ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение в целом относится к биспецифическим антигенсвязывающим молекулам. Кроме того, настоящее изобретение относится к полинуклеотидам, кодирующим такие биспецифические антигенсвязывающие молекулы, а также к векторам и клеткам-хозяевам, содержащим такие полинуклеотиды. Изобретение, кроме того, относится к способам получения биспецифических антигенсвязывающих молекул по изобретению и к способам применения этих биспецифических антигенсвязывающих молекул при лечении заболевания.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Би- или мультиспецифические антитела, способные к связыванию двух или более антигенов, известны в данной области техники. Такие мультиспецифические связывающие белки могут быть образованы путем слияния клеток гибридомы, химической конъюгации или методов рекомбинантных ДНК.
Биспецифические антитела представляют большой интерес для терапевтических применений, поскольку они дают возможность одновременного связывания и инактивации двух или более антигенов-мишеней, что, таким образом, устраняет необходимость в комбинированных терапиях. Другим перспективным применением биспецифических антител является применение в качестве молекул, привлекающих иммунные эффекторные клетки, например, для клеточной иммунотерапии рака. Для этой цели конструируют биспецифические антитела, связывающиеся с поверхностным антигеном на клетках-мишенях, и, например, с активирующим компонентом Т-клеточного рецепторного комплекса (TCR; от англ. "Т cell receptor"). Одновременное связывание такого антитела с обеими его мишенями усилит преходящее взаимодействие между клеткой-мишенью и Т-клеткой, вызывающее активацию любого цитотоксического Т лимфоцита (CTL; от англ. "cytotoxic T lymphocyte") и последующий лизис клетки-мишени. Следовательно, иммунный ответ перенаправляется на клетки-мишени независимо от презентирования пептидного антигена клеткой-мишенью или специфичности Т-клетки, необходимой для нормальной, ограниченной главным комплексом гистосовместимости (ГКГ) активации CTL. В данном контексте важно, чтобы CTL активировались только тогда, когда клетка-мишень презентирует им биспецифическое антитело, то есть, при имитации иммунологической конъюгации, а не просто при связывании антитела с T-клеточным антигеном.
В недавнем прошлом разработан ряд форматов рекомбинантного мультиспецифического антитела, включающих, например, слияния тетравалентного IgG с одноцепочечным вариабельным фрагментом (scFv) (см., например, Coloma & Morrison, Nat Biotechnol 15, 159-163 (1997)), антитела, подобные двойному вариабельному домену тетравалентного IgG (Wu et al., Nat Biotechnol 25, 1290-1297 (2007)) или бивалентные гибридные биспецифические IgG крысы/мыши (см., например, Lindhofer et al., J Immunol 155, 219-225 (1995)).
Также получено несколько биспецифических форматов, в которых основная структура антитела (IgA, IgD, IgE, IgG или IgM) больше не сохраняется. Примеры включают диатела (см., например, Holliger et al., Proc Natl Acad Sci USA 90, 6444-6448 (1995)), тандемные молекулы scFv (см., например, Bargou et al., Science 321, 974-977 (2008)) и различные их производные.
Множество форматов, находящихся в разработке, проявляет огромный потенциал, присущий биспецифическим антителам. Задача создания биспецифических антител, подходящих для конкретной цели, однако, никоим образом не является тривиальной и требует ряда разработок. Например, необходимо соответствующим образом выбрать валентность и геометрию антитела в зависимости от характеристик антигенов-мишеней и предназначенного эффекта. Как для всех терапевтических антител, необходимо сбалансировать эффективность и токсичность, что требует среди прочего минимизации иммуногенности и оптимизации фармакокинетических свойств антитела. Также необходимо учитывать желательность Fc-опосредованных эффектов. Кроме того, получение конструкций биспецифического антитела, имеющих клинически достаточное количество и чистоту, создает значительную проблему, поскольку гомодимеризация тяжелых цепей антитела и/или ошибочное спаривание тяжелых и легких цепей антитела различных специфичностей при совместной экспрессии снижает выход правильно собранной конструкции и приводит в результате к ряду нефункциональных побочных продуктов, от которых может быть трудно отделить желаемое биспецифическое антитело.
С учетом возрастающего числа возможных применений биспецифических антител, а также трудностей и недостатков, связанных с доступными в настоящее время биспецифическими антителами, остается необходимость в новых, усовершенствованных форматах таких молекул.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В первом аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, включающая первый Fab фрагмент, специфично связывающийся с первым антигеном, второй Fab фрагмент, специфично связывающийся со вторым антигеном, и домен Fc, состоящий из первой и второй субъединиц, способных к стабильной ассоциации; причем,
a) биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым и/или вторым антигеном,
b) первый Fab фрагмент, второй Fab фрагмент и первая субъединица домена Fc слиты друг с другом, и
c) в первом и/или втором Fab фрагменте произведена одна из следующих замен: (i) вариабельные домены легкой цепи (VL; от англ. "variable light") и тяжелой цепи (VH; от англ. "variable heavy") заменены друг другом, (ii) константный домен легкой цепи (CL; от англ. "constant light") и первый константный домен тяжелой цепи (СН; от англ. "constant heavy") CH1 заменены друг другом, или (iii) как вариабельные, так и константные домены VL-CL и VH-СН1 заменены друг другом,
при условии, что в первом и втором Fab фрагменте произведена не одна и та же замена.
В конкретных формах осуществления первый Fab фрагмент слит на его С-конце с N-концом второго Fab фрагмента, который, в свою очередь, слит на его С-конце с N-концом первой субъединицы домена Fc. В более конкретной форме осуществления первый Fab фрагмент слит на С-конце его тяжелой цепи с N-концом тяжелой цепи второго Fab фрагмента, который, в свою очередь, слит на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc. В других формах осуществления второй Fab фрагмент слит на его С-конце с N-концом первого Fab фрагмента, который, в свою очередь, слит на его С-конце с N-концом первой субъединицы домена Fc. В более конкретной форме осуществления второй Fab фрагмент слит на С-конце его тяжелой цепи с N-концом тяжелой цепи первого Fab фрагмента, который, в свою очередь, слит на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc. В других формах осуществления второй Fab фрагмент слит на его С-конце с N-концом первой субъединицы домена Fc, которая, в свою очередь, слита на ее С-конце с N-концом первого Fab фрагмента. В более конкретной форме осуществления второй Fab фрагмент слит на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc, которая, в свою очередь, слита на ее С-конце с N-концом тяжелой цепи первого Fab фрагмента.
В формах осуществления, в которых либо первый Fab фрагмент слит на С-конце его тяжелой цепи с N-концом тяжелой цепи второго Fab фрагмента, который, в свою очередь, слит на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc, либо второй Fab фрагмент слит на С-конце его тяжелой цепи с N-концом тяжелой цепи первого Fab фрагмента, который, в свою очередь, слит на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc, дополнительно легкая цепь Fab первого Fab фрагмента и легкая цепь Fab второго Fab фрагмента могут быть слиты друг с другом, необязательно посредством пептидного линкера.
В одной форме осуществления замена произведена в первом Fab фрагменте. В некоторых формах осуществления замена представляет собой замену вариабельных доменов VL и VH друг другом. В других формах осуществления замена представляет собой замену константных доменов CL и СН1 друг другом.
В одной форме осуществления биспецифическая антигенсвязывающая молекула по существу состоит из первого Fab фрагмента, второго Fab фрагмента, домена Fc и необязательно одного или более пептидных линкеров.
В конкретных формах осуществления биспецифическая антигенсвязывающая молекула включает третий Fab фрагмент, специфично связывающийся с первым или вторым антигеном. В одной форме осуществления третий Fab фрагмент слит со второй субъединицей домена Fc. В более конкретной форме осуществления третий Fab фрагмент слит на его С-конце с N-концом второй субъединицы домена Fc. В еще более конкретной форме осуществления третий Fab фрагмент слит на С-конце его тяжелой цепи с N-концом второй субъединицы домена Fc. В одной форме осуществления третий Fab фрагмент специфично связывается со вторым антигеном. В некоторых формах осуществления второй Fab фрагмент, третий Fab фрагмент и домен Fc являются частью молекулы иммуноглобулина. В такой конкретной форме осуществления молекула иммуноглобулина представляет собой молекулу иммуноглобулина класса IgG, более конкретно молекулу иммуноглобулина подкласса IgG1 или IgG4. В одной форме осуществления молекула иммуноглобулина представляет собой молекулу иммуноглобулина человека. В одной форме осуществления биспецифическая антигенсвязывающая молекула по существу состоит из первого Fab фрагмента, специфично связывающегося с первым антигеном, молекулы иммуноглобулина, специфично связывающейся со вторым антигеном, и необязательно одного или более пептидных линкеров.
В одной форме осуществления в Fab фрагментах, специфично связывающихся с одним и тем же антигеном, произведена одна и та же замена. В следующей форме осуществления замена произведена только в первом Fab фрагменте. В одной форме осуществления биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым антигеном. В одной форме осуществления биспецифическая антигенсвязывающая молекула не включает одноцепочечный Fab фрагмент.
В некоторых формах осуществления домен Fc включает модификацию, стимулирующую ассоциацию первой и второй субъединицы домена Fc. В такой конкретной форме осуществления аминокислотный остаток домена CH3 первой субъединицы домена Fc заменен аминокислотным остатком, имеющим больший объем боковой цепи, создавая посредством этого выпячивание внутри домена CH3 первой субъединицы, которое может быть расположено в углублении домена CH3 второй субъединицы, и аминокислотный остаток в домене CH3 второй субъединицы домена Fc заменен аминокислотным остатком, имеющим меньший объем боковой цепи, создавая посредством этого углубление внутри домена CH3 второй субъединицы, в котором может быть расположено выпячивание внутри домена CH3 первой субъединицы. В одной форме осуществления домен Fc представляет собой домен Fc IgG, в частности, домен Fc IgG1 или IgG4. В одной форме осуществления домен Fc представляет собой домен Fc человека. В некоторых формах осуществления домен Fc сконструирован таким образом, что он обладает измененным связывающим сродством с рецептором Fc и/или измененной эффекторной функцией по сравнению с не сконструированным доменом Fc.
Согласно другому аспекту изобретения предложен изолированный полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу по изобретению или ее фрагмент. Объем изобретения также включает полипептиды, кодируемые полинуклеотидами по изобретению. Кроме того, в изобретении предложен экспрессионный вектор, содержащий изолированный полинуклеотид по изобретению, и клетка-хозяин, содержащая изолированный полинуклеотид или экспрессионный вектор по изобретению. В некоторых формах осуществления клетка-хозяин представляет собой эукариотическую клетку, в частности, клетку млекопитающего.
В другом аспекте предложен способ получения биспецифической антигенсвязывающей молекулы по изобретению, включающий следующие стадии: а) культивирование клетки-хозяина по изобретению в условиях, подходящих для экспрессии биспецифической антигенсвязывающей молекулы, и b) выделение биспецифической антигенсвязывающей молекулы. Объем изобретения также включает биспецифическую антигенсвязывающую молекулу, полученную способом по изобретению.
Далее в изобретении предложена фармацевтическая композиция, содержащая биспецифическую антигенсвязывающую молекулу по изобретению и фармацевтически приемлемый носитель.
Объем изобретения также включает способы применения биспецифической антигенсвязывающей молекулы и фармацевтической композиции по изобретению. В одном аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула или фармацевтическая композиция по изобретению, применяемая в качестве лекарственного средства. В одном аспекте предложена биспецифическая антигенсвязывающая молекула или фармацевтическая композиция по изобретению, применяемая при лечении заболевания у индивидуума, нуждающегося в этом. В конкретной форме осуществления заболевание представляет собой рак.
Также предложено применение биспецифической антигенсвязывающей молекулы согласно изобретению для получения лекарственного средства для лечения заболевания у индивидуума, нуждающегося в этом; а также способ лечения заболевания у индивидуума, включающий введение индивидууму терапевтически эффективного количества композиции, содержащей биспецифическую антигенсвязывающую молекулу согласно изобретению в фармацевтически приемлемой форме. В конкретной форме осуществления заболевание представляет собой рак. В любой из вышеописанных форм осуществления индивидуум предпочтительно представляет собой млекопитающее, в частности, человека.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
ФИГ.1. Иллюстрация иллюстративных форматов биспецифических антигенсвязывающих молекул по изобретению. (А) формат "2+1", в котором фрагмент Crossfab отличающейся специфичности слит с N-концом Fab фрагмента, содержащегося в антителе ("2+1 IgG Crossfab (N-концевой)"). (В) формат "1+1", в котором фрагмент Crossfab отличающейся специфичности слит с N-концом Fab фрагмента, содержащегося в антителе, в котором отсутствует второй Fab фрагмент ("1+1 IgG Crossfab (N-концевой)"). (С) формат "2+1", такой как в (А), в котором порядок фрагмента Crossfab, и Fab фрагмент, с которым слит фрагмент Crossfab, инвертирован ("2+1" IgG Crossfab (N-концевой), инвертированный"). (D) формат "2+1", в котором фрагмент Crossfab отличающейся специфичности слит с С-концом субъединицы домена Fc, содержащейся в антителе ("2+1 IgG Crossfab (C-концевой)"). Черная точка: необязательная модификация в домене Fc, стимулирующая гетеродимеризацию.
ФИГ.2. (А, В) Электрофорез в ДСН-ПААГ (4-12% Трис-ацетат (А) или 4-12% Бис/Трис (В), NuPage Invitrogen, окрашивание Кумасси) "1+1 IgG Crossfab (N-концевой), Fc(впадина) P329G LALA/Fc(выступ), дикий тип" (анти-MCSP/анти-huCD3) (см. SEQ ID NO:1, 2, 3 и 4), не восстановленный (А) и восстановленный (В). (С) Аналитическая эксклюзионная хроматография (Superdex 200 10/300 GL GE Healthcare; 2 мМ 3-(N-морфолино)пропансульфоновая кислота (MOPS) pH 7,3, 150 мМ NaCl, 0,02% (масс/об) NaCl; нанесение 50 мкг образца) "1+1 IgG Crossfab (N-концевой), Fc(впадина) P329G 1-А1-А/Fc(выступ) дикий тип" (анти-MCSP/анти-huCD3).
ФИГ.3. (А, В) Электрофорез в ДСН-ПААГ (4-12% Бис/Трис, NuPage Invitrogen, окрашивание Кумасси) "2+1 IgG Crossfab (N-концевой)" (анти-MCSP/анти-huCD3) (см. SEQ ID NO:1, 3, 4 и 5), не восстановленный (А) и восстановленный (В). (С) Аналитическая эксклюзионная хроматография (Superdex 200 10/300 GL GE Healthcare; 2 мМ MOPS pH 7,3, 150 мМ NaCl, 0,02% (масс/об) NaCl; нанесение 50 мкг образца) "2+1 IgG Crossfab (N-концевой)" (анти-MCSP/анти-huCD3).
ФИГ.4. (А, В) Электрофорез в ДСН-ПААГ (4-12% Бис/Трис, NuPage Invitrogen, окрашивание Кумасси) "2+1 IgG Crossfab (N-концевой), инвертированный" (анти-СЕА/анти-huCD3) (см. SEQ ID NO:3, 8, 9 и 10), не восстановленный (А) и восстановленный (В). (С) Аналитическая эксклюзионная хроматография (Superdex 200 10/300 GL GE Healthcare; 2 мМ MOPS pH 7,3, 150 мМ NaCl, 0,02% (масс/об) NaCl; нанесение 50 мкг образца) "2+1 IgG Crossfab (N-концевой), инвертированный" (анти-СЕА/анти-huCD3).
ФИГ.5. (А, В) Анализ "2+1 IgG Crossfab (С-концевой)" (анти-с-Met/анти-Her3) (см. SEQ ID NO:11, 12, 13, 14), не восстановленный (А) и восстановленный (В) в капиллярном электрофорезе (КЭ) - геле с додецилсульфатом натрия (ДСН).
ФИГ.6. Одновременное связывание биспецифических конструкций с доменом D3 MCSP человека и CD3γ(G4S)5CD3ε-AcTev-Fc(выступ)-Avi/Fc(впадина) человека. (А) Постановка анализа Biacore; (В) измерение "2+1 IgG Crossfab (N-концевой)".
ФИГ.7. Уровни различных цитокинов, измеренные в супернатанте цельной крови после обработки 1 нМ различных биспецифических конструкций CD3-MCSP ("2+1 IgG Crossfab (N-концевой)", "(scFv)2") или соответствующих контрольных IgG в присутствии (А, В) ли в отсутствие (С, D) опухолевых клеток Colo-38 в течение 24 часов.
ФИГ.8. Уровень поверхностной экспрессии позднего активационного маркера CD25 Т клеток CD8+ яванского макака от двух различных животных (яванский макак Нестор, яванский макак Нобу) после 43 часов инкубации с указанными концентрациями биспецифической конструкции "2+1 IgG Crossfab (N-концевой)" (направленной на CD3 яванского макака и MCSP человека), в присутствии или в отсутствие опухолевых клеток-мишеней MV-3, экспрессирующих MCSP человека (отношение эффекторных клеток к клеткам-мишеням Е:Т (от англ. "Effector: Target)=3:1). В качестве контролей использовали референсные IgG (IgG против CD3 яванского макака, IgG против MCSP человека) или нефизиологический стимул фитогемагглютинин (ФГА)-М.
ФИГ.9. Цитолиз (измеренный на основании высвобождения лактатдегидрогеназы (ЛДГ)) опухолевых клеток MDA-MB-435 при совместном культивировании с общими Т-клетками человека (отношение Е:Т=5:1) и активации в течение 20 часов различными концентрациями биспецифических молекул "2+1 IgG Crossfab (N-концевой)" и "(scFv)2" и соответствующих IgG.
ФИГ.10. Цитолиз (измеренный на основании высвобождения ЛДГ) опухолевых клеток MDA-MB-435 при совместном культивировании с общими Т-клетками человека (отношение Е:Т=5:1) и активации в течение 21 часа различными концентрациями биспецифических конструкций и соответствующими IgG. Сравнивали биспецифические конструкции CD3-MCSP "2+1 IgG Crossfab (N-концевой)" и "1+1 IgG Crossfab (N-концевой)", молекулу "(scFv)2" и соответствующие IgG.
ФИГ.11. Цитолиз (измеренный на основании высвобождения ЛДГ) huMCSP-положительных клеток меланомы MV-3 при совместном культивировании с мононуклеарными клетками периферической крови (МКПК) человека (отношение Е:Т=10:1), обработанными различными биспецифическими конструкциями CD3-MCSP ("2+1 IgG Crossfab (N-концевой)" и "(scFv)2") в течение приблизительно 26 часов.
ФИГ.12. Иллюстративные конфигурации биспецифических антигенсвязывающих молекул по изобретению, имеющих сшитую легкую цепь. (A) Иллюстрация молекулы "2+1 IgG Crossfab (N-концевой), сшитая легкая цепь". (B) Иллюстрация молекулы "1+1 IgG Crossfab (N-концевой), сшитая легкая цепь". (C) Иллюстрация молекулы "2+1 IgG Crossfab (N-концевой), инвертированный, сшитая легкая цепь". (D) Иллюстрация молекулы "1+1 IgG Crossfab (N-концевой), инвертированный, сшитая легкая цепь".
ФИГ.13. Анализы КЭ-ДСН. Электрофореграмма, показанная в виде электрофореза в ДСН-ПААГ "2+1 IgG Crossfab (N-концевой), сшитая легкая цепь" (дорожка 1: восстановленный, дорожка 2: не восстановленный).
ФИГ.14. Аналитическая эксклюзионная хроматография "2+1 IgG Crossfab (N-концевой), сшитая легкая цепь" (конечный продукт). Нанесение 20 мкг образца "2+1 IgG Crossfab (N-концевой), сшитая легкая цепь".
ФИГ.15. Цитолиз (измеренный на основании высвобождения ЛДГ) MCSP-положительных опухолевых клеток MV-3 при совместном культивировании с МКПК человека (отношение Е:Т=10:1), обработанных различными биспецифическими конструкциями CD3-MCSP в течение приблизительно 44 часов. МКПК человека выделяли из свежей крови здоровых волонтеров.
ФИГ.16. Цитолиз (измеренный на основании высвобождения ЛДГ) MCSP-положительных опухолевых клеток Colo-38 при совместном культивировании с МКПК человека (отношение Е:Т=10:1), обработанных различными биспецифическими конструкциями CD3-MCSP в течение приблизительно 22 часов. МКПК человека выделяли из свежей крови здоровых волонтеров.
ФИГ.17. Цитолиз (измеренный на основании высвобождения ЛДГ) MCSP-положительных опухолевых клеток Colo-38 при совместном культивировании с МКПК человека (отношение Е:Т=10:1), обработанных различными биспецифическими конструкциями CD3-MCSP в течение приблизительно 22 часов. МКПК человека выделяли из свежей крови здоровых волонтеров.
ФИГ.18. Цитолиз (измеренный на основании высвобождения ЛДГ) MCSP-положительных клеток WM266-4 при совместном культивировании с МКПК человека (отношение Е:Т=10:1), обработанных различными биспецифическими конструкциями CD3-MCSP в течение приблизительно 22 часов. МКПК человека выделяли из свежей крови здоровых волонтеров.
ФИГ.19. Уровень поверхностной экспрессии раннего активационного маркера CD69 (А) и позднего активационного маркера CD25 (В) на Т-клетках CD8+ человека после 22 часов инкубации с 10 наномоль (нМ), 80 пикомоль (пМ) или 3 пМ различных биспецифических конструкций CD3-MCSP в присутствии или в отсутствие MCSP-экспрессирующих опухолевых клеток-мишеней Colo-38 человека (отношение Е:Т=10:1).
ФИГ.20. Анализы КЭ-ДСН. (А) Электрофореграмма, показанная в виде электрофореза в ДСН-ПААГ 1+1 IgG Crossfab (N-концевой); обмен VL/VH (LC007/V9): а) не восстановленный, b) восстановленный. (В) Электрофореграмма, показанная в виде электрофореза в ДСН-ПААГ 1+1 CrossMab; обмен CL/CH1 (LC007/V9): а) восстановленный, b) не восстановленный. (С) Электрофореграмма, показанная в виде электрофореза в ДСН-ПААГ 2+1 IgG Crossfab (N-концевой), инвертированный; обмен CL/CH1 (LC007/V9): а) восстановленный, b) не восстановленный. (D) Электрофореграмма, показанная в виде электрофореза в ДСН-ПААГ 2+1 IgG Crossfab (N-концевой); обмен VL/VH (M4-3 ML2/V9): а) восстановленный, b) не восстановленный. (Е) Электрофореграмма, показанная в виде электрофореза в ДСН-ПААГ 2+1 IgG Crossfab (N-концевой); обмен CL/CH1 (M4-3 ML2/V9): а) восстановленный, b) не восстановленный. (F) Электрофореграмма, показанная в виде электрофореза в ДСН-ПААГ 2+1 IgG Crossfab (N-концевой), инвертированный; обмен CL/CH1 (CH1A1A/V9): а) восстановленный, b) не восстановленный.
ФИГ.21. Уровень поверхностной экспрессии раннего активационного маркера CD69 (А) и позднего активационного маркера CD25 (В) на Т-клетках CD4+ или CD8+ человека после 24 часов инкубации с указанными концентрациями конструкций CD3/MCSP "1+1 CrossMab", "1+1 IgG Crossfab (N-концевой)" и "2+1 IgG Crossfab (N-концевой)". Анализ был проведен в присутствии или в отсутствие клеток-мишеней MV-3, как указано.
ФИГ.22. Цитолиз (измеренный на основании высвобождения ЛДГ) опухолевых клеток MKN-45 (А) или LS-174T (В) при совместном культивировании с МКПК человека (отношение Е:Т=10:1) и активации в течение 28 часов различными концентрациями конструкции "2+1 IgG Crossfab (N-концевой), инвертированный (VL/VH)" по сравнению с конструкцией "2+1 IgG Crossfab (N-концевой), инвертированный (CL/CH1)".
ФИГ.23. Цитолиз (измеренный на основании высвобождения ЛДГ) опухолевых клеток WM266-4 при совместном культивировании с МКПК человека (отношение Е:Т=10:1) и активации в течение 26 часов различными концентрациями конструкции "2+1 IgG Crossfab (N-концевой) (VL/VH)" по сравнению с конструкцией "2+1 IgG Crossfab (N-концевой) (CL/CH1)".
ФИГ.24. Цитолиз (измеренный на основании высвобождения ЛДГ) опухолевых клеток MV-3 при совместном культивировании с МКПК человека (отношение Е:Т=10:1) и активации в течение 27 часов различными концентрациями конструкции "2+1 IgG Crossfab (N-концевой) (VH/VL)" по сравнению с конструкцией "2+1 IgG Crossfab (N-концевой) (CL/CH1)".
ФИГ.25. Цитолиз (измеренный на основании высвобождения ЛДГ) MCSP-положительных опухолевых клеток WM266-4 человека (А) или опухолевых клеток MV-3 (В) при совместном культивировании с МКПК человека (отношение Е:Т=10:1) и активации в течение 21 часа различными концентрациями конструкции "2+1 IgG Crossfab (N-концевой)", "1+1 CrossMab" и "1+1 IgG Crossfab (N-концевой)", как указано.
ФИГ.26. Связывание биспецифических конструкций с CD3 человека, экспрессируемым клетками Jurkat (А), или с СЕА человека, экспрессируемым клетками LS-174T (В), определенное с помощью флуоресцентно-активируемого клеточного сортинга (FACS; от англ. "Fluorescence Activated Cell Sorting"). В качестве контроля также оценивали эквивалентную максимальную концентрацию референсных IgG и фоновое окрашивание за счет меченого второго антитела (ФИТЦ (флуоресцеинизотиоцианат)-конъюгированного F(ab')2 фрагмента коза против человека AffiniPure, специфичного к фрагменту Fcγ, Jackson Immuno Research Lab # 109-096-098).
ФИГ.27. Связывание биспецифических конструкций с CD3 человека, экспрессируемым клетками Jurkat, или с MCSP человека, экспрессируемым опухолевыми клетками WM266-4 (В), определенное с помощью FACS.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Определения
В данной работе использованы термины, обычно используемые в данной области техники, если ниже не определено иное.
Как используют в данной работе, термин "антигенсвязывающая молекула" относится в его самом широком смысле к молекуле, специфично связывающей антигенную детерминанту. Примерами антигенсвязывающих молекул являются иммуноглобулины и их производные, например, фрагменты.
Термин "биспецифическая" означает, что антигенсвязывающая молекула способна к специфичному связыванию с двумя отдельными антигенными детерминантами. В характерном случае биспецифическая антигенсвязывающая молекула содержит два антигенсвязывающих сайта, каждый из которых специфичен к различным антигенным детерминантам. В некоторых формах осуществления биспецифическая антигенсвязывающая молекула способна к одновременному связыванию двух антигенных детерминант, в частности, двух антигенных детерминант, экспрессирующихся на двух отдельных клетках.
Как используют в данной работе, термин "антигенная детерминанта" является синонимом "антигена" и "эпитопа" и относится к сайту (например, к непрерывному отрезку из аминокислот или к конформационной конфигурации, состоящей из различных участков несмежных аминокислот) на полипептидной макромолекуле, с которым связывается антигенсвязывающий фрагмент, образуя комплекс антигенсвязывающего фрагмента с антигеном. Полезные антигенные детерминанты могут быть обнаружены, например, на поверхностях опухолевых клеток, на поверхностях клеток, инфицированных вирусами, на поверхностях других пораженных клеток, на поверхности иммунных клеток, в свободном виде в сыворотке крови и/или во внеклеточном матриксе (ВКМ). Белки, относящиеся в данной работе к антигенам (например, ассоциированный с меланомой протеогликановый хондроитинсульфат (MCSP; от англ. "melanoma-associated chondroitin sulphate proteoglycan"), белок, активирующий фибробласты (FAP; от англ. "Fibroblast Activation Protein"), карцино-эмбриональный антиген (СЕА; от англ. "carcino-embryonic antigen"), рецептор эпидермального фактора роста (EGFR; от англ. "epidermal growth factor receptor"), CD33, CDS, c-Met, Her3) может представлять собой любую нативную форму белков из любого источника позвоночных, включая млекопитающих, таких как приматы (например, людей), и грызуны (например, мыши и крысы), если не указано иное. В конкретной форме осуществления антиген представляет собой белок человека. Если в данной работе сделана ссылка на конкретный белок, этот термин включает как "полноразмерный", непроцессированный белок, так и любую форму белка, являющуюся результатом процессинга в клетке. Этот термин также включает встречающиеся в природе варианты белка, например, варианты сплайсинга или аллельные варианты. Иллюстративные белки человека, полезные в качестве антигенов, включают, но не ограничены ими, следующие белки: ассоциированный с меланомой протеогликановый хондроитинсульфат (MCSP), также известный как протеогликановый хондроитинсульфат 4 (UniProt no. Q6UVK1, Национальный центр биотехнологической информации (NCBI; от англ. "National Center for Biotechnology Information"), № доступа NP_001888); белок, активирующий фибробласты (FAP), также известный как сепраза (Seprase; от англ. "Surface Expressing Protease" Uni Prot №№ Q12884, Q86Z29, Q99998, № доступа NCBI NP_004451); карцино-эмбриональный антиген (СЕА), также известный как молекула клеточной адгезии 5, родственная карцино-эмбриональному антигену (UniProt № Р06731, № доступа NCBI NP_004354); CD33, также известный как gp67 или Siglec-3 (UniProt № Р20138, №№ доступа NCBI NP_001076087, NP_001171079); рецептор эпидермального фактора роста (EGFR), также известный как ErbB-1 или Her1 (UniProt № Р0053, №№ доступа NCBI NP_958439, NP_958440), CD3, в частности, эпсилон-субъединица CD3 (UniProt № Р07766, № доступа NCBI NP_000724); c-Met, также известный как рецептор фактора роста гепатоцитов (UniProt № Р08581, №№ доступа NCBI NP_000236, NP_001120972) и Her3, также известный как ErbB-3 (UniProt № Р21860, №№ доступа NCBI NP_001973, NP_001005915). В некоторых формах осуществления биспецифическая антигенсвязывающая молекула по изобретению связывается с эпитопом первого антигена или второго антигена, являющимся консервативным среди первого антигена или второго антигена от различных видов.
Под "специфичным связыванием" подразумевают, что связывание является селективным для антигена, и его можно отличить от нежелательных или неспецифичных взаимодействий. Способность антитела к связыванию со специфичной антигенной детерминантой можно измерить либо посредством твердофазного иммуноферментного анализа (ELISA; от англ. "enzyme-linked immunosorbent assay"), либо другими методами, знакомыми специалистам в данной области техники, например, методом поверхностного плазменного резонанса (SPR; от англ. "surface plasmon resonance") (анализируемого на приборе BIAcore) (Liljeblad et al., Glyco J 17, 323-329 (2000)), и традиционными анализами связывания (Heeley, Endocr Res 28, 217-229 (2002)). В одной форме осуществления степень связывания антитела с посторонним белком составляет менее чем приблизительно 10% связывания антитела с антигеном, измеренного, например, с помощью SPR. В некоторых формах осуществления антитело или его фрагмент, связывающийся с антигеном, имеет константу диссоциации (KD), составляющую не более 1 мкМ, не более 100 нМ, не более 10 нМ, не более 1 нМ, не более 0,1 нМ, не более 0,01 нМ или не более 0,001 нМ (например, 10-8 М или менее, например, от 10-8 М до 10-13 M, например, от 10-9 М до 10-13 М).
"Сродство" относится к силе общей суммы нековалентных взаимодействий между отдельным связывающим сайтом молекулы (например, рецептора) и его партнером связывания (например, лигандом). Если не указано иное, как используют в данной работе, "связывающее сродство" относится к собственному связывающему сродству, отражающему взаимодействие 1:1 между членами связывающей пары (например, антигенсвязывающего фрагмента и антигена или рецептора и его лиганда). Сродство молекулы Х к ее партнеру Y может быть, как правило, представлено константой диссоциации (KD), представляющей собой отношение констант скоростей диссоциации и ассоциации (koff и kon, соответственно). Таким образом, эквивалентные значения сродства могут включать различные константы скоростей, если отношение констант скоростей остается тем же. Сродство можно измерить хорошо разработанными методами, известными в данной области техники, включающими методы, раскрытые в данной работе. Конкретным способом измерения сродства является поверхностный плазменный резонанс (SPR).
Термин "валентный", как используют в данной работе, обозначает наличие определенного числа антигенсвязывающих сайтов в антигенсвязывающей молекуле. Как таковой, термин "моновалентное связывание с антигеном" обозначает наличие одного (и не более чем одного) антигенсвязывающего сайта, специфичного к антигену антигенсвязывающей молекулы.
"Антигенсвязывающий сайт" относится к сайту, то есть к одному или более аминокислотных остатков антигенсвязывающей молекуле, обеспечивающему взаимодействие с антигеном. Например, антигенсвязывающий сайт антитела включает аминокислотные остатки из участков, определяющих комплементарность (CDR; от англ. "complementarity determining regions"). Нативная молекула иммуноглобулина в характерном случае имеет два антигенсвязывающих сайта, Fab фрагмент в характерном случае имеет единственный антигенсвязывающий сайт.
Как используют в данной работе, термин "антигенсвязывающий фрагмент" относится к молекуле полипептида, специфично связывающейся с антигенной детерминантой. Антигенсвязывающие фрагменты включают антитела и их фрагменты, как дополнительно определено в данной работе. Конкретные антигенсвязывающие фрагменты включают антигенсвязывающий домен антитела, содержащий вариабельную область тяжелой цепи антитела и вариабельную область легкой цепи антитела. В некоторых формах осуществления антигенсвязывающие фрагменты могут включать константные области антитела, как дополнительно определено в данной работе и известно в данной области техники. Полезные константные области тяжелой цепи включают любой из пяти изотипов: α, δ, ε, γ или μ. Полезные константные области легкой цепи включают любой из двух изотипов: κ и λ.
Как используют в данной работе, термины "первый" и "второй" по отношению к Fab фрагментам и т.д. используют для удобства различения, когда имеется более чем один тип фрагмента. Использование этих терминов не предназначено для определения конкретного порядка или ориентации биспецифической антигенсвязывающей молекулы, если явным образом не указано иное.
Как используют в данной работе, термин "одноцепочечный" относится к молекуле, включающей мономеры аминокислот, линейно связанные пептидными связями. Под одноцепочечным Fab фрагментом подразумевают молекулу Fab, в которой легкая цепь Fab и тяжелая цепь Fab соединены пептидным линкером с образованием одной пептидной цепи.
Термин "молекула иммуноглобулина" относится к белку, имеющему структуру встречающегося в природе антитела. Например, иммуноглобулины класса IgG представляют собой гетеротетрамерные гликопротеины, имеющие массу приблизительно 150000 Дальтон, состоящие из двух легких цепей и двух тяжелых цепей, связанных дисульфидными связями. От N-конца к С-концу каждая тяжелая цепь имеет вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которой следует шарнирная область (HR; от англ. "hinge region") и три константных домена (СН1, СН2 и CH3), также называемых константной областью тяжелой цепи. В случае иммуноглобулина класса IgE тяжелая цепь дополнительно имеет домен СН4. Следовательно, тяжелая цепь иммуноглобулина представляет собой полипептид, состоящий в направлении от N-конца к С-концу из следующих доменов: VH-CH1-HR-CH2-CH3-(CH4). Аналогично от N- к С-концу каждая легкая цепь имеет вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, за которым следует константный легкий домен (CL), также называемый константной областью легкой цепи. Следовательно, легкая цепь иммуноглобулина представляет собой полипептид, состоящий в направлении от N-конца к С-концу из следующих доменов: VL-CL. Тяжелая цепь иммуноглобулина может быть отнесена к одному из пяти типов, называемых α (IgA), δ (IgD), ε (IgE), γ (IqG) или μ (IgM), причем, некоторые из могут быть дополнительно разделены на подтипы, например, γ1 (IgG1), γ2 (IgG2), γ3 (IgG3), γ4 (IgG4), α1 (IgA1) и α2 (IgA2). Легкая цепь иммуноглобулина может быть отнесена к одному из двух типов, называемых каппа (κ) и лямбда (λ), на основе аминокислотной последовательности ее константного домена. Иммуноглобулин по существу состоит из двух Fab фрагментов и домена Fc, связанных посредством шарнирной области иммуноглобулина.
Термин "антитело" в данной работе используют в самом широком смысле, и он включает различные структуры антитела, включающие, но не ограниченные ими, моноклональные антитела, поликлональные антитела и фрагменты антител, если они проявляют желаемую антигенсвязывающую активность.
"Фрагмент антитела" относится к молекуле, отличающейся от интактного антитела, включающей участок интактного антитела, связывающий антиген, с которым связывается интактное антитело. Примеры фрагментов антитела включают, но не ограничены ими, Fv, Fab, Fab', Fab'-SH, F(ab')2, диатела, линейные антитела, одноцепочечные молекулы антител (например, scFv) и однодоменные антитела. Обзор некоторых фрагментов антител см. в статье Hudson et al., Nat Med 9, 129-134 (2003). Обзор фрагментов scFv см., например, в статье Pluckthun в The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp.269-315 (1994); см. также документ WO 93/16185; и патенты US №№5571894 и 5587458. Обсуждение Fab и F(ab')2 фрагментов, включающих остатки эпитопа связывания рецептора реутилизации, и имеющих увеличенный период полувыведения in vivo, см. в патенте US №5869046. Диатела представляют собой фрагменты антител, имеющие два антигенсвязывающих сайта, которые могут быть бивалентными или биспецифическими. См., например, следующие документы: ЕР 404097; WO 1993/01161; Hudson et al., Nat Med 9, 129-134 (2003); и Hollinger et al., Proc Natl Acad Sci USA 90, 6444-6448 (1993). Триатела и тетратела также раскрыты в статье Hudson et al., Nat Med 9, 129-134 (2003). Однодоменные антитела представляют собой фрагменты антител, содержащие полноразмерный вариабельный домен тяжелой цепи антитела или его участок, либо полноразмерный вариабельный домен легкой цепи антитела или его участок. В некоторых формах осуществления однодоменное антитело представляет собой однодоменное антитело человека (Domantis, Inc., Waltham, MA; см., например, патент US №6248516 В1). Фрагменты антител могут быть получены различными методами, включающими, но не ограниченными ими, протеолитическое расщепление интактного антитела, а также продуцирование рекомбинантными клетками-хозяевами (например, Е. coli или фагом), как раскрыто в данной работе.
"Fab фрагмент" относится к белку, состоящему из доменов VH и СН1 тяжелой цепи ("тяжелая цепь Fab") и доменов VL и CL легкой цепи ("легкая цепь Fab") иммуноглобулина. Fab фрагмент, слитый с другим белком, в его немодифицированной форме слит при С- или N-конце его тяжелой цепи. Следовательно, если вариабельные домены VH и VL заменены друг другом, Fab фрагмент слит при С-конце домена СН1 или при N-конце домена VL. Аналогично, если константные домены СН1 и CL заменены друг другом, Fab фрагмент слит при С-конце его домена CL или при N-конце домена VH, и, если полноразмерный Fab тяжелая цепь (VH-CH1) и легкая цепь Fab (VL-CL) заменены друг другом, Fab фрагмент слит при С- или N-конце его легкой цепи.
Под "слитым" подразумевают, что компоненты (например, Fab фрагмент и субъединицы домена Fc) связаны пептидными связями, либо непосредственно, либо посредством одного или более пептидных линкеров.
Термин "антигенсвязывающий домен" относится к участку антитела, включающему участок, специфично связывающийся с участком антигена и комплементарную ему. Антигенсвязывающий домен может быть представлен, например, одним или более вариабельных доменов антитела (также называемых вариабельными областями антитела). В частности, антигенсвязывающий домен включает вариабельную область легкой цепи антитела (VL) и вариабельную область тяжелой цепи антитела (VH).
Термин "вариабельная область" или "вариабельный домен" относится к домену тяжелой или легкой цепи антитела, вовлеченному в связывание антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела, как правило, имеют сходные структуры, причем, каждый домен содержит четыре консервативных каркасных участка (FR; от англ. "framework regions") и три гипервариабельных участка (HVR; от англ. "hypervariable regions"). См., например, Kindt et al., Kuby Immunology, 6th ed., W.H. Freeman and Co., page 91 (2007). Одного домена VH или VL может быть достаточно для придания антигенсвязывающей специфичности.
Термин "гипервариабельный участок" или "HVR", как используют в данной работе, относится к каждому из участков вариабельного домена антитела, который является гипервариабельным по последовательности и/или образует петли определенной структуры ("гипервариабельные петли"). Как правило, четырехцепочечные антитела содержат шесть HVR: три в VH (Н1, Н2, Н3) и три в VL (L1, L2, L3). HVR, как правило, содержат аминокислотные остатки из гипервариабельных петель и/или из участков, определяющих комплементарность (CDR), причем, последние обладают самой высокой вариабельностью последовательности и/или вовлечены в распознавание антигеном. За исключением CDR1 в VH, CDR, как правило, содержат аминокислотные остатки, образующие гипервариабельные петли. Гипервариабельные участки (HVR) также называют "участками, определяющими комплементарность" (CDR), и эти термины в данной работе используют взаимозаменяемо в отношении участков вариабельной области, образующих антигенсвязывающие участки. Данный конкретный участок описан авторами Kabat et al., U.S. Dept. of Health and Human Services, Sequences of Proteins of Immunological Interest (1983) и Chothia et al., J Mol Biol 196:901-917 (1987), где определения включают перекрывание подгрупп аминокислотных остатков, когда их сравнивают против каждой другой. Тем не менее, применение любого определения для отнесения к CDR антитела или его вариантов предназначено для включения в объем термина, определенного и используемого в данной работе. Соответствующие аминокислотные остатки, входящие в CDR, как определено в каждой из цитируемых выше ссылок, представлены ниже в таблице 1 в качестве сравнения. Точные числа остатков, входящих в конкретный CDR, варьируют в зависимости от последовательности и размера CDR. Специалисты в данной области техники могут рутинно определить, какие остатки составляют конкретный CDR, на основании аминокислотной последовательности вариабельной области антитела.
Kabat et al. также определили систему нумерации для последовательностей вариабельной области, применимую к любому антителу. Обычный специалист в данной области техники может однозначно применить данную систему "нумерации Kabat" к любой последовательности вариабельной области, не опираясь на какие-либо экспериментальные данные, кроме самой последовательности. Как используют в данной работе, "нумерация Kabat" относится к системе нумерации, представленной Kabat et al., U.S. Dept. of Health and Human Services, "Sequence of Proteins of Immunological Interest" (1983). Если не указано иное, ссылки на нумерацию конкретных положений аминокислотных остатков в вариабельной области антитела соответствуют системе нумерации Kabat.
Полипептидные последовательности перечня последовательностей не пронумерованы в соответствии с системой нумерации Kabat. Тем не менее, в пределах компетенции обычного специалиста в данной области техники находится преобразование нумерации последовательностей Перечня последовательностей в нумерацию Kabat.
"Каркас" или "FR" относится к остаткам вариабельного домена, отличающимся от остатков гипервариабельного участка (HVR). FR вариабельного домена, как правило, состоит из четырех доменов FR: FR1, FR2, FR3 и FR4.
Соответственно, последовательности HVR и FR, как правило, выглядят в виде следующей последовательности в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
"Класс" антитела или иммуноглобулина относится к типу константного домена или константной области, которой обладает его тяжелая цепь. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, lgG4, IgA1 и IgA2. Константные домены тяжелой цепи, соответствующие различным классам иммуноглобулинов, называют α, δ, ε, γ и μ, соответственно.
Термин "домен Fc" или "участок Fc" в данной работе используют для определения C-концевого участка тяжелой цепи иммуноглобулина, составляющего по меньшей мере часть константной области. Этот термин включает нативную последовательность Fc и варианты участков Fc. Хотя границы участка Fc тяжелой цепи IgG могут слегка варьировать, участок Fc тяжелой цепи IgG человека обычно определяют как имеющий протяженность от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. Тем не менее, C-концевой лизин (Lys447) участка Fc может присутствовать или отсутствовать. Если в данной работе не указано иное, нумерация аминокислотных остатков в участке Fc или константной области соответствует системе нумерации Евросоюза (ЕС), также называемой индексом ЕС, как описано в кн. Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991. "Субъединица" домена Fc, как используют в данной работе, относится к одному из двух полипептидов, образующих димерный домен Fc, то есть к полипептиду, содержащему C-концевые константные области тяжелой цепи иммуноглобулина, способные к стабильной самоассоциации. Например, субъединица домена Fc IgG содержит константную область СН2 IgG и CH3 IgG.
"Модификация, стимулирующая ассоциацию первой и второй субъединицы домена Fc" представляет собой манипуляцию с пептидным каркасом или посттрансляционные модификации субъединицы домена Fc, уменьшающие или предотвращающие ассоциацию полипептида, содержащего субъединицу домена Fc, с идентичным полипептидом с образованием гомодимера. Модификация, стимулирующая ассоциацию, как используют в данной работе, конкретно включает отдельные модификации, произведенные в каждой из двух субъединиц домена Fc, ассоциация которых желательна (то есть в первой и второй субъединице домена Fc), где эти модификации комплементарны друг другу, чтобы стимулировать ассоциацию двух субъединиц домена Fc. Например, модификация, стимулирующая ассоциацию, может изменять структуру или заряд одной или обеих субъединиц домена Fc таким образом, чтобы сделать их ассоциацию стерически или электростатически благоприятной, соответственно. Таким образом, (гетеро)димеризация происходит между полипептидом, содержащим первую субъединицу домена Fc, и полипептидом, содержащим вторую субъединицу домена Fc, которые могут быть неидентичными в том смысле, что другие компоненты, слитые с каждой из субъединиц (например, Fab фрагменты) являются неодинаковыми. В некоторых формах осуществления модификация, стимулирующая ассоциацию, содержит мутацию аминокислоты в домене Fc, в частности, замену аминокислоты. В конкретной форме осуществления модификация, стимулирующая ассоциацию, содержит отдельную мутацию аминокислоты, в частности, замену аминокислоты, в каждой из двух субъединиц домена Fc.
Термин "эффекторные функции" относится к биологическим активностям, присущим участку Fc антитела, варьирующим в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают следующие функции: связывание C1q и комплементзависимую цитотоксичность (КЗЦ), связывание рецептора Fc, антителозависимую клеточно-опосредованную цитотоксичность (АЗКЦ), антителозависимый клеточный фагоцитоз (АЗКФ), секрецию цитокинов, захват антигена антигенпрезентирующими клетками, опосредованный иммунным комплексом, понижающую регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора) и активацию В клеток.
Как используют в данной работе, термины "конструировать, сконструированный, конструирование" рассматривают как включающие любую манипуляцию с пептидным каркасом или посттрансляционные модификации природного или рекомбинантного полипептида или его фрагмента. Конструирование включает модификации аминокислотной последовательности, паттерна гликозилирования или группы боковой цепи индивидуальных аминокислот, а также комбинации этих методов.
Термин "мутация аминокислоты", как используют в данной работе, понимают как включающий замены, делеции, инсерции и модификации аминокислот. Для получения конечной конструкции может быть произведена любая комбинация замены, делеции, инсерции и модификации при условии, что конечная конструкция обладает желаемыми характеристиками, например, сниженным связыванием с рецептором Fc или повышенной ассоциацией с другим пептидом. Делеции и инсерции аминокислотной последовательности включают амино- и/или карбокси-концевые делеции и инсерции аминокислот. Конкретные мутации аминокислот представляют собой замены аминокислот. С целью изменения, например, характеристик связывания участка Fc особенно предпочтительны неконсервативные замены аминокислот, то есть замену одной аминокислоты другой аминокислотой, обладающей отличающимися структурными и/или химическими свойствами. Замены аминокислот включают замену неприродными аминокислотами или производными природных аминокислот из двадцати стандартных аминокислот (например, 4-гидроксипролином, 3-метилгистидином, орнитином, гомосерином, 5-гидроксилизином). Мутации аминокислот могут быть получены, используя генетические или химические методы, хорошо известные в данной области техники. Генетические методы могут включать сайт-направленный мутагенез, полимеразную цепную реакцию (ПЦР), генный синтез и тому подобное. Рассмотрено, что способы изменения группы боковой цепи аминокислоты методами, отличающимися от генно-инженерных методов, такими как химическая модификация, могут быть также полезны. В данной работе для указания одной и той же мутации аминокислоты можно использовать различные обозначения.
Например, замена пролина в положении 329 домена Fc глицином может быть указана как 329G, G329, 6329, P329G или Pro329Gly.
Как используют в данной работе, термин "полипептид" относится к молекуле, состоящей из мономеров (аминокислот), линейно связанных амидными связями (также известными как пептидные связи). Термин "полипептид" относится к любой цепи из двух или более аминокислот и не относится к определенной длине продукта. Таким образом, пептиды, дипептиды, трипептиды, олигопептиды, "белок", "аминокислотная цепь" или любой другой термин, используемый как относящийся к цепи из двух или более аминокислот, включен в определение "полипептид", а термин "полипептид" можно использовать вместо любого из этих терминов или взаимозаменяемо с ними. Термин "полипептид" также следует относить к продуктам постэкспрессионных модификаций полипептида, включающих без ограничения гликозилирование, ацетилирование, фосфорилирование, амидирование, дериватизацию известных защитных/блокирующих групп, протеолитическое расщепление или модификацию неприродными аминокислотами. Полипептид может быть выделен из природного биологического источника или получен с помощью рекомбинантной технологии, но необязательно транслирован с указанной нуклеиново-кислотной последовательности. Он может быть получен любым путем, включая химический синтез. Полипептид по изобретению может иметь размер, составляющий приблизительно 3 или более, 5 или более, 10 или более, 20 или более, 25 или более, 50 или более, 75 или более, 100 или более, 200 или более, 500 или более, 1000 или более или 2000 или более аминокислот. Полипептиды могут иметь определенную трехмерную структуру, хотя они необязательно имеют такую структуру. Полипептиды, имеющие определенную трехмерную структуру, называют свернутыми, а полипептиды, не обладающие определенной трехмерной структурой, но которые вероятнее могут принимать большое число различных конформаций, называют развернутыми.
Под "изолированным" полипептидом или его вариантом, либо их производным, подразумевают полипептид, не находящийся в его природной среде. Определенный уровень очистки не требуется. Например, изолированный полипептид может быть извлечен из его нативного или природного окружения. Полипептиды и белки, продуцируемые рекомбинантным путем, экспрессированные в клетках-хозяевах, для целей изобретения считают изолированными, как и нативные или рекомбинантные полипептиды, выделенные, фракционированные, либо частично или по существу очищенные любым подходящим методом.
Под "изолированной" молекулой нуклеиновой кислоты или полинуклеотидом подразумевают молекулу нуклеиновой кислоты, ДНК или РНК, извлеченную из ее природного окружения. Например, для целей изобретения изолированным считают рекомбинантный полинуклеотид, кодирующий полипептид, содержащийся в векторе. Следующие примеры изолированного полинуклеотида включают рекомбинантные полинуклеотиды, поддерживающиеся в гетерологичных клетках-хозяевах, или очищенные (частично или по существу) полинуклеотиды в растворе. Изолированный полинуклеотид включает молекулу полинуклеотида, содержащуюся в клетках, которые обычно содержат молекулу полинуклеотида, но эта молекула полинуклеотида присутствует внехромосомно или в хромосомной локализации, отличающейся от ее природной хромосомной локализации. Изолированные молекулы РНК включают транскрипты РНК in vivo или in vitro по настоящему изобретению, а также формы положительной и отрицательной нити и двунитевые формы. Изолированные полинуклеотиды или нуклеиновые кислоты согласно настоящему изобретению дополнительно включают такие молекулы, полученные синтетическим путем. Кроме того, полинуклеотид или нуклеиновая кислота может представлять собой или включать регуляторный элемент, такой как промотор, сайт связывания рибосомы или терминатор транскрипции.
Термин "вектор" или "экспрессионный вектор" является синонимом "экспрессионной конструкции" и относится к молекуле ДНК, используемой для введения определенного гена, с которым она операционно сцеплена, в клетку-мишень и его экспрессии в этой клетке. Этот термин включает вектор в виде самореплицирующейся нуклеиново-кислотной структуры, а также вектор, включенный в геном клетки-хозяина, в которую он введен. Экспрессионный вектор по настоящему изобретению включает экспрессионную кассету. Экспрессионные векторы дают возможность транскрипции больших количеств стабильной мРНК. Когда экспрессионный вектор находится внутри клетки-мишени, молекула рибонуклеиновой кислоты или белка, кодируемая геном, продуцируется за счет клеточного механизма транскрипции и/или трансляции. В одной форме осуществления экспрессионный вектор по изобретению включает экспрессионную кассету, содержащую полинуклеотидные последовательности, кодирующие биспецифические антигенсвязывающие молекулы по изобретению или их фрагменты.
Термины "клетка-хозяин", "линия клеток-хозяев" и "культура клеток-хозяев" используют взаимозаменяемо и относят к клеткам, в которые введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают "трансформанты" и "трансформированные клетки", включающие первичную трансформированную клетку и потомство, имеющее происхождение от нее, без учета числа пересевов. Потомство может быть не полностью идентичным родительской клетке по содержанию нуклеиновых кислот, но может содержать мутации. В данное изобретение включено мутантное потомство, обладающее такой же функцией или биологической активностью, как исходно трансформированная клетка, на основании ее скрининга или селекции. Клетка-хозяин представляет собой любой тип клеточной системы, которую можно использовать для получения биспецифических антигенсвязывающих молекул по настоящему изобретению. Клетки-хозяева включают культивируемые клетки, например, культивируемые клетки млекопитающих, такие как клетки СНО, клетки BHK, клетки NS0, клетки SP2/0, клетки миеломы YO, клетки миеломы мыши Р3Х63, клетки PER, клетки PER.C6 или клетки гибридомы, клетки дрожжей, клетки насекомых и клетки растений, среди которых перечислены лишь немногие, а также клетки, содержащиеся в трансгенном животном, трансгенном растении или в культивируемых тканях растения или животного.
"Активирующий рецептор Fc" представляет собой рецептор Fc, который после захвата доменом Fc антитела вызывает события передачи сигнала, стимулирующего клетку, несущую рецептор, к осуществлению эффекторных функций. Активирующие рецепторы Fc человека включают FcγRIIIa (CD16a), FcγRI (CD64), FcγRIIa (CD32) и FcαRI (CD89).
Антителозависимая клеточно-опосредованная цитотоксичность (АЗКЦ) представляет собой иммунный механизм, приводящий к лизису клеток-мишеней, покрытых пленкой из антител, иммунными эффекторными клетками. Клетки-мишени представляют собой клетки, с которыми специфично связываются антитела или их производные, содержащие участок Fc, как правило, посредством участка белка, являющегося N-концевым к участку Fc. Как используют в данной работе, термин "сниженная (или повышенная) АЗКЦ" определяют как снижение (повышение) числа клеток-мишеней, лизируемых в данное время, при данной концентрации антитела в среде, окружающей клетки-мишени, посредством определенного выше механизма АЗКЦ и/или повышение (снижение) концентрации антитела в среде, окружающей клетки-мишени, требующейся для достижения лизиса данного числа клеток-мишеней в данное время посредством механизма АЗКЦ. Снижение (повышение) АЗКЦ относится к АЗКЦ, опосредованной тем же антителом, продуцируемым тем же типом клеток-хозяев, при использовании тех же стандартных способов продуцирования, очистки, приготовления препарата и хранения (известных специалистам в данной области техники), но не сконструированным. Например, снижение АЗКЦ, опосредованной антителом, содержащим в его домене Fc аминокислотную последовательность, снижающую АЗКЦ, относится к АЗКЦ, опосредованной тем же антителом без данной замены аминокислоты в домене Fc. Подходящие анализы для измерения АЗКЦ хорошо известны в данной области техники (см., например, публикацию РСТ № WO 2006/082515 или публикацию заявки на патент РСТ № РСТ/ЕР2012/055393).
"Эффективное количество" агента относится к количеству, необходимому, чтобы привести в результате к физиологическому изменению в клетке или ткани, в которую его вводят.
"Терапевтически эффективное количество" агента, например, фармацевтической композиции, относится к количеству, эффективному в дозировках и в течение периодов времени, необходимых для достижения желаемого терапевтического или профилактического результата. Терапевтически эффективное количество агента, например, устраняет, уменьшает, замедляет, сводит к минимуму или предотвращает вредные эффекты заболевания.
"Индивидуум" или "субъект" представляет собой млекопитающее. Млекопитающие включают, но не ограничены ими, домашних животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и нечеловекообразных приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В частности, индивидуум или субъект представляет собой человека.
Термин "фармацевтическая композиция" относится к препарату, находящемуся в форме, предоставляющей возможность для эффективности биологической активности содержащегося в ней активного ингредиента, и не содержащему дополнительных компонентов, являющихся неприемлемо токсичными для субъекта, которому вводят этот препарат.
"Фармацевтически приемлемый носитель" относится к ингредиенту в фармацевтической композиции, отличающемуся от активного ингредиента и нетоксичному для субъекта. Фармацевтически приемлемый носитель включает, но не ограничен ими, буфер, наполнитель, стабилизатор или консервант.
Как используют в данной работе, "лечение" (и его грамматические варианты, такие как "лечить" или "лечащий") относится к клиническому вмешательству при попытке изменить естественное течение заболевания у индивидуума, подлежащего лечению, и его можно проводить либо для профилактики, либо в ходе клинической патологии. Желаемые эффекты лечения включают, но не ограничены ими, предотвращение возникновения или рецидива заболевания, частичное снятие симптомов, уменьшение каких-либо прямых или косвенных патологических последствий заболевания, предотвращение метастазов, снижение скорости прогрессирования заболевания, уменьшение интенсивности болезненного состояния или его временное облегчение и ремиссию или улучшение прогноза. В некоторых формах осуществления биспецифические антигенсвязывающие молекулы по изобретению применяют для замедления развития заболевания или замедления прогрессирования заболевания.
Термин "инструкция по применению" используют как относящийся к инструкциям, обычно включаемым в коммерческие упаковки терапевтических препаратов, содержащим информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предостережения, касающиеся применения таких терапевтических препаратов.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
В изобретении предложена биспецифическая антигенсвязывающая молекула, включающая первый Fab фрагмент, специфично связывающийся с первым антигеном, второй Fab фрагмент, специфично связывающийся со вторым антигеном, и домен Fc, состоящий из первой и второй субъединиц, способных к стабильной ассоциации; причем,
a) биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым и/или вторым антигеном,
b) первый Fab фрагмент, второй Fab фрагмент и первая субъединица домена Fc слиты друг с другом, и
c) в первом и/или втором Fab фрагменте произведена одна из следующих замен: (i) вариабельные домены VL и VH заменены друг другом, (ii) константные домены CL и СН1 заменены друг другом или (iii) оба вариабельных и константных домена VL-CL и VH-CH1 заменены друг другом при условии, что в первом и втором Fab фрагменте произведена не одна и та же замена.
Форматы биспецифической антигенсвязывающей молекулы
Компоненты биспецифической антигенсвязывающей молекулы могут быть слиты друг с другом в разнообразных конфигурациях. Иллюстративные конфигурации изображены на фиг.1.
В конкретных формах осуществления первый Fab фрагмент слит на его С-конце с N-концом второго Fab фрагмента, который, в свою очередь, слит на его С-конце с N-концом первой субъединицы домена Fc (см. примеры на фиг.1А и 1В). В одной такой форме осуществления второй Fab фрагмент слит с первой субъединицей домена Fc посредством шарнирной области иммуноглобулина. В следующей такой форме осуществления первый Fab фрагмент слит со вторым Fab фрагментом посредством пептидного линкера.
В одной форме осуществления первый Fab фрагмент слит на С-конце его тяжелой цепи с N-концом тяжелой цепи второго Fab фрагмента, который, в свою очередь, слит на С-конце его тяжелой цепи с (N-концом первой субъединицы домена Fc.
В других формах осуществления второй Fab фрагмент слит на его С-конце с N-концом первого Fab фрагмента, который, в свою очередь, слит на его С-конце с N-концом первой субъединицы домена Fc (см. пример на фиг.1C). В одной такой форме осуществления первый Fab фрагмент слит с первой субъединицей домена Fc посредством шарнирной области иммуноглобулина. В следующей такой форме осуществления второй Fab фрагмент слит с первым Fab фрагментом посредством пептидного линкера. В одной форме осуществления второй Fab фрагмент слит на С-конце его тяжелой цепи с N-концом тяжелой цепи первого Fab фрагмента, который, в свою очередь, слит на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc.
В некоторых формах осуществления, где либо первый Fab фрагмент слит на С-конце его тяжелой цепи с N-концом тяжелой цепи второго Fab фрагмента, который, в свою очередь, слит на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc, либо второй Fab фрагмент слит на С-конце его тяжелой цепи с N-концом тяжелой цепи первого Fab фрагмента, который, в свою очередь, слит на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc, дополнительно легкая цепь Fab первого Fab фрагмента и легкая цепь Fab второго Fab фрагмента слиты друг с другом, необязательно посредством пептидного линкера (см. примеры на фиг.12).
Согласно этим формам осуществления два Fab фрагмента, имеющих различную специфичность, слиты друг с другом, и один из них, в свою очередь, слит с субъединицей домена Fc. Данная конфигурация дает возможность для геометрии (например, расстояния, угла между Fab фрагментами), отличающейся от классического формата биспецифического иммуноглобулина, в котором два Fab фрагмента молекулы иммуноглобулина имеют различные специфичности. Например, авторы изобретения обнаружили, что данная конфигурация является более подходящей, чем классический формат биспецифического иммуноглобулина, для имитации иммунологической конъюгации между Т-клеткой и клеткой-мишенью, которая необходима, если биспецифическую антигенсвязывающую молекулу следует применять для захвата и перенаправления Т клеток (данные не представлены).
В других формах осуществления, второй Fab фрагмент is fused на его С-конце с N-концом первой субъединицы домена Fc, который, в свою очередь, слит на его С-конце с N-концом первого Fab фрагмента (см. пример на фиг.1D). В одной такой форме осуществления второй Fab фрагмент слит с первой субъединицей домена Fc посредством шарнирной области иммуноглобулина. В следующей такой форме осуществления первый Fab фрагмент слит с первой субъединицей домена Fc посредством пептидного линкера. В одной форме осуществления второй Fab фрагмент слит на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc, который, в свою очередь, слит на его С-конце с N-концом тяжелой цепи первого Fab фрагмента. Согласно этим формам осуществления два Fab фрагмента, имеющих различную специфичность, слиты с двумя концами субъединицы домена Fc. Опять же, данная конфигурация дает возможность для отличающейся геометрии, которая может иметь преимущество для конкретного применения. В одной форме осуществления биспецифическая антигенсвязывающая молекула по существу состоит из первого Fab фрагмента, второго Fab фрагмента, домена Fc и необязательно одного или более пептидных линкеров.
Биспецифическая антигенсвязывающая молекула согласно изобретению обеспечивает моновалентное связывание по меньшей мере с одним из антигенов, с которыми она связывается. Моновалентное связывание важно, например, в случаях, где следует ожидать интернализации антигена-мишени после связывания антигенсвязывающей молекулы, обладающей высоким сродством. В таких случаях присутствие более чем одного антигенсвязывающего фрагмента, специфичного к антигену-мишени, может усилить интернализацию антигена, посредством этого снижая его доступность. Кроме того, моновалентное связывание существенно, если нежелательно перекрестное связывание антигена-мишени. Например, в биспецифических антигенсвязывающих молекулах для захвата и перенаправления Т клеток бивалентное связывание с активирующим Т-клеточным антигеном, таким как CD3, может привести к активации Т-клетки даже в отсутствие клеток-мишеней.
Тем не менее, в других случаях может быть желательным бивалентное связывание, например, для повышения связывающего сродства, оптимизации направления к сайту-мишени или предоставления возможности перекрестного связывания антигена-мишени.
Соответственно, в конкретных формах осуществления биспецифическая антигенсвязывающая молекула включает третий Fab фрагмент, специфично связывающийся с первым или вторым антигеном. В одной форме осуществления третий Fab фрагмент слит со второй субъединицей домена Fc. В более конкретной форме осуществления третий Fab фрагмент слит на его С-конце с N-концом второй субъединицы домена Fc. В еще более конкретной форме осуществления третий Fab фрагмент слит на С-конце его тяжелой цепи с N-концом второй субъединицы домена Fc. В одной форме осуществления третий Fab фрагмент слит со второй субъединицей домена Fc посредством шарнирной области иммуноглобулина. В одной форме осуществления третий Fab фрагмент специфично связывается со вторым антигеном.
В некоторых формах осуществления второй Fab фрагмент, третий Fab фрагмент и домен Fc составляют часть молекулы иммуноглобулина. В формах осуществления, где третий Fab фрагмент специфично связывается со вторым антигеном, молекула иммуноглобулина представляет собой молекулу иммуноглобулина, специфично связывающуюся со вторым антигеном. В такой конкретной форме осуществления молекула иммуноглобулина представляет собой молекулу иммуноглобулина класса IgG, более конкретно молекулу иммуноглобулина подкласса IgG1 или IgG4. В одной конкретной форме осуществления молекула иммуноглобулина представляет собой молекулу иммуноглобулина IgG4, содержащую замену аминокислоты в положении S228 (нумерация Kabat), в частности, замену аминокислоты S228P. Эта замена аминокислоты снижает обмен Fab фрагментами in vivo антител IgG4 (см. Stubenrauch et al., Drug Metabolism and Disposition 38, 84-91 (2010)). В одной форме осуществления молекула иммуноглобулина представляет собой молекулу иммуноглобулина человека. В одной форме осуществления биспецифическая антигенсвязывающая молекула по существу состоит из первого Fab фрагмента, специфично связывающегося с первым антигеном, молекулы иммуноглобулина, специфично связывающейся со вторым антигеном и необязательно одного или более пептидных линкеров.
Согласно некоторым из вышеописанных форм осуществления легкая цепь первого Fab фрагмента и легкая цепь второго Fab фрагмента слиты друг с другом, необязательно посредством пептидного линкера. В зависимости от конфигурации первого и второго Fab фрагмента легкая цепь первого Fab фрагмента может быть слита на ее С-конце с N-концом легкой цепи второго Fab фрагмента, либо легкая цепь второго Fab фрагмента может быть слита на ее С-конце с N-концом легкой цепи первого Fab фрагмента. Слияние легкой цепи первого и второго Fab фрагмента дополнительно снижает ошибочное спаривание неспаренных тяжелых и легких цепей Fab, а также снижает число плазмид, необходимых для экспрессии некоторых из биспецифических антигенсвязывающих молекул по изобретению.
Согласно любой из вышеописанных форм осуществления компоненты биспецифической антигенсвязывающей молекулы (например, Fab фрагменты, субъединицы домена Fc) могут быть сшиты непосредственно или посредством различных линкеров, в частности, пептидных линкеров, содержащих одну или более аминокислот, в характерном случае приблизительно 2-20 аминокислот, раскрытых в данной работе или известных из уровня техники. Подходящий неиммуногенный пептидный линкер включает, например, следующие пептидные линкеры: (G4S)n, (SG4)n, (G4S)n или G4(SG4)n, где n, как правило, представляет собой число от 1 до 10, в характерном случае от 2 до 4. Особенно подходящим пептидным линкером для слияния легких цепей первого и второго Fab фрагмента друг с другом является (G4S)2. Кроме того, пептидные линкеры могут содержать шарнирную область иммуноглобулина (или ее участок). Такой иллюстративный линкер представляет собой EPKSC(D)-(G4S)2 (SEQ ID NO:72 и 73). В частности, где Fab фрагмент сшит с N-концом субъединицы домена Fc, он может быть сшит посредством шарнирной области иммуноглобулина или ее участка с дополнительным пептидным линкером или без него.
В некоторых формах осуществления биспецифическая антигенсвязывающая молекула включает полипептид, в котором область VL имеет общую карбокси-концевую пептидную связь с областью СН1, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с тяжелой цепью иммуноглобулина (VH-CH1-HR-CH2-CH3-(CH4)). В некоторых из этих форм осуществления биспецифическая антигенсвязывающая молекула дополнительно включает легкую цепь антитела (VL-CL) и/или полипептид, в котором область VH имеет общую карбокси-концевую пептидную связь с областью CL. В некоторых из этих форм осуществления биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором область VH имеет общую карбокси-концевую пептидную связь с областью CL, которая, в свою очередь, общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, общую карбокси-концевую пептидную связь с легкой цепью Fab (VL-CL).
В других формах осуществления биспецифическая антигенсвязывающая молекула включает полипептид, в котором тяжелая цепь Fab (VH-CH1) имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью VL, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью СН1, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с субъединицей домена Fc, включающей шарнирную область иммуноглобулина (HR-CH2-CH3-(CH4)). В некоторых из этих форм осуществления биспецифическая антигенсвязывающая молекула дополнительно включает легкую цепь антитела (VL-CL) и/или полипептид, в котором область VH имеет общую карбокси-концевую пептидную связь с областью CL. В некоторых из этих форм осуществления биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором легкая цепь Fab (VL-CL) имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью VH, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью CL.
В некоторых формах осуществления биспецифическая антигенсвязывающая молекула включает полипептид, в котором область VH имеет общую карбокси-концевую пептидную связь с областью CL, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с тяжелой цепью иммуноглобулина (VH-CH1-HR-CH2-CH3-(CH4)). В некоторых из этих форм осуществления биспецифическая антигенсвязывающая молекула дополнительно включает легкую цепь антитела (VL-CL) и/или полипептид, в котором область VL имеет общую карбокси-концевую пептидную связь с областью СН1. В некоторых из этих форм осуществления биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором область VL имеет общую карбокси-концевую пептидную связь с областью СН1, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с легкой цепью Fab (VL-CL).
В других формах осуществления биспецифическая антигенсвязывающая молекула включает полипептид, в котором тяжелая цепь Fab (VH-CH1) имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью VH, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью CL, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с субъединицей домена Fc, включающей шарнирную область иммуноглобулина (HR-CH2-CH3-(CH4)). В некоторых из этих форм осуществления биспецифическая антигенсвязывающая молекула дополнительно включает легкую цепь антитела (VL-CL) и/или полипептид, в котором область VL имеет общую карбокси-концевую пептидную связь с областью СН1. В некоторых из этих форм осуществления биспецифическая антигенсвязывающая молекула дополнительно включает полипептид, в котором легкая цепь Fab (VL-CL) имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью VL, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью СН1.
В других формах осуществления биспецифическая антигенсвязывающая молекула включает полипептид, в котором тяжелая цепь иммуноглобулина ((VH-CH1-HR-CH2-CH3-(CH4)) имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью VH, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью CL.
В одной форме осуществления биспецифическая антигенсвязывающая молекула дополнительно включает тяжелую цепь иммуноглобулина ((VH-CH1-HR-CH2-CH3-(CH4)). В другой форме осуществления биспецифическая антигенсвязывающая молекула дополнительно включает субъединицу домена Fc, необязательно включающую шарнирную область антитела ((HR)-CH2-CH3-(СН4)). В некоторых из этих форм осуществления биспецифическая антигенсвязывающая молекула дополнительно включает легкую цепь антитела (VL-CL) и/или полипептид, в котором область VL имеет общую карбокси-концевую пептидную связь с областью СН1.
Fab фрагменты
Антигенсвязывающая молекула по изобретению является биспецифической, то есть она включает по меньшей мере два антигенсвязывающих фрагмента, способных к специфичному связыванию с двумя отдельными антигенными детерминантами. В конкретной форме осуществления биспецифическая антигенсвязывающая молекула способна к одновременному связыванию с двумя отдельными антигенными детерминантами. Согласно изобретению антигенсвязывающие фрагменты представляют собой Fab фрагменты (то есть антигенсвязывающие домены, состоящие из тяжелой и легкой цепи, где каждая включает вариабельную и константную область). В одной форме осуществления данные Fab фрагменты являются человеческими. В другой форме осуществления данные Fab фрагменты являются гуманизированными. В другой форме осуществления данные Fab фрагменты включают константные области тяжелой и легкой цепи человека.
Согласно изобретению по меньшей мере один из Fab фрагментов представляет собой "Crossfab" фрагмент, где вариабельные и/или константные домены тяжелой и легкой цепи Fab обмениваются. Такие модификации предотвращают ошибочное спаривание тяжелых и легких цепей от различных Fab фрагментов, что, таким образом, улучшает выход и чистоту биспецифической антигенсвязывающей молекулы по изобретению при рекомбинантном продуцировании. Иными словами, проблема ошибочного спаривания тяжелой и легкой цепи при продуцировании биспецифического антитела преодолевается за счет обмена вариабельных и/или константных доменов тяжелой и легкой цепи внутри одного или более Fab фрагментов биспецифической антигенсвязывающей молекулы, так что фрагменты, имеющие различную специфичность, не имеют идентичное расположение доменов и, следовательно, не претерпевают "взаимный обмен" легких цепей.
Возможные замены включают следующие замены: (i) вариабельные домены тяжелой и легкой цепи Fab (VH и VL) заменены друг другом; (ii) константные домены тяжелой и легкой цепи Fab (CH1 и CL) заменены друг другом; или (iii) тяжелая и легкая цепь Fab (VH-CH1 и VL-CL) заменены друг другом.
Для достижения желаемого результата, то есть предотвращения ошибочного спаривания тяжелых и легких цепей, имеющих различную специфичность, в Fab фрагментах, обладающих различной специфичностью, должна быть произведена не одна и та же замена. Например, в Fab фрагменте, специфично связывающемся с первым антигеном, могут обмениваться вариабельные домены тяжелой и легкой цепи, тогда как в Fab фрагменте, специфично связывающемся со вторым антигеном, может обмениваться константная область тяжелой и легкой цепи. В качестве другого примера в Fab фрагменте, специфично связывающемся с первым антигеном, замена может не быть произведена, тогда как в Fab фрагменте, специфично связывающемся со вторым антигеном, вариабельные домены тяжелой и легкой цепи могут обмениваться.
В конкретной форме осуществления одна и та же замена произведена в Fab фрагментах, имеющих одинаковую специфичность (то есть в Fab фрагментах, специфично связывающихся с одним и тем же антигеном). Замена не обязательно должна быть произведена во всех Fab фрагментах, содержащихся в биспецифической антигенсвязывающей молекуле. Например, в формах осуществления, где имеется три Fab фрагмента, достаточно произвести замену только в Fab фрагменте, имеющем специфичность, отличающуюся от специфичности двух других Fab фрагментов. В частности, в формах осуществления, где биспецифическая антигенсвязывающая молекула включает третий Fab фрагмент, связывающийся с первым антигеном, замена произведена только во втором Fab фрагменте. Аналогично в формах осуществления, где биспецифическая антигенсвязывающая молекула включает третий Fab фрагмент, связывающийся со вторым антигеном, замена произведена только в первом Fab фрагменте.
В конкретных формах осуществления замена произведена в первом Fab фрагменте. В одной такой форме осуществления дополнительные замены не произведены. В некоторых формах осуществления замена представляет собой замену вариабельных доменов VL и VH друг другом. В других формах осуществления замена представляет собой замену константных доменов CL и СН1 друг другом. В других формах осуществления замена представляет собой замену и вариабельных, и константных доменов VL-CL и VH-CH1 друг другом.
В конкретной форме осуществления биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым антигеном. В одной форме осуществления биспецифическая антигенсвязывающая молекула не включает одноцепочечный Fab фрагмент.
В конкретном аспекте в настоящем изобретении предложена биспецифическая антигенсвязывающая молекула, включающая первый Fab фрагмент, специфично связывающийся с первым антигеном, второй Fab фрагмент, специфично связывающийся со вторым антигеном, и домен Fc, состоящий из первой и второй субъединицы, способных к стабильной ассоциации; причем
a) биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым антигеном,
b) первый Fab фрагмент слит на его С-конце с N-концом второго Fab фрагмента, который, в свою очередь, слит на его С-конце с N-концом первой субъединицы домена Fc,
c) в первом Fab фрагменте константные домены CL и СН1 заменены друг другом, и
d) биспецифическая антигенсвязывающая молекула необязательно включает третий Fab фрагмент, специфично связывающийся со вторым антигеном, слитый на его С-конце с N-концом второй субъединицы домена Fc.
В другом аспекте в настоящем изобретении предложена биспецифическая антигенсвязывающая молекула, включающая первый Fab фрагмент, специфично связывающийся с первым антигеном, второй Fab фрагмент, специфично связывающийся со вторым антигеном, и домен Fc, состоящий из первой и второй субъединицы, способных к стабильной ассоциации; причем
a) биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым антигеном,
b) первый Fab фрагмент слит на его С-конце с N-концом второго Fab фрагмента, который, в свою очередь, слит на его С-конце с N-концом первой субъединицы домена Fc,
c) в первом Fab фрагменте вариабельные домены VL и VH заменены друг другом, и
d) биспецифическая антигенсвязывающая молекула необязательно включает третий Fab фрагмент, специфично связывающийся со вторым антигеном, и слитый на его С-конце с N-концом второй субъединицы домена Fc.
В следующем конкретном аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, включающая первый Fab фрагмент, специфично связывающийся с первым антигеном, второй Fab фрагмент, специфично связывающийся со вторым антигеном, и домен Fc, состоящий из первой и второй субъединицы, способных к стабильной ассоциации; причем
a) биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым антигеном,
b) второй Fab фрагмент слит на его С-конце с N-концом первого Fab фрагмента, который, в свою очередь, слит на его С-конце с N-концом первой субъединицы домена Fc,
c) в первом Fab фрагменте константные домены CL и СН1 заменены друг другом, и
d) биспецифическая антигенсвязывающая молекула необязательно включает третий Fab фрагмент, специфично связывающийся со вторым антигеном, и слитый на его С-конце с N-концом второй субъединицы домена Fc.
В следующем аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, включающая первый Fab фрагмент, специфично связывающийся с первым антигеном, второй Fab фрагмент, специфично связывающийся со вторым антигеном, и домен Fc, состоящий из первой и второй субъединицы, способных к стабильной ассоциации; причем
a) биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым антигеном,
b) второй Fab фрагмент слит на его С-конце с N-концом первого Fab фрагмента, который, в свою очередь, слит на его С-конце с N-концом первой субъединицы домена Fc,
c) в первом Fab фрагменте вариабельные домены VL и VH заменены друг другом, и
d) биспецифическая антигенсвязывающая молекула необязательно включает третий Fab фрагмент, специфично связывающийся со вторым антигеном, и слитый на его С-конце с N-концом второй субъединицы домена Fc.
В следующем аспекте в изобретении предложена биспецифическая антигенсвязывающая молекула, включающая первый Fab фрагмент, специфично связывающийся с первым антигеном, второй Fab фрагмент, специфично связывающийся со вторым антигеном, и домен Fc, состоящий из первой и второй субъединицы, способных к стабильной ассоциации; причем
a) биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым антигеном,
b) второй Fab фрагмент слит на его С-конце с N-концом первой субъединицы домена Fc, который, в свою очередь, слит на его С-конце с N-концом первого Fab фрагмента,
c) в первом Fab фрагменте константные домены CL и СН1 заменены друг другом, и
d) биспецифическая антигенсвязывающая молекула необязательно включает третий Fab фрагмент, специфично связывающийся со вторым антигеном, и слитый на его С-конце с N-концом второй субъединицы домена Fc.
Домен Fc
Домен Fc биспецифической антигенсвязывающей молекулы состоит из пары полипептидных цепей, включающих домены тяжелой цепи молекулы антитела. Например, домен Fc молекулы иммуноглобулина G (IgG) представляет собой димер, каждая субъединица которого включает константные домены тяжелой цепи СН2 и CH3 IgG. Две субъединицы домена Fc способны к стабильной ассоциации друг с другом. Биспецифическая антигенсвязывающая молекула по изобретению включает не более чем один домен Fc.
В одной форме осуществления согласно изобретению домен Fc биспецифической антигенсвязывающей молекулы представляет собой домен Fc IgG. В конкретной форме осуществления домен Fc представляет собой домен Fc IgG1. В другой форме осуществления домен Fc представляет собой домен Fc IgG4. В более конкретной форме осуществления домен Fc представляет собой домен Fc IgG4, содержащий замену аминокислоты в положении S228 (нумерация Kabat), в частности, замену аминокислоты S228P. Эта замена кислоты снижает обмен Fab фрагментов антител IgG4 in vivo (см. Stubenrauch et al., Drug Metabolism and Disposition 38, 84-91 (2010)). В следующей конкретной форме осуществления домен Fc является человеческим. Иллюстративная последовательность области Fc IgG1 человека представлена в SEQ ID NO:71.
Модификации домена Fc, стимулирующие гетеродимеризацию Биспецифические антигенсвязывающие молекулы согласно изобретению включают различные Fab фрагменты, слитые с одной или с другой из двух субъединиц домена Fc, таким образом, две субъединицы домена Fc в характерном случае содержатся в двух неидентичных полипептидных цепях. Рекомбинантная совместная экспрессия этих полипептидов и последующая димеризация приводит к нескольким возможным комбинациям этих двух полипептидов. Таким образом, чтобы улучшить выход и чистоту биспецифических антигенсвязывающих молекул при рекомбинантной продуцировании, предпочтительно вводить в домен Fc биспецифической антигенсвязывающей молекулы модификацию, стимулирующую ассоциацию желаемых полипептидов.
Соответственно, в конкретных формах осуществления домен Fc содержит модификацию, стимулирующую ассоциацию первой и второй субъединицы домена Fc. Модификация может присутствовать в первой субъединице домена Fc и/или второй субъединице домена Fc.
Сайт наиболее протяженного белок-белкового взаимодействия между двумя субъединицами домена Fc IgG человека представляет собой домен CH3 домена Fc. Таким образом, в одной форме осуществления модификация находится в домене CH3 домена Fc.
В конкретной форме осуществления модификация представляет собой так называемую модификацию "выступ-во-впадину", включающую модификацию "выступ" в одной из двух субъединиц домена Fc и модификацию "впадина" в другой из двух субъединиц домена Fc.
Технология выступ-во-впадину раскрыта, например, в следующих документах: US 5731168; US 7695936; Ridgway et al., Prot Eng 9, 617-621 (1996) и Carter, J Immunol Meth 248, 7-15 (2001). В целом этот способ включает введение выпячивания ("выступа") на поверхности контакта первого полипептида и соответствующего углубления ("впадины") на поверхности контакта второго полипептида, так чтобы выпячивание могло быть расположено в углублении, чтобы стимулировать образование гетеродимера и затруднить образование гомодимера. Выпячивания конструируют путем замены низкомолекулярных боковых цепей аминокислот с поверхности контакта первого полипептида боковыми цепями большего объема (например, тирозином или триптофаном). Компенсаторные углубления, имеющие размер, идентичный или подобный размеру выпячиваний, создают на поверхности контакта второго полипептида путем замены боковых цепей аминокислот большего размера боковыми цепями меньшего размера (например, аланином или треонином).
Соответственно, в конкретной форме осуществления в домене CH3 первой субъединицы домена Fc биспецифической антигенсвязывающей молекулы аминокислотный остаток заменен аминокислотным остатком, имеющим больший объем боковой цепи, создавая посредством этого выпячивание внутри домена CH3 первой субъединицы, которое может быть расположено в углублении внутри домена CH3 второй субъединицы, и в домене CH3 второй субъединицы домена Fc аминокислотный остаток заменен аминокислотным остатком, имеющим меньший объем боковой цепи, создавая посредством этого углубление внутри домена CH3 второй субъединицы, в котором может быть расположено выпячивание, находящееся внутри домена CH3 первой субъединицы.
Выпячивание и углубление можно создать путем изменения нуклеиновой кислоты, кодирующей полипептиды, например, путем сайт-специфического мутагенеза или путем пептидного синтеза.
В конкретной форме осуществления в домене CH3 первой субъединицы домена Fc остаток треонина в положении 366 заменен остатком триптофана (T366W), и в домене CH3 второй субъединицы домена Fc остаток тирозина в положении 407 заменен остатком валина (Y407V). В одной форме осуществления во второй субъединице домена Fc дополнительно остаток треонина в положении 366 заменен остатком серина (T366S), и остаток лейцина в положении 368 заменен остатком аланина (L368A).
В следующей форме осуществления в первой субъединице домена Fc дополнительно остаток серина в положении 354 заменен остатком цистеина (S354C), и во второй субъединице домена Fc дополнительно остаток тирозина в положении 349 заменен остатком цистеина (Y349C). Введение этих двух остатков цистеина приводит в результате к образованию дисульфидного мостика между двумя субъединицами домена Fc, дополнительно стабилизирующего димер (Carter, J Immunol Methods 248, 7-15 (2001)).
В альтернативной форме осуществления модификация, стимулирующая ассоциацию первой и второй субъединицы домена Fc, включает модификацию, опосредующую электростатические направляющие эффекты, например, как раскрыто в публикации РСТ WO 2009/089004. В целом данный способ включает замену одного или более аминокислотных остатков в области контакта двух субъединиц домена Fc заряженными аминокислотными остатками, так что образование гомодимера становится электростатически неблагоприятным, а гетеродимеризация становится элекстростатически благоприятной.
Модификации домена Fc, изменяющие связывание рецептора Fc и/или эффекторную функцию
В некоторых формах осуществления домен Fc сконструирован таким образом, что он обладает измененным связывающим сродством с рецептором Fc и/или измененной эффекторной функцией по сравнению с не сконструированным доменом Fc.
Связывание с рецепторами Fc можно легко определить, например, с помощью ИФА (ELISA) или поверхностного плазмонного резонанса (SPR), используя стандартное оборудование, такое как прибор BIAcore (GE Healthcare), а сами рецепторы Fc могут быть получены путем рекомбинантной экспрессии. Такой подходящий анализ связывания раскрыт в данной работе. Альтернативно связывающее сродство доменов Fc или биспецифических антигенсвязывающих молекул, включающих домен Fc, к рецепторам Fc можно оценивать, используя известные линии клеток, экспрессирующих конкретные рецепторы Fc, такие как клетки NK, экспрессирующие рецептор FcγIIIa.
Эффекторную функцию домена Fc или биспецифической антигенсвязывающей молекулы, содержащей домен Fc, можно измерить способами, известными в данной области техники. Подходящие анализы in vitro для оценки активности АЗКЦ представляющей интерес молекулы раскрыты в следующих документах: публикации РСТ № WO 2006/082515 или в заявке на патент РСТ № РСТ/ЕР2012/055393, полностью включенных в данное описание посредством ссылки. Полезные эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (МКПК) и естественные киллерные клетки (NK; от англ. "Natural Killer"). Альтернативно или дополнительно активность АЗКЦ представляющей интерес молекулы можно оценивать in vivo, например, в модели на животных, например, как раскрыто в Clynes et al., Proc Natl Acad Sci USA 95, 652-656 (1998).
В некоторых формах осуществления изменено связывание домена Fc с компонентом комплемента, в частности, с C1q. Соответственно, в некоторых формах осуществления, где домен Fc сконструирован таким образом, что он обладает измененной эффекторной функцией, данная измененная эффекторная функция включает измененную КЗЦ. Анализы связывания C1q можно проводить, чтобы определить, способна ли биспецифическая антигенсвязывающая молекула связывать C1q, и, следовательно, обладать активностью КЗЦ. См., например, анализ ELISA связывания C1q и C3c в WO 2006/029879 и WO 2005/100402. Чтобы оценить активацию комплемента, можно проводить анализ КЗЦ (см., например, Gazzano-Santoro et al., J Immunol Methods 202, 163 (1996); Cragg et al., Blood 101, 1045-1052 (2003); и Cragg and Glennie, Blood 103, 2738-2743 (2004)).
а) Сниженное связывание рецептора Fc и/или сниженная эффекторная функция
Домен Fc придает биспецифической антигенсвязывающей молекуле благоприятные фармакокинетические свойства, включающие длительный период полувыведения в сыворотке, вносящий вклад в высокое накопление в ткани-мишени, и благоприятное отношение распределения между тканью и кровью. В то же время, эти свойства могут, тем не менее, привести к нежелательному более вероятному направлению биспецифической антигенсвязывающей молекулы в клетки, экспрессирующие рецепторы Fc, чем в предпочтительные клетки, несущие антиген. Кроме того, активация биохимических путей передачи сигнала рецептора Fc может привести к высвобождению цитокинов и к тяжелым побочным эффектам при системном введении.
Соответственно, в конкретных формах осуществления домен Fc биспецифической антигенсвязывающей молекулы сконструирован таким образом, что он обладает сниженным связывающим сродством к рецептору Fc и/или сниженной эффекторной функцией по сравнению с не сконструированным доменом Fc. В одной такой форме осуществления домен Fc (или биспецифическая антигенсвязывающая молекула, включающая этот домен Fc) проявляет менее 50%, предпочтительно менее 20%, более предпочтительно менее 10% и наиболее предпочтительно менее 5% связывающего сродства к рецептору Fc по сравнению с не сконструированным доменом Fc (или биспецифической антигенсвязывающей молекулой, включающей не сконструированный домен Fc), и/или менее 50%, предпочтительно менее 20%, более предпочтительно менее 10% и наиболее предпочтительно менее 5% эффекторной функции по сравнению с не сконструированным доменом Fc (или биспецифической антигенсвязывающей молекулой, включающей не сконструированный домен Fc). В одной форме осуществления домен Fc (или биспецифическая антигенсвязывающая молекула, включающая этот домен Fc) по существу не связывается с рецептором Fc и/или не индуцирует эффекторную функцию. В конкретной форме осуществления рецептор Fc представляет собой рецептор Fcγ. В одной форме осуществления рецептор Fc представляет собой рецептор Fc человека. В одной форме осуществления рецептор Fc представляет собой активирующий рецептор Fc. В конкретной форме осуществления рецептор Fc представляет собой активирующий рецептор Fcγ человека, более конкретно FcγRIIIa, FcγRI или FcγRIIa человека, наиболее конкретно FcγRIIIa человека. В одной форме осуществления эффекторная функция представляет собой одну или более функций, выбранных из группы КЗЦ, АЗКЦ, АЗКФ и секреции цитокинов. В конкретной форме осуществления эффекторная функция представляет собой АЗКЦ. В одной форме осуществления домен Fc проявляет по существу аналогичное связывающее сродство к неонатальному рецептору Fc (FcRn) по сравнению с не сконструированным доменом Fc. По существу аналогичное связывание с FcRn достигается, когда домен Fc (или биспецифическая антигенсвязывающая молекула, включающая этот домен Fc) проявляет более 70%, в частности, более, чем приблизительно 80%, более предпочтительно более, чем приблизительно 90% связывающего сродства не сконструированного домена Fc (или биспецифической антигенсвязывающей молекулы, включающей данный не сконструированный домен Fc) к FcRn.
В некоторых формах осуществления домен Fc биспецифической антигенсвязывающей молекулы содержит одну или более мутаций аминокислоты, снижающих связывающее сродство домена Fc к рецептору Fc и/или эффекторную функцию. В характерном случае в каждой из двух субъединиц домена Fc находится одна и та же одна или более чем одна мутация аминокислоты. В одной форме осуществления мутация аминокислоты снижает связывающее сродство домена Fc к рецептору Fc. В одной форме осуществления мутация аминокислоты снижает связывающее сродство домена Fc к рецептору Fc по меньшей мере в 2 раза, по меньшей мере в 5 раз или по меньшей мере в 10 раз. В формах осуществления, где присутствует более одной мутации аминокислоты, снижающей связывающее сродство домена Fc к рецептору Fc, комбинация этих мутаций аминокислот может снизить связывающее сродство домена Fc к рецептору Fc по меньшей мере в 10 раз, по меньшей мере в 20 раз или даже по меньшей мере в 50 раз. В конкретной форме осуществления рецептор Fc представляет собой рецептор Fcγ. В некоторых формах осуществления рецептор Fc представляет собой рецептор Fc человека. В некоторых формах осуществления рецептор Fc представляет собой активирующий рецептор Fc. В конкретной форме осуществления рецептор Fc представляет собой активирующий рецептор Fcγ человека, более конкретно FcγRIIIa, FcγRI или FcγRIIa человека, наиболее конкретно FcγRIIIa человека. Предпочтительно связывание с каждым из этих рецепторов снижено. В некоторых формах осуществления связывающее сродство к компоненту комплемента, в частности, связывающее сродство к C1q также снижено. В одной форме осуществления связывающее сродство к неонатальному рецептору Fc (FcRn) не снижено.
В некоторых формах осуществления домен Fc биспецифической антигенсвязывающей молекулы сконструирован таким образом, что он обладает сниженной эффекторной функцией по сравнению с не сконструированным доменом Fc. Сниженная эффекторная функция может включать, но не ограничена ими, одну. или более из следующих функций: сниженную комплементзависимую цитотоксичность (КЗЦ), сниженную антителозависимую клеточно-опосредованную цитотоксичность (АЗКЦ), сниженный антителозависимый клеточный фагоцитоз (АЗКФ), сниженную секрецию цитокинов, сниженный захват антигена антигенпрезентирующими клетками, опосредованный иммунным комплексом, сниженное связывание с клетками NK, сниженное связывание с макрофагами, сниженное связывание с моноцитами, сниженное связывание с полиморфоядерными клетками, сниженную прямую передачу сигнала, индуцирующего апоптоз, сниженное перекрестное связывание с антителами, связанными с мишенями, сниженное созревание дендритных клеток или сниженный Т-клеточный прайминг. В одной форме осуществления сниженная эффекторная функция представляет собой одну или более из группы следующих функций: сниженной КЗЦ, сниженной АЗКЦ, сниженного АЗКФ и сниженной секреции цитокинов. В конкретной форме осуществления сниженная эффекторная функция представляет собой сниженную АЗКЦ. В одной форме осуществления сниженная АЗКЦ составляет менее 20% АЗКЦ, индуцируемой не сконструированным доменом Fc (или биспецифической антигенсвязывающей молекулой, включающей не сконструированный домен Fc).
В одной форме осуществления мутация аминокислоты, снижающая связывающее сродство домена Fc к рецептору Fc и/или эффекторную функцию, представляет собой замену аминокислоты. В одной форме осуществления домен Fc содержит замену аминокислоты в положении, выбранном из группы Е233, L234, L235, N297, Р331 и Р329. В более конкретной форме осуществления домен Fc содержит замену аминокислоты в положении, выбранном из группы L234, L235 и Р329. В некоторых формах осуществления домен Fc содержит замены аминокислоты L234A и L235A. В одной такой форме осуществления домен Fc представляет собой домен Fc IgG1, в частности, домен Fc IgG1 человека. В одной форме осуществления домен Fc содержит замену аминокислоты в положении Р329. В более конкретной форме осуществления замена аминокислоты представляет собой Р329А или P329G, в частности, P329G. В одной форме осуществления домен Fc содержит замену аминокислоты в положении Р329, и дополнительную замену аминокислоты в положении, выбранном из Е233, L234, L235, N297 и Р331. В более конкретной форме осуществления дополнительная замена аминокислоты представляет собой Е233Р, L234A, L235A, L235E, N297A, N297D или P331S. В конкретных формах осуществления домен Fc содержит замены аминокислоты в положениях Р329, L234 и L235. В более конкретных формах осуществления домен Fc содержит мутации аминокислот L234A, L235A и P329G ("P329G LALA"). В одной такой форме осуществления домен Fc представляет собой домен Fc IgG1, в частности, домен Fc IgG1 человека. Комбинация замен аминокислот "P329G LALA" почти полностью запрещает связывание с рецептором Fcγ домена Fc IgG1, как раскрыто в заявке на патент РСТ № РСТ/ЕР2012/055393, полностью включенной в данное описание посредством ссылки. В документе РСТ/ЕР2012/055393 также описаны способы получения таких мутантных доменов Fc и способы определения их свойств, таких как связывание рецептора Fc или эффекторные функции.
Антитела 1964 проявляют сниженное связывающее сродство к рецепторам Fc и сниженные эффекторные функции по сравнению с антителами IgG1. Следовательно, в некоторых формах осуществления домен Fc биспецифических антигенсвязывающих молекул по изобретению представляет собой домен Fc IgG4, в частности, домен Fc IgG4 человека. В одной форме осуществления домен Fc IgG4 содержит замены аминокислот в положении S228, в частности, замену аминокислоты S228P. Чтобы дополнительно снизить его связывающее сродство к рецептору Fc и/или его эффекторную функцию, в одной форме осуществления домен Fc IgG4 содержит замену аминокислоты в положении L235, в частности, замену аминокислоты L235E. В другой форме осуществления домен Fc IgG4 содержит замену аминокислоты в положении Р329, в частности, содержит замену аминокислоты P329G. В конкретной форме осуществления домен Fc IgG4 содержит замены аминокислот в положениях S228, L235 и Р329, в частности, замены аминокислот S228P, L235E и P329G. Такие мутанты домена Fc IgG4 и их свойства связывания рецептора Fcγ раскрыты в заявке на патент РСТ № РСТ/ЕР2012/055393, полностью включенной в данное описание посредством ссылки.
В конкретной форме осуществления домен Fc, проявляющий сниженное связывающее сродство к рецептору Fc и/или сниженную эффекторную функцию по сравнению с нативным доменом Fc IgG1, представляет собой домен Fc IgG1 человека, содержащий замены аминокислот L234A, L235A и необязательно P329G, или домен Fc IgG4 человека, содержащий замены аминокислот S228P, L235E и необязательно P329G.
В некоторых формах осуществления N-гликозилирование домена Fc элиминировано. В одной такой форме осуществления домен Fc содержит мутацию аминокислоты в положении N297, в частности, замену кислоты, заменяющую аспарагин аланином (N297A) или аспарагиновой кислотой (N297D).
В дополнение к доменам Fc, раскрытым выше в данной работе и в заявке на патент РСТ № РСТ/ЕР2012/055393, домены Fc, обладающие сниженным связыванием с рецептором Fc и/или сниженной эффекторной функцией, также включают домены Fc, несущие замену одного или более следующих остатков домена Fc: 238, 265, 269, 270, 297, 327 и 329 (патент US №6737056). Такие мутанты Fc включают мутанты Fc, имеющие замены в двух или более следующих положений аминокислот: 265, 269, 270, 297 и 327, включая так называемый "DANA" мутант Fc, имеющий замену остатков 265 и 297 аланином (патент US №7332581).
Мутантные домены Fc могут быть получены путем делеции, замены, инсерции или модификации аминокислоты, используя генетические или химические способы, хорошо известные в данной области техники. Генетические способы могут включать сайт-специфический мутагенез кодирующей последовательности ДНК, ПЦР, генный синтез и тому подобное. Корректные нуклеотидные замены могут быть проверены, например, путем секвенирования.
b) Повышенное связывание рецептора Fc и/или повышенная эффекторная функция
Могут быть, наоборот, ситуации, где желательно сохранить или даже усилить связывание рецептора Fc и/или эффекторные функции биспецифических антигенсвязывающих молекул, например, когда биспецифическая антигенсвязывающая молекула направлена на высокоспецифичный опухолевый антиген. Следовательно, в некоторых формах осуществления домен Fc биспецифических антигенсвязывающих молекул по изобретению сконструирован таким образом, чтобы он обладал повышенным связывающим сродством к рецептору Fc. Повышенное связывающее сродство может представлять собой повышение связывающего сродства домена Fc к рецептору Fc по меньшей мере в 2 раза, по меньшей мере в 5 раз или по меньшей мере в 10 раз. В одной форме осуществления рецептор Fc представляет собой активирующий рецептор Fc. В конкретной форме осуществления рецептор Fc представляет собой рецептор Fcγ, в частности, рецептор Fcγ человека. В одной форме осуществления рецептор Fc выбран из группы FcγRIIIa, FcγRI и FcγRIIa. В конкретной форме осуществления рецептор Fc представляет собой FcγRIIIa.
В одной такой форме осуществления домен Fc сконструирован таким образом, что он обладает измененной олигосахаридной структурой по сравнению с не сконструированным доменом Fc. В такой конкретной форме осуществления домен Fc содержит повышенную долю нефукозилированных олигосахаридов по сравнению с не сконструированным доменом Fc. В более конкретной форме осуществления по меньшей мере приблизительно 50%, более конкретно по меньшей мере приблизительно 70% N-сшитых олигосахаридов в домене Fc биспецифической антигенсвязывающей молекулы не фукозилировано. Нефукозилированные олигосахариды могут относиться к гибридному или комплексному типу. В другой конкретной форме осуществления домен Fc содержит повышенную долю раздвоенных олигосахаридов по сравнению с не сконструированным доменом Fc. В более конкретной форме осуществления по меньшей мере приблизительно 35%, в частности, по меньшей мере приблизительно 50%, более конкретно по меньшей мере приблизительно 70% N-сшитых олигосахаридов в домене Fc биспецифической антигенсвязывающей молекулы является раздвоенными. Раздвоенные олигосахариды могут относиться к гибридному или комплексному типу. В другой конкретной форме осуществления домен Fc содержит повышенную долю раздвоенных, нефукозилированных олигосахаридов по сравнению с не сконструированным доменом Fc. В более конкретной форме осуществления по меньшей мере приблизительно 15%, более конкретно по меньшей мере приблизительно 25%, по меньшей мере приблизительно 35% или по меньшей мере приблизительно 50% N-сшитых олигосахаридов в домене Fc биспецифической антигенсвязывающей молекулы является раздвоенными и не фукозилированными. Раздвоенные, Нефукозилированные олигосахариды могут относиться к гибридному или комплексному типу.
Олигосахаридные структуры в домене Fc биспецифической антигенсвязывающей молекулы можно анализировать способами, хорошо известными в данной области техники, например, с помощью масс-спектроскопии (МС) путем времяпролетной ионизации с лазерной десорбцией из матрицы (MALDI-TOF; от англ. "matrix laser desorption ionization time-of-flight"), как описано в статье Umana et al., Nat Biotechnol 17, 176-180 (1999) или Ferrara et al., Biotechn Bioeng 93, 851-861 (2006). Процентное содержание нефукозилированных олигосахаридов представляет собой количество олигосахаридов, в которых отсутствуют остатки фукозы, относительно всех олигосахаридов, присоединенных к Asn297 (например, комплексных, гибридных и высокоманнозных структур), и его идентифицируют в образце, обработанном N-гликозидазой F, с помощью МС MALDI-TOF. Asn297 относится к остатку аспарагина, локализованному приблизительно в положении 297 в домене Fc (нумерация ЕС остатков области Fc); тем не менее, Asn297 может быть также локализован приблизительно на ±3 аминокислоты выше или ниже положения 297, то есть между положениями 294 и 300, за счет минорных вариаций последовательности в иммуноглобулинах. Процентное содержание раздвоенных или раздвоенных, нефукозилированных олигосахаридов определяют аналогично.
Модификация гликозилирования в домене Fc биспецифической антигенсвязывающей молекулы может быть результатом продуцирования биспецифической антигенсвязывающей молекулы в клетке-хозяине, с которой проводят манипуляции с целью экспрессии измененных уровней одного или более полипептидов, обладающих гликозилтрансферазной активностью.
В одной форме осуществления домен Fc биспецифической антигенсвязывающей молекулы сконструирован таким образом, что он имеет измененную структуру олигосахаридов по сравнению с не сконструированным доменом Fc за счет продуцирования биспецифической антигенсвязывающей молекулы в клетке-хозяине, обладающей измененной активностью одной или более гликозилтрансфераз. Гликозилтрансферазы включают, например, β(1,4)-N-ацетилглюкозаминилтрансферазу III (GnTIII), β(1,4)-галактозилтрансферазу (GalT), β(1,2)-N-ацетилглюкозаминилтрансферазу I (GnTI), β(1,2)-N-ацетилглюкозаминилтрансферазу II (GnTII) и α(1,6)-фукозилтрансферазу. В конкретной форме осуществления домен Fc биспецифической антигенсвязывающей молекулы сконструирован таким образом, что он содержит повышенную долю нефукозилированных олигосахаридов по сравнению с не сконструированным доменом Fc за счет продуцирования биспецифической антигенсвязывающей молекулы в клетке-хозяине, обладающей повышенной активностью β(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII). В еще более конкретной форме осуществления клетка-хозяин дополнительно обладает повышенной активностью α-маннозидазы II (ManII). Методология гликоинжиниринга, которую можно использовать для гликоинжиниринга биспецифических антигенсвязывающих молекул по настоящему изобретению более подробно раскрыта в следующих документах: Umana et al., Nat Biotechnol 17, 176-180 (1999); Ferrara et al., Biotechn Bioeng 93, 851-861 (2006); WO 99/54342 (патент US №6602684; EP 1071700); WO 2004/065540 (US Pat. Appl. Publ. No. 2004/0241817; EP 1587921), WO 03/011878 (публикация заявки на патент US №2003/0175884), содержание каждого из которых полностью явным образом включено в настоящее описание посредством ссылки.
В общем, любой тип культивируемой линии клеток, включая линии клеток, обсуждаемые в данной работе, можно использовать для создания клеточных линий, продуцирующих биспецифические антигенсвязывающие молекулы, имеющие измененный паттерн гликозилирования. Конкретные линии клеток включают клетки СНО, клетки BHK, клетки NS0, клетки SP2/0, клетки миеломы YO, клетки миеломы мыши Р3Х63, клетки PER, клетки PER.C6 или клетки гибридомы и другие клетки млекопитающих. В некоторых формах осуществления с клетками-хозяевами проведены манипуляции, в результате которых они экспрессируют повышенные уровни одного или более полипептидов, обладающих активностью β(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII). В некоторых формах осуществления с клетками-хозяевами проведены дополнительные манипуляции, в результате которых они экспрессируют повышенные уровни одного или более полипептидов, обладающих активностью α-маннозидазы II (ManII). В конкретной форме осуществления полипептид, обладающий активностью GnTIII, представляет собой слитый полипептид, включающий каталитический домен GnTIII и домен локализации в комплексе Гольджи гетерологичного Гольджи-резидентного полипептида. В частности, домен локализации в комплексе Гольджи представляет собой домен локализации в комплексе Гольджи маннозидазы II. Способы создания таких слитых полипептидов и их применения для продуцирования антител, обладающих повышенными эффекторными функциями, раскрыты в Ferrara et al., Biotechn Bioeng 93, 851-861 (2006) и WO 2004/065540, содержание которых полностью явным образом включено в настоящее описание посредством ссылки.
Клетки-хозяева, содержащие кодирующую последовательность биспецифической антигенсвязывающей молекулы по изобретению и/или кодирующую последовательность полипептида, обладающего активностью гликозилтрансферазы, и экспрессирующие биологически активные генные продукты, могут быть идентифицированы с помощью, например, гибридизации ДНК-ДНК или ДНК-РНК, присутствия или отсутствия функций "маркерного" гена, оценки уровня транскрипции, измеряемого на основании экспрессии соответствующих транскриптов мРНК в клетке-хозяине, или обнаружения генного продукта, измеряемого путем иммунологического анализа или на основании его биологической активности, то есть способами, хорошо известными в данной области техники. Активность GnTIII или Man II можно определить, например, путем использования пектина, связывающегося с продуктами биосинтеза GnTIII или ManII, соответственно. Примером такого пектина является лектин Е4-ФГА, преимущественно связывающийся с олигосахаридами, содержащими раздвоенные GlcNAc. Продукты биосинтеза (то есть определенные олигосахаридные структуры) полипептидов, обладающих активностью GnTIII или ManII, могут быть также обнаружены с помощью масс-спектрометрического анализа олигосахаридов, высвобожденных из гликопротеинов, продуцируемых клетками, экспрессирующими эти полипептиды. Альтернативно можно использовать функциональный анализ, в котором измеряют повышенную эффекторную функцию и/или повышенное связывание рецептора Fc, опосредованные биспецифическими антигенсвязывающими молекулами, продуцируемыми сконструированными клетками, содержащими полипептид, обладающий активностью GnTIII или ManII.
В другой форме осуществления домен Fc сконструирован таким образом, что он содержит повышенную долю нефукозилированных олигосахаридов по сравнению с не сконструированным доменом Fc за счет продуцирования биспецифической антигенсвязывающей молекулы в клетке-хозяине, обладающей сниженной активностью α(1,6)-фукозилтрансферазы. Клетка-хозяин, обладающая сниженной активностью α(1,6)-фукозилтрансферазы, может представлять собой клетку, в которой ген α(1,6)-фукозилтрансферазы прерван или иным образом инактивирован, например, нокаутирован (см. Yamane-Ohnuki et al., Biotech Bioeng 87, 614 (2004); Kanda et al., Biotechnol Bioeng 94(4), 680-688 (2006); Niwa et al., J Immunol Methods 306, 151-160 (2006)).
Другие примеры линий клеток, способных к продуцированию дефукозилированных биспецифических антигенсвязывающих молекул, включают клетки Lec13 CHO, обладающие дефицитом фукозилирования белков (Ripka et al., Arch Biochem Biophys 249, 533-545 (1986); заявка на патент US № US 2003/0157108 и WO 2004/056312, в частности, в Примере 11). Биспецифические антигенсвязывающие молекулы по настоящему изобретению альтернативно могут быть сконструирован методами гликоинжиниринга таким образом, что имеют укороченные остатки фукозы в домене Fc, согласно методам, раскрытым в следующих документах: ЕР 1176195 А1, WO 03/084570, WO 03/085119 и публикациях заявок на патент US №№2003/0115614, 2004/093621, 2004/110282, 2004/110704, 2004/132140, в патенте US №6946292 (Kyowa), например, в результате снижения или запрещения активности белка-переносчика гуанозиндифосфат (ГДФ)-фукозы в клетках-хозяевах, применяемых для продуцирования биспецифической антигенсвязывающей молекулы.
Биспецифические антигенсвязывающие молекулы по изобретению, сконструированные методами гликоинжиниринга, можно также продуцировать в экспрессионных системах, продуцирующих модифицированные гликопротеины, как заявлено в документе WO 2003/056914 (GlycoFi, Inc.) или WO 2004/057002 и WO 2004/024927 (Greenovation).
В одной форме осуществления домен Fc биспецифической антигенсвязывающей молекулы сконструирован таким образом, что он обладает повышенной эффекторной функцией по сравнению с не сконструированным доменом Fc. Повышенная эффекторная функция может включать, но не ограничена ими, одну или более из следующих функций: повышенную комплементзависимую цитотоксичность (КЗЦ), повышенную антителозависимую клеточно-опосредованную цитотоксичность (АЗКЦ), повышенный антителозависимый клеточный фагоцитоз (АЗКФ), повышенную секрецию цитокинов, повышенный захват антигена антигенпрезентирующими клетками, опосредованный иммунным комплексом, повышенное связывание с клетками NK, повышенное связывание с макрофагами, повышенное связывание с моноцитами, повышенное связывание с полиморфоядерными клетками, повышенную прямую передачу сигнала, индуцирующего апоптоз, повышенное перекрестное связывание с антителами, связанными с мишенями, повышенное созревание дендритных клеток или повышенный Т-клеточный прайминг.
В одной форме осуществления повышенная эффекторная функция представляет собой одну или более из группы следующих функций: повышенной КЗЦ, повышенной АЗКЦ, повышенного АЗКФ и повышенной секреции цитокинов. В конкретной форме осуществления повышенная эффекторная функция представляет собой повышенную АЗКЦ. В одной форме осуществления АЗКЦ, индуцируемая сконструированным доменом Fc (или биспецифической антигенсвязывающей молекулой, включающей сконструированный домен Fc), повышена по меньшей мере в 2раза по сравнению с АЗКЦ, индуцируемой не сконструированным доменом Fc (или биспецифической антигенсвязывающей молекулой, включающей не сконструированный домен Fc).
Антигены
Биспецифические антигенсвязывающие молекулы по изобретению могут связываться с рядом антигенов. В некоторых формах осуществления первый и/или второй антиген представляет собой антиген, ассоциированный с патологическим состоянием, например, антиген, презентированный на опухолевой клетке, на клетке, инфицированной вирусом, или в сайте воспаления. Подходящие антигены включают антигены клеточной поверхности (например, но не ограниченные ими, рецепторы клеточной поверхности), свободные антигены в сыворотке крови и/или антигены во внеклеточном матриксе. В конкретных формах осуществления антиген представляет собой антиген человека.
Неограничивающие примеры антигенов включают опухолевые антигены, например, следующие антигены: MAGE, MART-1/Melan-A, gp100, дипептидилпептидаза IV (DPP IV; от англ. "Dipeptidyl peptidase IV"), белок, связывающий аденозиндезаминазу (ADAbp; от англ. "adenosine deaminase-binding protein"), циклофилин b, колоректальный ассоциированный антиген (CRC)-C017-1A/GA733, канцероэмбриональный антиген (СЕА; от англ. "Carcinoembryonic Antigen") и его иммуногенные эпитопы САР-1 и CAP-2, etv6, aml1, простатоспецифический антиген (PSA; от англ. "Prostate Specific Antigen") и его иммуногенные эпитопы PSA-1, PSA-2 и PSA-3, простатоспецифический мембранный антиген (PSMA; от англ. "Prostate-specific Membrane Antigen"), T-клеточный рецептор/CD3-зета-цепь, семейство опухолевых антигенов MAGE (например, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-АН, MAGE-A12, MAGE-Xp2 (MAGE-B2), MAGE-Хр3 (MAGE-В3), MAGE-Xp4 (MAGE-B4), MAGE-C1, MAGE-C2, MAGE-C3, MAGE-C4, MAGE-C5), семейство опухолевых антигенов GAGE (например, GAGE-1, GAGE-2, GAGE-3, GAGE-4, GAGE-5, GAGE-6, GAGE-7, GAGE-8, GAGE-9), BAGE, RAGE, LAGE-1, NAG, GnT-V, MUM-1, CDK4, тирозиназа, р53, семейство MUC, HER2/neu, Her3, p21ras, RCAS1, α-фетопротеин, Е-кадхерин, α-катенин, β-катенин и γ-катенин, p120ctn, gp100 Pmel117, PRAME, NY-ESO-1, cdc27, белок аденоматозного полипоза толстой кишки (АРС; от англ. "adenomatous polyposis coli protein"), фодрин, коннексин 37, Ig-идиотип, р15, gp75, ганглиозиды GM2 и GD2, семейство опухолевых антигенов Smad, Imp-1, P1A, EBV-кодируемый ядерный антиген (EBNA)-1, гликогенфосфорилаза головного мозга, SSX-1, SSX-2 (HOM-MEL-40), SSX-1, SSX-4, SSX-5, SCP-1 и СТ-7, а также с-erbB-2; антигены ЕСМ, такие как синдекан, гепараназа, интегрины, остеопонтин, link, кадхерины, ламинин, ламинин типа EGF, лектин, фибронектин и его альтернативно сплайсированные домены (например, экстра-домен В), Notch, различные формы тенасцина (например, тенасцин С) и его альтернативно сплайсированные домены (например, домен А1 или А2 тенасцина-С) и матриксин; белок активации фибробластов (FAP; от англ. "Fibroblast Activation Protein"), рецептор эпидермального фактора роста (EGFR; от англ. "Epidermal Growth Factor Receptor"), CD2 (Т-клеточный поверхностный антиген), CD3 (гетеромультимер, ассоциированный с TCR), CD19, CD22 (B-клеточный рецептор), CD23 (низкоаффинный рецептор IgE), CD25 (рецептор α-цепи интерлейкина-2 (ИЛ-2)), CD30 (рецептор цитокина), CD33 (поверхностный антиген миелоидных клеток), CD40 (рецептор фактора некроза опухоли), IL-6R (рецептор ИЛ-6), CD20, меланома-ассоциированный протеогликановый хондроитинсульфат (MCSP; от англ. "Melanoma-associated Chondroitin Sulfate Proteoglycan"), рецептор инсулиноподобного фактора роста-1 (IGF-1R; от англ. "Insulin-like growth factor-1 receptor") и PDGFβR (рецептор β тромбоцитарного фактора роста (от англ. "platelet-derived growth factor receptor").
В конкретной форме осуществления первый и второй антиген выбраны из группы белка активации фибробластов (FAP), меланома-ассоциированного протеогликанового хондроитинсульфата (MCSP), рецептора эпидермального фактора роста (EGFR), Her3, c-Met, канцероэмбрионального антигена (СЕА), CD33 и CD3.
В конкретной форме осуществления первый антиген представляет собой CD3, в частности, CD3 человека или яванского макака, наиболее конкретно CD3 человека. В некоторых формах осуществления первый антиген представляет собой эпсилон-субъединицу CD3. В одной форме осуществления первый Fab фрагмент может конкурировать с моноклональным антителом Н2С (раскрытым в публикации РСТ № WO 2008/119567) за связывание эпитопа CD3. В конкретной форме осуществления первый Fab фрагмент может конкурировать с моноклональным антителом SP34 (раскрытым в статье Pessano et al., EMBO J 4, 337-340 (1985)) за связывание эпитопа CD3. В другой форме осуществления первый Fab фрагмент может конкурировать с моноклональным антителом V9 (раскрытым в статье Rodrigues et al., Int J Cancer Suppl 7, 45-50 (1992) и в патенте US №6054297) за связывание эпитопа CD3. В другой форме осуществления первый Fab фрагмент может конкурировать с моноклональным антителом FN18 (раскрытым в статье Nooij et al., Eur J Immunol 19, 981-984 (1986)) за связывание эпитопа CD3. В одной форме осуществления первый Fab фрагмент специфичен к CD3 и включает CDR1 тяжелой цепи SEQ ID NO:77, CDR2 тяжелой цепи SEQ ID NO:78, CDR3 тяжелой цепи SEQ ID NO:79, CDR1 легкой цепи SEQ ID NO:81, CDR2 легкой цепи SEQ ID NO:82 и CDR3 легкой цепи SEQ ID NO:83. В следующей форме осуществления Fab фрагмент, обладающий специфичностью к CD3, включает последовательность вариабельной области тяжелой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:80, и последовательность вариабельной области легкой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:84, либо ее варианты, сохраняющие функциональность. В другой форме осуществления первый Fab фрагмент специфичен к CD3 и включает CDR1 тяжелой цепи SEQ ID NO:104, CDR2 тяжелой цепи SEQ ID NO:105, CDR3 тяжелой цепи SEQ ID NO:106, CDR1 легкой цепи SEQ ID NO:108, CDR2 легкой цепи SEQ ID NO:109 и CDR3 легкой цепи SEQ ID NO:110. В следующей форме осуществления Fab фрагмент, специфичный к CD3, включает последовательность вариабельной области тяжелой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:107, и последовательность вариабельной области легкой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:111, либо их варианты, сохраняющие функциональность.
В конкретной форме осуществления согласно изобретению биспецифическая антигенсвязывающая молекула способна к одновременному связыванию с антигеном клетки-мишени, в частности, с антигеном опухолевой клетки, и CD3. В одной форме осуществления биспецифическая антигенсвязывающая молекула способна к перекрестному связыванию Т-клетки и клетки-мишени посредством одновременного связывания с антигеном клетки-мишени и CD3. В еще более конкретной форме осуществления такое одновременное связывание приводит в результате к лизису клетки-мишени, в частности, опухолевой клетки. В одной форме осуществления такое одновременное связывание приводит в результате к активации Т-клетки. В других формах осуществления такое одновременное связывание приводит в результате к клеточному ответу Т-лимфоцита, в частности, цитотоксического Т-лимфоцита, выбранному из группы, состоящей из следующих ответов: пролиферации, дифференциации, секреции цитокинов, высвобождению цитотоксической эффекторной молекулы, цитотоксической активности и секреции маркеров активации. В одной форме осуществления связывание биспецифической антигенсвязывающей молекулы с CD3 без одновременного связывания с антигеном-мишенью не приводит в результате к Т-клеточной активации.
В одной форме осуществления биспецифическая антигенсвязывающая молекула способна к перенаправлению цитотоксической активности Т-клетки на клетку-мишень. В конкретной форме осуществления такое перенаправление является независимым от ГКГ-опосредованного презентирования пептидного антигена клеткой-мишенью и/или от специфичности Т-клетки.
В частности, Т-клетка согласно любой из форм осуществления изобретения представляет собой цитотоксическую Т-клетку. В некоторых формах осуществления Т-клетка представляет собой CD4+ или CD8+ Т-клетку, в частности, CD8+ Т-клетку.
В одной форме осуществления первый антиген представляет собой c-Met, в частности, c-Met человека. В одной форме осуществления первый Fab фрагмент может конкурировать с моноклональным антителом 5D5 (раскрытым, например, в патенте US №7476724, полностью включенном в настоящее описание посредством ссылки) за связывание эпитопа c-Met. В одной форме осуществления первый Fab фрагмент специфичен к c-Met и включает CDR1 тяжелой цепи SEQ ID NO:63, CDR2 тяжелой цепи SEQ ID NO:64, CDR3 тяжелой цепи SEQ ID NO:65, CDR1 легкой цепи SEQ ID NO:67, CDR2 легкой цепи SEQ ID NO:68 и CDR3 легкой цепи SEQ ID NO:69. В следующей форме осуществления Fab фрагмент, специфичный к c-Met, включает последовательность вариабельной области тяжелой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:66, и последовательность вариабельной области легкой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:70, либо их варианты, сохраняющие функциональность.
В конкретных формах осуществления второй антиген представляет собой опухолеассоциированный антиген, в частности, антиген, презентированный на опухолевой клетке или клетке опухолевой стромы. В одной такой форме осуществления второй антиген выбран из группы следующих антигенов: белка активации фибробластов (FAP), меланома-ассоциированного протеогликанового хондроитинсульфата (MCSP), рецептора эпидермального фактора роста (EGFR), Her3, CD33 и канцероэмбрионального антигена (СЕА).
В одной форме осуществления второй антиген представляет собой меланома-ассоциированный протеогликановый хондроитинсульфат (MCSP). В другой форме осуществления второй и необязательно третий Fab фрагмент может конкурировать с моноклональным антителом LC007 (см. SEQ ID NO:18 и 22) за связывание с эпитопом MCSP. В одной форме осуществления Fab фрагмент, специфичный к MCSP, включает CDR1 тяжелой цепи SEQ ID NO:15, CDR2 тяжелой цепи SEQ ID NO:16, CDR3 тяжелой цепи SEQ ID NO:17, CDR1 легкой цепи SEQ ID NO:19, CDR2 легкой цепи SEQ ID NO:20 и CDR3 легкой цепи SEQ ID NO:21. В следующей форме осуществления Fab фрагмент, специфичный к MCSP, включает последовательность вариабельной области тяжелой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:18, и последовательность вариабельной области легкой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:22, либо их варианты, сохраняющие функциональность. В другой форме осуществления второй и необязательно третий Fab фрагмент может конкурировать с моноклональным антителом М4-3 ML2 (см. SEQ ID NO:99 и 103) за связывание с эпитопом MCSP. В одной форме осуществления Fab фрагмент, специфичный к MCSP, включает CDR1 тяжелой цепи SEQ ID NO:96, CDR2 тяжелой цепи SEQ ID NO:97, CDR3 тяжелой цепи SEQ ID NO:98, CDR1 легкой цепи SEQ ID NO:100, CDR2 легкой цепи SEQ ID NO:101 и CDR3 легкой цепи SEQ ID NO:102. В следующей форме осуществления Fab фрагмент, специфичный к MCSP, включает последовательность вариабельной области тяжелой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:99, и последовательность вариабельной области легкой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:103, либо их варианты, сохраняющие функциональность.
В другой форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:1, полипептидную последовательность SEQ ID NO:2, полипептидную последовательность SEQ ID NO:3 и полипептидную последовательность SEQ ID NO:4, либо их варианты, сохраняющие функциональность. В следующей форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:1, полипептидную последовательность SEQ ID NO:3, полипептидную последовательность SEQ ID NO:4 и полипептидную последовательность SEQ ID NO:5, либо их варианты, сохраняющие функциональность. В другой форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:4, полипептидную последовательность SEQ ID NO:5, полипептидную последовательность SEQ ID NO:6 и полипептидную последовательность SEQ ID NO:7, либо их варианты, сохраняющие функциональность. В другой форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:4, полипептидную последовательность SEQ ID NO:5, полипептидную последовательность SEQ ID NO:1 и полипептидную последовательность SEQ ID NO:85, либо их варианты, сохраняющие функциональность. В следующей форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:1, полипептидную последовательность SEQ ID NO:3, полипептидную последовательность SEQ ID NO:4 и полипептидную последовательность SEQ ID NO:86, либо их варианты, сохраняющие функциональность. В следующей форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:4, полипептидную последовательность SEQ ID NO:87, полипептидную последовательность SEQ ID NO:89 и полипептидную последовательность SEQ ID NO:90, либо их варианты, сохраняющие функциональность. В следующей форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:3, полипептидную последовательность SEQ ID NO:91, полипептидную последовательность SEQ ID NO:92 и полипептидную последовательность SEQ ID NO:93, либо их варианты, сохраняющие функциональность. В другой форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:87, полипептидную последовательность SEQ ID NO:91, полипептидную последовательность SEQ ID NO:93 и полипептидную последовательность SEQ ID NO:94, либо их варианты, сохраняющие функциональность.
В одной форме осуществления второй антиген представляет собой канцероэмбриональный антиген (СЕА). В другой форме осуществления второй и необязательно третий Fab фрагмент может конкурировать с моноклональным антителом СН1А1А за связывание эпитопа СЕА. См. публикацию РСТ WO 2011/023787, полностью включенную в данное описание посредством ссылки. В одной форме осуществления Fab фрагмент, специфичный к СЕА, включает CDR1 тяжелой цепи SEQ ID NO:39, CDR2 тяжелой цепи SEQ ID NO:40, CDR3 тяжелой цепи SEQ ID NO:41, CDR1 легкой цепи SEQ ID NO:43, CDR2 легкой цепи SEQ ID NO:44 и CDR3 легкой цепи SEQ ID NO:45. В следующей форме осуществления Fab фрагмент, специфичный к СЕА, включает последовательность вариабельной области тяжелой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:42, и последовательность вариабельной области легкой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:46, либо их варианты, сохраняющие функциональность.
В другой форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:3, полипептидную последовательность SEQ ID NO:8, полипептидную последовательность SEQ ID NO:9 и полипептидную последовательность SEQ ID NO:10, либо их варианты, сохраняющие функциональность. В другой форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:9, полипептидную последовательность SEQ ID NO:10, полипептидную последовательность SEQ ID NO:87 и полипептидную последовательность SEQ ID NO:95, либо их варианты, сохраняющие функциональность.
В одной форме осуществления второй антиген представляет собой Her3. В другой форме осуществления второй и необязательно третий Fab фрагмент может конкурировать с моноклональным антителом Mab 205.10 за связывание с эпитопом Her3. См. публикацию РСТ № WO 2011/076683, полностью включенную в данное описание посредством ссылки. В одной форме осуществления Fab фрагмент, специфичный к Her3, включает CDR1 тяжелой цепи SEQ ID NO:55, CDR2 тяжелой цепи SEQ ID NO:56, CDR3 тяжелой цепи SEQ ID NO:57, CDR1 легкой цепи SEQ ID NO:59, CDR2 легкой цепи SEQ ID NO:60 и CDR3 легкой цепи SEQ ID NO:61. В следующей форме осуществления Fab фрагмент, специфичный к Her3, включает последовательность вариабельной области тяжелой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:58, и последовательность вариабельной области легкой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:62, либо их варианты, сохраняющие функциональность.
В другой форме осуществления биспецифическая антигенсвязывающая молекула включает полипептидную последовательность SEQ ID NO:11, полипептидную последовательность SEQ ID NO:12, полипептидную последовательность SEQ ID NO:13 и полипептидную последовательность SEQ ID NO:14, либо их варианты, сохраняющие функциональность.
В одной форме осуществления второй антиген представляет собой рецептор эпидермального фактора роста (EGFR). В другой форме осуществления второй и необязательно третий Fab фрагмент может конкурировать с моноклональным антителом GA201 за связывание с эпитопом EGFR. См. публикацию РСТ № WO 2006/082515, полностью включенную в данное описание посредством ссылки. В одной форме осуществления Fab фрагмент, специфичный к EGFR, включает CDR1 тяжелой цепи SEQ ID NO:23, CDR2 тяжелой цепи SEQ ID NO:24, CDR3 тяжелой цепи SEQ ID NO:25, CDR1 легкой цепи SEQ ID NO:27, CDR2 легкой цепи SEQ ID NO:28 и CDR3 легкой цепи SEQ ID NO:29. В следующей форме осуществления Fab фрагмент, специфичный к EGFR, включает последовательность вариабельной области тяжелой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичный SEQ ID NO:26, и последовательность вариабельной области легкой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:30, либо их варианты, сохраняющие функциональность.
В одной форме осуществления второй антиген представляет собой белок активации фибробластов (FAP). В другой форме осуществления второй и необязательно третий Fab фрагмент может конкурировать с моноклональным антителом 3F2 за связывание с эпитопом FAP. См. публикацию РСТ WO 2012/020006, полностью включенную в данное описание посредством ссылки. В одной форме осуществления Fab фрагмент, специфичный к FAP, включает CDR1 тяжелой цепи SEQ ID NO:31, CDR2 тяжелой цепи SEQ ID NO:32, CDR3 тяжелой цепи SEQ ID NO:33, CDR1 легкой цепи SEQ ID NO:35, CDR2 легкой цепи SEQ ID NO:36 и CDR3 легкой цепи SEQ ID NO:37. В следующей форме осуществления Fab фрагмент, специфичный к FAP, включает последовательность вариабельной области тяжелой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:34, и последовательность вариабельной области легкой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:38, либо их варианты, сохраняющие функциональность.
В одной форме осуществления второй антиген представляет собой CD33. В одной форме осуществления Fab фрагмент, специфичный к CD33, включает CDR1 тяжелой цепи SEQ ID NO:47, CDR2 тяжелой цепи SEQ ID NO:48, CDR3 тяжелой цепи SEQ ID NO:49, CDR1 легкой цепи SEQ ID NO:51, CDR2 легкой цепи SEQ ID NO:52 и CDR3 легкой цепи SEQ ID NO:53. В следующей форме осуществления Fab фрагмент, специфичный к CD33, включает последовательность вариабельной области тяжелой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:50, и последовательность вариабельной области легкой цепи, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO:54, либо их варианты, сохраняющие функциональность.
Полинуклеотиды
Далее в изобретении предложены изолированные полинуклеотиды, кодирующие биспецифическую антигенсвязывающую молекулу, как описано в данной работе, или ее фрагмент. Полинуклеотиды, кодирующие биспецифические антигенсвязывающие молекулы по изобретению, могут экспрессироваться в виде одного полинуклеотида, кодирующего полноразмерную биспецифическую антигенсвязывающую молекулу, или в виде множественных (например, двух или более) полинуклеотидов, экспрессирующихся совместно. Полипептиды, кодируемые полинуклеотидами, экспрессируемыми совместно, могут связываться посредством, например, дисульфидных связей или других средств с образованием функциональной биспецифической антигенсвязывающей молекулы. Например, участок Fab фрагмента, представляющий собой легкую цепь, может кодироваться полинуклеотидом, отдельным от участка биспецифической антигенсвязывающей молекулы, включающего участок Fab фрагмента, представляющий собой тяжелую цепь, субъединицу домена Fc и необязательно другой Fab фрагмент (его часть). При совместной экспрессии полипептиды тяжелой цепи связываются с полипептидами легкой цепи с образованием Fab фрагмента. В другом примере участок биспецифической антигенсвязывающей молекулы, включающий одну из двух субъединиц домена Fc и необязательно один или более Fab фрагментов (или их частей), может кодироваться полинуклеотидом, отдельным от участка биспецифической антигенсвязывающей молекулы, включающего другую из двух субъединиц домена Fc и необязательно Fab фрагмент (его часть). При совместной экспрессии субъединицы домена Fc связываются с образованием домена Fc.
В одной форме осуществления изолированный полинуклеотид по изобретению кодирует первую субъединицу домена Fc, тяжелую цепь второго Fab фрагмента и тяжелую цепь первого Fab фрагмента. В более конкретной форме осуществления изолированный полинуклеотид кодирует полипептид, в котором область VL имеет общую карбокси-концевую пептидную связь с областью СН1, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с тяжелой цепью иммуноглобулина (VH-CH1-HR-CH2-CH3-(CH4)). В другой конкретной форме осуществления изолированный полинуклеотид кодирует полипептид, в котором тяжелая цепь Fab (VH-CH1) имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью VL, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью СН1, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с субъединицей домена Fc, включающей шарнирную область иммуноглобулина (HR-CH2-CH3-(CH4)). В другой конкретной форме осуществления изолированный полинуклеотид кодирует полипептид, в котором область VH имеет общую карбокси-концевую пептидную связь с областью CL, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с тяжелой цепью иммуноглобулина (VH-CH1-HR-CH2-CH3-(CH4)). В другой конкретной форме осуществления изолированный полинуклеотид кодирует полипептид, в котором тяжелая цепь Fab (VH-CH1) имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью VH, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью CL, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с субъединицей домена Fc, включающей шарнирную область иммуноглобулина (HR-CH2-CH3-(CH4)). В другой конкретной форме осуществления изолированный полинуклеотид кодирует полипептид, в котором тяжелая цепь иммуноглобулина ((VH-CH1-HR-CH2-CH3-(CH4)) имеет общую карбокси-концевую пептидную связь с пептидным линкером, который, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью VH, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью CL.
В следующих формах осуществления изолированный полинуклеотид по изобретению кодирует вторую субъединицу домена Fc и необязательно тяжелую цепь третьего Fab фрагмента. В конкретной форме осуществления изолированный полинуклеотид кодирует тяжелую цепь иммуноглобулина ((VH-CH1-HR-CH2-CH3-(CH4)). В другой конкретной форме осуществления изолированный полинуклеотид кодирует субъединицу домена Fc, необязательно включающую шарнирную область антитела ((HR)-CH2-CH3-(CH4)).
В следующих формах осуществления изолированный полинуклеотид по изобретению кодирует одну или более легких цепей, содержащихся в биспецифической антигенсвязывающей молекуле. В конкретной форме осуществления изолированный полинуклеотид кодирует легкую цепь иммуноглобулина (VL-CL). В другой конкретной форме осуществления изолированный полинуклеотид кодирует полипептид, в котором область VL имеет общую карбокси-концевую пептидную связь с областью СН1. В другой конкретной форме осуществления изолированный полинуклеотид кодирует полипептид, в котором область VH имеет общую карбокси-концевую пептидную связь с областью CL. В другой конкретной форме осуществления изолированный полинуклеотид кодирует полипептид, в котором область VH имеет общую карбокси-концевую пептидную связь с областью CL, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с легкой цепью Fab (VL-CL). В другой конкретной форме осуществления изолированный полинуклеотид кодирует полипептид, в котором легкая цепь Fab (VL-CL) имеет общую карбокси-концевую пептидную связь с областью VH, которая, в свою очередь, имеет общую карбокси-концевую пептидную связь с областью CL.
В другой форме осуществления настоящее изобретение направлено на изолированный полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу по изобретению или ее фрагмент, где полинуклеотид включает последовательность, кодирующую последовательность вариабельной области, представленную в SEQ ID NO:18, 22, 26, 30, 34, 38, 42, 46, 50, 54, 58, 62, 66, 70, 80, 84, 99, 103, 107 и 111. В другой форме осуществления настоящее изобретение направлено на изолированный полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу по изобретению или ее фрагмент, где полинуклеотид включает последовательность, кодирующую полипептидную последовательность, представленную в SEQ ID NO:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 и 95. В другой форме осуществления настоящее изобретение направлено на изолированный полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу по изобретению или ее фрагмент, где полинуклеотид включает последовательность, кодирующую последовательность вариабельной области, по меньшей мере приблизительно на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности в SEQ ID NO:18, 22, 26, 30, 34, 38, 42, 46, 50, 54, 58, 62, 66, 70, 80, 84, 99, 103, 107 или 111. В другой форме осуществления настоящее изобретение направлено на изолированный полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу по изобретению или ее фрагмент, где полинуклеотид включает последовательность, кодирующую полипептидную последовательность, по меньшей мере на 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности в SEQ ID NO:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95. Изобретение включает изолированный полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу по изобретению или ее фрагмент, где полинуклеотид включает последовательность, кодирующую последовательность вариабельной области SEQ ID NO:18, 22, 26, 30, 34, 38, 42, 46, 50, 54, 58, 62, 66, 70, 80, 84, 99, 103, 107 или 111 с консервативными заменами аминокислот. Изобретение также включает изолированный полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу по изобретению или ее фрагмент, где полинуклеотид включает последовательность, кодирующую полипептидную последовательность SEQ ID NO:1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94 или 95 с консервативными заменами аминокислот.
В некоторых формах осуществления полинуклеотид или нуклеиновая кислота представляет собой ДНК. В других формах осуществления полинуклеотид по настоящему изобретению представляет собой РНК, например, в форме мессенджер-РНК (мРНК). РНК по настоящему изобретению может быть однонитевой или двунитевой.
Рекомбинантные способы
Биспецифические антигенсвязывающие молекулы по изобретению могут быть получены, например, путем твердофазного пептидного синтеза (например, твердофазного синтеза Меррифильда) или рекомбинантного продуцирования. Для рекомбинантного продуцирования один или более полинуклеотидов, кодирующих биспецифическую антигенсвязывающую молекулу (фрагмент), например, как раскрыто выше, выделяют и встраивают в один или более векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такой полинуклеотид можно легко выделить и секвенировать, используя традиционные методы. В одной форме осуществления предложен вектор, предпочтительно экспрессионный вектор, содержащий один или более полинуклеотидов по изобретению. Для конструирования экспрессионных векторов, содержащих кодирующую последовательность биспецифической антигенсвязывающей молекулы (фрагмент) вместе с соответствующими транскрипционными/трансляционными контрольными сигналами можно использовать способы, хорошо известные специалистам в данной области техники. Эти способы включают методы рекомбинантных ДНК in vitro, синтетические методы и рекомбинацию/генетическую рекомбинацию in vivo. См., например, методы, раскрытые в следующих руководствах: Maniatis et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, N.Y. (1989); и Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates and Wiley Interscience, N.Y (1989). Экспрессионный вектор может представлять собой часть плазмиды, вируса, либо может представлять собой фрагмент нуклеиновой кислоты. Экспрессионный вектор включает экспрессионную кассету, в которой клонируют полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу (фрагмент) (то есть кодирующую область) в операционном сцеплении с промотором и/или другим контролирующим элементом транскрипции или трансляции. Как используют в данной работе, "кодирующая область" представляет собой участок нуклеиновой кислоты, состоящий из кодонов, транслируемых в аминокислоты. Хотя "терминирующий кодон" (TAG, TGA или ТАА) не транслируется в аминокислоту, его можно считать частью кодирующей области, если он присутствует, но любые фланкирующие последовательности, например, промоторы, сайты связывания рибосомы, транскрипционные терминаторы, интроны, 5' и 3' нетранслируемые участки и тому подобное, не являются частью кодирующей области. Две или более кодирующих областей могут присутствовать в одной полинуклеотидной конструкции, например, на одном векторе, или в отдельных полинуклеотидных конструкциях, например, на отдельных (различных) векторах. Кроме того, любой вектор может содержать одну кодирующую область или может содержать две или более кодирующих областей, например, вектор по настоящему изобретению может кодировать один или более полипептидов, которые отделяют от конечных белков после или во время трансляции посредством протеолитического расщепления. Кроме того, вектор, полинуклеотид или нуклеиновая кислота по изобретению может кодировать гетерологичные кодирующие области, либо слитые, либо не слитые с полинуклеотидом, кодирующим биспецифическую антигенсвязывающую молекулу (фрагмент) по изобретению или ее вариант или производное. Гетерологичные кодирующие области включают без ограничения специализированные элементы или мотивы, такие как секреторный сигнальный пептид или гетерологичный функциональный домен. Операционное сцепление существует, когда кодирующая область генного продукта, например полипептид, сцеплена с одной или более регуляторных последовательностей таким образом, чтобы поместить экспрессию генного продукта под влияние или контроль регуляторной последовательности (последовательностей). Два фрагмента ДНК (такие как кодирующая область полипептида и сцепленный с ней промотор) являются "операционно сцепленными", если индукция функции промотора приводит в результате к транскрипции мРНК, кодирующей желаемый генный продукт, и, если природа связи между двумя фрагментами ДНК не препятствует способности регуляторных последовательностей экспрессии к направлению экспрессии генного продукта, или не препятствует способности ДНК матрицы к транскрипции. Таким образом, промоторная область операционно сцеплена с нуклеиновой кислотой, кодирующей полипептид, если промотор способен осуществлять транскрипцию этой нуклеиновой кислоты. Промотор может представлять собой клеточно-специфичный промотор, направляющий существенную транскрипцию ДНК только в предопределенных клетках. Другие транскрипционные контрольные элементы, кроме промотора, например, энхансеры, операторы, репрессоры и сигналы терминации транскрипции, могут быть операционно сцеплены с полинуклеотидом, чтобы направлять клеточно-специфичную транскрипцию. Подходящие промоторы и другие транскрипционные контролирующие области раскрыты в данной работе. Различные транскрипционные контролирующие области известны специалистам в данной области техники. Эти области включают без ограничения транскрипционные контролирующие области, функционирующие в клетках позвоночных, такие как, но не ограниченные ими, промоторные и энхансерные сегменты из цитомегаловирусов (например, немедленно-ранний промотор в сочетании с интроном-А), вируса обезьян 40 (например, ранний промотор) и ретровирусов (таких как, например, вирус саркомы Рауса). Другие транскрипционные контролирующие области включают области, имеющие происхождение из генов позвоночных, таких как актин, белок теплового шока, гормон роста крупного рогатого скота и α-глобин кролика, а также другие последовательности, способные контролировать экспрессию генов в эукариотических клетках. Дополнительные подходящие транскрипционные контрольные области включают тканеспецифические промоторы и энхансеры, а также индуцибельные промоторы (например, промоторы, индуцируемые тетрациклинами). Аналогично ряд трансляционных контрольных элементов известен обычным специалистам в данной области техники. Эти элементы включают, но не ограничены ими, сайты связывания рибосом, кодоны инициации и терминации трансляции и элементы, имеющие происхождение из вирусных систем (в частности, участок внутренней посадки рибосомы, или IRES (от англ. "internal ribosome entry site"), также называемый последовательностью CITE). Экспрессионная кассета может также включать другие признаки, такие как точка начала репликации и/или элементы хромосомной интеграции, такие как длинные концевые повторы (ДКП) ретровирусов или инвертированные концевые повторы (ИКП) аденоассоциированных вирусов (ААВ).
Полинуклеотидные и нуклеиново-кислотные кодирующие области по настоящему изобретению могут быть связаны с дополнительными кодирующими областями, кодирующими секреторные или сигнальные пептиды, направляющие секрецию полипептида, кодируемого полинуклеотидом по настоящему изобретению. Например, если желательна секреция биспецифической антигенсвязывающей молекулы, ДНК, кодирующая сигнальную последовательность, может быть помещена выше нуклеиновой кислоты, кодирующей биспецифическую антигенсвязывающую молекулу по изобретению или ее фрагмент. Согласно сигнальной гипотезе белки, секретируемые клетками млекопитающих, имеют сигнальный пептид или секреторную лидерную последовательность, отщепляемую от зрелого белка, как только инициирован экспорт растущей белковой цепи через шероховатый эндоплазматический ретикулум. Обычным специалистам в данной области техники известно, что, как правило, в полипептидах, секретируемых клетками позвоночных, сигнальный пептид, отщепляемый от транслированного полипептида с образованием секретированной или "зрелой" формы полипептида, слит с N-концом полипептида. В некоторых формах осуществления используют нативный сигнальный пептид, например, сигнальный пептид тяжелой цепи или легкой цепи иммуноглобулина или функциональное производное такой последовательности, которая сохраняет способность к направлению секреции полипептида, операционно сцепленного с ним. Альтернативно можно использовать гетерологичный сигнальный пептид млекопитающих или его функциональное производное. Например, лидерная последовательность дикого типа может быть замещена лидерной последовательностью тканевого активатора плазминогена человека (ТРА; от англ. "tissue plasminogen activator") или β-глюкуронидазы мыши. Иллюстративные аминокислотные последовательности секреторных сигнальных пептидов приведены в SEQ ID NO:74-76.
ДНК, кодирующая короткую белковую последовательность, которую можно было бы использовать, чтобы облегчить последующую очистку (например, гистидиновая метка) или способствовать мечению биспецифической антигенсвязывающей молекулы, может быть включена внутри или на концах полинуклеотида, кодирующего биспецифическую антигенсвязывающую молекулу (фрагмент).
В следующей форме осуществления предложена клетка-хозяин, содержащая один или более полинуклеотидов по изобретению. В некоторых формах осуществления предложена клетка-хозяин, содержащая один или более векторов по изобретению. Полинуклеотиды и векторы могут включать любой из признаков, отдельно или в комбинации, раскрытых в настоящей работе в отношении полинуклеотидов и векторов, соответственно. В одной такой форме осуществления клетка-хозяин содержит (например, трансформирована или трансфицирована им) вектор, содержащий полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу (ее часть) по изобретению. Как используют в данной работе, термин "клетка-хозяин" относится к любому виду клеточной системы, которая может быть сконструирована для получения биспецифических антигенсвязывающих молекул по изобретению или их фрагментов. Клетки-хозяева, подходящие для репликации и для поддержания экспрессии биспецифических антигенсвязывающих молекул, хорошо известны в данной области техники. Такие клетки могут быть трансфицированы или трансдуцированы, как целесообразно для конкретного экспрессионного вектора, и большие количества клеток, содержащих вектор, можно выращивать для засева крупномасштабных ферментеров, чтобы получить достаточные количества биспецифической антигенсвязывающей молекулы для клинических применений. Подходящие клетки-хозяева включают прокариотические микроорганизмы, такие как Е. coli, или различные эукариотические клетки, такие как клетки яичника китайского хомячка (СНО), клетки насекомых или тому подобное. Например, полипептиды можно продуцировать в бактериях, в частности, когда нет необходимости в гликозилировании. После экспрессии полипептид может быть выделен из бактериальной клеточной суспензии в растворимой фракции, а затем может быть дополнительно очищен. В дополнение к прокариотам, подходящими хозяевами для клонирования или экспрессии векторов, кодирующих полипептид, являются эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи, включая штаммы грибов и дрожжей, биохимические пути гликозилирования которых "гуманизированы", что приводит в результате к продуцированию полипептида с частичным или полным человеческим паттерном гликозилирования. См. документы Gerngross, Nat Biotech 22,1409-1414 (2004) и Li et al., Nat Biotech 24, 210-215 (2006). Подходящие клетки-хозяева для экспрессии (гликозилированных) полипептидов также имеют происхождение от многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Идентифицированы различные бакуловирусные штаммы, которые можно использовать в сочетании с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda. Культуры клеток растений можно также использовать в качестве хозяев. См., например, патенты US №№5959177, 6040498, 6420548, 7125978 и 6417429 (описание технологии PLANTIBODIES™ для продуцирования антител в трансгенных растениях). Клетки позвоночных можно также использовать в качестве хозяев. Например, могут быть полезны линии клеток млекопитающих, адаптированные к росту в суспензии. Другими примерами полезных линий клеток-хозяев млекопитающих являются следующие линии: линия клеток почек обезьяны CV1, трансформированных SV40 (COS-7); линия клеток эмбриональных почек человека (клетки 293 или 293Т, как раскрыто, например, в Graham et al., J Gen Virol 36, 59 (1977)), клетки почек детенышей хомячка (ВНК), клетки Сертоли мыши (клетки ТМ4, как раскрыто, например, в Mather, Biol Reprod 23, 243-251 (1980)), клетки почек обезьяны (CV1), клетки почек африканской зеленой мартышки (VERO-76), клетки карциномы шейки матки человека (HELA), клетки почек собаки (MDCK), клетки печени серой крысы (BRL 3А), клетки легких человека (W138), клетки печени человека (Нер G2), клетки опухоли молочной железы мыши (ММТ 060562), клетки TRI (как описано, например, в Mather et al., Annals N.Y. Acad Sci 383, 44-68 (1982)), клетки MRC5 и клетки FS4. Другие полезные линии клеток-хозяев млекопитающих включают клетки яичника китайского хомячка (СНО), включающие клетки dhfr- CHO (Urlaub et al., Proc Natl Acad Sci USA 77, 4216 (1980)); и линии клеток миеломы, такие как YO, NS0, P3X63 и Sp2/0. Обзор некоторых линий клеток-хозяев млекопитающих, подходящих для продуцирования белка, см., например, в кн. Yazaki and Wu, Methods in Molecular Biology, Vol. 248 (B.K.C. Lo, ed., Humana Press, Totowa, NJ), pp.255-268 (2003). Клетки-хозяева включают культивируемые клетки, например, культивируемые клетки млекопитающих, клетки дрожжей, клетки насекомых, бактериальные клетки и клетки растений, среди которых названо лишь немного, а также клетки, содержащиеся внутри трансгенного животного, трансгенного растения или культивируемой ткани растения или животного. В одной форме осуществления клетка-хозяин представляет собой эукариотическую клетку, предпочтительно клетку млекопитающего, такую как клетка яичника китайского хомячка (СНО), клетка эмбриональных почек человека (HEK) или лимфоидная клетка (например, клетка Y0, NS0, Sp20).
В данной области техники известны стандартные методы экспрессии чужеродных генов в этой системе. Клетки, экспрессирующие полипептид, включающий либо тяжелую, либо легкую цепь антигенсвязывающего домена, такого как антитело, могут быть сконструированы таким образом, что они также экспрессируют другие из цепей антитела, так что экспрессируемый продукт представляет собой антитело, имеющее и тяжелую, и легкую цепь.
В одной форме осуществления предложен способ получения биспецифической антигенсвязывающей молекулы согласно изобретению, включающий культивирование клетки-хозяина, содержащей полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу, предложенную в данной работе, в условиях, подходящих для экспрессии биспецифической антигенсвязывающей молекулы, и выделение биспецифической антигенсвязывающей молекулы из клетки-хозяина (или из культуральной среды клетки-хозяина).
Компоненты биспецифической антигенсвязывающей молекулы генетически слиты друг с другом. Биспецифическая антигенсвязывающая молекула может быть сконструирована таким образом, что ее компоненты слиты непосредственно друг с другом или опосредованно с помощью линкерной последовательности. Композицию и длину линкера можно определить в соответствии со способами, хорошо известными в данной области техники, и можно тестировать его на эффективность. Примеры линкерных последовательностей между различными компонентами биспецифических антигенсвязывающих молекул находятся в последовательностях, предложенных в данной работе. При желании могут быть также включены дополнительные последовательности, чтобы включить сайт расщепления для разделения индивидуальных компонентов слияния, например, последовательность распознавания эндопептидазы.
В некоторых формах осуществления Fab фрагменты, составляющие часть биспецифических антигенсвязывающих молекул, включают по меньшей мере вариабельную область антитела, способную к связыванию антигенной детерминанты. Вариабельные области могут составлять часть природных или неприродных антител и их фрагментов, или могут быть получены из них. Способы получения поликлональных антител и моноклональных антител хорошо известны в данной области техники (см., например, руководство Harlow and Lane, "Antibodies, a laboratory manual", Cold Spring Harbor Laboratory, 1988). Неприродные антитела можно конструировать, используя твердофазный пептидный синтез, можно получать рекомбинантным путем (например, как раскрыто в патенте US №4186567) или можно получать, например, путем скрининга комбинаторных библиотек, содержащих вариабельные тяжелые цепи и вариабельные легкие цепи (см., например, патент US №5969108, McCafferty).
В биспецифических антигенсвязывающих молекулах по изобретению можно использовать любые виды антител животных, фрагмент антитела, антигенсвязывающий домен или вариабельную область. Неограничивающие антитела, фрагменты антител, антигенсвязывающие домены или вариабельные области, полезные в настоящем изобретении, могут иметь происхождение от мышей, приматов или человека. Если биспецифическая антигенсвязывающая молекула предназначена для применения у человека, можно использовать химерную форму антитела, в которой константные области антитела имеют происхождение от человека. Гуманизированная или полностью человеческая форма антитела может быть также получена в соответствии со способами, хорошо известными в данной области техники (см., например, патент US №5565332, Winter). Гуманизация может быть достигнута различными способами, включающими, но не ограниченными ими, следующие способы: (а) прививание CDR антитела не человеческого происхождения (например, донорного антитела) на каркасную область и константные области человека (например, реципиентное антитело) с сохранением или без сохранения принципиально важных каркасных остатков (например, остатков, важных для сохранения хорошего антигенсвязывающего сродства или функций антитела), (b) прививание только участков, определяющих специфичность (SDR; от англ. "specificity-determining regions"; или a-CDR; остатков, принципиально важных для взаимодействия антитела с антигеном) не человеческого происхождения на каркасную область и константные области человека, или (с) прививание полноразмерных вариабельных доменов не человеческого происхождения, но "маскировка" их человекоподобным участком путем замены поверхностных остатков. Обзор гуманизированных антител и способов их получения приведен, например, в статье Almagro and Fransson, Front Biosci 13, 1619-1633 (2008), и дополнительно раскрыт, например, в следующих документах: Riechmann et al., Nature 332, 323-329 (1988); Queen et al., Proc Natl Acad Sci USA 86, 10029-10033 (1989); патенты US №№5821337, 7527791, 6982321 и 7087409; Jones et al., Nature 321, 522-525 (1986); Morrison et al., Proc Natl Acad Sci 81, 6851-6855 (1984); Morrison and Oi, Adv Immunol 44, 65-92 (1988); Verhoeyen et al., Science 239, 1534-1536 (1988); Padlan, Molec Immun 31(3), 169-217 (1994); Kashmiri et al., Methods 36, 25-34 (2005) (раскрыто прививание SDR (a-CDR)); Padlan, Mol Immunol 28, 489-498 (1991) (раскрыто "изменение поверхности"); Dall'Acqua et al., Methods 36, 43-60 (2005) (раскрыта "перетасовка FR"); и Osbourn et al., Methods 36, 61-68 (2005) и Klimka et al., Br J Cancer 83, 252-260 (2000) (раскрыт метод "направленной селекции" перетасовки FR). Антитела человека и вариабельные области человека могут быть получены, используя различные методы, известные в данной области техники. Антитела человека в целом описаны в статье van Dijk and van de Winkel, Curr Opin Pharmacol 5, 368-74 (2001) и Lonberg, Curr Opin Immunol 20, 450-459 (2008). Вариабельные области человека могут составлять часть моноклональных антител человека, полученных методом гибридомы (см., например, кн. Monoclonal Antibody Production Techniques and Applications, pp.51-63 (Marcel Dekker, Inc., New York, 1987)), или могут быть получены из этих антител. Антитела человека и вариабельные области человека могут быть также получены путем введения иммуногена трансгенному животному, модифицированному таким образом, чтобы продуцировать интактные антитела человека или интактные антитела с вариабельными областями человека в ответ на антигенную стимуляцию (см., например, статью Lonberg, Nat Biotech 23, 1117-1125 (2005). Антитела человека и вариабельные области человека могут быть также получены путем выделения клона Fv последовательностей вариабельной области, выделенного из библиотеки фагового дисплея человеческого происхождения (см., например, статью Hoogenboom et al. в Methods in Molecular Biology 178, 1-37 (O'Brien et al., ed., Human Press, Totowa, NJ, 2001); and McCafferty et al., Nature 348, 552-554; Clackson et al., Nature 352, 624-628 (1991)). В характерном случае фаг выявляет фрагменты антитела, либо в виде одноцепочечных Fv (scFv) фрагментов, либо в виде Fab фрагментов.
В некоторых формах осуществления Fab фрагменты, полезные в настоящем изобретении, сконструированы таким образом, что обладают усиленным связывающим сродством, согласно, например, способам, раскрытым в публикации заявки на патент US №2004/0132066, полное содержание которой включено в настоящее описание посредством ссылки. Способность биспецифической антигенсвязывающей молекулы по изобретению к связыванию со специфичной антигенной детерминантой можно измерить либо посредством твердофазного иммуноферментного анализа (ELISA), либо другими методами, известными специалистам в данной области техники, например, методом поверхностного плазменного резонанса (анализируемого на системе BIACORE Т100) (Liljeblad, et al., Glyco J 17, 323-329 (2000)) и традиционными анализами связывания (Heeley, Endocr Res 28, 217-229 (2002)). Анализ конкурентного связывания можно использовать для идентификации антитела, фрагмента антитела, антигенсвязывающего домена или вариабельного домена, конкурирующего с референсным антителом за связывание с определенным антигеном. В некоторых формах осуществления такое конкурирующее антитело связывается с тем же эпитопом (например, линейным или конформационным эпитопом), который связывается референсным антителом. Подробные иллюстративные способы картирования эпитопа, с которым связывается антитело, приведены в статье Morris (1996) "Epitope Mapping Protocols" в кн. Methods in Molecular Biology vol. 66 (Humana Press, Totowa, NJ). При иллюстративном конкурентном анализе иммобилизованный антиген инкубируют в растворе, содержащем первое меченое антитело, связывающееся с антигеном, и второе немеченое антитело, тестируемое на способность к конкуренции с первым антителом за связывание с антигеном. Второе антитело может находиться в супернатанте гибридомы. В качестве контроля иммобилизованный антиген инкубируют в растворе, содержащем первое меченое антитело, но не содержащем второе немеченое антитело. После инкубации в условиях, пермиссивных для связывания первого антитела с антигеном, избыток несвязанного антитела удаляют и измеряют количество метки, ассоциированной с иммобилизованным антигеном. Если количество метки, ассоциированной с иммобилизованным антигеном, существенно снижено в тестируемом образце относительно контрольного образца, это указывает на то, что второе антитело конкурирует с первым антителом за связывание с антигеном. См. руководство Harlow and Lane (1988) Antibodies: A Laboratory Manual ch.14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY).
Биспецифические антигенсвязывающие молекулы, полученные, как описано в данной работе, могут быть очищены методами, известными в данной области техники, такими как высокоэффективная жидкостная хроматография (ВЭЖХ), ионообменная хроматография, гель-электрофорез, аффинная хроматография, эксклюзионная хроматография и тому подобное. Действительные условия, используемые для очистки конкретного белка, отчасти зависят от таких факторов, как суммарный заряд, гидрофобность, гидрофильность и т.д., и очевидны специалистам в данной области техники. Для очистки аффинной хроматографией можно использовать антитело, лиганд, рецептор или антиген, с которыми связывается биспецифическая антигенсвязывающая молекула. Например, для очистки биспецифических антигенсвязывающих молекул по изобретению аффинной хроматографией можно использовать матрикс с белком А или белком G. Последовательную аффинную хроматографию на белке А или G и эксклюзионную хроматографию можно использовать для выделения биспецифической антигенсвязывающей молекулы по существу, как раскрыто в Примерах. Чистоту биспецифической антигенсвязывающей молекулы можно определить любым из ряда хорошо известных аналитических способов, включающих гель-электрофорез, жидкостную хроматографию высокого давления и тому подобное. Например, было показано, что слитые белки тяжелой цепи, экспрессируемые, как раскрыто в Примерах, являются интактными и правильно уложенными, как продемонстрировано с помощью восстанавливающего электрофореза в ДСН-ПААГ (см., например, фиг.3). Было разделено три полосы, имеющие молекулярную массу, составляющую приблизительно Mr 25000, Mr 50000 и Mr 75000, соответствующую предсказанным молекулярным массам легких цепей, тяжелой цепи и слитого белка тяжелая цепь/тяжелая цепь Fab биспецифической антигенсвязывающей молекулы, соответственно.
Анализы
Биспецифические антигенсвязывающие молекулы, предложенные в настоящем изобретении, могут быть идентифицированы, подвергнуты скринингу или охарактеризованы на их физические/химические свойства и/или биологические активности с помощью различных анализов, известных в данной области техники.
Анализы сродства
Сродство биспецифической антигенсвязывающей молекулы к рецептору Fc или к антигену-мишени можно определить в соответствии со способами, представленными в Примерах, с помощью поверхностного плазмонного резонанса (SPR), используя стандартное оборудование, такое как прибор BIAcore (GE Healthcare), и рецепторов или белков-мишеней, которые могут быть получены с помощью рекомбинантной экспрессии. Альтернативно связывание биспецифических антигенсвязывающих молекул с различными рецепторами или антигенами-мишенями можно оценивать, используя линии клеток, экспрессирующие конкретный рецептор или антиген-мишень, например, с помощью проточной цитометрии (FACS). Конкретная иллюстративная форма осуществления измерения связывающего сродства раскрыта далее и в приведенных ниже Примерах.
Согласно одной форме осуществления Ко измеряют с помощью поверхностного плазменного резонанса, используя прибор BIACORE® T100 (GE Healthcare) при 25°С.
Для анализа взаимодействия между Fc-участком и рецепторами Fc His-меченый рекомбинантный Fc-рецептор иммобилизуют антителом против пента-His (Qiagen), иммобилизованным на микрочипах СМ5, и биспецифические конструкции используют в качестве аналитов. Кратко, биосенсорные чипы с карбоксиметилированным декстраном (СМ5, GE Healthcare) активируют N-этил-N'-(3-диметиламинопропил)-карбодиимида гидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) согласно инструкциям изготовителя. Антитело против пента-His разводят 10 мМ ацетатом натрия, рН 5,0, до 40 м кг/мл, после чего наносят при скорости тока 5 мкл/мин до достижения приблизительно 6500 резонансных единиц (RU; от англ. "resonance unit") связанного белка. После нанесения лиганда наносят 1 М этаноламин для блокирования непрореагировавших групп. Затем Fc-рецептор иммобилизуют в течение 60 с в количестве 4 или 10 нМ. Для кинетических измерений четырехкратные серийные разведения биспецифической конструкции (диапазон от 500 нМ до 4000 нМ) наносят в буфере HBS-EP (GE Healthcare, 10 мМ N-2-гидроксиэтилпиперазин-N-2-этансульфокислота (ГЭПЭС), 150 мМ NaCl, 3 мМ этилендиаминтетрауксусная кислота (ЭДТА), 0,05% сурфактант Р20, рН 7,4) при 25°С со скоростью тока 30 мкл/мин в течение 120 с.
Чтобы определить сродство к антигену-мишени, биспецифические конструкции иммобилизуют антителом, специфичным против Fab человека (GE Healthcare), иммобилизованным на поверхности активированного СМ5-сенсорного чипа, как описано для антитела против пента-His. Конечное количество связанного белка составляет приблизительно 12000RU. Биспецифические конструкции иммобилизуют в течение 90 с в количестве 300 нМ. Антигены-мишени пропускают через проточные ячейки в течение 180 с в диапазоне концентраций от 250 до 1000 нМ при скорости тока 30 мкл/мин. Мониторинг диссоциации проводят в течение 180 с.
Групповые различия показателя преломления корректируют путем вычитания ответа, полученного на референсной проточной ячейке. Ответ состояния равновесия использовали, чтобы вывести константу диссоциации KD путем приведения в соответствие нелинейной кривой изотермы связывания Лангмюра. Скорости ассоциации (kon) и скорости диссоциации (koff) вычисляют, используя простую модель связывания Лангмюра один к одному (программное обеспечение оценки BIACORE® T100 версия 1.1.1), путем одновременного приведения в соответствие сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (KD) вычисляют как отношение koff/kon. См., например, статью Chen et al., J Mol Biol 293, 865-881 (1999).
Анализы активности
Биологическую активность биспецифических антигенсвязывающих молекул по изобретению можно измерить с помощью различных анализов, известных в данной области техники, включающих анализы, описанные в Примерах. Биологические активности могут, например, включать индукцию пролиферации Т-клеток, индукцию передачи сигнала в Т-клетках, индукцию экспрессии маркеров активации в Т-клетках, индукцию секреции цитокинов Т-клетками, индукцию передачи сигнала в клетках-мишенях, таких как опухолевые клетки или клетки опухолевой стромы, ингибирование пролиферации клеток-мишеней, индукцию лизиса клеток-мишеней и индукцию регрессии опухоли и/или улучшения выживания.
Композиции, препараты и пути введения
В следующем аспекте в изобретении предложены фармацевтические композиции, содержащие любую из биспецифических антигенсвязывающих молекул, предложенных в данном изобретении, например, применяемые в любом из описанных ниже терапевтических способов. В одной форме осуществления фармацевтическая композиция содержит любую из биспецифических антигенсвязывающих молекул, предложенных в данном изобретении, и фармацевтически приемлемый носитель. В другой форме осуществления фармацевтическая композиция содержит любую из биспецифических антигенсвязывающих молекул, предложенных в данном изобретении, и по меньшей мере один дополнительный терапевтический агент, например, как раскрыто ниже.
Далее предложен способ получения биспецифической антигенсвязывающей молекулы по изобретению в форме, подходящей для введения in vivo, включающий следующие стадии: (а) получение биспецифической антигенсвязывающей молекулы согласно изобретению и (b) включение биспецифической антигенсвязывающей молекулы в препарат по меньшей мере с одним фармацевтически приемлемым носителем, в результате чего готовят препарат биспецифической антигенсвязывающей молекулы для введения in vivo.
Фармацевтические композиции по настоящему изобретению содержат терапевтически эффективное количество одной или более биспецифических антигенсвязывающих молекул, растворенных или диспергированных в фармацевтически приемлемом носителе. Выражения "фармацевтически или фармакологически приемлемый" относятся к молекулярным субстанциям и композициям, в целом нетоксичным для реципиентов при применяемых дозировках и концентрациях, то есть не производят вредную, аллергическую или иную нежелательную реакцию пи введении животному, такому как, например, человек, если это целесообразно. Получение фармацевтической композиции, содержащей по меньшей мере одну биспецифическую антигенсвязывающую молекулу и необязательно дополнительный активный ингредиент, известно специалистам в данной области техники в свете настоящего описание, как проиллюстрировано в кн. Remington's Pharmaceutical Sciences, 18th Ed. Mack Printing Company, 1990, включенной в настоящее описание посредством ссылки. Кроме того, понятно, что для введения животному (например, человеку) препараты должны соответствовать стандартам стерильности, пирогенности, общей безопасности и чистоты на основании требований бюро биологических стандартов Управления по контролю за пищевыми продуктами и лекарственными средствами США (FDA; от англ. Food and Drug Administration) или соответствующих государственных органов в других странах. Предпочтительные композиции представляют собой лиофилизированные препараты или водные растворы. Как используют в данной работе, "фармацевтически приемлемый носитель" включает любой и все из следующих веществ: растворителей, буферов, диспергирующих сред, покрытий, сурфактантов, антиоксидантов, консервантов (например, антибактериальных агентов, противогрибковых агентов), изотонических агентов, агентов, замедляющих всасывание, солей, консервантов, антиоксидантов, белков, лекарственных средств, стабилизаторов лекарственных средств, полимеров, гелей, связующих веществ, наполнителей, разрыхлителей, смазывающих веществ, подсластителей, корригентов, красителей, где такие подобные вещества и их комбинации должны быть известны обычным специалистам в данной области техники (см., например, кн. Remington's Pharmaceutical Sciences, 18th Ed. Mack Printing Company, 1990, pp.1289-1329, включенную в настоящее описание посредством ссылки). За исключением тех случаев, когда какой-либо традиционный носитель несовместим с активным ингредиентом, рассмотрено его применение в терапевтических и фармацевтических композициях.
Композиция может содержать различные типы носителей в зависимости от того, вводят ли ее в твердой, жидкой или аэрозольной форме, и необходимо ли, чтобы она была стерильной, для таких путей введения, как инъекция. Биспецифические антигенсвязывающие молекулы по настоящему изобретению (и любой дополнительный терапевтический агент) можно вводить внутривенно, внутрикожно, внутриартериально, интраперитонеально, внутрь повреждения, интракраниально, внутрисуставным путем, внутрь простаты, внутриселезеночно, интраренально, внутриплеврально, интратрахеально, интраназально, интравитреально, интравагинально, интраректально, внутрь опухоли, внутримышечно, подкожно, субконъюнктивально, интравезикулярно, в слизистую оболочку, интраперикардиально, интраумбиликально, внутриглазным путем, перорально, местно, локально, путем ингаляции (например, аэрозольной ингаляции), инъекции, инфузии, непрерывной инфузии, локализованной перфузии, непосредственно омывающей клетки-мишени, посредством катетера, посредством лаважа, в виде кремов, в жидких композициях (например, в липосомах), либо другим способом или посредством любой комбинации вышеописанных способов, как должно быть известно обычным специалистам в данной области техники (см., например, кн. Remington's Pharmaceutical Sciences, 18th Ed. Mack Printing Company, 1990, pp.1289-1329, включенную в настоящее описание посредством ссылки). Для введения полипептидных молекул, таких как биспецифические антигенсвязывающие молекулы по изобретению, чаще всего используют парентеральное введение, в частности, внутривенное введение.
Парентеральные композиции включают композиции, предназначенные для введения путем инъекции, например, подкожной, внутрикожной, внутрь повреждения, внутривенной, внутриартериальной, внутримышечной, подоболочечной или интраперитонеальной инъекции. Для инъекции биспецифические антигенсвязывающие молекулы по изобретению можно включать в препараты в виде водных растворов, предпочтительно в физиологически совместимых буферах, таких как раствор Хэнкса, раствор Рингера или физиологический солевой буфер. Этот раствор может содержать вспомогательные вещества, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты. Альтернативно биспецифические антигенсвязывающие молекулы могут находиться в форме порошка для восстановления подходящей основой, например, стерильной апирогенной водой, перед применением. Стерильные инъекционные растворы готовят путем включения биспецифических антигенсвязывающих молекул по изобретению в необходимом количестве в подходящую основу с различными другими необходимыми ингредиентами, перечисленными ниже. Стерилизация может быть легко выполнена, например, путем фильтрования через стерильные фильтрационные мембраны. Как правило, дисперсии готовят путем включения различных стерилизованных активных ингредиентов в стерильную основу, содержащую базовую дисперсионную среду и/или другие ингредиенты. В случае стерильных порошков для приготовления стерильных инъекционных растворов, суспензий или эмульсий предпочтительные способы получения представляют собой методы вакуумной сушки и сушки вымораживанием, в результате которых получают порошок активного ингредиента с добавлением любого желаемого дополнительного ингредиента из его жидкой среды, предварительно стерилизованной фильтрованием. Жидкую среду при необходимости следует соответствующим образом забуферить, и жидкий разбавитель перед инъекцией сначала делают изотоническим достаточным количеством физиологического раствора или глюкозы. Композиция должна быть стабильной в условиях получения и хранения, и должна быть предохранена против заражения микроорганизмами, такими как бактерии и грибы. Понятно, что загрязнение эндотоксином следует поддерживать минимально на безопасном уровне, например, менее 0,5 нг/мг белка. Подходящие фармацевтически приемлемые носители включают, но не ограничены ими, следующие вещества: буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включающие аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид; бензетония хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол и мета-крезол); низкомолекулярные (менее чем приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные сурфактанты, такие как полиэтиленгликоль (ПЭГ). Водные инъекционные суспензии могут содержать соединения, повышающие вязкость суспензии, такие как натриевая соль карбоксиметилцеллюлозы, сорбит, декстран или тому подобное. Необязательно суспензия может также содержать подходящие стабилизаторы или агенты, повышающие растворимость соединений, чтобы дать возможность приготовления высоко концентрированных растворов. Кроме того, суспензии активных соединений можно готовить в виде соответствующих масляных инъекционных суспензий. Подходящие липофильные растворители или основы включают нелетучие масла, такие как кунжутное масло, либо синтетические сложные эфиры жирных кислот, такие как этилолеаты или триглицериды, либо липосомы.
Активные ингредиенты могут быть заключены в микрокапсулы, полученные, например, методами коацервации или путем полимеризации на границе раздела фаз, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и полиметакрилатные микрокапсулы, соответственно, в коллоидные системы доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методы раскрыты в кн. Remington's Pharmaceutical Sciences (18th Ed. Mack Printing Company, 1990). Можно готовить препараты пролонгированного высвобождения. Подходящие примеры препаратов пролонгированного высвобождения включают полупроницаемые матриксы из твердых гидрофобных полимеров, содержащие полипептид, находящиеся в форме формованных изделий, например, пленок или микрокапсул. В конкретных формах осуществления пролонгированное всасывание инъекционной композиции может быть вызвано путем использования в композициях агентов, замедляющих всасывание, таких как, например, моностеарат алюминия, желатин или их комбинации.
В дополнение к композициям, раскрытым выше, биспецифические антигенсвязывающие молекулы можно также включать в депо-препарат. Такие препараты длительного действия можно вводить путем имплантации (например, подкожно или внутримышечно) или путем внутримышечной инъекции. Так, например, биспецифические антигенсвязывающие молекулы можно включать в препараты с подходящими полимерными или гидрофобными веществами (например, в виде эмульсии в приемлемом масле) или ионообменными смолами, либо в виде очень слаборастворимых производных, например, в виде очень слаборастворимой соли.
Фармацевтические композиции, содержащие биспецифические антигенсвязывающие молекулы по изобретению, могут быть получены традиционными способами смешивания, растворения, эмульгирования, инкапсуляции, заключения или лиофилизации. Фармацевтические композиции могут быть получены традиционным способом, используя один или более физиологически приемлемых носителей, разбавителей, наполнителей или вспомогательных веществ, облегчающих обработку белков для получения препаратов, которые можно применять фармацевтически. Точная композиция зависит от выбранного пути введения.
Биспецифические антигенсвязывающие молекулы можно включать в композицию в виде свободной кислоты или основания, в нейтральной или солевой форме. Фармацевтически приемлемые соли представляют собой соли, по существу сохраняющие биологическую активность свободной кислоты или основания. Эти соли включают соли присоединения кислоты, например, образованные со свободными аминогруппами белковой композиции, или образованные с неорганическими кислотами, такими как, например, соляная или фосфорная кислота, или с такими органическими кислотами, как уксусная, щавелевая, винная или миндальная кислота. Соли, образованные со свободными карбоксильными группами, могут быть также получены из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа; или из таких органических оснований, как изопропиламин, триметиламин, гистидин или прокаин. Фармацевтически соли склонны к большей растворимости в водных и других протонных растворителях, чем соответствующие формы свободного основания.
Терапевтические способы и композиции
Любую из биспецифических антигенсвязывающих молекул, предложенных в данном изобретении, можно применять в терапевтических способах. Биспецифические антигенсвязывающие молекулы по изобретению можно применять, например, при лечении раков.
Для применения в терапевтических способах биспецифические антигенсвязывающие молекулы по изобретению следует включать в препараты, дозировать и вводить способом, согласующимся с надлежащей медицинской практикой. Факторы, которые следует учитывать в данном контексте, включают конкретное расстройство, подлежащее лечению, конкретное млекопитающее, подлежащее лечению, клиническое состояние индивидуального пациента, причину расстройства, сайт доставки агента, способ введения, режим введения и другие факторы, известные практикующим медицинским работникам.
В одном аспекте предложены биспецифические антигенсвязывающие молекулы по изобретению, применяемые в качестве лекарственного средства. В следующих аспектах предложены биспецифические антигенсвязывающие молекулы по изобретению, применяемые при лечении заболевания. В некоторых формах осуществления предложены биспецифические антигенсвязывающие молекулы по изобретению, применяемые в способе лечения. В одной форме осуществления в изобретении предложена биспецифическая антигенсвязывающая молекула, как раскрыто в данном изобретении, применяемая при лечении заболевания у индивидуума, нуждающегося в этом. В некоторых формах осуществления в изобретении предложена биспецифическая антигенсвязывающая молекула, применяемая в способе лечения индивидуума, страдающего заболеванием, включающем введение этому индивидууму терапевтически эффективного количества биспецифической антигенсвязывающей молекулы. В некоторых формах осуществления заболевание, подлежащее лечению, представляет собой пролиферативное расстройство. В конкретной форме осуществления заболевание представляет собой рак. В некоторых формах осуществления способ дополнительно включает введение индивидууму терапевтически эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, противоракового агента, если заболевание, подлежащее лечению, представляет собой рак. "Индивидуум" согласно любой из описанных выше форм осуществления представляет собой млекопитающее, предпочтительно человека.
В следующем аспекте в изобретении предложено применение биспецифической антигенсвязывающей молекулы по изобретению при получении или приготовлении лекарственного средства. В одной форме осуществления лекарственное средство предназначено для лечения заболевания у индивидуума, нуждающегося в этом. В следующей форме осуществления лекарственное средство предназначено для применения в способе лечения заболевания, включающем введение индивидууму, страдающему заболеванием, терапевтически эффективного количества лекарственного средства. В некоторых формах осуществления заболевание, подлежащее лечению, представляет собой пролиферативное расстройство. В конкретной форме осуществления заболевание представляет собой рак. В одной форме осуществления способ дополнительно включает введение индивидууму терапевтически эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, противоракового агента, если заболевание, подлежащее лечению, представляет собой рак. "Индивидуум" согласно любой из описанных выше форм осуществления представляет собой млекопитающее, предпочтительно человека.
В следующем аспекте в изобретении предложен способ лечения заболевания. В одной форме осуществления способ включает введение индивидууму, страдающему заболеванием, терапевтически эффективного количества биспецифической антигенсвязывающей молекулы по изобретению. В одной форме осуществления данному индивидууму вводят композицию, содержащую биспецифическую антигенсвязывающую молекулу по изобретению в фармацевтически приемлемой форме. В некоторых формах осуществления заболевание, подлежащее лечению, представляет собой пролиферативное расстройство. В конкретной форме осуществления заболевание представляет собой рак. В некоторых формах осуществления способ дополнительно включает введение индивидууму терапевтически эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, противоракового агента, если заболевание, подлежащее лечению, представляет собой рак. "Индивидуум" согласно любой из описанных выше форм осуществления представляет собой млекопитающее, предпочтительно человека.
В некоторых формах осуществления заболевание, подлежащее лечению, представляет собой пролиферативное расстройство, в частности, рак. Неограничивающие примеры раков включают рак мочевого пузыря, рак головного мозга, рак головы и шеи, рак поджелудочной железы, рак легкого, рак молочной железы, рак яичника, рак матки, рак шейки матки, рак эндометрия, рак пищевода, рак ободочной кишки, колоректальный рак, рак прямой кишки, рак желудка, рак простаты, рак крови, рак кожи, чешуйчато-клеточную карциному, рак кости и рак почки. Другие расстройства клеточной пролиферации, которые можно лечить применением биспецифической антигенсвязывающей молекулы по настоящему изобретению, включают, но не ограничены ими, новообразования, имеющие следующие локализации: в брюшной полости, кости, молочной железе, пищеварительной системе, печени, поджелудочной железе, брюшине, эндокринных железах (надпочечнике, паращитовидной железе, гипофизе, семенниках, яичнике, тимусе, щитовидной железе), глазу, голове и шее, нервной системе (центральной и периферической), лимфатической системе, тазу, коже, мягкой ткани, селезенке, грудной полости и мочеполовой системе. Также включены предраковые состояния или повреждения и метастазы рака. В некоторых формах осуществления рак выбран из группы, состоящей из следующих раков: почечно-клеточного рака, рака кожи, рака легкого, колоректального рака, рака молочной железы, рака головного мозга, рака головы и шеи. Специалист в данной области техники легко распознает, что во многих случаях биспецифическая антигенсвязывающая молекула может не обеспечить излечение, но может обеспечить только частичную пользу. В некоторых формах осуществления физиологическое изменение, обладающее некоторой пользой, также рассматривают как терапевтически благоприятное. Таким образом, в некоторых формах осуществления количество биспецифической антигенсвязывающей молекулы, обеспечивающее физиологическое изменение, считают "эффективным количеством" или "терапевтически эффективным количеством". Субъект, пациент или индивидуум, нуждающийся в лечении, в характерном случае представляет собой млекопитающее, более конкретно человека.
В некоторых формах осуществления эффективное количество биспецифической антигенсвязывающей молекулы по изобретению вводят в клетку. В других формах осуществления терапевтически эффективное количество биспецифической антигенсвязывающей молекулы по изобретению вводят индивидууму для лечения заболевания.
Для предупреждения или лечения заболевания соответствующая дозировка биспецифической антигенсвязывающей молекулы по изобретению (при применении отдельно или в комбинации с одним или более других дополнительных терапевтических агентов) зависит от типа заболевания, подлежащего лечению, пути введения, массы тела пациента, типа биспецифической антигенсвязывающей молекулы, тяжести и хода заболевания независимо от того, вводят ли биспецифическую антигенсвязывающую молекулу для превентивных или терапевтических целей; от предшествующих или одновременных терапевтических вмешательств, истории болезни пациента и ответа на биспецифическую антигенсвязывающую молекулу, а также от мнения лечащего врача. Практикующий врач, ответственный за введение, в любом случае определит концентрацию активного ингредиента (ингредиентов) в композиции и соответствующую дозу (дозы) для индивидуального субъекта. В данной работе рассмотрены различные режимы дозирования, включающие, но не ограниченные ими, однократные или многократные введения в течение различных промежутков времени, болюсное введение и прерывистую инфузию.
Биспецифическую антигенсвязывающую молекулу целесообразно вводить пациенту за один период времени или на протяжении серии терапий. В зависимости от типа и тяжести заболевания кандидатом на начальную дозу для введения пациенту может быть доза, составляющая приблизительно от 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг - 10 мг/кг) биспецифической антигенсвязывающей молекулы, либо, например, путем одного или более отдельных введений, либо путем непрерывной инфузии. Одна типичная суточная доза может находиться в диапазоне приблизительно от 1 мкг/кг до 100 мг/кг или более в зависимости от факторов, упомянутых выше. Для повторных введений в течение нескольких суток или дольше в зависимости от состояния терапию, как правило, следует продолжать до тех пор, пока не произойдет желаемое подавление симптомов заболевания. Одна иллюстративная дозировка биспецифической антигенсвязывающей молекулы находится в диапазоне от приблизительно 0,005 мг/кг до приблизительно 10 мг/кг. В других неограничивающих примерах доза может также включать от приблизительно 1 микрограмм/кг массы тела, приблизительно 5 микрограммов/кг массы тела, приблизительно 10 микрограммов/кг массы тела, приблизительно 50 микрограммов/кг массы тела, приблизительно 100 микрограммов/кг массы тела, приблизительно 200 микрограммов/кг массы тела, приблизительно 350 микрограммов/кг массы тела, приблизительно 500 микрограммов/кг массы тела, приблизительно 1 миллиграмм/кг массы тела, приблизительно 5 миллиграммов/кг массы тела, приблизительно 10 миллиграммов/кг массы тела, приблизительно 50 миллиграммов/кг массы тела, приблизительно 100 миллиграммов/кг массы тела, приблизительно 200 миллиграммов/кг массы тела, приблизительно 350 миллиграммов/кг массы тела, приблизительно 500 миллиграммов/кг массы тела до приблизительно 1000 мг/кг массы тела или более на введение, а также любой диапазон, производный от данного диапазона. В неограничивающих примерах диапазона, производного от чисел, приведенных в данной работе, диапазон от приблизительно 5 мг/кг массы тела до приблизительно 100 мг/кг массы тела, от приблизительно 5 микрограммов/кг массы тела до приблизительно 500 миллиграммов/кг массы тела и т.д. можно вводить, основываясь на числах, раскрытых выше. Таким образом, пациенту можно вводить одну или более доз, составляющих приблизительно 0,5 мг/кг, 2,0 мг/кг, 5,0 мг/кг или 10 мг/кг (или любую их комбинацию). Такие дозы можно вводить периодически, например, каждую неделю или каждые три недели (например, таким образом, что пациент получает от приблизительно двух до приблизительно двадцати или, например, шесть доз биспецифической антигенсвязывающей молекулы). Можно вводить начальную, более высокую насыщающую дозу, за которой следует одна или более чем одна более низкая доза. Тем не менее, могут быть полезны другие режимы дозирования. За ходом этой терапии легко следить с помощью традиционных методов и анализов.
Биспецифические антигенсвязывающие молекулы по изобретению, как правило, применяют в количестве, эффективном для достижения предназначенной цели. Для применения при лечении или предупреждении болезненного состояния биспецифические антигенсвязывающие молекулы по изобретению или их фармацевтические композиции вводят или применяют в терапевтически эффективном количестве. Определение терапевтически эффективного количества находится в пределах надлежащей компетенции специалистов в данной области техники, в частности, в свете подробного описания, приведенного в данной работе.
Для системного введения терапевтически эффективная доза первоначально может быть установлена на основании анализов in vitro, таких как анализы на клеточной культуре. Затем дозу можно разрабатывать в моделях на животных до достижения уровня циркулирующей концентрации, включающего значение IC50, определенное в клеточной культуре. Такую информацию можно использовать, чтобы более точно определить полезные дозы у людей.
Начальные дозы также могут быть установлены на основании данных in vivo, например, в моделях на животных, используя методы, хорошо известные в данной области техники. Обычный специалист в данной области техники сможет легко оптимизировать введение людям на основании данных, полученных на животных.
Величину и интервал дозы можно регулировать индивидуально до получения уровней биспецифических антигенсвязывающих молекул в плазме, достаточных для поддержания терапевтического эффекта. Обычные дозировки для введения пациенту путем инъекции находятся в диапазоне от приблизительно 0,1 до 50 мг/кг/сутки, в характерном случае от приблизительно 0,5 до 1 мг/кг/сутки. Терапевтически эффективные уровни в плазме могут быть достигнуты путем введения многократных доз каждые сутки. Уровни в плазме можно измерить, например, с помощью ВЭЖХ.
В случаях локального введения или избирательного захвата эффективная локальная концентрация биспецифических антигенсвязывающих молекул может не быть связана с концентрацией в плазме. Специалист в данной области техники способен оптимизировать терапевтически эффективные локальные дозировки без ненужного экспериментирования.
Терапевтически эффективная доза биспецифических антигенсвязывающих молекул, раскрытых в данной работе, в целом обеспечивает терапевтическую пользу, не вызывая существенную токсичность. Токсичность и терапевтическую эффективность биспецифической антигенсвязывающей молекулы можно определить стандартными фармацевтическими методами на культуре клеток или подопытных животных. Анализы на культуре клеток и исследования на животных можно использовать для определения LD50 (дозы, летальной для 50% популяции) и ED50 (дозы, терапевтически эффективной в 50% популяции). Отношение дозы между токсическими и терапевтическими эффектами представляет собой терапевтический индекс, который может быть выражен в виде отношения LD50/ED50. Предпочтительны биспецифические антигенсвязывающие молекулы, проявляющие высокие терапевтические индексы. В одной форме осуществления биспецифическая антигенсвязывающая молекула согласно настоящему изобретению проявляет высокий терапевтический индекс. Данные, полученные из анализов культуры клеток и исследований на животных, можно использовать при разработке диапазонов дозировок, подходящих для применения у людей. Дозировка предпочтительно находится в пределах диапазона циркулирующих концентраций, включающего ED50 при небольшой или при отсутствии токсичности. Дозировка может варьировать в пределах этого диапазона в зависимости от ряда факторов, например, от применяемой лекарственной формы, от используемого пути введения, от состояния субъекта и тому подобного. Точную композицию, путь введения и дозировку может выбрать индивидуальный врач в свете состояния пациента (см., например, Fingl et al., 1975, в кн.: The Pharmacological Basis of Therapeutics, Ch. 1, p.1, полностью включенной в данное описание посредством ссылки).
Лечащий врач для пациентов, которых лечат биспецифическими антигенсвязывающими молекулами по изобретению, должен знать, как и когда остановить, прервать или отрегулировать введение вследствие токсичности, дисфункции органа и тому подобного. Напротив, лечащий врач должен также знать, как отрегулировать лечение до более высоких уровней, если клинический ответ был неадекватным (препятствующую токсичность). Величина вводимой дозы при лечении расстройства, представляющего интерес, варьирует в зависимости от тяжести состояния, подлежащего лечению, от пути введения и тому подобного. Тяжесть состояния можно, например, оценить отчасти стандартными способами прогностической оценки. Кроме того, доза и, возможно, частота дозы также варьирует в соответствии с возрастом, массой тела и ответом индивидуального пациента.
Другие агенты и терапии
Биспецифические антигенсвязывающие молекулы по изобретению можно вводить в комбинации с одним или более других агентов в терапии. Например, биспецифическую антигенсвязывающую молекулу по изобретению можно вводить совместно по меньшей мере с одним дополнительным терапевтическим агентом. Термин "терапевтический агент" включает любой агент, вводимый для лечения симптома или заболевания у индивидуума, нуждающегося в таком лечении. Такой дополнительный терапевтический агент может включать любые активные ингредиенты, подходящие для конкретного показания, подлежащего лечению, предпочтительно обладающие комплементарными активностями, не оказывающими вредного влияния друг на друга. В некоторых формах осуществления дополнительный терапевтический агент представляет собой иммуномодуляторный агент, цитостатический агент, ингибитор клеточной адгезии, цитотоксический агент, активатор клеточного апоптоза или агент, повышающий чувствительность клеток к индукторам апоптоза. В конкретной форме осуществления дополнительный терапевтический агент представляет собой противораковый агент, например, дезинтегратор микротрубочек, антиметаболит, ингибитор топоизомеразы, интеркалятор ДНК, алкилирующий агент, гормональную терапию, ингибитор киназы, антагонист рецептора, активатор апоптоза опухолевых клеток или антиангиогенный агент.
Такие другие агенты целесообразно присутствуют в комбинации в количествах, эффективных для предназначенной цели. Эффективное количество таких других агентов зависит от применяемого количества биспецифической антигенсвязывающей молекулы, от типа расстройства или лечения и от других факторов, обсуждаемых выше. Биспецифические антигенсвязывающие молекулы обычно применяют в таких же дозировках и путях введения, как раскрыто в данной работе, либо приблизительно от 1 до 99% дозировок, раскрытых в данной работе, либо в любой дозировке и любым путем, эмпирически/клинически определенным как подходящий.
Такие комбинированные терапии, отмеченные выше, включают комбинированное введение (где два или более терапевтических агентов включено в одну и ту же или в отдельные композиции) и отдельное введение, в случае которого введение биспецифической антигенсвязывающей молекулы по изобретению может происходить до, во время и/или после введения дополнительного терапевтического агента и/или адъюванта. Биспецифические антигенсвязывающие молекулы по изобретению можно также применять в комбинации с радиотерапией.
Изделия
В другом аспекте изобретения предложено изделие, содержащее вещества, полезные для лечения, предупреждения и/или диагностики расстройств, раскрытых выше. Изделие включает контейнер и этикетку или инструкцию по применению на контейнере или в сочетании с контейнером. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты для внутривенных (IV) растворов и т.д. Контейнеры могут быть сформованы из разнообразных материалов, таких как стекло или полимер. Контейнер удерживает композицию, которая находится в нем сама по себе или в комбинации с другой композицией, эффективной для лечения, предупреждения и/или диагностики состояния, и может иметь стерильный входной канал (например, контейнер может представлять собой пакет для внутривенного раствора или флакон, имеющий пробку, пробиваемую подкожной инъекционной иглой). По меньшей мере один активный агент в композиции представляет собой биспецифическую антигенсвязывающую молекулу по изобретению. На этикетке или в инструкции по применению указано, что композицию применяют для лечения выбранного состояния. Кроме того, изделие может включать следующие изделия: (а) первый контейнер с содержащейся в нем композицией, где композиция содержит биспецифическую антигенсвязывающую молекулу по изобретению; и (b) второй контейнер с содержащейся в нем композицией, где композиция содержит дополнительный цитотоксический или иной терапевтический агент. Изделие в данной форме осуществления изобретения может дополнительно включать инструкцию по применению, в которой указано, что композиции можно применять для лечения конкретного состояния. Альтернативно или дополнительно изделие может дополнительно включать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI; от англ. "bacteriostatic water for injection"), фосфатно-солевой буферный раствор, раствор Рингера и раствор декстрозы. Он может дополнительно включать другие материалы, желательные с коммерческой и потребительской точки зрения, включающие другие буферы, разбавители, фильтры, иглы и шприцы.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее приведены примеры способов и композиций по изобретению. Понятно, что с учетом общего описания, приведенного выше, можно выполнять на практике различные другие формы осуществления.
Общие способы
Методы рекомбинантных ДНК
Для манипуляции с ДНК использовали стандартные способы, описанные в руководстве Sambrook et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Молекулярно-биологические реагенты использовали согласно инструкциям изготовителя. Общая информация, касающаяся нуклеотидных последовательностей легких и тяжелых цепей иммуноглобулинов человека, приведена в книге Kabat, E.A. et al., (1991) Sequences of Proteins of Immunological Interest, 5th ed., NIH Publication No. 91-3242.
Секвенирование ДНК
Последовательности ДНК определяли путем двунитевого секвенирования.
Генный синтез
При необходимости желаемые генные сегменты были либо созданы с помощью ПЦР, используя соответствующие матрицы, либо синтезированы с помощью набора Geneart AG (Regensburg, Germany) из синтетических олигонуклеотидов и продуктов ПЦР с помощью автоматизированного генного синтеза. В случаях, где точная последовательность гена была недоступна, олигонуклеотидные праймеры были сконструированы на основании последовательностей из ближайших гомологов, и гены были выделены с помощью обратной транскрипции с полимеразной цепной реакцией (ОТ-ПЦР) из РНК, имеющей происхождение из соответствующей ткани. Генные сегменты, фланкированные уникальными сайтами расщепления эндонуклеазой рестрикции, были клонированы в стандартных клонирующих/секвенирующих векторах. Плазмидную ДНК очищали из трансформированных бактерий, и концентрацию определяли с помощью УФ спектроскопии. Последовательность ДНК субклонированных генных фрагментов подтверждали секвенированием ДНК. Генные сегменты конструировали с подходящими сайтами рестрикции, чтобы дать возможность субклонирования в соответствующих экспрессионных векторах. Все конструкции конструировали с 5'-концевой последовательностью ДНК, кодирующей лидерный пептид, направляющий секрецию белков в эукариотических клетках. Иллюстративные лидерные пептиды представлены в SEQ ID NO:74-76.
Выделение первичных общих Т-клеток человека из МКПК
Мононуклеарные клетки периферической крови (МКПК) были получены с помощью центрифугирования в градиенте плотности Histopaque из обогащенных препаратов лейкоцитов (лейкоцитарных пленок), полученных из местных банков крови или из свежей крови человека от здоровых доноров. Кратко, кровь разводили стерильным фосфатно-солевым буферным раствором (ФСБ) и осторожно наслаивали на градиент Histopaque (Sigma, H8889). После центрифугирования в течение 30 минут при 450 × g при комнатной температуре (при отключенном торможении) часть плазмы выше интерфазы, содержащей МКПК, отбрасывали. МКПК переносили в новые пробирки Falcon на 50 мл, и эти пробирки дополняли ФСБ до общего объема 50 мл; Смесь центрифугировали при комнатной температуре в течение 10 минут при 400 × g (при отключенном торможении). Супернатант отбрасывали, и осадок МКПК дважды промывали стерильным ФСБ (стадии центрифугирования при 4°С в течение 10 при 350 × g). Полученную в результате популяцию МКПК считали автоматически (ViCell) и хранили в среде RPMI1640, содержащей 10% фетальную сыворотку теленка (ФСТ) и 1% L-аланил-L-глутамин (Biochrom, K0302) при 37°С, 5% CO2 в термостате до начала анализа.
Обогащение Т-клетками из МКПК выполняли, используя набор для выделения Т-клеток Pan T-Cell Isolation Kit II (Miltenyi Biotec #130-091-156) согласно инструкциям изготовителя. Кратко, осадки клеток разводили в 40 мкл холодного буфера на 10 млн. клеток (ФСБ, содержащий 0,5% бычий сывороточный альбумин (БСА), 2 мМ ЭДТА, стерилизованный фильтрованием) и инкубировали с 10 мкл смеси биотин-антитело на 10 млн. клеток в течение 10 мин при 4°С. Добавляли 30 мкл холодного буфера и 20 мкл антибиотиновых магнитных гранул на 10 млн. клеток, и смесь инкубировали еще в течение 15 мин при 4°С. Клетки промывали путем добавления 10-20х действительного объема и последующей стадии центрифугирования при 300 × g в течение 10 мин. Вплоть до 100 млн. клеток ресуспендировали в 500 мкл буфера. Магнитное отделение немеченых общих Т-клеток человека выполняли, используя колонки LS (Miltenyi Biotec #130-042-401) согласно инструкциям изготовителя. Полученную в результате популяцию Т-клеток считали автоматически (ViCell) и хранили в среде AIM-V при 37°С, 5% CO2 в термостате до начала анализа (не дольше 24 ч).
Выделение первичных необученных Т-клеток человека из МКПК
Мононуклеарные клетки периферической крови (МКПК) были получены с помощью центрифугирования в градиенте плотности Histopaque из обогащенных препаратов лейкоцитов (лейкоцитарных пленок), полученных из местных банков крови или из свежей крови человека от здоровых доноров. Обогащение Т-клетками из МКПК выполняли, используя набор для выделения необученных Т-клеток Naive CD8+ T-Cell isolation Kit от фирмы Miltenyi Biotec (#130-093-244) согласно инструкциям изготовителя, но пропуская последнюю стадию выделения CD8+ Т-клеток (см. также описание выделения первичных общих Т-клеток человека).
Выделение первичных МКПК яванского макака из гепаринизированной крови
Мононуклеарные клетки периферической крови (МКПК) были получены с помощью центрифугирования в градиенте плотности из свежей крови яванских макак от здоровых доноров, как описано ниже: гепаринизированную кровь разводили 1:3 стерильным ФСБ, и среду Lymphoprep (Axon Lab #1114545) разводили до 90% стерильным ФСБ. Два объема разведенной крови наслаивали на один объем разведенного градиента плотности, и фракцию МКПК отделяли центрифугированием в течение 30 мин при 520 × g, без торможения, при комнатной температуре. Полосу МКПК переносили в новую пробирку Falcon на 50 мл и промывали стерильным ФСБ путем центрифугирования в течение 10 мин при 400 × g при 4°С. Одно низкоскоростное центрифугирование проводили, чтобы удалить тромбоциты (15 мин при 150 × g, 4°С), и полученную в результате популяцию МКПК автоматически считали (ViCell) и немедленно использовали для дальнейших анализов.
Клетки-мишени
Для оценки MCSP-направленных биспецифических антигенсвязывающих молекул использовали следующие линии опухолевых клеток: линию клеток меланомы человека WM266-4 (АТСС #CRL-1676), выделенную из метастатического сайта злокачественной меланомы и экспресирующую высокие уровни MCSP человека; и линию клеток меланомы человека MV-3 (любезно предоставленную Медицинским Центром Неймегенского университета (The Radboud University Nijmegen Medical Centre)), экспресирующую средние уровни MCSP человека.
Для оценки СЕА-направленных биспецифических антигенсвязывающих молекул использовали следующие линии опухолевых клеток: линию клеток рака желудка человека MKN45 (DSMZ #АСС 409), экспресирующую высокие уровни СЕА человека; линию клеток карциномы ободочной кишки женщин европеоидной расы LS-174T (ЕСАСС #87060401), экспресирующую от средних до низких уровней СЕА человека; линию клеток эпителиоидной карциномы поджелудочной железы человека Panc-1 (АТСС #CRL-1469), экспресирующую (очень) низкие уровни СЕА человека; и линию клеток карциномы ободочной кишки мыши МС38-huCEA, сконструированную в лаборатории для стабильной экспрессии СЕА человека.
Кроме того, линию Т-клеточной лейкемии человека Jurkat (ATCC #TIB-152) использовали для оценки связывания различных биспецифических конструкций на клетках CD3 человека.
Пример 1
Получение, очистка и характеризация биспецифических антигенсвязывающих молекул
Последовательности ДНК вариабельной области тяжелой и легкой цепи субклонировали в рамке считывания либо с константной тяжелой цепью, либо с вариабельной тяжелой цепью, предварительно встроенной в соответствующий экспрессионный вектор-реципиент млекопитающих. Экспрессия антитела была направлена промотором MPSV, и синтетическая последовательность полиА расположена на 3'-конце CDS. Кроме того, каждый вектор содержал последовательность EBV OriP.
Молекулы продуцировали путем котрансфекции клеток HEK293 EBNA экспрессионными векторами млекопитающих. Экспоненциально растущие клетки HEK293 EBNA трансфицировали, используя кальций-фосфатный способ. Альтернативно клетки HEK293 EBNA, растущие в суспензии, трансфицировали, используя полиэтиленимин (ПЭИ). Для получения конструкций "1+1 IgG Crossfab" клетки трансфицировали соответствующими экспрессионными векторами в отношении 1:1:1:1 ("вектор второй тяжелой цепи": "вектор первой легкой цепи": "вектор легкой цепи Crossfab": "вектор первой тяжелой цепи - тяжелой цепи Crossfab"). Для получения конструкций "2+1 IgG Crossfab" клетки трансфицировали соответствующими экспрессионными векторами в отношении 1:2:1:1 ("вектор второй тяжелой цепи": "вектор легкой цепи": "вектор первой тяжелой цепи-тяжелой цепи Crossfab": "вектор легкой цепи Crossfab". Для получения конструкции "2+1 IgG Crossfab (N-концевой), сшитая легкая цепь" клетки трансфицировали соответствующими экспрессионными векторами в отношении 1:1:1:1 ("вектор тяжелой цепи": "вектор легкой цепи": "вектор тяжелой цепи (CrossFab-Fab-Fc)": "вектор сшитой легкой цепи"). Для получения конструкции "1+1 CrossMab" клетки трансфицировали соответствующими экспрессионными векторами в отношении 1:1:1:1 ("вектор первой тяжелой цепи": "вектор второй тяжелой цепи": "вектор первой легкой цепи": "вектор второй легкой цепи").
Для трансфекции с использованием фосфата кальция клетки выращивали в виде прикрепленных монослойных культур в Т-флаконах, используя культуральную среду, представляющую собой модифицированную Дульбекко среду Игла (DMEM; от англ. Dulbecco Modified Eagle Medium) с добавлением 10% (об/об) ФСТ, и трансфицировали при достижении ими от 50 до 80% конфлюентности. Для трансфекции флакона Т150 15 млн. клеток высевали за 24 часа до трансфекции в 25 мл культуры в среде DMEM с добавлением ФСТ (при конечном объеме 10% об/об), и клетки помещали в термостат при 37°С в атмосферу 5% СО2 на ночь. Для каждого трансфицируемого флакона Т150 готовили раствор ДНК, CaCl2 и воды путем смешивания 94 мкг суммарной ДНК плазмидных векторов, разделенной в соответствующем отношении, воды до конечного объема 469 мкл и 469 мкл 1 М раствора CaCl2. К этому раствору добавляли 938 мкл раствора 50 мМ ГЭПЭС, 280 мМ NaCl, 1,5 мМ Na2HPO4 при рН 7,05, сразу смешивали в течение 10 с и оставляли стоять при комнатной температуре в течение 20 с. Суспензию разводили 10 мл DMEM с добавлением 2% (об/об) ФСТ и добавляли в Т150 вместо существующей среды. Затем добавляли дополнительно 13 мл среды для трансфекции. Клетки инкубировали при 37°С, 5% CO2 в течение приблизительно от 17 до 20 часов, затем среду заменяли 25 мл DMEM, 10% ФСТ. Кондиционированную культуральную среду собирали примерно через 7 суток после замены среды путем центрифугирования в течение 15 мин при 210 × g, стерильно фильтровали (фильтр 0,22 мкм) с добавлением азида натрия до конечной концентрации, составляющей 0,01% (масс/об) и хранили при 4°С.
Для трансфекции с использованием полиэтиленимина (ПЭИ) клетки HEK293 EBNA культивировали в суспензии в среде CD CHO без сыворотки. Для продуцирования во встряхиваемых колбах на 500 мл 400 млн. клеток HEK293 EBNA высевали за 24 часа до трансфекции. Для трансфекции клетки центрифугировали в течение 5 мин при 210 × g, и супернатант заменяли 20 мл подогретой среды CD CHO. Экспрессионные векторы смешивали в 20 мл среды CD CHO до конечного количества ДНК, составляющего 200 мкг. После добавления 540 мкл ПЭИ смесь перемешивали на вортексе в течение 15 с, а затем инкубировали в течение 10 мин при комнатной температуре. Затем клетки смешивали с раствором ДНК/ПЭИ, переносили во встряхиваемую колбу на 500 мл и инкубировали в течение 3 часов при 37°С в термостате с атмосферой 5% СО2. После периода инкубации добавляли 160 мл среды F17, и клетки культивировали в течение 24 часов. Через одни сутки после трансфекции добавляли 1 мМ вальпроевую кислоту и 7% Добавку 1 (Lonza). После культивирования в течение 7 суток супернатант собирали для очистки путем центрифугирования в течение 15 мин при 210 × g, раствор стерильно фильтровали (фильтр 0,22 мкм), добавляли азид натрия до конечной концентрации 0,01% масс/об и держали при 4°С.
Секретируемые белки очищали из супернатантов клеточных культур аффинной хроматографией на белке А с последующей стадией эксклюзионной хроматографии.
Для аффинной хроматографии супернатант наносили на колонку HiTrap ProteinA HP (объем колонки (CV; от англ. column volume) = 5 мл, GE Healthcare), уравновешенную 25 мл раствора 20 мМ фосфата натрия, 20 мМ цитрата натрия, рН 7,5, или 40 мл раствора 20 мМ фосфата натрия, 20 мМ цитрата натрия, 0,5 М хлорида натрия, рН 7,5. Несвязанный белок удаляли путем промывания по меньшей мере десятью объемами колонки раствора 20 мМ фосфата натрия, 20 мМ цитрата натрия, 0,5 М хлорида натрия рН 7,5 с последующей стадией дополнительной отмывки с использованием шести объемов колонки раствора 10 мМ фосфата натрия, 20 мМ цитрата натрия, 0,5 М хлорида натрия рН 5,45. затем колонку промывали 20 мл раствора 10 мМ морфолиноэтансульфоновой кислоты (МЭС), 100 мМ хлорида натрия, рН 5,0, и целевой белок элюировали в шести объемах колонки раствора 20 мМ цитрата натрия, 100 мМ хлорида натрия, 100 мМ глицина, рН 3,0. Альтернативно целевой белок элюировали, используя градиент на 20 объемов колонки от 20 мМ цитрата натрия, 0,5 М хлорида натрия, рН 7,5, до 20 мМ цитрата натрия, 0,5 М хлорида натрия, рН 2,5. Раствор белка нейтрализовали добавлением 1/10 0,5 М фосфата натрия, рН 8. Целевой белок концентрировали и фильтровали, после чего наносили на колонку HiLoad Superdex 200 (GE Healthcare), уравновешенную раствором 25 мМ фосфата калия, 125 мМ хлорида натрия, 100 мМ глицина, рН 6,7. Для очистки 1+1 IgG Crossfab колонку уравновешивали раствором 20 мМ гистидина, 140 мМ хлорида натрия, рН 6,0.
Концентрацию белка очищенных образцов белка определяли путем измерения оптической плотности (OD; от англ. "optical density") при 280 нм, используя коэффициент молярной экстинкции, вычисленный на основании аминокислотной последовательности. Чистоту и молекулярную массу биспецифических конструкций анализировали с помощью электрофореза в ДСН-ПААГ в присутствии и в отсутствие восстанавливающего агента (5 мМ 1,4-дитиотрейтола) и окрашивали Кумасси (SimpleBlue™ SafeStain от Invitrogen), используя гель системы NuPAGE® Pre-Cast (Invitrogen, США) согласно инструкциям изготовителя (4-12% трис-ацетатные гели или 4-12% Бис-Трис). Альтернативно чистоту и молекулярную массу молекул анализировали с помощью анализов КЭ-ДСН в присутствии и в отсутствие восстанавливающего агента, используя систему Caliper LabChip GXII (Caliper Lifescience) согласно инструкциям изготовителя.
Содержание агрегатов в образцах белка анализировали, используя колонку для аналитической эксклюзионной хроматографии Superdex 200 10/300GL (GE Healthcare) в подвижном буфере 2 мМ MOPS, 150 мМ NaCl, 0,02% (масс/об) NaN3, рН 7,3 при 25°С. Альтернативно содержание агрегатов в образцах антитела анализировали, используя колонку для аналитической эксклюзионной хроматографии TSKgel G3000 SW XL (Tosoh) в подвижном буфере 25 мМ K2HPO4, 125 мМ NaCl, 200 мМ L-аргинина моногидрохлорид, 0,02% (масс/об) NaN3, рН 6,7 при 25°С.
Конструкция "2+1 IgG Crossfab (С-концевой)" (см. SEQ ID NO:11, 12, 13 и 14) была получена с использованием плазмидных векторов, имеющих геномную организацию генов, включающих интронные последовательности, промотор CMV и сигнал полиаденилирования из гена гормона роста быка. Биспецифическую конструкцию транзитно экспрессировали в клетках HEK293F путем одновременной трансфекции необходимыми плазмидами посредством липофекции, используя различные отношения плазмид. Клетки выращивали в среде F17. Супернатант собирали через 5-7 суток после трансфекции. Перед очисткой собранный супернатант клеточной культуры стерильно фильтровали через мембрану с размером пор 0,2 мкм (Millipore). Для очистки биспецифическую молекулу иммобилизовали на смоле MabSelectSure (GE Healthcare), промывали 1x ФСБ и элюировали 20 мМ цитратом натрия при рН 3,0. Молекулу дополнительно очищали эксклюзионной хроматографией, используя колонку Superdex™ 200 GL (Amersham Bioscience), уравновешенную раствором 20 мМ гистидина, 140 мМ NaCl, рН 6,0. Характеризацию (оценку целостности антитела) биспецифической молекулы выполняли, используя анализ с помощью капиллярного электрофореза (КЭ-ДСН), используя микрожидкостную технологию Labchip (Caliper). Для анализа КЭ-ДСН готовили 5 мкл раствора белка, используя набор НТ Protein Express Reagent Kit согласно инструкциям изготовителя, и анализировали на системе LabChip GXII, используя микрочип НТ Protein Express Chip.
На фиг.2-5 представлены результаты электрофореза в ДСН-ПААГ и аналитической эксклюзионной хроматографии, и в таблице 2 представлены выходы, содержание агрегатов после белка А и конечное содержание мономеров в препаратах различных биспецифических конструкций. Важно, что биспецифические конструкции IgG Crossfab по настоящему изобретению проявляли содержание агрегатов, сниженное в 10-20 раз, после аффинной хроматографии на белке А по сравнению с соответствующими биспецифическими конструкциями, содержащими одноцепочечный Fab фрагмент вместо фрагмента Crossfab (данные не представлены).
На фиг.13 показан результат анализов в КЭ-ДСН биспецифической конструкции анти-CD3/анти-MCSP "2+1 IgG Crossfab (N-концевой), конструкции "сшитая легкая цепь" (см. SEQ ID NO:1, 4, 5 и 85). Для анализов использовали 2 мкг образца. На фиг.14 показан результат аналитической эксклюзионной хроматографии конечного продукта (нанесение 20 мкг образца).
На фиг.20 показаны результаты анализов КЭ-ДСН следующих конструкций: 1+1 IgG Crossfab (N-концевой); обмен VL/VH (LC007/V9), 1+1 CrossMab; обмен CL/CH1 (LC007/V9), 2+1 IgG Crossfab (N-концевой), инвертированный; обмен CL/CH1 (LC007/V9), 2+1 IgG Crossfab (N-концевой); обмен VL/VH (M4-3 ML2/V9), 2+1 IgG Crossfab (N-концевой); обмен CL/CH1 (M4-3 ML2/V9) и 2+1 IgG Crossfab (N-концевой), инвертированный; обмен CL/CH1 (CH1A1A/V9), и в таблице 2 показаны выходы, содержание агрегатов после белка А и конечное содержание мономеров препаратов различных биспецифических конструкций.
Пример 2
Одновременное связывание биспецифических конструкций с обоими антигенами-мишенями
Одновременное связывание конструкции "2+1 IgG Crossfab (N-концевой)" (SEQ ID NO:1, 3, 4, 5) с MCSP человека и с CD3e человека анализировали с помощью поверхностного плазменного резонанса (фиг.6).
Все эксперименты поверхностного плазменного резонанса (SPR) выполняли на приборе Biacore T100 при 25°C с буфером HBS-EP в качестве подвижного буфера (0,01 М ГЭПЭС рН 7,4, 0,15 М NaCl, 3 мМ ЭДТА, 0,005% сурфактант Р20, Biacore, Freiburg/Germany).
Анализ одновременного связывания биспецифической конструкции с опухолевым антигеном и с CD3ε человека выполняли путем прямого сочетания 1650 резонансных единиц (RU; resonance units) биотинилированного домена D3 MCSP на сенсорном чипе SA, используя стандартный метод сочетания. Постановка анализа показана на фиг.6А.
Конструкцию "2+1 IgG Crossfab (N-концевой)" иммобилизовали в течение 60 с при 200 нМ. Затем конструкцию CD3γ(G4S)5CD3ε-AcTev-Fc(выступ)-Avi/Fc(впадина) человека пропускали при концентрации 2000 нМ и скорости тока 40 мкл/мин в течение 60 с. Общие различия показателя преломления корректировали путем вычитания ответа, полученного на референсной проточной ячейке, где рекомбинантный CD3ε пропускали над поверхностью иммобилизованного домена D3 MCSP без иммобилизованной биспецифической конструкции.
Как показано на фиг.6В, эта конструкция была способна связывать опухолевый антиген и CD3 одновременно. Уровень связывания (RU) после нанесения CD3ε человека был выше, чем уровень связывания, достигнутый после нанесения одной конструкции, что отражает, что оба антигена, опухолевый антиген и CD3ε человека, связаны с биспецифической конструкцией.
Пример 3
Т-клеточная активация биспецифической конструкцией в присутствии и в отсутствие клеток-мишеней
Высвобождение цитокинов
Очищенную конструкцию "2+1 IgG Crossfab (N-концевой)" (SEQ ID NO:1, 3, 4, 5) и молекулу "(scFv)2", где обе конструкции направлены на MCSP человека и CD3 человека, анализировали на их способность индуцировать опосредованную Т-клетками секрецию цитокинов de novo в присутствии и в отсутствие опухолевых клеток-мишеней.
Кратко, помещали 280 мкл цельной крови от здорового донора на лунку 96-луночного планшета с глубокими лунками. 30000 опухолевых клеток-мишеней Colo-38, экспрессирующих MCSP человека, а также две биспецифические конструкции и контроли IgG добавляли при конечной концентрации 1 нМ. Клетки инкубировали в течение 24 ч при 37°С, 5% СО2, а затем центрифугировали в течение 5 мин при 350 × g. супернатант переносили в новый 96-луночный планшет с глубокими лунками для последующего анализа. Анализ СВА проводили в соответствии с инструкциями изготовителя для FACS Cantoll, используя комбинацию следующих наборов СВА FlexSet: гранзим В человека (BD #560304), интерферон-γ (ИФН) человека FlexSet (BD #558269), фактор некроза опухоли (ФНО) человека FlexSet (BD #558273), ИЛ-10 человека FlexSet (BD #558274), ИЛ-6 человека FlexSet (BD #558276), ИЛ-4 человека FlexSet (BD #558272), ИЛ-2 человека FlexSet (BD #558270).
На фиг.7 показаны уровни различных цитокинов, измеренные в супернатанте. Основным цитокином, секретируемым в присутствии опухолевых клеток Colo-38, был ИЛ-6, за которым следовал ИФН-γ. Кроме того, уровни гранзима В также сильно повысились после активации Т-клеток в присутствии клеток-мишеней. В целом обе биспецифические конструкции индуцировали высокие уровни секреции цитокинов в присутствии клеток-мишеней (фиг.7, А и В), но не индуцировали ее в отсутствие клеток-мишеней (фиг.7, С и D). Значительная секреция цитокинов Th2 (ИЛ-10 и ИЛ-4) отсутствовала после активации Т-клеток в присутствии (или в отсутствие) клеток-мишеней.
Экспрессия поверхностный маркеров активации
В другом эксперименте очищенные конструкции "2+1 IgG Crossfab (N-концевой)" (SEQ ID NO:4, 5, 6, 7), направленные на CD3 яванского макака и MCSP человека, анализировали на их потенциал повышающей регуляции поверхностного маркера активации CD25 на CD8+ Т-клетках в присутствии опухолевых клеток-мишеней. Кратко, опухолевые клетки-мишени, экспрессирующие MCSP человека, собирали буфером для диссоциации клеток, промывали и ресуспендировали в среде DMEM, содержащей 2% ФСТ и 1% GlutaMax. 30000 клеток на лунку высевали в 96-луночный планшет с круглым дном и добавляли разведение соответствующего антитела в указанных концентрациях (фиг.8). Молярность биспецифических конструкций и различных контролей IgG регулировали до одинакового значения. Эффекторные клетки МКПК яванского макака, выделенные из крови двух здоровых животных, добавляли до получения конечного отношения Е:Т 3:1. После инкубации в течение 43 ч при 37°С, 5% CO2 клетки центрифугировали при 350 × g в течение 5 мин и дважды промывали ФСБ, содержащим 0,1% БСА. Поверхностное окрашивание на CD8 (Miltenyi Biotech #130-080-601) и CD25 (BD #557138) выполняли согласно предложениям поставщика. Клетки дважды промывали 150 мкл/лунка ФСБ, содержащего 0,1% БСА, и фиксировали в течение 15 мин при 4°С, используя 100 мкл/лунка буфера фиксации (BD #554655). После центрифугирования образцы ресуспендировали в 200 мкл/лунка ФСБ с 0,1% БСА и анализировали, используя прибор FACS Cantoll (Software FACS Diva).
Как изображено на фиг.8, биспецифическая конструкция индуцирует зависимую от концентрации повышающую регуляцию CD25 на CD8+ Т-клетках только в присутствии клеток-мишеней. IgG против CD3 яванского макака (клон FN-18) также способно индуцировать повышающую регуляцию CD25 на CD8+ Т-клетках без перекрестного связывания с опухолевыми клетками-мишенями (см. данные, полученные с яванским макаком Нестором). Гиперактивация Т-клеток яванского макака при максимальной концентрации биспецифической конструкции (в отсутствие клеток-мишеней) отсутствовала.
В другом эксперименте конструкцию CD3-MCSP "2+1 IgG Crossfab (N-концевой), сшитая легкая цепь" (см. SEQ ID NO:1, 4, 5 и 85) сравнивали с конструкцией CD3-MCSP "2+1 IgG Crossfab (N-концевой)" (см. SEQ ID NO:1, 3, 4 и 5) на их потенциал повышающей регуляции раннего маркера активации CD69 или позднего маркера активации CD25 на CD8+ Т-клетках в присутствии опухолевых клеток-мишеней. Первичные МКПК человека (выделенные, как описано выше) инкубировали с указанными концентрациями биспецифических конструкций в течение по меньшей мере 22 ч в присутствии или в отсутствие MCSP-положительных клеток-мишеней Colo38. Кратко, 0,3 млн. первичных МКПК человека на лунку высевали в 96-луночный планшет с плоским дном, содержащий MCSP-положительные клетки-мишени (или среду). Конечное отношение эффектора к клетке-мишени (Е:Т) составляло 10:1. Клетки инкубировали с указанными концентрациями биспецифических конструкций и контролей в течение указанных периодов инкубации при 37°С, 5% CO2. Эффекторные клетки окрашивали на CD8 и CD69 или CD25 и анализировали с помощью FACS Cantoll.
На фиг.19 показан результат этого эксперимента. Значимые различия, обнаруженные для повышающей регуляции CD69 (А) или CD25 (В) между двумя молекулами 2+1 IgG Crossfab (со сшитой легкой цепью или без сшитой легкой цепи) отсутствовали.
Еще в одном другом эксперименте конструкции CD3-MCSP "2+1 IgG Crossfab (N-концевой)" (см. SEQ ID NO:1, 3, 4, 5) и "1+1 IgG Crossfab (N-концевой)" (см. SEQ ID NO:1, 3, 4, 86) сравнивали с биспецифической IgG-подобной конструкцией CD3-MCSP, в который один антигенсвязывающий фрагмент замещен фрагментом Crossfab ("1+1 CrossMab"; см. SEQ ID NO:4, 87, 88, 89) на их потенциал повышающей регуляции CD69 или CD25 на CD4+ или CD8+ Т-клетках в присутствии опухолевых клеток-мишеней. Анализ выполняли, как описано выше, в присутствии или в отсутствие опухолевых клеток MV-3, экспрессирующих MCSP, причем, период инкубации составлял 24 ч.
Как показано на фиг.21, конструкции "1+1 IgG Crossfab (N-концевой)" и "2+1 IgG Crossfab (N-концевой)" индуцировали более выраженную повышающую регуляцию маркеров активации, чем молекула "1+1 CrossMab".
Пример 4
Перенаправленная Т-клеточная цитотоксичность, опосредованная перекрестно связанными биспецифическими конструкциями, направленными на CD3 на Т-клетках и на MCSP на опухолевых клетках (анализ высвобождения ЛДГ)
Биспецифические конструкции анализировали на их потенциал индукции Т-клеточно-опосредованного апоптоза в опухолевых клетках-мишенях при перекрестном связывании конструкции посредством связывания антигенсвязывающих фрагментов с их соответствующими антигенами-мишенями на клетках.
В одной форме осуществления сравнивали конструкцию "2+1 IgG Crossfab (N-концевой)" (SEQ ID NO:1, 3, 4, 5), направленную на CD3 человека и MCSP человека, и соответствующую молекулу "(scFv)2". Кратко, клетки-мишени меланомы человека MDA-MB-435, экспрессирующие huMCSP, собирали буфером диссоциации клеток, промывали и ресуспендировали в среде AIM-V (Invitrogen #12055-091). 30000 клеток на лунку высевали в 96-луночный планшет с круглым дном и добавляли соответствующее разведение конструкции при указанной концентрации. Молярность всех конструкций и контролей регулировали до одинаковых значений. Эффекторные общие Т-клетки человека добавляли до получения конечного отношения Е:Т, составляющего 5:1. В качестве положительного контроля на активацию общих Т-клеток человека использовали 1 мкг/мл ФГА-М (Sigma #1-8902). Для нормализации максимальный лизис клеток-мишеней (=100%) определяли путем инкубации клеток-мишеней при конечной концентрации 1% Тритон Х-100. Минимальный лизис (=0%) относится к клеткам-мишеням, совместно инкубируемым с эффекторными клетками, но в отсутствие какой-либо конструкции или антитела. После ночной инкубации в течение 20 ч при 37°С, 5% CO2, высвобождение ЛДГ апоптических/некротических клеток-мишеней в супернатант измеряли с помощью набора для обнаружения ЛДГ (Roche Applied Science, #11 644 793 001) согласно инструкциям изготовителя.
Как изображено на фиг.9, конструкция "2+1 IgG Crossfab (N-концевой)" индуцирует апоптоз в клетках-мишенях сравнимо с молекулой "(scFv)2".
В следующем эксперименте очищенные конструкции "2+1 IgG Crossfab (N-концевой)" (SEQ ID NO:1, 3, 4, 5), "1+1 IgG Crossfab (N-концевой)" (SEQ ID NO:1, 2, 3, 4) и молекулу "(scFv)2" на их потенциал индукции Т-клеточно-опосредованного апоптоза в опухолевых клетках-мишенях при перекрестном связывании конструкции посредством связывания обоих антигенов-мишеней, CD3 и MCSP, на клетках. В качестве клеток-мишеней использовали клетки меланомы человека MDA-MB-435, экспрессирующие huMCSP, отношение Е:Т составляло 5:1, и время инкубации составляло 20 ч. Результаты представлены на фиг.10. Конструкция "2+1 IgG Crossfab (N-концевой)" индуцирует апоптоз в клетках-мишенях сравнимо с молекулой "(scFv)2". Сравнение моно- и бивалентных форматов "IgG Crossfab (N-концевой)" показывает, что бивалентный формат более эффективен в данном анализе.
Еще в одном другом эксперименте сравнивали очищенную конструкцию "2+1 IgG Crossfab (N-концевой)" (SEQ ID NO:4, 5, 6, 7), направленную на CD3 яванского макака и MCSP человека, и соответствующую конструкцию "(scFv)2", используя в качестве клеток-мишеней линию клеток меланомы человека (MV-3), экспрессирующих MCSP. Кратко, клетки MV-3 собирали буфером диссоциации клеток, промывали и ресуспендировали в среде DMEM, содержащей 2% ФСТ и 1% GlutaMax. 30000 клеток на лунку высевали в 96-луночный планшет с круглым дном и добавляли соответствующее разведение конструкции или референсного IgG при указанных концентрациях. Молярность биспецифической конструкции и различных контролей IgG регулировали до одинаковых значений. Эффекторные клетки МКПК яванского макака, выделенные из крови здоровых яванских макак, добавляли до получения конечного отношения Е:Т, составляющего 10:1. После инкубации в течение 26 ч при 37°С, 5% CO2 высвобождение ЛДГ апоптических/некротических клеток-мишеней в супернатант измеряли с помощью набора для обнаружения ЛДГ (Roche Applied Science, #11 644 793 001) согласно инструкциям изготовителя.
Как изображено на фиг.11, конструкция "2+1 IgG Crossfab (N-концевой)" более эффективна на основании EC50, чем молекула "(scFv)2".
В другой серии экспериментов конструкцию CD3-MCSP "2+1 IgG Crossfab (N-концевой), сшитая легкая цепь" (см. SEQ ID NO:1, 4, 5 и 85) сравнивали с конструкцией CD3-MCSP "2+1 IgG Crossfab (N-концевой)" (см. SEQ ID NO:1, 3, 4 и 5). Кратко, клетки-мишени (Colo-38 человека, MV-3 человека или клетки меланомы WM266-4) собирали буфером диссоциации клеток на сутки анализа (или трипсином за сутки до начала анализа), промывали и ресуспендировали в соответствующей культуральной среде (RPMI1640, включающей 2% ФСТ и 1% Glutamax). 20000-30000 клеток на лунку высевали в 96-луночный планшет с круглым дном и добавляли соответствующее разведение антитела, как указано (три повтора). МКПК в качестве эффекторных клеток добавляли до получения конечного отношения эффекторных клеток к клеткам-мишеням (Е:Т), составляющего 10:1. Молярность всех конструкций и контролей регулировали до одинаковых значений, время инкубации составляло 22 ч. Определение высвобождения ЛДГ и нормализацию выполняли, как описано выше.
На фиг.15-18 показан результат четырех анализов, выполненных с клетками меланомы MV-3 (фиг.15), с клетками Colo-38 (фиг.16 и 17) или с клетками WM266-4 (фиг.18). Как показано на фиг.15, конструкция со сшитой легкой цепью была менее эффективной по сравнению с конструкцией без сшитой легкой цепи в анализе с клетками MV-3 в качестве клеток-мишеней. Как показано на фиг.16 и 17, конструкция со сшитой легкой цепью была более эффективной по сравнению с конструкцией без сшитой легкой цепи в анализах с клетками Colo-38 с высокой экспрессией MCSP в качестве клеток-мишеней. Наконец, как показано на фиг.18, значительное различие между двумя конструкциями отсутствовало при использовании клеток WM266-4 с высокой экспрессией MCSP в качестве клеток-мишеней.
В другом эксперименте сравнивали две СЕА-направленные конструкции "2+1 IgG Crossfab (N-концевой), инвертированный", в которых во фрагменте Crossfab был проведен обмен либо областей V (VL/VH, см. SEQ ID NO:3, 8, 9, 10), либо областей С (CL/CH1, см. SEQ ID NO:9, 10, 87, 95). Анализ проводили, как описано выше, используя МКПК человека в качестве эффекторных клеток и клетки-мишени, экспрессирующие СЕА человека. Клетки-мишени (опухолевые клетки MKN-45 или LS-174T) собирали трипсином-ЭДТА (LuBiosciences #25300-096), промывали и ресуспендировали в среде RPMI1640 (Invitrogen #42404042), включающей 1% Glutamax (LuBiosciences #35050087) и 2% ФСТ. 30000 клеток на лунку высевали в 96-луночный планшет с круглым дном и добавляли биспецифические конструкции при указанных концентрациях. Молярность всех конструкций и контролей регулировали до одинаковых значений. Эффекторные клетки МКПК человека добавляли до получения конечного отношения Е:Т, составляющего 10:1, время инкубации составляло 28 ч. Значения EC50 вычисляли, используя программу GraphPad Prism 5.
Как показано на фиг.22, конструкция с обменом CL/CH1 проявляет несколько лучшую активность на обеих линиях клеток-мишеней, чем конструкция с обменом VL/VH. Вычисленные значения ЕС50 составляли 115 и 243 пМ на клетках MKN-45 и 673 и 955 пМ на клетках LS-174T для конструкции с обменом CL/CH1 и конструкции с обменом VL/VH соответственно.
Аналогично сравнивали две MCSP-направленные конструкции "2+1 IgG Crossfab (N-концевой)", в которых во фрагменте Crossfab был проведен обмен либо областей V (VL/VH, см. SEQ ID NO:3, 91, 92, 93), либо областей С (CL/CH1, см. SEQ ID NO:87, 91, 93, 94). Анализ проводили, как описано выше, используя МКПК человека в качестве эффекторных клеток и клетки-мишени, экспрессирующие MCSP человека. Клетки-мишени (WM266-4) собирали буфером диссоциации клеток (LuBiosciences #13151014), промывали и ресуспендировали в среде RPMI1640 (Invitrogen #42404042), включающей 1% Glutamax (LuBiosciences #35050087) и 2% ФСТ. 30000 клеток на лунку высевали в 96-луночный планшет с круглым дном и добавляли конструкции при указанных концентрациях. Молярность всех конструкций и контролей регулировали до одинаковых значений. Эффекторные клетки МКПК человека добавляли до получения конечного отношения Е:Т, составляющего 10:1, время инкубации составляло 26 ч. Значения ЕС50 вычисляли, используя программу GraphPad Prism 5.
Как изображено на фиг.23, две конструкции показывают сравнимую активность, причем, конструкция с обменом CL/CH1 имеет несколько более низкое значение ЕС50 (12,9 пМ для конструкции с обменом CL/CH1 по сравнению с 16,8 пМ для конструкции с обменом VL/VH).
На фиг.24 показан результат аналогичного анализа, проведенного с клетками-мишенями MV-3, экспрессирующими MCSP человека. Опять же, обе конструкции показывают сравнимую активность, причем, конструкция с обменом CL/CH1 имеет несколько более низкое значение EC50 (приблизительно 11,7 пМ для конструкции с обменом CL/CH1 по сравнению с приблизительно 82,2 пМ для конструкции с обменом VL/VH). Точные значения ЕС50 невозможно было вычислить, поскольку кривые цитолиза не достигали плато при высоких концентрациях соединений.
В следующем эксперименте конструкции CD3-MCSP "2+1 IgG Crossfab (N-концевой)" (см. SEQ ID NO:1, 3, 4, 5) и "1+1 IgG Crossfab (N-концевой)" (см. SEQ ID NO:1, 3, 4, 86) сравнивали с конструкцией CD3-MCSP "1+1 CrossMab" (см. SEQ ID NO:4, 87, 88, 89). Анализ проводили, как описано выше, используя МКПК человека в качестве эффекторных клеток и клетки-мишени WM266-4 или MV-3 (отношение Е:Т=10:1) и время инкубации 21 ч.
Как показано на фиг.25, конструкция "2+1 IgG Crossfab (N-концевой)" является наиболее эффективной конструкцией в данном анализе, за которой следует "1+1 IgG Crossfab (N-концевой)" и "1+1 CrossMab". Это ранжирование является даже более выраженным на клетках MV-3, экспрессирующих средние уровни MCSP, по сравнению с клетками WM266-4 с высокой экспрессией MCSP. Вычисленные значения EC50 на клетках MV-3 составляли 9,2, 40,9 и 88,4 пМ, на клетках WM266-4 33,1, 28,4 и 53,9 пМ для конструкций "2+1 IgG Crossfab (N-концевой)", "1+1 IgG Crossfab (N-концевой)" и "1+1 CrossMab" соответственно.
Пример 5
Связывание биспецифических конструкций с соответствующими антигенами-мишенями на клетках
Связывание различных биспецифических конструкций с CD3 на клетках Jurkat (ATCC #TIB-152) и с соответствующими опухолевыми антигенами: MCSP на клетках WM266-4 или СЕА на клетках LS174-T определяли с помощью FACS. Кратко, клетки собирали, считали и проверяли на жизнеспособность. 0,15-0,2 млн. клеток на лунку высевали в 96-луночный планшет с круглым дном и инкубировали с указанной концентрацией биспецифических конструкций и контролей в течение 30 мин при 4°С. Для лучшего сравнения конструкции нормализовали до одинаковой молярности. Клетки промывали один раз ФСБ, содержащим 0,1% БСА. После инкубации с ФИТЦ- или ФЭ-конъюгированным вторым антителом в течение 30 мин при 4°С, связанные конструкции обнаруживали, используя FACS Cantoll (программа FACS Diva). Использовали ФИТЦ- или ФЭ-конъюгированный фрагмент AffiniPure F(ab')2 коза против IgG человека, специфичный к фрагменту Fcγ (Jackson Immuno Research Lab #109-096-098/рабочий раствор 1:20, или #109-116-170/рабочий раствор 1:80, соответственно). Если не указано иное, клетки фиксировали 100 мкл/лунка буфера фиксации (BD #554655) в течение 15 мин при 4°С в темноте, центрифугировали в течение 6 мин при 400 × g и хранили в 200 мкл/лунка ФСБ, содержащего 0,1% БСА, до анализа. Значения ЕС50 вычисляли, используя программу GraphPad Prism 5.
На фиг.26 показано связывание биспецифических конструкций CD3/CEA "2+1 IgG Crossfab (N-концевой), инвертированный" либо с обменом VL/VH (см. SEQ ID NO:3, 8, 9, 10), либо с обменом CL/CH1 (см. SEQ ID NO:9, 10, 87, 95) во фрагменте Crossfab, c CD3 человека, экспрессируемым клетками Jurkat, или с СЕА человека, экспрессируемым клетками LS-174T. В качестве контроля также оценивали эквивалентную максимальную концентрацию соответствующих IgG и фоновое окрашивание за счет меченого второго антитела (ФИТЦ-конъюгированного фрагмента AffiniPure F(ab')2 коза против человека, специфичного к фрагменту Fcγ, Jackson Immuno Research Lab #109-096-098). Обе конструкции проявляли высокое связывание как с СЕА человека, так и с CD3 человека на клетках. Вычисленные значения ЕС50 составляли 4,6 и 3,9 нМ (CD3) и 9,3 и 6,7 нМ (СЕА) для конструкций "2+1 IgG Crossfab (N-концевой), инвертированный (VL/VH)" и "2+1 IgG Crossfab (N-концевой), инвертированный (CL/CH1)", соответственно.
На фиг.27 показано связывание конструкций CD3/MCSP "2+1 IgG Crossfab (N-концевой)" (см. SEQ ID NO:1, 3, 4, 5) и "2+1 IgG Crossfab (N-концевой), инвертированный" (см. SEQ ID NO:4, 87, 89, 90) с CD3 человека, экспрессируемым клетками Jurkat, или с MCSP человека, экспрессируемым клетками WM266-4. Хотя связывание обеих конструкций с MCSP на клетках было сравнимо высоким, связывание "инвертированной" конструкции CD3 было снижено по сравнению с другой конструкцией. Вычисленные значения ЕС50 составляли 6,1 и 1,66 нМ (CD3) и 0,57 и 0,95 нМ (MCSP) для конструкций "2+1 IgG Crossfab (N-концевой), инвертированный" и "2+1 IgG Crossfab (N-концевой)", соответственно.
Хотя описанное выше изобретение раскрыто в некоторых подробностях в качестве иллюстрации и примера в целях ясности понимания, описания и примеры не следует истолковывать как ограничивающие объем изобретения. Описания всей патентной и научной литературы, цитируемой в данной работе, полностью явным образом включены посредством ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К АНТИГЕНАМ, АКТИВИРУЮЩИМ Т-КЛЕТКИ, И ОПУХОЛЕВОМУ АНТИГЕНУ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2605390C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2019 |
|
RU2797268C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С HLA-A2/WT1 | 2018 |
|
RU2815176C2 |
| НОВЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, ОБЛАДАЮЩИЕ СПОСОБНОСТЬЮ СПЕЦИФИЧЕСКИ СВЯЗЫВАТЬСЯ С CD40 И FAP | 2018 |
|
RU2766234C2 |
| АНТИТЕЛА, НЕ СОДЕРЖАЩИЕ FC-ФРАГМЕНТА, ВКЛЮЧАЮЩИЕ ДВА FAB-ФРАГМЕНТА, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2617970C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ К PD1 И TIM3 | 2016 |
|
RU2729371C1 |
| АКТИВИРУЕМЫЕ ПРОТЕАЗОЙ СВЯЗЫВАЮЩИЕ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИЕ МОЛЕКУЛЫ | 2017 |
|
RU2830288C2 |

















































































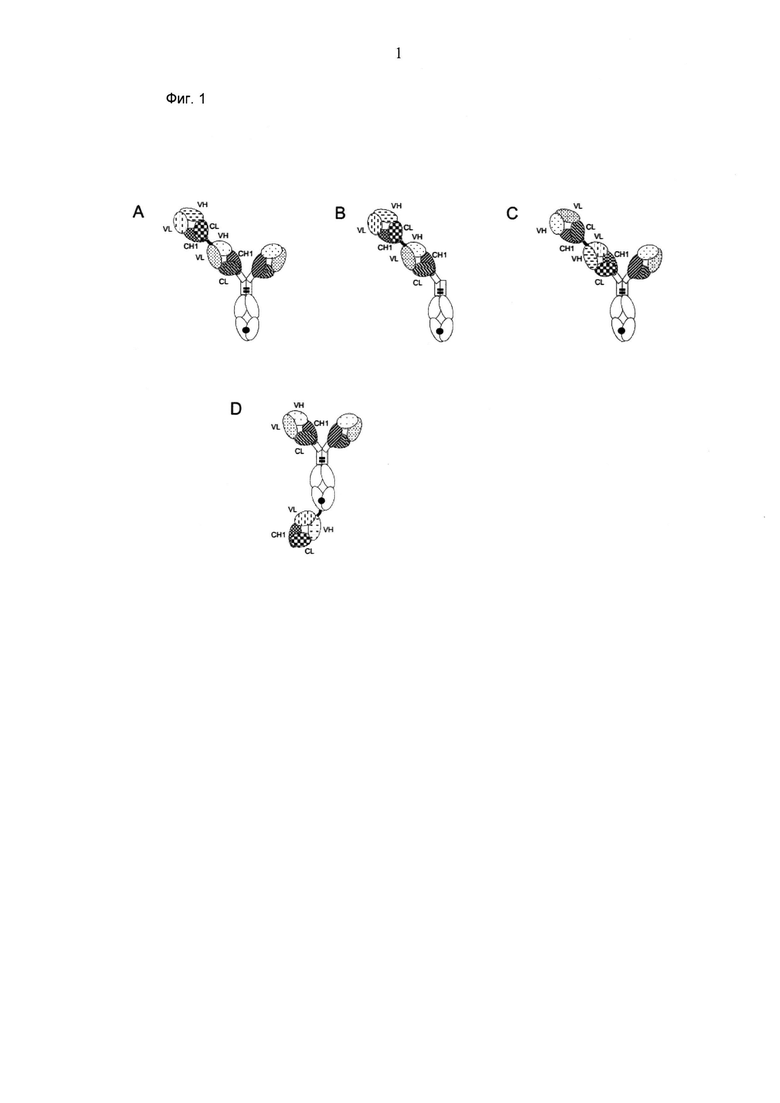
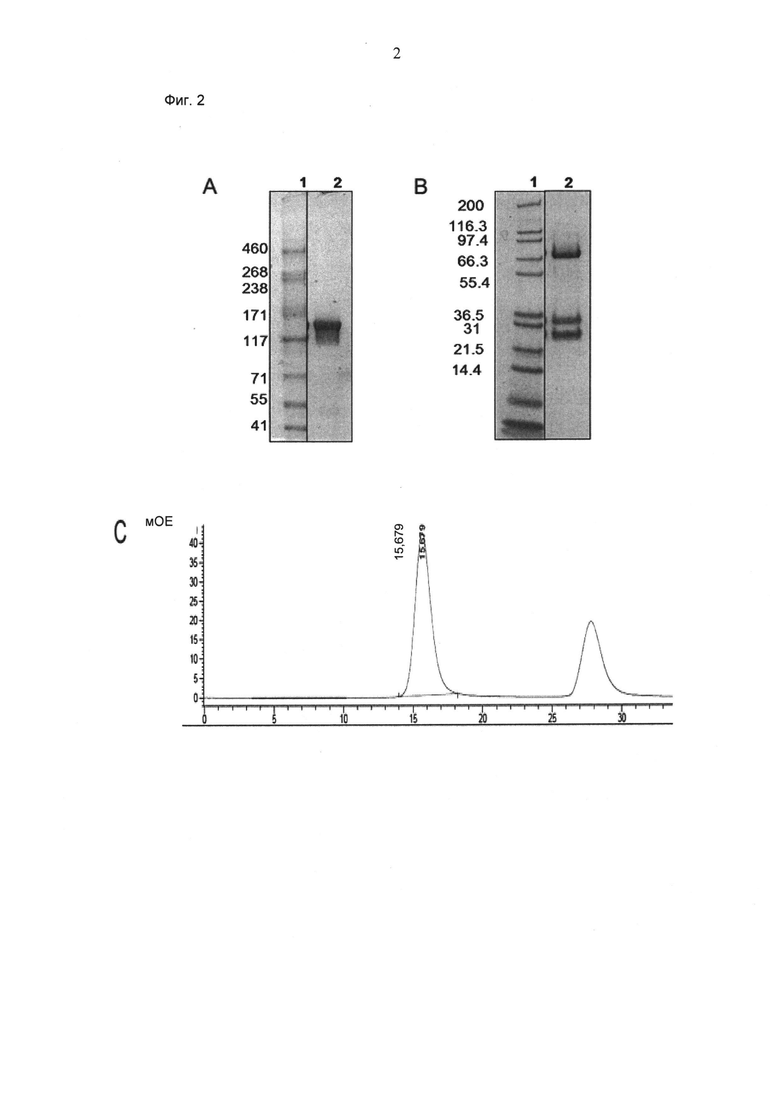

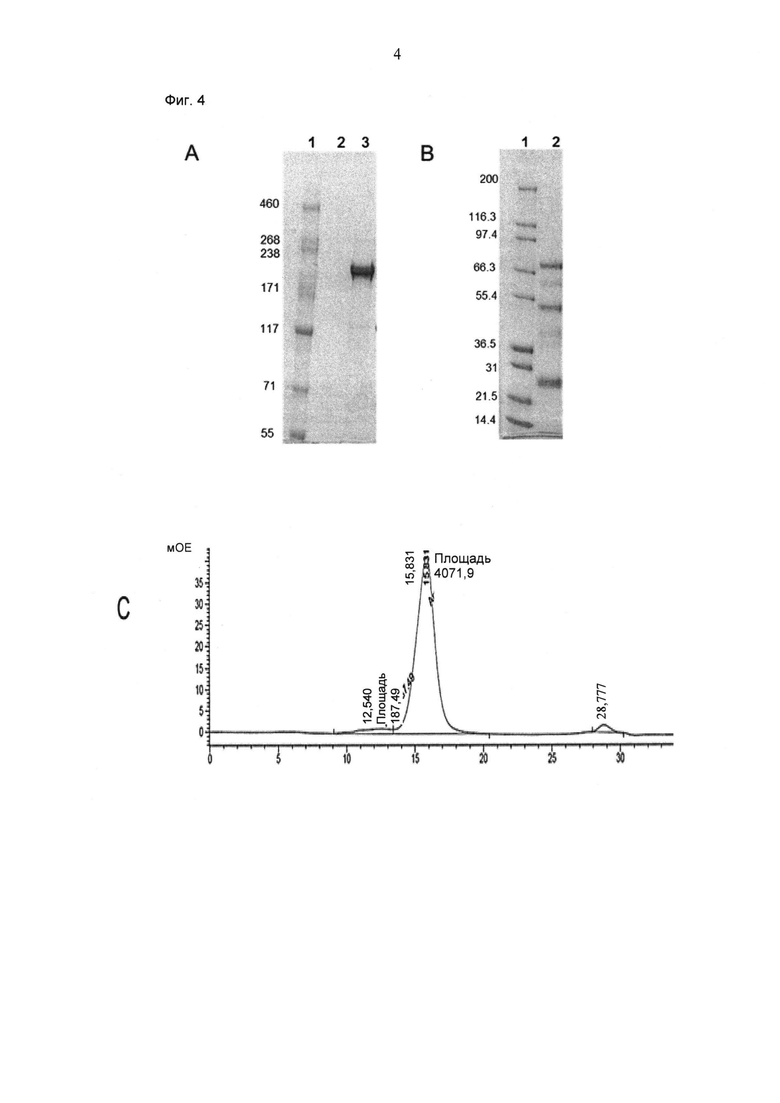



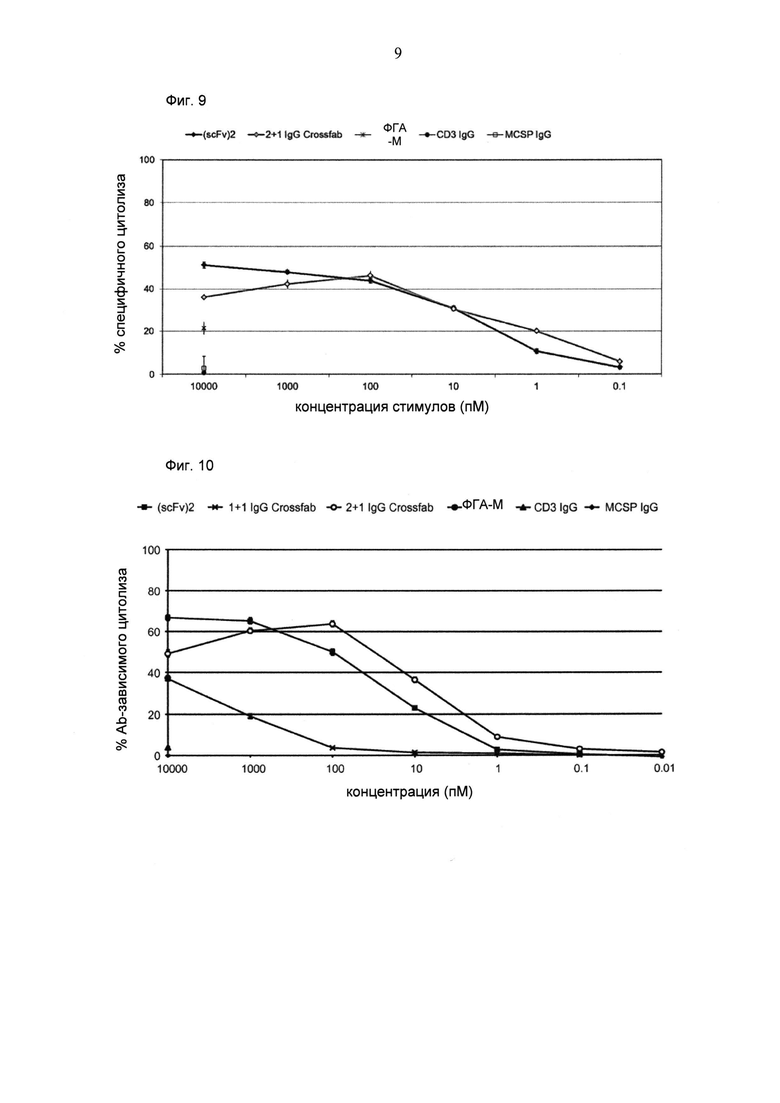
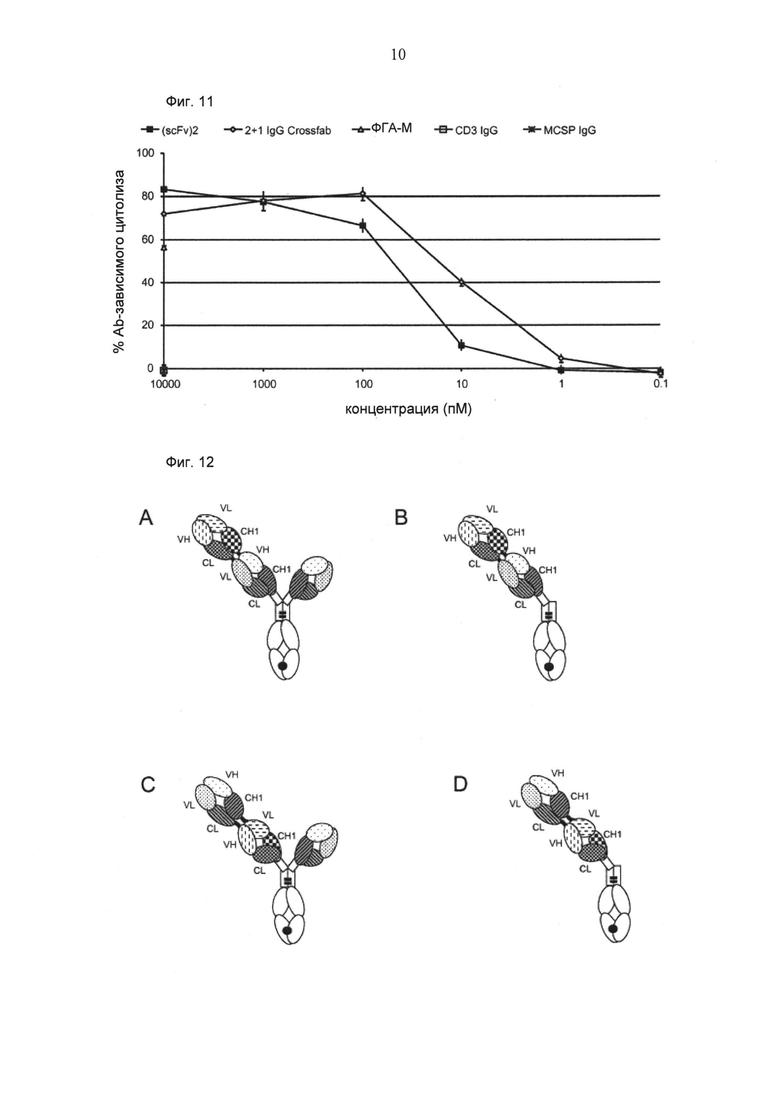

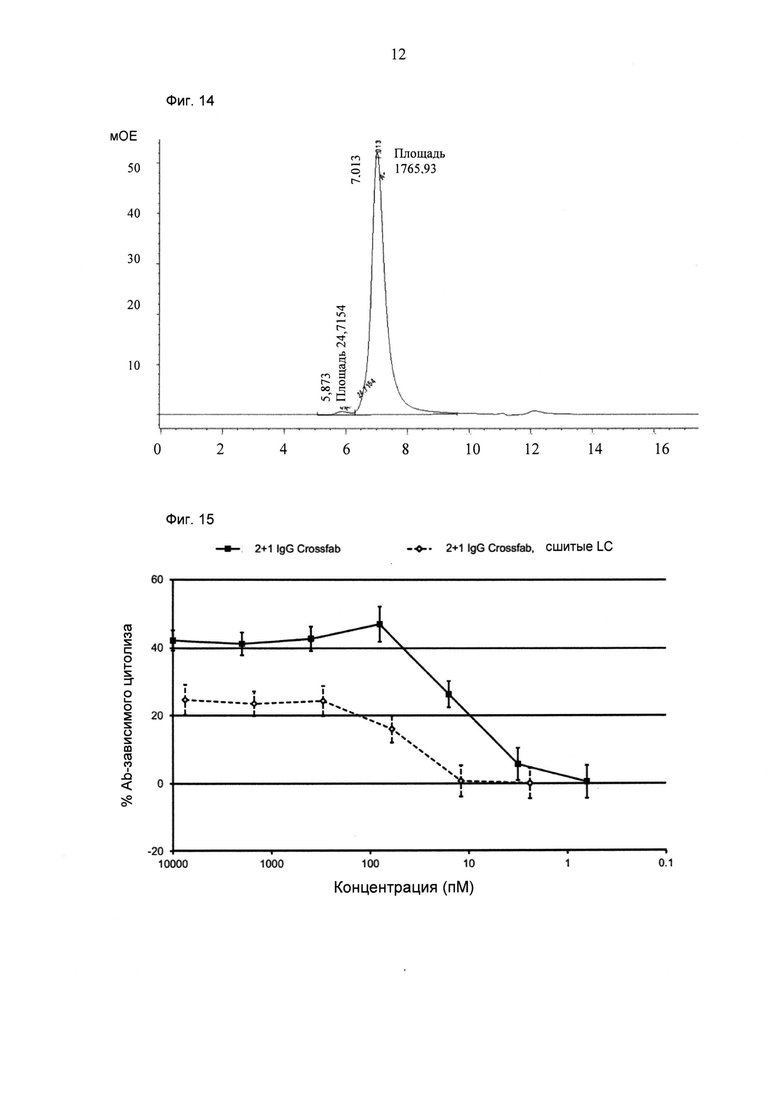

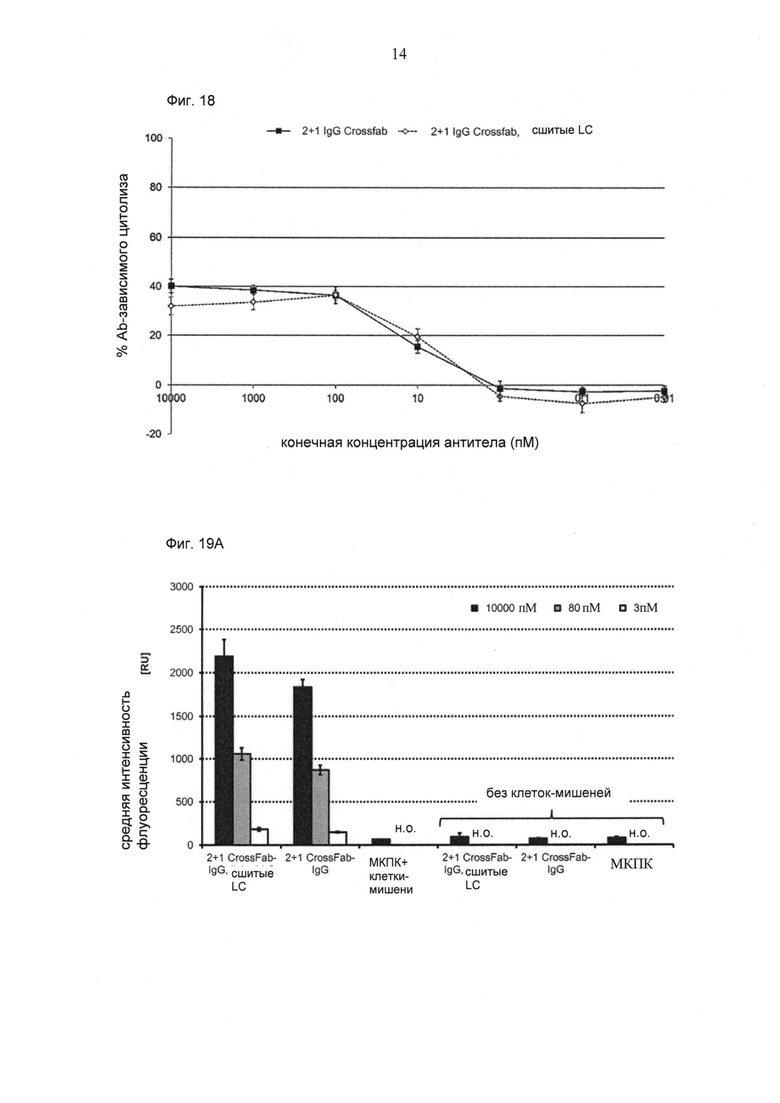

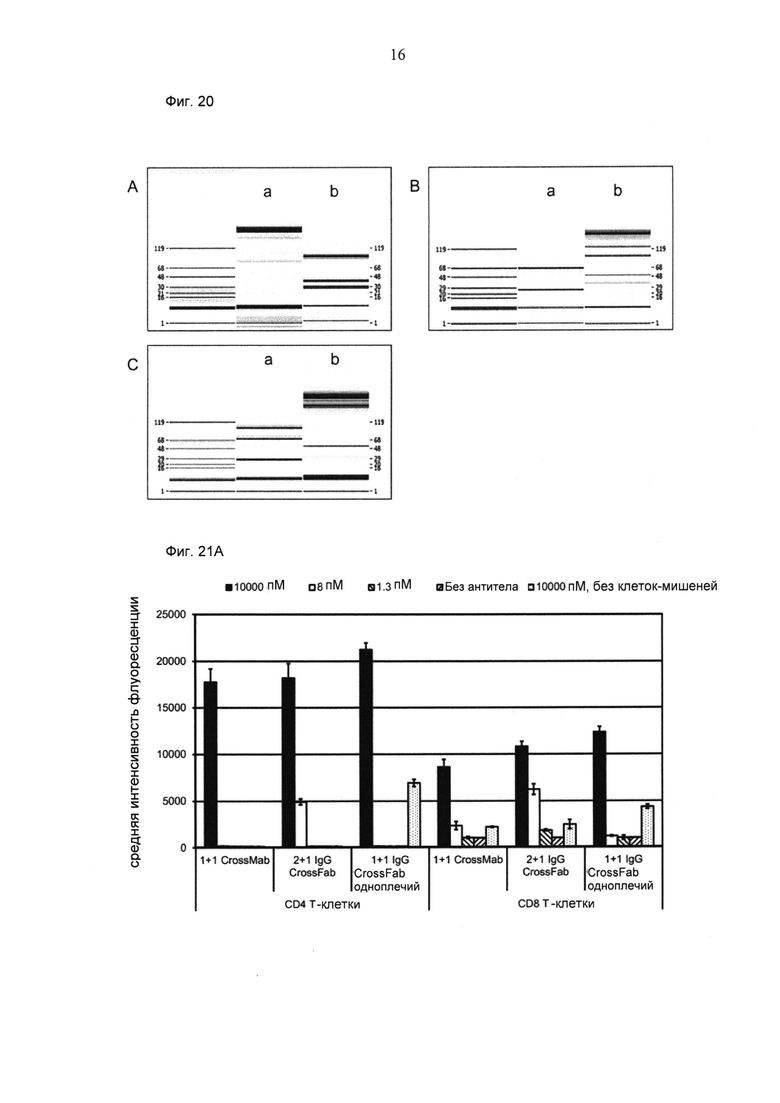

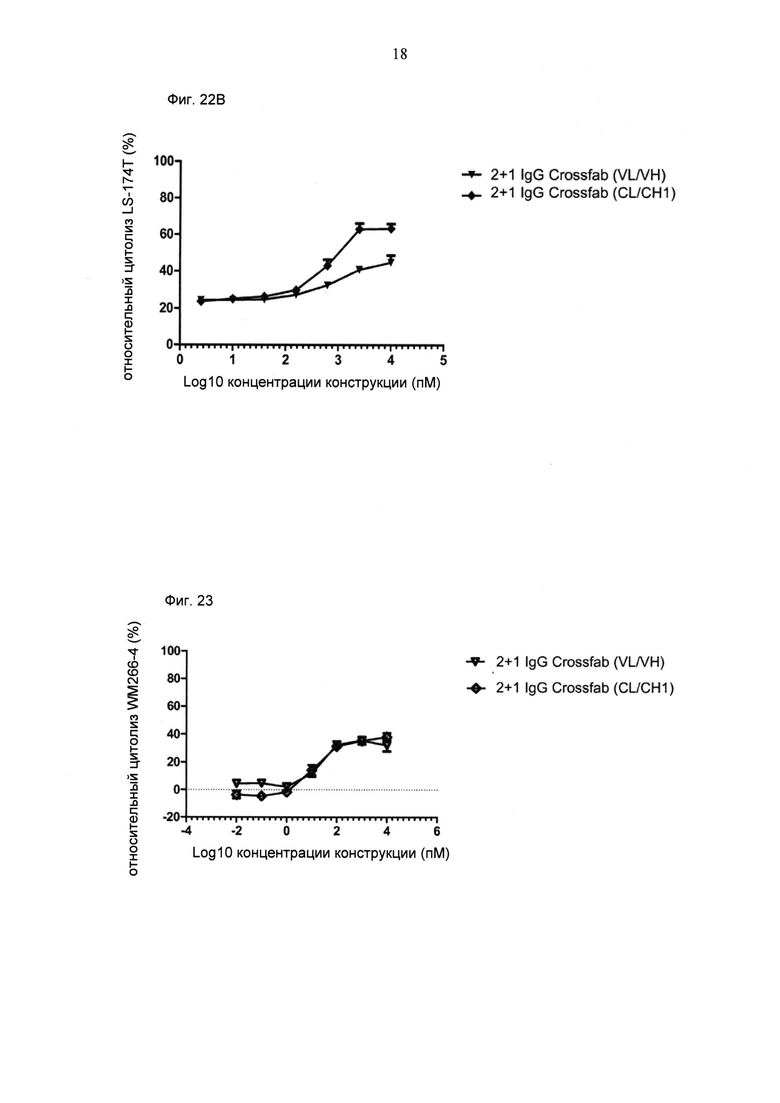
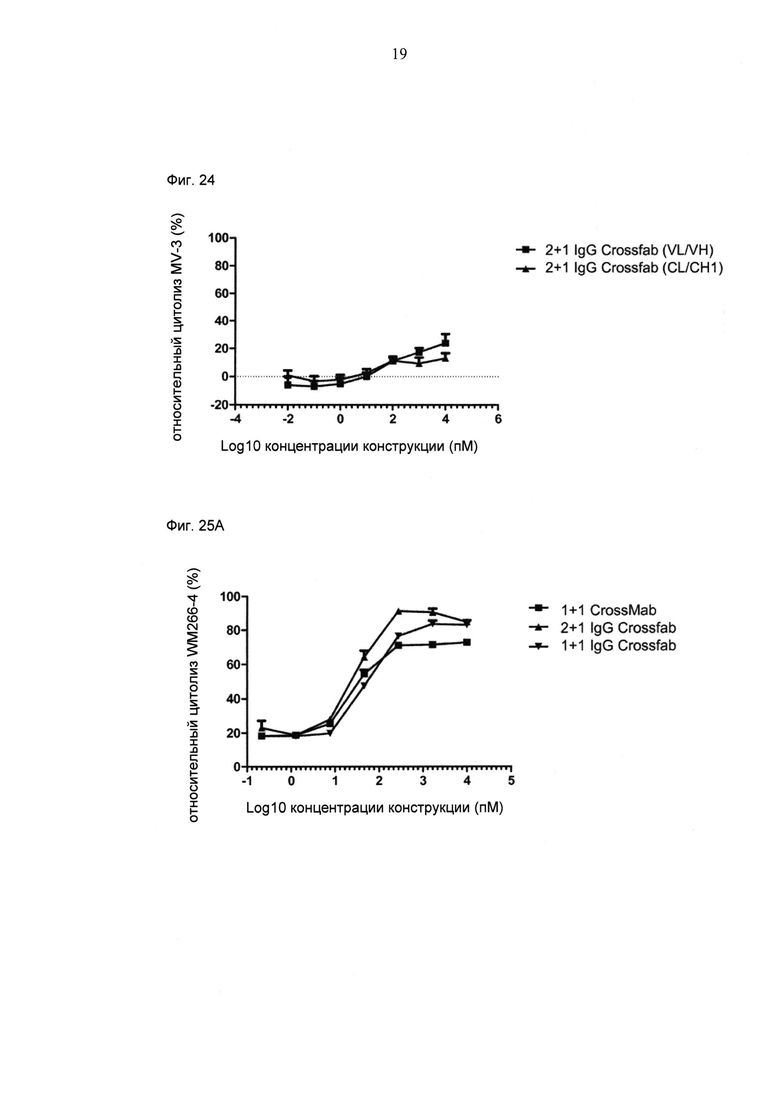



Настоящее изобретение относится к иммунологии. Предложен способ получения биспецифической антигенсвязывающей молекулы, включающей Fc домен и Fab фрагменты, специфично связывающиеся с первым и вторым антигенами, причем биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым и/или вторым антигенами. Один из Fab в составе указанной молекулы слит на С-конце своей тяжелой цепи с N-концом тяжелой цепи другого Fab фрагмента, который, в свою очередь, слит на С-конце своей тяжелой цепи с N-концом субъединицы домена Fc. При этом по меньшей мере в одном из Fab производят одну из следующих замен: (i) вариабельные домены легкой цепи (VL) и тяжелой цепи (VH) заменяют друг другом, или (ii) константный домен легкой цепи (CL) и первый константный домен тяжелой цепи (СН) СН1 заменяют друг другом, при условии, что в первом и втором Fab фрагментах производят не одну и ту же замену. Кроме того, домен Fc в составе указанной молекулы включает модификацию, стимулирующую ассоциацию субъединиц домена Fc. Данное изобретение позволяет получать биспецифические антигенсвязывающие молекулы с улучшенной активностью и может найти применение в получении терапевтических конструкций на основе антител. 24 з.п. ф-лы, 27 ил., 2 табл., 5 пр.
1. Способ получения биспецифической антигенсвязывающей молекулы, включающей первый Fab фрагмент, специфично связывающийся с первым антигеном, второй Fab фрагмент, специфично связывающийся со вторым антигеном, и домен Fc, состоящий из первой и второй субъединиц, способных к стабильной ассоциации; причем:
a) биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым и/или вторым антигенами,
b) (i) первый Fab фрагмент сливают на С-конце его тяжелой цепи с N-концом тяжелой цепи второго Fab фрагмента, который, в свою очередь, сливают на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc, или (ii) второй Fab фрагмент сливают на С-конце его тяжелой цепи с N-концом тяжелой цепи первого Fab фрагмента, который, в свою очередь, сливают на С-конце его тяжелой цепи с N-концом первой субъединицы домена Fc,
c) в первом и/или втором Fab фрагментах производят одну из следующих замен: (i) вариабельные домены легкой цепи (VL) и тяжелой цепи (VH) заменяют друг другом, или (ii) константный домен легкой цепи (CL) и первый константный домен тяжелой цепи (СН) СН1 заменяют друг другом, при условии, что в первом и втором Fab фрагментах производят не одну и ту же замену; и
d) где домен Fc включает модификацию, стимулирующую ассоциацию первой и второй субъединиц домена Fc;
при этом способ включает стадии а) культивирования клетки-хозяина, содержащей один или более изолированный полинуклеотид, кодирующий биспецифическую антигенсвязывающую молекулу, в условиях, подходящих для экспрессии биспецифической антигенсвязывающей молекулы, и b) выделение биспецифической антигенсвязывающей молекулы.
2. Способ по п. 1, где дополнительно легкую цепь первого Fab фрагмента и легкую цепь второго Fab фрагмента сливают друг с другом, необязательно посредством пептидного линкера.
3. Способ по п. 1, где производят замену в первом Fab фрагменте.
4. Способ по п. 1, где замена представляет собой замену вариабельных доменов VL и VH друг другом.
5. Способ по п. 1, где замена представляет собой замену константных доменов CL и СН1 друг другом.
6. Способ по п. 1, где биспецифическая антигенсвязывающая молекула по существу состоит из первого Fab фрагмента, второго Fab фрагмента, домена Fc и необязательно одного или нескольких пептидных линкеров.
7. Способ по п. 1, где биспецифическая антигенсвязывающая молекула включает третий Fab фрагмент, специфично связывающийся с первым или вторым антигеном.
8. Способ по п. 7, где третий Fab фрагмент сливают со второй субъединицей домена Fc.
9. Способ по п. 7 или 8, где третий Fab фрагмент сливают на его С-конце с N-концом второй субъединицы домена Fc.
10. Способ по п. 9, где третий Fab фрагмент сливают на С-конце его тяжелой цепи с N-концом второй субъединицы домена Fc.
11. Способ по п. 7, где третий Fab фрагмент специфично связывается со вторым антигеном.
12. Способ по п. 7, где второй Fab фрагмент, третий Fab фрагмент и домен Fc составляют часть молекулы иммуноглобулина.
13. Способ по п. 12, где молекула иммуноглобулина представляет собой молекулу иммуноглобулина класса IgG.
14. Способ по п. 12 или 13, где молекула иммуноглобулина представляет собой молекулу иммуноглобулина подкласса IgG1 или IgG4.
15. Способ по п. 12, где молекула иммуноглобулина представляет собой молекулу иммуноглобулина человека.
16. Способ по п. 12, где биспецифическая антигенсвязывающая молекула по существу состоит из первого Fab фрагмента, специфично связывающегося с первым антигеном, молекулы иммуноглобулина, специфично связывающейся со вторым антигеном, и необязательно одного или нескольких пептидных линкеров.
17. Способ по п. 1, где в Fab фрагментах, специфично связывающихся с одним и тем же антигеном, производят одну и ту же замену.
18. Способ по п. 1, где биспецифическая антигенсвязывающая молекула обеспечивает моновалентное связывание с первым антигеном.
19. Способ по п. 1, где производят замену только в первом Fab фрагменте.
20. Способ по п. 1, где биспецифическая антигенсвязывающая молекула не включает одноцепочечный Fab фрагмент.
21. Способ по п. 1, где в домене СН3 первой субъединицы домена Fc аминокислотный остаток заменяют аминокислотным остатком, имеющим больший объем боковой цепи, создавая посредством этого выпячивание внутри домена СН3 первой субъединицы, которое может быть расположено в углублении домена СН3 второй субъединицы, и в домене СН3 второй субъединицы домена Fc аминокислотный остаток заменяют аминокислотным остатком, имеющим меньший объем боковой цепи, создавая посредством этого углубление внутри домена СН3 второй субъединицы, внутри которого может быть расположено выпячивание внутри домена СН3 первой субъединицы.
22. Способ по п. 1, где домен Fc представляет собой домен Fc IgG.
23. Способ по п. 22, где домен Fc представляет собой домен Fc IgG1 или IgG4.
24. Способ по п. 1, где домен Fc представляет собой домен Fc человека.
25. Способ по п. 1, где домен Fc конструируют таким образом, что он обладает измененной аффинностью связывания с рецептором Fc и/или измененной эффекторной функцией по сравнению с несконструированным доменом Fc.
| Колосоуборка | 1923 |
|
SU2009A1 |
| SCHAEFER WOLFGANG et al | |||
| Immunoglobulin domain crossover as a generic approach for the production of bispecific IgG antibodies | |||
| Proceedings of the National Academy of Sciences, июнь 2011 г., 108(27): 11187-11192 | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| LIU YANSHUN, EISENBERG DAVID | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Protein science, 2002, 11(6): 1285-1299 | |||
| МОИСЕЕНКО В.М | |||
| Моноклональные антитела в лечении злокачественных опухолей | |||
| Практическая онкология, 2003, т | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

