, рассматриваемую последовательность длиной 90 оснований выравнивают относительно последовательности сравнения в 100 нуклеотидов для определения значения идентичности (%). На 5'-конце рассматриваемой последовательности есть делеций и, следовательно, выравнивание при помощи FASTDB не учитывает совпадения/выравнивания первых 10 оснований на 5'-конце. Эти 10 неспаренных оснований составляют 10% последовательности (число не совпадающих оснований на 5'- и 3'-конце/общее число оснований в последовательности сравнения), поэтому из значения идентичности (%), рассчитанного программой FASTDB, вычитают 10%. Если бы оставшиеся 90 оснований идеально совпали, значение идентичности составило бы 90%. В другом примере рассматриваемую последовательность длиной 90 оснований сравнивают с последовательностью сравнения длиной 100 оснований. В этот раз делеций представляют собой внутренние делеций и, поэтому, на 5' и 3' рассматриваемой последовательности нет оснований, не совпадающих/не выровненных с последовательностью сравнения. В этом случае процент идентичности, рассчитанный при помощи FASTDB, не корректируют вручную. Еще раз, только основания [за] 5' и 3' рассматриваемой последовательности, которые не совпадают/не выровнены, с последовательностью сравнения подвергают ручной коррекции. Никакая другая пользовательская коррекция не допустима для целей настоящего изобретения.
Утверждение, что полипептид имеет последовательность аминокислот, «идентичную» по меньшей мере например, на 95% последовательности аминокислот сравнения данного изобретения, здесь означает, что последовательность аминокислот рассматриваемого полипептида идентична последовательности сравнения, за тем исключением, что эта рассматриваемая последовательность может включать до пяти изменений аминокислот на каждые 100 аминокислот последовательности сравнения аминокислот. Другими словами, чтобы получить полипептид, имеющий последовательность аминокислот, по меньшей мере на 95% идентичную последовательности сравнения аминокислот, до 5% аминокислотных остатков рассматриваемой последовательности может быть подвергнуты инсерции, делеций или замене на другую аминокислоту. Эти изменения по отношению к опорной последовательности могут появиться в N-концевом или С-концевом положениях опорной последовательности аминокислот либо в любых положениях между этими концевыми положениями, и могут быть распределены по одному между остатками последовательности сравнения или собраны в одну или несколько непрерывных групп внутри последовательности сравнения.
На практике, является ли некоторый полипептид по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичным опорному полипептиду, можно определить обычным способом с применением известных в данной области компьютерных программ. Предпочтительный способ определения «лучшего общего совпадения» между последовательностью сравнения (последовательностью согласно настоящему изобретению) и рассматриваемой последовательностью, также известный как «полное выравнивание последовательностей», может быть осуществлен при помощи компьютерной программы FASTDB, основанной на алгоритме, описанном в Brutlag et al., Comp. Арр. Biosci. 6: 237-245 (1990). При выравнивании последовательностей последовательность сравнения и рассматриваемая последовательность одновременно представляют собой либо последовательности нуклеотидов либо последовательности аминокислот. Результаты указанного «полного выравнивания последовательностей» выражаются в процентном значении идентичности. Следующие параметры предпочтительны для выравнивания последовательностей ДНК при помощи программы FASTDB: Matrix (матрица) = РАМ 0, k-tuple (размер участка максимального совпадения) = 2, MismatchPenalty (штраф за несовпадение) = 1, Joining Penalty (штраф за соединение) = 20, Randomization Group Length (длина группы рандомизации) = 0, Cutoff Score (пороговое значение) = 1, Gap Penalty (штраф на введение гэпов (делеций)) = 5, Gap Size Penalty (штраф на размер гэпа) = 0.05, Window Size (размер сегмента) == 500 или длине рассматриваемой последовательности, если она короче 500 аминокислот.
Если рассматриваемая последовательность короче последовательности сравнения вследствие N-концевых или С-концевых делеций, должна быть проведена коррекция результатов «вручную». Это необходимо т.к. программа FASTDB не учитывает усечения N- или С-концов при расчете общего значения идентичности (%). Для рассматриваемых последовательностей, усеченных по сравнению с последовательностью сравнения на N- или С-конце, значение идентичности (%) корректируют путем расчета процентного отношения числа оснований последовательности сравнения, которые представляют собой N- или С-концы рассматриваемой последовательности и которые не совпадают/не выровнены, к общему числу оснований последовательности сравнения. Совпадение/выравнивание нуклеотида определяют по результатам выравнивания программой FASTDB. Полученный процент затем вычитают из значения идентичности (%), вычисленного ранее программой FASTDB с использованием указанных параметров, и получают конечное значение идентичности (%). Этот конечный процент идентичности и применяют затем в целях данного изобретения. Только остатки на N- и С-концах рассматриваемой последовательности которые не совпадают/не выровнены с последовательностью сравнения рассматривают в целях ручной корректировки процента идентичности, т.е., только положения остатков последовательности сравнения, находящиеся за самыми конечными остатками N- и С-концов рассматриваемой последовательности.
Например, рассматриваемую последовательность длиной 90 аминокислотных остатков выравнивают относительно последовательности сравнения в 100 остатков для определения значения идентичности (%). На N-конце рассматриваемой последовательности есть делеция и, следовательно, выравнивание при помощи FASTDB не показывает совпадения/выравнивания первых 10 оснований на N-конце. Эти 10 неспаренных оснований составляют 10% последовательности (число не совпадающих оснований на N- и С-концах/общее число оснований в последовательности сравнения), поэтому из значения идентичности (%), рассчитанного программой FASTDB, вычитают 10%. Если бы оставшиеся 90 оснований идеально совпали, значение идентичности составило бы 90%. В другом примере рассматриваемую последовательность длиной 90 оснований сравнивают с последовательностью сравнения длиной 100 оснований. В этот раз делеций представляют собой внутренние делеций и поэтому на N и С рассматриваемой последовательности нет оснований, не совпадающих/не выровненных с последовательностью сравнения. В этом случае значение идентичности (%), рассчитанное программой FASTDB, не корректируют вручную. Еще раз, только основания за N- и С-концами рассматриваемой последовательности (что показывает выравнивание программой FASTDB), которые не совпадают/не выровнены, с последовательностью сравнения подвергают ручной коррекции. Никакая другая пользовательская коррекция не допустима для целей настоящего изобретения.
Термин «нуклеиновая кислота, которая «гибридизуется в жестких условиях» с нуклеиновой кислотой данного изобретения, относится здесь к полинуклеотиду, который гибридизуется в ходе ночной инкубации при 42°С в растворе, содержащем 50% формамида, 5 × раствор SSC (750 мМ NaCl, 75 мМ лимоонокислый натрий), 50 мМ фосфата натрия (рН 7.6), 5 × раствор Денхардта, 10% декстран сульфат и 20 мкг/мл денатурированной расщепленной ДНК спермы лосося, с последующей промывкой фильтров в 0.1 × SSC при приблизительно 65°С.
Термин «домен, отвечающий за локализацию в комплексе Гольджи» относится здесь к последовательности аминокислот полипептида-резидента комплекса Гольджи, которая ответственна за «заякоревание» этого полипептида в положении внутри комплекса Гольджи. Обычно, домены локализации находятся на N- конце ферментов.
Термин «эффекторная функция» относится здесь к биологической активности, которая может быть приписана Fc-области (нативной последовательности Fc-области или варианту последовательности аминокислот Fc-области) антитела. Примеры эффекторной функции антител включаютбез огарничения: аффинность связывания Fc-рецептора, антителозависимую клеточную цитотоксичность (АЗКЦ), антителозависимый клеточный фагоцитоз (АЗКФ), секрецию цитокинов, опосредованное иммунными комплексами поглощение антигена антиген-презентирующими клетками, понижающая регуляция поверхностных клеточных рецепторов и т.д.
Термины «модифицировать», «модифицированный», «модификация» и «модификация гликозилирования» рассматриваются здесь как включающие любые манипуляции с паттерном (вариантом расположения остатков Сахаров) гликозилирования встречающихся в природе полипептидов и их фрагментов. Модификация гликозилирования включает метаболическую модификацию элементов клетки, участвующих в гликозилировании, включая генетические манипуляции путями синтеза олигосахаридов для достижения измененного гликозилирования гликопротеинов, эксперссируемых в клетках. Кроме того, модификация гликозилирования включает влияние мутаций и окружающей клетку среды на гликозилирование.
Термин «клетка-хозяин» охватывает любые клеточные системы, которые могут быть модифицированы для наработки модифицированных гликоформ требуемых белков, белковых фрагментов или пептидов, включая антитела, фрагменты антител и слитые белки. Обычно, на клетку-хозяина воздействуют таким образом, чтобы оптимизировать уровни экспрессии GnTIII в ней. Клетки-хозяева включают культивируемые клетки, например, культивируемые клетки млекопитающих, такие как клетки яичников китайского хомячка (СНО), клетки почек хомячков (ВНК), клетки мышиной миеломы NSO, клетки мышиной миеломы SP2/0, клетки миеломы YO, клетки миеломы мыши Р3Х63, клетки PER, клетки PER. С6 или клетки гибридом, клетки дрожжей, клетки насекомых, клетки растений, если назвать только некоторые, а также клетки трансгенных растений, трансгенных животных или культивируемых тканей растений или животных. Термин Fc-опосредованная клеточная цитотоксичность включает здесь антителозависимую клеточную цитотоксичность, опосредуемую растворимым Fc- белком слияния, который содержит Fc-область антитела человека. Это иммунный механизм, приводящий к лизису «клеток-мишеней антител» «эффекторными клетками иммунной системы человека», где:
«Эффекторные клетки иммунной системы человека» представляют собой популяцию лейкоцитов, имеющих на своей поверхности Fc-рецепторы, при помощи которых они связывают Fc-область антител или Fc- белков слияния и осуществляют эффекторную функцию. Такая популяция может включать, но не ограничиваться, мононуклеары периферической крови (МНПК) и/или естественные киллеры (NK-клетки). «Клетки-мишени антител» представляют собой клетки, связываемые антителами или Fc- белками слияния. Антитела или Fc-слитые белки связывают клетки-мишени через белковую часть N-конца Fc-области.
Термин «повышенная Fc-опосредованная клеточная цитотоксичность» определен здесь либо как повышение количества «клеток-мишеней антител», лизируемых по механизму Fc-опосредованной клеточной цитотоксичности (определенной выше) за данное время при заданной концентрации антитела или Fc- белка слияния в окружающей клетки-мишени среде, и/или уменьшение концентрации антитела или Fc-слитого белка в окружающей клетки-мишени среде, необходимое для достижения лизиса по механизму Fc-опосредованной клеточной цитотоксичности заданного количества «клеток-мишеней антител» за данное время. Повышение Fc-опосредованной клеточной цитотоксичности определяют по сравнению с клеточной цитотоксичностью, опосредуемой тем же антителом или Fc- белком слияния, продуцируемым клетками-хозяевами того же типа, при применении тех же стандартных способов получения, очистки и хранения, известных специалистам в данной области, но в отсутствии модификации клетки-хозяина описанными здесь способами для экспрессии гликозилтрансферазы GnTIII. Под «антителом, обладающим повышенной антителозависимой клеточной цитотоксичностью (АЗКЦ)» понимают антитело, обладающее повышенной АЗКЦ, что определено любым подходящим методом из известных среднему специалисту в данной области. Один из принятых анализов для определения АЗКЦ in vitro представляет собой следующее:
1) в анализе используют клетки-мишени установленно экспрессирующие целевой антиген, распознаваемый антиген-связывающей областью антитела;
2) в анализе в качестве эффекторных клеток используют мононуклеары периферической крови (МНПК) человека, полученные из крови выбранного случайным образом здорового донора;
3) анализ проводят согласно следующему протоколу
i) МНПК выделяют, используя стандартные процедуры центрифугирования в градиенте плотности и суспендируют в концентрации 5×106 клеток/мл в среде RPMI для клеточных культур;
ii) клетки-мишени выращивают стандартными методами культивирования тканей, собирают в экспоненциальной фазе роста (при жизнеспособности более 90%), промывают в среде RPMI для клеточных культур, метят 100 мккюри 51Cr, дважды промывают средой для клеточных культур и ресуспендируют в среде для клеточных культур при плотности 106 клеток/мл;
iii) переносят по 100 мкл приготовленной таким образом конечной суспензии клеток-мишеней в каждую лунку 96-луночного микропланшета;
iv) проводят серийные разведения антитела от 4000 нг/мл до 0.04 нг/мл в среде для клеточных культур и вносят по 50 мкл полученных растворов к клеткам-мишеням в 96-луночном микропланшете, исследуя различные концентрации антитела во всем указанном выше диапазоне в трипликатах.
v) в качестве контроля максимального высвобождения (MR), в 3 дополнительные лунки на планшете, содержащие с меченые клетки-мишени добавляют, вместо раствора антитела (п. iv выше), по 50 мкл 2% (по объему) водного раствора неионного детергента (Nonidet, Sigma, Сент-Луис);
vi) в качестве контроля спонтанного высвобождения (SR), в 3 дополнительные лунки, содержащие меченые клетки-мишени добавляют, вместо раствора антитела (п. iv выше), по 50 мкл среды RPMI для клеточных культур;
vii) затем 96-луночный микропланшет центрифугируют при 50×g в течение одной минуты и инкубируют в течение 1 часа при 4°С;
viii) в каждую лунку 96-луночного микропланшета добавляют по 50 мкл суспензии МНПК (п. i выше) для получения отношения количества эффекторных клеток к клеткам-мишеням 25:1, затем планшеты помещают в инкубатор при 5% содержании СО2 в атмосфере и выдерживают при 37°С в течение 1 часа;
ix) собирают свободный от клеток супернатант из каждой лунки и определяют экспериментальны высвобожденную радиоактивность (ER) при помощи счетчика гамма-частиц;
x) для каждой концентрации антитела рассчитывают процентную долю специфического лизиса по формуле: (ER-MR)/(MR-SR)×100, где ER- средняя радиоактивность, определенная (см. п. ix выше) для данной концентрации антитела, MR - средняя радиоактивность определенная для контролей максимального высвобождениями, п. v выше), a SR- средняя радиоактивность определенная для контролей спонтанного высвобождения (см. п. vi выше);
4) термин «повышенная АЗКЦ» определен либо как повышение максимальной процентной доли специфического лизиса, наблюдаемого в диапазоне концентраций антитела, исследованных, как описано выше, и/или снижение концентрации антитела, необходимой для достижения половины максимальной процентной доли специфического лизиса, наблюдаемого в диапазоне концентраций антитела, исследованных, как описано выше. Повышение АЗКЦ определяют относительно определенной при помощи описанного выше анализа АЗКЦ, опосредуемой тем же антителом, продуцируемым клетками-хозяевами того же типа, при использовании тех же стандартных способов получения, очистки и хранения, известных специалистам в данной области, но которое не было продуцировано клеткой-хозяином, модифицированной описанными здесь способами для экспрессии гликозилтрансферазы GnT III.
Термин анти-CD20 антитело здесь использован для обозначения антитела, которое специфически распознает расположенный на поверхности клетки негликозилированный фосфопротеин массой 35,000 Да, обозначаемый обычно как антиген узкой дифференцировки В-лимфоцитов человека Вр35, часто называемый CD20.
Настоящее изобретение основано на открытии того факта, что модификация продуцирующих антитела клеток для экспрессии нового слитого белка, обладающего активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) или, в качестве альтернативы, активностью бета-1,4-N-галактозилтрансферазы(GalT) и содержащего отвечающий за локализацию в комплексе Гольджи домен полипептида-резидента комплекса Гольджи, приводит к получению антител с повышенной аффинностью связывания Fc-рецептора и повышенной эффекторной функцией. В качестве альтернативы, посредством модификации продуцирующих антитела клеток для повышенной экспрессии молекулы нуклеиновой кислоты, кодирующей полипептид, обладающий каталитической активностью маннозидазы II, могут быть получены антитела с повышенной эффекторной функцией и/или связыванием Fc-рецептора. В предпочтительных воплощениях конструкции слияния, обладающие активностью GnTIII или GalT экспрессируются совместно с молекулами нуклеиновой кислоты, кодирующими ManII или GnTII.
Предпочтительно, изолированная нуклеиновая кислота имеет последовательность нуклеотидов, показанную на Фиг. 24 и SEQ ID NO: 12. В другом предпочтительном воплощении полипептид слияния содержит каталитический домен бета-1,4-N-ацетилглюкозаминилтрансферазы III, а отвечающий за локализацию в комплексе Гольджи домен является доменом бета-1,2-N-ацетилглюкозаминилтрансферазы I (GnTI). Предпочтительно, нуклеиновая кислота имеет последовательность нуклеотидов, показанную на Фиг. 25 и SEQ ID NO: 14. В качестве альтернативы может быть использован отвечающий за локализацию в комплексе Гольджи домен другого полипептида-резидента комплекса Гольджи. В другом предпочтительном воплощении отвечающий за локализацию в комплексе Гольджи домен выбирают из группы, состоящей из домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы II, домена локализации маннозидазы I и домена локализации альфа1-6 кор-фукозилтрансферазы.
В другом предпочтительном воплощении настоящее изобретение предусматривает изолированную нуклеиновую кислоту, содержащую последовательность, которая кодирует полипептид, имеющий последовательность аминокислот, показанную на Фиг. 24 и SEQ ID NO: 13 или на Фиг. 25 и SEQ ID NO: 15. Настоящее изобретение включает также изолированную нуклеиновую кислоту, которая в жестких условиях гибридизуется с гибридизационным зондом с последовательностью нуклеотидов, которая состоит из последовательности нуклеотидов, показанной на Фиг. 24 и SEQ ID NO: 12 или Фиг. 25 и SEQ ID NO: 14. Далее изобретение предусматривает изолированную нуклеиновую кислоту, содержащую последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичную последовательности нуклеотидов, показанной на Фиг. 24 и SEQ ID NO: 12 или Фиг. 25 и SEQ ID NO: 14. В другом воплощении изобретение предусматривает изолированную нуклеиновую кислоту, содержащую последовательность, кодирующую полипептид, имеющий аминокислотную последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичную последовательности аминокислот, показанной на Фиг. 24 и SEQ ID NO: 13 или Фиг. 25 и SEQ ID NO: 15. Изобретение также включает изолированную нуклеиновую кислоту, содержащую последовательность, которая кодирует полипептид, имеющий последовательность аминокислот на Фиг. 24 и SEQ ID NO: 13 или Фиг. 25 и SEQ ID NO: 15 с консервативными заменами аминокислот.
Далее настоящее изобретение предусматривает способ модификации профиля гликозилирования полипептида, продуцируемого клеткой-хозяином, включающий введение в указанную клетку-хозяина молекулы нуклеиновой кислоты или вектора данного изобретения. Предпочтительно, модифицированный полипептид представляет собой IgG или его фрагмент, содержащий Fc-область. В другом предпочтительном воплощении модифицированный полипептид представляет собой белок слияния, который включает область, эквивалентную Fc-области IgG человека.
Далее настоящее изобретение предусматривает клетку-хозяина, содержащую нуклеиновую кислоту и вектор экспрессии данного изобретения. В одном воплощении настоящее изобретение предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III, или, в качестве альтернативы, активностью бета-1,4-галактозилтрансферазы («GalT»), в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого указанной клеткой-хозяином, где указанный полипептид выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела, содержащего Fc-фрагмент и слитого белка, который содержит область, эквивалентную Fc-области иммуноглобулина. В предпочтительном воплощении полипептид, продуцируемый указанной клеткой-хозяином, представляет собой IgG или его фрагмент. Наиболее предпочтительно, полипептид, продуцируемый указанной клеткой-хозяином, представляет собой IgG1 или его фрагмент. В качестве альтернативы, полипептид, продуцируемый указанной клеткой-хозяином, представляет собой белок, который содержит область, эквивалентную Fc-области IgG человека, например, IgG1.
Модифицированные полипептиды, продуцируемые клеткой-хозяином данного изобретения, демонстрируют, в результате модификации, повышенную аффинность связывания Fc-рецептора и/или повышенную эффекторную функцию. Предпочтительно, повышенная аффинность связывания Fc-рецептора представляет собой повышенное связывание активирующего рецептора Fcγ, такого как рецептор FcγRIIIa. Повышенная эффекторная функция предпочтительно представляет собой повышение одной или более из следующих функций: повышенная антителозависимая клеточная цитотоксичность, повышенный антителозависимый клеточный фагоцитоз (АЗКФ), повышенная секреция цитокинов, повышенное опосредованное иммунными комплексами поглощение антигена антиген-презентирующими клетками, повышенная Fc-опосредованная клеточная цитотоксичность, повышенное связывание с NK-клетками, повышенное связывание с макрофагами, повышенное связывание с полиморфноядерными клетками, повышенное связывание с моноцитами, повышенное перекрестное связывание со связанными с (другими) мишенями антителами, повышенная прямая передача индуцирующих апоптоз сигналов, повышенное созревание дендритных клеток, повышенная активность праймирования Т-клеток (индукции дифференцировки).
В особенно предпочтительном воплощении клетка-хозяин данного изобретения представляет собой клетку СНО, клетку ВНК, клетку NSO, клетку SP2/0, клетку миеломы YO, клетку миеломы мыши Р3Х63, клетку PER, клетку PER.C6 или клетку гибридомы, а полипептид, продуцируемый указанной клеткой-хозяином, представляет собой анти-CD20 антитело, такое как IDEC-С2В8. В другом предпочтительном изобретении клетка-хозяин представляет собой антитело С225 против РЭФР (рецептор эпидермального фактора роста) человека.
В дополнение к нуклеиновой кислоте, кодирующей полипептид слияния данного изобретения, клетка-хозяин данного изобретения может содержать по меньшей мере один отрезок нуклеиновой кислоты, кодирующий молекулу антитела, сохраняющий свою функциональную Fc-область фрагмент антитела или белок слияния, который содержит область, эквивалентную Fc-области иммуноглобулина. В предпочтительных воплощениях этот по меньшей мере один отрезок нуклеиновой кислоты кодирует анти-CD20 антитело, химерное моноклональное антитело chCE7 против нейробластомы человека, моноклональное антитело chG250 против почечной клеточной карциномы, химерное моноклональное антитело ING-1 против карциномы ободочной кишки, легкого и груди, гуманизированное антитело 3622W94 против человеческого антигена 17-1А, гуманизированное антитело А33 против опухоли ободочной и прямой кишки, антитело R24 против меланомы человека, направленное к ганглиозиду GD3, химерное моноклональное антитело SF-25 против плоскоклеточной карциномы человека, антитело против РЭФР человека, антитело против рецептора к эпидермальному фактору роста РЭФР vIII человека, антитело против ПСМА (простат-специфический мембранный антиген), антитело против поверхностного антигена стволовой клетки простаты(PSCA), антитело против CD22 человека, антитело против CD30 человека, антитело против высокомолекулярного опухоль ассоциированного антигена меланомы человека(HMWMAA), антитело против ганглиозида GD3, антитело против ганглиозида GD2, антитело против ганглиозида GM2, антитело против ганглиозидов человека,, антитело против рецептора к эпидермальному фактору роста EGFRvIII (РЭФР)человека, антитело против интегрина, анти-CDSO антитело, анти-LeY антитело, антитело против муцина, анти-MUC18 антитело, антитело против CD33 человека, антитело против CD38 человека, антитело против CD40 человека, антитело против CD45 человека, антитело против CD52 человека, антитело против CD 138 человека, антитело против HLA (вариант HLA-DR), антитело против ЕрСАМ человека, антитело против СЕА человека, антитело против белка MUC1 человека, антитело против кора белка MUC1 человека, антитело против абберантно гликозилированного белка MUC1 человека, антитело против содержащих домен ED-B вариантов фибронектина человека или антитело против HER2/neu человека.
Настоящее изобретение также предусматривает способ продуцирования полипептида в клетке-хозяине, включающий (а) культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III, или, в качестве альтернативы, активностью бета-1,4-галактозилтрансферазы («GalT»), в условиях, которые допускают продуцирование клеткой полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела, сохранившего функциональную Fc-область, и белка слияния, который содержит область, эквивалентную Fc-области иммуноглобулина, где указанный полипептид, обладающий активностью GnTII или, в качестве альтернативы, активностью GalT, экспрессируется в количестве, достаточном для модификации олигосахаридов Fc-области указанного полипептида, продуцируемого клеткой-хозяином; и (b) выделение указанного полипептида. В предпочтительном воплощении полипептид слияния содержит каталитический домен бета-1,4-N-ацетилглюкозаминилтрансферазы. В особенно предпочтительном воплощении белок слияния также содержит отвечающий за локализацию в комплексе Гольджи домен какого-либо полипептида-резидента комплекса Гольджи. Предпочтительно, отвечающий за локализацию в комплексе Гольджи домен представляет собой домен локализации маннозидазы II или бета-1,2-N-ацетилглюкозаминилтрансферазы I (GnTI). В качестве альтернативы, отвечающий за локализацию в комплексе Гольджи домен выбирают из группы, состоящей из домена локализации маннозидазы I, домена локализации бета-1,2-N-ацетилглюкозаминилтрансферазы II и домена локализации альфа1-6 кор-фукозилтрансферазы. Полипептиды, продуцируемые способами согласно настоящему изобретению, обладают повышенной аффинностью связывания Fc-рецептора и/или повышенной эффекторной функцией. Повышенная эффекторная функция предспочтительно представляет собой повышение одной или более из следующих функций: Fc-опосредованная клеточная цитотоксичность (включая повышенную антителозависимую клеточную цитотоксичность), повышенный антителозависимый клеточный фагоцитоз (АЗКФ), повышенная секреция цитокинов, повышенное опосредованное иммунными комплексами поглощение антигена антиген-презентирующими клетками, повышенная Fc-опосредованная клеточная цитотоксичность, повышенное связывание с NK-клетками, повышенное связывание с макрофагами, повышенное связывание с моноцитами, повышенное связывание полиморфноядерными клетками, повышенное перекрестное связывание со связанными с (другими) мишенями антителами, повышенная прямая передача индуцирующих апоптоз сигналов, повышенное созревание дендритных клеток, повышенная активность праймирования Т-лимфоцитов. Повышенная аффинность связывания Fc-рецептора предпочтительно представляет собой повышенное связывание активирующего рецептора Fcγ, такого как рецептор FcγRIIIa.
В другом воплощении настоящее изобретение предусматривает полипептид, продуцируемый способами согласно настоящему изобретению, у которого увеличена доля разветвленных олигосахаридов в Fc-области указанного полипептида. В другом воплощении продуцируемый способами согласно настоящему изобретению полипептид имеет, в результате указанной модификации, повышенную долю нефукозилированных олигосахаридов в своем Fc-фрагменте. Указанные нефукозилированные олигосахариды могут быть олигосахаридами гибридного или сложного типа. В особенно предпочтительном воплощении полипептид, продуцируемый клеткой-хозяином и по способам данного изобретения, имеет увеличенную долю разветвленных (bisected) нефукозилированных олигосахаридов в Fc-области. Разветвленные нефукозилированные олигосахариды могут быть либо гибридными либо сложными. В частности, способы согласно настоящему изобретению могу применяться для продуцирования полипептидов, в которых по меньшей мере 15%, предпочтительно - по меньшей мере 20%, более предпочтительно - по меньшей мере 25%, еще более предпочтительно - по меньшей мере 30%), еще более предпочтительно - по меньшей мере 35%, еще более предпочтительно -по меньшей мере 40%, еще более предпочтительно - по меньшей мере 45%, еще более предпочтительно - по меньшей мере 50%, еще более предпочтительно - по меньшей мере 55%, еще более предпочтительно - по меньшей мере 60%, еще более предпочтительно - по меньшей мере 70%, еще более предпочтительно - по меньшей мере 80%, еще более предпочтительно - по меньшей мере 85%, еще более предпочтительно - по меньшей мере 90%, еще более предпочтительно - по меньшей мере 95%, еще более предпочтительно - по меньшей мере 96%, еще более предпочтительно - по меньшей мере 97%, еще более предпочтительно - по меньшей мере 98%, еще более предпочтительно - по меньшей мере 99% олигосахаридов в Fc-области полипептида представляют собой нефукозилированные олигосахариды. Способы согласно настоящему изобретению могу также применяться для продуцирования полипептидов, в которых по меньшей мере 15%, более предпочтительно - по меньшей мере 20%, еще более предпочтительно - по меньшей мере 25%, еще более предпочтительно - по меньшей мере 30%, еще более предпочтительно - по меньшей мере 35%, еще более предпочтительно - по меньшей мере 40%, еще более предпочтительно - по меньшей мере 45%, еще более предпочтительно - по меньшей мере 50%, еще более предпочтительно - по меньшей мере 55%, еще более предпочтительно - по меньшей мере 60%, еще более предпочтительно - по меньшей мере 70%, еще более предпочтительно -по меньшей мере 80%, еще более предпочтительно - по меньшей мере 85%, еще более предпочтительно - по меньшей мере 90%, еще более предпочтительно - по меньшей мере 95%, еще более предпочтительно - по меньшей мере 96%, еще более предпочтительно -по меньшей мере 97%, еще более предпочтительно - по меньшей мере 98%, еще более предпочтительно - по меньшей мере 99% олигосахаридов в Fc-области полипептида представляют собой разветвленные (bisected) олигосахариды. Далее, способы согласно настоящему изобретению могу применяться для продуцирования полипептидов, в которых по меньшей мере 15%, более предпочтительно - по меньшей мере 20%, еще более предпочтительно -по меньшей мере 25%, еще более предпочтительно - по меньшей мере 30%, еще более предпочтительно - по меньшей мере 35%, еще более предпочтительно - по меньшей мере 40%, еще более предпочтительно - по меньшей мере 45%, еще более предпочтительно - по меньшей мере 50%, еще более предпочтительно - по меньшей мере 55%, еще более предпочтительно - по меньшей мере 60%, еще более предпочтительно - по меньшей мере 70%, еще более предпочтительно - по меньшей мере 80%, еще более предпочтительно - по меньшей мере 85%, еще более предпочтительно - по меньшей мере 90%, еще более предпочтительно - по меньшей мере 95%, еще более предпочтительно - по меньшей мере 96%, еще более предпочтительно - по меньшей мере 97%, еще более предпочтительно - по меньшей мере 98%, еще более предпочтительно - по меньшей мере 99% олигосахаридов в Fc-области полипептида представляют собой разветвленные (bisected), нефукозилированные олигосахариды. Способы согласно настоящему изобретению могу также применяться для продуцирования полипептидов, в которых по меньшей мере 15%, более предпочтительно - по меньшей мере 20%, еще более предпочтительно - по меньшей мере 25%, еще более предпочтительно - по меньшей мере 30%, еще более предпочтительно - по меньшей мере 35% олигосахаридов в Fc-области полипептида являются разветвленными гибридными нефукозилированными олигосахаридами.
В другом воплощении, настоящее изобретение предусматривает антитело, модифицированное для повышения эффекторной функции и/или повышения аффинности связывания Fc-рецептора. Предпочтительно, повышенная эффекторная функция является одной из следующих: повышенная Fc-опосредованная клеточная цитотоксичность (включая повышенную антителозависимую клеточную цитотоксичность), повышенный антителозависимый клеточный фагоцитоз (АЗКФ), повышенная секреция цитокинов, повышенное опосредованное иммунными комплексами поглощение антигена антиген-презентирующими клетками, повышенное связывание с NK-клетками, повышенное связывание с макрофагами, повышенное связывание с моноцитами, повышенное связывание полиморфноядерными клетками, повышенное перекрестное связывание со связанными с (другими) мишенями антителами, повышенная прямая передача индуцирующих апоптоз сигналов, повышенное созревание дендритных клеток, повышенная активность праймирования Т-клеток (индукции дифференцировки). В предпочтительном воплощении повышенная аффинность связывания Fc-рецептора представляет собой повышенное связывание активирующего рецептора Fcγ, наиболее предпочтительно - с рецептором FcγRIIIa. Изобретение далее предусматривает фрагменты антитела, содержащие Fc-область и белки слияния, которые содержат область, эквивалентную Fc-области иммуноглобулина. Такие фрагменты антител и белки слияния проявляют повышенную аффинность связывания Fc-рецептора и/ил повышенную эффекторную функцию.
Настоящее изобретение далее предусматривает фармацевтические составы, содержащие антитела, сохранившие Fc-область фрагменты антител и белки слияния, имеющие область, эквивалентную Fc-области иммуноглобулина согласно настоящему изобретению и фармацевтически приемлемый носитель.
Настоящее изобретение далее предусматривает применение таких фармацевтических составов в способе лечения рака. В частности, настоящее изобретение предусматривает способ лечения рака, включающий назначение терапевтически эффективного количества фармацевтических составов по данному изобретению.
Настоящее изобретение также предусматривает клетку-хозяина, содержащую вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид слияния, где указанный полипептид обладает активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержит отвечающий за локализацию в комплексе Гольджи какого-либо белка-резидента комплекса Гольджи, а также вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид, где полипептид обладает активностью маннозидазы II (ManII). В предпочтительных воплощениях молекула нуклеиновой кислоты, кодирующая полипептид слияния, и молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью маннозидазы И, находятся в одном или в отдельных векторах экспрессии. В другом предпочтительном воплощении полипептид слияния содержит каталитический домен GnTIII. В дополнительном предпочтительном изобретении отвечающий за локализацию в комплексе Гольджи домен представляет собой домен локализации ManII, бета-1,2-N-ацетилглюкозаминилтрансферазы II, маннозидазы I или альфа 1,6-N кор-фукозилтрансферазы.
В дальнейшем предпочтительном воплощении, клетку-хозяина выбирают из группы, состоящей из клетки дрожжевой клетки, клетки насекомого или клетки растения. Далее изобретение предусматривает клетку-хозяина, содержащую вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо белка-резидента комплекса Гольджи, вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид, который обладает активностью маннозидазы II (ManII), и вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид, где полипептид обладает активностью бета-1,2-N-ацетилглюкозаминилтрансферазы II (GnTII). В предпочтительных воплощениях молекула нуклеиновой кислоты, кодирующая полипептид слияния, молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью ManII, и молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью GnTII, находятся в одном, или в отдельных векторах экспрессии. Также предпочтительным является, чтобы молекула нуклеиновой кислоты, кодирующая полипептид слияния находилась в одном векторе экспрессии, а молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью ManII, и молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью GnTII, - в другом векторе экспрессии. Также предпочтительным является, чтобы молекула нуклеиновой кислоты, кодирующая ManII, находилась в одном векторе экспрессии, а молекула нуклеиновой кислоты, кодирующая полипептид слияния, и молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью GnTII, - в другом векторе экспрессии. В другом воплощении GnTII находится в одном векторе экспрессии, а молекула нуклеиновой кислоты, кодирующая полипептид слияния, и молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью ManII - другом векторе экспрессии.
В дополнительном аспекте изобретение предусматривает клетку-хозяина, содержащую вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид слияния, где полипептид слияния обладает активностью бета-1,4-N-галактозилтрансферазы (GalT) и содержит отвечающий за локализацию в комплексе Гольджи какого-либо полипептида-резидента комплекса Гольджи, а также вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид, где полипептид обладает активностью маннозидазы II (ManII). В предпочтительных воплощениях молекула нуклеиновой кислоты, кодирующая полипептид слияния, и молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью маннозидазы П, находятся в одном или в отдельных векторах экспрессии. Предпочтительно, полипептид слияния содержит каталитический домен GalT. В дополнительном воплощении отвечающий за локализацию в комплексе Гольджи домен представляет собой домен локализации ManII, бета-1,2-N-ацетилглюкозаминилтрансферазы I, бета-1,2-N-ацетилглюкозаминилтрансферазы II, маннозидазы I или альфа 1,6-N кор-фукозилтрансферазы. В предпочтительном воплощении клетку-хозяина выбирают из группы, состоящей из клетки млекопитающего, дрожжевой клетки, клетки насекомого или клетки растения. Предпочтительно клетка-хозяин представляет собой клетку СНО, клетку ВНК, клетку NSO, клетку SP2/0, клетку миеломы YO, клетку миеломы мыши Р3Х63, клетку PER, клетку PER, клетку PER. С6 или клетку гибридомы.
В дополнительном аспекте изобретение предусматривает клетку-хозяина, содержащую вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид слияния, где полипептид слияния обладает активностью бета-1,4-N-галактозилтрансферазы (GalT) и содержит отвечающий за локализацию в комплексе Гольджи домен какого-либо белка-резидента комплекса Гольджи, а также вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид, где полипептид обладает активностью маннозидазы II (ManII), и вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид, который обладает активностью бета-1,2-N-ацетилглюкозаминилтрансферазы II (GnTII). Предпочтительно, все нуклеиновые кислоты находятся в одном векторе экспрессии. В другом способе реализации изобретения каждая из молекул нуклеиновой кислоты находится в отдельном векторе экспрессии. Изобретение далее предусматривает, что молекула нуклеиновой кислоты, кодирующая полипептид слияния, находится в одном векторе экспрессии, а молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью ManII, и молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью GnTII, - другом векторе экспрессии. Изобретение также предусматривает, что молекула нуклеиновой кислоты, кодирующая ManII, находится в одном векторе экспрессии, а молекула нуклеиновой кислоты, кодирующая полипептид слияния, и молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью GnTII - в другом векторе экспрессии. Изобретение также предусматривает, что молекула нуклеиновой кислоты, кодирующая GnTII, находится в одном векторе экспрессии, а молекула нуклеиновой кислоты, кодирующая о полипептид слияния, и молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью ManII - в другом векторе экспрессии. Далее в предпочтительном воплощении отвечающий за локализацию в комплексе Гольджи домен представляет собой домен локализации ManII, бета-1,2-N-ацетилглюкозаминилтрансферазы I, бета-1,2-N-ацетилглюкозаминилтрансферазы II, маннозидазы I или альфа 1,6-N кор-фукозилтрансферазы.
Изобретение далее предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью GnTIII, и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью ManII, в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, где полипептид, продуцируемый клеткой-хозяином, выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела и белка слияния, который содержит область, эквивалентную Fc-области иммуноглобулина.
Изобретение также предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью GnTIII, по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью ManII, и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью GnII, в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, где полипептид, продуцируемый клеткой-хозяином, выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела и белка слияния, который содержит область, эквивалентную Fc-области иммуноглобулина.
Настоящее изобретение дополнительно предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью GalT, и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью ManII, в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, где полипептид, продуцируемый клеткой-хозяином, выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела и белка слияния, который содержит область, эквивалентную Fc-области иммуноглобулина.
В отдельном воплощении настоящее изобретение также предусматривает клетку-хозяина, модифицированную для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью GalT, по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью ManII, и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью GnIIT, в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, где полипептид, продуцируемый клеткой-хозяином, выбирают из группы, состоящей из целой молекулы антитела, фрагмента антитела и белка слияния, который содержит область, эквивалентную Fc-области иммуноглобулина. Предпочтительно, полипептид, продуцируемый клеткой-хозяином проявляет, в результате модификации, повышенную аффинность связывания Fc-рецептора. В дополнительно предпочтительном способе реализации изобретения полипептид, продуцируемый клеткой-хозяином проявляет, в результате модификации, повышенную эффекторную функцию. Предпочтительно, повышенная эффекторная функция представляет собой одну или более из следующих: повышенная Fc-опосредованная клеточная цитотоксичность повышенное связывание с NK-клетками, повышенное связывание с макрофагами, повышенное связывание полиморфноядерными клетками, повышенное связывание с моноцитами, повышенное перекрестное связывание со связанными с (другими) мишенями антителами, повышенная прямая передача индуцирующих апоптоз сигналов, повышенное созревание дендритных клеток и/ил повышенная активность праймирования Т-клеток (индукции дифференцировки).
В дополнительном воплощении изобретение предусматривает способ продуцирования полпептида в клетке-хозяине, включающий культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью GnTIII, и по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью ManII, в условиях, которые допускают продуцирование полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела и белка слияния, который содержит область, эквивалентную Fc-области иммуноглобулина, где полипептид слияния экспрессируется в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, а также выделение этого полипептида. Предпочтительно, клетку-хозяина также модифицируют для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью GnTII. В дополнительно предпочтительном воплощении полипептид слияния содержит каталитический домен GnTIII. В дальнейшем предпочтительном воплощении полипептид слияния содержит также отвечающий за локализацию в комплексе Гольджи домен какого либо гетерологичного полипептида-резидента комплекса Гольджи. Предпочтительно, отвечающий за локализацию в комплексе Гольджи домен представляет собой домен локализации маннозидазы II, бета-1,2-N-ацетилглюкозаминилтрансферазы I, маннозидазы I, бета-1,2-N-ацетилглюкозаминилтрансферазы II или альфа 1,6-N кор-фукозилтрансферазы. Предпочтительно, полипептид обладает, в результате описанной выше модификации, повышенной эффекторной функцией.
Изобретение далее предусматривает способ продуцирования полипептида в клетке-хозяине, включающий культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью GalT, и по меньшей мере одной нуклеиновой кислоты, кодирующей обладающий активностью ManII полипептид, в условиях, которые допускают продуцирование полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела и белка слияния, который содержит область, эквивалентную Fc-области иммуноглобулина, где полипептид слияния экспрессируется в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, а также выделение этого полипептида. В дополнительном воплощении клетку-хозяина также модифицируют для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью GnTII. Предпочтительно, полипептид слияния содержит каталитический домен GalT. Также предпочтительно, полипептид слияния содержит также отвечающий за локализацию в комплексе Гольджи домен какого либо гетерологичного полипептида-резидента комплекса Гольджи. Предпочтительно, отвечающий за локализацию в комплексе Гольджи домен представляет собой домен локализации маннозидазы II, бета-1,2-N-ацетилглюкозаминилтрансферазы I, маннозидазы I, бета-1,2-N-ацетилглюкозаминилтрансферазы II или альфа 1,6-N кор-фукозилтрансферазы. Предпочтительно, полипептид обладает, в результате описанной выше модификации, повышенной эффекторной функцией. Конкретно, в предпочтительном воплощении, полипептид, продуцируемый клеткой-хозяином, имеет увеличенную долю разветвленных (bisected) нефукозилированных олигосахаридов в Fc-области полипептида. Предпочтительно, разветвленные нефукозилированные олигосахариды являются гибридными. Даже более предпочтительно, чтобы разветвленные нефукозилированные являлись сложными. В предпочтительном воплощении по меньшей мере приблизительно от 10% до 95% олигосахаридов в Fc-области полипептида являются разветвленными нефукозилированными. Особенно предпочтительно, чтобы приблизительно 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% олигосахаридов Fc-области гликомодифицированных полипептидов согласно настоящему изобретению представляли собой разветвленные и нефукозилированные олигосахариды.
В дополнительно предпочтительном воплощении настоящее изобретение предусматривает, что антитела, модифицированные согласно способам настоящего изобретения, обладают повышенной эффекторной функцией.
В дополнительном воплощении изобретение далее предусматривает фармацевтические составы, содержащие антитела, модифицированные согласно способам настоящего изобретения. Предпочтительно, чтобы фармацевтические составы данного изобретения содержали фармацевтически приемлемый носитель.
Изобретение далее предусматривает способ лечения раковых опухолей, включающий назначение терапевтически эффективного количества фармацевтического состава согласно настоящему изобретению нуждающемуся в лечении пациенту. Изобретение далее предусматривает способ продуцирования полипептида, обладающего повышенной Fc-опосредованной клеточной цитотоксичностью, в клетке-хозяине, включающий культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей GalT и по меньшей мере одной нуклеиновой кислоты, кодирующей ManII, в условиях, которые допускают продуцирование полипептида, выбранного из группы, состоящей из целой молекулы антитела и фрагмента антитела, который содержит Fc-область иммуноглобулина, где уровень экспрессии одного из полипептидов GalT и ManII, или их обоих, достаточен для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, и где полипептид обладает, в результате модификации, повышенной Fc-опосредованной клеточной цитотоксичностью, а также выделение полипептида, обладающего повышенной Fc-опосредованной клеточной цитотоксичностью. В дополнительно предпочтительном способе реализации изобретения уровень экспрессии GalT пориводит к появлению молекулы антитела или фрагмента антитела, которые содержат Fc-область иммуноглобулина, обладающую, повышенной Fc-опосредованной клеточной цитотоксичностью.
Изобретение далее предусматривает способ продуцирования полипептида, обладающего повышенной Fc-опосредованной клеточной цитотоксичностью, в клетке-хозяине, включающий культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей GalT, и по меньшей мере одной нуклеиновой кислоты, кодирующей ManII, в условиях, которые допускают продуцирование полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела который содержит Fc-область иммуноглобулина, где уровень экспрессии одного из полипептидов GalT и ManII, или их обоих, достаточен для модификации олигосахаридов в Fc-области продуцируемого клеткой-хозяином полипептида, и где полипептид обладает, в результате модификации, повышенной Fc-опосредованной клеточной цитотоксичностью, а также выделение полипептида, обладающего повышенной Fc-опосредованной клеточной цитотоксичностью. В предпочтительном воплощении описанная выше клетка-хозяин также содержит по меньшей мере одну нуклеиновую кислоту, кодирующую GnTIII, где GnTIII экспрессируется в количестве, достаточном для модификации олигосахаридов в Fc-области полипептида, продуцируемого клеткой-хозяином, где полипептид обладает, в результате модификации, повышенной Fc-опосредованной клеточной цитотоксичностью. Предпочтительно, уровень экспрессии одного или более из полипептидов GalT, ManII и GnTIII достаточен для образования разветвленных (bisected) олигосахаридов в Fc-области полипептида. Даже более предпочтительно, чтобы доля разветвленны[олигосахаридов в Fc-области полипептида составляла по меньшей мере приблизительно 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90 или 95 процентов. Предпочтительно, чтобы доля разветвленны [олигосахаридов в Fc-области составляла по меньшей мере 45 процентов. В предпочтительном воплощении разветвленные олигосахариды являются гибридными. Предпочтительно, клетка-хозяин представляет собой клетку млекопитающего, дрожжевую клетку или клетку растения. Даже более предпочтительно, чтобы клетка-хозяин представляла собой клетку растения.
В другом аспекте настоящее изобретение предусматривает способ продуцирования полипептида клеткой-хозяином, включающий:
a. культивирование клетки-хозяина, модифицированной для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей полипептид, обладающий активностью альфа-маннозидазы II с условиях, которые допускают продуцирование полипептида, выбранного из группы, состоящей из целой молекулы антитела, фрагмента антитела и белка слияния, который содержит область, эквивалентную Fc-области иммуноглобулина, где указанный полипептид, обладающий активностью альфа-маннозидазы II экспрессируется в количестве, достаточном для модификации олигосахаридов в Fc-области указанного полипептида, продуцируемого клеткой-хозяином; и
b. Выделение указанного полипептида, продуцируемого указанной клеткой-хозяином.
Настоящее изобретение также предусматривает полипептиды, в особенности - антитела, обладающие, в результате указанной модификации олигосахаридов, повышенной эффекторной функцией и/или повышенным связыванием Fc-рецептора, а равно и на их применение в терапевтических составах для лечения расстройств, в особенности - опухолей.
Идентификация и получение нуклеиновых кислот, кодирующих белок, рисунок гликозилирования которого желательно модифицировать.
Настоящее изобретение предусматривает способы получения и применения систем «клетка-хозяин» для получения гликоформ антител, фрагментов антител и белков слияния, которые содержат область, эквивалентную Fc-области иммуноглобулина, обладающих повышенной аффинностью связывания Fc-рецептора (предпочтительно -активирующего Fc-рецептора) и/или повышенными эффекторными функциями, включая антителозависимую клеточную цитотоксичность. Идентификация целевых эпитопов и создание имеющих возможное терапевтическое значение антител, рисунок (pattern) гликозилирования которых желательно модифицировать, а также выделение соответствующих кодирующих последовательностей нуклеиновых кислот находятся в рамках области охвата изобретения.
Для получения антител к целевым эпитопам могут быть использованы разнообразные известные в данной области процедуры. Такие антитела включают без огарничения: поликлональные, моноклональные, химерные, гуманизированные, полностью человеческие и одноцепочечные антитела, фрагменты Fab и фрагменты, полученные из экспрессионной библиотеки ScFv, Fab, VH, IgG. Такие антитела могут быть полезны в качестве, например, диагностических или терапевтических агентов. Нейтрализующие антитела, т.е. те антитела, которые конкурируют за связывание с лигандом, субстратом или адаптерной молекулой, представляют особенный интерес в качестве терапевтических агентов.
Для получения антител разнообразных животных (хозяев), включая, но не ограничиваясь, кроликов, мышей, крыс и т.д., иммунизируют путем инъекции целевым белком-мишенью. Можно получить большое количество поликлональных антител путем множественных подкожных (пк) или внутрибрюшинных (вб) инъекций соответствующего антигена и адъюванта. Для повышения иммунологического ответа можно применять, в зависимости от вида животного-хозяина, различные адъюванты, включая, но не ограничиваясь, адъювант Фрейнда (полный и неполный), минеральные гели, такие как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, полиспирты плюроника, полианионы, белки, сапонин, масляные эмульсии, гемоцианин морского блюдечка (KLH, ГМБ), динитрофенол и потенциально полезные адъюванты человека, такие как BCG (bacilli Calmette-Guerin) и Corinobacterium parvum. Животных иммунизируют против антигена, иммуногенных конъюгатов или производных путем смешивания, например, 100 г или 5 г белка или конъюгата (для кролика или мыши, соответственно) с тремя объемами полного адъюванта Фрейнда и последующего введения раствора путем подкожной инъекции во множественные сайты. Через месяц проводят поддерживающую иммунизацию путем инъекции во множественные сайты от 1/5 до 1/10 первоначального объема белка или конъюгата в адъюванте Фрейнда. Через 7-14 дней у животных забирают кровь и определяют титр антител. Поддерживающие иммунизации проводят до тех пор, пока титр не перестанет увеличиваться. Предпочтительно, поддерживающие иммунизации животного проводят одним и тем же антигеном, но конъюгированным с другим белком и/или через другое связывающее вещество. Моноклональные антитела к целевой мишени могут быть приготовлены с использованием любых технологий, которые предусматривают продуцирование молекул антител непрерывными клеточными линиями в культуре. Такие технологии включают без огарничения: гибридомную технологию, впервые описанную в Kohler, Milstein, Nature 256: 495-97 (1975), гибридомную технологию В-клеток человека (Kosbor et al., Immunology Today 4: 72 (1983); Cote et al., Proc. Natl. Acad. Set U. S. A. 80: 2026-30 (1983) и технологию EBV-гибридомы (Cole et al, Monoclonal Antibodies and Cancer Therapy 77-96 (Alan R. Liss, Inc., 1985)). В дополнение может быть задействована технология получения «химерных антител» (Morrison et al., Proc. Natl. Acad. Sci. U.S.A. 81: 6851-55 (1984); Neuberger et al., Nature 312: 604-08 (1984); Takeda et al., Nature 314: 452-54 (1985) путем сплайсинга (соединения) генов молекулы мышиного антитела соответствующей антагенной специфичности с генами молекулы человеческого антитела с подходящей биологической активностью. Эти технологии могут быть использованы также для получения химерных антител, состоящих из молекул антител других млекопитающих. В качестве альтернативы можно адаптировать для получения одноцепочечных антител желаемой специфичности технологию получения одноцепочечных антител (Патент США No. 4,946, 778). Изобретение далее предусматривает гуманизированные антитела, подвергнутые гликомодификации согласно способам настоящего изобретения. Технологии создания гуманизированных антител раскрыты, например в Патенте США U.S. Pat. No. 6,180, 320 to Queen et al, полные содержания которых полностью включены в описание изобретения путем ссылки.
Фрагменты антитела, которые содержат сайты специфического связывания целевого белка-мишени, могут быть получены по известным технологиям. Например, такие фрагменты включают без ограничения: F(ab')2-фрагменты, которые могут быть получены путем расщепления пепсином молекулы антитела, и Fab-фрагменты, которые могут быть получены путем уничтожения дисульфидных мостиков фрагментов F(ab')2. В качестве альтернативы, могут быть созданы экспрессионные библиотеки Fab(Huse et al., Science 246: 1275-81 (1989), что позволит быстро и легко идентифицированть моноклональные Fab-фрагменты желаемой специфичности к целевому белку-мишени.
После идентификации антитела или фрагмента антитела, паттерн гликозилирования которого желательно модифицировать, идентифицируют и выделяют кодирующую последовательность нуклеиновой кислоты, для чего используют хорошо известные в данной области технологии.
а. Получение клеточных линий для продуцирования белков с измененным паттерном (pattern - вариантом, рисуноком) гликозилирования.
Настоящее изобретение предусматривает системы экспрессии «клетка-хозяин» для получения белков с модифицированным паттерном гликозилирования. В частности, настоящее изобретение предусматривает системы «клетка-хозяин» для получения гликоформ белков повышенной терапевтической ценности. Поэтому, в одном аспекте изобретение предусматривает экспрессионные системы «клетка-хозяин» выбранные или модифицированные для экспрессии, например, белка слияния, обладающего активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащего отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи. В частности, такие экспрессионные системы «клетка-хозяин» могут быть модифицированы рекомбинантной молекулой нуклеиновой кислоты, кодирующей такой белок слияния, функционально связанной с конститутивной или регулируемой промоторной системой.
В одном особом воплощении, настоящее изобретение предусматривает клетку-хозяина, которая была модифицирована для экспрессии по меньшей мере одной нуклеиновой кислоты, кодирующей белок слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи. В одном аспекте клетка-хозяин модифицирована молекулой нуклеиновой кислоты, содержащей по меньшей мере один ген, кодирующий полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи.
В целом, любой тип культивируемой клеточной линии может быть использован в качестве основы для конструирования клеточных линий согласно настоящему изобретению. В предпочтительном воплощении в качестве основы для получения модифицированных клеток-хозяев данного изобретения использованы, дрожжевые клетки, клетки насекомых или клетки растений. (См. Ma, J. K.-С.et al., Nature Genetics 4: 794-805 (Октябрь 2003) и цитируемые там ссылки) (полное содержание которых включено в описание изобретения путем ссылки).
Предполагается, что изобретение включает модифицированные клетки-хозяева, экспрессирующие полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи.
Одна или несколько аминокислот, кодирующих полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, могут экспрессироваться под контролем конститутивного промотора или, в качестве альтернативы, регулируемой экспрессионной системы. Подходящие регулируемые экспрессионные системы включают без ограничения: тетрациклин-регулируемую экспрессионную систему, экдизон-индуцируемую систему экспрессии, систему экспрессии, содержащую lac-промтор, глюкокортикоид-индуцируемую систему экспрессии, индуцируемую температурой промотроную систему и металлотиониновую металл-индуцируемую систему экспрессии. Если в системе «клетка-хозяин» содержится несколько молекул нуклеиновой кислоты, кодирующих слитые белки, обладающие активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащие отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, некоторые из них могут экспрессироваться под контролем конститутивного промотора, в то время как другие экспрессируются под контролем регулируемого промотора. Под максимальным уровнем экспрессии подразумевают самый высокий из возможных уровень стабильной экспрессии белка слияния, который не оказывает значительного отрицательного воздействия на скорость роста клетки. Максимальный уровень экспрессии может быть определен в ходе рутинных экспериментов. Уровни экспрессии определяют широко известными в данной области методами, включая Вестерн-блот с использованием антитела, специфического к белку, обладающему активностью GnTIII, или антитела, специфического к участку - метке белка, присоединенному к полипептиду, обладающему активностью GnTIII, Нозерн-блот с использованием нуклеиновой кислоты-зонда, специфического к гену, кодирующему белок слияния, обладающий активностью GnTIII или нуклеиновой кислоты-зонда, специфичной к нуклеиновой кислоте, кодирующей участок белка - метку, слитый с полипептидом, обладающим активностью GnTIII, или измерение активности GnTIII. В качестве альтернативы возможно применение лектина, который связывается с биосинтетическими производными GnTIII, например, лецитин Е4-РНА. В качестве альтернативы можно применить функциональное исследование для измерения связывания Fc-рецептора или повышенной эффекторной функции, опосредуемой антителами, продуцируемыми клетками, которые были модифицированы нуклеиновой кислотой, кодирующей полипептид с активностью GnTIII. В качестве другой альтернативы, нуклеиновая кислота может быть функционально связана с репортерным геном (геном-репортером); уровни экспрессии полипептида слияния, обладающего активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III(GnTIII) и содержащего отвечающий за локализацию в комплексе Гольджи какого-либо гетерологичного полипептида-резидента комплекса Гольджи, определяют путем измерения сигнала, связанного с уровнем экспрессии репортерного гена. Репортерный ген может транскрибироваться совместно с нуклеиновой кислотой (кислотами), кодирующей белок слияния с образованием единой молекулы мРНК; их соответствующие кодирующие последовательности могут быть соединены либо внутренним сайтом посадки рибосом (IRES), либо энхансером кэп-независимой трансляции (CITE). Репортерный ген может транслироваться совместно с по меньшей мере одной молекулой нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III(GnTIII) и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, с образованием единой полипептидной цепи. Нуклеиновые кислоты, кодирующие полипептиды слияния согласно настоящему изобретению, могут быть функционально связаны с репортерным геном и поставлены под контроль единого (общего) промотора, и таким образом, чтобы нуклеиновая кислота, кодирующая полипептид слияния, и репортерный ген транскрибировались в молекулу РНК, которая путем альтернативного сплайсинга превращается в отдельные молекулы матричной РНК (мРНК); одна из полученных в результате молекул мРНК транслируется затем в репортерный белок, а другая транслируется в полипептид слияния.
Если экспрессируются несколько разных нуклеиновых кислот, кодирующих полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, они могут быть организованы таким образом, чтобы в результате транскрипции образовывалась одна или несколько молекул мРНК. Если они транскрибируются как единая молекула мРНК, соответствующие кодирующие последовательности могут быть соединены либо внутренним сайтом посадки рибосом (IRES), либо энхансером кэп-независимой трансляции (CITE). Их транскрипция может инициироваться общим промотором и приводить к образованию молекулы РНК, которая путем альтернативного сплайсинга превращается в несколько отдельных молекул матричной РНК (мРНК), каждая из которых затем транслируется в соответствующий закодированный ею белок слияния.
В других воплощениях настоящее изобретение предусматривает системы экспрессии «клетка-хозяин» для получения терапевтических антител, обладающих повышенной аффинностью связывания Fc-рецептора, в частности, активирующих Fc-рецепторов, и повышенной эффекторной функцией, включая антителозависимую клеточную цитотоксичность. Обычно, системы экспрессии «клетка-хозяин» модифицируют и/или выбирают для экспрессии нуклеиновых кислот, кодирующих антитело, измененные гликоформы которого желательно получить, одновременно с по меньшей мере одной нуклеиновой кислотой, кодирующей полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи. В одном воплощении в систему «клетка-хозяин» трансфецируют по меньшей мере одной нуклеиновой кислотой, кодирующей такой полипептид слияния. Обычно трансфецированные клетки подвергают отбору (селекции) для идентификации и выделения клонов, которые стабильно экспрессируют белок слияния данного изобретения.
Культивируемая клеточная линия любого типа может быть использована в качестве основы для конструирования клеточных линий согласно настоящему изобретению. В предпочтительном воплощении могут быть использованы клетки дрожжевые клетки, клетки насекомых или клетки растений. Обычно, такие клеточные линии модифицируют таким образом, чтобы они также содержали по меньшей мере одну, введенную путем трансфекции, нуклеиновую кислоту, кодирующую целую молекулу антитела, фрагмент антитела, содержащий Fc-область иммуноглобулина, или белок слияния, который включает область, эквивалентную Fc-области иммуноглобулина. Обычно такие клеточные
линии для получения антитела происходят от клонов, продуцирующих и секретирующих антитела с высокой специфической продуктивностью в диапазоне между 20 и 120 пг/(клетка⋅день). В альтернативном воплощении, в качестве основы для получения модифицированных клеток-хозяев данного изобретения использована гибридомная клеточная линия, экспрессирующая отдельное целевое антитело.
В одном воплощении нуклеиновые кислоты, кодирующие антитело, фрагмент антитела, или Fc-полипептид слияния, клонируют в векторы экспрессии антитела затем вводят путем трансфекции в клетки-хозяева и ищут и отбирают клеточные клоны с высокой и стабильной специфической продуктивностью выработки антител. Затем такие отобранные клоны трансфецируют векторами экспрессии гликопротеин-модифицирующих гликотрансфераз, содержащих нуклеиновые кислоты, кодирующие, например, (а) полипептид слияния с активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III(GnTII), или (b) полипептид слияния с активностью бета-1,4-галактозилтрансферазы (GalT), или (с) полипептид слияния с активностью альфа-маннозидазы II(ManII) комплекса Гольджи, или (d) полипептид слияния с активностью GnTIII а также полипептид с активностью ManII, или (е) полипептид слияния с активностью GalT а также полипептид с активностью ManII. Клоны затем подвергают селеции (антибиотиками) и среди них выбирают колонии клеток, отличающиеся стабильной экспрессией генов, кодирующих антитела, причем уровни экспрессии должны обеспечивать высокую специфическую продуктивность выработки антител, а также колонии стабильной экспрессией генов гликопротеин-модифицирующих гликотрансфераз, причем уровни экспрессии должны обеспечивать модификацию паттерна гликозилирования Fc-области, включая увеличение доли нефукозилированных олигосахаридов (которые могут быть разветвленными либо неразветвленными, гибридными или сложными), ассоциированной с повышением аффинности связывании Fc-рецептора, в частности, с увеличением аффинности связывания Fc-FcγRIII, а также повышением эффекторных функций, опосредуемых Fc-рецептором, включая, но не ограничиваясь, Fc-зависимую клеточную цитотоксичность. Методы селекции и отбора описаны ниже.
В другом воплощении две описанные выше трансфекции, а именно, трансфекцию векторами экспрессии антитела и трансфекцию векторами экспрессии гликопротеин-модифицирующих гликотрансфераз, проводят в обратном порядке, т.е. клетку-хозяина вначале трансфецируют векторами экспрессии гликопротеин-модифицирующих гликотрансфераз, а затем - векторами экспрессии антитела. При таком подходе среди клонов, полученных после первой трансфекции, могут быть выявлены клоны, дающие адекватный стабильный уровень экспрессии генов гликозилтрансфераз. Выявление таких клонов можно осуществлять любым из описанных ниже способов, либо, в качестве альтернативы, путем временной трансфекции репликатов таких клонов вектором экспрессии антитела и последующего применения описанных ниже способов скрининга с целью идентификации клонов со стабильным уровнем экспрессии генов гликозилтрансфераз, причем уровни экспрессии должны обеспечивать модификацию паттерна гликозилирования Fc-области и повышение аффинности связывания Fc-рецептора, включая рецепторы Fc-FcγRIII, а также повышение Fc-опосредуемых эффекторных функций, включая, но не ограничиваясь, Fc-зависимую клеточную цитотоксичность. Методы селекция и отбора описаны ниже.
В дальнейшем воплощении гены, кодирующий антитело, и гены гликозилтрансфераз трансфецируют совместно в рамках общей процедуры трансфекции в единственном векторе экспрессии или в отдельных векторах.
Обычно, по меньшей мере одна нуклеиновая кислота в системе «клетка-хозяин» кодирует полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) или, в качестве альтернативы, активностью бета-1,4-галактозилрансферазы, и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи. В качестве альтернативы нуклеиновая кислота кодирует полипептид с активностью альфа-маннозидазы II комплекса Гольджи.
Одна или несколько нуклеиновых кислот, кодирующих полипептид данного изобретения, могут экспрессироваться под контролем конститутивного промотора или, в качестве альтернативы, регулируемой экспрессионной системы. Подходящие регулируемые экспрессионные системы включают без огарничения: тетрациклин-регулируемую экспрессионную систему, экдизон-индуцируемую систему экспрессии, систему экспрессии, содержащую lac-промотор, глюкокортикоид-индуцирумую систему экспрессии, индуцируемую температурой промотроную систему и металлотиониновую металл-индуцируемую систему экспрессии. Если в системе «клетка-хозяин» содержится несколько разных молекул нуклеиновой кислоты, кодирующих слитые белки, обладающие активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащие отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, некоторые из них могут экспрессироваться под контролем конститутивного промотора, в то время как другие экспрессируются под контролем регулируемого промотора. Под максимальным уровнем экспрессии подразумевают самый высокий из возможных уровень стабильной экспрессии слитого белка, который не оказывает отрицательного воздействия на скорость роста клетки. Максимальный уровень экспрессии может быть определен в ходе рутинных экспериментов. Уровни экспрессии определяют общеизвестными в данной области методами, включая Вестерн-блот с использованием, например, антитела, специфического к белку, обладающему активностью GnTIII, или антитела, специфичного к отрезку-метке белка, слитого с полипептидом, обладающим активностью GnTIII, Нозерн-блот с использованием нуклеиновой кислоты-зонда, специфического, например, к гену, кодирующему белок слияния, обладающий активностью GnTIII или нуклеиновой кислоты-зонда, специфического к гену, кодирующему белковую метку, слитую с полипептидом, обладающим активностью GnTIII, или измерение активности GnTIII. В качестве альтернативы возможно применение лектина, который связывается с биосинтетическими производными GnTIII, например, лецитин Е4-РНА. В качестве альтернативы можно применить функциональное исследование для измерения повышенного связывания Fc-рецептора или повышенной эффекторной функции, опосредуемой антителами, продуцируемыми клетками, модифицированными нуклеиновой кислотой, кодирующей полипептид с активность GnTIII. В качестве другой альтернативы, нуклеиновая кислота может быть функционально связана с репортерным геном (геном-репортером); уровни экспрессии слитого полипептида определяют, измеряя сигнал, связанный с уровнем экспрессии репортерного гена. Репортерный ген может транскрибироваться совместно с нуклеиновой кислотой (кислотами), кодирующей белок слияния с образованием единой молекулы мРНК; их соответствующие кодирующие последовательности могут быть соединены либо через последовательность, кодируюущую внутренний сайт посадки рибосом (IRES), либо энхансер кэп-независимой трансляции (CITE). Репортерный ген может транслироваться совместно с по меньшей мере одной молекулой нуклеиновой кислоты, кодирующей полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III(GnTIII) и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, с образованием единой полипептидной цепи. Нуклеиновая кислота, кодирующая полипептид слияния, может быть функционально связана с репортерным геном и поставлена под контроль единого(общего) промотора, и таким образом, чтобы нуклеиновая кислота, кодирующая полипептид слияния, и репортерный ген транскрибировались в молекулу РНК, которая путем альтернативного сплайсинга превращается в две отдельные молекулы матричной РНК (мРНК); одна из полученных в результате молекул мРНК транслируется затем в репортерный белок, а другая транслируется в указанный полипептид слияния.
Если экспрессируются несколько разных нуклеиновых кислот, кодирующих полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащий отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, они могут быть организованы таким образом, чтобы в результате их транскрипции получалась одна или несколько молекул мРНК. Если они транскрибируются с образованием единая молекула мРНК, соответствующие кодирующие последовательности могут быть связаны либо внутренним сайтом посадки рибосом (IRES), либо энхансером кэп-независимой трансляции (СГГЕ). Их транскрипция может инициироваться общим промотором и приводить к образованию молекулы РНК, которая путем альтернативного сплайсинга превращается в несколько отдельных молекул матричной РНК (мРНК), каждая из которых затем транслируется в соответствующий закодированный белок слияния.
i. Системы экспрессии
Для конструирования векторов экспрессии, содержащих кодирующие последовательности целевых белков, и кодирующие последовательности полипептидов, обладающие активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащие отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, вместе с сигналами, контролирующими транскрипцию и трансляцию, можно применять хорошо известные специалистам в данной области методы. Эти методы включают in vitro технологии рекомбинантных ДНК, синтетические in vitro технологии и рекомбинацию/генетическую рекомбинацию in vivo. См., например, технологии, описанные в Maniatis et al, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory, Нью-Йорк (1989) и Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates & Wiley Interscience, Нью-Йор (1989). Для экспрессии кодирующих последовательностей белков слияния согласно настоящему изобретению могут быть использованы разнообразные системы «хозяин-вектор экспрессии». Предпочтительно использовать в качестве систем «клетка-хозяин» клетки млекопитающих, трансфецированные рекомбинантными векторами экспрессии в виде плазмидной ДНК или космидной ДНК, содержащими кодирующую последовательность целевого белка и кодирующую последовательность полипептида слияния. Наиболее предпочтительно использование в качестве систем «клетка-хозяин» дрожжевых клеток, клеток насекомых или клеток растений. Некоторые примеры экспрессионных систем и способов отбора описаны в следующих источниках и источниках, не которые в них содержатся ссылки: Borth et al., Biotechnol. Bioen. 71 (4): 266-73 (2000-2001), in Werner et al., Arzneimitteforschung/Drug Res. 48(8):870-80(1998), Andersenand, Krummen, Curr. Op. Biotechnol. 13: 117-123 (2002), Chadd, Chamow, Curr. Op. Biotechnol. 12: 188-194 (2001), и Giddings, Curr. Op. Biotechnol. 12: 450-454 (2001). В альтернативных воплощениях могут быть рассмотрены другие системы «клетка-хозяин», включая дрожжевые клетки трансформированные рекомбинантными дрожжевыми векторами экспрессии, содержащими кодирующую последовательность целевого белка и кодирующую последовательность полипептида слияния, клетки насекомых, инфицированные рекомбинантными вирусными векторами экспрессии (например, бакуловирусными), содержащими кодирующую последовательность целевого белка и кодирующую последовательность полипептида слияния, системы клеток растений, инфицированные рекомбинантными вирусными векторами экспрессии (например, вирус мозаики цветной капусты (ВМЦК, CaMV), вирус табачной мозаики (ВТМ, TMV)) или трансформированные плазмидными векторами экспрессии (например, Ti-плазмидой), содержащими кодирующую последовательность целевого белка и кодирующую последовательность полипептида слияния данного изобретения, или системы клеток животного, инфицированных рекомбинантными вирусными векторами экспрессии (например, аденовирус, вирус vaccinia), включая клеточные линии, модифицированные таким образом, чтобы содержать множественные копии ДНК, кодирующей целевой белок, и кодирующей последовательности полипептида слияния согласно данному изобретению (либо стабильно амплифицированные (CHO/dhfr), либо нестабильно амплифицированные в "double-minute" хромосомах (например, в мышиных клетках).
Для способов согласно настоящему изобретению стабильная экспрессия, в целом, предпочтительнее, чем временная экспрессия, поскольку она обычно дает более воспроизводимые результаты, а также более пригодна для крупномасштабного получения, т.е. получения гликомодифицированных антител, согласно настоящему изобретению, в клеточных линиях для крупномаштабного производства. Вместо содержащих вирусные точки начала репликации векторов экспрессии, клетки-хозяева могут быть трансформированы соответствующими кодирующими нуклеиновыми кислотами, контролируемыми соответствующими элементами контроля экспрессии (например, последовательностями промотора, энхансера, терминаторов транскрипции, сайтов полиаденилирования и т.д.), и селективным маркером. После введения чужеродной ДНК, модифицированные клетки можно оставить на 1-2 дня расти в обогащенной среде, а затем перевести на селективную среду. Селективный маркер в рекомбинантной плазмиде обеспечивает устойчивость к селекции и позволяет отбирать клетки, которые стабильно интегрировали такую плазмиду в свои хромосомы и растут с образованием колоний, которые, в свою очередь, можно клонировать и развить в клеточную линию.
Может быть использован ряд систем селекции (отбора), включая, но не ограничиваясь, гены тимидин киназы вируса простого герпеса (Wigler et al., Cell 11: 223 (1977)), гипоксантин-гуанин фосфорибозилтрансферазы (Szybalska & Szybalski, Proc. Natl. Acad. Sci. USA 48: 2026 (1962)) и аденин фосфорибозилтрансферазы (Lowy et al., Cell 22: 817 (1980)), которые могут применяться в клетках tk-, hgrpt- или aprt-, соответственно. Устойчивость к антиметаболитам также может быть положена в основу отбора по гену dhfr, который обеспечивает клетке устойчивость к метотрексату (Wigler et al., Natl. Acad. Sci. USA 77: 3567 (1989); O'Hare et al., Proc. Natl. Acad. Sci. USA78: 1527 (1981)), по гену gpt, который обеспечивает клетке устойчивость к микофеноловой кислоте (Mulligan & Berg, Proc. Natl. Acad. Sci. USA 78: 2072 (1981)), по гену neo, который обеспечивает клетке устойчивость к аминогликозиду G-418 (Colberre-Garapin et al, J. Mol. Biol. 150: 1 (1981)), и по гену hygro, который обеспечивает клетке устойчивость к гигромицину (Santerre et al., Gene30: 147 (1984). В последнее время были описаны дополнительные селективные гены, а именно: trpB, который позволяет клетке использовать индол вместо триптофана, hisD, который позволяет клетке использовать гистинол вместо гистидина (Hartman & Mulligan, Proc. Natl. Acad. Sci. USA 85: 8047 (1988)), глутамин-синтазная система и ODC (орнитин декарбоксилаза, ornithine decarboxylase) которая обеспечивает клетке устойчивость к ингибитору орнитин декарбоксилазы, 2-дифлуорометил- орнитину - DFMO (McConlogue, в: Current Comniunications in Molecular Biology, Cold Spring Harbor Laboratory ed. (1987)).
ii. Идентификация трансфектантов или трансформантов, которые экспрессируют белок с модифицированным рисунком гликозилирования.
Клетки-хозяева, которые содержат кодирующую последовательность и которые экспрессируют биологически активный продукт гена, могут быть идентифицированы при помощи по меньшей мере четырех следующих общих подходов: (а)ДНК-ДНК или ДНК-РНК гибридизация, (b) наличие или отсутствие функций «маркерного» гена, (с) оценка уровня транскрипции по экспрессии соответствующих транскриптов мРНК в клетке-хозяине, (d) обнаружение продукта гена по результатам иммуноанализа или по биологической активности этого продукта.
В первом подходе присутствие кодирующей последовательности целевого белка и кодирующей последовательности полипептида слияния данного изобретения, введенных в вектор экспрессии может быть обнаружено путем ДНК-ДНК или ДНК-РНК гибридизации с зондами, содержащими последовательности нуклеотидов, гомологичные соответствующим кодирующим последовательностям, либо их части, или их производные.
Во втором подходе рекомбинантная система экспрессии вектор/хозяин может быть идентифицирована и отобрана на основании наличия или отсутствия определенных функций «маркерного» гена (например, активности тимидин киназы, устойчивости к антибиотикам, устойчивости к метотрексату, фенотипу трансформации, образованию телец-включений в случае бакуловируса и т.д.). Например, если кодирующая последовательность целевого белка и кодирующая последовательность полипептида слияния согласно данному изобретению введены внутрь последовательности маркерного гена вектора, рекомбинанты, содержащие соответствующую кодирующую последовательность, могут быть идентифицированы по отсутствию функции маркерного гена. В качестве альтернативы, маркерный ген может быть помещен тандемно с кодирующими последовательностями под контроль общего или разных промоторов, используемых для контроля экспрессии кодирующих последовательностей. Экспрессия маркера в ответ на индукцию или отбор указывает на экспрессию кодирующей целевой белок последовательности и кодирующей полипептид слияния последовательности. В третьем подходе активность транскрипции кодирующей области целевого белка области и кодирующей последовательности полипептида слияния согласно данному изобретению можно оценить при помощи гибридизационных анализов. Например, можно выделить РНК и анализировать ее методом Нозерн-блот с использованием зонда, гомологичного кодирующей области целевого белка и кодирующей последовательности полипептида слияния согласно данному изобретению, или их определенным частям. В качестве альтернативы, можно выделить и анализировать при помощи таких зондов тотальную ДНК и РНК клетки-хозяина.
В четвертом подходе экспрессию белковых продуктов целевого белка и кодирующей последовательности полипептида слияния, обладающего активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) и содержащего отвечающий за локализацию в комплексе Гольджи домен какого-либо гетерологичного полипептида-резидента комплекса Гольджи, можно оценить иммунологическими методами, например, при помощи метода Вестерн-блот, модификаций иммуноанализа, таких как радиоиммунопреципитация, твердофазный иммуноанализ и т.п.Однако, решающий тест успешности экспрессионной системы включает определение биологически активных продуктов генов.
b. Получение и применение белков и фрагментов белков с измененными рисунками гликозилирования.
i. Получение и применение антител, обладающих повышенной эффекторной функцией, включая антителозависимую клеточную цитотоксичность.
В предпочтительных воплощениях настоящее изобретение предусматривает гликоформы антител и фрагментов антител, обладающих повышенной аффинностью связывания Fc-рецептора и/или эффекторной функцией, включая антителозависимую клеточную цитотоксичность.
Недавние клинические испытания неконъюгированных моноклональных антител (МКАТ, mAb) для лечения некоторых видов рака дали ободряющие результаты. Dillman, Cancer Biother. & Radiopharm. 12: 223-25 (1997), Deo et al, Immunology Today 18: 127 (1997). Химерные неконъюгированные IgG1 были одобрены для низкозлокачественной или фолликулярной В-клеточной лимфомы отличной от лимфомы Ходжкина (Dillman CancerBiother. & Radiopharm. 12: 223-25 (1997)), в то время как другие неконъюгированные МКАТ, гуманизированные IgG1, направленные к твердым опухолям груди, также продемонстрировали многообещающие результаты в фазе III клинических испытаний. Deo et al, Immunology Today 18: 127 (1997). Антигены этих двух МКАТ активно экспрессируются в клетках соответствующих опухолей и антитела опосредуют эффективное разрушение опухоли эффекторными клетками in vitro и in vivo. В отличие от этих, многие другие неконъюгированные МКАТ с высокой специфичностью к опухолям не в состоянии запустить эффекторные функции достаточно сильные, чтобы быть клинически полезными. Frost et al, Cancer 80: 317-33 (1997); Surfus et al., J.Immunother. 19: 184-91 (1996). Для некоторых из этих более слабых МКАТ в настоящее время исследуется дополнительная терапия цитокинами. Добавление цитокинов может стимулировать антителозависимую клеточную цитотоксичность (АЗКЦ) путем повышения активности и увеличения количества циркулирующих лимфоцитов. Frost et al., Cancer 80: 317-33 (1997); Surfus et al., J. Immunother. 19: 184-91 (1996). АЗКЦ, литическая атака антител на клетки-мишени, запускается связыванием рецепторов лейкоцитов к константной области(Рс) антител. Deo et al., Imunology Today 18: 127 (1997). Другой, но дополняющий предыдущий, подход к повышению активности АЗКЦ неконъюгированных IgG1 состоит в модификации Fc-области антител. Исследования модификации белков показало, что рецепторы FcγR взаимодействуют с доменом СН2 области нижней петли IgG. Lund et al., J. Immunol. 157: 4963-69 (1996). Однако, завязывание FcγR требует также присутствия олигосахаридов ковалентно прикрепленных
к консервативному Asn297 в области СН2 (Lund et al., J. Immunol 157: 4963 - 69 (1996); Wright and Morrison, Trends Biotech. 15: 26-31 (1997)), что позволяет предположить, что либо и олигосахарид и полипептид прямо участвуют в формировании сайта взаимодействия, либо что олигосахарид необходим для поддержания активной конформации полипептида СН2. Модификация структуры олигосахаридов может, следовательно, быть исследована как возможное средство для повышения аффинности взаимодействия.
Молекула IgG несет в Fc-области два N-связанных олигосахарида, по одному на каждой тяжелой цепи. Как и любой гликопротеин, антитело продуцируется в виде набора гликоформ, обладающих одинаковым полипептидным каркасом, но разными олигосахаридами, прикрепленными в сайтах гликозилирования. Олигосахариды, обнаруживаемые обычно в Fc-области IgG сыворотки принадлежат к типу сложных «двуантенных» (Wormald et al., Biochemistry 36: 130-38 (1997), с низким уровнем сиаловой кислоты на концах и N-ацетилглюкозамином в месте разветвления (разветвляющий, GlcNAc), а также с вариабельной степенью терминального галактозилирования и фукозилирования коровой части (базового фрагмента). Некоторые исследования предполагают, что минимальная углеводная структура, необходимая для связывания FcγR, ограничивается кором олигосахарида. Lund et al., J. Immunol. 157: 4963-69 (1996)
Клеточные линии, полученные из клеток мыши или хомяка и используемые для получения неконъюгированных МКАТ в промышленности и в научных целях, обычно прикрепляют олигосахаридные детерминанты к Fc-сайтам. Однако иммуноглобулины, экспрессируемые в этих клеточных линиях, лишены «разветвляющего» GlcNAc, найденного в небольших количествах в иммуноглобулинах плазмы. Lifely et al., Glycobiology 318: 813-22 (1995). Как показали недавние наблюдения, продуцируемые миеломой гуманизированные IgG1 (САМРАТН-1H), напротив, несут в некоторых из своих гликоформ разветвляющий GlcNAc. Lifely et al., Glycobiology 318: 813-22 (1995). Полученные в клетках крыс антитела достигали максимальной in vitro активности АЗКЦ близкой к максимальной активности продуцируемых в стандартных клеточных линиях антител САМРАТН-1Н, но при значительно более низких концентрациях антител. Высокий уровень антигена САМРАТН обычно присутствует в клетках лимфомы, и это химерное МКАТ обладает высокой активностью АЗКЦ в отсутствие «разветвляющего» GlcNAc. Lifely et al., Glycobiology 318: 813-22 (1995). При N-гликозилировании разветвляющий GlcNAc добавляет фермент бета-1,4-ацетилглюкозаминилтрансфераза III (GnT III). Schachter, Biochem. Cell Biol. 64: 163-81 (1986).
В более ранних исследованиях использовали единственную продуцирующая антитело клеточную линию СНО, которая была предварительно модифицирована для экспрессии, под внешней регуляцией, разных уровней клонированного гена фермента GnT III (Umana, P., et al., Nature Biotechnol. 17: 176-180 (1999)). Этот подход впервые установил жесткую связь между экспрессией GnT III и активностью АЗКЦ модифицированного антитела.
Другие антитела данного изобретения, обладающие повышенной аффинностью связывания Fc-рецептора и повышенной эффекторной функцией включают без ограничения: моноклональное антитело (chCE7) против нейробластомы человека, полученное способами данного изобретения, химерное антитело (chG250) против клеток карциномы почки человека, полученное способами данного изобретения, гуманизированное анти-HER2 моноклональное антитело (например, Trastuzumab (HERCEPTIN)), полученное способами данного изобретения, химерное моноклональное антитело (ENG-1) против клеток карциномы ободочной кишки, легкого и груди, полученное способами данного изобретения, гуманизированное моноклональные антитело (3622W94) против человеческого антигена 17-1А, полученное способами данного изобретения, гуманизированное антитело (А33) против опухоли ободочной и прямой кишки человека, полученное способами данного изобретения, антитело (R24) против меланомы человека, направленное к ганглиозиду GC3, полученное способами данного изобретения, а также химерное моноклональное антитело (SF-25) против плоскоклеточной карциномы легкого, полученное способами данного изобретения, моноклональное антитело (ВЕС2, InClone Systems, Merck KgaA) против мелкоклеточной карциномы легкого, полученное способами данного изобретения, моноклональное антитело (Bexxar, (tositumomab, Coulter Pharmaceuticals), Oncolym (Techniclone, Alpha Therapeutics)) против лимфомы не Ходжкина человека, полученное способами данного изобретения, моноклональное антитело (С225, ImClone Systems) против плоскоклеточной карциномы головы и шеи человека, приготовленное способами данного изобретения, моноклональное антитело (Panorex (edrecolomab), Centocor, Glaxo Wellcome) против карциномы ободочной и прямой кишки, приготовленное способами данного изобретения, моноклональное антитело (Theragyn, Antisoma) против карциномы яичника человека, полученное способами данного изобретения, моноклональное антитело (SmartM195, Protein Design Labs, Kanebo) против острой миелогенной лейкемической карциномы, полученное способами данного изобретения, моноклональное антитело (Cotara, Techniclone, Cambridge Antibody Technology) против злокачественной глиомы человека, полученное способами данного изобретения, моноклональное антитело (IDEC-Y2B8, IDEC Pharmaceuticals) против В-клеточной лимфомы человека отличной от лимфомы Ходжкина, полученное способами данного изобретения, моноклональное антитело (СЕА-Cide, Inimunomedics) против твердых опухолей человека, полученное способами данного изобретения, моноклональное антитело (Iodme131-MN-14,Immunomedics) против карциномы ободочной и прямой кишки, полученное способами данного изобретения, моноклональное антитело (MDX-210, Medarex, Novartis) против карциномы яичника, почки, груди и предстательной железы, полученное способами данного изобретения, моноклональное антитело (ТТМА, Pharmacie & Upjohn) против карциномы ободочной и прямой кишка, а также поджелудочной железы, полученное способами данного изобретения, моноклональное антитело (MDX-220, Medarex) против экспрессирующей TAG-72 карциномы человека, полученное способами данного изобретения, моноклональное антитело (MDX-447) против экспрессирующей ЭФРr карциномы человека, полученное способами данного изобретения, моноклональное антитело (Genentech) против VEGF (сосудистого эндотелиального фактора роста), полученное способами данного изобретения, моноклональное антитело (BrevaRex, AltaRex) против злокачественной меланомы и карциномы груди, легкого, предстательной железы и поджелудочной железы человека, полученное способами данного изобретения, а также моноклональное антитело (Monoclonal Antibody Conjugate, Immune) против острой миелогенной лейкемии человека, моноклональное антитело. Дополнительно, изобретение предусматривает фрагмент антитела и белки слияния, содержащие область, эквивалентную Fc-области иммуноглобулина.
ii. Получение и применение белков слияния, содержащих область, эквивалентную Fc-области иммуноглобулина, которая активизирует Fc-опосредуемую цитотоксичность.
Как обсуждалось выше, настоящее изобретение относится к способу повышения аффинности связывания Fc-рецептора и/или эффекторной функции терапевтических антител. Этого достигают путем модификации рисунка гликозилирования Fc-области таких антител, в частности, путем модификации продуцирующих антитело клеток для продуцирования полипептида, например, с активностью GnTIII, или с активностью GalT, или с активностью ManII, которые модифицирует олигосахариды, прикрепленные к Fc-области таких антител. Эту стратегию можно применять для повышения Fc-опосредуемой клеточной цитотоксичности против нежелательных клеток, опосредуемой любой молекулой, несущей область, эквивалентную Fc-области иммуноглобулина, а не только терапевтическими антителами, поскольку обусловленные модификацией гликозилирования изменения затрагивают только Fc-область и, следовательно, ее взаимодействия с Fc-рецептором на поверхности эффекторной клетки, вовлеченной в механизм АЗКЦ. Fc-содержащие молекулы, для которых применимы раскрытые здесь методы, включаютбез огарничения: (а) - растворимые белки слияния, полученные из нацеливающего домена белка, слитого с N-концом Fc-области (Chamov & Ashkenazi, Trends Biotech. 14: 52 (1996)), и (b) мембраносвязанные белки слияния, полученные из отвечает за локализацию в плазматической мембране трансмембранного домена II типа, соединенного с N-концом Fc-области Stabila, P F., Nature Biotech. 16: 1357 (1998)).
Для растворимых белков слияния (а) нацеливающий домен направляет связывание белка слияния с нежелательными клетками, такими как раковые клетки, т.е. аналогично связыванию терапевтических антител. Применение раскрываемых настоящим способов усиления эффекторной функции, включая активность Fc-опосредованную клеточную цитотоксичность, опосредуемой этими молекулами было бы, следовательно, идентичным способу, применяемому для терапевтических антител.
В случае мембраносвязанных белков слияния (b) нежелательные клетки в организме должны экспрессировать ген, кодирующий полипептид слияния. Этого можно достигнуть либо при помощи подходов генной терапии, т.е. путем трансфекции клеток in vivo плазмидным или вирусным вектором, который предписывает экспрессию гена, кодирующего белок слияния, в нежелательных клеткам, либо путем имплантации в организм клеток,, генетически модифицированных для экспрессии белка слияния на свою поверхность. Эти последние клетки могут быть имплантированы в организм внутри полимерной капсулы (терапия инкпсулированными клетками), в которой они не могут быть разрушены по механизму Fc-опосредованной клеточной цитотоксичности. Однако, если, в случае разрушения капсулы, присутствие освобожденных клеток оказывается нежелательным, они могут быть элиминированы по механизму Fc-опосредованной клеточной цитотоксичности. Stabila et al., NatureBiotech. 16: 1357 (1998). В этом случае раскрываемые настоящим способы возможно применять либо путем внедрения в вектор для генной терапии дополнительной экспрессионной кассеты, предписывающей адекватные или максимальные уровни экспрессии полипептида слияния согласно донному изобретению, либо путем модификации клеток для имплантации с целью экспрессирования в них адекватных или максимальных уровней полипептида слияния согласно данному изобретению.
Терапевтические приложения антител, фрагментов антител и полипептидов слияния, полученных согласно способам данного изобретения.
Антитела (т.е. антитела, фрагменты антител и белки слияния, содержащие область, эквивалентную Fc-области иммуноглобулина) согласно настоящему изобретению могут применяться самостоятельно для выявления и лизиса опухолевых клеток in vivo. Возможно также применение для лечения карциномы человека антител, конъюгированных с подходящими терапевтическими агентами. Например, возможно применение антител в комбинации со стандартными или обычными способами лечения, такими как химиотерапия, радиационная терапия, либо конъюгирование антател с терапевтическим лекарственным средством или токсином, а равно и с лимфкином или опухоль-ингибирующим фактором роста для доставки терапевтического агента к месту локализации карциномы.
Технологии конъюгирования таких трапевтических агентов с антителами хорошо известны (См., например., Amon et al., “Monoclonal Antibodies for Immunotargeting of Drugs in Cancer Therapy", в Monoclonal Antibodies and Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., "Antibodies For Drug Delivery", в Controlled Drug Delivery (2e изд.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", в Monoclonal Antibodies'84: Biological And Clinical Applications, Pinchera et al. (eds.), pp. 475-506 (1985); and Thorpe et al, "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol Rev., 62: 119-58 (1982)].
В качестве альтернативы, гликомодифицированные антитела могут быть связаны с веществами высокой энергии излучения, например, <131>I, который при локализации в опухоли, приводит к лизису клеток на расстоянии нескольких клеточных диаметров [См., например, Order, "Analysis, Results, and Future Prospective of The Therapeutic Use of Radiolabeled Antibody" Cancer Therapy", в Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985)]. Согласно другому воплощению, антитела данного изобретения могут быть конъюгированы с вторым антителом с целью получения гетероконъюгата антител для лечения опухолевых клеток, как описано в патенте США Segal, патент США №4,676,980.
Другие терапевтические приложения антител согласно данному изобретению включают конъюгирование или связывание, например, при помощи технологий рекомбинантной ДНК, с ферментами, способными превращать пролекарство в цитотоксическое лекарственное средство, и применение такого конъюгата антитело-фермент в комбинации с пролекарством для превращения пролекарства в итотоксическое лекарственное средство в сайте локализации опухоли. [См., например, Senter et al., "Anti-Tumor Effects of Antibody-alkaline
Phosphatase", Proc. Natl. Acad. Sci. USA 85: 4842-46 (1988); "Enhancement of the in vitro and in vivo Antitumor Activites of Phosphorylated Mitocycin С and Etoposide Derivatives by Monoclonal. Antibody-Alkaline Phosphatase Conjugates", Cancer Research 49: 5789-5792 (1989); and Senter, "Activation of Prodrugs by Antibody-Enzyme Conjugates: A New Approach to Cancer Therapy," FASEB J.4: 188-193 (1990)].
Еще одно применение терапевтических антител согласно данному изобретению включает применение, либо в присутствии комплемента, либо в составе конъюгата антитело-лекарственное средство или антитело-токсин, для удаления опухолевых клеток из костного мозга больных раком пациентов. Согласно этому подходу возможно очистить костный мозг ex vivo путем обработки антителом, а затем вернуть в организм пациента инфузионным путем. См., например, Ramsay et al., "Bone Marrow Purging Using Monoclonal Antibodies", J. Clin. Immunol., 8 (2): 81-88 (1988)
Далее, в терапии могут применяться химерные антитела, рекомбинантные иммунотоксины и другие рекомбинантные конструкции данного изобретения, обладающие специфичностью антиген - связывающей области желаемого моноклонального антитела. Например, одноцепочечные иммунотоксины данного изобретения могут применяться для лечения карциномы человека in vivo. Аналогично, белок слияния, содержащий по меньшей мере одну антиген-связывающую область антитела согласно данному изобретению присоединенную к по меньшей мере одной функционально активной части второго белка, обладающего противоопухолевой активностью, например, лимфокина или онкостатина, можно применять для лечения карцином человека in vivo. Далее, можно применять рекомбинантные технологии известные в данной области, для конструирования биспецифических антител, где одна из специфичностей связывания антитела направлена к опухоль-ассоциированному антигену, а вторая связывающая специфичность антитела совпадает со специфичностью молекулы, отличной от опухоль-ассоциированого антигена.
Настоящее изобретение предусматривает способ селективного лизиса опухолевых клеток, экспрессирующих антиген, который специфически связывается с моноклональным антителом согласно данному изобретению или его функциональным эквивалентом. Этот способ включает реакцию гликомодифицированного антитела согласно настоящему изобретению, или иммуноконъюгата (например, иммунотоксина), содержащего гликомодифицированное антитело настоящего изобретения, с указанными опухолевыми клетками. Эти опухолевые клетки могут представлять собой клетки карциномы. Дополнительно, это изобретение предусматривает способ лечения карцином (например, карцином человека) in vivo. Это способ включает назначение пациенту фармацевтически эффективного количества состава, содержащего по меньшей мере одно из гликомодифицированных антител согласно данному изобретению или иммуноконъюгаты (например, иммунотоксин), содержащие по меньшей мере одно из гликомодифицированных антител д согласно данному изобретению.
В дальнейшем аспекте изобретение предусматривает улучшенный способ лечения аутоиммунных заболеваний, полностью или частично вызываемыми патогенными антителами, основанный на уменьшении количества В-клеток и включающий назначение терапевтически эффективного количества иммунологически активного антитела человеку, нуждающемуся в лечении. Улучшение заключается в назначении терапевтически эффективного количества антитела, обладающего повышенной АЗКЦ и приготовленного согласно способам данного изобретения. В предпочтительном воплощении антитело представляет собой анти-CD20 антитело. Примеры аутоиммунных заболеваний включают без ограничения иммуно-опосредованные тромбоцитопении (такие как острые идиопатические тромбоцитопенические пурпуры и хронические идиопатические тромбоцитопенические пурпуры, дерматомиозит, хорею Сиденхама, волчаночный нефрит, ревматическую лихорадку, полигландулярные синдромы, пурпуру Шонлейна-Геноха, постстрептококковый нефрит, нодозную (узелковую) эритему, артериит Такаясу, болезнь Аддисона, мультиформная (экссудативная) эритема, нодозный полиартериит, анкилозирующий спондилит, синдром Гудпасчера (Goodpasture's), облитерирующий тромбоангиит, первичный желчный цирроз, тироидит Хашимото, тиротоксикоз, хронический активный гепатит, полимиозит (дермтаомиозит), полихондрит, обыкновенный пемфигус, грануломатоз Вегенера, мембранозная нефропатия, амиотрофический латеральный склероз, сухотку спинного мозга, полимиаглию, злокачественное малокровие, бстропрогрессирующий гломерулонефрит и фиброзирующий альвеолит, воспалительные реакции, такие как воспалительные кожные заболевания, включая псориаз и дерматит, например, атопический дерматит), системную склеродерму и склероз, реакции, ассоциированные с воспалительными кишечными заболеваниями (такими как болезнь Крона и язвенный колит), респираторный дистресс-синдром (включая респираторный дистресс-синдром взрослых-РДСВ), дерматит, менингит, энцефалит, увеит, колит, гломерулонефрит, аллергические состояния (такие как экзема, астма и другие состояния, сопровождающиеся инфильтрацией Т-лимфоцитами и хроническими воспалительными реакциями), атеросклероз, дефицит адгезии лимфоцитов, ревматоидный артрит, системную красную волчанку (СКВ), сахарный диабет (например, сахарный диабет I типа или инсулинозависимый сахарный диабет), множественный склероз, синдром Рейно, аутоиммунный тироидит, аллергический энцефаломиелит, синдром Съёргена, ювенильный диабет, а также иммунные реакции, ассоциированные с острой и замедленной гиперчувствительностью, опосредуемой цитокинами и Т-лимфоцитами, и обнаруживаемые обычно при туберкулезе, сакроидозе, полимиозите, грануломатозе и васкулите, злокачественное малокровие (болезнь Аддисона), заболевания, сопровождающиеся диапедезом лимфоцитов, воспалительные заболевания центральной нервной системы (ЦНС), синдром множественного поражения органов, гемолитическую анемию (включая без ограничения криоглобинемию или Кумбс-положительную анемию), миастению gravis, заболевания, опосредуемые комплексом антиген-антитело, иммунное повреждение клубочков, антивфосфолипидный синдром, аллергический неврит, болезнь Грейвза, миастенический синдром Ламберта-Итона, пузырчатку, аутоиммунные полиэндокринопатии, болезнь Райтера, синдром обездвиженности, болезнь Бехчета, гигантоклеточный артериит, иммунокомплексный нефрит, IgA-нефропатию, IgM-полинейропати, иммунную тромбоцитопеническую пурпуру (ИТП) или аутоиммунную тромбоцитопению и т.д. В этом аспекте изобретения антитела согласно изобретению применяют для того, чтобы очистить кровь от нормальных В-лимфоцитов на длительный период
В соответствии с осуществлением данного изобретения, пациентом может быть человек, лошадь, свинья, бык (корова), мышь, собака, кошка или птица. Другие теплокровные животные также включены в область охвата изобретение.
Настоящее изобретение предусматривает также способ лечения пациентов, страдающих от рака. Пациент может быть человеком, собакой, кошкой, мышью, крысой, кроликом, лошадью, козой, овцой, коровой, цыпленком. Рак может быть идентифицирован как рак груди, рак желчного пузыря, ретинобластома, папиллярная цистаденокарцинома яичника, опухоль Вилма или мелкоклеточный рак легкого, и в общих чертах характеризуется как группа клеток, имеющих на поверхности опухоль-ассоциированный антиген. Этот метод включает назначение пациенту достаточного для лизиса раковых клеток количества направленного к опухоли антитела, присоединенного к цитотоксическому агенту. Обычно соединение противоопухолевого антитела с цитотоксическим агентом проводят в условиях, которые позволяют соединенному таким образом антителу связывать свою мишень на клеточной поверхности. Направленное к опухоли антитело посредством связывания со своей мишенью прямо или опосредованно вызывает или содействует лизису клеток, способствуя, таким образом, излечению пациента.
Также предусмотрен способ ингибирования пролиферации клеток опухолей млекопитающих, который включает контакт клеток опухолей млекопитающих с гликомодифицированными антителами настоящего изобретения в достаточной концентрации или с иммуноконъюгатом, содержащим те же антитела, для подавления пролиферации клеток опухолей млекопитающих.
Далее настоящее изобретение предусматривает способы ингибирования роста клеток опухолей млекопитающих, лечения опухоли у пациента, пролиферативных заболевания у пациента. Эти способы включают назначение пациенту эффективного количества состава по данному изобретению.
Таким образом, очевидно, что настоящее изобретение включает в себя фармацевтические составы, комбинации и способы лечения карцином у людей. Например, изобретение включает фармацевтические составы для применения в лечении карцином у людей, содержащие фармацевтически эффективное количество антитела согласно настоящему изобретению и фармацевтически приемлемый носитель.
Составы могут содержать антитело или фрагменты антитела, либо неизмененные, конъюгированные с терапевтическим агентом (например, лекарственным веществом, токсином, ферментом или вторым антителом), либо в рекомбинантной форме (например, химерные антитела, фрагменты химерных антител, биспецифические антитела). Составы могут дополнительно содержать другие антитела или конъюгаты для лечения карцином (например, «коктейль» антител).
Составы данного изобретения, содержащие антитело, конъюгат антитела и иммунотоксин, могут быть назначены для введения обычными способами, включая, но не ограничиваясь, введением внутривенно, внутибрюшинно, орально, внутрь лимфатических узлов или непосредственно в опухоль. Предочтительным способом введения является внутривенный.
Возможны различные лекарственные формы составов данного изобретения, включая, но не ограничиваясь, жидкие растворы или суспензии, таблетки, пилюли, порошки, суппозитории, полимерные микрокапсулы или микровезикулы, липосомы, растворы для инфузионного и внутривенного введения.
[0193] Составы согласно данному изобретению также предпочтительно содержат обычные фармакологически приемлемые носители и адъюванты, известные в данной области, такие как альбумин сыворотки крови человека (ЧСА), ионообменные в-ва, окись алюминия, лецитин, буферные вещества, такие как фосфаты, глицин, сорбиновая кислота, сорбат калия, а также соли и электролиты, такие как протамина сульфат.
Наиболее эффективный способ введения и режим дозировки состава по данному изобретению зависят от тяжести и течения заболевания, состояния здоровья пациента и реакции на лечение, а также от соображений лечащего врача. Соответственно, дозировки состава следует титровать для каждого отдельного пациента. Тем не менее, эффективная доза состава по данному изобретению может варьировать в пределах от приблизительно 1 до приблизительно 2000 мг/кг.
Описанные здесь молекулы могут входить в разнообразные лекарственные формы, которые включают без ограничения жидкие растворы или суспензии, таблетки, пилюли, порошки, суппозитории, полимерные микрокапсулы или микровезикулы, липосомы, а также растворы для инфузионного и внутривенного введения. Предпочтительная форма зависит от способа введения и терапевтического приложения.
Наиболее эффективный способ введения и режим дозировки молекул согласно настоящему изобретению зависят от локализации подвергаемой лечению опухоли, тяжести и течения рака, состояния здоровья пациента и реакции на лечения, а также от соображений лечащего врача. Соответственно, дозировки состава следует титровать для каждого отдельного пациента.
Взаимное отношение дозировок для животных различных размеров и видов, а также для людей в мг/кг площади поверхности в Freireich, Е. J., et al. Cancer Chemother., Rep.50 (4): 219-244 (1966). Возможна корректировка режима дозировок с целью оптимизации ингибирования роста и лизиса клеток опухоли, например, возможно разделение дозы на порции для ежедневного введения или пропорциональное уменьшение дозы в зависимости от ситуации (например, несколько разделенных доз можно вводить ежедневно или соразмерно уменьшить в зависимости от определенной терапевтический ситуации).
Должно быть очевидно, что для достижения излечения дозу состава согласно данному изобретению, необходимую для излечения, можно уменьшать и далее, одновременно с оптимизацией графика введения.
В соответствии с осуществлением изобретения формацевтический носитель может представлять собой липидный носитель. Липидный носитель может представлять собой фосфолипид. Липидный носитель может также представлять собой детергент. Термин «детергент» означает здесь любое вещество, которое изменяет (обычно - снижает) поверхностное натяжение жидкости.
В одном из примеров данного изобретения детергент может представлять собой неионный детергент.Примеры неионных детергентов включаютбез огарничения: Polisorbate 80 (полисорбат 80, также известен как Tween 80 или полиоксиэтиленсорбитан), Brij и Triton (например, Triton WR-1339 и Triton А-20).
В качестве альтернативы, детергент может представлять собой ионный детергент. Пример ионного детергента включает, но не ограничивается, алкилтриметиламмония бромид.
Дополнительно, в соответствии с настоящим изобретением липидный носитель может представлять собой липосому. В данном приложении «липосома» - это любая ограниченная мембраной везикула, которая содержит любую молекулу по данному изобретению или их комбинацию.
Нижеследующие примеры дают более детальное объяснение изобретения. Следующие способы приготовления и примеры даны, чтобы дать специалистам в данной области возможность более ясно понять и осуществить настоящее изобретение. Настоящее изобретение, однако, не ограничивается рамками приведенных в примерах способов реализации, которые имеют целью проиллюстрировать только один аспект изобретения, функционально эквивалентные способы попадают в область охвата данного изобретения. Действительно, благодаря нижеследующему описанию и сопутствующим рисункам для специалистов в данной области станут очевидными разнообразные модификации изобретения, отличные от описанных. Такие модификации находятся в рамках приложенной формулы изобретения.
Примеры
ПРИМЕР 1
Материалы и методы
1. Конструирование векторов экспрессии антитела.
Вектор экспрессии анти-CD20 антитела pETR1502
Вектор экспрессии антиi-CD20 антитела С2В8, pETR1502, состоит из четырех отдельных независимых экспрессионных кассет (одна для легкой цепи антитела С2В8, одна - для тяжелой цепи антитела С2В8, одна - для гена устойчивости к неомицину и одна - для мышиного гена dhfr). Все гены находятся под контролем промотора вируса миелопролиферативной саркомы (MPSV) и содержат синтетический консенсусный сигнал полиаденилирования, полученный из сигнала полиаденилирования гена бета-глобина кролика.
КДНК, кодирующие вариабельные участки тяжелой цепи (VH) и легкой цепи(VL) анти-CD20 антитела С2В8 были собраны в ходе одностадийного процесса ПНР ((Kobayashi, N., et al., Biotechniques 23: 500-503 (1997))) из набора перекрывающихся однонитевых олигонуклеотидов. Исходную последовательность, кодирующую области VH и VL антитела С2В8 взяли из Международной публикации заявки на патенту (International Publication Number: WO 94/11026). Собранные фрагменты кДНК, кодирующие VH и VL субклонировали в вектор pBluescriptIIKS(+), в результате чего были получены плазмиды pBlue-C2B8VH и pBlue-C2B8VH, и затем секвенировали.
Вариабельные цепи антитела С2В8 амплифицировали из соответствующих плазмид pBlue-C2B8VH и pBlue-C2B8VH, используя праймеры, которые вводят сайт рестрикции AscI на 5'-конце и соответствующие сайты рестрикции в месте соединения вариабельной и константной областей (BsiWI для легкой цепи и NheI-для тяжелой цепи). Амплифицировали константные области IgG1 из библиотеки кДНК лейкоцитов человека (Quickclone, Clontech), используя праймеры, которые вводят подходящие сайты рестрикции на 5'- и 3'-концах (BsiWI и BamHI для константной легкой цепи, NheI и BamHI - для константной тяжелой цепи).
После подтверждения правильности последовательностей ДНК и тяжелые и легкие цепи антитела С2В8 комбинировали с промотором MPSV и сигналами полиаденилирования. На первом этапе были сконструированы два разных вектора экспрессии: один (pETR1315) для легкой цепи С2В8, а другой (pETR13156) - для тяжелой цепи. На втором этапе в вектор экспрессии pETR1315 ввели экспрессионную кассету, содержащую ген устойчивости к неомицину (ген устойчивости к неомицину, полученный из транспозона Tn5, находящийся под контролем минимального промотора MPSV), в результате чего получили плазмиду pETR1481. Экспрессионную кассету, содержащую ген dhfr под контролем промотора MPSV, встраивали (инсертировали) в вектор pETR1316, что давало плазмиду pETR1328. На завершающем этапе, оба экспрессионных модуля (легкая цепь С2В8 + ген устойчивости к неомицину и тяжелая цепь С2В8 + ген dhfr) объединяли в один вектор, получая в результате плазмиду pETR1502.
Вектор экспрессии анти-CD20 антитела pETR1520
Вектор pETR1520 представляет собой комбинацию вектора экспрессии анти-CD20 антитела С2В8 с ориджином (точкой инициации) репликации вируса Эпштейна-Барр (oriP) для репликации и поддержания эписомного вектора в клетках, продуцирующих ядерный антиген вируса Эпштейна-Барр (EBNA). Для конструирования вектора экспрессии антитела С2В8, pETR1520, модуль экспрессии антитела С2В8 из вектора pETR1416 поместили в виде HinDIII-фрагмента (т.е. фрагмента, полученного при помощи обработки рестриктазой HinDIII) в вектор pETR1507, содержащий oriP. Вектор pETR1416 подобен вектору pETR1502 за тем исключением, что в этой плазмиде ген устойчивости в неомицину находится под контролем полного, а не минимального промотора MPSV.
Вектор экспрессии антифибринонектинового антитела pETR1546
Это вектор идентичен вектору pETR1520 за тем исключением, что области, кодирующие тяжелую и легкую вариабельные цепи анти-CD20 антитела C2D8, были заменены на соответствующие кодирующие области антитела L19 человека, распознающего домен ED-B фибронектина. ДНК, кодирующие вариабельные области были синтезированы методом ПЦР с перекрывающимся удлинением при использовании синтетических олигонуклеотидов, основу которых составляет последовательность вариабельных областей антитела L19 (Pini, A. Et al. J. Biol. Chem. 273 (34): 21769-76 (1998)). Вектор экспрессии анти-РЭФР антитела (С225*) pURSI28
Это вектор идентичен вектору pETR1520 за тем исключением, что области, кодирующие тяжелую и легкую вариабельные цепи анти-CD20 антитела С2В8 были заменены на соответствующие кодирующие области антитела С225 - химерного антитела, распознающего рецептор к эпидермальному фактору роста человека. Фрагменты ДНК, кодирующие вариабельные области антитела С225 были синтезированы методом ПЦР с перекрывающимся удлинением при использовании синтетических олигонуклеотидов, основу которых составляет последовательность вариабельных областей антитела С225 (последовательность можно найти в опубликованной заявке на патент, международная публикация номер WO 96/40210, на Фиг. 16 и Фиг. 17 которой показаны последовательности тяжелой и легкой цепей соответственно).
2. Конструирование векторов экспрессии GnTIII-полипептида слияния pETR1166. Вектор для конститутивной экспрессии GnTIII.
Для конструирования вектора экспрессии GnTIII, pETR1166, посредством ПЦР амплифицировали GnTIII крысы, полученный из библиотеки кДНК почки крысы (Clontech). Для удобства последующего определения GnTIII методом Вестерн-блот сразу за стоп-кодоном гена GnTIII в направлении «против транскрипции»(ирзггеат) был добавлен С-концевой участок c-myc-эпитопа (последовательность аминокислот: PEQKLISEEDL). После подтверждения правильности последовательности GnTIII, ген помешали под контроль промотора MPSV и добавляли синтетический сигнал полиаденилирования бета-глобина кролика. Окончательный вектор экспрессии GnTIII содержал также отдельную селективную кассету с геном устойчивости к пуромицину также под контролем промотора MPSV и с синтетическим сигналом полиаденилирования бета-глобина кролика.
pETR1425: Замена первых 76 аминокислот GnTIII на первые 102 аминокислоты GnTI человека.
Конструирование гена этой гибридной гликозилтрансферазы производили путем последовательных перекрывающихся реакций ПЦР. В одной реакции «стержневую» область (stem region) GnTI амплифицировали, используя праймеры GAB-179 и GAB-180. В ходе этой реакции ПЦР на 5'-конце были введены консенсусная Kozak-последовательность и сайт рестрикции Ascl. Полученный в результате ПЦР фрагмент содержит участок перекрывания с геном Gnlll длиной 23 н.п., начинающийся в положении 229. В ходе второй реакции ПЦР с применением праймеров GAB-177 и GAP-178 был амплифицирован область Gnlll между положениями 229 и 380 (включая 229 и 380). В результате был получен фрагмент, содержащий уникальный сайт BstXI на 3'-конце и участок перекрывания со стержневой областью GnTI длиной 22 н.п.на 5'-конце. Оба фрагмента, полученные в результате ПЦР, очищали и использованы в качестве матриц в третьей ПЦР-реакции (праймеры: GAB-179 и GAB-178). Полученные в результате фрагменты очищали, расщепляли рестриктазой AcsI и лигировали с вектором pETR1001 (разрезенный AscI и SmaI), в результате чего получали плазмиду pETR1404. После того, как правильность последовательности вставки, была подтверждена, к вектору pETR1404 был добавлен промоторный элемент MPSV в виде AscI (частичное расщепление) / PmeI - фрагмента. В результате получили плазмиду pETR1423. Затем SphI/BstXI-фрагмент вектора pETR1166, вектора экспрессии, несущего исходный ген GnTIII крысы, заменяли на соответствующий фрагмент вектора pETR1425, содержащего ген полипептида слияния GnTI-GnTIII под контролем промотора MPSV и кассету, содержащую ген устойчивости к пуромицину (для селекции)
Последовательности праймеров:
GAB-177: GCGTGTGCCTGTGACCCCCGCGCCCCTGCTCCAGCCACTGTCCCC
GAB-178: GAAGGTTTCTCCAGCATCCTGGTACC
GAB-179: CTGAGGCGCGCCGCCACCATGCTGAAGAAGCAGTCTGCAGGGC
GAB-180: GGGGACAGTGGCTGGAGCAGGGGCGCGGGGGTCACAGGCACACGCGGC
pETR1506: Замена первых 76 N-концевых аминокислот GnTIII на первые 102 аминокислоты маннозидазы II человека.
Конструирование вектора pERT1506 проводили аналогично конструированию pETR1425. Стержневой участок маннозидазы II человека амплифицировали с праймерами GAB-252 и GAB-253, используя вектор pBlue-man в качестве матрицы. В ходе ПЦР на 5'-конце были введены сайт Fse и консенсусная Kozak-последовательность. Полученный в результате ПЦР фрагмент перекрывает ген GnTIII на участке длиной 23 п.н., начинающемся в положении 229. В ходе второй реакции ПЦР амплифицировали часть гена GnTIII (положения 229-460) с праймерами GAB-254 и GAB-255. Эта реакция ПЦР породила фрагмент, содержащий участок перекрытия с геном маннозидазы II длиной 43 н.п. на 5'-конце и уникальный сайт StuI на 5'-конце. Оба фрагмента очищали и использовали в качестве матриц в третьей ПЦР с праймарами GAB-252 и GAB-255. Полученный в результате фрагмент ввели в вектор pIC19H, что дало вектор pETR1484. После того, как правильность последовательности вставки была подтверждена, конструировали полный слитый ген путем лигирования FseI/StuI-фрагмента вектора pETR1484 с Fsel/BamHI-фрагментом вектора pETR1166 в вектор pETR12177(FseI/BamHI). Полученная в результате плазмида (pETR1500) содержала гибридный ген ManII-GnTIII (SEQ ID NO: 12) под контролем промотора MPSV. Для селекции плазмиды в клетке млекопитающего в нее была введена (в виде SpeI-фрагмента из вектора pETR1166) кассета, содержащая ген устойчивости к пуромицину, в результате чего получили плазмиду pETR1506.
Последовательности праймеров:
GAB-252:
GCTAGGCCGGCCGCCACCATGAAGTTAAGCCGCCAGTTCACCGTGTTCGG
GAB-253:
GGGGACAGTGGCTGGAGCAGGGGTGAGCCAGCACCTTGGCTGAAATTGCTTT
GTGAACTTTCGG
GAB-254:
TCCGAAAAGTTCACAAAGCAATTTCAGCCAAGGTGCTGGCTCACCCCTGCT
CCAGCCACTGTCCCC
GAB-255: ATGCCGCATAGGCCTCCGAGCAGGACCCC
pETR1519: Комбинация гибридного гена полипептида слияния ManII-GnTIII с ориджином репликации oriP вируса Эпштейна-Барр.
Используя праймеры GAB-261 и GAB-262 амплифицировали содержащий oriP фрагмент длиной 2 кб плазмиды рСЕР4 (Invitrogen). В ходе реакции ПЦР на обоих концах фрагмента были введены сайты SspI и EcoRI. Для секвенирования фрагмент с oriP был введен в вектор pICI19H. После того, как правильность последовательности были подтверждена, вводили oriP в виде SspI-фрагмента в вектор pETR 1001 (расщепляли рестриктазой BsmBI, перекрывающиеся 5'-концы заполняли, используя полимеразу Кленова). Полученная плазмида было обозначена как pETR1507. Экспрессионная кассета ManII-GnTI в виде SphI/NruI-фрагмента вектора pETR1510 была введена в вектор pETR1507, расщепленный теми же эндонуклеазами, что дало в результате плазмиду зETR1519.
Последовательности праймеров:
GAB-261: GCTAAATATTGAATTCCCTTTATGTGTAACTCTTGGCTGAAGC
GAB-262: TAGCAATATTGAATTCGCAGGAAAAGGACAAGCAGCGAAAATTCACGC
pETR1537: Комбинация гибридного гена полипептида слияния ManII-GnTIII и усеченного гена маркера клеточной поверхности CD4.
Вектор экспрессии pETR1506 модифицировли для дополнительной экспрессии усеченного гена маркера клеточной поверхности CD4. Вкратце, экспрессионная кассета гибридного гена полипептида слияния ManIII-GnTIII была преобразована из моноцистронной в бицистронную экспрессионную кассету путем вставки, в направлении «по ходу транскрипции» от стоп-кодона гена белка слияния ManIII-GnTIII, элемента IRES вируса полиомиелита и следующей за ним кДНК, кодирующей усеченный белок CD4 человека (содержащий лидерную последовательность CD4 человека, необходимую для секреции, и расположенные за ней трансмембранный и экстрацеллюлярный домены).
3. Трансфекция клеток млекопитающих векторами экспрессии полипептида слияния GnTIII и антител.
Трансфекция клеток ВКН
Клетки в экспоненциальной фазе роста (жизнеспособность 90-95%) высевали в соответствующее число флаконов Т75 в концентрации 0.9×106 клеток/мл за 24 часа до проведения электропорации. Использовали культуральную среду Invitrus (Cell Culture Technologies, Швейцария) дополненную 10% фетальной сыворотки крупного рогатого скота. Перед электропорацией подсчитывали количество клеток. Собирали 8×106 клеток путем центрифугирования (5 мин, 200g) и удаляли супернатант. Клетки ресуспендировали в 800 мл среды Invitrus и переносили в стерильные кюветы для электропорации (зазор 0.4 см), содержащие 8 мкг кольцевой плазмидной ДНК, затем инкубировали в течение 5 мин при комнатной температуре. Клетки подвергали электропорации в электропораторе Gene Pulser II (BioRad) при следующих условиях: 400В, 960 мкФ, два импульса в 30 сек. интервале. После электропорации клетки немедленно переносили во флакон 25Т, содержащий 5 мл среды для роста (Invitrus/20% об. фетальной сыворотки/1.25% об. ДМСО), инкубировали при 37°С в инкубаторе с атмосферой, содержащей 5% CO2. Для получения неизмененных (негликомодифицированных) антител клетки трансфецировали только векторами экспрессии антител. Для получения гликомодифицированных антител клетки подвергали котрансфекции двумя плазмидами - одной для экспрессии антитела, а другой, для экспрессии полипептида слияния GnTIII (SEQ ID NO: 15) - в соотношении 3:1, соответственно.
Трансфекция клеток HEK293-ENBA
Клетки HEK293-ENBA в экспоненциальной фазе роста трансфецировали кальций-фосфатным способом. Клетки выращивали в виде адгезивной монослойной культуры во флаконах Т со средой DMEM, дополненной 10% фетальной сыворотки, и трансфецировали, когда непрерывность слоя находилась в пределах 50-80. Для трансфекции флакона Т75 за 24 часа до трансфекции высевали 8 миллионов клеток в 14 мл культуральной среды DMEM, дополненной фетальной сывороткой (в конечном соотношении 10% об.) и 250 мкг/мл неомицина, затем клетки помещали на ночь при 37°С в инкубатор с атмосферой, содержащей 5% СО2. Для каждого подлежащего трансфекции флакона Т75 готовили раствор, состоящий из ДНК, CaCl2 и воды. Для этого смешивали 47 мкг тотальной ДНК плазмидного вектора, 235 мкл 1М раствора CaCl2 и доводили водой до конечного объема 469 мкл. К этому раствору добавляли 469 мкл раствора, содержащего 50 мМ HEPES, 280 мМ NaCl и 1.5 мМ Na2HPO4, с рН 7.05, немедленно перемешивали в течение 10 сек, и оставляли стоять при комнатной температуре в течение 20 сек. Суспензию разбавляли 12 мл среды DMEM, дополненной 2% фетальной сыворотки, и добавляли в Т75 вместо существующей среды. Клетки инкубировали при 37°С, 5% СО2 в течение приблизительно 17-20 часов, затем среду заменяли на 12 мл DMEM с 10% фетальной сыворотки. Для получения неизмененных (негликомодифицированных) антител клетки трансфецировали только векторами экспрессии антител. Для получения гликомодифицированных антител клетки подвергали котрансфекции двумя плазмидами - одной для экспрессии антитела, а другой, для экспрессии полипептида слияния GnTIII в соотношении 4:1, соответственно. На 5 день после трансфекции собирали супернатант, центрифугировали его в течение 5 мин при 1200 об/мин, после чего центрифугировали его второй раз (4 мин при 4000 об/мин). Супернатант хранили при 4°С.
Получение стабильных клеточных линий, экспрессирующих рекомбинантное анти-CD20 антитело
Клетки ВНК (ВНК21-13с) трансфецировали вектором экспрессии pETRT502 антителаС2В8, который содержит экспрессионную кассету гена устойчивости к неомицину, путем электропорации. Сначала отбирали устойчивые к неомицину клоны, чтобы получить набор клонов, которые интегрировали ДНК вектора pETR1502 в хоромосомы. Клоны затем исследовали на предмет продуцирования рекомбинантных антител при помощи твердофазного ИФА. Вкратце, через 24 часа после электропорации подсчитывали жизнеспособные клетки и определяли эффективность трансфекции путем подсчета флуоресцирующих клеток в параллельной контрольной электропорации вектором экспрессии pEYFP. Клетки разбавляли селективной средой Invitrus, содержащей 10% фетальной сыворотки и 1 мг/мл неомицина. Обычно восемь 96-луночных планшетов засевали жизнеспособными трансфецированными клетками в разных концентрациях (1×103, 5×102 и 1×102 клеток/лунку) и инкубировали при 37°С до тех пор, пока не становилось возможным идентифицировать клоны. Как только монослои клеток клонов становились почти непрерывными, их супернатанты анализировали методом твердофазного ИФА (ELISA). ELISA-положительные клоны переводили в емкости с большим объемом: вначале в 24-луночные планшеты, затем в 6-луночные планшеты и после этого во флаконы Т25. После 5 дней роста во флаконах Т25 определяли конечный титр антител при помощи набора для твердофазного ИФА. При помощи процедур электропорации и отбора был выделен клон клеток ВНК, экспрессирующий анти-CD20 антитело С2В8, ВНК-1502-28, который продуцировал в описанных выше условиях культивирования 13 мкг/мл антитела.
Получение стабильных клеточных линий млекопитающих, экспрессирующих рекомбинантное анти-CD20 антитело и полипептид слияния GnTIII.
Клон ВНК-1502-28, конститутивно экспрессирующий гены анти-CD20 моноклонального антитела и ген устойчивости к неомицину, трансфецировали вектором экспрессии pETR1537 путем электропорации. pETR1537 представляет собой вектор для конститутивной экспрессии гена ManII-GnTIII и усеченной формы CD4, причем экспрессия усеченной формы CD4 является IRES-зависимой. Вектор также содержит экспрессионную кассету гена устойчивости к пуромицину. Вначале отбирали устойчивые к пуромицину клоны для того, чтобы получить набор клонов, которые интегрировали ДНК вектора pETR1537 в свои хромосомы. Затем клоны исследовали на предмет экспрессии на поверхность усеченного CD4 (tCD4), который служит маркером уровня экспрессии бицистронной единицы экспрессии гена ManII-GnTIII+tCD4. Продуцирование рекомбинантыных антител отобранными клонами проверяли методом твердофазного ИФА.
Вкратце, ДНК вектора pETR1537 линеаризовали рестрикцией Xmnl. Параллельно проводили контрольную трансфекцию вектором экспрессии флуоресцентного белка EYFP. Эффективность трансфекции определяли через 24 часа, подсчитывая отношение числа клеток экспрессирующих EYFP к общему числу клеток. 58% клеток экспрессировали EYFP. Жизнеспособность клеток составляла 91%. Через 1 день после трансфекции проводили серийные разведения (1:100,1:500:1:1000 и 1:5000) клеток, трансфецированных векторами pETR1537 или pEYFP, и затем высевали в 96-луночные планшеты с селективной средой (конечный объем 0.2 мл, состав: Invitrus, 10% фетальной сыворотки, 20 мкг/мл пуромицина, 1 мг/мл неомицина). Клоны становились видимыми через 2 недели. Их размножали и исследовали на предмет экспрессии CD4(tCD4) и экспрессии антитела.
Для исследования уровня экспрессии tCD4 промывали буфером для FACS приблизительно 500,000 клеток и инкубировали на льду с 5 мл меченных Р1ТС(ФИТЦ, флуоресцин изотиоцианат) анти-CD4 ч антител (Becton Dickinson, Швейцария) в течение 10 минут. После двух промывок клетки ресуспендировали в 0.5 мл буфера для FACS и анализировали по методу «сортировки клеток с активацией флуоресценцией» (FACS) (Фиг. 17А-В). Таким образом был выделен клон с хорошим уровнем экспрессии tCD4 (ВНК-1502-28-11). После 5 дней роста в флаконе Т25 он продуцировал анти-CD20 антитела с конечными титром (определенным методом твердофазного ИФА) приблизительно равным 17 мкг/мл.
4. Получение и очистка неизмененных и гликомодифицированных антител.
Сбор культуральной среды
В случае с клетками ВНК, трансфецированными вектором экспрессии антитела или подвергнутыми котрансфекции вектором экспрессии антитела и вектором экспрессии полипептида слияния GnTIII, супернатант культуры собирали через 96 часов культивирования трансфецированных клеток после трансфекции. В зависимости от ожидаемой продуктивности для одного вектора проводили несколько(10-15) электропораций.
В случае с клетками HEK293-EBNA, трансфецированными вектором экспрессии антитела или подвергнутыми котрансфекции вектором экспрессии антитела и вектором экспрессии полипептида слияния GnTIII, культуральную среду заменяли на свежую культуральную среду приблизительно через 16 часов после трансфекции. Свежую культуральную среду собирали затем после дополнительных 120 часов культивирования трансфецированных клеток.
Для стабильной клеточной линии ВНК-1502-28-11 культуру высевали в концентрации 500,000 клеток/мл, а супернатант собирали после 4 дней культивирования, когда плотность клеток составляла 1.7×106 жизнеспособных клеток/мл при жизнеспособности равной 69%.
Очистка антитела
Моноклональное антитело очищали от культурального супернатанта посредством двух последовательных стадий хроматографии. Первая стадия состояла из хроматографии с использованием белка А с элюцией в градиенте рН, которая эффективно разделяет человеческий и бычий IgG. За этой стадией следовала стадия катионообменной [роматографии для замены буфера пробы на фосфатный буферный раствор (ФБР).
5. Анализ олигосахаридов
Олигосахариды отщепляли от иммобилизованных на мембране PVDF(Ha основе поливинилденфторида) или растворенных антител (высвобождали) ферментативным путем: при помощи расщепления гликозидазой PNGaseF.
Полученный в результате расщепления раствор, содержащий высвобожденные олигосахариды либо сразу готовили для анализа на масс-спектрометре MALDI/TOF-MS, либо подвергали перед подготовкой для анализа на масс-спектрометре MALDI/TOF-MS дальнейшему расщеплению эндогликозидазой EndoH.
Способ высвобождения олигосахаридов для иммобилизованных на мембране PVDF антител.
Лунки 96-луночного планшета, покрытого мембраной PVDF (Immobilon Р, Millipore, Бедфорд, Масачусетс) смачивали 100 мкл метанола, и прокачивали жидкость сквозь PVDF-мембрану под действием вакуума, созданного фильтрационной системой Multiscreen vacuum manifol (Millipore, Бедфорд, Масачусетс). PVDF-мембраны трижды промывали 300 мкл воды. Лунки затем промывали 50 мкл буфера RCM (8М мочевина, 360 мМ Tris, 3.2 мМ ЭДТА, рН 8.6). В ячейку, содержащую 10 мкл буфера RCM, помещали от 30 до 40 мкл антитела. Жидкость под вакуумом продавливали через мембрану, затем мембрану дважды последовательно промывали 50 мкл буфера RCM. Дисульфидные мостики разрушали посредством добавления 50 мкл дитиотрейтола в буфере RCM и инкубации при 37°С в течение одного часа.
После разрушения мостиков при помощи вакуума удаляли раствор дитиотрейтола из лунки. Лунки трижды промывали 300 мкл воды, а затем проводили карбометилирование остатков цистеина посредством добавления 50 мкл 0.1 М йодуксусной кислоты в буфере RCM и инкубировали 30 мин при комнатной температуре.
После карбометилирования ячейки сушили под вакуумом и затем трижды промывали 300 мкл воды. Затем PVDF-мембрану блокировали, для предотвращения адсорбции эндогликозидазы, посредством инкубирования 100 мкл 1% водного раствора поливинилпирролидона 360 при комнатной температуре в течение одного часа. Затем блокирующий реагент удаляли мягким вакуумом с тремя последующими промывками 300 мкл воды.
N-связанные олигосахариды высвобождали, добавляя 2.5 mU пептид-N-гликозидазыF (PNGaseF-рекомбинантная N-гликаназа, recombinant N-Glykanase, GLYKO, Новато, Калифорния) и 0.1 mU Сиалидазы (Sialidase, GLYKO, Новато, Калифорния), чтобы удалить любые заряженные остатка моносахаридов. Конечный объем - 25 мкл в растворе 20 мМ NaHCO3, рН 7.0. Расщепление проводили при 37°С в течение 3 часов.
Способ высвобождения для антител в растворе
От 40 до 50 мкл антитела смешивали с 2.5mU гликозидазы PNGaseF (Glyko, США) в 2 мМ Tris (рН 7.0) (конечный объем 25 микролиторов); смесь инкубировали при 37°С в течение 3 часов.
Применение расщепления ЭнодогликозидазойН высвобожденных гликозидазой PNGaseF олигосахаридов для установления соответствия между структурами гибридных разветвленных олигосахаридов пикам нейтральных олигосахаридов. полученных на масс-спектрометре MALDI/TOF-MS
Высвобожденные гликозидазой PNGaseF подвергали затем расщеплению Эндогликозидазой Н (EndoH, КФ 3.2.1.96). Для расщепления эндогликозидазой Н в раствор, полученный после расщепления гликозидазой PNGaseF (антитело в растворе, способ описан выше), добавляли 15 mU EndoH для получения конечного объема, равного 30 микролитрам, смесь инкубировали в течение 3 часов при 37°С. EndoH расщепляет олигосахариды между остатками N-ацетилглюкозамина хитобиозного кора N-связанных олигосахаридов. Фермент расщепляет только гликаны «олигоманнозного» типа (содержащие только N-ацетилглюкозамин и маннозу) и большинство гликанов гибридного типа, тогда как олигосахариды сложного типа этим ферментом не гидролизуются.
Приготовление проб для масс-спектроскопии на приборе MALDI/TOF-MS
Продукты ферментативного расщепления, содержащие высвобожденные олигосахариды, инкубировали в течение еще 3 часов при комнатной температуре после добавления уксусной кислоты (конечная концентрация 150 мМ), а затем пропускали через 0.6 мл катионообменной смолы (смола AG50W-X8, водородная форма, гранулы 100-200, BioRad, Швейцария), заключенной в хроматографической колонке Micro Bio-Spin (BioRad, Швейцария), для удаления катионов и белков. Один микролитр полученной в результате пробы наносили на пластину-мишень из нержавеющей стали и смешивали на ней с 1 мкл матрикса sDHB. Матрикс sDHB готовили следующим образом: 2 мг 2,5-дигидробензойной кислоты и 0.1 мг 5-метоксисалициловаой кислоты растворяли в 1 мг раствора состава: этанол/10 мМ водный р-р хлорида натрия (соотношение 1/1 по объему). Пробы высушивали воздухом, наносили 0.2 мкл этанола и в заключение позволяли рекристаллизоваться на воздухе.
MALDI/TOF-MS
Для получения спектров масс применяли масс-спектрометр MALDI-TOF, модель Voyager Elite (Perspective Biosystems). Прибор работал в линейном режиме с ускорением 20 кВ и задержкой 80 нс. Для установления соответствия масс ионов применяли внешнюю калибровку стандартными олигосахаридами. Для получения конечного спектра совмещали спектры, полученные от 200 лазерных импульсов 200 импульсов.
6. Приготовление МНПК
Мононуклеары периферической крови (МНПК) готовили с использованием Histopaque-1077 (Sigma Diagnostics Inc., Сент-Луис, M063178, США), строго следуя инструкциям производителя. Вкратце, у добровольцев, пробежавших в полную силу 1 минуту для увеличения процентного содержания естественных клеток-киллеров (NK-клеток), отбирали венозную кровь шприцами с гепарином. Кровь разбавляли ФБР, не содержащем Са и Mg, в соотношении 1:0.75-1.3и наносили на Histopaque-1077. Градиент центрифугировали при 400×g без перерывов в течение 30 минут при комнатной температуре (КТ). Жидкость, прилежащую к границе с Histopaque, содержащую МНПК, собирали, промывали ФБР (50 мл на клетки с двух градиентов), и отделяли МНПК посредством центрифугирования при 300×g в течение 10 минут при КТ. После ресуспендирования осадка в ФБР, МНПК подсчитывали и промывали второй раз посредством центрифугирования при 200×g в течение 10 минут при КТ. Затем клетки ресуспендировали в подходящей среде для последующих процедур.
7. Выделение NK-клеток
NK-клетки человека выделяли из МНПК, применяя процедуру негативного отбора с использованием магнитных гранул, не связывающих CD16- и С056-положительные клетки ((MACS system, Miltenyi Biotec GmbH, 51429 Бергиш Гладбах, Германия)). МНПК промывали один раз ледяным буфером MACS (ФБР, содержащий 2% фетальной сыворотки и 2 мМ ЭДТА), ресуспендировали в концентрации 20 млн клеток на мл смеси фетальной сыворотки и буфера MACS (1:1), а затем инкубировали в течение 10 мин при 4°С. Затем клетки осаждали и ресуспендировали в буфере MACS, содержащем 10% фетальной сыворотки, (80 мкл буфера на 10 миллионов клеток). Затем добавляли 20 мкл раствора Гаптен-антитело на 10 млн. клеток. Клетки инкубировали в течение 10 мин при 4°С, постоянно вращая пробирки. После двух промывок в буфере MACS (по меньшей мере в 10 маркировочных объемах буфера) После однократной промывки буфером MACS (в по меньшей мере десятикратном объеме) клетки ресуспендировали в буфере MACS, содержащем 10% фетальной сыворотки, в концентрации 10 миллионов клеток на 80 мкл и добавляли по 20 мкг анти-гаптен микрогранул (т.е. микрогранул, покрытых антителами к гаптену) на каждые 100 миллионов клеток. Пробирки инкубировали в течение 15 минут при 4°С, постоянно вращая пробирки. После одной промывки буфером MACS клетки ресуспендировали в концентрации до 100 миллионов клеток в 500 мкл буфера MACS и загружали в колонку LS MACD, которую помещали в магнит MINI-MACS и уравновешивали 3 мл буфера MACS. Колонку трижды промывали 3 мл буфера MACS.
Клетки собирали в проточной фазе и впоследствии использовали в качестве NK-клеток. Чистота, определенная по экспрессии CD56, составляла 88-95%.
8. Анализ АЗКЦ
МНПК или NK в качестве эффекторных клеток готовили, как описано выше. Соотношение эффекторных клеток к клеткам-мишеням составляло 25:1 или 10:1 для МНПК и NK-клеток, соответственно. Эффекторные клетки готовили в среде AIM-V в концентрации, подходящей для того, чтобы поместить по 50 мкл в каждую лунку 96-луночного пленшета. Клетки-мишени антител С2В8 представляли собой клетки SKW6.4 или В-лимфобластоидные клетки линии Namalwa, выращенные в среде DMEM, содержащей 10% фетальной сыворотки. Клетки-мишени промывали в ФБР, подсчитывали и ресуспендировали в среде AIM-V в концентрации 0.3 миллиона клеток на мл для того, чтобы поместить по 30'000 клеток в 100 мкл раствора в каждую лунку микропланшета. Антитела разбавляли в среде АГМ-V, добавляли по 50 мкл полученного раствора к предварительно нанесенным на планшет клеткам-мишеням и оставляли на 10 мин при КТ для связывания. Затем добавляли эффекторные клетки и инкубировали планшет в течение 4 часов при 37°С во влажной атмосфере, содержащей 5% СО2. Лизис клеток-мишеней, определяли, измеряя высвобождение лактат дегидрогеназы(ЛДГ) из поврежденных клеток при помощи набора для определения цитотоксичности Cytotoxicity Detection kit (Roche Diagnostics, Роткрейц, Швейцария). После четырехчасовой инкубации планшеты центрифугировали при 800×g. По 100 мкл супернатанта из каждой лунки переносили в новый прозрачный плоскодонный 96-луночный планшет. В каждую лунку добавляли по 100 мкл цветного субстрата из набора. Значения Vmax цветной реакции определяли на считывающем устройстве для твердофазного ИФА (длина волны 490 нм) в течение по меньшей мере 10 мин, используя программное обеспечение SOFTmax PRO software (Molecular Devices, Саннивейл, CA94089, США). Спонтанное высвобождений ЛДГ измеряли в лунках, содержащих только клетки-мишени и эффекторные клетки, без антител. Максимальное высвобождение определяли в лунках, содержащих только клетки-мишени и 1% Triton Х-100. Процент специфического антиген-опосредованного лизиса вычисляли следующим образом: ((x-SR)/MR-SR)*100, где х обозначает среднее Vmax для определенной концентрации антител, SR - среднее Vmax спонтанного высвобождения, MR - среднее Vmax максимального высвобождения.
9. Связывание рецептора FcγRIIIA на NK-клетках
Свежевыделенные NK-клетки центрифугировали 5 минут при 200х g и протравливали 0.09% (вес/объем) раствором молочной кислоты (140 мМ NaCl, 5 мМ KCl, рН 3.9), посредством инкубации в концентрации 3×105 клеток/мл при комнатной температуре в течение 5 минут удаления для NK-ассоциированных IgG (De Haas М., J.Immunol. 156: 2948 (1996)).
Клетки дважды промывали смесью ФБР, 0.1% БСА, 0.01% азида натрия и доводили концентрацию до 2×106 клеток/мл в смеси ФБР, 0.1% БСА, 0.01% азида натрия. 5×105 клеток инкубировали в течение 30 минут при 4°С с 0, 0.1, 0.3, 1, 3 и 10 мг/мл вариантов антитела. Затем клетки дважды промывали и определяли связывание антитела посредством инкубирования с конъюгатом флуоресцин изотиоцианина и IgG козы против F(ab')2 человека (1:200) и с конъюгатом антитела против CD56 человека и РЕ (фикоэритрина) (5 мкл/5×105 клеток) в течение 30 минут при 4°С (Shields R. Et al. J. Biol. Chem. 277 (30): 26733-26740 (2002)). IgG козы против F(ab')2 человека были получены в Jackson hnmunoReasearch, West Grove, Пенсильвания, а конъюгат анти-чCD56-PE -BD Pharmingen, Сан-Диего, Калифорния.
Интенсивность флуоресценции, соответствующей связавшимся вариантам антител, определяли для CD56+ клеток на приборе FACSCalibur (BD Bioscience, Сан-Хосе, Калифорния).
10. Связывание рецептора FcgammaRIIb на клетках лимфомы линии Raji.
В-клетки лимфомы линии Raji промывали 20 мин при 37°С в ФБР (концентрация 0.3 млн клеток/мл). Затем клетки ресуспендировали в концентрации 2.22 млн клеток/мл в смеси ФБР, 0.1% БСА, 0.01%) NaN3 и добавляли по 180 мкл полученной суспензии в каждую пробирку для FACS. К клеткам линии Raji добавляли неизмененные антитела L19 и гликомодифицированные антитела L19 в десятикратных разведениях (0, 0.1, 0.3, 1, 3, 10, 30 мкг/мл) и инкубировали при 4°С в течение 30 минут (конечная концентрация клеток 2 млн. клеток/мл). После двух промывок добавляли конъюгат флуоресцин изотиоцианина с IgG козы против F(ab')2 человека (Jackson ImmunoReasearch, West Grove, Пенсильвания) в соотношении 1:200 и инкубировали при 4°С в течение 30 минут. После одной промывки клетки ресуспендировали в 0.5 мл смеси ФБР, 0.1% БСА, 0.01% NaN3 и определяли интенсивность флуоресценции, относящейся к связавшимся вариантам антител, на приборе FACSCalibur (BD Bioscience, Сан-Хосе, Калифорния).
35
11. Анализ комплемент-зависимой клеточной цитотоксичности
Клетки-мишени подсчитывали, промывали в ФБР, ресуспендировали в среде AIM-V (Invitrogen) концентрации 1 миллион клеток/мл. По 50 мкл клеток вносили в лунки плоскодонного 96-луночного планшета. Антитела разводили в среде AIM-V и добавляли по 50 мкл к клеткам. Затем антителам оставляли для связывания с клетками в течение 10 минут при комнатной температуре. Свежеразмороженный комплемент сыворотки человека (Quidel) разбавляли в три раза средой AIM-V и добавляли в объеме 50 мкл в лунки. Комплемент кролика (Cedarlane Laboratories) готовили согласно описаниям производителя, разбавляли в три раза средой AJM-V и добавляли в объеме 50 мкл в лунки. В качестве контроля использовали прогретые в течение 30 минут при 56°С исходники комплемента.
Планшеты для анализа инкубировали в течение 2 часов при 37°С. Лизис клеток определяли по высвобождению ЛДГ. Вкратце, планшеты центрифугировали при 300×g в течение 3 мин. По 50 мкл супернатанта из каждой ячейки переносили в новый 96-луночный планшет и добавляли по 50 мкл реагента из набора для определения цитотокчсичности Cytotoxity Kit (Roche). В ходе кинетических измерений на считывающем устройстве для твердофазного ИФА определяли значение Vmax, соответствующее концентрации ЛДГ в супернатанте. Максимальное высвобождение определяли посредством инкубирования клеток с 1% Triton Х-100.
Результаты и обсуждение
Гликомодифицированные варианты анти-CD20 химерного антитела IgG1 (химерное антитело С2В8, также известное как rituximab) были получены посредством котрансфекции клеток млекопитающих векторами экспрессии антител и векторами экспрессии генов, кодирующих различные полипептиды с активностью GnTIII. Неизмененные (негликомодифицированные) варианты тех же антител получали посредством трансфекции клеток млекопитающих только вектором экспрессии гена антитела. Трансфецированные клетки культивировали по меньшей мере в течение 3 дней, затем секретированные рекомбинантные антитела очищали от культуральной среды путем аффинной хроматографии с использованием белка А. Экспрессия генов, кодирующих полипептид с активностью GnTIII не оказывала существенного влияния на жизнеспособность клеток, их рост или продуцирование антител по сравнению с клетками, не продуцирующими такие полипептиды.
Объектом анализа являлся паттерн гликозилирования очищенных антител. Эти антитела несут N-связанные олигосахариды, прикрепленные исключительно к остатку Asn297 Fc-области IgG1 человека. Олигосахариды отделяли от антител ферментативным путем (при помощи расщепления гликозидазой PNGaseF) и анализировали затем на масс-спектрометре MALDI/TOF-MS. Эта методика позволяет доли различных видов олигосахаридов в общей, исходной, популяции олигосахаридов, связанных с Fc-областью, а также установить соответствие структур различным пикам спектров (Umana, P. et al., NatureBiotechnol. 17: 176-180(1999)).
На Фиг. 1 показаны полученные при помощи MALDI/TOF-MS спектры(составы) нейтральных олигосахаридов химерного анти-CD20 антитела IgG1, продуцируемого клетками ВНК. Эти клетки были трансфецированы вектором экспрессии антитела pETR1502. На Фиг. 2 - Фиг. 4 показаны соответствующие спектры для того же антитела, продуцируемого клетками ВНК, модифицированными нуклеиновыми кислотами, кодирующими GnTIII дикого типа. Спектр на Фиг. 2 получен в результате применения нуклеиновой кислоты, кодирующей GnTIII дикого типа, экспрессируемой с вектора pETR1166. Спектр на Фиг. 3 получен в результате применения нуклеиновой кислоты, кодирующей полипептид слияния, содержащий на N-конце домен локализации GnTI, слитый с С-концевым каталитическим доменом GnTIII. Этот слитый ген экспрессировался с вектора pETR1425. Спектр на Фиг. 4 получен в результате применения нуклеиновой кислоты, кодирующей полипептид слияния, содержащий на N-конце домен локализации вкомплексе Гольджи альфа-маннозидазы II (ManII), слитый каталитическим доменом GnTIII. Этот слитый ген экспрессировался с вектора pETR1506.
Неизмененное антитело обладает типичным спектром (составом) олигосахаридов (Фиг. 1), в котором присутствуют пики с отношениями m/z 1485, 1648 и 1810, соответствующими двуантенным кор-фукозилированным сложным олигосахаридам с количеством остатков галактозидазы 0, 1 и 2 соответственно. Схожие спектры обнаруживают олигосахариды Fc-области немодифицированных антител класса IgG1, продуцируемых другими стандартными промышленными линиями клеток млекопитающих, такими как СНО и клетки миеломы мыши (Lifely, М.R. et al., Glycobiology 5: 813-822 (1995)). Модифицирование антител, продуцируемых клетками путем экспрессии GnTIII дикого типа приводит, в основном, к появлению разветвленных сложных двуантенных олигосахаридов (Фиг. 2) с пиками при соотношении m/z, равном 1689, 1851 и 2014, которые соответствуют разветвленным копиям пиков неразветвленных
фукозилированных олигосахаридов, обнаруженных в неизмененном антителе. Модифицирование продуцирующих антитела клеток путем экспрессии нуклеиновой кислоты, кодирующей полипептид слияния GnTI-GnTIII, каталитический домен GnTIII которого локализуется при помощи отвечающего за локализацию в комплексе Гольджи домена GnTI, также приводит, в основном, к появлению разветвленных сложных двуантенных олигосахаридов (обратите внимание на пики с m/z 1689 и 1851 на Фиг. 3) Однако, относительно GnTIII дикого типа, применение белка слияния GnTI-GnTIII приводит к увеличению доли разветвленных нефукозилированных и разветвленных гибридных структур (сравните отношение пиков с m/z 1664, 1810, 1826 и 1973 к общему на Фиг. 2 и Фиг. 3). Синтез разветвленных нефукозилированных и разветвленных гибридных структур является результатом конкуренции за субстраты, модифицируемые GnTI, между каталитическим доменом рекомбинантной GnTIII и (i) эндогенной альфа-1,6-кор-фукозилтрансферазой, (ii) альфа-маннозидазой II (ManII) комплекса Гольджи и (iii) GnTIIII; поскольку олигосахарид, модифицированный в ходе катализируемой GnTIII реакции добавлением N-ацетилглюкозамина в месте разветвления («разветвляющего GlcNAc»), не может после этого быть модифицирован тремя остальными ферментами. Следовательно, блокирующее действие ManII, добавляющей разветвляющий GlcNAc, также эффективно блокирует GnTII, поскольку GnTII действует по направлению пути синтеза N-связанных олигосахаридов «ниже», чем ManII. Пики с m/z 1664 и 1826 представляют не фукозилированные олигосахариды, а с m/z 1810 и 1973 - фукозилированные. Для подтверждения того, что увеличение этих пиков является следствием увеличения доли связанных с Fc-областью разветвленных неефукозилокованных и разветвленных гибридных олигосахаридов(Фиг. 8А), применяли расщепление гликозидазой EndoH, которая способна «различать» гибридные и сложные олигосахариды (см. ниже).
Модификация продуцирующих антитело клеток путем экспрессии нуклеиновой кислоты, кодирующей полипептид слияния ManII-GnTIII(SEQ ID NO: 12), где каталитический домен GnTIII локализуется через отвечающий за локализацию в комплексе Гольджи домен ManII, в отличие от других использованных здесь нуклеиновых кислот, кодирующих активность GnTIII, приводит, в основном, к появлению разветвленных нефукозилированных и разветвленных гибридных структур (обратите внимание на пики с m/z 1664, 1810, 1826 и 1973 на Фиг. 4). Следовательно, по сравнению с GnTIII дикого типа и со слитым GnTI- GnTIII (SEQ ID NOS: 14 и 15), слитый ManII-GnTIII был более эффективен в синтезе прикрепленных к Fc-области разветвленных нефукозилированных и разветвленных гибридных олигосахаридов (сравните отношения пиков с m/z 1664, 1810, 1826 и 1973 к общему на Фиг. 2, Фиг. 3 и Фиг. 4). Доли разветвленных нефукозилированных олигосахаридов Fc-области, полученных в результате экспрессии нуклеиновых кислот, кодирующих GnTIII дикого типа, белка слияния GnTI-GnTIII и белка слияния ManII-GnTIII составили приблизительно 4, 13 и 33% соответственно. В неизмененном (негликомодифицированном) антителе разветвленные олигосахариды обнаружены не были.
Повышение экспрессии конструкта слияния ManII-GnTIII в продуцирующих антитело клетках приводило к дальнейшему увеличению доли разветвленных нефукозилированных олигосахаридов. Это продемонстрировала экспрессия конструкта ManII-GnTIII с вектора (pETR1519), содержащего oriP для эписомной регуляции, в трансфецированных клетках HEK-ENBA. Известно, что эта экспрессионная система дает высокие уровни экспрессии, она также была использована для экспрессии генов антитела с вектора pETR1520. Спектр олигосахаридов очищенного неизмененного (негликомодифицированного) антитела, экспрессируемого в этой системе показан на Фиг. 5, на котором также вновь показан типичный спектр олигосахаридов с пиками неразветвленных фукозилированных олигосахаридов, содержащих 0,1 и 2 остатков галактозы (например, сравните Фиг. 1 и Фиг. 5., на которых показаны схожие спектры олигосахаридов неизмененного антитела, экспрессируемого на более высоких уровнях в клетках HEK-ENBA). Модификация продуцирующих антитело клеток нуклеиновой кислотой, кодирующей белко слияния ManII-GnTIII, экспрессируемый на более высоких уровнях в этой системе, приводит к получению антител, в которых большинство связанных с Fc-областью олигосахаридов были разветвленным нефукозилированным (см. Фиг. 6, на котором пики разветвленных нефукозилированных гибридных олигосахаридов с m/z 1664 и 1826 составляют в сумме 90% всех олигосахаридов).
Как упоминалось выше, эндогликозидазуН (EndoH) использовали для подтверждения соответствия разветвленных нефукозилированных и разветвленных гибридных структур различным пикам олигосахаридов, наблюдаемых на спектрах MALDI. Полученные на MALDI/TOF-MS спектры нейтральных олигосахаридов, образовавшихся при расщеплении гликозидазой PNGaseF и гликозидазами PNGaseF + EndoH гликанов, полученных из анти-CD20 химерного антитела класса IgG1, продуцируемого клетками НЕК293, гликомодифицированными путем повышенной экспрессией GnTIII(M2), показаны на Фиг. 7. Пик при m/z 1664 может быть поставлен в соответствие двум различным гликанам, а именно: нефукозилированному гибридному разветвленному и нефукозилированному сложному разветвленному. Разные структуры обнаруживают одинаковое отношение m/z вследствие одинакового состава моносахаридов (Фиг. 8В).
Расщепление эндолгикозидазой Н высвобожденных гликозидазаой PNGaseF олигосахаридов порождает новые структуры с основным пиком, сдвинутым с m/z 1664 на m/z1460 (Фиг. 7B). Разница соответствует массе остатка N-ацетилглюкозамина. Как упоминалось выше, эндогликозидаза Н не способна расщеплять олигосахариды сложного типа. Следовательно, после расщепления эндогликозидазой Н основной пик с m/z 1664 может поставлен в соответствие нефукозилированным олигосахаридам гибридного разветвленного типа.
Другие пики могут быть поставлены в соответствие сложным или гибридным разветвленным гликанам. После расщепления эндогликозидазой Н пик с m/z 1810 исчез, что говорит о том, что эти структуры относятся к типу фукозилированных гибридномых разветвленных. Удаление одного остатка GlcNAc и одного остатка фукозы (от фукозилированного альфа 1-6 кор-фукозилтрансферазой остатка GlcNAc на восстанавливающем конце) от пика с m/z 1810 порождает структуру с m/z 1460. Исчезновение пика с m/z 1502 после расщепления эндогликозидазой Н (остаток GlcNAc удален) и появление пика с m/z 1298 демонстрируют то факт, что пик 1502 может быть поставлен в соответствие олигосахаридам, относящимся к типу нефукозилированных гибридных разветвленных. Исчезновение пика с m/z 1647 после расщепления эндогликозидазой Н демонстрирует, что этот пик соответствует фукозилированной гибридной разветвленной структуре. Удаление остатка GlcNAc и одного остатка фукозы порождает структуру с m/z 1298. Пик с m/z 1826, соответствующий типу нефукозилированных гибридных разветвленных, исчез после расщепления Endo Н. Это породило структуру с m/z 1622. После расщепления эндогликозидазой Н пик на m/z 1053 может быть поставлен в соответствие олигосахаридам с высоким содержанием маннозы (m/z 1257), расщепленных эндогликозидазой Н. Расщепление эндогликозидазой Н, как и ожидалось, не оказывает влияния на пик на m/z 1689 (сложные разветвленные). В итоге, на основе данных, содержащихся в таблице 1., мы приходим к выводу, что 88% олигосахаридных структур несут разветвляющий» GlcNAc, из них 68% являются нефукозилированными разветвленными олигосахаридными структурами, 22% фукозилированными гибридными разветвленными и 6% - фукозилированными сложными разветвленными олигосахаридными структурами.

Балансы масс (в молярных долях, %):
a) Пики на m/z 1502 и 1647: 7+7%=14% (ожидаемый), Расщепление EndoH для обоих пиков порождает пик на m/z 1298 (после обработки EndoH получено 13%)
b) Пики на m/z 1664 и 1810: 47+13%=62% (ожидаемый), Расщепление EndoH порождает пик на m/z 1460 (после обработки EndoH получено 60%)
c) Пики на m/z 1826 и 1972: 4+3%=7%, Расщепление EndoH порождает пик на m/z 1622 (7%)
Резюме: Общее относительное процентное отношение структур
Приведенные выше данные (Фиг. 1 - Фиг. 6) показывают, что и уровень экспрессии GnTIII и конкретный домен локализации, который используется для направления каталитического домена GnTIII в комплекс Гольджи, влияют на конкуренцию между
каталитическим доменом рекомбинантной GnTIII, эндогенной кор-альфа-1,6-фукозилтрансферазой, ферментами ManII и GnTII за модифицированные GnTI олигосахаридные субстраты. Более высокая экспрессия обеспечивает GnTIII преимущество в этой конкуренции, приводя к повышению уровней разветвленных гибридных и разветвленных нефукозилированных олигосахаридов, а также к сопутствующему снижению уровней разветвленных сложных и разветвленных фукозилированных олигосахаридов. Ранее это было отмечено также и для GnTIII дикого типа (Umana, P.et al., Nature Biotechnol. 17: 176-180 (1999)). Хотя, несмотря на то, что локализация каталитического домена GnTIII через домен локализации GnTI или ManII приводит к сходному (с GnTIII дикого типа) общему уровню разветвленных олигосахаридов, она приводила к более эффективной, по сравнению с GnTIII дикого типа, конкуренции с эндогенной кор-альфа-1,6-фукозилтрансферазой, а также ферментами ManII и GnTII за модифицированные GnTI олигосахаридные субстраты.
Более высокую, по сравнению с GnTIII дикого типа, эффективность белка слияния GnTI-GnTIII в синтезе разветвленных гибридных и разветвленных нефукозилированных олигосахаридов можно объяснить более «ранним» положением (распределением) GnTI по отношению к GnTIII в комплексе Гольджи по направлению цис-транс транспорта субстрата. Точное распределение в комплексе Гольджи GnTI и ManII было установлено ранее методом количественной иммуноэлектронной микроскопии (Rabouille, С.et al. J Cell Sci. 108: 1617-27 (1995)). Оба фермента распределены вместе вдоль комплекса Гольджи, локализуются в основном в медиал-(срединньгх) и транс-цистернах стопки Гольджи, причем их уровень в срединных цистернах выше относительно транс-цистерн. Точное количественное распределение кор-альфа-1,6-фукозилтрансферазы и GnTIII дикого типа до сих пор не было определено. Вышеуказанное, однако, не объясняет, почему слитый ManII-GnTIII существенно более эффективен в синтезе разветвленных гибридных и разветвленных нефукозилированных олигосахаридов, чем слитый GnTI-GnTIII, поскольку пространственное распределение GnTI и ManII в субкомпартментах комплекса Гольджи идентично.
Более высокая эффективность белка слияния ManII-GnTIII указывает на существование относительно организованных функциональных реакционных субкомпартментов гликозилирования внутри физических субкомпартментов медиал- и транс-цистерн комплекса Гольджи. Считается, что так называемые «ферменты гликозилирования медиал- Гольджи», GnTI, GnTII и ManII, существуют в комплексе Гольджи в виде высокомолекулярных комплексов. Однако, если домены локализации позволяют этим ферментам образовывать часть этих комплексов, то же самое должно происходить со
слитыми GnTI-GnTIII и ManII-GnTIII. Экспрессия рекомбинантных белков слияния GnTI-GnTIII не приводила к изменению положения фермента GnTI дикого типа, сколько-нибудь существенному для синтеза олигосахаридов Fc-области, поскольку все применяемые здесь конструкты GnTIII приводили к тому, что большинство олигосахаридов было модифицировано ходе как реакций, катализируемых GnTI, так и реакций, катализируемых GnTIII.
Наши данные указывают на то, что благодаря свойствам домена локализации ManII, происходит более тонкое структурирование (объединение) каталитических доменов эндогенной GnTI и рекомбинантного белка слияния ManII-GnTIII. Известно, что организованное структурирование ферментов, катализирующих последовательные реакции какого-либо пути биосинтеза, облегчающее переход продуктов первой реакции ко второму каталитическому сайту (относительно диффузионного удаления этих продуктов от ферментов), встречается в других путях биосинтеза, таких как гликолиз и синтез полипептидов. Есть сообщения о том, что GnTI и ManII образуют «родственные олигомеры» (kin oligomers), по меньшей мере, при перемещении одного из эитх фекментов по эндоплазматическому ретикулуму (Nilsson, Т. et al. EMBOJ 13 (3). 562-74 (1994)). Было обнаружено, что по паре заряженных аминокислотных остатков в стержневых (каркасных) областях этих двух ферментов играют ключевую роль в этом «родственном распознавании». Остатки в GnTI несут заряд, противоположный заряду остатков ManII. Мы идентифицировали сходные остатки в эквивалентных положениях стержневых областей (stem regions) других ферментов гликозилирования комплекса Гольджи, вовлеченных в эту часть пути биосинтеза N-связанных олигосахаридов, а именно: кор-альфа-1,6-фукозилтрансфераза (такие же, как у GnTI заряды, вместо комплементарных им зарядов ManII), ManI и GnTII. Мы также установили, что эти остатки консервативны у разных видов. Хотя было сделано предположение, что эти остатки не являются необходимыми для встраивания в высокомолекулярные комплексы, образуемые ферментами, и даже для локализации в комплексе Гольджи (Opat, A.S. et al. J.Biol. Chem. 275(16): 11836-45 (2000)), они, возможно, вовлечены в более тонкое структурирование каталитических доменов в ходе биосинтеза олигосахаридов. Такое структурирование не обязательно является необратимым, оно может быть опосредовано временными, динамическими взаимодействиями между ферментами. Возможно, существуют дополнительные детерминанты структурирования в других положениях стержневой или каталитической области. Однако, любой вклад, каталитического домен ManII в специфическое структурирование GnTI-ManII, будет утрачен в рекомбинантном слитом полипептиде, несущем каталитический домен GnTIII.
Фиг. 9 - Фиг. 11 демонстрируют повышенную антителозависимую клеточную цитотоксичность (АЗКЦ), являющуюся результатом повышенной экспрессии нуклеиновой кислоты, кодирующей полипептид с активностью GnTIII, который локализуется в комплексе Гольджи при помощи различных доменов локализации, в продуцирующих антитело клетках. Повышенная АЗКЦ, являющаяся результатом повышенной экспрессии рекомбинантного полипептида с активностью GnTIII, который локализуется в комплексе Гольджи при помощи домена отвечающего за локализацию в комплексе Гольджи GnTI, показана на Фиг. 9. Спектр олигосахаридов контрольного антитела, использованного в анализе АЗКЦ (Фиг. 9), показан на Фиг. 1. Спектр олигосахаридов гликомодифицированного антитела, использованного в анализе АЗКЦ (Фиг. 9), показан на Фиг. 3. Повышенная АЗКЦ, являющаяся результатом повышенной экспрессии рекомбинантного полипептида с активностью GnTIII, который локализуется в комплексе Гольджи при помощи отвечающего за локализацию в комплексе Гольджи домена гликозидазы ManII, показана на Фиг. 10. Спектр олигосахаридов контрольного антитела, использованного в анализе АЗКЦ (Фиг. 10), показан на Фиг. 5. Спектр олигосахаридов гликомодифицированного антитела, использованного в анализе АЗКЦ (Фиг. 10), показан на Фиг. 6.
На Фиг. 11 показано, что экспрессия рекомбинантного полипептида с активностью GnTIII, локализованного в комплексе Гольджи при помощи отвечающего за локализацию в комплексе Гольджи домена ManII, приводит к повышенной, по сравнению с вызываемой полипептидом GnTHI дикого типа, обладающим собственным доменом, отвечающим за локализацию в комплексе Гольджи, активности АКЗЦ. Спектр олигосахаридов, антитела, гликомодицифицированного путем экспрессии GnTIII дикого типа и использованного в анализе АЗКЦ (Фиг. 11), показан на Фиг. 2. Спектр олигосахаридов антитела, гликомодицифицированного путем экспрессии полипептида слияния с активностью GnTIII, локализованного в комплексе Гольджи при помощи домена локализации ManII и использованного в анализе АЗКЦ (Фиг. 11), показан на Фиг. 4. Эти данные также показывают, что антитела с разветвленными олигосахаридами, включая разветвленные гибридные и разветвленные нефукозилированные олигосахариды, обладают повышенной, по сравнению с антителами со сложными фукозилированными неразветвленными олигосахаридами, активностью АЗКЦ. Следует отметить, что все разветвленные олигосахариды более активного антитела, использованного в анализе АЗКЦ, результаты которого показаны на Фиг. 10, представляют собой разветвленные нефукозилированные гибридные олигосахариды. Как упоминалось ранее, применение полипептида слияния с активностью GnTIII, локализованного в комплексе Гольджи при помощи домена локализации ManII, приводит к более эффективному синтезу нефукозилированных разветвленных олигосахаридов; на Фиг. 11 показано, что антитела с повышенными уровнями этих разветвленных нефукозилированных олигосахаридов более активны, по сравнению с антителами с более низкими уровнями этих олигосахаридов, в реакциях АЗКЦ. Повышение активности АЗКЦ связано с увеличением доли разветвленных нефукозилированных олигосахаридов в популяции олигосахаридов, связанных с Fc-областью, а значительное повешение активности наблюдается при увеличении этой доли на величину, большую, чем 15-20%.
Известно, что естественные киллеры (NK-клетки) являются важными участниками АЗКЦ. На поверхности этих клеток находится активирующий рецептор Fcgamma IIIA, также известный, как CD16a. Связывание Fc-области связанных с клетками-мишенями антител с рецепторами FcgammaRIIIA на поверхности NK-клеток необходимо для перекрестного связывания этих рецепторов на NK-клетках и последующей индукции АЗКЦ. Таким образом, важно оценить связывание антител, полученных описанными здесь способами с Fc-рецепторами, особенно - с рецепторами в нативной форме, т.е. в форме, в которой они существуют на поверхности эффекторных клеток иммунной системы человека.. Фиг. 12 демонстрирует, что гликомодифицированные антитела, полученные посредством экспрессии в продуцирующих антитела клетках нуклеиновой кислоты, кодирующей полипептид слияния с активностью GnTIII, обладают повышенной аффинностью связывания с активирующим Fc-рецептором человека FcgammaRIIIA. Как упоминалось выше для случаев анализа АЗКЦ, у этих антител повышен уровень разветвленных нефукозилированных олигосахаридов, что является результатом экспрессии полипептида слияния с активностью GnTIII в продуцирующих антитела клетках. NK-клетки, использовавшиеся в этом анализе были получены от донора с генотипом, не экспрессирующим рецептор FcgammaRIIc в NK-клетках донора (Metes, D. et al. J. Immunol. Methods 258(1-2): 85-95 (2001)). Таким образом, единственным Fc-рецептором на поверхности таких клеток является активирующий рецептор FcgammaRIIIA. На Фиг. 13 показано, что в анализе связывания определяется аффинность специфического связывания с этим рецептором. Это иллюстрирует конкуренция с фрагментом антитела, специфически блокирующего FcgammaRIIIA(Fab2 фрагмент 3G8).
Убедительным свидетельством сильного влияния усиленных взаимодействий между Fc-областью и Fc-рецепторами на результат лечения опухолей антителами стала связь между эффективностью гомозиготностью пациента по варианту гена, кодирующему рецептор FcgammaRIIIA с более высокой активностью («высокоаффинный» вариант), обнаруженная у получающих rituximab пациентов (Cartron, G. et al. Blood 99(3): 754-8 (2002)). Это был единственный параметр, для которого была обнаружена связь с существенным повышением объективной степени ответа и повышением доли молекулярных ответов. Источником повышенной, вследствие усиленных взаимодействий FcgammaRIIIA-Fc, эффективности могут быть функции, выполняемые различными типами клеток иммунной системы, включая естественные киллеры (NK), макрофаги и дендритные клетки. Перекрестное связывание активирующего рецептора FcgammaRIIIA на поверхности NK-клеток приводит к лизису опухолевых клеток по механизму АЗКЦ, который многие считают главным Fc-R-зависимым механизмом лизиса клеток in vivo(Maloney, D.G. et al.Semite. Oncol. 29(1 Suppl. 2): 2-9 (2002), Amigorena S., J Exp.Med. 195(1):Fl-3 (2002)), а также к антителозависимому клеточному фагоцитозу (Hazenbos, W. L. et al. J. Immunol. 161 (6): 3026-32(1998), Reff, M. E., Heard C., Crit Rev Oncol Hematol. 40(1): 25-35 (2001))и высвобождению цитокинов вблизи опухолевых клеток (Carson, W.Е. et al. Eur. J. Immunol. 31: 3016-3025 (2001)). Эти цитокины способны, в свою очередь, вызывать направленный на опухолевые клетки прямой цитолитический эффект, антиангиогенный эффект, который приводит к торможению роста опухоли вследствие прекращения снабжения ее кислородом и питательными веществами, а также к усилению презентации опухолевых антигенов, что является частью активного Т-клеточного иммунного ответа, направленного против опухолевых клеток. Дендритные клетки играют решающую роль в презентации антигенов Т-клеткам, а перекрестное связывание FcgammaRIIIA на их поверхности (например, связанных антителами умирающих опухолевых клеток, атакованных предварительно in vivo по механизму АЗКЦ) может приводить к повышению интенсивности созревания дендритных клеток, поглощения антигенов и их презентации Т-клеткам и перекрестному праймированию цитотоксичных Т-клеток, причем последнее потенциально является очень эффективным механизмом активации противоопухолевого иммунитета (AmigorenaS., J. Exp.Med. 195(1): F1-3 (2002), Kalergis, A.M. & Ravetch, J. V.J. Exp.Med. 195(12): 1653-1659 (2002), Selenko, N. et. aU. Clin. Immunol. 22(3): 124-130 (2002)). Перекрестное связывание связанных с клетками-мишенями антител с Fc-рецепторами на поверхности эффекторных клеток иммунной системы может также приводить к повышению прямого лизиса клеток-мишеней, например, путем апоптоза, индуцированного антитело-опосредованным перекрестным связыванием молекул антигенов-мишеней (Reff, М.Е. & Heard, С. Crit Rev Oncol Hematol. 40(1): 25-35 (2001), Cragg, M.S. et al. Blood 101 (3): 1045-1052 (2003)). Из всех эффекторных клеток иммунной системы только NK-клетки обладают исключительно активирующими поверхностными Fcgamma-рецепторами. В клетках других типов активирующие рецепторы FcgammaRIII присутствуют одновременно с ингибирующим рецептором FcgammaRIIb, а индукция противоопухолевых эффекторных функций является результатом преобладания активирующих сигналов над ингибирующими.
На Фиг. 15 показано, что повышение связывания Fc-рецептора селективно в отношении активирующего рецептора по сравнению ингибиторному FcgammaRIIb. Такая селективность важна, как объяснялось выше, для выполнения клетками иммунной системы, такими как NK-клетки, их эффекторных функций. Более того, повышение связывания, достигнутое путем гликомодифицирования Fc-области антитела описанными здесь способами, намного больше наблюдаемого обычно у гомозиготных по «высокоаффинному» варианту FcgammaRIIIA пациентов/доноров, получающих стандартное, неизмененное антитело (Фиг. 16), которое исследователи связали с повышенной активностью противораковых антител (Cartron, G. et al. Blood 99(3): 754-8 (2002)).
Связывающий домен активирующего рецептора FcgammaRIIIB почти полностью идентичен связывающему домену FcgammaRIIIA. Следовательно, приведенные выше данные также указывают на то, что описанные здесь гликомодифицированные антитела могут вызывать повышение эффекторных функций, опосредуемых эффекторными клетками, которые имеют на своей поверхности FcgammaRIIIB, такими как полиморфноядерные клетки, включая высвобождение токсических продуктов и фагоцитоз ((Reff, М.Е., Heard, С. Crit Rev Oncol Henaatol. 40(1): 25-35 (2001), Daeron, FM. Annu. Rev. Immunol. 15: 203-34 (1997), Ravetch, J.V., Bolland S. Annu. Rev. Immunol. 19: 275-90 (2001)).
На Фиг. 18 показан спектр олигосахаридов анти-CD20 антитела, продуцируемого клетками ВНК, растущими в суспензии и модифицированными для конститутивной экспрессии на высоком уровне как рекомбинантного антитела, так и полипептида слияния с активностью GnTIII. Спектр олигосахаридов показывает увеличение уровней разветвленных нефукозилированных и разветвленных гибридных олигосахаридов у антитела, полученного в клетке, модифицированной белком слияния GnTIII (См. также таблицу 2). Эти структуры не обнаружены в негликомодифицированных антителах, продуцируемых негликомодифицированными клетками ВНК (См. Фиг. 1). Модифицированные клетки, экспрессирующие конструкцию слияния GnTIII демонстрировании нормальный рост в суспензии и хорошую продуктивность наработки антител.
Относительное процентное содержание разных олигосахаридов гликомодифицированных моноклональных антител, продуцируемых клетками стабильной клеточной линии ВНК-1502-28-11 представлено в Таблице 2.
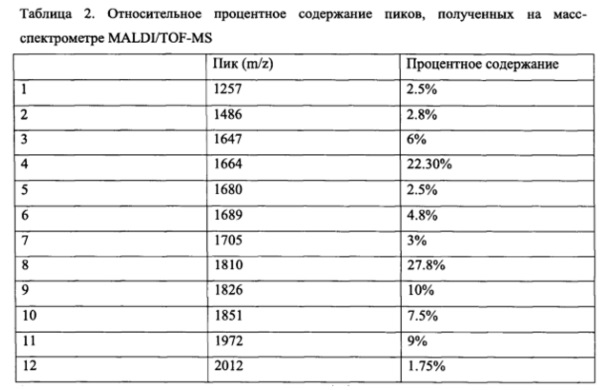
Всего разветвленных: 88.6% (4+5+6+7+8+9+10+11+12)
Всего нефукозилированных разветвленных: 37.8%(4+5+7+9)
Всего фукозилированных разветвленных: 50.8%(6+8+10+11+12)
Сложных разветвленных: 17%(6+7+10+12)
Гибридных разветвленных: 71.6%(4+5+9+11
Анализ олигосахаридов показал, что 88.6% структур несут разветвляющий остаток N-ацетилглюкозамина, 50.8% являются фукозилированными, а 37.8% - не фукозилированными. Расщепление эндогликозидазой Н высвобожденных гликозидазой PNG-ase продемонстрировало, что большинство пиков были получены от олигосахаридов гибридного разветвленного типа (Фиг. 19). Фиг. 20 показывает повышенную аффинность связывания гликомодифицированного антитела, продуцируемого клетками клеточной линии ВНК-1502-28-11, с активирующим Fc-рецептором FcgammaraIIIA на поверхности NK-клеток человека. Клеточные линии, растущие в суспензии и дающие конститутивную стабильную экспрессию генов как антитела так и полипептида слияния GnTIII, идеальны для крупномасштабного производства терапевтических антител. Применяя стандартные способы модификации клеток, можно произвести гликомодифицироание либо путем введения гена белка слияния GnTIII в клетки клеточной линии, содержащей гены антитела, либо путем введения генов антител в клетки клеточной линии, содержащие гены белка слияния GnTIII («прегликомодифицированная производственная клеточная линия»), либо путем одновременного введения генов антител и слитой GnTIII.
Анти-CD20 антитело, продуцируемое в клетках, модифицированных для экспрессии на высоком уровне нуклеиновой кислоты, кодирующей полипептид слияния с активностью GnTIII, локализуемый в комплексе Гольджи при помощи домена локализации ManII, исследовалось также на предмет комплемент-опосредованного лизиса (Зд.-КОЛ) - другой эффекторной функции, которая не связана с Fc-рецепторами на эффекторных клетках иммунной системы. Подавляющее большинство олигосахаридов этого гликомодифицированного антитела представляли собой разветвленные гибридные нефукозилированные олигосахаридов. У этих анти-CD20 антител наблюдали сниженную, по сравнению с неизмененными антителами, активность комплементопосредованного лизиса (Фиг. 21). Для некоторых приложений может быть желательным иметь антитела с повышенной АЗКЦ, но пониженным комплемент-опосредованным лизисом, например, для уменьшения побочных эффектов, таких как вызванный комплемент-опосредованным лизисом васкулит кровеносных сосудов в сайтах локализации опухолей. Другой существенный КОЛ-опосредованный побочный эффект наблюдали в ходе лечения анти-CD20 антителом (van der Kolk L.E. et al. Br J Haematol. 115 (4): 807-11 (2001)). Однако, приведенные выше спектры олигосахаридов показывают также, что возможно также модифицировать продуцирующие антитело клетки для экспрессии полипептида слияния GnTIII на среднем уровне, что приводит к средним уровням разветвленных гибридных нефукозилированных олигосахаридов (выше 15%), но со значительной долей сложных олигосахаридов в популяции связанных с Fc-областью гликомодифицированного антитела олигосахаридов. Такие сложные олигосахариды ассоциированы с нормальными, не пониженными, уровнями КОЛ. Следовательно, данные указывают на то, что антитела с повышенной АЗКЦ могут продуцироваться таким образом, чтобы КОЛ поддерживался на том же уровне, что и у немодифицированных антител.
Второе химерное антитело класса IgG2, С225, также известное как cemximab, которое распознает рецептор к эпидермальному фактору роста человека (РЭФР, EGFR) было гликомодифицированно описанными здесь способами. НА Фиг. 22 показаны спектры олигосахаридов немодифицированного анти-РЭФР антитела С225 и гликомодифицированного варианта того же антитела. Последнее продуцировалось клетками, экспрессирующими нуклеиновую кислоту, кодирующую полипептид слияния с активностью GnTIII и локализуемый в комплексе Гольджи при помощи домена локализации маннозидазыII. На Фиг. 23 показана повышенная в результате модификации АЗКЦ анти-РЭФР антитела. Гликомодифицированные антитела, полученные описанными здесь способами и обладающие повышенной АЗКЦ и повышенной аффинностью связывания активирующих Fc-рецепторов, являются перспективными молекулами для терапии рака и аутоиммунных заболеваний с использованием антител, поскольку они должны приводить к повышению эффективности, относительно соответствующих неизмененных (негликомодифицированных) вариантов тех же антител, такой терапии. Дополнительно должно стать возможным уменьшить терапевтические дозы гликомодифицированных антител по сравнению с неизмененными, что положительно скажется на экономических аспектах производства антител.
ПРИМЕР 2
Лечение иммуноопосредованной тромбоцитопении у пациента с хронической болезнью отторжения трасплантата («Graft-versus-host» disease).
Аутоиммнунная тромбоцитопения при хронической болезни отторжения трансплантата представляет собой пример связанной с В-лимфоцитами дерегуляции, приводящей к клиническому заболеванию. Для лечения иммуноопосредованной тромбоцитопении у пациента с хронической болезнью отторжения трансплантата химерное анти-CD20 моноклональное антитело, приготовленное способами настоящего изобретения и обладающие повышенной АЗКЦ, назначают пациенту, как описано в nRatanatharathom, V. et al, Ann. Intern. Med. 133(4): 275-79 (2000) (полное содержание источника включено в описание изобретения путем ссылки). Конкретно, пациенту назначают в течение 4 недель еженедельные инфузии антитела (375 мг/м2).Терапия с применением антител приводит к заметному снижению количества В-лимфоцитов в периферической крови и понижению уровней тромбоцит-ассоциированного антитела.
ПРИМЕР 3
Лечение тяжелой иммуноопосредованной истинной эритроцитарной аплазии и гемолитической анемии.
Иммуноопосредованная приобретенная истинная эритроцитарная аплазия является редким расстройством, часто ассоциированным с другими аутоиммунными явлениями. Для лечения приобретенной иммуноопосредованной истинной эритроцитарной аплазии пациенту назначают, как описано в Zecca, М. Et al., Blood 12: 3995-97 (1997) (полное содержание источника включено в описание изобретения путем ссылки), химерное анти-CD20 моноклональное антитело, приготовленное способами настоящего изобретения и обладающие повышенной АЗКЦ. Конкретно, пациент с иммуноопосредованной истинной эритроцитарной аплазией и гемолитической анемией получает еженедельно две дозы антитела, 375 мг/м2. После терапии с применением антител начинают заместительное лечение вводимым внутривенно гемеглобином. Это лечение приводит к значительному снижению уровня В-лимфоцитов в периферической крови и значительному увеличению числа ретикулоцитов, сопровождающемуся увеличением уровня гемоглобина.
ПРИМЕР 4
Материалы и методы
1. Конструирование векторов экспрессии белка слияния GalT
Вектор для конститутивной экспрессии GalT
Для конструирования вектора экспрессии GalT, кодирующую GalT к ДНК амплифицируют из библиотеки ДНК (Clonetech) путем перекрывающихся реакций ПЦР. Сразу же после стоп-кодона этого гена против направления транскрипции (upstream) добавляют С-концевой участок эпитопа с-myc для последующего удобного определения GalT по методу Вестерн-блот (последовательность аминокислот PEQKLISEEDL). После подтверждения правильной последовательности GalT ген помещают под контроль промотора MPSV и добавляют синтетический сигнал полиаденилирования бета-глобина кролика. Окончательный вектор экспрессии GalT также содержит отдельную экспрессионную кассету для отбора (селекции). Эта кассета содержит ген устойчивости к пуромицину, также находящийся под контролем промотора MPSV, и синтетический сигнал полиаденилирования бета-глобина кролика.
Замена аминокислот, кодирующих домен локализации GalT на аминоксилоты, кодирующие домен локализации GnTI человека.
Конструирование этого гена гибридной галактозилтрансферазы производят, например, путем перекрывающих реакций ПЦР, дающих в результате плазмиду, содержащую ген белка слияния GnTI-GalT под контролем промотора MPSV и кассету устойчивости к пуромицину для отбора.
Замена аминокислот, кодирующих домен локализации GalT на аминоксилоты, кодирующие домен локализации маннозидазы II человека.
Конструируют вектор экспрессии GalT. Полученная результате плазмида содержит гибридный ген ManII-GalT под контролем промотора MPSV
Комбинация гибридного гена белка слияния ManII-GalT с ориждином репликации oriP вируса Эпштейна-Барр
Фрагмент ДНК, содержащий oriP субколнируют, как описано в Примере 1, в гибридный вектор экспрессии ManII-GalT, описанный выше.
Комбинация гибридного гена белка слияния ManII-GalT с усеченным геном маркера клеточной поверхности CD4
Вектор экспрессии модифицируют, для того чтобы получить дополнительно экспрессию усеченного гена маркера клеточной поверхности CD4. Вкратце, экспрессионную кассету рекомбинантного гибридного гена белка слияния ManII-GalT преобразуют из моноцистронной в бицистронную путем вставки, в направлении «по ходу транскрипции» от стоп-кодона гена белка слияния ManII-GalT, элемента IRES вируса полиомиелита и следующей за ним кДНК, кодирующей усеченный белок CD4 человека (содержащий лидерную последовательность CD4 человека, необходимую для секреции, и расположенные за ней трансмембранный и экстрацеллюлярный домены).
3. Трансфекция клеток млекопитающих векторами экспрессии белка слияния GalT и антитела Трансфекция клеток ВНК
Клетки в экспоненциальной фазе роста (жизнеспособность 90-95%) культивировали, собирали и затем трансфецировали, как описано в Примере 1. Для получения гликомодифицированных антител клетки подвергали котрансфекции двумя плазмидами: одной для экспрессии антитела и другой - для экспрессии полипептида слияния GalT, в соотношении 3:1, соответственно
Трансфекция клеток HEK293-ENBA
Клетки HEK293-ENBA в экспоненциальной фазе роста трансфецировали, как описано в Примере 1. Для получения гликомодифицированных антител клетки подвергали котрансфекции двумя плазмидами: одной для экспрессии антитела и другой - для экспрессии полипептида слияния GalT, в соотношении 4:1, соответственно. На день 5 после трансфекции собирали супернатант, центрифугировали его вначале в течение 5 минут при 1200 об/мин, а затем в течение 10 минут при 4000 об/мин, и хранили при 4°С.
Получение стабильных клеточных линий, экспрессирующих рекомбинантное анти-CD20 антитело и слитый GalT
Клон ВНК-1502-28, конститутивно экспрессирующий гены моноклонального анти-СБ20 антитела и ген устойчивости к неомицину, трансфецируют вектором экспрессии путем электропорации. Вектор дает возможность конститутивной экспрессии гена ManII-GalT и усеченной формы CD4 человека, причем экспрессия последнего не зависит от IRES. Вектор также содержит экспрессионную кассету гена устойчивости к пуромицину. Устойчивые к пуромицину клоны вначале подвергали отбору для того, чтобы получить набор клонов, которые интегрировали в свои хромосомы векторную ДНК. Затем клоны исследовали на предмет экспрессии усеченного CD4(tCD4), который служит маркером уровня экспрессии бицистронной экспрессионной единицы гена ManII-GalT+tCD4. Продуцирование рекомбинантного антитела проверяли при помощи твердофазного иммуноферментного анализа.
Трансфекцию и последующее исследования уровня экспрессии tCD4 проводят, как описано в Примере 1.
4. Получение и очистка неизмененных и гликомодифицированных антител.
В случае клеток ВНК, трансфецированных вектором экспрессии или подвергнутых котрансфекции вектором экспрессии антитела и вектором экспрессии белка слияния GalT, клетки после трансфекции культивируют, а супернатант культуры собирают после 96 часов культивирования. В зависимости от ожидаемой продуктивности, проводят несколько (10-15) электропораций одним и тем же вектором.
В случае клеток HEK293-EBNA, трансфецированных вектором экспрессии или подвергнутых котрансфекции вектором экспрессии антитела и вектором экспрессии белка слияния GalT, культуральную среду заменяют свежей культуральной средой примерно через 16 часов после трансфекции, и свежую среду собирают после еще 120 часов культивирования трансфецированных клеток.
В случае стабильной клеточной линии ВНК-1502-28, культуры высевают в концентрации 500,000 клеток/мл и после 4 дней культивирования собирают супернатант.
Очистка антител
Моноклональное антитело очищают от культуральных супернатантов путем двух последовательных стадий хроматографии: хроматографии с использованием белка А и катионообменной хроматографии, как описано в Примере 1.
5. Анализ олигосахарида
Олигосахариды высвобождали из антител ферментативным путем: при помощи расщепления гликозидазой PNGase, при это антитела либо были иммобилизованы на мембране PVDF либо находились в растворе.
Полученный в результате расщепления раствор, содержащий высвобожденные олигосахариды либо сразу готовили для анализа на масс-спектрометре MALDI/TOF-MS, либо подвергали перед подготовкой для анализа на масс-спектрометре MALDI/TOF-MS дальнейшему расщеплению эндогликозидазой EndoH
Способ высвобождения олигосахаридов для иммобилизованных на мембране PVDF антител и Способ высвобождения олигосахаридов для антител в растворе проводят, как описано в Примере 1.
Применение расщепления гликозидазой EndoH высвобожденных гликозидазой PNGaseF олигосахаридов для установления соответствия между гибридными гликозилированными структурами олигосахаридов м пиками нейтральных олигосахаридов в спектре MALDI/TOF-MS
Высвобожденные гликозидазой PNGaseF олигосахариды подвергают затем расщеплению эндогликозидазой Н (КФ 3.2.1.96), как описано в Примере 1.
MALDI/TOF-MS
Готовят пробы, содержащие продукты ферментативного расщепления высвобожденных олигосахаридов, и затем анализируют их на масс-спектрометре MALDI/TOF-MS, как описано в Примере 1.
6. Приготовление и выделение клеток
Мононуклеары периферической крови (МНПК) готовят, используя Histopaque-1077 (Sigma Diagnosticslnc, Сент-Луис, М063178 США), неукоснительно следуя инструкциям производителя и протоколу, описанному в Примере 1.
NK-клетки человека выделяют из МНПК, применяя процедуру негативного отбора, как описано в Примере 1.
8. Анализ АЗКЦ
МНПК или NK-клетки, в качестве эффекторных клеток, готовят, как описано выше и анализируют их способность опосредовать цитотоксичность в реакциях антителозависимой клеточной цитотоксичности(АЗКЦ), как описано в Примере 1.
9. Связвание FcgammaRIIIA на NK-клетках и связывание FcgammaRIIb на клетках лимфомы линии Raji.
Связвание FcgammaRIIIA на свежевыделенных NK-клетках и связывание FcgammaRIIb на клетках лимфомы линии Raji определяют, как описано в Примере 1.
10. Анализ комплементзависимой клеточной цитотоксичности
Анализ комплемент зависимой цитотоксичности проводят, используя разведения антител, согласно способам, описанными в Примере 1.
Результаты и обсуждение
Анализы проводили с целью продемонстрировать влияние экспрессии генов, кодирующих полипептиды с активностью GalT на жизнеспособность клеток, рост клеток и продукцию антител, относительно клеток, не продуцирующих такие полипептиды. Варианты гликозилирования очищенных антител анализировали на масс-спектрометре MALDI/TOF-MS, как описано в Примере 1. Применяя эту методику возможно определить доли разных видов олигосахаридов в общей, исходной, популяции олигосахаридов, связанных с Fc-областью, а также возможно определить соответствие структур разным пикам спектра (Umana, P. et al., Nature Biotechnol. 17: 176- 180 (1999)).
Определяют спектр олигосахаридов неизмененного антитела. Конкретно, определяют, приводит ли модификация продуцирующих антитело клеток посредством экспрессии GalT дикого типа, к появлению, в основном, галактозилированных сложных двуантенных олигосахаридв с фукозилированным кором. Также определяют, приводит ли модификация продуцирующих антитело клеток посредством экспрессии нуклеиновой кислоты, кодирующей полипептид слияния GnTI-GalT, где каталитический домен GalT локализуется отвечающим за локализацию в комплексе Гольджи при помощи домена GnTI, к появлению, в основном, галактозилированных сложных двуантенных олигосахаридов, по сравнению с GalT дикого типа. Если синтезируются галактозилированные нефукозилированные и галактозилированные гибридные структуры, это может быть результатом конкуренции между GalT и другими гликозилтрансферазами или гликозидазами. Ожидается, олигосахарид, модифицированный в ходе катализируемой GalT реакции катализируемой GalT, добавлением остатка галактозы, не может быть далее модифицирован альфа-1,6-кор-ф фукозилтрансферазой, альфа-маннозидазой II (ManII) комплекса Гольджи и GnTII комплекса Гольджи.
Для того, чтобы оценить долю галактозилированных нефукозилированных и галактозилированных гибридных олигосахаридов, прикрепленных в Fc-области, применяют расщепление эндогликозидазой Н, которая «различает» гибридные и сложные олигосахариды.
Проводят тесты с целью определить, приводит ли модификация продуцирующих антитело клеток посредством экспрессии нуклеиновой кислоты, кодирующей полипептид слияния ManII-GalT, где каталитический домен GalT локализуется отвечающим за локализацию в комплексе Гольджи при помощи домена ManII, к появлению в основном галактозилированных нефукозилированных и галактозилированных гибридных олигосахаоридов. В частности, определяют является ли слитый ManII-GalT более эффективным, по сравнению с GalT дикого типа и гибридом GnTI-GalT, в синтезе прикрепленных Fc-области галактозилированных нефукозилированных и галактозилированных гибридных олигосахаридов.
Как упоминалось выше, эндогликозидазу Н (EndoH) применяют для подтверждения соответствия галактозилированных нефукозилированных и галактозилированных гибридных структур различным пикам олигосахаридов, наблюдаемых в спектрах MALDI. Определяют процентное содержание нефукозилированных гибридных структур, фукозилированных гибридных структур и фукозилированных сложных олигосахаридных структур, фукозилированных гибридных и фукозилированных сложных олигосахаридных структур среди остатков галактозы олигосахарида, которые несут остаток галактоз.
Определяют влияние уровня экспрессии GalT, и в особенности домена локализации, используемого для направления каталитического домена GalT в комплекс Гольджи, на конкуренцию между рекомбинантным каталитическим доменом GalT и эндогенной кор-альфа-1,6-фукозилтрансферазой, ManII и GnTII за модифицированные GalT олигосахаридные субстраты. Определяют уровень антителозависимой клеточной цитотоксичности (АЗКЦ), являющейся результатом повышенной экспрессии в продуцирующей антитело клетке нуклеиновой кислоты, кодирующей полипептид с активностью GalT, локализуемый в комплексе Гольджи различными доменами локализации.
Также определяют, обладают ли гликомодифицированные антитела, продуцируемые путем экспрессии в продуцирующих антитело клетках нуклеиновой кислоты, кодирующей полипептид слияния с активностью GalT, повышенной активностью связывания с активирующим Fc-рецептором FcgammaRIIIA человека или с ингибиторным рецептором FcgammaRIIb.
Конструкции GalT могут подавить активность эндогенной коровой кор-альфа-1,6-фукозилтрансферазой, гликомодифицируя Fc-область антитела и, следовательно, повышая АЗКЦ.
Определяют спектр олигосахаридов анти-CD20 антитела, полученного в клетках ВНК, растущих в суспензии и модифицированных для коститутивной экспрессии на высоком уровне как рекомбинантного антитела, так и полипептида слияния с активностью GalT. Также определяют относительное процентное содержание олигосахаридов гликомодифицированного моноклонального антитела, продуцируемого клетками стабильной клеточной линии ВНК-1502-28-11.
ПРИМЕР 5 Материалы и методы
1. Конструирование экспрессионных векторов ManII и GnTII
Для конструирования вектора экспрессии маннозидазы II (SEQ ID NO: 17) кДНК субклонировали в плазмиду (вектор экспрессии) в направлении «по ходу транскрипции» от промотора MPSV и в направлении «против хода транскрипции» от синтетического сигнала полиаденилирования бета-глобина кролика. Для экспрессии GnTIII использовали плазмиду (вектор экспрессии) с кДНК GnTII человека субклонированной в направлении «по ходу транскрипции» от промотора/энхансера цитомегаловируса человека и в направлении «против хода транскрипции» от сигнала полиаденилирования бычьего гормона роста.
Комбинация векторов экспрессии с точкой инициации репликации OriP вируса Эпштейна-Барра.
Фрагмент ДНК с OriP субклонировали, как описано в Примере 1 в вектор экспрессии ManII, описанный выше. В результате получили вектор экспрессии ManII pCLF9. Фрагмент ДНК с OriP Субклонировали, как описано в Примере 1, в вектор экспрессии GnTII, описанный выше. В результате получали вектор экспрессии GnTII pGnTII.
2. Трансфекция клеток HEK293-ENBA
Клетки HEK293-ENBA в экспоненциальной фазе роста трансфецировали, как описано в Примере 1. Для получения неизмененного антитела «Cwt» клетки трансфецировали вектором экспрессии антитела (pETR1520). Для получения гликомодифицированного антитела «Cbrt» клетки подвергали котрансфекции двумя плазмидами: одной для экспрессии антитела(pETR1520) и другой - для экспрессии полипептида слияния GnTIII(pETR1519), в соотношении 4:1, соответственно. Для получения гликомодифицированного антитела «Cm» клетки подвергали котрансфекции тремя плазмидами: одной для экспрессии антитела(pETR1520) одной для экспрессии полипептида слияния GnTIII(pETR1519), и еще одной - для экспрессии маннозидазы II (pCLF9) в соотношении 3:1:1, соответственно. Для получения гликомодифицированного антитела «Cmg» клетки подвергали котрансфекции четырьмя плазмидами: одной для экспрессии aHTHTena(pETR1520), одной для экспрессии полипептида слияния GnTIII(pETR1519), одной для экспрессии маннозидазы II (pCLF9) и ещй одной - для экспрессии GnTII (pGnTII) в соотношении 4:0.33:0.33:0.33, соответственно.
3. Получение и очистка неизмененных и гликомодифицированных антител
Культуральную среду описанных выше трансфецированных клеток заменяли свежей культуральной средой приблизительно на 16-й после трансфекции час культивирования, свежую среду собирали после еще 120 часов культивирования. Собранный супернатант, центрифугировали в течение 5 мин при 1200 об/мин, после чего центрифугировали его второй раз (4 мин при 4000 об/мин). Супернатант хранили при 4°С.
Очистка антител
Моноклональные антитела очищали от культуральных супернатантов, применяя две последовательные стадии хроматографии: хроматографию с использованием белка А и катионообменную хроматографию, как описано в Примере 1. Для антител cwt7, cbrt5 и cm1 дополнительно проводили вытесняющую хроматографию на колонке Superdex 200 (Amersham Pharmacia),добавляли фосфатный буферный раствор после стадии катионообменной хроматографии и собирали пик мономерных антител.
4. Анализ олигосахаридов
Олигосахариды высвобождали из антител, находящихся в растворе, ферментативным путем: при помощи расщепления гликозидазой PNGaseF, как описано в примере 1. Применение расщепления гликозидазой EndoH высвобожденных гликозидазой PNGaseF олигосахаридов для установления соответствия между гибридными гликозилированными структурами олигосахаридов м пиками нейтральных олигосахаридов на спектре MALDI/TOF-MS
Высвобожденные гликозидазой PNGaseF олигосахариды подвергают затем расщеплению эндогликозидазой Н (КФ 3.2.1.96), как описано в Примере 1.
MALDI/TOF-MS
Готовят пробы, содержащие продукты ферментативного расщепления высвобожденных олигосахаридов, и затем анализируют их на масс-спектрометре MALDI/TOF-MS, как описано в Примере 1.
5. Приготовление и выделение клеток
Мононуклеары периферической крови (МНПК) готовят, используя Histopaque-1077 (Sigma Diagnosticslnc, Сент-Луис, М063178 США), неукоснительно следуя инструкциям производителя и протоколу, описанному в Примере 1.
NK-клетки человека выделяют из МНПК, применяя процедуру негативного отбора, как описано в Примере 1.
35
7. Анализ АЗКЦ
МНПК, в качестве эффекторных клеток, готовят, как описано выше и анализируют их способность опосредовать цитотоксичность в Анализе антителозависимой клеточной цитотоксичности(АЗКЦ), как описано в Примере 1.
8. Связывание FcgammaRIIIA на NK-клетках
Связвание FcgammaRIIIA на свежевыделенных NK-клетках и связывание FcgammaRIIb на клетках лимфомы линии Raji определяют, как описано в Примере 1.
9. Анализ комплементзависимой цитотоксичности
Анализы комплемент зависимой цитотоксичности проводили, используя разведения антител, согласно способам, описанными в Примере 1, с последующей модификацией источника комплемента человека. Вкратце, нормальную сыворотку человека (НЧС) готовили из крови здоровых добровольцев. Кровь оставляли на 1 час для свертывания, затем центрифугировали при 1200g в течение 20 мин. Бесклеточный супернатант аликвотировали и хранили при -80°С. НЧС использовали в отношении 20% от конечного объема анализа.
Результаты и обсуждение
Гликомодифицированные варианты анти-CD20 химерных антител класса IgG1 (химерное антитело С2В8 также известное как rituximab) продуцировали культуры клеток млекопитающих, подвергнутые котрансфекции векторами экспрессии генов антитела и векторами экспрессии генов, кодирующих полипептиды с активностью GnTIII и маннозидазы II. Дополнительный гликомодифицированный вариант антитела был получен в культуре клеток млекопитающих, подвергнутых котрансфекции векторами экспрессии генов антитела и векторами экспрессии генов, кодирующих полипептиды с активностью GnTIII, маннозидазы II и GnTII. Для получения гликомодифицированного антитела «Cbrt» клетки подвергали котрансфекции двумя плазмидами: одной для экспрессии антитела(pETR1520) и другой - для экспрессии полипептида GnTIII(pETR1519). Для получения гликомодифицированного антитела «Cm» клетки подвергали котрансфекции тремя плазмидами: одной для экспрессии антитела(pETR1520) одной для экспрессии полипептида GnTIII(pETR1519), и еще одной - для экспрессии маннозидазы II (pCLF9). Для получения гликомодифицированного антитела «Cmg» клетки подвергали котрансфекции четырьмя плазмидами: одной для экспрессии антитела(pETRT520), одной для экспрессии полипептида GnTIII(pETR1519), одной - для экспрессии маннозидазы II (pCLF9) и еще одной - для экспрессии GnTII (pGnTII). Неизмененный (негликомодифицированный) вариант того же антитела («Cwt») был получен посредством трансфекции клеток млекопитающих исключительно вектором для экспрессии гена антитела. Трансфецированные клетки культивировали в течение 5 дней, затем секретированные рекомбинантные антитела очищали от культуральной среды. Экспрессия генов, кодирующих полипептиды с активностью GnTIII и ManII, не оказывала значительного влияния на жизнеспособность клеток, рост клеток или продукцию антитела по сравнению с клетками, не продуцирующими такие полипептиды с активностью гликозилтрансферазы или гликозидазы.
Объектом анализа являлся рисунок гликозилирования очищенных антител. Эти антитела несут N-связанные олигосахариды, прикрепленные исключительно к остатку Asn297 Fc-области IgG1 человека. Олигосахариды отделяли от антител ферментативным путем (при помощи расщепления гликозидазой PNGase) и исследовали затем на масс-спектрометре MALDI/TOF-MS. Применяя эту методику, зможно определить различные фракции олигосахаридов в общей, исходной, популяции олигосахаридов, связанных с Fc-областью, а также установить соответствие структур различным пикам спектра (Umana, P. et al., Nature Biotechnol. 17: 176-180 (1999)).
На Фиг. 26 показан полученный на масс-спектрометре MALDI/TOF-MS спектр нейтральных олигосахаридов неизмененного рекомбинантного анти-CD20 химерного антитела класса IgG1 С2В8 Cwt. Что касается неизмененного антитела, описанного ранее в Примере 1, Фиг. 5, Cwt обладал типичным спектром олигосахаридов с пиками в m/z 1485, 1648 и 1810 и состоял из двуантенных кор-фукозилированных сложных олигосахаридов с количеством остатков галактозы, равным 0, 1 и 2 соответственно. Модификация продуцирующих антитело клеток посредством экспрессии нуклеиновой кислоты, кодирующей полипептид слияния ManII-GnTIII, в котором каталитический домен локализуется в комплексе Гольджи при помощи домена локализации ManII, привела к продуцированию антитела (Cbrt), в котором большинство связанных с Fc-областью олигосахаридов были разветвленными нефукозилированными гибридными (см. Фиг 27). Как описано в Примере 1, эндогликозидазу Н (EndoH) применяли для подтверждения соответствия галактозилированных нефукозилированных и галактозилированных гибридных структур различным пикам олигосахаридов, наблюдаемых в спектрах MALDI.
Модификация продуцирующих антитело клеток посредством ко-экспрессии нуклеиновой кислоты, кодирующей полипептид слияния ManII-GnTIII, в котором каталитический домен локализуется в комплексе Гольджи при помощи домена локализации ManII, и нуклеиновой кислотой, кодирующей ManII, приводила к продуцированию антитела (Cm), в котором большинство связанных с Fc-областью олигосахаридов были разветвленными нефукозилированными сложными (см. Фиг 28). Модификация продуцирующих антитело клеток посредством ко-экспрессии нуклеиновой кислоты, кодирующей полипептид слияния ManII-GnTIII, в котором каталитический домен локализуется в комплексе Гольджи при помощи домена локализации ManII, а также нуклеиновой кислоты, кодирующей ManII и нуклеиновой кислоты, кодирующей GnTII, приводила к продуцированию антитела (Cmg), в котором большинство связанных с Fc-областью олигосахаридов являются разветвленными нефукозилированными сложными, причем доля разветвленных нефукозилированных сложных олигосахаридов Fc-области была даже выше, чем у антитела cm.
На Фиг 29 показаны данные, демонстрирующие повышенную антителозависимую клеточную цитотоксичность (АЗКЦ), являющуюся результатом экспрессии в продуцирующих антитело клетках нуклеиновой кислоты, кодирующей полипептид слияния ManII-GnTIII, в котором каталитический домен локализуется в комплексе Гольджи при помощи домена локализации ManII, где кодирующая ManII-GnTIII нуклеиновая кислота экспрессируется либо сама по себе (антитело Cbrt), либо коэкспрессируется одновременно с нуклеиновой кислотой, кодирующей ManII (антитело Cm). Таким образом, повышение уровня разветвленных нефукозилированных олигосахаридов гибридного либо сложного типа в Fc-области гликомодифицированных антител приводит к повышенной активности АЗКЦ.
Известно, что естественные киллеры (NK-клетки) являются важными участниками АЗКЦ. На поверхности этих клеток находится активирующий рецептор Fcgamma IIIA также известный, как CD16a. Связывание Fc-области связанных с клетками-мишенями антител с рецепторами FcgammaRIIIA на поверхности NK-клеток необходимо для перекрестного связывания этих рецепторов на NK-клетках и последующей индукции АЗКЦ. Следовательно, важно оценить связывание антител, полученных описанными здесь способами с Fc-рецепторами, особенно - с рецепторами в нативной форме, т.е. в форме, в которой они существуют на поверхности эффекторных клеток иммунной системы человека.. Фиг. 30 демонстрирует, что гликомодифицированные антитела, полученные путем экспрессии в продуцирующих антителоа клетках нуклеиновой кислоты, кодирующей полипептид слияния с активностью GnTIII, где кодирующая GnTIII нуклеиновая кислота экспрессируется либо сама по себе (антитело Cbrt), либо коэкспрессируется в продуцирующей антитело клетке одновременно с нуклеиновой кислотой, кодирующей ManII (антитело Cm),обладают повышенной аффинностью связывания с активирующим Fc-рецептором человека FcgammaRIIIA. Как упоминалось выше для случаев анализа АЗКЦ, у этих антител повышен уровень разветвленных нефукозилированных олигосахаридов, что является результатом экспрессии полипептида слияния с активностью GnTIII в продуцирующих антитела клетках.
Следовательно, повышение уровня разветвленных нефукозилированных олигосахаридов гибридного либо сложного типа в Fc-области т гликомодифицированных антител приводит к повышенной активности АЗКЦ. NK-клетки, используемые в этом анализе были получены от донора с генотипом, не допускающим экспрессию рецептора FcgammaRIIc в NK-клетках донора (Metes, D. et al. J. Immunol. Methods 255(1-2): 85-95 (2001)). Таким образом, единственным Fc-рецептором на поверхности таких клеток является активирующий рецептор FcgammaRIIIA
Связывающий домен активирующего рецептора FcgammaRIIIB почти полностью идентичен связывающему домену FcgammaRIIIA. Следовательно, приведенные выше данные также указывают на то, что описанные здесь гликомодифицированные антитела могут вызывать повышение эффекторных функций, опосредуемых эффекторными клетками, которые имеют на своей поверхности FcgarnmaRIIIB, такими как полиморфноядерные клетки, включая высвобождение токсических продуктов и фагоцитоз ((Reff, М. Е., Heard, С.Crit Rev Oncol Henaatol. 40(1): 25-35 (2001), Daeron, FM. Annu. Rev. Immunol. 15: 203-34 (1997), Ravetch, J.V., Bolland S. Annu. Rev. Immunol. 19: 275-90 (2001)).
Cbrt, анти-CD20 антитело, продуцируемое клетками, модифицированными для экспрессии нуклеиновой кислоты, кодирующей полипептид слияния с активностью GnTIII, который локализуется в комплексе Гольджи при помощи домена локализации ManII, исследовали также на предмет комплементопосредованного лизиса(Зд.-КОЛ) - другой эффекторной функции, которая не связана с Fc-рецепторами на эффекторных клетках иммунной системы. Подавляющее большинство олигосахаридов этого гликомодифицированного антитела принадлежат к типу разветвленных гибридных нефукозилированных олигосахаридов. У антител Cbrt наблюдали сниженную, по сравнению с неизмененными антителами Cwt, активность комплементопосредованного лизиса (Фиг. 31). Для некоторых приложений может быть желательным иметь антитела с повышенной АЗКЦ, но пониженным комплемент-опосредованным лизисом, например, уменьшения побочных эффектов, таких как васкулит кровеносных сосудов в сайтах локализации опухолей, опосредованный КОЛ. Другой КОЛ-опосредованный побочный эффект наблюдали в ходе лечения анти-CD20 антителом (van der Kolk L.E. et al. Br J Haematol. 115 (4): 807-11 (2001)). Однако, возможно получать гликомодифицированные антитела с повышенными активностью АЗКЦ и связыванием FcgammaRTII без значительного снижения активности КОЛ по сравнению с неизмененными антителами, как в случае с антителами Cm (Фиг. 31). Такие антитела желательно иметь для приложений, где максимальная элиминация клеток-мишеней требует как высокой АЗКЦ, так и высокой активации комплемента и активности КОЛ. Приведенные выше спектры олигосахаридов показывают, что возможно модифицировать клетки для получения антител, в которых большинство присоединенных к Fc-области олигосахаридов принадлежат к типу разветвленных нефукозилированных сложных вместо разветвленных нефукозилированных гибридных. Этого можно достичь путем коэкспрессии полипептида слияния GnTIII вместе с нуклеиновой кислотой, кодирующей ManII антитело Cm) или вместе с нуклеиновой кислотой, кодирующей Man II и нуклеиновой кислотой, кодирующей GnTII (антитело Cmg).
Гликомодифицированные антитела обладают повышенной активностью АЗКЦ и повышенной активностью связывания FcgammaRTII, которая ассоциирована с повышенными уровнями разветвленных нефукозилированных олигосахаридов, в то время как их активность КОЛ повышалась при увеличении доли сложных олигосахаридов относительно доли гибридных олигосахаридов.
В этом и предыдущих примерах описана экспрессия кодирующих полипептиды слияния нуклеиновых кислот, где полипептиды слияния локализуются в аппарате Гольджи и имеют каталитический домен, который конкурирует с эндогенными фукозилтрансферазами за олигосахаридные акцепторы, ранее модифицированные в ходе реакций, катализируемых GnTI. Рекомбинантные гликопротеины, продуцируемые модифицированными таким образом клетками, обладают повышенными уровнем нефукозилированных олигосахаридов. Этот пример демонстрирует, что ко-экспрессия в таких клетках-хозяевах нуклеиновых кислот, кодирующих ManII и/или GnTII одновременно с описанными выше нуклеиновыми кислотами, кодирующими слитые полипептиды, приводит к увеличению биосинтеза сложных олигосахаридов за счет гибридных олигосахаридов и, следовательно, увеличению синтеза гликопротеинов с повышенным, относительно гликопротеинов, продуцируемых в клетках, которые не коэкспрессируют нуклеиновые кислоты, кодирующие ManII и/или GnTII, уровнем нефукозилированных сложных олигосахаридов.
ПРИМЕР 6
Повышенная экспрессия альфа-маннозидазы
Молекулярное клонирование
Альфа-маннозидаза II человека
Ген, кодирующий альфа-маннозтдазу II человека ("hManII")(КФ 3.2.1.114, SEQ ID NO:17) под контролем промотора MPSV, клонировали в векторе экспрессии, содержащем элемент oriP. Полученный в результате вектор экспрессии pCLF9 показан на Фиг. 32А. Векторы экспрессии, кодирующие легкую и тяжелую цепи анти-CD20 моноклонального антитела представляли собой, соответственно, векторы pETR1842 и pETR1843 (Фиг. 32В и Фиг. 32С).
Слитый белок ManII-GalT
Слитый белок (SEQ ID NO: 20), состоящий из области CTS (цитоплазматическая + транспордная + «стержневая» области) маннозидазы 11 ч и каталитического домена бета-1,4-галактозилтрансферазы (М22921, аминокислоты 126-397) был сконструирован, как описано ниже. Область CTS маннозидазы Пч амплифицировали методом ПЦР из вектора pETR1484 (праймеры CF33, GAB252). Каталитический домен GalT (аминокислоты 126-397), используя праймеры CF31 и CF35, из вектора pBlueGalT. Область CTS маннозидазы IIч комбинировали с каталитическим доменом GalT, в результате чего получили белок слияния, контролируемый промотором MPSV(pCLF24). Полный ген GalT был получен из pBlueGalT. Кодирующий GalT ген был секвенирован(8Е0 ID NO: 16):
MRLREPLLSGSAAMPGASLQRACRLLVAVCALHLGVTLVYYLAGRDLSR
LPQLVGVSTPLQGGSNSAAAIGQSSGELRTGGARPPPPLGASSQPRPGGDS
SPWDSGPGPASNLTSVPVPHTTALSLPACPEESPLLVGPMLIEFNMPVDL
ELVAKQNPNVKMGGRYAPRDCVSPHKVAIIIPFRNRQEHLKYWLYYLHP
VLQRQQLDYGIYVrNQAGDTIFNRAKLLNVGFQEALKDYDYTCFVFSDV
DLIPMNDHNAYRCFSQPRHISVAMDKFGFSLPYVQYFGGVSALSKQQFLT
INGFPNNWGWGGEDDDIFNRLWRGMSISRPNAWGRCRMIRHSRDKK
NEPNPQRFDRIAHTKETMLSDGLNSLTYQVLDVQRYPLYTQITVDIGTPS
5' - часть GalT амплифицировали с праймерами CF31/CF38 и добавляли сайт рестрикции FseI перед геном. Секвенирование показало правильность последовательности; после чего последовательность была переведена в вектор pCLF24 путем расщепления рестриктазами FseI/DraIII, что дало вектор pCLF26. После добавления в векторы pCLF24 и pCLF26 OriP получили, соответственно векторы pCLF25 и pCLF27 (Фиг. 33А и Фиг. 33В).
Экспрессия альфа-маннозидазы и ManII-GalT в клетках HEK293-ENBA.
Клетки HEK293-ENBA трансфецировали кальций-фосфатным способом. Вкратце, для трансфекции Т150 за 24 часа до трансфекции 15 миллионов клеток высевали в 28 мл среды DMEM, содержащей 10% фетальной сыворотки и 250 мкг/мл неомицина, и инкубировали при 37°С в атмосфере с 5% СО2 в течение ночи.
Для каждого подлежащего трансфекции флакона Т150 готовили раствор ДНК, CaCl2 и воды: смешивали 94 мкг тотальной ДНК плазмидного вектора, 469 мкл 1М раствора CaCl2 и доводили объем смеси до 938 мкл путем добавления воды. К этому раствору добавляли 938 мкл раствора, содержащего 50 мМ HEPES, 280 мМ NaCl и 1.5 мМ Na2HPO4,, с рН 7.05 и немедленно перемешивали в течение 10 сек., после чего отставляли стоять при комнатной температуре в течение 20 сек. Суспензию разбавляли 24 мл среды DMEM, дополненной 2% фетальной сыворотки, и добавляли в Т150 вместо существующей среды. Клетки инкубировали при 37°С, 5% СО2 в течение приблизительно 17-20 часов, затем среду заменяли на 30 мл DMEM, содержащей 10% фетальной сыворотки.
Для исследование влияния альфа-маннозидазы II на конкуренцию кор-фукозилтрансферазы клетки трансфецировали векторами pETR1842, pETR1843 и pCLF9 в отношении 2:1:1 соответственно. Для экспрессии белка слияния ManII-GalT клетки трансфецировали векторами pETR1842, pETR1843 и pCLF25 в отношении 2:1:1 соответственно. На 5 день после трансфекции собирали супернатант, центрифугировали его в течение 5 мин при 1200 об/мин, после чего центрифугировали его второй раз (4 мин при 4000 об/мин). Супернатант фильтровали и хранили при 4°С.
Очистка моноклонального анти-CD20 антитела
Моноклональное антитело очищают от 30 мл супернатанта путем двух стадий хроматографии: хроматографии с использованием белка А для того, чтобы отделить маонклональное антитела от присутствующего в сыворотке антитела коровы, и следующей за ней катионообменной хроматографией для замены буфера пробы на ФБР.
Анализ олигосахаридов
Расщепление гликозидазой PNGaseF
Пробу моноклонального антитела (50 мкг) инкубировали с N-гликозидазойF (рекомбинантная, Roche, Швейцария) в концентрации O.lmU/мкл. Расщепление проводили в 0.2 мМ Tris-буфере (рН 7.0) в течение 3 часов при 37°С. Затем высвобожденные олигосахариды инкубировали с 150 мМ уксусной кислотой в течение 3 часов при комнатной температуре. Затем концентрацию соли в образцах уменьшали, используя 0.6 мл катионообменной смолы (смола AG50W-X8, водородная форма, размер гранул 100-200, BioRad, Геркулес, Калифорния), заключенной в хроматографической колонке Micro Bio-Spin (BioRad, Геркулес, Калифорния).
Расщепление эндогликозидазой Н.
Перед обработкой уксусной кислотой, высвобожденные гликозидазой PNGaseF расщепляли эндогликозидазой Н (КФ 3.2.1.96 Roche, Швейцария), ферментом, расщепляющим между остатками N-ацетилглюкозамина хитобиозного кора N-связанных олигосахаридов. Этот фермент расщепит гликаны «олигоманнозного» типа и большую часть гликанов гибридного типа, в то время как олигосахариды сложного типа не гидролизуются.
Олигосахариды расщепляли 0.2 mU/мкл эндогликозидазы Н в 2 мМ Tris-буфере. Расщепление проводили в 0.2 мМ Tris-буфере (рН 7.0) в течение 3 часов при 37°С. Затем высвобожденные олигосахариды инкубировали с 150 мМ уксусной кислотой в течение 3 часов при комнатной температуре. Затем концентрацию соли в образцах уменьшали, используя 0.6 мл катионообменной смолы (смола AG50W-X8, водородная форма, размер гранул 100-200, BioRad, Геркулес, Калифорния), заключенной в хроматографической колонке Micro Bio-Spin (BioRad, Швейцария)
Приготовление матрикса и образцов.
Матрикс на основе 2,5-дигидроксибензойной кислоты (sDHB) готовили следующим образом: 2 мг 2,5-дигидроксибензойной кислоты и 0.1 мг 5-метоксисалициловой кислоты растворяли в 1 мл смеси этанола и 10 мМ водного хлодира натрия (в объемном отношении 1:1). 1 мкл пробы наносили на мишень из нержавеющей стали и смешивали с 1 мкл матрикса на основе 2,5-дигидроксибензойной кислоты. Образцы высушивали воздухом и наносили 0.2 мкл этанола.
Анализ MALDI/TOF
Для получения спектров масс использовали масс-спектрометр MALDI/TOF марки Voyager Elite (Perspective Biosystems) с задержкой экстракции. Положительные ионы ускоряли при 20кВ после задержки 57нс. Установка работала в рефлекторном режиме. Для получения конечного спектра совмещали пять спектров по 40 ударов (200 ударов). Для присваивания масс ионам применяли внешнюю калибровку стандартными олигосахаридами.
Спектр олигосахаридов
Спектр олигосахаридов анти-CD20 антитела, продуцируемого в присутствии ManII показан на Фиг. 34. Олигосахариды, связанные с Fc-областью антитела представляют собой сложные структуры, в 48% которых отсутствует коровая фукоза. Альфа-маннозидаза II конкурирует с кор-фукозилтрансферазой, порождая 48% нефукозилированных олигосахаридных структур. В отсутствие альфа-маннозидазы II олигосахариды Fc-области антитела представлены исключительно фукозилированными структурами (антитело дикого типа).
Спектр олигосахаридов анти-CD20 антитела, продуцируемого в присутствии полипептида слияния ManII-GalT показан на Фиг. 35А-В. В случае белка слияния ManII-GalT, также как и для альфа- меннозидазы II, возрастает количество нефукозилированных олигосахаридных структур. Среди нефукзилированных структур высоко процентное содержание структур с высоким содержанием маннозы (m/z 1256, 1419 и 1581). В дополнение к этим 67 процентам нефукозилированных Сахаров было обнаружено 30% гибридных нефукозилированных структур (m/z 1622). Следовательно, проба, продуцированная в присутствии белка слияния ManII-GalT демонстрирует почти 100% содержание нефукозилированных структур.
Биологическая активность антител, продуцируемых в присутствии альфа-маннозидазы II или белка слияния ManII-GalT
Для того, чтобы определить влияние действия альфа-маннозидазы II и фермента ManII-GalT в конкуренции с кор-фукозилтрансферазой были проведены соответствующие биологические анализы. Пробы тестировали на предмет in vitro антителозависимой клеточной цитотоксичности (АЗКЦ) и связывания рецептора CD 16, экспрессируемого на поверхность модифицированной клеточной линии СНО (СНО-1708-37).
Связывание IeG на СНО-1708-37
Клетки линии СНО-1708-37 экспрессировали на свою поверхность рецептор FcγRTIIA(CD16) и цепь γ рецептора FcγRI. Экспрессию рецептора FcγRIIIA(CD16) оценивали при помощи анализа FACS, используя моноклональное антитело 3G8-FITC (Фиг. 36). Клетки СНО-1708-37 инкубировали в концентрации 1 млн/мл в ФБР, содержащим 0.1% БСА, с разными вариантами антител в разных концентрациях (10, 3, 1, 0.3, 0.1 мкг/мл) в трипликатах. Клетки инкубировали в течение 30 минут при 4°С и затем промывали ФБР, содержащим 0.1% БСА. Связывание антитела определяли, инкубируя его в соотношении 1:200 с конъюгатом флуоресцин изотиоцианина и фрагментом IgG козы против фрагмента F(ab')2 человека (JacksonlmmunoResearch, Вест Гроув, Пенсильвания) в течение 30 минут при 4°С. Интенсивность флуоресценции, связанную со связанными вариантами антитела определяли на приборе FASCalibur(BD Bioscience, Сан-Хосе, Пенсильвания), пропускающим живые клетки.
В эксперимент с анализом связывания вошли следующие варианты антитела:
Cwt 8(дикий тип 1)
ManII(альфа-маннозидаза II)
продуцируемое в присутствии альфа-маннозидазы II антитело (ManII) связывается с рецептором FcγRIIIA с более высокой аффинностью, чем антитела дикого типа.
Антителозависимая клеточная цитотоксичность in vitro (ADCC)
Мононуклеары периферической крови (МНПК) готовили с использованием Histopaque-1077(Sigma Diagnostics Inc., Сент-Луис, M063178, США), строго следуя инструкциям производителя. Вкратце, у добровольцев, бегавших в течение 1 минуты в полную силу для увеличения процента естественных киллеров (NK-клеток), отбирали венозную кровь шприцами с гепарином. Кровь разбавляли в ФБР, не содержащего Са и Mg, в соотношении 1:0.75-1.3, и наслаивали на Histopaque-1077. Градиент центрифугировали при 400×g без перерывов в течение 30 минут при комнатной температуре (КТ). Жидкость, прилежащую к границе, Histopaque-1077 содержащую МНПК, собирали, промывали ФБР (50 мл на клетки с двух градиентов) и собирали посредством центрифугирования при 300 × g в течение 10 минут при КТ. После ресуспендирования осадка в ФБР, МНПК подсчитывали и промывали второй раз посредством центрифугирования при 200 × g в течение 10 минут при КТ. Затем клетки ресуспендировали в подходящей среде для последующих процедур.
Клетки-мишени (клетки линии Raji) промывали в ФБР, подсчитывали и ресуспендировали в концентрации 1 млн клеток на мл в среде DMEM, содержащей 10% фетальной сыворотки и 1% Glutamax. К этому добавляли кальцин в отношении 1:100, и затем клетки инкубировали в течение 30 мин при 37°С. Затем клетки промывали в ФБР, подсчитывали и ресуспендировали в концентрации 0.3 млн клеток на мл в среде AIM-V. По 100 мкл этих клеток вносили лунки круглодонного 96-луночного планшета. Антитела разбавляли в среде AIM-V, добавляли по 50 мкл к клеткам-мишеням и оставляли на 10 мин при комнатной температуре для связывания. Эффекторные клетки МНПК готовили, как описано выше. Отношение эффекторных клеток к клетками-мишеням составляло 25:1. Эффекторные клетки готовили в концентрации 15 млн. клеток на мл в среде АГМ-V и вносили по 50 мкл в лунки. Планшеты инкубировали в течение 4 часов при 37°С во влажной атмосфере, содержащей 5% СО2. Клетки дважды промывали в ФБР и добавляли по 200 мкл боратного раствора. Лизис клеток-мишеней оценивали, измеряя кальцин, высвобожденный в среду после лизиса боратным раствором (Фиг. 37).
Спонтанное высвобождение измеряли в лунках, содержащих только клетки-мишени и эффекторные клетки(без антител). Максимальное высвобождение определяли по лункам, содержащим только клетки-мишени и 1% Triton Х-100. Процент специфического антиген-опосредованного лизиса вычисляли следующим образом: ((x-SR)/MR-SR)*100, где х обозначает среднее Vmax для определенной концентрации антител, SR - среднее Vmax спонтанного высвобождения, MR - среднее Vmax максимального высвобождения.
ПРИМЕР 7
Применение гликомодифицированных анти-РЭФР моноклональных антител в лечении псориаза.
Пациентов (людей), больных псориазом, возможно лечить гликомодифицированными анти-РЭФР моноклональными антителами, полученными согласно способам данного изобретения. В частности, пациент еженедельно получает инфузионным путем ударную дозу (400 мг/м2) гликомодифицированных антител. Поддерживающие дозы по 250 мг/м2 назначают еженедельно до достижения полного ответа.
ПРИМЕР 8
Применение гликомодифицированного анти-ErbB2 моноклонального антитела в лечении метастатического рака предстательной железы, метастатического рака груди, метастатического рака прямой и ободочной кишки, немелкоклеточного рака легкого (стадий IIIb или IV).
RhuMAb2C4 представляет собой полноразмерное гуманизированное моноклональное антитело (продуцируемое клетками СНО), направленно к ErbB2. RhuMAb2C4 блокирует связывание ErbB2 с другими членами семейства ErbB, ингибируя тем самым внутриклеточную передачу сигналов по пути, связанным с ErbB. RhuMAb2C4 не только ингибирует рост опухолей, в которых повышена экспрессия ErbB2, но также блокирует рост опухолей, которым необходима ErbB2-лигандзависимая передача сигнала.
Гликомодифицированную форму RhuMAb2C4, созданная согласно способам настоящего изобретения, можно применять для монотерпии пациентов с гормон-рефрактерным (андроген-независимым) раком предстательной железы, пациентов с метастатическим раком груди, пациентов с метастатическим раком прямой и ободочной кишки, пациентов с немелкоклеточным раком легкого в стадии IIIb или IV. В частности, гликомодифицированное RhuMAb2C4 назначают для внутривенного введения (i.v.) еженедельно или один раз в каждые три недели в дозе 2 или 4 мг/кг, соответственно, до прекращения прогрессирования заболевания. Антитело поставляется в виде мультидозовой жидкой лекарственной формы (дозы по 20 мл с концентрацией антитела 20 мг/мл или выше).
Должно быть очевидно, что данное изобретение может быть осуществлено способами, отличными от подробно описанных в вышеизложенном описании и примерах. В свете вышеизложенных идей возможны многочисленные модификации и варианты настоящего изобретения, которые, следовательно, находятся в рамках прилагаемой формулы.
Полное раскрытие всех цитируемых здесь публикаций (включая патенты, приложения к патентам, журнальные статьи, лабораторные руководства, книги и другие документы) включено в описание изобретения путем ссылки.
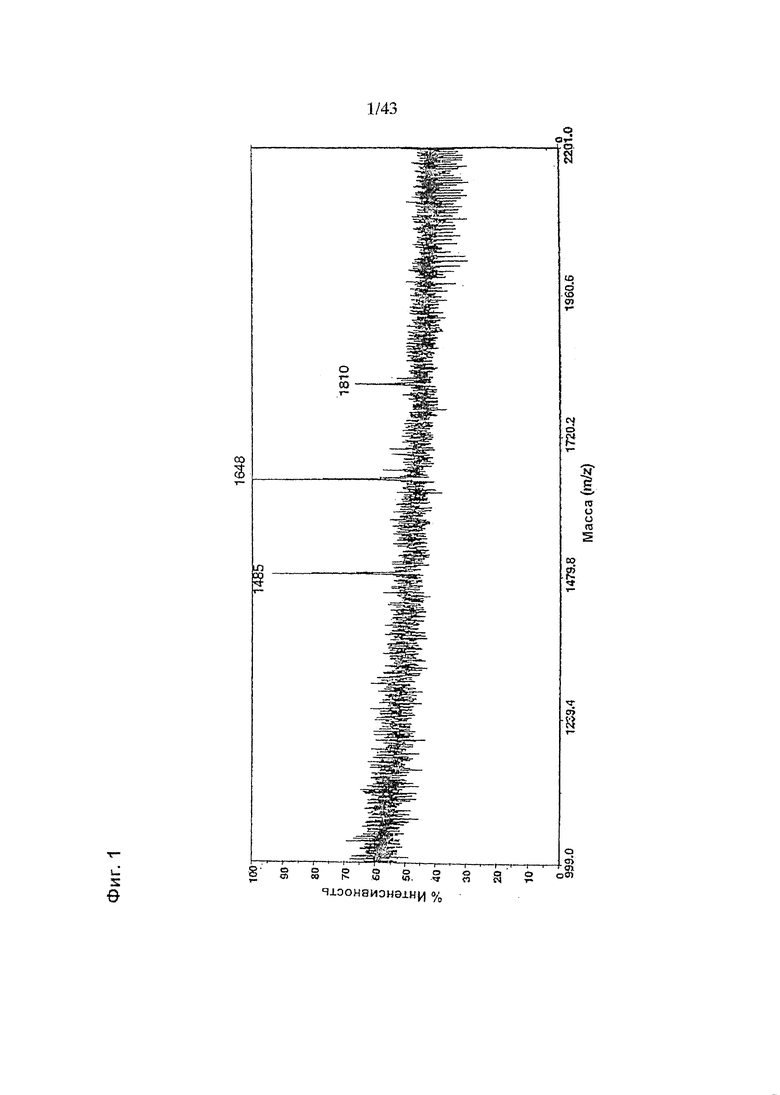
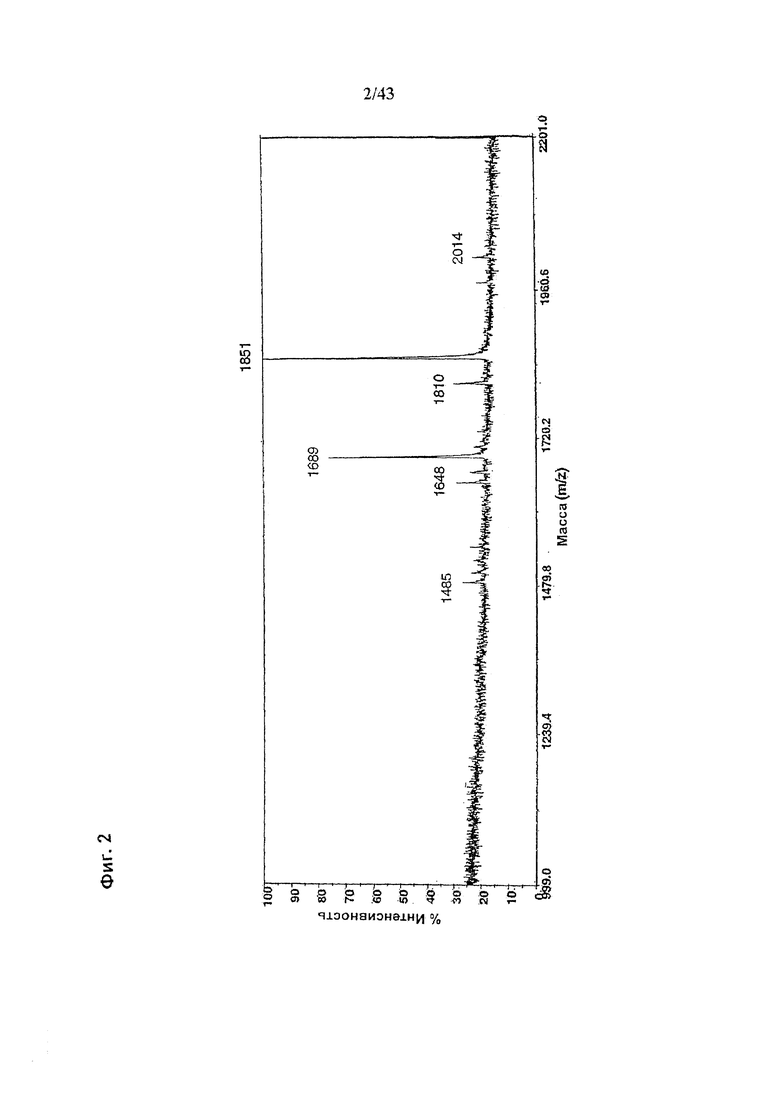
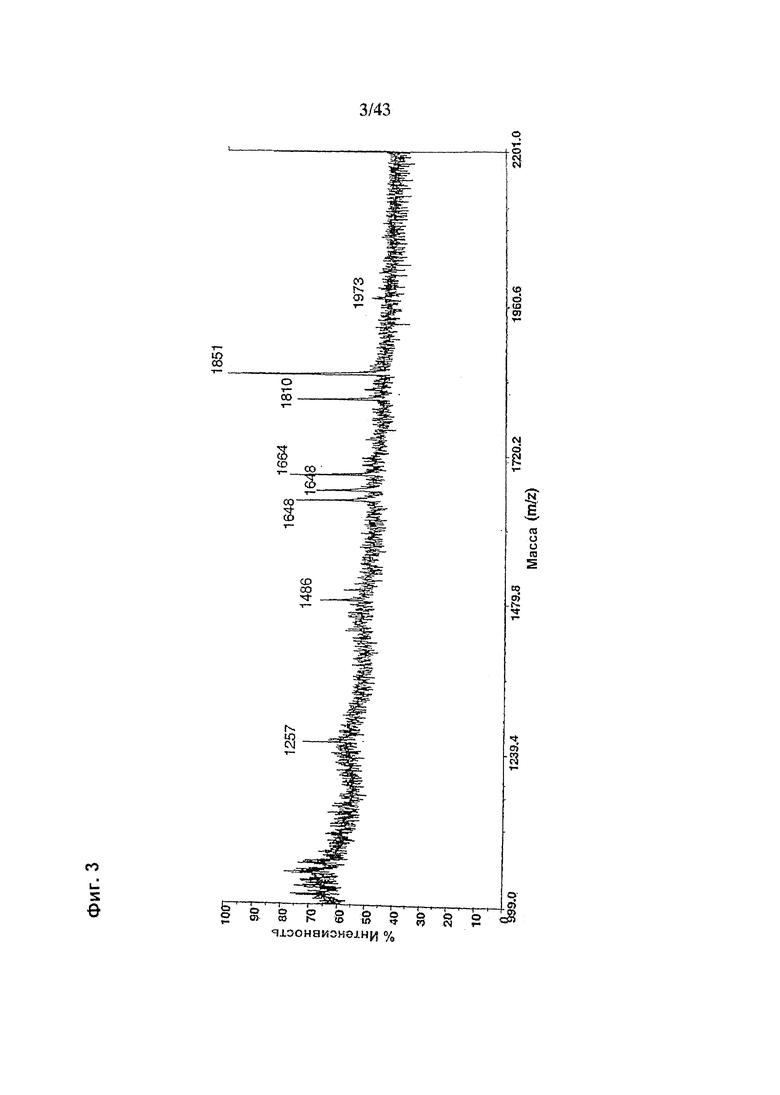
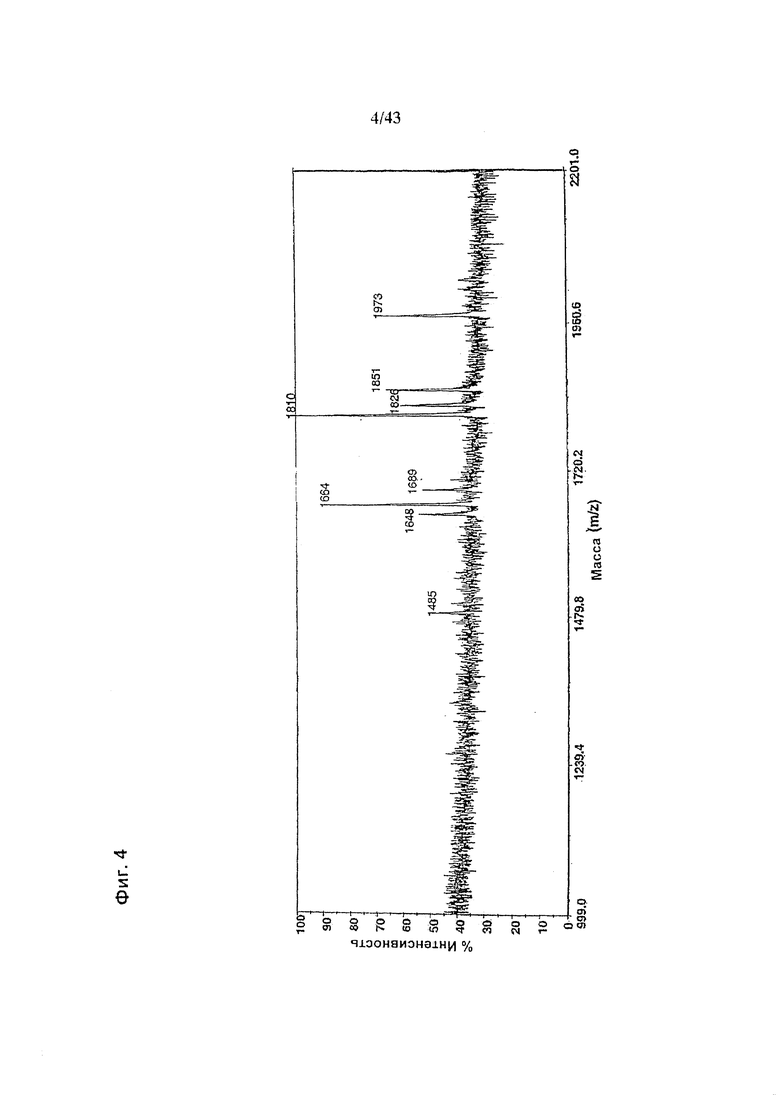
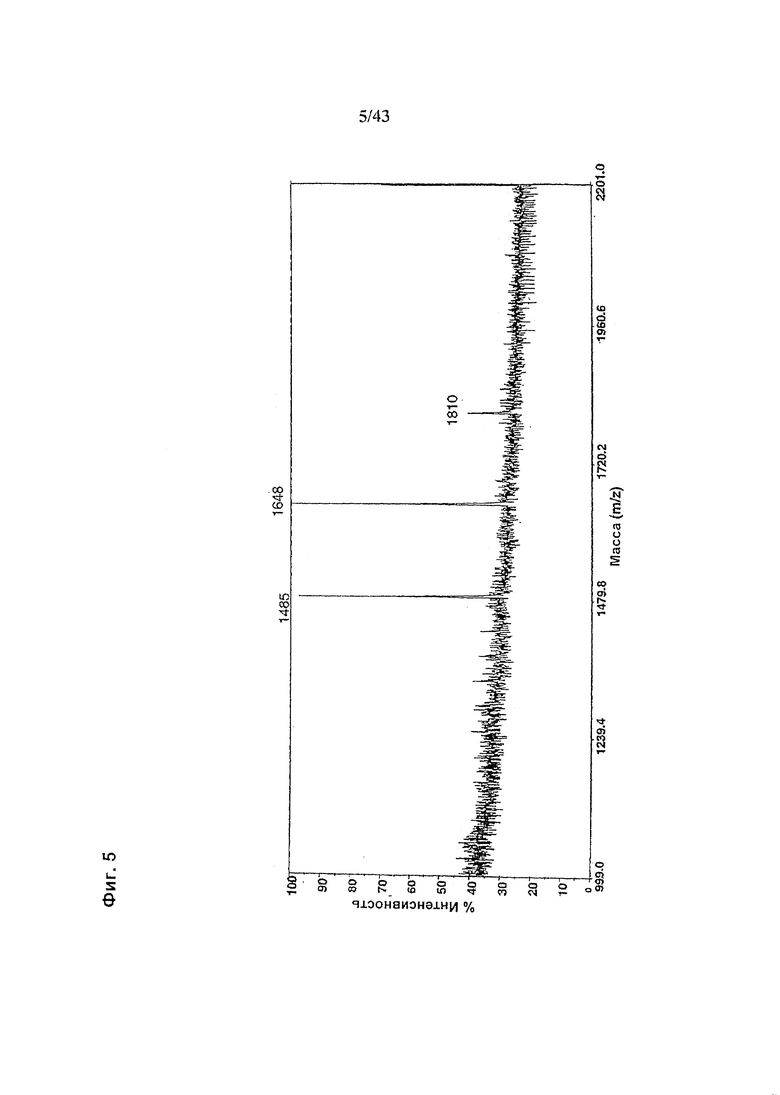
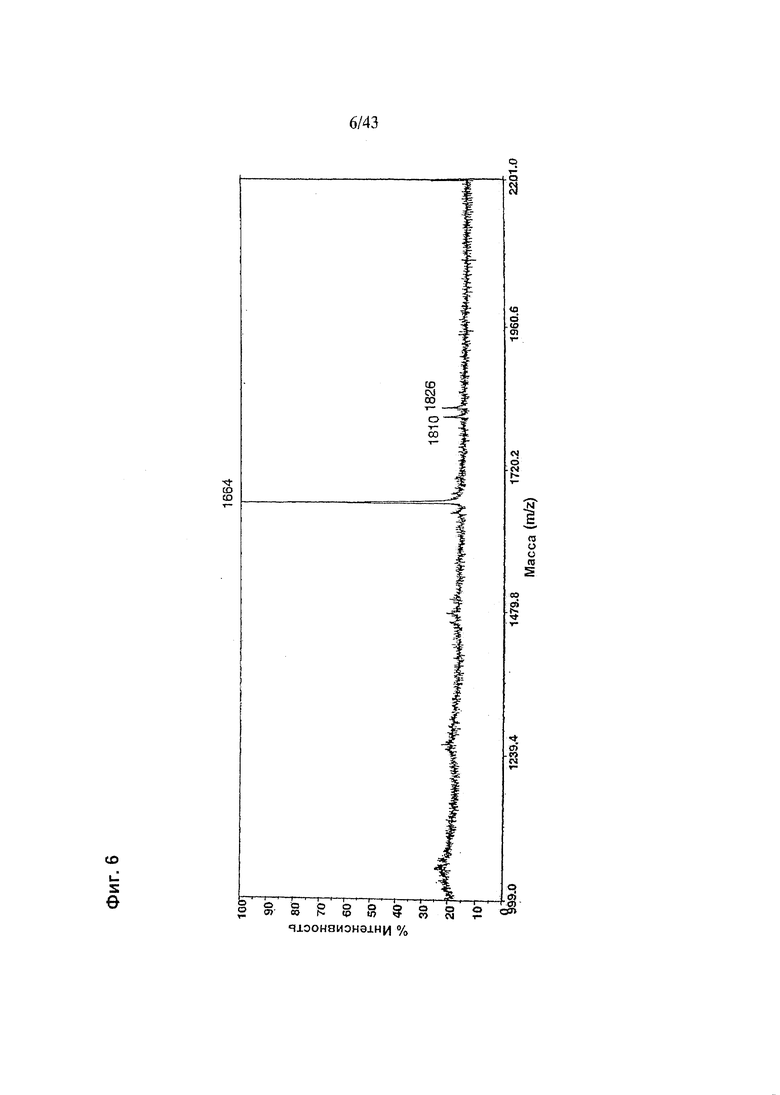
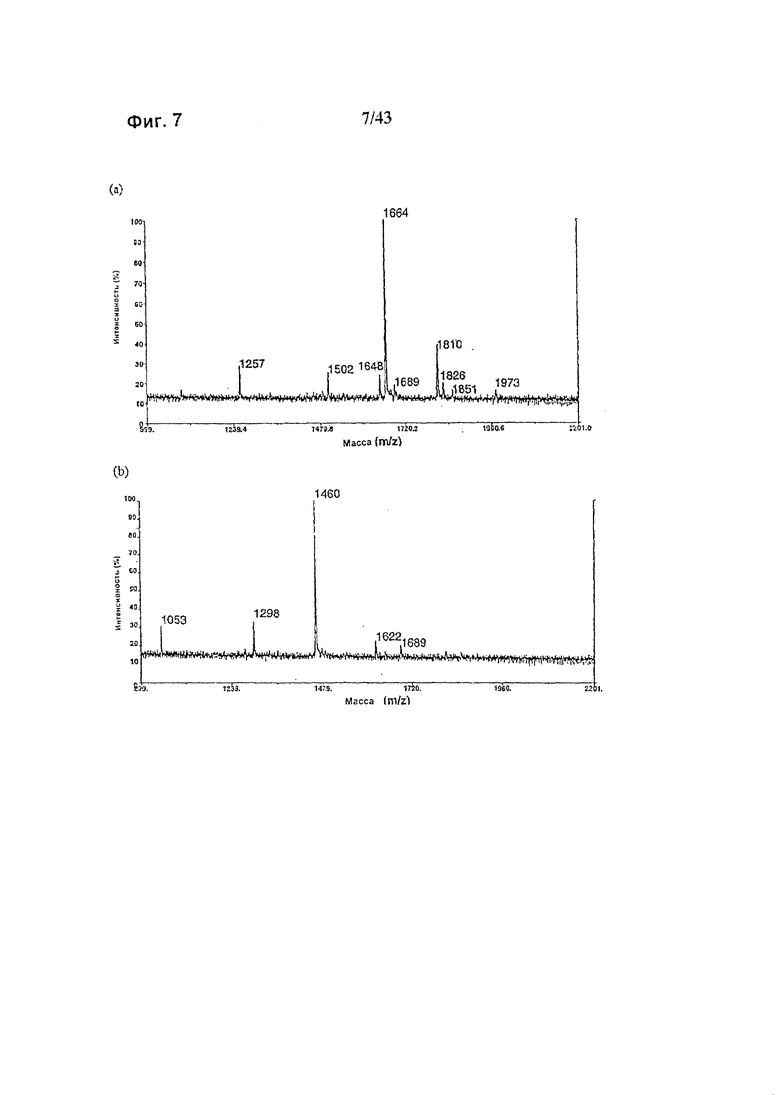

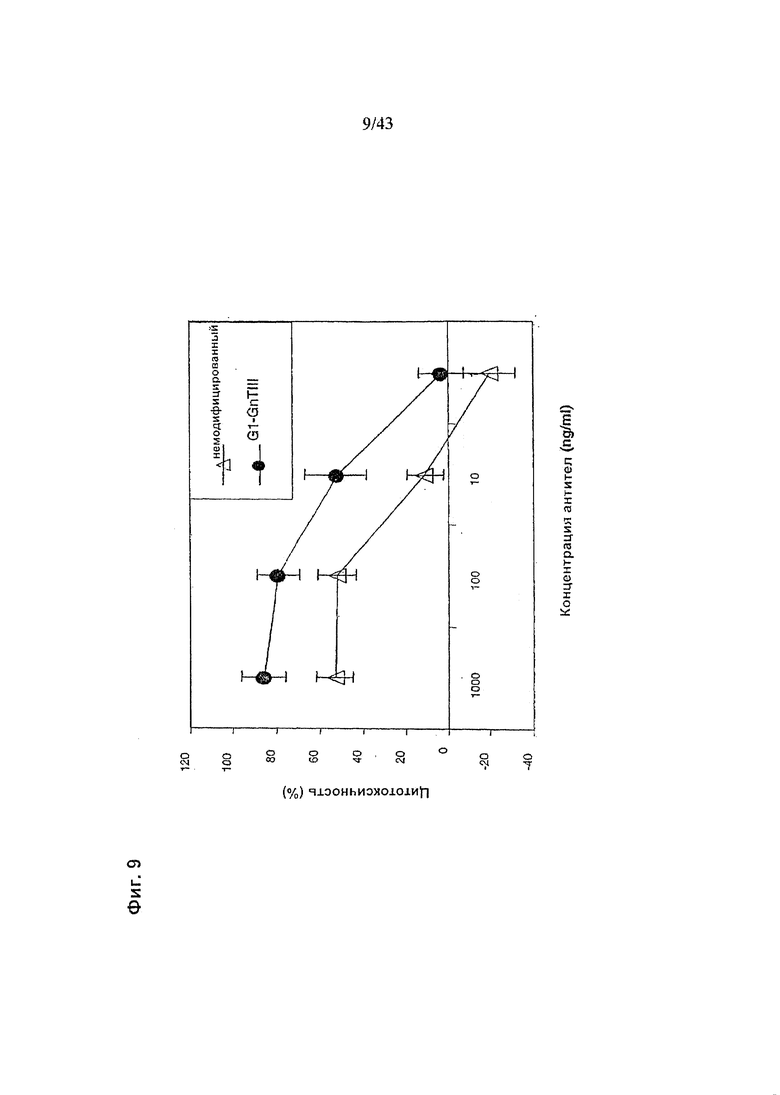
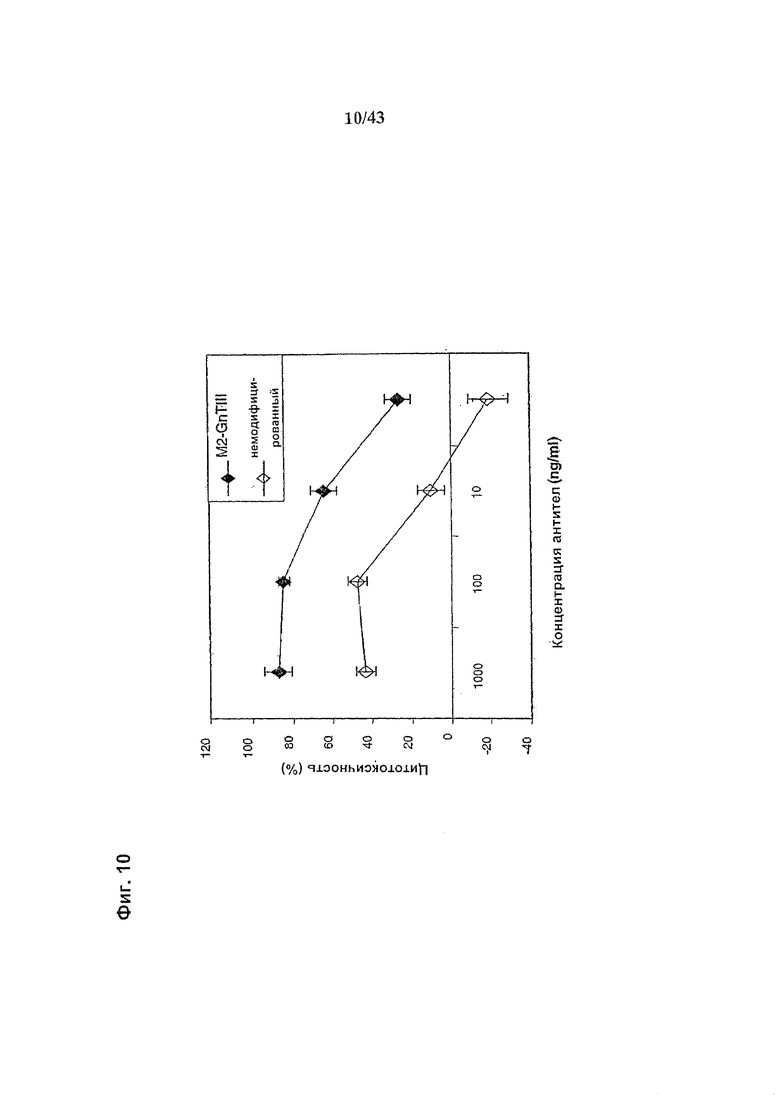
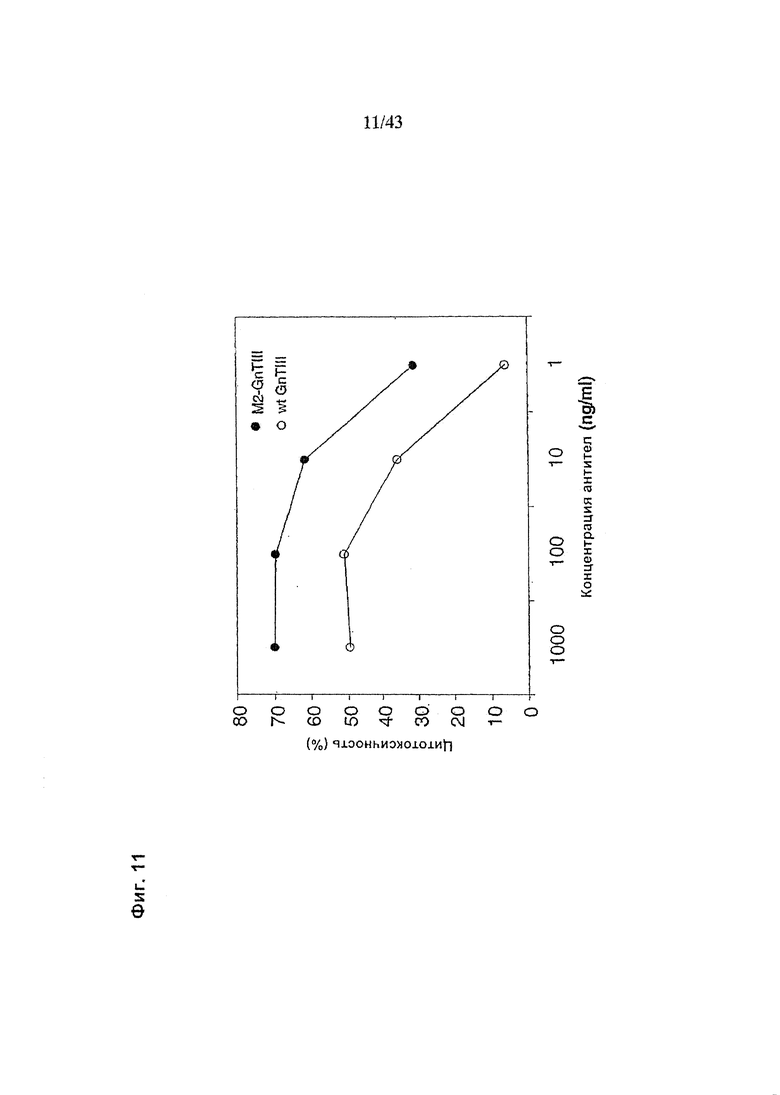
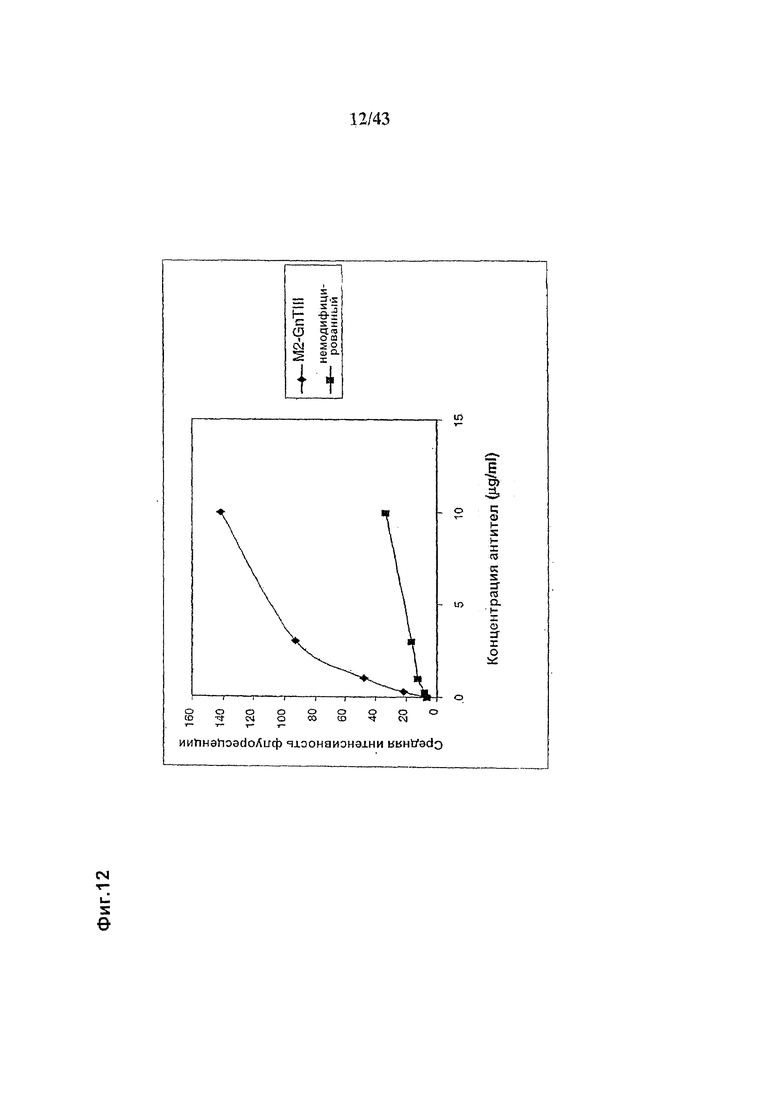
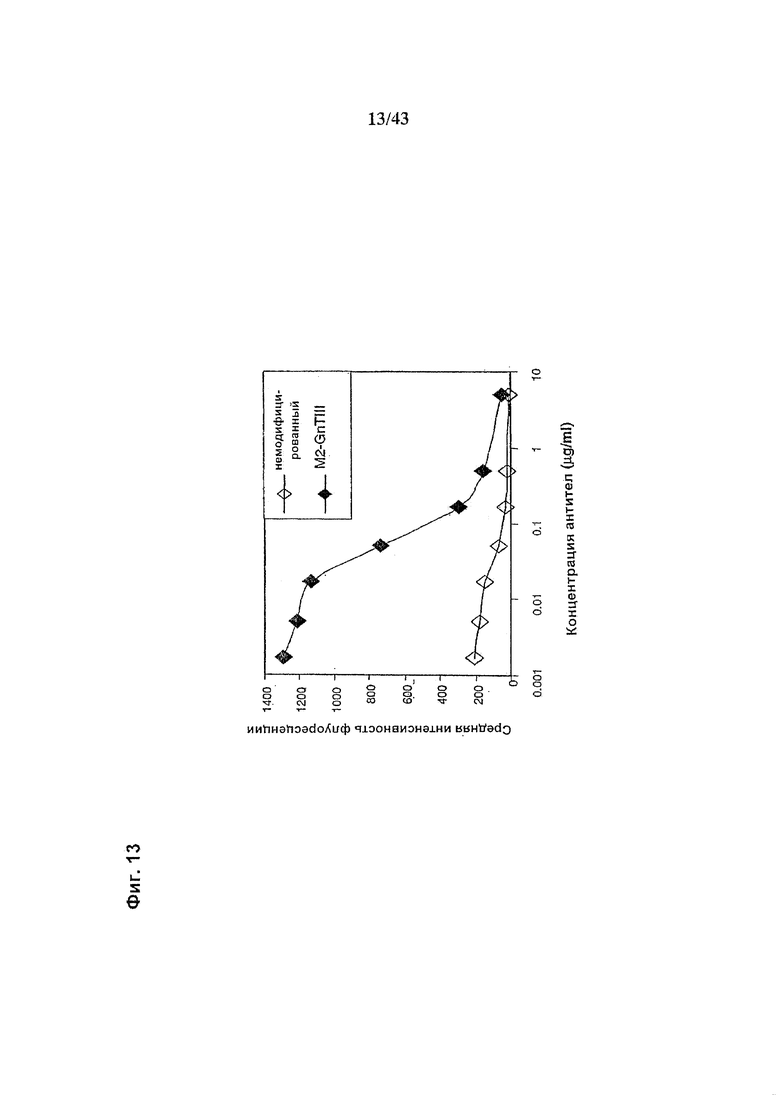
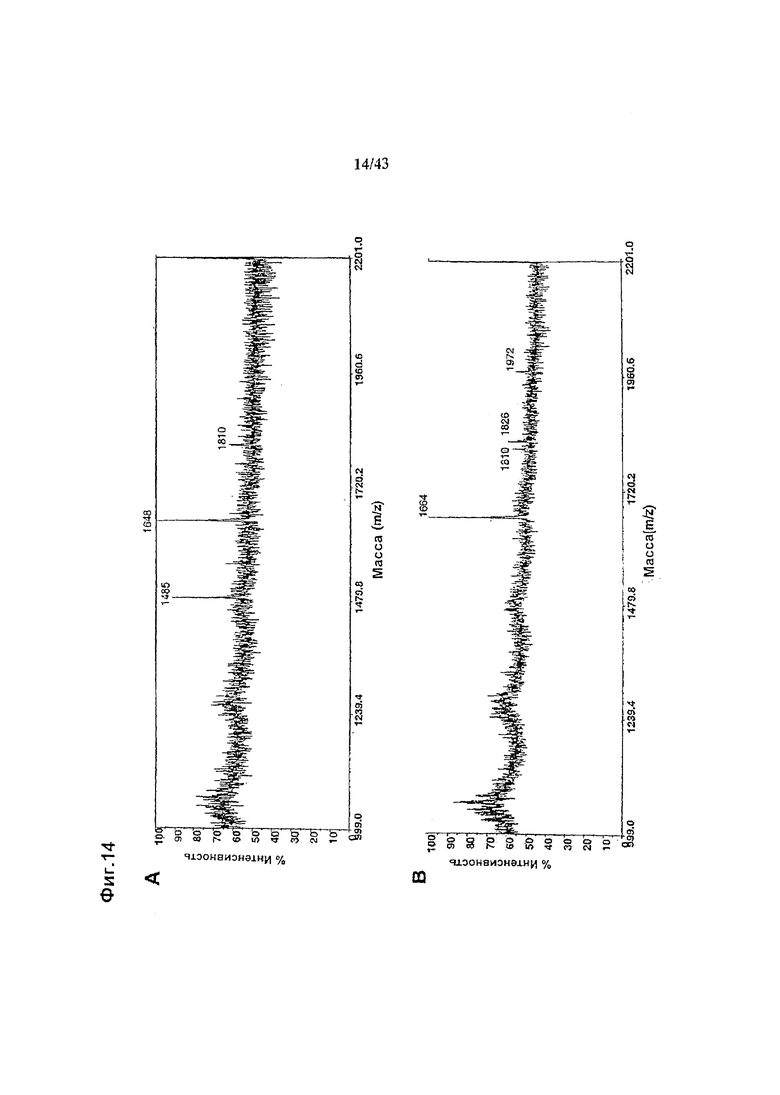
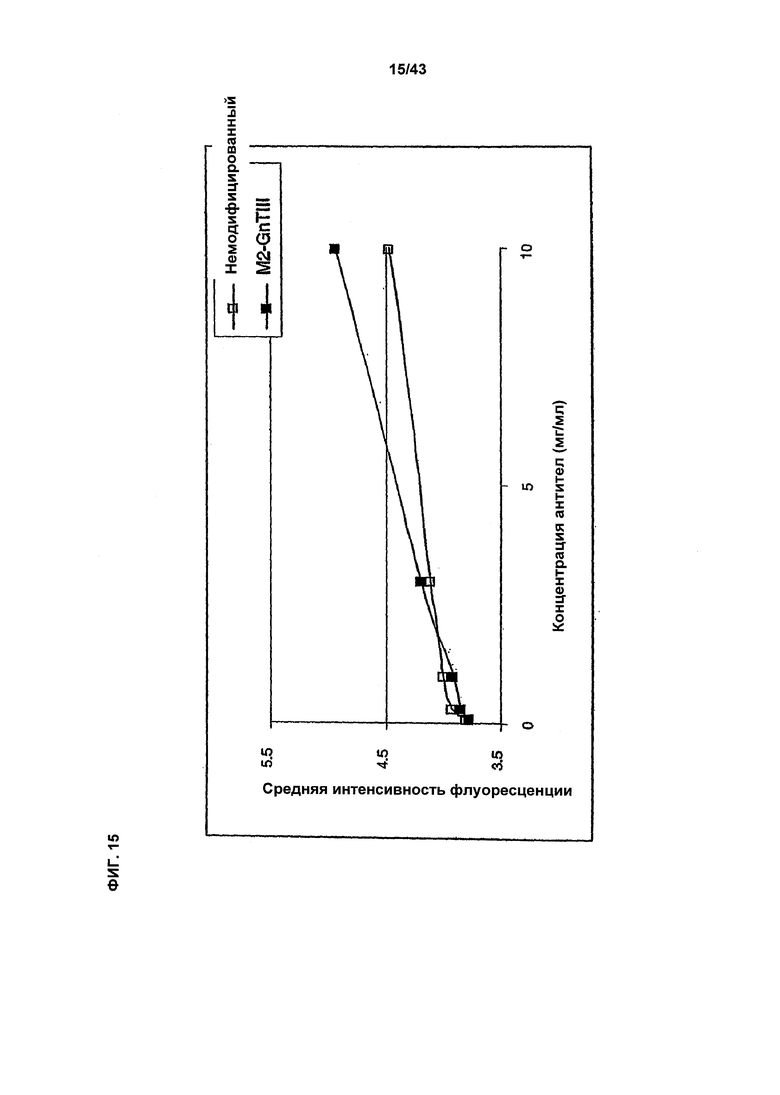

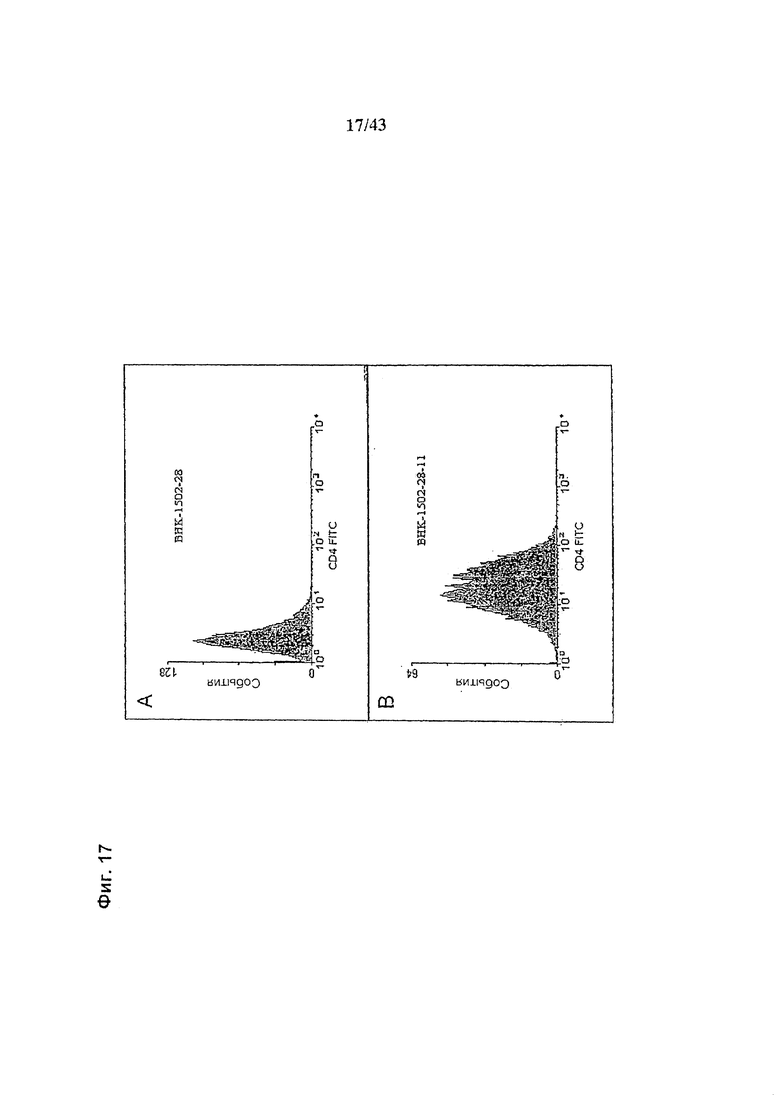
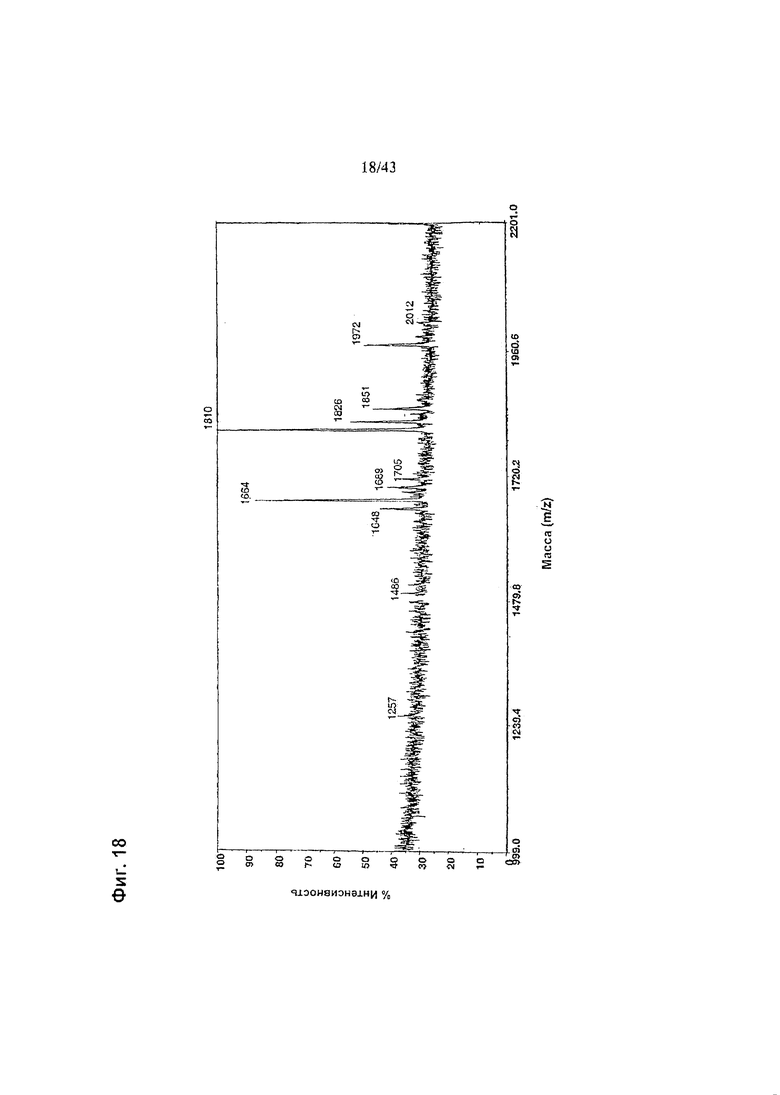
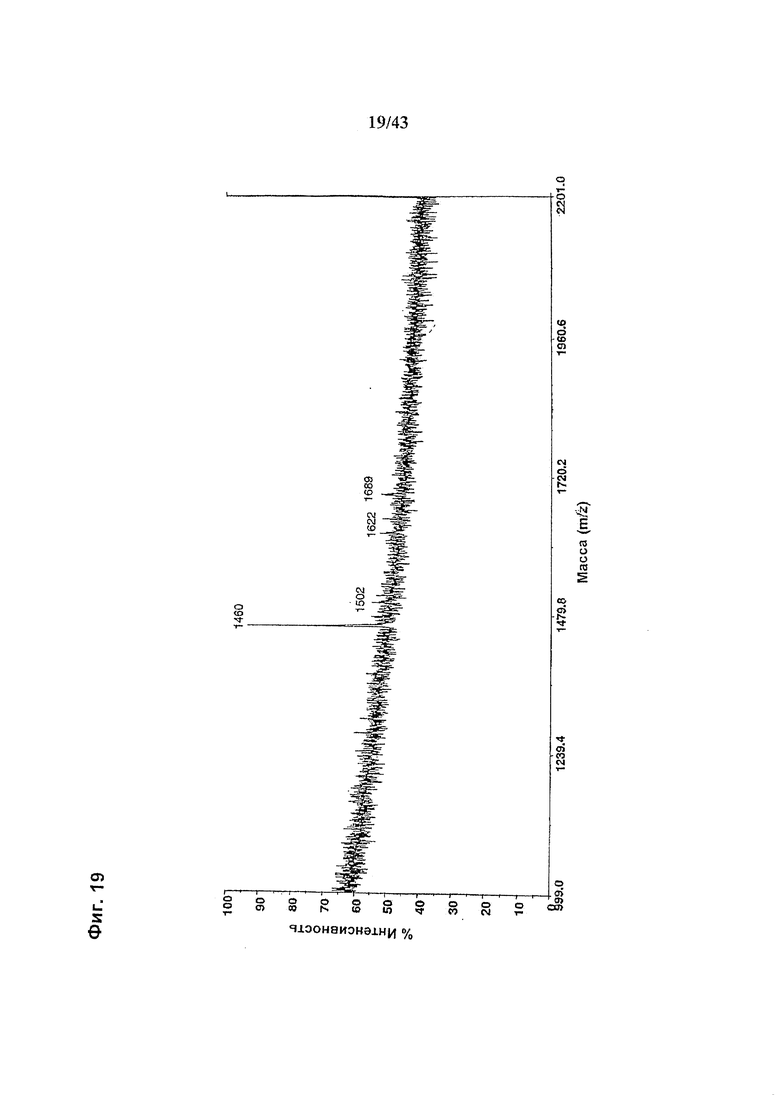

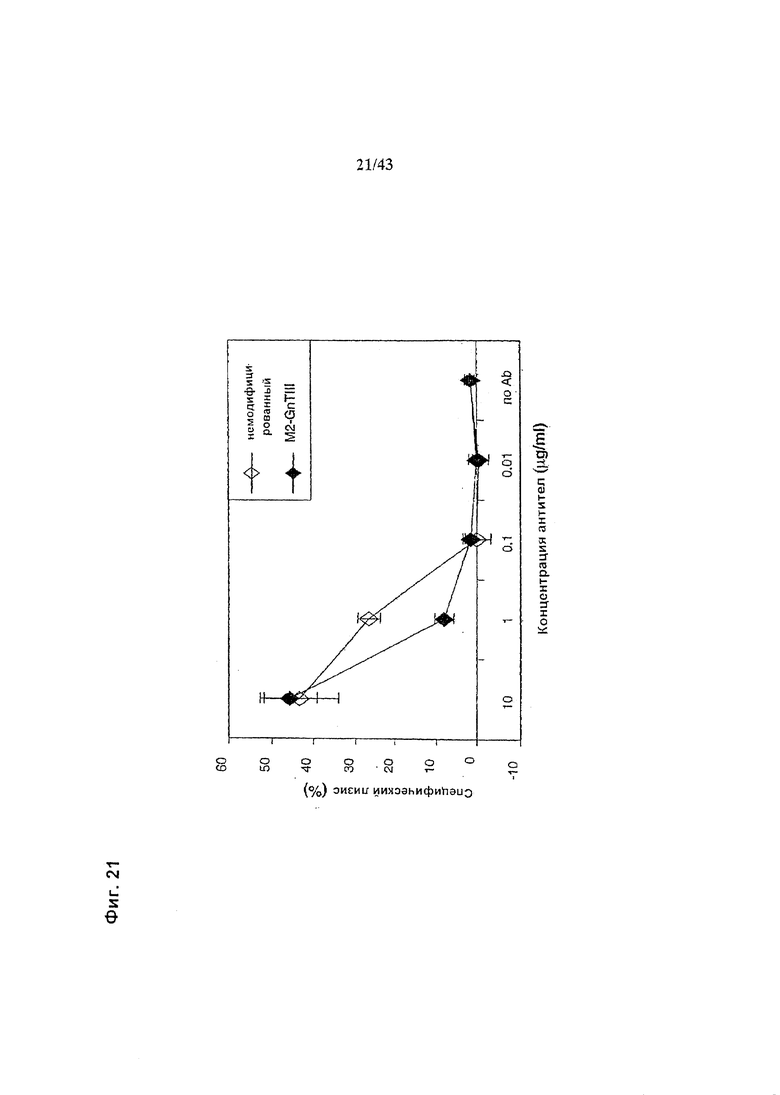
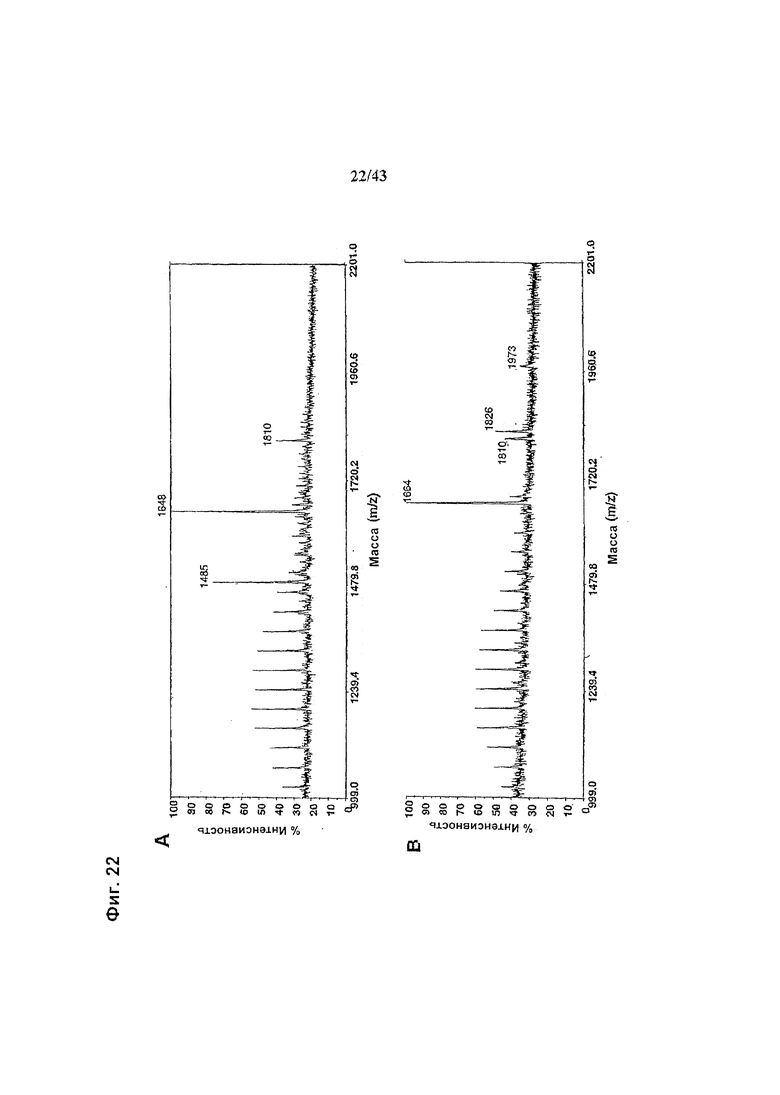
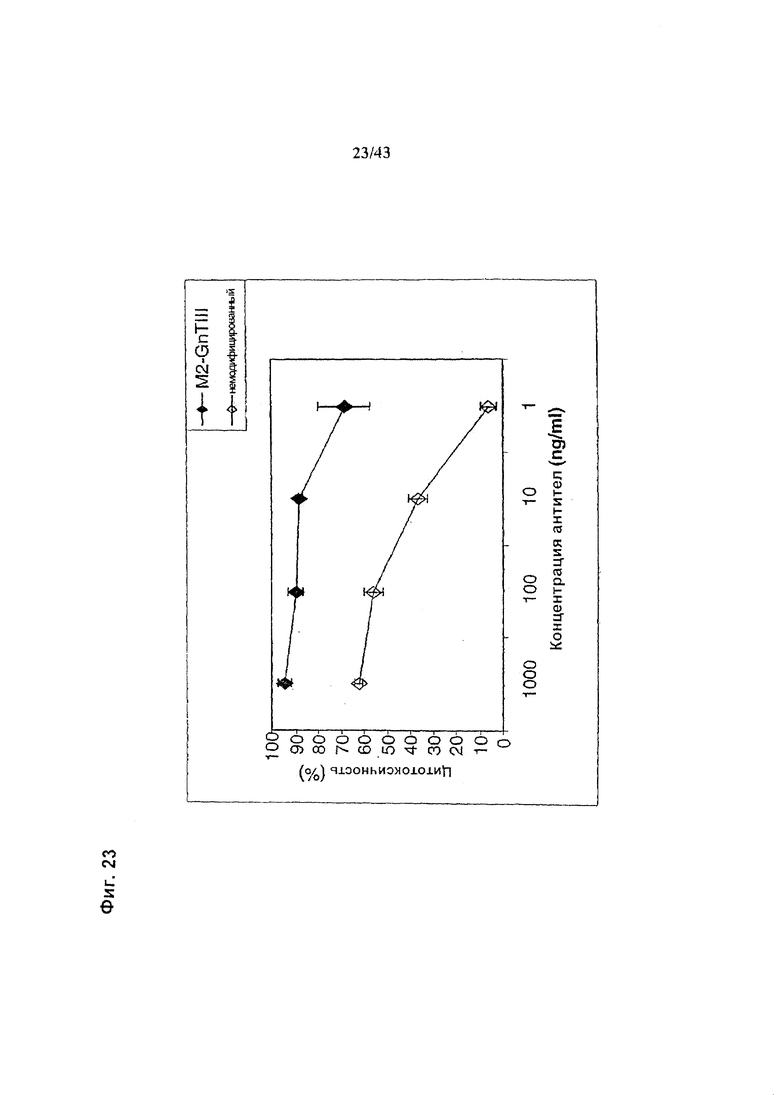


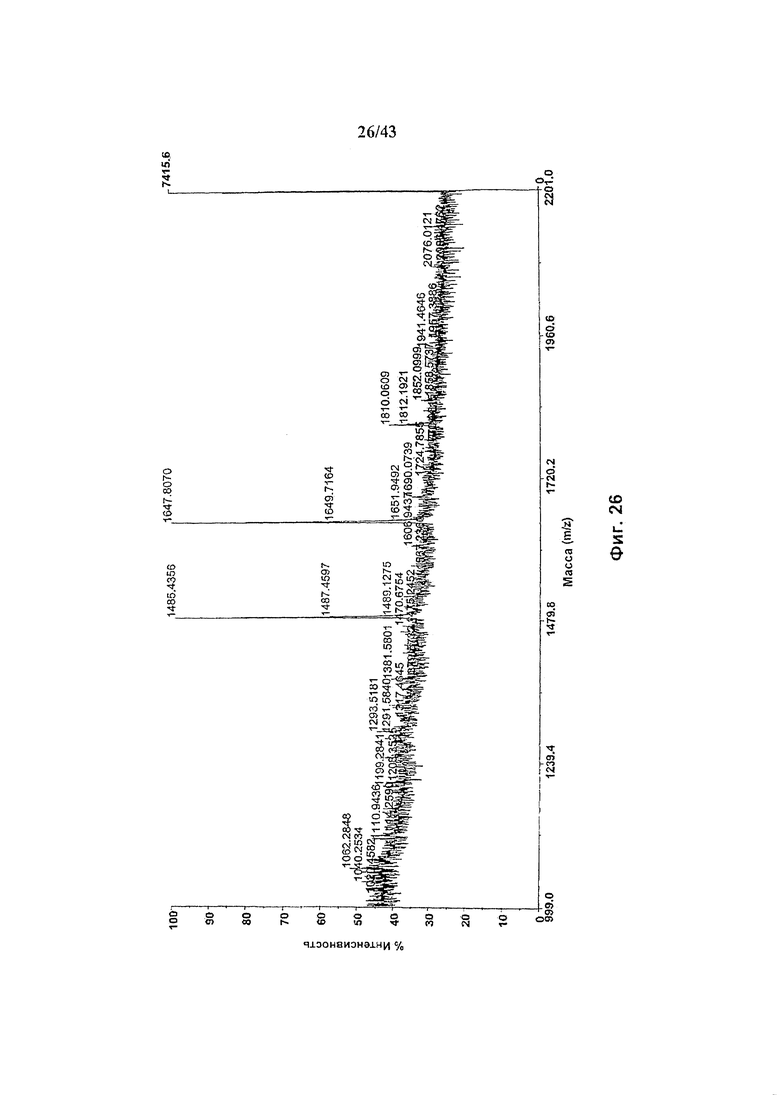
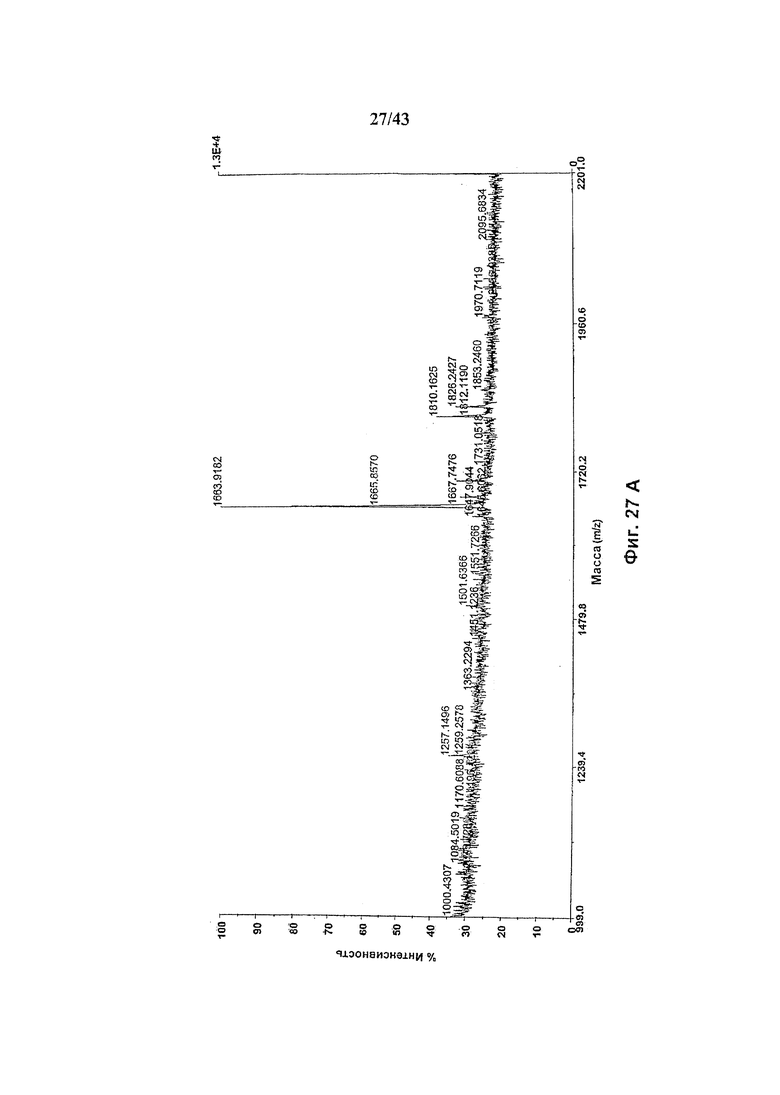
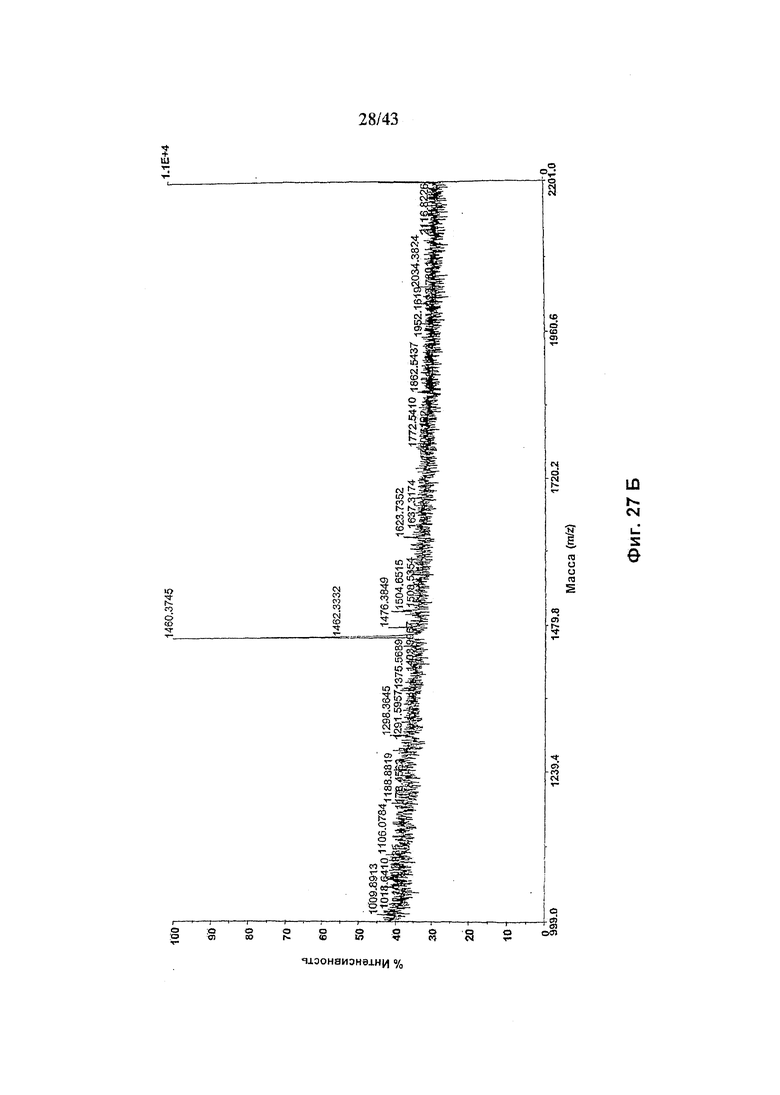
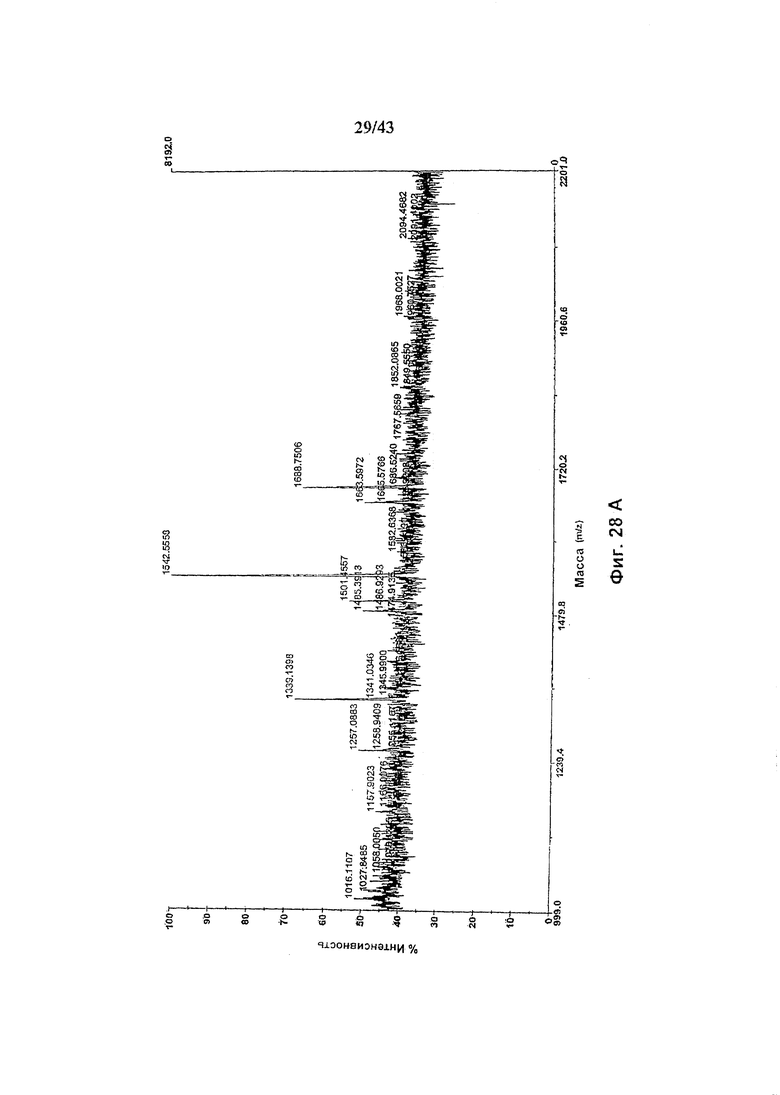

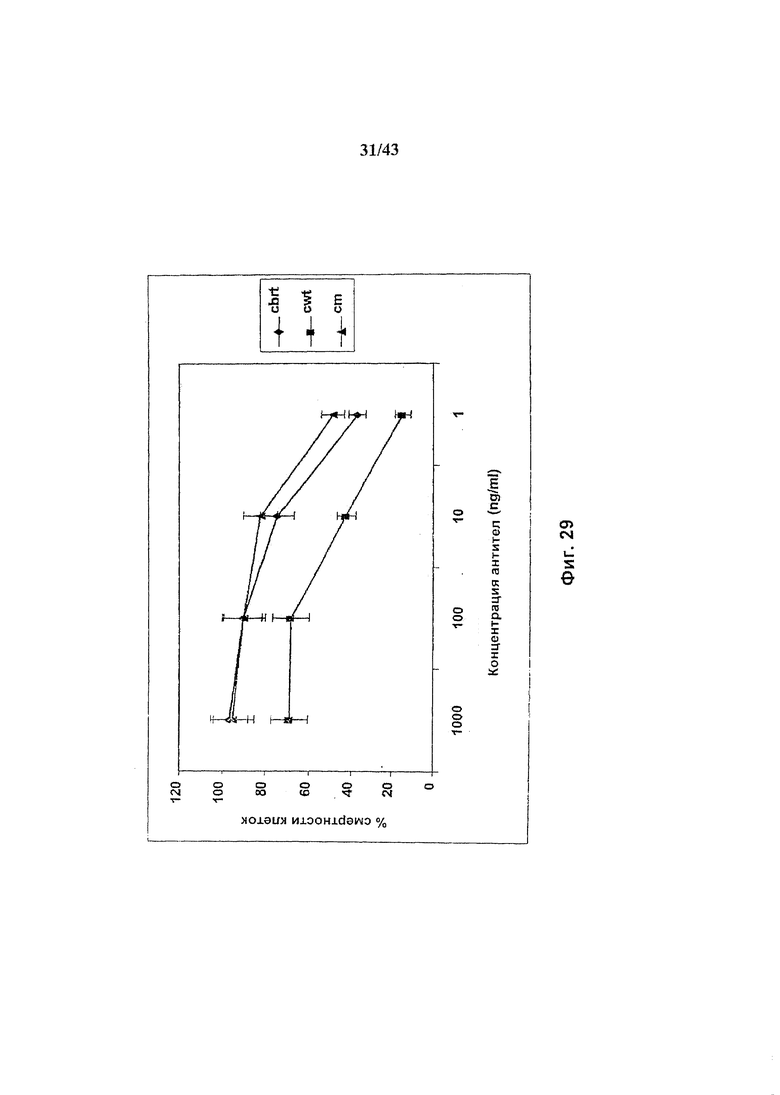
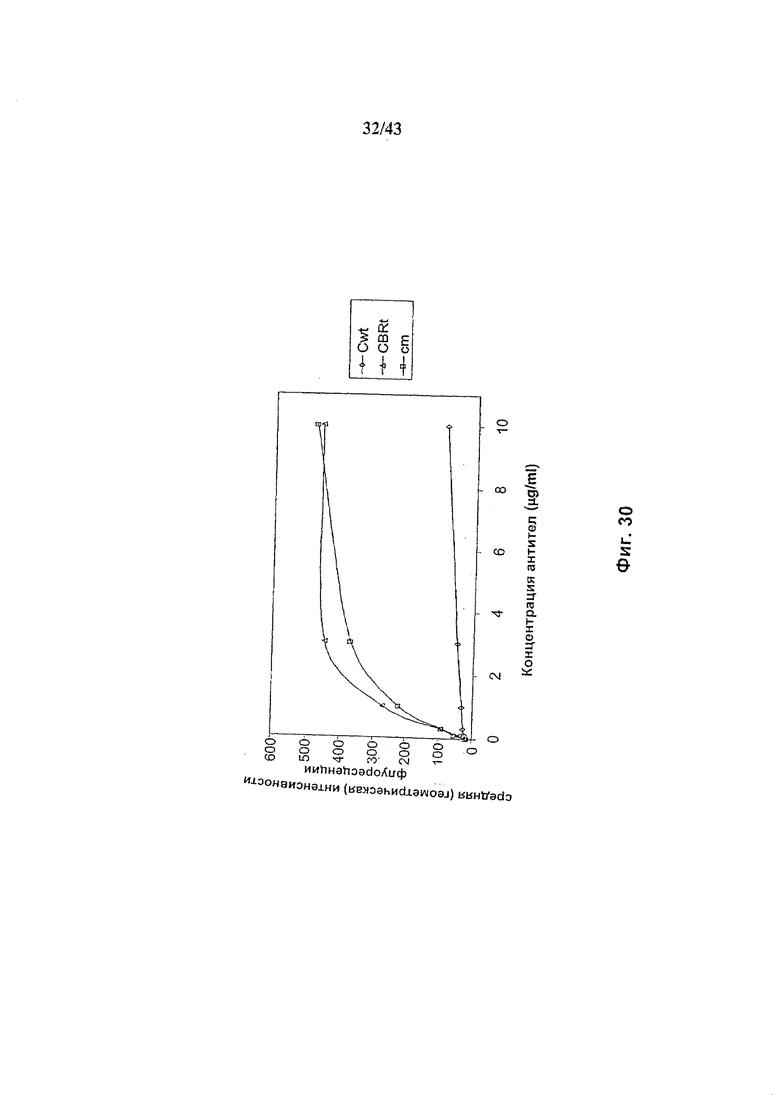
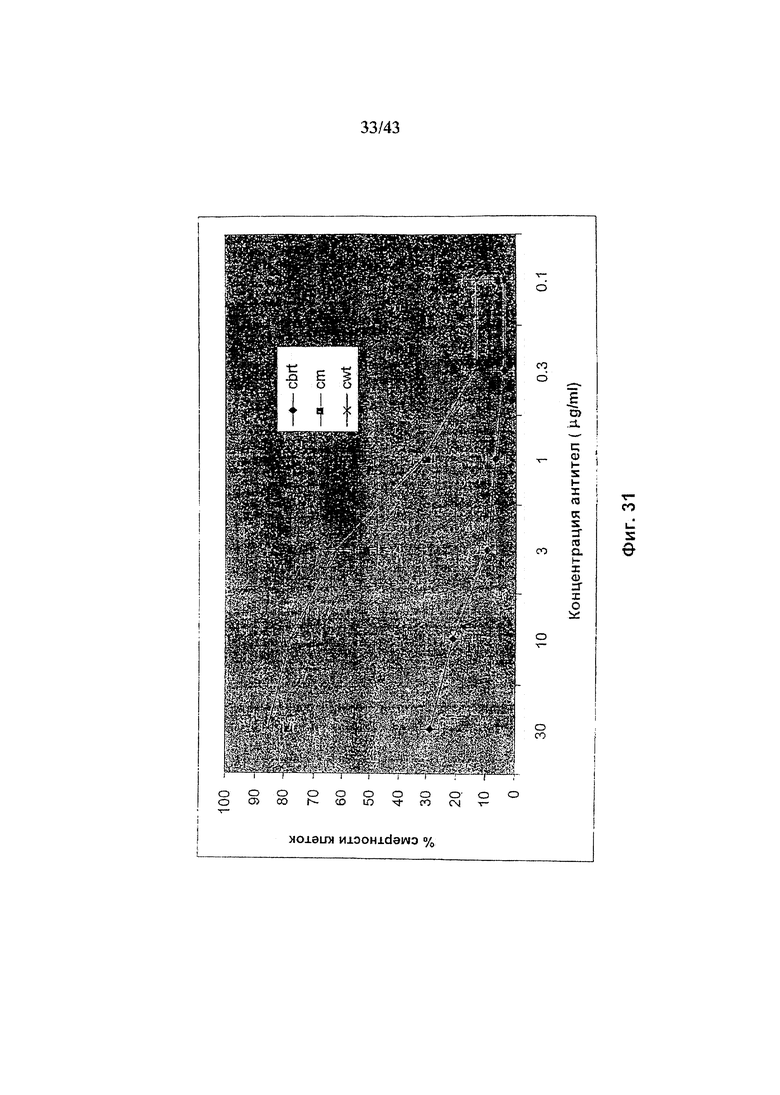

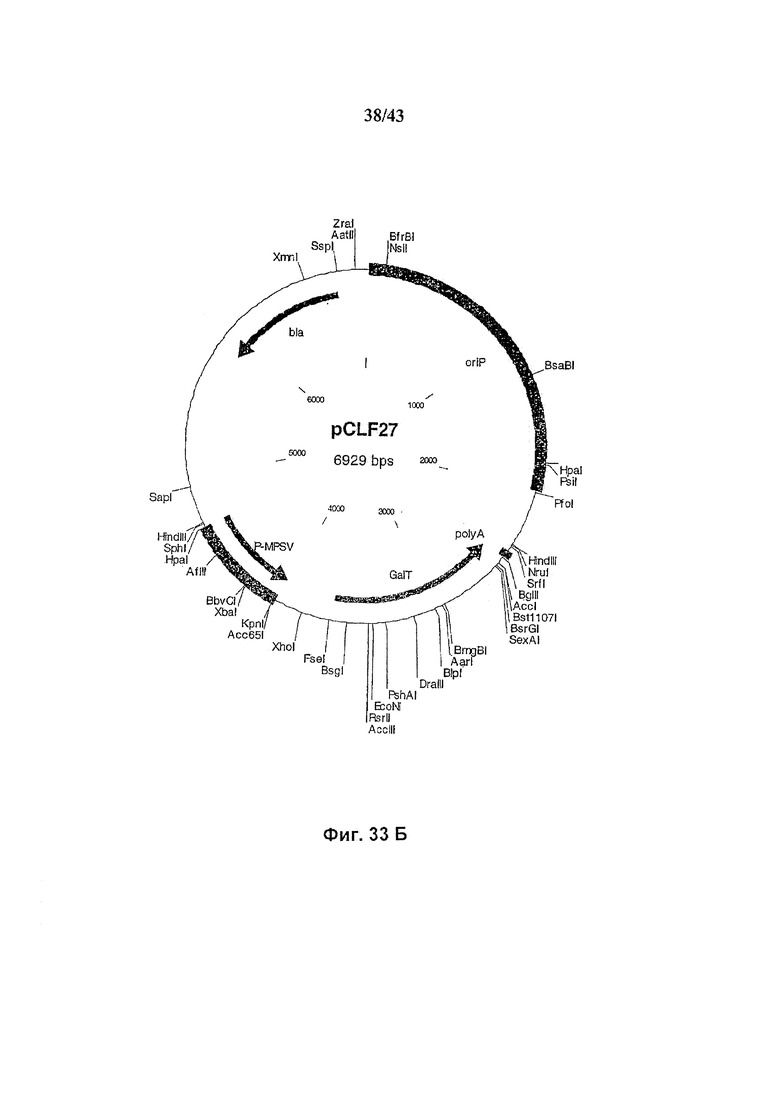
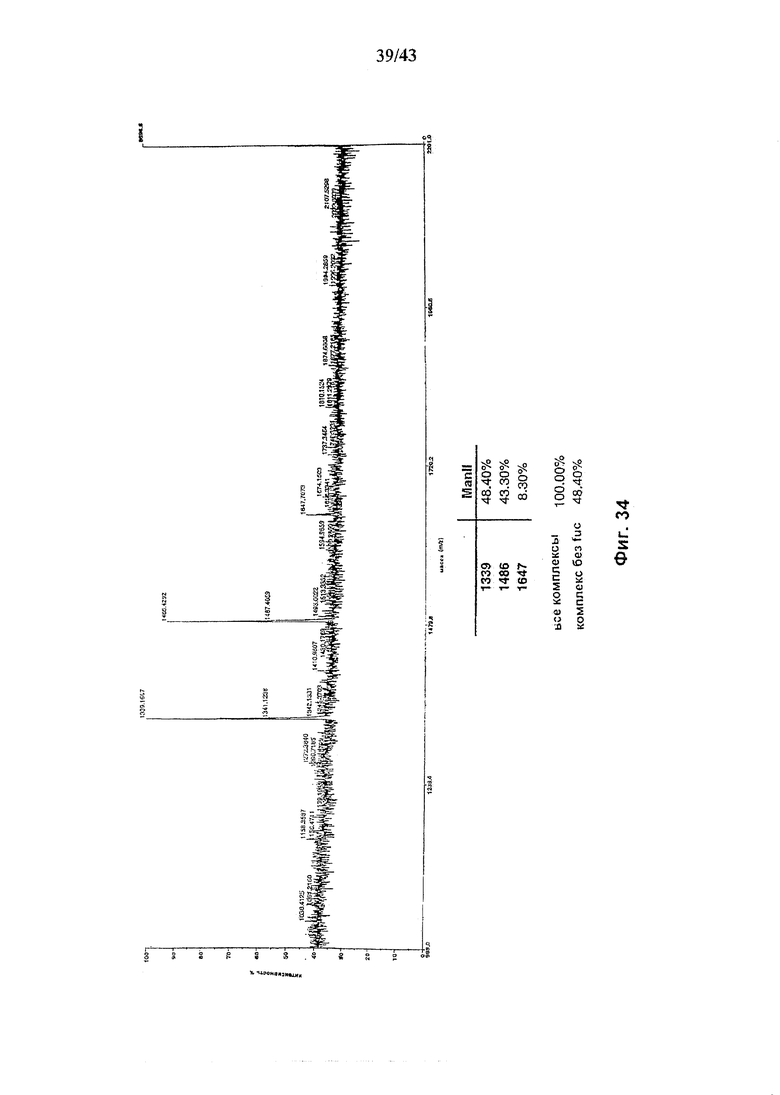
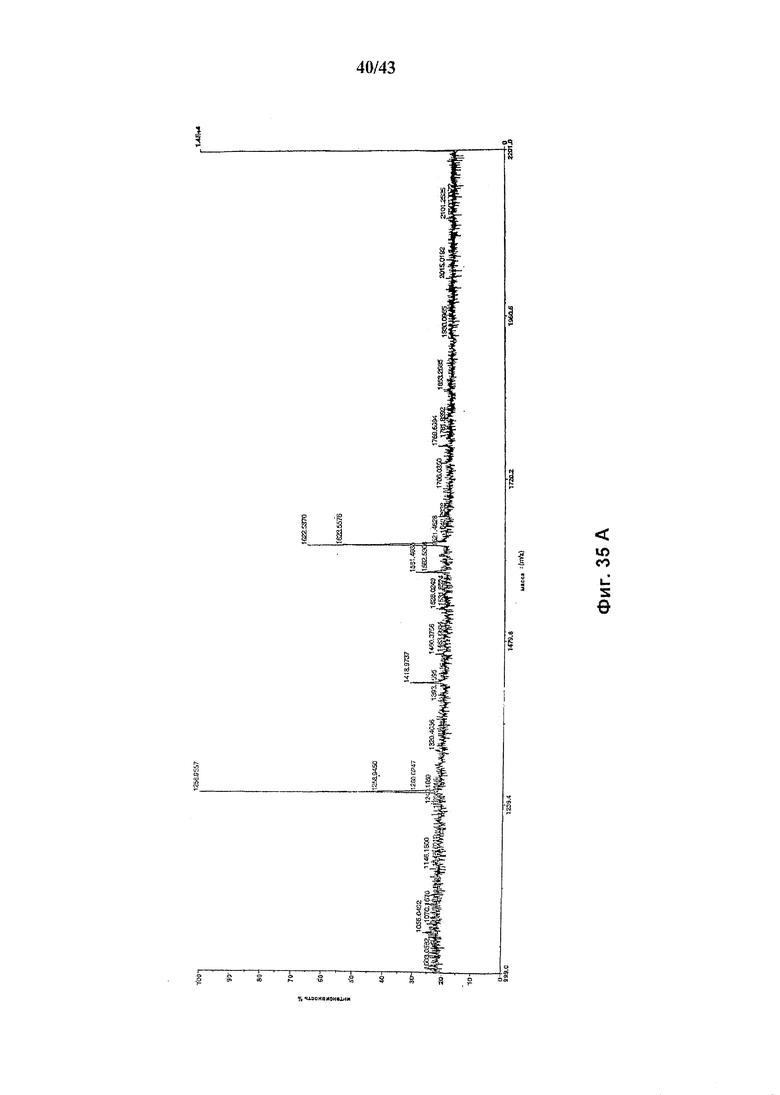
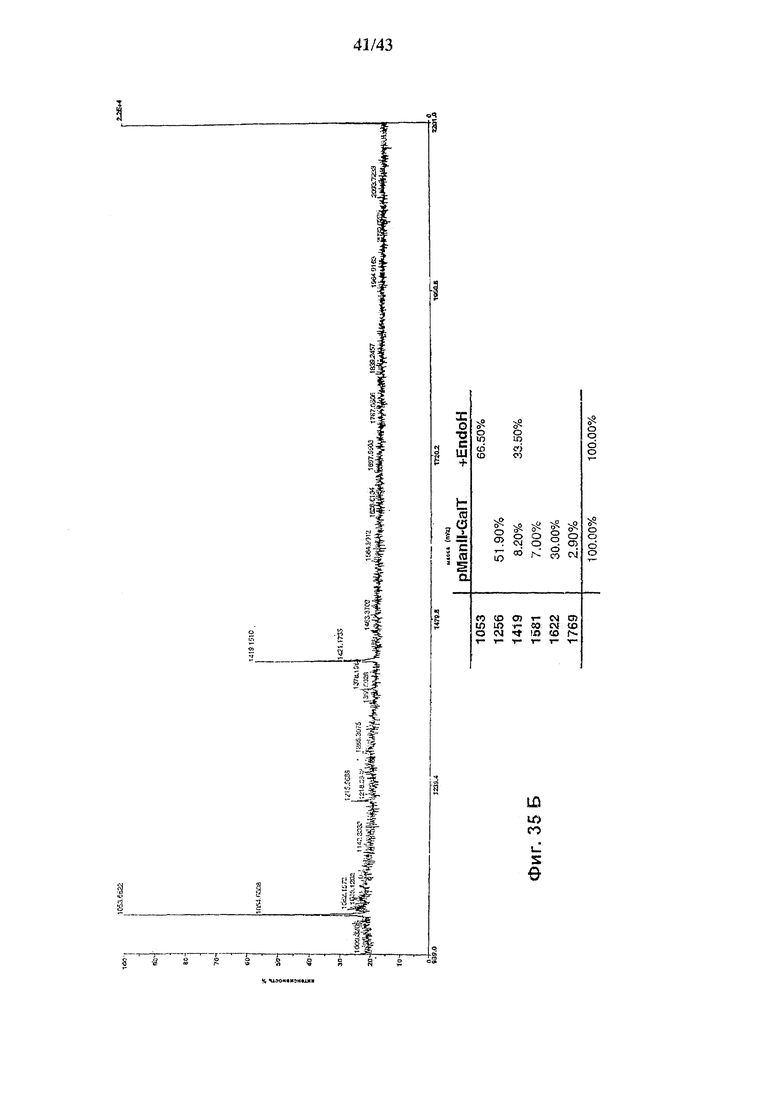
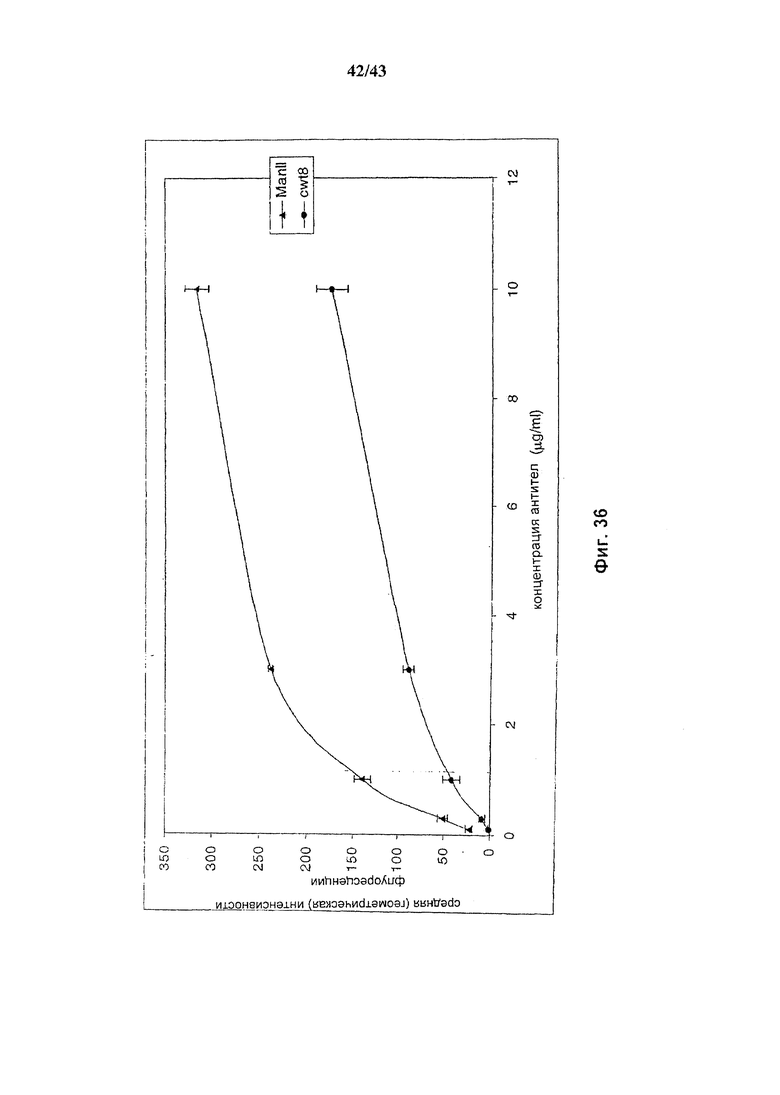
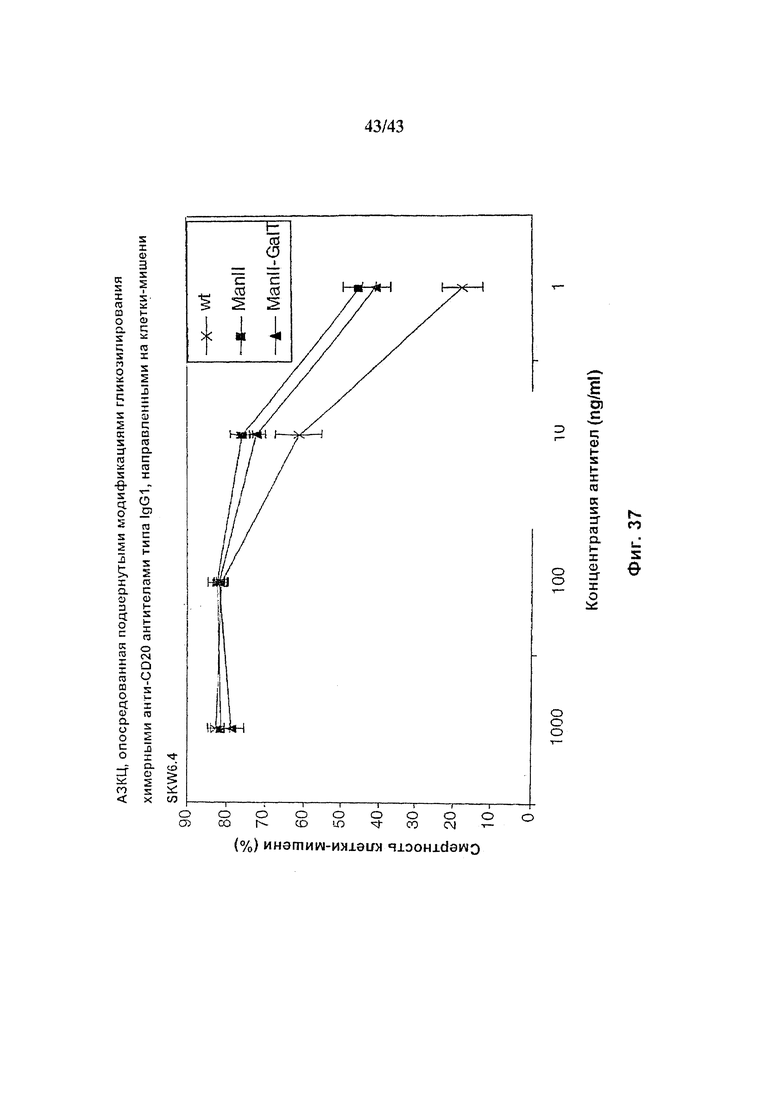
Изобретение относится к области биохимии, в частности к выделенной нуклеиновой кислоте, кодирующей полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) или бета-1,4-галактозилтрансферазы (GalT). Также раскрыты клетка-хозяин, содержащая указанную нуклеиновую кислоту, и вектор экспрессии, содержащий указанную выделенную нуклеиновую кислоту. Раскрыт способ модификации профиля гликозилирования полипептида указанной нуклеиновой кислотой. Изобретение позволяет модифицировать гликозилирование в клетке-хозяине с целью получения полипептидов с улучшенными терапевтическими свойствами. 6 н. и 18 з.п. ф-лы, 43 ил, 2 табл., 8 пр.
1. Выделенная нуклеиновая кислота, кодирующая полипептид слияния, обладающий активностью бета-1,4-N-ацетилглюкозаминилтрансферазы III (GnTIII) или бета-1,4-галактозилтрансферазы (GalT), причем указанный полипептид слияния содержит каталитический домен GnTIII млекопитающего или GalT млекопитающего и отвечающий за локализацию в комплексе Гольджи домен бета-1,2-N-ацетилглюкозаминилтрансферазы I (GnTI) млекопитающего или маннозидазы II млекопитающего (Man II),
причем указанный каталитический домен GnTIII млекопитающего содержит аминокислотную последовательность SEQ ID NO: 21,
причем указанный каталитический домен GalT млекопитающего содержит аминокислотную последовательность SEQ ID NO: 22,
причем указанный отвечающий за локализацию в комплексе Гольджи домен Man II млекопитающего содержит аминокислотную последовательность SEQ ID NO: 23, и
причем указанный отвечающий за локализацию в комплексе Гольджи домен GnTI млекопитающего содержит аминокислотную последовательность SEQ ID NO: 24.
2. Выделенная нуклеиновая кислота по п. 1, отличающаяся тем, что указанный полипептид слияния обладает активностью GnTIII и содержит каталитический домен GnTIII млекопитающего и отвечающий за локализацию в комплексе Гольджи домен Man II млекопитающего.
3. Выделенная нуклеиновая кислота по п. 2, отличающаяся тем, что указанный каталитический домен GnTIII млекопитающего содержит аминокислотную последовательность SEQ ID NO: 21, и при этом указанный отвечающий за локализацию в комплексе Гольджи домен Man II млекопитающего содержит аминокислотную последовательность SEQ ID NO: 23.
4. Выделенная нуклеиновая кислота по п. 3, отличающаяся тем, что указанный полипептид слияния имеет аминокислотную последовательность SEQ ID NO: 26.
5. Выделенная нуклеиновая кислота по п. 1, отличающаяся тем, что указанный полипептид слияния обладает активностью GnTIII и содержит каталитический домен GnTIII млекопитающего и отвечающий за локализацию в комплексе Гольджи домен GnTI млекопитающего.
6. Выделенная нуклеиновая кислота по п. 5, отличающаяся тем, что указанный каталитический домен GnTIII млекопитающего содержит аминокислотную последовательность SEQ ID NO: 21, и при этом указанный отвечающий за локализацию в комплексе Гольджи домен GnTI млекопитающего содержит аминокислотную последовательность SEQ ID NO: 24.
7. Выделенная нуклеиновая кислота по п. 6, отличающаяся тем, что указанный полипептид слияния имеет аминокислотную последовательность SEQ ID NO: 28.
8. Выделенная нуклеиновая кислота по п. 1, отличающаяся тем, что указанный полипептид слияния обладает активностью GalT и содержит каталитический домен GalT млекопитающего и отвечающий за локализацию в комплексе Гольджи домен Man II млекопитающего.
9. Выделенная нуклеиновая кислота по п. 8, отличающаяся тем, что указанный каталитический домен GalT млекопитающего содержит аминокислотную последовательность SEQ ID NO: 22, и при этом указанный отвечающий за локализацию в комплексе Гольджи домен Man II млекопитающего содержит аминокислотную последовательность SEQ ID NO: 23.
10. Выделенная нуклеиновая кислота по п. 1, отличающаяся тем, что указанный полипептид слияния обладает активностью GalT и содержит каталитический домен GalT млекопитающего и отвечающий за локализацию в комплексе Гольджи домен GnTI млекопитающего.
11. Выделенная нуклеиновая кислота по п. 10, отличающаяся тем, что указанный каталитический домен GalT млекопитающего содержит аминокислотную последовательность SEQ ID NO: 22, и при этом указанный отвечающий за локализацию в комплексе Гольджи домен GnTI млекопитающего содержит аминокислотную последовательность SEQ ID NO: 24.
12. Клетка-хозяин млекопитающего для продуцирования полипептида слияния, обладающего активностью GnTIII или GalT, причем указанная клетка-хозяин млекопитающего содержит выделенную нуклеиновую кислоту по любому и пп. 1-11, и при этом указанная клетка-хозяин млекопитающего не является эмбриональной клеткой человека.
13. Вектор экспрессии млекопитающего, содержащий выделенную нуклеиновую кислоту по любому из пп. 1-11.
14. Клетка-хозяин млекопитающего для продуцирования полипептида слияния, обладающего активностью GnTIII или GalT, причем указанная клетка-хозяин млекопитающего содержит вектор экспрессии млекопитающего по п. 13, и при этом указанная клетка-хозяин млекопитающего не является эмбриональной клеткой человека.
15. Способ модификации профиля гликозилирования полипептида, продуцированного клеткой-хозяином млекопитающего, причем указанный способ включает введение в указанную клетку-хозяина млекопитающего выделенной нуклеиновой кислоты по любому из пп. 1-11, причем указанный полипептид, продуцированный указанной клеткой-хозяином млекопитающего, выбран из группы, состоящей из целой молекулы антитела, фрагмента антитела, сохраняющего функциональную Fc-область, и белка слияния, содержащего область, эквивалентную Fc-области иммуноглобулина, и при этом указанная клетка-хозяин млекопитающего не является эмбриональной клеткой человека.
16. Способ по п. 15, отличающийся тем, что указанный полипептид, продуцированный указанной клеткой-хозяином млекопитающего, представляет собой IgG или его фрагмент, содержащий Fc-область IgG.
17. Способ по п. 15, отличающийся тем, что указанный полипептид, продуцированный указанной клеткой-хозяином млекопитающего, представляет собой IgG1 или его фрагмент, содержащий Fc-область IgG1.
18. Способ по п. 15, отличающийся тем, что указанный полипептид, продуцированный указанной клеткой-хозяином млекопитающего, представляет собой белок слияния, содержащий область, эквивалентную Fc-области IgG человека.
19. Способ по п. 15, отличающийся тем, что указанная клетка-хозяин млекопитающего выбрана из группы, состоящей из клетки СНО, клетки ВНК, клетки NS0, клетки SP2/0, клетки YO миеломы, клетки Р3Х63 миеломы мыши, клетки PER, клетки PER.C6 и клетки гибридомы.
20. Способ модификации профиля гликозилирования полипептида, продуцированного клеткой-хозяином млекопитающего, причем указанный способ включает введение в указанную клетку-хозяина млекопитающего вектора экспрессии млекопитающего по п. 13, причем указанный полипептид, продуцированный указанной клеткой-хозяином млекопитающего, выбран из группы, состоящей из целой молекулы антитела, фрагмента антитела, сохраняющего функциональную Fc-область, и белка слияния, содержащего область, эквивалентную Fc-области иммуноглобулина, и при этом указанная клетка-хозяин млекопитающего не является эмбриональной клеткой человека..
21. Способ по п. 20, отличающийся тем, что указанный полипептид, продуцированный указанной клеткой-хозяином млекопитающего, представляет собой IgG или его фрагмент, содержащий Fc-область IgG.
22. Способ по п. 20, отличающийся тем, что указанный полипептид, продуцированный указанной клеткой-хозяином млекопитающего, представляет собой IgG1 или его фрагмент, содержащий Fc-область IgG1.
23. Способ по п. 20, отличающийся тем, что указанный полипептид, продуцированный указанной клеткой-хозяином млекопитающего, представляет собой белок слияния, содержащий область, эквивалентную Fc-области IgG человека.
24. Способ по п. 20, отличающийся тем, что указанная клетка-хозяин млекопитающего выбрана из группы, состоящей из клетки СНО, клетки ВНК, клетки NS0, клетки SP2/0, клетки YO миеломы, клетки Р3Х63 миеломы мыши и клетки гибридомы.
| WO 1991012340 A1 от 22.08.1991;US 20020137134 A1 от 26.09.2002;US 5876714 A от 02.03.1999;WO 2000050608 A1 от 31.08.2000;RU 2133624 C1 от 27.07.1999. |

